
Submitted:
29 December 2023
Posted:
03 January 2024
You are already at the latest version
Abstract
The agricultural sector faces challenges due to climate change and the growing global population. Alkaline stress could adversely impact plant growth and crop production. This stressor diminishes water quality essential for crop cultivation, consequently impairing plant growth and overall productivity. Gamma-aminobutyric acid (GABA), a non-protein amino acid, may exhibit multifaceted roles in plant responses to adverse environmental conditions. Optimization of crop production in controlled environmental agriculture under artificial light attracted much attention. In the present study, we investigated the effects of different light spectra and GABA concentrations on Lollo Rosso lettuce plants under alkaline stress. Seedlings were placed under different light spectra [red (R), blue (B), white (W), 3R:1B, and 2R:1B:1far-red (FR)] and treated by GABA (0 and 50 µmol). Alkaline conditions were instituted by applying NaHCO3 at a concentration of 40 mM. The experiment incorporated a control treatment without NaHCO3. In general, R light exposure caused the highest growth performance of lettuce plants. GABA augmented biomass production in control and alkaline-treated plants across all light spectra. Additionally, it increased the concentrations of photosynthetic and protective pigments under alkaline conditions. GABA also improved the photosynthetic performance under all light spectra. Non-photochemical quenching and pigmentation were enhanced by exposure to B light, especially in plants treated with GABA. Alkaline stress induced an increase in soluble carbohydrate content in the lettuce leaves. These findings highlight the support of GABA application in facilitating coping plants to alkaline stress.
Keywords:
Abiotic stress
; Controlled environment agriculture
; Light quality
; Photosynthesis
; Stress tolerance
1. Introduction
In the face of climate change and an ever-expanding global population, the agricultural sector has been driven to embrace innovative practices to ensure the continuous supply of fresh produce while maintaining environmental sustainability. Plants confront various environmental constraints, including salinity, alkalinity, drought, extreme temperatures, heavy metal toxicity, flooding, ultraviolet radiation, and ozone exposure [1]. These abiotic stresses disrupt plants' growth, physiology, and overall functionality, posing significant challenges to crop yield and global food security. Key to this endeavor is a comprehensive understanding of plant responses and the effective management of stress factors that impact crop growth and development. Stresses pose a significant threat to crop growth and productivity. In this pursuit of resilient agriculture, a particular stressor that warrants attention is alkalinity stress [1]. Alkaline conditions in the soil and irrigation water can severely hamper plant growth and productivity [1,2]. Excessive alkalinity in water can disrupt the delicate pH balance essential for proper nutrient uptake in plants, leading to nutrient deficiencies and reduced growth. Moreover, elevated alkalinity can trigger oxidative stress by impairing the plant's ability to regulate reactive oxygen species (ROS), causing cellular damage and hindering overall physiological functions [1]. Studying alkalinity in hydroponic culture even when adjusting pH with acids, remains important for the following reasons: the alkalinity level affects the capacity of the solution to resist changes in pH. Understanding the alkalinity level provides insights into the system's buffering capacity and its ability to maintain a stable pH over time [3]. Alkalinity affects the availability, solubility, and precipitation of various nutrients. While adjusting pH with acids may address immediate pH concerns, studying alkalinity helps in understanding long-term nutrient interactions and potential nutrient imbalances that may arise due to alkalinity effects. By studying alkalinity, growers can optimize nutrient management strategies to ensure proper nutrient uptake and avoid imbalances. Alkalinity is a critical parameter for assessing water quality suitability for hydroponic systems. Water sources with high alkalinity may require additional treatments or adjustments to achieve optimal growing conditions [4]. Therefore, exploring novel strategies for alleviating this stress and promoting crop resilience is essential.
Plant growth and productivity is an intricate interplay between external and internal factors. Environmental factors like light, temperature, nutrients, and water profoundly influence crop growth [2]. Controlled environment agriculture (CEA) has been introduced to produce crops safely [5]. However, optimization of environmental conditions, including lighting and growing environments in CEA, is a challenging issue that has attracted the attention of scientists and plant growers [5]. Optimal manipulation of artificial light in CEA, especially in vertical and indoor farms, is critical to the successful and cost-effective production of crops in those environments. Moreover, removing the alkalinity problem in the irrigation water for crop production in CEA is vital in the region where water quality is not optimum.
Gamma-aminobutyric acid (GABA) is a natural non-protein amino acid that plays a crucial role in plants metabolism. While traditionally known for its significance as an inhibitory neurotransmitter in animals and humans, GABA's role as a signaling molecule in plants has gained attention in recent years. It is involved in various physiological and biochemical processes, mediating responses to environmental stresses [6]. When subjected to alkaline conditions, plants can synthesize and accumulate higher levels of GABA. This increase in GABA content helps mitigate the damaging effects of alkaline stress. GABA is a versatile molecule in this context, playing a dual role. First, it functions as an osmolyte, regulating cellular water balance and preventing water loss from plant cells under alkaline stress conditions [6,7]. Second, GABA acts as a signaling molecule, activating various stress-responsive genes and pathways. It aids in enhancing the plant's defence mechanisms against oxidative stress, reducing the production of harmful ROS, and maintaining cellular redox balance. Moreover, GABA enhances the activity of antioxidant enzymes, such as superoxide dismutase, catalase, and peroxidases, thus providing additional protection against oxidative damage induced by stresses. By modulating these physiological processes, GABA helps plants maintain cellular homeostasis and adapt to alkaline stress conditions, ultimately promoting better growth and survival [8,9]. GABA's involvement in mitigating the adverse effects of stress highlights its importance as a plant stress regulator. Understanding the mechanisms by which GABA confers plant protection under alkaline conditions can pave the way for developing more resilient crop varieties and sustainable agricultural practices to tackle environmental challenges.
Amidst the quest for strategies to bolster crop productivity in CEA systems under artificial light when water quality is not optimum, attempts to increase yields while facing such challenges have often met with limited success. Nevertheless, mitigating the impact of these stressors on plants remains a perpetual endeavor, prompting the exploration of innovative approaches.
The response of plants to light, in particular, hinges on diverse factors, encompassing light quality, environmental conditions, seasonal variations, genotype, cultivation techniques, and more. One such avenue of investigation lies in harnessing the potential of different light spectra in CEA systems. Light-emitting diodes (LEDs) are widely used in CEAs to compensate for the lack of sunlight in closed CEAs. Red (R), blue (B), and farred red (FR) attracted as much attention as the quality of light sources in indoor farming systems. However, the interaction between light quality and other environmental factors is still a matter of investigation and needs further elucidation.
The current scientific inquiry delves into GABA's positive influence in alleviating the adverse effects of alkalinity stress on Lollo Rosso lettuce plants under diverse light spectra. The study aimed to shed light on a novel approach that could potentially fortify the resilience and productivity of Lollo Rosso lettuce when confronted with alkalinity-induced stress in CEA, thus contributing to the broader understanding of plant stress responses and their implications for agricultural practices.
2. Materials and Methods
2.1. Plant materials and growth conditions
Eleven days old Lollo Rosso lettuce (Lactuca sativa) seedlings grown under white light were transferred to different cabinets equipped with various LED light qualities and irrigated with a Hoagland solution [10]. The plants were placed in a growth medium containing equal proportions of cocopeat and perlite. The average temperature of 25 ± 2 °C and relative humidity of 60±10% were kept in the growth cabinets.
2.2. Treatments
To investigate the effect of GABA and alkalinity stress under different light spectra, the seedlings were subjected to different light spectra, including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), a combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio. Light-emitting diode (LED) modules (Parcham Co, Tehran, Iran) were used as the light source to have different light spectra in growth cabinets. The plants were exposed to a 16/8 h light/dark period. The light intensity in each chamber was carefully fixed and maintained at 250 ± 20 µmol m-2 s-1 photosynthetic photon flux density (PPFD). The intensity and spectral quality of the light were meticulously monitored using a spectrometer (Sekonic C-7000, Tokyo, Japan). Following the light treatments, at four stages, the seedlings were pre-treated with two levels of GABA (0 and 50 µmol L-1) into the Hoagland's nutrient solution in a three-day interval time-scale before applying alkaline stress. Subsequently, alkalinity stress was induced using two levels of NaHCO3 (0 and 40 mM) every three days (a total of approximately 12 applications of alkalinity stress).
2.3. Morphological properties
Upon completion of the experiment and subsequent harvest, fresh weight (FW) and dry weight (DW) of different plant parts, including roots, stems, and leaves, were meticulously measured. Firstly, the leaves and stems were carefully detached from the plant base. Any excess substrate clinging to the roots was meticulously removed. Then, the FW of leaves, roots, and stems were determined. Subsequently, DW was recorded following placing plant materials in an oven at 75 °C for 72 h.
2.4. Assessment of fast induction of chlorophyll fluorescence
The fully grown leaves of Lollo Rosso lettuce plants underwent a 20-minute incubation in total darkness. To determine the polyphasic chlorophyll (Chl) fluorescence transients (OJIP test), a Fluorpen FP 100-MAX device (Photon Systems Drasov, Czech Republic) was employed. The assessment of OJIP transient parameters followed by well-established methodologies as previously reported in earlier studies [11,12,13,14].
2.5. Imaging of chlorophyll fluorescence quenching
Samples were collected from fully matured leaves of Lollo Rosso lettuce cultivated under different treatments. These samples were subjected to a dark-adaptation period of 20 min before being immediately utilized for the measurement of slow induction of chlorophyll fluorescence using a FluorCam (FluorCam FC 1000-H, Photon Systems Instruments, PSI, Drasov, Czech Republic). The FluorCam was equipped with a high-resolution CCD camera and four fixed LED panels, which were utilized to provide precise measuring pulses and induce saturating flashes. Maximum quantum efficiency of photosystem II (Fv/Fm) was calculated using a designed protocol [15]. In brief, chlorophyll fluorescence measurement commenced by subjecting the samples to short flashes in total darkness. Subsequently, high light intensity (3900 µmol m-2 s-1) was applied at the end of fast measuring flashes to halt the electron transport resulting from the reduction of quinone acceptors. This meticulous protocol allowed for recording two sets of fluorescence data: one averaged throughout short flashes in darkness (Fo), and the other recorded at the time of exposure to the saturating flash (Fm).
To determine non-photochemical quenching (NPQ), the maximum fluorescence in the light-adapted steady state (Fm') and dark-adapted state (Fm) were recorded and subsequently utilized for NPQ measure [16]. All relevant data and intricate calculations were meticulously performed using version 7 of the FluorCam software (PSI, Drasov, Czech Republic).
2.6. Pigments analysis
To investigate the impact of light spectra, GABA, and alkaline stress on photosynthetic pigment levels (including Chl a, b, total (Chla + Chlb), and carotenoids), the methodology outlined by Lichtenthaler’s protocol [17] was employed. To do so, leaf tissue samples (100 mg) were taken and homogenized in 10 mL of 80% acetone. The mixture was then incubated overnight at 4 °C in total darkness to avoid potential light interference. Following incubation, the supernatant was obtained by centrifuging the sample at 6000 rpm for 10 min at a constant temperature of 25 °C. Subsequently, spectrophotometric readings were taken at wavelengths of 646 nm, 663 nm, and 470 nm to assess the absorbance levels.
2.7. Measurement of total anthocyanin content
Total anthocyanin content was quantified using a methanol-formic acid (98:2, v/v) extraction solution. Initially, 25 ml of this solution was meticulously added to vials containing 500 mg of powdered lettuce leaves to facilitate the comprehensive extraction of anthocyanins. Subsequently, the vials underwent a brief ultrasonic bath for 2 min, followed by centrifugation (SIGMA 1-14K, Darmstadt, Germany) at 2500 rpm for 5 min at 25 °C. The conclusive step involved the quantification of anthocyanins, which was achieved by applying calibration curves derived from malvidin-3-glucoside [18].
2.8. Quantitative Analysis of Carbohydrates
Well-developed leaves were collected to determine the concentration of soluble carbohydrates in lettuce. The assessment of total soluble carbohydrates was conducted using the anthrone procedure [19]. Initially, samples of lettuce leaves were ground meticulously in liquid nitrogen. Then, 0.2 g of tissue was extracted and combined with 7 mL of 70% ethanol (w/v) for a duration of 5 min while being kept on ice. Subsequently, centrifugation at 6700× g for 10 min at 4 °C was carried out to separate the components effectively. To initiate the spectrophotometric analysis, 200 mL of the supernatant was mixed with 1 mL of an anthrone solution consisting of 0.5 g of anthrone, 250 mL of 95% H2SO4, and 12.5 mL of distilled water. The resulting mixture was subjected to spectrophotometric recording at 625 nm (PerkinElmer, Lambda 25, USA).
2.9. Statistical analysis
This study was conducted in a completely randomized factorial design, comprising three factors: pre-treatment with GABA at two levels (0 and 50 µM), light spectra at five levels, and alkalinity stress at two levels (0 and 40 mM) with three replicants for each treatment. After data collection, analysis of variance (ANOVA) and subsequent comparisons of means were conducted employing Duncan's multiple range test at a significance level of 0.01. The statistical analysis was undertaken expertly using the sophisticated SAS software (Statistical Analysis System, version 9.4), ensuring the utmost reliability and precision in the obtained results.
3. Results
3.1. Morphological characteristics of lettuce plant were affected by light spectra, alkaline stress and GABA feeding
The results of this investigation on the FW and DW of leaves stems, and roots of cultivated Lollo Rosso lettuce revealed a significant difference among the treatments (Figure 1, 2). In general, the FW and DW of the leaves exhibited a similar pattern among the treatments. The highest leaf FW and DW were obtained in the plants exposed to R light under 50 µmol GABA and in absence of alkanility. Leaf FW and DW were 5.3 and 6.7 times higher, respectively, compared to the FW and DW in plants exposed to B light under stress (40 mM NaHCO3) and no GABA application, which represented the lowest leaf FW and DW (Figure 1, A, B). Generally, the FW and DW of leaves in mature plants grown under treatments containing GABA and without stress showed higher levels than those under other treatments.
The highest stem FW was recorded in the plant grown under 2R:1B:1FR light treatment containing 50 µmol GABA with no stress (Figure 1, C). Concurrently, the stem FW in alkaline-exposed plants grown under B, 3R:1B, and W light treatments without GABA application did not exhibit a significant difference and allocated the lowest amount of stem FW.
The highest stem DW was observed in the plants grown under 2R:1B:1FR light in the absence of alkalinity and GABA applications, which was 13 times more than the stem DW of alkaline-exposed plants that were grown under W light without GABA application (Figure 1, D). Root FW and DW developed better under R light in plants exposed to GABA and alkaline conditions (Figure 1, E, F). These values are approximately 3.5 and 4.5 times higher than the FW and DW of roots recorded in plants grown under B light and alkaline stress without GABA application, respectively.
Figure 2.
Biomass partitioning in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) and 0 and 40 mM NaHCO3 as alkaline stress.
Figure 2.
Biomass partitioning in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) and 0 and 40 mM NaHCO3 as alkaline stress.

Biomass partitioning is a critical analysis that provides insights into a plant's resource allocation strategy and its response to environmental factors. Plants grown in the presence of GABA and under stress-free conditions directed their maximum biomass to the aerial organs, a trend observed under all light treatments. In contrast, plants subjected to stress treatment without GABA allocated more biomass to the underground organs across all light treatments. Furthermore, light treatments containing FR in the lighting environment devoted higher biomass to the stem compartment of the lettuce plant by limiting biomass allocation to the leaf or root organs (Figure 3).
3.2. Photosynthetic performance positively affected by GABA
To investigate the influence of different combinations of light spectra and GABA application under alkaline stress on the photosynthetic functionality of Lollo Rosso lettuce, polyphasic Chl a fluorescence (OJIP) analysis was conducted on dark-adapted leaves. The main objective of this study was to explore how different ratios of light spectra, along with GABA application, affected the photosynthetic performance of the plants under alkaline stress.
The maximum quantum yield of PSII (Fv/Fm) was significantly (P ≤ 0.001) influenced by the different treatments, as depicted in Figure 4. Fv/Fm showed a noticeable decline in plants exposed to alkaline stress without GABA application (Figure 4). Remarkably, no significant distinctions were observed between the 2R:1B:1FR and 3R:1B light spectra. In particular, the plants treated with 50 µmol GABA without alkaline stress exhibited a substantially higher Fv/Fm than those under other treatments. On the other hand, the B and R light treatment without GABA application and under 40 mM alkaline stress displayed the lowest Fv/Fm, while in the same conditions, plants exposed to combinational light recepies including 2R:1B:1FR and 3R:1B were less affected.
The performance index for the photochemical activity (PIabs) is an essential indicator of the PSII validity index. Under all light spectra, PIabs was at the highest in non-stress exposed plants treated with GABA, while its lowest was recorded in R and W light growth (Figure 5). Plants grown under light quality containing FR and B lights showed the highest PIabs. No significant differences in the PIabs were found among plants exposed to B and 2R:1B:1FR lights.
The investigation revealed that changes in the light spectrum significantly affected the energy fluxes at each reaction center (RC). Through the analysis of Chl a fluorescence output during rapid inductions, calculations were made to establish specific energy fluxes per RC related to energy absorption, commonly referred to as ABS/RC (Table 1). Among the treatments, the peak value for ABS/RC was detected in plants treated with monochromatic B light without GABA feeding under alkaline stress. This result indicated the highest recorded value, nearly three times more significant than the ABS/RC detected in plants grown under 3R:1B and 2R:1B:1FR, fed by 50 µmol GABA without alkaline stress.
Similarly, the results of the TR0/RC parameter, which represents trapped energy flux per RC, showed the highest level in the plants exposed to B light without GABA under alkaline stress, which led to 1.5 times higher TR0/RC than the plants exposed to 2R:1B:1FR light treatment and fed by 50 µmol GABA without alkaline stress. The ET0/RC serves as a pivotal metric, symbolizing the comprehensive efficacy of electron transportation, representing electron transport flux per reaction center. The highest ET0/RC was detected in plants exposed to B light under stress in 0 and 50 µmol GABA treatments (Table 1). The lowest level of this parameter was detected in plants grown under 2R:1B:1FR light treatment containing GABA and under non-stress conditions. The highest level of ET0/RC was 1.5 times higher than the lowest recorded level. The highest level of dissipated energy flux per each reaction center (DI0/RC) was obtained in plants grown under B light without GABA under alkaline stress, which was almost three times higher than the DI0/RC calculated from plants grown under 2R:1B:1FR light treatment containing GABA without stress conditions as the lowest of this parameter (Table 1).
The values of energy fluxes were derived from the analysis of OJIP transients. Light spectra included red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730), a combination of R and B light in a 3R:1B ratio, and a combination of R and B along with far red (FR, 700-750 nm) in a 2R:1B:1FR ratio, maintained at a fixed light intensity of 250 ± 10 µmol m-2 s-1. Fertigation was performed with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress for plants under each light spectrum.
In a column, means followed by the same letters are not significantly different at p ≤ 0.05, as determined by Duncan's multiple range test.
3.3. B light and GABA enhanced NPQ in lettuce leaves
The NPQ is a crucial physiological photoprotection mechanism obtained through fluorescence analysis, which is used to assess a plant's response to environmental stressors. Significant differences were detected among the NPQ of plants under different treatments (Figure 6). Under alkaline stress conditions, the highest NPQ was seen in plants exposed to B light and GABA under stress conditions. Generally, the highest and lowest NPQ under all light spectra were detected in plants exposed to alkaline stress with GABA feeding and in plants under alkaline stress without GABA feeding, respectively.
3.4. Photosynthetic pigments influenced by GABA feeding and different light spectra
The analysis of the obtained data revealed significant differences in the content of Chl a, Chl b, and total Chl. The highest Chl a content was detected in plants exposed to 2R.1B:1FR light and fed by 50 µmol L-1 GABA under stress-free conditions. This value was almost twice higher than the amount of Chl a, obtained from plants grown under 3R:1B with no GABA and subjected to stress (Figure 7. A). Similarly, Chl b concentration reached its peak in plants exposed to 2R.1B:1FR light and fed by 50 µmol L-1 GABA under no alkaline stress, exhibiting an approximately threefold increase compared to Chl b concentration in plants exposed to 3R:1B light with no GABA under alkaline stress (Figure 7. B). Consequently, the total Chl content yielded consistent results, with the highest and lowest concentrations observed in the plants cultivated under 2R.1B:1FR light with 50 µmol GABA and stress-free conditions.
Moreover, carotenoid accumulation significantly increased in stress-free plants and treated with 50 µmol GABA in W and 2R:1B:1FR light treatments. Conversely, the lowest carotenoid concentration was recorded in the leaves of plants grown under R light and treated with 50 µmol GABA under alkaline stress conditions (Figure 7. D).
3.5. Total anthocyanins were increased by GABA application and combinational light
The concentration of total anthocyanins in Lollo Rosso lettuce leaves exhibited a remarkable increase with the application of GABA (Figure 8). The leaves of plants grown under the 3R:1B and 2R:1B:1FR lights and treated with 50 µmol GABA supplementation under stress-free conditions showed the highest total anthocyanins. This amount was five times greater than the total anthocyanin content extracted from leaves of plants grown under W light treatment without GABA supplementation under stress conditions. Under all light qualities, an increase in the concentration of anthocyanins was detected in the leaves of lettuce plants treated with GABA and alkaline stress.
3.6. Carbohydrate levels were accumulated due to GABA feeding and red light exposure
Measurements of the soluble carbohydrate levels were conducted to investigate the impact of various light spectra and GABA feeding on the accumulation of soluble carbohydrates in Lollo Rosso lettuce leaves under alkaline conditions. The results demonstrated that the different treatments significantly influenced the carbohydrate content (Figure 9). Notably, carbohydrates were primarily accumulated in those plants exposed to light recipies containing R light. The highest concentration of soluble carbohydrates was observed in the two combined light treatments, 3R:1B and 2R:1B:1FR, both with the addition of GABA and under stress-free conditions, which showcased a remarkable 12-fold increase compared to the soluble carbohydrate content in plants grown under B light, without the addition of GABA and under stress conditions.
4. Discussion
4.1. Red light promoted the growth of lettuce, and GABA reduced the negative impact of alkaline stress on biomass production
The results of this study provide insights into how growing light quality and GABA feeding influenced the growth and morphological traits of Lollo Rosso lettuce under alkaline stress. R light induces leaf and root growth, while FR light induces stem growth in lettuce plants (Figure 1, Figure 2 and Figure 3). A R light's promotive effect on plant growth induction has been widely reported [20,21]. However, phytochromes become inactive when exposed to FR light due to phytochrome conversion. Due to these conversions, the phytohormone gibberellin would accumulate, leading to stem elongation [22]. This response is also common when plants compete for light, such as the dense canopy and plant population. This typical low light response of plants has been named shade avoidance response [23]. This response occurs when other light spectra, such as Chl pigments, absorb R and B lights, and FR is the primary light spectrum, penetrates inside the dense canopy. Consequently, it induces stem elongation, enabling plants to cope with low light exposure [24].
We observed significant effects on the FW and DW of the leaf, stem, and root by employing different light spectra along with GABA application. These findings shed light on potential strategies to enhance the resilience and productivity of lettuce in challenging alkaline conditions. The incorporation of GABA has demonstrated a noticeable enhancement in leaf production for plants cultivated under stress and non-stress light conditions. Light treatments incorporating R light and GABA application exhibited the most substantial increase in the growth of lettuce plants. Conversely, the findings regarding root FW and DW suggested that under stress conditions, the introduction of GABA resulted in a remarkable boost in root production. It can be recommended that GABA-induced mechanisms contribute to the plant's ability to resist and adapt to stressful conditions. This is by findings from literature that GABA, as an essential signaling molecule in plants, has multifunctional roles in stress response pathways [8,25,26,27]. Under alkaline stress conditions, plants experience disruptions in nutrient uptake, impaired ion balance, and oxidative damage, which can hamper growth and survival. Studies have demonstrated that exogenous application of GABA can mitigate the detrimental effects of alkaline stress by enhancing plant tolerance mechanisms [6,8,27].
Root systems serve as the primary interface between plants and the soil environment. GABA treatment has been found to stimulate lateral root development and elongation, enabling plants to explore a larger soil volume and efficiently absorb water and essential nutrients even under alkaline conditions [6]. Additionally, GABA-induced root growth helps to stabilize the plant in the soil, thereby reducing the risk of uprooting due to soil alkalinity-induced instability.
In the present study, GABA application caused a significant increase in leaf biomass production in plants grown under stress-free and alkaline stress conditions. Leaves are the primary sites of photosynthesis, and their productivity is critical for sustaining plant growth and yield [25,26]. Alkaline stress often reduces photosynthetic efficiency and chlorophyll degradation, decreasing leaf production and premature leaf senescence. However, GABA has been found to upregulate the expression of stress-responsive genes involved in photosynthesis, thus maintaining a higher leaf area and improved chlorophyll content under alkaline stress conditions. This enhanced leaf production improves carbon assimilation and plant vigor [28,29]. GABA signalling activates specific pathways, such as the GABA shunt, which produces reactive oxygen species (ROS) scavengers and osmolytes [26]. This helps alleviate oxidative stress and stabilise cell membranes, thus preserving cell integrity under alkaline stress. Additionally, GABA-triggered signalling cascades modulate the expression of stress-related genes, transcription factors, and hormone signalling pathways, orchestrating a comprehensive response that enhances plant tolerance to alkaline stress [6].
4.2. Red light and alkalinity down-regulated photosynthetic performance, while GABA application enhanced it in lettuce leaves
In the present study, growing plants under monochromatic R light reduced the photosynthetic performance of lettuce plants. In contrast, photosynthetic performance was enhanced by B light or when FR light was included in the growing light environment (Figure 4 and Figure 5). It has been reported that long-term exposure to monochromatic R light reduces the uploading of carbohydrates into the sink organs, which induces feedback inhibition on the photosynthetic performance of the plants [30]. The reduced photosynthetic performance due to monochrome R light exposure has been reported in chrysanthemum, basil, saffron, rose flower and many other species [12,13,30,31,32]. On the other hand, an enhanced photosynthetic performance due to B light has also been reported. Therefore, the photosynthetic apparatus would be down-regulated due to the accumulation of carbohydrates in the source organs (leaves). B light probably increases the expression and activity of Rubisco, fructose-1, 6-bisphosphatase and glyceraldehyde-phosphate dehydrogenase enzymes involved in the Calvin cycle [6]. R light increases the biomass of roots, but B light increases the growth of leaves. Therefore, blue light improves photosynthetic efficiency [30].
The present study also showed that the application of GABA reduced the adverse effects of alkaline stress on the photosynthetic performance of lettuce plants. The results obtained demonstrated a significant improvement in Fv/Fm and PIabs upon the addition of GABA. This enhancement suggests that GABA plays a pivotal role in promoting the overall photosynthetic efficiency of the organism [33]. Plants grown under FR and B lights showed the highest PIabs. Although the plants grown under B and R light showed the lowest Fv/Fm value under alkaline stress conditions, in the same conditions, plants exposed to FR and 3R:1B lights were less affected by alkaline stress. It has been shown that FR light can improve abiotic stress tolerance in diverse plant species [34,35].
As an indicator of photosynthetic functionality, Fv/Fm is a fundamental parameter used to assess the maximum efficiency of PSII, reflecting the organism's ability to convert light energy into chemical energy during photosynthesis [11]. The observed increase in Fv/Fm in the presence of GABA indicates that adding GABA enhances the organism's capacity to capture and utilize light energy efficiently, potentially leading to improved biomass production and growth. PIabs provides valuable insights into photosynthetic efficiency by measuring the overall performance of PSII under natural conditions [11]. The notable increase in PIabs following GABA supplementation further supports the notion that GABA positively influences the photosynthetic machinery, likely by facilitating electron transport or optimizing the energy conversion process, especially under stress conditions [36]. The observed effects of GABA on photosynthetic performance are consistent with previous studies that have explored the role of GABA in enhancing plant stress tolerance, growth, and yield [37,38,39]. GABA has been reported to act as a signaling molecule in various physiological processes, including stress responses, carbon and nitrogen metabolism, and modulation of ion transport. Consequently, the positive impact of GABA on photosynthetic efficiency may be attributed to its multifaceted role in cellular signaling and regulation of metabolic pathways [8,26,29].
4.3. Blue light and GABA enhanced tolerance mechanisms against alkaline stress
When plants experience stresses, such as excess light or drought conditions, they protect themselves from potential damage caused by overexcitation of the photosynthetic machinery [16]. One of the protective mechanisms plants employ is the conversion of excess absorbed light energy into heat, a process known as NPQ. The NPQ was investigated to understand the effect of light spectra and GABA on increasing the tolerance of lettuce plants to alkaline stress. The results revealed that the highest NPQ values were observed under B light across all conditions, both in the presence and absence of GABA. Furthermore, the introduction of GABA significantly augmented the NPQ in all light treatments under alkaline stress compared to stress-free conditions (Fig. 6). These compelling findings underscore the pivotal roles played by both B light and GABA in inducing plant tolerance to alkaline stress. One of the critical mechanisms by which GABA increases plant tolerance to alkaline stress is its role as an osmoregulator. Alkaline stress can disrupt the osmotic balance within plant cells, leading to cell dehydration and damage [6,26]. GABA helps maintain osmotic balance by acting as an osmolyte. It accumulates in the plant cells and increases the solute concentration, allowing them to retain water and maintain their turgor pressure, even under alkaline stress conditions [6]. This osmoregulatory function prevents cell dehydration and damage, thus enhancing the plant's ability to withstand stress. GABA also participates in mitigating oxidative damage caused by alkaline stress. Under stressful conditions, ROS accumulate in plant cells, leading to oxidative stress and cellular damage [40]. GABA acts as an antioxidant by scavenging ROS and inhibiting lipid peroxidation, protecting cell membranes and cellular components from oxidative harm. This antioxidant activity helps maintain plant cells' integrity and improves tolerance to alkaline stress. Furthermore, GABA plays a significant role in modulating the activity of ion channels and transporters under alkaline stress. It has been observed that GABA enhances the activity of specific ion transporters, particularly those involved in the uptake of essential nutrients like potassium (K+) and calcium (Ca2+). This increased uptake of essential nutrients helps maintain ion homeostasis and ensures proper cellular functions even in high alkaline conditions [8,28,29,40]. Additionally, GABA can regulate ion fluxes across cell membranes, which aids in maintaining the pH balance and minimizing the impact of alkaline stress on cellular processes.
It was suggested that B light plays a significant role in increasing plant tolerance to alkaline stress through various physiological and biochemical mechanisms [2,41,43]. When exposed to B light, plant photoreceptors, such as cryptochromes and phototropins, are activated, initiating a signaling cascade that triggers responses to combat alkaline stress [41]. One crucial effect of B light is its influence on stomatal regulation. Under alkaline stress, plants tend to close their stomata, reducing water loss through transpiration [2,43]. B light optimizes the stomatal aperture, ensuring proper gas exchange and minimizing water loss, thus mitigating the adverse effects of alkaline stress [2]. Moreover, B light exposure induces the upregulation of antioxidant enzymes and molecules, which help scavenge harmful ROS produced under alkaline stress. This protection against oxidative damage is vital for preserving the integrity of plant cells and their photosynthetic machinery [42]. In addition to these effects, B light influences gene expression, activating stress-responsive genes. These genes are involved in the synthesis of stress-related proteins, osmolytes, and other protective molecules, bolstering the plant's ability to withstand alkaline stress [40,43]. Furthermore, B light can modulate ion channels and transporters in plant cells, maintaining proper ion homeostasis under alkaline stress conditions. This regulation is crucial for preserving cellular turgor pressure and osmotic balance, which are essential for plant growth and survival under stressful conditions [43].
4.4. GABA induced pigment and carbohydrate accumulation in lettuce plants
In the current study, GABA induced the highest concentrations of photosynthetic pigments, including Chl a, b, total, and carotenoid. It has been suggested that GABA positively affects the production and accumulation of chlorophyll a, chlorophyll b, and carotenoids in plants [26]. GABA's role as a signaling molecule and stress-responsive compound is crucial in this enhancement. Under stress conditions, GABA stimulates the expression of genes involved in the synthesis of photosynthetic pigments [8]. This upregulation increases enzymatic activity in the chlorophyll and carotenoid biosynthetic pathways, resulting in higher levels of chlorophyll a, chlorophyll b, and carotenoids. Additionally, GABA's antioxidant properties protect these pigments from oxidative damage, ensuring their stability and longevity within the plant cells under stressful conditions. Consequently, GABA positively influences the efficiency of photosynthesis, leading to improved plant growth and productivity under stress conditions [7].
Yu et al., (2022); reported GABA’s positive affects on the total anthocyanin production and accumulation in plants [44]. The results of the current study confirmed this effect under all light treatments under both stress and non-stress conditions. Anthocyanins are pigments responsible for various plant tissues' vibrant R, purple, and B colors [7,26]. When GABA is applied or accumulated in plant cells, it acts as a signaling molecule that triggers the upregulation of genes involved in anthocyanin biosynthesis. This increased gene expression leads to enhanced production of anthocyanins. GABA can also influence the activity of enzymes responsible for anthocyanin synthesis, further boosting their accumulation [45]. The overall result is a significant increase in the levels of anthocyanins, which not only add vibrant colors to the plants but also serve as powerful antioxidants, protecting the plant from oxidative stress and promoting its resilience to environmental challenges [26].
The observed increase in soluble carbohydrate content due to GABA application in alkaline stress (Figure 9) can be attributed to a well-known adaptive response known as osmotic adjustment. Alkaline stress alters the pH of the soil, making essential nutrients less available to plants. This disruption in nutrient uptake leads to an imbalance in the plant's osmotic potential, causing water to move out of the plant cells [2]. As a result, the plant experiences a water deficit or dehydration, which can harm its survival. Plants initiate osmotic adjustment to counteract this water loss and maintain cellular turgor pressure. Osmotic adjustment is a process in which the plant accumulates solutes, such as soluble carbohydrates (e.g., sugars and starches) and other organic compounds, in the cytoplasm of their cells [1]. GABA can act as an osmolyte, accumulating in plant cells when water availability is limited. This accumulation helps maintain turgor pressure and prevents wilting under drought stress. On the other hand, GABA can influence ion transport, affecting the uptake of ions like potassium (K+) and calcium (Ca2+), which are essential for osmotic regulation [7]. GABA can also influence stomatal closure, reducing water loss through transpiration, which is another mechanism of osmotic adjustment [25,26]. Additionally, GABA can contribute to the accumulation of soluble carbohydrates in leaves by its influence on carbon and nitrogen metabolism, which can provide a source of energy and osmotic solutes to support plant growth and survival under challenging conditions [8,28,29].
5. Conclusions
Growing Lollo Rosso lettuce plants under red light and supplying GABA enhanced biomass gain under alkaline and non-stress conditions. FR inclusion in the growing light environment led to the direction of biomass toward the stem. The best photosynthetic performance was recorded in plants treated by GABA, especially when exposed to alkaline stress. The highest NPQ was detected in plants grown under B light and provided by 50 µmol GABA in both stress and non-stress conditions. This indicates enhancement of plant tolerance mechanisms through exposure to B light and feeding with GABA. The highest concentration of pigments was recorded in plants fed by GABA. Plants grown in environment with high proportion of R light in their overall spectrum or provided by GABA showed an elevated soluble carbohydrate content in their leaves. Although this carbohydrate accumulation caused a feedback inhibition response on photosynthesis, this is considered an adaptive response called osmotic adjustment, preserving water potential, supporting metabolic processes, and enhancing the plant's survival and tolerance to alkaline stress as the result of GABA application. These findings highlight the significant role of GABA in improving photosynthetic efficiency, and plant tolerance to alkaline stress in CEA production of lettuce plants under artificial lights.
References
- Fang, S.; Hou, X.; Liang, X. Response Mechanisms of Plants Under Saline-Alkali Stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Shamsabad, M.R.M.; Esmaeilizadeh, M.; Roosta, H.R.; Dehghani, M.R.; Dąbrowski, P.; Kalaji, H.M. The effect of supplementary light on the photosynthetic apparatus of strawberry plants under salinity and alkalinity stress. Sci. Rep. 2021, 12, 13257. [Google Scholar] [CrossRef]
- Anderson, T.S.; Martini, M.R.; De Villiers, D.; Timmons, M.B. Growth and tissue elemental composition response of Butterhead lettuce (Lactuca sativa, cv. Flandria) to hydroponic conditions at different pH and alkalinity. Hortic. 2017, 3, 41. [Google Scholar] [CrossRef]
- Boyd, C.E.; Tucker, C.S.; Somridhivej, B. Alkalinity and hardness: Critical but elusive concepts in aquaculture. J. World Aquacult. Soc. 2016, 47, 6–41. [Google Scholar] [CrossRef]
- Lastochkina, O.; Aliniaeifard, S.; SeifiKalhor, M.; Bosacchi, M.; Maslennikova, D.; Lubyanova, A. Novel approaches for sustainable horticultural crop production: Advances and prospects. Hortic. 2022, 8, 910. [Google Scholar] [CrossRef]
- Li, Y.; Liu, B.; Peng, Y.; Liu, C.; Zhang, X.; Zhang, Z.; Liang, W.; Ma, F.; Li, C. Exogenous GABA alleviates alkaline stress in Malus hupehensis by regulating the accumulation of organic acids. Sci. Hortic. 2020, 261. [Google Scholar] [CrossRef]
- Ullah, A.; Ali, I.; Noor, J.; Zeng, F.; Bawazeer, S.; Eldin, S.M.; Asghar, M.A.; Javed, H.H.; Saleem, K.; Ullah, S.; Ali, H. Exogenous γ-aminobutyric acid (GABA) mitigated salinity-induced impairments in mungbean plants by regulating their nitrogen metabolism and antioxidant potential. Front. Plant Sci. Front. Plant Sci. 2023, 13, 1081188. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Jalil, S.U.; Chopra, P.; Chhillar, H.; Ferrante, A.; Khan, N.A.; Ansari, M.I. Role of GABA in plant growth, development and senescence. Plant Gene 2021, 26, 100283. [Google Scholar] [CrossRef]
- Ramos-Ruiz, R.; Martinez, F.; Knauf-Beiter, G. The effects of GABA in plants. Cogent Food Agr. 2019, 5, 1670553. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil: University of California. Circular. Calif. Agric. Exp. Stn. 1938, 347, 32. [Google Scholar]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi-Nezhad, S.M. How glycine betaine induces tolerance of cucumber plants to salinity stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef]
- Hosseini, A.; Zare Mehrjerdi, M.; Aliniaeifard, S.; Seif, M. Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiol. Mol. Biol. Pla. 2019, 25, 741–752. [Google Scholar] [CrossRef]
- Moradi, S.; Kafi, M.; Aliniaeifard, S.; Salami, S.A.; Shokrpour, M.; Pedersen, C.; Moosavi-Nezhad, M.; Wrobel, J.; Kalaji, H.M. Blue light improves photosynthetic performance and biomass partitioning toward harvestable organs in saffron (Crocus sativus L.). Cells 2021, 10, 1994. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 25, 445–483. [Google Scholar]
- Aliniaeifard, S.; Malcolm Matamoros, P.; van Meeteren, U. Stomatal malfunctioning under low VPD conditions: Induced by alterations in stomatal morphology and leaf anatomy or in the ABA signaling? Physiol. Plantarum 2014, 152, 688–699. [Google Scholar] [CrossRef]
- Shomali, A.; Aliniaeifard, S.; Bakhtiarizadeh, M.R.; Lotfi, M.; Mohammadian, M.; Sadi, M.S.V.; Rastogi, A. Artificial neural network (ANN)-based algorithms for high light stress phenotyping of tomato genotypes using chlorophyll fluorescence features. Plant Physiol. Bioch. 2023, 201, 107893. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Wagner, G.J. Content and Vacuole/Extravacuole Distribution of Neutral Sugars, Free Amino Acids, and Anthocyanin in Protoplasts. Plant Physiol. 1979, 64, 88–93. [Google Scholar] [CrossRef]
- Van Doorn, W.G. Water relations of cut flowers: An update. Hortic. Rev. 2012, 40, 55–106. [Google Scholar]
- Zhang, Y.; Ji, J.; Song, S.; Su, W.; Liu, H. Growth, nutritional quality and health-promoting compounds in Chinese Kale grown under different ratios of red: Blue LED lights. Agronomy 2020, 10, 1248. [Google Scholar] [CrossRef]
- Oh, H.E.; Yoon, A.; Park, Y.G. Red light enhances the antioxidant properties and growth of Rubus hongnoensis. Plants 2021, 10, 2589. [Google Scholar] [CrossRef]
- Jin, W.; Urbina, J.L.; Heuvelink, E.; Marcelis, L.F. Adding far-red to red-blue light-emitting diode light promotes yield of lettuce at different planting densities. Front. Plant Sci. 2021, 11, 609977. [Google Scholar] [CrossRef]
- Martinez-Garcia, J.F.; Gallemi, M.; Molina-Contreras, M.J.; Llorente, B.; Bevilaqua, M.R.; Quail, P.H. The shade avoidance syndrome in Arabidopsis: The antagonistic role of phytochrome A and B differentiates vegetation proximity and canopy shade. PLoS ONE 2014, 9, 109275. [Google Scholar] [CrossRef]
- Ortiz-Alcaide, M.; Llamas, E.; Gomez-Cadenas, A.; Nagatani, A.; Martinez-Garcia, J.F.; Rodriguez-Concepcion, M. Chloroplasts modulate elongation responses to canopy shade by retrograde pathways involving HY5 and abscisic acid. Plant Cell 2019, 31, 384–398. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Aliniaeifard, S.; Hassani, B.; Niknam, V.; Lastochkina, O. Diverse role of γ-aminobutyric acid in dynamic plant cell responses. Plant Cell Rep. 2019, 38, 847–867. [Google Scholar] [CrossRef]
- Ramos-Ruiz, R.; Martinez, F.; Knauf-Beiter, G. The effects of GABA in plants. Cogent Food Agric. 2019, 5. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef]
- Barbosa, J.M.; Locy, R.D.; Barger, T.W.; Singh, N.K.; Cherry, J.H. GABA Increases the Rate of Nitrate Uptake and Utilization in Arabidopsis Roots. Plant Tolerance to Abiotic Stresses in Agricu: Role of Genetic Engineering 2000, 53–63. [CrossRef]
- Pan, Y.; Shen, Y.; Zhang, H.; Ran, X.; Xie, T.; Zhang, Y.; Yao, C. Fine-tuned regulation of photosynthetic performance via γ-aminobutyric acid (GABA) supply coupled with high initial cell density culture for economic starch production in microalgae. Bioresources Bioproc. 2022, 9, 1–18. [Google Scholar] [CrossRef]
- Moosavi-Nezhad, M.; Alibeigi, B.; Estaji, A.; Gruda, N.S.; Aliniaeifard, S. Growth, biomass partitioning, and photosynthetic performance of chrysanthemum cuttings in response to different light spectra. Plants 2022, 11, 3337. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, ply052. [Google Scholar] [CrossRef]
- Cheng, P.; Yue, Q.; Zhang, Y.; Zhao, S.; Khan, A.; Yang, X.; He, J.; Wang, S.; Shen, W.; Qian, Q.; Du, W. Application of γ-aminobutyric acid (GABA) improves fruit quality and rootstock drought tolerance in apple. J. Plant Physiol. 2023, 280, 153890. [Google Scholar] [CrossRef] [PubMed]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Enviro. 2021, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Malekzadeh Shamsabad, M.R.; Esmaeilizadeh, M.; Roosta, H.R.; Dąbrowski, P.; Telesinski, A.; Kalaji, H.M. Supplemental light application can improve the growth and development of strawberry plants under salinity and alkalinity stress conditions. Sci. Rep. 2022, 12, 9272. [Google Scholar] [CrossRef] [PubMed]
- Seifikalhor, M.; Aliniaeifard, S.; Bernard, F.; Seif, M.; Latifi, M.; Hassani, B.; Didaran, F.; Bosacchi, M.; Rezadiist, H.; Li, T. γ-Aminobutyric acid confers cadmium tolerance in maize plants by concerted regulation of polyamine metabolism and antioxidant defense systems. Sci. Rep. 2020, 10, 3356. [Google Scholar] [CrossRef] [PubMed]
- Kalhor, M.S.; Aliniaeifard, S.; Seif, M.; Asayesh, E.J.; Bernard, F.; Hassani, B.; Li, T. Enhanced salt tolerance and photosynthetic performance: Implication of γ -amino butyric acid application in salt-exposed lettuce (Lactuca sativa L.) plants. Plant Physiol. Bioch. 2018, 130, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Kaspal, M.; Kanapaddalagamage, M.H.; Ramesh, S.A. Emerging roles of γ aminobutyric acid (GABA) gated channels in plant stress tolerance. Plants 2021, 10, 2178. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Wu, X.; Gong, B.; Huo, R.; Zhao, L.; Li, J.; Li, J.; Lu, G.; Gao, H. GABA Metabolism, Transport and Their Roles and Mechanisms in the Regulation of Abiotic Stress (Hypoxia, Salt, Drought) Resistance in Plants. Metabolites 2023, 13, 347. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P. GABA shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Carvalho, R.F.; Campos, M.L.; Azevedo, R.A. The role of phytochrome in stress tolerance. J. Integr. Plant Biol. 2011, 53, 920–929. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biote. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Wang, F.F.; Wang, M.H.; Zhang, M.K.; Qin, P.; Cuthbertson, A.G.S.; Lei, C.L.; Qiu, B.L.; Yu, L.; Sang, W. Blue light stimulates light stress and phototactic behavior when received in the brain of Diaphorina citri. Ecotox. Environ. Saf. 2023, 251, 114519. [Google Scholar] [CrossRef]
- Yu, G.; Chen, F.; Wang, Y.; Chen, Q.; Liu, H.; Tian, J.; Wang, M.; Ren, C.; Zhao, O.; Yang, F.; Sheng, Y. Exogenous γ-aminobutyric acid strengthens phenylpropanoid and nitrogen metabolism to enhance the contents of flavonoids, amino acids, and the derivatives in edamame. Food Chem. X 2022, 16, 100511. [Google Scholar] [CrossRef]
- Ren, T.; Zheng, P.; Zhang, K.; Liao, J.; Xiong, F.; Shen, Q.; Ma, Y.; Fang, W.; Zhu, X. Effects of GABA on the polyphenol accumulation and antioxidant activities in tea plants (Camellia sinensis L.) under heat-stress conditions. Plant Physiol. Bioch. 2021, 159, 363–371. [Google Scholar] [CrossRef]
Figure 1.
Morphological characteristics in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
Figure 1.
Morphological characteristics in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
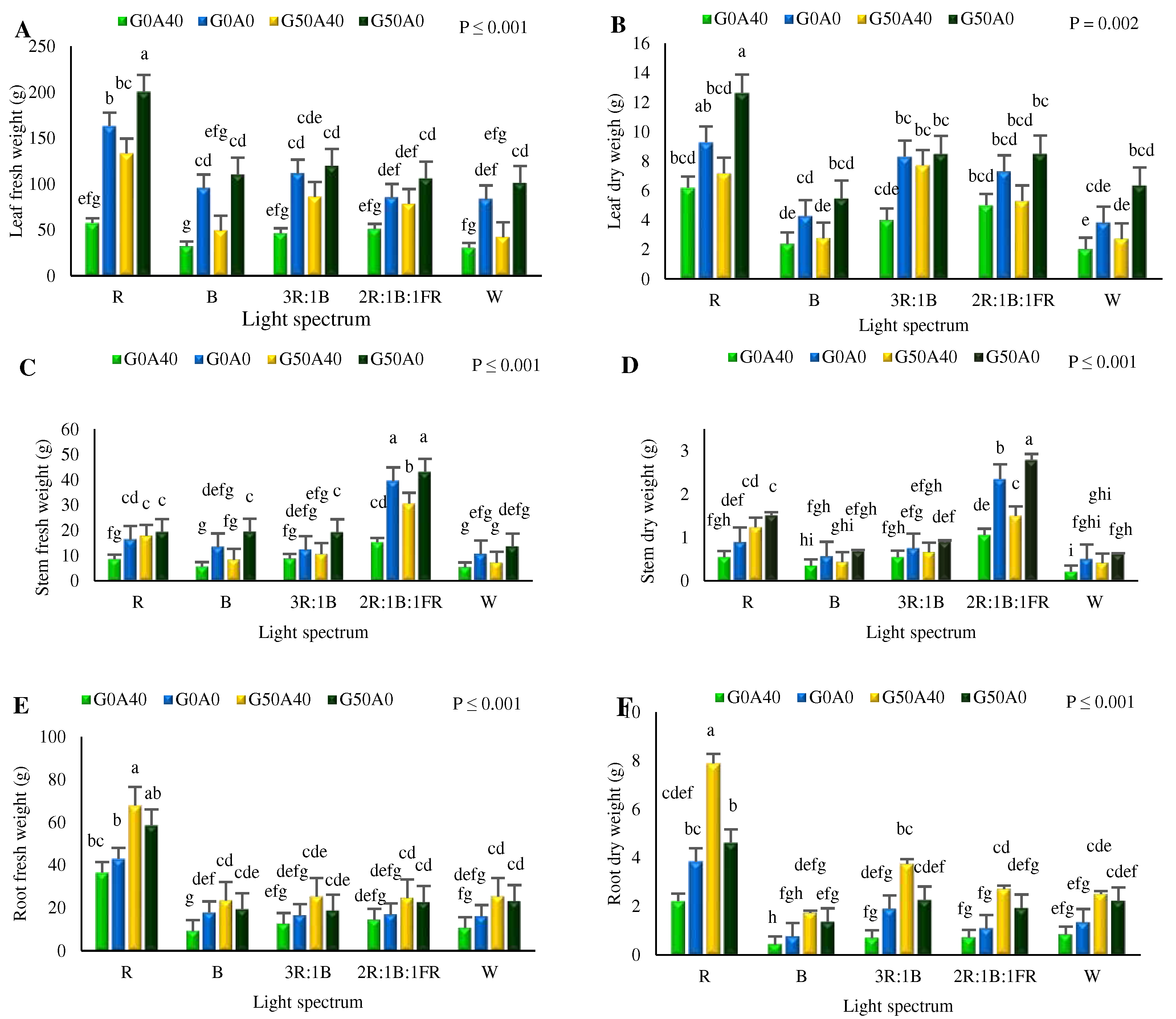
Figure 3.
Biomass partitioning in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) and 0 and 40 mM NaHCO3 as alkaline stress.
Figure 3.
Biomass partitioning in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) and 0 and 40 mM NaHCO3 as alkaline stress.
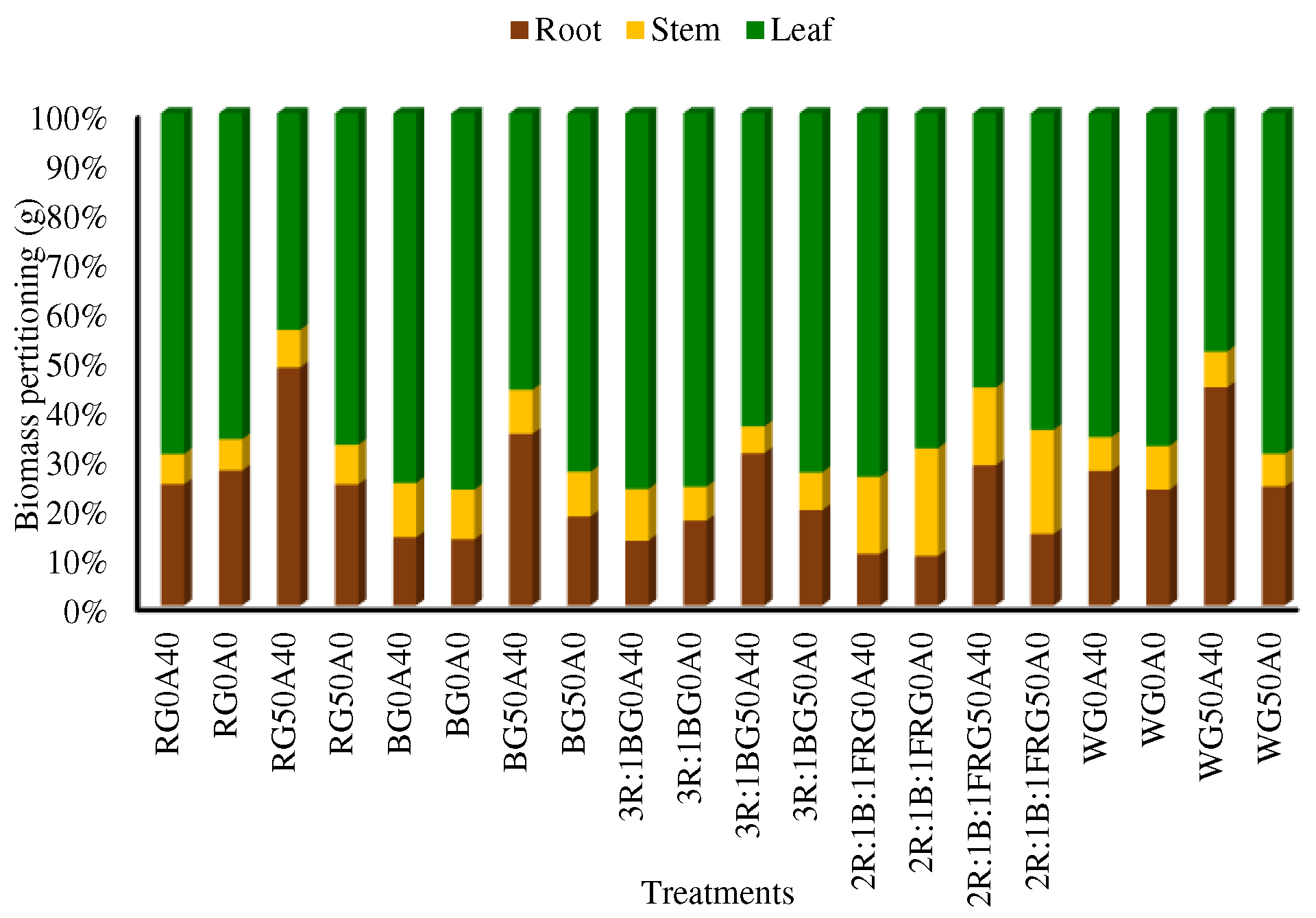
Figure 4.
The corresponding image of the maximum quantum yield of photosystem II (Fv/Fm) in Lollo Rosso lettuce plants under different light spectra, including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
Figure 4.
The corresponding image of the maximum quantum yield of photosystem II (Fv/Fm) in Lollo Rosso lettuce plants under different light spectra, including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
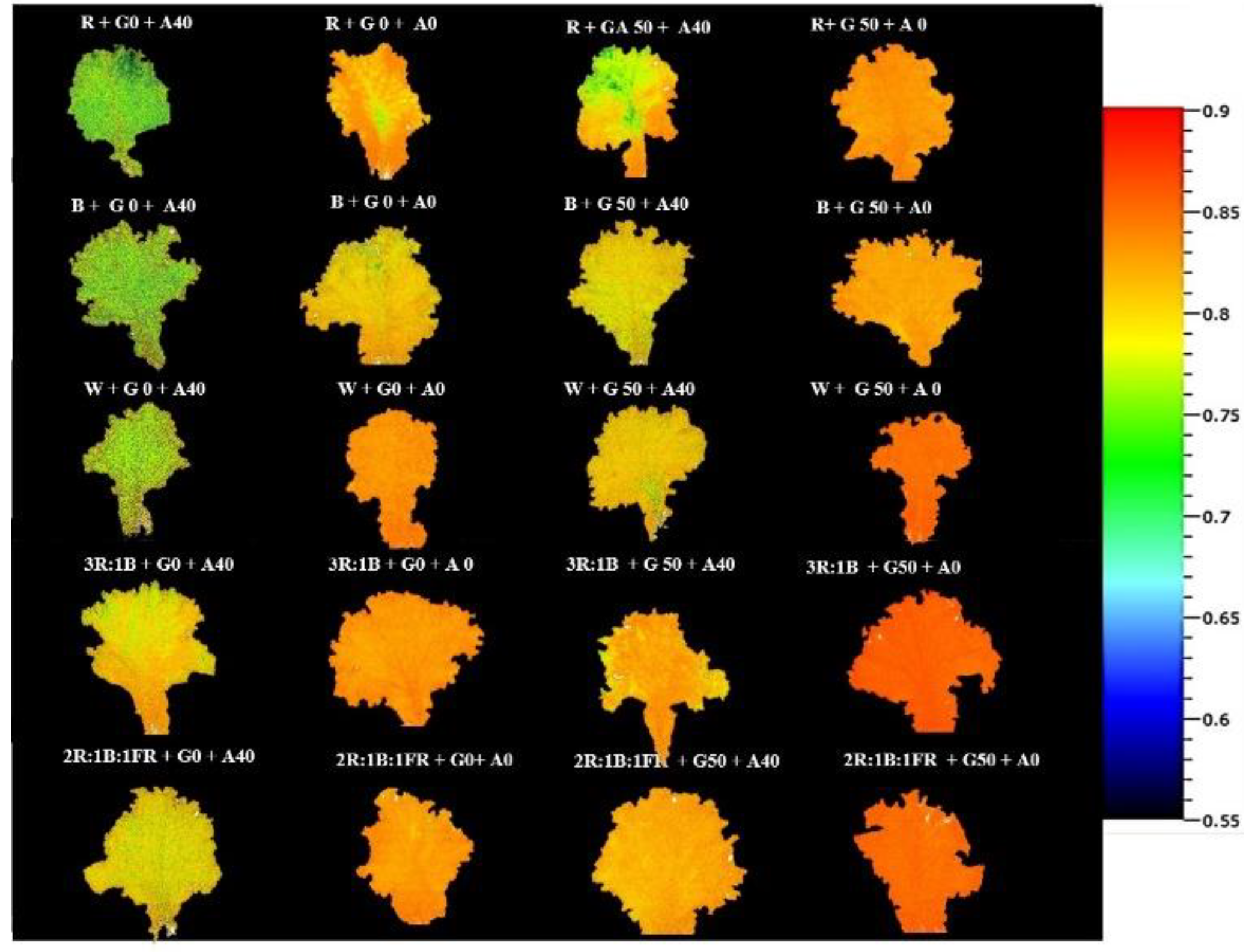
Figure 5.
Performance index per absorbed light (PIabs) in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
Figure 5.
Performance index per absorbed light (PIabs) in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress.
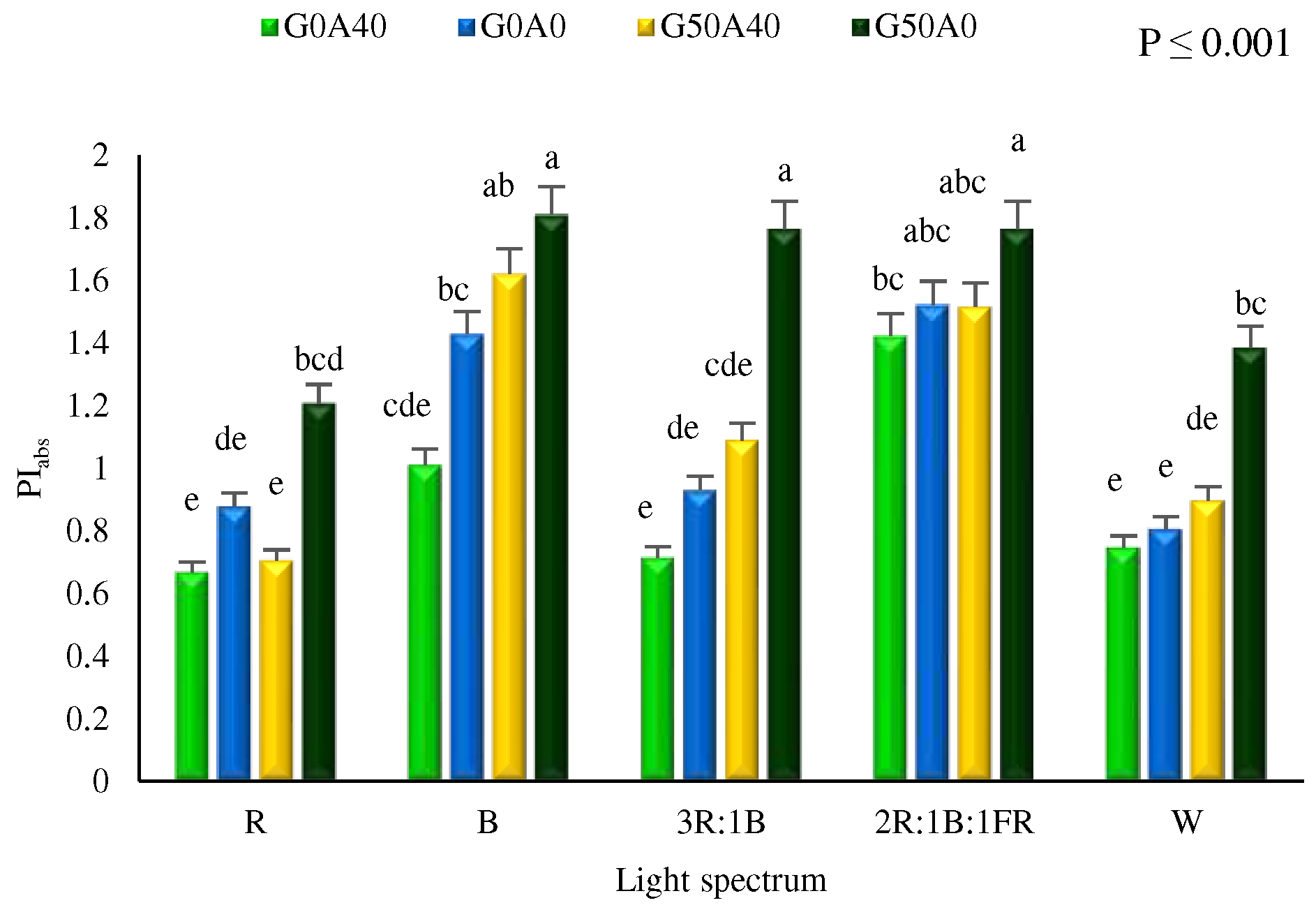
Figure 6.
The Non-Photochemical Quenching (NPQ) of chlorophyll fluorescence in Lollo Rosso lettuce plants under different light spectra, including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
Figure 6.
The Non-Photochemical Quenching (NPQ) of chlorophyll fluorescence in Lollo Rosso lettuce plants under different light spectra, including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
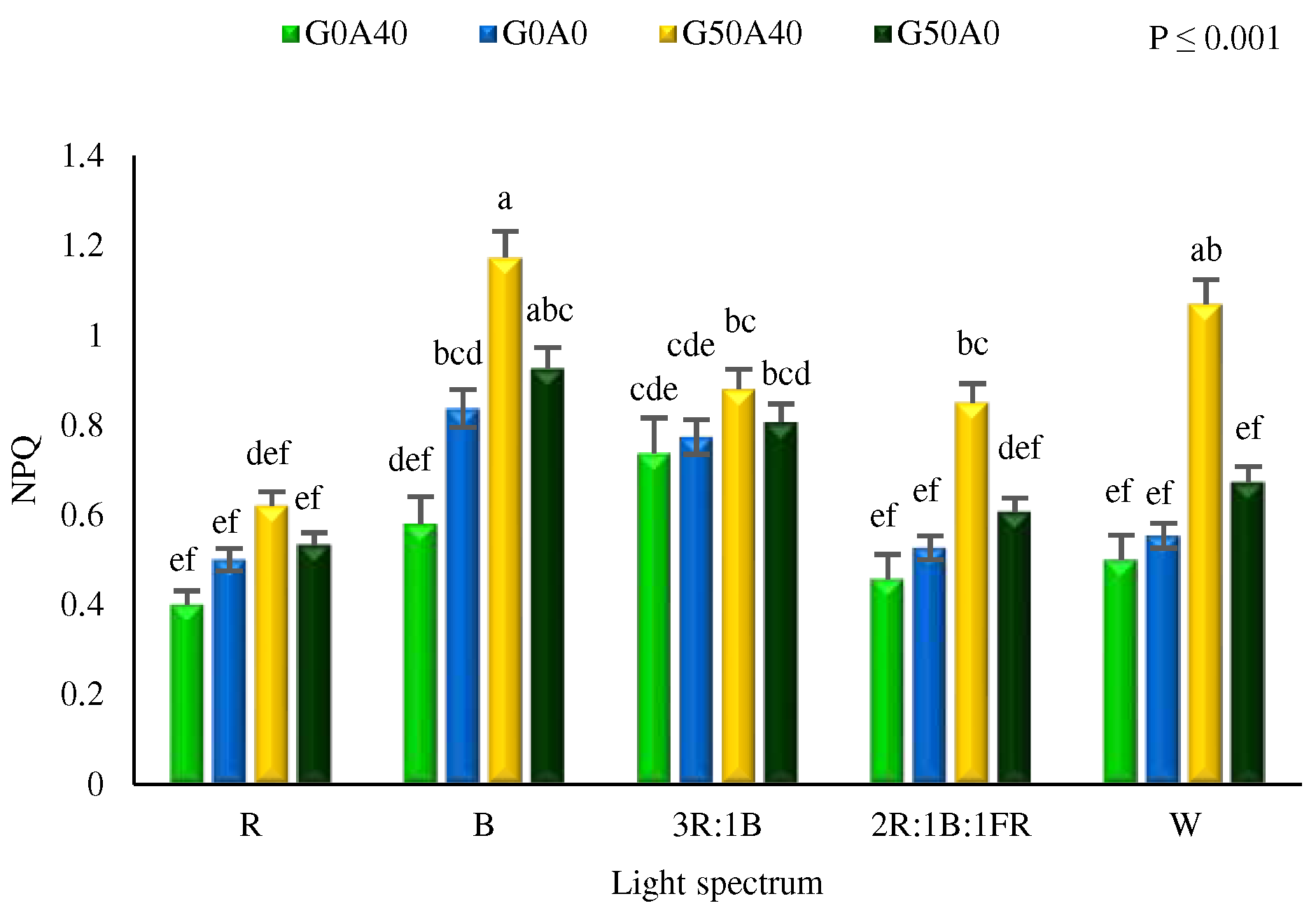
Figure 7.
Chlorophyll a (A; Chl a), Chlorophyll b (B; Chl b), total chlorophyll (C; Total Chl), and Carotenoid (D) contents in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
Figure 7.
Chlorophyll a (A; Chl a), Chlorophyll b (B; Chl b), total chlorophyll (C; Total Chl), and Carotenoid (D) contents in Lollo Rosso lettuce plants under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
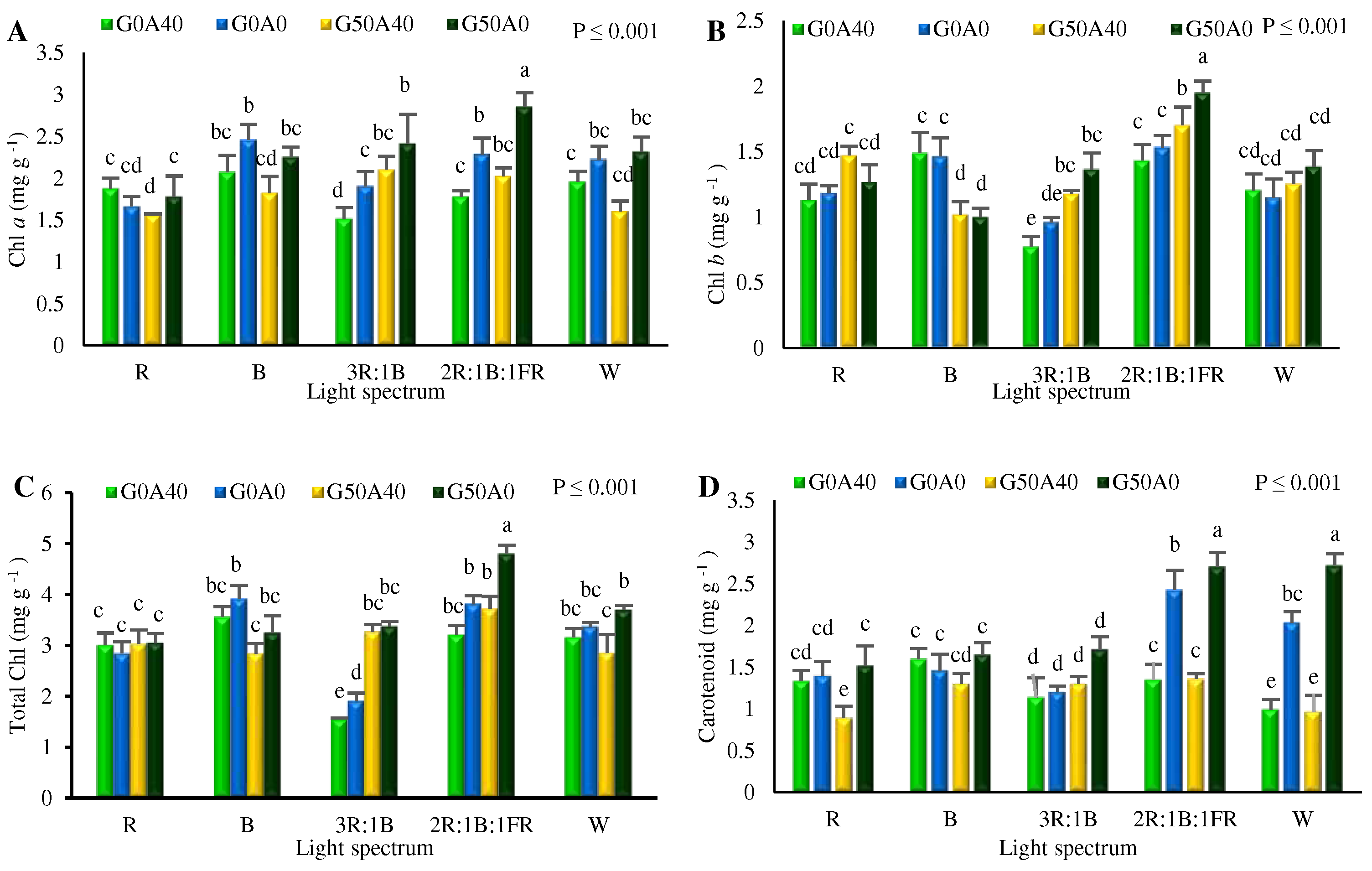
Figure 8.
Total anthocyanins content in Lollo Rosso lettuce leaves under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol L-1 γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
Figure 8.
Total anthocyanins content in Lollo Rosso lettuce leaves under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol L-1 γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
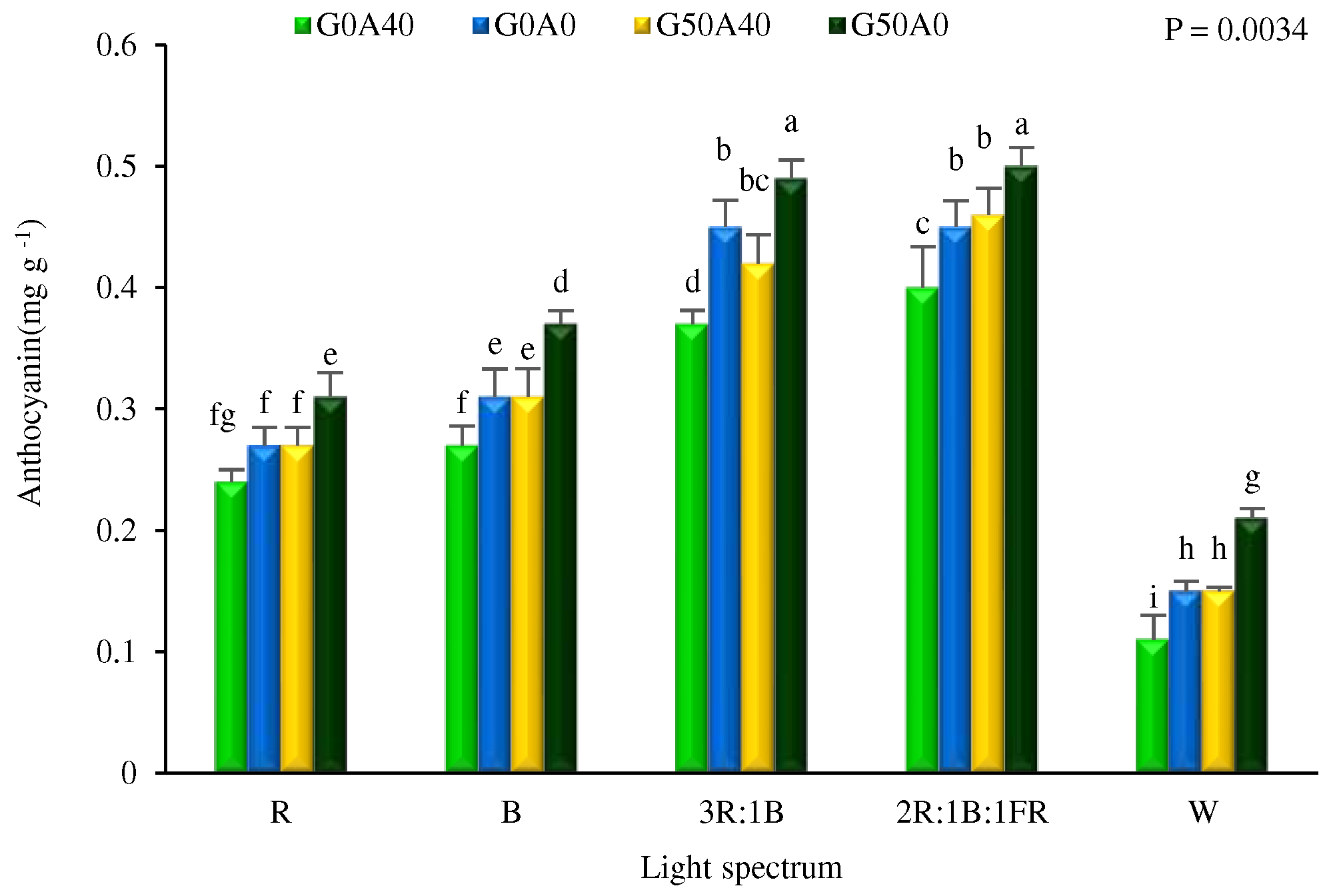
Figure 9.
Soluble carbohydrate content in Lollo Rosso lettuce leaves under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
Figure 9.
Soluble carbohydrate content in Lollo Rosso lettuce leaves under different light spectra including red (R, 600–700 nm), blue (B, 400–500 nm), white (W, 400-730 nm), combination of R and B light in 3 R:1B ration, and combination of R and B together with far red (FR, 700-750 nm) in 2R:1B:1FR ratio under fixed light intensity of 250 ± 10 µmol m-2 s-1. Plants under each light spectra were fertigated with Hoagland solution containing 0 and 50 µmol γ-aminobutyric acid (GABA) under 0 and 40 mM NaHCO3 as alkaline stress. Different letters indicate that values are significantly different at p < 0.01 according to Duncan’s multiple range tests. Bars represent the mean value of three replications ± standard deviation.
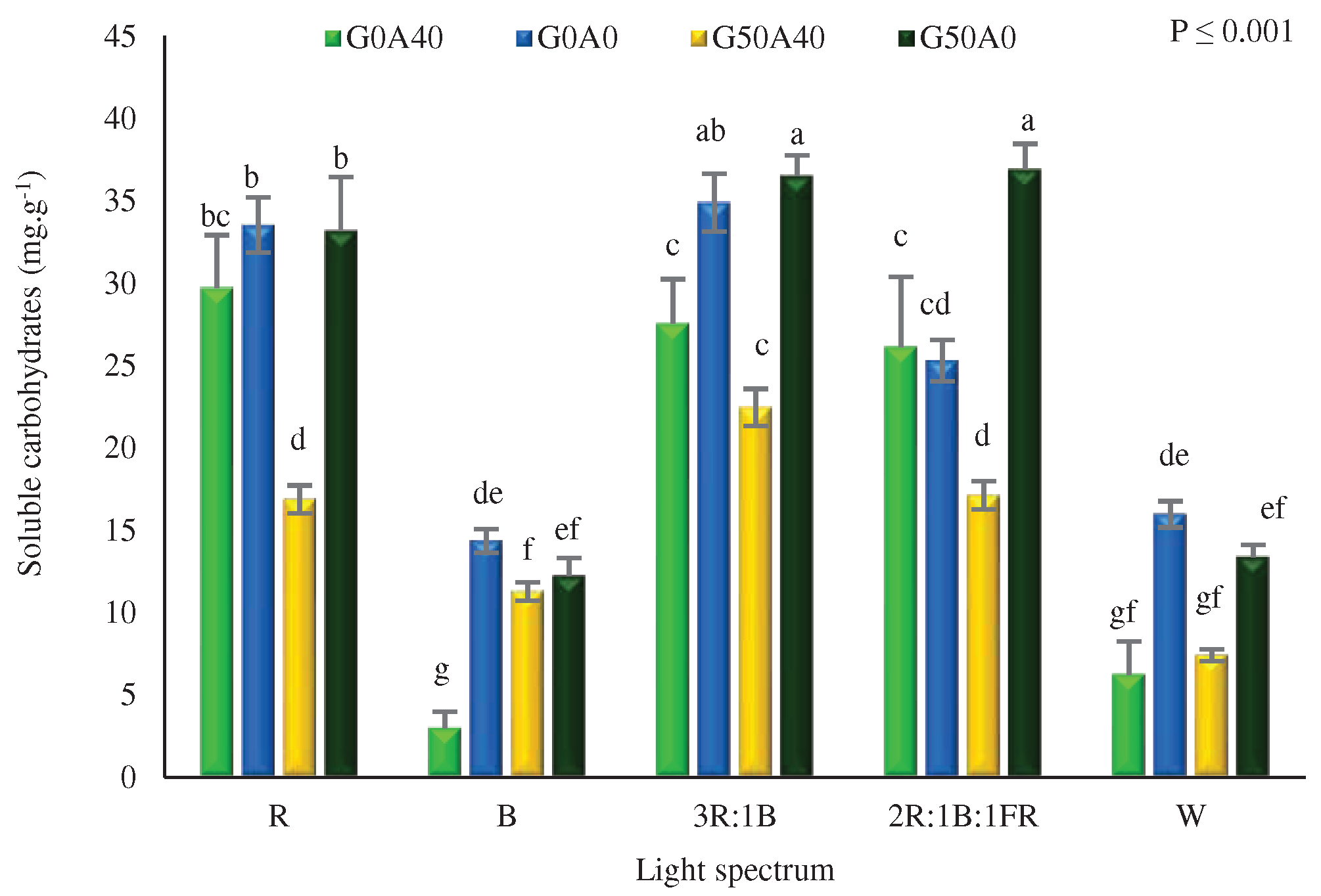

Table 1.
Energy Fluxes in Lollo Rosso lettuce under various light spectra and alkaline stress conditions.
Table 1.
Energy Fluxes in Lollo Rosso lettuce under various light spectra and alkaline stress conditions.
| Light spectrum | GABA (µmolL-1) | Alkalinity stress(mM) | ABS/RC | TR0/RC | ET0/RC | DI0/RC |
|---|---|---|---|---|---|---|
| R | 0 | 0 | 3.8f | 3.2f | 1.8e | 0.9f |
| R | 0 | 40 | 4.2e | 3.3e | 1.9d | 1e |
| R | 50 | 0 | 3.4g | 3.1g | 1.7f | 0.8g |
| R | 50 | 40 | 3.8f | 3.2f | 1.8e | 0.9f |
| B | 0 | 0 | 5.4b | 3.6b | 2.1b | 1.3b |
| B | 0 | 40 | 5.7a | 3.7a | 2.2a | 1.4a |
| B | 50 | 0 | 5c | 3.5c | 2c | 1.2c |
| B | 50 | 40 | 5.4b | 3.6b | 2.2a | 1.3b |
| 3R:1B | 0 | 0 | 2.6i | 3h | 1.6h | 0.6i |
| 3R:1B | 0 | 40 | 3.4g | 3.1g | 1.8g | 0.8g |
| 3R:1B | 50 | 0 | 2.2j | 2.9i | 1.4j | 0.5j |
| 3R:1B | 50 | 40 | 3h | 3h | 1.6h | 0.7h |
| 2R:1B:1FR | 0 | 0 | 2.6i | 2.8j | 1.5i | 0.6i |
| 2R:1B:1FR | 0 | 40 | 3h | 2.9i | 1.7f | 0.7h |
| 2R:1B:1FR | 50 | 0 | 2.2j | 2.7k | 1.4j | 0.5j |
| 2R:1B:1FR | 50 | 40 | 2.6i | 2.8j | 1.5i | 0.6i |
| W | 0 | 0 | 4.6d | 3.4d | 2c | 1.1d |
| W | 0 | 40 | 5c | 3.5c | 2.1b | 1.2c |
| W | 50 | 0 | 4.2e | 3.3e | 1.9d | 1e |
| W | 50 | 40 | 4.6d | 3.4d | 2c | 1.1d |
| P Value | - | - | 0.014 | 0.025 | 0.0148 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



