
Submitted:
02 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
This comprehensive review is devoted to an under-exploited family of phenolic compounds, the coumarins, and the most relevant strands in which they are involved in some spirit beverages – wine spirit, brandy, whiskey, sugar cane spirits (rum and cachaça) and grape marc spirit – with great importance worldwide in terms of production, trade and consumption. It gathers the key discoveries on the topic considering the production process of each spirit beverage, the related sources of coumarins (different kinds of wood used in the ageing stage) along with the factors that govern them and can influence the sensory properties of the aged beverages. An overview of the analytical methods available for their identification/quantification is also included, as well as the corresponding trends for the advancement of knowledge in this field. Moreover, the remarkable role of coumarins as nutraceuticals, their importance as chemical markers for authenticity purposes, and their relationship with food safety of these spirit beverages are also addressed, highlighting the current gaps and issues, and providing clues for future research.
Keywords:
coumarins
; wine spirit
; brandy
; grape marc spirit
; whiskey
; rum
; cachaça
; ageing
; nutraceuticals
; authenticity
; food safety
1. Introduction
According to the European Regulation [1], spirit beverages are classified into 44 categories; some of which are marketed without ageing while others undergo an ageing stage during their production process and are marketed as aged spirits. Among them, wine spirit, brandy, whiskey, sugar cane spirits (rum and cachaça) and grape marc spirit are traditionally aged in wooden barrels, and have great importance worldwide in terms of production, trade and consumption [2,3,4] - Figure 1. The spirit beverages accounts for c.a. 37% of the global alcoholic drinks market, the market's largest segment with a value of 408.8 billion USD in 2023 [4]. In addition, the latter is projected to grow c.a. 23% in the 2023-2028 period, reaching 1,373.9 billion USD; specifically, the spirits segment is expected to grow by 2.2% in 2024 and keep up this trend until 2029 [4,5]. Evidence supports the notion that premium spirit beverages have a favourable impact on driving market expansion. This helps mitigate market constraints caused by the adverse effects of excessive alcohol intake on consumer health [5,6]. Indeed, moderate alcohol consumption behaves as a stimulant, due to the direct absorption into the bloodstream from the stomach, while excessive consumption (≥ 15 drinks per week) can affect the central nervous system and cause chronic diseases [7,8]. In almost every world's cultures, spirit beverages have been appreciated due to their composition and sensory properties, and because they can be stored for long periods; their consumption is socially acceptable and they play an important role in the diet [9,10]. Therefore, the six types of aged spirit beverages aforementioned, enriched in wood compounds that enhance the distillates quality through the sensory and nutraceutical features imparted, can have a prominent role, contributing to the development of the related value chains, meeting consumer preferences, and driving market growth.
The freshly distilled spirits have high ethanol content (80-96%) and are rich in volatile compounds derived from the raw materials and/or formed during the operations composing their production process up to distillation (described in Section 2), but are devoid of phenolic compounds other than some volatile phenols, and of some furanic aldehydes [11,12,13,14,15,16,17,18,19,20]. Thus, contact with wood during the ageing stage is a core aspect in the manufacture of these beverages, as it confers them unique physicochemical and sensory characteristics, resulting from many phenomena involving the distillate and wood compounds, as well as the oxygen that enters the barrel by diffusion or is applied by micro-oxygenation [14,18,19,21,22,23,24].
Specifically, phenolic compounds (detailed in Section 3), released from the wood into the distillate, play a notable role in the distinctive characteristics acquired by these spirits - colour, aroma, flavour (including bitterness) and mouthfeel sensations (such as astringency) [22, 25,26,27,28,29,30] - which are not significantly modified during storage in bottle [31], and can therefore be perceived by the consumer.
Although many studies have been carried out to gain knowledge on the phenolics involved and their relationship with the factors that govern ageing, as well as their repercussions, the information available in the literature on coumarins is limited. The main sources of coumarins are examined in Section 4. According to our knowledge, research on coumarins began in the 1960s-1970s with the works of Baldwin et al. [32] in whiskey, Bricout [33] and Joseph and Marche [34] in wine spirit, and Otsuka and Zenibayashi [35] in brandy, rum and whiskey. However, it wasn't until the late 1980s that new studies were conducted, particularly in wine spirit [36,37,38]. Recently, this topic has been addressed for wine spirit [39,40,41,42], cachaça [43,44,45], whiskey [22,46,47], brandy [48], grape marc spirit [22] and rum [22]. This chronology can be ascribed to:
i) the evolution of analytical methods supported by new technologies (described in Section 5);
ii) growing attention paid by the consumer and the scientific community to nutraceutical quality, even in alcoholic beverages, because there is greater awareness that the presence of bioactive compounds, such as coumarins, can partially mitigate the harmful effects of alcohol [49]. From this perspective, scientific outcomes are mainly related to wine [50,51], and only a few works have been devoted to whiskey [52], wine spirit [53,54,55], brandy [54,56], and rum [57] (described in Section 7). This could be an important driver for the six related industries (wine spirit, brandy, whiskey, rum, cachaça, and grape marc spirit);
iii) health concerns on the part of the consumer, industries and food science researchers, leading to an interest in food safety. However, little is known about the relationship between coumarins and the food safety of spirits (disclosed in Section 8);
iv) the importance of chemical markers for authenticity purposes, which is a key aspect in the production and trade of spirits, in order to protect crucial players in each value chain: the producers (the Designations of Origin, if applicable) and the consumers [58,59]. Coumarins are potential chemical markers - for example, they are associated with oak wood versus chestnut wood in the ageing of wine spirits [38,40] - but this requires knowledge on their contents and their variation with the ageing conditions for the different spirit beverages (discussed in Section 6 and Section 9).
Despite coumarins are minority phenolic compounds in these matrices [19], a thorough understanding of the above-mentioned topics is imperative to understand their importance, to explore/manipulate and control the ageing stage, and consequently the production process, in order to shape the final product profile according to the target consumer, while guaranteeing quality, sustainability, authenticity and food safety. This review aims to summarise, for the first time, the existing knowledge on coumarins in the most relevant spirit beverages, focusing on the issues highlighted above.
2. Spirit beverages: aged and unaged
According to the European Regulation [1], the spirit beverage is an alcoholic drink meant for human consumption and produced by direct distillation of fermented and/or macerated products or indirectly by the addition of other spirit drinks, with specific sensory properties, and a minimum alcoholic strength by volume of 15% vol., with the exception of egg-based liqueur.
The spirits beverages addressed in this work are, as aforementioned, wine spirit, brandy, whiskey, rum and cachaça, and grape marc spirit. Wine spirit and grape marc spirit are a derived product and a sub-product of wine, respectively [60], whiskey is made from cereals, such as malt, barley, maize, wheat and rye [14], and rum and cachaça are produced from sugar cane [13].
2.1. Wine spirit
Wine spirit is obtained by the process of distilling wine, fortified wine intended for distillation, or a wine distillate with an alcohol content of less than 86% vol., with a methanol content of less than 200 g/hL of 100% alcohol, and a volatile substances content of 125 g/hL of 100% alcohol or more. The minimum alcoholic strength of wine spirit is 37.5% vol., and no alcohol is allowed to be added. The addition of caramel to adjust the colour is permitted, but the flavouring is not allowed. If it is aged, a minimum of six months is required [1].
The wine spirit production process (Figure 2) begins with the harvest of grapes in the vineyard, knowing that some changes start to occur in the grapes, such as the oxidation of phenolic compounds and the release of methanol by the pectins [61]. The grapes are crushed and pressed, and the must, without addition of sulphur dioxide [62,63], is placed in a tank for alcoholic fermentation to take place, under controlled temperature and density (or specific gravity). Thereafter, the wine is distilled and traditionally aged in wooden barrels for at least six months. After ageing, finishing operations are performed, which include blending, dilution, cooling and filtration, and finally bottling.
In Europe, the three main Protected Designations of Origin (PDO) of wine spirits are Cognac, Armagnac and Lourinhã. The difference between wine spirits from these Designations of Origin lies essentially in the terroir, the distillation stage (process and type of distillation system used) and the ageing stage [60,61,62,63]. Cognac and Lourinhã wine spirits must be aged for at least two years in wooden barrels [64,65] while Armagnac wine spirit must be aged for at least one year in wooden barrels [66].
2.2. Brandy
The name “brandy”, which was originally "brandwijn" (burnt wine), is ascribed to the Dutch, who began distilling wines from the Cognac region to preserve them in the 16th century [62]. This spirit beverage is produced from wine spirit, with or without addition of wine distillate (distilled at less than 94.8% vol. and not exceeding 50% of the alcohol content of the final product). The brandy must be aged for a minimum of six months in oak barrels. The limits set for methanol and the volatile substances are the same as for wine spirits. The minimum alcoholic strength is 36% vol., and no alcohol can be added. Brandy must not be flavoured, and the addition of caramel is permitted to adjust the colour [1,67].
The brandy production process starts with the harvest of grapes in the vineyard. The grapes are crushed, pressed, and the must, with the addition of Saccharomyces cerevisiae yeasts and without addition of sulphur dioxide [17,63], is placed in a tank for alcoholic fermentation to take place, with temperature and density control. The wine is then distilled, and the distillate is aged. After ageing, the finishing operations are carried out (blending, dilution, cooling and filtration) and finally bottling.
The major difference between the technological process of producing brandy and wine spirit lies in the ageing and finishing stages.
Brandy de Jerez or Sherry brandy is the best known, and has a unique and dynamic ageing system consisting of Criaderas and Solera [1,68,69]. The brandy is stored in barrels of different groups/scales, depending on its age. The Solera contains the longest-aged brandy and is located at ground level [1,70]. The scale above the Solera is called the First Criadera, which contains brandy that is younger than the Solera but older than the scale above - the Second Criadera - and so on, up to the last scale, which contains the youngest brandy [1,70] (Figure 3).
2.3. Whiskey
The name “whiskey”, established in the 18th century, derives from the translation of aqua vitae (distilled alcohol in Latin), which was converted into “uisge baetha” (Gaelic) by Irish monks, and later on this expression gave rise to the terms “uiskie” (shortened form) and “whiskey” [71].
Whiskey is produced solely by distilling a cereal mash, which may or may not include malted cereal grains. It could be saccharified by the diastase of the malt, along with the action of other natural enzymes, and fermented by yeasts [1]. The cereals predominantly used for the manufacturing of whiskey are barley (malted and unmalted), corn, rye and wheat. Cereals grains do not undergo natural fermentation and thus need to be converted into a fermentable substrate. This process involves the modification of their structural components (by malting or by milling and cooking) to release the starch, which is then converted into sugar by enzymes under controlled temperature [72,73] (Figure 4). Temperature control is essential for the reactions between amino acids and sugars to take place, giving rise to a variety of flavour compounds and a suitable alcohol yield. The liquid formed at this stage is called wort [73]. The wort is fermented through the action of yeasts, essentially from the Saccharomyces cerevisiae species [72]. A fermented liquid is obtained, known as wash, which is then distilled in column stills (continuous system) or in alembics (discontinuous), depending on the type of whiskey to be produced. Each distillate must have an alcohol content of less than 94.8%, as well as the aroma and flavour of the cereal(s) used [1]. The distillate is placed into wooden barrels to age and acquire the intended sensory properties [72,73]. The ageing conditions depend on the production region; for example, in Europe, it lasts no less than three years in wooden barrels with a maximum capacity of 700 L and normally reused [1], while in the USA (Bourbon and Tennessee), a minimum of two years in new oak barrels (subject to charring) with a maximum volume of 190 L is required [74]. In addition, only water and caramel can be added to the final distillate, although maintaining the sensory properties resulting from the production process. The minimum alcoholic strength is 40% vol. [1].
Whiskeys are produced all over the world, but the most widely recognised are from Scotland, Ireland, USA, Canada, and Japan. They are basically categorised according to the cereal (type and proportion of grains) and the production process used [72] - Figure 5.
2.4 Sugar cane spirits
Sugar cane spirits result from the distillate obtained from the molasses or juice of sugar cane (Saccharum officinarum L.) - Figure 6.
2.4.1. Rum
The distillate must be obtained at less than 96% vol., allowing the sensory properties of the raw material to be perceived. The minimum alcoholic strength is 37.5% vol., and no alcohol may be added. These spirit beverages cannot be flavoured, however the addition of caramel to adjust the colour is permitted [1].
According to Nicol [13], the “rum” designation has three possible etymological origins: i) from the Latin name for sucrose - Saccharum; ii) from the American words used in Devon - “Rumbullion” and “Rumbustion” (associated with the great tumult caused by the imposition of laws and taxes on sugar and molasses, considered one of the causes of the American Revolution); iii) from the Spanish name for this beverage - “ron”.
The raw material used for rum production are molasses and/or juice from sugar cane. The sugar cane is peeled and sliced before it goes to the mill. To maintain the quality of the cane, the duration between cutting and milling should not exceed 24 hours [13]. The stages of the pre-fermentation process are milling, extraction, clarification/decantation, filtration and dilution. Sulphuric acid is added to the diluted molasses and juice to reduce the pH to 4.5 - 4.8. Additionally, ammonium sulphate is also added as a nitrogen source to promote yeast development [12,13,75]. For fermentation purposes, Saccharomyces cerevisiae, Saccharomyces. Bayanus or Schizosacharomyces pombe yeasts are typically added. The fermentation’s time is only 24 hours [12,13,75]. Distillation is carried out in single or double alembics and distillation columns, after which the distillate is aged. Rum can be classified into two groups according to its colour and flavour: white rum, which is filtered to remove the colour acquired during ageing; and aged rum [27].
Rum is mainly produced in the Caribbean islands (Jamaica, Cuba, Trinidad and Tobago, among others) and Central America countries [13]. There is also the Madeira’s Rum, which is a Protected Geographical Indication (PGI); the beverage is obtained by the alcoholic fermentation and distillation of sugar cane juice produced in this Portuguese island [76].
2.4.2. Cachaça
The “cachaça” designation has two possible origins: from the Iberian word “cachazza”, a cheap beverage consumed in Spain and Portugal; from the feminine of “cachaço”, part of the pig, because this liquid was traditionally used to tenderise wild pig meat [12].
According to Brazilian legislation [77], cachaça is a typical spirit beverage, exclusively produced in Brazil, with an alcoholic strength ranging from 38% to 48% vol., and it is produced by distilling fermented sugar cane juice. Furthermore, sugar (up to 30 g/L) is allowed to be added for taste correction. The stages of the pre-fermentative process are the same as for rum. For fermentation purposes, only Saccharomyces cerevisiae yeasts are added and the fermentation’s time varies from 20 to 30 hours [12,13,75]. Single or double alembics and distillation columns are used for distillation (Figure 6). To be classified as aged cachaça, it must contain at least 50% of its volume aged in wooden barrels with a maximum capacity of 700 L for at least one year [77].
2.5. Grape marc spirit
Grape marc is a by-product of winemaking composed of skins, stems and seeds that can be reused as raw material, giving rise to a new product, called grape marc spirit [78,79]. According to the European regulation [1], the grape marc spirit is produced solely from the distillation, directly by steam or by the addition of water, of fermented grape marc, and each distillation is carried out at less than 86% vol. A quantity of 25 kg of wine lees can be added to 100 kg of grape marc for distillation. The methanol content of grape marc spirit should be less than 1000 g/hL of 100% alcohol, and the volatile substances contents should be 140 g/hL or more of 100% alcohol. The minimum alcoholic strength is 37.5% vol., and no alcohol may be added. This spirit beverage cannot be flavoured, and addition of caramel is only allowed to adjust the colour.
It is worth mentioning that, in the traditional winemaking process, red grape marc is obtained after alcoholic fermentation and contains residual sugars and some ethanol, while white grape marc is obtained after pressing and only contains residual sugars (Figure 7). Moreover, red grape marc usually has lower methanol content than the white one due to the distribution of this alcohol (formed from the pectins of the grape skin) between the must/red wine and the grape marc [80]. According to Belchior [81], grape marc needs to be slightly oxygenated before ensiling in order to favour yeasts' multiplication during fermentation. The grape marc is ensiled for the shortest possible time (a few months) in horizontal tunnels or stainless-steel tanks and fermented under anaerobic conditions (Figure 7). Sometimes the grape marc is sprayed with an aqueous solution of tartaric acid at a concentration of 5 to 10% (w/w) before storage to lower the pH level, preventing bacterial contamination and reducing methanol production. The liquid formed during fermentation is stored to acidify other grape marcs [16,78,82]. The fermented grape marc can be distilled by two different processes: discontinuous and continuous. In both processes there are two phases: distillation of the fermentate and rectification of the distillate [16,78,82,83]. As the distillate is rich in methanol, a demethylation column can be included in the system to reduce its content to the legal limit [16,78,80,82].
3. Coumarins in the context of phenolic composition
The name “coumarin” derives from the French term “coumarou” for the tonka bean that are the seeds of Dipteryx odorata (Aubl.) Forsyth f. (or Coumarouna odorata Aubl.), in which coumarin was first isolated by Vogel [85,86]. This compound has a sweet odour that is easily identified as the aroma of freshly mown hay; as a result, it has been used in perfumes since the late 19th century [87,88].
Coumarins are important secondary metabolites that can be found abundantly in the plants (detailed in Section 4). Numerous studies have been conducted on the isolation, structural characterization, synthesis, and biological activity of an extensive variety of natural coumarins, derived from trees and plants, including some used in the production of beverages [89,90,91,92,93], as well as coumarins obtained by chemical synthesis [94]. Regarding its chemical structure, natural coumarins are unsaturated lactones that belong to the C6C3 chemical class, that is, a class of benzopyrones (1,2-benzopyrones or 2H-1-benzopyran-2-ones; consisting of a benzene ring fused to an α-pyrone ring), which can be thought of structurally as ortho-hydroxy-cinnamic acid derivatives (Figure 8). The majority of natural coumarins, including umbelliferone, scopoletin, aesculetin, among others, include an oxygenated substituent at position 7 [95]. This substituent can exist either in its free form, as seen in hydroxylated umbelliferone, or in combination with other compounds (methyl, ethyl, saccharides, etc.) in different derivatives. These compounds exhibit a conjugated system that is rich in electrons, resulting in favourable charge-transport features [91,96].
Several methodologies for classifying coumarins can be found in the literature. Among them, the most comprehensive, which includes coumarins with the greatest complexity and substituted coumarins, often found in combination with other heterocycles [88], categorises these compounds into the following groups: simple coumarins, furanocoumarins, dihydrofuranocoumarins, pyranocoumarins (both linear and angular), phenylcoumarins, bicoumarins, and coumarin glycosides, as depicted in Figure 9 [97].
Simple hydroxycoumarins, furanocoumarins and isofuracoumarins, pyranocoumarins, bicoumarins, and dihydroisocoumarins are the most important compounds of this chemical family isolated from plants. In plants, coumarins often exist in the free form due to their polar nature, and a significant number of these compounds have the ability to undergo sublimation. Additionally, they can also be present in the form of glycosides, including psoralen-correlated compounds. These entities exhibit a distinct blue fluorescence owing to their absorption of UV light, and they possess a high degree of photosensitivity, making them susceptible to modification by natural light. These properties are exploited for separation and analysis purposes, as well as in unconventional therapeutic approaches such as photochemotherapy and the chemical sensor industry (detailed in Section 5 and Section 7).
Simple coumarins are synthesised biogenetically by the transformation of shikimic acid into cinnamic acid. The underlying primary enzymatic transformation is the C-2 hydroxylation, which results in the cleavage (β-oxidation) of the side chain, or alternatively, chain isomerization followed by lactonization, leading to the formation of umbelliferone (Figure 10). Shikimate dehydrogenase (AroE) promotes the transformation of shikimate and NADP+ into 3-dehydroshikimate, NADPH, and H+. The shikimate pathway is a metabolic system consisting of seven steps and it is involved in the synthesis of phenylalanine, tyrosine, and tryptophan.
Shikimic acid is also the source of furanocoumarins and pyranocoumarins, which have a wide range of biological activity and are present in many natural products and synthetic drugs [98].
Furanocoumarins consist of a furan (or dihydrofuran) ring fused with the coumarin skeleton and can be classified as linear or angular; a good example can be seen in the structure of psoralen [99]. These structures are derived from umbelliferone by the addition of a prenyl group [100]. In general, furanocoumarins are biosynthesized involving two pathways, the phenylpropanoid and the mevalonic acid ones, by a coupling of dimethylallyl pyrophosphate (DMAPP) with umbelliferone, as well as through the formation of a prenylated simple coumarin intermediate [99].
Pyranocoumarins typically consist of a pyran (or dihydropyran) ring that is usually fused with the aromatic ring of the coumarin skeleton, as in xanthyletin [99]. Based on where the isopentenyl pyrophosphate is condensed to further cyclize and produce the heterocycle, these coumarins can be classified as linear or angular. The synthesis of these complex coumarins might possibly be the outcome of the cyclization of a prenylated simple coumarin [101]. Phenylalanine, a compound generated by the shikimate biosynthetic pathway, undergoes conversion by phenylalanine ammonia lyase (PAL) into trans-cinnamic acid, which gives rise to the core metabolite 4’-coumaroyl-S-CoA. This intermediate can subsequently be converted into a variety of phenylpropanoids via 6’-hydroxylation, trans > cis isomerization of the exocyclic double bond, and the final lactonization/cyclization step (Figure 11). The initial and crucial step in biosynthesis is 6'-ortho-hydroxylation, which is carried out by the 2-oxoglutarate-dependent dioxygenase F6'H1.
Some works have been devoted to clarify the biosynthesis pathways of scopoletin at a molecular level. Scopoletin, like umbelliferone, undergoes benzene ring alterations, and studies conducted on Arabidopsis thaliana demonstrated that scopoletin is produced through the phenylpropanoid pathway via ortho-hydroxylation of cinnamate, p-coumarate, caffeate, and ferulate [102,103]. Similarly, these coumarins, as well as esculetin, can be produced via the phenylpropanoid route (Figure 12), specifically generated from trans-cinnamic acid, but instead of undergoing through the lactonization process of the caffeic acid derivative, 2’-hydroxy-caffeic acid, they can be synthesised via umbelliferone [104]. Cinnamic acid also plays a role in the synthesis of scopoletin, as an intermediate, but this time through caffeic acid ester derivatives. Besides the synthesis of coumarins, the phenylpropanoid route is responsible for the synthesis of other phenolic compounds, such as flavonoids and phenolic acids.
Furthermore, bicoumarins deserve attention as they are a category of great interest for pharmaceutical applications, particularly as anticoagulants. Bicoumarin connects two coumarin moieties by an orthoester structure, being the dicoumarol the first isolated compound [105]. Dicoumarol, in particular, consists of two cyclic β-ketoesters linked by a methylenic bridge, and was isolated from plant sources, in which the two coumarin units are linked at C3-C3’ via methylene group [106] (Figure 13). One possible route for the biogenesis of this compound might be the hydroxylation of the carbon atom at 4-position of coumarin framework, which then captures a molecule of formaldehyde, and is condensed with another molecule of 4-hydroxycoumarin, and finally enolises the keto group that forms dicoumarol [107,108].
Dicoumarol, and related compounds with substituent groups (R) on carbon atoms at 3, 5, 6, 7 and 8-positions, are often addressed due to their unique molecular structures, which may include two intramolecular O–H···O hydrogen bonds (Figure 13b) and different biological features depending on type of substituents on the central methylene linkage (3-position). The parent molecule, the 4-hydroxycoumarin, which may be represented as one of three tautomeric structures (Figure 14), effectively depicts the structural changes occurring in dicoumarol. Some studies have shown that coumarin form A (Figure 14) is the primary tautomer in both the solid state and in solution in polar solvents [109,110], which is relevant in the context of spirit beverages.
In general, nature exhibits a certain degree of creativity; given the proper building blocks, it is capable of producing the most varied compounds; bicoumarins are an example of this matter. A wide variety of bicoumarins are available, each of which is composed of coumarins connected by carbon-carbon bonds or an ether linkage. Nevertheless, these links are located on different carbon atoms, such as C3-C3', C3-C6', C3- C7, C3-C8', C4-C4', C8-C8', C5-C6', C6-C6 C6-C8', C5-C8', C7-C7', C7-C8', C8-C8', or C6-C6'-linked bicoumarins. Spirobicoumarins, monoterpene-unit linked bicoumarins, and C3-C3'-linked with methylene and methine group-linked bicoumarins are further examples of bicoumarin sets [105].
Being aware of the metabolic pathways of compounds such as coumarins is useful for a variety of reasons, including nutraceutical and safety evaluations (detailed in Section 7 and Section 8). Determining how coumarins are metabolised can be useful for identifying the risks associated with their consumption and defining safe levels of exposure. In terms of coumarin metabolism, two major processes have been identified: 7-hydroxylation (umbelliferone) and the opening of the lactone ring with the loss of carbon dioxide (Figure 15). This reaction takes place on the intermediate coumarin 3,4-epoxide produced in the phase I of the metabolic process. Under aqueous conditions, this epoxide releases carbon dioxide to create ortho-hydroxyphenyl acetaldehyde (o-HPA), which can subsequently be metabolised to the corresponding acid (o-HPAA) and alcohol (o-HPE). A series of intermediates can be formed when glutathione nucleophilically attacks the 3,4-epoxide, including the 4-HDHC-GSH (4-hydroxy-3,4-dihydrocoumarin-3-mercapturic acid) or 3-hydroxycoumarin.
Understanding coumarins’ metabolic pathways is crucial for determining the toxicity of the metabolites generated through excretion, as well as for extrapolating the formation of coumarins in spirit beverages through various chemical processes such as oxidation, reduction, hydroxylation, radical reactions, electrophilic or nucleophilic additions, and metal-catalysed additions (mainly copper and iron) [28,114,115,116]. These processes may occur during the prolonged ageing of beverages in wooden barrels and may cause changes in the coumarins profile.
4. Sources of coumarins
Coumarins have been isolated from hundreds of plant species belonging to more than 40 families [117], and in several parts/tissues: leaves [118], flowers [119], fruits [117,120], seeds [121], stems [122], wood [38,123,124], inner bark [112] and roots [125,126]. Indeed, phenolic compounds play a significant role in the intricate defence mechanism of plants, and their biosynthesis is triggered by abiotic stress, such as UV radiation and ozone [108,127,128,129], and biotic stresses such as infections and wounding [108,130,131]. For these reasons, coumarins are considered phytoalexins; some of these compounds have been found in high concentrations, while others, as is usually the case with coumarins, even in low concentrations are involved in the plant's defence [104,132].
Coumarins are found in the raw materials used in the production of spirit beverages addressed in this work: i) grapes [133] and wine [22,134] in grape marc spirit, wine spirit and brandy manufacture; ii) cereal seeds [135,136] in whiskey manufacture; iii) sugar cane [137] in rum and cachaça manufacture. However, they are not released from the wine, the grape marc or the wash into the steam during distillation, and consequently, they are not present in the corresponding distillates. Thus, the only source of coumarins is the wood used in their ageing. According to the literature, among the many coumarins found in plants, those present in wood used for the ageing of spirit beverages are shown in Table 1, and their structures are elucidated in Figure 16.
The results attained for coumarins in these studies clearly reveal the differences imparted by the botanical species. Moreover, variation in their concentrations within each kind of wood is observed. It reflects the influence of some well-known factors, such as the geographical origin and the single-tree [124,139,143,144,145,146,147,148], the wood age [149,150,151], as well as the analytical methods used in their quantification. Details on the last topic are addressed in Section 5.
Nevertheless, scopoletin was the most plentiful compound in the majority of the wood types examined, followed by umbelliferone, aesculetin and aesculin in the oak wood, and coumarin in the Brazilian kinds of wood. On the other hand, among the oaks, higher levels of scopoletin were found in American oak wood, followed by Q. robur wood, and lower levels were exhibited by Q. pyrenaica wood. Regarding the other species, cherry wood presented an intermediate scopoletin content, while very low concentrations were observed in Portuguese chestnut, amendoim, cabreúva-parda and canela-sassafrás, and it was not detected in the remaining species. Umbelliferone was also more abundant in American oak wood, its levels were similar in the other kinds of oak wood, and it was not detected in the other species. Scopoletin and umbelliferone have been reported as chemical markers of oak wood [19,152]. Aesculetin contents in different oak wood were quite similar, except in Q. faginea which was richer in this compound, and higher contents were associated with cherry wood and mulberry wood from Serbia. Aesculin was quantified in Q. robur and Q. sessiliflora wood, which presented lower contents than mulberry and myrobalan plum wood. Finally, the wood from Brazilian species can be classified into three groups according to the coumarin content: jatobá, with the highest amount; pequi, amendoim and cabreúva-parda, with intermediate levels; canela-sassafrás and castanheira, with the lowest content.
The effect of the wood type on the concentration of coumarins in the spirit beverages under study is therefore predictable, and will be discussed in Section 6.
Additionally, it is noteworthy the potential use of waste from these kinds of wood resulting from the barrel making or production of fragments in the cooperage industry (closely associated to the aged beverages industries) as sources of coumarins for other purposes, such as natural preservatives and fortifying agents in the food industries [153,154] and as bioactive compounds for the pharmaceutical industry [97,155,156,157,158,159,160].
5. Methods for identification and quantification of coumarins
A range of analytical methods are commonly employed to identify and quantify coumarins, as well as other compounds found in spirit beverages. Different techniques can be applied, depending on criteria such as the characteristics of the sample, the desired level of sensitivity, and the equipment available.
The identification of natural coumarins is mostly based on their fluorescence, as they produce a blue hue when exposed to UV light (as mentioned in Section 3), and it is precisely because of this property that they are frequently characterised by spectrometric techniques. Table 2 presents a concise overview of the excitation and emission wavelengths for prevalent coumarins that occur in wood and aged spirit beverages. This feature provides the basis for the development of identification and quantification techniques [161]. On the other hand, coumarins are photosensitive and can change with natural light, so these characteristics are used for their isolation and analysis as well [117,161].
The most common methods for the separation, identification and/or quantification of coumarins include: Thin-layer Chromatography (TLC); Gas Chromatography (GC); High-Performance Liquid Chromatography (HPLC); Ultra-Performance Liquid Chromatography (UPLC); Liquid Chromatography-Mass Spectrometry (LC-MS); Capillary Electrophoresis (CE); UV-Vis spectrophotometry; Fluorescence Spectroscopy.
5.1. Sample preparation
A variety of methods for extracting coumarins has been reported in the literature. Extraction can be carried out on fresh or dried material, with solvents of different polarities, depending on the coumarins’ structure [117]; methanol, ethanol, benzene, chloroform, diethyl and petroleum ethers or combinations are often used for this purpose. Thus, coumarins in the free form and as glycosides are extracted exhaustively with ethanol and its aqueous solutions, while compounds with medium polarity are soluble in chloroform, ethyl acetate and acetone [162,163]. Petroleum ether provides a good yield of furanocoumarins that can be isolated in the crystalline form. According to [162,163], several studies were devoted to the solvents used for extraction (with different concentrations, temperatures and extraction times) and the techniques applied (Soxhlet apparatus, ultrasonication, ultrasound-assisted extraction, microwave solvent extraction, accelerated solvent extraction, and pressurised liquid extraction.
Regarding the spirit beverages under study, different sample preparations have been reported (Table 3).
5.2. Analysis
Several methodologies have been developed for coumarins’ analysis. The selection of the appropriate technique is contingent upon several factors, including the particular coumarins of interest, the intricacy of the sample matrix, and the availability of suitable equipment. Often, a variety of methods are used together to accurately identify and quantify the amount of coumarins in a sample.
The first phase for the analysis is the process of separation, which was initially carried out by paper chromatography, thin layer chromatography and colourimetric tests. Currently, high-performance liquid chromatography (HPLC) is the most widely used method [168]. The chromatographic techniques mostly used to analyse coumarins are described below.
5.2.1. Thin-layer Chromatography
A simple procedure used for the separation of coumarins in spirit beverages is the thin-layer chromatography (TLC) - Table 4. It is a cost-effective, fast and widely-used technique. It resorts to a thin layer of stationary phase (typically silica gel or alumina) placed on a flat support such as glass or plastic. For quantification purposes, TLC must be combined with other analytical techniques [162,168,169].
In general terms, TLC includes the following four steps: sample application, separation, development and visualisation. Visualisation techniques include ultraviolet (UV) light, chemical staining, and exposure to additional detection techniques. The capacity to examine and identify coumarins on the TLC plate depends on the detection methods used, as well as the parallel elution of a standard for comparative purposes. Staining or chemical derivatization may also be used to detect some compounds that do not have natural fluorescence.
Although more advanced chromatographic techniques, such as HPLC or GC, are preferred for separation and subsequent quantitative and high-resolution analyses, allowing the automation of some steps, TLC remains a valuable tool, particularly when performing rapid qualitative analyses [168].
5.2.2. High-performance Liquid Chromatography
High-Performance Liquid Chromatography (HPLC) is a sophisticated chromatographic method used to separate, identify, and quantify components of mixtures. HPLC is extremely adaptable and can be used to analyse a wide range of substances, from small molecules to polymers. HPLC has been the most widely used method for identification and quantifying coumarins and its derivatives [163,168], particularly in spirit beverages (Table 4). An advantage of this technique is the possibility of coupling it to different spectroscopic or spectrometric detectors; for coumarins analysis, the most suitable/accurate one is the fluorescence detector (FDL) [168], taking advantage of the properties of these phenolic compounds (reported in Section 3). It should be noted that the separation of simple coumarins may not be accurate due to the similarity of chemical structure and polarity [168].
Ultra-Performance Liquid Chromatography (UPLC) was launched in 2004 [170] and represents a significant breakthrough in liquid chromatography, expanding on the concepts of HPLC and introducing improvements that allowed for even faster and more efficient separations. The UPLC technique is a viable alternative for the determination of coumarins, since it allows the development and application of a method with a shorter running time, lower solvent consumption and good separation/resolution of the chromatographic peaks [168].
5.2.3. Gas Chromatography
Gas Chromatography (GC) is a powerful analytical method employed to separate and analyse volatile and semi-volatile compounds in a mixture. The process relies on the concept of differential partitioning of sample components between a stationary phase (usually a liquid or polymer) and a moving phase (an inert gas like helium or nitrogen) that flows through a chromatographic column. Since this technique requires volatile analytes, it has mostly been used to identify and quantify furanocoumarins [162,163]. Although the matrices studied were essentially plant parts/tissues, it may be of interest to analyse coumarins in aged spirits.
5.2.4. Spectrometric detectors
Along with the chromatographic methods for the separation, spectrophotometry/spectrofluorimetry has also been used for coumarins analysis in spirit beverages (Table 4). The use of a spectrophotometer relies on the absorption capacity of coumarins in the ultraviolet range related to the transitions of π electrons from bonding to antibonding molecular orbitals. However, it requires a reference sample for a more accurate determination [162].
Technological advancements have led to the development of new equipment, which has improved the precision and accuracy of existing techniques used to detect and quantify coumarins even in very low concentration. These techniques include HPLC with diode array detection (DAD) and fluorescence detector (FLD) coupled to mass spectrometry (MS), GC or LC coupled to ESI-MS, MS-time of flight (TOF), MS-ion trap and MS-MS tandem quadrupole (Q)-TOF [43,170,171,172,173,174,175,176,177,178].
5.3. Analytical trends
The growing demand for more sensitive, selective, and efficient methodologies has propelled advancements in analytical techniques for the structural elucidation of organic molecules. Some of the cutting-edge analytical methodologies that will undoubtedly have an impact on the determination of coumarins in wood extracts and beverages are those that are currently being used for other organic substrates. High-Resolution Mass Spectrometry (HRMS), for example, enables exact determination of molecular weights, resulting in enhanced accuracy for identifying chemical formulas and assisting in the understanding of molecular structures [179,180]. Tandem Mass Spectrometry (MS/MS or MSn) procedures yield fragmentation patterns that can be employed to infer the structural characteristics of the compound [173,177]. Employing multiple stages of fragmentation (MSn) can unveil further intricate details. On the other hand, Nuclear Magnetic Resonance Spectroscopy (NMR), including 2D-NMR (two-dimensional NMR) and 2D homonuclear and heteronuclear correlation investigations, offers improved resolution and detailed information for determining molecular structure [181,182]. Ion Mobility Spectrometry (IMS) is a technique that separates ions in a gas phase depending on their mobility, which helps to distinguish between structural isomers, and might be very useful for chiral coumarins derivatives [183,184]. Another cutting-edge methodology is the integration of liquid chromatography with nuclear magnetic resonance (LC-NMR), which enables the concurrent separation and determination of the molecular structure of substances [185]. Computational techniques might be very useful as well, such as density functional theory (DFT) and molecular dynamics simulations, that are progressively employed to forecast and explain spectroscopic data, facilitating the determination of molecular structure [186]. The use of advanced data processing techniques such as Data Mining and Cheminformatics, along with machine learning algorithms, is employed and allows the interpretation of extensive analytical data [187]. These methodologies can accelerate the process of determining the structure of compounds, which can aid in identifying new coumarins in spirit beverages, and determining their metabolism. The employment of miniaturised analytical techniques and lab-on-a-chip equipment also might enable quicker analyses with smaller sample volumes, contributing to enhanced efficiency in structural elucidation [188,189].
These trends, taken together, can contribute to a more thorough and efficient approach to the advancement of knowledge on organic compounds, including coumarins.
6. Influence of ageing factors on the coumarins contents of spirit beverages
Scientific studies conducted over the last three decades have proven that the physicochemical phenomena that take place during the ageing of spirit beverages, particularly the release of wood compounds into the distillate, are governed by several factors, including the characteristics of the distillate, the ageing technology (traditional vs alternative), the kind of wood and its heat treatment, the barrel size, the cellar conditions (temperature, relative humidity and air circulation) and the ageing period [14,19,190]. In this context, the influence of some remarkable factors on the coumarins contents in the spirit beverages under consideration will be emphasised.
6.1. Wood
The wood from the European oak species Quercus robur L. is predominantly used for the ageing of wine spirit [19,62], brandy [191] and some grape marc spirits [18], while Quercus alba L., from North America, is commonly used in the ageing of Bourbon and Tennessee whiskeys [14,23,192], brandy [70,191] and rum [13,27,193].
Other botanical species, such as Quercus sessiliflora Salisb., Quercus pyrenaica Willd. and Quercus faginea Lam., grown in Mediterranean countries, are used in brandy ageing [21] and grape marc spirit ageing [18]. Chestnut wood (Castanea sativa Mill.) is also a valuable resource for this purpose in wine spirit [19] and brandy [21].
As far as whiskey is concerned, the production of this beverage in Scotland and Ireland is mainly based on barrels that have already been used for the ageing of Bourbon whiskey, Sherry wine and Brandy de Jerez, Port wine and Madeira wine, imported from the USA, Spain and Portugal, respectively [14,192], and therefore the American and European oak species also influences its chemical composition, specifically the phenolic one, including the coumarins contents.
In contrast, cachaça in Brazil is aged in oak wooden barrels but also in barrels made of wood from autochthonous species [43,45,190] such as those specified in Table 1.
As shown in Table 1, great variability exists on the coumarins contents in these kinds of wood, which has an impact on their concentrations in the spirits beverages aged with it (Table 5, Table 6 and Table 7).
According to the findings of these research teams (Table 5), for the two kinds of wine spirits (Armagnac and Lourinhã), scopoletin is quantitatively the most important coumarin, followed by umbelliferone, which is in line with the contents found in the wood used in their ageing (Table 1). Scopoletin concentration is particularly high in wine spirits aged in oak barrels, especially those made with American and French species, which was also noted by Winstel et al. [22] in commercial spirits. Conversely, their levels are much lower in the wine spirits aged in chestnut barrels. Hence, these results support the hypothesis that scopoletin and umbelliferone act as chemical markers of oak wood.
Low contents of aesculetin and 4-methylumbelliferone were quantified in Armagnac spirits. However, Winstel et al. [22] recorded the presence of these compounds, as well as of fraxetin in higher concentrations in Cognacs. Such differences, as above mentioned, can be assigned to the variability of wood chemical composition and to the analytical methods. Interestingly, as far as we know, 4-methylumbelliferone was only detected in oak wood by Winstel et al. [22], although its content was not disclosed.
Despite the extensive literature on brandies, including Brandy de Jerez, few studies have been devoted to analysing coumarins and their variation depending on ageing conditions, such as the type of wood used. The work of Mattivi et al. [123] showed similar contents of scopoletin in Italian brandies aged in oak barrels from Quercus robur and from Quercus sessiliflora (Table 6).
Considering that the raw material to produce wine spirit and brandy is the same (wine), it is interesting to compare the scopoletin content of the corresponding aged beverages. Regarding the wine spirits aged in Q. robur wooden barrels (Table 4), a considerable difference to Armagnac is observed, while a similar concentration then in the Lourinhã wine spirit studied by Patrício et al. [39] is found. However, a marked difference is noted for the Lourinhã wine spirit aged in Q. sessiliflora wooden barrels. Since the analytical method applied in the studies of Patrício et al. [39] and Canas [19] was the same, these discrepancies should be mainly ascribed to the wood variability.
Regarding whiskey, the research has shown that coumarins are present in this aged spirit, but to the best of our knowledge, there is no information available on their concentrations depending on the kind of wood. Indeed, Aylott et al. [194] studied five blended Scotch whiskies and reported a range of scopoletin concentration between 180 and 760 μg/L; the wood was not specified and, as these are blends, it is difficult to understand the effect of this ageing factor. Collins et al. [47] examined 63 commercial American whiskeys (including Tennessee whiskeys, Bourbon whiskeys and rye whiskeys, with different ageing periods); despite the differentiation made between the kinds of whiskey based on several compounds, including scopoletin and aesculetin, the wood species involved has not been unveiled. Similarly, Mignani et al. [46] conducted a trial aiming to distinguish single-malt Scotch whiskeys based on 18 samples; the analytical techniques used allowed to deduce that coumarins were associated with the fluorescent spectra obtained, although no results were presented on their identification and quantification. Winstel et al. [22] analysed nine commercial whiskeys aged in oak wood, including four Bourbon whiskeys, and quantified scopoletin, umbelliferone, aesculetin, 4-methylumbelliferone and fraxetin, but their contents were not disclosed.
Information on coumarins contents in rum according to the wood used is scarce. Winstel et al. [22] analysed three commercial rums, reporting an average content of scopoletin of 1500 μg/L. However, there is more substantial information about cachaça. Several studies were performed on cachaça, including wood from oak species and from species grown in Brazil (Table 7). Notwithstanding, it should be emphasised that some of the native species included in these studies do not correspond to the kinds of wood investigated (Table 1), and most of the experimental designs/available information do not allow a relationship to be established between the kind of wood used and the coumarin contents found in this spirit beverage. Santiago et al. [44] assessed the chemical composition of cachaça aged for 12 months in used barrels (200 L) made of European oak and new barrels (20 L) from four exotic kinds of wood. Results obtained after 12 months (Table 7) revealed very low contents of coumarins in the aged cachaça, and only the amburana and balsamo barrels released coumarins (quantifiable concentrations) into the distillate; the former was richer in these compounds, presenting a higher level of coumarin than 4-methylumbelliferone. However, these outcomes are not comparable due to the different ageing conditions used in those essays: barrel usage and barrel size. According to the literature, the pool of wood extractable compounds of used barrels is substantially lower than that of new barrels [195], the reason why oak barrels did not exert the expected effect. On the other hand, 20 L barrels have quite higher surface to volume ratio than 200 L barrels, promoting greater extraction of wood compounds as well as favouring other phenomena occurring during ageing [196].
Bortoletto et al. [29] reported their results as percentage of peak area of coumarin in cachaça aged for 12 months in amburana (Amburana cearensis (Allemão) A.C.Sm.), castanheira (Bertholletia excelsa Humboldt & Bonpland) and cabreúva (Myrocarpus frondosus Allem.) 220 L toasted barrels: 19.08%, 0.51%, and not detected, respectively. Same for hydrocoumarin in cachaça aged in these kinds of barrels: 8.08%, 0.69% and 0.55% respectively. The ageing in amburana barrels imparted the highest contents of coumarins, castanheira conferred intermediate levels, and cabreúva induced the lowest ones, which contradicts the results obtained by Silva et al. [43] for the wood of cabreúva and castanheira species (Table 1). This discrepancy may derive from the wood variability and the different analytical methods applied.
Finally, despite Bettin et al. [197] and de Aquino et al. [25] analysed many cachaça samples, from the market and from local producers, and presented concentration levels of two coumarins (scopoletin and coumarin), the relationship with the kinds of wood used in their ageing was not revealed. dos Anjos et al. [198] studied cachaça aged in 200 L oak barrels (unidentified species) for 12 months, and found that the levels of coumarin and 4-methylumbelliferone were lower than the limit of detection of the method.
To the best of our knowledge, no information is available in the literature on the coumarins’ contents of grape marc spirit depending on the type of wood used for its ageing.
6.2. Heat treatment
It is unquestionably accepted by the scientific community that the heat treatment of the wood, carried out during the cooperage process, plays a decisive role in the set of extractable compounds that can be released into the distillate during ageing [19,23,195]. In Europe, a fire with wood scraps is used to bend the staves and then toast their inner surface, while in North America heated steam is used to bend the staves, followed by a fire or a gas burner for charring (in a controlled manner) the inner surface of the wood [23,164,195,199,200,201]. Thus, in barrels made by the European technique, there is normally 6-8 mm of toasted wood, and in barrels made using the American technique there is normally 2-3 mm of charred wood followed by 3-5 mm of toasted wood. The aim of the bending phase is to give the barrel a concave shape without the staves breaking. The toasting phase is carried out intentionally to change the structure [202], physical properties [203] and chemical composition of the wood [164,200,201,204], thus imparting a particular character to the distillate aged in it. Though toasting procedures differ according to the cooperage, the toasting level is usually categorised as light, medium or heavy. The char (active charcoal) layer is required for some distillates, such as whiskey and cachaça, to decrease pungency and remove off-flavours associated with some constituents, such as sulphur compounds, through adsorption and oxidation [190,200,205,206,207].
Hydrothermolysis of the wood biopolymers (cellulose, hemicelluloses and lignin) and tannins mainly occur during toasting and leads to the formation of low molecular weight compounds [164,208], which can be easily extracted by the distillate during ageing, thus influencing its chemical composition and sensory properties. Cellulose gives rise to 5-hydroxymethylfurfural and 5-methylfurfural, hemicelluloses originate furfural, while lignin is the precursor of several phenolic compounds, including coumarins [200]. Ellagic acid and gallic acid result from the degradation of ellagitannins and gallotannins, respectively [209]. Subsequently, several reactions take place in the liquid medium, determining the features of the aged spirits [26,28,177,190,192]. Regarding coumarins, in a recent study, Sampaio et al. [210] showed that scopoletin is thermolabile, and underwent degradation in a temperature range from 182.32 to 303 °C, with mass loss of 99.98%. Besides, as far as we know, the effect of this technological factor on their contents has only been reported for wine spirits - Figure 17.
The results showed that toasting had a significant effect on the average concentration of umbelliferone in wine spirit aged for four years in 250 L barrels; an increase in concentration was observed as the intensity of wood toasting rose, reflecting the accumulation of this compound in the wood due to its remarkable thermal stability [211], which could then be extracted by the wine spirit. The concentration of scopoletin was slightly affected, but the highest level was associated with medium toasting, suggesting that lower temperatures favoured its formation, and as the toasting temperature increased, degradation phenomena exceeded the formation ones [210]. According to the works of Sampaio et al. [210] and Sun [211], the umbelliferone demonstrated greater thermal stability than scopoletin.
6.3. Ageing time
In the regulations of the European Union and other countries in which aged spirits are produced, a minimum ageing time is always defined. For those addressed in the present review, this requirement is specified in Section 2. Actually, the results of extensive research on this subject have clearly shown that the ageing stage requires a certain period of time for multiple physicochemical phenomena to occur, slowly and continuously, in order to confer specific characteristics to the distillates, contributing to the sensory fullness of the final products [14,18,19,21,190]. It is also recognised that these phenomena basically fall into two categories:
- Additive - including the ones that introduce or give rise to new compounds in the distillate undergoing ageing, such as extraction of wood compounds and lignin hydroalcoholysis;
- Subtractive - encompassing the ones that remove or modify some constituents of the distillate undergoing ageing, such as evaporation of volatile compounds, adsorption/degradation by the charred surface of the barrel, sorption in wood, and oxidation and hydrolysis reactions, among other chemical transformations.
Consequently, the chemical composition of the aged spirits evolved progressively over the ageing time as a result of the balance between additive and subtractive phenomena. Concerning coumarins, some available data in the literature (for whiskey, wine spirit and cachaça) are shown in Figure 18.
The results reveal that scopoletin, coumarin and 4-methylumbelliferone contents increase in the first years of ageing, which can be ascribed to the prevalence of addictive phenomena, being the higher concentration gradient between the wood and the distillate (devoid of these phenolic compounds) the most plausible driving force [212]. However, different patterns are observed depending on the compound, the kind of distillate, the wood and their interaction. Interestingly, after many years of ageing, the scopoletin content in wine spirit decreased sharply (Figure 18c), maybe reflecting the predominance of subtractive phenomena over additive ones. This behaviour may contribute to reducing the bitter taste of the aged wine spirit, in which coumarins seem to be involved [22]. The formation of coumarin glycosides often occurs through the process of hydroxylation and isomerization of trans-hydroxycinamic acid and analogous molecules, a reaction that can also occur during the ageing stage [115]. Due to the glycosidic bond, coumarin glycosides taste bitter. Bitterness comes from sugar moieties connected to the coumarin core. In plant defence mechanisms, bitterness discourages herbivores and insects from overeating [213]. When the sugar moiety is removed, coumarins have a slightly bitter taste in their aglycone form [22].
In addition, Winstel et al. [22] monitored the evolution of scopoletin, umbelliferone, 4-methylumbelliferone, esculetin, coumarin and fraxetin in ten vintages of Cognac (between 1970 and 2015) from the same producer. The wine spirits were aged in 350 L used barrels made from oak wood, with five replicates. The outcomes show that scopoletin and esculetin had higher concentrations in the oldest spirits (338 and 264 μg/L, respectively). Compared with the aforementioned previous work [37], the opposite behaviour found for scopoletin during ageing might be related to the ageing conditions and/or to the analytical method used. The coumarin concentration also tended to increase over time but with some fluctuation. Regarding fraxetin, the evolution pattern was similar to a bell-shaped curve; lower concentrations were observed in the 2015 and 1973 samples (19 μg/L and 118 μg/L, respectively), and the highest concentration was found in the 1995 sample (204 μg/L). A similar trend was registered for 4-methylumbelliferone, which exhibited lower concentrations in 2015 (69 μg/L) and in 1973 (27 μg/L), and the highest one in 2008 (307 μg/L). Their behaviour in older spirits has been reported as a possible result of degradation, but differences in ageing practices and those associated with barrel suppliers over time have led the authors to consider the need for further studies to confirm it. Umbelliferone was only quantified, at low concentrations (varying between 5.5 and 67.8 μg/L), in four old vintages (1995, 1993, 1990, and 1973), which was assigned to its low level in the oak wood and to its slow extraction kinetics.
6.4. Ageing technology
Traditionally, the spirit beverages are aged in wooden barrels for several months or years, in consonance with the producing region regulation as above mentioned. Although the high quality of the final products obtained through this technology, it presents the following drawbacks: i) time-consuming; ii) costly; iii) low production efficiency and profitability for the producer; iv) substantial loss of the spirit beverage by evaporation, which is more pronounced in tropical areas such as those related to sugar cane spirits production; v) significant demand for wood, which is a limited-supply natural resource [23,24,44,215]. Several research teams have therefore been looking for alternative technologies towards more sustainable production processes; the most widely exploited consists of using stainless steel tanks with wood pieces inside and micro-oxygenation as an option [28,30,41,42,69,114,216,217,218,219,220]. In these studies, the assessment of the technologies' performance has been made by analysing the physicochemical characteristics (basic chemical parameters, volatile compounds, phenolic compounds, mineral elements) and sensory properties acquired by the distillates during ageing. Among the phenolics, phenolic acids, phenolic aldehydes and tannins have been often scrutinised, which has not been the case with coumarins. The results available for coumarins are presented below.
Oliveira-Alves et al. [42] compared the scopoletin and umbelliferone contents in wine spirits obtained from the same Lourinhã distillate aged for 12 months in 250 L barrels of chestnut wood (B), in 50 L demijohns with staves of the same wood combined with three micro-oxygenation modalities (oxygen was applied with a flow rate of 2 mL/L/month during 15 days - O15, 30 days - O30, and 60 days - O60, followed by 0.6 mL/L/month until the end of the trial) and one control modality using nitrogen (N), with two replicates. The outcomes are shown in Figure 19.
The wine spirits aged through the alternative technology, regardless of the micro-oxygenation modality, were significantly richer in umbelliferone than those aged by the traditional one and the control. On the contrary, scopoletin level was significantly higher in the wine spirits from the barrels and the control. The oxygen applied (alternative technology) seems to have contributed to the balance between additive and subtractive phenomena involving umbelliferone. As for scopoletin, the outcomes suggest that micro-oxygenation contributed to shifting this balance towards the subtractive phenomena, thus decreasing its content in the corresponding wine spirits.
These results corroborate those of a previous project [41,165] in which a Lourinhã distillate was aged in 250 L barrels and in 1000 L stainless steel tanks with wood staves inside combined with a single micro-oxygenation flow rate (2 mL/L/month) for 18 months. It should be stressed that they suggest a different sensitivity of these coumarins to oxidation, since the direct supply of oxygen through micro-oxygenation favours oxidation reactions involving phenolic compounds [28]: scopoletin seems to have enhanced susceptibility to oxidation compared to umbelliferone.
7. The role of coumarins in the nutraceutical quality of spirit beverages
Besides the contribution of coumarins to the sensory properties of spirit beverages according to the ageing conditions, they are involved in the nutraceutical component of these beverages' quality. Nutraceutical quality is related to the health benefits of foods and beverages [49,221,222,223]. Indeed, moderate consumption of alcoholic beverages as part of a healthy lifestyle has been associated with lower cardiovascular risk compared to abstinence or heavy drinking [224]. The health benefits of moderate consumption of aged spirits can be assigned to their phenolic composition, which has remarkable biological functions, such as free radical scavenging, inhibiting lipid peroxidation, and reducing platelet aggregation and thrombosis [225], in opposition to the ethanol-induced damage [6].
Concerning specifically the spirit beverages covered by this review, evidence exists on the relationship between the in vitro antioxidant properties and lignin-derived phenolic compounds extracted from wooden barrels to wine spirits [31,53,54,55,226], brandy [54,56], whiskey [52] and rum [57]. In addition, consumption of whiskey (100 mL/day) transiently increases the total phenolic content and enhances the plasma antioxidant capacity, thus potentially lowering the risk of coronary heart disease [227]. The work of Suzuki et al. [228] revealed that compounds extracted from wooden barrels during the ageing of whiskey increased the cytoprotective protein, the enzyme heme oxygenase-1 (HO-1), in human endothelial cells, demonstrating that upregulation of HO-1 protein level might possibly contribute to the maintenance of blood vessel function.
The health effects of phenolic compounds, and particularly those found in aged spirit beverages, depend on their chemical composition, their potential interactions with ethanol and the presence of other bioactive compounds [8]. Among them, phenolic aldehydes (such as sinapaldehyde, coniferaldehyde and vanillin), phenolic acids (such as vanillic acid and ellagic acid) and coumarins (such as esculetin) identified in whiskey play an important role as antioxidants, and may partly contribute to the protection of blood vessels by triggering the activation of HO-1 gene [228,229,230]. Similar effects have been reported for wine spirit by Duriez et al. [231] and Umar et al. [232].
Coumarins are nutraceutical compounds due to their ability to exert non-covalent interactions with protein structures and free radical scavenging activity [233,234]; in general, they scavenge reactive oxygen species (ROS) via hydrogen atom transfer mechanism or electron transfer to peroxyl radical [235]. The health effects of simple coumarins, the most plentiful in wood and aged spirit beverages (as specified in Section 4 and Section 6), have been thoroughly investigated in vitro and in vivo studies, as shown in Table 8.
Apart from the activities identified, the high bioavailability of coumarins [117] is an essential feature to guarantee their biological effectiveness. Actually, the oral intake does not result in complete uptake of bioactive compounds into the gastrointestinal tract, and a certain percentage is not absorbed [223]. Bioavailability includes bioaccessibility and bioactivity: i) bioaccessibility corresponds to the fraction of a compound released from the food matrix during digestion process in the luminal content, being accessible for absorption in the small intestine or biotransformed by the gut microbiota; ii) bioactivity contemplates the phenomena involving the absorbed compounds or their metabolites in the target tissue, resulting in biological activity on the body [275,276]. However, in the light of current knowledge, compound concentrations cannot usually be determined directly at the site of action or target tissue. Thus, bioavailability assessment consists of determining the concentration of the bioactive compound in the blood (how fast and how much of a compound appears in the blood after a specific dose is administered) or urine [277].
Interestingly, strategies have been widely investigated to enhance the bioavailability and transport of pharmaceuticals by conjugating with coumarins. In a pharmacological study, bioavailability evaluation of the prodrug system showed that coumarin-based prodrug meptazinol produced a fourfold increase in oral bioavailability over the parent drug meptazinol in rats [278]. The pharmaceutical sertraline, an antidepressant drug, replaced the chloride anion for coumarin 3-carboxylate (bioactive compound), revealing that its antidepressant action was improved compared to the native drug [279]. Coumarins can also be used to minimise the permeability glycoprotein-mediated efflux (P-glycoprotein) and enhance bioavailability of some drugs, such as placlitaxel (anticancer drug), which inhibits breast cancer stem cell growth [280]. The P-glycoprotein is known to cause multidrug resistance phenotype in cancer cells, being a major obstacle in cancer treatment [281]. In an experimental study, the introduction of a sulfamide moiety into coumarin derivatives provided greater RAF/MEK (rapidly accelerated fibrosarcoma/mitogen-activated protein kinase) inhibitory activity concomitantly with an acceptable pharmacokinetic profile (51% bioavailability in mouse) [282].
In summary, coumarins are one of the most important groups of natural compounds with diverse pharmacological properties, and therefore they have been extensively studied in the field of medicinal chemistry and as therapeutic agents. However, studies involving beverages, and especially spirit beverages, have received less attention.
8. Do coumarins have impact on food safety?
As aforementioned, simple coumarins are the most plentiful in wood and aged spirits. The majority of these coumarins are considered not hazardous to human health at the concentrations detected in beverages and edible plants [283]. Nevertheless, studies dealing with higher dosage of coumarin extracted from natural sources revealed chronic toxic effects, particularly hepatotoxicity [234]. As a result, it is necessary to quantify coumarins, specifically the coumarin, in spirit beverages using precise and accurate analytical methods (described in Section 5).
Clinical studies have shown that coumarin is probably not a carcinogen to humans, which have led to its classification as a Group 3 chemical with only limited toxicity [284]. Previously, coumarin was used in flavours and as a chrome plating brightener until 1954, when the FDA classified it as a carcinogen and banned its use in foodstuffs [284]. In 1956, in vivo studies revealed that the coumarin incorporated in the diet of rats and dogs caused the initial damage to their liver tissue [285]. Contrary to these findings, the in vivo studies for mutagenic and genotoxic potential suggest that coumarin is not a genotoxic agent, because the dose–response relationships for coumarin-induced toxicity and carcinogenicity are non-linear, with tumour formation occurring only at high doses (> 280 mg/kg/day) which are associated with pulmonary and hepatic toxicity [112,286].
Although the majority of coumarins are not intrinsically toxic, with the exception of coumarin, some derivatives and higher doses may raise concerns related to food safety. Regarding data on food toxicology, which is focused on the study of compounds present in food that can potentially cause adverse effects on consumer health, the Chemical Safety Data Sheets (SDS) and PubChem data available confirm that the most common coumarins found in wood raw material and aged spirit beverages have little harmful effect on human beings (Table 9).
Table 16 have a notable impact, and even minor differences in structure lead to different actions and varied affinities for biological targets: hydroxyl, methoxy, and glycosidic groups in various positions on the coumarin skeleton. Certain structural modifications during metabolic degradation can result in more toxic metabolites, the reason why it is critical to identify and understand the metabolites that can be generated in the human system. Umbelliferone, for example, is a coumarin degradation metabolite that is also a known human metabolite of 7-methoxycoumarin and coumarin.
Specific tissues or organs may be highly susceptible to the impacts of particular substances due to their physiological activities, high metabolic activity, or exposure. As in the instance of coumarin, which is predominantly metabolised in the liver through cytochrome P-450. In phase I, metabolic events such as oxidation, reduction, and hydrolysis are used to modify functional groups in order to prepare the substrate for subsequent conjugation reactions in phase II. The cytochrome P-450 enzyme in the liver hydroxylates coumarin and similar compounds [298], and the most prevalent hydroxylation pathways happens at 3 and 7-positions to produce 3-hydroxycoumarin (3-OHC) and 7-hydroxycoumarin (7-OHC), respectively and for coumarin [299,300]. The presence of a significant amount of 3-OHC is thought to facilitate the formation of the cytotoxic byproduct o-HPA (Figure 15), which could potentially contribute to or be accountable for coumarin-induced toxicity.
Coumarin demands extra attention when compared to other naturally occurring coumarins, not only because its lower LD50 (more toxic), but also because it is commonly found in cinnamon-containing food and drinks, which is not applicable to the spirit beverages addressed in this review. It is noteworthy that the influence on food safety mainly depends on the coumarins concentrations in specific food products and their compliance with regulatory criteria than to the presence of coumarins in general. To ensure food safety, as with any food component, moderation is key, and it is advised to follow dietary guidelines and limits set by local authorities for this purpose.
9. Coumarins and spirit beverages’ authenticity
Assuring authenticity is a pivotal aspect in spirit beverages value chains, and has therefore been a major concern and challenge for all their players, including the regulatory/governmental agencies and researchers worldwide. It allows protecting producers, producing regions and countries, as well as consumers, from inauthentic spirit beverages. This type of beverages has become a target for adulteration and fraud through practices that include supplementing with ingredients or aromas to boost the product at a lower price, and adding of non-specified additives to increase volume [58]. According to Popping [301], there are two types of inauthenticity events: i) misrepresentation that a beverage is within the contractual arrangement between trading partners, namely breach of contract; ii) misrepresentation that a beverage is within the legal obligations of the region of intended trade, namely noncompliance (Figure 20).
The global market and the growing variability and availability of alcoholic beverages from different countries have made consumers increasingly aware of the importance of consuming certified products (in terms of quality and food safety) [302]. In this regard, Europe has taken the lead position by creating, in 1992, a robust system of geographical indications to protect the name of a product originating from a specific region and resulting from a traditional production process. Currently, this regime comprises: Protected Designations of Origin - in which all production stages must take place; Protected Geographical Indications (PGI) - at least one of the production, processing or preparation phases must take place in the region. Concerning the spirit beverages, the rules are set by Regulation (EU) [1]. Other producing countries, such as the USA, Canada and Brazil also have specific regulations for spirit drinks.
For this purpose, more advanced analytical methods, as well as the identification of compounds that act as chemical markers, are required for the certification of spirit drinks and the strengthening of legislation. Several techniques, such as spectrophotometry [303,304,305], synchronous fluorescence spectrometry [306], GC [307,308,309], HPLC [310], capillary electrophoresis [311], nuclear magnetic resonance (NMR) spectroscopy [312,313,314], and electrospray ionisation mass spectrometry (EIS-MS) [315,316,317] have been used to determine the indicators of authenticity in the spirit beverages, such as alcohol strength [318], sugars [315], volatile compounds [319], colouring compounds [318] and wood-derived compounds [320].
Regarding the wood-derived compounds of the spirit beverages under analysis, resulting from the ageing stage of the corresponding distillates, some studies pointed out the role of coumarins, phenolic aldehydes and phenolic acids as chemical markers of wine spirits, brandies, whiskeys, rums and cachaças [41,167,192,306,311,321]. Among coumarins, scopoletin has been considered as a possible marker of ageing in oak barrels [19,36,322].
Furthermore, coumarins contribute to antioxidant activity reported among alcoholic beverages, which is also thought to be a marker of authenticity. Ziyatdinova et al. [307], studying the adulteration of brandies, reported that the antioxidant activity of authentic brandies was 15-fold higher than that of adulterated ones. Hence, the literature suggests that coumarins can be used as authenticity markers of aged spirits, but further research is needed to provide robust outcomes for quality assessment and authenticity control.
10. Concluding Remarks and Perspectives
A comprehensive overview of the state-of-the-art and contemporary approach on coumarins in spirit beverages is presented for the first time. A connection between these findings and crucial aspects of everyday life, particularly in terms of authenticity and food safety concerns, was established. Six renowned types of spirit beverages were chosen (wine spirit, brandy, whiskey, rum, cachaça, and grape marc spirit) due to their enhancement with wood compounds during the ageing process, and were thoroughly discussed. The ageing process improves the quality of the distillate by enhancing its sensory and nutraceutical properties, in which coumarins are involved. Given the dynamic interplay between additive and subtractive processes, the chemical composition of aged spirits undergoes significant changes over time. Consequently, when taking into account differentiated products, consumer health concerns, food science, and food safety in the market's largest segment of the global alcoholic drink market, comparing the effects of different ageing time, wood sources, heat treatment, and ageing technology on specific phenolic composition of spirit beverages is worthwhile. A balance of its phenolic composition and certain coumarins, can play a major role in developing value chains, meeting consumer preferences, and driving market growth.
In order to understand the occurrence of coumarins in the addressed spirit beverages, it was considered essential to elucidate their production processes and the type of wood used in the ageing stage. Furthermore, significant information revealing that different kinds of wood provide considerably different coumarins contents have been collected in this review. Nevertheless, only simple coumarins have been identified in wood and aged spirits: coumarin, scopoletin, umbelliferone, 4-methylumbelliferone, aesculin, aesculetin and fraxetin. Among them, scopoletin was the most plentiful in the majority of wood species, while umbelliferone was more abundant in American oak wood. Though, the concentrations of coumarins in aged beverages are significantly lower in comparison to those reported from wood sources.
This review further looked at coumarin metabolic pathways for nutraceutical and safety purposes. As a prime instance, it was clarified that umbelliferone is metabolised into coumarin, which is then metabolised into 7-hydroxycoumarin and eliminated by the human system, and that all these metabolites exhibit interactions and increased biological activities. Therefore, further research is needed for a comprehensive insight into the metabolites generated after coumarins ingestion. Although the majority of coumarins are generally non-toxic, except coumarin itself, some derivatives and higher amounts may trigger food safety issues. Coumarins' significant bioavailability is an important feature that ensures their biological efficacy; thus, detailed information about how the gastrointestinal system absorbs these compounds and what happens to the other portion of bioactive substances from oral ingestion is required.
A growing demand for more sensitive, selective, and efficient procedures has driven improvements in analytical techniques for molecular structural elucidation, and some of the cutting-edge analytical methods being used for other organic substrates will likely influence the identification of novel coumarins in wood extracts and spirit beverages. Among these novel approaches that are becoming more accessible, democratised, and disseminated in the scientific community, this review highlights Tandem Mass Spectrometry (MS/MS or MSn), Ion Mobility Spectrometry (IMS), liquid chromatography with nuclear magnetic resonance (LC-NMR), as well as computational techniques that may be very useful, along with the use of advanced data processing techniques such as Data Mining and Cheminformatics, along with machine learning algorithms. The authenticity of spirit beverages is of the utmost importance for consumers, producers, governmental authority, and the agrifood sector, the reason why the role of coumarins in this topic was contextualised. Despite their low concentrations, the results support scopoletin and umbelliferone as chemical markers for ensuring authenticity of aged spirit beverages, which has been a major concern and challenge for all players and researchers worldwide. Given the broad biological function exhibited by the coumarins chemical family, further research will be necessary in this matter.
Author Contributions
Conceptualization, S.C., S.A., S.L. and T.A.F.; investigation, S.A., S.L., T.A.F. and S.C.; writing—original draft preparation, S.A., S.L., T.A.F. and S.C.; writing—review and editing, S.C., T.A.F., S.A. and S.L.; supervision, S.C.; funding acquisition, S.A., T.A.F. and S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Foundation for Science and Technology (FCT) through the projects UIDB/00239/2020 [CEF], UIDP/00100/2020 [CQE], UIDB/00100/2020 [CQE], contract CEECIND/02725/2018, and UIDB/05183/2020 [MED].
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Reg. EU nº 2019/787 of the European Parliament and of the Council of 17 April 2019 on the Definition, Description, Presentation and Labelling of Spirit Drinks, the Use of the Names of Spirit Drinks in the Presentation and Labelling of other Foodstuffs, the Protection of Geographical Indications for Spirit Drinks, the Use of Ethyl Alcohol and Distillates of Agricultural Origin in Alcoholic Beverages, and Repealing Regulation (EC) No 110/2008; OJEU. L130 European Union. 2019, pp. 1–54. Available online: https://eur-lex.europa.eu/eli/reg/2019/787/2022-08-15 (accessed on 13 September 2023).
- Tsakiris, A.; Kallithrakab, S.; Kourkoutasc, Y. Grape brandy production, composition and sensory evaluation. J. Sci. Food Agric. 2014, 94, 404–414. [Google Scholar] [CrossRef]
- Krstić, J.D.; Kostić-Stanković, M.M.; Veljović, S.P. Traditional and innovative aging technologies of distilled beverages: The influence on the quality and consumer preferences of aged spirit drinks. J. Agric. Sci. 2021, 66, 209–230. [Google Scholar] [CrossRef]
- Statista. Alcoholic Drinks - Worldwide 2023. Available online: https://www.statista.com/outlook/cmo/alcoholic-drinks/worldwide (accessed on 13 September 2023).
- Alcoholic Beverages Global Market Analysis, Insights and Forecast, 2018-2029. Fortune Business Insights 2023. Report FBI 107439. Available online: www.fortunebusinessinsights.com/alcoholic-beverages-market-107439 (accessed on 13 September 2023).
- World Health Organization (WHO). Global status report on alcohol and health 2018. Available online: https://www.who.int/publications/i/item/9789241565639 (accessed on 13 September 2023).
- Wardencki, W. Alcoholic Beverages. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering. Encyclopedia of Analytical Science, 3rd ed.; Worsfold, P., Poole, C., Townshend, A., Miró, M., Eds.; Academic Press: Cambridge, USA, 2019; pp. 67–76. [Google Scholar] [CrossRef]
- Kang, Q.; Sun, J.; Wang, B.; Sun, B. Wine, beer and Chinese Baijiu in relation to cardiovascular health: the impact of moderate drinking, Food Sci. Hum. Wellness. 2023, 12(1), 1–13. [Google Scholar] [CrossRef]
- Cravero, 2020 Cravero, M.C.; Laureati, M.; Spinelli, S.; Bonello, F.; Monteleone, E.; Proserpio, C.; Lottero, M.R.; Pagliarini, E.; Dinnella, C. Profiling individual differences in alcoholic beverage preference and consumption: New insights from a large-scale study. Foods 2020, 9(8), 1131. [CrossRef]
- Lin, M.; Yang, B.; Dai, M.; Xu, Y.; Li, X.; Sun, B. East meets west in alcoholic beverages: Flavor comparison, microbial metabolism and health effects. Food Biosci. 2023, 56, 103385. [Google Scholar] [CrossRef]
- Pino, J.; Martí, M.P.; Mestres, M.; Pérez, J.; Busto, O.; Guasch, J. Headspace solid-phase microextraction of higher fatty acid ethyl esters in white rum aroma. J. Chromat. A. 2002, 954, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Faria, B.; Loyola, E.; López, M.G.; Dufour, J.-P. Cachaça, Pisco and Tequila. In Fermented Beverage Production; Lea, A. G. H., Geoffrey, A., Lea, H., Piggott, J.R, Eds.; Springer Science and Business Media: New York, U.S.A., 2003; pp. 335–364. [Google Scholar] [CrossRef]
- Nicol, D.A. Rum. In Fermented Beverage Production; Lea, A. G. H., Geoffrey, A., Lea, H., Piggott, J.R, Eds.; Springer Science and Business Media: New York, U.S.A., 2003; pp. 263–287. [Google Scholar] [CrossRef]
- Russell, I. Whisky. Technology, production and marketing; Elsevier: London, UK, 2003. [Google Scholar]
- López-Vásquez, C.; Bollaín, M.H.; Moser, S.; Orriols, I. Characterization and differentiation of monovarietal grape pomace distillate from native varieties of Galicia. J. Agric. Food Chem. 2010, 58, 9657–9665. [Google Scholar] [CrossRef] [PubMed]
- Da Porto, C. Grappa: production, sensory properties and market development. In Alcoholic beverages. Sensory evaluation and consumer research; Piggott, J., Ed.; Woodhead Publishing: Philadelphia, U.S.A., 2012; pp. 299–314. [Google Scholar] [CrossRef]
- Louw, L.; Lambrechts, M.G. Grape-based brandies: production, sensory properties and sensory evaluation. In Alcoholic beverages. Sensory evaluation and consumer research; Piggott, J., Ed.; Woodhead Publishing: Philadelphia, U.S.A., 2012; pp. 281–298. [Google Scholar] [CrossRef]
- Rodríguez-Solana, R.; Salgado, J.M.; Domínguez, J.M.; Cortés-Diéguez, S. First approach to the analytical characterization of barrel-aged grape marc distillates using phenolic compounds and colour parameters. Food Technol. Biotechnol. 2014, 52, 391–402. [Google Scholar] [CrossRef]
- Canas, S. Phenolic composition and related properties of aged wine spirits: Influence of barrel characteristics. A review. Beverages 2017, 3, 55. [Google Scholar] [CrossRef]
- Puentes, C.; Joulia, X.; Vidal, J.-P.; Esteban-Decloux, M. Simulation of spirits distillation for a better understanding of volatile aroma compounds behavior: Application to Armagnac production. Food Bioprod. Process. 2018, 112, 31–62. [Google Scholar] [CrossRef]
- García-Moreno, M.V.; Sánchez-Guillén, M.M.; Ruiz de Mier, M.; Delgado-Gonzaléz, M.J.; Rodríguez-Dodero, M.C.; García-Barroso, C.; Guillén-Sánchez, D.A. Use of alternative wood for the ageing of Brandy de Jerez. Foods 2020, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Winstel, D.; Gautier, E.; Marchal, A. Role of oak coumarins in the taste of wines and spirits: Identification, quantitation, and sensory contribution through perceptive interactions. J. Agric. Food Chem. 2020, 68(28), 7434–7443. [Google Scholar] [CrossRef] [PubMed]
- Gollihue, J.; Pook, V.G.; DeBolt, S. Sources of variation in bourbon whiskey barrels: a review. J. Inst. Brew. 2021, 127, 210–223. [Google Scholar] [CrossRef]
- Canas, S.; Caldeira, I.; Fernandes, T.A.; Anjos, A.; Belchior, A.P.; Catarino, S. Sustainable use of wood in wine spirit production. In Improving Sustainable Viticulture and Winemaking Practices; Costa, J.M., Catarino, S., Escalona, J.M., Piergiorgio, C., Eds.; Elsevier: London, UK, 2022a; pp. 259–280. [Google Scholar]
- de Aquino, F.W.; Rodrigues, S.; Nascimento, R.F.; Casemiro, A.R.S. Simultaneous determination of aging markers in sugar cane spirits. Food Chem. 2006, 98, 569–574. [Google Scholar] [CrossRef]
- Fujieda, M.; Tanaka, T.; Suwa, Y.; Koshimizu, S.; Kouno, I. Isolation and structure of whiskey polyphenols produced by oxidation of oak wood ellagitannins. J. Agric. Food Chem. 2008, 56, 7305–7310. [Google Scholar] [CrossRef]
- Pino, J.A.; Tolle, S.; Gök, R.; Winterhalter, P. Characterisation of odour-active compounds in aged rum. Food Chem. 2012, 132, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Canas, S.; Danalache, F.; Anjos, O.; Fernandes, T.A.; Caldeira, I.; Santos, N.; Fargeton, L.; Boissier, B.; Catarino, S. Behaviour of low molecular weight compounds, iron and copper of wine spirit aged with chestnut staves under different levels of microoxygenation. Molecules 2020, 25, 5266. [Google Scholar] [CrossRef]
- Bortoletto, A.M.; Silvello, G.C.; Alcarde, A.R. Aromatic profiling of flavor active compounds in sugarcane spirits aged in tropical wooden barrels. Braz. J. Food Technol. 2021, 24, e2019071. [Google Scholar] [CrossRef]
- Rodríguez-Solana, R.; Rodríguez-Freigedo, S.; Salgado, J.M.; Domínguez, J.M.; Cortés-Diéguez, S. Optimisation of accelerated ageing of grape marc distillate on a micro-scale process using a Box–Benhken design: influence of oak origin, fragment size and toast level on the composition of the final product. Aust. J. Grape Wine Res. 2017, 23, 5–14. [Google Scholar] [CrossRef]
- Oliveira-Alves, S.; Lourenço, S.; Anjos, O.; Fernandes, T.A.; Caldeira, I.; Catarino, S.; Canas, S. Influence of the storage in bottle on the antioxidant activities and related chemical characteristics of wine spirits aged with chestnut staves and micro-oxygenation. Molecules 2022, 27, 106. [Google Scholar] [CrossRef]
- Baldwin, S.; Black, R.A.; Andreasen, A.A.; Adams, S.L. Aromatic congener formation in maturation of alcoholic distillates. J. Agr. Food Chem. 1967, 15, 381–385. [Google Scholar] [CrossRef]
- Bricout, J. Analyse de quelques constituants dérivés du chêne dans les vieilles eaux-de-vie d'armagnac. Ann. Technol. Agric. 1971, 20, 217–224. [Google Scholar]
- Joseph, E.; Marche, M. Contribution à l'étude du vieillissement du cognac. Identification de la scopolétine, del'aesculétine, de l'ombelliférone, de la b-méthyl-ombelliférone, de l'aesculine et de la scopoline, hétérosides provenant du bois. Conn. Vigne Vin 1972, 6, 1–58. [Google Scholar] [CrossRef]
- Otsuka, K.I.; Zenibayashi, J. On the Determination of Scopoletin in Aged Distilled Liquors. Agric. Biol. Chem. 1974, 38, 1079–1080. [Google Scholar] [CrossRef]
- Salagoity-Auguste, M.-H.; Tricard, C.; Sudraud, P. Dosage simultané des aldéhydes aromatiques et des coumarines par chromatographie liquide haute performance. J. Chromat. 1987, 392, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Tricard, C.; Salagoity, M.H.; Sudraud, P. La scopolétine: un marqueur de la conservation en fûts de chêne. Conn. Vigne Vin 1987, 21(1), 33–41. [Google Scholar] [CrossRef]
- Puech, J.-L.; Moutounet, M. Liquid chromatographic determination of scopoletin in hydroalcoholic extract of oak wood and in matured distilled alcoholic beverages. J. Assoc. Off. Anal. Chem. 1988, 71(3), 512–514. [Google Scholar] [CrossRef] [PubMed]
- Patrício, I.; Canas, S.; Belchior, A.P. Effect of brandies’ agitation on the kinetics of extraction/oxidation and diffusion of wood extractable compounds in experimental model. Ciência Téc. Vitiv. 2005, 20, 1–15. [Google Scholar]
- Canas, S.; Silva, V.; Belchior, A.P. Wood related chemical markers of aged wine brandies. Ciência Téc. Vitiv. 2008a, 23, 45–52. [Google Scholar]
- Canas, S.; Anjos, O.; Caldeira, I.; Belchior, A.P. Phenolic profile and colour acquired by the wine spirit in the beginning of ageing: Alternative technology using microoxygenation vs traditional technology. LWT – Food Sci. Technol. 2019, 111, 260–269. [Google Scholar] [CrossRef]
- Oliveira-Alves S.C., Lourenço S., Fernandes T.A., Anjos O., Caldeira I., Canas S., Catarino S. Quantification of coumarins in wine spirit aged by different technologies using chestnut wood. In Livro de resumos do XV Encontro de Química dos Alimentos: Estratégias para a Excelência, Autenticidade, Segurança e Sustentabilidade Alimentar, Funchal, Portugal, 5-8 September 2021 (07 September 2021). 8 September.
- da Silva, A.A.; Nascimento, E.S.P.; Cardoso, D.R.; Franco, D.W. Coumarins and phenolic fingerprints of oak and Brazilian woods extracted by sugarcane spirit. J. Sep. Sci. 2009, 32, 3681–3691. [Google Scholar] [CrossRef] [PubMed]
- Santiago, W.D.; Cardoso, M.G.; Nelson, D.L. Cachaça stored in casks newly constructed of oak (Quercus sp.), amburana (Amburana cearensis), jatoba (Hymenaeae carbouril), balsam (Myroxylon peruiferum) and peroba (Paratecoma peroba): alcohol content, phenol composition, colour intensity and dry extract. J. Inst. Brew. 2017, 123, 232–241. [Google Scholar] [CrossRef]
- Lima, C.M.G.; Benoso, P.; Pierezan, M.D.; Santana, R.F.; de Souza Hassemer, G.; da Rocha, R.A.; Nora, F.M.D.; Verruck, S.; Caetano, D.; Simal-Gandara, J. A state-of-the-art review of the chemical composition of sugarcane spirits and current advances in quality control. J. Food Compos. Anal. 2022, 106, 104338. [Google Scholar] [CrossRef]
- Mignani, A.G.; Ciaccheri, L.; Gordillo, B.; Mencaglia, A.A.; González-Miret, M.L.; Heredia, F.J.; Culshaw, B. Identifying the production region of single-malt Scotch whiskies using optical spectroscopy and pattern recognition techniques. Sensor. Actuat. B Chem. 2012, 171-172, 458–462. [Google Scholar] [CrossRef]
- Collins, T.S.; Zweigenbaum, J.; Ebeler, S.E. Profiling of nonvolatiles in whiskeys using ultra high pressure liquid chromatography quadrupole time-of-flight mass spectrometry (UHPLC–QTOF MS). Food Chem. 2014, 163, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Sádecká, J.; Uríčková, V.; Májek, P.; Jakubíková, M. Comparison of different fluorescence techniques in brandy classification by region of production. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 216, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. A practical guide for designing effective nutraceutical combinations in the form of foods, beverages, and dietary supplements against chronic degenerative diseases. Trends Food Sci Technol. 2019, 88, 179–193. [Google Scholar] [CrossRef]
- Li, L.; Sun, B. Grape and wine polymeric polyphenols: Their importance in enology. Crit Rev Food Sci Nutr 2017, 59, 563–579. [Google Scholar] [CrossRef]
- Romanet, R.; Sarhane, Z.; Bahut, F.; Uhl, J.; Schmitt-Kopplin, P.; Nikolantonaki, M.; Gougeon, R.D. Exploring the chemical space of white wine antioxidant capacity: A combined DPPH, EPR and FT-ICR-MS study. Food Chem. 2021, 355, 129566. [Google Scholar] [CrossRef]
- Koga, K.; Taguchi, A.; Koshimizu, S.; Suwa, Y.; Yamada, Y.; Shirasaka, N.; Yoshizumi, H. Reactive oxygen scavenging activity of matured whiskey and its active polyphenols. J. Food Sci. 2007, 72, S212–S217. [Google Scholar] [CrossRef]
- Canas, S.; Casanova, V.; Belchior, A.P. Antioxidant activity and phenolic content of Portuguese wine aged brandies. J. Food Compos. Anal. 2008b, 21, 626–633. [Google Scholar] [CrossRef]
- Ziyatdinova, G.; Salikhova, I.; Budnikov, H. Chronoamperometric estimation of cognac and brandy antioxidant capacity using MWNT modified glassy carbon electrode. Talanta 2014, 125, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Nocera, A.; Ricardo-da-Silva, J.M.; Canas, S. Antioxidant activity and phenolic composition of wine spirit resulting from an alternative ageing technology using microoxygenation: a preliminary study. Oeno One 2020, 54, 485–496. [Google Scholar] [CrossRef]
- Schwarz, M.; Rodríguez, M.C.; Martínez, C.; Bosquet, V.; Guillén, D.; Barroso, C.G. Antioxidant activity of Brandy de Jerez and other distillates, and correlation with their polyphenolic content. Food Chem. 2009, 116, 29–33. [Google Scholar] [CrossRef]
- Regalado, E.L.; Tolle, S.; Pino, J.A.; Winterhalter, P.; Menendez, R.; Morales, A.R.; Rodriguez, J.L. Isolation and identification of phenolic compounds from rum aged in oak barrels by highspeed countercurrent chromatography/high performance liquid chromatography-diode array detection-electrospray ionization mass spectrometry and screening for antioxidant activity. J. Chromatogr. A 2019, 41, 7358–7364. [Google Scholar] [CrossRef]
- Kamiloglu, S. Authenticity and traceability in beverages. Food Chem. 2019, 277, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Sikorska, E. Fluorescence spectroscopy and chemometrics in analysis of beverages. In Quality Control in the Beverage Industry; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier: London, UK, 2019; Volume 17: The Science of Beverages, pp. 161–203. [Google Scholar] [CrossRef]
- Belchior, A.P.; Canas, S.; Caldeira, I.; Carvalho, E. Aguardentes vinícolas - Tecnologias de produção e envelhecimento. Controlo de qualidade; Publindústria, Edições Técnicas: Porto, Portugal, 2015; pp. 1–181. [Google Scholar]
- Cantagrel, R.; Galy, B. From Vine to Cognac. In Fermented Beverage Production; Lea, A. G. H., Geoffrey, A., Lea, H., Piggott, J.R, Eds.; Springer Science and Business Media: New York, U.S.A., 2003; pp. 195–212. [Google Scholar] [CrossRef]
- Lurton, L.; Ferrari, G.; Snakkers, G. Cognac: production and aromatic characteristics. In Alcoholic beverages. Sensory evaluation and consumer research; Piggott, J., Ed.; Woodhead Publishing: Philadelphia, U.S.A., 2012; pp. 242–266. [Google Scholar] [CrossRef]
- Bertrand, A. Armagnac and Wine-Spirit. In Fermented Beverage Production; Lea, A. G. H., Geoffrey, A., Lea, H., Piggott, J.R, Eds.; Springer Science and Business Media: New York, U.S.A., 2003; pp. 213–238. [Google Scholar] [CrossRef]
- Décret nº 2009-1146 du 21 septembre 2009 relatif à l’appellation d’origine contrôlée “Cognac” ou ”Eau-de-vie de Cognac” ou “Eau-de-vie des Charentes”; JOFR nº 0221 du 24 septembre 2009, pp 15619, text nº 25, Available online:. Available online: https://www.legifrance.gouv.fr/jorf/id/JORFTEXT000021065172 (accessed on 17 October 2023).
- Decreto-Lei nº 323/94, de 29 de dezembro, do Ministério da Agricultura, Estatuto da Região Demarcada das Aguardentes Vínicas da Lourinhã; Diário da República nº 300/1994, Série I-A, 7486-7489. Available online: https://diariodarepublica.pt/dr/detalhe/decreto-lei/323-1994-322096 (accessed on 17 October 2023).
- Décret nº 2009-1285 du 23 octobre relatif aux appellations d’origine contrôlées “Armagnac”, ”Blanche Armagnac”, “Bas Armagnac”, “Haut Armagnac”et “Armagnac Ténarèze”; NOR : AGRT0912388D version consolidée au 25 octobre 2009. Available online: https://www.legifrance.gouv.fr/loda/id/JORFTEXT000021190654 (accessed on 17 October 2023).
- Bamforth, C.W. Fermented Beverages. In Encyclopedia of Agriculture and Food Systems. Alfen, N.K.V., Eds.; Elsevier: London, UK, 2014; pp. 124–136. [Google Scholar] [CrossRef]
- Schwarz, M.; Rodríguez, M.C.; Sanchez, M; Guillen, D.A.; Barroso, C.G. Development of an accelerated aging method for Brandy. LWT - Food Sci. 2014, 59(1), 108–114. [CrossRef]
- Schwarz, M.; Rodríguez-Dodero, M.C.; Soledad Jurado, M.; Puertas, B.; Barroso, C.G.; Guillén, D.A. Analytical characterization and sensory analysis of distillates of different varieties of grapes aged by an accelerated method. Foods 2020, 9, 277. [Google Scholar] [CrossRef]
- Durán-Guerrero, E.; Castro, R.; García-Moreno, M.V.; Rodríguez-Dodero, M.C.; Schwarz, M.; Guillén-Sánchez, D. Aroma of Sherry products: A review. Foods 2021, 10, 753. [Google Scholar] [CrossRef]
- Wiśniewska, P.; Dymerski, T.; Wardencki, W.; Namieśnik, J. Chemical composition analysis and authentication of whisky. J. Sci. Food Agric. 2014, 95(11), 2159–2166. [Google Scholar] [CrossRef]
- Piggott, J.R.; Conner, J.M. Whiskies. In Fermented Beverage Production; Lea, A.G.H., Geoffrey, A., Lea, H., Piggott, J.R., Eds.; Springer Science and Business Media: New York, U.S.A., 2003; pp. 239–262. [Google Scholar] [CrossRef]
- Jack, F.R. Whiskies: composition, sensory properties and sensory analysis. In Alcoholic beverages. Sensory evaluation and consumer research; Piggott, J., Ed.; Woodhead Publishing: Philadelphia, U.S.A., 2012; pp. 379–392. [Google Scholar] [CrossRef]
- Authenticated U. S. Government Information GPO. Title 27—Alcohol, Tobacco Products and Firearms. 27 CFR Ch. I (4–1–11 Edition). Available online: https://www.govinfo.gov/content/pkg/CFR-2011-title27-vol1/pdf/CFR-2011-title27-vol1-chapI.pdf (accessed on 20 October 2023).
- Faria, J.B. Sugar cane spirits: cachaça and rum production and sensory properties. In Alcoholic beverages. Sensory evaluation and consumer research; Piggott, J., Ed.; Woodhead Publishing: Philadelphia, U.S.A., 2012; pp. 348–358. [Google Scholar] [CrossRef]
- Decreto Legislativo Regional nº 18/2021/M, Região Autónoma da Madeira, Confirma, define e caracteriza o «Rum da Madeira» e estabelece as regras relativas à sua produção e comercialização; Diário da República nº 145 de 28 de julho de 2021, 1ª Série, 76-83. Available online: https://diariodarepublica.pt/dr/detalhe/decreto-legislativo-regional/18-2021-168475297 (accessed on 24 October 2023).
- Portaria MAPA Nº 539, de 26 de dezembro de 2022, do Ministério de Estado da Agricultura, Pecuária e Abastecimento, Estabelece os padrões de identidade e qualidade da aguardente de cana e da cachaça, DOU - Diário Oficial da União, Edição 243, Seção 1, pp. 12. Available online: https://www.in.gov.br/en/web/dou/-/portaria-mapa-n-539-de-26-de-dezembro-de-2022-453828778 (accessed on 30 October 2023).
- Da Porto, C. Grappa and grape-spirit production. Crit. Rev. Biotechnol. 1998, 18(1), 13–24. [Google Scholar] [CrossRef]
- Muhlack, R.A.; Potumarthi, R.; Jeffery, D.W. Sustainable wineries through waste valorisation: a review of grape marc utilisation for value-added products. Waste Manag. 2017, 72, 99–118. [Google Scholar] [CrossRef]
- Belchior A.P., Carvalho E.C. Factores que condicionam o teor de metanol dos bagaços. Vin. Port. Doc. 1980, 10, 1–9.
- Belchior, A.P. Qualidade e composição química de aguardentes de bagaço. Vin. Port. Doc. 1977, 7, 1–15. [Google Scholar]
- Silva, M.L.; Macedo, A.C.; Malcata, F.X. Review: steam distilled spirits from fermented grape pomace. Food Sci. Tech Int. 2000, 6(4), 285–300. [Google Scholar] [CrossRef]
- Orriols, I.; Cortés, S.M.; Fornos, D. Carcactéristiques des distillats de marc du commerce “Orujo de galicia” d’Espagne. In Les eaux-de-vie traditionelles d’origine viticole. Bertrand, A., Ed.; Lavoisier Tec & Doc: Paris, France, 2008; pp. 173–179. [Google Scholar]
- Reg. EU nº 110/2008 of the European Parliament and of the Council of 15 January 2008 on the on the Definition, Description, Presentation, Labelling and the Protection of Geographical Indications of spirit drinks and repealing Council Regulation (EEC) No 1576/89; OJEU. L39 European Union. 2008, pp. 16–54. Available online: https://eur-lex.europa.eu/eli/reg/2008/110(1)/oj/por (accessed on 30 October 2023).
- Vogel, A. Darstellung von Benzoesaure aus der Tonka-Boline und aus den MeliIoten-oder Steinklee-Blumen. Ann. Phys. 1820, 64, 161–166. [Google Scholar] [CrossRef]
- Stefanachi, A.; Leonetti, F.; Pisani, L.; Catto, M.; Carotti, A. Coumarin: A Natural, Privileged and Versatile Scaffold for Bioactive Compounds. Molecules 2018, 23, 250. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Roleira, F.; Milhazes, N.; Santana, L.; Uriarte, E. Simple coumarins and analogues in medicinal chemistry: occurrence, synthesis and biological activity. Curr. Med. Chem. 2005, 12(8), 887–916. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Roleira, F.; Milhazes, N.; Uriarte, E.; Santana, L. Simple coumarins: Privileged scaffolds in medicinal chemistry. Front. Med. Chem. Biol. Inter. 2009, 4, 23–85. [Google Scholar] [CrossRef]
- Dean, F.M. Naturally occurring coumarins. In Fortschritte der Chemie Organischer Naturstoffe/Progress in the Chemistry of Organic Natural Products/Progrès Dans La Chimie Des Substances Organiques Naturelles. Fortschritte der Chemie Organischer Naturstoffe/Progress in the Chemistry of Organic Natural Products/Progrès Dans La Chimie Des Substances Organiques Naturelles; Zechmeister, L., Ed.; Springer: Wien, Austria, 1952; Volume 9, pp. 225–291. [Google Scholar] [CrossRef]
- Murray, R.D.H.; Mendez, J.; Brown, S.A. The Natural Coumarins: Occurrence, Chemistry and Biochemistry; John Wiley & Sons: New York, USA, 1982. [Google Scholar]
- Murray, R.D.H. Naturally Occurring Plant Coumarins. In Fortschritte der Chemie organischer Naturstoffe / Progress in the Chemistry of Organic Natural Products. Fortschritte der Chemie organischer Naturstoffe / Progress in the Chemistry of Organic Natural Products; Herz, W., Kirby, G.W., Steglich, W., Tamm, C., Eds.; Springer: Wien, Austria, 1991; Volume 58, pp. 83–316. [Google Scholar] [CrossRef]
- Keating, G.J.; O’Kennedy, R. The chemistry and occurrence of coumarins, 1st ed; John Wiley & Sons: West Sussex, England, 1997. [Google Scholar]
- Iranshahi, M.; Askari, M.; Sahebkar, A.; Hadjipavlou-Litina, D. Evaluation of antioxidant, anti-inflammatory and lipoxygenase inhibitory activities of the prenylated coumarin umbelliprenin. DARU 2009, 17(2), 99–103. Available online: https://ikee.lib.auth.gr/record/226183/files/Litina.pdf.
- O’Kennedy, R.; Thornes, R.D. (Eds.) Coumarins: Biology, Applications, and Mode of Action; JohnWiley & Sons: New York, USA, 1997. [Google Scholar]
- Cai, Y.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–88. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.D.H. Naturally Occurring Plant Coumarins. In Fortschritte der Chemie organischer Naturstoffe / Progress in the Chemistry of Organic Natural Products. Fortschritte der Chemie organischer Naturstoffe / Progress in the Chemistry of Organic Natural Products; Herz, W., Kirby, G.W., Moore, R.E., Steglich, W., Tamm, C., Eds.; Springer: Wien, Austria, 1997; Volume 72, pp. 1–119. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on natural coumarin lead compounds for their pharmacological activity. BioMed. Res. Int. 2013, 963248, 1–14. [Google Scholar] [CrossRef]
- Gaudino, E.C.; Tagliapietra, S.; Martina, K.; Palmisanob, G.; Cravotto, G. Recent advances and perspectives in the synthesis of bioactive coumarins. RSC Adv. 2016, 6, 46394–46405. [Google Scholar] [CrossRef]
- Sarker, S.D.; Nahar, L. Progress in the chemistry of naturally occurring coumarins. In Progress in the chemistry of organic natural products; Kinghorn, A., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Springer International Publishing: Cham, Swiss, 2017; vol 106, pp 241–304. [Google Scholar] [CrossRef]
- Zou, Y.; Teng, Y.; Li, J.; Yan, Y. Recent advances in the biosynthesis of coumarin and its derivatives. GreenChE 2023. [Google Scholar] [CrossRef]
- Bruneton, J. Pharmacognosie -Phytochimie Plantes Médicinales. Tecnique et Documentation, 2nd ed.; Lavoisier: Paris, France, 1993. [Google Scholar]
- Kai, K.; Mizutani, M.; Kawamura, N.; Yamamoto, R.; Tamai, M.; Yamaguchi, H.; Skata, K.; Shimizu, B. Scopoletin is biosynthesized via ortho-hydroxylation of feruloyl CoA by a 2-oxolutarate-dependent dioxygenase in Arabidopsis thaliana. Plant J. 2008, 55, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Shimizu, B.; Mizutani, M.; Watanabe, K.; Sakata, K. Accumulation of coumarins in Arabidopsis thaliana. Phytochemistry 2006, 67, 379–86. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Hehn, A.; Larbat, R.; Doerper, S.; Gontier, E.; Kellner, S.; Matern, U. Biosynthesis of coumarins in plants: A major pathway still to be unravelled for cytochrome P450 enzymes. Phytochem. Rev. 2006, 5, 293–308. [Google Scholar] [CrossRef]
- Hussain, H.; Hussain, J.; Al-Harrasi, A.; Krohn, K. The chemistry and biology of bicoumarins. Tetrahedron 2012, 68(12), 2553–2578. [Google Scholar] [CrossRef]
- Basa, S.C. Natural bicoumarins. Phylochemistry 1988, 27(7), 1933–1941. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; John Wiley & Sons Ltd: West Sussex, England, 2002. [Google Scholar] [CrossRef]
- Hussain, M.I.; Qamar Abbas, S.; Reigosa, M.J. Activities and novel applications of secondary metabolite coumarins. Planta Daninha 2018, 36, e018174040. [Google Scholar] [CrossRef]
- Arndt, F.; Loewe, L.; Ün, R.; Ayça, E. Cumarindiol und Cumarin-Chromon Tautomerie. Chem Ber. 1951, 84, 319–29. [Google Scholar] [CrossRef]
- Farmer, V.C. Spectra and structure of 4-hydroxycoumarins. Spectrochim. Acta 1959, 15, 870–82. [Google Scholar] [CrossRef]
- Lake, B.G.; Gaudin, H.; Price, R.J.; Walters, D.G. Metabolism of (3-carbon-14) coumarin to polar and covalently bound products by hepatic microsomes from the rat, Syrian hamster, gerbil and humans. Food Chem. Toxicol. 1992, 30(2), 105–115. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G. Coumarin metabolism, toxicity and carcinogenicity: relevance for human risk assessment. Food Chem. Toxicol. 1999, 37(4), 423–453. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Katsutani, N.; Maruyama, T.; Kawamura, T.; Yamazaki, H.; Murayama, N.; Tong, W.; Yamazoe, Y.; Hirose, A. Combined risk assessment of food-derived coumarin with in Silico approaches. Food Safety 2022, 10(3), 73–82. [Google Scholar] [CrossRef]
- Catarino, S.; Thanasi, V.; Morin, G.; Anjos, O.; Fernandes, T.A.; Caldeira, I.; Fargeton, L.; Boissier, B.; Canas, S. Shedding light on metals release from chestnut wood to wine spirit using ICP-MS. Foods 2022, 11, 3617. [Google Scholar] [CrossRef]
- Prusty, J.S.; Kumar, A. Coumarins: antifungal effectiveness and future therapeutic scope. Mol. Divers 2020, 24, 1367–1383. [Google Scholar] [CrossRef]
- Kirillova, M.V.; Fernandes, T.A.; André, V.; Kirillov, A.M. Mild C−H functionalization of alkanes catalyzed by bioinspired copper(II) cores. Org. Biomol. Chem. 2019, 17, 7706–7714. [Google Scholar] [CrossRef]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins — An important class of phytochemicals. Phytochemicals - isolation, characterisation and role in human health. In Phytochemicals - Isolation, Characterisation and Role in Human Health; Rao, A.V., Rao, L.G., Eds; InTechOpen, London, UK, 2015; pp. 113–140. [CrossRef]
- Fraissinet-Tachet, L.; Baltz, R.; Chong, J.; Kauffmann, S.; Fritig, B.; Saindrenan, P. Two tobacco genes induced by infection, elicitor and salicylic acid encode glucosyltransferases acting on phenylpropanoids and benzoic acid derivatives, including salicylic acid. FEBS Lett. 1998, 437, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Nitao, J.K.; Zangerl, A.R. Floral development and chemical defense allocation in wild parsnip (Pastinaca Sativa). Ecology 1987, 68, 521–529. Available online: http://www.jstor.org/stable/1938457. [CrossRef]
- Tiwari, R.; Mishra, S.; Danaboina, G.; Jadaun, G.P.S.; Kalaivani, M.; Kalaiselvan, V.; Dhobi, M.; Raghuvanshi, R.S. Comprehensive chemo-profiling of coumarins enriched extract derived from Aegle marmelos (L.) Correa fruit pulp, as an anti-diabetic and anti-inflammatory agent. Saudi. Pharm. J. 2023, 31, 101708. [Google Scholar] [CrossRef]
- Ehlers, D.; Pfister, M.; Bork, W.-R.; Toffel-Nadolny, P. HPLC analysis of tonka bean extracts. Z. Lebensm.-Unters. Forsch. 1995, 201, 278–282. [Google Scholar] [CrossRef]
- Rohde, A.; Morreel, K.; Ralph, J.; Goeminne, G.; Hostyn, V.; De Rycke, R.; Kushnir, S.; van Doorsselaere, J.; Joseleau, J.-P.; Vuylsteke, M.; van Driessche, G.; van Beeumen, J.; Messens, E.; Boerjan, W. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism. Plant Cell 2004, 16, 2749–2771. [Google Scholar] [CrossRef] [PubMed]
- Mattivi, F.; Versini, G.; Sarti, S. La presenza di scopoletina come indicatore della modalità di invecchiamento dei distillati. Riv. Vitic. Enol. 1989, 2, 3–10. Available online: https://openpub.fmach.it/retrieve/e1dbfeaa-784b-4ac9-e053-1705fe0a1c61/RIV.%20ENOLOGIA%201989.pdf.
- Canas, S.; Leandro, M.C.; Spranger, M.I.; Belchior, A.P. Influence of botanical species and geographical origin on the content of low molecular weight phenolic compounds of woods used in Portuguese cooperage. Holzforschung 2000a, 54, 255–261. [Google Scholar] [CrossRef]
- Latté, K.P.; Kayser, O.; Tan, N.; Kaloga, M.; Kolodziej, H. Unusual coumarin patterns of Pelargonium species forming the origin of the traditional herbal medicine umckaloabo. Z. Naturforsch., C, J. Biosci. 2000, 55, 528–533. [Google Scholar] [CrossRef]
- Maier, W.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Secondary products in mycorrhizal roots of tobacco and tomato. Phytochemistry 2000, 54, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Eckey-Kaltenbach, H.; Ernst, D.; Heller, W.; Sandermann, H. Jr. Biochemical plant responses to ozone (IV. Cross-induction of defensive pathways in parsley (Petroselinum crispum L.) plants). Plant Physiol 1994, 104, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.C.; Parry, A.; Tena, M.; Jorrin, J.; Edwards, R. Abiotic elicitation of coumarin phytoalexins in sunflower. Phytochemistry 1995, 38, 1185–1191. [Google Scholar] [CrossRef]
- Hussain, M.I.; Syed, Q.A.; Khattak, M.N.K.; Hafez, B.; Reigosa, M.J.; El-Keblawy. A. Natural product coumarins: biological and pharmacological perspectives. Biologia 2019, 74, 863–888. [CrossRef]
- Sharan, M.; Taguchi, G.; Gonda, K.; Jouke, Y.; Shimosaka, M.; Hayashida, N.; Okazaki, M. Effects of methyl jasmonate and elicitor on the activation of phenylalanine ammonia-lyase and the accumulation of scopoletin and scopolin in tobacco cell cultures. Plant Sci. 1998, 132, 13–19. [Google Scholar] [CrossRef]
- de la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic Compounds. In Postharvest physiology and biochemistry of fruits and vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 253–271. [Google Scholar] [CrossRef]
- de Lira, S.P.; Seleghim, M.H.R.; Williams, D.E.; Marion, F.; Hamill, P.; Jean, F.; Andersen, R.J.; Hajdu, E.; Berlinck, R.G.S. A SARS-coronovirus 3CL protease inhibitor isolated from the marine sponge Axinella cf. corrugata: structure elucidation and synthesis. J. Braz. Chem. Soc. 2007, 18 (2). [Google Scholar] [CrossRef]
- Garrido, J.; Borges, F. Wine and grape polyphenols - a chemical perspective. Food Res. Int. 2013, 54, 1844–1858. [Google Scholar] [CrossRef]
- Hroboňová, K.; Sádecká, J. Coumarins content in wine: Application of HPLC, fluorescence spectrometry, and chemometric approach. J. Food Sci. Technol. 2020, 57, 200–209. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Cacco, G.; Sorgonà, A.; Marabottini, R.; Paolacci, A.R.; Ciaffi, M.; Badiani, M. The inhibitory effects of coumarin on the germination of durum wheat (Triticum turgidum ssp. durum, cv. Simeto) seeds. J. Chem. Ecol. 2006, 32, 489–506. [Google Scholar] [CrossRef]
- Saleh, A.M.; Kebeish, R. Coumarin impairs redox homeostasis in wheat aleurone layers. J. Plant Res 2018, 131, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Bini, A.P.; Rossi, G.D.; Poeschl, Y; Serra, M.C.D.; Camargo, L.E.A.; Monteiro-Vitorello, C.B.; van Sluys, M.-A.; van Dam, N.M.; Uthe H.; Creste, S. Molecular, biochemical and metabolomics analyses reveal constitutive and pathogen-induced defense responses of two sugarcane contrasting genotypes against leaf scald disease. Plant Physiol. Biochem. 2023, 203, 108033. [CrossRef]
- Cadahia, E.; Muñoz, L.; Fernández de Simón, B.; García-Vallejo, M.C. Changes in low molecular weight phenolic compounds in Spanish, French, and American oak woods during natural seasoning and toasting. J. Agric. Food Chem. 2001, 49, 1790–1798. [Google Scholar] [CrossRef]
- Smailagić, A.; Veljović, S.; Gašić, U.; Zagorac, D.D.; Stanković, M.; Radotić, K.; Natić, M. Phenolic profile, chromatic parameters and fluorescence of different woods used in Balkan cooperage. Ind. Crop. Prod. 2019, 132, 156–167. [Google Scholar] [CrossRef]
- Fernández de Simón, B.; Sanz, M.; Cadahía, E.; Poveda, P.; Broto, M. Chemical characterization of oak heartwood from Spanish forests of Quercus pyrenaica (Wild.). Ellagitannins, low molecular weight phenolic, and volatile compounds. J. Agric. Food Chem. 2006, 54, 8314–8321. [Google Scholar] [CrossRef]
- Sanz, M.; Cadahia, E.; Esteruelas, E.; Muñoz, A.M.; Fernández de Simón, B.; Hernández, T.; Estrella, I.; Pinto, E. Phenolic compounds in chestnut (Castanea sativa Mill.) heartwood. Effect of toasting at cooperage. J. Agric. Food Chem. 2010a, 58, 9631–9640. [Google Scholar] [CrossRef]
- Sanz, M.; Cadahia, E.; Esteruelas, E.; Muñoz, A.M.; Fernández de Simón, B.; Hernández, T.; Estrella, I. Phenolic compounds in cherry (Prunus avium) heartwood with a view to their use in cooperage. J. Agric. Food Chem. 2010b, 58, 4907–4914. [Google Scholar] [CrossRef] [PubMed]
- Masson, G.; Guichard, E.; Fournier, N.; Puech, J.-L. Stereoisomers of b-methyl-g-octalactone. II. Contents in the wood of French (Quercus robur and Quercus petraea) and American (Quercus alba) oaks. Am. J. Enol. Vitic. 1995a, 46, 424–428. [Google Scholar] [CrossRef]
- Masson, G.; Moutounet, M.; Puech, J.-L. Ellagitannin content of oak wood as a function of species and of sampling position in the tree. Am. J. Enol. Vit. 1995b, 46, 262–268. [Google Scholar] [CrossRef]
- Snakkers, G.; Nepveu, G.; Guilley, E.; Cantagrel, R. Variabilités géographique, sylvicole et individuelle de la teneur en extractibles de chênes sessiles français (Quercus petraea Liebl.): polyphénols, octalactones et phénols volatils. Ann. For. Sci. 2000, 57, 251–260. [Google Scholar] [CrossRef]
- Doussot, F.; De Jeso, B.; Quideau, S.; Pardon, P. Extractives content in cooperage oak wood during natural seasoning and toasting; influence of tree species, geographic location, and single-tree effects. J. Agric. Food Chem. 2002, 50, 5955–5961. [Google Scholar] [CrossRef]
- Jordão, A.M.; Ricardo da Silva, J.M.; Laureano, O. Ellagitannins from Portuguese oak wood (Quercus pyrenaica Willd.) used in cooperage: Influence of geographical origin, coarseness of the grain and toasting level. Holzforschung 2007, 61, 155–160. [Google Scholar] [CrossRef]
- Alañón, M.E.; Pérez-Coello, M.; Díaz-Maroto, I.J.; Martín-Alvarez, P.J.; Vila-Lameiro, P.; Díaz-Maroto, M.C. Influence of geographical location, site and silvicultural parameters, on volatile composition of Quercus pyrenaica Willd. Wood used in wine aging. For. Ecol. Manag. 2011, 262, 124–130. [Google Scholar] [CrossRef]
- Peng, S.; Scalbert, A.; Monties, B. Insoluble ellagitannins in Castanea sativa and Quercus petraea woods. Phytochemistry 1991, 30, 775–778. [Google Scholar] [CrossRef]
- Hamada, J.; Pétrissans, A.; Mothe, F.; Ruelle, J.; Pétrissans, M.; Gérardin, P. Intraspecific variation of European oak wood thermal stability according to radial position. Wood Sci. Technol. 2017, 51, 785–794. [Google Scholar] [CrossRef]
- Traoré, M.; Kaal, J.; Cortizas, A.M. Variation of wood color and chemical composition in the stem cross-section of oak (Quercus spp.) trees, with special attention to the sapwood-heartwood transition zone. Spectrochim. Acta A Mol. Biomol. Spectrosc 2023, 285, 121893. [Google Scholar] [CrossRef]
- Canas, S.; Belchior, A.P.; Spranger, M.I.; Bruno de Sousa, R. HPLC method for the quantification of phenolic acids, phenolic aldehydes, coumarins and furanic derivatives in different kinds of toasted wood used for the ageing of brandies. Anal. Methods 2011, 3, 186–19. [Google Scholar] [CrossRef]
- Annunziata, F.; Pinna, C.; Dallavalle, S.; Tamborini, L.; Pinto, A. An overview of coumarin as a versatile and readily accessible scaffold with broad-ranging biological activities. Int. J. Mol. Sci. 2020, 21, 4618. [Google Scholar] [CrossRef]
- Olvera-Aguirre, G.; Piñeiro-Vázquez, Á.T.; Sanginés-García, J.R.; Sánchez Zárate, A.; Ochoa-Flores, A.A.; Segura-Campos, M.R.; Vargas-Bello-Pérez, E.; Chay-Canul, A.J. Using plant-based compounds as preservatives for meat products: a review. Heliyon 2023, 9(6), e17071. [Google Scholar] [CrossRef]
- Piller, N.B. A comparison of the effectiveness of some anti-inflammatory drugs on thermal oedema. Br. J. Exp. Pathol. 1975, 56, 554–560. [Google Scholar]
- Huang, G.J.; Deng, J.S.; Liao, J.C.; Hou, W.C.; Wang, S.Y.; Sung, P.J.; Kuo, Y.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory activity of imperatorin from Glehnia littoralis. J. Agric. Food Chem. 2012, 60(7), 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Fylaktakidou, K.C.; Hadjipavlou-Litina, D.J.; Litinas, K.E.; Nicolaides, D.N. Natural and synthetic coumarin derivatives with anti-inflammatory/antioxidant activities. Curr. Pharm. Des. 2004, 10(30), 3813–3833. [Google Scholar] [CrossRef] [PubMed]
- Baek, N.I.; Ahn, E.M.; Kim, H.Y.; Park, Y.D. Furanocoumarins from the root of Angelica dahurica. Arch. Pharm. Res. 2000, 23(5), 467–470. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, S.; Maggio, A.M.; Faraone, N.; Spadaro, V.; Morris-Natschke, S.L.; Bastow, K.F.; Lee, K.H.; Bruno, M. The cytotoxic properties of natural coumarins isolated from roots of Ferulago campestris (Apiaceae) and of synthetic ester derivatives of aegelinol. Nat. Prod. Commun. 2009, 4(12), 1701–1706. [Google Scholar]
- Patil, A.D.; Freyer, A.J.; Eggleston, D.S.; Haltiwanger, R.C.; Bean, M.F.; Taylor, P.B.; Caranfa, M.J.; Breen, A.L.; Bartus, H.R.; Johnson, R.K.; et al. The inophyllums, novel inhibitors of HIV-1 reverse transcriptase isolated from the Malaysian tree, Calophyllum inophyllum Linn. J. Med. Chem. 1993, 36(26), 4131–4138. [Google Scholar] [CrossRef]
- Izquierdo, M.E.F.; Granados, J.Q.; Mir, M.V.; Martinez, M.C.L. Comparison of methods for determining coumarins in distilled beverages. Food Chem. 2000, 70 (2), 251–258. [Google Scholar] [CrossRef]
- Lozhkin, A.V.; Sakanyan, E.I. Structure of chemical compounds, methods of analysis and process control. Natural coumarins: methods of isolation and analysis. Pharm. Chem. J. 2006, 40(6), 337–346. [Google Scholar] [CrossRef]
- Skalicka-Woźniak, K.; Głowniak, K. Coumarins - Analytical and preparative techniques. In Encyclopedia of Analytical Chemistry: Applications, Theory and Instrumentation; 2014; pp. 1–26. [Google Scholar] [CrossRef]
- Canas, S.; Belchior, A.P.; Falcão, A.; Gonçalves, J.A.; Spranger, M.I.; Bruno de Sousa, R. Effect of heat treatment on the thermal and chemical modifications of oak and chestnut wood used in brandy ageing. Ciência Téc. Vitiv. 2007, 22 (1), 5–14. [Google Scholar]
- Lourenço, S.; Anjos, O.; Caldeira, I.; Oliveira Alves, S.; Santos, N.; Canas, S. Natural blending as a novel technology for the production process of aged wine spirits: Potential impact on their quality. Appl. Sci. 2022, 12, 10055. [Google Scholar] [CrossRef]
- Zacaroni, L.M.; Cardoso, M.G.; Saczk, A.A.; de Moraes, A.R.; dos Anjos, J.P.; Machado, A.M.R.; Nelson, D.L. Determination of phenolic compounds and coumarins in sugar cane spirit aged in different species of wood. Anal. Lett. 2011, 44, 2061–2073. [Google Scholar] [CrossRef]
- Bernardes, C.D.; Barbeira, P.J.S. Different chemometric methods for the discrimination of commercial aged cachaças. Food Anal. Methods 2016, 9, 1053–1059. [Google Scholar] [CrossRef]
- LončarM.; Jakovljević, M.; Šubarić, D.; Pavlić, M.; Služek, V.B.; Cindrić, I.; Molnar, M. Coumarins in food and methods of their determination: Review. Foods 2020, 9, 645. [CrossRef]
- Abu-Mustafa, E.A.; El-Bay, F.K.A.; Fayez, M.B.E. Natural coumarins XI. The distribution of coumarins in the Ammi Majus plant and a possible pattern for their biogenetic evolution. Planta Med. 1970, 18, 90–97. [Google Scholar] [CrossRef]
- Krieger, S.; Hayen, H.; Scmitz, O.J. Quantification of coumarin in cinnamon and woodruff beverages using DIP-APCI-MS and LC-MS. Anal Bioanal Chem. 2013, 405, 8337–8345. [Google Scholar] [CrossRef]
- Ballin, N.Z.; Sorensen, A.T. Coumarin content in cinnamon containing food products on the danish market. Food Control 2014, 38, 198–203. [Google Scholar] [CrossRef]
- Moreira, L.M.; de Melo, M.M., Martis, P.A.; Lyon, J.P.; Romani, A.P.; Codognoto, L.; dos Santos, S.C.; de Oliveira H.P.M. Photophysical properties of coumarin compounds in neat and binary solvent mixtures: evaluation and correlation between solvatochromism and solvent polarity parameters. J. Braz. Chem. Soc. 2014, 25(5), 873–881. [CrossRef]
- Shen, Y.; Han, C.; Liu,B.; Lin, Z.; Zhou, X.; Wang, C.; Zhu, Z. Determination of vanillin, ethyl vanillin, and coumarin in infant formula by liquid chromatography-quadrupole linear ion trap mass spectrometry. J. Dairy Sci. 2014, 97, 679–686. [CrossRef]
- McAdam, K.; Enos, T.; Goss, C.; Kimpton, H.; Faizi, A.; Edwards, S.; Wright, C.; Porter, A.; Rodu, B. Analysis of coumarin and angelica lactones in smokeless tobacco products. Chem. Cent. J. 2018, 12, 142. [Google Scholar] [CrossRef]
- Arigó, A.; Rigano, F.; Ruuso, M.; Trovato, E.; Dugo, P.; Mondello, L. Dietary intake of coumarins and furocoumarins through citrus beverages: a detailed estimation by HPLC-MS/MS method combined with the linear retention index system. Foods 2021, 10, 1533. [Google Scholar] [CrossRef]
- Aznar, R.; Rodríguez-Pérez, C.; Rai, D.K. Comprehensive characterization and quantification of antioxidant compounds in finger lime (Citrus australasica L.) by HPLC-QTof_MS and UPLC-MS/MS. Appl. Sci. 2022, 12, 1712. [Google Scholar] [CrossRef]
- Fernandes, T.A.; Antunes, A.M.M.; Caldeira, I.; Anjos, O.; de Freitas, V.; Fargeton, L.; Boissier, B.; Catarino, S.; Canas, S. Identification of gallotannins and ellagitannins in aged wine spirits: A new perspective using alternative ageing technology and high-resolution mass spectrometry. Food Chem. 2022, 382, 132322. [Google Scholar] [CrossRef] [PubMed]
- Razuvaeva, Y.G.; Toropova, A.A.; Salchak, S.M.; Olennikov, D.N. Coumarins of Ferulopsis hystrix: LC-MS profiling and gastroprotective and antioxidant activities of skimmin and peucenidin. Appl. Sci. 2023, 13, 9653. [Google Scholar] [CrossRef]
- Ren, Z.; Nie, B.; Liu, T.; Yuan, F.; Feng, F.; Zhang, Y.; Zhou, W.; Xu, X.; Yao, M.; Zhang, F. Simultaneous determination of coumarin and its derivatives in tobacco products by liquid chromatography-tandem mass spectrometry. Molecules 2016, 21(11), 1511. [Google Scholar] [CrossRef]
- Kabbash, E.M.; Abdel-Shakour, Z.T.; El-Ahmady, S.H.; Wink, M.; Ayoub, I.M. Comparative metabolic profiling of olive leaf extracts from twelve different cultivars collected in both fruiting and flowering seasons. Sci. Rep. 2023, 13, 612. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Ma, X.; Fedotov, D.; Kjaerulff, L.; Frydenvang, K.; Coriani, S.; Hansen, P.R.; Kongstad, K.T.; Staerk, D. Structure Elucidation of Prenyl- and Geranyl-Substituted Coumarins in Gerbera piloselloides by NMR Spectroscopy, Electronic Circular Dichroism Calculations, and Single Crystal X-ray Crystallography. Molecules 2020, 25(7), 1706. [Google Scholar] [CrossRef] [PubMed]
- Tabago, M.K.A.G.; Calingacion, M.N.; Garcia, J. Recent advances in NMR-based metabolomics of alcoholic beverages. Food Chem.: Mol. Sci. 2021, 2, 100009. [Google Scholar] [CrossRef] [PubMed]
- Hartner, N.T.; Wink, K.; Raddatz, C.R.; Thoben, C.; Schirmer, M.; Zimmermann, S.; Belder, D. Coupling droplet microfluidics with ion mobility spectrometry for monitoring chemical conversions at nanoliter scale. Anal Chem. 2021, 93(40), 13615–13623. [Google Scholar] [CrossRef]
- Faleh, A.B.; Warnke, S.; Rizzo, T.R. Combining ultrahigh-resolution ion-mobility spectrometry with cryogenic infrared spectroscopy for the analysis of glycan mixtures. Anal. Chem. 2019, 91(7), 4876–4882. [Google Scholar] [CrossRef]
- Salgado, A. Recent advances in nuclear magnetic resonance spectroscopy detection compatible with on-flow operational regimes: New uses of NMR detectors hyphenated to LCs and other separation techniques and in reaction monitoring. In Handbooks in Separation Science, Liquid Chromatography, 3rd ed.; Fanali, S.; Chankvetadze, B.; Haddad, P.R.; Poole, C.F.; Riekkola, M.-L., Eds.; Elsevier, London, UK, Volume 1, 2023, pp. 743–794. [CrossRef]
- Gawad, S.A.A.; Sakr, M.A.S. Spectroscopic investigation, DFT and TD-DFT calculations of 7-(Diethylamino) Coumarin (C466), J. Mol. Struct. 2022, 1248, 131413. [Google Scholar] [CrossRef]
- Rampogu, S.; Shaik, M.R.; Khan, M.; Khan, M.; Hwan Oh, T.; Shaik, B. CBPDdb: a curated database of compounds derived from Coumarin–Benzothiazole–Pyrazole. Database 2023, 2023, baad062. [Google Scholar] [CrossRef]
- Chen, H.M.; Nasseri, S.A.; Rahfeld, P.; Wardman, J.F.; Kohsiek, M.; Withers, S.G. Synthesis and evaluation of sensitive coumarin-based fluorogenic substrates for discovery of α-N-acetyl galactosaminidases through droplet-based screening. Org. Biomol. Chem. 2021, 19(4), 789–793. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, A.; Kapoor, A.; Kumar, P.S.; Ponnuchamy, M.; Sivasamy, B.; Vo, D.N. Lab-on-a-chip technologies for food safety, processing, and packaging applications: a review. Environ. Chem. Lett. 2022, 20, 901–927. [Google Scholar] [CrossRef]
- Corbion, C.; Smith-Ravin, J.; Marcelin, O.; Bouajila, J. An overview of spirits made from sugarcane juice. Molecules 2023, 28, 6810. [Google Scholar] [CrossRef]
- Dhiman, A.K.; Attri, S. Production of brandy. In Handbook of Enology: Principles, Practices and Recent Innovations; Josh V.K. Ed.; Asiatech Publisher, INC.: New Delhi, India, 2011; Chapter 35, pp. 60-119. [Google Scholar]
- Roullier-Gall, C.; Signoret, J.; Hemmler, D.; Witting, M.A.; Kanawati, B.; Schäfer, B.; Gougeon, R.D.; Schmitt-Koppli, P. Usage of FT-ICR-MS metabolomics for characterizing the chemical signatures of barrel-aged whisky. Front. Chem. 2018, 6, 29. [Google Scholar] [CrossRef]
- Quesada Granados, J.; Merelo Guervós, J.J.; Oliveras López, M.J.; González Penãlver, J.; Olalla Herrera, M.; Blanca Herrera, R.; López Martinez, M.C. Application of artificial aging techniques to samples of rum and comparison with traditionally aged rums by analysis with artificial neural nets. J. Agric. Food Chem. 2002, 50, 1470–1477. [Google Scholar] [CrossRef]
- Aylot, R.I.; Clyne, A.H.; Fox, A.P.; Walker, D.A. Analytical strategies to confirm Scotch whisky authenticity. Analyst 1994, 119, 1741–1746. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Puech, J.-L. Wood maturation of distilled beverages. Trends Food Sci. Technol. 1998, 9, 95–101. [Google Scholar] [CrossRef]
- Canas, S.; Vaz, M.; Belchior, A.P. Influence de la dimension du fût dans les cinétiques d’extraction/oxydation des composés phénoliques du bois pour les eaux-de-vie Lourinhã. In Les Eaux-de-vie Traditionnelles D’origine Viticole; Bertrand, A., Ed.; Lavoisier—Tec & Doc: Paris, France, 2008c; pp. 143–146.
- Bettin, S.; Isique, W.; Franco, D.; Andersen, M.; Knudsen, S.; Skibsted, L. Phenols and metals in sugar-cane spirits. Quantitative analysis and effect on radical formation and radical scavenging. Eur. Food Res. Technol. 2002, 215, 169–175. [Google Scholar] [CrossRef]
- Dos Anjos, J.P.; das Graças Cardoso, M.; Saczk, A.A.; Dórea, H.S.; Santiago, W.D.; Machado, A.M.R.; Zacaroni, L.M.; Nelson, D.L. Evolution of the concentration of phenolic compounds in cachaça during aging in an oak (Quercus sp.) barrel. J. Braz. Chem. Soc. 2011, 22, 1307–1314. [Google Scholar] [CrossRef]
- Nishimura, K.; Matsuyama, R. Maturation and maturation chemistry. In The science and technology of whiskies; Piggott J.R., Sharp R., Duncan R.E.B., Eds.; Longman Scientific & Technical, Essex, England, 1989; pp. 235–263.
- Sarni, F.; Moutounet, M.; Puech, J.-L.; Rabier, P. Effect of heat treatment of oak wood extractable compounds. Holzforschung 1990, 44, 461–466. [Google Scholar] [CrossRef]
- Le Floch, A.; Jourdes, M.; Teissedre, P.-L. Polysaccharides and lignin from oak wood used in cooperage: Composition, interest, assays: A review. Carbohydr. Res. 2015, 417, 94–102. [Google Scholar] [CrossRef]
- Hale, M.D.; McCafferty, K.; Larmie, E.; Newton, J.; Swan, J.S. The influence of oak seasoning and toasting parameters on the composition and quality of wine. Am. J. Enol. Vitic. 1999, 50, 495–502. [Google Scholar] [CrossRef]
- Acuña, L.; Gonzalez, D.; de la Fuente, J.; Moya, L. Influence of toasting treatment on permeability of six wood species for enological use. Holzforschung 2014, 68, 447–454. [Google Scholar] [CrossRef]
- Collins, T.S.; Miles, J.L.; Boulton, R.B.; Ebeler, S.E. Targeted volatile composition of oak wood samples taken during toasting at a commercial cooperage. Tetrahedron 2015, 71, 2971–2982. [Google Scholar] [CrossRef]
- Clyne, J.; Conner, J.M.; Paterson, A.; Piggott, J.R. The effect of cask charring on Scotch whisky maturation. Int. J. Food Sci. Technol. 1993, 28, 69–81. [Google Scholar] [CrossRef]
- Whiters, S. J.; Piggott, J. R.; Leroy, G.; Conner, J.M.; Paterson, A. Factors affecting pungency of malt distillates and ethanol-water mixtures. J. Sens. Studies 1995, 10, 273–283. [Google Scholar] [CrossRef]
- Ochiai, N.; Sasamoto, K.; MacNamara, K. Characterization of sulfur compounds in whisky by full evaporation dynamic headspace and selectable one-dimensional retention time locked gas chromatography-mass. J. Chromat. A 2012, 1270, 296–304. [Google Scholar] [CrossRef]
- Fengel, D.; Wegner, G. Wood. Chemistry, Ultrastructure, Reactions; Walter de Gruyter: Berlin, Germany, 1989; pp. 1–612. [Google Scholar]
- Gagić, T.; Knez, Z.; Skerget, M. Hydrothermal hydrolysis of sweet chestnut (Castanea sativa) tannins. J. Serbian Chem. Soc. 2019, 84, 1–14. [Google Scholar] [CrossRef]
- Sampaio, P,A.; Serafim, S.C.; Nascimento Menezes, P.M.; Valenca Pereira, E.C.; Sousa de Sá. P.G.; Teixeira de Alencar Filho, J.M.; Gonçalves de Oliveira Júnior, R.; Rolim Neto, P.J.; da Silva, J.M.; Rolim, L.A. Development and characterization of the zeolite imidazolate framework for a modified release of the drug scopoletin. J. Drug Deliv. Sci. Technol. 2022, 61, 102131. [CrossRef]
- Sun, C.; Huang, Z.; Liu, L.; Li, M.; Zheng, H. Umbelliferone as a Small Molecular Peroxidase Mimic towards Sensitive Detection of H2O2 and Glucose. Anal. Sci. 2018, 34(8), 933–938. [Google Scholar] [CrossRef] [PubMed]
- Canas, S.; Belchior, A.P.; Mateus, A.M.; Spranger, M.I.; Bruno de Sousa, R. Kinetics of impregnation/evaporation and release of phenolic compounds from wood to brandy in experimental model. Ciência Téc. Vitiv. 2002, 17, 1–14. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7(10), 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Salagoity-Auguste, M.-H. La scopolétine: un marqueur de la conservation en fûts de chêne. Vigne Vin Publ. Intern. 1992, 105–113. [Google Scholar]
- Ceccherini, G.; Duveiller, G.; Grassi, G.; Lemoine, G.; Avitabile, V.; Pilli, R.; Cescatti, A. Abrupt increase in harvested forest area over Europe after 2015. Nature 2020, 583, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Canas, S.; Caldeira, I.; Belchior, A.P. Comparison of alternative systems for the ageing of wine brandy. Wood shape and wood botanical species effect. Ciência Tec. Vitiv. 2009, 24, 91–99. [Google Scholar]
- Bortoletto, A.M.; Alcarde, A.R. Aging marker profile in cachaça is influenced by toasted oak chips. J. Inst. Brew. 2015, 121, 70–77. [Google Scholar] [CrossRef]
- Caldeira, I.; Anjos, O.; Belchior, A.P.; Canas, S. Sensory impact of alternative ageing technology for the production of wine brandies. Ciência Tec. Vitiv. 2017, 32, 12–22. [Google Scholar] [CrossRef]
- Taloumi, T.; Makris, D.P. Accelerated aging of the traditional Greek distillate Tsipouro using wooden chips. Part I: Effect of static maceration vs. ultrasonication on the polyphenol extraction and antioxidant activity. Beverages 2017, 3(1), 5. [Google Scholar] [CrossRef]
- Granja-Soares, J.; Roque, R.; Cabrita, M.J.; Anjos, O.; Belchior, A.P.; Caldeira, I.; Canas, S. Effect of innovative technology using staves and micro-oxygenation on the sensory and odorant profile of aged wine spirit. Food Chem. 2020, 333, 127450. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Jain, S.; Singh, J. Nutraceutical—Medicine of the future. J. Glob. Biosci. 2015, 4, 2790–2794. [Google Scholar]
- McClements, D.J.; Xiao, H. Designing food structure and composition to enhance nutraceutical bioactivity to support cancer inhibition. Semin. Cancer Biol. 2017, 46, 215–226. [Google Scholar] [CrossRef]
- Oliveira-Alves, S.C.; Serra, A.T.; Bronze, M.R. Food matrices that improve the oral bioavailability of pharmaceuticals and nutraceuticals. In Nutraceuticals and Natural Product Pharmaceuticals; Galanakis, C.M., Ed.; Academic Press: Cambridge, USA, 2019; pp. 197–233. [Google Scholar] [CrossRef]
- Holmes, M.V.; Dale, C.E.; Zuccolo, L.; Silverwood, R.J.; Guo, Y.; Ye, Z.; Prieto-Merino, D.; Dehghan, A.; Trompet, S.; Wong, A.; Cavadino, A.; Drogan, D.; Padmanabhan, S.; Li, S.; Yesupriya, A.; Leusink, M.; Sundstrom, J.; Hubacek, J.A.; Pikhart, H.; Swerdlow, D.I.; Panayiotou, A.G.; Borinskaya, S.A.; Finan, C.; Shah, S.; Kuchenbaecker, K.B.; Shah, T.; Engmann, J.; Folkersen, L.; Eriksson, P.; Ricceri, F.; Melander, O.; Sacerdote, C.; Gamble, D.M.; Rayaprolu, S.; Ross, O.A.; McLachlan, S.; Vikhireva, O.; Sluijs, I.; Scott, R.A.; Adamkova, V.; Flicker, L.; Bockxmeer, F.M.v.; Power, C.; Marques-Vidal, P.; Meade, T.; Marmot, M.G.; Ferro, J.M.; Paulos-Pinheiro, S.; Humphries, S.E.; Talmud, P.J.; Leach, I.M.; Verweij, N.; Linneberg, A.; Skaaby, T.; Doevendans, P.A.; Cramer, M.J.; Harst, P.v.d.; Klungel, O.H.; Dowling, N.F.; Dominiczak, A.F.; Kumari, M.; Nicolaides, A.N.; Weikert, C.; Boeing, H.; Ebrahim, S.; Gaunt, T.R.; Price, J.F.; Lannfelt, L.; Peasey, A.; Kubinova, R.; Pajak, A.; Malyutina, S.; Voevoda, M.I.; Tamosiunas, A.; Maitland-van der Zee, A.H.; Norman, P.E.; Hankey, G.J.; Bergmann, M.M.; Hofman, A.; Franco, O.H.; Cooper, J.; Palmen, J.; Spiering, W.; Jong, P.A.d.; Kuh, D.; Hardy, R.; Uitterlinden, A.G.; Ikram, M.A.; Ford, I.; Hypponen, E.; Almeida, O.P.; Wareham, N.J.; Khaw, K.-T.; Hamsten, A.; Husemoen, L.L.N.; Tjonneland, A.; Tolstrup, J.S.; Rimm, E.; Beulens, J.W.J.; Verschuren, W.M.M.; Onland-Moret, N.C.; Hofker, M.H.; Wannamethee, S.G.; Whincup, P.H.; Morris, R.; Vicente, A.M.; Watkins, H.; Farrall, M.; Jukema, J.W.; Meschia, J.; Cupples, L.A.; Sharp, S.J.; Fornage, M.; Kooperberg, C.; LaCroix, A.Z.; Dai, J.Y.; Lanktree, M.B.; Siscovick, D.S.; Jorgenson, E.; Spring, B.; Coresh, J.; Li, Y.R.; Buxbaum, S.G.; Schreiner, P.J.; Ellison, R.C.; Tsai, M.Y.; Patel, S.R.; Redline, S.; Johnson, A.D.; Hoogeveen, R.C.; Hakonarson, H.; Rotter, J.I.; Boerwinkle, E.; Bakker, P.I.W.d.; Kivimaki, M. Association between alcohol and cardiovascular disease: Mendelian randomisation analysis based on individual participant data. BMJ 2014, 349, 4164–4164. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Farzei, M.H.; Khodarahmi, R. Polyphenols and their benefits: a review. Int. J. Food Prop. 2017, 20, S2057–S2069. [Google Scholar] [CrossRef]
- Da Porto, C.; Calligaris, S.; Celotti, E.; Nicoli, M.C. Antiradical properties of commercial cognacs assessed by the DPPH(.) test. J. Agric. Food Chem. 2000, 48(9), 4241–4245. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.G.; Pedersen, M.W.; Gardner, P.T.; Morrice, P.C.; Jenkinson, A. McE.; McPhail, D.B.; Steele, G.M. The effect of whisky and wine consumption on total phenol content and antioxidant capacity of plasma from healthy volunteers. Eur. J. Clin. Nutr. 1998, 52(10), 733–736. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.: Nemoto, A.; Tanaka, I.; Koshimizu, S.; Suwa, Y.; Ishihara, H. Induction of heme oxygenase-1 by whisky congeners in human endothelial cells. J. Food Sci. 2010, 75 (6), H163–H166. [CrossRef]
- Owuor, E.D.; Kong, A.N. Antioxidants and oxidants regulated signal transduction pathways. Biochem. Pharmacol. 2002, 64, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Piggott, J.R. Whisky. In Current Developments in Biotechnology and Bioengineering. Food and Beverages Industry; Pandey, A., Sanromán, M.A., Du, G., Soccol, C.R., Dussap, C.-G., Eds.; Elsevier: London, UK, 2017; pp. 435–450. [Google Scholar] [CrossRef]
- Duriez P., Cren, C., Luc, G., Fruchart, J.C., Rolando, Ch. and Teissier E., 2001. Ingestion of cognac significantly increases plasma phenolic and ellagic acid concentrations and plasma antioxidant capacity in humans. In: Proceedings of 26th World Congress of OIV - Section Wine and health, 358-369.
- Umar, A.; Guerin, V.; Renard, M.; Boisseau, M.; Garreau, C.; Begaud, B.; Molimard, M.; Moore, N. Effects of Armagnac extracts on human platelet function in vitro and on rat arteriovenous shunt thrombosis in vivo. Thromb. Res. 2003, 110, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.P.; Pereira, T.M.; Vitorio, F.; Nadur, N.F.; Lacerda, R.B.; Kümmerle, A.E. A importância das cumarinas para a química medicinal e o desenvolvimento de compostos bioativos nos últimos anos. Quim. Nova 2021, 44(2), 180–197. [Google Scholar] [CrossRef]
- Garg, S.S.; Gupta, J.; Sharma, S.; Sahu, D. An insight into the therapeutic applications of coumarin compounds and their mechanisms of action. Eur. J. Pharm. Sci. 2020, 152, 105424. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Ding, Y.; Yan, X.T.; Kim, Y.-H.; Jan, H.-D. Scopoletin and scopolin isolated from Artemisia iwayomogi suppress differentiation of osteoclastic macrophage RAW 264.7 cells by scavenging reactive oxygen species. J. Nat. Prod. 2013, 76, 615–662. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Fang, F.; Song, M.; Li, R.; Ma, Z.; Ma, S. Umbelliferone reverses depression-like behavior in chronic unpredictable mild stress-induced rats by attenuating neuronal apoptosis via regulating ROCK/Akt pathway. Behav. Brain Res. 2017, 317, 147–156. [Google Scholar] [CrossRef]
- Wang, D.; Wang, X.; Tong, W.; Cui, Y.; Li, X.; Sun, H. Umbelliferone alleviates lipopolysaccharide-induced inflammatory responses in acute lung injury by down-regulating TLR4/MyD88/NF-κB signaling. Inflammation 2019, 42(2), 440–448. [Google Scholar] [CrossRef]
- Lee, B., Yeom, M., Shim, I.; · Lee, H.; Hahm, D.-H. Umbelliferone modulates depression-like symptoms by altering monoamines in a rat post-traumatic stress disorder model. J. Nat. Med. 2020, 74, 377–386. [CrossRef]
- Naowaboot, J.; Somparn, N.; Saentaweesuk, S.; Pannangpetch, P. Umbelliferone improves an impaired glucoseand lipid metabolism in high-fat diet/streptozotocin-induced type 2 diabetic rats. Phytother. Res. 2015, 29, 1388–1395. [Google Scholar] [CrossRef]
- Ramesh, B.; Pugalendi, K.V. Antihyperglycemic effect of umbelliferone in streptozotocin-diabetic rats. J. Med. Food 2006, 9(4), 562–566. [Google Scholar] [CrossRef]
- Yu, S.M.; Hu, D.H.; Zhang, J.J. Umbelliferone exhibits anticancer activity via the induction of apoptosis and cell cycle arrest in HepG2 hepatocellular carcinoma cells. Mol. Med. Rep. 2015, 12(3), 3869–3873. [Google Scholar] [CrossRef] [PubMed]
- Jesus, R.L.C.; Silva, I.L.P.; Araújo, F.A.; Moraes, R.A.; Silva, L.B.; Brito, D.S.; Lima, G.B.C.; Alves, Q.L.; Silva, D.F. 7-Hydroxycoumarin induces vasorelaxation in animals with essential hypertension: Focus on potassium channels and intracellular Ca2+ mobilization. Molecules 2022, 27, 7324. [Google Scholar] [CrossRef] [PubMed]
- Hajime, M.; Shuichi, Y.; Makoto, N.; Masanori, Y.; Ikuko, K.; Atsushi, K.; Mutsuo, S.; Keiichi, T. Inhibitory effect of 4-methylesculetin on hyaluronan synthesis slows the development of human pancreatic cancer in vitro and in nude mice. Int. J. Cancer 2007, 120(12), 2704–2709. [Google Scholar] [CrossRef]
- Urakawa, H.; Nishida, Y.; Wasa, J.; Arai, E.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan synthesis in breast cancer cells by 4-methylumbelliferone suppresses tumorigenicity in vitro and metastatic lesions of bone in vivo. Int. J. Cancer 2012, 130(2), 454–466. [Google Scholar] [CrossRef] [PubMed]
- Edward, M.; Quinn, J.A.; Pasonen-Seppänen, S.M.; McCann, B.A.; Tammi, R.H. 4-Methylumbelliferone inhibits tumour cell growth and the activation of stromal hyaluronan synthesis by melanoma cell-derived factors. Br. J. Dermatol. 2010, 162(6), 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, D.V.; Nikolskaya, A.I.; Goriainov, S.V.; Astakhova, A.A.; Sergeeva, M.G. Inhibitor of hyaluronic acid synthesis 4-methylumbelliferone as an anti-inflammatory modulator of LPS-mediated astrocyte responses. Int. J. Mol. Sci. 2020, 21, 8203. [Google Scholar] [CrossRef] [PubMed]
- Sim, M.-O.; Ham, J.R.; Lee, H.-I.; Seo, K.-I.; Lee, M.-K. Long-term supplementation of umbelliferone and 4-methylumbelliferone alleviates high-fat diet induced hypertriglyceridemia and hyperglycemia in mice. Chem. Biol. Interact. 2014, 216, 9–16. [Google Scholar] [CrossRef]
- Orhan, I.; Tosun, F.; Sener, B. Coumarin, anthroquinone and stilbene derivatives with anticholinesterase activity. Z. Naturforsch. C. J. Biosci. 2008, 63(5-6), 366–370. [Google Scholar] [CrossRef]
- Shin, E.; Choi, K.-M.; Yoo, H.-S.; Lee, C.-K.; Hwang, B.Y.; Lee, M.K. Inhibitory effects of coumarins from the stem barks of Fraxinus rhynchophylla on adipocyte differentiation in 3T3-L1 cells. Biol. Pharm. Bull. 2010, 33(9), 1610–1614. [Google Scholar] [CrossRef]
- Prabakaran, D.; Ashokkumar, N. Protective effect of esculetin on hyperglycemia-mediated oxidative damage in the hepatic and renal tissues of experimental diabetic rats. Biochimie 2013, 95(2), 366–373. [Google Scholar] [CrossRef]
- Hsia, C.-W.; Lin, K.-C.; Lee, T.-Y.; Hsia, C.-H.; Chou, D.-S.; Jayakumar, T.; Velusamy, M.; Chang, C.-C.; Sheu, J.-R. Esculetin, a coumarin derivative, prevents thrombosis: Inhibitory signaling on PLCγ2–PKC–AKT activation in human platelets. Int. J. Mol. Sci. 2019, 20, 2731. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-C.; Lee, H.-J.; Hu, C.-C.; Shun, H.-I.; Tseng, T.-H. Enhancement of esculetin on Taxol-induced apoptosis in human hepatoma HepG2 cells. Toxicol. Appl. Pharmacol. 2006, 210, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Kok, S.H.; Yeh, C.C.; Chen, M.L.; Kuo, M.Y.P. Esculetin enhances TRAIL-induced apoptosis through DR5 upregulation in human oral cancer SAS cells. Oral Oncol. 2009, 45, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Witaicenis, A.; Seito, L.N.; Di Stasi, L.C. Intestinal antiinflammatory activity of esculetin and 4-methylesculetin in the trinitrobenzenesulphonic acid model of rat colitis. Chem. Biol. Interact. 2010, 186(2), 211–218. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Li, L.; Yu, H.; Wu, Y.; Li, H. Coumarins ameliorate diabetogenic action of dexamethasone via Akt activation and AMPK signaling in skeletal muscle. J. Pharmacol. Sci. 2019, 139(3), 151–157. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, X.; Qin, S.; Zhou, S.; Li, J.; Gao, Y. Esculin ameliorates cognitive impairment in experimental diabetic nephropathy and induces anti-oxidative stress and anti-inflammatory effects via the MAPK pathway. Mol. Med. Rep. 2018, 17, 7395–7402. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Lee, W.; Jung, Y.; Lee, J.H.; Lee, S.; Eom, D.W.; Jeon, Y.; Yoo, H.H.; Jin, M.J.; Song, K.I.; Kim, W.J.; Ham, J.; Kim, H.J.; Kim, S.N. Protective effect of esculin on streptozotocin-induced diabetic renal damage in mice. J. Agric. Food Chem. 2014, 62(9), 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Y.; Wang, X.; Zhang, H.; He, Z.; Zhi, W.; Liu, F.; Niu, X. Gastroprotective effect of esculin on ethanol-induced gastric lesion in mice Weifeng. Fundam. Clin. Pharmacol. 2017, 31, 174–184. [Google Scholar] [CrossRef]
- Zhang, M.; Xin, X.; Lai, F.; Zhang, X.; Li, X.; Wu, H. Cellular tansport of esculin and its acylated derivatives in Caco-2 Cell monolayers and their antioxidant properties in vitro. J. Agric. Food Chem. 2017, 65, 7424–7432. [Google Scholar] [CrossRef]
- Niu, X.; Wang, Y.; Li, W.; Zhang, H.; Wang, X.; Mu, Q.; He, Z.; Yao, H. Esculin exhibited anti-inflammatory activities in vivo and regulated TNF-α and IL-6 production in LPS-stimulated mouse peritoneal macrophages in vitro through MAPK pathway. Int. Immunopharmacol. 2015, 29, 779–786. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Wang, X.; He, Z.; Liu, F.; Zhi, W.; Zhang, H.; Niu, X. Esculin attenuates endotoxin shock induced by lipopolysaccharide in mouse and NO production in vitro through inhibition of NF-κB activation. Eur. J. Pharmacol. 2016, 791, 726–734. [Google Scholar] [CrossRef]
- Liu, A.; Shen, Y.; Du, Y.; Chen, J.; Pei, F.; Fu, W.; Qiao, J. Esculin prevents lipopolysaccharide/D-galactosamine-induced acute liver injury in mice. Microb. Pathog. 2018, 125, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Pari, L.; Rajarajeswari, N. Efficacy of coumarin on hepatic key enzymes of glucose metabolism in chemical induced type 2 diabetic rats. Chem. Biol. Interact. 2009, 181(3), 292–296. [Google Scholar] [CrossRef]
- Abul Qais, F.; Ahmad, I. Mechanism of non-enzymatic antiglycation action by coumarin: A biophysical study. New J. Chem. 2019, 43, 12823. [Google Scholar] [CrossRef]
- Chuang, J.Y.; Huang, Y.F.; Lu, H.F.; Ho, H.C.; Yang, J.S.; Li, T.M.; Chang, N.W.; Chung, J.G. Coumarin induces cell cycle arrest and apoptosis in human cervical cancer HeLa cells through a mitochondria and caspase-3 dependent mechanism and NF-κB down-regulation. In Vivo 2007, 21(6), 1003–1009. Available online: https://iv.iiarjournals.org/content/invivo/21/6/1003.full.pdf.
- Elinos-Báez, C.M.; León, F.; Santos, E. Effects of coumarin and 7OH-coumarin on bcl-2 and Bax expression in two human lung cancer cell lines in vitro. Cell Biol Int. 2005, 29(8), 703–708. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gonzalez, J.S.; Prado-Garcia, H.; Aguilar-Cazares, D.; Molina-Guarneros, J.A. Apoptosis and cell cycle disturbances induced by coumarin and 7-hydroxycoumarin on human lung carcinoma cell lines. Lung Cancer 2004, 43, 275–283. [Google Scholar] [CrossRef]
- Luchini, A.C.; Rodrigues-Orsi, P.; Cestari, S.H.; Seito, L.N.; Witaicenis, A.; Pellizzon, C.H.; Di Stasi, L.C. Intestinal anti-inflammatory activity of coumarin and 4-hydroxycoumarin in the trinitrobenzenesulphonic acid model of rat colitis. Biol. Pharm. Bull. 2008, 31(7), 1343–1350. [Google Scholar] [CrossRef]
- Napiroon, T.; Bacher, M.; Balslev, H.; Tawaitakham, K.; Santimaleeworagun, W.; Vajrodaya, S. Scopoletin from Lasianthus lucidus Blume (Rubiaceae): a potential antimicrobial against multidrug-resistant Pseudomonas aeruginosa. J. Appl. Pharmaceut. Sci. 2018, 8, 1–6. [Google Scholar] [CrossRef]
- Armenia, A.; Hidayat, R.; Meiliani, M.; Yuliandra, Y. Blood pressure lowering effect of scopoletin on oxidative stress-associated hypertensive rats. J. Res. Pharm. 2019, 23(2), 249–258. [Google Scholar] [CrossRef]
- Jang, J.H.; Park, J.E.; Han, J.S. Scopoletin inhibits α-glucosidase in vitro and alleviates postprandial hyperglycemia in mice with diabetes. Eur. J. Pharmacol. 2018, 834, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Park, J.E.; Han, J.S. Scopoletin increases glucose uptake through activation of PI3K and AMPK signaling pathway and improves insulin sensitivity in 3T3-L1 cells. Nutr. Res. 2020, 74, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Jamuna, S.; Karthika, K.; Paulsamy, S.; Thenmozhi, K.; Kathiravan, S.; Venkatesh, R. Confertin and scopoletin from leaf and root extracts of Hypochaeris radicata have anti-inflammatory and antioxidant activities. Ind. Crop Prod. 2015, 70, 221–230. [Google Scholar] [CrossRef]
- Nam, H.; Kim, M.-M. Scopoletin has a potential activity for anti-aging via autophagy in human lung fibroblasts. Phytomedicine 2015, 22(3), 362–368. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29(11), 751–60. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Marques, M.C.; Hacke, A.; Loubet Filho, P.S.; Cazarin, C.B.B.; Mariutti, L.R.B. Trust your gut: Bioavailability and bioaccessibility of dietary compounds. Curr. Res. Food Sci. 2022, 5, 228–233. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Cruz-Martins, N.; López-Jornet, P.; Lopez, E.P.-F.; Harun, N.; Yeskaliyeva, B.; Beyatli, A.; Sytar, O.; Shaheen, S.; Sharopov, F.; Taheri, Y.; Docea, A.O.; Calina, D.; Cho, W.C. Natural coumarins: Exploring the pharmacological complexity and underlying molecular mechanisms. Oxid. Med. Cell. Longev. 2021, 6492346. [Google Scholar] [CrossRef]
- Xie, Q.; Wang, X.; Wang, X.; Jiang, Z.; Qiu, Z. Design, synthesis, and bioavailability evaluation of coumarin-based prodrug of meptazinol. Bioorganic Med. Chem. Lett. 2005, 15(22), 4953–4956. [Google Scholar] [CrossRef]
- Escudero, G.E.; Laino, C.H.; Echeverría, G.A.; Piro, O.E.; Martini, N.; Rodríguez, A.N.; Martínez Medina, J.J.; López Tévez, L.L.; Ferrer, E.G.; Williams, P.A.M. Improving the antidepressant action and the bioavailability of sertraline by co-crystallization with coumarin 3-carboxylate. Structural determination. Chem. Biol. Interact. 2016, 249, 46–55. [Google Scholar] [CrossRef]
- Tripathi, A.; Misra, K. Inhibition of P-glycoprotein mediated efflux of Paclitaxel by coumarin derivatives in cancer stem cells: An in silico approach. Comb. Chem. High Throughput Screen. 2016, 19(6), 497–506. [Google Scholar] [CrossRef] [PubMed]
- Karthika, C.; Sureshkumar, R.; Zehravi, M.; Akter, R.; Ali, F.; Ramproshad, S.; Mondal, B.; Tagde, P.; Ahmed, Z.; Khan, F.S.; Rahman, M.H.; Cavalu, S. Multidrug resistance of cancer cells and the vital role of P-glycoprotein. Life (Basel). 2022, 12(6), 897. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Hyohdoh, I.; Furuichi, N.; Ozawa, S.; Watanabe, F.; Matsushita, M.; Sakaitani, M.; Ori, K.; Takanashi, K.; Harada, N.; Tomii, Y.; Tabo, M.; Yoshinari, K.; Aoki, Y.; Shimma, N.; Iikura, H. The sulfamide moiety affords higher inhibitory activity and oral bioavailability to a series of coumarin dual selective RAF/MEK inhibitors. Bioorganic Med. Chem. Lett. 2013, 23(23), 6223–6227. [Google Scholar] [CrossRef]
- Eggleston, W. Coumarins. In Encyclopedia of Toxicology, 4th ed.; Wexler, P., Ed.; Academic Press: Cambridge, USA, 2024; pp. 293–297. [Google Scholar] [CrossRef]
- Clark, G.S. Coumarin. An aroma chemical profile. Perfum. flavor. 1995, 20, 23–34. Available online: https://img.perfumerflavorist.com/files/base/allured/all/document/2016/02/pf.9545.pdf.
- Hazleton, L.W.; Murer, H.K.; Thiessen, R.Jr.; Tusing, T.W.; Zeitlin, B.R. Toxicity of coumarin. J. Pharmaco.l Exp. Ther. 1956, 118(3), 348–358. Available online: https://jpet.aspetjournals.org/content/118/3/348.long.
- Carlton, B.D.; Aubrun, J.-C.; Simon, G.S. Effects of coumarin following perinatal and chronic exposure in Sprague-Dawley rats and CD-1 mice. Fundam. Appl. Toxicol. 1996, 30, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Opdyke, D.L.J. Monographs on fragrance raw materials: Coumarin. Food Chem. Toxicol. 1974, 12(3), 385–388. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Iwaki, R. Coumarin derivatives for medicinal purposes. XVII. Yakugaku Zasshi 1963, 83(12), 1124–1128. [Google Scholar] [CrossRef]
- Shimamoto, K.; Takaori. S. Pharmakologische untersuchungen mit einem Melilotus-extrakt [Pharmacologic studies with a Melilotus extract]. Arzneimittelforschung 1965, 15(8), 897–899.
- Jenner, P.M.; Hagan, E.C.; Taylor, J.M.; Cook, E.L.; Fitzhugh, O.G. Food flavourings and compounds of related structure I. Acute oral toxicity. Food Cosmet. Toxicol. 1964, 2, 327–343. [Google Scholar] [CrossRef]
- Nieschulz, O.; Schmersahl, P. Uber choleretische wirkstoffe aus Artemisia abrotanum L [On choleretic agents from Artemisia abrotanum L]. Arzneimittelforschung 1968, 18(10), 1330–1336. [Google Scholar]
- Mishkinsky, J.S.; Goldschmied, A.; Joseph, B.; Ahronson, Z.; Sulman, F.G. Hypoglycaemic effect of Trigonella foenum graecum and Lupinus termis (leguminosae) seeds and their major alkaloids in alloxan-diabetic and normal rats. Arch. Int. Pharmacodyn. Ther. 1974, 210(1), 27–37. [Google Scholar]
- KSRNAM Kiso to Rinsho. Clinical Report; Yubunsha Co., Ltd.: Tokyo, Japan, 1971, Volume 5, pp. 1619. [Google Scholar]
- PHTXA6 Pharmacology and Toxicology. English translation of FATOAO; New York, USA, 1958, Volume 21, pp. 555.
- Yamagami, I.; Suzuki, Y.; Ito, K. Pharmacological studies on the components of Fraxinus japonica Blume. Nihon Yakurigaku Zasshi. 1968, 64(6), 714–729. [Google Scholar]
- Pencheva, I.; Kostovs, I.; Konstantinov, S.; Naidenova, E.; Karaivanova, M.; Manolov, I. Cardioprotective efficacy of new esculin metal complexes. Acta Pharm. 1998, 48(2), 127–131. [Google Scholar]
- United Nations (ONU). United Nations Purple Book – Globally Harmonized System of Classification and Labelling of Chemicals (GHS), 8th revised ed.; ONU: New York, USA; Geneva, Swiss, 2019; p. 117. Available online: https://unece.org/fileadmin/DAM/trans/danger/publi/ghs/ghs_rev08/ST-SG-AC10-30-Rev8e.pdf.
- Lindberg, R.L.; Negishi, M. Alteration of mouse cytochrome P450coh substrate specificity by mutation of a single amino-acid residue. Nature 1989, 339(6226), 632–634. [Google Scholar] [CrossRef]
- Raunio, H.; Rahnasto-Rilla, M. CYP2A6: genetics, structure, regulation, and function: Review. Drug Metab. Drug Interact. 2012, 27(2), 73–88. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, O.; Rautio, A.; Raunio, H.; Pasanen, M. CYP2A6: a human coumarin 7-hydroxylase. Toxicology 2000, 144(1-3), 139–147. [Google Scholar] [CrossRef]
- Popping, B.; Buck, N.; Bánáti, D.; Brereton, P.; Gendel, S.; Hristozova, N.; Chaves, S.M.; Saner, S.; Spink, J.; Willis, C.; Wunderlin, D. Food inauthenticity: Authority activities, guidance for food operators, and mitigation tools. Compr. Rev. Food Sci. Food Saf. 2022. [Google Scholar] [CrossRef] [PubMed]
- Danezis, G.P.; Tsagkaris, A.S.; Camin, F.; Brusic, V.; Georgiou, C.A. Food authentication: Techniques, trends & emerging approaches. TrAC, Trends Anal. Chem. 2016, 85(Part A), 123–132. [Google Scholar] [CrossRef]
- Martins, A.R.; Talhavini, M.; Vieira, M.L.; Zacca, J.J.; Braga, J.W.B. Discrimination of whisky brands and counterfeit dentification by UV–Vis spectroscopy and multivariate data analysis. Food Chem. 2017, 229, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewska, P.; Boqué, R.; Borràs, E.; Busto, O.; Wardencki, W.; Namieśnik, J.; Dymerski, T. Authentication of whisky due to its botanical origin and way of production by instrumental analysis and multivariate classification methods. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2017, 173, 849–853. [Google Scholar] [CrossRef]
- Kiefer, J.; Cromwell, A.L. Analysis of single malt Scotch whisky using Raman spectroscopy. Anal. Methods 2017, 9(3), 511–518. [Google Scholar] [CrossRef]
- Žiak, Ľ.; Sádecká, J.; Májek, P.; Hroboňová, K. Simultaneous determination of phenolic acids and scopoletin in brandies using synchronous fluorescence spectrometry coupled with partial least squares. Food Anal. Methods 2014, 7, 563–570. [Google Scholar] [CrossRef]
- Ziyatdinova, G.; Salikhova, I.; Skorobogatova, N.; Chibisova, M.; Budnikov, H. New electrochemistry-based approaches to brandy quality evaluation using antioxidant parameters. Food Anal. Methods 2015, 8, 1794–1803. [Google Scholar] [CrossRef]
- Stupak, M.; Goodall, I.; Tomaniova, M.; Pulkrabova, J.; Hajslova, J. A novel approach to assess the quality and authenticity of Scotch whisky based on gas chromatography coupled to high resolution mass spectrometry. Anal. Chim. Acta 2018, 1042, 60–70. [Google Scholar] [CrossRef]
- Heinz, H.A.; Elkins, J.T. Comparison of unaged and barrel aged whiskies from the same Mash Bill using gas chromatography/mass spectrometry. J. Brew. Distilling 2019, 8(1), 1–6. [Google Scholar] [CrossRef]
- Yang, K.; Somogyi, A.; Thomas, C.; Zhang, H.; Cheng, Z.; Xu, S.; Miller, C.; Spivey, D.; Blake, C.; Smith, C.; Dafoe, D.; Danielson, N.D.; Crowder, M.W. Analysis of barrel-aged Kentucky Bourbon whiskey by ultrahigh resolution mass spectrometry. Food Anal. Methods 2020, 13(12), 2301–2311. [Google Scholar] [CrossRef]
- Heller, M.; Vitali, L.; Oliveira, M.A.L.; Costa, A.C.O.; Micke, G.A. A rapid sample screening method for authenticity control of whiskey using capillary electrophoresis with online preconcentration J. Agric. Food Chem. 2011, 59, 6882–6888. [Google Scholar] [CrossRef]
- Fotakis, C.; Zervou, M. NMR metabolic fingerprinting and chemometrics driven authentication of Greek grape marc spirits. Food Chem. 2016, 196, 760–768. [Google Scholar] [CrossRef]
- Monakhova, Y.B.; Kuballa, T.; Lachenmeier, D.W. Nontargeted NMR analysis to rapidly detect hazardous substances in alcoholic beverages. Appl. Magn. Reson. 2012, 42(3), 343–352. [Google Scholar] [CrossRef]
- Kuballa, T.; Hausler, T.; Okaru, A.O.; Neufeld, M.; Abuga, K.O.; Kibwage, I.O.; Rehm, J.; Luy, B.; Walch, S.G.; Lachenmeier, D.W. Detection of counterfeit brand spirits using 1H NMR fingerprints in comparison to sensory analysis. Food Chem. 2018, 245, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Tosato, F.; Correia, R.M.; Oliveira, B.G.; Fontes, A.M.; Franca, H.S.; Coltro, W.K.; Romao, W. Paper spray ionization mass spectrometry allied to chemometric tools for quantification of whisky adulteration with additions of sugarcane spirit. Anal. Methods 2018, 10(4), 1952–1960. [Google Scholar] [CrossRef]
- Moller, J.K.; Catharino, R.R.; Eberlin, M.N. Electrospray ionization mass spectrometry fingerprinting of whisky: immediate proof of origin and authenticity. Analyst 2005, 130, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Kew, W.; Mackay, C.L.; Goodall, I.; Clarke, D.; Uhrín, D. Complementary ionization techniques for the analysis of Scotch whisky by high resolution mass spectrometry. Anal. Chem. 2018, 90(19), 11265–11272. [Google Scholar] [CrossRef] [PubMed]
- Stupak, M.; Kocourek, V.; Kolouchova, I.; Hajslova, J. Rapid approach for the determination of alcoholic strength and overall quality check of various spirit drinks and wines using GC e MS. Food Control 2017, 80, 307–313. [Google Scholar] [CrossRef]
- Wiśniewska, P.; Dymerski, T.; Wardencki, W.; Namieśnik, J. Chemical composition analysis and authentication of whisky: Review. J. Sci. Food Agric. 2015, 95(11), 2159–2166. [Google Scholar] [CrossRef]
- Kelly, T.J.; O’Connor, C.; Kilcawley, K.N. Sources of Volatile Aromatic Congeners in whiskey. Beverages 2023, 9, 64. [Google Scholar] [CrossRef]
- Panossian, A.; Mamikonyan, G.; Torosyan, M.; Gabrielyan, E.; Mkhitaryan, S. Analysis of Aromatic Aldehydes in brandy and wine by high-performance capillary electrophoresis. Anal. Chem. 2001, 73(17), 4379–4383. [Google Scholar] [CrossRef]
- Rentzsch, M.; Wilkens, A.; Winterhalter, P. Non-flavonoid phenolic compounds. In Wine Chemistry and Biochemistry Moreno-Arribas, M.V., Polo, M.C., Eds; Springer: New York, USA, 2009; pp. 509–527. [Google Scholar] [CrossRef]
Figure 1.
Global market exchange rates of alcoholic beverages from 2018 to 2023. Adapted from Statista [4].
Figure 1.
Global market exchange rates of alcoholic beverages from 2018 to 2023. Adapted from Statista [4].
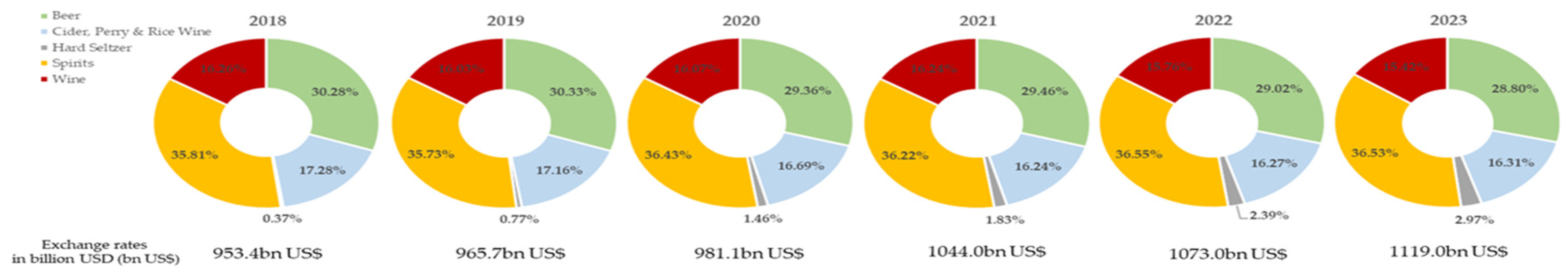
Figure 2.
Scheme of wine spirit production process.
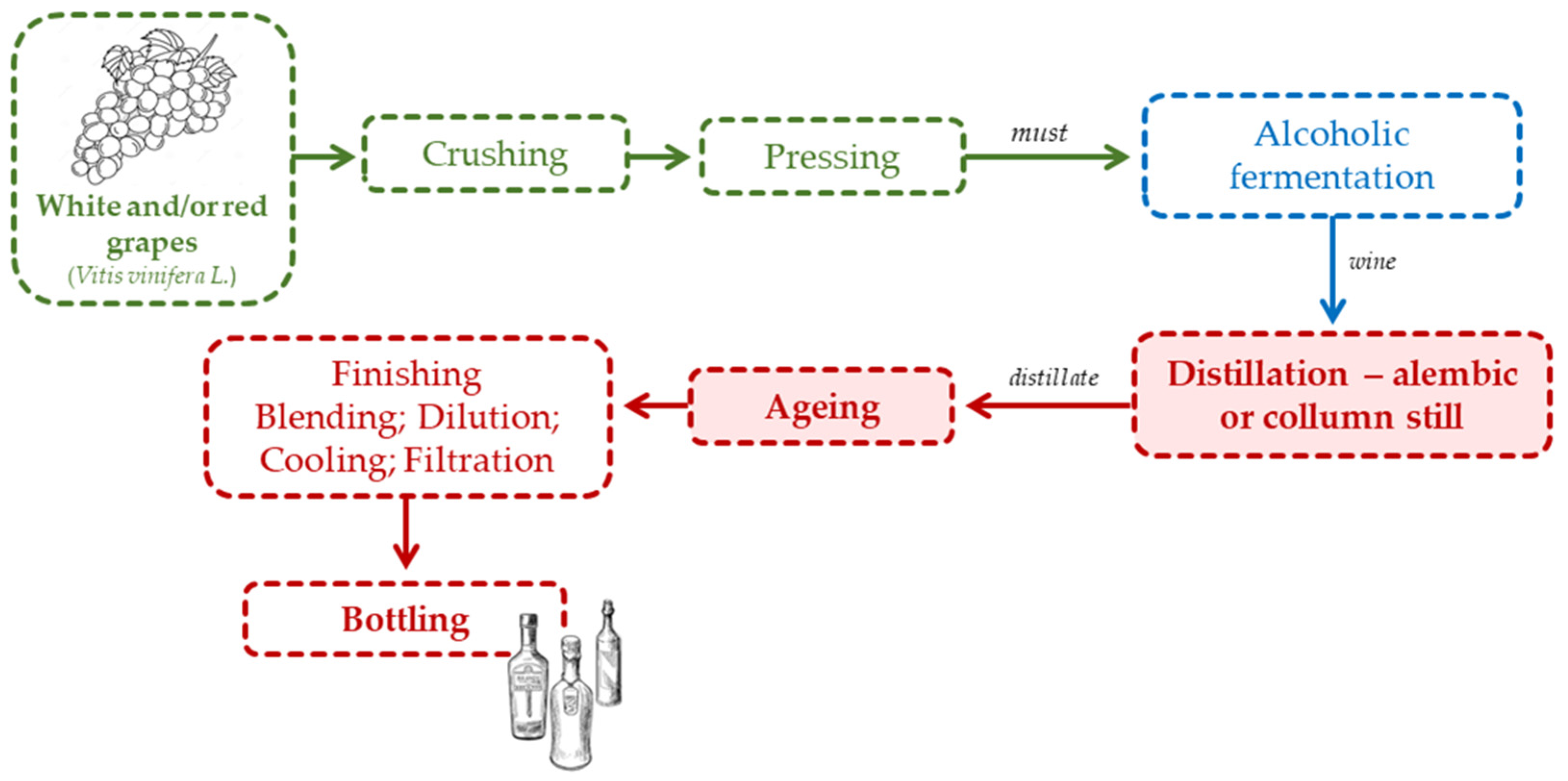
Figure 3.
Scheme of Brandy de Jerez ageing process. Adapted from Durán-Guerrero et al. [70].
Figure 3.
Scheme of Brandy de Jerez ageing process. Adapted from Durán-Guerrero et al. [70].
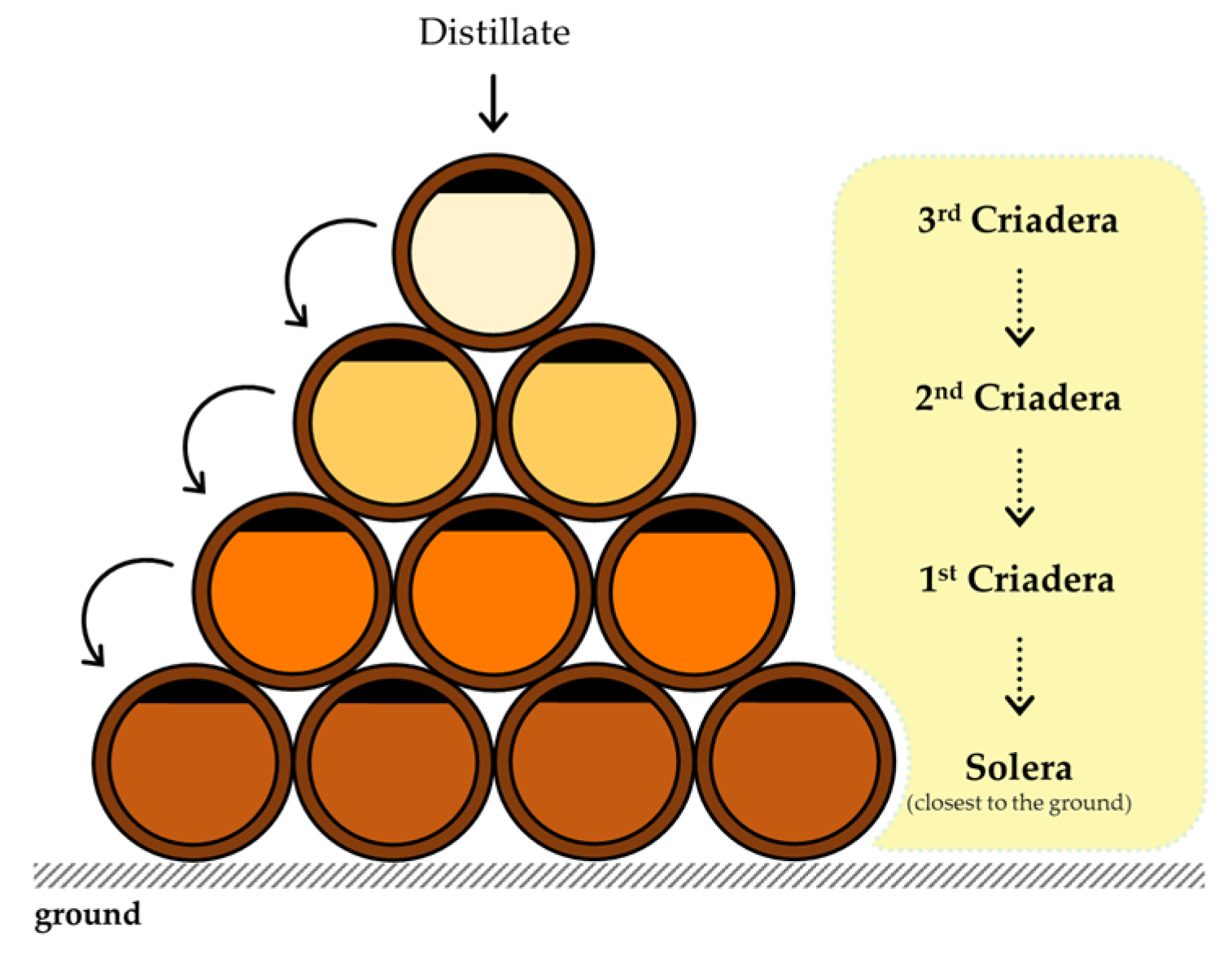
Figure 4.
Scheme of whiskey production process.
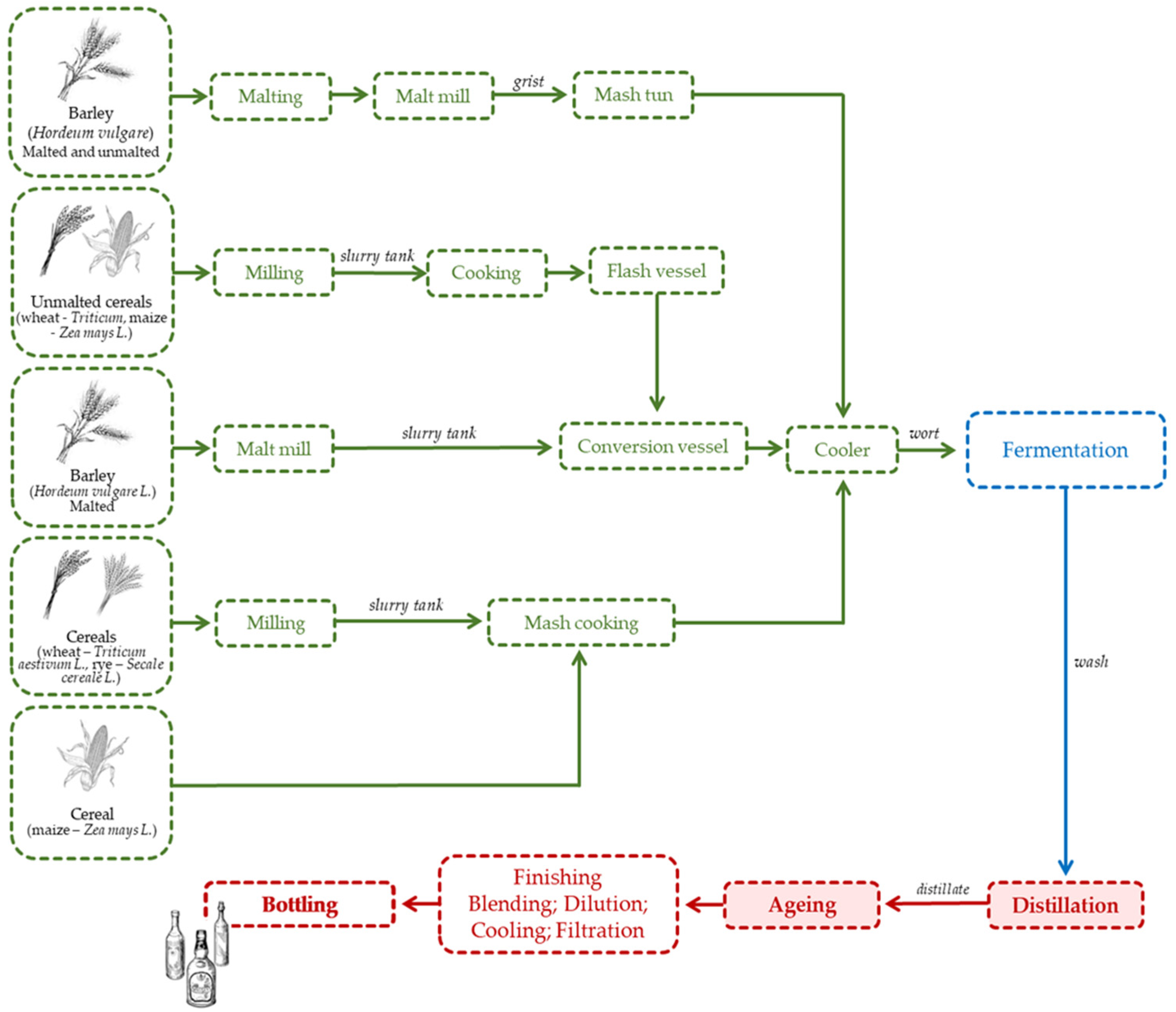
Figure 5.
Main types of whiskey.
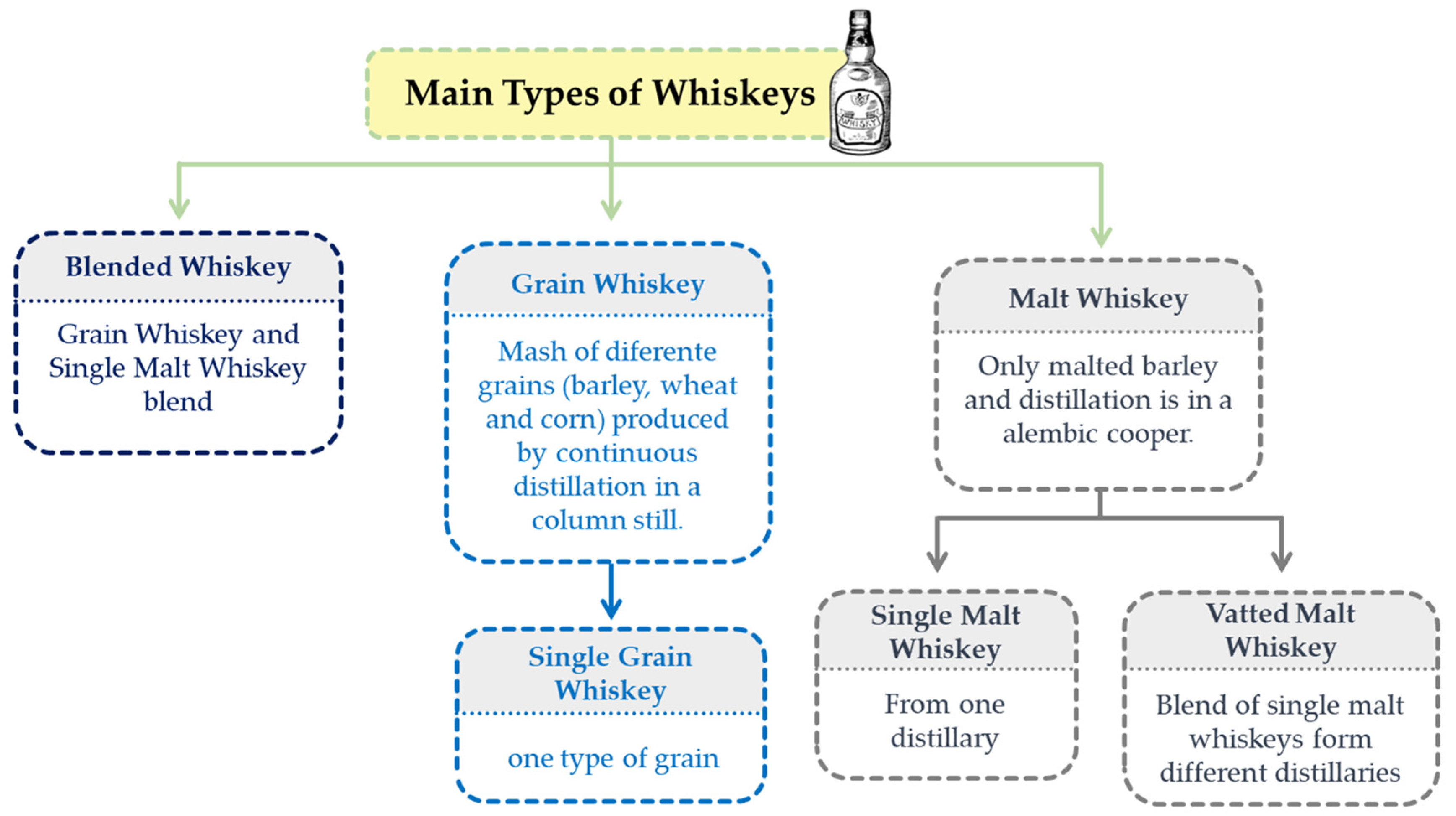
Figure 6.
Scheme of rum and cachaça production process.
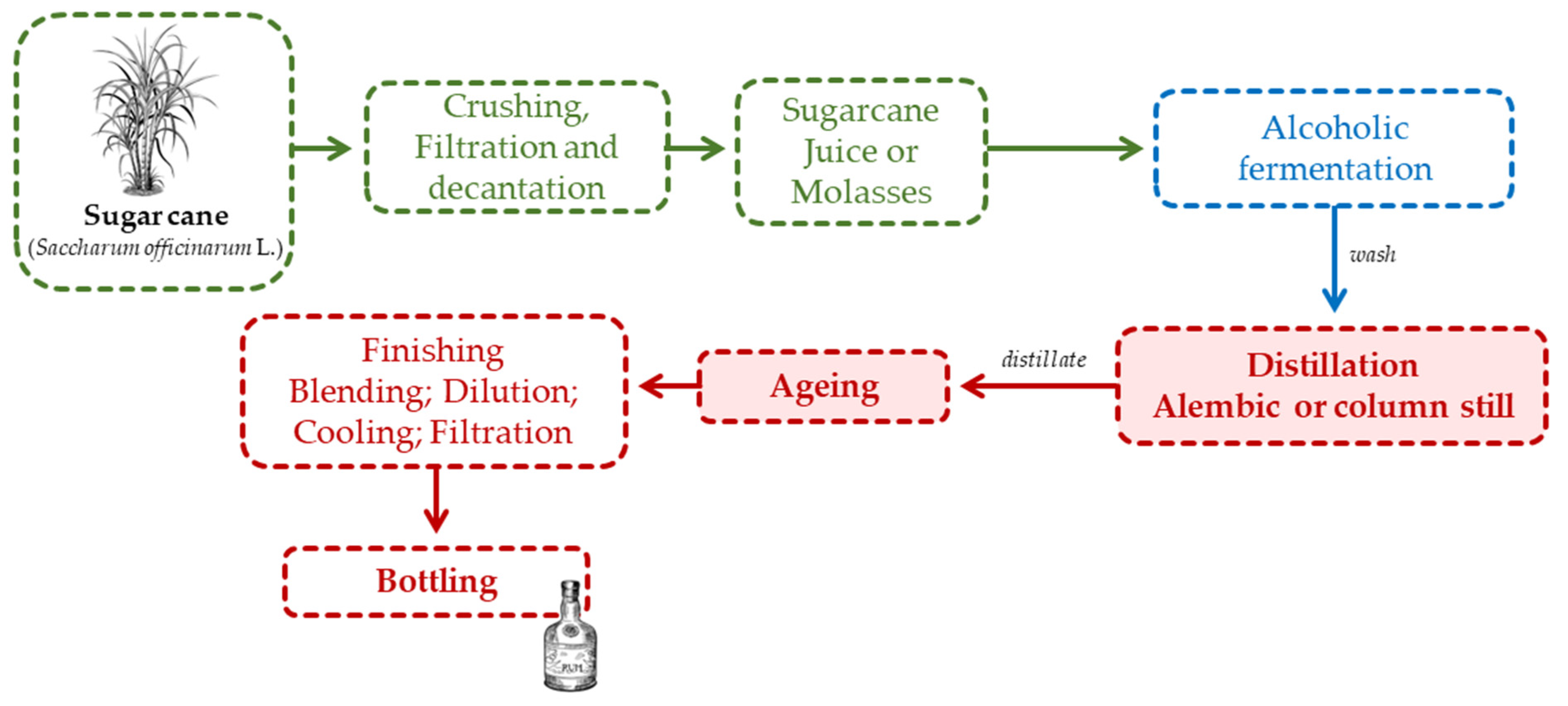
Figure 7.
Scheme of grape marc spirit production process.
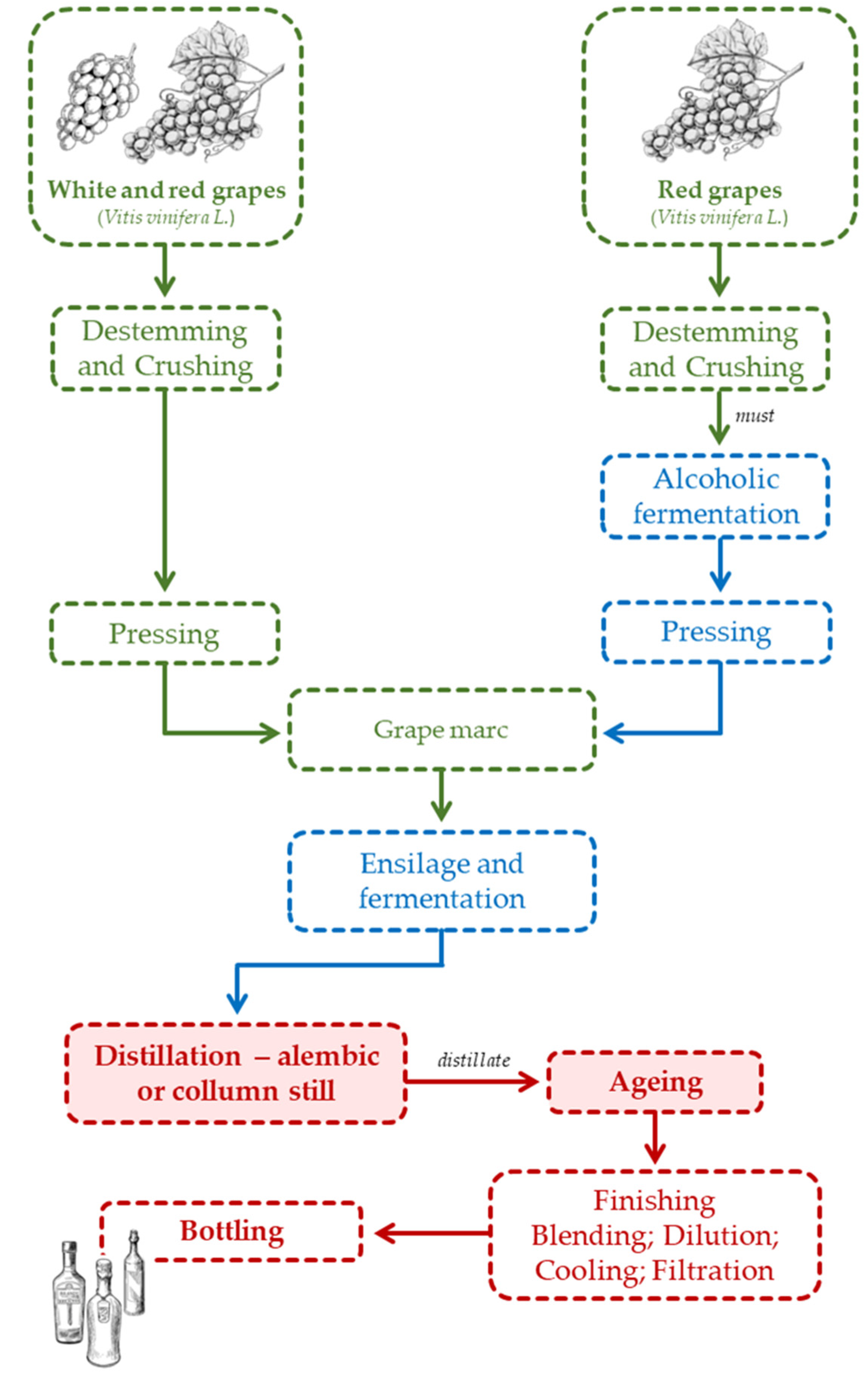
Figure 8.
Chemical structure of coumarin and the IUPAC numbering system for its framework.
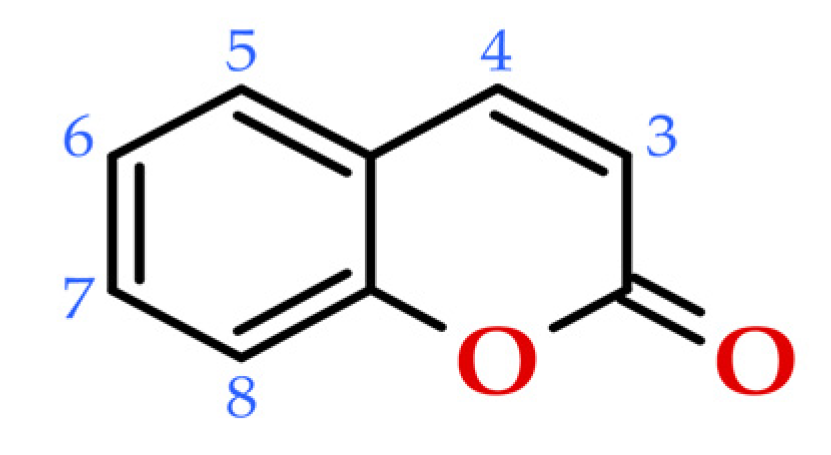
Figure 9.
Classification and generic structures of the main categories of plant-derived coumarins (some examples are given in green).
Figure 9.
Classification and generic structures of the main categories of plant-derived coumarins (some examples are given in green).
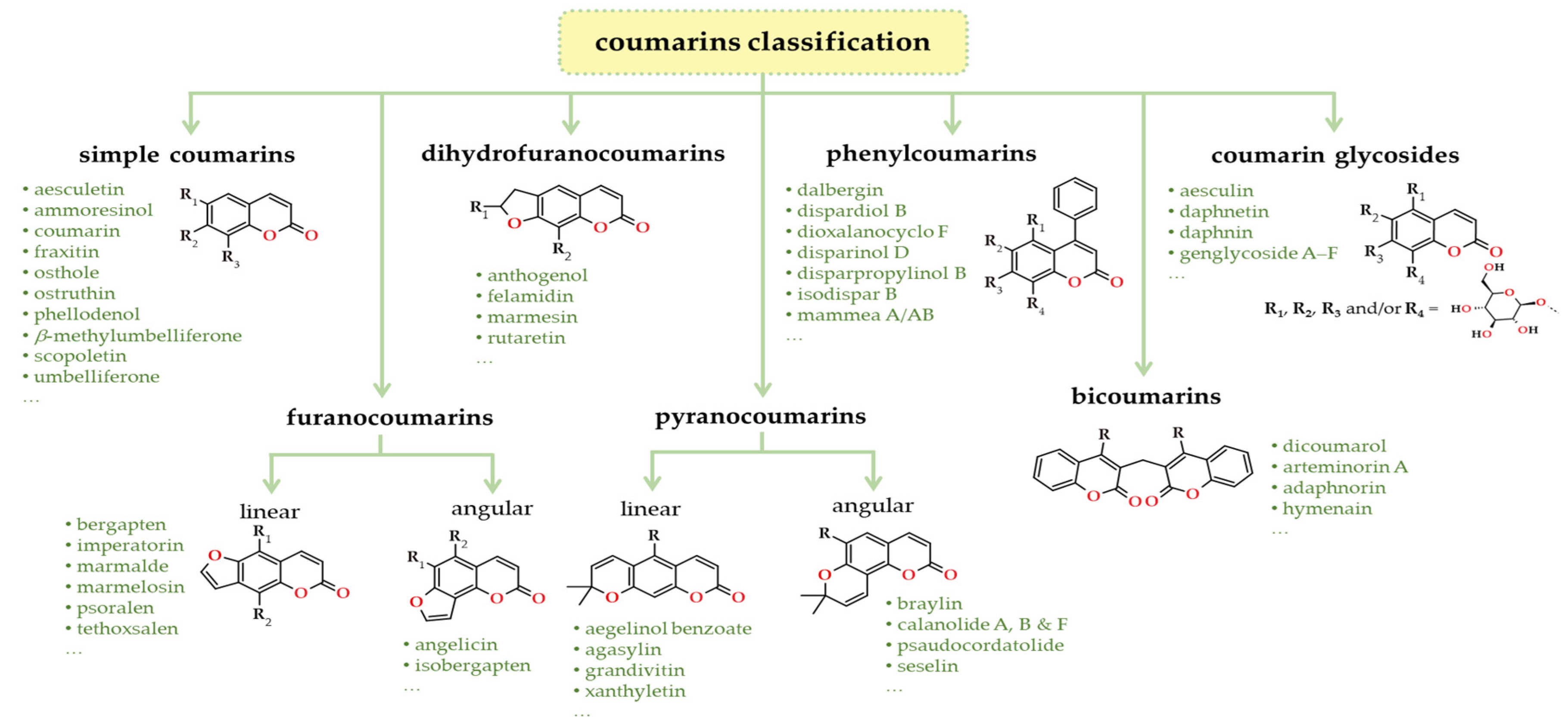
Figure 10.
Phenylpropanoids' general biosynthetic route. PAL: l-phenylalanine ammonia-lyase; TAL: tyrosine ammonia lyase.
Figure 10.
Phenylpropanoids' general biosynthetic route. PAL: l-phenylalanine ammonia-lyase; TAL: tyrosine ammonia lyase.
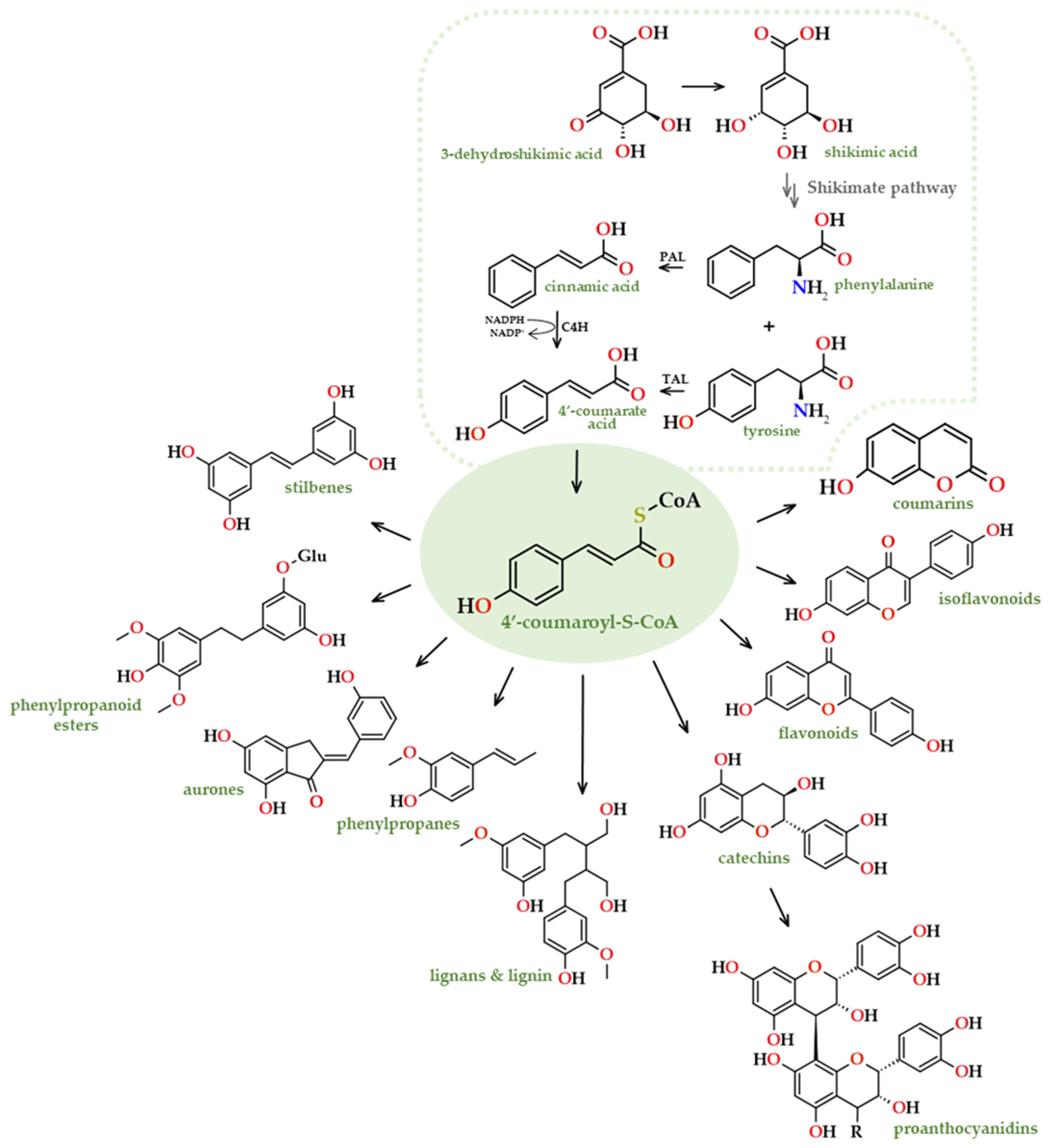
Figure 11.
Proposed radical mechanism for 7-hydroxycoumarin (umbelliferone) biosynthesis from 4'-coumaroyl-S-CoA (adapted from [102]). F6'H1, a 2-oxoglutarate-dependent dioxygenase, comprises the first step.
Figure 11.
Proposed radical mechanism for 7-hydroxycoumarin (umbelliferone) biosynthesis from 4'-coumaroyl-S-CoA (adapted from [102]). F6'H1, a 2-oxoglutarate-dependent dioxygenase, comprises the first step.
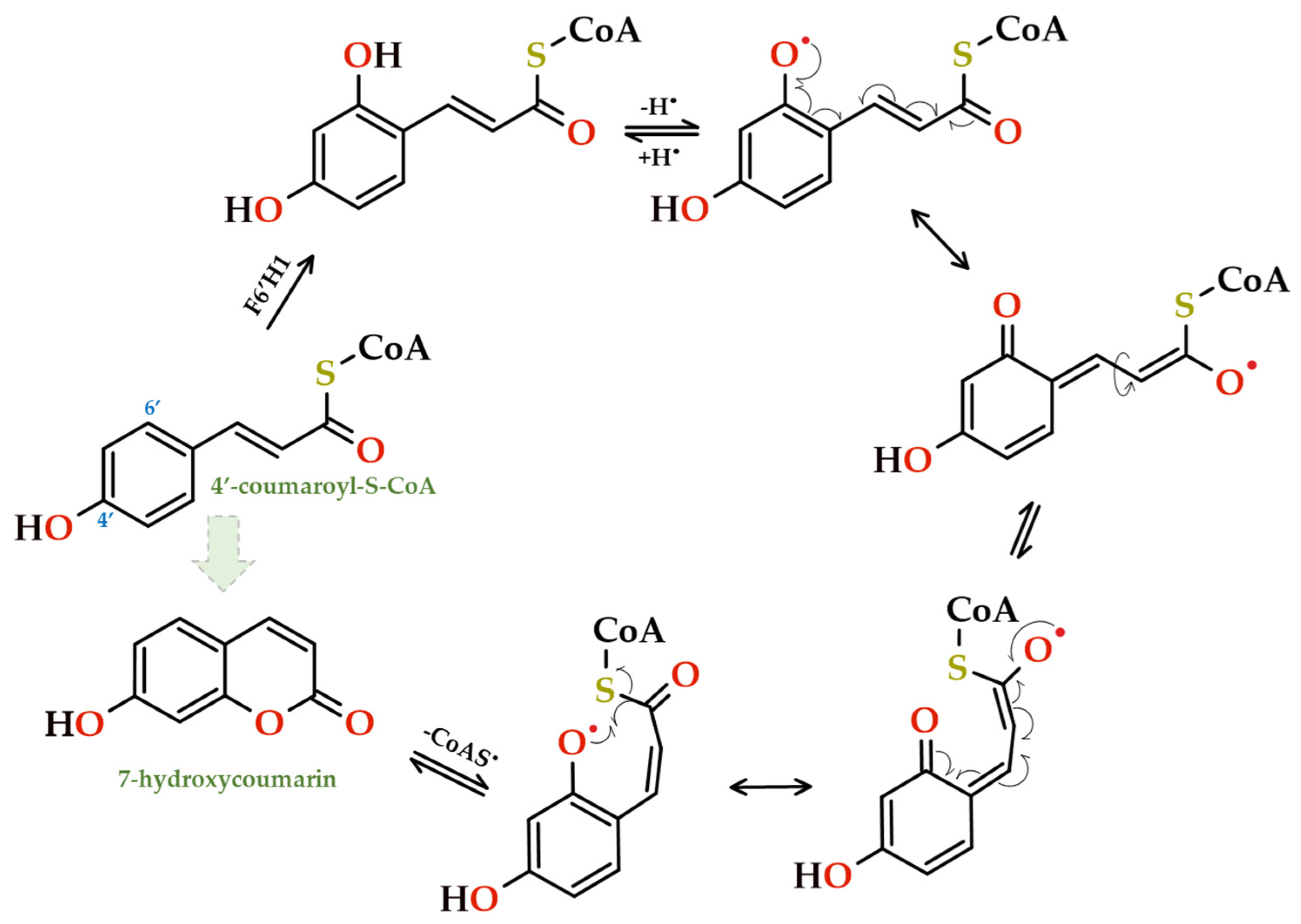
Figure 12.
Schematic illustration of the phenylpropanoid pathway leading to coumarins (particularly esculetin and scopoletin) and the formation of flavonoids and other phenolic compounds. PAL: l-phenylalanine ammonia-lyase.
Figure 12.
Schematic illustration of the phenylpropanoid pathway leading to coumarins (particularly esculetin and scopoletin) and the formation of flavonoids and other phenolic compounds. PAL: l-phenylalanine ammonia-lyase.
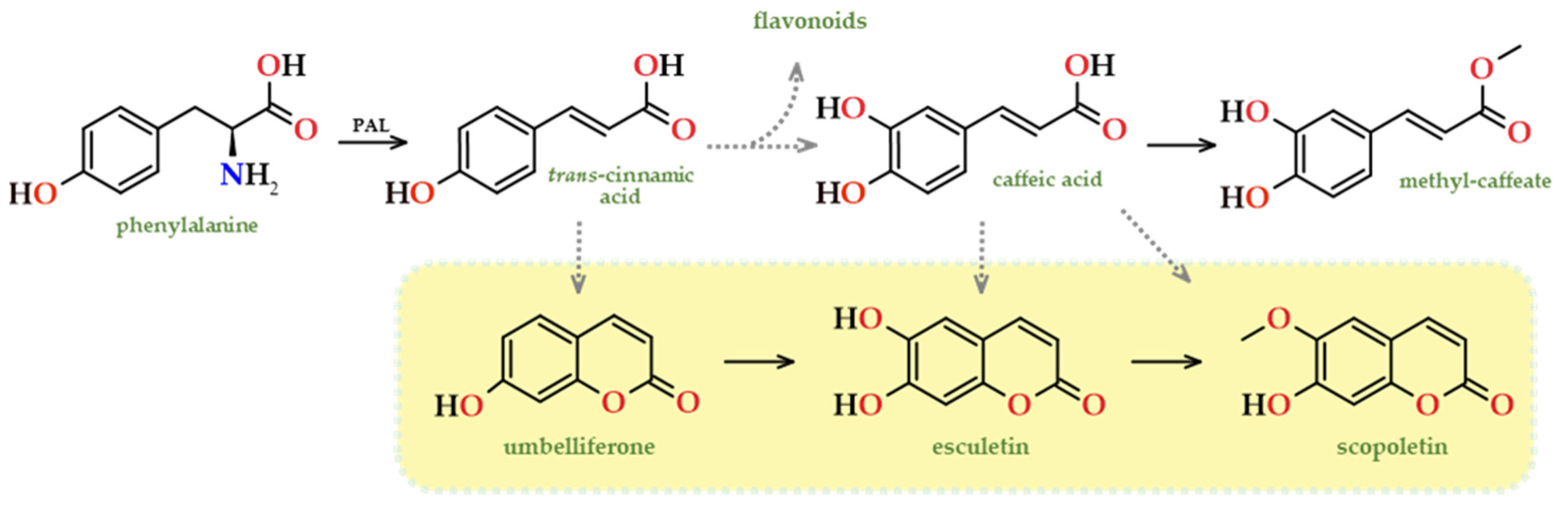
Figure 13.
Structures of dicoumarol.
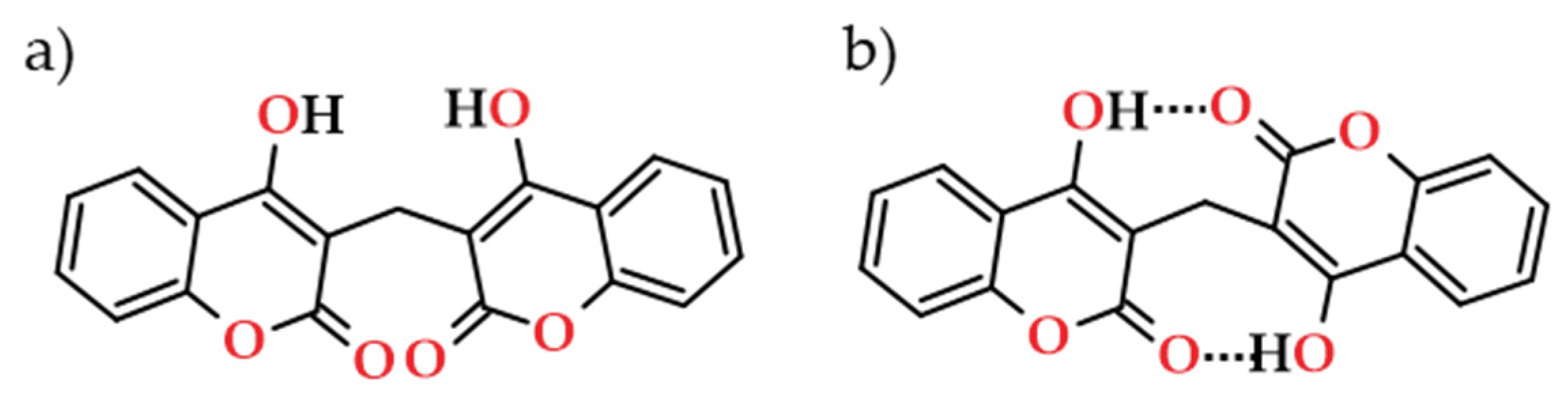
Figure 14.
Tautomeric structures of 4-hydroxycoumarin (A, B and C).
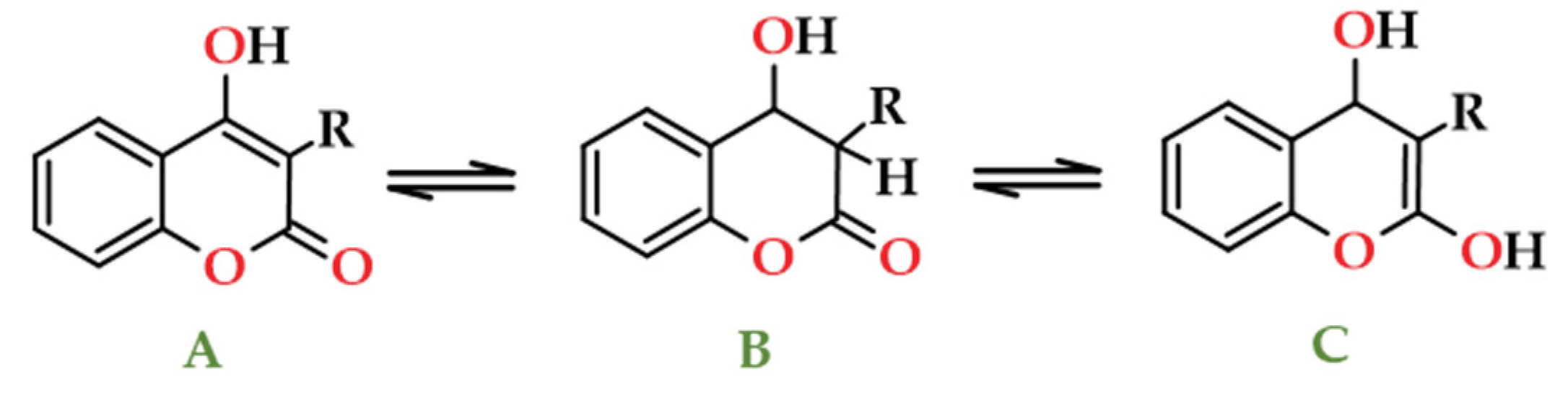
Figure 15.
Illustrative routes of coumarin metabolism (adapted from [111,112,113]. GSH: glutathione.
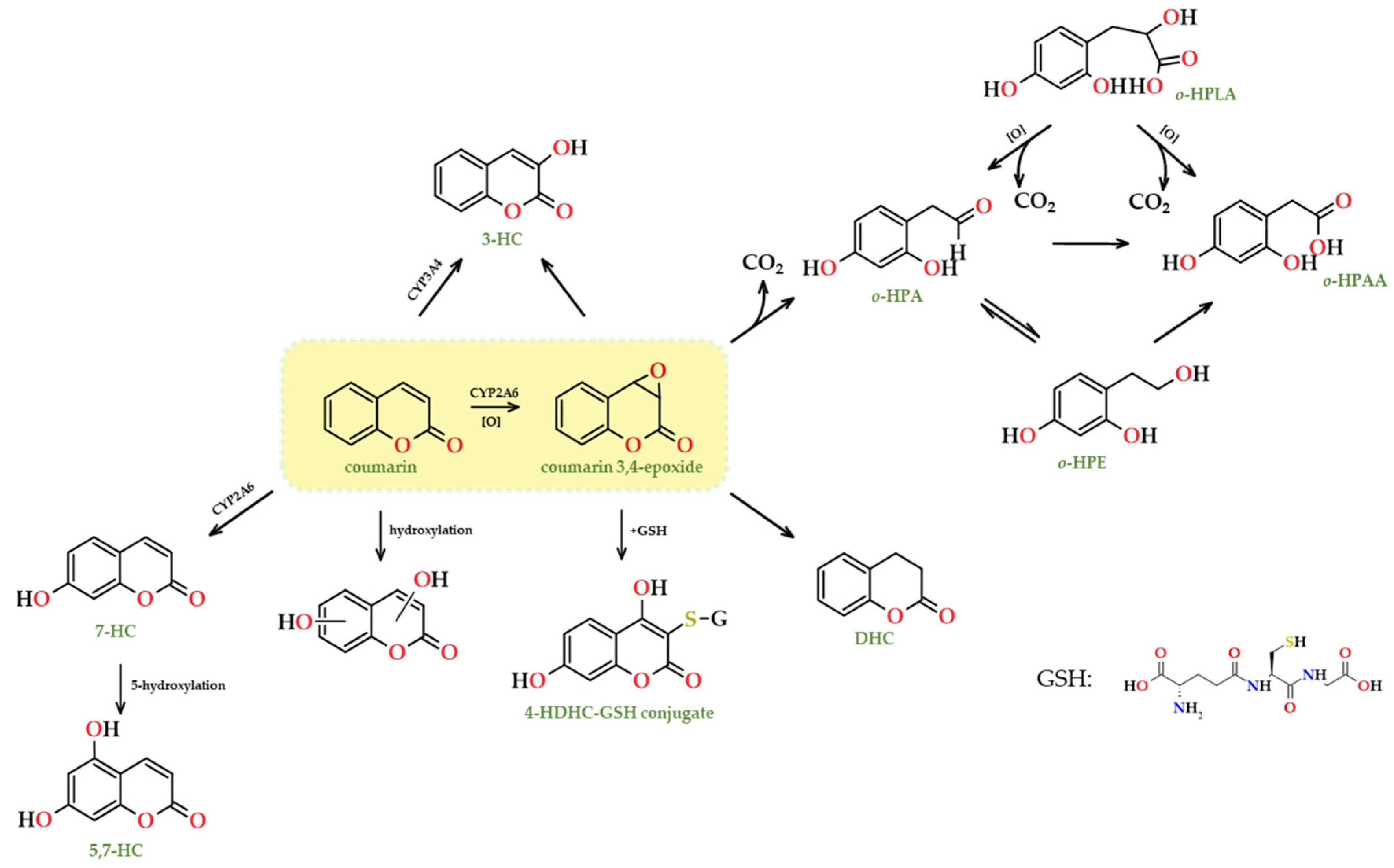
Figure 16.
Chemical structure of the most prevalent coumarins in wood used for the ageing of spirit beverages.
Figure 16.
Chemical structure of the most prevalent coumarins in wood used for the ageing of spirit beverages.
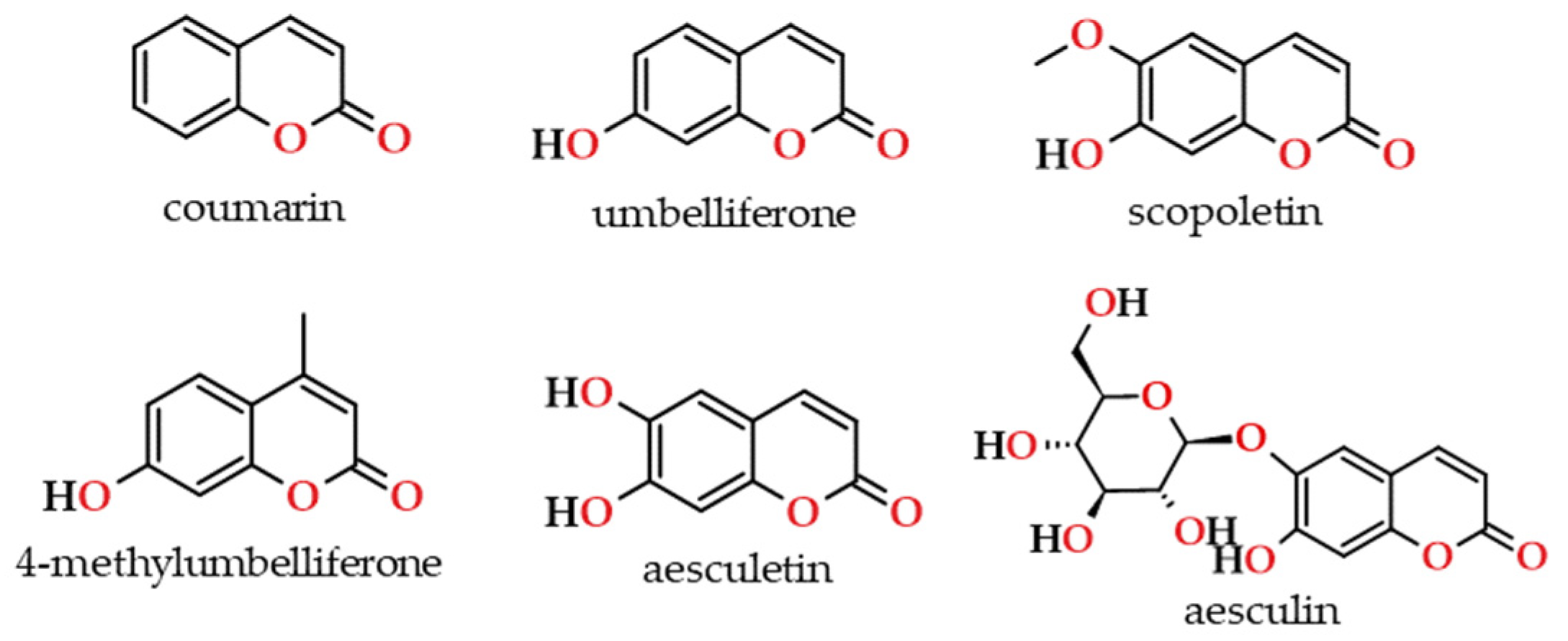
Figure 17.
Average concentrations of coumarins in aged wine spirits according to the toasting level of the barrels; LT - light toasting, MT - medium toasting, HT - heavy toasting [adapted from Canas 19].
Figure 17.
Average concentrations of coumarins in aged wine spirits according to the toasting level of the barrels; LT - light toasting, MT - medium toasting, HT - heavy toasting [adapted from Canas 19].
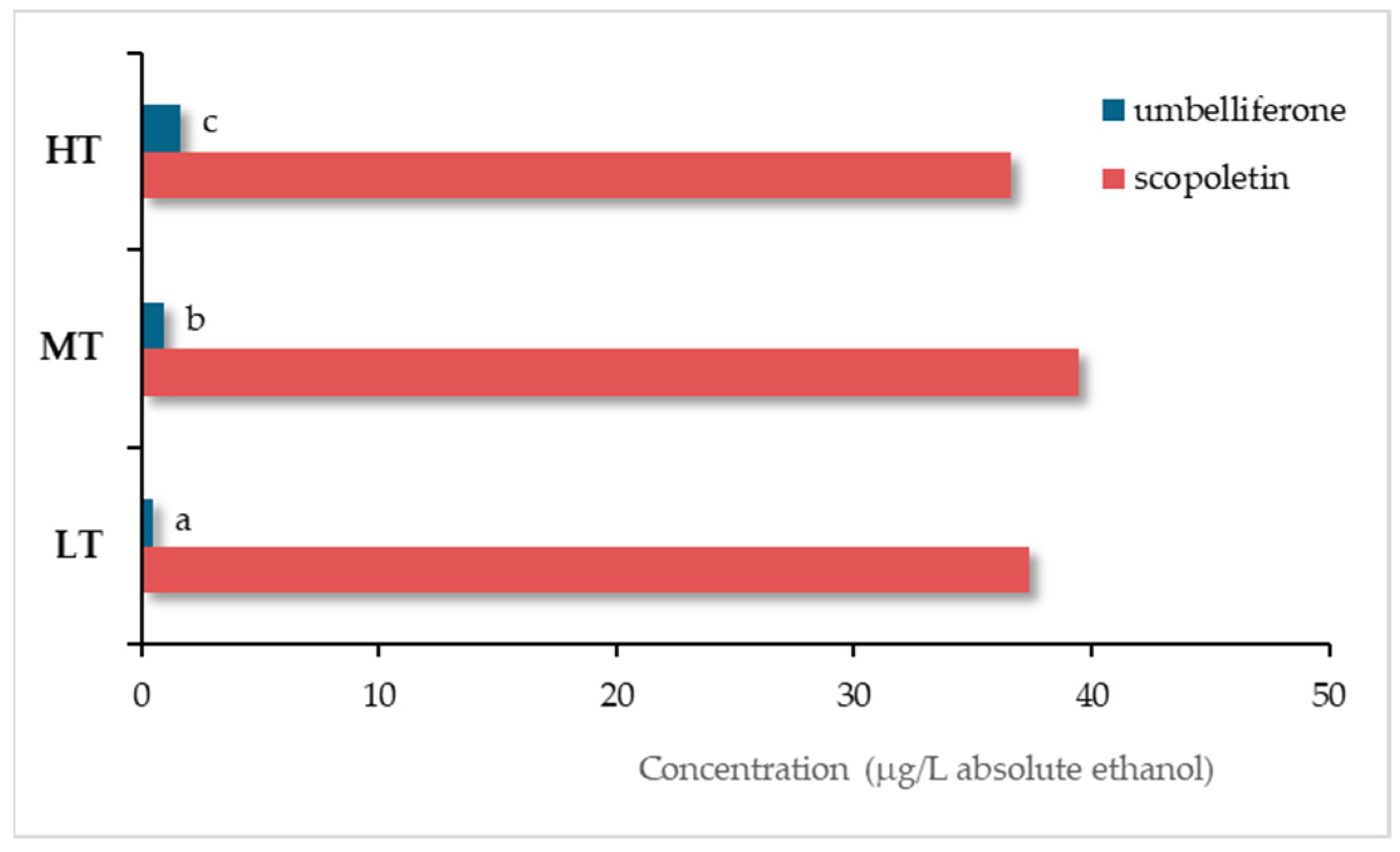
Figure 18.
Evolution of coumarins contents in spirit beverages over the ageing time: (a) whiskey [adapted from Otsuka and Zenibayaashi, [35]]; (b) wine spirit [adapted from Salagoity, [214]; (c) wine spirit [adapted from Tricard et al. [37]; (d) cachaça [adapted from Santiago et al. [44].
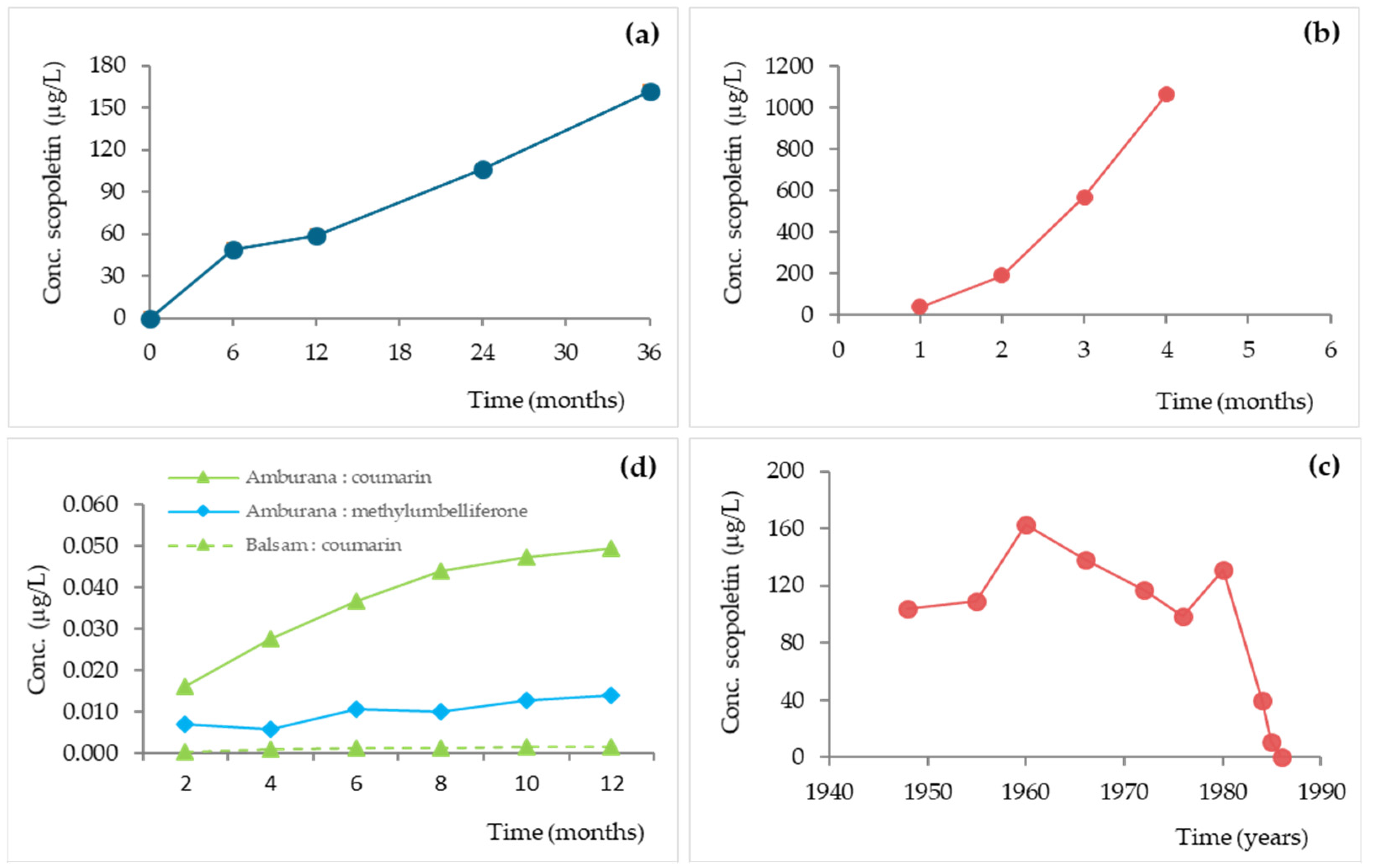
Figure 19.
Average contents of coumarins in wine spirits aged in barrels (B), in demijohns with wood staves and micro-oxygenation (O15, O30 and O60) and nitrogen (N, control).
Figure 19.
Average contents of coumarins in wine spirits aged in barrels (B), in demijohns with wood staves and micro-oxygenation (O15, O30 and O60) and nitrogen (N, control).
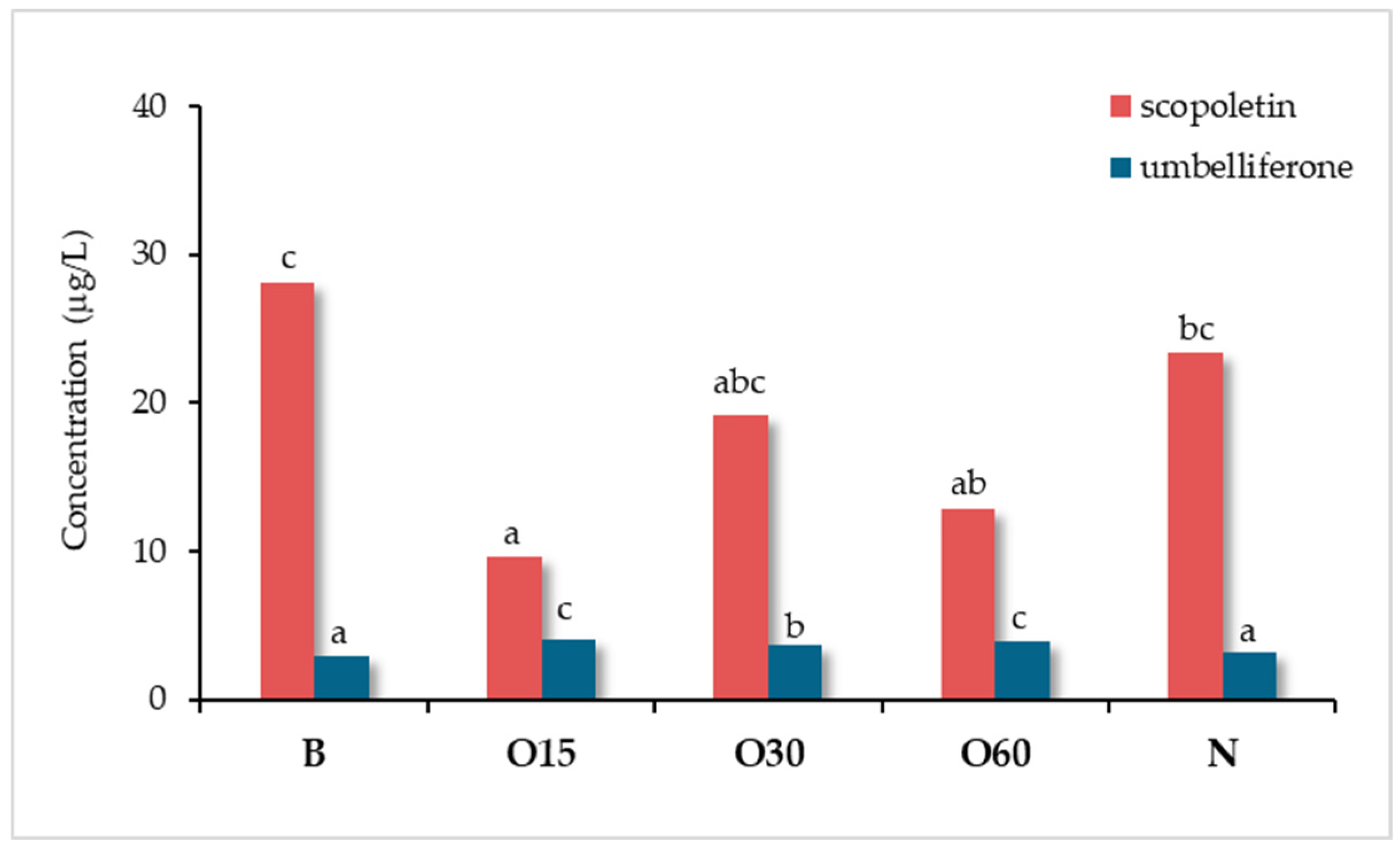
Figure 20.
Beverages inauthenticity events. Adapted from Popping [301].
Figure 20.
Beverages inauthenticity events. Adapted from Popping [301].
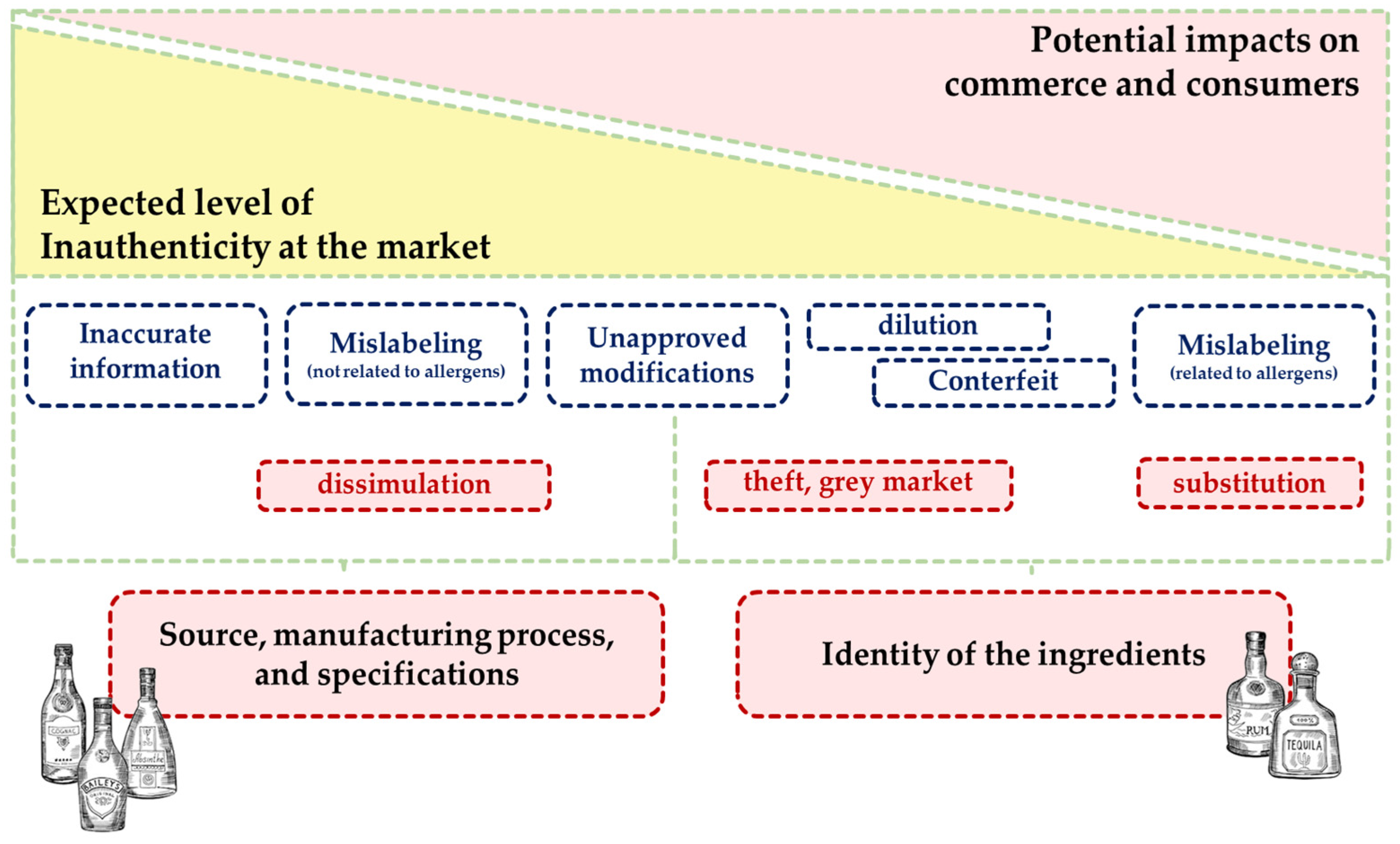

Table 1.
Sources of coumarins.
| Source | Species’ country of origin | Compound | Mean content | References |
|---|---|---|---|---|
| Oak Quercus robur L. |
France Limousin |
scopoletin umbelliferone aesculetin |
65 μg/L 4.7 μg/g FW 5.28 μg/g DW 0.006 μg/g DW 1.68 μg/g FW |
[38] [123] [124] [124] [138] |
| Bulgaria | scopoletin | 125 μg/L | [38] | |
| Spain | aesculetin | 2.1 μg/g FW | [138] | |
| Croatia | aesculin | 0.44 μg/g FW | [139] | |
| Serbia | aesculin | 0.22 μg/g FW | [139] | |
| Bosnia and Herzegovina | aesculin | 0.11 μg/g FW | [139] | |
| Oak Quercus sessiliflora Salisb. |
France Allier |
scopoletin umbelliferone aesculetin |
55 μg/L 3.6 μg/g FW 3.45 μg/g DW 0.006 μg/g DW 1.97 μg/g FW |
[38] [123] [124] [124] [138] |
| Spain | aesculetin | 4.2 μg/g FW | [138] | |
| Serbia | aesculin | 0.44 μg/g FW | [139] | |
| Portugal | scopoletin umbelliferone |
1.37 μg/g DW 0.006 μg/g DW |
[138] |
|
| Oak Quercus faginea Lam. |
Spain |
aesculetin |
4.9 μg/g FW |
[138] |
| Oak Quercus alba L., Quercus bicolor Willd. Quercus lyrata Walt. Quercus stellata Wanghen. |
North America | scopoletin umbelliferone aesculetin |
425 ug/L 25.15 ug/g DW 0.007-0.025 μg/g DW 1.45 μg/g FW |
[38] [124] [124] [138] |
| Chestnut Castanea sativa Mill. |
Portugal | scopoletin umbelliferone |
1.07 μg/g DW nd |
[124] |
| France | scopoletin | 6.73-16.7 μg/g FW | [141] | |
| Cherry Prunus avium L. |
Spain | scopoletin | 18.8 μg/g FW | [142] |
| Serbia | aesculetin | 102.30 μg/g FW | [139] | |
| Myrobalan plum Prunus cerasifera Ehrh. |
Serbia | aesculin | 1.87 μg/g FW | [139] |
| Mulberry Morus alba L. |
Serbia |
aesculin | 2.31 μg/g FW | [139] |
| aesculetin | 29.15 μg/g FW | [139] | ||
| Amendoim Pterogyne sp. |
Brazil | scopoletin coumarin |
0.007 mg/L 0.067 mg/L |
[43] |
| Cabreúva-parda Myrocarpus frondosus Allem. |
Brazil | scopoletin coumarin |
0.018 mg/L 0.050 mg/L |
[43] |
| Canela-Sassafrás Ocotea pretiosa (Vell.) Rohwer |
Brazil | scopoletin coumarin |
0.028 mg/L 0.011 mg/L |
[43] |
| Jatobá Hymenaea courbaril L. |
Brazil | scopoletin coumarin |
nd 9.15 mg/L |
[43] |
| Pequi Caryocar brasiliense Cambess. |
Brazil | scopoletin coumarin |
nd 0.071 mg/L |
[43] |
| Castanheira Bertholletia excelsa Humb. & Bonpl. |
Brazil | scopoletin coumarin |
nd 0.007 mg/L |
[43] |
1 DW - dry weight; FW - fresh weight; nd - not detected.
Table 2.
Excitation and emission wavelengths of prevalent coumarins in wood and aged spirit beverages.
Table 2.
Excitation and emission wavelengths of prevalent coumarins in wood and aged spirit beverages.
| Compound | λex (nm) | λem (nm) |
|---|---|---|
| 4-methylumbelliferone | 218 | 454 |
| coumarin | 280 | 393 |
| aesculin | 335 - 337 | 409 |
| umbelliferone | 340, 380 | 467 |
| scopoletin | 345 - 347 | 430, 460 |
| aesculetin | 347 - 353, 340 | 467 |
Table 3.
Sample preparations by spirit beverage.
| Spirit Beverage | Sample preparation | References |
|---|---|---|
| Wine Spirit | Coumarins should be extracted since this matrix is complex and rich in phenolic compounds. An ether extraction was therefore carried out | [36] |
| Samples directly analysed (without preparation) | [38] | |
| Adding an internal standard to the samples, filtering them through a 0.45 µm membrane and analysing them by direct injection | [39,40,152,164,165] | |
| De-alcoholised samples (to 8% alcohol) and filtered through a 0.45 µm membrane | [22] | |
| Brandy | Samples directly analysed (without preparation) | [38] |
| Samples filtered at 0.20 µm membrane and directly injected | [123] | |
| De-alcoholised samples (to 8% alcohol) and filtered through a 0.45 µm membrane | [22] | |
| Whiskey | Samples extracted with ethyl acetate | [35] |
| Samples directly analysed (without preparation) | [38] | |
| De-alcoholised samples (to 8% alcohol) and filtered through a 0.45 µm membrane | [22] | |
| Sugar cane spirit | Samples extracted with ethyl acetate (Rum) | [35] |
| SPE extraction (Cachaça) | [43] | |
| Samples filtered at 0.45 µm polyethylene membrane and directly injected | [166] | |
| For fluorescence detection, the samples were not prepared; for UV-vis detection, the samples were diluted 30 times with an ethanol/water solution (40:60 % v/v) (Cachaça) | [167] | |
| De-alcoholised samples (to 8% alcohol) and filtered through a 0.45 µm membrane (Rum) | [22] |
Table 4.
Analytical methods used for coumarins determination in spirit beverages.
| Spirit Beverage | Analytical methods | References | |
|---|---|---|---|
| Separation | Detection | ||
| Wine Spirit | HPLC | [36,38,39,40,152,164,165] | |
| HMRS with ESI/ HESI II | [22] | ||
| Brandy | HPLC | FLD | [38,123] |
| HMRS with ESI/ HESI II | [22] | ||
| Fluorescence Spectrometry | [48] | ||
| Whiskey | TLC | Fluorometer | [35] |
| HPLC | FLD | [38] | |
| HMRS with ESI/ HESI II | [22] | ||
| Sugar cane spirit | TLC | Fluorometer | [35] |
| HPLC | FLD | [43] | |
| HPLC | DAD | [166] | |
| UV-Vis spectrophotometry and spectrofluorimetry | [167] | ||
| HPLC | HMRS with ESI/ HESI II | [22] | |
HPLC: High-performance liquid chromatography; TLC: Thin-layer Chromatography; DAD: Diode Array Detector; FLD: Fluorescence Detector; HMRS: High-resolution mass spectrometry; ESI/HESI II: Heated electrospray ionisation.
Table 5.
Coumarins contents in wine spirits according to the kind of wood used in their ageing.
| Origin | Wood species | Compound | Mean content (μg/L) |
References |
|---|---|---|---|---|
| Armagnac | Oak | scopoletin umbelliferone 4-methylumbelliferone aesculetin |
301.1 2.8 2.8 1.9 |
[36] [36] [36] [36] |
| Lourinhã | Oak Q. robur |
scopoletin umbelliferone |
88.0 37.12 1.0 0.95 |
[39] [19] [39] [19] |
| Oak Q. sessiliflora |
scopoletin umbelliferone |
19.74 0.78 |
[19] [19] |
|
| Oak Q. pyrenaica |
scopoletin umbelliferone |
10.33 0.98 |
[19] [19] |
|
| Oak Q. alba Q. bicolor Q. lyrata Q. stellata |
scopoletin umbelliferone |
164.77 1.48 |
[19] [19] |
|
| Chestnut C. sativa |
scopoletin umbelliferone |
9.0 8.63 5.0 0.92 |
[39] [19] [39] [19] |
Table 6.
Coumarins contents in brandies according to the kind of wood used in their ageing.
| Origin | Wood species | Compound | Mean content (μg/L) |
References |
|---|---|---|---|---|
| Italy | Oak Q. robur |
scopoletin | 102.1 | [123] |
| Oak Q. sessiliflora |
scopoletin | 101.5 | [123] |
Table 7.
Coumarins contents in cachaças according to the kind of wood used in their ageing.
| Origin | Wood species | Compound | Mean content (μg/L) |
References |
|---|---|---|---|---|
| Brazil | European oak | 4-methylumbelliferone coumarin |
nd nd |
[44] |
| Amburana Amburana cearensis (Allemão) A.C.Sm. |
4-methylumbelliferone coumarin |
0.014 0.049 |
[44] | |
| Balsam Myroxylon peruiferum L. f. |
4-methylumbelliferone coumarin |
nd 0.002 |
[44] | |
| Jatoba Hymenaeae carbouril L. |
4-methylumbelliferone coumarin |
nd nd |
[44] | |
| Peroba Paratecoma peroba (Record) Kuhlm. |
4-methylumbelliferone coumarin |
nd nd |
[44] |
1 nd - not detected.
Table 8.
Coumarins and related pharmacological activities examined in vitro and in vivo studies.
| Compound | Pharmacological activity |
Research Method | Experimental | Doses | References |
|---|---|---|---|---|---|
| umbelliferone |
neuroprotective effects | chronic unpredictable mild stress (CUMS) | male Sprague-Dawley rats | 15 mg/kg 30 mg/kg |
[236] |
| anti-inflammatory | inflammatory cytokines in hippocampus | male Sprague-Dawley rats | 15 mg/kg 30 mg/kg |
[236] | |
| lipopolysaccharide (LPS)-induced acute lung injury (ALI) | BALB/c mice | 40 mg/kg |
[237] | ||
| antidepressant | post-traumatic stress disorder model (PTSD) | male Sprague-Dawley rats | 60 mg/kg | [238] | |
| antidiabetic | streptozotocin (STZ)-induced diabetes | male Wistar rats | 30 mg/kg | [239,240] | |
| anticancer | cell cycle analysis and apoptosis detection | HepG2 HCC cells |
50 μM | [241] | |
| antihypertensive | vascular activity assay | male spontaneously hypertensive rats (SHR) | 300 μM | [242] | |
|
4-methylumbelliferone |
anticancer | human pancreatic cancer cells (KP1-NL) transplanted into the hypodermis of nude mice | male nude mice | 3 mg/g | [243] |
| anticancer | breast cancer xenograft models | BALB/C nu/nu mice | 0.5 mg/g | [244] | |
| antitumor | inhibition of fibroblast and melanoma cells | human melanoma cell line, C8161 and MV3 | 0.5 mM | [245] | |
| anti-inflammatory | inflammatory cytokines in astrocytes cultures | rat astrocytes (glial cells) | 400 μM | [246] | |
| antidiabetic | glucose level and glucose tolerance test in mice fed a high-fat diet |
male C57BL/6J mice | 0.2 g/kg | [247] | |
| neuroprotective | acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) inhibition | in vitro assay | 1 mg/mL | [248] | |
|
aesculetin |
antiadipogenic | PPARg Expression | 3T3-L1 preadipocyte cells | 100 μM |
[249] |
| antidiabetic | streptozotocin (STZ)-induced diabetes | male albino rats (Wistar strain) | 40 mg/kg | [250] | |
| antithrombotic | platelet aggregation | human assay | 50 μM | [251] | |
| antitumor | mitogen-activated protein kinase (MAPK) assay | HepG2 cells | 100 μM |
[252] | |
| anticancer | trail-induced apoptosis | oral cancer SAS cells | 10 μg/mL | [253] | |
| anti-inflammatory | colitis induced by trinitrobenzenesulphonic acid (TNBS) | male Wistar rats | 5 mg/kg | [254] | |
|
aesculin |
antidiabetic | dexamethasone-induced insulin resistance | male Institute of Cancer Research (ICR) mice | 40 mg/kg | [255] |
| streptozotocin (STZ)-induced diabetes | male C57BL/6J | 20 mg/kg | [256] | ||
| renoprotective | STZ-induced diabetic renal damage | male ICR mice | 20 mg/kg | [257] | |
| gastroprotective | gastric mucosal injury index | male Kunming mice | 20 mg/kg | [258] | |
| antioxidant | AAPH-induced erythrocyte hemolysis assay | erythrocytes | 50 μM | [259] | |
| anti-inflammatory | carrageenan induced paw oedema | male and female Kunming mice | 20 mg/kg | [260] | |
| endotoxin shock induced by lipopolysaccharide (LPS) | male and female Kunming mice | 20 mg/kg | [261] | ||
| liver injury induced by lipopolysaccharide/D-Galactosamine (LPS/D-Gal) | BALB/c mice | 40 mg/kg | [262] | ||
| coumarin |
antidiabetic | animal model of induced type 2 diabetes mellitus (NIDDM) | male albino Wistar rats | 100 mg/kg | [263] |
| α-glucosidase inhibition assay | in vitro assay | 20 mM | [264] | ||
| anticancer | inhibition of proliferation and induction of apoptosis | human cervical cancer HeLa cells | 100 μM | [265] | |
| inhibition of proliferation | human lung cancer cell lines, A427 and Calu-1 | 1.0 mM | [266] | ||
| inhibition of proliferation | human lung cancer cell lines, A427 and SK-LU-1 | 100 μg/mL | [267] | ||
| anti-inflammatory | colonic damage induced by trinitrobenzenesulfonic acid (TNBS) | Male Wistar rats | 25 mg/kg | [268] | |
|
scopoletin |
antibacterial | minimum inhibitory concentrations (MIC) | Pseudomonas aeruginosa | 0.66 μg/mL | [269] |
| antihypertensive | multiple model of hypertension (blood pressure) | male Wistar-Kyoto rats | 10 mg/kg | [270] | |
| antidiabetic | streptozotocin (STZ)-induced diabetes | male ICR mice | 10 mg/kg | [271] | |
| α-glucosidase/α-amylase inhibition assay | in vitro assay | 100 μM | [271] | ||
| glucose uptake assay | 3T3-L1 preadipocyte cells | 20 μM | [272] | ||
| anti-inflammatory | carrageenan induced paw oedema | female Sprague-Dawley rats | 10 mg/kg | [273] | |
| anti-ageing | autophagy assay | human lung fibroblast cells (IMR 90) |
16 μM | [274] |
Table 9.
Bioaccumulative potentiala and toxicological information of most prevalent coumarins in wood and aged spirit beverages.
Table 9.
Bioaccumulative potentiala and toxicological information of most prevalent coumarins in wood and aged spirit beverages.
| Compound | n-octanol/water (log KOW)b | Test Type/Routec | Organism | Dose | References | Remarks |
|---|---|---|---|---|---|---|
| coumarin | 1.39 | LD50 / oral | rat | 293 mg/kg | [287] | May cause irritation; Harmful if swallowed: causes liver injury and somnolence |
| LD50 / oral | mouse | 196 mg/kg | [288] | |||
| LD50 / intraperitoneal | mouse | 220 mg/kg | [289] | |||
| LD50 / subcutaneous | mouse | 242 mg/kg | [288] | |||
| LD50 / oral | guinea pig | 202 mg/kg | [290] | |||
| umbelliferone | 1.03 (TOXNET) | LD50 / intravenous | mouse | 450 mg/kg | [291] | Causes liver effects, effects on plasma proteins, hypoglycemia, and changes in clotting factors in 45-day intermittent intraperitoneal studies of rats |
| scopoletin | nd | LD50 / oral | rat | 3800 mg/kg | [292] | Irritant |
| 4-methylumbelliferone | nd | LD50 / intravenous | mouse | 350 mg/kg | [291] | Irritant |
| LD50 / oral | rat | 3850 mg/kg | [293] | |||
| LD50 / intraperitoneal | rat | 2550 mg/kg | [293] | |||
| LD50 / subcutaneous | rat | 7200 mg/kg | [293] | |||
| LD50 / oral | mouse | 2850 mg/kg | [293] | |||
| LD50 / intraperitoneal | mouse | 1250 mg/kg | [294] | |||
| aesculetin | 0.55 | LD50 / intraperitoneal | mouse | 1500 mg/kg | [295] | Irritant |
| aesculin | nd | LD50 / intraperitoneal | mouse | 1900 mg/kg | [296] | Irritant |
a Data collected from the safety data sheets available from Thermo Fisher Scientific, Sigma-aldrich, and Carl Roth, and also from PubChem. b (pH value: 7 at 25 °C). c The LD50 (lethal dose, 50%) is a toxicological metric that expresses the dosage of a drug that is deadly to 50% of a population (usually laboratory animals) exposed to it. Parameters in the European Union toxicity classification Oral LD50 for rats: Category 1: Very Toxic: LD50 ≤ 5 milligrams per kilogram of body weight (mg/kg); Category 2: Toxic: 5 mg/kg < LD50 ≤ 50 mg/kg; Category 3: Harmful: 50 mg/kg < LD50 ≤ 300 mg/kg; Category 4: Warning: 300 mg/kg < LD50 ≤ 2000 mg/kg [297].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



