
Submitted:
31 December 2023
Posted:
02 January 2024
You are already at the latest version
Abstract
Exosomes are small subtypes of extracellular vesicles (EVs) naturally released by different types of cells into their environment. Their physiological roles appear to be multiple, and many aspects of their biological activities remain to be understood. These vesicles can transport and deliver a variety of cargoes, including proteins, metabolites, nucleic acids, and lipids. Consequently, it is argued that exosomes may serve as unconventional secretory vesicles, playing a crucial role as important vectors for intercellular communication and the maintenance of homeostasis. Exosome production and content can vary under several stresses or modifications in the cell microenvironment, influencing cellular responses both qualitatively and quantitatively. Thus, the analysis of EVs from multiple biofluids, due to its ease of implementation, has become a booming tool for monitoring various pathologies, particularly cancer, as we shall outline further on. During infectious processes, exosomes are described as double-edged swords, displaying both beneficial and detrimental effects. Indeed, during a viral infection, the production of EVs and circulating exosomes is often enhanced to signal danger, stimulate immunity, and possibly serve as pathological biomarkers. Conversely, viruses can disrupt or hijack the exosome content and production. In this review, we discuss the particular and ambiguous functions of exosomes in infectious contexts. Furthermore, we explore the vectoring capacity of exosomes in light of the extensive literature examining their function in mounting adaptive immune responses. Exosomes provide an interesting platform for antigen presentation and could therefore serve as novel therapeutic targets or be used in vaccine strategies.
Keywords:
Exosomes
; extracellular vesicles
; cargoes
; pathological biomarkers
; infection
; viruses
; flaviviruses
; antigen presentation
; vaccine
1. Introduction
Cellular communication is an essential aspect of multicellular organism physiology. It relies on signals emitted by cells, capable of circulating to specifically reach target cells equipped to receive the message. The mechanisms of signal production and reception have been extensively explored for decades, notably in the field of immune cell interactions, where coordinated immune responses are required to eliminate pathogens or tumor cells. Intercellular signaling between immune cells was initially thought to occur either through cell-cell contact or soluble factors (such as cytokines) until the late 1990s when it was identified that extracellular vesicles (EVs) were involved in dendritic and T cells crosstalk [1]. Since then, their biological functions were clarified and EVs have emerged as key players in intercellular communication. They provide a unique facility for transporting a wide variety of signaling molecules to recipient cells. Research efforts in the field have revealed multiple roles for EVs including that of antigen-presenting device [2]. Their characteristics also opened up attractive perspectives for their use as biomarkers, and their possible engineering turned them into therapeutic tools for drug delivery [3]. In recent years, the development of methods for EVs purification, imaging, size measurement and content analysis through lipidomic, proteomic and other studies have revealed that EVs and their cargoes are highly diverse. Their biogenesis, production and contents differ according to cell type, metabolic activity, normal and pathological conditions. This diversity makes understanding the biological activities of EVs a complex task. In this review, we have chosen to focus on exosomes among EVs. In light of the latest data in the literature, our aim is to provide insight into known features of exosomes, and how their specific loading can lead to specific physiological responses, thus constituting indicators of health and pathological situations as biomarkers, particularly in the well-documented field of cancer pathologies. We will then focus on infectious diseases, considering the role of exosomes in the organism's response to viral infections and how viruses, notably flaviviruses, hijack these transport systems. In the final section, we will review what is known about exosomes as tools for antigen presentation in promising vaccine strategies.
2. Exosomes are small, specialized extravesicular vesicles (EVs)
Extracellular vesicles (EVs) refer to all lipid vesicles released by cells. Despite an ever-changing nomenclature, EVs are characterized by their biogenesis pathways, morphology, molecular composition and size [4,5]. According to a substantial body of literature, the size of EVs serves as a reliable indicator for determining their nature and origin. This led to the identification of three subcategories of EVs: firstly, blebs and apoptotic bodies, with sizes reaching up to 5 µm; secondly, microvesicles, also called ectosomes, ranging between 100 and 500 nm, or even up to 1,000 nm; thirdly, exosomes, exhibiting diameters varying between 30 and 150 nm [6,7]. The first two subcategories are produced by shedding from the cell surface, with blebs generated in the advanced stages of apoptosis, and their membrane is derived from the plasma membrane of the donor cell. In contrast, exosomes are formed through biogenesis from endosomal membranes and are released after exocytosis, via cell secretion pathways. EVs are produced continuously from a wide variety of cell types and the capacity of cells to release vesicles appears to be a highly conserved mechanism throughout evolution [8,9]. The consequence of this mechanism is that large quantities of EVs can be detected in extracellular compartments. In humans, exosomes can particularly be found in almost all body fluids including blood, cerebrospinal fluid, saliva, breast milk and urine [6,10,11,12]. When first discovered in the late 1980s, exosomes were initially considered as cellular waste. Subsequently, it has been shown that EVs play a crucial role in intercellular communication, both in physiological and pathological conditions, in order to maintain homeostasis at cellular and systemic levels [13,14]. This role is fulfilled by their ability to encapsulate and transport diverse and specific complex cargoes, including proteins, lipids, metabolites, nucleic acids with various types of RNAs and even short DNA molecules. This characteristic positions them as true unconventional secretory systems [15]. EVs have been described as acting either proximally to the site of their production, exhibiting autocrine or paracrine signaling, or distantly from their secretion site, serving as stable conveyors of signaling molecules with endocrine action [16]. Exosome interaction with the surface of recipient cells, and their eventual uptake may elicit pleiotropic responses with functional and/or phenotypic changes [17,18]. The in vitro biological effects of exosome exposure have been extensively documented. However, the exact physiological or pathological roles of exosomes in vivo are yet to be fully understood, presenting a significant area for further research. Given their potential as molecular signatures and prognostic biomarkers, they have become a tool to enhance our understanding of several pathologies. Their production and behavior in the biology of viral infections deserve particular attention. Moreover, their ability to vectorize different molecules and present antigens makes them powerful new tools in the field of vaccinology.
2.1. Exosomes biogenesis
Exosomes are small EVs that have their own biogenesis pathway within cells before being released into the extracellular environment. Krylova and Feng recently extensively reviewed the literature on the biogenesis mechanisms of exosomes [19]. The vesicle formation occurs in cellular compartments called multivesicular bodies (MVBs), which are produced by the maturation of endosomal compartments (Figure 1) [20]. Endosomes are intracellular convergence sites for vesicular trafficking originating mainly from the cell surface [21]. A broad range of substances can pass through the endomembrane compartments [22,23]. Those internalized by receptor-dependent endocytic pathways transit with their receptors in early endosomes [24]. The endosomal compartment also includes recycling components engaged in centrifugal vesicular trafficking back to the plasma membrane, as well as components undergoing maturation into late endosomes with a more acidic content, in proximity to autophagosomes. The remodeling of late endosomes into MVBs occurs after the invagination of their membrane. This results in the formation of a multitude of small vesicles called intraluminal vesicles (ILVs) [25,26]. During this remodeling process, ILVs can be loaded with numerous cellular components such as cytosolic proteins, metabolites, nucleic acids, and lipids. An exocytosis mechanism enabling an MVB to fuse with the cell membrane leads to the release of ILVs into the extracellular compartment. They then become exosomes and join the pool of EVs. Otherwise, MVBs proceed to the lysosomes to degrade their contents. Routing to the lysosome enables ILV components to be degraded, which in turn allows turnover of membrane receptors [27,28]. The whole process requires an intricate protein machinery called the Endosomal Sorting Complex Required for Transport (ESCRT) which reshapes the endosomal membrane topology [29,30]. During MVB formation, ESCRT allows intraluminal budding of vesicles with the sorting of embedded cargo proteins. The ESCRT is composed of approximately twenty proteins including the ALG-2 Interacting protein X (ALIX), the Tumor susceptibility gene 101 (TSG101) and the Charged multivesicular body protein 4a (CHMP4) [31]. The molecular actors of ESCRT are organized into four cooperative complexes called ESCRT-0, -I, -II, -III, which are conserved from yeast to mammals [31]. The ubiquitination of proteins present on the endosomal membrane drives the inward budding. ILVs containing ubiquitinated proteins are those destined to be degraded by the lysosome. Deubiquitination of these proteins by the de-ubiquitylating enzymes (DUBs) will rescue the ILVs from degradation. The specific interaction with Rab proteins and soluble NSF attachment protein receptor (SNARE) complexes is crucial. It directs the subsequent addressing of multivesicular bodies (MVBs) either to lysosomes for degradation or towards fusion with the plasma membrane, resulting in the extracellular release of exosomes [32,33,34,35,36]. Numerous studies have also shown that phospho-inositides, ceramides and, more generally, the lipid composition of the membrane are key factors for the vesicles biogenesis in MVBs [37,38]. It's also worth mentioning that ESCRT-independent mechanisms have been identified for an alternative formation of ILVs, and that vesicle release could also occur from lysosomes [39,40,41]. In addition, a process involving tetraspanin enrichment appears to be important for membrane remodeling and ILVs formation [42,43]. Tetraspanins are a family of transmembrane proteins including CD9, CD63 and CD81, which are abundantly found in MVBs and ILVs membranes [44,45]. They are known to help select cargo to be packed into ILVs, and to direct exosome capture by target receptor cells [43,46]. The tetraspanins CD9, CD63 and CD81 are therefore frequently used as markers for identifying EVs from exosomal origin [47] but we must point out that the attested presence of CD9 and CD81 also on the plasma membrane makes CD63 the most relevant tetraspanin marker for exosomes [48].
2.2. Exosomes composition
During their formation, ILVs are loaded with both membrane and soluble cytosolic proteins. While some proteins are shared by all exosomes and associated with the ILV biosynthetic pathway, others are unique and indicative of the cell type from which the exosomes originate [1,50,51]. In addition, the content of exosomes reflects the physiological state and composition of the producing cell, influencing specific biological activities directed towards recipient cells [52,53]. Of note, the hypothesis that intercellular communication can take place via EVs is based on the ability of cells to control EVs’ production, both quantitatively and qualitatively. This suggests that cells direct the loading and then the content of ILVs. They must also control the vesicular traffic that will enable the secretion of exosomes carrying specific, selectively addressed messages [42]. A schematic representation of an exosome with some of its typical, but not exhaustive, compounds and cargoes is shown in Figure 1B. Several of the identified proteins represent a molecular signature of exosomes and abundance of Syntenin-1, TSG101 and ALIX suggests that these proteins could serve as robust exosome markers [54]. Regarding common denominators, the tetraspanins mentioned above in the biogenesis pathways of ILVs (i.e., CD9, CD63 and CD81), with a preponderance of CD63 are widely considered as easily accessible exosomal markers because of their high representation in the exosome membrane [55]. They play an important role in cell contact, membrane fusion or uptake events. Within the membrane, various adhesion molecules like integrins are also among the proteins identified [49]. Depending on their relative abundance on the exosome's surface, these membrane adhesion factors may reflect pathological situations. This has been shown, for example, in the case of cancer and tumor progression. In this way, integrin display patterns are linked to the organotropism of tumor exosomes and their uptake contributes to the establishment of pre-metastatic niches [56,57]. Major histocompatibility complex (MHC) type I molecules are present in variable levels in the membrane of exosomes. This may depend on cell type and local microenvironment. MHC class II and co-stimulatory molecules, such as ICAM-1, are more restricted to exosomes secreted by professional antigen-presenting cells (APC). Both are required for mature exosomes to prime naive T cells [58]. The presence of functional MHC-antigenic peptide complexes, specifically, indicates a potential role for exosomes in adaptive immune responses [14,59]. This point will be specifically discussed in section 5 of this review. About exosome protein content, the previously mentioned ALIX and TSG101, are systematically present. Various chaperones appear to be able to be enclosed in exosomes and follow this alternative secretion pathway to take part in extracellular networks. These include members of the heat-shock proteins (HSPs) and J-domain proteins [60]. A correlation between HSP70 levels in exosomes and cancer progression had suggested that analysis of exosome profiles and their contents are a way of estimating disease severity, and that exosomes are potentially also actors in the disease progression [61]. Of note, the presence of proteins traditionally considered abundant in exosomes, such as cytoskeletal proteins, GAPDH, and even HSPs, has become a topic of controversy. These uncertainties about exosome content arise from advancements in techniques for separating small vesicles and the availability of increasingly powerful analytical methods for studying their proteomes, such as mass spectrometry. Finally, depending on the context and the cell type, certain proteins will be specifically embedded but the exact mechanisms of exosome loading remain elusive. Thus, cytokines and various mediators seem to be able to use the exosomal vectorisation pathway. This has been documented for interleukin-1β (IL-1β), which can be loaded and transported via exosomes in inflammatory and pyroptotic contexts [62,63]. IL-6, IL-10 and TNF-alpha could also be transported in this way, giving these mediators a longer lifespan in body fluids and making again exosomes suitable markers for prognostic and diagnostic [64]. The diversity of exosomal contents also suggests a broad and still poorly understood trafficking between cellular compartments. For example, MVBs and autophagosomes have been shown to interact, allowing cargo exchange. This proximity also favors the release of exosomes [65].
Besides proteins, exosomes contain lipids which are involved in maintaining the exosomal membrane shape and in the biogenesis of exosomes [66]. Sphingomyelin, phosphatidylserine, glycosphingolipids and regulated concentrations of cholesterol are the hallmarks of ILVs and exosome membranes. Thus, MVBs rich in cholesterol would be more likely to undergo lysosomal digestion, while lower cholesterol levels would be associated with the exocytosis pathway [67]. Exosomes also carry nucleic acids like short DNA molecules, small RNAs and especially miRNAs [68,69]. The deep sequencing RNA technique confirmed that many types of non-coding RNAs were loaded into ILVs, and that their composition depended on cell type and context. Several heterogeneous nuclear ribonucleoproteins (HNRNPs) and RNA-binding proteins have been shown to promote RNA sorting and loading in ILVs. They could also contribute to RNA function in target cells. These mechanisms were extensively reviewed by Corrado et al. [70].
Remarkably, and in the light of numerous data in the literature, it is clear that exosome composition undergoes significant changes under different physiological and pathological conditions. This variability, particularly in terms of miRNAs, is regarded as both prognostic and diagnostic biomarkers. Consequently, the quantity and content of exosomes derived from patients' plasma emerge as a valuable tool for monitoring the progression of diverse pathologies [71], from autoimmune diseases such as rheumatoid arthritis [72,73], to cancers [74]. These aspects will be elaborated further below.
2.3. Exosomes biological activities according to their mode of interaction with recipient cell
The enrichment of exosomes with various components underscores their intricate nature and the potential for functional diversity. There is extensive literature on the messenger functions of exosomes, particularly during pathophysiological situations like immune responses, tumor development or host-pathogen interactions [75]. Recipient cells, whether near or distant from the releasing cell, can interact due to the ability of exosomes to circulate in various body fluids and to be elements naturally bioavailable. Vesicle-mediated transport of biological mediators intuitively suggests that they will be protected from degradation by their membrane envelope. However, this feature is not fully verified in vivo. While EVs can remain stable in serum for days, their half-life in circulation is notably brief. Studies in mice indicate that exogenous EVs exhibit a very short half-life (2–10 minutes) in the blood circulation [76]. This brief circulation time could, however, be attributed to a rapid uptake of circulating EVs by blood cells like monocytes and by endothelial cells. Indeed, several studies have shown that small EVs have no difficulty in crossing endothelial and blood-brain barriers, sometimes by transcytosis, and thus diffuse widely into tissues [77,78,79].
Exosomes appear to be able to interact with their target cells in various ways [35,50]. In particular, they can act directly by contact with cell surfaces. This is the case for exosomes that carry MHC-peptide complexes and can induce the activation of TCR receptors on T lymphocytes, as already mentioned [58]. Another example is given by exosomes that have been loaded with FasL ligands in melanoma cells, which are able to induce apoptosis in recipient lymphocytes [80].
Furthermore, exosomes can deliver their contents directly to target cells. Their membrane-based nature allows them to fuse with the plasma membrane [81]. Recipient cells can also internalize exosomes through an uptake process that can be visualized through several imaging techniques [82]. The mechanisms by which recipient cells take up exosomes have been a subject of extensive research for over 20 years [83,84]. Although macropinocytosis may be a potential entry mechanism, the capture of exosomes appears to be quite specific. However, the recognition process by which exosomes are identified by their target cells remains poorly understood, and specialized receptors for this purpose have not been well identified yet [85]. It has been proposed that T-cell immunoglobulin- and mucin-domain-containing molecule 4 (TIM4) could bind to exosomes through the phosphatidylserine exposed on their surface [86,87]. Other studies have suggested that exosome endocytosis may result from interaction with heparan-sulfate proteoglycans [88], or syndecan 4 [89]. Upon internalization, exosomes are believed to have several potential routes and outcomes. They might follow a recycling pathway, acquiring new cargo along the way before being returned to the external environment. Alternatively, they may take a pathway leading to lysosomal digestion. Finally, exosomes can deliver their cargo to the recipient cell by fusing, dependant on pH, with endosomal membranes [90]. The factors and mechanisms that dictate the fate of exosomes after internalization remain to be fully elucidated.
The biological effect of exosomes on cell phenotype and behavior is particularly well studied in pathological contexts, in order to identify their positive or negative role in pathological progression, with a view to identifying therapeutic players or targets. Their role in cancerous processes has notably received particular attention, enabling the scientific community to unravel the mechanisms of exosome capture and action. For example, and as previously mentioned, EVs derived from malignant cells, whose content was modified, compared to exosomes produced from healthy cells, have been shown to play a key role in liver pre-metastatic niche initiation during pancreatic cancer [56,57]. Moreover, exosome transfer from highly-metastatic to poorly-metastatic tumor models has been shown to increase the metastatic behavior of the latter model [91]. It remains to be elucidated how exosomes shape the microenvironment of those pre-metastatic niches. Selective uptake of exosomes by Kupffer cells in the liver was proposed to induce the activation of fibrotic pathways and a pro-inflammatory environment that supports metastasis formation [92]. Mechanisms such as vascular permeability impairment or local immunosuppression may be involved. Indeed, exosomal hepatocyte growth factor (HGF) receptor was shown to skew hematopoietic stem cells towards pro-vasculogenic phenotypes and C-C motif chemokine ligand 2 (CCL2) facilitates uptake of tumor exosomes by immune cells, which ultimately leads to enrichment of myeloid suppressive cells at future metastatic sites [93,94].
Although simple contact between exosomes and their target can result in phenotypic changes in recipient cells, it seems that exosome capture is most often required to observe the biological effect of this signaling pathway. The transfer of exosome contents to cells has been demonstrated on numerous occasions, in particular for proteins. The mechanisms were extensively reviewed by Mathieu et al. [35]. Small mRNAs and miRNAs that are carried by exosomes have also been shown to be transferred into recipient cells. Once inside, they retain full functionality and have the capacity to modify the behavior of the recipient cells through gene expression modulation [95]. Moreover, the transfer of oncomir-like microRNAs through exosomes, produced by cancer cells, has been shown to induce reprogramming of the recipient cells and promote prometastatic behavior [96]. Certain miRNAs loaded into EVs derived from tumor cells are suspected to be involved in immune escape by promoting anergy (activation-induced non-responsiveness) of CD8+ T lymphocytes [97]. Indeed, a decrease in cytokine production and granzyme B secretion was observed after internalization of tumor exosomes by lymphocytes. In addition, they are suspected to contribute to resistance against anti-cancer drugs. This mechanism has been demonstrated in diverse cancer types, encompassing hematological malignancies, as well as colorectal, breast, prostate, liver, and lung cancers [98]. In their study, Patel et al. demonstrated that, when exposed to gemcitabine, the primary chemotherapeutic agent for pancreatic cancer, exosomes produced by pancreatic cancer cell lines led to an increased IC50 (half maximal inhibitory concentration) of native cancer cells after treatment with these exosomes [99]. Thus, in the context of cancer, exosomes would play determinant roles in the migration and proliferation capacities of tumor cells, in angiogenesis and in the remodeling of the tumor's immune microenvironment [100,101]. In this way, given the duality of EVs in terms of their pro-tumor effects, but also their strong ability to mediate communication in a microenvironment, the strategies for dealing with EVs in cancer are based, on the one hand, on inhibiting EV trafficking as a new method to improve new therapeutic combinations [102] and, on the other hand, on engineering EVs to use them as vectors for therapies [103].
Among other processes linked to the induction of phenotypic changes, exosomes were shown to contribute to the polarization of macrophages [104]. Exosomes have also been described as players in epithelial-mesenchymal transition (EMT) in a variety of pathological conditions, including fibrosis and cancer [105,106]. However, miRNA contents of exosomes, depending on the emitting cells, would mediate potentially antagonistic effects on EMT, revealing both promotion and inhibition role [107,108].
In conclusion, the vast amount of data on the biological effects of exosomes produced in response to pathological contexts reveals the multifaceted nature of exosome-mediated cellular communication. The evolving understanding of the role of exosomes makes these extracellular vesicles potential key players in disease progression. Exploiting this knowledge could find applications in terms of innovative diagnostic approaches and targeted therapeutic interventions, paving the way for further exploration of personalized medicine and precision therapeutics.
3. Exosomes as biomarkers of pathologies in human medicine
In the past few years, the use of exosomes as biomarkers and diagnostic tools has surged. Exosomes can be easily isolated from almost all body fluids, such as blood, urine [109], saliva and even amniotic fluid. This versatility positions exosomes as a potential tool for fetal sex determination [110]. The amount of circulating exosomes can serve as an indicator of pathological conditions. Exosome production has been reported to increase in the aftermath of a variety of cancers, in HIV-positive patients, as well as in acute kidney and cardiovascular diseases. Notably, this increase occurs simultaneously with the onset of the disease, reinforcing the potential of exosomes as early biomarkers [111].
In pathophysiological conditions associated with neurological diseases, viral infections, or cancer, the content of exosomes may undergo significant alterations. Many cancer studies now focus on circulating EVs in an attempt to establish correlations between the quantity or content of EVs and treatment responses, the presence of metastasis, or disease progression [112,113,114]. Analysis of EVs has emerged as a promising tool for cancer surveillance, surpassing traditional biopsy methods. EVs can be harvested in a minimally invasive manner from multiple biofluids, employing a technique known as liquid biopsy [111]. Another of their significant advantages lies in the easily detectable and analyzable nucleic acids they encase. For example, double-stranded DNAs in exosomes can be used as predictive clinical biomarkers for cancer diagnosis and prognosis, as they may harbor mutations such as KRAS and TP53 in pancreatic cancer [115]. The other family of candidates on the scene of exosomal biomarkers is miRNAs. Indeed, circulating exosomal miR-17–5p and miR-92a-3p have been associated with the pathological stage and grade of colon cancers [116], whereas the miR-21 found in cerebrospinal fluid can serve as a biomarker for glioblastoma development [117]. Furthermore, circular RNAs (circRNAs), which proceed from mRNA splicing, may also serve this purpose. Their circular structure provides effective protection from exonuclease degradation, thus conferring greater stability compared to miRNAs [118]. Their relevance in cancer diagnosis has been investigated in pancreatic cancers [119].
Exosomes have been recently proposed as interesting tools for therapeutic drug monitoring. Nephrotoxicity is a notorious side effect associated with numerous drugs, including antimicrobials, anti-cancer therapies and non-steroidal anti-inflammatory drugs. Their deleterious effects on the kidneys are currently monitored through the dosage of plasmatic biomarkers, which, unfortunately, lack sensitivity and specificity. Urinary exosomes could offer a good alternative. Differences in their surface proteins and miRNA content may correlate with specific renal diseases, thus facilitating diagnosis and adequate care [120].
Although exosomes present irrefutable advantages, their use in the clinical setting is yet to be popularized. Indeed, several subtypes of EVs of unequal clinical relevance may be found in different quantities in body fluids. If clinically-relevant EVs are present in lower abundance, they might go unnoticed by detection methods, leading to potential false-negative results [121]. Moreover, isolation methods and cargo analysis are often labor-intensive, and current techniques not only lack standardization but also require costly laboratory equipment that need precise stewardship; a lack of practicability that is currently incompatible with widespread use. The development of microfluidic technologies and ongoing miniaturization efforts may address these challenges [122]. It is noteworthy that exosome-based diagnostic tools have already been introduced in clinical settings, demonstrating promising results in guiding clinicians to establish proper therapeutic interventions, as illustrated in this study on high-grade prostate cancer [123].
4. Exosomes and biology of infection
The particular biological context generated during infections has prompted investigations into whether exosomes produced and circulating exhibit a modified profile, from a quantitative and qualitative point of view. Numerous studies have shown that infections impact the biogenesis of exosomes in infected cells, leading to an increase in their production and modification of their content. This has been demonstrated in the case of infection by very different families of both RNA and DNA viruses during in vitro studies or analysis of biological samples, e.g. patient plasma [124,125,126,127,128,129]. In particular, an increase in exosome quantity has been revealed in the case of infection with Dengue (DENV) and Zika (ZIKV) flaviviruses. This was observed in vitro from the first 24 h of infection, and was accompanied by a change in EV size [130]. As previously mentioned, exosomes are small EVs of endosomal origin that allow unconventional secretion of factors and play crucial roles in intercellular communication. Their ability to traffic from and to plasma membranes, across the endosomal pathway, has led to suspicions of possible involvement of exosomes in dissemination and pathogenesis of intracellular pathogens (viruses, parasites, and intracellular bacteria). However, their active and increasingly documented role in mounting the immune response provides beneficial effects during the course of an infection. In this paragraph, we will attempt to review the literature on this double-edged capacity of exosomes during virus infection. The main aspects of this duality are summarized in a schematic representation (Figure 2).
4.1. Exosomes are small, specialized extravesicular vesicles (EVs)
Many viruses exploit intracellular trafficking mechanisms to complete their multiplication cycle. They initially interact with the cell surface and commonly follow pathways leading to endosomes. The assembly and budding of enveloped viruses share many similarities with the biogenesis of small EVs, as both processes rely on cellular membranes. Consequently, it is unsurprising that viruses interfere with the biogenesis of MVBs and the production of EVs. In particular, the ESCRT system, required for the formation of ILVs, has been found as an essential tool for the production of enveloped viruses. Numerous studies have shown that many viruses hijack the ESCRT system to promote their own budding and, thus, the efficiency of their production and dissemination [131,132]. Flaviviruses are particularly concerned, and numerous studies have shown the dependence of their viral cycle on ESCRT-dependent membrane remodeling [133,134,135].
Under these conditions, it can be hypothesized that MVB formation and the release of exosomes is negatively impacted, and this defect may favor the virus at the expense of homeostasis control. The hijacking of exosome biogenesis might additionally confer the ability to viruses to target crucial components within exosomes, including their own viral nucleic acids, as well as viral proteins or even complete virions. This was notably shown with the hepatitis B virus (HBV) [136] and since the role of EVs in infections has been studied, examples of viruses using host exosomes as transport systems for their own factors have become widespread and are regularly and recently the subject of comprehensive reviews [137,138]. This ability has led to a Trojan Horse hypothesis that exosomes actively contribute to viral pathogenesis and sometimes facilitate viral persistence [139]. Hence, Hepatitis C Virus (HCV), like other Flaviviridae family members (e.g. flaviviruses like DENV, ZIKV or West Nile virus) enter through endocytosis and release their genome by a “back fusion” phenomenon which is comparable to the one observed for exosomes [140,141]. But a particularity of HCV is that its genome can be included in ILVs, following endocytosis of the virus and its transit through endosomes. This viral genetic material is then released into exosomes [141]. Strikingly, these exosomes were shown to act as pseudo viral particles that can infect efficiently recipient cells [142]. Similarly, exosomes derived from epithelial cells infected with Chikungunya virus (CHIKV), a member of the alphavirus genus, have been shown to display viral RNA and proteins. These exosomes were able to infect naive epithelial cells [143]. This unexpected mode of transmission raises questions about a general ability of viruses to use exosomes as alternative routes of entry into cells [144]. This mimicry with a physiological mode of EV transport and capture could indeed diversify the cellular tropism of viruses and contribute to their escape from the immune system. During Human Immunodeficiency Virus (HIV) infection, HIV-associated RNAs (including TAR, Trans-activation response element) have been detected in infected T cell-derived exosomes. These RNAs were able to down-regulate apoptotic signals in recipient cells thus presenting an improved support of HIV replication [145]. This detrimental effect can also be expressed as an accentuation of pro-tumor profiles, especially in Head and Neck Squamous Cell Carcinoma (HNSCC) cancers. Indeed, it has been shown that exosomes from infected T cells carrying TAR (trans-activation response element) RNA increase the proliferation of cancer cells and induce proto-oncogene expression in recipient cells [146]. Among viral elements embedded in exosomes are also miRNAs which, when delivered to the target cells, will exert numerous adverse effects. For example, miRNAs encoded by Epstein-Barr virus (EBV) produced by infected cells are able to inhibit genes involved in antiviral activities and inflammation in neighboring non-infected cells through exosomal transfer [147]. During Japanese Encephalitis virus (JEV) infection, activated microglia release let-7a/b miRNAs through extracellular vesicles [148]. These miRNAs activate caspases in uninfected neuronal cells, potentially contributing to bystander neuronal death. Additionally, in COVID-19 patients, SARS-CoV-2 (Severe Acute Respiratory Syndrome Coronavirus 2) RNA was detected in plasma-derived exosomes, along with a distinct proteomic signature. Based on the proteomic analysis, these circulating exosomes could potentially contribute to various processes like coagulation, inflammation, and immuno-modulation during SARS-CoV-2 infection [149].
Viral proteins also use exosomes as effective delivery systems. For example, during HIV infection, the viral membrane protein Gag and the virulence factor Nef have been identified in exosomes derived from infected cells [150,151]. Of note, exosomal Nef was shown to restore infectivity of Nef(-) HIV virions in recipient cells. Nef in exosomes also triggers apoptosis in CD4+ T cells and endothelial cells, contributing to the pathophysiology of the infection [152]. Of further note, Nef limits the effect of RNA interference in recipient cells due to a modification of the miRNA content of exosomes [153]. Other viruses are known to induce the incorporation of their proteins into exosomes, which facilitates their replication or increases the virus susceptibility of recipient cells. Such mechanism has notably been reported for glycoprotein B of cytomegalovirus (CMV) [154] as well as E2 protein of HCV [142]. Mishra et al. reviewed the data concerning the role of exosomes during Dengue fever. Thus, exosomes produced in response to DENV infection are also thought to facilitate cell-to-cell viral transmission by carrying various viral factors and to promote immune evasion. The various cargoes carried by exosomes could modulate immunity in T lymphocytes and platelets, and their action on endothelial cells may contribute to pathophysiology, notably vascular permeabilization during hemorrhagic forms of the infectious disease [155].
Among the effects produced by the enclosure of viral factors, in line with the Trojan horse hypothesis, it appears that hijacking of exosomes by viruses is an excellent means of promoting their incognito trafficking. Firstly, their capacity for prolonged circulation and access to extracellular spaces implies their superiority in facilitating cell-to-cell communication and efficient diffusion of their cargo. Secondly, the relatively small size and consistent shape of exosomes enable them to effectively evade immune defenses. As such, exosomes from APCs were shown to express membrane regulators of complement (CD46, CD55 and CD59) on their surface, preventing the activation of opsonin and coagulation factors [156]. Exosomes are thus protected from destruction by the complement cascade, warranting their stability and large-scale distribution in biofluids. In general, EVs have demonstrated their ability to regulate complement activity, albeit with variable effects, but contributing to the pro- and anti-inflammatory immune balance [157,158].
In addition, some infectious agents have the ability to influence exosome biogenesis to promote or hinder the presence of host-specific proteins. HIV has been shown to promote the availability of its major entry receptors, CCR5 and CXCR4, on exosomes secreted by infected cells. The transfer of these proteins into receptor cells is associated with increased viral susceptibility of these cells [159]. In another study, it was shown that galectin-9-containing exosomes released from EBV-infected cells promoted apoptosis of CD4+ T cells with deleterious Th-1 suppressive effects [160,161]. Finally, antigen presentation is essential to mount a proper immune response during infection. Surprisingly, Herpes Simplex Virus-1 is able to counteract this mechanism, limiting antigen presentation by addressing peptide/HLA-DR complexes to MVBs and exosomes [162].
The ability of viruses to exploit exosomes is often auspicious from a therapeutic standpoint. Indeed, cancer cells infected by oncolytic adenoviruses, which induce systemic anti-tumor immunity by specific targeting of malignant cells, secrete exosomes loaded with viral genome and those exosomes can infect other cancer cells [163,164]. Although it was previously unknown that adenoviruses could hijack exosomes, those findings may explain how oncolytic viruses achieve targeting of distant tumor cells and induce systemic responses while evading the antiviral immune response.
The study of the roles played by exosomes in infection biology shows many examples of proviral effects of exosome hijacking. However, many studies show a much more ambiguous role for exosomes. For instance, they also appear to be part of antiviral cellular responses or at least involved in the host response to pathogen multiplication.
4.2. Exosomes contribute to infection resolution
Exosomes are known for their aptitude to induce immune responses and participate in immunomodulatory mechanisms. These functions have been extensively described in tumor processes [165]. The exosomes released during infection could play a similar role in modulating immune responses [166].
First, as stated above, exosomes from infected cells were shown to carry viral nucleic acids. This may lead to the stimulation of pattern recognition receptors in recipient cells (PRRs), such as cyclic GMP-AMP synthase stimulator of interferon genes (cGAS-STING) or RIG-I like receptors (RLRs), as demonstrated in the case of cancer-derived exosomes [167]. The downstream signaling pathways of these PRRs are part of the innate immune system, triggering the expression of inflammatory genes and type I interferon response, which are deleterious for the pathogen replication and dissemination [168]. Such mechanisms were notably reported during HCV infection, illustrating the dual role played by exosomes that can have proviral and antiviral activity [169]. Secondly, exosomes can participate in antigen presentation, influencing the adaptive immune response [2]. This ability will be developed more specifically below (section 5), as it contributes to the suitability of exosomes for vaccine development.
Supporting their ability to perform antiviral signaling, exosomes have been identified as carriers of various chemokines and cytokines important for infection resolution, like IL2, IL4, IL17, IL21, IL22, IL33, IFNα, IFNγ, TGFβ, TNFα, and other antiviral mediators [170,171]. An exosome-mediated antiviral activity through IFNα and other mediators delivery was reported in the case of both HBV [172] and HIV infections [173]. Exosomes produced during DENV infection carry the interferon-induced transmembrane protein 3 (IFITM3), which promotes antiviral activities from cell to cell [174]. The cells that release such exosomes in response to infection are of several types and this activity follows stimulation of innate immunity pathways. Thus, interferon ɑ-stimulated macrophages address to the neighboring cells exosomes that contain the antiviral mediator apolipoprotein B mRNA editing enzyme catalytic polypeptide-like 3G (APOBEC3G). Uptake of these exosomes triggers antiviral activity in the recipient cells [173]. Cytokine encapsulation in exosomes can be achieved by stimulating keratinocytes with Poly(I:C) [175]. In addition, exosomes produced by TLR3-activated human brain microvascular endothelial cells were shown to carry mRNA and proteins of interferon-stimulated genes (ISG), ISG15, ISG56 and Mx2, which could be transferred to recipient macrophages [176]. As final examples, exosomes derived from respiratory syncytial virus (RSV)-infected cells and carrying several viral proteins were shown to trigger the production of various chemokines like monocyte chemoattractant protein-1 (MCP-1), Interferon gamma-induced protein (IP-10), and (regulated upon activation normal T cell expressed and secreted) RANTES by monocytes [124]. And EVs released by DENV-infected macrophages that were characterized by the encapsulation of viral proteins and various miRs elicited increased production of ICAM, TNF-α, IP-10, IL-10, RANTES and MCP-1 triggering an antiviral defense program in endothelial cells [177].
As mentioned previously, many viruses can exploit exosome biogenesis pathways to their advantage, transporting viral material and enhancing their dissemination. Exosome production by infected cells therefore tends to be pro-viral. Unsurprisingly, type I interferon was found to be able to limit exosome secretion both in vitro and in vivo, thereby attenuating their pro-viral effects. The mechanism is linked to the induction of ISG15, which triggers ISGylation of proteins crucial to exosome biogenesis, notably TSG101. This process significantly reduced exosome secretion by promoting lysosomal degradation of MVBs [178,179].
Ultimately, increasing studies explore how miRNAs transported in exosomes can modulate gene expression in recipient cells to improve cell ability to limit infection. Exosomes derived from human trophoblasts carry miRNAs from chromosome 19 clusters, such as miR-512-3p, miR-516-5p, and miR-517-3p. Upon exposure to these miRNAs, recipient cells undergo autophagy, conferring resistance to various viruses including hepatitis C virus (HCV) [180]. Similarly, miR-483-3p, which is found at high levels in the serum of mice infected with the H5N1 influenza virus, has been shown to mediate the expression of proinflammatory cytokines in the vascular endothelial cells [181]. This could perhaps establish an antiviral state that promotes the resolution of the infection.
In conclusion, viral infections lead to a quantitative and qualitative modification of the released exosomes. Exosome signaling must be considered as a fully-fledged response of the host, playing a bidirectional regulatory effect on host-pathogen interaction. Hijacking of the system enables pathogens to spread viral material and escape immune surveillance, while exosomes with modified contents negatively regulate pathogens by transmitting antiviral mediators and immunomodulators from cell to cell, including monocyte-macrophages, NK cells, T and B lymphocytes.
5. Exosomes and antigen presentation, perspectives in vaccinology
5.1. Exosomes and antigen presentation
A particular feature of exosomes that is receiving increasing attention is their immunomodulatory functions and their involvement in the presentation of antigens to the adaptive immune system (Figure 3). Exosomes derived from mature dendritic cells (DCs) were found to be enriched in MHC class I and II molecules, as well as ICAM-1 [182]. EVs secreted by mature DCs are known to carry MHC-peptide complexes on their surface, thus enabling a greater potency in stimulating T cells, compared to exosomes isolated from immature DCs [183]. The presentation of pathogen-derived peptides by exosomes has been demonstrated in the case of bacterial and viral antigens. For example, it has been shown that LPS-activated monocyte-derived dendritic cells (mo-DCs) interaction with activated bystander T-cells induced changes in the morphological characteristics of activated mo-DCs. This led to the release of exosomes carrying MHC class II-peptide, ICAM-1, and a high amount of miR155. miR155 is a well-established central regulator of T-cell responses, possessing the capacity to activate antigen-specific CD8+ T cells [184]. Another study demonstrated that upon encountering Escherichia coli, DCs generated a substantial quantity of EVs, both carrying antigens derived from the previously-phagocytosed bacteria, along with MHC class II molecules. This investigation emphasized the potential effectiveness of exosomes in presenting antigens and triggering an adaptive immune response, when specifically packed with pathogen-derived peptides, loaded onto MHC molecules [185].
Interestingly, exosomes that carry MHC-peptide complexes or antigens may act as indirect antigen presentation platforms, following internalization by other cells [2]. Indeed, migrating and resident DCs were demonstrated to exchange antigens through exosomes, thereby allowing DCs that were not directly exposed to the antigen to participate in T cell priming [186]. Following EV capture, antigens can either be driven into the endosomal pathway, subsequently degraded, and loaded onto MHC class II molecules; or they can be released into the cytoplasm upon fusion of the plasmic and exosomal membranes, subsequently degraded, and loaded onto MHC class I molecules. Greater capacity for antigen presentation and efficient T cell activation can thus be achieved by transferring antigens to DCs via exosome capture. In addition, it is sometimes useful to promote cell-mediated immunity through antigen presentation to MHC class I [187].
This ability to participate in the establishment of a specific immune response via EVs may also happen during viral infections. As above-mentioned, exosomes that are produced during a viral infection are frequently loaded with viral factors that can be exhibited outwards, thus enhancing their immunogenicity. Along DCs, various cell types may produce exosomes capable of triggering an immune response. For example, infection with the HTLV-1 retrovirus is accompanied by the detection of exosomes carrying viral proteins in the cerebrospinal fluid, which remains barren of viral particles. PBMCs and CD4+CD25+ T cells have been shown to be the main source of production of such exosomes, which contain the HTLV-1 Tax protein. Remarkably, these exosomes were shown to sensitize an HTLV-1-specific immune response, and to induce infiltration of HTLV-1 Tax-specific cytotoxic T lymphocytes (CTLs) into the central nervous system of patients [188]. A study has shown that exosomes isolated from mild COVID-19 patients carry SARS-CoV-2 spike peptides and other immunomodulatory molecules, such as MHC class II molecules, CD86 and others. These exosomes exhibit the ability to effectively regulate antigen-specific CD4+ T-cell responses and induce IL-2 secretion in vitro. Proteomic analysis was conducted on exosomes from these patients, revealing a correlation between the protein repertoire and an enhanced immune response signature, supporting what has been found in vitro [189].
Concerning flaviviruses, exosomes originating from ZIKV- and TBEV (tick-borne encephalitis virus)- infected cells were found to display the viral envelope protein on their surface, suggesting their ability to induce an immune response [reviewed in: [190]]. Furthermore, our research has revealed that the non-structural protein-1 (NS1) of both DENV and ZIKV was found associated onto the surface of EVs produced during infection [130]. However, the ability of these vesicles, which circulate abundantly in the body, to stimulate an immune response against the NS1 of these two flaviviruses has not yet been investigated.
The idea that exosomes could be contributors to viral antigen presentation led to the exploration of the possibility of manipulating exosomal addressing. A study of exosome engineering for this purpose showed some success with the expression in exosomes of the envelope protein of bovine leukemia virus. This work paved the way for the use of exosomes in vaccine strategies [191].
5.2. Exosomes in vaccinology
Exosomes exhibit intriguing structural features and properties that make them promising biocompatible nanocarriers [192]. They also have the advantage of being modifiable during biogenesis. Deliberate incorporation of specific antigens during vesicle formation makes them useful for directing antigen presentation to the immune system. As a result, exosomes can serve as vectors for therapeutic products. Of note is the clinical application of exosomes derived from allogeneic bone marrow mesenchymal stem cells, such as EXOFLOTM, which has proved effective in attenuating the cytokine storm and associated tissue damage in patients with COVID-19 disease [193]. They offer a delivery system for RNAs, which are protected from degradation by the enclosing lipid membrane. This encapsulation system makes it possible to enhance the stability of the drug, thus considerably improving its bioavailability and delivery to target cells [192]. The strategies, approaches and technologies for engineering exosomes are plethoric, and the bibliography on the subject is considerable including the following recent reviews [194,195,196]. Engineering exosomes to make them suitable vaccination platforms requires the assembly of targets of interest (DNA, RNA or proteins), at the surface or within EVs. Several methods have been explored to date, each more or less suitable depending on the nature of the target to be integrated. For example, electroporation can be used to load exosomes with RNA or other hydrophobic therapeutic cargoes. Alternatively, RNAs can also be fused to RNA-binding proteins, such as HuR, TAT and L7Ae, allowing them to be loaded into exosomes [197]. This approach enhances the tissue biodistribution potential of effective hydrophobic molecules [192]. However, there are still many challenges to overcome in order to exploit their therapeutic potential.
However, vaccine development based on exosomes is a promising application [198]. The potential advantages are as follows: (1) a more stable conformation of antigens in different conditions; (2) enhanced molecular distribution as exosomes can circulate in fluids, reaching distal organs; (3) more efficient association with antigen-presenting cells due to the expression of adhesion molecules on exosomes surface; (4) and exosomes, acting as inherent carriers of immunomodulatory mediators, already known to transport antigens between cells and play a role in cross-priming. The latter process involves the activation of naïve T cells by antigen-presenting cells that have acquired their specific antigens, not through direct contact, as commonly occurs, but from another cell, as mentioned previously and in Figure 3 [199]. Pathogens are notorious for evading the immune system, a challenge that can be addressed through the use of cocktails and multi-epitope vaccines (MEVs) [200]. In this context, exosomes could be exploited to present multiple antigens, thus serving as vaccine-design exosomes [197].
Exosomes have thus emerged as potential candidates for the development of vaccines or adjuvants against viruses. For instance, exosomes from B cells infected by Epstein-Barr virus (EBV) have demonstrated the ability to protect uninfected B-cells from EBV infection [201]. More recently, engineered exosomes of full-length HCV-NS3 protein appeared immunogenic when injected in mice, proven by the detection of NS3-specific memory CD8+ T lymphocyte pool [202]. Anticoli et al. have also demonstrated the efficacy of DNA-vectors expressing diverse fusion products, including Ebola Virus VP24, VP40 and NP, Influenza virus NP, Crimean-Congo Hemorrhagic Fever NP, and West Nile Virus NS3. These vectors have been shown to elicit antigen-specific CD8+T cell response associating with a cytotoxic activity against the peptide loaded into exosomes or antigen-expressing syngeneic cells, in mouse model [203]. Exosomes could be also considered as valuable adjuvants in vaccine strategies. This was shown for recombinant hepatitis B antigen (HBsAg) [204]. The results of this study suggested that the isolated exosomes carrying HBsAg provoked a pro-inflammatory profile in spleen cells of healthy mice by stimulating the production of cytokines such as TNF-α, and IL-1β. The vaccination of mice with exosomes combined with a solution of hepatitis B recombinant antigen induced a humoral immune response. However, the increase in IFN-γ secretion has demonstrated that exosomes triggered an immunomodulator effect on the cellular immune response.
Concerning HIV-1, Khatua et al. have found that exosomes secreted by CD4+ H9 T cells and mature monocyte-derived DC encapsidated A3G (APOBEC3G) and A3F, which are part of a cellular defense system against retroviruses. They indeed were able to inhibit L1 (non-LTR retrotransposons LINE-1) and Alu retrotransposition [173]. Therefore, DC derived exosomes are a major source of potential vaccines due to their high stability during storage as well as their rich composition in immunoregulatory molecules [205]. Moreover, it was demonstrated that HIV-1 Gag-specific exosome-targeted T cell-based vaccine stimulated effector CTL responses giving a long-term immunity against Gag/HLA-A2 expressing B16 melanoma in transgenic HLA-A2 mice [206].
More recently, in the face of the global COVID-19 crisis, there has been a flurry of research into innovative vaccination strategies, notably by exploiting the exosome approach [207,208]. Two of these strategies are summarized in Figure 4. One strategy, following the general implementation of mRNA vaccines, was to use the encapsulation and nucleic acid carrying properties of exosomes to turn them into mRNA vectorization tools. This use of exosomes to vectorize mRNAs for vaccine purposes has been successfully tested and shown to promote an immune response [209]. Thus, for example, Popowski et al. developed a system based on lung-derived exosomes carrying SARS-CoV-2 Spike mRNA. These exosomes have shown extensive distribution in the bronchioles and parenchyma of mice. In addition, they elicited significant immunoglobulin G (IgG) and secretory IgA (SIgA) production, demonstrating the relevance of such an approach for obtaining a good adaptive immune response [210]. Other work showed that bovine-milk derived exosomes loaded with mRNA encoding the SARS-CoV-2 receptor-binding domain (RBD) stimulated the production of neutralizing antibodies against RBD in mice [211].
It is interesting to note that exosome-based vaccines against coronaviruses have been investigated since 2007 [212]. Thus, it was demonstrated that exosomes containing the Spike (S) proteins of SARS-CoV, which transmembrane and cytoplasmic domains were replaced by those of the vesicular stomatitis virus glycoprotein (VSV-G), induced high levels of neutralizing antibodies [212]. In the context of the COVID-19 pandemic, this strategy of modifying exosomes as antigen-presenting platforms has been relaunched by a growing number of start-ups and private or semi-private biotech companies worldwide, interested in developing extracellular vesicle-based therapies and virus- and adjuvant-free vaccines based on recombinant exosomes [213]. In this respect, an extremely promising vaccine candidate that has passed pre-clinical trials proposes the inhalation delivery of exosomes of pulmonary origin, which carry the recombinant SARS-CoV-2 RBD. The process of this second strategy, related to a use of the exosome as an antigen-presenting device is described in Figure 4. The engineering purified exosomes had the advantage of being remarkably stable after lyophilization. They have been shown both to enhance RBD retention in the airways and lung parenchyma, and to induce RBD-specific IgG antibodies in mice. They also enabled a mucosal IgA response and an effective T cell response that was able to clear the virus in a challenge test [214]. To the best of our knowledge, this promising work has given new momentum to ongoing research by start-ups into exosome-based vaccine strategies against the Chikungunya, Zika, Dengue and West Nile viruses.
6. Challenges and issues
Exosomes are a promising platform for cell-free-based vaccine development. However, there are some challenges to consider. One issue is the formulation, with the necessity to produce reliable batches of exosomes with appropriate levels of antigens to provide protection [199] while controlling the immunogenicity of “non-antigen” components originating from the cell line used to produce the exosomes.
Similarly, one limitation is that exosomes are heterogeneous in nature and contain a large number of diverse proteins and materials from host cells, making it difficult to tailor their cargoes compared to liposomes [215]. While immunogens and adjuvants can be easily packaged in liposomes, incorporating desired components into exosomes has proven challenging at times [216]. Furthermore, when exosomes are injected into a patient, they will compete with endogenous exosomes, potentially affecting the desired therapeutic effects [215]. Therefore, exosomes from specific cell types could be detrimentally affected. Exosomes from DCs could be the most promising, however, they can be blocked from entering bone marrow derived dendritic cells (BMDC) or by binding of competitive ligands to the cell receptors [217].
The use of DCs exosomes as vaccine carriers is challenging because they are short-lived and sensitive to CTL-mediated elimination. Therefore, the antigen presentation by peptide-pulsed DCs has limited longevity in vivo. However, Luketic et al. have pointed out that antigen presentation by exosomes released from peptide-pulsed dendritic cells is not suppressed by the presence of active cytotoxic lymphocyte T (CTL) [218]. Indeed, they found that following DC immunization, even if the majority of injected DCs disappeared from the draining lymph nodes within 7 days, antigen presentation has persisted for at least 14 days.
7. Concluding remarks
The global health crisis caused by SARS-CoV-2 has reinforced the need to diversify vaccine strategies against viral infections. This, in turn, has renewed interest in exosomes as particularly attractive tools, serving as biocompatible nanocarriers. We therefore set out to review the literature on the biology of exosomes, which mediate signaling between cells and play a key role in both physiological and pathological processes. We pointed out their considerable importance as potential biomarkers, diagnostic and therapeutic tools for contemporary medicine. Their role in viral infections is increasingly being studied, and we have pointed out that it is often dual, complicating the prospects for their therapeutic use. However, advances in the engineering of these vesicles for custom loading with protein or mRNA confirm that they are interesting carriers for new therapeutic and for being vaccine candidates, unveiling novel strategies in viral infection control. The recent demonstration of their effective use in preclinical models indicates that they constitute original platforms for antigen presentation or mRNA vectorization systems. This paves the way of exploring these new approaches to the control of flavivirus infections, such as DENV or ZIKV, for which this task has turned challenging. The transition to clinical trials will of course be necessary to validate these approaches and to verify the issues inherent in cell-derived material and noted in point 6.
Author Contributions
Conceptualization, D.E.S., M.K., A.M., A.L., U.H., G.L., W.V., and P.K-T.; Methodology; resources; writing—original draft preparation, D.E.S., M.K., A.M., A.L., U.H., G.L., W.V., and P.K-T.; Figure drawings, D.E.S., G.L., and M.K.; Writing—review and editing, P.K.-T.; Visualization, P.K.-T. and W.V.; Supervision, P.K.-T.; Funding acquisition, P.K.-T. and W.V. All authors have read and agreed to the published version of the manuscript.
Funding
D.E.S. has PhD degree scholarships from Réunion University (Ecole doctorale STS) funded by DIRED/2021-1115 Région Réunion and M.K. has a PhD degree scholarship from Nantes University (Ecole doctorale BS) funded by the French ministry MEESR. U.H. received financial support from ITMO Cancer of Aviesan on funds administered by Inserm.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
All figures were created using Biorender.com (accessed on the 1st of December).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ALIX | ALG-2 Interacting protein X |
| APCs | Antigen-Presenting Cells |
| APOBEC3G | Apolipoprotein B mRNA editing enzyme catalytic polypeptide-like 3G |
| BMDC | Bone marrow derived dendritic cells |
| CCL2 | C-C motif chemokine ligand 2 |
| cGAS-STING | cyclic GMP-AMP synthase stimulator of interferon genes |
| CHIKV | Chikungunya virus |
| CHMP4 | Charged multivesicular body protein 4a |
| circRNAs | circular RNAs |
| CMV | Cytomegalovirus |
| CTL | Cytotoxic Lymphocyte T |
| DC | Dendritic cell |
| DENV | Dengue virus |
| DUBs | de-ubiquitylating enzymes |
| EBV | Epstein-Barr virus |
| EMT | Epithelial-mesenchymal transition |
| ESCRT | Endosomal Sorting Complex Required for Transport |
| EV | Extracellular vesicles |
| HBsAg | Hepatitis B antigen |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| HGF | hepatocyte growth factor |
| HIV | Human Immunodeficiency virus |
| HLA-DR | Human Leukocyte Antigen DR isotype |
| HNRNPs | Heterogeneous Nuclear Ribonucleoproteins |
| HNSCC | Head and Neck Squamous Cell Carcinoma |
| HSP70 | Heat Shock Protein 70 |
| ICAM-1 | Intercellular Adhesion Molecule 1 |
| IC50 | half maximal Inhibitory Concentration |
| IFITM3 | Interferon-induced transmembrane protein 3 |
| IFNα | Interferon alpha |
| IFNγ | Interferon gamma |
| IgG | immunoglobulin G |
| IL-1β | Interleukin-1β |
| ILVs | Intraluminal vesicles |
| IP-10 | Interferon gamma-induced protein |
| ISGs | Interferon-stimulated genes |
| JEV | Japanese Encephalitis virus |
| L1 | non-LTR retrotransposons LINE-1 |
| LPS | Lipopolysaccharides |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MEVs | Multi-epitopic vaccines |
| MHC | Major Histocompatibility Complex |
| mo-DCs | monocyte-derived dendritic cells |
| MVBs | Multivesicular bodies |
| NS1 | Non-Structural protein-1 |
| PRRs | Pattern Recognition Receptors |
| RANTES | Regulated upon activation normal T cell expressed and secreted |
| RBD | Receptor-binding domain |
| RLRs | RIG-I like receptors |
| RSV | Respiratory syncytial virus |
| SIgA | Secretory IgA |
| SNARE | Soluble NSF attachment protein receptor |
| SARS-Cov2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| STING | Stimulator of interferon genes |
| TAR | Trans-activation response element |
| TBEV | Tick-borne encephalitis virus |
| TGFβ | Tumor Growth Factor β |
| TIM4 | T-cell immunoglobulin- and mucin-domain-containing molecule 4 |
| TNFα | Tumor Necrosis Factor α |
| TSG101 | Tumor susceptibility gene 101 |
| VLP | Virus-like particles |
| VSV-G | vesicular stomatitis virus glycoprotein |
| WNV | West Nile virus |
| ZIKV | Zika virus |
References
- C. Théry, L. Zitvogel, et S. Amigorena, « Exosomes: composition, biogenesis and function », Nat. Rev. Immunol., vol. 2, no 8, p. 569-579, août 2002. [CrossRef]
- M. F. S. Lindenbergh et W. Stoorvogel, « Antigen Presentation by Extracellular Vesicles from Professional Antigen-Presenting Cells », Annu. Rev. Immunol., vol. 36, p. 435-459, avr. 2018. [CrossRef]
- A. A. Danilushkina, C. C. Emene, N. A. Barlev, et M. O. Gomzikova, « Strategies for Engineering of Extracellular Vesicles », Int. J. Mol. Sci., vol. 24, no 17, p. 13247, août 2023. [CrossRef]
- A. Hendrix et al., « Extracellular vesicle analysis », Nat. Rev. Methods Primer, vol. 3, no 1, p. 56, juill. 2023. [CrossRef]
- G. van Niel, G. D’Angelo, et G. Raposo, « Shedding light on the cell biology of extracellular vesicles », Nat. Rev. Mol. Cell Biol., vol. 19, no 4, p. 213-228, avr. 2018. [CrossRef]
- L. M. Doyle et M. Z. Wang, « Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis », Cells, vol. 8, no 7, p. 727, juill. 2019. [CrossRef]
- R. Kalluri et V. S. LeBleu, « The biology, function, and biomedical applications of exosomes », Science, vol. 367, no 6478, p. eaau6977, févr. 2020. [CrossRef]
- B. Cappe, P. Vandenabeele, et F. B. Riquet, « A guide to the expanding field of extracellular vesicles and their release in regulated cell death programs », FEBS J., oct. 2023. [CrossRef]
- B. L. Deatherage et B. T. Cookson, « Membrane vesicle release in bacteria, eukaryotes, and archaea: a conserved yet underappreciated aspect of microbial life », Infect. Immun., vol. 80, no 6, p. 1948-1957, juin 2012. [CrossRef]
- C. de la Torre Gomez, R. V. Goreham, J. J. Bech Serra, T. Nann, et M. Kussmann, « “Exosomics”-A Review of Biophysics, Biology and Biochemistry of Exosomes With a Focus on Human Breast Milk », Front. Genet., vol. 9, p. 92, 2018. [CrossRef]
- A. Yokoi et T. Ochiya, « Exosomes and extracellular vesicles: Rethinking the essential values in cancer biology », Semin. Cancer Biol., vol. 74, p. 79-91, sept. 2021. [CrossRef]
- M. Izadpanah, A. Seddigh, S. Ebrahimi Barough, S. A. S. Fazeli, et J. Ai, « Potential of Extracellular Vesicles in Neurodegenerative Diseases: Diagnostic and Therapeutic Indications », J. Mol. Neurosci. MN, vol. 66, no 2, p. 172-179, oct. 2018. [CrossRef]
- D. Petroni, C. Fabbri, S. Babboni, L. Menichetti, G. Basta, et S. Del Turco, « Extracellular Vesicles and Intercellular Communication: Challenges for In Vivo Molecular Imaging and Tracking », Pharmaceutics, vol. 15, no 6, p. 1639, juin 2023. [CrossRef]
- C. Théry, « Exosomes: secreted vesicles and intercellular communications », F1000 Biol. Rep., vol. 3, juill. 2011. [CrossRef]
- J. Meldolesi, « Unconventional Protein Secretion Dependent on Two Extracellular Vesicles: Exosomes and Ectosomes », Front. Cell Dev. Biol., vol. 10, p. 877344, juin 2022. [CrossRef]
- G. van Niel, D. R. F. Carter, A. Clayton, D. W. Lambert, G. Raposo, et P. Vader, « Challenges and directions in studying cell–cell communication by extracellular vesicles », Nat. Rev. Mol. Cell Biol., vol. 23, no 5, p. 369-382, mai 2022. [CrossRef]
- T. Mori, L. Giovannelli, A. R. Bilia, et F. Margheri, « Exosomes: Potential Next-Generation Nanocarriers for the Therapy of Inflammatory Diseases », Pharmaceutics, vol. 15, no 9, p. 2276, sept. 2023. [CrossRef]
- S. Li, Z. Lin, X. Jiang, et X. Yu, « Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools », Acta Pharmacol. Sin., vol. 39, no 4, p. 542-551, avr. 2018. [CrossRef]
- S. V. Krylova et D. Feng, « The Machinery of Exosomes: Biogenesis, Release, and Uptake », Int. J. Mol. Sci., vol. 24, no 2, p. 1337, janv. 2023. [CrossRef]
- J. Gruenberg, « Life in the lumen: The multivesicular endosome », Traffic, vol. 21, no 1, p. 76-93, janv. 2020. [CrossRef]
- N. Naslavsky et S. Caplan, « The enigmatic endosome – sorting the ins and outs of endocytic trafficking », J. Cell Sci., vol. 131, no 13, p. jcs216499, juill. 2018. [CrossRef]
- A. Norris et B. D. Grant, « Endosomal microdomains: Formation and function », Curr. Opin. Cell Biol., vol. 65, p. 86-95, août 2020. [CrossRef]
- J. A. Solinger et A. Spang, « Sorting of cargo in the tubular endosomal network », BioEssays, vol. 44, no 12, p. 2200158, déc. 2022. [CrossRef]
- S. Sigismund, L. Lanzetti, G. Scita, et P. P. Di Fiore, « Endocytosis in the context-dependent regulation of individual and collective cell properties », Nat. Rev. Mol. Cell Biol., vol. 22, no 9, p. 625-643, sept. 2021. [CrossRef]
- A.-K. Pfitzner et al., « Vps60 initiates alternative ESCRT-III filaments », J. Cell Biol., vol. 222, no 11, p. e202206028, nov. 2023. [CrossRef]
- M. Colombo, G. Raposo, et C. Théry, « Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles », Annu. Rev. Cell Dev. Biol., vol. 30, no 1, p. 255-289, oct. 2014. [CrossRef]
- Y. Kuchitsu et T. Taguchi, « Lysosomal microautophagy: an emerging dimension in mammalian autophagy », Trends Cell Biol., p. S0962892423002386, déc. 2023. [CrossRef]
- J. Huotari et A. Helenius, « Endosome maturation: Endosome maturation », EMBO J., vol. 30, no 17, p. 3481-3500, août 2011. [CrossRef]
- J. G. Carlton et B. Baum, « Roles of ESCRT-III polymers in cell division across the tree of life », Curr. Opin. Cell Biol., vol. 85, p. 102274, déc. 2023. [CrossRef]
- J. Schöneberg, I.-H. Lee, J. H. Iwasa, et J. H. Hurley, « Reverse-topology membrane scission by the ESCRT proteins », Nat. Rev. Mol. Cell Biol., vol. 18, no 1, p. 5-17, janv. 2017. [CrossRef]
- Y. Rivera-Cuevas et V. B. Carruthers, « The multifaceted interactions between pathogens and host ESCRT machinery », PLOS Pathog., vol. 19, no 5, p. e1011344, mai 2023. [CrossRef]
- S. L. N. Maas, X. O. Breakefield, et A. M. Weaver, « Extracellular Vesicles: Unique Intercellular Delivery Vehicles », Trends Cell Biol., vol. 27, no 3, p. 172-188, mars 2017. [CrossRef]
- M. A. Karim, D. R. Samyn, S. Mattie, et C. L. Brett, « Distinct features of multivesicular body-lysosome fusion revealed by a new cell-free content-mixing assay », Traffic Cph. Den., vol. 19, no 2, p. 138-149, févr. 2018. [CrossRef]
- N. P. Hessvik et A. Llorente, « Current knowledge on exosome biogenesis and release », Cell. Mol. Life Sci., vol. 75, no 2, p. 193-208, janv. 2018. [CrossRef]
- M. Mathieu, L. Martin-Jaular, G. Lavieu, et C. Théry, « Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication », Nat. Cell Biol., vol. 21, no 1, p. 9-17, janv. 2019. [CrossRef]
- F. Ghorbaninezhad et al., « Dendritic cell-derived exosomes: A new horizon in personalized cancer immunotherapy? », Cancer Lett., vol. 562, p. 216168, mai 2023. [CrossRef]
- X. Jin, T. Xia, S. Luo, Y. Zhang, Y. Xia, et H. Yin, « Exosomal lipid PI4P regulates small extracellular vesicle secretion by modulating intraluminal vesicle formation », J. Extracell. Vesicles, vol. 12, no 4, p. 12319, avr. 2023. [CrossRef]
- S. M. Crivelli et al., « Function of ceramide transfer protein for biogenesis and sphingolipid composition of extracellular vesicles », J. Extracell. Vesicles, vol. 11, no 6, p. e12233, juin 2022. [CrossRef]
- D. Wei et al., « RAB31 marks and controls an ESCRT-independent exosome pathway », Cell Res., vol. 31, no 2, p. 157-177, févr. 2021. [CrossRef]
- R. Horbay, A. Hamraghani, L. Ermini, S. Holcik, S. T. Beug, et B. Yeganeh, « Role of Ceramides and Lysosomes in Extracellular Vesicle Biogenesis, Cargo Sorting and Release », Int. J. Mol. Sci., vol. 23, no 23, p. 15317, déc. 2022. [CrossRef]
- B. Tancini et al., « Lysosomal Exocytosis: The Extracellular Role of an Intracellular Organelle », Membranes, vol. 10, no 12, p. 406, déc. 2020. [CrossRef]
- A. C. Dixson, T. R. Dawson, D. Di Vizio, et A. M. Weaver, « Context-specific regulation of extracellular vesicle biogenesis and cargo selection », Nat. Rev. Mol. Cell Biol., vol. 24, no 7, p. 454-476, juill. 2023. [CrossRef]
- D. Perez-Hernandez et al., « The Intracellular Interactome of Tetraspanin-enriched Microdomains Reveals Their Function as Sorting Machineries toward Exosomes », J. Biol. Chem., vol. 288, no 17, p. 11649-11661, avr. 2013. [CrossRef]
- T. Zheng, S. Xu, et J. Xu, « A Review of the Roles of Specialized Extracellular Vesicles, Migrasomes, and Exosomes in Normal Cell Physiology and Disease », Med. Sci. Monit., vol. 29, avr. 2023. [CrossRef]
- J.-M. Escola, M. J. Kleijmeer, W. Stoorvogel, J. M. Griffith, O. Yoshie, et H. J. Geuze, « Selective Enrichment of Tetraspan Proteins on the Internal Vesicles of Multivesicular Endosomes and on Exosomes Secreted by Human B-lymphocytes », J. Biol. Chem., vol. 273, no 32, p. 20121-20127, août 1998. [CrossRef]
- H. Wei et al., « Regulation of exosome production and cargo sorting », Int. J. Biol. Sci., vol. 17, no 1, p. 163-177, 2021. [CrossRef]
- W. Suwakulsiri et al., « Comparative proteomic analysis of three major extracellular vesicle classes secreted from human primary and metastatic colorectal cancer cells: Exosomes, microparticles, and shed midbody remnants », PROTEOMICS, p. 2300057, juill. 2023. [CrossRef]
- M. Mathieu et al., « Specificities of exosome versus small ectosome secretion revealed by live intracellular tracking of CD63 and CD9 », Nat. Commun., vol. 12, no 1, p. 4389, juill. 2021. [CrossRef]
- J. Kowal et al., « Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes », Proc. Natl. Acad. Sci., vol. 113, no 8, févr. 2016. [CrossRef]
- S. Gurung, D. Perocheau, L. Touramanidou, et J. Baruteau, « The exosome journey: from biogenesis to uptake and intracellular signalling », Cell Commun. Signal., vol. 19, no 1, p. 47, avr. 2021. [CrossRef]
- R. J. Simpson, S. S. Jensen, et J. W. E. Lim, « Proteomic profiling of exosomes: Current perspectives », PROTEOMICS, vol. 8, no 19, p. 4083-4099, oct. 2008. [CrossRef]
- M. P. Zaborowski, L. Balaj, X. O. Breakefield, et C. P. Lai, « Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study », BioScience, vol. 65, no 8, p. 783-797, août 2015. [CrossRef]
- D. K. Jeppesen et al., « Reassessment of Exosome Composition », Cell, vol. 177, no 2, p. 428-445.e18, avr. 2019. [CrossRef]
- F. G. Kugeratski et al., « Quantitative proteomics identifies the core proteome of exosomes with syntenin-1 as the highest abundant protein and a putative universal biomarker », Nat. Cell Biol., vol. 23, no 6, p. 631-641, juin 2021. [CrossRef]
- Z. Andreu et M. Yáñez-MÃ3, « Tetraspanins in Extracellular Vesicle Formation and Function », Front. Immunol., vol. 5, sept. 2014. [CrossRef]
- I. Casari, J. A. Howard, E. E. Robless, et M. Falasca, « Exosomal integrins and their influence on pancreatic cancer progression and metastasis », Cancer Lett., vol. 507, p. 124-134, juin 2021. [CrossRef]
- A. Hoshino et al., « Tumour exosome integrins determine organotropic metastasis », Nature, vol. 527, no 7578, p. 329-335, nov. 2015. [CrossRef]
- E. Segura et al., « ICAM-1 on exosomes from mature dendritic cells is critical for efficient naive T-cell priming », Blood, vol. 106, no 1, p. 216-223, juill. 2005. [CrossRef]
- G. Raposo et al., « B lymphocytes secrete antigen-presenting vesicles. », J. Exp. Med., vol. 183, no 3, p. 1161-1172, mars 1996. [CrossRef]
- J. E. A. Braun, « Extracellular chaperone networks and the export of J-domain proteins », J. Biol. Chem., vol. 299, no 2, p. 102840, févr. 2023. [CrossRef]
- M. Linder et E. Pogge von Strandmann, « The Role of Extracellular HSP70 in the Function of Tumor-Associated Immune Cells », Cancers, vol. 13, no 18, p. 4721, sept. 2021. [CrossRef]
- Y. Qu, L. Franchi, G. Nunez, et G. R. Dubyak, « Nonclassical IL-1β Secretion Stimulated by P2X7 Receptors Is Dependent on Inflammasome Activation and Correlated with Exosome Release in Murine Macrophages », J. Immunol., vol. 179, no 3, p. 1913-1925, août 2007. [CrossRef]
- Y. Liao et al., « The Ras GTPASE -activating-like protein IQGAP1 bridges Gasdermin D to the ESCRTsystem to promote IL -1β release via exosomes », EMBO J., vol. 42, no 1, p. e110780, janv. 2023. [CrossRef]
- B. Weber et al., « Diagnostic and Prognostic Potential of Exosomal Cytokines IL-6 and IL-10 in Polytrauma Patients », Int. J. Mol. Sci., vol. 24, no 14, p. 11830, juill. 2023. [CrossRef]
- M. P. Bebelman, P. Bun, S. Huveneers, G. van Niel, D. M. Pegtel, et F. J. Verweij, « Real-time imaging of multivesicular body–plasma membrane fusion to quantify exosome release from single cells », Nat. Protoc., vol. 15, no 1, p. 102-121, janv. 2020. [CrossRef]
- T. Skotland, K. Sandvig, et A. Llorente, « Lipids in exosomes: Current knowledge and the way forward », Prog. Lipid Res., vol. 66, p. 30-41, avr. 2017. [CrossRef]
- R. C. Piper et D. J. Katzmann, « Biogenesis and Function of Multivesicular Bodies », Annu. Rev. Cell Dev. Biol., vol. 23, no 1, p. 519-547, nov. 2007. [CrossRef]
- K. O’Brien, K. Breyne, S. Ughetto, L. C. Laurent, et X. O. Breakefield, « RNA delivery by extracellular vesicles in mammalian cells and its applications », Nat. Rev. Mol. Cell Biol., vol. 21, no 10, p. 585-606, oct. 2020. [CrossRef]
- L. Geis-Asteggiante et al., « Differential Content of Proteins, mRNAs, and miRNAs Suggests that MDSC and Their Exosomes May Mediate Distinct Immune Suppressive Functions », J. Proteome Res., vol. 17, no 1, p. 486-498, janv. 2018. [CrossRef]
- C. Corrado, M. M. Barreca, C. Zichittella, R. Alessandro, et A. Conigliaro, « Molecular Mediators of RNA Loading into Extracellular Vesicles », Cells, vol. 10, no 12, p. 3355, nov. 2021. [CrossRef]
- M. Omrani et al., « Global trend in exosome isolation and application: an update concept in management of diseases », Mol. Cell. Biochem., mai 2023. [CrossRef]
- R. Heydari et al., « Exosomes as Rheumatoid Arthritis Diagnostic Biomarkers and Therapeutic Agents », Vaccines, vol. 11, no 3, p. 687, mars 2023. [CrossRef]
- F. Tavasolian et al., « Exosomes: Effectual players in rheumatoid arthritis », Autoimmun. Rev., vol. 19, no 6, p. 102511, juin 2020. [CrossRef]
- J. Silva et al., « Vesicle-related microRNAs in plasma of nonsmall cell lung cancer patients and correlation with survival », Eur. Respir. J., vol. 37, no 3, p. 617-623, mars 2011. [CrossRef]
- M. Yáñez-Mó et al., « Biological properties of extracellular vesicles and their physiological functions », J. Extracell. Vesicles, vol. 4, p. 27066, 2015. [CrossRef]
- Y. Takahashi et al., « Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection », J. Biotechnol., vol. 165, no 2, p. 77-84, mai 2013. [CrossRef]
- H. M. Ramos-Zaldívar et al., « Extracellular vesicles through the blood–brain barrier: a review », Fluids Barriers CNS, vol. 19, no 1, p. 60, juill. 2022. [CrossRef]
- N. García-Romero et al., « DNA sequences within glioma-derived extracellular vesicles can cross the intact blood-brain barrier and be detected in peripheral blood of patients », Oncotarget, vol. 8, no 1, p. 1416-1428, janv. 2017. [CrossRef]
- M. Abdelsalam, M. Ahmed, Z. Osaid, R. Hamoudi, et R. Harati, « Insights into Exosome Transport through the Blood–Brain Barrier and the Potential Therapeutical Applications in Brain Diseases », Pharmaceuticals, vol. 16, no 4, p. 571, avr. 2023. [CrossRef]
- G. Andreola et al., « Induction of Lymphocyte Apoptosis by Tumor Cell Secretion of FasL-bearing Microvesicles », J. Exp. Med., vol. 195, no 10, p. 1303-1316, mai 2002. [CrossRef]
- M. Somiya et S. Kuroda, « Reporter gene assay for membrane fusion of extracellular vesicles », J. Extracell. Vesicles, vol. 10, no 13, p. e12171, nov. 2021. [CrossRef]
- D. Levy, M. A. Do, J. Zhang, A. Brown, et B. Lu, « Orchestrating Extracellular Vesicle With Dual Reporters for Imaging and Capturing in Mammalian Cell Culture », Front. Mol. Biosci., vol. 8, p. 680580, juin 2021. [CrossRef]
- L. A. Mulcahy, R. C. Pink, et D. R. F. Carter, « Routes and mechanisms of extracellular vesicle uptake », J. Extracell. Vesicles, vol. 3, no 1, p. 24641, janv. 2014. [CrossRef]
- L. Ginini, S. Billan, E. Fridman, et Z. Gil, « Insight into Extracellular Vesicle-Cell Communication: From Cell Recognition to Intracellular Fate », Cells, vol. 11, no 9, p. 1375, avr. 2022. [CrossRef]
- A. Gonda, J. Kabagwira, G. N. Senthil, et N. R. Wall, « Internalization of Exosomes through Receptor-Mediated Endocytosis », Mol. Cancer Res., vol. 17, no 2, p. 337-347, févr. 2019. [CrossRef]
- M. Miyanishi, K. Tada, M. Koike, Y. Uchiyama, T. Kitamura, et S. Nagata, « Identification of Tim4 as a phosphatidylserine receptor », Nature, vol. 450, no 7168, p. 435-439, nov. 2007. [CrossRef]
- B. Sims et al., « Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells », Int. J. Nanomedicine, vol. Volume 12, p. 4823-4833, juill. 2017. [CrossRef]
- H. C. Christianson, K. J. Svensson, T. H. van Kuppevelt, J.-P. Li, et M. Belting, « Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity », Proc. Natl. Acad. Sci., vol. 110, no 43, p. 17380-17385, oct. 2013. [CrossRef]
- J. Ochieng et al., « Extracellular histones are the ligands for the uptake of exosomes and hydroxyapatite-nanoparticles by tumor cells via syndecan-4 », FEBS Lett., vol. 592, no 19, p. 3274-3285, oct. 2018. [CrossRef]
- E. Bonsergent, E. Grisard, J. Buchrieser, O. Schwartz, C. Théry, et G. Lavieu, « Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells », Nat. Commun., vol. 12, no 1, p. 1864, mars 2021. [CrossRef]
- A. Zomer et al., « In Vivo Imaging Reveals Extracellular Vesicle-Mediated Phenocopying of Metastatic Behavior », Cell, vol. 161, no 5, p. 1046-1057, mai 2015. [CrossRef]
- B. Costa-Silva et al., « Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver », Nat. Cell Biol., vol. 17, no 6, p. 816-826, juin 2015. [CrossRef]
- H. Peinado et al., « Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET », Nat. Med., vol. 18, no 6, p. 883-891, juin 2012. [CrossRef]
- L. G. Lima et al., « Tumor microenvironmental cytokines bound to cancer exosomes determine uptake by cytokine receptor-expressing cells and biodistribution », Nat. Commun., vol. 12, no 1, p. 3543, juin 2021. [CrossRef]
- H. Valadi, K. Ekström, A. Bossios, M. Sjöstrand, J. J. Lee, et J. O. Lötvall, « Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells », Nat. Cell Biol., vol. 9, no 6, p. 654-659, juin 2007. [CrossRef]
- X. You et al., « Exosomal miR-663b exposed to TGF-β1 promotes cervical cancer metastasis and epithelial-mesenchymal transition by targeting MGAT3 », Oncol. Rep., vol. 45, no 4, p. 12, avr. 2021. [CrossRef]
- V. Vignard et al., « MicroRNAs in Tumor Exosomes Drive Immune Escape in Melanoma », Cancer Immunol. Res., vol. 8, no 2, p. 255-267, févr. 2020. [CrossRef]
- N. Milman, L. Ginini, et Z. Gil, « Exosomes and their role in tumorigenesis and anticancer drug resistance », Drug Resist. Updat., vol. 45, p. 1-12, juill. 2019. [CrossRef]
- G. K. Patel et al., « Exosomes confer chemoresistance to pancreatic cancer cells by promoting ROS detoxification and miR-155-mediated suppression of key gemcitabine-metabolising enzyme, DCK », Br. J. Cancer, vol. 116, no 5, p. 609-619, févr. 2017. [CrossRef]
- S. A. A. Kooijmans, R. M. Schiffelers, N. Zarovni, et R. Vago, « Modulation of tissue tropism and biological activity of exosomes and other extracellular vesicles: New nanotools for cancer treatment », Pharmacol. Res., vol. 111, p. 487-500, sept. 2016. [CrossRef]
- K. Qian, W. Fu, T. Li, J. Zhao, C. Lei, et S. Hu, « The roles of small extracellular vesicles in cancer and immune regulation and translational potential in cancer therapy », J. Exp. Clin. Cancer Res., vol. 41, no 1, p. 286, sept. 2022. [CrossRef]
- A. F. Teixeira, Y. Wang, J. Iaria, P. ten Dijke, et H.-J. Zhu, « Simultaneously targeting extracellular vesicle trafficking and TGF-β receptor kinase activity blocks signaling hyperactivation and metastasis », Signal Transduct. Target. Ther., vol. 8, no 1, p. 456, déc. 2023. [CrossRef]
- T. Briolay, T. Petithomme, M. Fouet, N. Nguyen-Pham, C. Blanquart, et N. Boisgerault, « Delivery of cancer therapies by synthetic and bio-inspired nanovectors », Mol. Cancer, vol. 20, no 1, p. 55, mars 2021. [CrossRef]
- A. Pritchard et al., « Lung Tumor Cell-Derived Exosomes Promote M2 Macrophage Polarization », Cells, vol. 9, no 5, p. 1303, mai 2020. [CrossRef]
- Z. Zhang et al., « Exosomes derived from human adipose mesenchymal stem cells ameliorate hepatic fibrosis by inhibiting PI3K/Akt/mTOR pathway and remodeling choline metabolism », J. Nanobiotechnology, vol. 21, no 1, p. 29, janv. 2023. [CrossRef]
- J. Jiang, J. Li, X. Zhou, X. Zhao, B. Huang, et Y. Qin, « Exosomes Regulate the Epithelial–Mesenchymal Transition in Cancer », Front. Oncol., vol. 12, p. 864980, mars 2022. [CrossRef]
- Y. Yao, R. Chen, G. Wang, Y. Zhang, et F. Liu, « Exosomes derived from mesenchymal stem cells reverse EMT via TGF-β1/Smad pathway and promote repair of damaged endometrium », Stem Cell Res. Ther., vol. 10, no 1, p. 225, déc. 2019. [CrossRef]
- Z. Lang et al., « Hepatocyte-derived exosomal miR-146a-5p inhibits hepatic stellate cell EMT process: a crosstalk between hepatocytes and hepatic stellate cells », Cell Death Discov., vol. 9, no 1, p. 304, août 2023. [CrossRef]
- J. Lin et al., « Exosomes: Novel Biomarkers for Clinical Diagnosis », Sci. World J., vol. 2015, p. 1-8, 2015. [CrossRef]
- S. Keller, J. Ridinger, A.-K. Rupp, J. W. Janssen, et P. Altevogt, « Body fluid derived exosomes as a novel template for clinical diagnostics », J. Transl. Med., vol. 9, no 1, p. 86, déc. 2011. [CrossRef]
- N. Ludwig, T. L. Whiteside, et T. E. Reichert, « Challenges in Exosome Isolation and Analysis in Health and Disease », Int. J. Mol. Sci., vol. 20, no 19, p. 4684, sept. 2019. [CrossRef]
- S. R. Douglas, K. T. Yeung, J. Yang, S. L. Blair, O. Cohen, et B. P. Eliceiri, « Identification of CD105+ Extracellular Vesicles as a Candidate Biomarker for Metastatic Breast Cancer », J. Surg. Res., vol. 268, p. 168-173, déc. 2021. [CrossRef]
- D. Osti et al., « Clinical Significance of Extracellular Vesicles in Plasma from Glioblastoma Patients », Clin. Cancer Res., vol. 25, no 1, p. 266-276, janv. 2019. [CrossRef]
- Q. Sabbagh et al., « The von Willebrand factor stamps plasmatic extracellular vesicles from glioblastoma patients », Sci. Rep., vol. 11, no 1, p. 22792, nov. 2021. [CrossRef]
- B. K. Thakur et al., « Double-stranded DNA in exosomes: a novel biomarker in cancer detection », Cell Res., vol. 24, no 6, p. 766-769, juin 2014. [CrossRef]
- F. Fu, W. Jiang, L. Zhou, et Z. Chen, « Circulating Exosomal miR-17-5p and miR-92a-3p Predict Pathologic Stage and Grade of Colorectal Cancer », Transl. Oncol., vol. 11, no 2, p. 221-232, avr. 2018. [CrossRef]
- J. C. Akers et al., « miR-21 in the Extracellular Vesicles (EVs) of Cerebrospinal Fluid (CSF): A Platform for Glioblastoma Biomarker Development », PLoS ONE, vol. 8, no 10, p. e78115, oct. 2013. [CrossRef]
- Y. Wang et al., « Exosomal circRNAs: biogenesis, effect and application in human diseases », Mol. Cancer, vol. 18, no 1, p. 116, déc. 2019. [CrossRef]
- J. Li et al., « Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis », J. Exp. Clin. Cancer Res., vol. 37, no 1, p. 177, déc. 2018. [CrossRef]
- Z. Zhou et al., « Urinary exosomes: a promising biomarker of drug-induced nephrotoxicity », Front. Med., vol. 10, p. 1251839, sept. 2023. [CrossRef]
- C. He, S. Zheng, Y. Luo, et B. Wang, « Exosome Theranostics: Biology and Translational Medicine », Theranostics, vol. 8, no 1, p. 237-255, 2018. [CrossRef]
- A. Makler et W. Asghar, « Exosomal biomarkers for cancer diagnosis and patient monitoring », Expert Rev. Mol. Diagn., vol. 20, no 4, p. 387-400, avr. 2020. [CrossRef]
- R. Tutrone et al., « Clinical utility of the exosome based ExoDx Prostate(IntelliScore) EPI test in men presenting for initial Biopsy with a PSA 2–10 ng/mL », Prostate Cancer Prostatic Dis., vol. 23, no 4, p. 607-614, déc. 2020. [CrossRef]
- H. S. Chahar, T. Corsello, A. S. Kudlicki, N. Komaravelli, et A. Casola, « Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells », Sci. Rep., vol. 8, no 1, p. 387, janv. 2018. [CrossRef]
- Y. Peng, Y. Yang, Y. Li, T. Shi, Y. Luan, et C. Yin, « Exosome and virus infection », Front. Immunol., vol. 14, p. 1154217, mars 2023. [CrossRef]
- Z. Li et al., « Porcine Hemagglutinating Encephalomyelitis Virus Co-Opts Multivesicular-Derived Exosomes for Transmission », mBio, vol. 14, no 1, p. e03054-22, févr. 2023. [CrossRef]
- Y. Sato et al., « Epstein–Barr virus tegument protein BGLF2 in exosomes released from virus-producing cells facilitates de novo infection », Cell Commun. Signal., vol. 20, no 1, p. 95, déc. 2022. [CrossRef]
- C. Zhou et al., « Exosomes Carry microRNAs into Neighboring Cells to Promote Diffusive Infection of Newcastle Disease Virus », Viruses, vol. 11, no 6, p. 527, juin 2019. [CrossRef]
- S. Kopcho, M. McDew-White, W. Naushad, M. Mohan, et C. M. Okeoma, « SIV Infection Regulates Compartmentalization of Circulating Blood Plasma miRNAs within Extracellular Vesicles (EVs) and Extracellular Condensates (ECs) and Decreases EV-Associated miRNA-128 », Viruses, vol. 15, no 3, p. 622, févr. 2023. [CrossRef]
- D. E. Safadi et al., « Extracellular Vesicles Are Conveyors of the NS1 Toxin during Dengue Virus and Zika Virus Infection », Viruses, vol. 15, no 2, p. 364, janv. 2023. [CrossRef]
- Y. Ju, H. Bai, L. Ren, et L. Zhang, « The Role of Exosome and the ESCRT Pathway on Enveloped Virus Infection », Int. J. Mol. Sci., vol. 22, no 16, p. 9060, août 2021. [CrossRef]
- C. Martin, G. Ligat, et C. E. Malnou, « The Yin and the Yang of extracellular vesicles during viral infections », Biomed. J., p. 100659, sept. 2023. [CrossRef]
- C. Thepparit, S. Khongwichit, K. Ketsuwan, S. Libsittikul, P. Auewarakul, et D. R. Smith, « Dengue virus requires apoptosis linked gene-2-interacting protein X (ALIX) for viral propagation », Virus Res., vol. 261, p. 65-71, févr. 2019. [CrossRef]
- P.-T.-H. Tran, A. I. Chiramel, M. Johansson, et W. Melik, « Roles of ESCRT Proteins ALIX and CHMP4A and Their Interplay with Interferon-Stimulated Gene 15 during Tick-Borne Flavivirus Infection », J. Virol., vol. 96, no 3, p. e01624-21, févr. 2022. [CrossRef]
- L. N. Carpp, R. Galler, et M. C. Bonaldo, « Interaction between the yellow fever virus nonstructural protein NS3 and the host protein Alix contributes to the release of infectious particles », Microbes Infect., vol. 13, no 1, p. 85-95, janv. 2011. [CrossRef]
- Q. Wu et al., « Presence of Intact Hepatitis B Virions in Exosomes », Cell. Mol. Gastroenterol. Hepatol., vol. 15, no 1, p. 237-259, 2023. [CrossRef]
- N. Raab-Traub et D. P. Dittmer, « Viral effects on the content and function of extracellular vesicles », Nat. Rev. Microbiol., vol. 15, no 9, p. 559-572, sept. 2017. [CrossRef]
- F. Rey-Cadilhac, F. Rachenne, D. Missé, et J. Pompon, « Viral Components Trafficking with(in) Extracellular Vesicles », Viruses, vol. 15, no 12, p. 2333, nov. 2023. [CrossRef]
- S. J. Gould, A. M. Booth, et J. E. K. Hildreth, « The Trojan exosome hypothesis », Proc. Natl. Acad. Sci., vol. 100, no 19, p. 10592-10597, sept. 2003. [CrossRef]
- A. M. Nour, Y. Li, J. Wolenski, et Y. Modis, « Viral Membrane Fusion and Nucleocapsid Delivery into the Cytoplasm are Distinct Events in Some Flaviviruses », PLoS Pathog., vol. 9, no 9, p. e1003585, sept. 2013. [CrossRef]
- A. Longatti, B. Boyd, et F. V. Chisari, « Virion-Independent Transfer of Replication-Competent Hepatitis C Virus RNA between Permissive Cells », J. Virol., vol. 89, no 5, p. 2956-2961, mars 2015. [CrossRef]
- V. Ramakrishnaiah et al., « Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells », Proc. Natl. Acad. Sci., vol. 110, no 32, p. 13109-13113, août 2013. [CrossRef]
- B. C. T. Le et al., « Characterization and Involvement of Exosomes Originating from Chikungunya Virus-Infected Epithelial Cells in the Transmission of Infectious Viral Elements », Int. J. Mol. Sci., vol. 23, no 20, p. 12117, oct. 2022. [CrossRef]
- D. Cortes-Galvez, J. A. Dangerfield, et C. Metzner, « Extracellular Vesicles and Their Membranes: Exosomes vs. Virus-Related Particles », Membranes, vol. 13, no 4, p. 397, mars 2023. [CrossRef]
- A. Narayanan et al., « Exosomes Derived from HIV-1-infected Cells Contain Trans-activation Response Element RNA », J. Biol. Chem., vol. 288, no 27, p. 20014-20033, juill. 2013. [CrossRef]
- L. Chen et al., « Exosomes derived from HIV-1-infected cells promote growth and progression of cancer via HIV TAR RNA », Nat. Commun., vol. 9, no 1, p. 4585, nov. 2018. [CrossRef]
- C. Fan et al., « The emerging role of Epstein-Barr virus encoded microRNAs in nasopharyngeal carcinoma », J. Cancer, vol. 9, no 16, p. 2852-2864, 2018. [CrossRef]
- S. Mukherjee, I. Akbar, B. Kumari, S. Vrati, A. Basu, et A. Banerjee, « Japanese Encephalitis Virus-induced let-7a/b interacted with the NOTCH - TLR 7 pathway in microglia and facilitated neuronal death via caspase activation », J. Neurochem., vol. 149, no 4, p. 518-534, mai 2019. [CrossRef]
- E. Barberis et al., « Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection », Front. Mol. Biosci., vol. 8, p. 632290, févr. 2021. [CrossRef]
- Y. Fang, N. Wu, X. Gan, W. Yan, J. C. Morrell, et S. J. Gould, « Higher-Order Oligomerization Targets Plasma Membrane Proteins and HIV Gag to Exosomes », PLoS Biol., vol. 5, no 6, p. e158, juin 2007. [CrossRef]
- T. D. Campbell, M. Khan, M.-B. Huang, V. C. Bond, et M. D. Powell, « HIV-1 Nef protein is secreted into vesicles that can fuse with target cells and virions », Ethn. Dis., vol. 18, no 2 Suppl 2, p. S2-14-9, 2008.
- M. Lenassi et al., « HIV Nef is Secreted in Exosomes and Triggers Apoptosis in Bystander CD4 + T Cells », Traffic, vol. 11, no 1, p. 110-122, janv. 2010. [CrossRef]
- M. Aqil, A. R. Naqvi, A. S. Bano, et S. Jameel, « The HIV-1 Nef Protein Binds Argonaute-2 and Functions as a Viral Suppressor of RNA Interference », PLoS ONE, vol. 8, no 9, p. e74472, sept. 2013. [CrossRef]
- N. Plazolles, J.-M. Humbert, L. Vachot, B. Verrier, C. Hocke, et F. Halary, « Pivotal Advance: The promotion of soluble DC-SIGN release by inflammatory signals and its enhancement of cytomegalovirus-mediated cis -infection of myeloid dendritic cells », J. Leukoc. Biol., vol. 89, no 3, p. 329-342, oct. 2010. [CrossRef]
- R. Mishra, S. Lata, A. Ali, et A. C. Banerjea, « Dengue haemorrhagic fever: a job done via exosomes? », Emerg. Microbes Infect., vol. 8, no 1, p. 1626-1635, janv. 2019. [CrossRef]
- A. Clayton, C. L. Harris, J. Court, M. D. Mason, et B. P. Morgan, « Antigen-presenting cell exosomes are protected from complement-mediated lysis by expression of CD55 and CD59 », Eur. J. Immunol., vol. 33, no 2, p. 522-531, févr. 2003. [CrossRef]
- E. Karasu, S. U. Eisenhardt, J. Harant, et M. Huber-Lang, « Extracellular Vesicles: Packages Sent With Complement », Front. Immunol., vol. 9, p. 721, avr. 2018. [CrossRef]
- M. W. A. Hussain et al., « Exosomes for Regulation of Immune Responses and Immunotherapy », J. Nanotheranostics, vol. 3, no 1, p. 55-85, mars 2022. [CrossRef]
- R. Kulkarni et A. Prasad, « Exosomes Derived from HIV-1 Infected DCs Mediate Viral trans-Infection via Fibronectin and Galectin-3 », Sci. Rep., vol. 7, no 1, p. 14787, nov. 2017. [CrossRef]
- J. Klibi et al., « Blood diffusion and Th1-suppressive effects of galectin-9–containing exosomes released by Epstein-Barr virus–infected nasopharyngeal carcinoma cells », Blood, vol. 113, no 9, p. 1957-1966, févr. 2009. [CrossRef]
- C. Zhang et al., « Galectin-9 promotes a suppressive microenvironment in human cancer by enhancing STING degradation », Oncogenesis, vol. 9, no 7, p. 65, juill. 2020. [CrossRef]
- S. Temme, A. M. Eis-Hübinger, A. D. McLellan, et N. Koch, « The Herpes Simplex Virus-1 Encoded Glycoprotein B Diverts HLA-DR into the Exosome Pathway », J. Immunol., vol. 184, no 1, p. 236-243, janv. 2010. [CrossRef]
- H. Saari et al., « Extracellular vesicles provide a capsid-free vector for oncolytic adenoviral DNA delivery », J. Extracell. Vesicles, vol. 9, no 1, p. 1747206, sept. 2020. [CrossRef]
- Y. Kakiuchi et al., « Local oncolytic adenovirotherapy produces an abscopal effect via tumor-derived extracellular vesicles », Mol. Ther., vol. 29, no 10, p. 2920-2930, oct. 2021. [CrossRef]
- J. Dai et al., « Exosomes: key players in cancer and potential therapeutic strategy », Signal Transduct. Target. Ther., vol. 5, no 1, p. 145, août 2020. [CrossRef]
- W. Zhang, X. Jiang, J. Bao, Y. Wang, H. Liu, et L. Tang, « Exosomes in Pathogen Infections: A Bridge to Deliver Molecules and Link Functions », Front. Immunol., vol. 9, p. 90, févr. 2018. [CrossRef]
- Y. Kitai et al., « DNA-Containing Exosomes Derived from Cancer Cells Treated with Topotecan Activate a STING-Dependent Pathway and Reinforce Antitumor Immunity », J. Immunol., vol. 198, no 4, p. 1649-1659, févr. 2017. [CrossRef]
- A. Ablasser et S. Hur, « Regulation of cGAS- and RLR-mediated immunity to nucleic acids », Nat. Immunol., vol. 21, no 1, p. 17-29, janv. 2020. [CrossRef]
- A. Longatti, « The Dual Role of Exosomes in Hepatitis A and C Virus Transmission and Viral Immune Activation », Viruses, vol. 7, no 12, p. 6707-6715, déc. 2015. [CrossRef]
- B. J. Barnes et C. C. Somerville, « Modulating Cytokine Production via Select Packaging and Secretion From Extracellular Vesicles », Front. Immunol., vol. 11, p. 1040, mai 2020. [CrossRef]
- A. G. Souza et L. M. Colli, « Extracellular Vesicles and Interleukins: Novel Frontiers in Diagnostic and Therapeutic for Cancer », Front. Immunol., vol. 13, p. 836922, mars 2022. [CrossRef]
- J. Li et al., « Exosomes mediate the cell-to-cell transmission of IFN-α-induced antiviral activity », Nat. Immunol., vol. 14, no 8, p. 793-803, août 2013. [CrossRef]
- A. K. Khatua, H. E. Taylor, J. E. K. Hildreth, et W. Popik, « Exosomes Packaging APOBEC3G Confer Human Immunodeficiency Virus Resistance to Recipient Cells », J. Virol., vol. 83, no 2, p. 512-521, janv. 2009. [CrossRef]
- X. Zhu et al., « IFITM3-containing exosome as a novel mediator for anti-viral response in dengue virus infection: Antiviral effect of IFITM3 mediated by exosome », Cell. Microbiol., vol. 17, no 1, p. 105-118, janv. 2015. [CrossRef]
- A. A. Rana et al., « Poly(I:C) induces controlled release of IL-36γ from keratinocytes in the absence of cell death », Immunol. Res., vol. 63, no 1-3, p. 228-235, déc. 2015. [CrossRef]
- L. Sun, X. Wang, Y. Zhou, R.-H. Zhou, W.-Z. Ho, et J.-L. Li, « Exosomes contribute to the transmission of anti-HIV activity from TLR3-activated brain microvascular endothelial cells to macrophages », Antiviral Res., vol. 134, p. 167-171, oct. 2016. [CrossRef]
- M. L. Velandia-Romero et al., « Extracellular vesicles of U937 macrophage cell line infected with DENV-2 induce activation in endothelial cells EA.hy926 », PloS One, vol. 15, no 1, p. e0227030, 2020. [CrossRef]
- J. R. Edgar, P. T. Manna, S. Nishimura, G. Banting, et M. S. Robinson, « Tetherin is an exosomal tether », eLife, vol. 5, p. e17180, sept. 2016. [CrossRef]
- C. Villarroya-Beltri et al., « ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins », Nat. Commun., vol. 7, no 1, p. 13588, nov. 2016. [CrossRef]
- E. Delorme-Axford et al., « Human placental trophoblasts confer viral resistance to recipient cells », Proc. Natl. Acad. Sci., vol. 110, no 29, p. 12048-12053, juill. 2013. [CrossRef]
- T. Maemura, S. Fukuyama, et Y. Kawaoka, « High Levels of miR-483-3p Are Present in Serum Exosomes Upon Infection of Mice With Highly Pathogenic Avian Influenza Virus », Front. Microbiol., vol. 11, p. 144, févr. 2020. [CrossRef]
- A. L. Hodge, A. A. Baxter, et I. K. H. Poon, « Gift bags from the sentinel cells of the immune system: The diverse role of dendritic cell-derived extracellular vesicles », J. Leukoc. Biol., vol. 111, no 4, p. 903-920, mars 2022. [CrossRef]
- E. Segura, S. Amigorena, et C. Théry, « Mature dendritic cells secrete exosomes with strong ability to induce antigen-specific effector immune responses », Blood Cells. Mol. Dis., vol. 35, no 2, p. 89-93, sept. 2005. [CrossRef]
- M. F. S. Lindenbergh et al., « Bystander T-Cells Support Clonal T-Cell Activation by Controlling the Release of Dendritic Cell-Derived Immune-Stimulatory Extracellular Vesicles », Front. Immunol., vol. 10, p. 448, mars 2019. [CrossRef]
- M. F. S. Lindenbergh, R. Wubbolts, E. G. F. Borg, E. M. Van ’T Veld, M. Boes, et W. Stoorvogel, « Dendritic cells release exosomes together with phagocytosed pathogen; potential implications for the role of exosomes in antigen presentation », J. Extracell. Vesicles, vol. 9, no 1, p. 1798606, sept. 2020. [CrossRef]
- A. Montecalvo et al., « Exosomes As a Short-Range Mechanism to Spread Alloantigen between Dendritic Cells during T Cell Allorecognition », J. Immunol., vol. 180, no 5, p. 3081-3090, mars 2008. [CrossRef]
- Y. Matsuzaka et R. Yashiro, « Regulation of Extracellular Vesicle-Mediated Immune Responses against Antigen-Specific Presentation », Vaccines, vol. 10, no 10, p. 1691, oct. 2022. [CrossRef]
- M. R. Anderson et al., « Viral antigens detectable in CSF exosomes from patients with retrovirus associated neurologic disease: functional role of exosomes », Clin. Transl. Med., vol. 7, no 1, p. e24, déc. 2018. [CrossRef]
- E. Pesce et al., « Exosomes Recovered From the Plasma of COVID-19 Patients Expose SARS-CoV-2 Spike-Derived Fragments and Contribute to the Adaptive Immune Response », Front. Immunol., vol. 12, p. 785941, janv. 2022. [CrossRef]
- J. M. Reyes-Ruiz et al., « The Regulation of Flavivirus Infection by Hijacking Exosome-Mediated Cell–Cell Communication: New Insights on Virus–Host Interactions », Viruses, vol. 12, no 7, p. 765, juill. 2020. [CrossRef]
- A. De Gassart et al., « Exosomal sorting of the cytoplasmic domain of bovine leukemia virus TM Env protein », Cell Biol. Int., vol. 33, no 1, p. 36-48, janv. 2009. [CrossRef]
- J. L. Hood, « Post isolation modification of exosomes for nanomedicine applications », Nanomed., vol. 11, no 13, p. 1745-1756, juill. 2016. [CrossRef]
- V. Sengupta, S. Sengupta, A. Lazo, P. Woods, A. Nolan, et N. Bremer, « Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19 », Stem Cells Dev., vol. 29, no 12, p. 747-754, juin 2020. [CrossRef]
- C. Yang, Y. Xue, Y. Duan, C. Mao, et M. Wan, « Exosomes and their engineering strategies, delivery systems, and biomedical applications », J. Control. Release Off. J. Control. Release Soc., p. S0168-3659(23)00782-4, déc. 2023. [CrossRef]
- P. Gangadaran et al., « The emerging role of exosomes in innate immunity, diagnosis and therapy », Front. Immunol., vol. 13, p. 1085057, janv. 2023. [CrossRef]
- J. Rädler, D. Gupta, A. Zickler, et S. E. Andaloussi, « Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading », Mol. Ther., vol. 31, no 5, p. 1231-1250, mai 2023. [CrossRef]
- D. Jafari et al., « Designer Exosomes: A New Platform for Biotechnology Therapeutics », BioDrugs, vol. 34, no 5, p. 567-586, oct. 2020. [CrossRef]
- M. N. Huda et M. Nurunnabi, « Potential Application of Exosomes in Vaccine Development and Delivery », Pharm. Res., vol. 39, no 11, p. 2635-2671, nov. 2022. [CrossRef]
- J. S. Schorey, Y. Cheng, P. P. Singh, et V. L. Smith, « Exosomes and other extracellular vesicles in host–pathogen interactions », EMBO Rep., vol. 16, no 1, p. 24-43, janv. 2015. [CrossRef]
- J. Davod, D. N. Fatemeh, H. Honari, et R. Hosseini, « Constructing and transient expression of a gene cassette containing edible vaccine elements and shigellosis, anthrax and cholera recombinant antigens in tomato », Mol. Biol. Rep., vol. 45, no 6, p. 2237-2246, déc. 2018. [CrossRef]
- H. Vallhov et al., « Exosomes Containing Glycoprotein 350 Released by EBV-Transformed B Cells Selectively Target B Cells through CD21 and Block EBV Infection In Vitro », J. Immunol., vol. 186, no 1, p. 73-82, janv. 2011. [CrossRef]
- S. Anticoli, E. Falcone, A. Ruggieri, et M. Federico, « Engineered exosomes boost the HCV NS3-specific CD8+ T lymphocyte immunity in humans », Trials Vaccinol., vol. 5, p. 105-110, 2016. [CrossRef]
- S. Anticoli et al., « An Exosome-Based Vaccine Platform Imparts Cytotoxic T Lymphocyte Immunity Against Viral Antigens », Biotechnol. J., vol. 13, no 4, p. 1700443, avr. 2018. [CrossRef]
- S. Jesus, E. Soares, M. T. Cruz, et O. Borges, « Exosomes as adjuvants for the recombinant hepatitis B antigen: First report », Eur. J. Pharm. Biopharm., vol. 133, p. 1-11, déc. 2018. [CrossRef]
- J. M. Pitt et al., « Dendritic Cell–Derived Exosomes as Immunotherapies in the Fight against Cancer », J. Immunol., vol. 193, no 3, p. 1006-1011, août 2014. [CrossRef]
- R. Wang, Y. Xie, T. Zhao, X. Tan, J. Xu, et J. Xiang, « HIV-1 Gag-specific exosome-targeted T cell-based vaccine stimulates effector CTL responses leading to therapeutic and long-term immunity against Gag/HLA-A2-expressing B16 melanoma in transgenic HLA-A2 mice », Trials Vaccinol., vol. 3, p. 19-25, 2014. [CrossRef]
- Y. Matsuzaka et R. Yashiro, « Extracellular Vesicle-Based SARS-CoV-2 Vaccine », Vaccines, vol. 11, no 3, p. 539, févr. 2023. [CrossRef]
- M. Cacciottolo et al., « Exosome-Based Multivalent Vaccine: Achieving Potent Immunization, Broadened Reactivity, and Strong T-Cell Responses with Nanograms of Proteins », Microbiol. Spectr., vol. 11, no 3, p. e00503-23, juin 2023. [CrossRef]
- S. J. Tsai et al., « Exosome-mediated mRNA delivery in vivo is safe and can be used to induce SARS-CoV-2 immunity », J. Biol. Chem., vol. 297, no 5, p. 101266, nov. 2021. [CrossRef]
- K. D. Popowski et al., « Inhalable dry powder mRNA vaccines based on extracellular vesicles », Matter, vol. 5, no 9, p. 2960-2974, sept. 2022. [CrossRef]
- Q. Zhang et al., « Intraduodenal Delivery of Exosome-Loaded SARS-CoV-2 RBD mRNA Induces a Neutralizing Antibody Response in Mice », Vaccines, vol. 11, no 3, p. 673, mars 2023. [CrossRef]
- S. Kuate, J. Cinatl, H. W. Doerr, et K. Überla, « Exosomal vaccines containing the S protein of the SARS coronavirus induce high levels of neutralizing antibodies », Virology, vol. 362, no 1, p. 26-37, mai 2007. [CrossRef]
- V. Karn et al., « Extracellular Vesicle-Based Therapy for COVID-19: Promises, Challenges and Future Prospects », Biomedicines, vol. 9, no 10, p. 1373, oct. 2021. [CrossRef]
- Z. Wang et al., « Exosomes decorated with a recombinant SARS-CoV-2 receptor-binding domain as an inhalable COVID-19 vaccine », Nat. Biomed. Eng., vol. 6, no 7, p. 791-805, juill. 2022. [CrossRef]
- A. Schwab et al., « Extracellular vesicles from infected cells: potential for direct pathogenesis », Front. Microbiol., vol. 6, oct. 2015. [CrossRef]
- R. C. Lai, R. W. Y. Yeo, K. H. Tan, et S. K. Lim, « Exosomes for drug delivery — a novel application for the mesenchymal stem cell », Biotechnol. Adv., vol. 31, no 5, p. 543-551, sept. 2013. [CrossRef]
- M. Marcus et J. Leonard, « FedExosomes: Engineering Therapeutic Biological Nanoparticles that Truly Deliver », Pharmaceuticals, vol. 6, no 5, p. 659-680, avr. 2013. [CrossRef]
- L. Luketic et al., « Antigen Presentation by Exosomes Released from Peptide-Pulsed Dendritic Cells Is not Suppressed by the Presence of Active CTL », J. Immunol., vol. 179, no 8, p. 5024-5032, oct. 2007. [CrossRef]
Figure 1.
Insights into exosome biogenesis and composition. A. Exosomes Biogenesis. The early endosomes produced by the endocytic pathways can mature into multivesicular bodies (MVB). During their formation, the intraluminal vesicles (ILVs) are loaded with cytosolic material like proteins, lipids and nucleic acids. MVBs have two possible fates. They are either directed to lysosomes, enabling degradation of ILV constituents, in particular membrane components, or released by exocytosis, enabling externalization of ILVs, which are then called exosomes. Created with Biorender.com. B. Schematic representation of an exosome and its standard cargo. Exosomes are delimited by a phospholipid bilayer membrane containing proteins from the tetraspanin family, such as CD63, CD9 or CD81. The intercellular adhesion molecule 1 (ICAM-1) and major histocompatibility complex MHC I and/or II are detected in variable proportions in the membrane, depending on the emitting cells, and contribute to the biological or pathological functions of exosomes. The ALIX protein is identified within these vesicles, serving as one of the primary intravesicular markers. Additional intra-exosomal markers exist like the tumor susceptibility gene 101 (TSG101) which plays a critical role in exosome biogenesis and secretion. A variety of chaperones, including members of the heat shock protein 70 (HSP70) family, could use this unconventional secretion pathway. In addition to proteins, these vesicles can carry nucleic acids, including DNA, RNA such as miRNA. Adapted from [49].
Figure 1.
Insights into exosome biogenesis and composition. A. Exosomes Biogenesis. The early endosomes produced by the endocytic pathways can mature into multivesicular bodies (MVB). During their formation, the intraluminal vesicles (ILVs) are loaded with cytosolic material like proteins, lipids and nucleic acids. MVBs have two possible fates. They are either directed to lysosomes, enabling degradation of ILV constituents, in particular membrane components, or released by exocytosis, enabling externalization of ILVs, which are then called exosomes. Created with Biorender.com. B. Schematic representation of an exosome and its standard cargo. Exosomes are delimited by a phospholipid bilayer membrane containing proteins from the tetraspanin family, such as CD63, CD9 or CD81. The intercellular adhesion molecule 1 (ICAM-1) and major histocompatibility complex MHC I and/or II are detected in variable proportions in the membrane, depending on the emitting cells, and contribute to the biological or pathological functions of exosomes. The ALIX protein is identified within these vesicles, serving as one of the primary intravesicular markers. Additional intra-exosomal markers exist like the tumor susceptibility gene 101 (TSG101) which plays a critical role in exosome biogenesis and secretion. A variety of chaperones, including members of the heat shock protein 70 (HSP70) family, could use this unconventional secretion pathway. In addition to proteins, these vesicles can carry nucleic acids, including DNA, RNA such as miRNA. Adapted from [49].
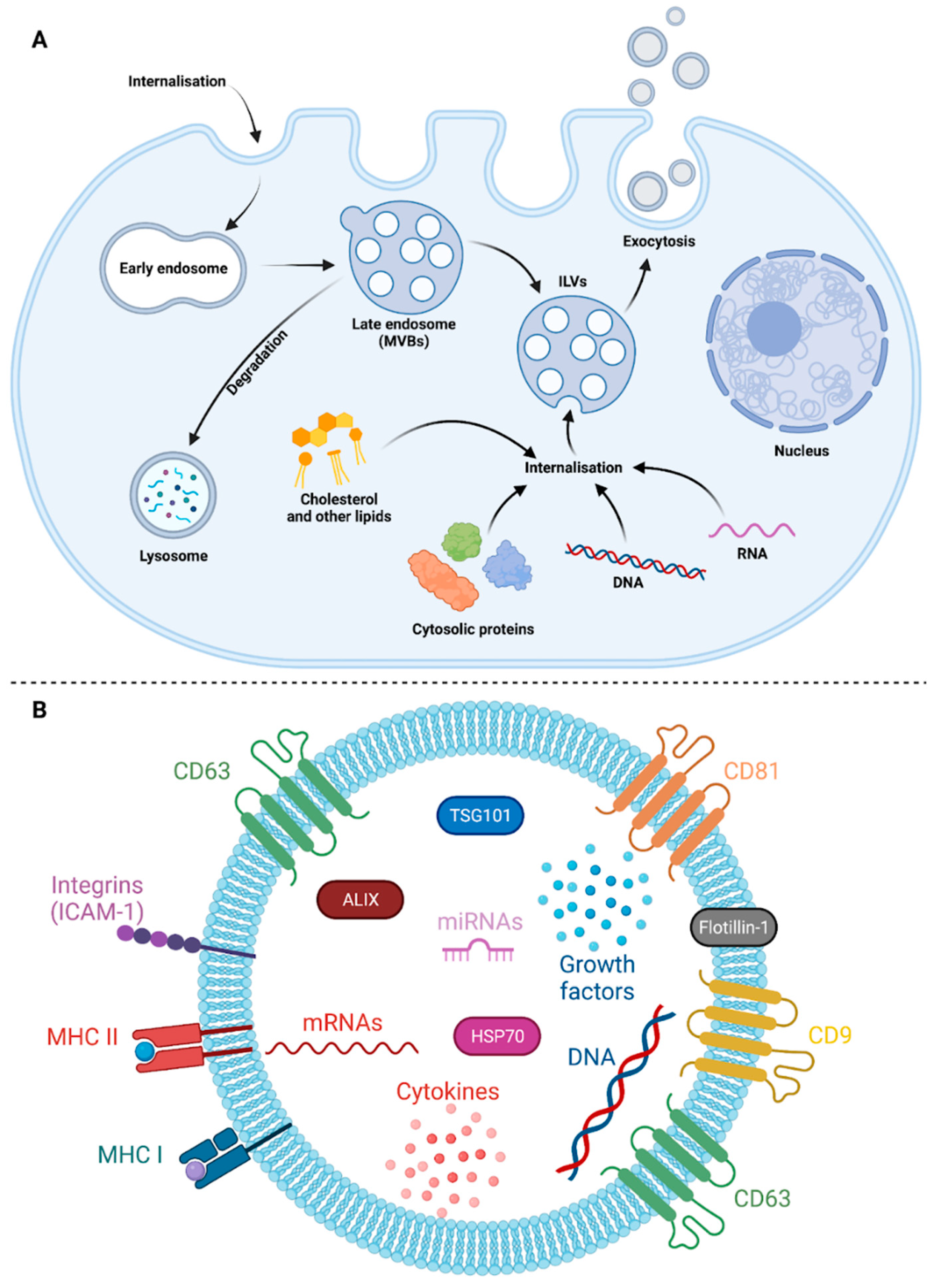
Figure 2.
Exosomes are double-edged swords during infection. On the one hand, exosomes may have detrimental effects during infection by promoting viral replication, impairing immune response and increasing susceptibility, ultimately leading to enhanced viral spreading. On the other hand, exosomes can participate in controlling infection through their immunomodulatory effects, including antigen presentation, delivery of cytokines and antiviral mediators, miRNA delivery and engagement of the stimulator of interferon genes. Created with Biorender.com.
Figure 2.
Exosomes are double-edged swords during infection. On the one hand, exosomes may have detrimental effects during infection by promoting viral replication, impairing immune response and increasing susceptibility, ultimately leading to enhanced viral spreading. On the other hand, exosomes can participate in controlling infection through their immunomodulatory effects, including antigen presentation, delivery of cytokines and antiviral mediators, miRNA delivery and engagement of the stimulator of interferon genes. Created with Biorender.com.
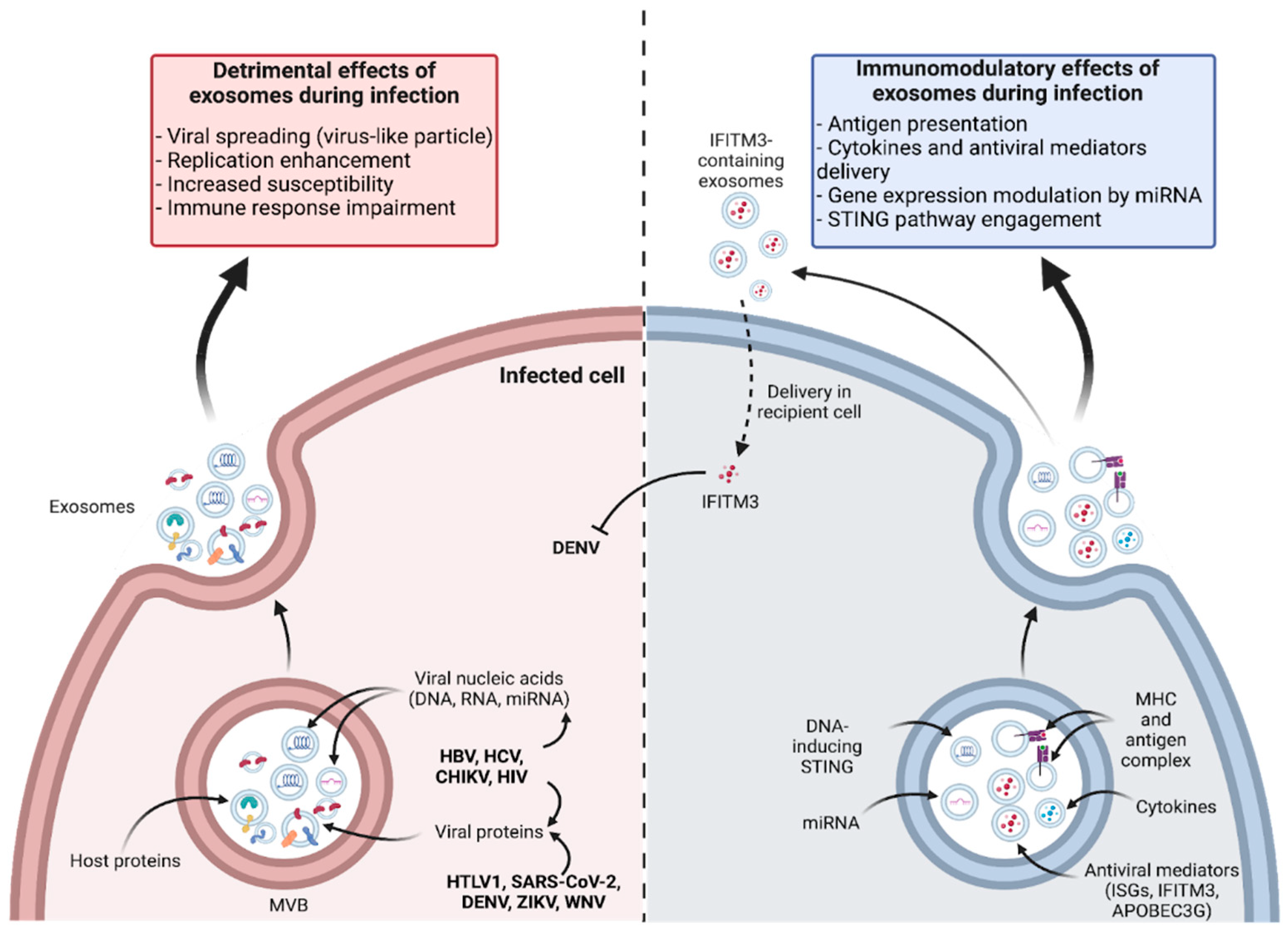
Figure 3.
Antigen presentation by exosomes. Having been primed with non-self antigens, dendritic cells may emit exosomes wherein the non-self antigens have been enclosed. Upon interaction with naïve dendritic cells, exosomes may deliver the non-self antigens, in a process called cross-priming: although the naive cells have never been directly exposed to them, they can now present the exosome-derived antigens to CD4+ T cells and CD8+ T cells. CD4+T cells will activate naïve B cells and induce clonal expansion and differentiation. Cellular immunity response will thus be set up leading to Th1 memory and Th2-like cytokines. A particularity of exosome and antigen presentation is that B-cell-derived exosomes can present allergen peptides and activate allergen-specific T cells.
Figure 3.
Antigen presentation by exosomes. Having been primed with non-self antigens, dendritic cells may emit exosomes wherein the non-self antigens have been enclosed. Upon interaction with naïve dendritic cells, exosomes may deliver the non-self antigens, in a process called cross-priming: although the naive cells have never been directly exposed to them, they can now present the exosome-derived antigens to CD4+ T cells and CD8+ T cells. CD4+T cells will activate naïve B cells and induce clonal expansion and differentiation. Cellular immunity response will thus be set up leading to Th1 memory and Th2-like cytokines. A particularity of exosome and antigen presentation is that B-cell-derived exosomes can present allergen peptides and activate allergen-specific T cells.
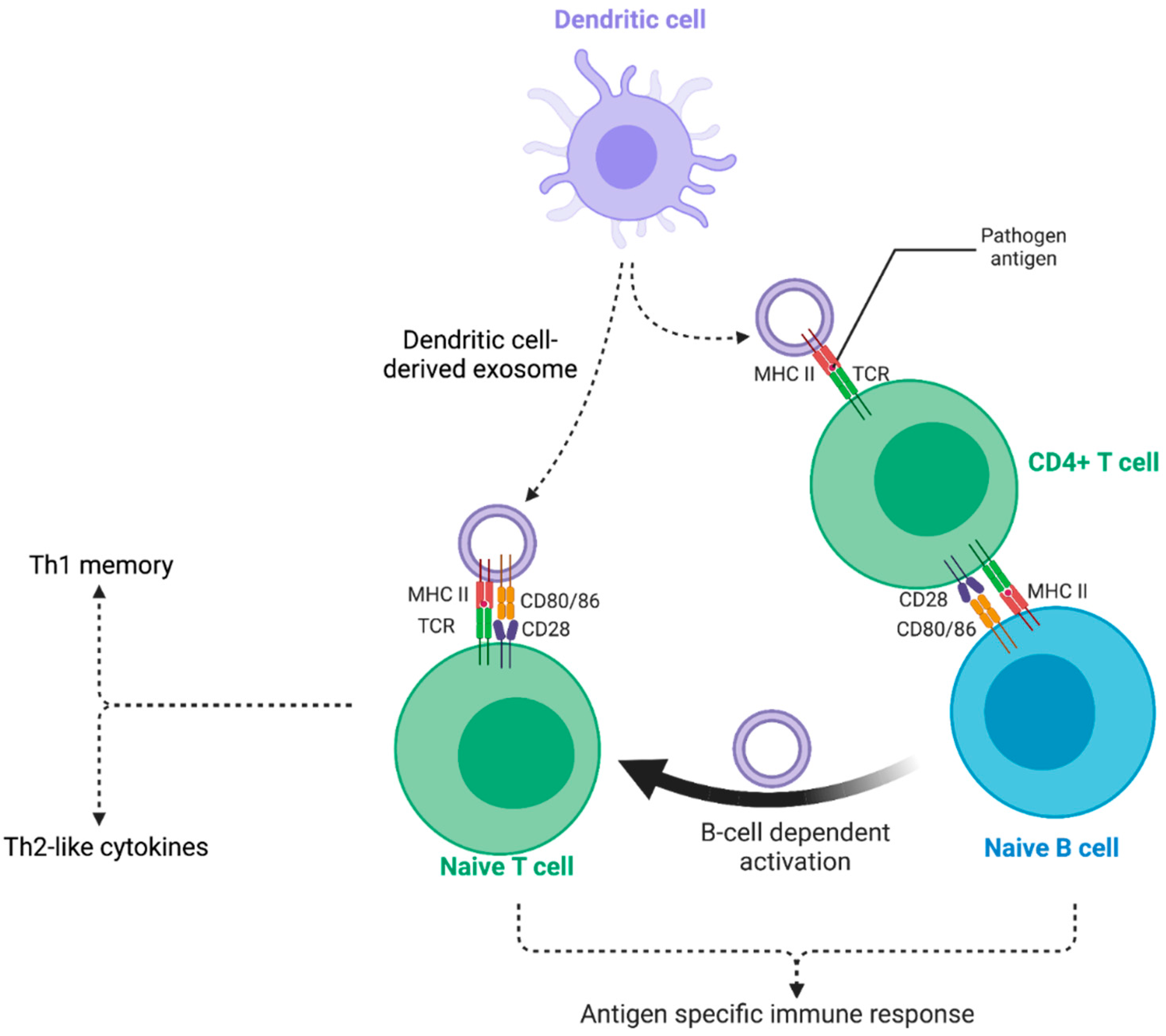
Figure 4.
Exosomes modified according to two strategies have been proposed as candidate vaccines against SARS-CoV-2. They are either designed to carry viral mRNA or engineered to expose a viral antigen, such as the Spike RBD. Both of these candidate vaccines demonstrated protection against SARS-CoV-2 infection in mice.
Figure 4.
Exosomes modified according to two strategies have been proposed as candidate vaccines against SARS-CoV-2. They are either designed to carry viral mRNA or engineered to expose a viral antigen, such as the Spike RBD. Both of these candidate vaccines demonstrated protection against SARS-CoV-2 infection in mice.
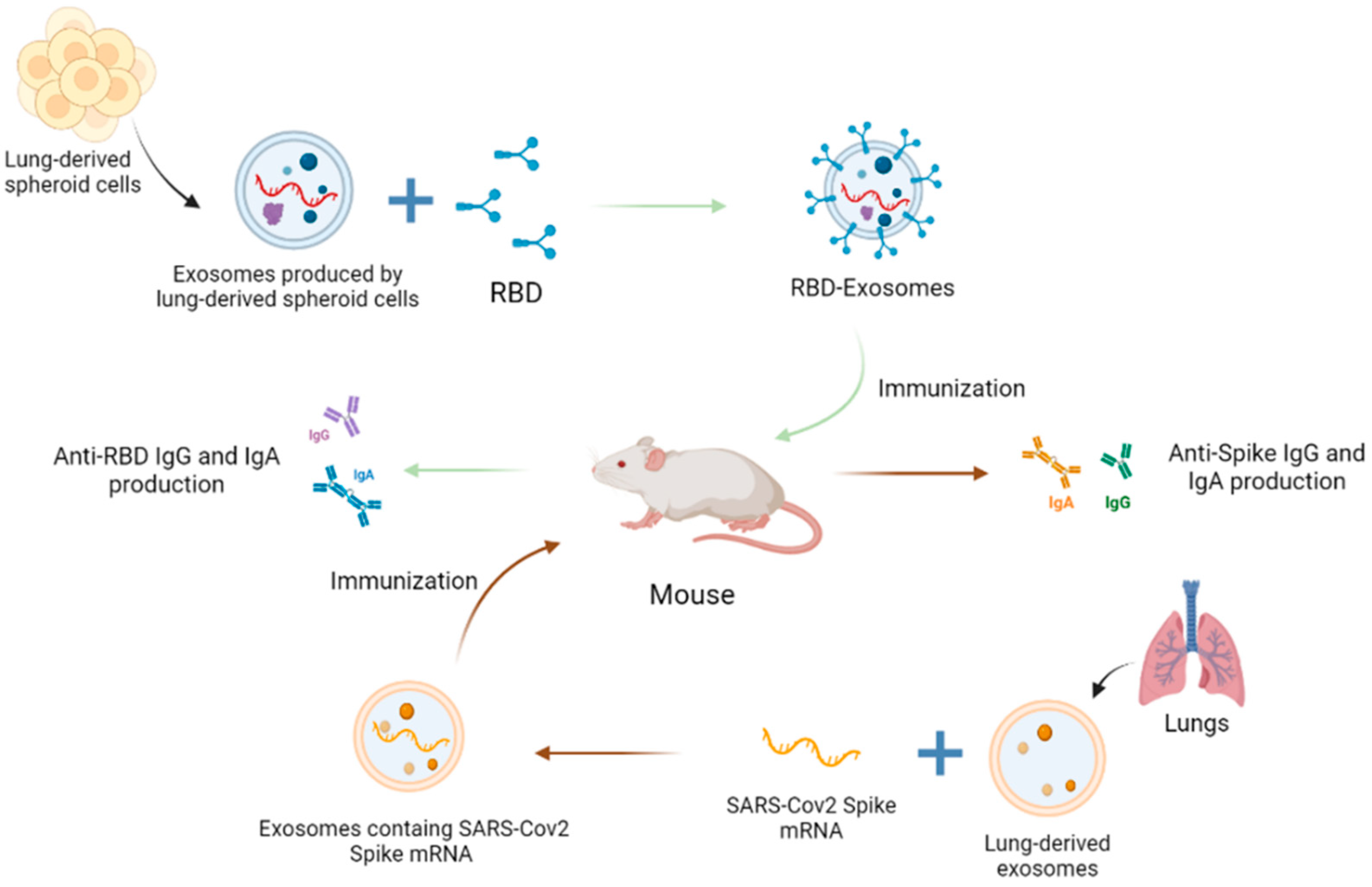
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



