
Submitted:
02 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
Orchids are among the plants most threatened by anthropic impact and environmental changes. Therefore, all known orchid species are protected in several countries by regional, national and international legislation. Several studies cast doubts on the effectiveness of legislation to ensure protection of wild orchids. We assessed the vitality of four orchid populations in a coastal area in Northern Italy, by monitoring vegetative and reproductive traits of orchid populations growing both in protected sites comprised in the Natura 2000 network and in non-protected sites. We also monitored the level of environmental threat to orchid vitality. Early flowering deceptive species (Ophrys sphegodes and Anacamptis morio) exhibited high vegetative vitality and experienced similar levels of environmental threat in protected and non-protected areas. However, their reproductive success was strongly jeopardized probably by failed pollination that impeded fruit set. Late-flowering, partially or totally rewarding species (Anacamptis pyramidalis and Anacamptis coriophora) were more strongly impacted by spring mowing, ungulate herbivory and alien species. Only for A. coriophora were herbivory and alien species invasion lower at protected vs. non-protected sites which ensured higher population vitality at protected sites. We conclude that environmental protection is ineffective for preserving orchids without targeted actions against specific environmental threats.
Keywords:
environmental threats
; Natura 2000
; population vitality
; protected areas
; reproductive traits
; vegetative traits
1. Introduction
In the past few decades, the impact of human activities on natural ecosystems has led to a significant increase in extinction rate. According to the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) report, more than 500,000 species have insufficient habitat for long-term survival due to habitat loss and fragmentation [1]. Orchidaceae is one of the largest and most widely distributed families of Angiosperm plants, with more than 28,000 species and 763 genera [2]. During the evolutionary process, orchids have adapted to different environments and these species are absent only from desert and polar regions [3]. Although orchids occupy a wide range of habitats, several of them are extremely rare. This group of plants is the most threatened by anthropic impact and environmental changes. As a matter of fact, more than half (56.5%) of the only 948 orchid species estimated worldwide using Global Red List Criteria are considered threatened [4]. Therefore, all orchid species are protected in several countries by regional, national and international legislation such as the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) [5,6]. Habitat degradation, weed invasion, herbivory, illegal harvesting, pollinator decline, and climate change are the main threats to orchid survival [7]. All of these factors can negatively affect population dynamics and the long-term viability of orchids. The flowering and survival of many European orchid species is closely related to the resource status of plants which is in turn influenced by appropriate site management [8]. Moreover, these species are expected to be at a greater risk of extinction as they are dependent on interactions with mycorrhizal fungi and pollinators which are also being affected by habitat loss and climate change [9,10]. Orchids are particularly sensitive to environmental changes and are often the first to disappear in response to anthropogenic disturbances, making these species relevant bioindicators of the ecological quality of ecosystems [11,12]. Consequently, orchids pose unique challenges for in-situ conservation which is a focus target for many European countries where habitat fragmentation has led to a decline in habitat connectivity and reduced fitness of different species [13,14].
For the reasons explained above, establishing an extensive network of protected areas is one of the main tools to protect endangered species, habitats, and ecosystems and neutralize biodiversity loss [15]. Based on the Habitats Council Directive 92/43/EC, the European Union created the Natura 2000 Network, the largest coordinated multinational network of protected areas in the world which includes more than 18 % of the EU's land area and more than 8 % of its marine territory [16,17]. The role of protected areas in the prevention of species extinction is still uncertain in several cases. Studies have stated that there is a marked difference between the conservation expectations and the actual effectiveness of these areas [18,19]. This is mainly due to the absence of systematic management planning, that can furthermore vary under different socio-economic contexts [20]. In Italy and overall in Europe, several rare orchid species are hosted in dry grasslands, especially the semi-natural dry grasslands habitat listed in Annex I of the Habitats Directive (92/43/EEC) as 6210(*): ‘semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia)’ [21]. Unfortunately, the abandonment of traditional agricultural practices for high-intensity management and urbanization has caused significant ecological and structural changes in these habitats as the decrease in niche availability and the enhancement of the dominance of few species with impacts on weak competitors such as orchids and, in general, a decline in plant species richness [22]. As the Natura 2000 network guarantees nature protection 'taking into account economic, social and cultural requirements and regional and local characteristics’, many environments (e.g., grasslands or scrublands) fall within urban or man-made areas where are considered as typical green spaces. Consequently, a sustainable economic and ecologic management of these spaces is not always appropriately planned taking due account to the ecological requirements of the species and environments. In the Po Delta (Northern Italy) during the period of strong urbanization for seaside tourism in the 1960s, many natural areas and habitats (e.g. pine forests, wet and dry meadows and grasslands) were destroyed or incorporated as urban green spaces covered by natural vegetation. Unfortunately, these areas are managed with urban policies implying for example several mowing phases during the year because unmowed meadows with taller grass are commonly regarded as synonymous of mess, not cared or non-aesthetic [23,24]. During the years, these meadows were enriched with non-native plants from neighbouring gardens, but they still represent the natural habitat of wild plants including several orchid species. Hence, these meadows, whether they are part of the Natura 2000 network or not, often represent relict or fragmented habitats that must be protected with proper management for biodiversity conservation and environmental sustainability [25,26].
In this study, we monitored populations of four native orchid species both in protected and unprotected areas of semi-dry grasslands in the Po Delta which is considered one of the most important natural areas in Europe. However, this territory has been strongly modified by human activities related to urbanization and agricultural development [27] and because of this the Po Delta Regional Park, in Emilia-Romagna, encompasses several protected areas that aim to conserve biodiversity in a highly anthropic environment and prevent ecosystem degradation and species extinction [28]. Therefore, the objective of this study was to establish the vitality of these orchid populations by monitoring both the vegetative and reproductive traits. Moreover, through the recording of threats to orchid vitality we compare protected and unprotected areas to test the effectiveness of the actual protection based on the assumption that orchids of protected areas will have higher conservation status.
2. Results
2.1. Vegetative and reproductive traits
There were overall modest differences between protected and non-protected sites in terms of vegetative traits (Figure 2). O. sphegodes presented larger rosettes in protected sites (Figure 2I); A. morio had higher stem in protected sites as well (Figure 2B). Conversely, A. pyramidalis had higher SLA and higher LDMC in non-protected sites (Figure 2N,S). A. coriophora presented a contrasting pattern of vegetative traits in relation to protection level, with more leaves in protected sites but taller stems and higher SLA in non protected sites (Figure 2D,H,P).
There were strong differences among species with respect to reproductive traits. Indeed, all species flowered, with only A. morio presenting more flowers in non-protected sites (Table 1). However, both O. sphegodes and A. morio produced occasional fruits if any because the great majority of flowers were not fecundated and rotted without setting fruits during ripening in these two species. For this reason, number of fruits, seed mass and number of embryos could be recorded only for A. pyramidalis and A. coriophora (Table 1). These two species presented similar patterns of reproductive traits in relation to protection level, with equal performance in protected vs. non-protected sites for most traits. There were more fruits in protected sites for A. pyramidalis, and heavier seeds as well as more embryos in protected sites for A. coriophora (Table 1).
2.2. Vitality of orchid individuals and populations
The vitality of O. sphegodes and A. morio individuals were much higher in terms of vegetative traits than of reproductive traits with negligible differences related to protection level (Figure 3A,B). The A. pyramidalis individuals also presented higher vegetative vitality than reproductive vitality although with less strong difference between traits (Figure 3C). Conversely, A. coriophora individuals has higher reproductive vitality than vegetative vitality (Figure 3D). The reproductive vitality of A. pyramidalis individuals was higher in non-protected sites than in protected sites while the reverse was true for A. coriophora (Figure 3C,D).
Population vitality was higher in terms of vegetative traits compared with reproductive traits for O. sphegodes, A. morio, and although to a lower extent for A. pyramidalis. The reverse was true for A. coriophora (Table 2). For O. sphegodes and A. morio the Q values suggested prosperous population condition in term of vegetative traits, while for A. pyramidalis and A coriophora the Q values suggested moderately prosperous to equilibrium condition in terms of vegetative traits (Table 2). Both O. sphegodes and A. morio presented very low Q values in terms of reproductive traits indicating depressed population condition independent of protection level. A. pyramidalis had somewhat higher reproductive Q values in both protected and non-protected sites compared to O. sphegodes and A. morio, indicating equilbrium condition (Table 2). A. coriophora had much higher reproductive Q values in protected sites, indicating prosperous population condition, than in non-protected sites where the population condition was in equilibrium (Table 2).
2.3. Environmental threats
3. Discussion
In terms of vegetative traits, both O. sphegodes and A. morio presented high performance independent of protection level. The populations of both species were classified as prosperous either in protected and in non-protected areas. In contrast, the vegetative performance of A. pyramidalis and A. coriophora was somewhat lower. Consequently, the populations of A. pyramidalis and A. coriophora were classified as moderately prosperous in non-protected areas and even in equilibrium in protected areas. Such rather strong difference in vegetative performance between O. sphegodes and A. morio, on one side, and A. pyramidalis and A. coriophora, on the other side, was related to different phenology of the four species. Indeed, both O. sphegodes and A. morio have wintergreen rosettes and aboveground stems developing early in spring [29,30] while A. pyramidalis and A. coriophora both lack overwintering rosettes and their aboveground organs start developing more than a month later [31,32]. For this reason, the four species reacted differently to mowing. Although mowing is believed to be advantageous for orchid fitness, its occurrence does not automatically ensure vitality of orchid populations [33,34,35]. Mowing practice conducted outside the growing period have been found to improve vitality of orchid populations. Early mowing alleviates the competitive pressure from species that can outcompete orchids because of higher growth potential [36]. Late mowing removes the old plant biomass, reducing shading of orchids in the subsequent year and increasing the light available for photosynthesis [37,38]. In our study area, mowing is generally practiced intensively from late April to mid May in order to ameliorate green areas for public use. Because of their early phenology, O. sphegodes and A. morio were unharmed by mowing that took place when the growing season of both species had already ended. Conversely, A. pyramidalis and A. coriophora were more or less heavily damaged by mowing during their growing season. In particular, many individuals of A. pyramidalis were wiped out at the most strongly mowed sites. To our knowledge, there are no well defined prescriptions for regulating the mowing timing in the Po Delta Park. Hence, environmental protection in our study area was unsuccessful for preventing damage to orchid populations caused by inappropriate seasoning of mowing practice. Although there were overall few differences in vegetative traits between protected and non-protected areas for both A. pyramidalis and A. coriophora, the slightly lower vegetative performance in protected areas was prevalently associated with lower SLA. As SLA represents a powerful proxy of photosynthetic capacity in vascular plants [39,40], reduced SLA may constrain photosynthetic activity ultimately limiting plant growth. Smaller plant size may be detrimental to vitality because medium-size and big plants contribute more to the persistence of orchid populations [41]. High photosynthetic rates can also improve the reproductive success of orchid species [42,43]. We have no mechanistic explanation as to why the vegetative vitality of A. pyramidalis and A. coriophora was, albeit slightly, worse in the protected areas. However, it has to be considered that small-scale environmental variations associated, for example, with mosaic-like patterns of canopy height can influence the vegetative performance of orchids independent of protection status [44,45].
The reproductive success of O. sphegodes and A. morio was practically nil, in spite of their high vegetative performance. So, the populations of both species were classified as depressed in terms of reproductive traits. The lack of reproductive success in O. sphegodes and A. morio was totally unrelated to the threats detected in our surveys and protection level as well. Failed sexual reproduction in some orchids species has been linked to adverse weather conditions in the previous growing season. For example, Kirillova and Kirillov [46] observed less flowers and smaller seeds in Platanthera bifolia the year after a dry summer season. Although our study area did experience extremely hot dry weather during summer 2022, viz. the year preceding our sampling (https://www.arpae.it/it/temi-ambientali/clima/clima), it is very unlikely that this was the cause hampering sexual reproduction in O. sphegodes and A. morio in 2023. Indeed, summer drought causes no problems to species which produce leaves in the autumn and remain green as they have no above ground organs [47]. The observed failure of sexual reproduction in O. sphegodes and A. morio was determined by lack of fruit setting even if both species flowered vigorously. We did not observe any apparent sign of pest occurrence on the stems of these orchids. Therefore, we believe that failed pollination was the most likely cause of lacking fructification in O. sphegodes and A. morio, for example due to temporal displacement in the phenology of plant and pollinator [48]. Indeed, both O. sphegodes [29] and A. morio [49] require a very narrow specialized set of pollinators to be available for successful pollination. Strictly deceptive orchid species, like O. sphegodes and A. morio, usually flower early and present lower fructification rates with even < 15% of the flowers produce capsules [50,51,52]. Thus, their strategy seems to be based on a low-risk approach that ensures the production of a smaller number of fruits before the shading effect of the surrounding plant community and the potential impact of mowing [37]. However, if sexual reproduction fails for several consecutive years this can considerably reduce population size because vegetative multiplication occurs rather occasionally through survival or splitting of the old tuber [53].
Both A. pyramidalis and A. coriophora set fruit containing viable seeds that produced embryos, even if the reproductive vitality was overall higher in A. coriophora. This finding was consistent with the pollination strategy of the two species, with A. pyramidalis possessing a generalized deceptive behaviour because their flowers contain nectar in the spur [54] while A. coriophora acts as a fully rewarding species [32]. All of the recorded types of threat recorded in our survey besides mowing, viz. waste dumping, herbivory and IAS, could to some extent lower the reproductive vitality of A. pyramidalis and A. coriophora although the populations of either species never attained depressed condition in terms of reproductive traits. In addition to direct and indirect effects of waste dumping on environmental pollution [55], waste dump is susceptible to bring about habitat loss and fragmentation, both representing major threats to the viability of orchid populations [56]. Our data do not allow us to evaluate whether and to what extent waste dumping impacted the vitality of the orchid populations. However, as the objective of our study was to assess the effectiveness of environmental protection in ensuring the vitality of orchid populations, we can state that the threat associated with waste dumping was unrelated to protection level for any of the four species investigated. In our study area herbivory mainly consists in browsing by fallow deer (Dama dama). While grazing practices by sheep and cattle have been shown to contribute to the preservation of orchids in semi-dry grasslands, not only by appropriate biomass removal but also by selective defoliation and creation of gaps suitable for orchid seed germination [57,58], browsing by deer has detrimental effects on several plant species [59]. For example, browsing by the North-American white-tailed deer (Odocoileus virginianus) negatively impacts the vegetative growth of the rare orchid Platanthera integrilabia [60] which in turn hampers the reproductive success of the species [42,43]. Browsing by white-tailed deer has also been found to hamper population viability of the herb Panax quinquefolius, through removal of foliage, flowers and fruits [61]. In general, herbivory has negative effects on floral traits, plant attractiveness to pollinators and eventually on the plant reproductive success. The damage can either derive by direct effects of browsing reproductive organs or by indirect effects on reproduction determined by defoliation which can lead to decreased seed production, reduced leaf growth and flowering in subsequent years [62,63]. Moreover, intensive herbivory by ungulates can enhance invasion of alien species because many invasive plants are adapted to habitat disturbance so that their survival is favoured as ungulates eliminate native plants which are more palatable than invasive species [64]. Besides possible interactions with herbivory [65], IAS alters ecological interactions since invasive species are superior competitors for light and space compared with native species [66]. Wild orchids often tolerate IAS, but their survival is negatively impacted once the cover of alien species exceeds 20% [67]. Invasive alien species also hamper the reproductive success of native species by reducing population fitness through reduced seed set, germination rate and seedling establishment [68]. So, besides out-competing native orchids thanks to their superior competitive ability, IAS can exert a negative impact on orchid reproduction for example by producing novel chemical compounds [69] or by reducing the diversity of mycorrhizal fungi [70,71]. Environmental protection was effective in lowering threat brought about by herbivory and IAS in sites hosting A. coriophora populations but not in the sites hosting A. pyramidalis populations. This was because the A. coriophora sites were located in areas less frequented by fallow deer. In these areas environmental protection was furthermore able to act as a filter against IAS [72,73]. Population vitality of A. pyramidalis unexpectedly was lower in protected than in non-protected areas. Such seemingly paradoxical finding may be due to uninvestigated micro-environmental factors that negatively affected reproduction of A. pyramidalis at the protected sites [74,75].
We conclude that environmental protection per sé is unlikely to effectively preserve endangered plant species [76], in particular orchids [77]. Targeted actions against specific environmental threats, targeted actions translated into explicit prescriptions in the regulations of protected areas, must be definited to achieve effective protection of orchids.
4. Materials and methods
4.1. Study area
The study area is located in the north-easternmost part of the region Emilia-Romagna (Northern Italy), along the North Adriatic coast (about 44°32’-44°56’N; 12°08’-12°16’E; Figure 1). Most of this area is within the territory of the Po Delta Regional Park Emilia-Romagna. However, the park does not cover a continuous area but is divided into six stations separated by non-protected areas. The Regional Park has a total surface area of 54,000 ha and hosts sixteen sites within the Natura 2000 network, either sites of community importance or special areas of conservation (https://www.parks.it/parco.delta.po.er/Epar.php). In addition, the northern part of this area hosts the National Nature Reserve Bosco della Mesola. According to the Köppen classification of climates, the climate in the study area is temperate-continental with mean annual temperature of about14 °C, mean temperature of the coldest month (January) of about 3 °C and mean annual temperature of the warmest month (July) of about 24 °C. Mean total annual precipitation is 600-700 mm, with a short weak period of summer aridity [78].
Several populations of terrestrial orchids are native to the study area [79]. Most of them are settled in semi-dry grasslands rich in annual species, such as Silene conica, Cerastium semidecandrum, Vulpia membranacea and Medicago minima. These grasslands generally lie on sandy flat areas corresponding to ancient dunes levelled by atmospheric agents. Syntaxonomically, these grasslands are most similar to communities of the Sileno conicae – Cerastietum semidecandri and can therefore be included in the priority habitat 2130 (Fixed coastal dunes with herbaceous vegetation, ‘grey dunes’). However, the presence of some representative perennial species such as Sanguisorba minor, Scabiosa columbaria, Fumana procumbens and Helianthemum apenninum, besides the richness of orchid species outlines some resemblance to the habitat 6210 (http://vnr.unipg.it/habitat/collaboratori.jsp).
4.2. Sampling and laboratory analyses
We selected for sampling four orchid species occurring in at least three protected sites and three non-protected sites within the study area. They were: Ophrys sphegodes Mill.; Anacamptis morio (L.) R.M. Bateman, Pridgeon et M.W. Chase; Anacamptis pyramidalis (L.) Rich.; Anacamptis coriophora (L.) R.M. Bateman, Pridgeon et M.W. Chase. There were 15 sampling sites in total, some of them hosting more than one species. Seven of these sites were in protected areas and eight in non-protected areas (Figure 1). We surveyed the sites several times during the growing season in order to capture, for each of the four species, the appropriate phenological phases for sampling. For each species, at the peak of the flowering period we randomly chose 30 sound individuals in three protected sites and three non-protected sites. Only for A. coriophora could we locate an additional site in a non-protected area. In a minority of cases the site hosted small orchid populations with fewer than thirty individuals. In those cases all of the individuals occurring at the site were sampled. The individuals selected were tagged by means of a numbered wooden stick inserted in the ground about 1.5 cm apart of the stem. The height of the stem and the diameter of the basal rosette were measured in the field with a ruler and the number of flowers was counted on each individual. We also counted the number of leaves on each selected individual. One healthy rosette leaf for each individual was harvested and carried to the laboratory for subsequent analyses. For each species, at the peak of the fruit ripening we counted the number of fruits present on each of the tagged individuals. Afterwards, all fruits of a given individual were harvested, sealed in a plastic bag wrapped with wet paper and carried to the laboratory for subsequent analyses.
In the laboratory, the harvested leaves were rehydrated overnight. The day after, they were weighed and scanned (CanoScan LiDE 120, Canon) in order to determine fresh weight and leaf area, respectively. The same leaves were weighed again after being dried at 105 °C for 72 h in order to determine dry weight. All harvested fruits were carefully empted of the seeds. The latter were pooled on a paper sheed and weighed. A subsample of 100 seeds was picked from the pool and examined under a stereo microscope for counting the embryos.
4.3. Data compilation and statistical analyses
The data obtained by field measurements and subsequent laboratory analyses were arranged in two groups:
- Vegetative traits: stem height (measured in the field); number of leaves (counted in the field); rosette diameter (measured in the field); specific leaf area (SLA, the ratio between leaf area and leaf fresh mass as obtained by laboratory scanning and weighing); leaf dry matter content (LDMC, the ratio between dry mass and fresh mass as obtained by laboratory weighings).
- Reproductive traits: number of flowers (counted in the field); number of fruits (counted in the field); seed mass (as obtained by laboratory weighing); number of embryos (counted in the laboratory under the stereo microscope).
The four species differed greatly from each other in terms of all vegetative and reproductive traits because of intrinsic differences depending on evolutionary legacy. Hence, it would be meaningless to assess interspecific differences based on rough data. We therefore run separate one-way ANOVAs with protection level of the sampling sites (protected vs. non-protected) as factorial factor, for all traits in the four species.
For comparing the vitality of orchid individuals in terms of both vegetative and reproductive traits, we calculated a normalized index (Qindividual) based on fractions of maximum values [80]. Normalization was performed as in formula (1)
Qindividual = Xi / Xmax
Where Xi is the rough value of a given trait for an individual of a given orchid species in a given site and Xmax is the maximum rough value across the whole set of the above values. The Qindividual values were then pooled separately for vegetative traits and reproductive traits. To this aim, we calculated the mean values of Qindividual for the five vegetative traits and the four reproductive traits, respectively, for all individuals of a given orchid species in a given site. These mean values were statistically analysed by two-way ANOVAs with trait (vegetative vs. reproductive), protection level (protected vs. non-protected) and their interaction as factorial factors.
The vitality of the orchid populations in protected vs. non-protected sites was assessed by splitting the mean Qindividual values of vegetative and reproductive traits into three categories [27]. Vitality class a: mean Qindividual > 0.66; vitality class b: 0.33 > mean Qindividual < 0.66; vitality class c: mean Qindividual < 0.33. The overall vitality of orchid populations was eventually estimated by calculating the fraction of individuals within classes a and b, according to formula (2):
Q = (a + b) / 2
The Q values, in the range from 0 to 0.5, were classified as follows. Prosperous: Q > 0.333; equilibrium: 0.167 > Q < 0.333; depressed: Q < 0.167.
Whenever we visited the sites, we assessed visually the level of four types of potential or real risks to the vitality of the orchis populations, viz. mowing, waste dumping (waste), browsing by herbivores (herbivory) and invasion of alien species (IAS). At the end of the season we summarized the levels of the four risk types at each site in four categories: 1: absent; 2: weak; 3: moderate; 4: high.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org. Supplementary File S1: Supplementary file S1 - ANOVAs of the risk categories.
Author Contributions
Conceptualization, Lisa Scramoncin and Lisa Brancaleoni; Investigation, Lisa Scramoncin, Renato Gerdol and Lisa Brancaleoni; Writing – original draft, Lisa Scramoncin and Renato Gerdol; Writing – review & editing, Renato Gerdol. All authors have read and agreed to the published version of the manuscript.
Funding
This publication was co-funded by the European Social Fund (ESF)—National Operational Programme (NOP) on Research and Innovation 2014–2020. 2021-PON-DM-1062-MW-RIC.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
We thank Dr. Massimiliano Costa, Director of the Po Delta Regional Park Emilia-Romagna, for authorizing sampling in the protected areas.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPBES. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. 2019. Available online: https://www.ipbes.net/global-assessment (accessed on 27 December 2023).
- Christenhusz, M.J.M.; Byng, J.W. The number of known plant species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [CrossRef]
- Fay, M.F.; Chase, M.W. Orchid biology: from Linnaeus via Darwin to the 21st century. Ann. Bot. 2009, 104, 359–364. [CrossRef]
- Khapugin, A. A global systematic review of publications concerning the invasion biology of four tree species. Hacquetia 2019, 18, 233–270. [CrossRef]
- Ticktin, T.; Charitonidou, M.; Douglas, J.; Halley, J.M.; Hernández-Apolinar, M.; Liu, H.; Mondragón, D.; Pérez-García, E.A.; Tremblay, R.L.; Phelps, J. Wild orchids: A framework for identifying and improving sustainable harvest. Biol. Conserv. 2023, 277, 109816. [CrossRef]
- Fay, M.F. Orchid conservation: how can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [CrossRef]
- Reiter, N.; Whitfield, J.; Pollard, G.; Bedggood, W.; Argall, M.; Dixon, K.; Davis, B.; Swarts, N. Orchid re-introductions: an evaluation of success and ecological considerations using key comparative studies from Australia. Plant Ecol. 2016, 217, 81–95. [CrossRef]
- Kull, T. Population Dynamics of North Temperate Orchids. In Orchid biology VIII: Reviews and Perspectives, 1st ed.; Kull, T.; Arditti, J., Eds.; Kluwer Academic Publishers: Dordrecht, 2002; pp. 139–165, ISBN 978-1-4020-0580-0.
- Štípková, Z.; Kindlmann, P. Factors determining the distribution of orchids – a review with examples from the Czech Republic. Eur. J. Environ. Sci. 2021, 11, 21–30. [CrossRef]
- Wraith, J.; Pickering, C. Quantifying anthropogenic threats to orchids using the IUCN Red List. Ambio 2018, 47, 307–317. [CrossRef]
- Bianco, P.M. Le orchidee come indicatori di qualità degli habitat. Biol. Ital. 2012, 42, 35–48.
- Gale, S.W.; Fischer, G.A.; Cribb, P.J.; Fay, M.F. Orchid conservation: bridging the gap between science and practice. Bot. J. Linn. Soc. 2018, 186, 425–434. [CrossRef]
- Newman, B.J.; Ladd, P.; Brundrett, M.; Dixon, K.W. Effects of habitat fragmentation on plant reproductive success and population viability at the landscape and habitat scale. Biol. Conserv. 2013, 159, 16–23. [CrossRef]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N.D. Effectiveness of terrestrial protected areas in reducing habitat loss and population declines. Biol. Conserv. 2013, 161, 230–238. [CrossRef]
- Abeli, T.; D’Agostino, M.; Orsenigo, S. et al. IDPlanT: the Italian database of plant translocation. Plant Biosyst. 2021. [CrossRef]
- Orlikowska, E.H.; Roberge, J.M.; Blicharska, M.; Mikusiński, G. Gaps in ecological research on the world's largest internationally coordinated network of protected areas: A review of Natura 2000. Biol. Conserv. 2016, 200, 216–227. [CrossRef]
- European Commission Natura 2000. 2020. Available online: https://ec.europa.eu/environment/nature/natura2000/index_en.htm (accessed on 27 December 2023).
- Hill, R.; Miller, C.; Newell, B.; Dunlop, M.; Iain, J.G. Why biodiversity declines as protected areas increase: the effect of the power of governance regimes on sustainable landscapes. Sustain. Sci. 2015, 10, 357–369. [CrossRef]
- Gray, C.; Hill, S.; Newbold, T.; Hudson, L.N; Borger, L.; Contu, S.; Hpskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [CrossRef]
- Pulido-Chadid, K.; Virtanen, E.; Geldmann, J. How effective are protected areas for reducing threats to biodiversity? A systematic review protocol. Environ. Evid. 2023, 12, 18. [CrossRef]
- Olmeda, C.; Šefferová, V.; Underwood, E.; Millan, L.; Gil, T.; Naumann, S. (compilers). EU Action plan to maintain and restore to favourable conservation status the habitat type 6210 Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (*important orchid sites). 2019. Available online: https://www.ecologic.eu/17536 (accessed on 27 December 2023).
- Slaviero, A.; Del Vecchio, S.; Pierce, S.; Fantinato, E.; Buffa, G. Plant community attributes affect dry grassland orchid establishment. Plant Ecol. 2016, 217, 1533–1543. [CrossRef]
- Ignatieva, M.; Hedblom, M. An alternative urban green carpet. How can we move to sustainable lawns in a time of climate change? Science 2018, 362, 148–149. [CrossRef]
- Ignatieva, M.; Haase, D.; Dushkova, D.; Haase, A. Lawns in cities: from a globalised urban green space phenomenon to sustainable nature-based solutions. Land 2020, 9, 73. [CrossRef]
- Sehrt M.; Bossdorf O.; Freitag M.; Bucharova A. Less is more! Rapid increase in plant species richness after reduced mowing in urban grasslands. Basic Appl. Ecol. 2020, 42, 47–53. [CrossRef]
- Paudel, S.; States, S.L. Urban green spaces and sustainability: Exploring the ecosystem services and disservices of grassy lawns versus floral meadows. Urban For. Urban Green. 2023, 84, 127932. [CrossRef]
- Simeoni, U.; Corbau, C. A review of the Delta Po evolution (Italy) related to climatic changes ad human impacts. Geomorphology 2009, 107, 64–71. [CrossRef]
- Committee on the Environment, Agriculture and Local and Regional Affairs, Protection of European deltas Report. 2005. Available online: https://assembly.coe.int/nw/xml/XRef/X2H-Xref-ViewHTML.asp?FileID=10868&lang=EN (accessed on 27 December 2023).
- Jacquemyn, H.; Hutchings, M.J. Biological flora of the British Isles: Ophrys sphegodes. J. Ecol. 2015, 103, 1680-1696. [CrossRef]
- Teibert, C.F. Reproductive success of Anacamptis morio (Orchidaceae) in der Donau-Auen National Park, Austria. MSc Thesis, 2018. University of Natural Resources and Life Sciences, Vienna, Austria.
- Robbirt, K.M. Phenological responses of British orchids and their pollinators to climate change: an assessment using herbarium and museum collections. MSc Thesis, 2012. University of East Anglia, Norwich, UK.
- Lanzino, M.; Palermo, A.M.; Pellegrino, G. The effect of inflorescence display size and flower position on pollination success in two deceptive and one rewarding orchid. Plant Biol. 2023, 25, 396–402. [CrossRef]
- Willems, J.H. Population dynamics of Spiranthes spiralis in South Limburg, The Netherlands. Mém. Soc. Royale Bot. Belgium 1998, 11, 115–121.
- Moen, A.; Oien, D.I. Ecology and survival of Nigritella nigra, a threatened orchid species in Scandinavia. Nord. J. Bot. 2002, 22, 435–461. [CrossRef]
- Damgaard, C.; Moeslund, J.E.; Wind, P. Changes in the abundance of Danish orchids over the past 30 years. Diversity 2020, 12, 244. [CrossRef]
- Djordjević, V.; Aćić, S.; Kabaš, E.; Lazarević, P.; Tsiftsis, S.; Lakušić, D. The orchids of wetland vegetation in the Central Balkans. Diversity 2023, 15, 26. [CrossRef]
- Pierce, S.; Vagge, I.; Brusa, G.; Cerabolini, B.E.L. The intimacy between sexual traits and Grime’s CSR strategies for orchids coexisting in semi-natural calcareous grassland at the Olive Lawn. Plant Ecol. 2014, 215, 495–505. [CrossRef]
- Meekers, T.; Hutchings, M.J.; Honnay, O.; Jacquemyn, H. Biological flora of the British Isles: Gymnadenia conopsea s.l. J. Ecol. 2012, 100, 1269–1288. [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [CrossRef]
- Tonin, R.; Gerdol, R.; Tomaselli, M.; Petraglia, A.; Carbognani, M.; Wellstein, C. Intraspecific functional trait response to advanced snowmelt suggests increase of growth potential but decrease of seed production in snowbed plant species. Front. Plant Sci. 2019, 10, 289. [CrossRef]
- Ackerman, J.D.; Tremblay, R.L.; Pérez, M.E.; Madden, H.; Bechtold, M.; Boeken, M. Small populations on small islands: What chance does an orchid have? Int. J. Plant Sci. 2020, 181, 667–685. [CrossRef]
- Willems, J.H.; Melser, C. Population dynamics and life-history of Coeloglossum viride (L.) Hartm.: an endangered orchid species in The Netherlands. Bot. J. Linn. Soc. 1998, 126, 83–93. [CrossRef]
- Zhang, S.; Hu, H.; Zhou, Z.; Xu, K.; Yan, N.; Li, S. Photosynthesis in relation to reproductive success of Cypripedium flavum. Ann. Bot. – London 2005, 96, 43–49. [CrossRef]
- Landi, M.; Frignani, F.; Lazzeri, C.; Angiolini, C. Abundance of orchids on calcareous grasslands in relation to community species, environmental, and vegetational conditions. Russ. J Ecol 2009, 40, 486–494. [CrossRef]
- Catorci, A.; Cesaretti, S.; Gatti, R. Effect of long-term abandonment and spring grazing on floristic and functional composition of dry grasslands in a central Apennine farmland. Pol. J. Ecol. 2013, 61, 505–518.
- Kirillova, I.A.; Kirillov, D.V. Impact of weather conditions on the population dynamics and reproductive success of Platanthera bifolia (L.) Rich. in the Komi Republic. Contemp. Probl. Ecol. 2023, 16, 819–830. [CrossRef]
- Wells, T.C.E.; Rothery, P.; Cox, R.; Bamford, S. Flowering dynamics of Orchis morio L. and Herminium monorchis (L.) R.Br. at two sites in eastern England. Bot. J. Linn. Soc. 1998, 126, 39–48. [CrossRef]
- Rafferty, N.E.; Ives, A.R. Effects of experimental shifts in flowering phenology on plant-pollinator interactions. Ecol. Lett. 2011, 14, 69–74. [CrossRef]
- Jersáková, J.; Kindlmann, P. Patterns of pollinator-generated fruit set in Orchis morio (Orchidaceae). Folia Geobot. 1998, 33, 377–390. [CrossRef]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [CrossRef]
- Pellissier, L.; Vittoz, P.; Internicola, A.I; Gigord, L.D.B. Generalized food-deceptive orchid species flower earlier and occur at lower altitudes than rewarding ones. J. Plant Ecol. 2010, 3, 243–250. [CrossRef]
- Hornemann, G.; Michalski, S.G.; Durka, W. Short-term fitness and long-term population trends in the orchid Anacamptis morio. Plant Ecol. 2012, 213, 1583–1595. [CrossRef]
- Arditti, J. Fundamentals of Orchid Biology. John Wiley & Sons Ltd.: New York, USA, 1992; 691 p.
- Nazarov, V.V., Efetov, K.A. On the role of Zygaenidae (Lepidoptera) in pollination of Anacamptis pyramidalis (Orchidaceae). Zool. Zhurn. 1993, 72, 54–67.
- Taylor, R.; Allen, A. In Protecting Groundwater for Health: Managing the Quality of Drinking-water Sources; Schmoll, O.; Howard, G.; Chilton, J.; Chorus, I., Eds.; IWA: London, UK, 2006; p. 697.
- Gijbels, P.; De Hert, K.; Jacquemyn, H.; Honnay, O. Reduced fecundity and genetic diversity in small populations of rewarding versus deceptive orchid species: a meta-analysis. Plant Ecol. Evol. 2015, 148, 153–159. [CrossRef]
- Barbaro, L.; Dutoit, T.; Grossi, J.L. Influence des facteurs agro-écologiques sur les assemblages d'orchidées dans les pelouses calcicoles du Vercors (Préalpes, France). Bot. Helv. 2003, 113, 63–79.
- Metera, E.; Sakowski, T.; Sloniewski, K.; Romanowicz, B. Grazing as a tool to maintain biodiversity of grassland - a review. Anim. Sci. Pap. Rep. 2010, 28, 315–334.
- Maron, J.L.; Crone, E. Herbivory: effects on plant abundance, distribution and population growth. Proc. R. Soc. B 2006, 273, 2575–2584. [CrossRef]
- Wooten, S.; Call, G.; Dattilo, A.; Cruse-Sanders, J.; Boyd, J.N. Impacts of forest thinning and white-tailed deer herbivory on translocation of the rare terrestrial orchid Platanthera integrilabia. Diversity 2020, 12, 412. [CrossRef]
- McGraw, J.B.; Furedi, M.A. Deer browsing and population viability of a forest understory plant. Science 2005, 307, 920–922. [CrossRef]
- Vallius, E. Factors affecting fruit and seed production in Dactylorhiza maculata (Orchidaceae), Bot. J. Linn. Soc. 2001, 135, 89–95. [CrossRef]
- Moreira, X.; Castagneyrol, B.; Abdala-Roberts, L.; Traveset, A. A meta-analysis of herbivore effects on plant attractiveness to pollinators. Ecology 2019, 100, e02707. [CrossRef]
- Vavra, M.; Parks, C.J.; Wisdom, M. J. Biodiversity, exotic plant species, and herbivory: The good, the bad, and the ungulate, Forest Ecol. Manag. 2007, 246, 66–72. [CrossRef]
- Whigham, D.E. Ecology of woodland herbs in temperate deciduous forests. Annu. Rev. Ecol. Evol. S. 2004, 35, 583–621. [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species' traits and environment. Glob. Change Biol. 2012, 18, 1725–1737. [CrossRef]
- Martín-Forés, I.; Bywaters, S.L.; Sparrow, B.; Guerin, G.R. Simultaneous effect of habitat remnancy, exotic species, and anthropogenic disturbance on orchid diversity in South Australia. Conserv. Sci. Pract. 2022, 4, e12652. [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [CrossRef]
- Kottawa-Arachchi, J.D.; Gunasekara, R.S. Research priorities and future directions in conservation of wild orchids in Sri Lanka: A review. Nat. Conserv. Res. 2020, 5, 34–45. [CrossRef]
- Bonnardeaux, Y.; Brundrett, M.; Batty, A.; Dixon, K.; Koch, J.; Sivasithamparam, K. Diversity of mycorrhizal fungi of terrestrial orchids: compatibility webs, brief encounters, lasting relationships and alien invasions. Mycol. Res. 2007, 111, 51–61. [CrossRef]
- Li, T.; Wu, S.; Yang, W.; Selosse, M.A.; Gao, J. How mycorrhizal associations influence orchid distribution and population dynamics Front. Plant Sci. 2021, 12, 647114. [CrossRef]
- Foxcroft, L.C.; Jarošík, V.; Pyšek, P.; Richardson, D.M.; Mathieu, R. Protected-area boundaries as filters of plant invasions. Conserv. Biol. 2010, 25, 400–405. [CrossRef]
- Le, H.; Zhao, C.; Xu, W.; Deng, Y.; Xie, Z. Anthropogenic activities explained the difference in exotic plants invasion between protected and non-protected areas at a northern subtropics biodiversity hotspot. J. Environ. Manage. 2023, 345, 118939. [CrossRef]
- Jones, P. Aspects of the population biology of Liparis loeselii (L.) Rich. var. ovata Ridd. ex Godfery (Orchidaceae) in the dune slacks of South Wales, UK. Bot. J. Linn. Soc. 1998, 126, 123–139. [CrossRef]
- Huang, B.Q.; Yang, X.Q.; Yu, F.H.; Luo, Y.B.; Tai, Y.D. Surprisingly high orchid diversity in travertine and forest areas in the Huanglong valley, China, and implications for conservation. Biodivers. Conserv. 2008, 17, 2773–2786. [CrossRef]
- Heywood, W.H. Conserving plants within and beyond protected areas - still problematic and future uncertain. Plant Divers. 2019, 41, 36–49. [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction Ann. Bot. – London 2009, 104, 543–556. [CrossRef]
- Alessandrini, A.; Balboni, G.; Brancaleoni, L.; Gerdol, R.; Nobili, G.; Pellizzari, M.; Piccoli, F.; Ravaglioli, M. The Vascular Flora of the Bosco della Mesola Nature Reserve (Northern Italy). Springer Nature Switzerland AG: Cham, Switzerland, 2021; Geobotany Studies; 107 p. [CrossRef]
- Piccoli, F.; Pellizzari, M.; Alessandrini, A. Flora del Ferrarese. Istituto per i Beni Artistici Culturali e Naturali della Regione Emilia-Romagna, Longo Ed.: Ravenna, Italy, 2014; 321 p.
- Zlobin, Y.; Kovalenko, I.; Klymenko, H.; Kyrylchuk, K.; Bondarieva, L.; Tykhonova, O.; Zubtsova, I. Vitality analysis algorithm in the study of plant individuals and populations. Open Agric. 2021, 15, 119–129. [CrossRef]
Figure 2.
Mean (+ 1 SE) values of vegetative traits (A-D: stem height; E-H: number of leaves; I-K: rosette diameter; L-P: specific leaf area, SLA; Q-T: leaf dry matter content, LDMC) in the four orchid species at protected (P) and non-protected (NP) sites. The asterisks indicate significant (p < 0.05) differences between P and NP obtained by one-way ANOVAs. n.a. = not available.
Figure 2.
Mean (+ 1 SE) values of vegetative traits (A-D: stem height; E-H: number of leaves; I-K: rosette diameter; L-P: specific leaf area, SLA; Q-T: leaf dry matter content, LDMC) in the four orchid species at protected (P) and non-protected (NP) sites. The asterisks indicate significant (p < 0.05) differences between P and NP obtained by one-way ANOVAs. n.a. = not available.
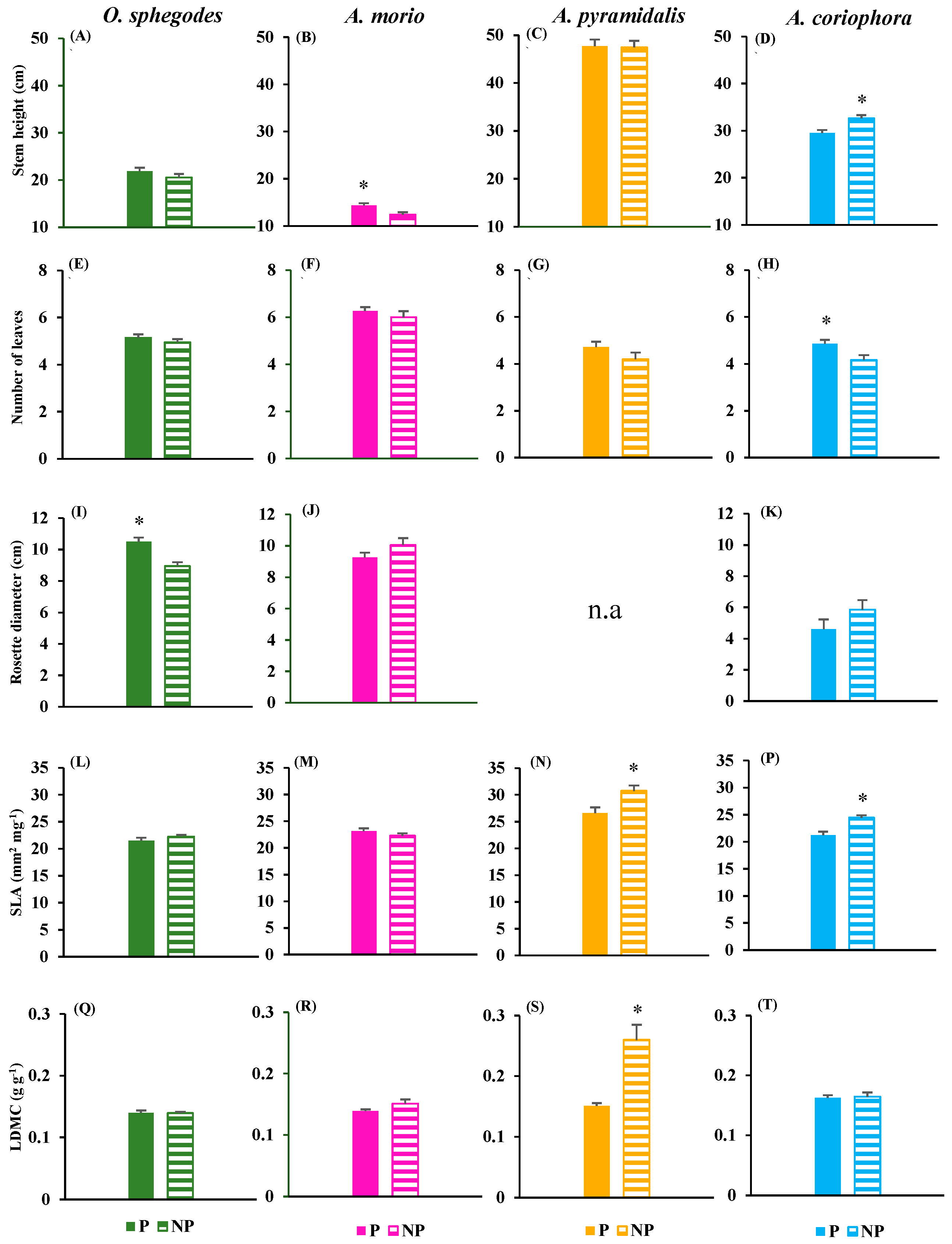
Figure 3.
Mean (+ 1 SE) values of Qindividual for vegetative and reproductive traits in the four orchid species (A-D) at protected (P) and non-protected (NP) sites. Significant (p < 0.05) effects of trait (T), protection level (P) and their interaction, obtained by two-way ANOVAs, are indicated by letters within each panel. The asterisks indicate significant (p < 0.05) differences between protected and non-protected sites for any combination of trait and protection level.
Figure 3.
Mean (+ 1 SE) values of Qindividual for vegetative and reproductive traits in the four orchid species (A-D) at protected (P) and non-protected (NP) sites. Significant (p < 0.05) effects of trait (T), protection level (P) and their interaction, obtained by two-way ANOVAs, are indicated by letters within each panel. The asterisks indicate significant (p < 0.05) differences between protected and non-protected sites for any combination of trait and protection level.
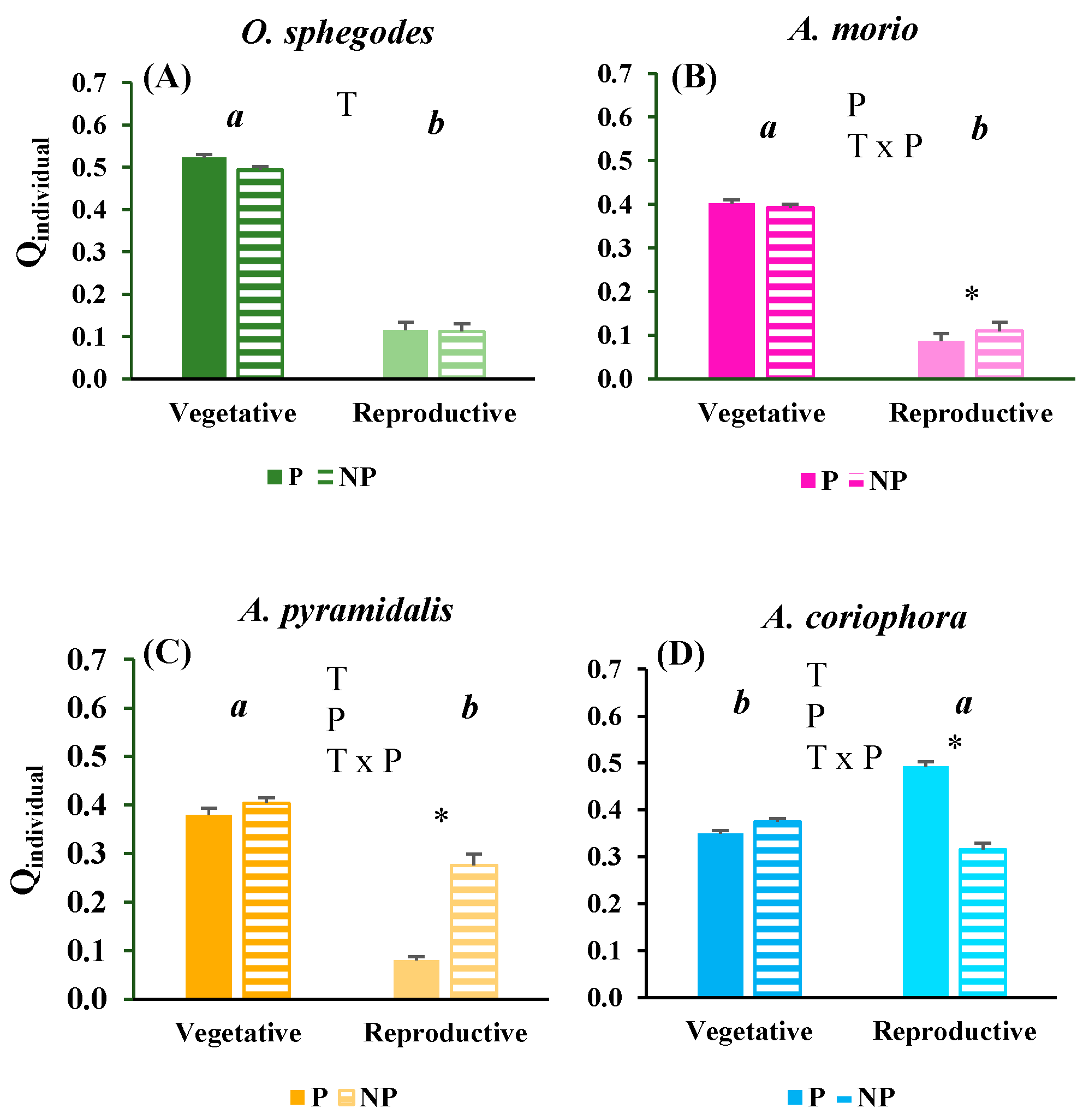
Figure 4.
Mean values of four types of environmental risks (mowing, waste dumping, waste; browsing by herbivores, herbivory; invasion of alien species, IAS) to the four orchid species (A-D) at protected (P) and non-protected (NP) sites. The asterisks indicate significant (p < 0.05) differences between protected and non-protected sites obtained by one-way ANOVAs. For clarity of presentation the values are percentages of the maximum value (for details of the raw data and ANOVAs results see Supplementary File S1).
Figure 4.
Mean values of four types of environmental risks (mowing, waste dumping, waste; browsing by herbivores, herbivory; invasion of alien species, IAS) to the four orchid species (A-D) at protected (P) and non-protected (NP) sites. The asterisks indicate significant (p < 0.05) differences between protected and non-protected sites obtained by one-way ANOVAs. For clarity of presentation the values are percentages of the maximum value (for details of the raw data and ANOVAs results see Supplementary File S1).
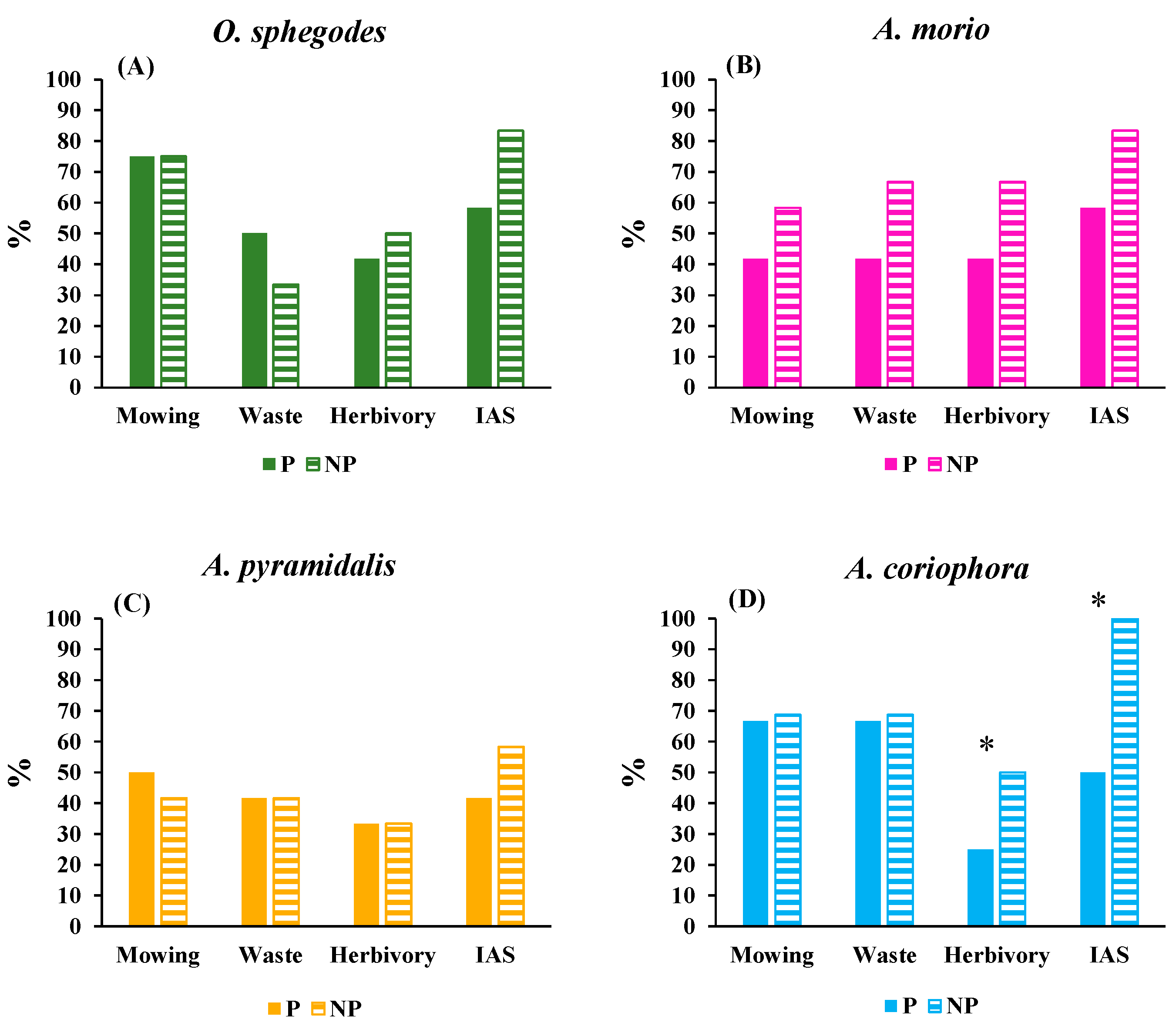
Figure 1.
Map of the study area. The pink larger dots are the sampling sites in protected areas, the white smaller dots are the sampling sites in non-protected sites.
Figure 1.
Map of the study area. The pink larger dots are the sampling sites in protected areas, the white smaller dots are the sampling sites in non-protected sites.
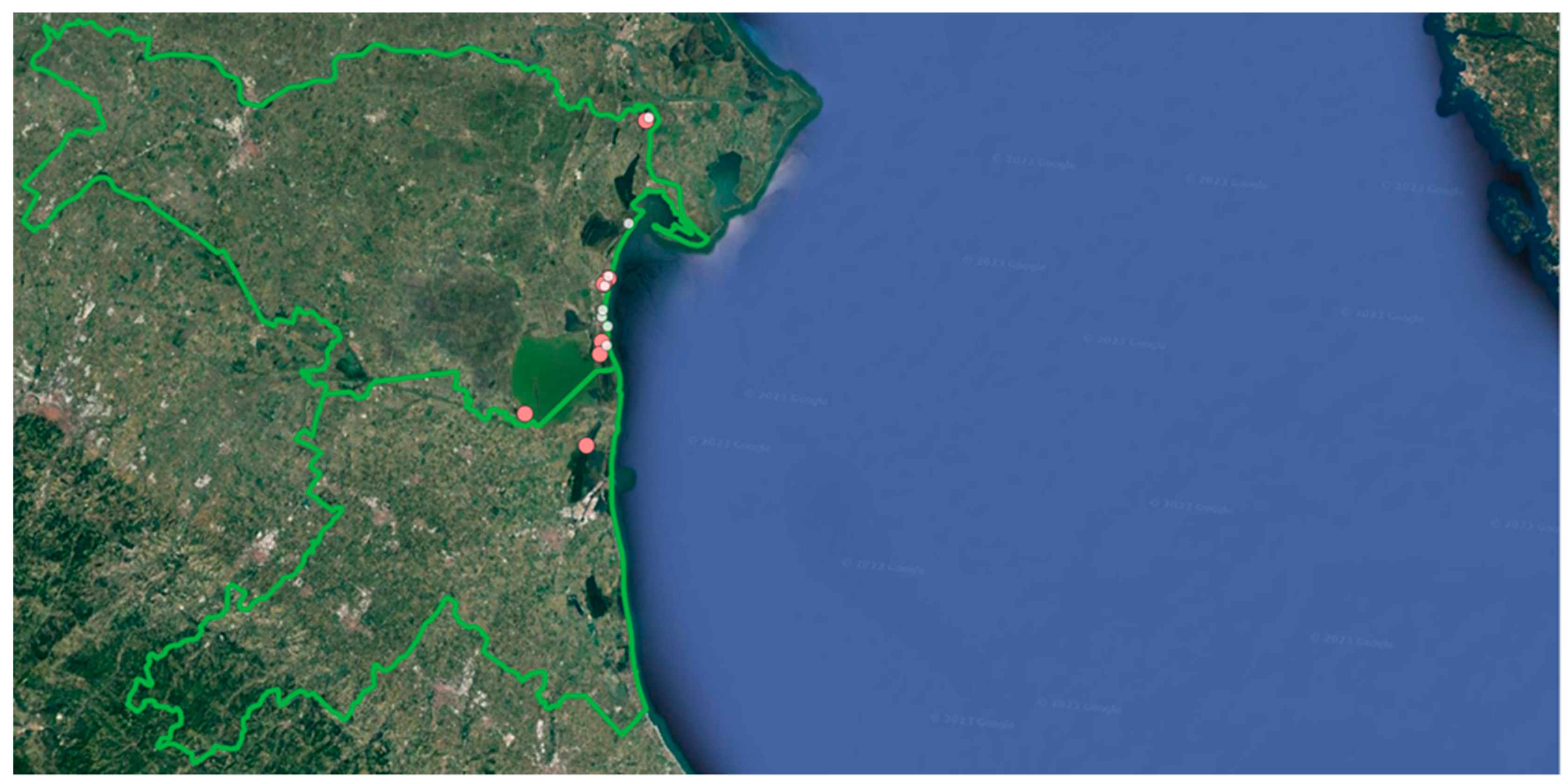

Table 1.
Mean (± 1 SE) values of reproductive traits in the four orchid species at protected (P) and non-protected (NP) sites. The asterisks indicate significant differences between P and NP obtained by one-way ANOVAs. n.a. = not available.
Table 1.
Mean (± 1 SE) values of reproductive traits in the four orchid species at protected (P) and non-protected (NP) sites. The asterisks indicate significant differences between P and NP obtained by one-way ANOVAs. n.a. = not available.
| Trait | O. sphegodes | A. morio | A. pyramidalis | A. coriophora | ||
|---|---|---|---|---|---|---|
| Number of flowers | P NP |
4.60±0.20 4.48±0.19 |
7.61±0.37 9.66±0.46* |
44.20±2.97 49.47±2.78 |
22.89±0.79 20.93±0.74 |
|
| Number of fruits | P | n.a | n.a | 2.79 ± 0.42 * | 15.80 ± 0.70 | |
| NP | n.a | n.a | 1.36 ± 0.22 | 13.86 ± 0.91 | ||
| Seed mass (mg) | P | n.a | n.a | 0.78 ± 0.12 | 5.30 ± 0.30 * | |
| NP | n.a | n.a | 0.69 ± 0.11 | 2.00 ± 0.20 | ||
| Number of embryos | P | n.a | n.a | 42.62 ± 5.98 | 96.37 ± 1.90 * | |
| NP | n.a | n.a | 39.78 ± 5.42 | 48.45 ± 4.52 | ||
Table 2.
Q values indicating population vitality in the four orchid species in terms of vegetative (V) and reproductive (C) traits at protected (P) and non-protected (NP) sites.
Table 2.
Q values indicating population vitality in the four orchid species in terms of vegetative (V) and reproductive (C) traits at protected (P) and non-protected (NP) sites.
| Species | Protection | Trait | Q |
|---|---|---|---|
| O. sphegodes | P | CV | 0.494 |
| NP | CV | 0.500 | |
| P | CR | 0.079 | |
| NP | CR | 0.082 | |
| A. morio | P | CV | 0.428 |
| NP | CV | 0.383 | |
| P | CR | 0.058 | |
| NP | CR | 0.076 | |
| A. pyramidalis | P | CV | 0.303 |
| NP | CV | 0.395 | |
| P | CR | 0.220 | |
| NP | CR | 0.211 | |
| A. coriophora | P | CV | 0.311 |
| NP | CV | 0.350 | |
| P | CR | 0.483 | |
| NP | CR | 0.267 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



