
Submitted:
02 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
This study explored techniques with enteric methane (CH4) abating potential, ranging from chemical additives including 3-NOP, nitrate, and halogenated compounds to natural feed additives such as tannins, oils, seaweed extracts, and probiotics. Most of the evaluated strategies exert their influence either by promoting the growth of ruminal microbes that compete with methanogens for metabolic hydrogen as their resource or by directly consuming CH4. Additionally, externally supplied probiotics, including propionate-forming bacteria and methane-oxidizing bacteria, are a promising tool. However, challenges persist in implementing these strategies effectively. High doses of chemical additives and plant based secondary metabolites often achieve significant CH4 reduction, but they are associated with negative impact on dry matter intake and digestibility, thus compromising overall animal performance. On the other hand, using lower doses to maintain animal performance may not achieve beneficial CH4 reduction. Furthermore, concerns about the impact of certain additives on human health have led to a shift towards more natural alternatives, but their efficacy and feasibility in vivo necessitate further evaluation. Comprehensive research that integrates data from diverse global experiments on ruminants through meta-analytical approaches is essential. Such research will help in identify strategies that either reduce methane emissions and/or enhance production performance, ensuring sustainable livestock farming practices. In conclusion, mitigating CH4 emissions from ruminant livestock is a complex challenge with wide-ranging implication.
Keywords:
ruminants methane grazing
Introduction
Current scenarios of global warming show strong evidence of more erratic fluctuations in weather patterns in the future as compared to the past and present, and this is attributable to the current agricultural and industrial practices that generate high levels of greenhouse gases into the atmosphere (Horrigan et al, 2022). About one-third of the total anthropogenic methane (CH4) emissions in the atmosphere are from the microbial fermentation in ruminants (Akapali, 2018). Strategies with CH4 reducing potential while at least maintaining or improving production performance necessitate assertation. Therefore, the focus of this review is to evaluate the overview of rumen ecology, ruminal microbiota in relation to strategies with CH4 abatement potential (chemical additives including 3-NOP, nitrate, and halogenated compounds to natural feed additives such as tannins, oils, seaweed extracts, and probiotics), CH4 modulation by such strategies and effect on animal productivity.
Overview of rumen ecology
The rumen ecosystem comprises a rich consortium microbiota exhibiting symbiotic and parasitic relationships. The isolation and analysis of ruminal microbiota showed the presence of bacteria species (1010 to 1011cells/ml, over 50 genera), anaerobic fungi (103 to 105 zoospores/ml, over five genera), and protozoa (104 to 106cells/ml, over 25 genera) in rumen fluid sample (Cieslak et al., 2013; Adejoro, 2019) which are responsible for degradation of the digesta, and production of energy for the host animal from the volatile fatty acids (Mizhari and Jami, 2018). The rumen washout aids the translocation of microbes to the lower gut where they get digested and provide microbial protein to the host animal. The concentration and distribution of the various rumen microorganisms is hugely influenced by diet. Plant polysaccharides such as cellulose, hemicellulose, and starch are broken down by enzymes and ruminal microbes into principal components of utilization. By-products from hydrolysis of the pentose and hexose sugars are utilized by succinate, lactate and furamate intermediates to generate volatile fatty acids (VFAs), which are then absorbed or deaminated in the ruminant system. Equally, protein and lipids go through a redox reaction in the rumen as depicted in Figure 1 and discussed below.
Bacteria is the dominant group in the ruminal microbiota and the major rumen fibre degrading (fibrolytic) bacteria species include, fibrobacter succinogens, Ruminococcus albus, and Ruminococcus flavefaciens etc. (Russel, 2002; Adejoro, 2019). Equally, structural carbohydrate degrading bacteria species such as Butyrivibros fibrisolvens, and Eubacterium cellulosolvens have been reported in the rumen and contain extracellular cellulase with minute cellulolytic efficacy and these bacteria species should be attached to the substrate to carry out the cellulolytic activity owing to their enzymes being retained in their cell wall (Shreck, 2013). Protein degrading bacteria species include Streptococcus bovis, Bacteroides amylophilus, and Bacteroides fibrisolvens, whereas lipid associated species include Anaerovibrio lypotica. On the other hand, amylolytic bacteria species include Bacteroides ruminicola, and Bacteroides amylophilus (Krause et al., 2003). Homoacetogens are acetate-producing bacterial group with 23 diverse bacterial genera and are present in the rumen with concentration ranging from 107 to 108 cells/g (Drake et al., 2007). These can grow heterotrophically by utilising sugars and autotrophically by utilising metabolic hydrogen and carbon dioxide in the rumen (Fonty et al., 2007). Autotrophic growth of these bacterial species is catalysed by a hydrogenase enzyme through the Wood-Ljungdhal pathway (Ragsdale and Pierce, 2008)
Anaerobic fungi make up about 10% of the microbiota (Rezaeina et al., 2004), and attach to the plant tissue where they degrade about 34% of the lignified fraction that is indigestible to other microorganisms in the rumen (Lowe et al., 1987). The persistency and abundance of anaerobic fungi is less than that of bacteria because the growth rate of fungi is much slower than the ruminal dilution rate (Krause et al., 2003). Anaerobic fungi are characterised by broad range of highly active enzymes, filamentous growth, and exo-acting cellulase activity. This class of ruminal microorganisms include Neocallimastix frontalis, Caecomycens (previously named Sphaeromonas) communis, and Piromycens (previously named Priromonas) communis. Moreover, Neocallimastix frontalis are the well-studied and notable exceedingly active fungal species against crystalline cellulose (Nagpal et al., 2004). Howsoever, the abundance and activity of anaerobic fungi is regulated to a certain extent by some bacteria and protozoa species in the rumen. The solubility of crystalline cellulose, release of endoglucanase, and production of short chain fatty acids was reduced when chitinolytic Clostridium tertium was co-cultured with anaerobic fungi (Hodrová et al., 1995). Also, when anaerobic fungi were incubated with protozoa, it was evident that protozoa are capable to prey and digest fungi (Morgavi et al., 1994). In this view, the presence of some protozoal and bacterial species limit fungi to exhibit their full lignified plant tissue degradation potential.
Of the two existing subclasses of protozoal population in the rumen, only the ciliates are found in the rumen contents of adult animals as the abundance of flagellates diminishes as animal grows. Rumen fluid collected from defaunated sheep revealed an increase in numbers of Methanovebribacter, and Selenomonas ruminantiun species (Newbold, 1996). Similarly, Lactobacillus plantarum vanished in the rumen due to protozoa predation (Sharp et al., 1994). Therefore, it is prudent to conclusively state that protozoa have the substantial capacity to engulf bacteria as their nitrogen source. Rumen fluid dilution rate is faster than the protozoal population outflow and this renders protozoa as microorganisms with adverse effect to the ruminant host animal due to the outflow of bacteria to the lower gut being reduced, thus a substantial setback in total nitrogen recycling. Howsoever, the authors (Devillard et al., 2003) postulated that even though not well understood, the protozoa group may have the capacity to digest fibrous fraction of plant tissue in the rumen.
Impacts of methane reducing strategies on ruminal microbiota
The diversity of microbiomes in the rumen is the key element making the ruminants to adapt in a vast array of dietary conditions (McCann et al., 2014). The research on the concept of host animal’s individual microbiome has gained interest due to the demonstrated influence of the diversity and abundance of these communities on the rumen fermentation pathways associated with enteric CH4 emission (Jami and Mizhari, 2012; Henderson et al., 2015; Weimer, 2015). To examine the rumen ecology, the fundamental use of 16S ribosomal RNA (16S rRNA) as a genetic marker is used, allowing for the identification and quantification of different microbial communities present in the rumen, given that 16S rRNA is a type of ribosomal RNA found in prokyaryotic cells (bacteria and archaea). Furthermore, other techniques include but are not limited to; metagenomic sequencing, quantitative polymerase chain reaction, fluorescence in situ hybridisation, microbiota cutting, next generation gene sequencing and bioinformatics analysis.e.tc.
Dietary additives
Tannins
Tannins exert intricate and diverse effects on the microbial populations residing within the digestive systems of ruminants. These effects are depending upon the specific source of tannins or dietary components rich in tannins. For example, in a study of Min et al. (2014) involving goats, the consumption of pine bark (PB) powder led to an increased relative abundance of Clostridia populations compared to the control diet. Conversely, the population of Bacteroidetes, decreased under the same conditions. Smith et al. (2003) revealed that enhancing the oxidative stress response in Escherichia coli allowed these bacteria to counteract the inhibitory effects of condensed tannins. Tabacco et al. (2006) demonstrated the complex nature of tannins, showing that chestnut tannins improved fermentation quality, proteolysis, and the degradation of protein in alfalfa silage. Beyond abundance shifts, tannins also exert influence on the diversity and composition of rumen microbial communities. Terré et al. (2016) found that the introduction of quebracho tannins into the diets of dairy cows resulted in a reduction in the diversity of rumen bacterial communities. Similarly, Patra and Saxena (2011) observed a reduction in the population of methanogenic archaea in sheep rumens when tannins were included in their diets. Furthermore, tannins play a role in regulating the activity and functions of microbial communities. A study by Min et al. (2018) demonstrated that the inclusion of tannins in the diets of goats led to a reduction in the activity of cellulolytic bacteria, resulting in decreased digestibility of fibre. In contrast, Wang et al. (2017) reported an increase in the activity of proteolytic bacteria in sheep rumens due to tannin inclusion, leading to improved utilization of protein and retention of nitrogen. The influence of tannins on rumen microbial communities is a complex phenomenon influenced by various factors such as the origin and concentration of tannins, dietary composition, and the specific microbial species present. This complexity highlights the need for a comprehensive understanding when considering the utilization of tannins to modulate microbial dynamics in the digestive systems of ruminants.
Oils
The study by Gruninger et al. (2021) reported a decrease in both the amplified sequence variants and phylogenic diversity in the rumen fluid collected at and analysed for different time intervals in response to oil inclusion in the diets of Angus beef heifers. Chestnut oil inclusion in the diet of sheep reduced protozoa microbial populations (Liu et al., 2011). Linseed reduced the abundance of Ruminococcaceae in a study of Popova et al. (2019). The study of Guyander et al. (2015) reported an increase in Selenomonas abundance in dairy cows fed linseed. These studies agree with the notion stating that fatty acids are toxic to cellulolytic microbes while in favour of succinate reducing microbes to aid the formation of propionate in ruminants (Zened et al.,2012; Maia et al., 2007). This is an indication that the addition of oil treatment into the diet of ruminants modulates the rumen microbiota favouring the proliferation of members of the order Selenomonadales (propionate producers; metabolic hydrogen consumers) while reducing cellulolytic Ruminococcaceae (hydrogen producers). Therefore, the supplementation of oils facilitates the diversion of hydrogen from the methane formation pathways to the pathway forming propionate the percussor of glucose in ruminants.
Saponin
The findings of Patra and Yu (2013) revealed in vitro potency of Quillaja saponin in reducing protozoal counts. These findings from Patra and Yu (2013) are in accord with other studies including an in vitro study of Guo et al., (2010), and in vivo studies in sheep (Zhou et al., 2011) and goats (Mao et al., 2010) where tea seed saponin inhibited protozoal growth. Similarly, the use of Yucca shidigera saponins had the inhibitory effect in ruminal ciliate protozoa in vitro (Pen et al., 2006), sheep (Pen et al., 2007), dairy cattle (Lovett et al., 2006), and beef cattle (Lila et al., 2005). However, other studies reported no effect of Yucca shidigera saponins on protozoal count in vivo (Singer et al., 2008; Holtshausen et al., 2009). The study of Ramírez-Restrepo et al. (2016) reported a decrease in abundance of R. flavefaciens, whereas F. succinogens and R. albus increased in steers fed tea seed saponin. The effect of saponin on the ruminal microbiome from the currently reviewed studies represent a complex picture as there are conflicting results. However, it is clear that fundamentally saponins reduce protozoal counts and their effects are influenced by type of saponin, animal species and microbial species targeted.
Nitrate
When nitrate was added to diets of nitrate-adapted steers there was an increase in abundance of nitrate reducing bacteria including C. fetus, S. ruminantium, and M. succiniproducens (Zhao et al., 2015). Similarly, Lin et al. (2013) reported high abundance of S. ruminantium even though it was 0.55% higher than the study of Zhao et al. (2015), which could be attributed to dietary and animal differences between the studies. The supplementation of nitrate increased the abundance of orders Coriobacteriales and Burkholderiales (Ashida et al., 2010). These orders contain taxa which is known to exhibit nitrate reducing capacity and these are in line with high nitrite concentration in nitrate-fed dairy cows in the study of Guyader et al. (2015). The supply of nitrate to ruminants exerts its influence on microbiota through being an alternative hydrogen sink. This modulates methanogenesis and/or dihydride which are energetically costly to the animal towards production pertaining metabolic hydrogen reductants. The translocation of metabolic hydrogen towards the reduction of nitrate is beneficial for ruminants, but the process is regulated by the ruminal conditions including microbiota abundance and composition, to name yet a few.
3-. Nitroxy propanol
There were no changes on the population of methanogens in response to supplementation of 3-NOP to diets of sheep (Martínez-Fernández et al., 2014) and beef cattle (Romero-Perez et al., 2014). However, the compound decreased methanogen population in dairy cows (Haisan et al., 2014). Also, the abundance of Methanobrevibacter genus (hydrogenotropic methanogens) and Methanomassiliicoccaceae family (methylotropic methanogens) were reduced by 3-NOP in a study of Martinez-Fernandez et al. (2018). Furthermore, in a similar study the ratio of Methanobrevibacter genus to both Methanomassiliicoccaceae and Methanosphaera was reduced in response to the dose supply of 3-NOP to beef cattle, suggesting a greater effect of 3-NOP in hydrogenotropic methanogens. The amount of metabolic hydrogen released does not increase in 3-NOP treated groups compared to the control groups (Haisan et al., 2014; Martinez-Fernandez et al., 2018). This could be an indication of that the substantial concentration of hydrogen in the rumen of animals subjected to 3-NOP is redirected to other reduced end products other than CH4 and H2, such as microbial cell mass (Ungerfeld, 2015).
Seaweed
There is scanty literature reporting the use of genomics and metagenomics approaches to examine the effects of seaweed on the composition, abundance, and function of the ruminal microbiome (Abbott et al., 2020). When Ascophyllum nodosum was added to the Rusitec in vitro fermentation system there were noticeable decrease in growth of Fibrobacter succinogens while Prevottella bryantii increased (Wang et al., 2009). The abundance of Methanobrevibacter reduced in a Rusitec in vitro fermentation system in a study of Abbott et al. (2020) and there was a shift in dominance towards Methanomicrobium on day 13 of the in vitro trial evaluating the effect of seaweed on ruminal microbiome. Dietary administration of seaweed led to a decrease in the levels of Butyrivibrio fibrisolvens, Butyrivibrio proteoclasticus, and Prevotella ruminicola, while Anaerovibrio lipolytica increased in abundance (Choi et al., 2021). Roque et al. (2019) reported that Asparagopsis taxiformis and other studies evaluating other seaweeds reported similar trend of modulating the metabolic functionality of the methanogens and abundance. Since knowledge about the ruminal microbiome's response to seaweeds is still in its early stages, metagenomic studies are imperative to gain insight into whether the effects of these seaweeds could be manipulated to improve the productivity and health of ruminants.
Dietary manipulation
The 16S rRNA gene libraries in the study of Fernando et al. (2010) revealed two distinct microbial populations in beef steers: one group fed roughage-based diet alone, and the other supplemented with concentrate diet. Furthermore, the researchers reported only 24 common operational taxonomic units (OUTs) found between the rumen contents of steers assigned to two diets out of 398 and 315 OUTs in roughage based and concentrate-supplemented steers, respectively. Consequently, steers fed roughage-based diet had a higher abundance of Fibrobacteres, while concentrate-supplemented steers had a higher number of Bacteroidetes. Metzler-Zebel et al. (2013) found that concentrate supplementation of up to 60% DM in the diet fed to goats reduced the abundance of Prevotella and Fibrobacter succinogens, while the abundance of Lactobacillus species increased.
Mitigating enteric methane emission
There is a global interest on finding effective solutions to ameliorate enteric CH4 emission arising from ruminant livestock. Hence, extensive research has been conducted to evaluate various approaches to mitigate enteric methane emissions. Notwithstanding numerous strategies previously demonstrating successful evidence of efficacy towards reducing enteric CH4 emission in conjunction with improved animal productivity, most of these strategies are allied with costs and pose both environmental and human health risks. Therefore, it is imperative to understand existing strategies to facilitate creation of better and efficient solutions towards the curb of CH4 emission from ruminants.
Plant-based antimethanogenic compounds
Tannins
Tannins are plant secondary metabolites having pH-dependent binding effects with the substrate (proteins) in the rumen (Poudel et al., 2023). There are two forms of tannins based on the molecular weight, which are hydrolysable (HT), and condensed tannins (CT). In most forage species commonly fed to ruminants, HT are less common compared to CT (Aboagye and Beauchemin, 2019). The source, molecular weight and structure of the tannins are factors influencing their reactivity and impact on digestibility in ruminants, which are extensively reviewed elsewhere (Naumann et al., 2017). Table 1 shows summarised effects of HT and CT from various plats including Schinopsis lorentzii, Schinopsis balansae, Prosopis cineraria, Quercus robur, Ficus infectoria, Psidium guajava, Acacia mearnsii, Castanea sativa, Vitis vinifera, Onobrychis viciifolia, Schinopsis balansae, and Tea waste on dry matter digestibility, nitrogen balance and CH4 production in ruminant livestock.
Tannin-protein binding complexes reduce ruminal digestion of protein thereby improving protein bypass in the host animal. Yanza et al. (2021) postulated that the bypass protein is not absorbed in the small intestine because protein-tannin molecule bonds are resistant to enzymes in the small intestines and therefore it is directly excreted in faeces. In consistency with such, in the study by Dschaak et al. (2010), milk urea nitrogen (N) was reduced, whereas the concentration of faecal N increased, which is partly explained by the poor absorption of protein in the small intestine due to tannin supply in dairy cows. Moreover, Orlandi et al. (2015) reported a linear increase in N retention, and faecal N concentrations while ammonia concentrations in the rumen decreased in steers supplemented with Acarcia mearnsi. Similarly, the findings from Wischer et al. (2014) revealed an increase in faecal N concentration in sheep supplemented with tannin from valonia and chestnut. These studies are in congruent with the observation of increased faecal N content in relation to tannin supply among studies summarized in Table 1. Also, methane and ammonia production were reduced in response to tannin inclusion in diets fed to Polish Holstein-Frisian dairy cows in a study by Cieslak et al. (2012). Therefore, it is noticeable with caution underlying the low dose, that tannins improve nitrogen efficiency and alleviates ammonia toxicity incidents in ruminants. Authors (Silanikove et al., 2001; Woodward et al., 2004; Lima et al., 2019) have reported that tannins have a toxic effect on ruminal microorganism, particularly fibrolytic bacterial species thereby adversely affecting fibre degradation in the rumen which is evident in reduced dry matter digestibility in Table 1. In view of tannin type (CT and HT), the CT have the greatest capacity to reduce ruminal degradability compared to HT owing to CT having strong affinity for particulate phase of digesta and is inherently resistant to hydrolysis, whereas HT is easily degraded by the ruminal microbes (Yanza et al., 2021). Howsoever, protein bypass, and the reduction of enteric gaseous emissions to the environment in response to tannin supply are noticeable benefits in ruminants (Hassanpour et al., 2011).
Terpenes
The terpene group is one of the most diverse natural products (Tiwari and Rana 2015). The group has a 5-carbon chain length with the biological activity in ruminants ranging from herbivory tolerance to ruminal microbiota suppression. The ethnoveterinary benefits of terpenes include the control of infections caused by fungi, bacteria arthropods, and gastrointestinal nematodes (Silver et al., 2020). Ghamkhar et al. (2018) reported that terpenes reduce enteric methane emission in vitro. Equally, the inclusion of terpen-based Pinus densiflora (50 mg/L) and Mentha canadensis (70 mg/L) extracts in diets evaluated for methane emission in vitro using rumen fluid from steers offered Timothy hay and commercial concentrate at 60:40 forage to concentrate ratio, showed a decrease on CH4 production (Lee et al. 2020). Furthermore, it was conclusively advised that the inclusion level threshold of terpenes on ruminant diets should be 30 mg/L (Lee et al., 2020). However, in vivo studies using terpens as CH4 mitigating strategy is scarce. Therefore, In vivo studies of terpenes need to expand to realize the potential of terpenes in alleviating CH4 emissions in ruminants.
Oils
The study of Chillard et al. (2009) show a reduction in dry matter intake (DMI) in response to linseed oil inclusion in lactating dairy cows. Moreover, such was reflected on the suppression of milk yield by 15% in the linseed compared to the control treatment. Also, milk fat content was lower for a diet containing the linseed oil compared to other treatments, such would be due to the oils supressing DMI owing to reduced rumen degradability and passage rate. The reduction in ruminal degradability could be possibly attributed to oils coating feed particles and creating a physical barrier that reduces microbial attachment to the digesta in the rumen. Such ultimately influences the rumen fill which is the physical distension of the rumen and is implicated to regulate dry matter intake. Therefore, it could be possible that dry matter intake was supressed through the slow passage clearance owing to reduced degradability. In a study with non-lactating Holstein dairy cows, linseed oil inclusion reduced CH4 emission by 22% compared to the control treatment and by 5% more than the nitrate fed group. When Sunflower oil was added to high versus low roughage-based rations of dairy cattle in the study by Bayat et al. (2017), the methane emission (g/d and g/kg) was reduced, respectively. Also, the dry matter intake was noticeable lower in lactating dairy cows fed high roughage-based diet compared to the other group (Bayat et al., 2017). The observed reduced dry matter intake in lactating dairy cows fed low concentrate diet could be attributed to interacting factors which are prolonged retention times of digesta associated with the high fibre content (carbon: nitrogen ratio) found in roughage-based diet compared to high concentrate diets and the physical barrier created by the oils in the digesta preventing access by the ruminal microbes for microbial fermentation. Chestnut oil inclusion in the diet of sheep reduced CH4 emission with no adverse effects on the growth performance of Rideau Arcott sheep (Liu et al., 2011). Origanuum oil as an additive in diets formulated for sheep (small Tailed Han × Dorper male castrate sheep) resulted in linear decrease of CH4 production. Therefore, inclusion of oils in ruminant diets have both the positive and negative feedback, there is variability in response by ruminants based on the inclusion levels and the source of the oils (Giannenas et al., 2011). The use of oils in ruminant diets suppresses dry matter intake at higher doses while reducing or at least maintaining methane emission.
Saponin
Saponins are glucosides with foaming characteristics and defaunating capacity directly targeting ciliate protozoa in the rumen which have positive symbiotic relationship with methanogens, lowering the abundance of methanogens associated with protozoa, thus exerting their influence on reducing enteric methane production (Benchaar et al., 2007). In an in vitro study by Patra and Yu (2013), in response to a dose rate of 0.6 and 1.2 g/L of quillaja saponin they reported a 12% and 25% reduction in methane production, respectively. However, Goel et al. (2008) reported no change in methane production even though the protozoal counts were adversely affected. Notable, the relationship between the methanogens and protozoa is not unidirectional which is reflected by the lack of correlation in reduction of protozoal count and methanogenesis. Tea saponin reduced in vitro CH4 production whereas it increased (+14%) CH4 yield (g/kg DMI) in lactating dairy cows in vivo (Guyader et al., 2017). From the similar study, tea saponin reduced dry matter intake and subsequently milk yield. However, tea saponin had no effect on enteric methane production in beef steers fed conventional finishing feedlot-diet (Ramírez-Restrepo et al., 2016). On the other hand, tea saponin increased the production of propionate, lowering acetate to propionate ratio and ultimately exerting its influence on reduced CH4 production in sheep (Zhou et al., 2011). Even though average daily gain and gain to feed ratio were improved in Boer goats receiving diets supplemented with 3g/d of tea saponin there were no improvements in performance metrices (daily gain and gain to feed ratio) when the dose rate was increased to 6g/d (Hu et al., 2006). The inconsistency in the results from these studies suggest that the threshold above which the supplementation of saponin adversely affecting the production performance of ruminants warrants further documented. Additionally, Figure 2 below depicts the elucidated effect of saponin in ruminants.
Ether extract
The crude fat containing other compounds extracted by diethyl or petroleum ether prior to hydrolysis is referred to as ether extract (EE). These are predominantly not fermented by the ruminal microbes, and mainly the unsaturated fatty acids part of EE has been reported to inhibit methanogenic system of microbes (Lourenço et al., 2010). The hydrogenation of unsaturated fatty acids enhances the production of propionate and supress bacterial cellulolytic activity by specifically inhibiting protozoa thus reducing the production of enteric methane (Horan et al., 2022). Similarly, previous study of Batta et al. (2016) reported that glycerol resulting from hydrogenation of unsaturated fat inhibit the methanogenic bacterial cellulolytic activity. The study by Lee et al. (2003 reported that EE tended to lower methane production in an in vitro trial. Given that the literature evaluating the effect of EE on CH4 emission is scanty, such exacerbate narrowly defining the mode of action of EE pertaining its regulatory effect on enteric CH4 in ruminants. Howsoever, when EE was added as one of the explanatory variables in regression models developed to ascertain efficacy of CH4 reducing strategies the predictive ability of the regression improved Bina et al. 2022)
Halogenated methane analogue
Seaweeds (e.g., A. Armata and A. taxiforms) contain bromochloromethane (BCM), a compound that inhibits cobamide-dependent methyl transferase step in enteric methane formation pathway in the rumen. This compound has been reported to reduce CH4 yield and improve feed efficiency in sheep and cattle (May et al., 1995; Goel et al., 2009). In contrary to this view, results from studies conducted in beef cattle are consistent in dose-dependent reduction of DMI (-7.4% to -11.3%) and CH4 yield (-90.6% to -95.2%) in response to BCM (Johnson et al., 1972; McCrabb et al., 1997). However, BCM in sheep and goats had little (-0.5%) to no detrimental effect on DMI while CH4 yield was reduced by 33.6% to 86.9% (Sawyer et al., 1974; Abecia et al., 2012). Methane mitigating potential of seaweed (containing BCM) has been evaluated in beef steers (Kinley et al., 2020), dairy cows (Roque et al., 2019) and sheep (Li et al., 2018) with consistent CH4 reduction. When A. Armata was used in dairy cows at the dose rate of 1% in dry matter basis there was a 37% and 50% decrease in dry matter intake (DMI) and CH4 yield (g/ kg DMI), respectively (Roque et al., 2019). In a study by Kinley et al. (2020) beef steers in a feedlot subjected to A. taxiforms had a linear decrease in DMI and CH4 yield by -0.4% and -98%, respectively. However, in a study by Li et al. (2018) when A. taxiforms was used in sheep at higher dosage (3% vs 0.2%) compared to beef steers in a study by Kinley et al. (2020), there were no changes in DMI whereas CH4 yield reduction was lower in magnitude (80.7% vs 98%). Beside the study of Kinley et al. (2020) the results from studies evaluating the efficacy of halogenated CH4 analogues on enteric CH4 from ruminant livestock are consistent in reducing CH4.
Chemical Antimethanogenic Feed Additives
3-. nitoroxy-propanol
3-nitroxy-propanol (3-NOP) is an analogue of enzyme methyl-coenzyme-M, known for its inhibitory role for enzyme methyl-coenzyme-M reductase (Duval and Kinderman, 2012; Pitta et al. 2018). The Methyl-coenzyme M is a cofactor involved in the transfer of methyl group to methyl-coenzyme M reductase (MCR) at the end step of methane formation in the rumen. The MCR in the tetrapyrrole derivative known as cofactor F430 contains nickel and for it to be in the active state it should be in a form of nickel (Ni+) oxidation state (Jayanegara et al., 2017). The active site of MCR is directly inactivated through oxidation by 3-nitroxy-propanol (3-NOP), forming nitrate, nitrite, and 1,3-propane-diol (Duin et al., 2016; Jayanegara et al., 2017). There are two major metabolic stages of 3-NOP in vivo, first step is the oxidation of the primary alcohol to a carboxylic group (i.e., 3-nitroxy-propionic acid; NOPA) and lastly the nitrate ester cleavage to form 3-hydroxy-propionic acid; HPA (Figure 2).
Figure 2.
The chemical structures of 3-NOP and the metabolites (NOP and HPA) in the blood plasma (Adapted from Thiel et al., 2019).
Figure 2.
The chemical structures of 3-NOP and the metabolites (NOP and HPA) in the blood plasma (Adapted from Thiel et al., 2019).
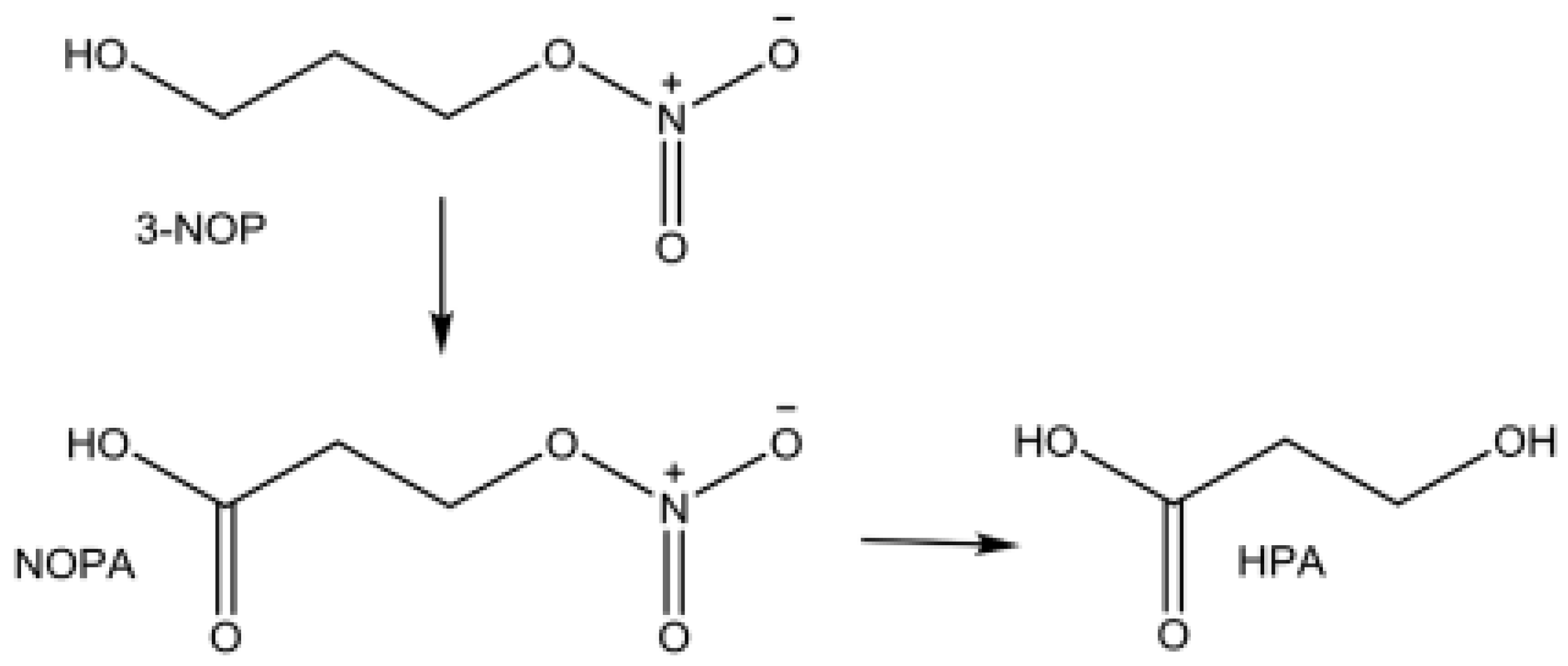
In a study by Garcia et al. (2022), lactating Holstein Friesian dairy cows had a 54% decrease in CH4 emission per digestible organic matter in response to 3-NOP at the supplementation rate of 100 mg/kg DM. Short-term supply of 3-NOP on DMI in lactating dairy cows had no effects even at the highest inclusion level of 120 mg/kg MD (Melgar et al., 2020). There was a linear dose-dependent decrease in CH4 corrected for DMI in Angus heifers fed high forage diet supplemented with incremental levels of 3-NOP, and at the highest inclusion level there was a 33% reduction of CH4 g/kg DMI (Romero-Perez et al. 2014). The 3-nitroxy-propanol has never manifested any toxicity in ruminants when used as CH4 abatement strategy in sheep (Martinez-Fernandez et al., 2013), dairy cattle (Reynolds et al., 2014; Garcia et al., 2022), and beef cattle (Romero-Perez et al., 2014). In a meta-analysis by Jayanegara et al. (2017) it was predicted that the inclusion of 3-NOP at 200 mg/kg DM may reduce CH4 emissions by 33.4-42.1% in ruminants. The use of 3-NOP at the threshold of 200 mg/kg DM and beyond warrants further studies both in vitro and in vivo on reducing CH4 emissions and to facilitate the adoption of this dose it should not induce reduced in vitro and in vivo dry matter digestibility and intake in ruminants (Vyas et al., 2016).
Nitrate
Nitrate (NO3-) provides non-protein nitrogen to the ruminal microbiota and outcompetes the methanogenesis pathway for metabolic hydrogen (H2). Nitrate has a strong affinity for H2 than carbon dioxide (CO2) in the rumen system (Latham et al., 2016; Almeida et al., 2021). Hence, the energy from the consumed substrate by the ruminant usually lost in methanogenesis is redirected to microbial growth in the rumen system. However, the use of nitrate in ruminants as antimethanogenic agent has limited application due to toxicity through methemoglobinemia. Nitrite is formed post-ingestive of NO3- and when absorbed into the bloodstream of the ruminant it oxidizes the iron (Fe2+) in haemoglobin forming methemoglobin (Met-Hb). However, to circumvent such, animals are gradually introduced to NO3- to attain persistency. Also, nitrite as an intermediate of nitrate reduction has a direct toxic effect to methanogens (van Zijderveld et al., 2010 and Zhou et al., 2012). When the dose rate of 0.9 g NO3-/kg0.75/d was evaluated in a growth trial of Thai native × Angola-Nubian goat kids either fed rice straw or Pongola grass there was an overall increase in DMI by 3% (Yuangklang et al., 2022). In the study by Adejoro et al. (2020) there was a 20% improvement in average daily gain with 17% reduction in methane (g/kg DM intake) of lambs supplied with nitrate. The authors concluded that the energy usually lost in methane formation could have shifted towards growth performance in lambs receiving nitrate-based diets. Other studies evaluating nitrate supplementation reported consistent reduction in methane production in beef cattle (Hulshof et al., 2012; Granja-Salcedo et al., 2019), dairy cows (van Zijderveld et al., 2011; Olijhoek et al., 2016), and sheep (Nolan et al, 2010; Guyader et al., 2016). The variability in the magnitude of reduction in methane production could be attributed to quite several possible factors which include but are not limited to; differences in animal species, age of trial animals, basal diets, and NO3- dose rate.
Diet Manipulation
Dry matter intake is the principial variable influencing enteric methane emissions in ruminants (Ellis et al., 2007; Hristov et al., 2013) and grazing is the most common and economical system of feeding for herbivores (Pepeta et al., 2022). However, this system is unavoidable subject to seasonal variation of quantity and quality of pastures, which in turn affect production performance and carbon footprint of animals reared under this system. During the seasons where forage availability is low with poor quality such as in dry winter season in the tropics, supplementation is of paramount importance to at least maintain production performance of ruminants in these regions. When 0 vs 5 kg/head of cow per day was supplemented to grazing Holstein-Frisian lactating dairy cows in a study by Muñoz et al. (2015) there were no changes in CH4 g/kg DMI (CH4 yield) and CH4 g/kg milk yield (CH4 intensity). This could be attributed to the fact that in the study by Muñoz et al. (2015) animals were grazing pastures with temperate grass (Lollium perene) which is of good quality. Therefore, supplementation of concentrate increased both the milk yield and CH4 through improved dry matter intake, such that the CH4 g/kg milk yield remained unchanged. Conversely, in lactating Jersey dairy cows grazing “medium quality” Pennisetum clandestinum-dominant pastures in summer in the tropics, CH4 yield, and intensity decreased linearly while CH4 g/day (production) increased with incremental levels of concentrate supplementation (van Wyngaard et al., 2018). Similarly, Jiao et al. (2014) reported congruent results in lactating Holstein-Frisian dairy cows grazing Lollium perene and supplemented with incremental levels of concentrate diets. Both studies by the authors (Jiao et al., 2014 and van Wyngaard et al., 201), reported induced increase in total dry matter intake with the proportion of forage consumed decreasing when concentrate supplementation levels were increasing. In this view, the addition of concentrate diets in dairy rations shifted the concentration of neutral detergent fibre (NDF) to starch ratio to a lower proportion resulting in reduced CH4 production (Knapp et al., 2014). Also, the controversy of reduced CH4 yield and intensity with increased CH4 production in a study of van Wyngaard et al. (2018) could be attributed to the shift in the rumen system caused by the increase in starch to NDF ratio not being enough for propionate production rather favored the production of butyrate. Medium-high quality forages are characterized with higher amounts of starch and carbohydrates, altering the abundance and functionality of the microbiota in the rumen. From such, there is a noticeable increase in proteobacteria phylum and Succini vibrionaceae family which are hydrogen-consumers to form succinate a propionate precursor, thus reducing enteric CH4 production (Karekar et al., 2022). Brazilian semi-arid non descriptive goats had reduced CH4 production and yield with increasing total dry matter intake and average daily gain in response to increasing levels of concentrate supplementation (Barbosa et al., 2017). Conversely, goats and sika deer had increased CH4 production and yield in response to increasing levels of concentrate supplementation in consumed diets (Na et al., 2017). Goats and sika deer are intermediate and concentrate eaters, respectively, thus they have low stomach capacity, large opening with fast passage rate of digesta (Na et al., 2017). Therefore, the indigested material in the rumen of concentrate and intermediate feeders passes to the omasum at a faster rate for complete methanogenesis to occur due to short retention time. High residual feed intake groups of Holstein-Friesian and Jersey cows when both were fed high concentrate diet (61% of total diet, DM basis) compared to high fibre diet (68% of total diet, DM basis) had lower CH4 (L/day) of 540 vs 635 and 493 vs 499, respectively (Olijhoek et al., 2017). Reduction in forage to concentrate ratio in diets of ruminants (grazers) is noted to reduce CH4 production without adversely affecting production performance and it is advisable that this strategy should be applied with caution to avoid acidosis occurrence especially in animals fed medium-high quality forages.
Exogenous Supply of Microbes
Exogenous supply of microbes commonly referred to as direct fed microbials are living organisms, either alone or in a mixture, that encourage the growth of beneficial microorganisms in the rumen and offer positive effects when given to animals (Tseten et al., 2022). Several rumen bacteria are believed to engage in competition with methanogens for the available hydrogen supply by stimulating processes like propionogenesis, acetogenesis, and the reduction of nitrate/nitrite or sulfate, which can serve as an alternative way to consume metabolic hydrogen (Ungerfeld, 2015). Possible ruminal pathways modulated by exogenously supplied microbes is depicted in Figure 3.
Acetogens
Several attempts have been made to isolate homoacetogens from the rumen and analyze their role as an alternative hydrogen sink, including Acetitomaculum ruminis, Eubacterium limosum, Blautia schinkii, and Blautia producta (Henderson et al., 2010). Furthermore, in vitro studies have also suggested that acetogenesis could serve as an alternative to methanogenesis in eliminating metabolic hydrogen [H] from the rumen (Morvan et al., 1996). However, their abundance and affinity towards [H] are generally lower than hydrogenotrophic methanogens (Le Van et al., 1998; Lopez et al., 1999). As Lopez et al. (1999) have concluded, high concentrations of acetogenic bacteria cannot compete against methanogens for [H] disposal, making it unclear whether homoacetogens could play a pivotal role in the ruminal ecosystem (Henderson et al., 2010). Homoacetogens, found in diverse environments including the rumen, possess the ability to produce metabolic acetate through heterotrophic and autotrophic growth. Stimulating their autotrophic growth is considered a competitive strategy against methanogenesis, as both pathways share common substrates. While they exhibit variability in abundance based on factors like diet and animal age, a negative correlation between homoacetogens and methanogens in the rumen of adult ruminants has been observed (Henderson et al., 2010). Efforts to utilize homoacetogens to mitigate ruminal methane production as direct fed microbials (DFMs) have encountered limitations. Methanogens hold advantages in terms of [H] threshold and energy yield. Furthermore, the ability of acetogens to grow heterotrophically affects their potential as effective methane reducers. However, the feasibility of utilizing these acetogens as DFMs depends on their performance under rumen conditions.
Propionate-forming bacteria
Methane emission (g/kg DMI) was reduced in a study of Vyas et al. (2014) using a combination of Propionibacterium jensenii P54 with P169 and P5 strains of Propionibacterium acidipropionic in steers fed roughage-based diets. However, when the same strains were used in beef heifers fed grain rich diet they were ineffective on CH4 emission (Vyas et al., 2014). In the study of Philippeau et al. (2017) Propionibacteria failed to elicit changes on CH4 emission in dairy cows receiving high-grain diet as opposed to high fibre diet where there was a decrease on CH4 emission. These studies suggest that the effect of Propionibacteria is masked or negated by the general high production of propionate in ruminants fed high grain-based diets. However, this strategy has a great potential of utilisation in ruminants feeding on roughage-based diets including grazing ruminants.
Methylotrophs
Methanotrophs are methylotrophs (microorganism utilising organic mono-carbon compounds as energy source) that classically utilise CH4 thereby preventing its emission from ruminants to the environment. Generally, methylotrophs utilise compounds such as methanol (CH3OH) and methylamine (CH3NH2) as their carbon sources rendering them suitable competitors with methanogens for substrate. Attainment of meticulous better apprehension of biological pathways entailed in the metabolism of methylotrophs may provide novelty in biological control of enteric CH4. There is dearth of literature regarding the use of these compounds as probiotics in ruminants. Methanotrophs utilize monooxygenase as their enzyme that converts CH4 to CH3OH through oxidation (Tseten et al., 2022). The latter is then further oxidized to formaldehyde and the enzyme involved as a catalyst is CH3OH dehydrogenase. Thereafter, formaldehyde is assimilated into either serine or ribulose-5-monophosphate pathway aiding biomass synthesis (Kalyuzhnhaya et al., 2015). Howsoever, there is only one study by Kajikawa et al. (2003) reporting methanotrophy and it was an in vitro study with minute evidence of between 0.2-0.5%. Other methylotrophs such as Methanosphaera spp. are considered important in ruminants fed forage-based diets (Jayanathan et al., 2011). However, the literature reporting in vivo studies of methylotrophy remains scanty, therefore there is a need to develop isolation, screening and in vivo validation studies to meticulously understand the potential of methylotrophs in lessening enteric CH4 emission.
Conclusions
Chemical additives like 3-NOP, nitrate, and halogenated compounds have demonstrated significant methane reduction in both in vitro and in vivo experiments. Such is achieved by either fostering the growth of microbes that compete for resources with methane producers or by directly inhibiting methane production. The focus has shifted towards natural feed additives such as oils, seaweed extracts, and other plant-derived compounds due to human health concerns. Additionally, the use of probiotics as animal dietary supplements has gained importance. One of the approaches is introducing microbes that compete for hydrogen with methane-producing organisms, like propionate-forming bacteria and acetogens. The other relies on methylotrophs, particularly methane-oxidizing bacteria, which directly consume methane produced during rumen fermentation. Developing efficient strategies to mitigate methane emissions while enhancing animal performance is crucial for agricultural sustainability. However, there are conflicting observations regarding the use of organic and inorganic strategies in ruminants to reduce methane. In many cases, when used in high doses to achieve significant methane reduction, these strategies negatively impact dry matter intake and digestibility, potentially reducing overall production performance. Conversely, when used at lower doses to maintain intake, digestibility, and performance, methane reduction is often insufficient. Incorporating data from various global experiments conducted in ruminants using meta-analytical approaches to identify potential benefits of these strategies in mitigating methane emissions and improving production performance, is mandatory and needs urgent attention.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, X.X. and Y.Y.; methodology, X.X.; software, X.X.; validation, X.X., Y.Y. and Z.Z.; formal analysis, X.X.; investigation, X.X.; resources, X.X.; data curation, X.X.; writing—original draft preparation, X.X.; writing—review and editing, X.X.; visualization, X.X.; supervision, X.X.; project administration, X.X.; funding acquisition, Y.Y. All authors have read and agreed to the published version of the manuscript.”, please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research received no external funding.
Acknowledgments
In this section you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
Declare conflicts of interest or state “The authors declare no conflict of interest.” Authors must identify and declare any personal circumstances or interest that may be perceived as inappropriately influencing the representation or interpretation of reported research results. Any role of the funders in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results must be declared in this section. If there is no role, please state “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results”.
References
- Abecia, L., Toral, P.G., Martín-García, A.I., Martínez, G., Tomkins, N.W., Molina-Alcaide, E., Newbold, C.J. and Yáñez-Ruiz, D.R., 2012. Effect of bromochloromethane on methane emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy goats. Journal of dairy scien. [CrossRef]
- Aboagye, I.A. and Beauchemin, K.A., 2019. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals, 9(11), p.856. [CrossRef]
- Adejoro, F.A., 2019. The use of condensed tannins and nitrate to reduce enteric methane emission and enhance utilization of high forage diets in sheep. Doctoral Dissertation; University of Pretoria.
- Adejoro, F.A., Hassen, A., Akanmu, A.M. and Morgavi, D.P., 2020. Replacing urea with nitrate as a non-protein nitrogen source increases lambs' growth and reduces methane production, whereas acacia tannin has no effect. Animal Feed Science and Technology, 259, p.114360. [CrossRef]
- Akapali, M., 2018. Seasonal variation in forage availability and grazing behavior of cattle in selected peri-urban areas in the Northern region of Ghana. Doctoral Dissertation; University of Ghana.
- Almeida, A.K., Hegarty, R.S. and Cowie, A., 2021. Meta-analysis quantifying the potential of dietary additives and rumen modifiers for methane mitigation in ruminant production systems. Animal Nutrition, 7(4), pp.1219-1230. [CrossRef]
- Al-Rawi, A.A., 2022. Outlook threat at the future climate changes on livestock resources. GSC Advanced Research and Reviews, 10(1), pp.001-006. Benchaar, C.; Wang, Y.; Chaves, A.V.; Mc Allister, T.A.; Beauchemin, K.A. 2007. Use of plant extracts in ruminant nutrition. In: Acharya, S.N., Thomas, J.E, Eds.; Advances in Medicinal Plant Research; Research Signpost: Kerala, India, pp. 465-489. [CrossRef]
- Bakkali, F., Averbeck, S., Averbeck, D. and Idaomar, M., 2008. Biological effects of essential oils–a review. Food and chemical toxicology, 46(2), pp.446-475. [CrossRef]
- Barbosa, A.L., Voltolini, T.V., Menezes, D.R., de Moraes, S.A., Nascimento, J.C.S. and de Souza Rodrigues, R.T., 2018. Intake, digestibility, growth performance, and enteric methane emission of Brazilian semiarid non-descript breed goats fed diets with different forage to concentrate ratios. Tropical animal health and production, 50(2), pp.283-289. [CrossRef]
- Bayat, A.R., Ventto, L., Kairenius, P., Stefański, T., Leskinen, H., Tapio, I., Negussie, E., Vilkki, J. and Shingfield, K.J., 2017. Dietary forage to concentrate ratio and sunflower oil supplement alter rumen fermentation, ruminal methane emissions, and nutrient utilization in lactating cows. Translational Animal Science, 1(3), pp.277-286. [CrossRef]
- Bina, J.D., Tonsor, G.T., Schulz, L.L. and Hahn, W.F., 2022. Regional and plant-size impacts of COVID-19 on beef processing. Food Policy, 108, p.102247. [CrossRef]
- Blumenthal, D.M., Mueller, K.E., Kray, J.A., Ocheltree, T.W., Augustine, D.J. and Wilcox, K.R., 2020. Traits link drought resistance with herbivore defense and plant economics in semi-arid grasslands: The central roles of phenology and leaf dry matter content. Journal of Ecology, 108(6), pp.2336-2351. [CrossRef]
- ce, 95(4), pp.2027-2036.
- Cieslak, A., Szumacher-Strabel, M., Stochmal, A., Oleszek, W., 2013. Plant components with specific activities against rumen methanogens. Animal 7 Suppl 2, 253–65. [CrossRef]
- Czerkawski, J.W., Blaxter, K.L. and Wainman, F.W., 1966. The metabolism of oleic, linoleic and linolenic acids by sheep with reference to their effects on methane production. British Journal of Nutrition, 20(2), pp.349-362. [CrossRef]
- Demeyer, D.I., 1975. Methanogenesis, an integrated part of carbohydrate fermentation and its control. Digestion and Metabolism in the Ruminant.
- Devillard, E., Bera-Maillet, C., Flint, H.J., Scott, K.P., Newbold, C.J., Wallace, R.J., Jouany, J.P. and Forano, E. (2003) Characterization of XYN10B, a modular xylanase from the ruminal protozoan Polyplastron multivesiculatum, with a family 22 carbohydrate-binding module that binds to cellulose. Biochem. J. 373, 495-503. [CrossRef]
- Drake HL, Gößner AS, and Daniel SL., 2008. Old acetogens, new light. Annal. NY Acad. Sci. 1125: 100-128. [CrossRef]
- Dschaak, C.M.; Williams, C.M.; Holt, M.S.; Eun, J.S.; Young, A.J.; Min, B.R. Effects of supplementing condensed tannin extract on intake, digestion, ruminal fermentation, and milk production of lactating dairy cows. J. Dairy Sci. 2011, 94, 2508–2519. [Google Scholar] [CrossRef] [PubMed]
- Duin EC, Wagner T, Shima S, Prakash D, Cronin B, Yanez-Ruiz DR, Duval S, Rumbeli R, Stemmler RT, Thauer RK, et al. 2016. Mode of action uncovered for the specific reduction of methane emissions from ruminants by the small molecule 3-nitrooxypropanol. Proc Natl Acad Sci USA. 113:6172–6177. [CrossRef]
- Ellis, J.L., Kebreab, E., Odongo, N.E., McBride, B.W., Okine, E.K. and France, J., 2007. Prediction of methane production from dairy and beef cattle. Journal of dairy science, 90(7), pp.3456-3466. [CrossRef]
- Fonty G, Joblin K, Chavarot M, Roux R, Naylor G, and Michallon F., 2007. Establishment and development of ruminal hydrogenotrophs in methanogen-free lambs. Appl. Environ. Microbiol. 73: 6391-6403. [CrossRef]
- Ghamkhar, K., Rochfort, S., Banik, B.K. and Revell, C., 2018. Candidate metabolites for methane mitigation in the forage legume biserrula. Agronomy for Sustainable Development, 38(3), pp.1-10. [CrossRef]
- Goel, G., Makkar, H.P.S. and Becker, K., 2008. Changes in microbial community structure, methanogenesis and rumen fermentation in response to saponin-rich fractions from different plant materials. Journal of applied microbiology, 105(3), pp.770-777. [CrossRef]
- Granja-Salcedo YT, Fernandes RM, Araujo RC, Kishi LT, Berchielli TT, Resende FD, Berndt A and Siqueira GR., 2019. Long-Term Encapsulated Nitrate Supplementation Modulates Rumen Microbial Diversity and Rumen Fermentation to Reduce Methane Emission in Grazing Steers. Front. Microbiol. 10:614. [CrossRef]
- Granja-Salcedo, Y.T., Fernandes, R.M., Araujo, R.C.D., Kishi, L.T., Berchielli, T.T., Resende, F.D.D., Berndt, A. and Siqueira, G.R., 2019. Long-term encapsulated nitrate supplementation modulates rumen microbial diversity and rumen fermentation to reduce methane emission in grazing steers. Frontiers in Microbiology, 10, p.614. [CrossRef]
- Gruninger, R.J., Zhang, X.M., Smith, M.L., Kung Jr, L., Vyas, D., McGinn, S.M., Kindermann, M., Wang, M., Tan, Z.L. and Beauchemin, K.A., 2022. Application of 3-nitrooxypropanol and canola oil to mitigate enteric methane emissions of beef cattle results in distinctly different effects on the rumen microbial community. Animal Microbiome, 4(1), p.35. [CrossRef]
- Gruninger, R.J., Zhang, X.M., Smith, M.L., Kung, L., Vyas, D., McGinn, S.M., Kindermann, M., Wang, M., Tan, Z.L. and Beauchemin, K.A., 2022. Application of 3-nitrooxypropanol and canola oil to mitigate enteric methane emissions of beef cattle results in distinctly different effects on the rumen microbial community. Animal Microbiome, 4(1), pp.1-17. [CrossRef]
- Guyader, J., Doreau, M., Morgavi, D.P., Gérard, C., Loncke, C., Martin, C., 2016. Long-term effect of linseed plus nitrate fed to dairy cows on enteric methane emission and nitrate and nitrite residuals in milk. Animal 10, 1173–1181. [CrossRef]
- Guyader, J., Eugène, M., Doreau, M., Morgavi, D.P., Gérard, C. and Martin, C., 2017. Tea saponin reduced methanogenesis in vitro but increased methane yield in lactating dairy cows. Journal of Dairy Science, 100(3), pp.1845-1855. [CrossRef]
- Haisan, J., Sun, Y., Guan, L.L., Beauchemin, K.A., Iwaasa, A., Duval, S., Barreda, D.R. and Oba, M., 2014. The effects of feeding 3-nitrooxypropanol on methane emissions and productivity of Holstein cows in mid lactation. Journal of dairy science, 97(5), pp.3110-3119. [CrossRef]
- Henderson, G., Cox, F., Ganesh, S., Jonker, A., Young, W. and Janssen, P.H., 2015. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Scientific reports, 5(1), p.14567. Addisu, S., 2016. Effect of dietary tannin source feeds on ruminal fermentation and production of cattle; a review. Online J. Anim. Feed Res, 6(2), pp.45-56.
- Henderson, G., Naylor, G. E., Leahy, S. C., & Janssen, P. H. (2010). Presence of novel, potentially homoacetogenic bacteria in the rumen as determined by analysis of formyltetrahydrofolate synthetase sequences from ruminants. Applied and Environmental Microbiology, 76(7), 2058-2066. [CrossRef]
- Hodrová, B., Kopecný, J., Petr, O., 1995. Interaction of the rumen fungus Orpinomyces 168 joyonii with Megasphaera elsdenii and Eubacterium limosum. Lett. Appl. Microbiol. 21, 34–37. [CrossRef]
- Horrigan, L., Lawrence, R.S. and Walker, P., 2002. How sustainable agriculture can address the environmental and human health harms of industrial agriculture. Environmental health perspectives, 110(5), pp.445-456. [CrossRef]
- Hristov, A.N., Oh, J., Firkins, J.L., Dijkstra, J., Kebreab, E., Waghorn, G., Makkar, H.P.S., Adesogan, A.T., Yang, W., Lee, C., Gerber, P.J., Henderson, B., Tricarico, J.M., 2013. SPECIAL TOPICS—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 91, 5045–5069. [CrossRef]
- Hu, W., Liu, J., Wu, Y., Guo, Y. and Ye, J., 2006. Effects of tea saponins on in vitro ruminal fermentation and growth performance in growing Boer goat. Archives of Animal Nutrition, 60(1), pp.89-97. [CrossRef]
- Hulshof, R.B.A., Berndt, A., Gerrits, W.J.J., Dijkstra, J., van Zijderveld, S.M., Newbold, J.R., Perdok, H.B., 2012. Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J. Anim. Sci. 90, 2317–2323. [CrossRef]
- Jami, E. and Mizrahi, I., 2012. Composition and similarity of bovine rumen microbiota across individual animals. PloS one, 7(3), p.e33306. [CrossRef]
- Jayanegara A, Sarwono KA, Kondo M, Matsui H, Ridla M, Laconi EB, Nahrowi. Use of 3-nitrooxypropanol as feed additive for mitigating enteric methane emissions from ruminants: a meta-analysis. Ital J Anim Sci 2017;17:650e6.
- Jiao, H.P., Dale, A.J., Carson, A.F., Murray, S., Gordon, A.W., Ferris, C.P., 2014. Effect of concentrate feed level on methane emissions from grazing dairy cows. J. Dairy Sci. 97, 7043–7053. [CrossRef]
- Johnson, E.D., Wood, A.S., Stone, J.B. and Moran Jr, E.T., 1972. Some effects of methane inhibition in ruminants (steers). Canadian Journal of Animal Science, 52(4), pp.703-712. [CrossRef]
- Karekar, S., Stefanini, R. and Ahring, B., 2022. Homo-Acetogens: Their Metabolism and Competitive Relationship with Hydrogenotrophic Methanogens. Microorganisms, 10(2), p.397. [CrossRef]
- Kinley RD, Fredeen AH. In vitro evaluation of feeding North Atlantic stormtoss seaweeds on ruminal digestion. J Appl Phycol. (2015) 27:2387.e2393. [CrossRef]
- Knapp, J. R., G. L. Laur, P. A. Vadas, W. P. Weiss, and J. M. Tricarico. 2014. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 97:3231–3261. [CrossRef]
- Krause, D.O., Denman, S.E., Mackie, R.I., Morrison, M., Rae, A.L., Attwood, G.T. and McSweeney, C.S., 2003. Opportunities to improve fiber degradation in the rumen: microbiology, ecology, and genomics. FEMS microbiology reviews, 27(5), pp.663-693. [CrossRef]
- Latham EA, Anderson RC, Pinchak WE, Nisbet DJ. Insights on alterations to the rumen ecosystem by nitrate and nitrocompounds. Front Microbiol 2016;7. [CrossRef]
- Le Van, T. D., Robinson, J. A., Ralph, J., Greening, R. C., Smolenski, W. J., Leedle, J. A., ... & Wallace, R. J. (1998). Assessment of reductive acetogenesis with indigenous ruminal bacterium populations and Acetitomaculum ruminis. Applied and Environmental Microbiology, 64(9), 3429-3436. [CrossRef]
- Lee, H.J., Lee, S.C., Kim, J.D., Oh, Y.G., Kim, B.K., Kim, C.W. and Kim, K.J., 2003. Methane production potential of feed ingredients as measured by in vitro gas test. Asian-Australasian Journal of Animal Sciences, 16(8), pp.1143-1150. [CrossRef]
- Lee, S.J., Lee, Y.J., Eom, J.S., Kim, H.S., Choi, Y.Y., Jo, S.U., Kang, S.N., Park, H.Y., Kim, D.H. and Lee, S.S., 2020. Effects of the appropriate addition of antioxidants from Pinus densiflora and Mentha canadensis extracts on methane emission and rumen fermentation. Animals, 10(10), p.1888. [CrossRef]
- Li X, Norman HC, Kinley RD, Laurence M, Wilmot M, Bender H, et al. 2016., Asparagopsis taxiformis decreases enteric methane production from sheep. Anim Prod Sci. 58:681–88. [CrossRef]
- Lima, P.R.; Apdini, T.; Freire, A.S.; Santana, A.S.; Moura, L.M.L.; Nascimento, J.C.S.; Rodrigues, R.T.S.; Dijkstra, J.; Garcez Neto, A.F.; Queiroz, M.A.Á.; 2019. Dietary supplementation with tannin and soybean oil on intake, digestibility, feeding behavior, ruminal protozoa and methane emission in sheep. Anim. Feed Sci. Technol. 249, 10–17. [CrossRef]
- Lopez, S., McIntosh, F. M., Wallace, R. J., & Newbold, C. J. (1999). Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Animal Feed Science and Technology, 78(1-2), 1-9. [CrossRef]
- Lowe, S.E., Theodorou, M.K. and Trinci, A.P. (1987) Cellulases and xylanase of an anaerobic rumen fungus grown on wheat straw, wheat straw holocellulose, cellulose, and xylan. Appl. Environ. Microbiol. 53, 1216-1223. [CrossRef]
- Martinez-Fernandez, G., Duval, S., Kindermann, M., Schirra, H.J., Denman, S.E. and McSweeney, C.S., 2018. 3-NOP vs. halogenated compound: methane production, ruminal fermentation and microbial community response in forage fed cattle. Frontiers in microbiology, 9, p.1582. [CrossRef]
- May, C., Payne, A.L. and Stewart, P.L., 1995. A delivery system for agents International Patent Application No. PCT/AU95/00733.
- McCann, J.C., Wickersham, T.A. and Loor, J.J., 2014. High-throughput methods redefine the rumen microbiome and its relationship with nutrition and metabolism. Bioinformatics and biology insights, 8, pp.BBI-S15389. [CrossRef]
- McCrabb, G.J., Berger, K.T., Magner, T., May, C. and Hunter, R.A., 1997. Inhibiting methane production in Brahman cattle by dietary supplementation with a novel compound and the effects on growth. Australian Journal of Agricultural Research, 48(3), pp.323-329. [CrossRef]
- Mizrahi, I. and Jami, E., 2018. The compositional variation of the rumen microbiome and its effect on host performance and methane emission. Animal, 12(s2), pp.s220-s232. [CrossRef]
- Moraïs, S. and Mizrahi, I., 2019. The road not taken: the rumen microbiome, functional groups, and community states. Trends in Microbiology, 27(6), pp.538-549. [CrossRef]
- Morgavi, D.P., Sakurada, M., Mizokami, M., Tomita, Y., Onodera, R., 1994. Effects of ruminal protozoa on cellulose degradation and the growth of an anaerobic ruminal fungus, piromyces sp. strain OTS1, in vitro. Appl. Environ. Microbiol. 60, 3718–3723. [CrossRef]
- Morvan, B., Bonnemoy, F., Fonty, G., & Gouet, P. (1996). Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and methanogenic archaea from digestive tract of different mammals. Current Microbiology, 32(2), 129-133. [CrossRef]
- Muñoz, C., S. Hube, J. M. Morales, T. Yan, and E. M. Ungerfeld. 2015. Effects of concentrate supplementation on enteric methane emissions and milk production of grazing dairy cows. Livest. Sci. 175:37–46. [CrossRef]
- Na, Y., Li, D.H. and Lee, S.R., 2017. Effects of dietary forage-to-concentrate ratio on nutrient digestibility and enteric methane production in growing goats (Capra hircus hircus) and Sika deer (Cervus nippon hortulorum). Asian-Australasian Journal of Animal Sciences, 30(7), p.967. [CrossRef]
- Nagpal, R., Puniya, A.K., Griffith, G.W., Goel, G., Puniya, M., Sehgal, J.P., Singh, K., 2009. Anaerobic rumen fungi: potential and applications. Agric. Important Micro-Organisms II, 375–393.
- Naumann, H.D., Tedeschi, L.O., Zeller, W.E. and Huntley, N.F., 2017. The role of condensed tannins in ruminant animal production: advances, limitations and future directions. Revista Brasileira de Zootecnia, 46, pp.929-949. [CrossRef]
- Newbold, C.J., Ushida, K., Morvan, B., Fonty, G. and Jouany, J.P. (1996) The role of ciliate protozoa in the lysis of methanogenic archaea in rumen £uid. Lett. Appl. Microbiol. 23, 421-425. [CrossRef]
- Nolan, J. V., Hegarty, R.S., Hegarty, J., Godwin, I.R., Woodgate, R., 2010. Effects of dietary nitrate on fermentation, methane production and digesta kinetics in sheep. Anim. Prod. Sci. 50, 801–806. [CrossRef]
- Olijhoek, D. and Lund, P., 2017. Methane production by ruminants. Department of Animal science AU-Foulum. Aarhus University, Denmark.
- Olijhoek, D.W., Hellwing, A.L.F., Brask, M., Weisbjerg, M.R., Hojberg, O., Larsen, M.K., Dijkstra, J., Erlandsen, E.J., Lund, P., 2016. Effect of dietary nitrate level on enteric methane production, hydrogen emission, rumen fermentation, and nutrient digestibility in dairy cows. J. Dairy Sci. 99, 6191–6205. [CrossRef]
- Orlandi, T.; Kozloski, G. V.; Alves, T.P., 2015. Mesquita, F.R.; Ávila, S.C. Digestibility, ruminal fermentation and duodenal flux of amino acids in steers fed grass forage plus concentrate containing increasing levels of Acacia mearnsii tannin extract. Anim. Feed Sci. Technol. 210, 37–45. [CrossRef]
- Patra, A.K., Saxena, J., 2010. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 71, 1198– 1222. [CrossRef]
- Patra, A.K., Yu, Z., 2013. Effective reduction of enteric methane production by a combination of nitrate and saponin without adverse effect on feed degradability, fermentation, or bacterial and archaeal communities of the rumen. Bioresour. Technol. 148, 352–360. [CrossRef]
- Pepeta, B.N., Moyo, M., Adejoro, F.A., Hassen, A. and Nsahlai, I.V., 2022. Techniques Used to Determine Botanical Composition, Intake, and Digestibility of Forages by Ruminants. Agronomy, 12(10), p.2456. [CrossRef]
- Poudel, S., Zeller, W.E., Fike, J. and Pent, G., 2023. Condensed Tannins Attributes: Potential Solution to Fescue Toxicosis?. Agriculture, 13(3), p.672. [CrossRef]
- Ragsdale SW, and Pierce E., 2008. Acetogenesis and the wood–ljungdahl pathway of CO2 fixation. Biochim. Biophys. Acta 1784: 1873- 1898. [CrossRef]
- Ramírez-Restrepo, C.A., Tan, C., López-Villalobos, N., Padmanabha, J., Wang, J. and McSweeney, C.S., 2016. Methane production, fermentation characteristics, and microbial profiles in the rumen of tropical cattle fed tea seed saponin supplementation. Animal Feed Science and Technology, 216, pp.58-67. [CrossRef]
- Rezaeian, M., Beakes, G.W., Parker, D.S., 2004. Distribution and estimation of anaerobic zoosporic fungi along the digestive tracts of sheep. Mycol. Res. 108, 1227–1233. [CrossRef]
- Roger, V., Fonty, G., Andre, C. and Gouet, P., 1992. Effects of glycerol on the growth, adhesion, and cellulolytic activity of rumen cellulolytic bacteria and anaerobic fungi. Current microbiology, 25(4), pp.197-201. [CrossRef]
- Roque BM, Salwen JK, Kinley R, Kebreab E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J Clean Prod. (2019) 234:132–8. [CrossRef]
- Russell, J.B., 2002. Rumen Microbiology and Its Role in Ruminant Nutrition.
- Sawyer, M.S., Hoover, W.H. and Sniffen, C.J., 1974. Effects of a ruminal methane inhibitor on growth and energy metabolism in the ovine. Journal of Animal Science, 38(4), pp.908-914. [CrossRef]
- Sharp, R., Hazlewood, G.P., Gilbert, H.J. and O’Donnell, A.G. (1994) Unmodi¢ed and recombinant strains of Lactobacillus plantarum are rapidly lost from the rumen by protozoal predation. J. Appl. Bacteriol. 76, 110-117. [CrossRef]
- Shreck, A., 2013. Use of alkaline treated crop residues as partial grain replacements for finishing cattle. Doctoral Dissertation; University of Nebraska-Lincoln.
- Silanikove, N.; Perevolotsky, A.; Provenza, F.D. 2001, Use of tannin-binding chemicals to assay for tannins and their negative postingestive effects in ruminants. Anim. Feed Sci. Technol. 91, 69–81. [CrossRef]
- Silva, J.J.M.D., Campanharo, S.C. and Paschoal, J.A.R., 2021. Ethnoveterinary for food-producing animals and related food safety issues: A comprehensive overview about terpenes. Comprehensive Reviews in Food Science and Food Safety, 20(1), pp.48-90. [CrossRef]
- Thiel, A., Schoenmakers, A.C.M., Verbaan, I.A.J., Chenal, E., Etheve, S. and Beilstein, P., 2019. 3-NOP: mutagenicity and genotoxicity assessment. Food and Chemical Toxicology, 123, pp.566-573. [CrossRef]
- Tiwari, R. and Rana, C.S., 2015. Plant secondary metabolites: a review. International Journal of Engineering Research and General Science, 3(5), pp.661-670.
- Van Wyngaard, J.D.V., Meeske, R. and Erasmus, L.J., 2018. Effect of concentrate feeding level on methane emissions, production performance and rumen fermentation of Jersey cows grazing ryegrass pasture during spring. Animal Feed Science and Technology, 241, pp.121-132. [CrossRef]
- van Zijderveld, S.M., Fonken, B., Dijkstra, J., Gerrits, W.J.., Perdok, H.B., Fokkink, W., Newbold, J. R., 2011. Effects of a combination of feed additives on methane production, diet digestibility, and animal performance in lactating dairy cows. J. Dairy Sci. 94, 1445–1454. [CrossRef]
- Van Zijderveld, S.M., Gerrits, W.J., Apajalahti, J.A., Newbold, J.R., Dijkstra, J., Leng, R.A., Perdok, H.B., 2010. Nitrate and sulfate: effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 93, 5856– 5866. [CrossRef]
- Vyas, D., McGinn, S.M., Duval, S.M., Kindermann, M.K. and Beauchemin, K.A., 2016. Optimal dose of 3-nitrooxypropanol for decreasing enteric methane emissions from beef cattle fed high-forage and high-grain diets. Animal Production Science, 58(6), pp.1049-1055. [CrossRef]
- Weimer, P.J., 2015. Redundancy, resilience, and host specificity of the ruminal microbiota: implications for engineering improved ruminal fermentations. Frontiers in microbiology, 6, p.296. [CrossRef]
- Wischer, G.; Greiling, A.M.; Boguhn, J. 2014, Steingass, H.; Schollenberger, M.; Hartung, K.; Rodehutscord, M. Effects of long-term supplementation of chestnut and valonea extracts on methane release, digestibility, and nitrogen excretion in sheep. Animal 8, 938–948. [CrossRef]
- Woodward, S.L.; Waghorn, G.C.; Laboyrie, P.G. 2004, Condensed tannins in birdsfoot trefoil (Lotus corniculatus) reduce methane emissions from dairy cows. Proc. N. Z. Soc. Anim. Prod. 64, 160–164.
- Yanza, Y.R., Fitri, A., Suwignyo, B., Hidayatik, N., Kumalasari, N.R., Irawan, A. and Jayanegara, A., 2021. The utilisation of tannin extract as a dietary additive in ruminant nutrition: a meta-analysis. Animals, 11(11), p.3317. [CrossRef]
- Yuangklang, C., Paengkoum, P., Paengkoum, S. and Schonewille, J.T., 2020. Nitrate supplementation of rations based on rice straw but not Pangola hay, improves growth performance in meat goats. Asian-Australasian Journal of Animal Sciences. [CrossRef]
- Zhang, L., Tian, H., Shi, H., Pan, S., Chang, J., Dangal, S.R., Qin, X., Wang, S., Tubiello, F.N., Canadell, J.G. and Jackson, R.B., 2022. A 130-year global inventory of methane emissions from livestock: Trends, patterns, and drivers. Global Change Biology, 28(17), pp.5142-5158. [CrossRef]
- Zhou, Y.Y., Mao, H.L., Jiang, F., Wang, J.K., Liu, J.X. and McSweeney, C.S., 2011. Inhibition of rumen methanogenesis by tea saponins with reference to fermentation pattern and microbial communities in Hu sheep. Animal Feed Science and Technology, 166, pp.93-100. [CrossRef]
- Zhou, Z., Yu, Z., Meng, Q., 2012. Effects of nitrate on methane production, fermentation, and microbial populations in in vitro ruminal cultures. Bioresour. Technol. 103, 173–179. [CrossRef]
Figure 1.
Fundamental pathways of the utilisation of consumed dietary sources by micro-organisms in the rumen to end products absorbed to the blood stream or emitted to the environment (Adopted from Moraïs and Mizrahi, 2019).
Figure 1.
Fundamental pathways of the utilisation of consumed dietary sources by micro-organisms in the rumen to end products absorbed to the blood stream or emitted to the environment (Adopted from Moraïs and Mizrahi, 2019).
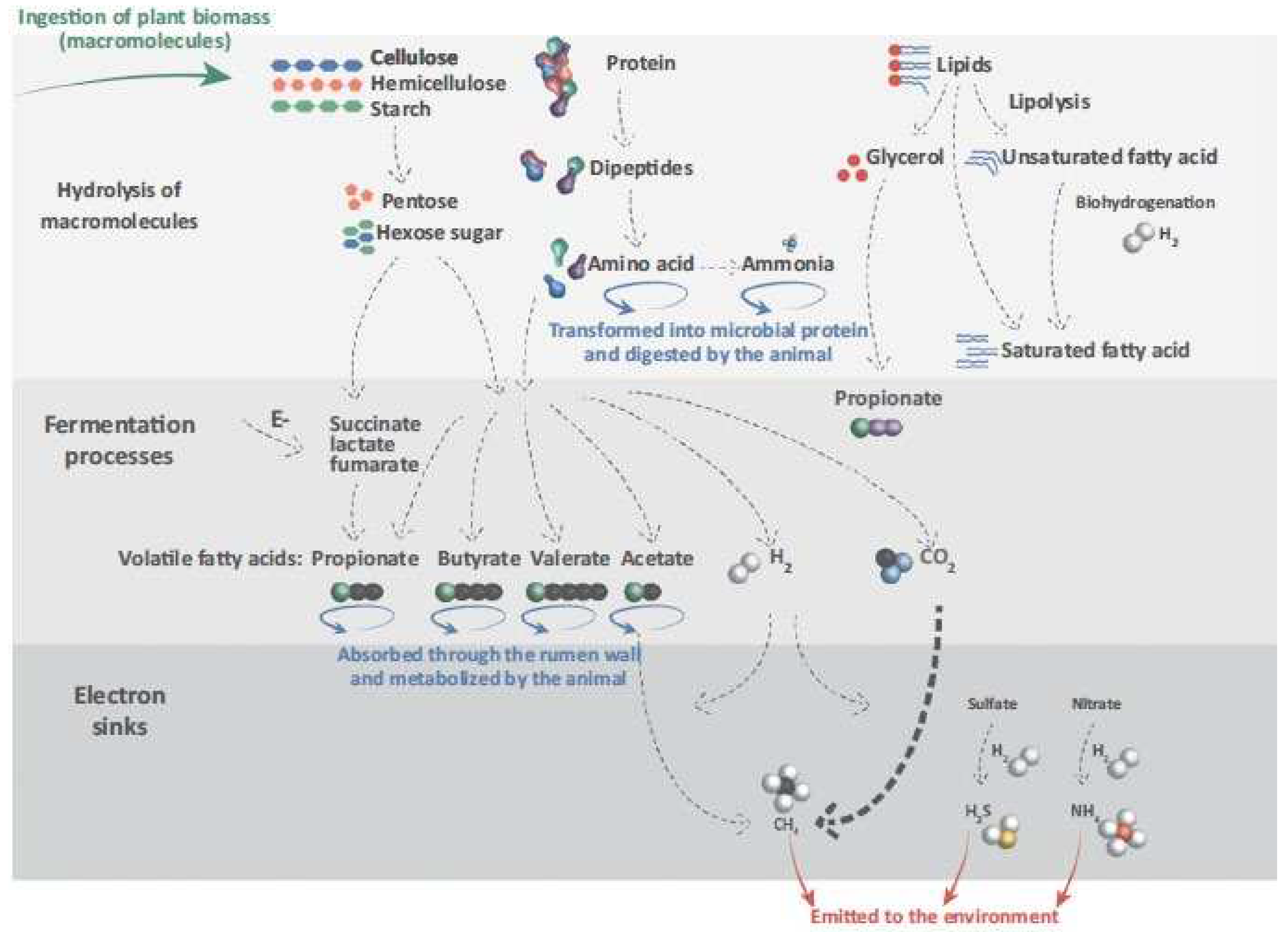
Figure 2.
Meticulously summarised biological pathway of dietary additive or supplementation of saponins in ruminants on CH4 reduction and animal productivity (Pepeta, personal communication).
Figure 2.
Meticulously summarised biological pathway of dietary additive or supplementation of saponins in ruminants on CH4 reduction and animal productivity (Pepeta, personal communication).
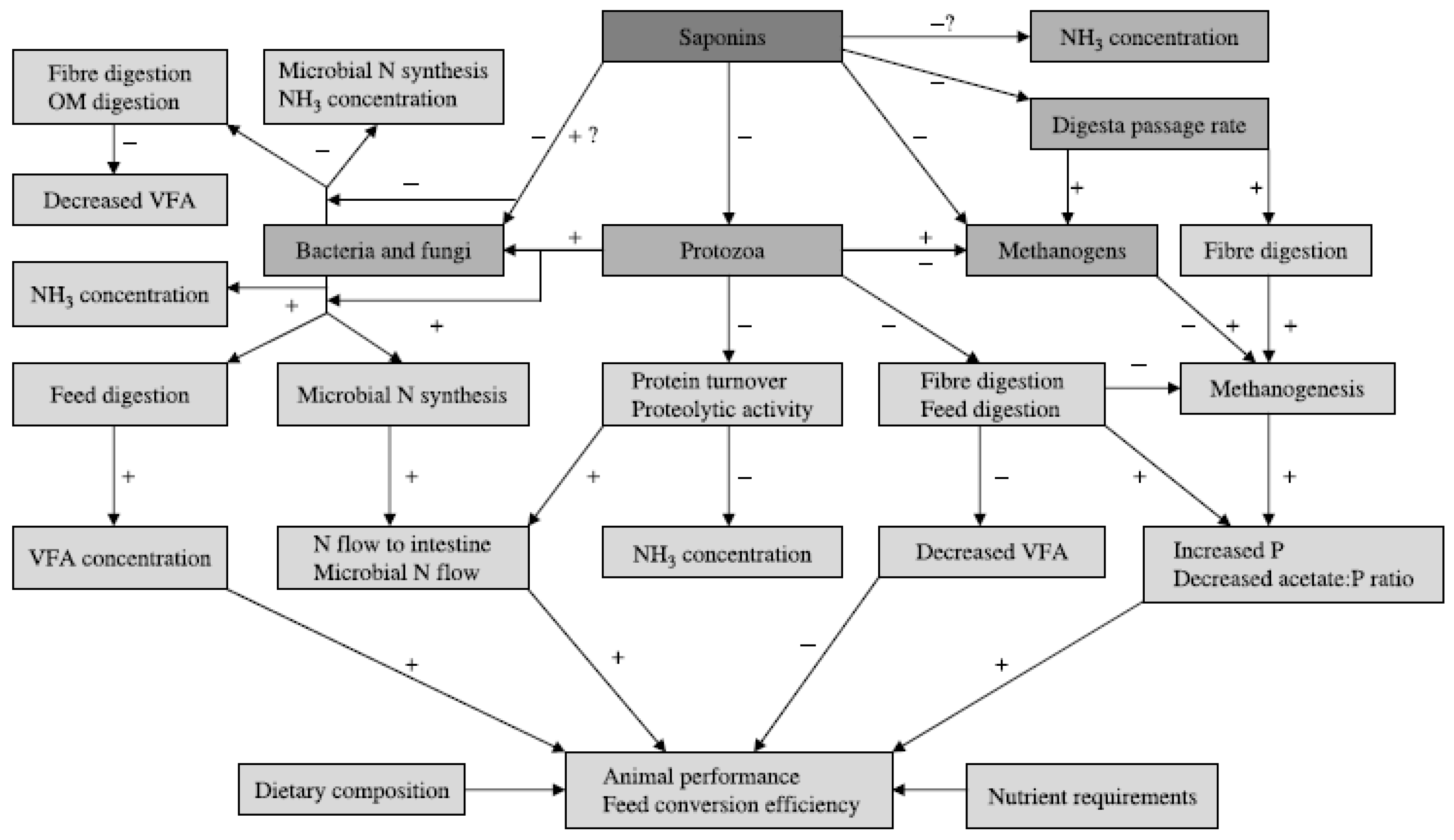
Figure 3.
Modulated pathways of ruminal fermentation due to exogenously supplied bacteria (adopted from Jeyanathan et al., 2013).
Figure 3.
Modulated pathways of ruminal fermentation due to exogenously supplied bacteria (adopted from Jeyanathan et al., 2013).
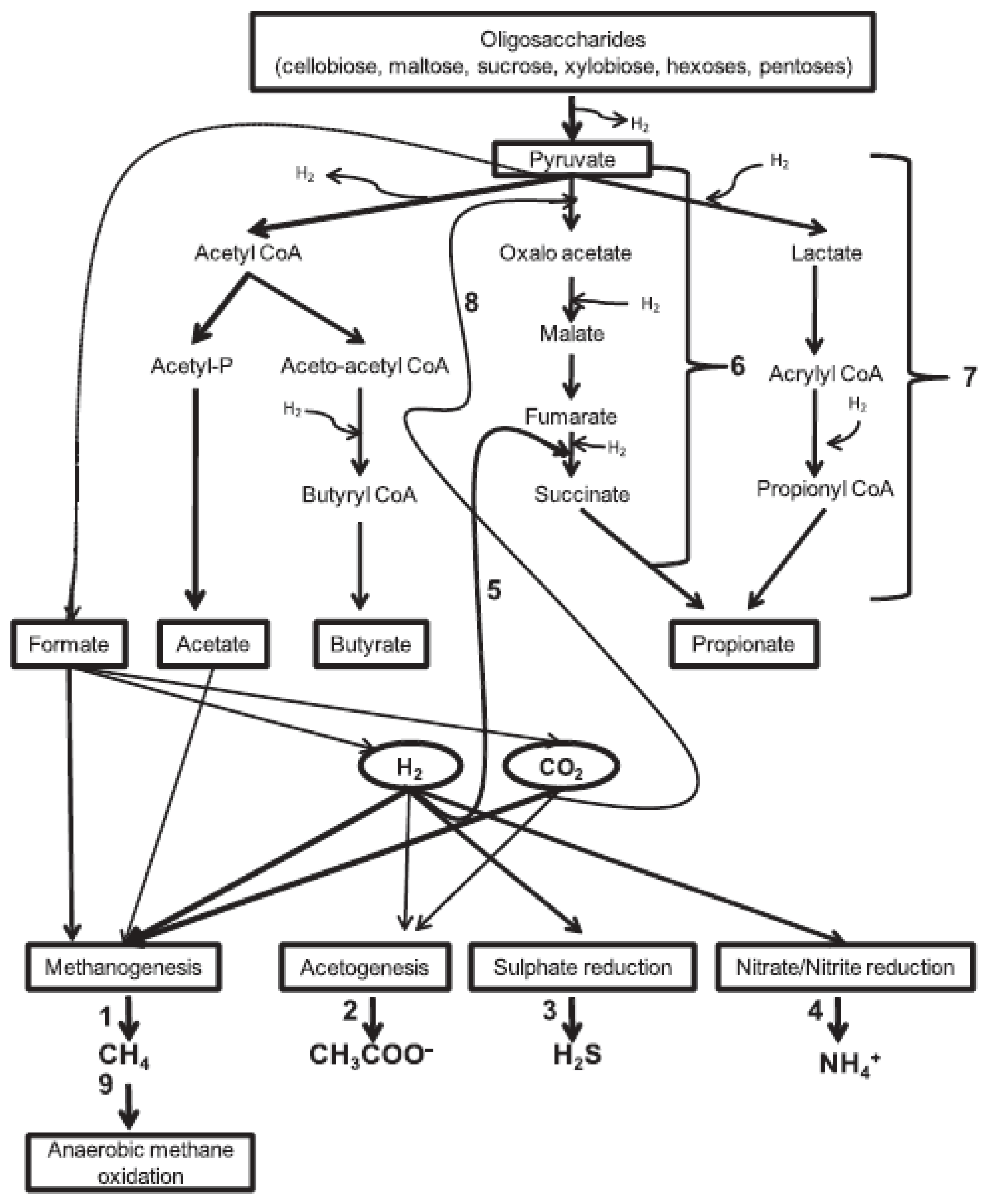

Table 1.
Effect of tannins on dry matter digestibility, nitrogen balance parameters and methane production in ruminants.
Table 1.
Effect of tannins on dry matter digestibility, nitrogen balance parameters and methane production in ruminants.
|
Tannin source |
Tannin type |
Inclusion level (g/kg) |
Animal species |
Diet CP (g/kg) |
F: C ratio |
Changes relative to the control treatment (%) |
Reference |
||||
|
Digestibility |
Urinary N |
Faecal N |
Retained N |
CH 4 |
|||||||
| Schinopsis lorentzii | CT | 10 | Dairy heifers | 265 | 50/50 | -1.20 | -15.87 | +6.90 | +190 | n.d | Ahnert et al., 2015 |
| Prosopis cineraria | HT | 18.40 | Lambs | 177 | TMR | -3.57 |
-27.59 | +3.33 | -1.59 | -22 | Bhatt et al., 2020 |
| Schinopsis balansae | CT | 40 | Castrate goats | 150 | 51/49 | -6.92 | -35.26 | +35.13 | +22.85 | n.d | Al-Kindi et al., 2016 |
| Schinopsis balansae | CT | 10 | Beef steers | 140 | TMR | -6.28 | -9.76 | +32.14 | -1.82 | +8.62 | Ebert et al., 2017 |
| Quercus robur | CT | 8.70 | Dairy cows | 131 | TMR | -0.59 | -12.15 | +3.29 | +6.67 | -1.06 | Focant et al., 2019 |
| Ficus infectoria and Psidium guajava (mixture of 1:1 ratio) | CT | 15 | Lambs | n.d | 70/30 | -1.55 | -20.41 | +30.25 | +82.07 | -14.34 | Pathak et al., 2017 |
| Acacia mearnsii | CT | 40 | Wethers | 146 | 50/50 | -20.25 | -47.98 | 41.27 | -5.88 | -31.83 | Adejoro et al., 2019 |
| Castanea sativa | HT | 15 | Beef steers | 171 | 50/50 | n.d | -6.74 | n.d | n.d | +3.24 | Aboagye et al., 2018 |
| Vitis vinifera | CT | 15.5 | Goat kids | 127 | 55/45 | +4.92 | -17.53 | +52.09 | -26.94 | +1.19 | Sinz et al., 2021 |
| Tea waste | CT | 15 | Dairy goats | 164 | TMR | -2.04 | -32.94 | +5.56 | +0.7 | +10.02 | Sundod et al., 2023 |
| Onobrychis viciifolia | CT | 8.8 | Dairy cows | 172 | TMR | -5.45 | +3.10 | +15.13 | +145.45 | -2.71 | Huyeen et al., 2016 |
| Schinopsis balansae | CT | 40 | Beef heifers | 103 | TMR | -19.10 | n.d | n.d | n.d | -56.26 | Piñeiro-Vázquez et al., 2018 |
N; nitrogen, F:C; forage to concentrate, CH4; methane, CT; condensed tannins, HT; hydrolysable tannin, n.d; no data, TMR; total mixed ration; negative and positive values respectively imply either a decrease or an increase in digestibility, nitrogen balance parameters and methane production in response to tannin supply treatments compared to the control groups of ruminant livestock.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



