
Submitted:
31 December 2023
Posted:
03 January 2024
You are already at the latest version
Abstract
Fibrosis represents a process characterized by excessive deposition of extracellular matrix (ECM) proteins. It often represents the evolution of pathological conditions, causes organ failure, and can, in extreme cases, compromises the functionality of the organs to the point of causing death. In recent years, considerable efforts have been made to understand the molecular mechanisms underlying fibrotic evolution and to identify possible therapeutic strategies. Great interest has been aroused by the discovery of a molecular association between epithelial to mesenchymal plasticity (EMP), in particular epithelial to mesenchymal transition (EMT), and fibrogenesis, which has led to the identification of complex molecular mechanisms closely interconnected with each other, which could explain EMT-dependent fibrosis. However, the result remains unsatisfactory from a therapeutic point of view. In recent years, advances in epigenetics, which is based on chromatin remodeling through various histone modifications or through the intervention of non-coding RNAs (ncRNAs), have provided more information on the fibrotic process, and this could represent a promising path forward for the identification of innovative therapeutic strategies for organ fibrosis. In this review, we summarize current research on epigenetic mechanisms involved in organ fibrosis, with a focus on epigenetic regulation of EMP/EMT-dependent fibrosis.
Keywords:
epigenetic
; inflammation
; fibrosis
; DNA methylation
; histone modification
; ncRNA
1. Introduction
Fibrosis, characterized by the deposition of connective tissue in a tissue or organ, represents a reaction to an injury and has reparative or pathological significance. The fibrotic evolution of a tissue or organ can have very negative consequences, leading to the inability to perform normal physiological functions and resulting in a pathological condition with high mortality [1,2,3,4]. Fibrosis is often associated with pathologies characterized by a chronic inflammatory state, such as autoimmune diseases or tumors. In these circumstances, the prolonged release of growth factors and/or pro-inflammatory factors such as transforming growth factor-β (TGF-β) or various cytokines mediate the activation of a cellular transformation process called epithelial-mesenchymal plasticity (EMP) [5]. When the EMP is activated, the epithelial cells, which have a phenotype of adherent cells closely connected to each other and not invasive, become transformed, assuming a hybrid epithelial/mesenchymal phenotype and/or a completely mesenchymal phenotype; in this case, the process is defined as epithelial-to-mesenchymal transition (EMT) [5]. These cells acquire much higher migratory capabilities and are able to deposit extracellular matrix (ECM) proteins. The triggering of various cascades of molecular, interconnected events leads to an exacerbation of the inflammatory state or to tumor proliferation and metastasis, with serious consequences [6,7,8]. Despite the fact that fibrosis appears to be a partly reversible process in various clinical studies [9], unfortunately, therapeutic options are still very limited. In recent years, very innovative studies have demonstrated how epigenetic modifications, by triggering or inhibiting gene transcription depending on the circumstances, can reprogram gene expression by adapting it to exposure to various risk factors [10]. This has been demonstrated, for example, in idiopathic pulmonary fibrosis (IPF) or in patients with non-alcoholic fatty liver disease, in which biopsy samples show higher expression of DNA methyltransferase, suggesting that DNA methylation could represent a predisposing factor for the onset of these pathologies [11]. The application of sequencing technology has demonstrated that the activation of fibroblasts, involved in collagen deposition during fibrogenesis, depends on various epigenetic modifications affecting the DNA to be transcribed [12]. Furthermore, epigenetic modifications appear to be largely involved in the modifications of epithelial cells towards the mesenchymal phenotype, a process essentially mediated by EMT [13,14].
Some authors have described in detail the mechanisms through which the main epigenetic modifications would act, inducing a regulation of the fibrotic evolution of the inflammatory processes and determining the transcription of pro-fibrotic genes [12]. In addition, epigenetics could explain the reversibility of the fibrosis [10]. However, although recent discoveries tend towards the involvement of epigenetic modifications in EMP/EMT-dependent fibrosis, the elucidation of the mechanisms involved still seems far from clear. This review aims to collect the latest discoveries made by studying the involvement of epigenetic modifications in the activation of EMP/EMT-dependent fibrosis, with the aim of suggesting new therapeutic perspectives.
2. Epigenetic mechanisms
Each phase of gene expression can undergo epigenetic modifications, thus leading to the synthesis or inhibition of certain downstream proteins. In the next paragraphs, we will analyze the main epigenetic modifications so that the reader can subsequently understand the role of epigenetics in EMP/EMT-dependent fibrosis regulation.
2.1. DNA methylation
Methylation of deoxyribonucleic acid (DNA) is the process of adding a methyl group to a nucleotide within the DNA. DNA methylation and demethylation represent heritable epigenetic marks that do not involve an alteration of the DNA sequence but can, however, determine a large modification of gene expression [15]); the methyl group is commonly donated by S-adenosyl-l-methionine (AdoMet) and occurs through the activity of the DNA methyltransferase (DNMT) family of enzymes [16]. The phenomenon of DNA methylation is an essential process for the physiological development of the individual and plays a key role in processes widely studied in recent years, such as genomic imprinting and inactivation of the elements. It follows that when DNA methylation is dysregulated, it can trigger the neoplastic transformation of cells [17]. The DNMT family comprises various elements: DNMT1, DNMT2, DNMT3A, DNMT3B, and DNMT3L [18]. The degree of DNA methylation affects gene expression, as methylation represents a means of inhibiting gene expression [19], which is restored following the reverse phenomenon, hypomethylation [20]. Gene repression is achieved through the activity of methyl-binding proteins such as methyl-Cap binding protein 2 (MeCP2) [21]; these proteins, through this domain, are able to bind to methylated CpG sequences [22]. Gene repression caused by methylation depends on several parameters, represented by the density of methylation, the frequent methylation gap of the CpG group within the gene, and also by the distance of the methylated group from the promoter region [23]. The methylation of the promoter region or gene has different effects; excess promoter methylation silences the gene, while a reduction in promoter methylation causes increased gene expression [24]. On the contrary, at the gene level, an excess of methylation causes active transcription of the gene itself, while the methylation of the gene has a meaning that is not yet known [25].
2.2. Histone modifications
Histone proteins are essential in stabilizing DNA by making DNA sites involved in gene transcription accessible. Wrapping DNA around histone proteins creates nucleosomes, each made up of about 150 base pairs of DNA sequence wrapped around a core of histone proteins [26]. Each nucleosome is made up of two repeating subunits containing four histones: H2A, H2B, H3, and H4 [26]. The role of the H1 protein is instead to stabilize the internucleosomal DNA without being part of the nucleosome itself. Histone proteins, through post-translational modifications, are able to determine the transition from open chromatin, called euchromatin, which is actively transcribed, to a tightly packed chromatin structure called heterochromatin. In this form, the DNA cannot be transcribed, resulting in gene silencing [27]. Through the modification of histones and the consequent variable interaction with DNA, a stringent control is carried out on gene transcription and activation [27].
Several post-translational modifications of histones have been identified, which include acetylation, methylation, phosphorylation, and ubiquitylation. These represent the best-known changes. In recent years, other histone modifications have been identified, such as GlcNAcylation, citrullination, krotonylation, and isomerization, which have yet to be studied in depth [28].
The following paragraphs are designed to explain the different mechanisms of the best-known histone modification.
2.2.1. Acetylation
Acetylation is one of the histone modifications identified several years ago and is best known. It influences transcriptional regulation through the neutralization of the positive charge present on the lysine residues of histone proteins. The neutralization of the charge makes the interactions between histones and chromatin less stable; therefore, binding with transcription factors becomes possible, allowing gene transcription [29].
In this way, histone acetylation intervenes in various mechanisms relating to cellular physiology, such as the regulation of the cell cycle, proliferation, and programmed cell death; furthermore, it appears to be responsible for or involved in many other cellular processes, including cell differentiation and DNA replication and repair [29]. Consequently, an imbalance in the mechanism of histone modification via acetylation is associated with tumorigenesis and dissemination via metastasis [30]. Acetyl groups are added to the lysine residues of histones H3 and H4 with consequent neutralization of the positive charge, thanks to the activity of histone acetyltransferases (HATs); the demethylation process, which leads to gene inactivation, occurs on the contrary through enzymes called deacetylases (HDAC). Histone acetylation was predominantly found in promoter regions and is associated with enhanced promoter activation [31]. Interestingly, low levels of acetylation are also found in all transcribed genes, but the function of this remains unclear to date.
2.2.2. Methylation
Methylation affects both the lysine and arginine residues of histones H3 and H4, with different outcomes on transcription. Arginine methylation promotes gene transcription, leading to the formation of euchromatin [32], while lysine methylation can have either a positive or negative impact on transcription depending on the site involved in methylation. This variability could depend on the fact that methylation does not cause an alteration of the histone charge nor directly influence histone-DNA interactions, unlike the mechanism of acetylation. In addition, more than one methyl group can be added to lysine residues, and this leads to further functional diversity depending on the site and mode of methylation. For example, both mono- and tri-methylation on K4 of histone H3 are an index of activation of gene transcription, but with substantial differences; in fact, the addition of a single methyl group typically identifies transcription enhancement, while the addition of three methyl groups to the lysine K4 residue of histone H3 identifies the promoter region [33]. To give an idea of the variability arising from methylation with more than one methyl group on gene activation, we report, for example, that tri-methylation of the lysine residue K36 on histone H3 is an activation marker associated with regions transcribed in the body of the genes. On the contrary, tri-methylation on residues K9 and K27 of histone H3 represents signals with a gene repression function. In particular, the tri-methylation of K9 and K27 has very particular outcomes, as the tri-methylation of K27 represents a temporary stop signal in the promoter of embryonic stem cell development genes, including the Hox and Sox genes [34]. On the contrary, the tri-methylation on the K9 residue is a permanent signal for heterochromatin formation in gene-poor, tandem repeat-rich chromosomal regions, such as satellite repeats or telomeres. This tri-methylation pattern on K9 also identifies retrotransposons and specific zinc finger [34] gene families. Further complicating this process of clarifying the meaning of methylation is the discovery that both tri-methylations of K9 and K27 are present on the inactive X chromosome; respectively, K9 tri-methylation is prevalent in the coding regions of active genes, while K27 tri-methylation is observed in intergenic and silenced coding regions [35]. The methylation process requires the activation of a specific set of protein enzymes with various substrates and cofactors. Methylation of an arginine residue requires arginine methyltransferase activity, while lysine requires a histone methyltransferase [36]. Recently, it has been discovered that histone methylation is also a reversible process through the action of histone demethylases [37].
2.2.3. Phosphorylation
The process leading to histone phosphorylation is a fundamental intermediate step in the condensation of chromosomes during cell division, in the regulation of gene transcription, and in the repair of DNA damage [38]. Unlike acetylation and methylation, histone phosphorylation has effects that depend on establishing interactions between histones that have undergone other modifications previously, as well as representing a necessary modification so that other effector proteins can be activated, triggering the activation of a transcriptional cascade. Phosphorylation affects all central histones, with different effects on each. Phosphorylation of serine residues 10 and 28 of histone H3 and serine residue T120 of histone H2A is involved in chromatin compaction during the phases of cell replication via mitosis. These are evolutionarily conserved cell cycle and cell growth markers across all eukaryotes. The phosphorylation of the S139 residue in histone H2A identifies, however, a docking point for the interaction with proteins involved in the repair of DNA damage [39]. Phosphorylation of histone H2B is not as well understood but appears to facilitate chromatin condensation during apoptosis, DNA fragmentation, and cell necrosis or physiological death [40].
2.2.4. Ubiquitylation
All histone core proteins can undergo a process of ubiquitination, but studies conducted over the years have shown that H2A and H2B are the proteins that are most frequently ubiquitinated in the nucleus [41]. Histone ubiquitylation appears to play a major role in DNA damage repair. In fact, the addition of a single ubiquitin residue at the level of the histone proteins H2A, H2B, and H2AX is found at the sites of DNA double-strand breaks. Overall, researchers believe that histone ubiquitination is associated with activation of gene expression, but it was demonstrated that the presence of a single ubiquitin, fundamental in the molecular event represented by the post-translational modification of histones, has different effects on H2A and H2B; in fact, monoubiquitylated H2A is also associated with gene silencing, while if the interaction with ubiquitin concerns H2B, transcription activation is triggered [41]. Polyubiquitination is less common but equally involved in DNA repair processes, providing a recognition site for proteins actively involved in repair processes. Like other histone modifications, the monoubiquitination of H2A and H2B is reversible and occurs thanks to the activation of ubiquitin ligase and reversed following binding with deubiquitinating enzymes [42].
2.3. Non-coding RNA
A noncoding RNA (ncRNA) is a functional RNA molecule that is regularly transcribed from DNA but is not used in the process of transduction or protein formation [43]. ncRNAs are actively involved in epigenetic processes, and it has been widely demonstrated that ncRNAs are able to regulate gene expression at both transcriptional and post-transcriptional levels. A classification of the ncRNAs involved in epigenetic modifications was carried out by identifying two broad categories: short ncRNAs, with a number of nucleotides less than 30, and long ncRNAs (lncRNAs), which include those RNAs with a number of nucleotides greater than 200 [43]. The three main classes of short noncoding RNAs are microRNAs (miRNAs), short interfering RNAs (siRNAs), and piwi-interacting RNAs (piRNAs). Both core groups have been shown to play a role in heterochromatin formation, histone modification, DNA methylation, and gene knockdown [44].
2.3.1. Short ncRNAs
MicroRNAs (miRNAs) generally work by binding to a specific target messenger RNA via a complementary sequence; this binding induces fragmentation and degradation of the mRNA, with consequent blocking of the translation phase. miRNAs and DNA methylation are the two epigenetic factors that have emerged in recent years as correlated in the regulation of gene expression [45]. Having highlighted a close correlation between these two epigenetic mechanisms, several researchers have investigated a possible role in pathological mechanisms. It emerged that miRNAs, by carrying out a negative regulation of gene expression, play a key role both in physiological conditions that require control over gene transcription, e.g., during development, and in pathological conditions such as tumorigenesis, diseases characterized by chronic inflammation, and psychiatric disorders [45]. Aberrant DNA methylation appears to be closely related to miRNAs in a feedback mechanism, manifesting in both general genome changes and localized changes at the gene promoter region, and, without a doubt, this represents a fundamental event in gene transcription implicated in the onset of cancer [46]. Mutual regulation of miRNAs and DNA methylation ultimately appears to occur in human tumors. MiRNAs regulate DNA methylation by acting on the transcription of genes involved in the synthesis of DNA methyltransferases, or methylation-related proteins. On the other hand, aberrant methylation of both miRNAs has often been found in human tumors [46]. Therefore, understanding the mechanisms underlying the mutual regulation of miRNAs and DNA methylation could lead to the identification of new therapeutic approaches efficient in counteracting the onset of cancer or its propagation.
The siRNAs function in a similar way as miRNAs to mediate post-transcriptional gene silencing as a result of mRNA degradation [47]. siRNAs represent small RNA molecules whose function is to prevent gene expression and subsequent protein synthesis. siRNAs repress the expression of a gene by binding to the mRNA and inducing its degradation. This mechanism is called RNA interference (RNAi), although it is worth remembering that siRNA-mediated RNAi is not the only type of RNAi. RNAi is perhaps one of the greatest promises in the field of gene therapy [47]. The idea is to block the expression of mutated and damaged genes in order to prevent the synthesis of harmful protein. siRNAs are obtained from a long double-stranded RNA molecule (double-stranded RNA, or dsRNA), which is cut into many small fragments (i.e., siRNAs) by the Dicer endoribonuclease. The siRNA thus obtained are added to the so-called RISC complex (RNA-induced silencing complex) to obtain the RISC-siRNA complex. In the initial, still inactive RISC-siRNA complex, the siRNAs are still double-stranded. Once activated, the siRNA loses one of the two strands and therefore becomes free to bind to the target messenger, i.e., the mRNA whose translation into protein is to be prevented [48]. By binding to the target mRNA, the RISC-siRNA complex induces its degradation. This inactivation process is highly specific, as even a single base of difference between the sequence of the mRNA and the siRNA prevents binding between the two [48].
The piRNAs are small RNAs that interact with the largely germline-specific Piwi subfamily of Argonaute proteins [49]. PIWI-interacting RNAs (piRNAs) are a new type of small noncoding RNAs (sncRNAs), which are 26–31 nucleotides in length and bind to PIWI proteins. Originally identified in Drosophila germ cells, piRNAs have also been identified in adult cells. PiRNAs have also recently been detected in human tumor cells [50], and it is currently believed that they can also influence gene expression in somatic cells. Their primary function is to silence transposon elements and maintain genome-wide stability [49,50]. The role of these molecules in driving the formation of heterochromatin that has no transcriptional activity is suggested by the finding that recruitment of Piwi to a reporter locus induces methylation of serine K9 in histone H3, which results in the inability to bind RNA polymerase II, directly driving chromatin changes [51]. Interestingly, some scholars have demonstrated that in C. elegans, H3K9 methylation is a transgenerational phenomenon that is conserved for at least two generations [52].
Currently, the molecular details underlying the function of piRNAs in the formation of heterochromatin have yet to be clarified, and, therefore, new developments are expected to better fit these small RNA molecules into the context of the most recent therapeutic research.
2.3.2. Long ncRNAs
Many lncRNAs act by forming complexes with proteins that modify the conformation of chromatin. In this way they expose and make available or cover catalytic sites in specific positions of the genome, influencing gene expression [53,54]. Most non-coding RNA transcription processes concern the lncRNA group. LncRNAs have various activities, now widely demonstrated, which concern chromatin remodeling, transcriptional and post-transcriptional regulation and act as precursors of siRNAs with the function of gene silencing [55,56]. LncRNA, seem to be involved also in the organization of protein complexes, cell-cell signaling interaction, and allosteric regulation of proteins structure [57]. A schematic overview of the epigenetic modification is reported in Figure 1.
3. The dynamic balance between EMT and EMP
EMT is a dynamic complex process during which epithelial cells reduce their epithelial properties gradually dissolving cell-cell junctions and rebuild cell-matrix connections acquiring characteristics typical of mesenchymal cells [58,59,60]. EMT was initially recognized to be ubiquitous throughout every aspect of life activity contributing to embryonal processes like gastrulation, neural crest formation, or heart development [58,61]. The program is also crucial for physiological processes like wound healing [62] and tissue homeostasis [62]. Importantly, pathological reactivation of the EMT process plays a fundamental role in diseases like organ fibrosis or cancer progression to metastasis. EMT is classified into three functional types: (a) type I which is linked to embryonic morphogenesis; (b) type II which is part of normal wound healing, though in chronic cases, enhanced myofibroblast activation coupled with the deposition of high levels of extracellular matrix proteins can lead to organ destruction via fibrosis; and (c) type III, which denotes malignant epithelial cells that acquire migratory phenotype, enabling them to invade and metastasize [63,64,65]. EMT is a phenomenon reversible, and the resulting cells shift back, from motile, multipolar mesenchymal types to polarized epithelial types via mesenchymal-epithelial transition (MET) process [66]. Therefore, until now, EMT was considered as an “all or nothing” program where the cells can exist in an epithelial morphology or a mesenchymal state. Interestingly, novel insights have shown that the cells that undergo to EMT present multiple intermediate phenotypes. This new concept, recently named as EMP, defines the capacity of the cells to interconvert between several states along the epithelial–mesenchymal spectrum thereby acquiring hybrid epithelial/mesenchymal phenotypic features [67,68]. Intriguingly, this cellular plasticity is very pliable, and epithelial cells often undergo partial remodelling and display mixed combination of epithelial and mesenchymal features during EMT [69] (Figure 2). Indeed, such cellular transformations and the resultant heterogeneity are believed to equip the cells of flexibility to face several physiological (embryonic development, wound healing) and pathological (organ fibrosis, cancer) conditions [70]. The dynamics of EMT/EMP and MET are regulated by a complex network of transcription factors (TFs) [71]. These mediators trigger within epithelial cells a transcription program in varied combinations to initiate EMT programs, orchestrating molecular interactions that result in a large number of intracellular cascades [72]. These changes in transcription, sometimes seen as gene reprogramming, involve three TFs families, Snail (Snail1) and Slug/Snail2, ZEB1 and ZEB2, and Twist [71,73]. All these TFs share the ability to repress epithelial genes like the E-cadherin encoding gene CDH1 via binding to E-Box motifs in their cognate promoter regions [58]. In parallel, the EMT-TFs, directly or indirectly, activate genes associated with a mesenchymal phenotype, including VIM (Vimentin), FN1 (Fibronectin), and CDH2 (N-cadherin) [58,71]. Upon induction of an epithelial plasticity response, they are considered as “master” drivers of the EMT program, conferring cellular shift among the epithelial-mesenchymal spectrum [69,72]. Interestingly, after expanding the knowledge and characterization of the EMT pathway, other than the signaling molecules regulating EMT, it becomes clear that activation and execution of EMT occur as a result of genetic and epigenetic processes. The study of epigenetic regulation is an important aspect of modulation of EMT [68,74] and different chromatin modifiers, non-coding RNAs, RNA splicing events, and DNA methylation/demethylation are major players in the epigenetic regulation of EMT [68]. Currently, there is a wealth of evidence in the literature supporting the role of chromatin remodeling in regulating EMT during tumor progression and metastasis. Similarly, recent discoveries have attributed a key role to epigenetic modifications also in the activation of the EMP process. Numerous evidences have demonstrated an altered expression of the main epigenetic modifications underlying the delicate balance between EMP and EMT, including histone modification, DNA methylation, and non-coding RNA, which could facilitate cancer metastasis [74].
4. Role of EMP/EMT in organ fibrosis
In recent years, our knowledge of the fibrotic process has been remarkably increased by the characterization of cellular mediators, key inflammatory and profibrogenic cytokines, molecular factors, and the evolution of new pathogenetic scenarios. A major determinant of fibrosis is the continuous spread of fibroblasts and myofibroblasts which suggests the question of how this cellular system can be fed [75]. Experiments conducted in the last 2 years have shown that cellular plasticity, which also includes the phenomenon of EMP, is not limited exclusively to development; it also characterizes cells that undergo reprogramming that occurs during the repair of tissue damage, during fibrotic processes, and during carcinogenesis [69]. However, knowledge of the molecular mechanisms involved in the cell's ability to modify its phenotype by evolving into another cell type is still at the beginning. The mechanisms underlying EMT are much more explored and known, and numerous studies have been conducted to evaluate the key role of EMT in fibrosis. In the context of identifying cellular drivers of fibrosis, various in vitro and in vivo studies have reported that EMT is a key mechanism during fibrogenesis, contributing substantially to the increase of interstitial fibroblasts and myofibroblasts and interrupting its progression can have a profound impact on the onset and progression of related diseases, particularly fibrosis [64,76,77]. Indeed, an intense build-up of fibrotic tissue can lead to chronic fibrotic diseases involving multiple organs, including the lung, liver, kidney, heart, skin, and salivary glands [78]. The following paragraph summarizes the most recent discoveries deriving from evaluating the molecular mechanisms underlying the process of EMT-dependent fibrosis in various organs in pathological conditions in order to identify potential therapeutic targets.
4.1. Contribution of epithelium to the fibrotic organ process via EMT activation
In the last few years, important findings have demonstrated that liver epithelial cells undergo the EMT process contributing to their transformation into myofibroblasts. Indeed, hepatocyte in which Snail1 gene was deleted by using Cre-loxP technology, showed a reduction of the EMT factors and a decrease in the severity of the inflammatory response compared to the control [79]. Moreover, Rowe et al. have examined a panel of genes known to contribute to the progression of liver fibrosis, including interstitial collagen types I and III and fibroblast markers, demonstrating that in the absence of Snail1, the liver expressed fewer profibrogenic gene levels such as Col1a1, Col2a1, Col3a1, vimentin, and FSP1 [79]. These data demonstrate that the hepatocyte Snail1 gene is a potent inducer in the progression of hepatic fibrosis. However, the involvement of hepatocytes and cholangiocytes in liver EMT is controversial and requires further study. Indeed, explorations have been performed to determine the origin of hepatic myofibroblasts activated in response to the type of liver injury. In particular, hepatic stellate cells (HSCs) are currently considered to be the main source of myofibroblasts after liver injury. In a mouse model subjected to hepatotoxic CCL4 liver injury, activated HSCs transformed almost totally into myofibroblasts, whereas the cholestatic bile duct ligation treatment stimulated preferentially portal fibroblasts [80,81]. Interestingly, in a quiescent state, HSCs expressed more epithelial factors than mesenchymal, but although HSCs are not strictly epithelial cells, they can undergo an EMT process in the presence of severe injurious stimulation [82].
Nowadays, it is experimentally proven that the activation of an EMT program occurs in a variety of pulmonary fibrosis diseases [83]. A study highlighted the contribution of the bronchial epithelial cells that, when treated with TGF-β1, are able to acquire myofibroblast phenotypes, and thereby lead to peribronchial fibrosis. This process involves airway epithelium remodeling and is a key event in the pathogenesis of asthma [84]. In idiopathic pulmonary fibrosis (IPF), alveolar epithelial cells (AECs) undergo an EMT program, inducing the formation of fibroblastic foci and thus triggering the fibrotic destruction of the lung architecture [85,86]. Interestingly, pleural mesothelial cells also undergo a special type of EMT, mesothelial-to-mesenchymal transition (MMT), during IPF pathogenesis. In the MMT process, the mesothelial cells, during serosal inflammation, acquire the mesenchymal phenotype and complete their transformation into myofibroblasts, thus contributing to the progression of parenchymal fibrosis that results in a progressive decline in lung function [87]. Indeed, an important report demonstrated the presence of pleural mesothelial cells exhibiting mesenchymal markers in the lung parenchyma of patients with IPF after fibrogenic stimulation in vivo and a correlation with disease severity and the degree of fibrosis [87].
By now, it has been widely accepted that tubular epithelial cells (TECs) undergoing an EMT program contribute to the development of severe fibrosis in chronic renal failure [88]. However, during the EMT process, the proportion of TECs that transform themselves into myofibroblasts is still uncertain. Actually, a new concept of partial EMT (pEMT) was proposed to explain the contribution of renal epithelial cells to renal fibrogenesis [76,89,90]. During the process of fibrosis, TECs seem to acquire some of the phenotypic characteristics of fibroblasts conserving also epithelial markers, rather than converting into a “fully fibroblastic phenotype” [76,89,90]. Therefore, TECs in this partial mesenchymal and epithelial phenotype remain attached to the basement membrane during the fibrotic process. Recently, it was demonstrated that Snail1 is able to trigger the pEMT process in TECs, relaying crucial signals to fibrogenic cytokine release and promoting differentiation into myofibroblasts, thus contributing to the exacerbation of the inflammatory response [89].
An increasing number of studies showed that EMT was also involved in inflammatory bowel disease (IBD), such as ulcerative colitis (UC) and Crohn’s disease (CD) [91]. In IBD patients, persistent intestinal inflammatory factors injure intestinal epithelial cells, determining reparative reactions that lead to the triggering of the EMT process and perpetuating a severe fibrotic condition [91,92]. Confirming this, the presence of high levels of tumor necrosis factor-like ligand 1A (TL1A) in the intestinal specimens of patients with UC and CD has been detected, which represents a potent inducer of EMT in intestinal fibrosis. As expected, the TGF-β1/Smad3 pathway may be involved in TL1A-induced EMT [92,93,94].
Recent discoveries have also shown that uncontrolled fibrosis is present in the heart and is triggered by EMT and its special type, endothelial to mesenchymal transition (EndMT) [33,95]. Epicardial EMT was activated after injury during pathological states such as myocardial infarction, atherosclerosis, and valve dysfunction to initiate angiogenesis and healing [96]. Under TGF-β stimuli, cardiac fibroblasts transdifferentiate into myofibroblasts, acquiring a phenotype similar to that of smooth muscle cells; furthermore, fibroblasts can also originate from endothelial cells through EndMT, giving rise to the progression of cardiac fibrosis [97]. In addition, TGF-β-driven EMT responsible for cardiac fibroblast formation appears to be triggered by the Hippo pathway, an evolutionarily conserved kinase cascade [98], as seen in a recent in vitro finding [99]. In addition, the process of cardiac fibrosis seems to be regulated by the C-Ski protein, identified as an inhibitory regulator of TGFβ signaling [100].
A recent research field focused on the demonstration of an intense fibrotic process in the salivary glands (SGs) derived from Sjögren’s Syndrome (SS) patients; fibrogenesis observed in SGs can be considered the end result of chronic, intense inflammatory reactions induced by a variety of stimuli in this autoimmune disease [7,8,101,102]; these pathological conditions lead to glandular failure and constitute risk factors for initial cell transformation and for cancer progression. Interesting research has evidenced that the SS patients with an intense fibrosis in SGs showed positively correlation between stimulated saliva flow and the degree of fibrosis, whereas the patients with higher fibrotic indices tended to have a greater decrease in saliva flow [101]. In all cases, SGs fibrosis is linked with an evident impairment of organ function that leads to progressive atrophy and a decrease in quality of life for patients [103].
In this context, using technology to create transgenic mice that conditionally overexpress active TGF-β1, experimental data have confirmed that the overexpression of active TGF-β1 leads to an abnormal accumulation of ECM proteins and severe hyposalivation and acinar atrophy in the mutated mice [104]. More recently, studies have demonstrated an exuberant upregulation of TGF-β1 in SS SGs that induces the loss of epithelial features and the acquisition of mesenchymal features in SG epithelial cells via the trigger of the TGF-β1/Smad/Snail signaling pathway [105]. Indeed, TGF-β1 regulates EMT through two main pathways: the canonical Smad-dependent pathway and a non-Smad signaling pathway [7,8,105]. Recent investigations have also demonstrated that TGF-β1 can mediate the non-Smad pathway thereby leading to the development of a pathological fibrotic state [7,8,105].
5. Epigenetics regulation of EMP/EMT-dependent fibrosis
The studies conducted in order to evaluate a possible connection between epigenetics, EMP, and fibrosis are leading to the first discoveries, made, for example, in airway persistent inflammation [106], but require further investigation. On the contrary, the field of studies conducted on the role of epigenetic modifications in EMT-dependent fibrosis is much more flourishing. One of the hot topics of the last few years has been the association between epigenetic regulation and fibrotic processes triggered by EMT [13]. Epigenetic modulation of tissue-stromal crosstalk involves several types of alterations, including DNA methylation, histone modifications, and non-coding RNA, all of which have been shown to play crucial roles in the drive EMT process [13]. The following paragraphs illustrate the current knowledge concerning the epigenetic aberrations involved in the fibrotic evolution induced by EMT during pathological processes.
5.1. DNA methylation in EMT-dependent fibrosis
DNA methylation, catalyzed by DNMTs, represents one of the best-characterized mechanisms of epigenetic gene modulation and regulation under the control of the EMT [107]. Altered transcription of EMT-related genes in accordance with the dynamic changes of DNA methylation, reinforcing the role of epigenetic mechanisms in the development of EMT [74]. Interesting evidence suggests that DNA methylation is the main regulator of CDH1, with DNMTs mediating methylation and the ten-eleven translocation (TETs) family becoming involved in subsequent demethylation [74]. Notably, altered methylation of the CDH1 promoter leads to diminished E-cadherin protein expression in numerous fibrotic conditions [108,109]. Direct methylation of transcription factors by DNMT1 concurs with the progression of EMT in renal epithelial cells, demonstrating a link with fibrogenesis in the kidneys [110]. Indeed, in arsenic-triggered renal fibrosis, treatment with a DNA methylation inhibitor reversed the EMT findings obtained by the use of the human kidney cell line, suggesting that the fibrotic modifications are induced by DNA methylation [111].
Therefore, interesting studies have evidenced the essential role of the DNMTs in cardiac fibrosis, resulting in the activation of the EndMT process. The inhibition of cytokine signaling 3 (SOCS3) mediated by DNMT1 determines the downregulation of STAT3 that induces cardiac fibroblast activation and collagen accumulation in cardiac fibrosis in diabetic cardiomyopathy [112,113]. The deregulation of SOCS3 triggers the transdifferentiation of the cardiac fibroblasts into myofibroblasts related to STAT3, probably induced by EndMT [112].
The transcriptional regulation driven by the DNA methylation pattern has a pivotal role in liver fibrosis [10]. In particular, DNA methylation is involved in the differentiation of HSC during hepatic diseases characterized by a severe and progressive fibrotic process. These data were confirmed downregulating DNMT3a and DNMT3b gene expression by the use of siRNA; in this case, DNA methylation was decreased, and HSC activation was subsequently suppressed [114]. Beyond DNMTs, another intrigant protein implicated in liver DNA methylation processes is glycine N-methyltransferase (GNMT). GNMT is the most abundant methyltransferase in the liver and hepatocytes. GNMT influences epigenetic regulatory determinants by competing with DNMT to regulate transmethylation flux [114].
Several studies have mentioned altered epigenetic modulations linked to the evolution of chronic respiratory diseases, such as chronic obstructive pulmonary disease and lung fibrosis [113,115]. Indeed, DNA methylation modifications have been widely demonstrated in the pathogenesis of pulmonary fibrosis. DNA methylation interacts with histone modifications and miRNA to induce or silence gene expression, determining pathological fibrotic progression [113,115] (Figure 3).
5.2. The involvement of histone modifications in EMT-dependent fibrosis
Histone modifications were linked with the dysregulation of EMT, leading to fibrotic processes [116]. Histone acetylation and deacetylation are histone modifications that have been extensively studied [29] and have recently linked to the activation of the EMT program in cancer and fibrosis. In fact, cancers with stem cell properties disseminate and form distant metastases by reactivating the developmental EMT program. Recent studies have demonstrated that epigenetic histone H2BK5 acetylation is important in the regulation of EMT [14]. For example, in trophoblast stem cells, H2BK5 acetylated promotes the expression of key genes involved in the maintenance of an epithelial phenotype. These trophoblast stem cells share similar H2BK5 acetylated-regulated gene expression when compared with stem-like claudin-low breast cancer cells, thus linking EMT-dependent development and EMT observed in cancer cells [14]. In particular, HATs and HDACs control gene silencing and activation through the modulation of the acetylation status of histones, thus orchestrating gene expression to induce liver fibrosis [116,117]. HDACs, firstly identified in liver fibrosis by de Ruijter et al., can enhance the cellular migration and ECM deposition by tissue myofibroblasts [118,119]. In liver fibrosis, the addition and removal of the methyl group contribute to hepatic fibrogenesis via EMT [114].
HDACs are widely studied in the pathogenesis of IPF as novel therapeutic targets for drug development. Epigenetic histone modifications through deacetylation could explain the persistently activated state of IPF fibroblasts [120], which indicated a “cancer-like” upregulation. According to this hypothesis, almost all Class I and Class II HDACs enzymes resulted overexpressed and this could be responsible of the abnormal repression of pro-apoptotic genes [120,121]. Supporting the role of histone acetylation in EMT-dependent fibrotic diseases, HDAC7 has also been demonstrated to regulate collagen deposition and other ECM protein accumulation in fibroblasts derived from patients affected by systemic sclerosis, and indeed, siRNA-mediated depletion of HDAC7 reduced ECM production in these cells, resulting in an evident decrease in fibrogenesis [10]. Therefore, histone modification may also be a key determinant of fibrotic progression in renal diseases. An interesting study has demonstrated that HDAC8 leads to the development of renal fibrosis through the activation of partial EMT [122]. Recent studies show that inhibition of HDACs activity decreases the activation process of renal fibroblasts and interstitial fibrosis in obstructive nephropathy [123]. Additionally, histone acetylation has been implied in pulmonary and cardiac fibrosis [114], performed through the activity of p300 HAT [124,125]. An interesting discovery enriches the molecular scenario by demonstrating that p300 HAT induces the fibrotic process in IPF and cardiac fibrosis via EMT and EndMT, respectively [126] (Figure 3).
5.3. Epigenetic involvement of ncRNAs in EMT-related fibrosis
NcRNAs were also recently recognized as potential key inductors of fibrogenesis [127]. Noncoding RNAs are micro-sequences of RNA transcribed but not translated into proteins and are grouped into short ncRNAs and long ncRNAs [128,129]. Increasing evidence has highlighted the involvement of noncoding RNAs in the EMT process. Several ncRNAs are able to modulate multiple transcription factors, such as Snail, and ZEB families, as drivers of the epithelial plasticity leading to the EMT process [130]. In particular, miR-200, recognized as a master regulator of epithelial phenotype, targets two key transcriptional suppressors of the cell adherence and cell polarity genes, ZEB1 and ZEB2 [131,132]. Adding new data, ncRNA have recently gained attention for their biological importance and involvement in the regulation of various conditions characterized by severe fibrosis. LncRNAs and miRNAs, for example, have been widely investigated in the context of pulmonary fibrosis [133]. Liu and colleagues [133] discovered that lung epithelial cells undergoing the EMT process and exhibit increased levels of lncRNA. In addition, current evidence has underlined the involvement of lncRNAs in the pathologic process of kidney fibrosis [134]. A growing number of experimental studies demonstrated that lncRNAs highly contribute to the development of the fibrotic process in the kidney, and several lncRNAs are linked to different cell types in various phases of kidney disease [134,135]. ncRNAs that regulate the EMT program were correlated with the pathological progress of liver diseases, contributing to the development of fibrosis, and many effective ncRNAs have been identified as possible therapeutic targets for fibrosis and for the diagnosis of liver disease [136]. In this context, ncRNA modulates the EMT process in hepatic cells that are undergoing the transformation process into myofibroblasts, resulting in severe liver fibrosis [136]. In cardiac fibrosis, accumulating studies have elucidated the regulatory role of ncRNAs on gene transcription. Recent investigations demonstrated that the expression of ncRNAs, and, in particular, lncRNAs, was strongly correlated with the onset, progression and treatment of cardiac fibrosis [127,137,138] (Figure 3).
6. Conclusion and future perspective
This article presents a comprehensive summary of the role of epigenetics in regulating EMP/EMT-dependent fibrosis and on its impact on several chronic inflammatory diseases. The purpose is to arouse growing interest in the scientific world and clarify the molecular mechanisms underlying these regulations of gene expression. This could identify factors predisposing to the onset of diseases related to the triggering of the EMP/EMT program, such as cancer or inflammatory diseases. After decades of studies, EMT is now considered a key physiological process active in embryogenesis and a pathological mechanism triggered in fibrotic diseases and cancer. Furthermore, recently, the meaning of EMT has also been revised by scientists, who have identified various degrees of cellular transformation towards a mesenchymal phenotype, so much so that they have coined the term EMP, underscoring cellular plasticity and the ability of an epithelial cell to assume intermediate phenotypes. Recent studies have unveiled the role of epigenetics in the control of EMP/EMT dynamics and, in general in cellular plasticity. Several epigenetic modifications have been identified capable of modifying gene transcription during the process of EMP/EMT related to fibrogenesis, adding even more complexity to the process of EMP/EMT-dependent fibrosis, which is finely regulated by various molecularly interconnected mechanisms. We hope that having summarized in this review the most recent discoveries in this very innovative field will provide new perspectives on molecular aspects and therapeutic approaches intended to regulate and make reversible the EMP/EMT-dependent fibrosis process.
Author Contributions
all authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version for publication. M.S. and S.L. had full access to the data collected in the review, take responsibility for their integrity and performed a critical reading. All authors have read and agreed to the published version of the manuscript.
Funding
this research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
the authors declare no conflict of interest.
References
- Antar, S.A.; Ashour, N.A.; Marawan, M.E.; Al-Karmalawy, A.A. Fibrosis: Types, Effects, Markers, Mechanisms for Disease Progression, and Its Relation with Oxidative Stress, Immunity, and Inflammation. Int. J. Mol. Sci. 2023, 24, 4004. [Google Scholar] [CrossRef] [PubMed]
- Mehal, W.Z.; Iredale, J.; Friedman, S.L. Scraping fibrosis: expressway to the core of fibrosis. Nat. Med. 2011, 17, 552–553. [Google Scholar]
- Wick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.F.; Feichtinger,J. ; Zelger, B.; Sgonc, R.; Wolfram, D. The immunology of fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Enderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: from mechanisms to medicines. Nature. 2020, 587, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Subhadarshini, S.; Markus, J.; Sahoo, S.; Jolly, M.K. Dynamics of Epithelial-Mesenchymal Plasticity: What Have Single-Cell Investigations Elucidated So Far? ACS Omega. 2023, 8, 11665–11673. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Wu, X.Q.; Zhang, D.D.; Wang, Y.N.; Guo, Y.; Li, P.; Xiong, Q.; Zhao, Y.Y. Deciphering the cellular mechanisms underlying fibrosis-associated diseases and therapeutic avenues. Pharmacol. Res. 2021, 163, 105316. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Ribatti, D.; Lisi, S. Organ Fibrosis and Autoimmunity: The Role of Inflammation in TGFβ-Dependent EMT. Biomolecules. 2021, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Lisi, S. Towards a Unified Approach in Autoimmune Fibrotic Signalling Pathways. Int. J. Mol. Sci. 2023, 24, 9060. [Google Scholar] [CrossRef]
- Povero, D.; Busletta, C.; Novo, E.; di Bonzo, L.V.; Cannito, S.; Paternostro, C.; Parola, M. Liver fibrosis: a dynamic and potentially reversible process. Histol. Histopathol. 2010, 25, 1075–1091. [Google Scholar]
- Liu, Y.; Wen, D.; Ho, C.; Yu, L.; Zheng, D.; O'Reilly, S.; Gao, Y.; Li, Q.; Zhang, Y. Epigenetics as a versatile regulator of fibrosis. J. Transl. Med. 2023, 21, 164. [Google Scholar] [CrossRef]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: from mechanisms to medicines. Nature. 2020, 587, 555–566. [Google Scholar] [CrossRef]
- Xue, T.; Qiu, X.; Liu, H.; Gan, C.; Tan, Z.; Xie, Y.; Wang, Y.; Ye, T. Epigenetic regulation in fibrosis progress. Pharmacol Res. 2021, 173, 105910. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Song, K.; Tu, B.; Sun, H.; Zhou, Y.; Xu, S.S.; Lu, D.; Sha, J.M.; Tao, H. New aspects of the epigenetic regulation of EMT related to pulmonary fibrosis. Eur. J. Pharmacol. 2023, 956, 175959. [Google Scholar] [CrossRef] [PubMed]
- Mobley, R.J.; Abell, A.N. Controlling Epithelial to Mesenchymal Transition through Acetylation of Histone H2BK5. J. Nat. Sci. 2017, 3, e432. [Google Scholar] [PubMed]
- Ciechomska, M.; O'Reilly, S. Epigenetic Modulation as a Therapeutic Prospect for Treatment of Autoimmune Rheumatic Diseases. Mediators Inflamm. 2016, 2016, 9607946. [Google Scholar] [CrossRef] [PubMed]
- Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, S.; Van Emburgh, B.O.; Robertson, K.D. DNA methylation in development and human disease. Mutat. Res. 2008, 647, 30–38. [Google Scholar] [CrossRef]
- Del Castillo Falconi, V.M.; Torres-Arciga, K.; Matus-Ortega, G.; Díaz-Chávez, J.; Herrera, L.A. DNA Methyltransferases: From Evolution to Clinical Applications. Int. J. Mol. Sci. 2022, 23, 8994. [Google Scholar] [CrossRef] [PubMed]
- Dowson, C.; O'Reilly, S. DNA methylation in fibrosis. Eur. J. Cell Biol. 2016, 95, 323–330. [Google Scholar] [CrossRef]
- Karouzakis, E.; Gay, R.E.; Michel, B.A.; Gay, S.; Neidhart, M. DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheum. 2009, 60, 3613–3622. [Google Scholar]
- Nan, X.; Ng, H.H.; Johnson, C.A.; Laherty, C.D.; Turner, B.M.; Eisenman, R.N.; Bird, A. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature. 1998, 393, 386–389. [Google Scholar] [CrossRef]
- Hendrich, B.; Tweedie, S. The methyl-CpG binding domain and the evolving role of DNA methylation in animals. Trends Genet. 2003, 19, 269–277. [Google Scholar] [CrossRef]
- Bian, E.B.; Huang, C.; Wang, H.; Chen, X.X.; Tao, H.; Zhang, L.; Lv, X.W.; Li, J. The role of methyl-CpG binding protein 2 in liver fibrosis. Toxicology. 2013, 309, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; He, L.; Yuan, H.; Jin, Y.; Yao, Y. Association between RUNX3 promoter methylation and non-small cell lung cancer: a meta-analysis. J. Thorac. Dis. 2014, 6, 694–705. [Google Scholar] [PubMed]
- Zhang, X.; Hu, M.; Lyu, X.; Li, C.; Thannickal, V.J.; Sanders, Y.Y. DNA methylation regulated gene expression in organ fibrosis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2389–2397. [Google Scholar] [CrossRef]
- Onufriev, A.V.; Schiessel, H. The nucleosome: from structure to function through physics. Curr. Opin. Struct. Biol. 2019, 56, 119–130. [Google Scholar] [CrossRef]
- Hergeth, S.P.; Schneider, R. The H1 linker histones: multifunctional proteins beyond the nucleosomal core particle. EMBO Rep. 2015, 16, 1439–1453. [Google Scholar] [CrossRef]
- Cavalieri, V. The Expanding Constellation of Histone Post-Translational Modifications in the Epigenetic Landscape. Genes 2021, 12, 1596. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.Y. , Denu, J.M., Allis, C.D. Histone acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef]
- Audia, J.E. , Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb Perspect Biol. 2016, 8, a019521. [Google Scholar] [CrossRef]
- Berndsen, C.B.; Denu, J.M. Catalysis and substrate selection by histone/protein lysine acetyltransferases. Curr. Opin. Struct. Biol. 2008, 18, 682–689. [Google Scholar] [CrossRef]
- Ng, S.S.; Yue, W.W.; Oppermann, U.; Klose, R.J. Dynamic protein methylation in chromatin biology. Cell Mol. Life Sci. 2009, 66, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Jiang, J.; Han, Y.; Shi, M.; Li, X.; Wang, Y.; Dong, Z.; Yang, C. Functional Characterization of the Lysine-Specific Histone Demethylases Family in Soybean. Plants (Basel). 2022, 11, 1398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ramlee, M.K.; Brunmeir, R.; Villanueva, C.J.; Halperin, D.; Xu, F. Dynamic and distinct histone modifications modulate the expression of key adipogenesis regulatory genes. Cell Cycle. 2012, 11, 4310–4322. [Google Scholar] [CrossRef] [PubMed]
- Rougeulle, C.; Chaumeil, J.; Sarma, K.; Allis, C.D.; Reinberg, D.; Avner, P.; Heard, E. Differential histone H3 Lys-9 and Lys-27 methylation profiles on the X chromosome. Mol. Cell Biol. 2004, 24, 5475–5484. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Litt, M.; Felsenfeld, G. Methylation of histone H4 by arginine methyltransferase PRMT1 is essential in vivo for many subsequent histone modifications. Genes Dev. 2005, 19, 1885–1893. [Google Scholar] [CrossRef] [PubMed]
- Greer, E.L.; Shi, Y. Histone methylation: a dynamic mark in health, disease and inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Kschonsak, M.; Haering, C.H. Shaping mitotic chromosomes: From classical concepts to molecular mechanisms. Bioessays. 2015, 37, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Lowndes, N.F.; Toh, G.W. DNA repair: the importance of phosphorylating histone H2AX. Curr Biol. 2005, 15, R99–R102. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.T.; Lee, S.Y.; Xu, Y.M.; Zheng, D.; Cho, Y.Y.; Zhu, F.; Kim, H.G.; Li, S.Q.; Zhang, Z.; Bode, A.M.; Dong, Z. Phosphorylation of histone H2B serine 32 is linked to cell transformation. J. Biol. Chem. 2011, 286, 26628–26637. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yan, Q. Histone ubiquitination and deubiquitination in transcription, DNA damage response, and cancer. Front. Oncol. 2012, 2, 26. [Google Scholar] [CrossRef]
- Segala, G.; Picard, D. H2B monoubiquitination: t'ub or not t'ub for inducible enhancers. Transcription. 2017, 8, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Kaikkonen, M.U.; Lam, M.T.; Glass, C.K. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell. 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, M.; Mohan, M. MicroRNAs: history, biogenesis, and their evolving role in animal development and disease. Vet. Pathol. 2014, 54, 759–774. [Google Scholar] [CrossRef]
- Wang, S.; Wu, W.; Claret, F.X. Mutual regulation of microRNAs and DNA methylation in human cancers. Epigenetics. 2017, 12, 187–197. [Google Scholar] [CrossRef]
- Lam, J.K.; Chow, M.Y.; Zhang, Y.; Leung, S.W. siRNA Versus miRNA as Therapeutics for Gene Silencing. Mol. Ther. Nucleic Acids. 2015, 4, e252. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.V.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA interference: biology, mechanism, and applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657–685. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.N.; Li, Y.; Xia, S.Q.; Zhang, Y.Y.; Zheng, J.H.; Li, W. PIWI Proteins and PIWI-Interacting RNA: Emerging Roles in Cancer. Cell. Physiol. Biochem. 2017, 44, 1–20. [Google Scholar] [CrossRef]
- Tang, X.; Xie, X.; Wang, X.; Wang, Y.; Jiang, X.; Jiang, H. The combination of piR-823 and eukaryotic initiation factor 3 B (EIF3B) activates hepatic stellate cells via upregulating TGF-β1 in liver fibrogenesis. Med. Sci. Monit. 2018, 24, 9151–9165. [Google Scholar] [CrossRef]
- Le Thomas, A.; Rogers, A.K.; Webster, A.; Marinov, G.K.; Liao, S.E.; Perkins, E.M.; Hur, J.K.; Aravin, A.A.; Tóth, K.F. Piwi induces piRNA-guided transcriptional silencing and establishment of a repressive chromatin state. Genes Dev. 2013, 27, 390–399. [Google Scholar] [CrossRef]
- Liberman, N.; Wang, S.Y.; Greer, E.L. Transgenerational epigenetic inheritance: from phenomena to molecular mechanisms. Curr. Opin. Neurobiol. 2019, 59, 189–206. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional classification and experimental dissection of long noncoding RNAs. Cell. 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (lncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non-Coding RNA, 2021; 7, 36. [Google Scholar]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Kazimierczyk, M.; Wrzesinski, J. Long Non-Coding RNA Epigenetics. Int. J. Mol. Sci. 2021, 22, 6166. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: long non-coding RNA functions in di-verse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.Y.J.; Jackson, R.A.A.; Thiery, J.P.P. EMT: 2016. Cell. 2013, 166, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Monkman, J.; Toh, A.K.L.; Nagaraj, S.H.; Thompson, E.W. Targeting epithelial-mesenchymal plasticity in cancer: clinical and preclinical advances in therapy and monitoring. Biochem. J. 2017, 474, 3269–3306. [Google Scholar] [CrossRef]
- Yuan, S.; Norgard, R.J.; Stanger, B.Z. Cellular plasticity in cancer. Cancer Discov, 2019; 9, 837–851. [Google Scholar]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell. 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- 62. Arnoux ,V.; Nassour, M.; L'Helgoualc'h, Hipskind R.A.; Savagner, P. Erk5 controls Slug expression and keratinocyte activation during wound healing. Mol. Biol. Cell. 2018; 19, 4738–4749.
- Acloque, H.; Adams, M.S.; Fishwick, K.; Bronner-Fraser, M.; Nieto, M.A. Epithelial-mesenchymal transitions: the importance of changing cell state in development and disease. J. Clin. Invest. 2009, 119, 1438–1449. [Google Scholar] [CrossRef] [PubMed]
- 64. Kalluri, R, Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Invest, 2009; 119, 1420–1428.
- Willis, B.C.; duBois, R.M.; Borok, V. Epithelial origin of myofibroblasts during fibrosis in the lung. Proc. Am. Thorac. Soc 2006, 3, 377–382. [Google Scholar] [CrossRef]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J. P. Mesenchymal-epithelial Transition in Development and Reprogramming. Nat. Cel Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Haerinck, J.; Goossens, S.; Berx, G. The epithelial–mesenchymal plasticity landscape: principles of design and mechanisms of regulation. Nat. Rev. Genet. 2023, 24, 590–609. [Google Scholar] [CrossRef]
- Dong, B.; Qiu, Z.; Wu, Y. Tackle Epithelial-Mesenchymal Transition With Epigenetic Drugs in Cancer. Front. Pharmacol. 2020, 11, 596239. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. EMT transition states during tumor progression and metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef]
- Yang, J.; Antin, P.; Berx, G.; Blanpain, C.; Brabletz, T.; Bronner, M.; et al. Guidelines and Definitions for Research on Epithelial-Mesenchymal Transition. Nat. Rev. Mol. Cel Biol. 2020, 21, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Dongre, A. , Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019; 20, 69–84. [Google Scholar]
- Puisieux, A.; Brabletz, T.; Caramel, J. Oncogenic roles of EMT-inducing transcription factors. Nat. Cell Biol. 2014, 16, 488–494. [Google Scholar] [CrossRef]
- Lamouille, S. , Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014; 15, 178–196. [Google Scholar]
- Liu, Q.L.; Luo, M.; Huang, C.; Chen, H.N.; Zhou, Z.G. Epigenetic Regulation of Epithelial to Mesenchymal Transition in the Cancer Metastatic Cascade: Implications for Cancer Therapy. Front Oncol. 2021, 11, 657546. [Google Scholar] [CrossRef]
- Verstappe, J.; Berx, G. A role for partial epithelial-to-mesenchymal transition in enabling stemness in homeostasis and cancer. Semin Cancer Biol. 2023, 90, 15–28. [Google Scholar] [CrossRef]
- Lovisa, S.; LeBleu, V.S.; Tampe, B.; Sugimoto, H.; Vadnagara, K.; Carstens, J.L; Wu, C.C. , Hagos, Y.; Burckhardt, B.C.; Pentcheva-Hoang, T.; Nischal, H.; Allison, J.P.; Zeisberg, M.; Kalluri, R. Epithelial-to-mesenchymal transition induces cell cycle arrest and parenchymal damage in renal fibrosis. Nat. Med. 2015, 21, 998–1009. [Google Scholar]
- Di Gregorio, J.; Robuffo, I.; Spalletta, S.; Giambuzzi, G.; De Iuliis, V.; Toniato, E.; Martinotti, S.; Conti, P.; Flati, V. The Epithelial-to-Mesenchymal Transition as a Possible Therapeutic Target in Fibrotic Disorders. Front. Cell Dev. Biol. 2020, 8, 607483. [Google Scholar] [CrossRef] [PubMed]
- Marconi, G.D.; Fonticoli, L.; Rajan, T.S.; Pierdomenico, S.D.; Trubiani, O.; Pizzicannella, J.; Diomede, F. Epithelial-Mesenchymal Transition (EMT): The Type-2 EMT in Wound Healing, Tissue Regeneration and Organ Fibrosis. Cells. 2021, 10, 1587. [Google Scholar] [CrossRef]
- Rowe, R.G.; Lin, Y.; Shimizu-Hirota, R.; Hanada, S.; Neilson, E.G.; Greenson, J.K.; Weiss, S.J. Hepatocyte-derived Snail1 propagates liver fibrosis progression. Mol. Cell Biol. 2011, 31, 2392–2403. [Google Scholar] [CrossRef]
- Karin, D.; Koyama, Y.; Brenner, D.; Kisseleva, T. The characteristics of activated portal fibroblasts/myofibroblasts in liver fibrosis. Differentiation. 2016, 92, 84–92. [Google Scholar] [CrossRef]
- Iwaisako, K.; Jiang, C.; Zhang, M.; Cong, M.; Moore-Morris, T.J.; Park, T.J.; Liu, X.; Xu, J.; Wang, P.; Paik, Y.H.; Meng, F.; Asagiri, M.; Murray, L.A.; Hofmann, A.F.; Iida, T.; Glass, C.K.; Brenner, D.A.; Kisseleva, T. Origin of myofibroblasts in the fibrotic liver in mice. Proc. Natl. Acad. Sci. U S A. 2014, 111, E3297–305. [Google Scholar] [CrossRef]
- Michelotti, G.A.; Xie, G.; Swiderska, M.; Choi, S.S.; Karaca, G.; Kruger, L.; Premont, R.; Yang, L.; Syn, W.K.; Metzger, D.; Diehl, A.M. Smoothened is a master regulator of adult liver repair. J. Clin. Invest. 2013, 123, 2380–2394. [Google Scholar] [CrossRef]
- Salton, F.; Volpe, M.C.; Confalonieri, M. Epithelial-Mesenchymal Transition in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Medicina (Kaunas). 2019, 55, 83. [Google Scholar] [CrossRef]
- Yang, Z.C.; Yi, M.J.; Ran, N.; Wang, C.; Fu, P.; Feng, X.Y.; Xu, L.; Qu, Z.H. Transforming growth factor-beta1 induces bronchial epithelial cells to mesenchymal transition by activating the snail pathway and promotes airway remodeling in asthma. Mol. Med. Rep. 2013, 8, 1663–1668. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hirai, S.; Tanaka, Y.; Sumi, T.; Miyajima, M.; Mishina, T.; Yamada, G.; Otsuka, M.; Hasegawa, T.; Kojima, T.; Niki, T.; Watanabe, A.; Takahashi, H.; Sakuma, Y. Fibroblastic foci, covered with alveolar epithelia exhibiting epithelial-mesenchymal transition, destroy alveolar septa by disrupting blood flow in idiopathic pulmonary fibrosis. Lab. Investig. 2017, 97, 232–242. [Google Scholar] [CrossRef]
- Liu, L.; Sun, Q.; Davis, F.; Mao, J.; Zhao, H.; Ma, D. Epithelial-mesenchymal transition in organ fibrosis development: current understanding and treatment strategies. Burns Trauma. 2022, 10, tkac011. [Google Scholar] [CrossRef]
- Zolak, J.S.; Jagirdar, R.; Surolia, R.; Karki, S.; Oliva, O.; Hock, T.; Guroji, P.; Ding, Q.; Liu, R.M.; Bolisetty, S.; Agarwal, A.; Thannickal, V.J.; Antony, V.B. Pleural mesothelial cell differentiation and invasion in fibrogenic lung injury. Am. J. Pathol. 2013, 182, 1239–1247. [Google Scholar] [CrossRef]
- Luo, G.H.; Lu, Y.P.; Yang, L.; Song, J.; Shi, Y.J.; Li, Y.P. Epithelial to mesenchymal transformation in tubular epithelial cells undergoing anoxia. Transplant Proc. 2008, 40, 2800–2803. [Google Scholar] [CrossRef]
- Grande, M.T.; Sanchez-Laorden, B.; Lopez-Blau, C.; De Frutos, C.A.; Boutet, A.; Arevalo, M. .; Rowe, R.G.; Weiss, S.J.; López-Novoa, J.M.; Nieto, M.A. Snail1-induced partial epithelial-to-mesenchymal transition drives renal fibrosis in mice and can be targeted to reverse established disease. Nat. Med. 2015, 21, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Zhuang, S. New Insights Into the Role and Mechanism of Partial Epithelial-Mesenchymal Transition in Kidney Fibrosis. Front. Physiol. 2020, 11, 569322. [Google Scholar] [CrossRef] [PubMed]
- Park Kim, J.; Lee, Y.J.; Bae, S.U.; Lee, H.W. Inflammatory bowel disease-associated intestinal fibrosis. J. Pathol. Transl. Med. 2023, 57, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wenxiu, J.; Mingyue, Y.; Fei, H.; Yuxin, L.; Mengyao, W.; Chenyang, L.; Jia, S.; Hong, Z.; Shih, D.Q.; Targan, S.R.; Xiaolan, Z. Effect and mechanism of TL1A expression on epithelial-mesenchymal transition during chronic colitis-related intestinal fibrosis. Mediat. Inflamm. 2021, 2021, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Luan, F.; Zhao, Y.; Hao, H.; Zhou, Y.; Han, W.; Fu, X. Epithelial-mesenchymal transition: An emerging target in tissue fibrosis. Exp. Biol. Med. 2016, 241, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Masia, D.; Gisbert-Ferrandiz, L.; Bauset, C.; Coll, S.; Mamie, C.; Scharl, M.; Esplugues, J.V.; Alós, R.; Navarro, F.; Cosín-Roger, J.; Barrachina, M.D.; Calatayud, S. Succinate activates EMT in intestinal epithelial cells through SUCNR1: a novel protagonist in fistula development. Cell. 2020, 9, 1104. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.N.; Feng, Q. Cardiac repair by epicardial EMT: current targets and a potential role for the primary cilium. Pharmacol. Ther. 2018, 186, 114–129. [Google Scholar] [CrossRef]
- Zeisberg, E. M.; Tarnavski, O.; Zeisberg, M.; Dorfman, A. L.; McMullen, J. R. .; Gustafsson, E.; Chandraker, A., Yuan, X; Pu, W.T.; Roberts, A.B.; Neilson, E.G.; Sayegh, M.H.; Izumo, S.; Kalluri, R. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat. Med. 2007, 13, 952–961. [Google Scholar]
- Travers, J.; Kamal, F.; Robbins, J.; Yutzey, K.; Blaxall, B. Cardiac fibrosis: the fibroblast awakens. Circ. Res. 2016, 118, 1021–1040. [Google Scholar] [CrossRef]
- Mia, M.M.; Singh, M.K. New Insights into Hippo/YAP Signaling in Fibrotic Diseases. Cells. 2022, 11, 2065. [Google Scholar] [CrossRef]
- Aharonov, A.; Shakked, A.; Umanski, K.; Savidor, A.; Genzelinakh, A.; Kain, D.; Lendengolts, D. , Revach, O.Y., Morikawa, Y.; Dong, J.; Levin, Y.; Geiger, B.; Martin, J.F.; Tzahor, E. ERBB2 drives YAP activation and EMT-like processes during cardiac regeneration. Nat. Cell Biol 2020, 22, 1346–1356. [Google Scholar]
- Ling, J.; Cai, Z.; Jin, W.; Zhuang, X.; Kan, L.; Wang, F.; Ye, X. Silencing of c-Ski aug-ments TGF-b1-induced epithelial-mesenchymal transition in cardiomyocyte H9C2 cells. Cardiol. J. 2019, 26, 66–76. [Google Scholar] [CrossRef]
- Bookman, A. A. M.; Shen, H.; Cook, R. J.; Bailey, D.; McComb, R. J.; Rutka, J. A.; Slomovic, A.R.; Caffery, B. Whole stimulated salivary flow: Correlation with the pathology of inflammation and damage in minor salivary gland biopsy specimens from patients with primary Sjögren’s syndrome but not patients with sicca. Arthritis Rheum. 2011, 63, 2014–2020. [Google Scholar] [CrossRef] [PubMed]
- Llamas-Gutierrez, F.J.; Reyes, E.; Martínez, B.; Hernández-Molina, G. Histopathological environment besides the focus score in Sjögren's syndrome. Int. J. Rheum. Dis. 2014, 17, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Altrieth, A.L.; O'Keefe, K.J.; Gellatly, V.A.; Tavarez, J.R.; Feminella, S.M.; Moskwa, N.L.; Cordi, C.V.; Turrieta, J.C.; Nelson, D.A.; Larsen, M. Identifying fibrogenic cells following salivary gland obstructive injury. Front Cell Dev Biol. 2023, 11, 1190386. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.E.; Zheng, C.; Swaim, W.D.; Cho, A.; Nagineni, C.N.; Eckhaus, M.A.; Flanders, K.C.; Ambudkar, I.S.; Baum, B.J.; Kulkarni, A.B. Conditional overexpression of TGF-beta1 disrupts mouse salivary gland development and function. Lab. Invest. 2010, 90, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Lorusso, L.; Ingravallo, G.; Ribatti, D.; Lisi, S. TGFβ1-Smad canonical and -Erk noncanonical pathways participate in interleukin-17-induced epithelial-mesenchymal transition in Sjögren's syndrome. Lab. Invest. 2020, 100, 824–836. [Google Scholar] [CrossRef]
- Brasier, A.R.; Qiao, D.; Zhao, Y. The Hexosamine Biosynthetic Pathway Links Innate Inflammation With Epithelial-Mesenchymal Plasticity in Airway Remodeling. Front. Pharmacol. 2021, 12, 808735. [Google Scholar] [CrossRef] [PubMed]
- Galle, E.; Thienpont, B.; Cappuyns, S.; Venken, T.; Busschaert, P.; Van Haele, M.; Van Cutsem, E.; Roskams, T.; van Pelt, J.; Verslype, C.; Dekervel, J.; Lambrechts, D. DNA methylation-driven EMT is a common mechanism of resistance to various therapeutic agents in cancer. Clin. Epigenetics. 2020, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Bücker, L.; Lehmann, U. CDH1 (E-cadherin) Gene Methylation in Human Breast Cancer: Critical Appraisal of a Long and Twisted Story. Cancers (Basel). 2022, 14, 4377. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; van Tilborg, A.A.; Zwarthoff, E.C. DNA methylation-based biomarkers in bladder cancer. Nat Rev Urol. 2013, 10, 327–335. [Google Scholar] [CrossRef]
- Bechtel, W.; McGoohan, S.; Zeisberg, E.M.; Müller, G.A.; Kalbacher, H.; Salant, D.J.; Müller, C.A.; Kalluri, R.; Zeisberg, M. Methylation determines fibroblast activation and fibrogenesis in the kidney. Nat Med. 2010, 16, 544–550. [Google Scholar] [CrossRef]
- Chang, Y.W.; Singh, K.P. Arsenic induces fibrogenic changes in human kidney epithelial cells potentially through epigenetic alterations in DNA methylation. J. Cell. Physiol. 2019, 234, 4713–4725. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Shi, P.; Zhao, X.D.; Xuan, H.Y.; Gong, W.H.; Ding, X.S. DNMT1 deregulation of SOCS3 axis drives cardiac fibroblast activation in diabetic cardiac fibrosis. J. Cell Physiol. 2021, 236, 3481–3494. [Google Scholar] [CrossRef] [PubMed]
- Effendi, W.I.; Nagano, T. Epigenetics Approaches toward Precision Medicine for Idiopathic Pulmonary Fibrosis: Focus on DNA Methylation. Biomedicines. 2023, 11, 1047. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Zhou, H. , Li, Y., Zheng, Q.; Ding, M.; Zhou, H. Li,X. Epigenetic modification in liver fibrosis: Promising therapeutic direction with significant challenges ahead. Acta Pharmaceutica Sinica B, 2023; in press. [Google Scholar]
- Avci, E.; Sarvari, P.; Savai, R.; Seeger, W.; Pullamsetti, S.S. Epigenetic Mechanisms in Parenchymal Lung Diseases: Bystanders or Therapeutic Targets? Int. J. Mol. Sci. 2022, 23, 546. [Google Scholar] [CrossRef]
- Ning, L.; Rui, X.; Bo, W.; Qing, G. The critical roles of histone deacetylase 3 in the pathogenesis of solid organ injury. Cell Death Dis. 2021, 12, 734. [Google Scholar] [CrossRef]
- Ghoneim, M.; Fuchs, H. , Musselman, C. Histone tail conformations: a fuzzy affair with DNA. Trends Biochem Sci. 2021, 46, 564–578. [Google Scholar]
- de Ruijter, A.J.; van Gennip, A.H.; Caron, H.N.; Kemp, S.; van Kuilenburg, A.B. Histone deacetylases (HDACs): characterization of the classical HDAC family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [CrossRef]
- Claveria-Cabello, A,; Colyn, L. ; Arechederra, M.; Urman, J.M.; Berasain, C.; Avila, M.A.; Fernandez-Barrena, M.G. Epigenetics in Liver Fibrosis: Could HDACs be a Therapeutic Target? Cells. 2020, 9, 2321. [Google Scholar] [CrossRef]
- Huang, S.K.; Scruggs, A.M.; Donaghy, J. ; Horowitz. J.C.; Zaslona. Z.; Przybranowski. S.; White, E.S.; Peters-Golden, M. Histone modifications are responsible for decreased Fas expression and apoptosis resistance in fibrotic lung fibroblasts. Cell Death Dis 2013, 4, e621. [Google Scholar]
- Sanders, Y.Y.; Hagood, J.S.; Liu, H.; Zhang, W.; Ambalavanan, N.; Thannickal, V.J. Histone deacetylase inhibition promotes fibroblast apoptosis and ameliorates pulmonary fibro-sis in mice. Eur. Respir. J. 2014, 43, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zou, J.; Tolbert, E.; Zhao, T.C.; Bayliss, G.; Zhuang, S. Identification of histone deacetylase 8 as a novel therapeutic target for renal fibrosis. FASEB J. 2020, 34, 7295–7310. [Google Scholar] [CrossRef]
- Pang, M.; Kothapally, J.; Mao, H.; Tolbert, E.; Ponnusamy, M.; Chin, Y.E.; Zhuang, S. Inhibition of histone deacetylase activity attenuates renal fibroblast activation and interstitial fibrosis in obstructive nephropathy. Am. J. Physiol. Renal. Physiol. 2009, 297, F996–F1005. [Google Scholar] [CrossRef]
- Rubio, K. , Molina-Herrera, A.; Pérez-González, A.; Hernández-Galdámez, H.V.; Pi-ña-Vázquez, C., Araujo-Ramos, T.; Singh, I. EP300 as a Molecular Integrator of Fibrotic Transcriptional Programs. Int. J. Mol. Sci. 2023, 24, 12302. [Google Scholar]
- Lim, Y.; Jeong, A.; Kwon, D.H.; Lee, Y.U.; Kim, Y.K.; Ahn, Y.; Kook, T.; Park, W.J.; Kook, H. P300/CBP-Associated Factor Activates Cardiac Fibroblasts by SMAD2 Acetylation. Int. J. Mol. Sci. 2021, 22, 9944. [Google Scholar] [CrossRef]
- Chu, L.; Xie, D.; Xu, D. Epigenetic Regulation of Fibroblasts and Crosstalk between Cardiomyocytes and Non-Myocyte Cells in Cardiac Fibrosis. Biomolecules 2023, 13, 1382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, G.; Zhang, Y.; Zhang, M.; Zhou, J.; Gao, W.; Xuan, X.; Yang, X.; Yang, D.; Tian, Z.; Ni, B.; Tang, J. Critical effects of long non-coding RNA on fibrosis diseases. Exp. Mol. Med. 2018, 50, e428. [Google Scholar] [CrossRef]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef]
- Wu, C.; Bao, S.; Sun, H.; Chen, X.; Yang, L.; Li, R.; Peng, Y. Noncoding RNAs regulating ferroptosis in cardiovascular diseases: novel roles and therapeutic strategies. Mol. Cell Biochem. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Khanbabaei, H.; Ebrahimi, S.; García-Rodríguez, J.L.; Ghasemi, Z.; Pourghadamyari, H.; Mohammadi, M.; Kristensen, L.S. Non-coding RNAs and epithelial mesenchymal transition in cancer: molecular mechanisms and clinical implications. J. Exp. Clin. Cancer. Res. 2022, 41, 278. [Google Scholar] [CrossRef]
- Davalos, V.; Moutinho, C.; Villanueva, A.; Boque, R.; Silva, P.; Carneiro, F.; Esteller, M. Dynamic epigenetic regulation of the microRNA-200 family mediates epithelial and mesenchymal transitions in human tumorigenesis. Oncogene. 2012, 31, 2062–2074. [Google Scholar] [CrossRef]
- Tang, O.; Chen, X.M.; Shen, S.; Hahn, M.; Pollock, C.A. MiRNA-200b represses transforming growth factor-β1-induced EMT and fibronectin expression in kidney proximal tubular cells. Am. J. Physiol. Renal. Physiol. 2013, 304, F1266–F1273. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Xu, Q.; Yao, W.; Wu, Q.; Yuan, J.; Yan, W.; Xu, T.; Ji, X.; Ni, C. Long non-coding RNA-ATB promotes EMT during silica-induced pulmonary fibrosis by competitively binding miR-200c. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 420–431. [Google Scholar] [CrossRef]
- Lin, J.; Jiang, Z.; Liu, C.; Zhou, D.; Song, J.; Liao, Y.; Chen, J. Emerging Roles of Long Non-Coding RNAs in Renal Fibrosis. Life (Basel). 2020, 10, 131. [Google Scholar] [CrossRef]
- Wang, P.; Luo, M.L.; Song, E.; Zhou, Z.; Ma, T.; Wang, J.; Jia, N.; Wang, G.; Nie, S.; Liu, Y.; Hou, F. Long noncoding RNA lnc-TSI inhibits renal fibrogenesis by negatively regulating the TGF-beta/Smad3 pathway. Sci. Transl. Med. 2018, 10, eaat2039. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jiang, Z.; Li, Y.; Zhang, Y.; Han, Y.; Gao, L. Non-coding RNAs regulating epithelial-mesenchymal transition: Research progress in liver disease. Biomed Pharmacother. 2022, 150, 112972. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Huang, C.; Lin, X.; Li, J. MicroRNA-29 family, a crucial therapeutic target for fibrosis diseases. Biochimie. 2013, 95, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, M.; Uchida, S. Long Non-Coding RNAs in Cardiac and Pulmonary Fibroblasts and Fibrosis. Non-Coding RNA 2022, 8, 53. [Google Scholar]
Figure 1.
A schematic overview of epigenetic modifications. Epigenetic regulation involves DNA methylation, histone modification and non-coding RNA. DNMT family members mediate DNA methylation, which suppresses gene transcription by adding a methyl group to the position of cytosine. Histone methylation catalyzed by HMTs and HDMTs, and histone acetylation regulated by HATs and HDACs. Non-coding RNA includes miRNA, circRNA and lncRNA. BET (Bromodomain and extraterminal); BRMDs (bromodomains); DNMT (DNA methyltransferases); HATs (Histone acetylases); HDACs (Histone deacetylases); HDMTs (Histone demethylases); HMTs (Histone methyltransferases); lncRNA (long non-coding RNA); TET (Ten-eleven translocation).
Figure 1.
A schematic overview of epigenetic modifications. Epigenetic regulation involves DNA methylation, histone modification and non-coding RNA. DNMT family members mediate DNA methylation, which suppresses gene transcription by adding a methyl group to the position of cytosine. Histone methylation catalyzed by HMTs and HDMTs, and histone acetylation regulated by HATs and HDACs. Non-coding RNA includes miRNA, circRNA and lncRNA. BET (Bromodomain and extraterminal); BRMDs (bromodomains); DNMT (DNA methyltransferases); HATs (Histone acetylases); HDACs (Histone deacetylases); HDMTs (Histone demethylases); HMTs (Histone methyltransferases); lncRNA (long non-coding RNA); TET (Ten-eleven translocation).
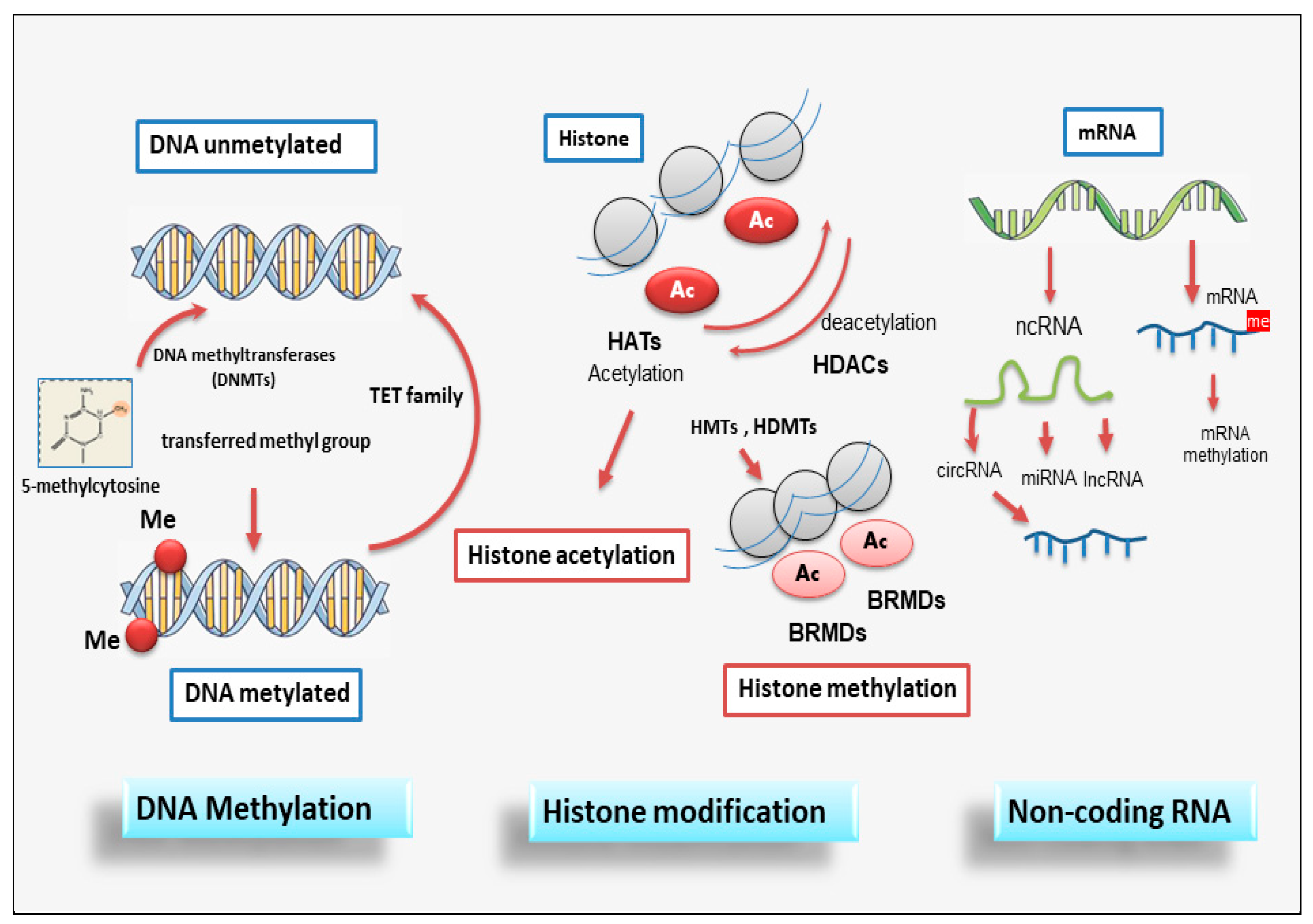
Figure 2.
Schematic representation of epithelial to mesenchymal plasticity (EMP). EMP, in response to epithelial injury, allows cells to convert between multiple states across the epithelial to mesenchymal transformation, acquiring hybrid epithelial/mesenchymal phenotypic features.
Figure 2.
Schematic representation of epithelial to mesenchymal plasticity (EMP). EMP, in response to epithelial injury, allows cells to convert between multiple states across the epithelial to mesenchymal transformation, acquiring hybrid epithelial/mesenchymal phenotypic features.
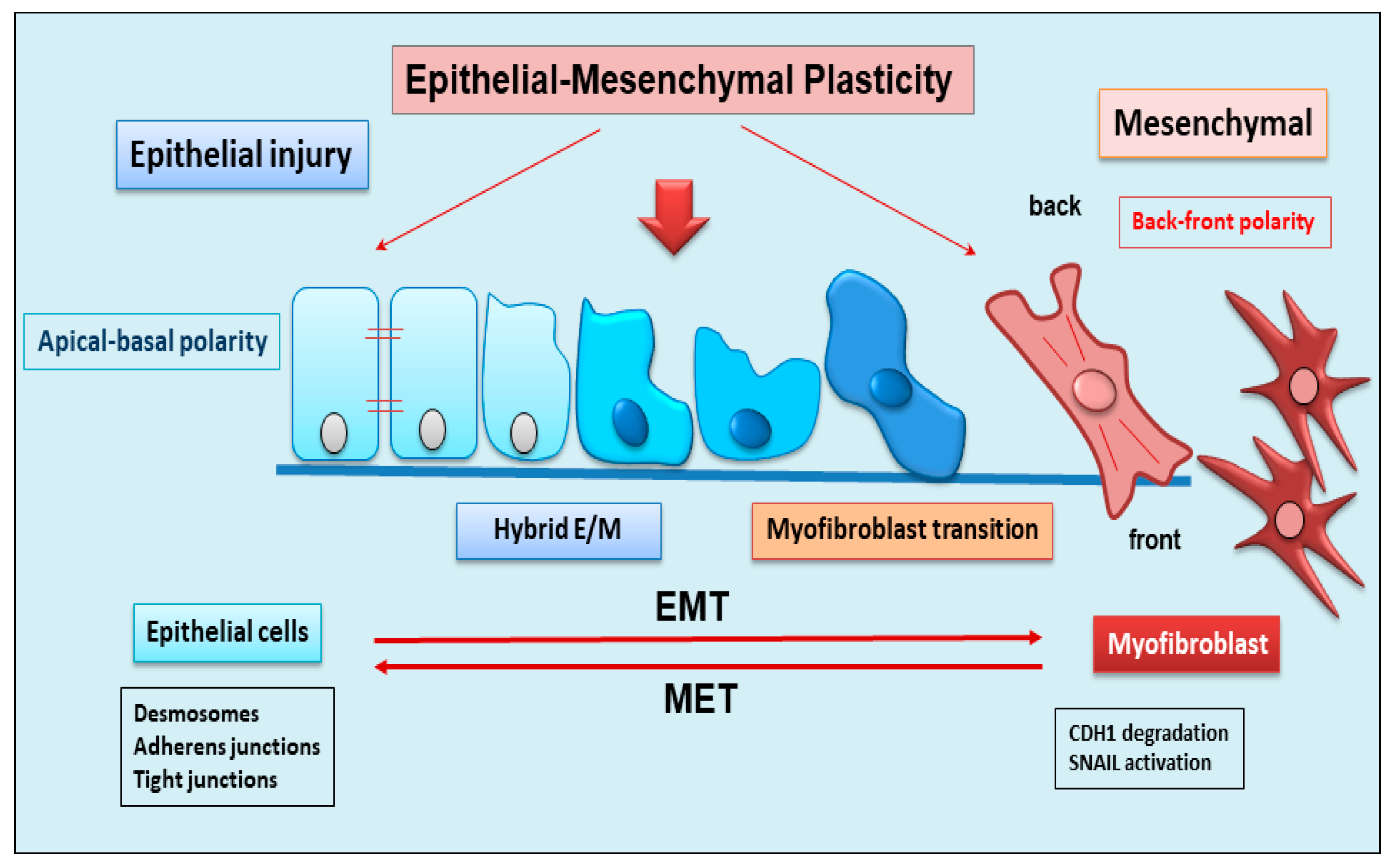
Figure 3.
Scheme of the epigenetic modifications in EMP/EMT-dependent fibrotic diseases. DNMT (DNA methyltransferases); HATs (Histone acetylases); HDACs (Histone deacetylases); EndMT (Endothelial-mesenchymal transition); EMT (Epithelial-mesenchymal transition); SOCS3 (cytokine signaling 3).
Figure 3.
Scheme of the epigenetic modifications in EMP/EMT-dependent fibrotic diseases. DNMT (DNA methyltransferases); HATs (Histone acetylases); HDACs (Histone deacetylases); EndMT (Endothelial-mesenchymal transition); EMT (Epithelial-mesenchymal transition); SOCS3 (cytokine signaling 3).
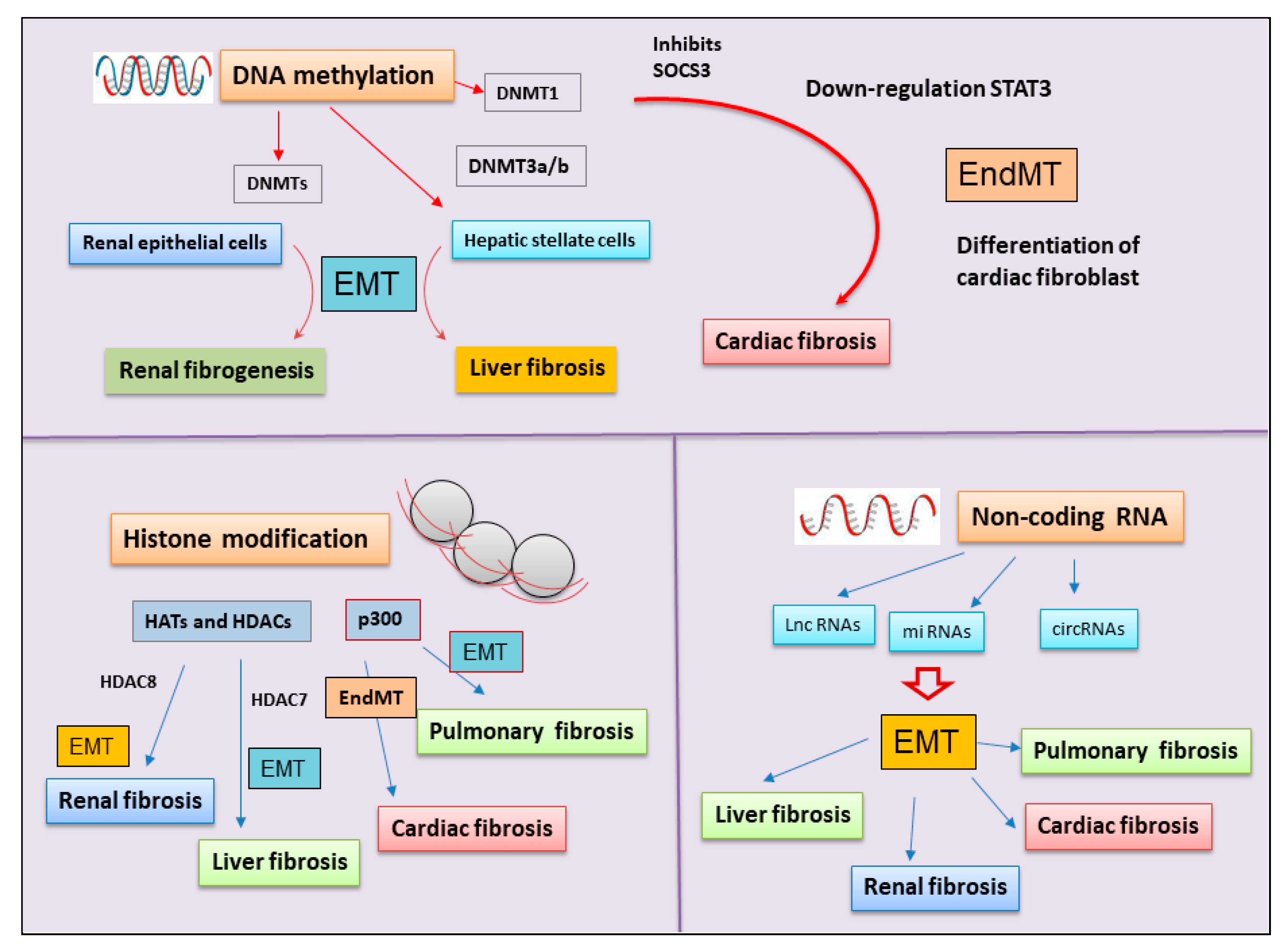
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



