
Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
The Gram-negative, strictly anaerobic bacterium Megasphaera elsdenii was first isolated from the rumen in 1953 and is common in the mammalian gastrointestinal tract. Its ability to use either lactate or glucose as its major energy source for growth has been well documented, although it can also ferment amino acids to ammonia and branched-chain fatty acids, which are growth factors for other bacteria. The ruminal abundance of M. elsdenii usually increases in animals fed grain-based diets due to its ability to use lactate (the product of rapid ruminal sugar fermentation), especially at low ruminal pH (< 5.5). M. elsdenii has been proposed as a potential dietary probiotic to prevent ruminal acidosis in feedlot cattle and high-producing dairy cows. However, this bacterium has also been associated with milk fat depression (MFD) in dairy cows, although proving a causative role has remained elusive. This review summarizes the unique physiology of this intriguing bacterium, and its role of in ruminal community function and in the health and productivity of the host animal. In addition to its effects in the rumen, the ability of M. elsdenii to produce C2-C7 carboxylic acids – potential precursors for industrial fuel and chemical production, is examined.
Keywords:
carboxylate platform
; lactic acid
; Megasphaera
; rumen
; ruminants
; volatile fatty acids
1. Introduction
Ruminant animals including cattle, sheep, and goats, are characterized by a digestive tract in which the foregut is compartmentalized into four chambers. The first chamber, the rumen, accounts for around 65% of foregut volume and contributes to more than 65% of digestion that occurs in the entire digestive tract of adult animals [1] (p.230). Digestion that takes place in the rumen is exclusively attributed to its microbial community, which contains a high density and high diversity of microbes, estimated at up 109 -to 1011 bacterial cells mL-1, 104 to 106 protozoa mL-1 [2] (p.34) and 102 to 104 fungal zoospores mL-1 [3] of rumen contents. Of particular importance is the digestion of polysaccharides within the plant cell wall, which cannot be digested by enzymes secreted by the animal. Ruminal digestion of feeds results in the production of microbial cells and volatile fatty acids (VFA), which are respectively the major source of amino acids and energy for the host [4,5]. Since Hungate began to study the rumen microbial ecosystem in the 1940s, around 450 bacterial species have been isolated from the rumen [6], but the total number of species is estimated to be > 4000; thus, the majority of rumen bacterial diversity remains uncharacterized.
One ruminal bacterium, Megasphaera elsdenii, has been studied in particular for its capacity to ferment lactate, produced by various sugar-fermenting bacteria in cattle and sheep fed high-grain diets [7], which makes this species potentially important in controlling ruminal acidosis. This in turn has created expectations regarding its potential use as a probiotic. In addition to its importance in rumen microbial ecology, M. elsdenii has also attracted attention for its ability to produce commercially useful, short- and medium- chain carboxylic acids (“volatile fatty acids”, VFA) [8]. The objective of this review is to highlight the principal metabolic aspects and major growth characteristics of M. elsdenii, focusing on its effects on ruminant production and its potential application as an industrial producer of carboxylic acids.
2. Classification and morphology
M. elsdenii is a Gram-negative, strictly anaerobic, coccus-shaped, nonmotile bacterium that inhabits the rumen of the cattle and other ruminants [7], as well as the large intestine of humans [9] and pigs [10]. It is classified within phylum Firmicutes (syn. Bacillota) and class Clostridia [11], although it was previously classified in class Negativicutes. Megasphaera is one of a few genera within its phylum to have a porous pseudo-outer membrane that results in a negative Gram-staining reaction, although details of its lipopolysaccharide structure remain sketchy [12]. Within the genus Megasphaera are included several species: M. elsdenii, M. hominis, M. cerevisiae, M. micronuciformis, M. paucivorans and M. sueciensis, which have been isolated from various sources, including the rumen, human clinical specimens, and spoiled beer [13].
The name Megasphaera originated from the distinctive morphology of this bacterium, i.e., a large cell (mega = Latin for “large”) having a spherical shape (coccus) [14] (Figure 1). Huhtanen and Gall [7], who first described the bacterium now known as M. elsdenii, isolated it from calves or adult cattle fed high grain diets; noted their relatively large size; and termed them as RO-C8 type LC (“large coccus”).
However, the name Megasphaera is a bit misleading: Although M. elsdenii’s cell size (2-2.5 μm diameter) exceeds that of the streptococci and staphylococci (0.7-1.5 μm) known at the time of its first description, it hardly compares to those of the largest coccoid bacteria discovered since, such as Thiovulum majus (18 μm [15]), or the gigantic Thiomargarita namibiensis (750 μm [16]).
Huhtanen and Gall [7] grew M. elsdenii in vitro and observed that it fermented lactate and produced C2-C6 VFA (acetic, propionic, butyric, valeric and caproic), as well as CO2. Subsequently, Elsden et al. [17] isolated a bacterium (also termed the LC organism) from the rumen of sheep and presented similar descriptions of its morphology and physiology. Gutierrez et al. [18] also described a rumen isolate from bloated cattle fed high-grain diets as “LC type” and proposed the name Peptostreptococcus elsdenii. Cells generally presented in pairs and short chains (4 to 8 cells), which produced C2-C5 VFA, CO2 and H2 when fermenting lactate. The authors proposed that P. elsdenii probably was important in stabilizing ruminal chemistry due to its capacity to ferment lactic acid. Later, Rogosa [14] proposed its reclassification from Peptostreptococcus elsdenii to Megasphaera elsdenii, based on differences between the ruminal isolates [17,19] and previously described members of the Gram-positive, actively proteolytic genus Peptostreptococcus, [20]. The LC-1 strain of Gutierrez represents the original lineage of the species’ currently recognized type strain, ATCC 25940 (= DSM 20460).
From an evolutionary perspective, Piknova et al. observed that strains of M. elsdenii isolated from lambs, calf and sheep had highly similar 16S rRNA gene sequences [21]. These authors posited that low genetic variability among M. elsdenii strains suggests a recent divergence from common ancestor. Similarly, Shetty et al. [13] observed that two strains of Megasphaera isolated from human gut showed high similarity in 16S rRNA gene sequence between M. hominis and M. elsdenii. Figure 2 shows a rooted phylogenetic tree with all species of genus Megasphaera, in which all rumen-derived strains (M. elsdenii) clustered together compared to those from monogastric or beverage sources.
At this writing, there are 53 M. elsdenii genomes in various states of assembly in the NCBI database, only three of which have been completed [22,23,24]. One strain of the close relative M. indica has also been fully sequenced. (Table 1).
These genomes contain a single circular chromsome of ~ 2.2 Mbp, encode for ~2200 proteins whose coding regions average ~53 mol % G+C, and lack extrachromosomal elements. The modest genome size is toward the lower end for bacterial genomes (range 0.6-14.3 Mbp) and is among the smallest for phylum Firmicutes, whose genome sizes span 1.5 to 5.5 Mbp [25]. The relatively small genome of M. elsdenii is consistent with its modest catabolic and biosynthetic capacity and its strong symbiotic associations with its host and the host’s microbiota, all of which are factors that have selected for smaller genome size in the bacterial world.
The genes for the type strain identified by Marx et al. [22] encode proteins which can be grouped in subsystems, according to Shetty et al. [13], from which the more abundant are: 1) amino acids and derivatives (273 proteins, 11% of total) ; 2) carbohydrate metabolism (201 proteins, 9%); 3) cofactors, vitamins, prosthetic groups and pigments (141 proteins, 6.4%); 4) protein metabolism (136 proteins, 6%); 5) RNA metabolism (115 proteins, 5%); 6) cell wall and capsule (108 proteins, 4.9%), and 7) DNA metabolism (94 proteins, 4%). Such a distribution reflects its niche specialization. For example, its proportion of genes associated with amino acid metabolism are substantially higher than in the ruminal cellulolytic specialist Fibrobacter succinogenes, which neither imports significant amounts of, nor catabolizes, amino acids [26].
Shetty et al. [13] reported that the genome of the ruminal isolate M. elsdenii DSM 20460 had low similarity to genomes of human gut isolates, where only 252 of the former’s proteins displayed more than 99% similarity those from humans. Additionally, they also found that more than 400 proteins encoded by the genes from human gut isolates were not observed in the ruminal isolate; these include genes related to bile resistance; various sensory and regulatory systems; stress response systems; membrane transporters; and resistance to antibiotics, as well as the presence of diverse and unique sets of Carbohydrate-Active enzymes (CAZymes) amongst these isolates, with a higher collection of CAZymes in the human gut isolates that are not found in rumen isolates’ genomes. The authors attribute these differences to differences in host diet suggestive of host-specific adaptation by these isolates (i.e., a much more extensive fiber degradation in the rumen, carried out mostly be specialist taxa).
3. Abundance in the rumen
Considering that all strains of M. elsdenii isolated from the rumen up to now prefer to ferment lactate rather than glucose or other sugars, and that lactate concentration in the rumen is higher in animals fed high-grain diets due to active starch degradation, the abundance of M. elsdenii in the rumen tends to be higher in starch-fed animals [27,28,29]. Because lactate is almost absent from the rumen of animals fed forage-based diets, M. elsdenii usually is not found, or is detected in much lower abundance, in forage-fed ruminants.
Early studies quantifying the abundance of M. elsdenii in the rumen relied on plate counting on lactate agar media [18,27,30] (Table 2).
In some studies the authors did not measure M. elsdenii numbers, but instead they enumerated total lactate-fermenting bacteria, which the authors assumed mostly comprised M. elsdenii. Mackie and Gilchrist [30] observed that in animals fed forage-based diets (only 10% grain) the total numbers of lactate-fermenting bacteria represented about 0.2% of total cultivable bacteria, while in animals fed high-grain diets (71% grain) lactate-fermenting bacteria represented 22.3% of total cultivable rumen bacteria, which supports the association betweeen lactate producers and lactate fermenters. The involvement of M. elsdenii in ruminal lactate fermentation is supported by observation of a positive relationship between the relative abundances of M. elsdenii and Gram-positive rods (probably lactobacilli) in 8-week-old calves [27] [Hobson et al., 1958]. Similarly, Yang et al. [31] found that, in biofilms formed in the grains of barley and corn incubated in the rumen of dairy cows the lactobacilli were dominant members associated to starch digestion, which were followed by an increased abundance of genus Megasphaera.
Since the development of culture-independent molecular methods, quantification has been achieved using qPCR based on the 16S rRNA gene, and by massive parallel sequencing of 16S rRNA genes in community DNA. While early qPCR quantifications were expressed in cell densities (cells mL-1) [32,33], virtually all recent studies have expressed abundance as “relative abundance” (RA), i.e., as a percentage of total bacterial 16 rRNA gene copies. Such quantifications can be misleading due to the wide variation in copy number of these genes across taxa. All three completed M. elsdenii genomes contain 7 copies of the 16S rRNA gene. Because ~80% of sequenced bacterial genomes have fewer than 7 copies of this gene [34], using 16S rRNA gene copy numbers in community analysis will likely overestimate M. elsdenii abundance. The extent of this overestimation is unclear because the average copy number of the bacterial community in the rumen is unknown. Nevertheless, 16S-based quantification can be useful for comparison of abundances across animals and treatments, particularly within individual studies.
Table 3 presents M. elsdenii abundance in the rumen of animals determined by these culture-independent methods [35,36,37,38,39,40,41,42,43]. Relative abundance (RA) in the rumen is typically well under 1% of total bacteria, except in animals experiencing metabolic or production problems, particularly milk fat depression. However, the single study of the ruminal bacterial transcriptome [43] yielded much higher RA than did parallel 16S gene sequencing methods, suggesting that M. elsdenii may be more metabolically active than its low 16S RNA-gene based RA values would suggest.
Several studies [44,45,46] have compared population sizes of M. elsdenii relative to a baseline or control treatment, without an indication of abundance relative to the total bacterial community. Fernando et al. [44] reported an 11-fold increase in M. elsdenii RA in beef steers within 21 d of switching from a hay diet to a concentate diet. Khafipour et al. [45] reported that in Holstein cows, induction of mild or severe subacute ruminal acidosis (SARA) by grain feeding increased the RA of M. elsdenii 16- and ~1000-fold, respectively, while induction of mild SARA by alfalfa pellet feeding actually decreased its RA approximately 16-fold. However, McCann et al. [46] observed no significant change in M. elsdenii RA upon SARA induction. He et al. [47] reported a 2-fold increase in the relative abundance of M. elsdenii in non-lactating Saanen goats under SARA (elicited by high-concentrate feeding) compared to herdmates not subjected to SARA induction.
4. Isolation and cultivation
Despite claims to the contrary [48], enrichment and isolation of M. esldenii is rather straightforward. Considering that this species is a lactate fermenter, ruminal samples from animals fed grain-based diets are the recommended environmental source for enrichment. Use of lactate as energy source for enrichment and isolation is recommended, based on the bacterium’s preference for lactate as well as the fact that (unlike glucose) lactate is fermented by few species of ruminal bacteria. Media such as LTY (lactate/trypticase/yeast extract medium) provides sources of energy, amino acids and vitamins [49]. Because another ruminal bacterium, Selenomonas ruminantium var. lactilytica, also can ferment lactate, selection of M. elsdenii is facilitated by addition of monensin, to which the former species is much more sensitive. Because M. elsdenii produces H2 [27,49], addition of NaMoO4 plus 1,8-dihydroxyanthraquinone and 0.01 mM 2-bromoethanesulfonic acid plus chloroform, will inhibit the growth of symbiotic hydrogenotrophic sulfate-reducers and methanogens, respectively [50].
Unlike most ruminal bacteria, M. elsdenii does not require a highly negative environmental redox potential. As a result, it is not necessary to add chemical reducing agents such as cysteine to an anoxic culture medium, as long as requirements for sulfur are met by addition of trypticase and/or yeast extract; in their absence, small (sub-millimolar) amounts of cysteine or other sulfur sources may be required for maximum growth. However, larger additions of cysteine should be avoided, as M. elsdenii can ferment cysteine to H2S, which combines with ferrous ions in the medium to produce black FeS precipitates that interfere with visual detection or quantitative measurement of cell growth (Figure 3).
5. Nutrient metabolism and growth
Growth requirements. Although M. elsdenii’s unique ability to use either lactate or glucose as a fermentable energy source was discovered at its first isolation [7], a more detailed characterization of its growth requirements dcveloped more slowly. Bryant and Robinson [51]] observed that factors found in yeast extract, such as biotin, pantothenate and pyridoxine stimulated the growth of M. elsdenii, while acetate was required when glucose was used as energy source. Although Hungate [2] (p.88) indicated that M. elsdenii required some amino acids for growth, Forsberg [52] observed that strains B159 and T81 could grow in a casein-free medium containing minerals, ammonium as N source, a sulfur source, and acetate, plus glucose as carbon and energy source, along with the above-mentioned vitamins. This observation is consistent with Lee et al. [53], who reported the presence of all genes for the biosynthesis of the 20 common protein amino acids. Subsequently, Wallace [54] observed that amino acid catabolism, while substantial, is of minor energetic significance. Rychlik et al. [55] observed that all strains of M. elsdenii they evaluated were capable of growth on free amino acids but could not grow on peptides, which suggests that M. elsdenii is deficient in peptidase activity. Unlike many ruminal bacteria, M. elsdenii does not require branched-chain volatile fatty acids (BCVFA) as growth factors [2] (p.88], producing any as needed for branched-chain lipid biosynthesis through amino acid fermentation. Additionally, M. elsdenii tests negative for many classical diagnostic tests (production of indole or acetoin; gelatin liquefaction; nitrate reduction) [2] (p.84).
Growth studies in chemostats have suggested that growth yields of M. elsdenii B159 are atypically high (0.24 – 0.58 g cells [g glucose]-1) [56,57]. However, such values are substantially overestimated because they do not take into account the catabolism of the relatively high concentrations of amino acids in the complex growth media [58], or the non-catabolic conversion of glucose to intracellular glycogen [49]. Growth yields on a glycogen-free basis for strain T81 on glucose or lactate, in media with low concentrations of amino acids, were reportedly 0.079 and 0.024 g cells (g substrate) -1, respectively [49].
Utilization of lactate and sugars. The distinguishing metabolic feature of M. elsdenii is its ability to utilize both glucose and lactate as fermentable energy sources, via well-established fermentation pathways.
Glucose is fermented via the Embden-Meyerhof pathway to pyruvate and thence to acetate, H2 and CO2 via a classical phosphoroclastic pathway involving pyruvate dehydrogenase, with ATP produced by substrate-level phosphorylation. Acetate (C2) is further converted to butyrate (C4) and caproate (C6) by chain elongation (CE, also called reverse β-oxidation; Figure 4). Genomics-based metabolic modeling [53] (discussed further in Section 9 below) suggests that during fermentation of glucose, crotonyl-CoA --a key intermediate in the CE process -- is produced from pyruvate via a bifurcating pathway with AcCoA and succinate as alternative intermediates (see Section 9 below).
Lactate is subjected to dismutation, with an oxidative path to acetate (via pyruvate, and generating ATP) and a reductive path to propionate (via an acrylyl CoA intermediate, the so-called acrylate pathway). Although the organism produces a NAD-independent D-lactate dehydrogenase (iD-LDH), an active lactate racemase (LR) assures that both D- and L-lactate are utilized. Both acetate (C2) and propionate (C3) can be further converted via CE to produce valerate (C5) and occasionally heptanoate (C7) [8].
The recent whole genome completed for M. elsdenii NCIMB 702410 [23] supports the known capacity of this bacterium to ferment both substrates. Glucose fermentation is enabled by the genes for all 10 enzymes involved in EMP pathway as well as genes coding enzymes necessary for lactate metabolism, including lactate racemase (4 genes), lactate permease (2 genes), and lactate utilization protein (1 gene). Contrarily, genes coding enzymes associated with the metabolism of lactose, mannose, xylose, trehalose, cellobiose, and sucrose were not found in the genome of M. elsdenii, which agrees with data presented by Marounek et al. [59], who observed that some strains were unable to ferment or presented weak reaction for those sugars. Also, genes encoding glucanases, hemicellulases, xylanases, and amylase activity were absent in the genome of M. elsdenii NCIMB 702410, consistent with its known inability to degrade complex polymers from the diet, such as cellulose, hemicellulose, and starch. Thus, the whole genome of M. elsdenii confirms its classification as a secondary fermenter in the rumen.
Much attention has been devoted to comparing M. elsdenii‘s behavior toward lactate versus sugars. Russell and Baldwin [60] reported that M. elsdenii B159 fermented glucose and maltose (sugars derived from hydrolysis of starch), as well as sucrose and lactate, but not xylose or cellobiose (sugars derived from structural carbohydrates). Although strain B159 could ferment glucose, maltose, and lactate simultaneously, lactate fermentation yielded slower growth (μmax 0.21 h-1, versus 0.45 h-1 or 0.55 h-1 on glucose or maltose, respectively). Using chemostats, Russell and Baldwin [61] observed that strain B159 displayed higher affinity for glucose than for lactate (Ks = 0.111 vs. 0.37 mM), but this was not confirmed in later studies.
Marounek et al. [59] showed that lactate fermentation by M. elsdenii strains proceeded 5 to 6-times faster than glucose fermentation. Similarly, Hino et al. [62] observed that M. elsdenii NIAH102 fermented lactate 6 times faster than glucose, likely due to the fact that 5- to 6-times more lactate, on a molar basis, was needed to support a similar extent of growth as glucose.
Weimer and Moen [49] observed that complete consumption of lactate by M. elsdenii T81was observed at initial concentrations of up to 210 mM while maximum glucose consumption was ~80 mM, and much of that glucose was stored intracellularly as glycogen, rather than fermented. Moreover, when glucose and DL-lactate were available simultaneously strain T81 used lactate first, and glucose utilization began only after lactate was almost completely consumed. Hino et al. [63] inferred that glucose is not fermented by M. elsdenii at higher rates until lactate racemase (LR) activity becomes minimal, which only happens when lactate is present at very low concentrations. These authors suggested that preference for lactate developed from its greater availability, and that intracellular LR activity seems to be the major means of controlling glucose fermentation. Preference for lactate is consistent with the constitutive synthesis of iD-LDH, priming the bacterium for immediate use when lactate is suddenly available during rapid fermentation of cereal grains.
M. elsdenii’s ability to ferment both glucose and lactate is likely of benefit in that feeding of highly digestible starch will yield both glucose (from starch depolymerization) and lactate (preferentially produced from glucose fermentation by streptococci or other community members). Moreover, M. elsdenii can accumulate large amounts of glycogen intracellularly for later use [49], which may provide a means of effectively competing against other community members for available glucose despite a lack of an innate capacity for high-flux glucose fermentation to lactate. Glycogen reserves also provide utilizable energy to support an unusually high maintenance requirement (m = 0.187 g glucose [g cells . h]-1 [56]).
End product formation. The distribution of products of both glucose and lactate fermentation by M. elsdenii are summarized in Table 4. As noted above, the two substrates yield starkly different product profiles. Glucose fermentation yields VFA with primarily even carbon numbers (acetate, butyrate, caproate), while lactate yields primarily acetate, propionate and valerate. In addition to VFA, M. elsdenii also produces fermentation gases, including CO2 and a moderate amounts of H2 (0.27 mol [mol substrate]-1 by strain T81 on both glucose and lactate [49] and 0.036 mol H2 [mol lactate]-1 by strain LC-S [17]).
Reducing equivalents generated from sugar oxidation are disposed of via reduction of protons to H2, or by CE of acetate. CE can be further stimulated by exogenous addition of acetate [63], which is stoichiometrically converted to butyrate with only modest additional caproate production [49]. Though H2 production diverts some reducing equivalents from the CE pathway, it appears necessary to provide substrate flux and entropic balance in the exergonic synthesis of VFA, in a manner similar to that in the well-studied asaccharolytic ethanol-utilizing CE bacterium, Clostridium kluyveri [64].
Lactate fermentation proceeds through an acrylyl CoA intermediate, with the necessary reducing equivalents generated by oxidation of some lactate via lactate dehydrogenase and the pyruvate dehydrogenase system, to produce acetate and ATP. A balanced dismutation for the simultaneous oxidation and reduction of lactate would yield the stoichiometry 3 Lactate → 2 Propionate + 1 Acetate + 1 CO2. In practice, however, similar molar amounts of acetate and propionate are obtained, with the excess reducing equivalents produced from lactate oxidation being used for CE of some propionate to valerate. Because acetate is also a potential substrate for CE, some butyrate is also produced. Lactate racemase (LR) is a key enzyme in lactate metabolism by M. elsdenii. Not only does it allow utilization of both D- and L-isomers of lactate (both of which are produced by the ruminal community), but it also serves as a key metabolic control point. Hino and Kuroda [65] observed that LR was not synthetized by M. elsdenii growing on glucose as an energy source, which explains why this bacterium does not produce propionate when fermenting glucose in the absence of lactate, which is the inducer of the LR [66]. Thus, it can be inferred that the contribution of M. elsdenii to propionate production in the rumen only will occur when sufficient lactate is available to stimulate LR synthesis. This property is very different from that in classical propionic acid bacteria, which produce propionate from glucose using the succinate pathway. Additionally, contrary to most other ruminal bacteria in which LDH is induced by substrate, in M. elsdenii this enzyme is constitutive.
Fermentation end product values vary widely among strains (Table 4), though some differences may be due to differences in growth conditions, particularly substrate concentration and incubation time. Higher substrate concentrations and longer incubation times promote a more “complete” fermentation with VFA chain elongation, enhancing the relative proportions of valerate and caproate.
The pH of the growth medium also can influence the end products generated by M. elsdenii. Counotte et al. [67] observed that the fermentation of DL-lactate to propionate via the acrylate pathway decreased with decreasing ruminal pH, with a resultant increase in butyrate production due to CE of acetate.
6. Resistance to environmental stress
pH. The pH is one of the most important characteristics of the chemical environment that affects living cells, because it affects cell membranes and their functionality [68] as well as intracellular properties such as enzyme activities [57] and cell viability [69], as well as the thermodynamics and kinetics of many catabolic reactions [70]. Most ruminal microbes can be classified as neutrophilic organisms, defined as those that thrive at pH 6.5 to 7.5 [71]. Many rumen microbial species, especially fibrolytic organisms, are sensitive to low ruminal pH (<6.0), but some species such as Streptococcus bovis and Lactobacillus spp. – both of which produce lactate -- are resistant to low pH [72].
Ruminants evolved to consume fibrous feedstuffs, which are slowly digested by ruminal microorganisms, and which stimulate secretion of well-buffered saliva, keeping the rumen pH close to neutral (pH range from 6.2 to 7.0 in forage-fed animals [1] (p.246], [72] (p.9), and 5.8-6.5 in well-adapted grain-fed cattle [73]. However, the increased demand for animal protein (meat and milk) by humankind in the last decades has pushed for improved animal performance, which can most easily be achieved by increasing the grain content of the diet. However, this strategy can decrease ruminal pH below 6.0 for many hours per day, because starch in grains is rapidly fermented (primarily to lactate) in the rumen. This can have negative effects on rumen microbial populations and their activities, and can lead to digestive disturbances such as rumen acidosis and associated diseases (bloat, rumenitis, liver abscess, laminitis, etc.) [74].
Russell et al. [75] suggested that at low extracellular pH bacteria would have insufficient energy to export protons through the cell membrane to establish a protonmotive force, a principal reason why many bacteria display poor growth when pH is decreased. In this way, these authors observed that M. elsdenii B159 grew in medium containing glucose as energy source only at pH > 6.1. Contrarily, Hobson et al. [27] observed that growth of the LC organism (M. elsdenii) occurred at pH values ranging from 4.8 to 8.1. Therion et al. [76] reported that M. elsdenii ATCC 25940 displayed more sensitivity to low pH (5.0 to 5.5) when fermenting glucose than when fermenting lactate.
Weimer and Moen [49] verified that growth rate (0.66 h-1) of M. elsdenii T81 on DL-lactate was not affected when the initial culture pH ranged from 5.0 to 6.6, but at pH 4.65 the growth rate was dramatically reduced (to 0.17 h-1). Rapid growth at pH 5.0, would help prevent lactate accumulation in the rumen even though this pH value is considered suboptimal for rumen function. By contrast, Waldrip and Martin [77] noted that M. elsdenii B159 displayed similar rates of L-lactate uptake when growing at pH 6.0 to 8.0, and higher rates at pH 4.0 to 5.0. The authors suggested that M. elsdenii probably takes up L-lactate by a proton motive force-driven mechanism.
Antibiotics. Antibiotics are used on farms to treat sick animals as well as to improve animal performance (i.e, as growth promoters) or prophylactically to prevent digestive disturbances such as ruminal acidosis [78]. Thus, the rumen microbial community is often exposed to antibiotics, and resistance mechanisms are critical for survival under these conditions. Antibiotic resistance can be an intrinsic characteristic of a microorganism (genes in DNA) or acquired from other organisms by horizontal gene transfer via plasmids or transposons [79]. The resistance of M. elsdenii to more common antibiotics used as growth promoters in ruminants diets, such as ionophores (e.g., monensin), is a critical aspect in its role in lactate fermentation in the rumen of feedlot cattle, because other lactate fermenters in the rumen, like S. ruminantium, are known to be highly sensitive to these agents. Rychlik et al. [55] evaluated monensin resistance of some M. elsdenii strains and concluded that monensin probably does not decrease M. elsdenii numbers in vivo at normal therapeutic doses.
Most of the common rumen bacterial species are sensitive to tetracycline [80,81], but Flint et al. [82] isolated strains of M. elsdenii resistant to this antibiotic, two of which had a plasmid of approximately 5-8 kilobase pairs (kbp) that carried a tetracycline (Tc) resistance marker. Additionally, Nagaraja and Taylor [83] showed that M. elsdenii B159 was totally resistant to eight antimicrobial feed additives (lasalocid, monensin, narasin, salinomycin, avoparcin, thiopeptin, tylosin, and virginiamycin). The susceptibility and resistance of M. elsdenii to thirteen other antibiotics have also been reported by El Akkad and Hobson [80] and by Wang et al. [84].
Organic acid toxicity. M. elsdenii T81 cannot initiate growth at lactate concentrations >210 mM at pH 6.8 [49], and is also sensitive to high extracellular VFA concentrations [85]. This inhibition is likely due in part to the well-known intracellular accumulation of VFA in response to passive migration of protonated VFA into cells, which upon ionization at higher intracellular pH results in high intracellular concentrations of VFA anions [86]. Other factors probably contribute to toxicity, as VFA toxicity increases with chain length despite similar pKa values for these VFAs. Other organic acids can also be inhibitory; for example, Prabhu et al. [66] have reported that the growth rate of M. elsdenii T81 (=ATCC 17753) was reduced 4-fold by 30 mM acrylate, a normally intracellular intermediate in lactate fermentation.
7. Relationships with others ruminal microorganisms
The rumen is a complex habitat in which factors related to diets and to the host are major determinants of the resident microbial community. Interactions among community members are also important as modulators of ruminal ecology [87,88]. These interactions -- mutualism, competition, predation and amensalism -- all contribute to rumen microbial community structure. Considering that M. elsdenii is not a primary fermenter in the rumen, as it cannot degrade polymeric carbohydrates from the diet (including structural or nonstructural carbohydrates), this bacterium depends on primary degraders which supply glucose or lactate that M. elsdenii can use as energy and carbon source. Thus, in order to understand better the role of M. elsdenii in the rumen it is important to consider its relationship with others ruminal microorganisms.
Interactions with starch and sugar fermenters. Assuming that M. elsdenii is probably the major lactate fermenter in the rumen and that rumen lactate primarily originated from Streptococcus bovis growing at high glucose concentrations, Russell et al. [89] examined the interaction between these two species in continuous culture as a function of pH and dilution (growth) rate. Binary coculture of the two species always displayed higher populations of S. bovis than of M. elsdenii, which was attributed to the former’s higher growth rates and the higher affinity for maltose, and its lower maintenance coefficient. Increasing dilution rate (from 0.12 to 0.36 h-1 at pH 6.6) caused a dramatic increase in the S. bovis/M. elsdenii ratio (from 3.5 to 23). Lactate production by S. bovis was modest at pH 6.6 and 5.7, and at lower dilution rates (0.12 and 0.22 h-1), but became significant at pH 5.7 and high dilution rates (0.36 h-1). In this last condition (high lactate concentration) a lower S. bovis/M. elsdenii ratio was observed, which can be explained by enhanced growth of M. elsdenii fermenting lactate produced by S. bovis. However, at pH 5.4 the S. bovis/M. elsdenii ratio increased until no M. elsdenii cells were detected, indicating the sensitivity of this bacterium to very low pH and a higher resistance of S. bovis to acidic conditions.
With the goal of examining the potential impact of M. elsdenii in both acute ruminal acidosis and SARA, Chen et al. [90] performed more systematic studies on the interactions among M. elsdenii, S. bovis, S. ruminatium, Lactobacillus fermentum and Butyrivibrio fibrisolvens in batch cultures fed 3 different levels of starch at both pH 5.5 and pH 6.5. Under these in vitro SARA-simulating conditions, lactate consumption was largely able to keep pace with its production, apparently due to the contributions of both M. elsdenii and B. fibrisolvens, with the latter species displaying more positive correlations to production of the degradation products propionate, butyrate and formate. Monocultures of M. elsdenii continuously fed 15, 30, or 90 mM lactate displayed increased growth rates with increased lactate concentration, except for the very poor growth at 90 mM lactate and pH 5.5; these results run counter to the widely held perception that M. elsdenii is a superior lactate fermenter under SARA conditions. Examination of gene expression under the different growth conditions led Chen et al. [90] to suggest that under the more extreme conditions M. elsdenii loses much of its capacity to keep pace with lactate production under ARA conditions, but that limiting the negative effects of acidosis may be attainable by controlling ruminal pH to a range where M. elsdenii can effectively operate. This conclusion is in accord with the study of Kung and Hessian [91], who showed that inoculation of M. elsdenii B195 into in vitro ruminal fermentations of rapidly fermentable carbohydrates substantially attenuated lactate accumulation and pH decrease.
Yang et al. [31] examined biofilms formed on the grains of barley and corn incubated in nylon bags inserted into the rumen of dairy cows. Genus Lactobacillus was dominant taxon associated with starch digestion after 12 h of incubation, which was followed by an increase abundance of genus Megasphaera. These results improve our understating of the rumen microbial community, not only by establishment of a tight relationship between these two genera but also by demonstrating that M. elsdenii is not restricted to liquid fraction of rumen contents. Thus, studying M. elsdenii population in the rumen needs to evaluate its growth in the both the liquid and solid fractions of rumen contents.
Competition with lactate utilizers. Selenomonas ruminantium subsp. lactilytica can be regarded as the chief known competitor of M. elsdenii for lactate produced in the rumen. However, the two species differ markedly in their catabolic pathways and their regulation (Table 5). Selenomonas ruminantium subsp. lactilytica conducts a mixed acid fermentation of glucose to acetate and propionate at low growth rates, but switches to a homolactic fermentation at high growth rates [56], and upon glucose exhaustion can ferment lactate to propionate, with ancillary production of acetate and CO2 [92]. Unlike M. elsdenii, Selenomonas ruminantium subsp. lactilytica uses the succinate pathway for propionate production. The genome of S. ruminantium lactilytica TAM6421 Is substantially larger and more complex than is that of M. elsdenii [93] (Table 5).
The current consensus is that M. elsdenii is the major lactate user in the rumen of animals fed high-grain diets, while Selenomonas ruminantium subsp. lactilytica is a significant but secondary contributor. This notion is supported by physiological characteristics of pure cultures; by enzymatic data; and by labeling studies that can distinguish between the different routes of propionate synthesis (succinate versus acrylate pathways).
Like the non-lactilytic subspecies S. ruminantium subsp. ruminantium, Selenomonas ruminantium subsp. lactilytica is an outstanding glucose fermenter. Russell and Baldwin [60] showed that the lactilytic strain HD4 was capable of rapid growth, a high catabolic growth yield, and very low maintenance coefficient on glucose (Table 5).
Russell and Baldwin [60] also observed that this strain displayed catabolite repression of lactate utilization when glucose or sucrose was added to growth medium. By contrast M. elsdenii displays lower growth yields on glucose and a much higher maintenance coefficient; is not subject to catabolite depression of lactate utilization by glucose; and displays superior growth properties on lactate. Counotte et al. [67] observed that pure cultures of M. elsdenii fermented DL-lactate more rapidly than did Selenomonas ruminantium subsp. lactiltyica (5.49 x10-14 mol cell-1 h-1 versus 2.42 x 10-14 mol cell-1 h-1). Moreover, Fan et al. [94] reported that, on lactate, M.elsdenii BE2-2083 displayed higher maximum specific growth rates than did Selenomonas ruminantium subsp. lactilytica HD4 at both pH 6.5 and pH 5.5 as well as shorter lag times before initiating growth.
In addition to differences in pathway regulation noted above, the two species differ in enzymatic capabilities. M, elsdenii has a lactate racemase that allows it to use both the D- and L-isomers of lactate. Because Selenomonas ruminantium subsp. lactiltyica lacks both an L-LDH and lactate racemase, its lactate fermentation is restricted to the D-isomer. Additionally, D-lactate uptake is greatly enhanced by several dicarboxylic acids (malate, fumarate and/or aspartate) that are intermediates or metabolites of the succinate pathway; this suggests that lactate utilization is more closely linked to (i.e., more dependent on) glucose metabolism.
Labeling studies also support the dominance of M. elsdenii. Early work revealed that increasing the levels of degradable starch in the diet (which facilitates lactate production) increased the relative contribution of the acrylate pathway towards propionate production from glucose [95,96]. Later, Counotte et al. [67] used 14C-labeled lactate to demonstrate that M. elsdenii accounted for 60-95% of lactate fermented in binary cocultures with S. ruminantium subsp. lactilytica. Moreover, labeling experiments with mixed ruminal inocula in vitro indicated that ~74% of the lactate was converted via the acrylate path (characteristic of M. elsdenii), with the remainder converted via the succinate path (characteristic of S. ruminantium subsp. lactilytica). These results are supported by more recent work of Fan et al. [94], who used 13C-NMR to show that, in batch-mode binary co-cultures with S. ruminantium subsp. lactilytica, M. elsdenii accounted for 82% and 75% of 13C-D-lactate catabolism at pH 6.5 and 5.5, respectively. These percentages likely change within the rumen over time: The ability of S. ruminantium subsp. lactilytica to ferment sugars in preference to D-lactate suggests that its propionate formation in the rumen is maximal shortly after starch feeding (when glucose is likely most abundant), while propionate production by M. elsdenii occurs later, after lactate has accumulated.
An additional twist on the relationship between M. elsdenii and S. ruminantium subsp. lactilytica is suggested by their response to lipopolysaccharides (LPS), cell outer membrane components widely produced by Gram-negative bacteria, including those in the rumen. Sarmikasoglu et al. [97] have reported that mixed LPS from the rumen reduced the growth rates of pure cultures of S. ruminantium subsp. lactilytica HD4 and Streptococcus bovis JB-1 on glucose (i.e., during lactate production) but did not affect growth of M. elsdenii T81 on lactate. Because strain HD4 is also lactilytic and strain T81 can grow on glucose, additional tests are needed to determine if these inhibition differences are inherent featues of the bacteria (independent of substrate), or are substrate-dependent.
While the interactions described above appear to be a sound model for lactate consumption patterns in the rumen, it is wise to recall that the rumen contains many bacterial species whose characteristics and functions remain to be elucidated. In this regard, two lactate-utilizing candidate species – phylogenetically related to Butyrivibrio fibrisolvens and Anaerococcus prevotii -- have recently been identified from metagenomic analysis of ruminal lactate enrichment cultures [98]. Isolation, characterization and quantification of these species in ruminal contents – along with elucidation of their interactions with conventional lactate utilizers such as M. elsdenii -- will ultimately improve our understanding of ruminal lactate metabolism.
8. Nutritional Importance
The nutritional importance of M. elsdenii can be evaluated from four different perspectives on ruminant production: 1) as a contributor to improved ruminant performance; 2) as a causative agent of milk fat depression in high-producing dairy cows; 3) as a probiotic to stimulate ruminal development in pre-weaning animals; and 4) as a probiotic to prevent ruminal acidosis in animals fed high-grain diets.
Milk production efficiency. Shabat et al. [100] have reported that in the rumen of cows that display high milk production efficiency, there was an increase in the relative abundance of genus Megasphaera (based on 16S rRNA gene copy number measured by Illumina sequencing), as well as an increase in the expression of genes associated with the acrylate pathway known to be characteristic of M. elsdenii lactate fermentation. On this basis, the authors ascribed a role for M. elsdenii in the elevated propionate levels observed in the rumen of these higher-efficiency cows. However, M. elsdenii abundance in this study was very low [<0.08% of 16S rRNA gene copy number], and thus would seem unlikely to have contributed substantially to production of propionate, which is produced by many ruminal bacteria, particularly members of the abundant genus Prevotella [101] Moreover, several other studies that compared cows of divergent efficiency did not reveal differences in M. elsdenii abundance [102,103]. It is worth recalling, though, that relative abundance does not equal relative activity (as indicated by the transcriptome study of Park et al. [43]). The high maintenance requirements of M. elsdenii (i.e., high catabolism without growth) may translate to an outsized fermentation and VFA production activity that may partially explain its contribution to propionate in the Shabat et al. [100] study. The potential importance of M. elsdenii in improving milk production efficiency, while intriguing, remains uncertain and warrants further study.
Milk fat depression. Fat is one of the most important milk components and has been used for decades as way to determine milk price in various countries, yet it is also the component of highest variability. In Holsteins, the fat content in milk can drop below 3.0%, the commonly held standard for so-called milk fat depression (MFD). MFD is most common in high-producing dairy cows fed high-starch (low fiber) diets, especially when supplemented with polyunsaturated fats, and when supplemented with the ionophore monensin. It is known that MFD is caused by the negative effect of certain trans-10 fatty acids -- particularly trans-10, cis-12 conjugated linoleic acid (t10-c12-CLA) -- on de novo fat synthesis in the mammary gland, which has its production increased in the rumen of MFD cows [104]. The current challenge is to identify the major organisms which effectively produce these fatty acids in vivo.
Although Butyrivibrio fibrisolvens is the ruminal bacterium more frequently associated to lipid metabolism in the rumen [105], presenting potential to hydrolyze lipids and hydrogenate polyunsaturated fatty acids, this bacterium does not synthetize t10-c12-CLA [106], nor can it grow at ruminal pH lower than 5.95 [57], conditions under which trans-10 fatty acids production in the rumen increases. In view of this, Kim et al. [107] [isolated M. elsdenii YJ-4 from the rumen of a cow fed with 90% grains, and observed in vitro that this strain, as well as M. elsdenii T81 produce considerably more t10-c12-CLA (>6 µg and > 4 µg mg protein-1, respectively) than other strains evaluated (B159, AW106 and JL1). Later, Kim et al. [108] inferred that the production of t10-c12-CLA could be increased when ruminal fluid was enriched with lactate.
In accord with these pure culture results, some studies [36,37] have shown that dairy cows displaying MFD presented higher relative abundance of M. elsdenii (up to 4% of total bacterial 16S rRNA gene copy number, compared to just 0.02% in cows before the challenge to MFD or in cows that did not present MFD). Based on these results, Weimer et al. [50] conducted three experiments in which M. elsdenii was dosed directly into the rumen of cannulated dairy cows; in some experiments, the dosed M. elsdenii strains were those that had been previously isolated from the same individual cow during an episode of MFD. The authors did not find any effect of dosing M. elsdenii in the rumen on milk production and composition, nor on rumen pH, total VFA and lactate in the rumen, although after dosing the butyrate proportion was increased. The authors also observed that in almost all animals the M. elsdenii abundance in rumen dropped to low baseline levels (<0.02% of 16S rRNA gene copy number) within 24 h of dosing, even upon multiple dosings, which could explain the lack of effect of dosing on ruminal parameters and, possibly, milk composition. At this point, the relationship between the ruminal abundance of M. elsdenii and MFD is associative, but a causative role has not yet been demonstrated.
Dosing to stimulate ruminal development. Two studies have been published that aimed to stimulate ruminal development by inoculation of M. elsdenii into preweaned calves. Muya et al. [109] found that M. elsdenii NCIMB 41125 dosed at 14 days of age increased ruminal butyrate and plasma β-hydroxybutyrate (BHBA), and also improved both intake of starter feed and rumen development (measured as increase in absorptive area), compared to non-dosed calves. In the second study [41] Lactipro® (2 × 108 cfu/mL of M. elsdenii NCIMB 41125) was administered to dairy calves and, although M. elsdenii shifted the abundance of some less abundant bacteria, there was no increase in M. elsdenii abundance, and no effect on ruminal and blood parameters.
Dosing to control lactic acidosis. The ability of M. elsdenii to consume lactate produced from starch or sugars by Streptococcus bovis and other rapid fermenters has fostered research aimed at using M. elsendii as a probiotic to attenuate the effects of acute lactic acidosis (ALA), a serious metabolic condition that contributes to disruption of rumen microbial ecology, reduced ruminal fiber digestion, damage to the rumen epithelia and the liver, and inflammation of the hoof wall (laminitis). Because ALA undesirably compromises both animal health and productivity, many of these studies have examined the effect of dosing on both. This research has yielded a wealth of sometimes contradictory data that appear to highly dependent on experimental conditions. A recent meta-analysis by Susanto et al. included 32 published studies (27 of which were peer-reviewed) with cattle and sheep [110]. These studies varied with respect to diet, and route of addition (oral vs ruminal cannula) and included a wide range of dosing levels (log10 cfu of 7.0 to 13.3); however, frequency or timing of supplemenation was not analyzed.. The authors concluded that supplementation with M. elsdenii decreased ruminal lactate concentration and the proportion of acetate in VFA, while increasing ruminal pH, methane production and the proportion of C3-C5 in VFA. The same meta-analysis reported that M. elsdenii supplementation reduced the occurrence of health problems (scours, bloat, and liver abscesses) and improved some metrics of animal performance (average daily gain (ADG), hot carcass weight). Interestingly, dry matter intake decreased with M. elsdenii supplementation, but feed conversion ratio did not change, raising the question of whether supplementation would compromise production. We discuss here some of the studies used in the meta-analysis in more detail to illustrate the range of responses observed.
Robinson et al. [111] dosed a strain of M. elsdenii into feedlot steers and observed lower ruminal lactate concentrations, higher ruminal pH, and 24% greater DMI when compared to control animals. Contrarily, Klieve et al. [33] did not observe any differences on rumen pH and lactate concentrations between steers inoculated with M. elsdenii and control steers, because M. elsdenii had also established in the rumen of animals from control group upon increase grain addition in the diet. However, grain-fed cattle inoculated with M. elsdenii displayed a 100-fold increase in abundance in the first 4 days following inoculation and also established a population of M. elsdenii 7-10 d earlier compared to uninoculated cattle.
Later, McDaniel [112], using feedlot steers fed high-concentrate diet dosed intraruminally with 1.62×108 CFU/mL of M. elsdenii strain NCIMB 41125, observed that inoculated steers had higher ruminal pH and lower lactate concentrations than non-dosed steers. In the same way, Leeuw et al. [113] dosed feedlot steers with M. elsdenii NCIMB 41125 and found 5.6% better ADG during weeks 3 - 5 of feedlot as well as lower morbidity for dosed steers compared to the non-dosed group. Later, Henning et al. [114] administered M. elsdenii to feedlot cattle prior to feeding high-concentrate diets and observed increase on dry matter intake, ADG, and feed efficiency. Henning et al. [115] then evaluated 9 strains of M. elsdenii in vivo and observed high variability among strains regarding the capacity to control ruminal pH, with some strains preventing lactate accumulation and a drop in ruminal pH during an acidosis challenge. Aikman et al. [116] administered M. elsdenii to lactating dairy cows and observed a decrease in the amount of time that ruminal pH was < 5.6 compared to cows that were not supplemented. However, Zebeli et al. [117] intraruminally inoculated mid-lactation dairy cows (fed a diet containing 45% concentrate) with 35 mL d-1 of M. elsdenii ATCC 25940 (MEGA) culture that contained 108 cfu mL-1 of bacteria, and observed only slight changes in plasma metabolites and milk composition.
Evaluating the persistence of orally administered M. elsdenii in the rumen of beef cattle fed a barley diet, Klieve et al. [118] noted that the populations of M. elsdenii of 103 - 107 cells mL-1 were observed in all inoculated steers after 3 d, but were also detected in 70% of uninoculated steers (though at a density lower than 1 x 106 cells mL-1). For both groups of steers M. elsdenii populations rapidly increased by day 14 (densities of 107 –108 cells mL-1) and remained stable for the remainder of the experiment. The authors mentioned that the rapid acquisition of M. elsdenii for both groups of steers could question the need to inoculate with the bacterium.
Higher ADG and hot carcass weight were observed by Drouillard et al. [119] in steers dosed orally with 200 mL of M. elsdenii NCIMB 41125 (1011 cfu mL-1) than steers of control group. In same way, Ye and Eastridge [120] observed higher milk and fat yield in dairy cows (> 3 lactations) dosed with 200 mL Lactipro® (1 x 108 cfu mL-1) compared to a control group.
Arik et al. [121] dosed M. elsdenii (ATCC 17753, 200 mL of 2.4 x 1010 cfu mL-1) in the rumen of cannulated Holstein heifers fed wheat or corn-based concentrate and found that M. elsdenii inoculation helped to prevent SARA by decreasing S. bovis populations and increasing protozoal populations in the rumen. Sedighi and Alipour [122] used in vitro and in vivo studies to evaluate 4 strains of M. elsdenii¸ and observed that M. elsdenii inoculation increased in vitro gas production and pH values, as well as decreased lactate concentration. In the in vivo study the authors found that oral dosing of M. elsdenii SA3 to dairy cows decreased ruminal lactate compared to control and had a tendency to increase milk fat yield.
M. elsdenii NCIMB 44125 was also orally dosed (100 mL of 2 x 108 cfu mL-1) into cull beef cows in two simultaneous studies in which the authors found increase on absorptive surface area (ASA) in rumen wall and increase on papillae area: ASA ratio [123] as well as a tendency to increase average daily gain and carcass weight compared to the undosed control group [124]. These authors also used the same strain of M. elsdenii (50 mL, 2 x 108 cfu mL-1) which was orally dosed in early weaned beef calves, and observed that dosed animals presented higher ADG during first 21 d of study as well as and a tendency to have higher percentage of intramuscular fat and higher marbling scores than undosed calves. The authors observed a lower liver score for dosed animals, which suggest the benefits of M. elsdenii in preventing liver abscesses.
Mazon et al. [125] evaluated M. elsdenii NCIMB (100 mL, 2x108 cfu mL-1) in lactating dairy cows dosed 4 d and 1 d before a SARA challenge. Dairy cows dosed 4 d before acidosis challenge presented higher ruminal pH and experienced shorter and less intense acidosis and, as a consequence of this, had higher dry matter intake and higher milk yield than undosed cows.
More recently, Lopes et al. [126] observed that dosing 20 mL of Lactipro® (M. elsdenii, 1 x 1010 cfu mL-1) into feedlot Nellore bulls permitted reducing the length of the adaptation to a feedlot diet by 6 d compared to the undosed control group.
Thus, the potential benefit of M. elsdenii in preventing ruminal acidosis appears to depend on its dosage and probably on the capacity of inoculated (dosed) strain to persist in the rumen after dosing as well as of challenge of ruminal acidosis, especially the occurrence of favorable conditions in the rumen (adequate pH and enough lactate) to persist. Unfortunately, very few studies have quantified the dosed strains in the rumen over time following inoculation. In the same way, the development of stable commercial products for frequent oral dosing to animals is essential for establishing the strain in the rumen, or at least maintaining a sufficient abundance between dosings in order to achieve the goal of preventing ruminal acidosis. Finally, it merits mention that any success of probiotic M. elsdenii in controlling acute lactic acidosis may be attenuated somewhat by the conversion of lactic acid to other VFA (particularly butyric), with a resulting increased propensity to trigger SARA [47].
The effectiveness of M. elsdenii in mitigating rumen acidification also might be hypothesized to control certain foodborne pathogens such as Escherichia coli O157:H7, whose pathogenicity has been reported to be induced by ruminal acidosis prior to shedding from the hindgut [127]. However, oral drenching of finishing steers with M. elsdenii did not significantly affect the prevalance of E. coli O157:H7 in anorectal swabs, suggesting that they did not impact this particular metric of food safety [128].
9. VFA production
M .elsdenii and the related ruminal isolates M. hexanoica [129] and M. indica [130] are among a select group of so-called chain-elongating (CE) bacteria that extend the length of short-chain fatty acids by successively adding acetyl units, thus converting acetate (C2) to butyrate (C4) and then caproate (C6), or converting propionate (C3) to valerate (C5) (Figure 4). This CE occurs as part of energy metabolism (catabolism), and differs from the nearly universally distributed anabolic synthesis of long-chain fatty acids for cellular lipids, in that in CE the carboxylate product is produced in large amounts and is exported from the cell, and does not proceed beyond production of medium-chain carboxylates (MCC, <C8). Because MCCs can serve as feedstocks for producing alkanes, alkenes or alcohols using chemical or electrochemical methods, there is substantial interest in this “carboxylate platform” for fermentative production of these MCCs from sugars or from primary fermentation products such as lactate or ethanol [8], or from complex feedstocks such as food waste or cellulosic biomass [131].
As noted above, M.elsdenii has the unusual property of producing even-chain fatty acids (C2, C4, C6) by fermentation of sugars, but primarily odd-chain fatty acids (C3, C5) from lactate [49]. Because lactic acid is readily produced by fermentation of sugars by various lactic acid bacteria (LAB), conversion of low-quality sugar-containing wastes via a lactic acid intermediate can be carried out by LAB/M. elsdenii co-cultures, yielding valerate as the primary product at titers of 3.3 g/L [132], approximately twice that reported for pure cultures of M. elsdenii of nonruminal origin on pure sugars [133]. Alternatively, M. elsdenii can produce caproate to high titers directly from fermentation of glucose and other sugars. Roddick and Britz [134] have reported production of up to 4.3 g butyric and 19 g caproic acids per L by M. elsdenii ATCC 25940 in fermenters employing ion-exchange resins to adsorb VFA end products. Choi et al. [135] obtained production of caproate by M. elsdenii NCIMB 702410 at titers of 28.42 g L-1 from sucrose, in a two-phase reactor. More recently, Nelson et al. [85] grew M. elsdenii NCIMB 702410 in fed-batch pertractive reactors to continuously extract VFA products, and demonstrated the equivalent of 36.5 g butyrate L-1 and 20.7 g caproate L-1, among the highest titers reported for microbial production of this acids mixture.
The CE process can be stimulated by exogenous addition of VFA such as acetate as electron acceptors, if the amount of electron donors is in excess. As noted above, this phenomenon was first noted by Hino et al. [63] upon acetate addition to cultures of M. elsdenii NIAH102 growing on glucose. More recently, Jeon et al. [136] reported that addition of C2-C4 VFA to cultures of the related species M. hexanoica MH (Figure 2)enhanced production of valeric, caproic, heptanoic and octanoic acids (C5-C8), resulting in titers of 5.7, 9.7, 3.2 and 1.2 g L-1, respectively. Using lactate as electron donor and acetate as electron acceptor, M. hexanoica MH has been reported to produce titers of up to 3.7 g caproate L-1, and 1.5 g caprylate (C8) L-1 [137]. Additionally, only low concentrations of odd-chain acids (C5, C7) were produced despite the use of the odd-chain electron donor, lactate, further distinguishing this species from M. elsdenii.
The potential of M.elsdenii as a carboxylate producer has attracted the interest of metabolic modelers. Lee et al. [53] developed a model, dubbed iME375, from the reconstructed metabolic pathways encoded in the genome of strain DSM 20460, with particular attention paid to carboxylate biosynthesis. A bifurcated pathway for synthesis of the essential CE intermediate crotonyl-CoA from glucose was identified (Figure 5). Succinate is a key intermediate in the pathway, and -- unlike in several ruminal bacterial species -- its direction toward crotonyl-CoA is not diverted toward propionate, due to the absence of genes encoding methylmalonyl-CoA mutase and methylmalonyl CoA decarboxylase [22,23].
Metabolic flux analysis indicated that the specific rate of caproate synthesis declined with increasing growth rate, due to decreased flux through the AcAcCoA arm of the bifurcated pathway; by contrast, there was nearly constant flux through the succinate arm of the pathway, which became dominant only μ > 0.4 h-1. Caproate production could be stimulated by exogenous supply of several pathway precursors, including acetate, succinate or butyrate. The authors concluded that balancing the two arms of the pathway is key to maximizing caproate productivity, while simple gene knockouts likely would not improve productivity. Further genomic analysis suggested that CE reactions downstream of crotonyl-CoA display substantial redundancy in the form of separately encoded isoenzymes, not surprising given the important role of CE in M. elsdenii catabolism.
The iME375 represents a useful step forward in optimizing VFA production by M. elsdenii, but as noted by Panikov [58] it has substantial limitations, in that the underlying data did not take into account the substantial contribution of highly abundant supplementary nutrients (particularly amino acids and yeast extract) that are known to alter both fundamental growth parameters (Yg, m) and the flux of pathway intermediates. Regardless, there is general agreement [53,58] that metabolic modeling will play a major role in advancing our understanding of M. elsdenii metabolism and VFA productivity.
Two scenarios are readily envisioned for use of M. elsdenii for industrial VFA production. First, this bacterium could be used either in purê culture using lactate- or sugar-containing fermentation broths (producing mixtures of primarily odd- or even-chain VFAs, respectively), or in binary mixed culture with Streptococcus bovis or other lactic acid producers using sugar-rich feedstocks (e.g., cheese whey or effluents of wet-fractionated herbage) to produce a spectrum of VFA products. Valerate, the major product of these latter fermentations (but not readily produced by most other CE cultures), has already been shown to be a platform chemical for synthesis of automotive fuel [138]. However, the low titers of valerate in these fermentations (~3 g/L) would have to be substantially improved for economic feasibility. The second scenario would involve incorporation of M. elsdenii into mixed culture reactor microbiomes that convert organic wastes to mixtures of medium-chain carboxylates (MCC, containing 5-8 C atoms). Such processes use complex, naturally-derived “open-culture” microbial communities that allow for a wide range of feedstocks and operate without the need to sterilize feedstocks or equipment [131]. These diverse bacterial communities can sustain considerable levels of MCC productivity, but generally produce low proportions of odd-chain MCCs [139], making them attractive targets for bioaugmentation with M. elsdenii. Ultimately, it should be possible to tune the fermentation to the desired mixture of VFA products whose subsequent chemical conversion would yield the desired type and proportions of fuel compounds (alkanes, alcohols and ketones of the desired chain lengths). The ability of M. elsdenii to contribute to MCC production in open-culture reactor microbiomes was noted by Scarborough et al. [140], who reported that M. elsdenii represented 46.3 and 14.2 %, of the bacterial community (based on 16S rRNA gene abundance) in anaerobic bioreactors fed thin stillage from ethanol fermentations and maintained at pH 5.0 or 5.5, respectively. These bioreactors produced substantial yields of MCCs, and M. elsdenii abundance was highest during periods of maximum odd-chain VFA production, consistent with this species’ characteristic capabilities in this regard.
Finally, M. elsdenii may also be coupled to conventional anaerobic digestion to produce biogas. A recent example is provided by Luo et al. [141], who investigated the effect of exogenous addition of M. elsdenii ATCC 25940 on the anaerobic fermentation of a synthetic food waste in a leach bed reactor (LR), and on subsequent biogas formation in an downstream upflow anaerobic sludge blanket (UASB) reactor. Comprehensive mass balance analysis showed that M. elsdenii augmentation improved volatile solids removal in the LR (from 59 to 65%), while improving methanogenesis from both the LR off-gas and the VFA-rich leachate. These metrics were further improved by supplementation of the LR with acetate (carbon conversion efficiencies of 32, 38, and 43 per cent for control, M, elsdenii-amended, and M. elsdenii-amended with acetate, respectively).The improvements were ascribed to more efficient redirection of lactate from the primary fermentation toward CE of acetate to produce butyrate by the M. elsdenii, along with increased H2 production during CE.
10. Final considerations
Despite its somewhat unusual metabolic properties and apparently outsized influence on its ruminant host, Megasphaera elsdenii remains relatively little studied. Our understanding of this bacterium would be greatly enhanced by focusing on several areas.
1) Development of a facile genetic system, including the introduction and maintenance of stable extrachromosomal elements.
2) Characterization of population dynamics throughout the ruminant’s feeding cycle. In the case of studies using this species as a probiotic, this would include measurement of population sizes during the entire “dosing cycle”, from the time of inoculation or feeding onward, as well as identification of factors that limit persistence in the rumen.
3) Examination of more nuanced interactions with the host, such as sensing/production of metabolites or signaling molecules that facilitate interactions between M. elsdenii and both its host and other ruminal microbes.
4) Continued efforts to elucidate the relationship between M. elsdenii abundance and milk fat depression, to identify a mechanism that would explain a relationship that currently is merely correlative.
5) Exploitation of VFA chain-elongating capabilities of M. elsdenii in pure culture, or in mixed communities containing other CE bacteria, with the goal of increasing VFA yield and titer, and of more effectively tuning the proportions of individual carboxylate products – particularly toward valerate, at whose production M. elsdenii is very adept.
Author Contributions
Conception: LSC; Writing, LSC and PJW. Both authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Van Soest, P.J. Nutritional Ecology of the Ruminant. 2nd ed. Cornell University Press: New York, 1994.
- Hungate, R.E. The Rumen and Its Microbes. Academic Press: New York, 1966.
- Joblin, K.N. Isolation, enumeration and maintenance of rumen anaerobic fungi in roll tubes. Appl. Environ. Microbiol. 1981, 42, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Kay, R. Digestion of protein in the intestines of adult ruminants. Proc. Nutrition Soc. 1969, 28, 140–151. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Won, M.Y.; Oyama, L.B.; Courtney, S.J.; Creevey, C.J.; Huws, S.A. Can rumen bacteria communicate to each other? Microbiome 2020, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, C.N.; Gall, L.S. Rumen organisms II: two lactate utilizers and six miscellaneous types. J. Bacteriol. 1953, 65, 554–559. [Google Scholar] [CrossRef]
- Agler, M.; Wrenn, B.A.; Zinder, S.H.; Angenent, L.T. Waste to bioproduct conversion with undefined mixed cultures: the carboxylate platform. Trends Biotechnol. 2011, 29, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, P.T.; Sutter, V.L.; Attebery, H.R.; Bricknell, K.S.; Finegold, S.M. Isolation of Acidaminococcus fermentans and Megasphaera elsdenii from normal human feces. Appl. Microbiol. 1974, 27, 274–275. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, D.; Wiesmayr, S.; Ledinek, M. Peptostreptococcus elsdenii from the caecum of pigs. J. Gen. Microbiol. 1970, 64, 123–126. [Google Scholar] [CrossRef]
- Yutin, N.; Galperin, M.Y. A genomic update on clostridial phylogeny: Gram-negative spore formers and other misplaced clostridia. Environ. Microbiol. 2013, 15, 2631–2641. [Google Scholar] [CrossRef]
- Sarmikasoglou, E.; Faciola, A.P. Ruminal bacteria lipopolysaccharides: an immunological and microbial outlook. J. Animal Sci. Biotechnol. 2022, 13, 41. [Google Scholar] [CrossRef]
- Shetty, S.A.; Marathe, N.P.; Laniekar, V.; Ranade, D.; Shouche, Y.S. Comparative genome analysis of Megasphaera sp. reveals niche specialization and its potential role in the human gut. Plos One. 2013, 8, 1–13. [Google Scholar] [CrossRef]
- Rogosa, M. Transfer of Peptostreptococcus elsdenii Gutierrez et al. to a new genus, Megasphaera [M. elsdenii (Gutierrez et al.) comb. nov.]. Int. J. Syst. Bacteriol 1972, 21, 187–189. [Google Scholar] [CrossRef]
- Bucci, M. Cell motility: Bigger and faster. Nat. Chem. Biol. 2015, 11, 381. [Google Scholar] [CrossRef]
- Levin, P.A.; Angert, E.R. Small but mighty: cell size and bacteria. Cold Spring Harbor Perspect. Biol. 2015, 7, a019216. [Google Scholar] [CrossRef] [PubMed]
- Elsden, S.R.; Gilchrist, F.M.C.; Lewis, D.; Volcani, B.E. Properties of a fatty acid forming organism isolated from the rumen of sheep. J. Bacteriol. 1956, 72, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Davis, R.E.; Lindahl, I.L.; Warwick, E.J. Bacterial changes in the rumen during the onset of feed-lot bloat of cattle and characteristics of Peptostreptococcus elsdenii n. sp. Appl. Microbiol. 1959, 7, 16–22. [Google Scholar] [CrossRef]
- Elsden, S.R; Lewis, D. The production of fatty acids by a gram-negative coccus. Biochem. J. 1953, 55, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Kluyver, A.J.; Van Niel, C.B. Prospects for a natural system of classification of bacteria. Zentralbl. Bakteriol. Parisitenk. Abt. 1935, 94, 369–403. [Google Scholar]
- Piknova, M.; Bires, O.; Javorsky, P.; Pristas, P. Limited genetic variability in Megasphaera elsdenii strains. Folia Microbiol. 2006, 51, 299–302. [Google Scholar] [CrossRef]
- Marx, H.; Graf, A.B.; Tatto, N.E.; Thallinger, G.G.; Mattanovich, D; Saueri, M. Genome sequence of the ruminal bacterium Megasphaera elsdenii. J. Bacteriol. 2011, 193, 5578–5579. [Google Scholar] [CrossRef]
- Hatmaker, E.A.; Klingeman, D.M.; O’Dell, K.B.; Riley, L.A.; Papanek, B.; Guss, A.M. Complete genome sequences of two Megasphaera elsdenii strains, NCIMB 702410 and ATCC 25940. Microbiol. Res. Announc. 2019, 8, e01430–18. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Ghosh, T.S.; Das, B. Whole-genome sequence of a Megasphaera elsdenii strain isolated from the gut of a healthy Indian adult subject. Genome Announc. 2017, 5, e01033–17. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gutierrez, C.A.; Aylward, F.O. Genome size distributions in bacteria and archaea are strongly linked to evolutionary history at broad phylogenetic scales. PLoS Genet. 2022, 18, e1010220. [Google Scholar] [CrossRef] [PubMed]
- Suen, G.; Weimer, P.J.; Stevenson, D.M.; Aylward, F.O.; Boyum, J. The complete genome sequence of Fibrobacter succinogenes S85 reveals a cellulolytic and metabolic specialist. PLoS ONE, 2011, 6, e18814. [Google Scholar] [CrossRef] [PubMed]
- Hobson, P.N.; Mann, S.O.; Oxford, A.E. Some studies on the occurrence and properties of a large Gram-negative coccus from the rumen. J. Gen. Microbiol. 1958, 19, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.S.; Bryant, M.P. The rumen bacteria. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Springer Dordrecht, United Kingdom, 1988; pp.10-72.
- Stewart, C.S.; Fonty, G.; Gouet, P.H. The establishment of rumen microbial communities. Anim. Feed Sci. Technol. 1988, 21, 69–97. [Google Scholar] [CrossRef]
- Mackie, R.I.; Gilchrist, F.M.C. Changes in lactate-producing and lactate-utilizing bacteria in relation to pH in the rumen of sheep during stepwise adaptation to a high-concentrate diet. Appl. Environ. Microbiol. 1979, 38, 422–430. [Google Scholar] [CrossRef]
- Yang, H.E.; Zotti, C.A.; McKinnon, J.J.; McAllister, T.A. Lactobacilli are prominent members of the microbiota involved in the ruminal digestion of barley and corn. Front. Microbiol. 2018, 9, 718. [Google Scholar] [CrossRef] [PubMed]
- Ouwerkerk, D.; Klieve, A.V.; Forster, R.J. Enumeration of Megasphaera elsdenii in rumen contents by real-time Taq nuclease assay. J. Appl. Microbiol. 2022, 97, 753–758. [Google Scholar] [CrossRef]
- Klieve, A.V.; Hennessy, D.; Ouwerkerk, D.; Forster, R.J.; Mackie, R.I.; Attwood, G.T. Establishing populations of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 2003, 95, 621–630. [Google Scholar] [CrossRef]
- Větrovský, T.; Baldrian, P. The variability of the 16S rRNA gene in bacterial genomes and its consequences for bacterial community analyses. PLoS One. 2013, 8, e57923. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J.; Stevenson, D.M.; Mertens, D.R. Shifts in bacterial community composition in the rumen of lactating dairy cows under milk-fat depression conditions. J. Dairy Sci. 2010, 93, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Palmonari, A.; Stevenson, D.M.; Mertens, D.R.; Cruywagen, C.W.; Weimer, P.J. pH dynamics and bacterial community composition in the rumen of lactating dairy cows. J. Dairy Sci. 2010, 93, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Mizrahi, I. Similarity of the ruminal bacteria across individual lactating cows. Anaerobe. 2012, 2012 18, 338e343. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Rico, D.; Preston, S.; Risser, J.; Harvatine, K. Rapid changes in key ruminal microbial populations during the induction of and recovery from diet-induced milk fat depression in dairy cows. Br. J. Nutr. 2015, 114, 358–367. [Google Scholar] [CrossRef]
- Yohe, T.T.; Enger, B.D.; Wang, L.; Tucker, H.L.M.; Ceh, C.A.; Parsons, C.L.M.; Yu, Z.; Daniels, K.M. Does early-life administration of a Megasphaera elsdenii probiotic affect long-term establishment of the organism in the rumen and alter rumen metabolism in the dairy calf? J. Dairy Sci. 2018, 101, 1747–1751. [Google Scholar] [CrossRef]
- de Melo, H.S.A.; Ítavo, L.C.V.; de Castro, A.P.; Ítavo, C.C.B.F.; de Araújo Caldas, R.; Mateus, R.G.; Niwa, M.V.G.; de Moraes, G.J.; da Silva Zornitta, C.; Gurgel, A.L.C.; Benchaar, C. Bacterial species in the ruminal content of steers fed oilseeds in the diet. Trop. Anim. Health. Prod. 2022, 54, 396. [Google Scholar] [CrossRef]
- Park, T.; Cersosimo, L.M.; Radloff, W.; Zanton, G.I.; Li, W. The rumen liquid metatranscriptome of post-weaned dairy calves differed by pre-weaning ruminal administration of differentially-enriched, rumen-derived inocula. Anim. Microbiome, 2022, 4, 4. [Google Scholar] [CrossRef]
- Fernando, S.C.; Purvis, H.T. II,, Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; DeSilva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol., 2010, 76, 7482–7490. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl. Environ. Microbiol. 2009, 75, 7115–7124. [Google Scholar] [CrossRef] [PubMed]
- McCann, J.C.; Luan, S.; Cardoso, F.C.; Derakhshani, H.; Khafipour, E.; Loor, J.J. Induction of subacute ruminal acidosis affects the ruminal microbiome and epithelium. Front Microbiol. 2016, 7, 701. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Fan, Y.; Wang, H. Lactate uptake in the rumen and its contributions to subacute rumen acidosis of goats induced by high-grain diets. Front. Vet. Sci. 2022, 9, 964027. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Yang, Y.; Lin, C.; Zhang, J.; Pan, J.; Luo, L.; Wu, G.; Awasthi, M.K.; Yan, B. Caproic acid production from anaerobic fermentation of organic waste – Pathways and microbial perspective. Renew. Sust. Energy Revs., 2023, 175, 113181. [Google Scholar] [CrossRef]
- Weimer, P.J.; Moen, G.N. Quantitative analysis of growth and volatile fatty acid production by the anaerobic ruminal bacterium Megasphaera elsdenii T81. Appl. Microbiol. Biotechnol. 2013, 97, 4075–4081. [Google Scholar] [CrossRef]
- Weimer, P.J.; da Silva Cabral, L.; Cacite, F. Effects of ruminal dosing of Holstein cows with Megasphaera elsdenii on milk fat production, ruminal chemistry, and bacterial strain persistence. J. Dairy Sci. 2015, 98, 8078–8092. [Google Scholar] [CrossRef] [PubMed]
- Bryant, M.P.; Robinson, I.M. Some nutritional characteristics of predominant culturable ruminal bacteria. J. Bacteriol. 1962, 84, 605–614. [Google Scholar] [CrossRef]
- Forsberg, C.W. Nutritional characteristics of Megasphaera elsdenii. Can. J. Microbiol. 1978, 24, 981–985. [Google Scholar] [CrossRef]
- Lee, N.R.; Lee, C.H.; Lee, D.Y.; Park, J.B. Genome-scale metabolic network reconstruction and in silico analysis of hexanoic acid producing Megasphaera elsdenii. Microorganisms 2020, 8, 539. [Google Scholar] [CrossRef]
- Wallace, R.J. Catabolism of amino acids by Megasphaera elsdenii LC1. Appl. Environ. Microbiol., 1986, 51, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, J.R.; LaVera, R.; Russell, J.B. Amino acid deamination by ruminal Megasphaera elsdenii strains. Curr. Microbiol. 2002, 45, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Baldwin, R.L. Comparison of maintenance energy expenditures and growth yields among several rumen bacteria grown on continuous culture. Appl. Environ. Microbiol. 1979, 37, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Dombrowski, D.B. Effect of pH on the efficiency of growth by pure cultures of rumen bacteria in continuous culture. Appl. Environ. Microbiol. 1980, 39, 604–610. [Google Scholar] [CrossRef]
- Panikov, N.S. True and illusory benefits of modeling: comment on “Genome-scale metabolic network reconstruction and in silico analysis of hexanoic acid producing Megasphaera elsdenii. Microorganisms 2020, 8, 539”. Microorganisms 2020, 8, 1742. [Google Scholar] [CrossRef] [PubMed]
- Marounek, M.; Fliegrova, K.; Bartos, S. Metabolism and some characteristics of ruminal strains of Megasphaera elsdenii. Appl. Environ. Microbiol. 1989, 55, 1570–1573. [Google Scholar] [CrossRef]
- Russell, J.B.; Baldwin, R.L. Substrate preferences in rumen bacteria: evidence of catabolite regulatory mechanisms. Appl. Environ. Microbiol. 1978, 36, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Baldwin, R.L. Comparison of substrate affinities among several rumen bacteria: a possible determinant of rumen bacterial competition. Appl. Environ. Microbiol. 1979, 37, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Hino, T.; Shimada, K.; Maruyama, T. Substrate preference in a strain of Megasphaera elsdenii, a ruminal bacterium, and its implications in propionate production and growth competition. Appl. Environ. Microbiol. 1994, 60, 1827–1831. [Google Scholar] [CrossRef]
- Hino, T.; Miyazaki, K.; Kuroda, S. Role of extracellular acetate in the fermentation of glucose by a ruminal bacterium, Megasphaera elsdenii. J. Gen. Appl. Microbiol. 1991, 37, 121–129. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol Rev. 1977, 31, 100–180. [Google Scholar] [CrossRef] [PubMed]
- Hino, T.; Kuroda, S. Presence of lactate dehydrogenase and lactate racemase in Megasphaera elsdenii grown on glucose or lactate. Appl. Environ. Microbiol. 1993, 59, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, R.; Altman, E.; Eiteman, M.A. Lactate and acrylate metabolism by Megasphaera elsdenii under batch and steady-state conditions. Appl. Environ. Microbiol. 2012, 78, 8564–8570. [Google Scholar] [CrossRef]
- Counotte, G.H.; Prins, R.A.; Janssen, R.H.A.M.; DeBie, M.J.A. Role of Megasphaera elsdenii in the fermentation of DL-[2-13C]-lactate in the rumen of dairy cattle. Appl. Environ. Microbiol. 1981, 42, 649–655. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M. Brock - Biology of Microorganisms, 11th ed.; Prentice-Hall International: Upper Saddle River, NJ, USA, 2006; p. 157. [Google Scholar]
- Moat, A.G.; Foster, J.W. Microbial Physiology. 3rd ed. John Wiley & Sons: New York, 1995, p.26.
- Jin, Q.; Kirk, M.F. pH as a primary control in environmental microbiology: 1. Thermodynamic perspective. Front. Environ. Sci. 2018, 6, 21. [Google Scholar] [CrossRef]
- Tortora, G.; Funke, B.; Case, C. Microbiology: An Introduction. 12thed. Benjamin-Cummings: Boston. 2015, p.152.
- Russell, J.B. Rumen Microbiology and Its Role in Ruminant Nutrition. James B. Russell Publications, Ithaca, USA, 2002. Available at https://www.ars.usda.gov/research/software/download/?softwareid=409.
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: the current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Nocek, J.E. Bovine acidosis: implications on laminitis. J. Dairy Sci. 1997, 80, 1005–1028. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Sharp, W.M.; Baldwin, R.L. The effect of pH on maximum bacterial growth rate and its possible role as a determinant of bacterial competition in the rumen. J. Anim. Sci. 1979, 48, 251–255. [Google Scholar] [CrossRef]
- Therion, J.T.; Kistner, A.; Kornelius, J.H. Effect of pH on growth rates of rumen amylolytic and lactilytic bacteria. Appl. Environ. Microbiol. 1982, 44, 428–434. [Google Scholar] [CrossRef]
- Waldrip, H.M.; Martin, S.A. Effects of an Aspegillus oryzae fermentation extract and other factors on lactate utilization by the ruminal bacterium Megasphaera elsdenii. J. Anim. Sci. 1993, 71, 2770–2776. [Google Scholar] [CrossRef]
- Noyes, N.R.; Yang, X.; Linke, L.M.; Magnuson, R.J.; Cook, S.; Zaheer, R.; Yang, H.; Woerner, D.R.; Geornaras, I.; Mcart, J.A.; et al. Characterization of the resistome in manure, soil and wastewater from dairy and beef production systems. Sci. Reports 2016, 6, 24645. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.; McAllister, T.A. Antimicrobial usage and resistance in beef cattle production. J. Anim. Sci. Biotechnol. 2016, 7, 1–22. [Google Scholar] [CrossRef] [PubMed]
- El Akkad, I.; Hobson, P. Effect of antibiotics on some rumen and intestinal bacteria. Nature. 1966, 209, 1046–1047. [Google Scholar] [CrossRef]
- Fulghum, R.S.; Baldwin, B.B.; Williams, P.P. Antibiotic susceptibility of anaerobic ruminal bacteria. Appl. Microbiol. 1968, 16, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Bisset, J.; Stewart, C.S. The isolation of tetracycline-resistant strains of strictly anaerobic bacteria from the rumen. Lett. Appl. Microbiol. 1988, 5, 113–115. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Taylor, M.B. Susceptibility and resistance of ruminal bacteria to antimicrobial feed additives. Appl. Environ. Microbiol. 1987, 53, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Wang, C. L.; Baldwin, B.B.; Fulghum, R.S.; Williams, P.P. Quantitative antibiotic sensitivities of ruminal bacteria. Appl. Microbiol. 1969, 18, 677–679. [Google Scholar] [CrossRef]
- Nelson, R.S.; Peterson, D.J.; Karp, E.M.; Beckham, G.T.; Salvachua, D. Mixed carboxylic acid production by Megasphaera elsdenii from glucose and lignocellulosic hydrolysate. Fermentation 2017, 3, 10. [Google Scholar] [CrossRef]
- Russell, J.B. Another explanation for the toxicity of fermentation acids at low pH: anion accumulation versus uncoupling. J. Appl. Microbiol. 1992, 73, 363–370. [Google Scholar] [CrossRef]
- Wolin, M.J. The rumen fermentation: a model for microbial interactions in anaerobic ecosystems. Adv. Microbial Physiol., 1979, 3, 49–77. [Google Scholar] [CrossRef]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Cotta, M.A.; Dombrowski, D.B. Rumen bacteria competition in continuous culture: Streptococcus bovis versus Megasphaera elsdenii. Appl. Environ. Microbiol. 1981, 41, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shen, Y.; Wang, C.; Ding, L.; Zhao, F.; Wang, M.; Fu, J.; Wang, H. Megasphaera elsdenii lactate degradation pattern shifts in rumen acidosis models. Front. Microbiol. 2019, 10, 162. [Google Scholar] [CrossRef]
- Kung, L.J.; Hessian, A.O. Preventing in vitro lactate accumulation in ruminal fermentations by inoculation with Megasphaera elsdenii. J. Anim. Sci. 1995, 73, 250–256. [Google Scholar] [CrossRef]
- Ricke, S.C.; Martin, S.A.; Nisbet, D.J. Ecology, metabolism, and genetics of ruminal selenomonads. Crit. Rev. Microbiol. 1996, 22, 27–65. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Yamada-Narita, S.; Abe, N.; Onodera, T.; Eiichiro Kan, E.; Kojima, S.; Miyazaki, T.; Yamamoto, Y.; Oguchi, A.; Ankai, A.; et al. Complete genome sequence of Selenomonas ruminantium subsp. lactilytica will accelerate further understanding of the nature of the class Negativicutes. FEMS Microbiology Letters 2015, 362, fnv050. [Google Scholar] [CrossRef]
- Fan, Y.; Xia, G.; Jin, Y.; Wang, H. Ambient pH regulates lactate catabolism pathway of the ruminal Megasphaera elsdenii BE2-2083 and Selenomonas ruminantium HD4. J. Appl. Microbiol. 2022, 132, 2661–2672. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Wood, W.A.; Emery, R.S. Conversion of lactate-C14 to propionate by the rumen microbiota. J. Bacteriol. 1962, 83, 907–913. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Wood, W.A.; Emery, R.S. Conversion of glucose-C14 to propionate by the rumen microbiota. J. Bacteriol. 1963, 85, 1346–1349. [Google Scholar] [CrossRef]
- Sarmikasoglou, E.; Ferrell, J.; Vinyard, J.R.; Flythe, M.D.; Tuanyok, A.; Faciola, A.P. Effects of ruminal lipopolysaccharides on growth and fermentation end products of pure cultured bacteria. Sci. Reports 2023, 12, 15932. [Google Scholar] [CrossRef]
- Bandarupalli, V.V.K.; St-Pierre, B. Metagenomics-based analysis of candidate lactate utilizers from the rumen of beef cattle. Microorganisms 2023, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Miura, H.; Horguchi, M.; Matsumoto, T. Nutritional interdependence among rumen bacteria, Bacteroides amylophilus, Megasphaera elsdenii, and Ruminococcus albus. Appl. Environ. Microbiol. 1980, 40, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Shabat, S.K.B.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Miller, M.E.B.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef] [PubMed]
- Betancour-Murillo, C.L.; Aguilar-Marin, S.B.; Jovel, J. Prevotella: A key player in ruminal metabolism. Microorganisms. 2022, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Jewell, K.A.; McCormick, C.; Odt, C.L.; Weimer, P.J.; Suen, G. Ruminal bacterial community composition in dairy cows is dynamic over the course of multiple lactations and correlates with feed efficiency. Appl. Environ. Microbiol. 2015, 81, 4697–4710. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J.; Cox, M.S.; Vieira de Paula, T.; Lin, L.; Hall, M.B.; Suen, G. Transient changes in milk production efficiency and bacterial community composition resulting from near-total exchange of ruminal contents between high- and low-efficiency Holstein cows. J. Dairy Sci. 2017, 100, 7165–7182. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–27. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C.; Wallace, R.J.; Moate, P.J.; Mosley, E.E. Board-invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J Anim. Sci. 2008, 86, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Liu, R.H.; Bond, D.; Russell, J.B. Effect of linoleic acid concentration on the conjugated linoleic acid (CLA) production of Butyrivibrio fibrisolvens A38. Appl. Environ. Microbiol. 2000, 66, 5226–5230. [Google Scholar] [CrossRef]
- Kim, Y.J.; Liu, R.H.; Rychlik, J.L.; Russell, J.B. Enrichment of a ruminal bacterium (Megasphaera elsdenii YJ-4) that produces the trans-10, cis-12 isomer of conjugated linoleic acid (CLA). J. Appl. Microbiol. 2002, 92, 976–982. [Google Scholar] [CrossRef]
- Kim, T.W.; Choi, N.J.; Hwangbo, J.; Hsu, J.T.; Lee, S.S.; Song, M.K.; Seo, L.J.; Kim, Y.J. Production of trans-10, cis-12 conjugated linoleic acid by Megasphaera elsdenii YJ-4: physiological roles in the rumen. Asian-Austr. J. Anim. Sci 2005, 18, 1425–1429. [Google Scholar] [CrossRef]
- Muya, M.C.; Nherera, F.V.; Miller, K.A.; Aperce, C.C.; Moshidi, P.M.; Erasmus, L.J. Effect of Megasphaera elsdenii NCIMB 41125 dosing on rumen development, volatile fatty acid production and blood β-hydroxybutyrate in neonatal dairy calves. J. Anim. Physiol. Anim. Nut. 2015, 99, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Susanto, I.; Wiryawab, K.G.; Suharti, S.; Retnani, Y.; Zahera, R. Jayanegara. Evaluation of Megasphaera elsdenii supplementation on rumen fermentation, production performance, carcass traits and health of ruminants: a meta-analysis. Anim. Biosci. 2023, 36, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.A.; Smolenski, W.J.; Greening, R.C.; Ogilvie, M.L.; Bell, R.L.; Barsuhn, K.; Peters, J.P. Prevention of acute acidosis and enhancement of feed intake in the bovine by Megasphaera elsdenii 407A. J. Anim. Sci 1992, 70, 310 (Abstract). [Google Scholar]
- McDaniel, M.R. The effects of dosing feedlot cattle with Megasphaera elsdenii strain NCIMB 41125 prior to the introduction of a grain rich diet. M.S. Thesis, Kansas State University, 2009. https://krex.k-state.edu/bitstream/handle/2097/1666/MichaelMcDaniel2009.pdf;sequence=1.
- Leeuw, K.J.; Siebrits, F.K.; Henning, P.H.; Meissner, H.H. Effect of Megasphaera elsdenii NCIMB 41125 drenching on health and performance of steers fed high and low roughage diets in the feedlot. S. Afr. J. Anim. Sci. 2009, 39, 337–348. [Google Scholar] [CrossRef]
- Henning, P.H.; Horn, C.H.; Steyn, D.G.; Meissner, H.H.; Hagg, F.M. The potential of Megasphaera elsdenii isolates to control ruminal acidosis. Anim. Feed Sci. Technol. 2010, 157, 13–19. [Google Scholar] [CrossRef]
- Henning, P.H.; Horn, C.H.; Leeuw, K.J.; Meissner, H.H.; Hagg, F.M. Effect of ruminal administration of the lactate-utilizing strain Megasphaera elsdenii (Me) NCIMB 41125 on abrupt or gradual transition from forage to concentrate diets. Anim. Feed Sci. Technol. 2010, 157, 20–29. [Google Scholar] [CrossRef]
- Aikman, P.C.; Henning, P.H.; Humphries, D.J.; Hom, C.H. Rumen pH and fermentation characteristics in dairy cows supplemented with Megasphaera elsdenii NCIMB 41125 in early lactation. J. Dairy Sci. 2011, 94, 2840–2849. [Google Scholar] [CrossRef]
- Zebeli, Q.; Terrill, S.J.; Mazzolari, A.; Dunn, S.M.; Yang, W.Z.; Ametaj, B.N. Intraruminal administration of Megasphaera elsdenii modulated rumen fermentation profile in mid-lactation dairy cows. J. Dairy Res. 2012, 79, 16–25. [Google Scholar] [CrossRef]
- Klieve, A.V.; McLennan, S.R.; Ouwerkerk, D. Persistence of orally administered Megasphaera elsdenii and Ruminococcus bromii in the rumen of beef cattle fed a high grain (barley) diet. Anim. Prod. Sci. 2012, 52, 297–304. [Google Scholar] [CrossRef]
- Drouillard, J.S.; Henning, P.H.; Meissner, H.H.; Leeuw, K.J. Short communication: Megasphaera elsdenii on the performance of steers adapting to a high-concentrate diet, using three or five transition diets. S. Afr. J. Anim. Sci. 2012, 42, 195–199. [Google Scholar] [CrossRef]
- Ye, D.; Eastridge, M.L. Oral administration of Megasphaera elsdenii to Jersey cows during early lactation. Profess. Anim. Sci. 2018, 34, 67–74. [Google Scholar] [CrossRef]
- Arik, H.D.; Gulsen, N.; Hayirli, A.; Alatas, M.S. Efficacy of Megasphaera elsdenii inoculation in subacute ruminal acidosis in cattle. J. Anim. Physiol. Anim. Nutr. 2018, 103, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, R.; Alipour, D. Assessment of probiotic effects of isolated Megasphaera elsdenii strains in Mehraban sheep and Holstein lactating cows. Anim. Feed Sci. Technol. 2019, 248, 126–131. [Google Scholar] [CrossRef]
- DeClerck, J.C.; Reeves, N.R.; Miller, M.F.; Johnson, B.J.; Ducharme, G.A.; Rathmann, R.J. Influence of dietary roughage level and Megasphaera elsdenii on feedlot performance and carcass composition of thin cull beef cows fed for a lean market. Transl. Anim. Sci. 2020, 29, 159–169. [Google Scholar] [CrossRef] [PubMed]
- DeClerck, J.C.; Wade, Z.E.; Reeves, N.R.; Miller, M.F.; Johnson, B.J.; Ducharme, G.A.; Rathmann, R.J. Influence of Megasphaera elsdenii and feeding strategies on feedlot performance, compositional growth, and carcass parameters of early weaned, beef calves. Transl. Ani. Sci. 2020, 4, 863–875. [Google Scholar] [CrossRef]
- Mazon, G.; Campler, M.R.; Holcomb, C.; Bewley, J.M.; Costa, J.H.C. Effects of a Megasphaera elsdenii oral drench on reticulorumen pH dynamics in lactating dairy cows under subacute ruminal acidosis challenge. Anim. Feed Sci. Technol. 2020, 261, 1–9. [Google Scholar] [CrossRef]
- Lopes, A.L.; Santos, F.A.P.; Meschiatti, M.; Oliveira, M.O.; Fernandes, J.J.R.; Drouillard, J.S.; Cappellozza, B.I. Effects of Megasphaera elsdenii administration on performance and carcass traits of finishing Bos indicus feedlot cattle. Transl. Anim. Sci. 2021, 28, 1–9. [Google Scholar] [CrossRef]
- Diez-Gonzalez, F.; Callaway, T.R.; Kizoulis, M.G.; Russell, J.B. Grain feeding and the dissemination of acid-resistant Escherichia coli from cattle. Science. 1998, 281, 1666–1668. [Google Scholar] [CrossRef]
- Maher, J.M.; Drouillard, J.S.; Baker, A.N.; Veloso, V. d.A.; Kang, Q.; Kasstner, J.J.; Gregg, S.E. Impact of the probiotic organism Megasphaera elsdenii on Escherichia coli O157:H7 prevalence in finishing cattle. J. Food Protection 2023, 86, 100133. [Google Scholar] [CrossRef]
- Jeon, B.S.; Kim, S.; Sang, B.I. Megasphaera hexanoica sp. nov., a medium-chain carboxylic acid-producing bacterium isolated from a cow rumen. Int. J. Syst. Evol. Microb. 2017, 67, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Lanjekar, V.B.; Marathe, N.P.; Ramana, V.V.; Shouche, Y.S.; Ranade, D.R. Megasphaera indica sp. nov., an obligate anaerobic bacteria isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2014, 64, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Holtzapple, M.T.; Lonkar, S.; Granda, C. Producing biofuels via the carboxylate platform. Chem. Eng. Progr. 2015, 111, 52–57. [Google Scholar]
- Weimer, P.J.; Digman, M.F. Fermentation of alfalfa wet-fractionation liquids to volatile fatty acids by Streptococcus bovis and Megasphaera elsdenii. Biores. Technol. 2013, 142, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Araoka, R.; Kajihara, Y.; Ito, T.; Miyamoto, H.; Kodama, H. Valerate production from Megasphaera elsdenii isolated from pig feces. J. Biosci. Bioeng. 2018, 125, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Roddick, F.A.; Britz, F.A. Production of hexanoic acid by free and immobilised cells of Megasphaera elsdenii: influence of in-situ product removal using ion exchange resin. J. Chem. Technol. Biotechnol. 1997, 69, 383–391. [Google Scholar] [CrossRef]
- Choi, K.; Jeon, BS.; Kim, B.C.; Oh, M.K.; Um, Y.; Sang, B.I. In situ biphasic extractive fermentation for hexanoic acid production from sucrose by Megasphaera elsdenii NCIMB 702410. Appl. Biochem. Biotechnol. 2013, 171, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.S.; Choi, O.; Um, Y.; Sang, B.I. Production of medium-chain carboxylic acids by Megasphaera sp. MH with supplemental electron acceptors. Biotechnol. Biofuels 2016, 9, 129. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Veiga, M.C.; Kennes, C. Effect of pH and medium composition on chain elongation with Megasphaera hexanoica producing C4-C8 fatty acids. Front. Microbiol. 2023, 14, 1281103. [Google Scholar] [CrossRef]
- Lange, J.P.; Price, R.; Ayoub, P.M.; Louis, J.; Petrus, L.; Clarke, L.; Gosselink, H. Valeric biofuels: a platform of cellulosic transportation fuels. Angew. Chem. Int. Ed. 2010, 49, 4479–4483. [Google Scholar] [CrossRef]
- Urban, C.; Xu, J.; Strauber, H.; Dantas, T.R.S.; Muhlenberg, J.; Hartig, C.; Angenent, L.T.; Harnisch, F. Production of drop-in fuels from biomass at high selectivity by combined microbial and electrochemical conversion. Energy Environ. Sci. 2017, 10, 2231–2244. [Google Scholar] [CrossRef]
- Scarborough, M.J.; Lynch, G.; Dickson, M.; McGee, M.; Donohue, T.J.; Noguera, D.R. Increasing the economic value of lignocellulosic stillage through medium-chain fatty acid production. Biotech. Biofuels. 2018, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Yan, B.; Xu, S.; Zhou, J.; Liang, J.; Zhao, J., Tyangi; Wong, J.W.C. Regulation of acidogenic fermentation through exogenous additives for promoting carbon conversion of food waste in two-phase anaerobic system. Biores. Technol. 2023, 368, 128368. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Photomicrograph of crystal violet-stained cells of Megasphaera elsdenii T81 grown in lactate medium. Cells present singly or in pairs, or rarely as chains of 4-8 cells. Bar represents 5 μm. Photo courtesy of J. McClure, USDA-Agricultural Research Service.
Figure 1.
Photomicrograph of crystal violet-stained cells of Megasphaera elsdenii T81 grown in lactate medium. Cells present singly or in pairs, or rarely as chains of 4-8 cells. Bar represents 5 μm. Photo courtesy of J. McClure, USDA-Agricultural Research Service.
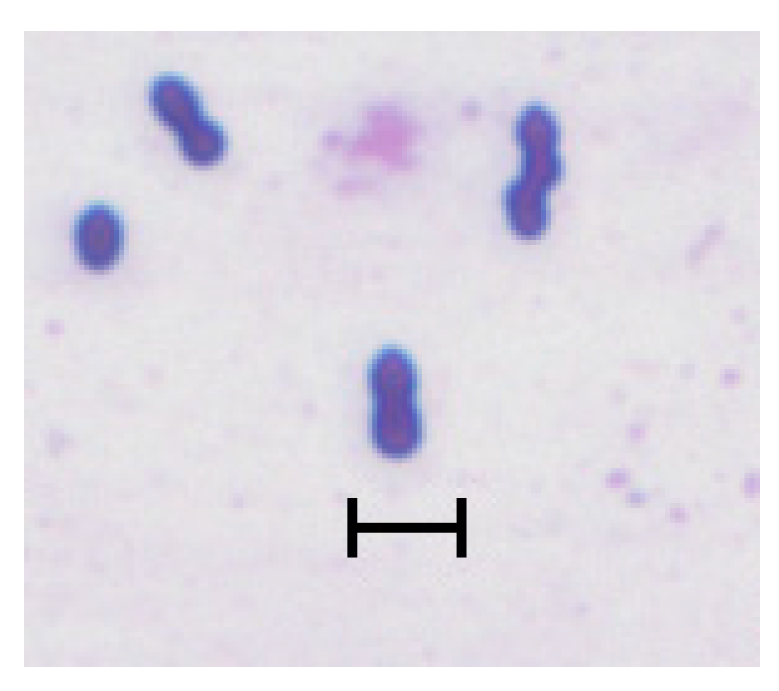
Figure 2.
Unrooted phylogenetic tree for M. elsdenii and related species based on 16S rRNA gene sequences. The tree was constructed using Mr. Bayes software, with Butyrivibrio fibrisolvens as the outgroup. M. elsdenii strains ATCC 25940 and NCIMB 702410 (not shown) were indistinguishable from the type strain DSM 20640T.
Figure 2.
Unrooted phylogenetic tree for M. elsdenii and related species based on 16S rRNA gene sequences. The tree was constructed using Mr. Bayes software, with Butyrivibrio fibrisolvens as the outgroup. M. elsdenii strains ATCC 25940 and NCIMB 702410 (not shown) were indistinguishable from the type strain DSM 20640T.
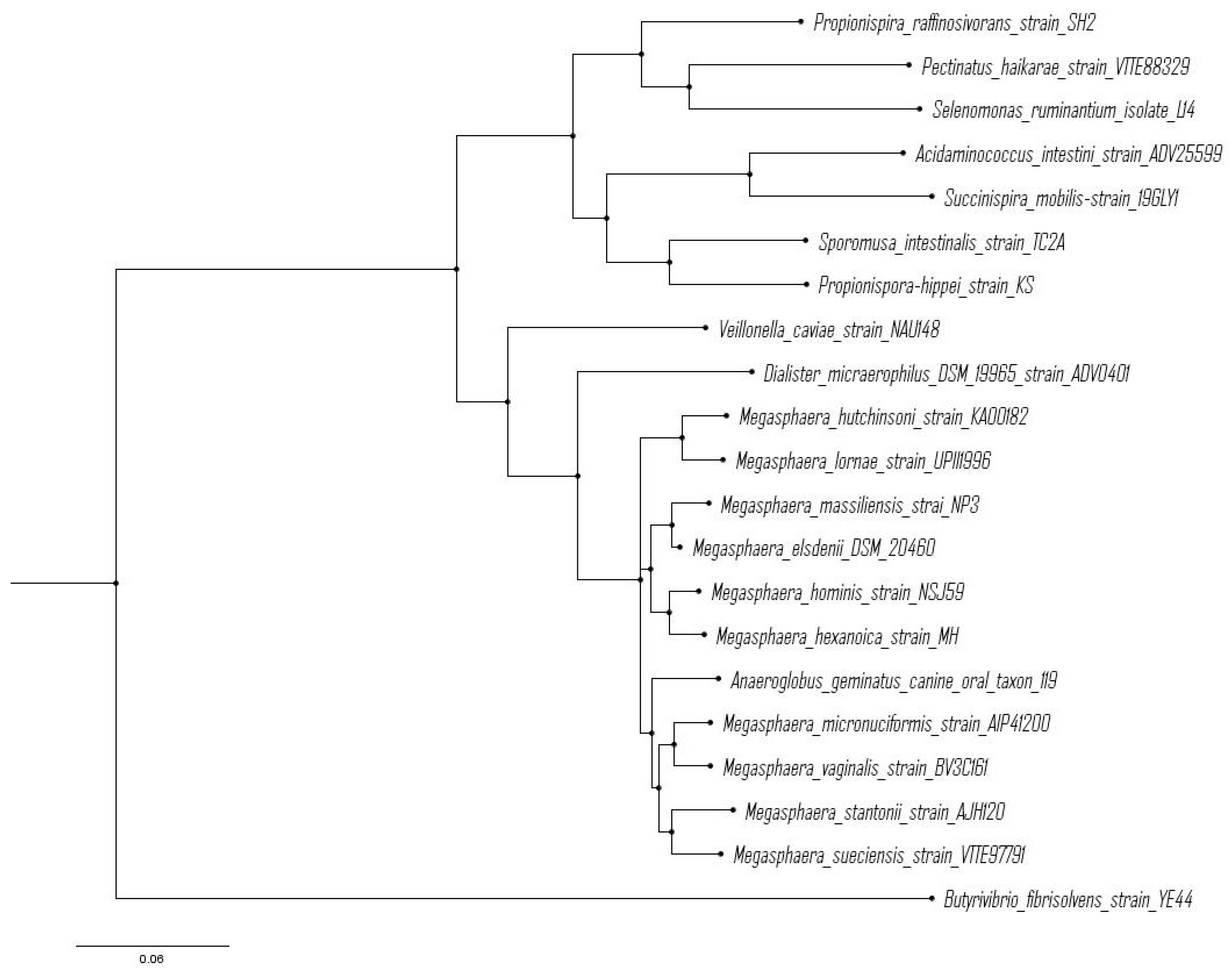
Figure 3.
Cultures of M. elsdenii B159 grown in LTY medium [49] containing (left to right) 0, 1 or 3 mM of added cysteine reducing agent. Blackening of medium is due to FeS precipitate formed following fermentation of cysteine to H2S.
Figure 3.
Cultures of M. elsdenii B159 grown in LTY medium [49] containing (left to right) 0, 1 or 3 mM of added cysteine reducing agent. Blackening of medium is due to FeS precipitate formed following fermentation of cysteine to H2S.

Figure 4.
Simplified fermentation pathway for glucose and lactate by Megasphaera elsdenii, showing the key roles of acrylyl-CoA in the production of odd-chain fatty acids, and acetyl-CoA (AcCoA) in the formation of both odd- and even-chain fatty acids. Glucose suppresses NAD-independent lactate dehydrogenase (iLDH) and lactate racemase (LR), preventing significant production of odd-chain fatty acids from glucose.
Figure 4.
Simplified fermentation pathway for glucose and lactate by Megasphaera elsdenii, showing the key roles of acrylyl-CoA in the production of odd-chain fatty acids, and acetyl-CoA (AcCoA) in the formation of both odd- and even-chain fatty acids. Glucose suppresses NAD-independent lactate dehydrogenase (iLDH) and lactate racemase (LR), preventing significant production of odd-chain fatty acids from glucose.
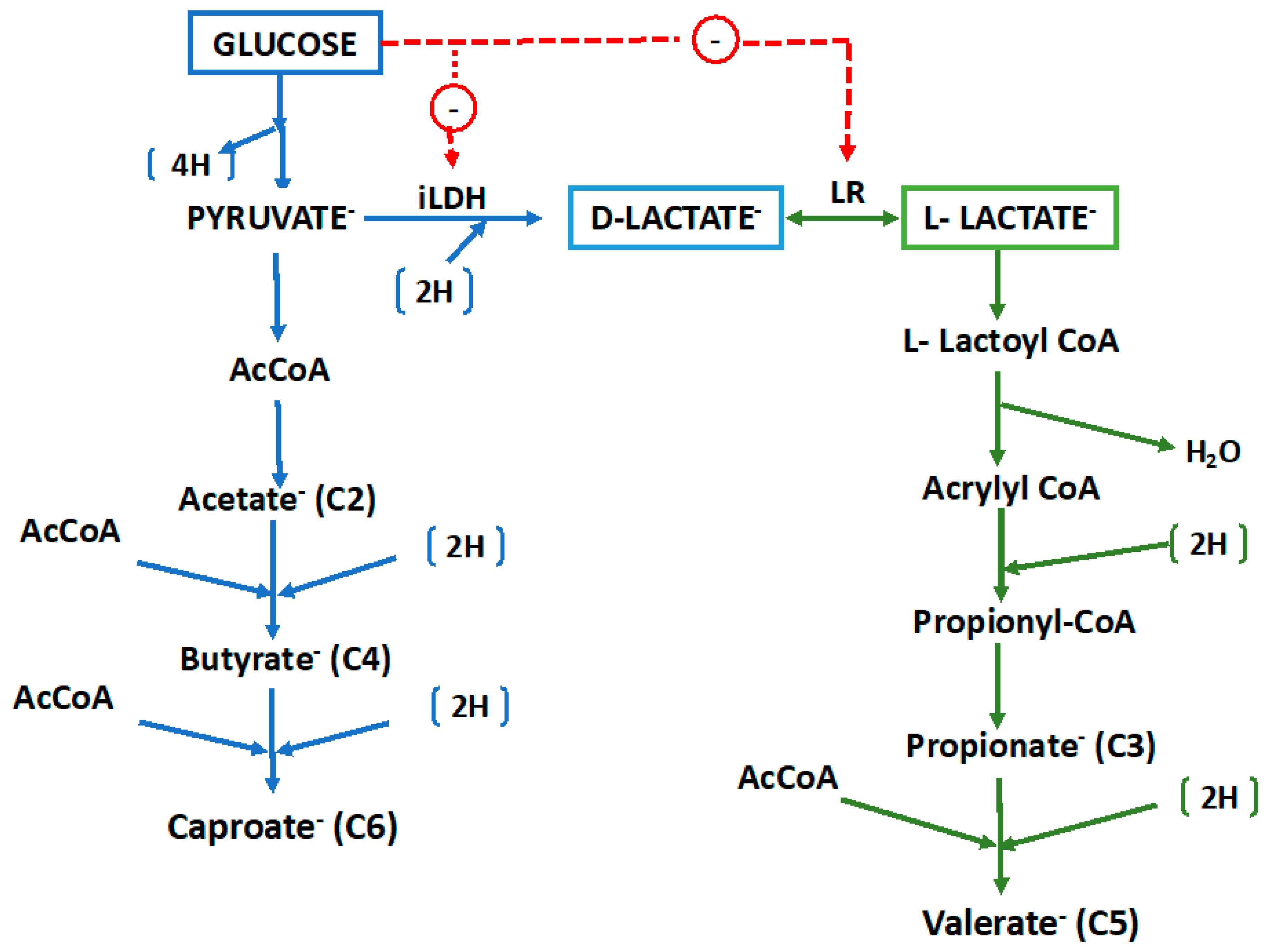
Figure 5.
Bifurcated pathway for the biosynthesis of crotonyl-CoA, the key intermediate in chain-elongation of even-chain volatile fatty acids, by M. elsdenii. Crotonyl-CoA is reduced to butyryl-CoA for butyrate production, or further elongated for caproate production. Adapted from Lee et al. [53].
Figure 5.
Bifurcated pathway for the biosynthesis of crotonyl-CoA, the key intermediate in chain-elongation of even-chain volatile fatty acids, by M. elsdenii. Crotonyl-CoA is reduced to butyryl-CoA for butyrate production, or further elongated for caproate production. Adapted from Lee et al. [53].
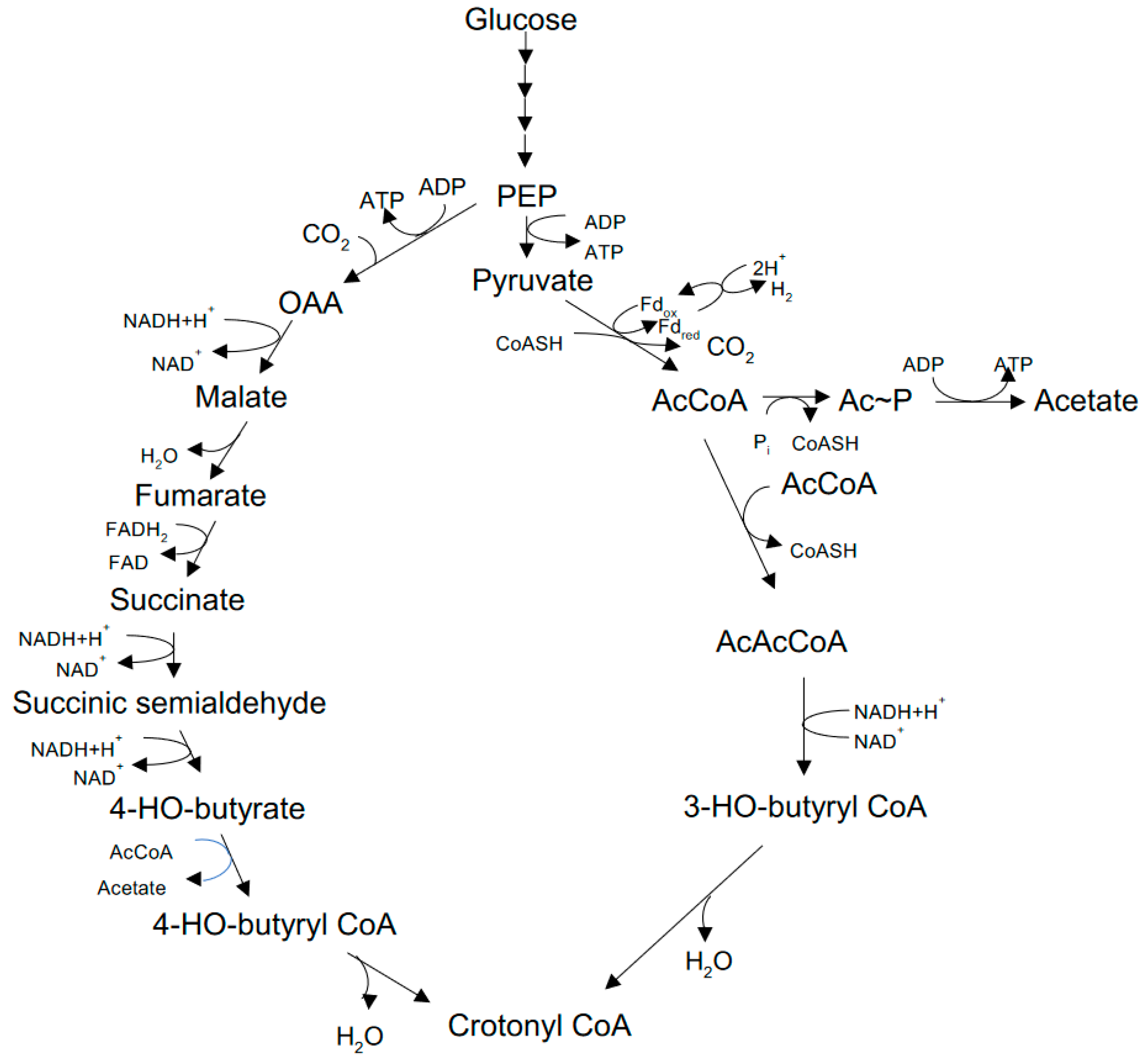

Table 1.
Sequenced genomes of Megasphera elsdenii.
| Strain | Source | Genome size (bp) | Putative number of genes | Average G+C content | Reference |
|---|---|---|---|---|---|
| DSM 20640T a | Rumen | 2,474,718 | 2220 | 54 | [22] |
| ATCC 25940 a | Rumen | 2,478,842 | 2194 | 52.8 | [23] |
| NCIMB 702410 | Rumen | 2,566,193 | 2280 | 52.7 | [23] |
| indica | Human feces | 2,429,033 | 2184 | 53.2 | [24] |
a These strains, though from different culture collections, are considered to have been derived from the same isolate.
Table 2.
Total concentration of the M. elsdenii (LC) or lactate-fermenting bacteria (LFB) in cattle fed with different diets.
Table 2.
Total concentration of the M. elsdenii (LC) or lactate-fermenting bacteria (LFB) in cattle fed with different diets.
| Organism designation | Comparison | Counts/mL | Reference |
|---|---|---|---|
| Large coccus | Young calves (3-9weeks) Adult cattle and sheep |
109 to 109 absent to 105 |
[27] |
| Large coccus | Before bloat Moderate bloat |
< 1 x104 to 2 x107 2 x 106 to 2 x108 |
[18] |
| Total lactate fermenting bacteria | Forage diet High grain diets |
4.8 x 106 1.89 x 1011 |
[30] |
Table 3.
Relative abundance of M. elsdenii in the rumen.
| Animal | n | % RA a | Method b | Notes | Reference |
|---|---|---|---|---|---|
| Holstein cows | 2 | 0.0001 - 0.0011 | qPCR | TMR, 27.5% NDF, 18% CP | [35] |
| Holstein cows | 2 | 0.002 - 0.40 | qPCR | High concentrate diet. RA declined upon monensin feeding and withdrawal | [36] |
| Holstein cows | 2 2 |
0.018 - 0.032 0.320 - 1.946 |
qPCR | Cows not fat-depressed Fat-depressed cows |
[37] |
| Holstein cows | 16 | 0.03 - 0.01 | qPCR | 30% forage/70% concentrate diet | [38] |
| Holstein calf to adult | 5 | 0.0003 - 0.10 | 16S 454 | Calf development study, diet varied over 5 time points from 1 d to 730 d age | [39] |
| Holstein cows | 8 | 0.0001- 0.0005 0.0025 - 0.0038 |
qPCR | Control or after recovery from MFD After induction of MFD |
[40] |
| Holstein heifer calves | 6 | 0 to 0.70 | 16S Illumina | Dosed with M. elsdenii probiotic at d14; RA declined to below uninoculated controls by d84 | [41] |
| Crossbred steers | 6 | 0.02 to 0.10 | 16S Illumina | 6 diets varying in oilseed source, sampled 14 d after each dietary switch | [42] |
| Holstein bull calves (post-weaning) | 5 5 |
0.379 0.790 |
RNA-seq | Inoculated with protozoal enrichment culture Uninoculated controls |
[43] |
a Relative abundance, expressed as a percentage of total 16S rRNA gene copy number. b Quantification method: qPCR, quantitative polymerase chain reaction; 16S Illumina, 16S rRNA gene sequencing on the Illumina platform. 16S 454, 16S rRNA gene sequencing on the 454 pyrotag sequencing platform. RNA-seq, metatranscriptome analysis on the Illumina platform.
Table 4.
Fermentation products of various strains of M. elsdenii grown in batch culture on D-glucose or DL-lactate.
Table 4.
Fermentation products of various strains of M. elsdenii grown in batch culture on D-glucose or DL-lactate.
| Strain | Time (h) | From Glucose | From Lactate | Reference | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Si d (mM) | Mol % product e | Si d (mM) | Mol % product e | |||||||||||||
| For (C1) |
Ace (C2) | Pro (C3) | But (C4) | Val (C5) |
Cap (C6) | For (C1) |
Ace (C2) | Pro (C3) | But (C4) | Val (C5) |
Cap (C6) | |||||
| LC | nra | nr | 26.7 | 22.4 | 22.4 | 28.6 | [18] | |||||||||
| LC-3 | 168 | 27.8 | 4.0 | 7.2 | 1.8 | 14.3 | 11.1 | 61.6 | 62 | 0 | 18.7 | 32.0 | 21.6 | 27.7 | 0 | [17] |
| LC-S | nra | 12.9 | 22.1 | 26.3 | 22.1 | 27.9 | 1.6 | [17] | ||||||||
| nra | nra | 55.5 | + | 33.9 | 1.3 | 52.1 | 6.7 | 5.9 | 111 | 35.5 | 38.2 | 13.7 | 12.6 | - | [27] | |
| B159 b | 72 | 27.8 | 20.4 | - | 77.5 | 2.0 | + | 17.7 | 31.3 | 37.5 | 12.5 | 18.8 | - | [52] | ||
| L8 | 20 | 10 | 27.9 | - | - | 48.6 | 2.7 | 20.7 | 20 | - | 29.7 | 30.6 | 18.0 | 21.6 | - | [59] |
| LC-1 | 4.2 | - | - | 59.4 | 19.8 | 16.7 | - | 51.3 | 38.0 | 6.8 | 3.8 | - | ||||
| J1 | 19.4 | - | - | 37.4 | 21.6 | 21.6 | - | 24.3 | 27.4 | 39.4 | - | 8.8 | ||||
| AW106 | 0.7 | 29.2 | - | 47.4 | 11.7 | 10.9 | 9.9 | 40.1 | 29.2 | 7.0 | 4.7 | - | ||||
| NIAH102 | 10 | 11.1 | 32.0 | - | 61.5 | 0.6 | 6.0 | [63] | ||||||||
| T81 c | nr | 35 | 43.1 | 50.4 | 3.4 | [66] | ||||||||||
| T81 c | 48 | 41 | - | 16.4 | - | 45.5 | 0.7 | 37.3 | 100 | 36.8 | 37.4 | 13.1 | 10.7 | 0.1 | [49] | |
a nr, not reported, b = ATCC 17752, c = ATCC 17753, d initial substrate concentration, e Molar amount of each VFA as a percentage of total moles of product; most studies do not report amount of substrate consumed, necessary for calculation of product yield per unit substrate consumed. For, formate; Ace, acetate; Pro, propionate; But, butyrate; Val, valerate; Cap, caproate.
Table 5.
Comparison of Megasphaera elsdenii and Selenomonas ruminantium lactilytica.
| Property | M. elsdenii | S. ruminantium lactilytica |
| Phylum | Firmicutes | Firmicutes |
| Class | Clostridia | Negativicutes |
| Gram reaction | Negative | Positive |
| Motility | Nonmotile | Motile |
| Genome | Single chromosome, ~2.5 Mbp | Single chromosome 3,003,680 bp, 9 plasmids 2619 -285,449 bp. Total genome size 3,631,933 bp |
| Preferred growth substrate | D- or L-Lactate | D-lactate |
| Other growth substrates | Glucose, Fructose, Maltose, Amino acids | Glucose, Cellobiose, Mannose, Xylose, Arabinose, Mannitol, Glycerol |
| Lactate fermentation pathway | Acrylate | Succinate |
| Lactate utilization enzymes | NAD-independent L-lactate dehydrogenase, Lactate racemase | NAD-independent D-lactate dehydrogenase. No L-LDH, No Lactate racemase |
| VFA fermentation productsa | From lactate, C2, C3, C4, C5 From glucose: C2, C4, C6 |
From lactate: C2, C3 From glucose: Lactate, Acetate |
| Response to monensin | Relatively resistant | Sensitive |
| mb on glucose | 0.187 | 0.021 |
a C2, acetate; C3, propionate; C4, butyrate; C5, valerate; C6, caproate. b maintenance coefficient (g glucose . g cells-1 . h-1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



