
Submitted:
31 December 2023
Posted:
03 January 2024
You are already at the latest version
Abstract
Prostate cancer (PC) is the second most diagnosed cancer among men. It was observed that early diagnosis of disease is highly beneficial for survival of cancer patients. Therefore, extension and increasing quality of life of PC patients can be achieved by broadening the Cancer Screening programs that are aimed at the identification of cancer manifestation in patients at earlier stages before they demonstrate well-understood signs of the disease. Therefore, there is an urgent need in standard, sensitive, robust, and commonly available screening and diagnosis tools for identification of early signs of cancer pathologies. In this respect, “Holy Grail” of cancer researchers and bioengineers for decades have been molecular sensing probes that would allow diagnosis, prognosis, and monitoring of cancer diseases by their interaction with cell-secreted and cellassociated PC biomarkers, e.g. PSA and PSMA correspondinly. At present, most PSA tests are performed at centralized laboratories using high throughput Total PSA immune analyzers which are suitable for dedicated laboratories and are not readily available for the broad health screening. Therefore, the current trend in detection of PC is developments of the portable biosensors for mobile laboratories and individual use. Phage display since its conception by George Smith in 1985 has emerged as a premier tool in molecular biology with widespread applications. This review describes how the paradigm of molecular evolution and phage display revolutionized the methods of early diagnosis and monitoring of PC.
Keywords:
Phage display
; landscape phage
; molecular evolution
; affinity selection
; recombinant antibodies
; PC
; prostate specific antigen (PSA)
; prostate-specific matrix antigen (PSMA)
; Enzyme-linked immunosorbent assay (ELISA)
; phage ELISA
; phage capture assay
; electrochemical biosensor
; total prostate-specific antigen (t-PSA)
; free prostate-specific antigen (F-PSA)
; electrochemical impedance spectroscopy
; label-free immunosensor
1. Introduction
Prostate cancer (PC) is the second most diagnosed cancer among men [1] It was observed that early diagnosis of disease is highly beneficial for survival of cancer patients [2,3]. Therefore, extension and increasing quality of life of PC patients can be achieved by broadening the Cancer Screening programs that are aimed at the identification of cancer manifestation in patients at earlier stages, before they demonstrate well-understood signs of the disease[2,3,4,5,6,7]. Significant social impact and economical effect of PC screening was estimated taking into consideration that the cost for treating the advanced disease is much higher than the scanning cost [8]. Therefore, there is an urgent need in standard, sensitive, robust, and commonly available screening tools for identification of early signs of cancer pathologies [9]. In this respect, “Holy Grail” of cancer researchers and bioengineers for decades have been molecular sensing probes that would allow diagnosis, prognosis and monitoring of cancer diseases by their interaction with tumor-associated cancer cells and/or blood-solubilized PC biomarkers, such as PSA, PCA3, sarcosine oxidase andothers [2,10,11,12,13,14,15,16,17]. At present, most PSA tests are performed at centralized laboratories using high throughput Total PSA immune analyzers. There are clear practical advantages of using these fully automated analyzers, including lower detection limits and high throughput of samples. Meantime, several authors noted a persistent disagreement among PSA results obtained by different commercial immunoassays [18,19,22]. This serious drawback of the PSA immunoanalizers can be attributed to the use of capture and tracer antibodies with different epitope specificities and affinities [20] Furthermore, , the important limitation of the PSA analyzers is that they are suitable for dedicated laboratories and are not readily available for the broad health care community[19]. Therefore, the emerging trend in screening and detection of PC is using the portable biosensors for mobile laboratories and individual use [14,17,21,22,23]. Phage display since its conception by George Smith in 1985 has emerged as a premier tool in molecular biology with widespread applications. This review describes how the paradigm of molecular evolution and phage display revolutionized the methods of early diagnosis and monitoring of PC.
2. Advanced Phage-Driven Analytical Tools for Diagnosis of PC
New urgent requirements for fast, sensitive, accurate, and inexpensive tools for early diagnosis of PC devalue the traditional PSA detection methods [17], such as enzyme-linked sorbent immunoassay (ELISA), radioimmunoassay immunoradiometric assay, and time-resolved immunofluorescence assay which require complex operation, hard miniaturization and can reveal limited sensitivity [24,25,26,27]. Modern immunoassays and biosensors require a biorecognition probe, which is attached to the interface of the analytical device, bind the target biological ligand and participate in generating a measurable signal [14,15,28,29], as illustrated in Figure 1. For example, in electrochemical biosensors the signal can be displayed in impedimetric, amperometric, or potentiometric formats.
PC detection techniques can be divided into two major categories: 1) cancer cell imaging techniques and 2) cancer cells-secreted soluble biomarker detection techniques [16]. Most analytic platforms rely on the use of monoclonal antibodies as biorecognition probes, such as Fab′ 8G8F5 monoclonal antibody (mAb) shown in in Figure 2 as a complex with PSA. However, their broad application is limited by high cost, low specificity, less-than-op timal affinity, and sensitivity to components of body fluids [7,16,17,22,23,33,34,35,36,37,38,39]. Harnessing the power of molecular evolution, phage display technique offered a new way for generating a rich repertoire of binding probes for any protein ligand or receptor[40,41,42,43,44].. The idea of phage display as a molecular evolution tool lies in the genetic fusion of a foreign protein to the virion structural proteins and its preservation through the viral replication. Considering desirable characteristics of different display systems, filamentous bacteriophages M13 and fd were commonly preferred as suitable vectors for generating peptide phage-displayed libraries (Figure 3). As complementary to p3-type phage display vectors, which were designed to discover diagnostic peptides and antibodies, the p8-type phage technology was developed with the goal of creation of diagnostic and detection nanoprobes by resurfacing of the whole phage particles [31,45,46,47,48,49,50,51,52,53,54].
2.1. Selection of Phage Probes against PC Cell-Associated Antigens
2.1.1. Selection of PC Cell-Binding Phages from f8-Type (landscape) Libraries
Since it was proven that malignant transformation of cells is linked with expression of cell antigens, the tumor cell-specific phage-displayed peptides and antibodies were considered as prospective versatile, diagnostic and therapeutic reagents[55,56]. The first PC cell-targeting landscape phages were discovered by Victor Romanov with colleagues [57]. It was shown that the phage displaying N-terminal 8-mer peptide DPRATPGS inserted in all 4000 domains of major coat protein p8, selected from p8-type (landscape) library f8/8 by it’s affinity selection (biopanning) against PC cells LNCaP (Figure 3,4) blocked spreading of LNCaP cells and their relatives C4-2 and C4-2b [45]. Later, the major principles and methods of phage sselection have been used in publications of other groups, but some details of the protocol were modified with the purpose of increasing specificity and selectivity of discovered phage probes towards the target cancer cells.
Figure 5.
Selection of landscape phage interacting with PC cell-associated antigens.. The most common phage survey strategy is affinity selection, called ‘biopanning’ which enriches for phage particles whose displayed peptides bind the target cells in culture or whole tissues in living animals. To use biopanning for selection of landscape phages against a variety of different PC, the researchers add the library to the immobilized target cells, wash away non-bound phage, elute bound phage particles, and amplify them. After 2-4 rounds of selection, propagate individual clones, and analyze them. This procedure was named biopanning, because it reminds panning — the process of extraction of gold particles from sand – panning.
Figure 5.
Selection of landscape phage interacting with PC cell-associated antigens.. The most common phage survey strategy is affinity selection, called ‘biopanning’ which enriches for phage particles whose displayed peptides bind the target cells in culture or whole tissues in living animals. To use biopanning for selection of landscape phages against a variety of different PC, the researchers add the library to the immobilized target cells, wash away non-bound phage, elute bound phage particles, and amplify them. After 2-4 rounds of selection, propagate individual clones, and analyze them. This procedure was named biopanning, because it reminds panning — the process of extraction of gold particles from sand – panning.

In continuation of this pioneering work, Prashanth Jayanna et al. used PC3 cells as target cells as they imitate the profile of advanced prostate tumors[58]. To increase repertoire of binding phages, f8/9 (9-mer) library was used in addition to f8/8 (8 -mer) library [52,59]. To isolating phages high selectivity towards PC-specific antigens, the libraies were depleted against plastic, serum and normal fibroblast cells before being allowed to interact with the target cells. Relative affinity of selected clones towards targeted and control cells was estimated using selectivity assay is based on interaction of phage particles with PC-3M cells in comparisons with other control cells The affinity of DTDSHVNL to PC3 cells was ~9 times higher than to either of the control cells and 32 times higher than to serum(cell-free media) whereas the affinity of DTPYLDTG to PC3 cells was ~8 times higher than to either of the control cells and 15 times higher than to serum. The other clones analyzed showed high affinity to target cells as well control cancer cells but not to normal epithelial cells or serum leading us to asume that these probes may be directed against a universal cancer receptor. A single clone DVVYALSDD isolated from 9-mer library demonstrated an affinity to PC3 cells that was almost eighty times higher than to the control cells and 600 times than to serum (cell-free media). Surprisingly, the other clones analyzed showed high affinity to target as well as normal epithelial cells but not to control cancer cells or serum indicating that they may directed towards a receptor that is common to both tumor and normal cells. A phage bearing an un-related streptavidin-avid peptide (VPEGAFSS) was used a control to demonstrate specificity of our phage probes.
To extend the panel of PC imaging phage probes, Olusegun Fagbohun et al. have screened landscape phage library f8/8 against metastatic PC cells PC-3M [60]. The most selective for PC-3M cells Phage EPTHSWAT was able to penetrate the PC-3M cells as revealed by immunofluorescence microscopy (Figure 6). Selectivity of the PC-specific phages EPTHSWAT towards PC-3M cells was studied by phage capture assayand demonstrated 35-fold greater binding than the non-relevant control phage VPEGAFSS. Furthermore, phage EPTHSWAT showed a statistically significant higher interaction with PC-3M cells than with other cells RWPE-1, HT-29, and serum. This high interaction of the phage particles with PC-3M cells might be due to pspecific hage interaction with an overexpressed PC cell antigen.
2.1.2. Selection of PC Cell Binders from p3-Type Phage-Displayed Antibody Libraries
Polyclonal antibodies purified from the serum of an immunized animal (i.e., mouse, rabbit, goat, lama, etc.) and monoclonal antibodies (mAb) secreted by immortalized B cells from the spleen of an immunized animal are commonly used in immunological assays. Their dominant role in immunochemical applications was faded after appearance of phage-displayed antibodies which are currently commonly used for discovery and detection of cancer-specific antigens and biomarkers [29,43,61,63]. To isolate antibodies with desired specificities, phage library selections must be performed on tumor-derived antigen sources. The phage display strategy for the selection of rabbit monoclonal antibodies that recognize PC tumor-associated antigens was reported by Mikhail Popkov et al[63]. Researchers immunized rabbits with either human PC cell line LNCap or DU145. Chimeric rabbit/human Fab libraries were generatedthrough oligo(dT)-primed, reverse transcription of RNA from animal’s spleen and bone marrow[64]. The antibody variable domains VL and VH were amplified, fused to human constant domains CL and CH1 and cloned into the phagemid vector pComb3X. Constructed by this way phage-displayed chimeric rabbit/human Fab libraries were screened against human PC cells DU145 using a novel whole-cell panning protocol resulting in discovery of clones bound selectively to DU145 cells but not to primary human prostate epithelial cell line PrEC, as detected by flow cytometry. In summary, this work first demonstrated the potential of immune antibody libraries for identification of imaging phage probes interacting with tumor-associated cell surface antigens.
2.2. Selection of Phage Probes against Prostate Specific Antigen (PSA)
2.2.1. PSA as a PC Biomarker
PSA is a serum marker that is commonly used for the diagnosis of prostatic diseases. Normally, It is produced by epithelial cells of the prostate and exist mostly in two molecular forms: free PSA (f-PSA), 10%−30% of PSAs, and the PSA-α1-antichymotrypsin complex (PSA-ACT), 70%−90% of PSAs [65,66,67,68]. The sum of f-PSA and PSA-ACT is called as the total PSA (t-PSA) is regarded in clinical medicine as the important index for early diagnosis of PC, evaluation of curative effect and monitoring of post operation [7,69,70,71,72,73]. Thus, specific detection of certain subforms could permit discrimination between benign and malignant cases. In general, the content of t-PSA in serum of healthy people is lower than 4 ng/mL, the level accepted as a threshold value in the clinical test of PC. When the content of t-PSA in serum is more than 10 ng/mL, the risk of PC is high, thus the accuracy rates of diagnosis of PC can reach 70 − 80% [74,75,76,77]. Therefore, joint detection of the ratio of f-PSA/t-PSA and the level of t-PSA can more accurately discriminate PC. and prostate diseases.
2.2.2. Selection of p3-Type Phage Displayed Peptides against PSA
To obtain peptide ligands specificly recognizing different forms of PSA , phage-displayed linear and cyclic peptide libraries were screened against PSA-coated microplate wells or PSA supported by immobilized anti-total PSA mAbs[78]. In the pioneering work, Ping Wu et all. [79,80,81] discovered PSA-binding peptides by screening p3-type cyclic and linear peptide phage display libraries. A p3-fused cyclic peptide with four bridged cysteine residues showed the highest affinity for PSA. The binding specificity was characterized by competition with monoclonal anti-PSA antibodies of known epitope specificities. The peptides bound to the same region as mAbs specific for free PSA indicating that they bind close to the active site of the enzyme. These results demonstrated that peptides binding to PSA and modulating its enzyme activity can be developed by phage display technique. However, when discovered peptides were tested in sandwich capturing PSA assays with the anti-PSA 5D5A5 mAb the lowest concentrations of detectable PSA were 0.2-2 mg/ml, not sensitive enough to allow PSA to be quantified in sera from patients with prostatic di-seases where the concentration of PSA was superior to 2–4 ng/ml. In the recent study of Wang et al.[82] the elution strategy in the biopanning of p3-type phage displayed peptide library Ph.D.–12 (New England Biolabs) against PSA was optimized by by additional BSA pre-screening and serum interference. PSA-specific phage expressing peptide TSIANYIGLALR showed the best affinity and specificity against PSA, was conjugated through C-terminal GGGGSK-biotin linker to streptavidin and used this construct as an signal amplifier in the, sandwiched ELISA system. The system could detect total prostate-specific antigen (tPSA) with the linear range of 0.25-200 ng/mL and the detection limit of 0.18 ng/ml demonstrating a good prospect of using peptide-streptavidin conjugates as substitute signaling antibodies in t-PSA.
2.2.3. Development and Affinity Maturation of p3-Type Phage Displayed Antibodies against PSA
The power of directed-evolution and phage display was succeeded by Muller et al. in enhancing affinity and sensitivity of immunoassay while maintaining its selectivity [83]. The original f-PSA assay based on the use of the high off rate 4D4 Mab as a tracer was less than ideal regarding sensitivity and low-end robustness of the assay. Attempts to use the 4D4 Mab for capturing was also not successful. Using phage-display library-derived mutant L3-2 Fab with reduced off-rate dissociation constant, both configurations were possible and improved assay performances. In comparison with the wild-type scFv, the best binders showed an enhancement of sensitivity in sandwich immunoassay.
2.2.4. Selection of PSA-Binding p8-Type Multivalent Landscape Phage Probes
In the p8-type phage display system, called Landscape Phage, the dense array of foreign peptides on the body of the phage composes a unique organic landscape, in which the structure and function of individual peptides can be influenced by interactions with neighboring proteins (Figure 3). Therefore, each landscape phage can be treated as a unique nanomaterial with novel and emergent properties that cannot be observed by use of an individual synthetic peptide alone. In many applications, including detection and imaging of PC cells, the extreme multivalency of landscape phages is a great advantage . It was proved that the landscape libraries represent an inexhaustible rich source of substitute antibodies—filaments that bind protein and glycoprotein antigens with nanomolar affinities and high specificity [26,46,48,50,51,84,85,86,87,88,89,90]. The foreign amino acids that form the bispecific `active site' of a landscape phage comprise up to 25% by weight of the particle and subtend up to 50% of its surface area which can accommodate hundreds of bound protein antigens. The phage structure is extraordinarily robust, being resistant to heat, organic solvents, urea, acid, or alkali, and can tolerate different modifications that increase efficacy of their use as detection and diagnostic probes [91]. Purified phages can be stored indefinitely at moderate temperatures without losing their infectivity and binding activity [92,93,94,95]. More detailed information regarding the evolution of Landscape Phage Detector paradigm, starting from its appearance in 1996 as a distinct part of the Phage Display concept[40,96], use the landscape phage as Phage Substitute Antibodies in the first Phage Biosensor and development of the landscape phage-based biosensors for liquid biopsy of PC can be found in the references[46,48,50,57,60,84,88,93,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112].
Target-specific landscape-phage probes can be prepared as described in commonly available protocols [26,51], as illustrated in (Figure 8). Thus, the specific phage probes against t-PSA were selected from the f8/8 landscape phage library[88,113]. Through three rounds of bio panning and phage capture assay, a novel phage clone P1 and P5 displaying octamers ERNSVSPS and ATRSANGM with the best affinity and selectivity for t-PSA was successfully obtained and then used as the capture probe to establish both ELISA and DPV assay systems. (Figure 10)
3. Development of Phage-Driven Biosensors for Detection of Different forms of PSA
The critical factor that determines the efficacy of early cancer detection is the analytical platform that converts invisible molecular binding events into the optical or electrical signal (Figure 1).. The common methods used for the detection of cancer-specific antigens (bio-markers) include immunohistochemistry, enzyme-linked immunosorbent assay (ELISA), and different types of immunosensors [84,114,115]. To date, most analytical platforms for detection of cancer biomarkers use monoclonal antibodies (mAb) as sensing probes [10,15,21,35,37,116,117,118,119,120,121,122,123] There is an urgent need in robust inexpensive highly sensitive and easily available sensing probes, such as landscape phage substitute antibodies [40,46]. It was shown that due to presentation of diagnostic peptide in ~4,000 copies on the surface of the phage particles, that form a dense interface in detection platforms, significantly increase their affinity and selectivity towards the analyte markers[48,49,124,125,126], as illustrated in Figure 9.
3.1. Landscape Phage-Driven Enzyme-Linked Immunosorbent Assay (Phage ELISA)
ELISA is normally designed in two different formats, broadly categorized as direct, for the detection of antigens, and indirect, for the detection of antigen-specific antibodies [127,128]. Within these broad categories, there are multiple variants, including the sandwich direct ELISA, in which the immobilized capture antibody binds the water-soluble target antigen to form a complex, which is detected by detector antibody that binds the captured antigen and produce a visible signal after adding a chromogenic substrat. Landscape phage can be used in direct ELISA as substitutes both for capture and detection antibodies [62].. The phage-based substitute antibody leverages the multivalency of the p8 display system for enhanced analyte capture, whereas the landscape phage-based signaling antibody benefits from phage multivalency to achieve ignal amplification, as illustrated in Figure 1. The details about the affinity selection of t-PSA- and f-PSA-specific octapeptide-fusion phages can be found in [26,106,113]. Shortly, phages selected in biopanning procedures against immobilized recombinant t-PSA and f-PSA showed the best affinity and selectivity as capture probes in a model ELISA and demonstrated good sensitivity and reliability in the t-PSA and f-PSA analysis in real serum samples. This work first proved that phage-based immunoassay can be applicable to clinical diagnosis of prostatic cancer. It is prospective that the obtained specific landscape phage would be used as a novel probe to replace traditional antibodies in practical PSA immunoassay.
3.2. Phage-Driven Electrochemical Immunosensors for Detection of PSA
Electrochemical immunosensors attracted attention of bioengineers as bioanalytical platforms for PC detection because of their high sensitivity, specificity, simple operation and easy miniaturization. Among numerous electrochemical methods, the electrochemical impedance spectroscopy (EIS) not only inherits the normal advantages of electrochemical immunosensor, but also shows ultra-high sensitivity [37,129,130,131,132,133,134]. Like other immunoassays, the EIS technology are based on the specific immunological recognition of the ligand with the antigen [135,136]. Considering the unique properties of landscape phages, Lei Han et al. constructed first phage-based dual f-PSA/t-PSA ratio assay [88]. As sensing probes for construction of the sensor’s interface (Figure 9) the researchers used landscape phages fused with octapeptides ERNSVSPS and ATRSANGM that have been discovered through screening the f8/8 landscape phages library against f-PSA and t-PSA respectively (Figure 10) [26,113]. The fabricated immunosensors showed high specificity, ultra-low limit of detection, wide linear detection range, excellent reusability, high reproducibility, and good stability that corresponds to high stability and biocompatibility of fibrous phage interface, which is determined by multivalency of surface-displayed octapeptides. Specifically, the phage-driven sensors demonstrated wide linear ranges (0.02 pg mL−1 – 200 ng mL-1 for f-PSA; 0.02 pg mL−1 – 200 ng mL−1 for t-PSA. Moreover, the immunosensors showed remarkably lower limits of detection (3 fg mL−1 f-PSA, S/N = 3; 4 fg mL−1 t-PSA, S/N = 3) than antibody- and aptamer-based methods reviewed in [113]. The results of this study provided a novel avenue to construct the phage-based sensors for the dual detection of f-PSA and t-PSA and analysis of f-PSA/t-PSA ratio in human blood. Authors suggested that phage clones for different epitopes of PSA can be obtained by biopanning and more phage-based immunosensors would be developed.
4. Conclusions
To summarize, the fabrication of the specific phage-based immunosensors for ultrasensitive detection of f-PSA and t-PSA in human sera is a encouraging example of effective harnessing the power of molecular evolution and phage display for creation of smart materials that can be used for extension and improving the quality of life of millions PC patients.
References
- Chhikara, B.S.; Parang, K. Global Cancer Statistics 2022: the trends projection analysis. Chemical Biology Letters 2023, 10. [Google Scholar]
- Hugosson, J.; Roobol, M.J.; Mansson, M.; Tammela, T.L.J.; Zappa, M.; Nelen, V.; Kwiatkowski, M.; Lujan, M.; Carlsson, S.V.; Talala, K.M.; et al. A 16-yr Follow-up of the European Randomized study of Screening for Prostate Cancer. Eur Urol 2019, 76, 43–51. [Google Scholar] [CrossRef]
- Hawkes, N. Cancer survival data emphasize importance of early diagnosis. BMJ 2019, 364, l408. [Google Scholar] [CrossRef]
- Crosby, D.; Bhatia, S.; Brindle, K.M.; Coussens, L.M.; Dive, C.; Emberton, M.; Esener, S.; Fitzgerald, R.C.; Gambhir, S.S.; Kuhn, P.; et al. Early detection of cancer. Science 2022, 375, 1244-+. [Google Scholar] [CrossRef]
- Shah, N.; Ioffe, V. Early Detection of Prostate Cancer: AUA/SUO Guideline Part I: Prostate Cancer Screening. Letter. J Urol 2023, 210, 731. [Google Scholar] [CrossRef]
- Moghul, M.; Cazzaniga, W.; Croft, F.; Kinsella, N.; Cahill, D.; James, N.D. Mobile Health Solutions for Prostate Cancer Diagnostics-A Systematic Review. Clin Pract 2023, 13, 863–872. [Google Scholar] [CrossRef]
- Duffy, M.J. Tumor Markers in Clinical Practice: A Review Focusing on Common Solid Cancers. Medical Principles and Practice 2013, 22, 4–11. [Google Scholar] [CrossRef]
- Jatho, A.; Mugisha, N.M.; Kafeero, J.; Holoya, G.; Okuku, F.; Niyonzima, N. Mobile cancer prevention and early detection outreach in Uganda: Partnering with communities toward bridging the cancer health disparities through "asset-based community development model". Cancer Medicine 2020, 9, 7317–7329. [Google Scholar] [CrossRef]
- Zhu, M.; Liang, Z.; Feng, T.; Mai, Z.; Jin, S.; Wu, L.; Zhou, H.; Chen, Y.; Yan, W. Up-to-Date Imaging and Diagnostic Techniques for Prostate Cancer: A Literature Review. Diagnostics (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Nasimi, H.; Madsen, J.S.; Zedan, A.H.; Malmendal, A.; Osther, P.J.S.; Alatraktchi, F.A. Protein biomarker detection in prostate cancer: A comprehensive review of electrochemical biosensors. Sensors and Actuators Reports 2023, 6. [Google Scholar] [CrossRef]
- Guliy, O.I.; Staroverov, S.A.; Dykman, L.A. Heat Shock Proteins in Cancer Diagnostics. Applied Biochemistry and Microbiology 2023, 59, 395–407. [Google Scholar] [CrossRef]
- Fenton, J.J.; Weyrich, M.S.; Durbin, S.; Liu, Y.; Bang, H.; Melnikow, J. Prostate-Specific Antigen-Based Screening for Prostate Cancer: Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2018, 319, 1914–1931. [Google Scholar] [CrossRef]
- Fenton, J.J.; Weyrich, M.S.; Durbin, S.; Liu, Y.; Bang, H.; Melnikow, J. Prostate-Specific Antigen-Based Screening for Prostate Cancer Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA-J. Am. Med. Assoc. 2018, 319, 1914–1931. [Google Scholar] [CrossRef]
- Singh, B.; Ma, S.L.; Hara, T.O.; Singh, S. Nanomaterials-Based Biosensors for the Detection of Prostate Cancer Biomarkers: Recent Trends and Future Perspective. Advanced Materials Technologies 2023, 8. [Google Scholar] [CrossRef]
- Liu, X.; Wang, D.; Chu, J.S.; Xu, Y.; Wang, W.J. Sandwich pair nanobodies, a potential tool for electrochemical immunosensing serum prostate-specific antigen with preferable specificity. Journal of Pharmaceutical and Biomedical Analysis 2018, 158, 361–369. [Google Scholar] [CrossRef]
- Kumar, V.; Kukkar, D.; Hashemi, B.; Kim, K.H.; Deep, A. Advanced Functional Structure-Based Sensing and Imaging Strategies for Cancer Detection: Possibilities, Opportunities, Challenges, and Prospects. Advanced Functional Materials 2019, 29. [Google Scholar] [CrossRef]
- Garg, S.; Sachdeva, A.; Peeters, M.; McClements, J. Point-of-Care Prostate Specific Antigen Testing: Examining Translational Progress toward Clinical Implementation. Acs Sensors 2023, 8, 3643–3658. [Google Scholar] [CrossRef]
- Panteghini, M. Implementation of standardization in clinical practice: not always an easy task. Clinical Chemistry and Laboratory Medicine 2012, 50, 1237–1241. [Google Scholar] [CrossRef]
- Ferraro, S.; Bussetti, M.; Rizzardi, S.; Braga, F.; Panteghini, M. Verification of Harmonization of Serum Total and Free Prostate-Specific Antigen (PSA) Measurements and Implications for Medical Decisions. Clinical Chemistry 2021, 67, 543–553. [Google Scholar] [CrossRef]
- Stenman, U.H.; Paus, E.; Allard, W.J.; Andersson, I.; Andrès, C.; Barnett, T.R.; Becker, C.; Belenky, A.; Bellanger, L.; Pellegrino, C.M.; et al. Summary report of the TD-3 workshop:: Characterization of 83 antibodies against prostate-specific antigen. Tumor Biology 1999, 20, 1–12. [Google Scholar] [CrossRef]
- Srinivasan, B.; Nanus, D.M.; Erickson, D.; Mehta, S. Highly portable quantitative screening test for prostate-specific antigen at point of care. Current Research in Biotechnology 2021, 3, 288–299. [Google Scholar] [CrossRef]
- Stephan, C.; Kramer, J.; Meyer, H.A.; Kristiansen, G.; Ziemer, S.; Deger, S.; Lein, M.; Loening, S.A.; Jung, K. Different prostate-specific antigen assays give different results on the same blood sample: an obstacle to recommending uniform limits for prostate biopsies. Bju International 2007, 99, 1427–1431. [Google Scholar] [CrossRef]
- Dukle, A.; Nathanael, A.J.; Panchapakesan, B.; Oh, T.H. Role of Paper-Based Sensors in Fight against Cancer for the Developing World. Biosensors-Basel 2022, 12. [Google Scholar] [CrossRef]
- Graves, H.C.; Wehner, N.; Stamey, T.A. Ultrasensitive Radioimmunoassay of Prostate-Specific Antigen. Clinical Chemistry 1992, 38, 735–742. [Google Scholar] [CrossRef]
- Myrtle, J.F.; Shackelford, W.; Bartholomew, R.M.; Wampler, J. PROSTATE-SPECIFIC ANTIGEN - QUANTITATION IN SERUM BY IMMUNORADIOMETRIC ASSAY. Clinical Chemistry 1983, 29, 1216–1216. [Google Scholar]
- Lang, Q.L.; Wang, F.; Yin, L.; Liu, M.J.; Petrenko, V.A.; Liu, A.H. Specific Probe Selection from Landscape Phage Display Library and Its Application in Enzyme-Linked Immunosorbent Assay of Free Prostate-Specific Antigen. Analytical Chemistry 2014, 86, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Vehniäinen, M.; Jansén, T.; Meretoja, V.; Saviranta, P.; Pettersson, K.; Lövgren, T. Dual-label time-resolved immunofluorometric assay of free and total prostate-specific antigen based on recombinant Fab fragments. Clinical Chemistry 2000, 46, 658–666. [Google Scholar] [CrossRef]
- Peltomaa, R.; Benito-Peña, E.; Barderas, R.; Moreno-Bondi, M.C. Phage Display in the Quest for New Selective Recognition Elements for Biosensors. Acs Omega 2019, 4, 11569–11580. [Google Scholar] [CrossRef] [PubMed]
- Kierny, M.R.; Cunningham, T.D.; Kay, B.K. Detection of biomarkers using recombinant antibodies coupled to nanostructured platforms. Nano Reviews & Experiments 2012, 3. [Google Scholar] [CrossRef]
- Ménez, R.; Michel, S.; Muller, B.H.; Bossus, M.; Ducancel, F.; Jolivet-Reynaud, C.; Stura, E.A. Crystal structure of a ternary complex between human prostate-specific antigen, its substrate acyl intermediate and an activating antibody. J Mol Biol 2008, 376, 1021–1033. [Google Scholar] [CrossRef]
- Petrenko, V.A.; Jayanna, P.K. Phage protein-targeted cancer nanomedicines. Febs Letters 2014, 588, 341–349. [Google Scholar] [CrossRef]
- Petrenko, V.A.; Smith, G.P.; Mazooji, M.M.; Quinn, T. Alpha-helically constrained phage display library. Protein Eng 2002, 15, 943–950. [Google Scholar] [CrossRef]
- Ji, S.; Lee, M.; Kim, D. Detection of early-stage prostate cancer by using a simple carbon nanotube@paper biosensor. Biosens. Bioelectron. 2018, 102, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Singh, J.; Goyat, R.; Saharan, Y.; Chaudhry, V.; Umar, A.; Ibrahim, A.A.; Akbar, S.; Ameen, S.; Baskoutas, S. Nanomaterials-based biosensor and their applications: A review. Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Pillay, T.S.; Muyldermans, S. Application of Single-Domain Antibodies ("Nanobodies") to Laboratory Diagnosis. Annals of Laboratory Medicine 2021, 41, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, C.; McNeil, C.J.; Rawson, K.; Nilsson, O. Disposable Noncompetitive Immunosensor for Free and Total Prostate-Specific Antigen Based on Capacitance Measurement. Analytical Chemistry 2004, 76, 5649–5656. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, Y.; Sadeghi, M.; Ehzari, H.; Derakhshankhah, H. Label-free electrochemical immunosensor based on antibody-immobilized Fe-Cu layered double hydroxide nanosheetas an electrochemical probe for the detection of ultra trace amount of prostate cancer biomarker (PSA). Microchemical Journal 2023, 195. [Google Scholar] [CrossRef]
- Nakhjavani, S.A.; Tokyay, B.K.; Soylemez, C.; Sarabi, M.R.; Yetisen, A.K.; Tasoglu, S. Biosensors for prostate cancer detection. Trends in Biotechnology 2023, 41, 1248–1267. [Google Scholar] [CrossRef] [PubMed]
- Saerens, D.; Frederix, F.; Reekmans, G.; Conrath, K.; Jans, K.; Brys, L.; Huang, L.; Bosmans, E.; Maes, G.; Borghs, G.; et al. Engineering camel single-domain antibodies and immobilization chemistry for human prostate-specific antigen sensing. Analytical Chemistry 2005, 77, 7547–7555. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P.; Petrenko, V.A. Phage display. Chemical Reviews 1997, 97, 391–410. [Google Scholar] [CrossRef]
- Smith, G.P. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Guliy, O.I.; Evstigneeva, S.S.; Dykman, L.A. Recombinant antibodies by phage display for bioanalytical applications. Biosens Bioelectron 2023, 222, 114909. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P. Phage Display: Simple Evolution in a Petri Dish (Nobel Lecture). Angew Chem Int Ed Engl 2019, 58, 14428–14437. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A.; Smith, G.P.; Gong, X.; Quinn, T. A library of organic landscapes on filamentous phage. Protein Eng 1996, 9, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A.; Smith, G.P. Phages from landscape libraries as substitute antibodies. Protein Eng 2000, 13, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A. Landscape phage as a molecular recognition interface for detection devices. Microelectronics Journal 2008, 39, 202–207. [Google Scholar] [CrossRef]
- Petrenko, V.A.; Vodyanoy, V.J. Phage display for detection of biological threat agents. J Microbiol Methods 2003, 53, 253–262. [Google Scholar] [CrossRef]
- Petrenko, V.A.; Sorokulova, I.B. Detection of biological threats. A challenge for directed molecular evolution. A challenge for directed molecular evolution. J Microbiol Methods 2004, 58, 147–168. [Google Scholar] [CrossRef]
- Nanduri, V.; Sorokulova, I.B.; Samoylov, A.M.; Simonian, A.L.; Petrenko, V.A.; Vodyanoy, V. Phage as a molecular recognition element in biosensors immobilized by physical adsorption. Biosens. Bioelectron. 2007, 22, 986–992. [Google Scholar] [CrossRef]
- Brigati, J.R.; Samoylova, T.I.; Jayanna, P.K.; Petrenko, V.A. Phage display for generating peptide reagents. Curr Protoc Protein Sci 2008, Chapter 18, Unit 18 19. [CrossRef]
- Kuzmicheva, G.A.; Jayanna, P.K.; Sorokulova, I.B.; Petrenko, V.A. Diversity and censoring of landscape phage libraries. Protein Engineering Design & Selection 2009, 22, 9–18. [Google Scholar]
- Petrenko, V.A.; Gillespie, J.W. Paradigm shift in bacteriophage-mediated delivery of anticancer drugs: from targeted "magic bullets' to self-navigated "magic missiles'. Expert Opinion on Drug Delivery 2017, 14, 373–384. [Google Scholar] [CrossRef]
- Horikawa, S.; Bedi, D.; Li, S.Q.; Shen, W.; Huang, S.C.; Chen, I.H.; Chai, Y.T.; Auad, M.L.; Bozack, M.J.; Barbaree, J.M.; et al. Effects of surface functionalization on the surface phage coverage and the subsequent performance of phage-immobilized magnetoelastic biosensors. Biosens. Bioelectron. 2011, 26, 2361–2367. [Google Scholar] [CrossRef]
- Scott, A.M.; Welt, S. Antibody-based immunological therapies. Current Opinion in Immunology 1997, 9, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Boon, T. Tumor antigens recognised by T lymphocytes. European Journal of Cancer 1999, 35, S216–S216. [Google Scholar] [CrossRef]
- Romanov, V.I.; Durand, D.B.; Petrenko, V.A. Phage display selection of peptides that affect prostate carcinoma cells attachment and invasion. Prostate 2001, 47, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Jayanna, P.K.; Deinnocentes, P.; Bird, R.C.; Petrenko, V.A. Landscape Phage Probes for PC3 Prostate Carcinoma cells. In Proceedings of the Nanotechnology Conference and Trade Show (Nanotech 2008), Boston, MA, USA, Jun 01-05 2008; pp. 457–460. [Google Scholar]
- Kuzmicheva, G.A.; Jayanna, P.K.; Eroshkin, A.M.; Grishina, M.A.; Pereyaslavskaya, E.S.; Potemkin, V.A.; Petrenko, V.A. Mutations in fd phage major coat protein modulate affinity of the displayed peptide. Protein Eng Des Sel 2009, 22, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Fagbohun, O.A.; Kazmierczak, R.A.; Petrenko, V.A.; Eisenstark, A. Metastatic prostate cancer cell-specific phage-like particles as a targeted gene-delivery system. J Nanobiotechnology 2013, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A.; Brigati, J.R. Phage as Bispecific Probes. In Immunoassay and Other Bioanalytical Techniques; Van Emon, J.M., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Popkov, M.; Rader, C.; Barbas, C.F. Isolation of human prostate cancer cell reactive antibodies using phage display technology. Journal of Immunological Methods 2004, 291, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Rader, C.; Ritter, G.; Nathan, S.; Elia, M.; Gout, I.; Jungbluth, A.A.; Cohen, L.S.; Welt, S.; Old, L.J.; Barbas, C.F. The rabbit antibody repertoire as a novel source for the generation of therapeutic human antibodies. Journal of Biological Chemistry 2000, 275, 13668–13676. [Google Scholar] [CrossRef]
- Stenman, U.H.; Leinonen, J.; Alfthan, H.; Rannikko, S.; Tuhkanen, K.; Alfthan, O. A COMPLEX BETWEEN PROSTATE-SPECIFIC ANTIGEN AND ALPHA-1-ANTICHYMOTRYPSIN IS THE MAJOR FORM OF PROSTATE-SPECIFIC ANTIGEN IN SERUM OF PATIENTS WITH PROSTATIC-CANCER - ASSAY OF THE COMPLEX IMPROVES CLINICAL SENSITIVITY FOR CANCER. Cancer Research 1991, 51, 222–226. [Google Scholar] [PubMed]
- Stamey, T.A.; Yang, N.; Hay, A.R.; McNeal, J.E.; Freiha, F.S.; Redwine, E. PROSTATE-SPECIFIC ANTIGEN AS A SERUM MARKER FOR ADENOCARCINOMA OF THE PROSTATE. New England Journal of Medicine 1987, 317, 909–916. [Google Scholar] [CrossRef]
- Rowe, E.W.J.; Laniado, M.E.; Walker, M.M.; Patel, A. Prostate cancer detection in men with a 'normal' total prostate-specific antigen (PSA) level using percentage free PSA: a prospective screening study. Bju International 2005, 95, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Brux, B.; Lein, M.; Rudolph, B.; Kristiansen, G.; Hauptmann, S.; Schnorr, D.; Loening, S.A.; Sinha, P. Molecular forms of prostate-specific antigen in malignant and benign prostatic tissue: Biochemical and diagnostic implications. Clinical Chemistry 2000, 46, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Q.; Zhang, Y.; Xu, H.G. Different free prostate-specific antigen to total prostate-specific antigen ratios using three detecting systems. Journal of Clinical Laboratory Analysis 2018, 32. [Google Scholar] [CrossRef]
- Lilja, H.; Oldbring, J.; Rannevik, G.; Laurell, C.B. SEMINAL VESICLE-SECRETED PROTEINS AND THEIR REACTIONS DURING GELATION AND LIQUEFACTION OF HUMAN-SEMEN. Journal of Clinical Investigation 1987, 80, 281–285. [Google Scholar] [CrossRef]
- Loeb, S.; Lilja, H.; Vickers, A. Beyond prostate-specific antigen: utilizing novel strategies to screen men for prostate cancer. Current Opinion in Urology 2016, 26, 459–465. [Google Scholar] [CrossRef]
- McJimpsey, E.L. Molecular Form Differences Between Prostate-Specific Antigen (PSA) Standards Create Quantitative Discordances in PSA ELISA Measurements. Scientific Reports 2016, 6. [Google Scholar] [CrossRef]
- Stephan, C.; Jung, K.; Lein, M.; Sinha, P.; Schnorr, D.; Loening, S.A. Molecular forms of prostate-specific antigen and human kallikrein 2 as promising tools for early diagnosis of prostate cancer. Cancer Epidemiology Biomarkers & Prevention 2000, 9, 1133–1147. [Google Scholar]
- Filella, X.; Truan, D.; Alcover, J.; Quintó, L.; Molina, R.; Luque, P.; Coca, F.; Ballesta, A.M. Comparison of several combinations of free, complexed, and total PSA in the diagnosis of prostate cancer in patients with urologic symptoms. Urology 2004, 63, 1100–1103. [Google Scholar] [CrossRef]
- Grossklaus, D.J.; Shappell, S.B.; Gautam, S.; Smith, J.A.; Cookson, M.S. RATIO OF FREE-TO-TOTAL PROSTATE SPECIFIC ANTIGEN CORRELATES WITH TUMOR VOLUME IN PATIENTS WITH INCREASED PROSTATE SPECIFIC ANTIGEN. The Journal of Urology 2001, 165, 455–458. [Google Scholar] [CrossRef]
- Jung, K.; Elgeti, U.; Lein, M.; Brux, B.; Sinha, P.; Rudolph, B.; Hauptmann, S.; Schnorr, D.; Loening, S.A. Ratio of Free or Complexed Prostate-specific Antigen (PSA) to Total PSA: Which Ratio Improves Differentiation between Benign Prostatic Hyperplasia and Prostate Cancer? Clinical Chemistry 2000, 46, 55–62. [Google Scholar] [CrossRef]
- Djavan, B.; Zlotta, A.; Kratzik, C.; Remzi, M.; Seitz, C.; Schulman, C.C.; Marberger, M. PSA, PSA density, PSA density of transition zone, free/total PSA ratio, and PSA velocity for early detection of prostate cancer in men with serum PSA 2.5 to 4.0 ng/ml. Urology 1999, 54, 517–522. [Google Scholar] [CrossRef]
- Ferrieu-Weisbuch, C.; Michel, S.; Collomb-Clerc, E.; Pothion, C.; Deléage, G.; Jolivet-Reynaud, C. Characterization of prostate-specific antigen binding peptides selected by phage display technology. Journal of Molecular Recognition 2006, 19, 10–20. [Google Scholar] [CrossRef]
- Wu, P.; Leinonen, J.; Koivunen, E.; Lankinen, H.; Stenman, U.H. Identification of novel prostate-specific antigen-binding peptides modulating its enzyme activity. European Journal of Biochemistry 2000, 267, 6212–6220. [Google Scholar] [CrossRef]
- Wu, P.; Zhu, L.; Stenman, U.H.; Leinonen, J. Immunopeptidometric assay for enzymatically active prostate-specific antigen. Clinical Chemistry 2004, 50, 125–129. [Google Scholar] [CrossRef]
- Koivunen, E.; Wang, B.C.; Dickinson, C.D.; Ruoslahti, E. PEPTIDES IN CELL-ADHESION RESEARCH. Extracellular Matrix Components 1994, 245, 346–369. [Google Scholar]
- Wang, Y.B.; Wang, M.Y.; Yu, H.P.; Wang, G.; Ma, P.X.; Pang, S.; Jiao, Y.M.; Liu, A.H. Screening of peptide selectively recognizing prostate-specific antigen and its application in detecting total prostate-specific antigen. Sensors and Actuators B-Chemical 2022, 367. [Google Scholar] [CrossRef]
- Muller, B.H.; Savatier, A.; L'Hostis, G.; Costa, N.; Bossus, M.; Michel, S.; Ott, C.; Becquart, L.; Ruffion, A.; Stura, E.A.; et al. In Vitro Affinity Maturation of an Anti-PSA Antibody for Prostate Cancer Diagnostic Assay. J Mol Biol 2011, 414, 545–562. [Google Scholar] [CrossRef]
- Han, L.; Xia, H.Q.; Yin, L.; Petrenko, V.A.; Liu, A.H. Selected landscape phage probe as selective recognition interface for sensitive total prostate-specific antigen immunosensor. Biosens. Bioelectron. 2018, 106, 1–6. [Google Scholar] [CrossRef]
- Kuzmicheva, G.A.; Jayanna, P.K.; Sorokulova, I.B.; Petrenko, V.A. Diversity and censoring of landscape phage libraries. Protein Eng Des Sel 2009, 22, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A.; Vodyanoy, V.J. Phage display for detection of biological threat agents. Journal of Microbiological Methods 2003, 53, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Knez, K.; Noppe, W.; Geukens, N.; Janssen, K.P.F.; Spasic, D.; Heyligen, J.; Vriens, K.; Thevissen, K.; Cammue, B.P.A.; Petrenko, V.; et al. Affinity Comparison of p3 and p8 Peptide Displaying Bacteriophages Using Surface Plasmon Resonance. Analytical Chemistry 2013, 85, 10075–10082. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, D.; Yan, L.; Petrenko, V.A.; Liu, A.H. Specific phages-based electrochemical impedimetric immunosensors for label-free and ultrasensitive detection of dual prostate-specific antigens. Sensors and Actuators B-Chemical 2019, 297. [Google Scholar] [CrossRef]
- Qi, H.; Wang, F.; Petrenko, V.A.; Liu, A. Peptide Microarray with Ligands at High Density Based on Symmetrical Carrier Landscape Phage for Detection of Cellulase. Analytical Chemistry 2014, 86, 5844–5850. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.R.; Kelly, K.A.; Mahmood, U.; Weissleder, R.; Deutscher, S.L. In vivo selection of phage for the optical imaging of PC-3 human prostate carcinoma in mice. Neoplasia 2006, 8, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Paramasivam, K.; Shen, Y.Z.; Yuan, J.S.; Waheed, I.; Mao, C.B.; Zhou, X. Advances in the Development of Phage-Based Probes for Detection of Bio-Species. Biosensors-Basel 2022, 12. [Google Scholar] [CrossRef]
- Petrenko, V.A. Landscape Phage: Evolution from Phage Display to Nanobiotechnology. Viruses-Basel 2018, 10. [Google Scholar] [CrossRef]
- Horikawa, S.; Bedi, D.; Li, S.; Shen, W.; Huang, S.; Chen, I.H.; Chai, Y.; Auad, M.L.; Bozack, M.J.; Barbaree, J.M.; et al. Effects of surface functionalization on the surface phage coverage and the subsequent performance of phage-immobilized magnetoelastic biosensors. Biosens Bioelectron 2011, 26, 2361–2367. [Google Scholar] [CrossRef]
- Huang, S.; Yang, H.; Lakshmanan, R.S.; Johnson, M.L.; Chen, I.; Wan, J.; Wikle, H.C.; Petrenko, V.A.; Barbaree, J.M.; Cheng, Z.Y.; et al. The effect of salt and phage concentrations on the binding sensitivity of magnetoelastic biosensors for Bacillus anthracis detection. Biotechnol Bioeng 2008, 101, 1014–1021. [Google Scholar] [CrossRef]
- Sorokulova, I.B.; Olsen, E.V.; Chen, I.H.; Fiebor, B.; Barbaree, J.M.; Vodyanoy, V.J.; Chin, B.A.; Petrenko, V.A. Landscape phage probes for Salmonella typhimurium. Journal of Microbiological Methods 2005, 63, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P. Phage Display: Simple Evolution in a Petri Dish (Nobel Lecture). Angew. Chem.-Int. Edit. 2019, 58, 14428–14437. [Google Scholar] [CrossRef]
- Bhasin, A.; Drago, N.P.; Majumdar, S.; Sanders, E.C.; Weiss, G.A.; Penner, R.M. Viruses Masquerading as Antibodies in Biosensors: The Development of the Virus BioResistor. Accounts Chem Res 2020, 53, 2384–2394. [Google Scholar] [CrossRef] [PubMed]
- Jayanna, P.K.; Bedi, D.; Deinnocentes, P.; Bird, R.C.; Petrenko, V.A. Landscape phage ligands for PC3 prostate carcinoma cells. Protein Eng Des Sel 2010, 23, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Jayanna, P.K.; Bedi, D.; Gillespie, J.W.; DeInnocentes, P.; Wang, T.; Torchilin, V.P.; Bird, R.C.; Petrenko, V.A. Landscape phage fusion protein-mediated targeting of nanomedicines enhances their prostate tumor cell association and cytotoxic efficiency. Nanomedicine 2010, 6, 538–546. [Google Scholar] [CrossRef]
- Lang, Q.; Wang, F.; Yin, L.; Liu, M.; Petrenko, V.A.; Liu, A. Specific probe selection from landscape phage display library and its application in enzyme-linked immunosorbent assay of free prostate-specific antigen. Anal Chem 2014, 86, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Liu, P.; Petrenko, V.A.; Liu, A. A Label-Free Electrochemical Impedance Cytosensor Based on Specific Peptide-Fused Phage Selected from Landscape Phage Library. Sci Rep 2016, 6, 22199. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Han, L.; Wang, F.; Petrenko, V.A.; Liu, A. Gold nanoprobe functionalized with specific fusion protein selection from phage display and its application in rapid, selective and sensitive colorimetric biosensing of Staphylococcus aureus. Biosens Bioelectron 2016, 82, 195–203. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.B.; Han, L.; Cai, Y.Y.; Ren, H.; Ma, T.X.; Li, X.Q.; Petrenko, V.A.; Liu, A.H. Colorimetric Assay of Bacterial Pathogens Based on Co3O4 Magnetic Nanozymes Conjugated with Specific Fusion Phage Proteins and Magnetophoretic Chromatography. ACS Appl. ACS Appl. Mater. Interfaces 2020, 12, 9090–9097. [Google Scholar] [CrossRef]
- Qi, H.; Lu, H.; Qiu, H.J.; Petrenko, V.; Liu, A. Phagemid vectors for phage display: properties, characteristics and construction. J Mol Biol 2012, 417, 129–143. [Google Scholar] [CrossRef]
- Qi, H.; Wang, F.; Petrenko, V.A.; Liu, A. Peptide microarray with ligands at high density based on symmetrical carrier landscape phage for detection of cellulase. Anal Chem 2014, 86, 5844–5850. [Google Scholar] [CrossRef]
- Wang, F.; Liu, P.; Sun, L.; Li, C.; Petrenko, V.A.; Liu, A. Bio-mimetic nanostructure self-assembled from Au@Ag heterogeneous nanorods and phage fusion proteins for targeted tumor optical detection and photothermal therapy. Sci Rep 2014, 4, 6808. [Google Scholar] [CrossRef]
- Yin, L.; Luo, Y.; Liang, B.; Wang, F.; Du, M.; Petrenko, V.A.; Qiu, H.J.; Liu, A. Specific ligands for classical swine fever virus screened from landscape phage display library. Antiviral Res 2014, 109, 68–71. [Google Scholar] [CrossRef]
- Han, L.; Liu, P.; Petrenko, V.A.; Liu, A.H. A Label-Free Electrochemical Impedance Cytosensor Based on Specific Peptide-Fused Phage Selected from Landscape Phage Library. Sci Rep 2016, 6, 10. [Google Scholar] [CrossRef]
- Shen, W.; Li, S.; Park, M.-K.; Zhang, Z.; Cheng, Z.; Petrenko, V.A.; Chin, B.A. Blocking Agent Optimization for Nonspecific Binding on Phage Based Magnetoelastic Biosensors. Journal of The Electrochemical Society 2012, 159, B818. [Google Scholar] [CrossRef]
- Li, S.; Lakshmanan, R.S.; Petrenko, V.A.; Chin, B.A.; Blois, H.; Cao, B.; Chin, B.; Deutscher, S.L.; Gea, M.; Iris, F.; et al. Phage-based Pathogen Biosensors. In Phage Nanobiotechnology, Petrenko, V., Smith, G.P., O'Brien, P., Craighead, H., Kroto, H., Eds.; The Royal Society of Chemistry: 2011; p. 0.
- Lakshmanan, R.S.; Guntupalli, R.; Hu, J.; Kim, D.J.; Petrenko, V.A.; Barbaree, J.M.; Chin, B.A. Phage immobilized magnetoelastic sensor for the detection of Salmonella typhimurium. J Microbiol Methods 2007, 71, 55–60. [Google Scholar] [CrossRef]
- Fu, L.; Li, S.; Zhang, K.; Chen, I.-H.; Petrenko, V.A.; Cheng, Z. Magnetostrictive Microcantilever as an Advanced Transducer for Biosensors. Sensors 2007, 7, 2929–2941. [Google Scholar] [CrossRef]
- Han, L.; Xia, H.; Yin, L.; Petrenko, V.A.; Liu, A. Selected landscape phage probe as selective recognition interface for sensitive total prostate-specific antigen immunosensor. Biosensors and Bioelectronics 2018, 106, 1–6. [Google Scholar] [CrossRef]
- Ferraro, S.; Bussetti, M.; Panteghini, M. Serum Prostate-Specific Antigen Testing for Early Detection of Prostate Cancer: Managing the Gap between Clinical and Laboratory Practice. Clinical Chemistry 2021, 67, 602–609. [Google Scholar] [CrossRef]
- Sandúa, A.; Sanmamed, M.F.; Rodríguez, M.; Ancizu-Marckert, J.; Gúrpide, A.; Perez-Gracia, J.L.; Alegre, E.; González, A. PSA reactivity in extracellular microvesicles to commercial immunoassays. Clinica Chimica Acta 2023, 543. [Google Scholar] [CrossRef]
- Liu, A.; Zhao, F.; Zhao, Y.; Shangguan, L.; Liu, S. A portable chemiluminescence imaging immunoassay for simultaneous detection of different isoforms of prostate specific antigen in serum. Biosensors and Bioelectronics 2016, 81, 97–102. [Google Scholar] [CrossRef]
- Andreeva, I.P.; Grigorenko, V.G.; Egorov, A.M.; Osipov, A.P. Quantitative Lateral Flow Immunoassay for Total Prostate Specific Antigen in Serum. Analytical Letters 2016, 49, 579–588. [Google Scholar] [CrossRef]
- Barbosa, A.I.; Castanheira, A.P.; Edwards, A.D.; Reis, N.M. A lab-in-a-briefcase for rapid prostate specific antigen (PSA) screening from whole blood. Lab on a Chip 2014, 14, 2918–2928. [Google Scholar] [CrossRef]
- Woodrum, D.L.; French, C.M.; Hill, T.M.; Roman, S.J.; Slatore, H.L.; Shaffer, J.L.; York, L.G.; Eure, K.L.; Loveland, K.G.; Gasior, G.H.; et al. Analytical performance of the Tandem®-R free PSA immunoassay measuring free prostate-specific antigen. Clinical Chemistry 1997, 43, 1203–1208. [Google Scholar] [CrossRef]
- Galkin, A.; Komar, A.; Gorshunov, Y.; Besarab, A.; Soloviova, V. NEW MONOCLONAL ANTIBODIES TO THE PROSTATE-SPECIFIC ANTIGEN: OBTAINING AND STUDYING BIOLOGICAL PROPERTIES. Journal of Microbiology Biotechnology and Food Sciences 2019, 9, 573–577. [Google Scholar] [CrossRef]
- Zhang, X.M.; Soori, G.; Dobleman, T.J.; Xiao, G.G. The application of monoclonal antibodies in cancer diagnosis. Expert Review of Molecular Diagnostics 2014, 14, 97–106. [Google Scholar] [CrossRef]
- Lee, S.; Xie, J.; Chen, X.Y. Peptide-Based Probes for Targeted Molecular Imaging. Biochemistry 2010, 49, 1364–1376. [Google Scholar] [CrossRef]
- Han, L.; Liu, P.; Petrenko, V.A.; Liu, A.H. A Label-Free Electrochemical Impedance Cytosensor Based on Specific Peptide-Fused Phage Selected from Landscape Phage Library. Scientific Reports 2016, 6. [Google Scholar] [CrossRef]
- Brigati, J.; Williams, D.D.; Sorokulova, I.B.; Nanduri, V.; Chen, I.H.; Turnbough, C.L., Jr.; Petrenko, V.A. Diagnostic probes for Bacillus anthracis spores selected from a landscape phage library. Clin Chem 2004, 50, 1899–1906. [Google Scholar] [CrossRef]
- Knez, K.; Noppe, W.; Geukens, N.; Janssen, K.P.; Spasic, D.; Heyligen, J.; Vriens, K.; Thevissen, K.; Cammue, B.P.; Petrenko, V.; et al. Affinity comparison of p3 and p8 peptide displaying bacteriophages using surface plasmon resonance. Anal Chem 2013, 85, 10075–10082. [Google Scholar] [CrossRef]
- Smith, G.P.; Petrenko, V.A.; Matthews, L.J. Cross-linked filamentous phage as an affinity matrix. J Immunol Methods 1998, 215, 151–161. [Google Scholar] [CrossRef]
- Goldberg, M.E.; Djavadi-Ohaniance, L. Methods for measurement of antibody/antigen affinity based on ELISA and RIA. Current Opinion in Immunology 1993, 5, 278–281. [Google Scholar] [CrossRef]
- Kubota, S.; Kawaki, H.; Takigawa, M. ELISA of CCN Family Proteins in Body Fluids Including Serum and Plasma. In CCN Proteins: Methods and Protocols; Takigawa, M., Ed.; Springer New York: New York, NY, 2017; pp. 127–138. [Google Scholar]
- Arévalo, F.J.; González-Techera, A.; Zon, M.A.; González-Sapienza, G.; Fernández, H. Ultra-sensitive electrochemical immunosensor using analyte peptidomimetics selected from phage display peptide libraries. Biosens. Bioelectron. 2012, 32, 231–237. [Google Scholar] [CrossRef]
- Luo, Z.B.; Qi, Q.G.; Zhang, L.J.; Zeng, R.J.; Su, L.S.; Tang, D.P. Branched Polyethylenimine-Modified Upconversion Nanohybrid-Mediated Photoelectrochemical Immunoassay with Synergistic Effect of Dual-Purpose Copper Ions. Analytical Chemistry 2019, 91, 4149–4156. [Google Scholar] [CrossRef]
- Yu, Z.Z.; Tang, Y.; Cai, G.N.; Ren, R.R.; Tang, D.P. Paper Electrode-Based Flexible Pressure Sensor for Point-of-Care Immunoassay with Digital Multimeter. Analytical Chemistry 2019, 91, 1222–1226. [Google Scholar] [CrossRef]
- Achi, F.; Attar, A.M.; Lahcen, A.A. Electrochemical nanobiosensors for the detection of cancer biomarkers in real samples: Trends and challenges. Trac-Trends in Analytical Chemistry 2024, 170. [Google Scholar] [CrossRef]
- Guo, X.F.; Kulkarni, A.; Doepke, A.; Halsall, H.B.; Iyer, S.; Heineman, W.R. Carbohydrate-Based Label-Free Detection of Escherichia coli ORN 178 Using Electrochemical Impedance Spectroscopy. Analytical Chemistry 2012, 84, 241–246. [Google Scholar] [CrossRef]
- Daniels, J.S.; Pourmand, N. Label-free impedance biosensors: Opportunities and challenges. Electroanalysis 2007, 19, 1239–1257. [Google Scholar] [CrossRef]
- Li, L.; Zhang, S.P.; Yu, L.Z.; Zhang, W.Z.; Wei, Y.; Feng, D.X. Electrochemical Immunosensor for Detection of Prostate Specific Antigen Based on CNSs/Thi@AuNPs Nanocomposites as Sensing Platform. International Journal of Electrochemical Science 2022, 17. [Google Scholar] [CrossRef]
- Hou, L.; Tang, Y.; Xu, M.; Gao, Z.; Tang, D. Tyramine-Based Enzymatic Conjugate Repeats for Ultrasensitive Immunoassay Accompanying Tyramine Signal Amplification with Enzymatic Biocatalytic Precipitation. Analytical Chemistry 2014, 86, 8352–8358. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a): Sandwich PSA ELISA (A) vs Phage PSA ELISA (B): The capture antibody (A), or phage (B) immobilized onto ELISA plates bind the analyte protein, and detection antibodies linked to enzyme are added to catalyze the appearance of a colored or fluorescent product. (b): PSA biosensors. Molecular interface linked to a transducer binds the analyte and generates a signal: changes in mass, capacitance, resistance, surface plasmon resonance, reflectometric interference, etc.
Figure 1.
(a): Sandwich PSA ELISA (A) vs Phage PSA ELISA (B): The capture antibody (A), or phage (B) immobilized onto ELISA plates bind the analyte protein, and detection antibodies linked to enzyme are added to catalyze the appearance of a colored or fluorescent product. (b): PSA biosensors. Molecular interface linked to a transducer binds the analyte and generates a signal: changes in mass, capacitance, resistance, surface plasmon resonance, reflectometric interference, etc.
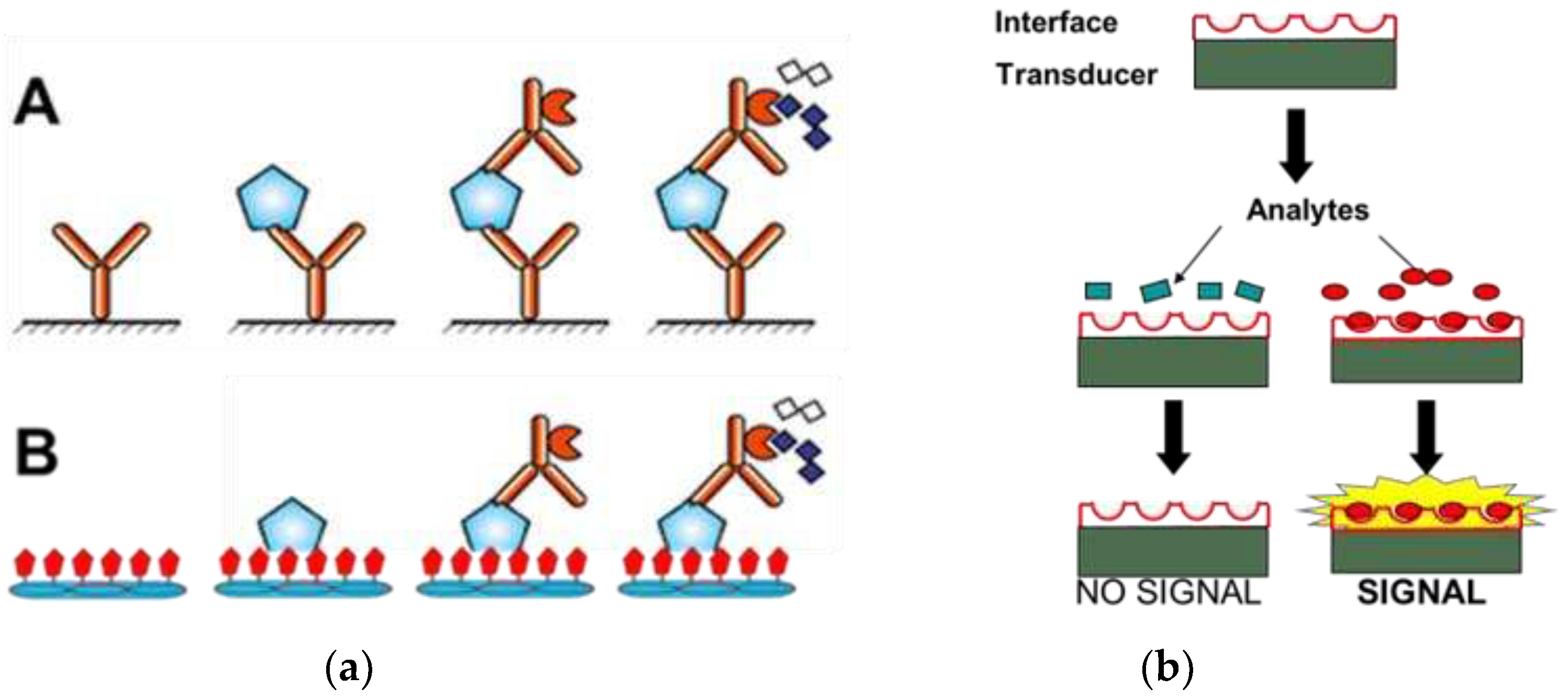
Figure 2.
Overall view of the complex structure of Fab′ 8G8F5 (H- blue, L – red) with –PSA (cyan)-substrate (magenta). Adapted from [30]. PDB ID: 2ZCK.
Figure 2.
Overall view of the complex structure of Fab′ 8G8F5 (H- blue, L – red) with –PSA (cyan)-substrate (magenta). Adapted from [30]. PDB ID: 2ZCK.

Figure 3.
Left: Electron microscopy image of the wild-type phage fd. Blue and red arrows depict the sharp and blunt ends of the phage capsid with attached minor coat proteins p3/p6 and p7/p9, respectively (five copies each). Major coat protein p8 (~2700 copies) forms the tubular capsid around viral single-stranded DNA (scale bar: 100 nm). Right: Peptide phage-displayed libraries. There are two essential types of phage display — display in the minor coat protein p3, and display in the major coat protein p8. . P3 type display is used when the desired product is the displayed peptide or antibody. In the alternative type 8 phage display, a fragment of foreign DNA is inserted into the gene VIII and a foreign peptide is fused to every pVIII subunit. In the p8 display, the whole phage virion is the goal of the discovery. Adapted with modifications from[31].
Figure 3.
Left: Electron microscopy image of the wild-type phage fd. Blue and red arrows depict the sharp and blunt ends of the phage capsid with attached minor coat proteins p3/p6 and p7/p9, respectively (five copies each). Major coat protein p8 (~2700 copies) forms the tubular capsid around viral single-stranded DNA (scale bar: 100 nm). Right: Peptide phage-displayed libraries. There are two essential types of phage display — display in the minor coat protein p3, and display in the major coat protein p8. . P3 type display is used when the desired product is the displayed peptide or antibody. In the alternative type 8 phage display, a fragment of foreign DNA is inserted into the gene VIII and a foreign peptide is fused to every pVIII subunit. In the p8 display, the whole phage virion is the goal of the discovery. Adapted with modifications from[31].

Figure 4.
Vectors and libraries. In the nucleotide sequences corresponding to the part of recombinant gene gpVIII encoding the N-terminal part of the major coat protein, randomized structures are designated as nnk, where n = A, T, G, or C, and k = G or T. Restriction sites for PstI and BamHI are underlined. N-terminal amino acid structures of mature recombinant pVIII proteins in libraries indicated by capital single letters according to amino acid abbreviations. Randomized amino acids are designated in small letters (a-h in the f8/8 library and a-i in the f8/9 library). Amino acids are numbered as in vector phage f8-5 .[32].
Figure 4.
Vectors and libraries. In the nucleotide sequences corresponding to the part of recombinant gene gpVIII encoding the N-terminal part of the major coat protein, randomized structures are designated as nnk, where n = A, T, G, or C, and k = G or T. Restriction sites for PstI and BamHI are underlined. N-terminal amino acid structures of mature recombinant pVIII proteins in libraries indicated by capital single letters according to amino acid abbreviations. Randomized amino acids are designated in small letters (a-h in the f8/8 library and a-i in the f8/9 library). Amino acids are numbered as in vector phage f8-5 .[32].

Figure 6.
Selectivity and specificity of phage probes. Phage probes selected from preliminary screening assays were incubated with target PC3 cells, control cells or serum treated wells of a 96-well cell culture plate. Phage associated with cells or serum were titered in bacteria and the ratio of phage output to phage input was expressed as recovery % to obtain the measure of the selectivity of a particular clone. The % recovery of the control phage bearing unrelated peptide relative to selected phage probe was indicative of the probe’s specificity. Results are the average of three replicates. A – f8/8 library, B – f8/9 library. zdapted from [58,99].
Figure 6.
Selectivity and specificity of phage probes. Phage probes selected from preliminary screening assays were incubated with target PC3 cells, control cells or serum treated wells of a 96-well cell culture plate. Phage associated with cells or serum were titered in bacteria and the ratio of phage output to phage input was expressed as recovery % to obtain the measure of the selectivity of a particular clone. The % recovery of the control phage bearing unrelated peptide relative to selected phage probe was indicative of the probe’s specificity. Results are the average of three replicates. A – f8/8 library, B – f8/9 library. zdapted from [58,99].

Figure 7.
immunofluorescence microscopic demonstration of phage EPTHSWAT interaction with PC-3M cells at 15 min and 1 h in comparison with the control non-relevant phage VPEGAFSS. Adapted with modifications from [60].
Figure 7.
immunofluorescence microscopic demonstration of phage EPTHSWAT interaction with PC-3M cells at 15 min and 1 h in comparison with the control non-relevant phage VPEGAFSS. Adapted with modifications from [60].

Figure 8.
Schematic illustration of bio panning for t-PSA (f-PSA and PSA-ACT). The f8/8 landscape phage library was added to the dishes with different immobilized forms of PSA. Unbound phages were washed away, and bound phages were eluted and used as a sub-library in the next round of biopanning. After three rounds, the individual phage clones were propagated, and their DNA segments corresponding to gpVIII were sequenced to determine corresponding phage-displayed peptide sequences. Detailed procedures can be found in [26,51,62].
Figure 8.
Schematic illustration of bio panning for t-PSA (f-PSA and PSA-ACT). The f8/8 landscape phage library was added to the dishes with different immobilized forms of PSA. Unbound phages were washed away, and bound phages were eluted and used as a sub-library in the next round of biopanning. After three rounds, the individual phage clones were propagated, and their DNA segments corresponding to gpVIII were sequenced to determine corresponding phage-displayed peptide sequences. Detailed procedures can be found in [26,51,62].

Figure 9.
The phages can be conjugated to the gold electrode surface using carbodiimide chemistry (A), or immobilized to the gold sensor through physical adsorption [50] (B) and analyzed by atomic force microscope (A) or electron microscopy (B). As shown in Figure 1B, a mass of filamentous phages were observed on the gold surface. The phages or phages bundles were covalently attached to the gold surface to generate an intercrossing random network. Adapted with modifications from [50,88].
Figure 9.
The phages can be conjugated to the gold electrode surface using carbodiimide chemistry (A), or immobilized to the gold sensor through physical adsorption [50] (B) and analyzed by atomic force microscope (A) or electron microscopy (B). As shown in Figure 1B, a mass of filamentous phages were observed on the gold surface. The phages or phages bundles were covalently attached to the gold surface to generate an intercrossing random network. Adapted with modifications from [50,88].

Figure 10.
Specificity of Phage Binding to f-PSA and t-PSA. The specificity (or selectivity) of discovered phages, as their ability to distinguish f-PSA from t-PSA and other cancer biomarkers, was established using phage capture assay. The protein samples were incubated separately in the wells of the 96-well plate. After blocking and washing of the wells, candidate phages were added and incubated with the biomarkers at 4 °C overnight. Wells were washed to remove unbound phages. Then bound phages were eluted and tittered, and the phage recovery was calculated to compare the captured phages by different target Specificity of different phage binding to different targets.: Adapted from [26,84].
Figure 10.
Specificity of Phage Binding to f-PSA and t-PSA. The specificity (or selectivity) of discovered phages, as their ability to distinguish f-PSA from t-PSA and other cancer biomarkers, was established using phage capture assay. The protein samples were incubated separately in the wells of the 96-well plate. After blocking and washing of the wells, candidate phages were added and incubated with the biomarkers at 4 °C overnight. Wells were washed to remove unbound phages. Then bound phages were eluted and tittered, and the phage recovery was calculated to compare the captured phages by different target Specificity of different phage binding to different targets.: Adapted from [26,84].
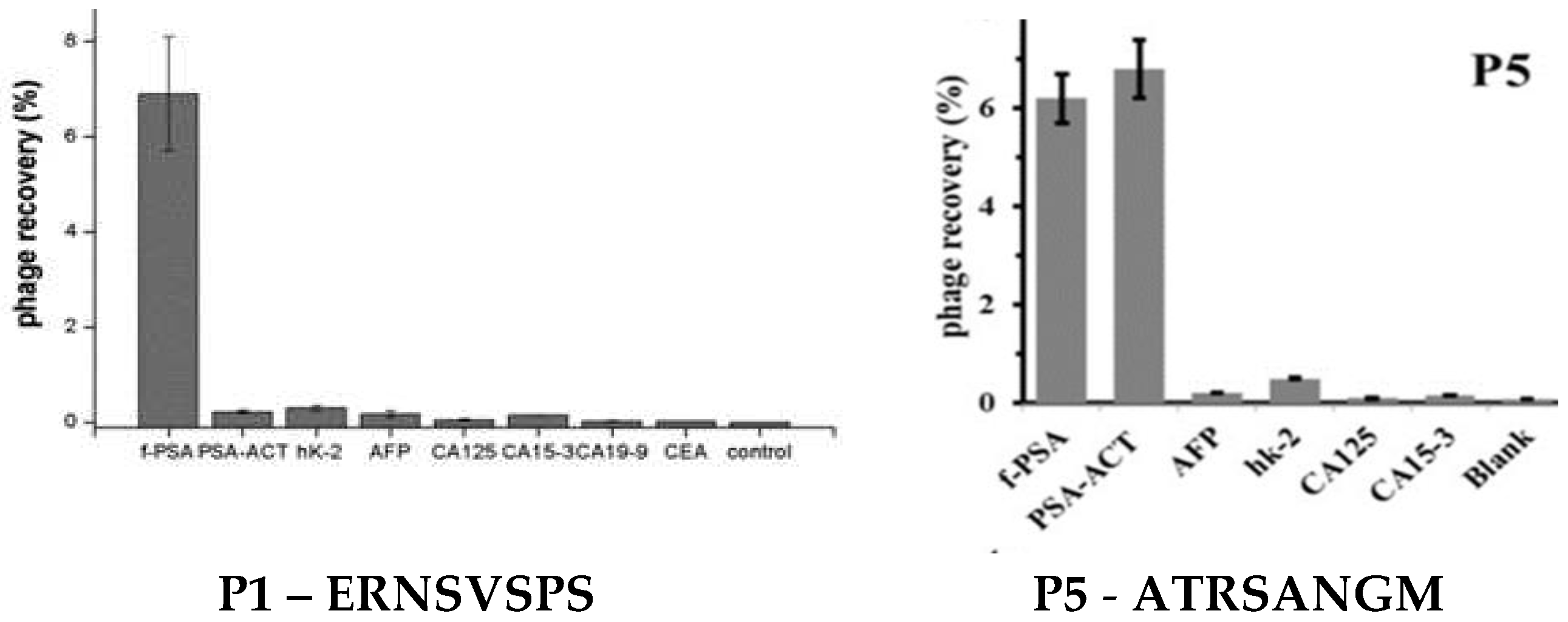
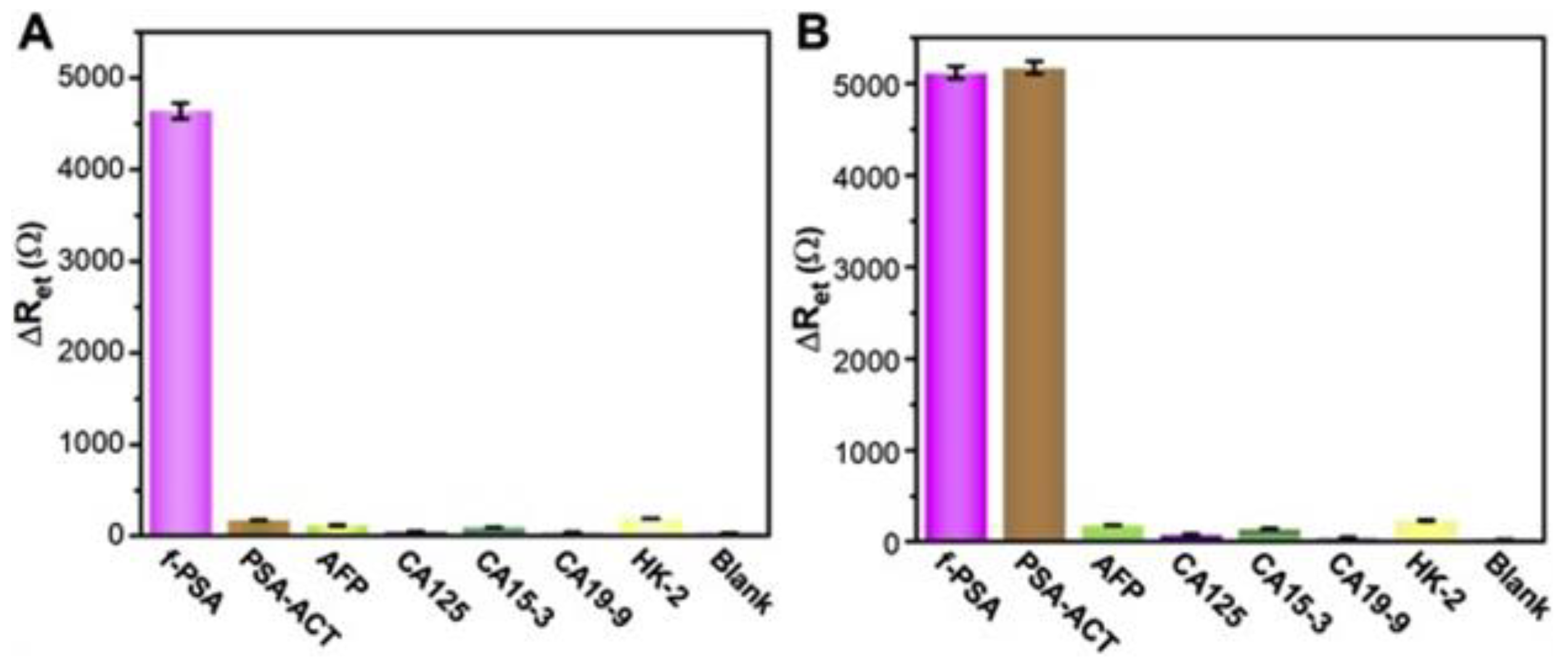
Figure 11.
The selectivity assay of P1- ERNSVSPS -immunosensor (A) and P5- ATRSANGM immunosensor (B) for f-PSA and t-PSA (total of f-PSA and PSA-ACT) in comparison with other common cancer biomarkers (AFP, CA125, CA15-3, CA19-9 and hK-2) in the serum as controls. The samples were dropped onto the phage-covered immunosensors., and incubated for same time. EIS assay was performed for the above immunosensors as described [88].
Figure 11.
The selectivity assay of P1- ERNSVSPS -immunosensor (A) and P5- ATRSANGM immunosensor (B) for f-PSA and t-PSA (total of f-PSA and PSA-ACT) in comparison with other common cancer biomarkers (AFP, CA125, CA15-3, CA19-9 and hK-2) in the serum as controls. The samples were dropped onto the phage-covered immunosensors., and incubated for same time. EIS assay was performed for the above immunosensors as described [88].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



