
Submitted:
01 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
Comparing to males, females require much more morphine to produce comparable levels of analgesia, suggesting a difference in the endogenous hyperalgesia and analgesia modulation between sexes. Aside from hormones, emerging evidence suggests sex-differential intrinsic neural regulation of pain generation and maintenance. According to the International Association for the Study of Pain (IASP) and American College of Gastroenterology (ACG), up to 25 % of the population having visceral pain at any one time, and in the United States 10-15 percent of adults suffering from irritable bowel syndrome (IBS). Here we analyzed literatures on clinical reports of sex differences in visceral pain focusing on IBS, other form of bowel dysfunction and comorbidities, and summarized animal models that provided means to investigate the underlying molecular mechanisms in sexual dimorphism of visceral pain. Neurons and nonneuronal cells (glia and immune cells) in the peripheral and central nervous systems all contribute to sex-dependent nociception and nociplasticity in the painful signal processing. Emotion is another factor in pain perception and appears to have sexual dimorphism.
Keywords:
pain
; visceral
; sex
; clinical
; preclinical
; molecule
Introduction
Defined by The International Association for the Study of Pain (IASP), 2019, chronic pain syndromes are classified into 7 categories: chronic primary pain, chronic cancer pain, chronic posttraumatic and postsurgical pain, chronic neuropathic pain, chronic headache and orofacial pain, chronic visceral pain, and chronic musculoskeletal pain [1]. Patients with chronic visceral pain and irritable bowel syndrome (IBS) also often experience comorbidities including psychiatric disorders and somatic pain [2,3,4]. Visceral and somatic pain derived from numerous factors such as inflammation, injury, and/or metabolic disorders are unpleasant experience in human and animals that greatly affect well-beings and reduce quality of life. Sexual dimorphism of pain has been documented extensively that are influenced by an intricate interplay of biological, psychological, and social factors [5,6,7,8,9,10,11,12,13,14,15,16]. Historically, the concept of sexual dimorphism in pain was rooted more in anecdotal evidence and cultural norms than in empirical research, dating back to the 1800s, largely based on clinical observations and cultural perceptions [17]. The rise of gender equity and gender-specific medicine marks a pivotal shift in pain research, especially the women’s health movement including advocates like Barbara Seaman (1935-2008) catalyzes a deeper exploration into gender differences in health and disease, which leads to initiatives to improve the management of chronic diseases including chronic pain in a gender-based fashion [18,19].
Pain perception, a term used in human, is a complex sensory and emotional experience. Due to a variety of psychosocial and culture elements, sexual dimorphism of pain in human has been documented with contradictory outcomes [12,20]. A recent review by Mogil JS [12] has summarized a great deal of examples that demonstrate the influence on pain perception by the testing environment. For example, women have higher pain ratings tested by men while men report less pain when tested by women. These types of sex biases are replicated by additional groups but not by all studies. Imbalanced social status and peer pressure between participants and experimenters also affect the outcomes of pain ratings. In addition, “tough” boys and “brave” man occur in pain tests [12,20]. In terms of visceral pain, a recent review by Kim YS and Kim N describes that IBS with constipation (C-IBS) is more prevalent in women than men and the treatment efficacy of IBS in men and women is also different, however, relatively few men are enrolled in most relevant clinical trials [21]. Diarrhea predominant IBS (IBS-D) appears more prevalent in men than women [8]. While, cultural differences in certain countries may affect the global epidemiological outcomes in sex/gender differences in IBS [11]. With the rising societal issues in opioid crisis in treatment of pain, more objective and better diagnosis are essential in the clinical settings. Recent scientific endeavors that increasingly focus on elucidating the biological underpinnings of these sex-specific differences can pave a way toward a better understanding of the pain process and suggest molecular targets for unbiased treatment approaches.
Peripheral nociceptor activation is a root cause of pain that are classified as nociceptive pain, neuropathic pain and nociplastic pain. The nociceptive pain, including visceral pain and somatic pain, are types of pain that occur when pain receptors in the tissues are activated by tissue inflammation or injury. The neuropathic pain are results of nerve impairment. The nociplastic pain, on the other hand, are functional pain disorders without apparent external tissue damage. The generation and maintenance of pain involve sensitization of primary afferent pathways including alterations of the activity of sensory neurons (nociceptors) and central sensitization, with participation of glial and immune cells. The dorsal root ganglia (DRG) contains pain-sensing neurons to convey sensory signals from the peripheral organs (e.g., the pelvic organs and limbs) to the central nervous system (CNS). In general, DRG neurons are classified, in a broad category, into small, medium and large neurons with the small neurons participating in promotion of pain sensation. Changes in the neurochemical coding in sensory neurons and synaptic activity in the CNS, modulated by nonneuronal cells, are fundamental process in pain initiation and persistence. Emerging evidence point out that molecular mechanisms other than the female sex hormone may underlie the sexual dimorphism of pain regulation since different stages of the estrus cycle have no greater impacts on nociception and analgesic responsiveness in females than that occurring intrinsically in males [22,23]. Recent progress derived from experimental animals shed lights on the role of sensory neuron subtypes in visceral and somatic pain sexual dimorphism [24,25,26]. In this review, we focus on the clinical and laboratory findings in visceral pain sexual dimorphisms and their comorbidities and summarize the as yet discovered potential underlying sensory biological mechanisms.
Clinical Findings in Visceral Pain Sexual Dimorphism
Comparison of colonic functionality in adult men and women demonstrates a significant effect of sex on rectal discomfort threshold, in which noxious sigmoid stimulation causes the most sensitivity in women with IBS and the least sensitivity in healthy women when compared to men with respective health conditions, while there are no significant differences in rectal discomfort thresholds between healthy men and men with IBS [5], inferring less sensitive perception of noxious stimulation in men with or without IBS. However, a recent study where 280 young healthy men and women (equal numbers) undergo rectal balloon distensions to measure visceral sensory and pain thresholds shows no sex differences nor IBS-related risk factors [27]. Another study comprising equal ratio of males and females in IBS and healthy groups (70% and 72% are women in each group) demonstrate that patients with IBS have lower sensory thresholds to rectal distention compared with healthy subjects, whereas the threshold for first sensation is similar in both groups [28], appearing as that sex composition in both groups does not skew the results. An internet-based nationwide survey of 88,607 people including 44815 male (50.6%) and 42176 female (47.6%) in the US to assess the prevalence of IBS and comorbidities in a 7-day recall period covers a variety of pain modalities and symptoms such as anal pain, belly pain or discomfort, bladder pain, bloating, bowel incontinence, constipation, diarrhea, difficulty swallowing, excess gas, heartburn or acid reflux/regurgitation, nausea or vomiting, pelvic pain, rectal pain, revealing that 6.1% met Rome IV IBS criteria, among which women (1.39-2.06 fold higher comparing to men) and every day smokers (1.32-1.73 fold higher comparing to nonsmokers) have higher odds for having all 3 IBS subtypes (IBS-M, IBS-C, IBS-D). Many comorbidities are associated with increased odds for the 3 subtypes, including prior gastroenteritis, fibromyalgia, gallstones, peptic ulcer disease, and thyroid disorder, and diabetic people tends to have IBS-D and IBS-M but not IBS-C [29].
For disease progression, meta-data analysis points to that woman tends to report IBS-C than man, while man has a higher reporting rate of IBS-D than woman [30,31]. Another analysis reports that female sex, younger age, and psychological distress during infectious enteritis are risking factors in the development of post-infectious (PI) IBS [32]. A study in 231 patients (131 males and 100 females) identified with acute gastroenteritis adds additional risking factors including longer duration of antibiotics use in PI-IBS, and the percentages of developing PI-IBS assessed by phone interview are 10.7% (14 out of 131) males and 20% (20 out of 100) females [33]. However, two IBS patient cohorts with 231 patients in Rome II and 141 patients in Rome III category who undergo rectal barostat testing in combination with questionnaires for anxiety, depression, somatization, and abuse demonstrate that pain threshold in Rome II cohort is positively associated with female gender, while pain threshold in Rome III cohort is positively associated with male gender [34]. The conflicting results are affected by a variety of factors such as hormonal levels altered in aging, psychological pain perception threshold, early life adversity, availability of medical cares, diet selections, societal roles, tendency to report, and geographical and cultural differences [11,21,35,36]. As for the role of sex hormones in visceral pain and pain at large, numerous original research and review articles have acknowledged them [15,37,38,39,40], however, the specific modulatory effects of sex hormones in pain as initiators, exacerbators, or endogenous protectors require more detailed investigation. Transgender patients who have pain-related conditions such as headaches, fibromyalgia, temporomandibular (TMD) myalgia, and visceral pain have provided additional information on the role of hormones and the effectiveness of hormone replacement therapy in pain management [41].
Physician’s attitudes and practices also play a role in identifying and grading the severity of visceral pain [42]. This is no different from assessment of other pain modalities in which the gender biases of the participants and testers are apparent [12,20,43,44]. A systemic review and meta-analyses of sex differences in healthy children between 0 and 18 years old in response to experimental pain, encompassing various pain-inducing tasks like cold pressor, heat, and pressure pain, demonstrates that there are no significant differences between boys and girls on pain-related outcomes from majority of studies, however, some studies show that girls have higher cold pressor pain intensity and lower heat pain threshold than boys, but not mechanical detection and pain stimuli [45,46]. A 3-year follow-up interview on a cohort consisting 95 females and 48 males, aged 5-23 years, with initially reported pain shows that all genders report continuing pain with females significantly more likely than males to report, especially for patients whose pain associate to psychosocial factors [47]. Interestingly, in a study comprising 73 children (37 boys, 36 girls) of 4-12 years old undergoing cold pressor pain task monitored by either father or mother, fathers give their sons higher pain ratings than their daughters, whereas mothers’ ratings do not differ among boys and girls [43].
RNAseq analysis of DRG from more than 50 patients for the first time reveals profound sex-differential molecular signatures in patients with neuropathic pain and provides more objective measurements at the molecular levels. In summary, male DRG samples have higher expression levels of IL1B, TNF, CXCL14, OSM, EGRB, TRPV4, LIF, CCL3/4 and female DRG have higher levels of CCL1, CCL19, CCL21, PENK, TRPA1, ADORA2B, GLRA3 in pain when compared to non-pain controls [48]. These factors are closely related to pain, nociception or inflammation [49]. Strikingly, the commonly upregulated genes associated with pain in both sexes are only a handful including ACAN, CPXM1, DIO2, FNDC1, HAMP, IQGAP3, LAIR2, LILRA5, LMNB1, LRRC24, LY6G5C, SIGLEC7 and TGM [48], suggesting an intrinsic sex-differential sensory biological process in pain. Of noting, 80% of the subjects are older than 55 years [48], which suggests that these sex-dependent changes in pain-associated molecules are unlikely results of sex hormonal fluctuations in females due to the hormonal decline in elder populations.
Visceral Pain Preclinical Models with Sexual Dimorphism
To understand the underlying molecular mechanisms in pain sexual dimorphism in order to identify key molecules/pathways for potential drug targets, experimental animal models have been developed vigorously to resemble the phenomenon seen in humans. Colonic inflammation has been used to recapitulate inflammatory bowel disease (IBD)-associated bowel dysfunction and post inflammatory visceral pain, one subtypes of IBS. Early life inflammation or stress, as well as adult stress and genetic models are also implemented to study mechanisms of IBS (Table 1).
The male-predominant visceral pain is reported in mice with colonic inflammation or enema treatment by organic chemicals in multiple models. Zymosan-induced behavioral visceral hypersensitivity is detected in male mice but not female mice, which is accompanied by a higher number of mechanosensitive colorectal afferents per mouse following zymosan treatment of male mice but not female mice [50]. A single dose of intracolonic 2, 4, 6-trinitrobenzenesulfonic acid (TNBS) installation induces colitis in both sexes on day 3 with male mice more severe than female mice [51] and by day 7 colonic hypersensitivity is more pronounced in male mice than in female mice, which is mediated by Piezo2 in nociceptive neurons [24]. Similarly, dextran sodium sulfate (DSS) treatment to induce colonic inflammation affects male mice stronger than female mice showing less inflammatory infiltrates, less crypt damage, and lower TNFα production in the colon of female mice than male mice [52], with sex-differential changes in the immune responses in the colon [53]. Histamine enema increases visceral hypersensitivity in male mice but not female mice and deletion of histamine N-methyltransferase (HNMT) from enteric glia protects males from histamine-driven visceral hypersensitivity, with no effects on female mice [54]. Isovalerate, a bacterially derived short-chain fatty acid, into gut also increases visceral hypersensitivity in male mice but not female mice [26]. These male-preferred colonic inflammatory pain (nociceptive pain) in experimental mice are similar to those observed in clinics that IBS-D is more prevalent in men than women [30].
Female-predominant visceral pain in experimental animals are seen in rats undergoing early life stress (ELS) [55,56] or serotonin transporter (SERT) knockout [57,58]. Specifically, visceromotor responses (VMR) to colorectal distention (CRD) reveals that abdominal contractions in response to visceral distention is significantly increased in female rats exposed to unpredictable early life (PND 8-12) odor-shock presentation when compared to female rats from the predictable ELS or odor-only group, which are not seen in male rats [55]. Female SERT KO rats also have visceral hypersensitivity comparing to female wildtype rats, however, male SERT rats do not [57]. Female but not male SERT KO rats demonstrate higher action potential of colonic afferent neurons comparing to wildtype rats of the same sex [57], suggesting a female-predominant role of the serotonin system in the development of IBS. This notion is further confirmed by application of 5-HT3 receptor antagonists or agonist showing 5-HT3 receptor signaling is essential in visceral hypersensitivity in female SERT-KO rats [58]. A study in rat TMD disorder that has IBS comorbidity also shows stronger and longer-lasting visceral pain behaviors in female rats than male rats [59]. These rat models are valuable tools to investigate the molecular mechanisms of female-preferred non-inflammation related nociplastic visceral pain.
The etiology of visceral pain (inflammation vs non-inflammation) appears to be a determining factor in sexual dimorphisms, however, species (rats vs mice) seem also have a correlation in which male mice and female rats are more prone to disease development (Table 1). Some other examples with exceptions may add more information into the models (Table 1). At baseline, colonic sensitivity is higher in female than male, regardless of species being rats [56,60] or mice [24], suggesting the baseline colonic sensitivity is not species dependent. Mustard oil treatment (induce colonic inflammation) increases visceral pain in both male rats and female rats with female rats have a greater increase than male rats [60]. Interestingly, the baseline spontaneous activity of thoracolumbar colonic afferents in normal male rats is greater than normal female rats, however, mustard oil treatment increases spontaneous activity in female rats but not male rats [60]. These results suggest that female rats, comparing to male rats, are easier to develop visceral hypersensitivity in response to inflammation, contrary to mice in which inflammation-induced visceral pain often occurs in male mice. Other findings in rats and mice show different sexual dimorphic changes in visceral inflammation and pain, comparing to the above categories (Table 1). In response to induction of bladder inflammation, female rats and male rats show similar changes in the urinary bladder [61]. Both female and male rats demonstrate visceral hypersensitivity after early life maternal separation (MS) [62]. Another study combining neonatal MS with early life TNBS colitis in rats shows that male adult rats have hyperactivity behaviors while female adult rats that have undergone the same procedure did not show abnormalities in behavioral tests [63]. Sex differences in microbiota in the gut have been suggested [64,65], while absence of gut microbiota (germ-free mice) causes visceral hypersensitivity in both male and female mice [66]. Maternal separation (nociplastic pain) also induces visceral hypersensitivity in male mice, in which female mice are not assessed [67].
Sex Differences in Visceral Pain Comorbidity
Visceral pain comorbidities occur in both women and men with different etiologies. Studies in women patients exploring the associations between lower urinary tract (LUT) and gastrointestinal symptoms shows that nocturia is significantly more prevalent in gastroenterological (GI) patients with fecal incontinence [68], and female patients with functional constipation or IBS report urinary frequency and incontinence, nocturia, straining to void, or incomplete voiding [68,69,70]. Studies in men shows that 3 or fewer bowel movements per week is associated with nocturia, incomplete bladder emptying, and urinary hesitancy, and more than 10 bowel movements per week was associated with nocturia only [71]. A comparison between men (n=927) and women (n=1073) demonstrates lower prevalence of chronic constipation in men than in women (15.3 vs 26.3%), but both men and women with overactive bladder report chronic constipations or fecal incontinence with similar percentage [72]. Girls and boys seen at the urology clinic for voiding dysfunction and at the GI clinic for functional constipation also have similar levels of symptoms [73]. Constipated children (9.4% of boys and 12.4% of girls) are 6.8 times more likely to have LUT dysfunction than those not constipated [74]. In a population who have adrenoleukodystrophy, the prevalence of urinary and bowel symptoms is similar in men (75.0%) and women (78.8%), with the bladder and bowel disease onset time occurring a decade earlier in males [75]. Patients with IBS also demonstrate comorbid chronic pain response to pressure, heat, or cold-pressor applied to upper extremities (neck, back and shoulder), with a higher percentage in females than in males (54.1% vs. 39.5%) [76]. Sex differences are more pronounced in cold-pressure tolerance, cold pressor intensity rating, but not pressure pain threshold [76]. Nonabdominal chronic pain in adolescents (mean age 16.1 years old) with IBS occurs more often in girls than in boys (22.6% vs 13.5%), and nearly one third of the IBS cases report nonabdominal chronic pain [77].
Animal models to study visceral pain comorbidities mainly are using experimental rodents (Table 1). In rats, normal females, males, or ovariectomized females with colitis demonstrate bladder hyperactivity [78,79,80,81,82,83]. Male rats are also used to study prostate-to-bladder cross-sensitization [84]. In mice, males that have colitis are detected for bladder hypersensitivity, and vice versa, male mice that have bladder inflammation exhibit colonic hypersensitivity [85,86,87,88]. Male mice are also used to assess the effects of colitis on somatic and psychological disorders [89,90]. In addition, male colitic rats and mice have been evaluated to show somatic mechanical and thermal pain [87,91,92,93,94], and increased calcium (Ca2+) activity in paw innervating afferent neurons in male mice [94]. Colitis-induced pain comorbidity in female mice has not been rigorously examined, which could be due to the less effectiveness of inducing colonic inflammation and colonic hypersensitivity in female mice [24,26,50,52]. One of the studies using DSS colitic model demonstrates that referred pain post DSS inflammation are comparable in male mice and female mice, except that female mice have more pronounced licking and freezing behaviors than male mice [16]. We assessed hind paw mechanical sensitivity in female mice following TNBS-induced colitis, which showed that the onset of the referred mechanical pain in female mice were delayed when compared to those in male mice (Figure 1). On day 7 following colitis induction, female mice did not exhibit robust colonic mechanical hypersensitivity [24] but showed strong referred pain (Figure 1). Similarly, DSS-induced less colonic inflammation in female mice than in male mice [52], however, the referred pain sensitivity in both sexes are similar following DSS treatment [16]. One of the explanations could be that the colon was the primary organ injured in colitis, while the hind paw was not directly injured. The underlying mechanisms of the differential behaviors of the injured vs uninjured organs in female mice are not known. It necessitates comparison of the neurochemical coding between the injured primary afferent neurons and the uninjured neurons.
Cellular and Molecular Mechanisms in Pain Sexual Dimorphism
Evidences in sexual dimorphisms in visceral pain and pain at large have inspired rigorous studies to uncover the underlying molecular mechanisms to shed lights on drug discoveries for sex-oriented effective treatment. Sensory neurons are primary targets for this type of studies and nonneuronal cells also spark considerable research interests. Neural circuits and neurotransmitters are important components in transducing pain signals thus peripheral damages can be timely sensed and remedied.
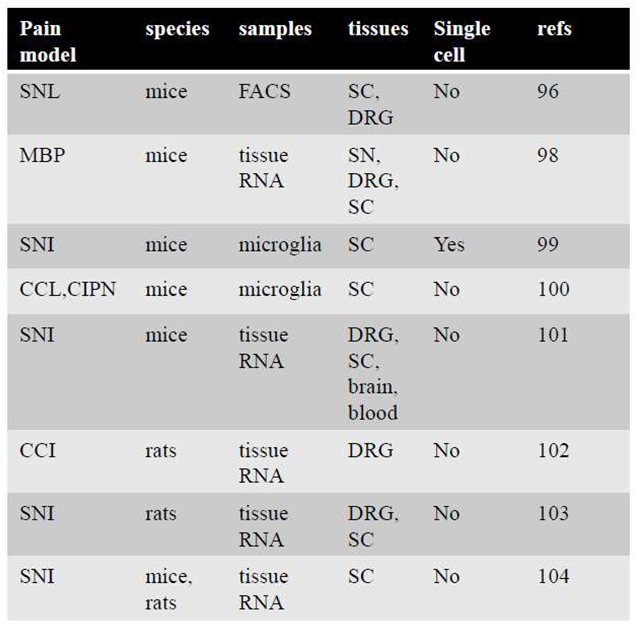
Transcriptome analysis is performed to reveal 540 sex-dependent differentially expressing genes (DEG) in DRG sensory neurons [95]. In female mice, a distinct set of genes predominantly expressed are linked to immune processes, nervous system development, extracellular matrix organization, and inflammatory pathways. These genes include those related to the complement system, chemokines, cytokines, and growth factor receptors, with a more prominent expression in trigeminal ganglion (TG) than DRG neurons [95]. This pattern suggests a potential connection to the higher prevalence of certain pain conditions such as migraine in females, possibly due to enhanced inflammatory and immune responses related to TG. Male mice demonstrate a different profile of predominantly expressed genes, focusing on proteasome subunits, mitochondrial function, and oxidative phosphorylation [95]. These findings point to potential protective mechanisms in male sensory neurons against nerve injury, highlighting sex-specific resilience pathways. Pain related genes in DRG of male and female mice such as those related to transcriptional and translational machinery are also different [95]. Another transcriptomic analysis demonstrates that there are no striking differences in gene expression between male and female mice and immune cell infiltration into DRG are sexually different in nerve injury pain [96]. DRG gene-expression patterns in chronic constriction nerve injury (CCI) also reveals sexual dimorphism showing that 859 genes, including Kcna4, Ccna2d3, and Scn8a, are upregulated in male mice in CCI, but not in female mice. However, 146 genes (including Ahr, Fos, and Socs3) are elevated in female mice in CCI [97]. A number of other studies [98,99,100,101,102,103,104] have provided considerable sizes of datasets from RNAseq to reveal sex-differential changes in molecular signatures in DRG, spinal cord, sciatic nerves, and brain tissues from mice and rats in pain, which is summarized in Table 2, to provide a guidance for in-depth study of their physiological roles in pain sexual dimorphisms.
Nociceptors:
CGRP (Calcitonin Gene-Related Peptide) and Nav1.8 are largely co-localized in nociceptive neurons to sense pain signals [105,106]. CGRP is produced in DRG neurons and can release peripherally to promote neurogenic inflammation or centrally to cause spinal central sensitization. Following colitis induction in male rats, CGRP is increased in DRG neurons, primarily in uninjured bladder afferent neurons and also in the spinal cord [107,108]. CGRP8–37, a CGRP antagonist, attenuates colitis or nerve growth factor (NGF)-induced visceral hypersensitivity in male rats [109]. When chemogenetically activating peripheral glia that results in visceral and somatic hypersensitivity in male but not female mice, the expression of CGRP is accompanying to increase in DRG neurons of male mice but not female mice [110]. In an acute arthritis pain model or cyclophosphamide-induced bladder pain, CGRP participates in peripheral and spinal sensitization and pain in male rats [111,112]. Following nerve injury, blockade of CGRP receptor inhibits neuropathic pain behaviors in rats of both sexes [113]. CGRP is also implicated in migraine pathophysiology that largely affects women. Anti-CGRP antibody can attenuates colitis-induced bladder hypersensitivity in female rats [81]. In hyperalgesic priming induced by activation of interleukin 6 signaling, intrathecal injection of CGRP receptor antagonists olcegepant or CGRP8-37 blocks and reverses hyperalgesic priming and neuropathic pain only in female mice but not male mice, and CGRP-sequestering monoclonal antibody blocks IL-6-induced mechanical hypersensitivity and hyperalgesic priming in female mice and rats [114]. These findings suggest that the sex-differential role of CGRP in pain is context-dependent and animal model specific.
The sex differential function of Nav1.8 ion channel is also suggested by recent studies. Optogenetic activation of Nav1.8-lineage sensory fibers elicits nerve fiber firing much easier in female mice than male mice at baseline [26]. Conditional deletion of Piezo2 from Nav1.8-lineage neurons reduces baseline colonic mechanical sensitivity in female mice but not male mice [24]. These findings suggest that female mice may have a narrower sensory homeostatic range at baseline (weaker inhibitory feedback when nociceptive neurons are activated and less strength to maintain activity when nociceptors are inhibited) than male mice. During pain states, Piezo2 conditional deletion from nociceptors attenuates TNBS-induced colonic hypersensitivity in male mice but not female mice [24], has no impact on complete Freund’s adjuvant (CFA)-induced knee swelling in female mice [25], but reduces knee swelling associated knee hyperalgesia in female mice, noting that male mice in this CFA context is not measured [25]. Conversely by sex, nociceptors lack of Piezo2 are associated with reduced experimental osteoarthritis-induced joint pain and referred mechanical allodynia of the hind paw as well as NGF-induced knee swelling and hyperalgesia in male mice when compared to control [25]. Spared nerve injury (SNI) induces mechanical hypersensitivity and cold allodynia similarly in both male and female mice, however, female mice demonstrate higher expression levels of ATF3 and HMGB1 expression in Nav1.8-lineage nociceptors than male mice [115]. Interestingly, Nav1.8-Cre based Toll-like receptor 4 (TLR4) deletion attenuates acutely (day 1-5 post SNI) but not chronically (day 7 post SNI) mechanical hypersensitivity in female mice but has no effects on mechanical pain in male mice, conversely, TLR4 conditional deletion reduces cold allodynia up to day 7 post SNI as measured in male mice but not female mice [115]. It appears that the sex-differential roles of nociceptors in regulation of pain is diverse by measured pain modalities and the time course examined after injury or stress. Visceral pain and somatic pain could be processed differently, as well as the mechanisms underlying inflammatory nociceptive pain, neuropathic pain, nociplastic pain, or referred pain and pain comorbidities.
Nonneuronal cells:
Microglia in the spinal cord are mostly studied for their roles in pain in a sex-dependent manner. In visceral pain, inhibition of microglia by minocycline, MAC-1-conjugated saponin (a method of depleting microglia), in female rats reduces visceral pain comorbidity [82]. Nerve injury induces mechanical allodynia in both male and female mice, however, inhibition of microglia by pharmacological approaches or chemogenetics reduces the development of the neuropathic pain in male mice but not female mice [116,117,118]. Conditional deletion of Orai1 channel from microglia attenuates microglia development in the spinal cord and mitigates pain hyperalgesia following nerve injury in male mice but not in female mice [119]. Chemogenetic activation of microglia elicits mechanical allodynia exclusively in male mice although the reactive microglia-dominant molecules are upregulated in microglia in both sexes [120]. The male-predominant role of microglia in neuropathic pain is conserved in rats and mice [121], however, it is different from visceral pain that microglia has a role in female rats [82]. The female-preferential role of microglia in rats is also shown by a study in which lipopolysaccharide (LPS) treatment induces greater microglia activation and interleukin IL-1β upregulation in the periaqueductal gray of female rats compared with male rats [122]. The male-predominant role of microglia in mice with pain is likely due to sex-differential development of microglia by which male mice have delayed gene expression relative to females and exposure of adult male mice to LPS accelerates microglial development in male mice [123]. Astrocytes, on the other hand, sex-independently regulate inflammatory and neuropathic pain in mice [124]. Satellite glial cells (SGCs) compose the largest glial populations in DRG. Chemogenetic activation of SGCs inhibits acute (15 min to 1 h after clozapine N-oxide (CNO) treatment) mechanical hyperalgesia in male mice (female mice are not examined) [125], and facilitates chronic (1-3 days with daily CNO injection) mechanical pain and colonic hypersensitivity in male mice but not female mice [110]. Strikingly, the number of Proteolipid protein Plp1-expressing SGCs are significantly less in DRG of female mice than male mice [110]. RNAseq information comparing DRG neurons and immune cells in DRG and the spinal cord shows that female and male mice develops nerve injury pain with no apparent changes in gene-expression profiles in DRG neurons and in the amount of immune cell activation in the spinal cord [96]. Additional information is needed to pinpoint the sex differential role of nonneuronal cells in visceral pain and pain at large.
Sexual Dimorphisms of Brain Circuits and Emotional Effects on Visceral Pain and Pain at Large
CGRP8-37 administration into the central nucleus of the amygdala, a structure in the brain involved with the experiencing of emotions, in chronic neuropathic rats reduces mechanical hypersensitivity in both sexes but shows female-predominant effects on emotional-affective responses (ultrasonic vocalizations) and anxiety-like behaviors (open field test) [113], suggesting an emotional influence in pain perception. The sex-differences in emotion-related cognitive processes are also seen in patients with IBS and healthy subjects [126]. In a task-free resting-state functional magnetic resonance imaging analysis, female healthy subjects and IBS patients have a frequency power distribution skewed toward high frequency to a greater extent in the amygdala and hippocampus compared with male subjects [127]. These variations in brain function point to a fundamental difference in how men and women process and perceive visceral pain, necessitating a sex-specific approach in understanding and managing these conditions.
Numerous brain areas are implicated in the regulation of emotions and chronic pain, including the thalamus, amygdala, medial prefrontal cortex, and periaqueductal gray (PAG) [128]. An increasing number of studies show that no neuron exists in a vacuum and that the connections among different brain regions are more important than the divisions inside the brain [129]. The fundaments to deciphering the intricate expressions of pain and emotion is to pinpoint particular or shared circuits that control them. In clinical trials, the default mode, central, visual, and sensorimotor modules in patients with chronic pain are analyzed using graph theory, modular analysis, and machine learning. Sexual differentiation is observed in the anterior cingulate cortex (ACC), amygdala, locus coeruleus, ventral tegmental area, and PAG, among other brain regions that link to pain and emotions. A recent study shows that ACC is important for both chronic pain and the emotional reactions associated with it [130]. In a cross-sectional study on resting-state functional connectivity (RSFC) among older adults, sex differences is evident in the associations of thermal pain with RSFC between the ACC and amygdala, and between the ACC and PAG in older females compared to older males [131]. Studies on the functional connectivity and structural connectivity between the descending antinociceptive system and the subgenual ACC (sgACC) also show that women’s brain wiring may allow for higher engagement of the descending modulation system to mediate pain habituation than man. Specifically, women show more functional connectivity than men between the sgACC and the PAG, raphe nucleus, medial thalamus, and anterior midcingulate cortex (aMCC) [132]. There have also been discoveries suggesting that women with chronic pain associated to ankylosing spondylitis, but not men, have altered sgACC circuitry. Women’s sgACC functional connectivity to the default mode and sensorimotor networks are higher than men’s [133].
One important biological process for the experience of pain and control of emotions is synaptic plasticity. Excitatory transmission and plasticity in the ACC have been shown in preclinical investigations to have a significant role in emotions connected to chronic pain. Using a 64-channel multielectrode (MED64) system to record synaptic plasticity in the ACC, long-term potentiation (LTP) does not exhibit a sex-related difference, however, long-term depression (LTD) is higher in the ACC in male mice than female mice [134]. Furthermore, the ACC in schizophrenia exhibits gender disparities in GABAergic gene expression, while it is still unclear if pain and pain-related emotions are influenced by sex-difference GABAergic gene expression [135].
Concluding Remarks:
Men and women respond differently to pain medications, such as morphine, in managing visceral pain and other pain modalities. These disparities, mediated at both central and peripheral levels, underscore the complexity of pain management and the necessity for personalized medication strategies based on sex. The caveat of confirming sexual dimorphism of pain in humans are largely interfered by emotions, geographical and societal status, and testing environments. Objective measurements of visceral pain and other pain formats are necessary to decipher pain sexual dimorphisms. Animal models provide a more reproducible states to investigate the underlying sex-differential pain mechanisms. Mice and rats are popular experimental animals for study of visceral pain and pain at large, however, majority of studies have been focusing on using males. Limited research data using both sexes thus far suggests that male mice are prone to be inflamed in the colon and have visceral pain when compared to female mice. Female but not male rats are reported to have visceral pain under other types of pathophysiology such as stress. It is certain that more experiments should be enforced to ensure sufficient body of information being generated to better understand sex differences in pain. Sensory neurons, immune cells and central neural circuits especially those involved in emotion have sex differences at baseline and during diseases. However, how these evolving landscapes contribute to sex-differential pain generation and maintenance are still ambiguous and necessitates further investigations.
Author Contributions
NT wrote the draft of the manuscript and performed some of the experiments. LYQ designed and revised the manuscript. NT and LYQ analyzed the data and approved the final version. The authors thank Mr. Shanwei Shen for his technical support in data collection.
Funding
This project was supported by grants NIH R01 DK118137 (LYQ); R01 DK121131 (LYQ); Virginia’s Commonwealth Health Research Board (CHRB) 236-06-18 (LYQ); NIH P30 DA033934 (LYQ).
Data Availability Statement
All data are included in this paper.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Kennedy, R.; Abd-Elsayed, A. The International Association for the Study of Pain (IASP) Classification of Chronic Pain Syndromes. Abd-Elsayed, A. (eds) Pain 2019. [CrossRef]
- Mayer, E.A.; Ryu, H.J.; Bhatt, R.R. The neurobiology of irritable bowel syndrome. Mol Psychiatry 2023, 28, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Engel, F.; Stadnitski, T.; Stroe-Kunold, E.; Berens, S.; Schaefert, R.; Wild, B. Pain and psyche in a patient with irritable bowel syndrome: chicken or egg? A time series case report. BMC Gastroenterol 2021, 21, 309. [Google Scholar] [CrossRef]
- Riedl, A.; Schmidtmann, M.; Stengel, A.; Goebel, M.; Wisser, A.S.; Klapp, B.F.; Monnikes, H. Somatic comorbidities of irritable bowel syndrome: a systematic analysis. J Psychosom Res 2008, 64, 573–582. [Google Scholar] [CrossRef]
- Chang, L.; Mayer, E.A.; Labus, J.S.; Schmulson, M.; Lee, O.Y.; Olivas, T.I.; Stains, J.; Naliboff, B.D. Effect of sex on perception of rectosigmoid stimuli in irritable bowel syndrome. Am J Physiol Regul Integr Comp Physiol 2006, 291, R277–R284. [Google Scholar] [CrossRef] [PubMed]
- Simren, M.; Abrahamsson, H.; Bjornsson, E.S. Lipid-induced colonic hypersensitivity in the irritable bowel syndrome: the role of bowel habit, sex, and psychologic factors. Clin Gastroenterol Hepatol 2007, 5, 201–208. [Google Scholar] [CrossRef]
- Chang, L.; Heitkemper, M.M. Gender differences in irritable bowel syndrome. Gastroenterology 2002, 123, 1686–1701. [Google Scholar] [CrossRef]
- Choghakhori, R.; Abbasnezhad, A.; Amani, R.; Alipour, M. Sex-Related Differences in Clinical Symptoms, Quality of Life, and Biochemical Factors in Irritable Bowel Syndrome. Digest Dis Sci 2017, 62, 1550–1560. [Google Scholar] [CrossRef]
- Hong, J.Y.; Labus, J.S.; Kilpatrick, L.A.; Stains, J.; Heendeniya, N.; Smith, S.R.; Katibian, D.; Tillisch, K.; Mayer, E.A. Patients With Irritable Bowel Syndrome Show Sex Related Differences in Resting-State Functional Connectivity. Gastroenterology 2014, 146, S847. [Google Scholar] [CrossRef]
- Goodman, W.A.; Erkkila, I.P.; Pizarro, T.T. Sex matters: impact on pathogenesis, presentation and treatment of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol 2020, 17, 740–754. [Google Scholar] [CrossRef]
- Camilleri, M. Sex as a biological variable in irritable bowel syndrome. Neurogastroent Motil 2020, 32, e13802. [Google Scholar] [CrossRef]
- Mogil, J.S. Qualitative sex differences in pain processing: emerging evidence of a biased literature. Nat Rev Neurosci 2020, 21, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Stephens, K.E.; Zhou, W.; Ji, Z.; Chen, Z.; He, S.; Ji, H.; Guan, Y.; Taverna, S.D. Sex differences in gene regulation in the dorsal root ganglion after nerve injury. BMC Genomics 2019, 20, 147. [Google Scholar] [CrossRef]
- Presto, P.; Mazzitelli, M.; Junell, R.; Griffin, Z.; Neugebauer, V. Sex differences in pain along the neuraxis. Neuropharmacology 2022, 210, 109030. [Google Scholar] [CrossRef] [PubMed]
- Bartley, E.J.; Fillingim, R.B. Sex differences in pain: a brief review of clinical and experimental findings. Br J Anaesth 2013, 111, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Francis-Malave, A.M.; Martinez Gonzalez, S.; Pichardo, C.; Wilson, T.D.; Rivera-Garcia, L.G.; Brinster, L.R.; Carrasquillo, Y. Sex differences in pain-related behaviors and clinical progression of disease in mouse models of colonic pain. Pain 2023, 164, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Classification of chronic pain. Descriptions of chronic pain syndromes and definitions of pain terms. Prepared by the International Association for the Study of Pain, Subcommittee on Taxonomy. Pain Suppl 1986, 3, S1–S226. [Google Scholar]
- Mazure, C.M.; Jones, D.P. Twenty years and still counting: including women as participants and studying sex and gender in biomedical research. BMC Womens Health 2015, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Legato, M.J. The International Society for Gender Medicine History and Highlights PREFACE. International Society for Gender Medicine: History and Highlights 2017, Xiii-Xvi.
- Samulowitz, A.; Gremyr, I.; Eriksson, E.; Hensing, G. "Brave Men" and "Emotional Women": A Theory-Guided Literature Review on Gender Bias in Health Care and Gendered Norms towards Patients with Chronic Pain. Pain Res Manag 2018, 2018, 6358624. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, N. Sex-Gender Differences in Irritable Bowel Syndrome. J Neurogastroenterol Motil 2018, 24, 544–558. [Google Scholar] [CrossRef]
- Gregus, A.M.; Levine, I.S.; Eddinger, K.A.; Yaksh, T.L.; Buczynski, M.W. Sex differences in neuroimmune and glial mechanisms of pain. Pain 2021, 162, 2186–2200. [Google Scholar] [CrossRef]
- Beery, A.K. Inclusion of females does not increase variability in rodent research studies. Curr Opin Behav Sci 2018, 23, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Madar, J.; Tiwari, N.; Smith, C.; Sharma, D.; Shen, S.; Elmahdi, A.; Qiao, L.Y. Piezo2 regulates colonic mechanical sensitivity in a sex specific manner in mice. Nat Commun 2023, 14, 2158. [Google Scholar] [CrossRef] [PubMed]
- Obeidat, A.M.; Wood, M.J.; Adamczyk, N.S.; Ishihara, S.; Li, J.; Wang, L.; Ren, D.; Bennett, D.A.; Miller, R.J.; Malfait, A.M.; et al. Piezo2 expressing nociceptors mediate mechanical sensitization in experimental osteoarthritis. Nat Commun 2023, 14, 2479. [Google Scholar] [CrossRef]
- Bayrer, J.R.; Castro, J.; Venkataraman, A.; Touhara, K.K.; Rossen, N.D.; Morrie, R.D.; Maddern, J.; Hendry, A.; Braverman, K.N.; Garcia-Caraballo, S.; et al. Gut enterochromaffin cells drive visceral pain and anxiety. Nature 2023, 616, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Icenhour, A.; Labrenz, F.; Roderigo, T.; Siebert, C.; Elsenbruch, S.; Benson, S. Are there sex differences in visceral sensitivity in young healthy men and women? Neurogastroenterol Motil 2019, 31, e13664. [Google Scholar] [CrossRef]
- Posserud, I.; Syrous, A.; Lindström, L.; Tack, J.; Abrahamsson, H.; Simrén, M. Altered rectal perception in irritable bowel syndrome is associated with symptom severity. Gastroenterology 2007, 133, 1113–1123. [Google Scholar] [CrossRef]
- Almario, C.V.; Sharabi, E.; Chey, W.D.; Lauzon, M.; Higgins, C.S.; Spiegel, B.M.R. Prevalence and Burden of Illness of Rome IV Irritable Bowel Syndrome in the United States: Results From a Nationwide Cross-Sectional Study. Gastroenterology 2023, 165, 1475–1487. [Google Scholar] [CrossRef]
- Adeyemo, M.A.; Spiegel, B.M.; Chang, L. Meta-analysis: do irritable bowel syndrome symptoms vary between men and women? Aliment Pharmacol Ther 2010, 32, 738–755. [Google Scholar] [CrossRef]
- Lungaro, L.; Costanzini, A.; Manza, F.; Barbalinardo, M.; Gentili, D.; Guarino, M.; Caputo, F.; Zoli, G.; De Giorgio, R.; Caio, G. Impact of Female Gender in Inflammatory Bowel Diseases: A Narrative Review. J Pers Med 2023, 13. [Google Scholar] [CrossRef]
- Barbara, G.; Grover, M.; Bercik, P.; Corsetti, M.; Ghoshal, U.C.; Ohman, L.; Rajilic-Stojanovic, M. Rome Foundation Working Team Report on Post-Infection Irritable Bowel Syndrome. Gastroenterology 2019, 156, 46–58. [Google Scholar] [CrossRef]
- Park, J.H.; Kang, S.H.; Kim, J.S.; Moon, H.S.; Sung, J.K.; Jeong, H.Y. Contribution of sex and gender roles to the incidence of post-infectious irritable bowel syndrome in a prospective study. Sci Rep 2023, 13, 19467. [Google Scholar] [CrossRef]
- Grinsvall, C.; Tornblom, H.; Tack, J.; Van Oudenhove, L.; Simren, M. Relationships between psychological state, abuse, somatization and visceral pain sensitivity in irritable bowel syndrome. United European Gastroenterol J 2018, 6, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Oka, P.; Parr, H.; Barberio, B.; Black, C.J.; Savarino, E.V.; Ford, A.C. Global prevalence of irritable bowel syndrome according to Rome III or IV criteria: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol 2020, 5, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Greuter, T.; Manser, C.; Pittet, V.; Vavricka, S.R.; Biedermann, L.; on behalf of Swiss Ibdnet. Gender Differences in Inflammatory Bowel Disease. Digestion 2020, 101 Suppl 1, 98–104. [Google Scholar] [CrossRef]
- Smith, Y.R.; Stohler, C.S.; Nichols, T.E.; Bueller, J.A.; Koeppe, R.A.; Zubieta, J.K. Pronociceptive and antinociceptive effects of estradiol through endogenous opioid neurotransmission in women. J Neurosci 2006, 26, 5777–5785. [Google Scholar] [CrossRef] [PubMed]
- Aranda, G.; Fernandez-Rebollo, E.; Pradas-Juni, M.; Hanzu, F.A.; Kalko, S.G.; Halperin, I.; Mora, M. Effects of sex steroids on the pattern of methylation and expression of the promoter region of estrogen and androgen receptors in people with gender dysphoria under cross-sex hormone treatment. J Steroid Biochem Mol Biol 2017, 172, 20–28. [Google Scholar] [CrossRef]
- Mulak, A.; Tache, Y.; Larauche, M. Sex hormones in the modulation of irritable bowel syndrome. World J Gastroenterol 2014, 20, 2433–2448. [Google Scholar] [CrossRef]
- Heitkemper, M.M.; Jarrett, M. Gender differences and hormonal modulation in visceral pain. Curr Pain Headache Rep 2001, 5, 35–43. [Google Scholar] [CrossRef]
- Athnaiel, O.; Cantillo, S.; Paredes, S.; Knezevic, N.N. The Role of Sex Hormones in Pain-Related Conditions. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Lacy, B.E.; Rosemore, J.; Robertson, D.; Corbin, D.A.; Grau, M.; Crowell, M.D. Physicians' attitudes and practices in the evaluation and treatment of irritable bowel syndrome. Scand J Gastroenterol 2006, 41, 892–902. [Google Scholar] [CrossRef]
- Moon, E.C.; Chambers, C.T.; Larochette, A.C.; Hayton, K.; Craig, K.D.; McGrath, P.J. Sex differences in parent and child pain ratings during an experimental child pain task. Pain Res Manag 2008, 13, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Chaloner, A.; Greenwood-Van Meerveld, B. Early life adversity as a risk factor for visceral pain in later life: importance of sex differences. Front Neurosci 2013, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Blankenburg, M.; Boekens, H.; Hechler, T.; Maier, C.; Krumova, E.; Scherens, A.; Magerl, W.; Aksu, F.; Zernikow, B. Reference values for quantitative sensory testing in children and adolescents: developmental and gender differences of somatosensory perception. Pain 2010, 149, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Boerner, K.E.; Birnie, K.A.; Caes, L.; Schinkel, M.; Chambers, C.T. Sex differences in experimental pain among healthy children: A systematic review and meta-analysis. Pain 2014, 155, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.L.; McGrath, P.A.; Brown, S.C.; Katz, J. Children with chronic pain: impact of sex and age on long-term outcomes. Pain 2007, 128, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.R.; Shiers, S.; Caruso, J.P.; Tavares-Ferreira, D.; Sankaranarayanan, I.; Uhelski, M.L.; Li, Y.; North, R.Y.; Tatsui, C.; Dussor, G.; et al. RNA profiling of human dorsal root ganglia reveals sex differences in mechanisms promoting neuropathic pain. Brain 2023, 146, 749–766. [Google Scholar] [CrossRef] [PubMed]
- LaCroix-Fralish, M.L.; Austin, J.S.; Zheng, F.Y.; Levitin, D.J.; Mogil, J.S. Patterns of pain: meta-analysis of microarray studies of pain. Pain 2011, 152, 1888–1898. [Google Scholar] [CrossRef]
- Guo, T.; Liu, J.; Chen, L.; Bian, Z.; Zheng, G.; Feng, B. Sex differences in zymosan-induced behavioral visceral hypersensitivity and colorectal afferent sensitization. Am J Physiol Gastrointest Liver Physiol 2023. [Google Scholar] [CrossRef]
- Kozik, A.J.; Nakatsu, C.H.; Chun, H.; Jones-Hall, Y.L. Age, sex, and TNF associated differences in the gut microbiota of mice and their impact on acute TNBS colitis. Exp Mol Pathol 2017, 103, 311–319. [Google Scholar] [CrossRef]
- Babickova, J.; Tothova, L.; Lengyelova, E.; Bartonova, A.; Hodosy, J.; Gardlik, R.; Celec, P. Sex Differences in Experimentally Induced Colitis in Mice: a Role for Estrogens. Inflammation 2015, 38, 1996–2006. [Google Scholar] [CrossRef]
- Hases, L.; Birgersson, M.; Indukuri, R.; Archer, A.; Williams, C. Colitis Induces Sex-Specific Intestinal Transcriptomic Responses in Mice. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- McClain, J.L.; Morales-Soto, W.; Gonzales, J.; Parmar, V.; Demireva, E.Y.; Gulbransen, B.D. Sexually Dimorphic Effects of Histamine Degradation by Enteric Glial Histamine N-Methyltransferase (HNMT) on Visceral Hypersensitivity. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Louwies, T.; Greenwood-Van Meerveld, B. Sex differences in the epigenetic regulation of chronic visceral pain following unpredictable early life stress. Neurogastroenterol Motil 2020, 32, e13751. [Google Scholar] [CrossRef]
- Chen, J.H.; Sun, Y.; Ju, P.J.; Wei, J.B.; Li, Q.J.; Winston, J.H. Estrogen augmented visceral pain and colonic neuron modulation in a double-hit model of prenatal and adult stress. World J Gastroenterol 2021, 27, 5060–5075. [Google Scholar] [CrossRef]
- Galligan, J.J.; Patel, B.A.; Schneider, S.P.; Wang, H.; Zhao, H.; Novotny, M.; Bian, X.; Kabeer, R.; Fried, D.; Swain, G.M. Visceral hypersensitivity in female but not in male serotonin transporter knockout rats. Neurogastroenterol Motil 2013, 25, e373–e381. [Google Scholar] [CrossRef] [PubMed]
- El-Ayache, N.; Galligan, J.J. 5-HT(3) receptor signaling in serotonin transporter-knockout rats: a female sex-specific animal model of visceral hypersensitivity. Am J Physiol Gastrointest Liver Physiol 2019, 316, G132–G143. [Google Scholar] [CrossRef]
- Da Silva, J.T.; Hernandez-Rojas, L.G.; Mekonen, H.K.; Hanson, S.; Melemedjian, O.; Scott, A.J.; Ernst, R.K.; Seminowicz, D.A.; Traub, R.J. Sex differences in visceral sensitivity and brain activity in a rat model of comorbid pain: a longitudinal study. Pain 1097. [Google Scholar] [CrossRef]
- Ji, Y.; Tang, B.; Cao, D.Y.; Wang, G.; Traub, R.J. Sex differences in spinal processing of transient and inflammatory colorectal stimuli in the rat. Pain 2012, 153, 1965–1973. [Google Scholar] [CrossRef]
- Qiao, L.Y.; Gulick, M.A. Region-specific changes in the phosphorylation of ERK1/2 and ERK5 in rat micturition pathways following cyclophosphamide-induced cystitis. Am J Physiol Regul Integr Comp Physiol 2007, 292, R1368–R1375. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Zhang, H.; Sun, H.; Zhou, L.; Chen, Y.; Xuan, L.; Jiang, Y.; Xu, S. Maternal Separation Induced Visceral Hypersensitivity from Childhood to Adulthood. J Neurogastroenterol Motil 2017, 23, 306–315. [Google Scholar] [CrossRef]
- Hasegawa, R.; Saito-Nakaya, K.; Gu, L.; Kanazawa, M.; Fukudo, S. Maternal separation and TNBS-induced gut inflammation synergistically alter the sexually differentiated stress response in rats. Biopsychosoc Med 2023, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Woo, S.Y.; Raza, S.; Ho, D.; Jeon, S.W.; Chang, Y.; Ryu, S.; Kim, H.L.; Kim, H.N. Association between gut microbiota and anxiety symptoms: A large population-based study examining sex differences. J Affect Disord 2023, 333, 21–29. [Google Scholar] [CrossRef] [PubMed]
- McGee, J.S.; Huttenhower, C. Of mice and men and women: Sexual dimorphism of the gut microbiome. Int J Womens Dermatol 2021, 7, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Pujo, J.; De Palma, G.; Lu, J.; Galipeau, H.J.; Surette, M.G.; Collins, S.M.; Bercik, P. Gut microbiota modulates visceral sensitivity through calcitonin gene-related peptide (CGRP) production. Gut Microbes 2023, 15, 2188874. [Google Scholar] [CrossRef] [PubMed]
- Tao, E.; Long, G.; Yang, T.; Chen, B.; Guo, R.; Ye, D.; Fang, M.; Jiang, M. Maternal Separation Induced Visceral Hypersensitivity Evaluated via Novel and Small Size Distention Balloon in Post-weaning Mice. Front Neurosci 2021, 15, 803957. [Google Scholar] [CrossRef] [PubMed]
- Wyndaele, M.; De Winter, B.Y.; Pelckmans, P.A.; De Wachter, S.; Van Outryve, M.; Wyndaele, J.J. Exploring associations between lower urinary tract symptoms (LUTS) and gastrointestinal (GI) problems in women: a study in women with urological and GI problems vs a control population. BJU Int 2015, 115, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Wyndaele, M.; De Winter, B.Y.; Pelckmans, P.; Wyndaele, J.J. Lower bowel function in urinary incontinent women, urinary continent women and in controls. Neurourol Urodyn 2011, 30, 138–143. [Google Scholar] [CrossRef]
- Carter, D.; Beer-Gabel, M. Lower urinary tract symptoms in chronically constipated women. Int Urogynecol J 2012, 23, 1785–1789. [Google Scholar] [CrossRef] [PubMed]
- Thurmon, K.L.; Breyer, B.N.; Erickson, B.A. Association of bowel habits with lower urinary tract symptoms in men: findings from the 2005-2006 and 2007-2008 National Health and Nutrition Examination Survey. J Urol 2013, 189, 1409–1414. [Google Scholar] [CrossRef]
- Coyne, K.S.; Cash, B.; Kopp, Z.; Gelhorn, H.; Milsom, I.; Berriman, S.; Vats, V.; Khullar, V. The prevalence of chronic constipation and faecal incontinence among men and women with symptoms of overactive bladder. BJU Int 2011, 107, 254–261. [Google Scholar] [CrossRef]
- Wolfe-Christensen, C.; Manolis, A.; Guy, W.C.; Kovacevic, N.; Zoubi, N.; El-Baba, M.; Kovacevic, L.G.; Lakshmanan, Y. Bladder and bowel dysfunction: evidence for multidisciplinary care. J Urol 2013, 190, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, C.; Sousa, A.S.; Fraga, L.G.; Veiga, M.L.; Bastos Netto, J.M.; Barroso, U., Jr. Constipation and Lower Urinary Tract Dysfunction in Children and Adolescents: A Population-Based Study. Front Pediatr 2016, 4, 101. [Google Scholar] [CrossRef] [PubMed]
- Corre, C.S.; Grant, N.; Sadjadi, R.; Hayden, D.; Becker, C.; Gomery, P.; Eichler, F.S. Beyond gait and balance: urinary and bowel dysfunction in X-linked adrenoleukodystrophy. Orphanet J Rare Dis 2021, 16, 14. [Google Scholar] [CrossRef]
- Stabell, N.; Stubhaug, A.; Flaegstad, T.; Nielsen, C.S. Increased pain sensitivity among adults reporting irritable bowel syndrome symptoms in a large population-based study. Pain 2013, 154, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Stabell, N.; Stubhaug, A.; Flaegstad, T.; Mayer, E.; Naliboff, B.D.; Nielsen, C.S. Widespread hyperalgesia in adolescents with symptoms of irritable bowel syndrome: results from a large population-based study. J Pain 2014, 15, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Pezzone, M.A.; Liang, R.; Fraser, M.O. A model of neural cross-talk and irritation in the pelvis: implications for the overlap of chronic pelvic pain disorders. Gastroenterology 2005, 128, 1953–1964. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.M.; Gulick, M.A.; Yu, S.J.; Grider, J.R.; Murthy, K.S.; Kuemmerle, J.F.; Akbarali, H.I.; Qiao, L.Y. Up-regulation of brain-derived neurotrophic factor in primary afferent pathway regulates colon-to-bladder cross-sensitization in rat. J Neuroinflammation 2012, 9, 30. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Kawamorita, N.; Oguchi, T.; Funahashi, Y.; Tyagi, P.; Chancellor, M.B.; Yoshimura, N. Pelvic organ cross-sensitization to enhance bladder and urethral pain behaviors in rats with experimental colitis. Neuroscience 2015, 284, 422–429. [Google Scholar] [CrossRef]
- Noor-Mohammadi, E.; Ligon, C.O.; Mackenzie, K.D.; Stratton, J.; Shnider, S.J.; Greenwood-Van Meerveld, B. Antinociceptive Effects of an Anti-CGRP Antibody in Rat Models of Colon-Bladder Cross-Organ Sensitization. J Pharmacol Exp Ther 2023, 387, 4–14. [Google Scholar] [CrossRef]
- Majima, T.; Funahashi, Y.; Kawamorita, N.; Takai, S.; Matsukawa, Y.; Yamamoto, T.; Yoshimura, N.; Gotoh, M. Role of microglia in the spinal cord in colon-to-bladder neural crosstalk in a rat model of colitis. Neurourol Urodyn 2018, 37, 1320–1328. [Google Scholar] [CrossRef]
- Malykhina, A.P.; Qin, C.; Greenwood-van Meerveld, B.; Foreman, R.D.; Lupu, F.; Akbarali, H.I. Hyperexcitability of convergent colon and bladder dorsal root ganglion neurons after colonic inflammation: mechanism for pelvic organ cross-talk. Neurogastroenterol Motil 2006, 18, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, Y.; Takahashi, R.; Mizoguchi, S.; Suzuki, T.; Takaoka, E.; Ni, J.; Wang, Z.; DeFranco, D.B.; de Groat, W.C.; Tyagi, P.; et al. Bladder overactivity and afferent hyperexcitability induced by prostate-to-bladder cross-sensitization in rats with prostatic inflammation. J Physiol 2019, 597, 2063–2078. [Google Scholar] [CrossRef] [PubMed]
- Grundy, L.; Harrington, A.M.; Castro, J.; Garcia-Caraballo, S.; Deiteren, A.; Maddern, J.; Rychkov, G.Y.; Ge, P.; Peters, S.; Feil, R.; et al. Chronic linaclotide treatment reduces colitis-induced neuroplasticity and reverses persistent bladder dysfunction. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Pan, X.Q.; Villamor, A.N.; Asfaw, T.S.; Chang, S.; Zderic, S.A.; Malykhina, A.P. Lack of transient receptor potential vanilloid 1 channel modulates the development of neurogenic bladder dysfunction induced by cross-sensitization in afferent pathways. J Neuroinflammation 2013, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Lamb, K.; Zhong, F.; Gebhart, G.F.; Bielefeldt, K. Experimental colitis in mice and sensitization of converging visceral and somatic afferent pathways. Am J Physiol Gastrointest Liver Physiol 2006, 290, G451–G457. [Google Scholar] [CrossRef] [PubMed]
- Atmani, K.; Wuestenberghs, F.; Baron, M.; Boulete, I.; Guerin, C.; Bahlouli, W.; Vaudry, D.; do Rego, J.C.; Cornu, J.N.; Leroi, A.M.; et al. Bladder-colon chronic cross-sensitization involves neuro-glial pathways in male mice. World J Gastroenterol 2022, 28, 6935–6949. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Hassan, A.M.; Koyani, C.N.; Mayerhofer, R.; Reichmann, F.; Farzi, A.; Schuligoi, R.; Malle, E.; Holzer, P. Behavioral and molecular processing of visceral pain in the brain of mice: impact of colitis and psychological stress. Front Behav Neurosci 2015, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, M.; Kempter, E.; Eleslambouly, T.; Lowry, C.A.; Langgartner, D.; Reber, S.O. Intranasal Mycobacterium vaccae administration prevents stress-induced aggravation of dextran sulfate sodium (DSS) colitis. Brain Behav Immun 2019, 80, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Price, D.D.; Caudle, R.M.; Verne, G.N. Visceral and somatic hypersensitivity in TNBS-induced colitis in rats. Dig Dis Sci 2008, 53, 429–435. [Google Scholar] [CrossRef]
- Zhou, Q.; Price, D.D.; Caudle, R.M.; Verne, G.N. Visceral and somatic hypersensitivity in a subset of rats following TNBS-induced colitis. Pain 2008, 134, 9–15. [Google Scholar] [CrossRef]
- Jain, P.; Materazzi, S.; De Logu, F.; Rossi Degl'Innocenti, D.; Fusi, C.; Li Puma, S.; Marone, I.M.; Coppi, E.; Holzer, P.; Geppetti, P.; et al. Transient receptor potential ankyrin 1 contributes to somatic pain hypersensitivity in experimental colitis. Sci Rep 2020, 10, 8632. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Han, S.; Huang, Q.; He, S.Q.; Ford, N.C.; Zheng, Q.; Chen, Z.; Yu, S.; Dong, X.; Guan, Y. Calcium imaging in population of dorsal root ganglion neurons unravels novel mechanisms of visceral pain sensitization and referred somatic hypersensitivity. Pain 2021, 162, 1068–1081. [Google Scholar] [CrossRef]
- Mecklenburg, J.; Zou, Y.; Wangzhou, A.; Garcia, D.; Lai, Z.; Tumanov, A.V.; Dussor, G.; Price, T.J.; Akopian, A.N. Transcriptomic sex differences in sensory neuronal populations of mice. Sci Rep 2020, 10, 15278. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.M.; Malek, N.; Edye, M.; Jager, S.B.; McMurray, S.; McMahon, S.B.; Denk, F. Sex differences in peripheral not central immune responses to pain-inducing injury. Sci Rep 2017, 7, 16460. [Google Scholar] [CrossRef]
- Vacca, V.; Marinelli, S.; Pieroni, L.; Urbani, A.; Luvisetto, S.; Pavone, F. 17beta-estradiol counteracts neuropathic pain: a behavioural, immunohistochemical, and proteomic investigation on sex-related differences in mice. Sci Rep 2016, 6, 18980. [Google Scholar] [CrossRef] [PubMed]
- Chernov, A.V.; Hullugundi, S.K.; Eddinger, K.A.; Dolkas, J.; Remacle, A.G.; Angert, M.; James, B.P.; Yaksh, T.L.; Strongin, A.Y.; Shubayev, V.I. A myelin basic protein fragment induces sexually dimorphic transcriptome signatures of neuropathic pain in mice. J Biol Chem 2020, 295, 10807–10821. [Google Scholar] [CrossRef]
- Tansley, S.; Uttam, S.; Urena Guzman, A.; Yaqubi, M.; Pacis, A.; Parisien, M.; Deamond, H.; Wong, C.; Rabau, O.; Brown, N.; et al. Single-cell RNA sequencing reveals time- and sex-specific responses of mouse spinal cord microglia to peripheral nerve injury and links ApoE to chronic pain. Nat Commun 2022, 13, 843. [Google Scholar] [CrossRef]
- Fiore, N.T.; Yin, Z.R.; Guneykaya, D.; Gauthier, C.D.; Hayes, J.; D'Hary, A.; Butovsky, O.; Moalem-Taylor, G. Sex-specific transcriptome of spinal microglia in neuropathic pain due to peripheral nerve injury. Glia 2022, 70, 675–696. [Google Scholar] [CrossRef]
- Parisien, M.; Samoshkin, A.; Tansley, S.N.; Piltonen, M.H.; Martin, L.J.; El-Hachem, N.; Dagostino, C.; Allegri, M.; Mogil, J.S.; Khoutorsky, A.; et al. Genetic pathway analysis reveals a major role for extracellular matrix organization in inflammatory and neuropathic pain. Pain 2019, 160, 932–944. [Google Scholar] [CrossRef]
- Stephens, K.E.; Chen, Z.Y.; Sivanesan, E.; Raja, S.N.; Linderoth, B.; Taverna, S.D.; Guan, Y. RNA-seq of spinal cord from nerve-injured rats after spinal cord stimulation. Mol Pain 2018, 14. [Google Scholar] [CrossRef]
- Ahlström, F.H.G.; Mätlik, K.; Viisanen, H.; Blomqvist, K.J.; Liu, X.; Lilius, T.O.; Sidorova, Y.; Kalso, E.A.; Rauhala, P.V. Spared Nerve Injury Causes Sexually Dimorphic Mechanical Allodynia and Differential Gene Expression in Spinal Cords and Dorsal Root Ganglia in Rats. Mol Neurobiol 2021, 58, 5396–5419. [Google Scholar] [CrossRef] [PubMed]
- Ghazisaeidi, S.; Muley, M.M.; Tu, Y.S.; Finn, D.P.; Kolahdouzan, M.; Pitcher, G.M.; Kim, D.; Sengar, A.S.; Ramani, A.K.; Brudno, M.; et al. Conserved transcriptional programming across sex and species after peripheral nerve injury predicts treatments for neuropathic pain. Brit J Pharmacol 2023, 180, 2822–2836. [Google Scholar] [CrossRef] [PubMed]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat Neurosci 2015, 18, 145–153. [Google Scholar] [CrossRef]
- Chiu, I.M.; Heesters, B.A.; Ghasemlou, N.; Von Hehn, C.A.; Zhao, F.; Tran, J.; Wainger, B.; Strominger, A.; Muralidharan, S.; Horswill, A.R.; et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature 2013, 501, 52. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.Y.; Grider, J.R. Colitis induces calcitonin gene-related peptide expression and Akt activation in rat primary afferent pathways. Exp Neurol 2009, 219, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Tiwari, N.; Madar, J.; Mehta, P.; Qiao, L.Y. Beta 2-adrenergic receptor mediates noradrenergic action to induce cyclic adenosine monophosphate response element-binding protein phosphorylation in satellite glial cells of dorsal root ganglia to regulate visceral hypersensitivity. Pain 2022, 163, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Delafoy, L.; Gelot, A.; Ardid, D.; Eschalier, A.; Bertrand, C.; Doherty, A.M.; Diop, L. Interactive involvement of brain derived neurotrophic factor, nerve growth factor, and calcitonin gene related peptide in colonic hypersensitivity in the rat. Gut 2006, 55, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, N.; Smith, C.; Sharma, D.; Shen, S.; Mehta, P.; Qiao, L.Y. Satellite glial cells drive colonic and somatic mechanical pain via upregulating Piezo2 in DRG neurons Cell Reports 2023, SSRN.
- Han, J.S.; Li, W.; Neugebauer, V. Critical role of calcitonin gene-related peptide 1 receptors in the amygdala in synaptic plasticity and pain behavior. J Neurosci 2005, 25, 10717–10728. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.C.; Xia, C.M.; Liu, M.; Shen, S.; Yu, S.J.; Chung, C.; Qiao, L.Y. Endogenous PI3K/Akt and NMDAR act independently in the regulation of CREB activity in lumbosacral spinal cord in cystitis. Exp Neurol 2013, 250, 366–375. [Google Scholar] [CrossRef]
- Presto, P.; Neugebauer, V. Sex Differences in CGRP Regulation and Function in the Amygdala in a Rat Model of Neuropathic Pain. Front Mol Neurosci 2022, 15, 928587. [Google Scholar] [CrossRef]
- Paige, C.; Plasencia-Fernandez, I.; Kume, M.; Papalampropoulou-Tsiridou, M.; Lorenzo, L.E.; David, E.T.; He, L.; Mejia, G.L.; Driskill, C.; Ferrini, F.; et al. A Female-Specific Role for Calcitonin Gene-Related Peptide (CGRP) in Rodent Pain Models. J Neurosci 2022, 42, 1930–1944. [Google Scholar] [CrossRef] [PubMed]
- Szabo-Pardi, T.A.; Barron, L.R.; Lenert, M.E.; Burton, M.D. Sensory Neuron TLR4 mediates the development of nerve-injury induced mechanical hypersensitivity in female mice. Brain Behav Immun 2021, 97, 42–60. [Google Scholar] [CrossRef] [PubMed]
- Inyang, K.E.; Szabo-Pardi, T.; Wentworth, E.; McDougal, T.A.; Dussor, G.; Burton, M.D.; Price, T.J. The antidiabetic drug metformin prevents and reverses neuropathic pain and spinal cord microglial activation in male but not female mice. Pharmacol Res 2019, 139, 1–16. [Google Scholar] [CrossRef]
- Saika, F.; Matsuzaki, S.; Kobayashi, D.; Ideguchi, Y.; Nakamura, T.Y.; Kishioka, S.; Kiguchi, N. Chemogenetic Regulation of CX3CR1-Expressing Microglia Using Gi-DREADD Exerts Sex-Dependent Anti-Allodynic Effects in Mouse Models of Neuropathic Pain. Front Pharmacol 2020, 11, 925. [Google Scholar] [CrossRef] [PubMed]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat Neurosci 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [PubMed]
- Tsujikawa, S.; DeMeulenaere, K.E.; Centeno, M.V.; Ghazisaeidi, S.; Martin, M.E.; Tapies, M.R.; Maneshi, M.M.; Yamashita, M.; Stauderman, K.A.; Apkarian, A.V.; et al. Regulation of neuropathic pain by microglial Orai1 channels. Sci Adv 2023, 9, eade7002. [Google Scholar] [CrossRef] [PubMed]
- Saika, F.; Matsuzaki, S.; Kishioka, S.; Kiguchi, N. Chemogenetic Activation of CX3CR1-Expressing Spinal Microglia Using Gq-DREADD Elicits Mechanical Allodynia in Male Mice. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Mapplebeck, J.C.S.; Dalgarno, R.; Tu, Y.; Moriarty, O.; Beggs, S.; Kwok, C.H.T.; Halievski, K.; Assi, S.; Mogil, J.S.; Trang, T.; et al. Microglial P2X4R-evoked pain hypersensitivity is sexually dimorphic in rats. Pain 2018, 159, 1752–1763. [Google Scholar] [CrossRef] [PubMed]
- Doyle, H.H.; Eidson, L.N.; Sinkiewicz, D.M.; Murphy, A.Z. Sex Differences in Microglia Activity within the Periaqueductal Gray of the Rat: A Potential Mechanism Driving the Dimorphic Effects of Morphine. J Neurosci 2017, 37, 3202–3214. [Google Scholar] [CrossRef]
- Hanamsagar, R.; Alter, M.D.; Block, C.S.; Sullivan, H.; Bolton, J.L.; Bilbo, S.D. Generation of a microglial developmental index in mice and in humans reveals a sex difference in maturation and immune reactivity. Glia 2018, 66, 460. [Google Scholar] [CrossRef]
- Chen, G.; Luo, X.; Qadri, M.Y.; Berta, T.; Ji, R.R. Sex-Dependent Glial Signaling in Pathological Pain: Distinct Roles of Spinal Microglia and Astrocytes. Neurosci Bull 2018, 34, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Xie, A.X.; Madayag, A.; Minton, S.K.; McCarthy, K.D.; Malykhina, A.P. Sensory satellite glial Gq-GPCR activation alleviates inflammatory pain via peripheral adenosine 1 receptor activation. Sci Rep 2020, 10, 14181. [Google Scholar] [CrossRef]
- Labus, J.S.; Gupta, A.; Coveleskie, K.; Tillisch, K.; Kilpatrick, L.; Jarcho, J.; Feier, N.; Bueller, J.; Stains, J.; Smith, S.; et al. Sex differences in emotion-related cognitive processes in irritable bowel syndrome and healthy control subjects. Pain 2013, 154, 2088–2099. [Google Scholar] [CrossRef]
- Hong, J.Y.; Kilpatrick, L.A.; Labus, J.; Gupta, A.; Jiang, Z.G.; Ashe-McNalley, C.; Stains, J.; Heendeniya, N.; Ebrat, B.; Smith, S.; et al. Patients with Chronic Visceral Pain Show Sex-Related Alterations in Intrinsic Oscillations of the Resting Brain. Journal of Neuroscience 2013, 33, 11994–12002. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, M.C.; Ceko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nature Reviews Neuroscience 2013, 14, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Han, G.S.; Domaille, D.W. Connecting the dynamics and reactivity of arylboronic acids to emergent and stimuli-responsive material properties. J Mater Chem B 2022, 10, 6263–6278. [Google Scholar] [CrossRef]
- Li, X.H.; Matsuura, T.; Xue, M.; Chen, Q.Y.; Liu, R.H.; Lu, J.S.; Shi, W.; Fan, K.; Zhou, Z.; Miao, Z.; et al. Oxytocin in the anterior cingulate cortex attenuates neuropathic pain and emotional anxiety by inhibiting presynaptic long-term potentiation. Cell Rep 2021, 36, 109411. [Google Scholar] [CrossRef]
- Monroe, T.B.; Fillingim, R.B.; Bruehl, S.P.; Rogers, B.P.; Dietrich, M.S.; Gore, J.C.; Atalla, S.W.; Cowan, R.L. Sex Differences in Brain Regions Modulating Pain Among Older Adults: A Cross-Sectional Resting State Functional Connectivity Study. Pain Med 2018, 19, 1737–1747. [Google Scholar] [CrossRef]
- Wang, G.; Erpelding, N.; Davis, K.D. Sex differences in connectivity of the subgenual anterior cingulate cortex. Pain 2014, 155, 755–763. [Google Scholar] [CrossRef]
- Osborne, N.R.; Cheng, J.C.; Rogachov, A.; Kim, J.A.; Hemington, K.S.; Bosma, R.L.; Inman, R.D.; Davis, K.D. Abnormal subgenual anterior cingulate circuitry is unique to women but not men with chronic pain. Pain 2021, 162, 97–108. [Google Scholar] [CrossRef]
- Liu, R.H.; Xue, M.; Li, X.H.; Zhuo, M. Sex difference in synaptic plasticity in the anterior cingulate cortex of adult mice. Mol Brain 2020, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Bristow, G.C.; Bostrom, J.A.; Haroutunian, V.; Sodhi, M.S. Sex differences in GABAergic gene expression occur in the anterior cingulate cortex in schizophrenia. Schizophr Res 2015, 167, 57–63. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of hind paw mechanical sensitivity in male (a) and female (b) mice after TNBS-induced colitis. Male: control n=6, colitis n=7; female day 4: control n=5; colitis n=10; female day 7 and 21: control n=7, colitis n=7. Two-way ANOVA with Tukey’s multiple comparison test. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001.
Figure 1.
Comparison of hind paw mechanical sensitivity in male (a) and female (b) mice after TNBS-induced colitis. Male: control n=6, colitis n=7; female day 4: control n=5; colitis n=10; female day 7 and 21: control n=7, colitis n=7. Two-way ANOVA with Tukey’s multiple comparison test. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001.
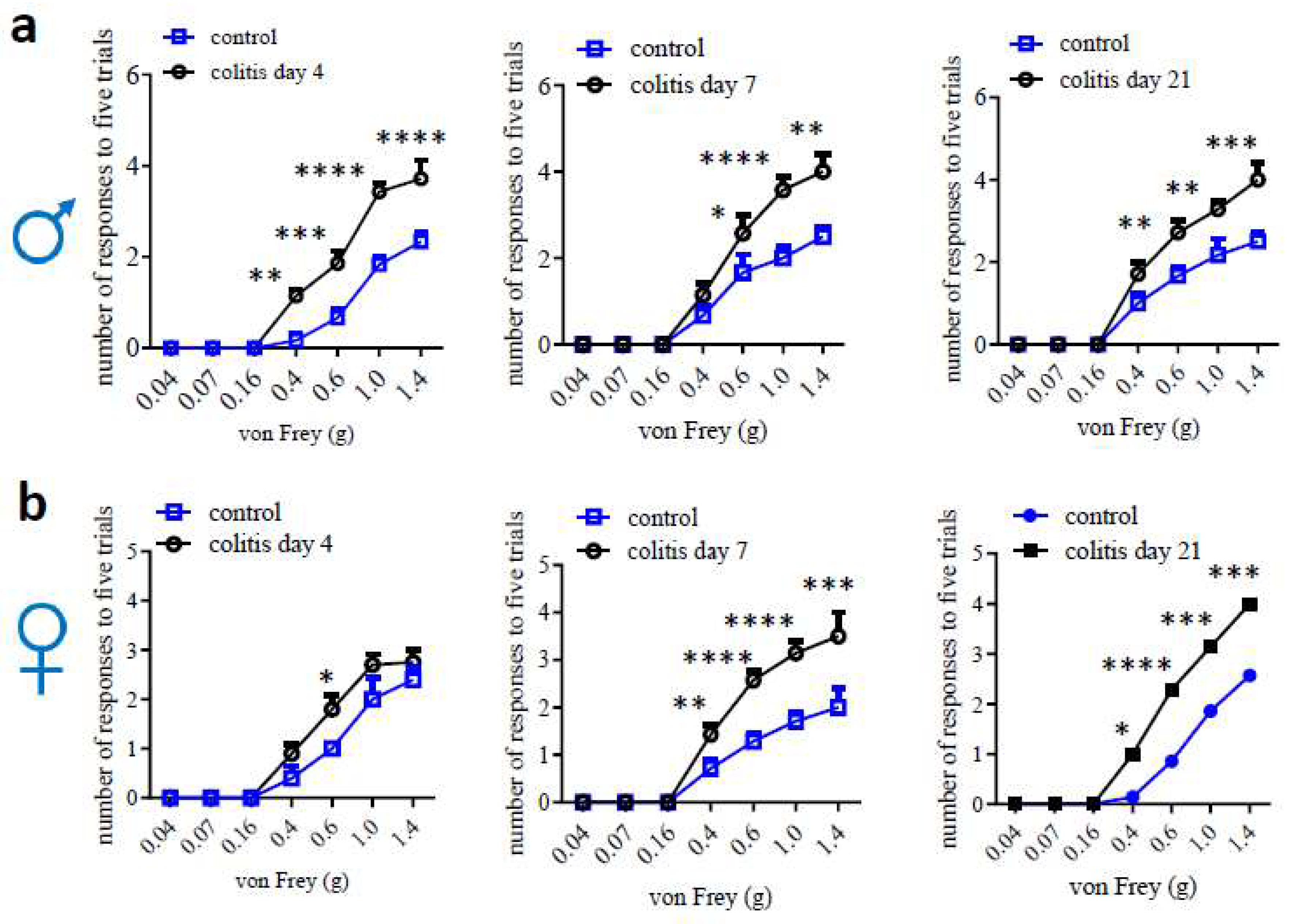

Table 1.
Sex differential outcomes of colonic inflammation, hypersensitivity and comorbidities. In general, colitis and associated visceral hypersensitivity (VH) are more severe in male (M) than in female (F) rodents. At baseline and VH not related to inflammation, F (especially rats) have higher visceral sensitivity than M. Under certain situation, both sexes develop VH and colitis-associated referred hypersensitivity and pain.
Table 1.
Sex differential outcomes of colonic inflammation, hypersensitivity and comorbidities. In general, colitis and associated visceral hypersensitivity (VH) are more severe in male (M) than in female (F) rodents. At baseline and VH not related to inflammation, F (especially rats) have higher visceral sensitivity than M. Under certain situation, both sexes develop VH and colitis-associated referred hypersensitivity and pain.
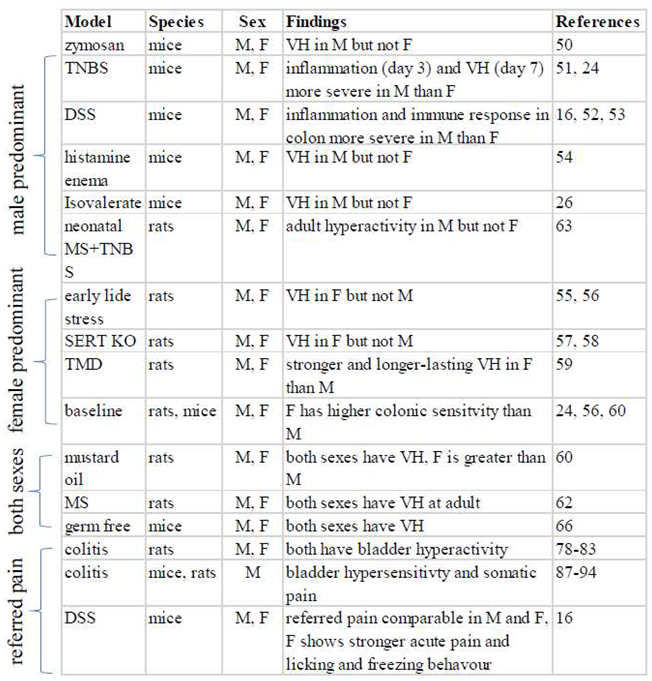 |
Table 2.
Transcriptome analysis of tissues involved in pain processing to shed lights on sex differences at molecular levels. SC: spinal cord; DRG: dorsal root ganglia; SN: sciatic nerves.
Table 2.
Transcriptome analysis of tissues involved in pain processing to shed lights on sex differences at molecular levels. SC: spinal cord; DRG: dorsal root ganglia; SN: sciatic nerves.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



