
Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Rosavin, a phenylpropanoid in Rhodiola rosea's rhizome, an adaptogen, is known for enhancing the body's response to environmental stress. It significantly affects cellular metabolism in health and many diseases, particularly influencing bone tissue metabolism. In vitro, rosavin inhibits osteoclastogenesis, disrupts F-actin rings formation, and lowers osteoclastogenesis-related gene expression (cathepsin K, CTR, TRAF6, TRAP, MMP-9). It also impedes NFATc1, c-Fos, NF-κB, and MAPK signaling pathways, and blocks phosphorylation processes crucial for bone resorption. Moreover, rosavin promotes osteogenesis, osteoblast differentiation and increases Runx2 and OCN expression. In vivo studies show its effectiveness in enhancing BMD in PMOP mice, re-straining osteoclast maturation, and increasing active osteoblast percentage in bone tissue. It modulates mRNA expressions (increasing EEF2 and decreasing HDAC1, thereby activating os-teoprotective epigenetic mechanisms) and alters various serum markers, including decrease of CTX-1, TRAcp5b, RANKL, M-CSF, TRAP, and increase of ALP and OCN. Additionally, when combined with zinc and probiotics, it reduces pro-osteoporotic MMP-3, IL-6, TNF-α, and en-hances anti-osteoporotic IL-10 and TIMP3 expressions. This paper aims to systematically review rosavin's impact on bone tissue metabolism, exploring its potential in osteoporosis prevention and treatment, and suggesting future research directions.
Keywords:
Keywords: rosavin
; adaptogens
; bone metabolism
; osteoblasts
; osteoporosis
; Rhodiola rosea
; bone loss
; osteoclastogenesis
; osteogenesis
1. Introduction
Rhodiola rosea belongs to the Crassulaceae plant family. It has been used in traditional medicine for over 2,000 years as an adaptogen, increasing the body's immunity and adaptive capabilities in unfavorable environmental conditions. Rhodiola rosea stimulates the nervous system, improves physical endurance, reduces fatigue, has antidepressant and anti-inflammatory properties [1,2,3,4,5]. This plant requires specific growing conditions and occurs naturally in specified regions of the Earth, especially in Northern Europe, Northern Asia (Siberia) and North America [6]. Large commercial cultivations of this plant are currently present in China and Ukraine. Due to many years of observations of the health benefits of Rhodiola rosea, especially its root extracts, it has become the subject of numerous scientific studies in the field of biochemistry, physiology, pathophysiology, and medicine. Phytochemical studies have identified approximately 140 chemical compounds in Rhodiola rosea root, divided into six groups: phenylpropanoids, phenylethanoid derivatives, flavonoids, monoterpene derivatives, triterpenes, and phenolic acids [7]. Subsequent studies have shown that the health-promoting properties of Rhodiola rosea root are primarily attributed to rosavin, a phenylpropanoid and salidroside, a phenylethanoid derivative [8]. Initially, assessing the individual impact of each compound on human health was difficult. Recent research trends include laboratory studies in cell cultures and animal models assessing the independent effects of salidroside and rosavin on various physiological and pathophysiological processes, including cellular metabolism, inflammation, and regeneration. Current research interest is more towards salidroside, considered to be the main and most characteristic representative of the group of phenylethanoid derivatives, next to the well-studied tyrosol [9,10]. However recently, rosavin, one of several phenylpropanoids found in Rhodiola rosea root, along with others such as rosin and rosarin, is also gaining attention in scientific research. Studies have confirmed the neuroprotective effect of rosavin in animal models of Alzheimer's disease [11] and in cerebral ischemia [12], its role in limiting respiratory tract damage in sepsis [13], its anticancer effect in small cell lung cancer [14] and its anti-inflammatory effect in fibrosis liver [15] and acute colitis [16]. It has also been shown that rosavin affects the metabolism of bone tissue. With the still increasing number of patients suffering from degenerative bone diseases such as osteoporosis, osteoarthritis and rheumatoid arthritis, and the associated health care costs worldwide, it is important to investigate new osteoprotective molecules that may become the future standard of care for patients with reduced bone mineral density (BMD) [17,18,19,20]. The aim of the work is to critically systematize the current knowledge about the influence of rosavin on the metabolism of bone tissue, taking into account potential future directions of research in this area.
2. Biochemical Structure of Rosavin
Rosavin is a phytochemical belonging to the group of phenylpropanoids [1]. This group of chemical compounds contains a six-carbon aromatic phenyl group and a three-carbon propene tail. Phenylpropanoids are synthesized from the amino acid phenylalanine via the shikimate pathway, which primarily occurs in bacteria, fungi, and plants (the phenylpropanoid synthesis pathway does not occur in animals) [21]. The chemical formula of rosavin is C20H28O10 [22] and its molecular weight is 428.4 g/mol [23]. In its solid form, rosavin appears as a white powder and is characterized by good solubility in water. The chemical structure of rosavin is depicted in Figure 1.
3. Impact of Rosavin on Bone Tissue Metabolism
In the study conducted by Zhang et al., the aim was to investigate the effect of rosavin on osteoclastogenesis in mouse models of variectomy-induced osteoporosis (OVX), which is analogous to postmenopausal osteoporosis (PMOP), as well as in in vitro cell culture conditions [24]. For in vitro tests, standard cell models for osteoclastogenesis were used, including bone marrow mesenchymal stem cells (BMMSCs) and murine macrophage cells (RAW264.7 cells). Five cell colonies were created for each cell type: the first colony served as the control culture, receiving only dedicated medium. The remaining four colonies were exposed to receptor activator for nuclear factor κ B ligand (RANKL) (100 ng/ml) and macrophage colony-stimulating factor (M-CSF) (30 ng/ml). Additionally, three of these colonies received supplementation with increasing concentrations of rosavin (1.25 μM, 2.5 μM and 5 μM respectively). After seven days of incubation, the morphology of the cell colonies and the number of tartrate-resistant acid phosphatase positive cells (TRAP-positive cells), which in this study corresponded to osteoclasts, were evaluated. It was observed that with increasing concentrations of rosavin, the number of TRAP-positive cells decreased and at a concentration of 5 μM, the morphology of both BMMSCs and RAW264.7 colonies closely resembled that of the control colony [24]. Subsequent analyses confirmed the positive impact of rosavin on osteogenesis in BMMSCs cell cultures. After 20 days of incubation with rosavin, alkaline phosphatase (ALP) staining and alizarin red staining were performed, which are used to identify osteoblasts in the examined tissue. Compared to the control group, it was demonstrated that rosavin significantly stimulated osteogenesis by increasing the population of osteoblast cells [24]. BMMSCs cultures with rosavin also exhibited increased expression of mouse runt-related transcription factor 2 (Runx2) and osteocalcin (OCN), which are widely recognized as typical markers of ongoing osteogenesis [25,26]. In the same study, it was noted that as the concentration of rosavin increased in the respective BMMSCs cultures, the number and size of F-actin rings decreased. The formation of F-actin rings is observed during osteoclastogenesis and is a physiological process for this cell line, without which the bone resorption process cannot proceed [27]. The influence of rosavin, directly proportional to its concentration, was also evident through a reduced surface area of resorbed bone tissue compared to the control group that did not receive rosavin supplementation [24]. Zhang et al. also demonstrated that rosavin inhibits osteoclastogenesis most effectively in BMMSCs and RAW264.7 colonies when administered to cells one day after the addition of RANKL and M-CSF [24]. The effectiveness of osteoclastogenesis inhibition decreases with a delayed intervention of rosavin in these colonies. Rosavin added on the 3rd day in the case of RAW264.7 colonies and on the 5th day in the case of BMMSCs no longer inhibits osteoclast formation. Western blot analysis confirmed these observations [24]. A greater decrease in the expression levels of osteoclastogenesis-related genes, such as cathepsin K, calcitonin receptor (CTR), tumor necrosis factor receptor associated factor 6 (TRAF6), TRAP and matrix metallopeptidase 9 (MMP-9), was noted in colonies that received rosavin on the first day after the administration of RANKL and M-CSF compared to those receiving it on the 3rd and 5th day [24]. From other studies on bone tissue metabolism, it is known that the reduction in the expression of these genes in bone tissue is due to a decrease in the cellular expression of nuclear factor of activated T-cell cytoplasmic 1 (NFATc1), which is considered a key transcription factor determining osteoclastogenesis [28,29]. Blocking NFATc1 inhibits osteoclast maturation, rendering precursor cells insensitive to the action of RANKL [30,31]. In the study, rosavin reduced the presence of NFATc1 in BMMSCs in a manner directly proportional to the concentration, following the principle that "the earlier the intervention, the more effective," which correlates with the aforementioned results of rosavin’s osteoprotective effect on bone tissue. Surprisingly, rosavin did not show any changes in the expression levels of receptor activator of nuclear factor-κB (RANK) and colony-stimulating factor-1 receptor (c-fms) in RAW264.7 colonies treated with M-CSF (what was determined by using Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction, qRT-PCR) [24]. Increased expression of these receptors is observed in osteoporosis and is responsible for intensifying pathological processes related to osteoclastogenesis [32,33]. Therefore, in this particular case, rosavin did not exhibit a potential inhibitory effect on osteoclastogenesis. Another analysis revealed that rosavin inhibits the signaling pathways induced by RANKL, which are typical for osteoclastogenesis and osteoporosis, including the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) [34,35] and mitogen-activated protein kinase (MAPK) pathways [36]. 5 μM of rosavin leads to the inhibition of cellular phosphorylation of human protein 56 (p56) and p56 translocation into the nucleus of RAW264.7 cells, blocking the biological effect of the NF-κB signaling pathway and osteoclastogenesis [24]. Rosavin also blocks the MAPK pathway by inhibiting the phosphorylation of its components, including extracellular signal-regulated kinase (ERK), human protein 38 (p38) and c-Jun N-terminal kinase (JNK), which may have potential significance in the treatment of osteoporosis [24]. Next to these findings, the inhibition of the MAPK pathway by rosavin was also indirectly observed. An effect of the MAPK pathway is the increased cellular expression of proto-oncogene c-Fos. Rosavin significantly reduces the expression of c-Fos in RAW264.7 cells stimulated with RANKL, providing indirect evidence of the effectiveness of blocking the MAPK pathway, which is responsible for bone tissue resorption [24]. A somewhat surprising result of the above-mentioned analysis is the rosavin’s inhibition of the phosphorylation of the inhibitory subunit of NF-κB alpha (IκBα). Phosphorylated IκBα is responsible for inhibiting NF-κB signaling pathway. Excess amount of dephosphorylated form of IκBα should result in activation of the NF-κB signaling pathway. However, as it turns out, this action has weaker biological effects than inhibiting p56 phosphorylation, so it does not change the final qualification of rosavin's metabolism manifestation, which is the inhibition of bone tissue resorption [24]. Mice with OVX-induced osteoporosis, which is analogous to PMOP, are a standard animal model in osteoporosis studies [37]. On such animal model another study was conducted to assess the effect of rosavin on bone tissue. The experimental group of mice received daily intraperitoneal injections of rosavin (10 mg/kg) for 6 weeks. Compared to the control group that did not receive the drug, rosavin led to an increase in BMD and trabecular bone surface in cross-sectional views of the distal femur and radiological studies. Bone loss in mice receiving rosavin was significantly limited [24]. This was confirmed by TRAP staining of prepared histological specimens from the femurs, which showed a statistically significantly smaller number of TRAP-positive cells corresponding to osteoclasts. Rosavin also reduced the mice blood serum levels of markers of increased bone turnover, such as cross-linked C-telopeptide of type I collagen (CTX-1) and tartrate-resistant acid phosphatase 5b (TRAcp5b). In mice receiving rosavin, an increased blood serum level of ALP and OCN, which are considered serum bone formation markers by many researchers, was observed [38,39]. Rosavin's osteoprotective action is also evident at the epigenetic level through the inhibition of the cellular activity of histone deacetylase 1 (HDAC1) [40]. HDAC1 belongs to the class of classical histone deacetylases that contain Zn2+ ions in the catalytic center. Based on many studies, it has been established that the overexpression of HDAC1 leads to the acetylation of many histones, resulting in the pathological silencing of the transcription of many genes and leading to the development of cancer or chronic inflammation [41,42,43]. Regarding bone tissue, it was observed that rosavin, by reducing the expression of HDAC1, increases the expression of eukaryotic translation elongation factor 2 (EEF2) in BMMSCs. Rosavin thus influences the inhibition of the deacetylation process of EEF2 [40]. EEF2, in turn, under physiological conditions, is responsible for bone formation processes and is postulated to have an anti-osteoporotic effect [40,44]. In a dedicated study on the HDAC1/EEF2 axis, it was observed that rosavin reduced the levels of RANKL, M-CSF, and TRAP in the blood serum of OVX mice [40]. In histological specimens of bone tissue from OVX mice receiving rosavin, there was a smaller number of osteoclasts, and the signs of bone resorption were less pronounced compared to OVX mice that did not receive intraperitoneal injections of rosavin. Furthermore, rosavin significantly increased the expression of bone formation markers such as ALP, OCN and Runx2, as well as the number of osteoblasts (by increasing the expression of the surface marker for osteoblasts - osteoprotegerin, OPG) in the bone tissue of OVX mice [40]. To confirm the involvement of rosavin in the HDAC1/EEF2 axis in osteoporosis, Zhang et al. decided to observe how the expression of HDAC1 and EEF2 mRNA changes in OVX mice and OVX mice, which received rosavin [40]. It turned out that mice that did not receive rosavin had increased mRNA expression for HDAC1 and decreased expression for EEF2, while the addition of rosavin reversed this effect [40]. The above observations have become the basis for another in vitro study on the following cells: a mouse clonal, osteoblast-like cell line (MC3T3-E1) and BMMSCs (corresponding to osteoclasts). It was demonstrated that HDAC1 stimulates the proliferation of osteoclasts by reducing the duration of the G0/G1 phase (acceleration of cell divisions) and inhibits cell divisions of osteoblasts. HDAC1 also stimulates the apoptosis of osteoblasts but inhibits the apoptosis of osteoclasts. Increased expression of EEF2 has the opposite effect to HDAC1 in BMMSCs and MC3T3-E1 cells, including inducing cell cycle arrest in the G0/G1 phase in osteoclasts, ultimately reducing their population [40]. A series of additional experiments on MC3T3-E1 cell culture showed that rosavin increased the expression of EEF2 in these cells, resulting in an increased number of ALP-positive cells (corresponding to osteogenesis) and increased calcium nodules, corresponding to the activation of the calcium ion deposition process presented typically in bone formation [40]. In contrast, in BMMSCs culture, which received M-CSF and RANKL, rosavin reduced the expression of genes for HDAC1 and decreased the expression of typical promoters for osteoclastogenesis, such as TRAP, NFATc1 and cathepsin K [40]. This manifested itself in the inhibition of differentiation and maturation processes of osteoclasts in the histological specimens observed. The impact on the HDAC1/EEF2 axis is a very promising potential direction for anti-osteoporotic drug action, also because this axis regulates intracellular signaling pathways for MAPK and NF-κB in the bone tissue of OVX mice [40]. Increased presence of EEF2 (in response to rosavin supplementation) in the bone tissue of OVX mice inhibits MAPK and NF-κB-mediated osteoclastogenesis and reduces bone tissue resorption [40]. In summary, rosavin as a regulator of the HDAC1/EEF2 axis positively influences bone tissue metabolism, which may have potential significance in the treatment of reduced BMD. The scheme of rosavin action on the HDAC1/EEF2 axis with the biological effects on bone metabolism are presented in Figure 2. Oral supplementation of rosavin (100mg/day) in combination with zinc (20mg/day) and a probiotic complex (CNS, Pharm Korea Co. Ltd.) exhibits protective effects on rat bone tissue in a model of osteoarthritis (OA) induced by intra-articular (right knee) injection of 3mg monosodium iodoacetate, compared to the control group and rats receiving only Celecoxib (13.5mg/day) [45]. Kwon et al. demonstrated this through the observation of post-mortem preparations of the distal femoral end of rats (by using micro computed tomography, micro-CT) receiving the aforementioned supplementation. Quantitative micro-CT results from the control group and Celecoxib-treated group did not differ from each other and showed pronounced bone structure deterioration, confirmed by quantitative assessment parameters of bone tissue such as object volume (Obj.V) and Obj.V/total volume (TV). Both of these parameters were higher and osteoporotic changes were inhibited in rats receiving rosavin with zinc and the probiotic complex. It is worth noting that besides osteoporosis, osteoarthritis often involves the thinning of bone tissue structure, which is sometimes observed in radiological studies in the form of subchondral cysts [46,47]. Additionally, osteoarthritis during exacerbations is associated with increased expression of inflammatory cytokines in periarticular tissues (synovial membrane), such as interleukin 1β (IL-1β), tumour necrosis factor α (TNF-α) and interleukin 6 (IL-6), which, apart from inflammation, also stimulate osteoclastogenesis and bone resorption. This is significant due to the proximity between the synovial membrane and the subchondral bone tissue layer [48]. Although Kwon et al. did not analyze the isolated impact of rosavin on bone tissue quality, considering the knowledge gathered so far, the authors regard the results of the cited study as promising, but certainly requiring further, more detailed research. It is worth adding that Kwon et al.'s study also demonstrated the effectiveness of rosavin, zinc and probiotic supplementation in reducing lower limb pain related to the musculoskeletal system and inhibiting inflammatory cytokines responsible for the development of both osteoarthritis and osteoporosis i.e. matrix metallopeptidase 3 (MMP-3) IL-6 and TNF-α [45,48,49]. Furthermore, the aforementioned complex stimulates the secretion of anti-inflammatory interleukin 10 (IL-10) and tissue inhibitor of metalloproteinase 3 (TIMP3) by synovial membrane cells, which exhibit anabolic and protective effects on various tissues, including bone tissue [50,51].
4. Discussion
Despite the promising research results regarding the anti-osteoporotic, anabolic effects of rosavin on bone tissue, the number of publications addressing this issue remains limited. This is even more intriguing since osteoporosis is currently one of the major global health problems, incurring significant financial and social costs [17,18,19]. Therefore, it seems justified to make greater efforts to explore new methods of prevention or treatment for this disease. In the opinion of the authors of this paper, rosavin may prove to be a highly promising potential drug for osteoporosis. The literature gathered above strongly confirms rosavin’s involvement in the metabolic processes of bone tissue: inhibiting its catabolism and promoting anabolism. In in vitro studies, rosavin inhibits osteoclastogenesis, blocks the formation of F-actin rings, reduces the expression levels of osteoclastogenesis-related genes (cathepsin K, CTR, TRAF6, TRAP, MMP-9), inhibits the activity of NFATc1, c-Fos and intracellular signaling pathways NF-κB and MAPK [24]. Inhibition of the NF-κB signaling pathway occurs through rosavin’s inhibition of p56 phosphorylation and its translocation to the cell nucleus, while the reduced biological activity of the MAPK pathway results from the inhibition of ERK, p38 and JNK phosphorylation [24]. In terms of anabolism, rosavin stimulates osteogenesis by increasing the population of osteoblasts in cell cultures and increasing the expression of Runx2 and OCN [24]. The summary of rosavin’s biological manifestations in in vitro studies is presented in Table 1. In in vivo studies, rosavin increases the BMD of the distal end of the femur in a mouse model of PMOP, as confirmed radiologically and histologically. It reduces the number of osteoclasts while increasing osteoblasts in osteoporotic bone tissue. In the blood serum, rosavin decreases the levels of CTX-1, TRAcp5b, RANKL, M-CSF, TRAP and increases the levels of ALP and OCN [24]. In mouse bone tissue cells receiving rosavin, there is a decrease of HDAC1 mRNA expression and an increase of EEF2 mRNA expression (rosavin’s influence on epigenetic osteoprotective mechanisms) [40]. Additionally, an increase in the number of calcium nodules in microscope examination is observed [40]. In a rat model, rosavin in combination with zinc and probiotics reduces the expression of MMP-3, IL-6 and TNF-α and increases the expression of TIMP3 and IL-10 in the synovial membrane of knee joints. Rosavin also affects the inhibition of osteoporotic processes in bone tissue (results confirmed in Quantitative micro-CT examination) [45]. The summary of rosavin’s biological manifestations in in vivo studies is presented in Table 2. It is worth to notice that not all research results cited in this study align with the osteoprotective profile of rosavin. This suggests that there is still much to discover about the multifaceted impact of rosavin on the complex network of metabolic and inflammatory processes within bone tissue. For example, rosavin inhibits the phosphorylation of IκBα subunit in RAW264.7 cells, what potentially is responsible for activating the NF-κB signaling pathway what is connected with osteoclastogenesis [24]. In the same cell culture, rosavin also does not decrease the expression of RANK and c-fms receptors, which is surprising taking into consideration its other anti-osteoporotic properties [24]. However, considering the cited publications, it should be acknowledged that rosavin’s osteoprotective action is probably significantly stronger overall than the individual examples promoting osteoclastogenesis. Moreover, it is not known what the resultant impact of these processes will be on other cell lines, and in a broader context, on the entire human system (potential side effects, systemic complications, etc.). In considering the potential use of rosavin as a treatment for osteoporosis, it is also important to determine whether it will exhibit osteoprotective properties throughout the entire course of the disease. Some presented research results suggest that rosavin exhibits its reparative action on bone tissue metabolism and increases BMD more effectively the earlier it is administered (1 day>3 days>5 days) after the presence of inflammatory and osteoclastogenesis-promoting factors is discovered (RANKL and M-CSF) [24]. This may raise legitimate concerns about the effectiveness of rosavin in patients who have been suffering from osteoporosis for many years. In this situation, rosavin should be considered more as a potential preventive rather than therapeutic measure. More detailed research is highly needed on this matter. It is worth noting that the biological manifestation of rosavin’s action in animal models occurred independently of its administration route - its influence on bone tissue metabolism was observed both through intraperitoneal injections and dietary supplementation [24,40,45]. The potential effectiveness of oral rosavin administration is another advantage, as it has been used in humans as an oral dietary supplement in the form of Rhodiola rosea extract successfully for hundreds of years. Oral administration is generally considered the most acceptable form of drugs administration for patients compared to subcutaneous, intramuscular, or intravenous injections, according to many studies [52]. However, it should be noted that there is still insufficient knowledge in the international literature regarding rosavin’s ability to penetrate enterocytes through the endothelium into the bloodstream. We do not know exactly how rosavin’s chemical structure changes in the human gastrointestinal tract, what metabolism it undergoes after absorption into the bloodstream and, more importantly, what the therapeutic dose of rosavin administered orally or intravenously is optimal to show biological effects on bone tissue. So far, such studies have not been conducted, which are necessary in the context of any considerations regarding the use of rosavin as a useful compound in the treatment of osteopenia or osteoporosis [1]. Another aspect worth considering is the assessment of the collective interaction of rosavin and salidroside on bone tissue metabolism. Current international studies focus on the individual impact of each of these substances on osteogenesis. However, it is worth noting that both of these compounds occur together in nature as components of Rhodiola rosea root and, as mentioned, have been taken together orally as a dietary supplement or adaptogens for many years [2,3,4]. Since research shows that rosavin and salidroside taken separately have anti-osteoporotic effects [53,54,55], it would be necessary to determine whether the combined action of these substances will undergo an additive effect or whether some biological actions of rosavin and salidroside will counteract each other. Conducting such studies would be valuable to gain a better understanding of the biological properties of rosavin and ways to potentially enhance its effectiveness by placing it in complex combinations with other substances, including salidroside. In the opinion of the authors of this paper, the next step toward establishing the potential osteoprotective action of rosavin on human bone tissue is to conduct research on human bone tissue cells, including osteoblasts and osteoclasts. So far, the cited results primarily concern animal models, which, while still promising, require further verification on human bone tissue. Such studies could determine safe rosavin doses in cell cultures and, more importantly, the therapeutic concentrations of rosavin that are optimal for human health. Considering all the above information (collected during the literature review and discussion), rosavin should be considered as a highly promising compound of plant origin with anti-osteoporotic properties. Rosavin has a multi-faceted protective effect on the metabolism of bone tissue, both at the level of cellular regulation of immunity, as well as through epigenetic processes. However, much more specialized laboratory and pharmacological tests are needed to consider this molecule useful in the prevention and treatment of osteoporosis in humans.

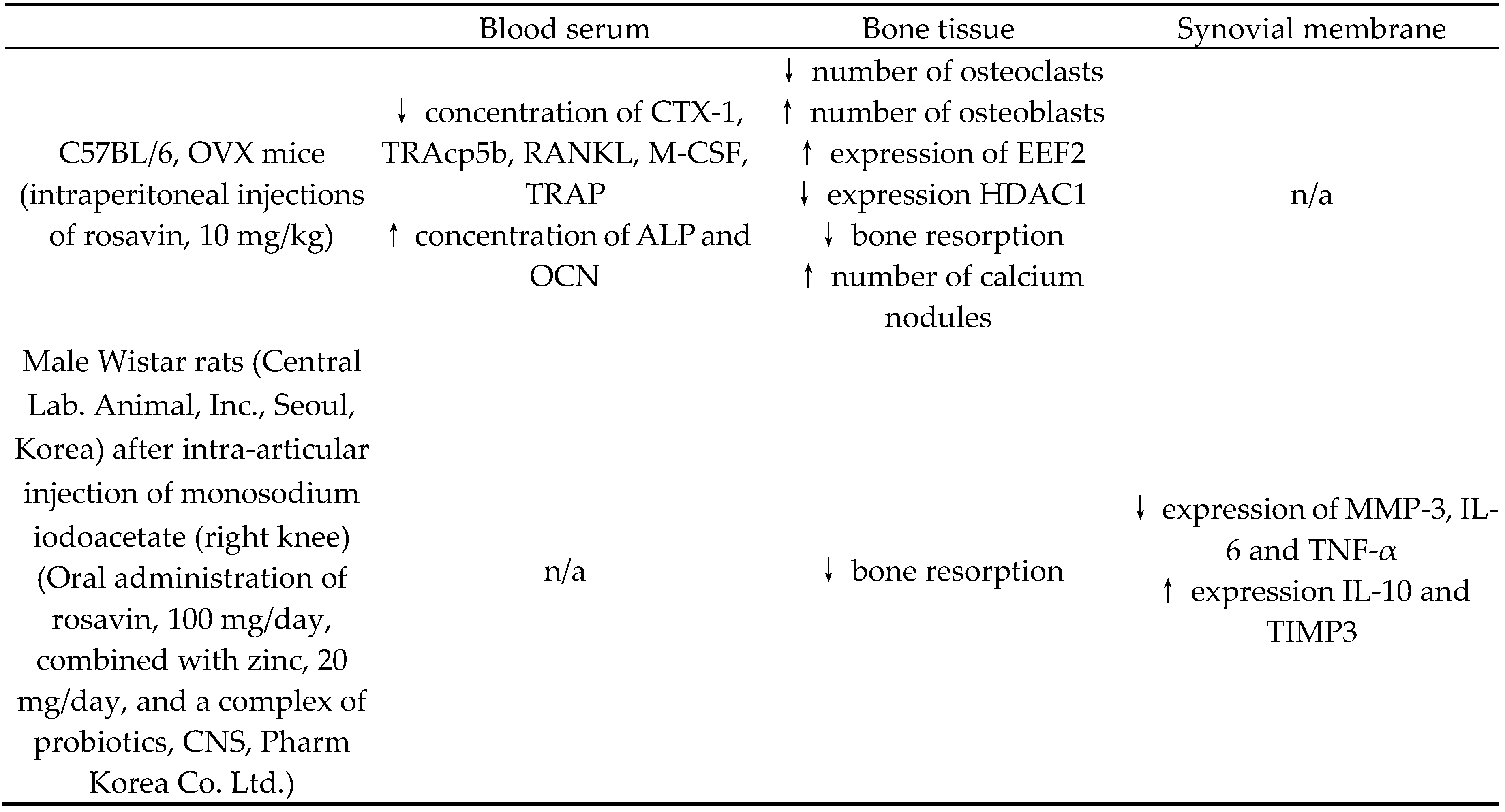
| Blood serum | Bone tissue | Synovial membrane | |
| C57BL/6, OVX mice(intraperitoneal injections of rosavin, 10 mg/kg) | ↓ concentration of CTX-1, TRAcp5b, RANKL, M-CSF, TRAP↑ concentration of ALP and OCN | ↓ number of osteoclasts↑ number of osteoblasts↑ expression of EEF2↓ expression HDAC1↓ bone resorption↑ number of calcium nodules | n/a |
| Male Wistar rats (Central Lab. Animal, Inc., Seoul, Korea) after intra-articular injection of monosodium iodoacetate (right knee)(Oral administration of rosavin, 100 mg/day, combined with zinc, 20 mg/day, and a complex of probiotics, CNS, Pharm Korea Co. Ltd.) | n/a | ↓ bone resorption | ↓ expression of MMP-3, IL-6 and TNF-α↑ expression IL-10 and TIMP3 |
Author Contributions
Conceptualization, P.W., P.T. and A.L-G.; Methodology, P.W., P.T., A.L-G., Ł.A.P. and D.P.; Software, P.W., P.T. and Ł.A.P.; Validation, P.W., P.T., A.L-G, Ł.A.P., D.P. and M.-R.M.-S.; Formal Analysis, P.W., A.L-G, Ł.A.P., D.P. and M.-R.M.-S.; Investigation, P.W., D.P., M.-R.M.-S. and D.S.; Resources, Ł.A.P., D.P., M.-R.M.-S. and D.S.; Data Curation, P.W., P.T. and D.S.; Writing – Original Draft Preparation, P.W. P.T., A.L-G., Ł.A.P. and D.P.; Writing – Review & Editing, P.W., Ł.A.P., D.P., M.-R.M.-S. and D.S.; Visualization, P.W., P.T. and A.L-G.; Supervision, P.W., P.T., Ł.A.P. and D.S.; Project Administration, P.W., Ł.A.P., M.-R.M.-S. and D.S.; Funding Acquisition, P.W., Ł.A.P., M.-R.M.-S. and D.S.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
For this type of study formal consent is not required.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Wang, S.; Feng, Y.; Zheng, L.; He, P.; Tan, J.; Cai, J.; Wu, M.; Ye, X. Rosavin: Research Advances in Extraction and Synthesis, Pharmacological Activities and Therapeutic Effects on Diseases of the Characteristic Active Ingredients of Rhodiola rosea L. Molecules 2023, 28, 7412. [CrossRef]
- Tinsley, G. M., Jagim, A. R., Potter, G. D. M., Garner, D., & Galpin, A. J. (2023). Rhodiola rosea as an adaptogen to enhance exercise performance: a review of the literature. The British journal of nutrition, 1–13. Advance online publication. [CrossRef]
- Ivanova Stojcheva, E.; Quintela, J.C. The Effectiveness of Rhodiola rosea L. Preparations in Alleviating Various Aspects of Life-Stress Symptoms and Stress-Induced Conditions—Encouraging Clinical Evidence. Molecules 2022, 27, 3902. 2022; 27, 3902. [CrossRef]
- Rhodiola rosea: A possible plant adaptogen. Altern. Med. Rev. 2001, 6, 293–302.
- European Medicines Agency; Committee on Herbal Medicinal Products (HMPC). Assessment Report on Rhodiola rosea L., Rhizoma Et Radix; Document Reference. EMA/HMPC/232100/2011; European Medicines Agency: Amsterdam, The Netherlands, 2011.
- Khanna, K., Mishra, K. P., Ganju, L., & Singh, S. B. (2017). Golden root: A wholesome treat of immunity. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie, 87, 496–502. [CrossRef]
- Bernatoniene, J.; Jakstas, V.; Kopustinskiene, D.M. Phenolic Compounds of Rhodiola rosea L. as the Potential Alternative Therapy in the Treatment of Chronic Diseases. Int. J. Mol. Sci. 2023, 24, 12293. [CrossRef]
- Li, Y., Pham, V., Bui, M., Song, L., Wu, C., Walia, A., Uchio, E., Smith-Liu, F., & Zi, X. (2017). Rhodiola rosea L.: an herb with anti-stress, anti-aging, and immunostimulating properties for cancer chemoprevention. Current pharmacology reports, 3(6), 384–395. [CrossRef]
- Zhang, X., Xie, L., Long, J., Xie, Q., Zheng, Y., Liu, K., & Li, X. (2021). Salidroside: A review of its recent advances in synthetic pathways and pharmacological properties. Chemico-biological interactions, 339, 109268. [CrossRef]
- Guo, Y., Zhao, Y., Zheng, C., Meng, Y., & Yang, Y. (2010). Synthesis, biological activity of salidroside and its analogues. Chemical & pharmaceutical bulletin, 58(12), 1627–1629. [CrossRef]
- Huang, M., Sui, R., Zhang, L., Zhu, Y., Yuan, X., Jiang, H., & Mao, X. (2023). Rosavin thwarts amyloid-β-induced macromolecular damages and neurotoxicity, exhibiting anti-Alzheimer's disease activity in Wister rat model. Inflammopharmacology, 10.1007/s10787-023-01320-y. Advance online publication. [CrossRef]
- Zou, H., Li, L., Yang, Z., Tang, L., & Wang, C. (2023). Rosavin protects the blood-brain barrier against ischemia/reperfusion-induced cerebral injury by regulating MAPK-mediated MMPs pathway. Clinical and experimental pharmacology & physiology, 50(8), 664–676. [CrossRef]
- Gao, T., Li, J., Shi, L., & Hu, B. (2023). Rosavin inhibits neutrophil extracellular traps formation to ameliorate sepsis-induced lung injury by regulating the MAPK pathway. Allergologia et immunopathologia, 51(4), 46–54. [CrossRef]
- Liu, R., Jiang, C., Zhao, Z., Lv, Y., & Wang, G. (2023). Rosavin exerts an antitumor role and inactivates the MAPK/ERK pathway in small-cell lung carcinoma in vitro. Acta pharmaceutica (Zagreb, Croatia), 73(2), 269–280. [CrossRef]
- Albadawy, R., Hasanin, A. H., Agwa, S. H. A., Hamady, S., Aboul-Ela, Y. M., Raafat, M. H., Kamar, S. S., Othman, M., Yahia, Y. A., & Matboli, M. (2022). Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death. International journal of molecular sciences, 23(17), 10148. 17. [CrossRef]
- Park, J. S., Choi, J., Kwon, J. Y., Jung, K. A., Yang, C. W., Park, S. H., & Cho, M. L. (2018). A probiotic complex, rosavin, zinc, and prebiotics ameliorate intestinal inflammation in an acute colitis mouse model. Journal of translational medicine, 16(1), 37. 1. [CrossRef]
- Williams, S. A., Daigle, S. G., Weiss, R., Wang, Y., Arora, T., & Curtis, J. R. (2021). Economic Burden of Osteoporosis-Related Fractures in the US Medicare Population. The Annals of pharmacotherapy, 55(7), 821–829. [CrossRef]
- Si, L., Winzenberg, T. M., Jiang, Q., Chen, M., & Palmer, A. J. (2015). Projection of osteoporosis-related fractures and costs in China: 2010-2050. Osteoporosis international : a journal established as result of cooperation between the European Foundation for Osteoporosis and the National Osteoporosis Foundation of the USA, 26(7), 1929–1937. [CrossRef]
- Hernlund, E., Svedbom, A., Ivergård, M., Compston, J., Cooper, C., Stenmark, J., McCloskey, E. V., Jönsson, B., & Kanis, J. A. (2013). Osteoporosis in the European Union: medical management, epidemiology and economic burden. A report prepared in collaboration with the International Osteoporosis Foundation (IOF) and the European Federation of Pharmaceutical Industry Associations (EFPIA). Archives of osteoporosis, 8(1), 136. [CrossRef]
- Lajas, C., Abasolo, L., Bellajdel, B., Hernández-García, C., Carmona, L., Vargas, E., Lázaro, P., & Jover, J. A. (2003). Costs and predictors of costs in rheumatoid arthritis: a prevalence-based study. Arthritis and rheumatism, 49(1), 64–70. [CrossRef]
- Dong, N. Q., & Lin, H. X. (2021). Contribution of phenylpropanoid metabolism to plant development and plant-environment interactions. Journal of integrative plant biology, 63(1), 180–209. [CrossRef]
- Masi, F., Chianese, G., Hofstetter, R. K., Cavallaro, A. L., Riva, A., Werz, O., & Taglialatela-Scafati, O. (2023). Phytochemical profile and anti-inflammatory activity of a commercially available Rhodiola rosea root extract. Fitoterapia, 166, 105439. [CrossRef]
- National Center for Biotechnology Information (2023). PubChem Compound Summary for CID 9823887, Rosavin. Retrieved , 2023 from https://pubchem.ncbi.nlm.nih.gov/compound/Rosavin. 15 December.
- Zhang, W. , Zhang, W., Huo, L., Chai, Y., Liu, Z., Ren, Z., & Yu, C. (2021). Rosavin suppresses osteoclastogenesis in vivo and in vitro by blocking the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK) signaling pathways. Annals of translational medicine, 9(5), 383. [CrossRef]
- Komori, T. (2010). Regulation of osteoblast differentiation by Runx2. Advances in experimental medicine and biology, 658, 43–49. [CrossRef]
- da Silva Sasso, G. R. , Florencio-Silva, R., de Pizzol-Júnior, J. P., Gil, C. D., Simões, M. J., Sasso-Cerri, E., & Cerri, P. S. (2023). Additional Insights Into the Role of Osteocalcin in Osteoblast Differentiation and in the Early Steps of Developing Alveolar Process of Rat Molars. The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society, 71(12), 689–708. [CrossRef]
- Matsubara, T., Kinbara, M., Maeda, T., Yoshizawa, M., Kokabu, S., & Takano Yamamoto, T. (2017). Regulation of osteoclast differentiation and actin ring formation by the cytolinker protein plectin. Biochemical and biophysical research communications, 489(4), 472–476. 4. [CrossRef]
- Zhao, Q. , Wang, X., Liu, Y., He, A., & Jia, R. (2010). NFATc1: functions in osteoclasts. The international journal of biochemistry & cell biology, 42(5), 576–579. [CrossRef]
- Takayanagi, H. (2007). The role of NFAT in osteoclast formation. Annals of the New York Academy of Sciences, 1116, 227–237. [CrossRef]
- Omata, Y. , Tachibana, H., Aizaki, Y., Mimura, T., & Sato, K. (2023). Essentiality of Nfatc1 short isoform in osteoclast differentiation and its self-regulation. Scientific reports, 13(1), 18797. [CrossRef]
- Kim, J. H., Kim, K., Youn, B. U., Jin, H. M., Kim, J. Y., Moon, J. B., Ko, A., Seo, S. B., Lee, K. Y., & Kim, N. (2011). RANKL induces NFATc1 acetylation and stability via histone acetyltransferases during osteoclast differentiation. The Biochemical journal, 436(2), 253–262. 2. [CrossRef]
- McClung, M. (2007). Role of RANKL inhibition in osteoporosis. Arthritis research & therapy, 9 Suppl 1(Suppl 1), S3. [CrossRef]
- Ko, Y. , Lee, G., Kim, B., Park, M., Jang, Y., & Lim, W. (2020). Modification of the RANKL-RANK-binding site for the immunotherapeutic treatment of osteoporosis. Osteoporosis international : a journal established as result of cooperation between the European Foundation for Osteoporosis and the National Osteoporosis Foundation of the USA, 31(5), 983–993. [CrossRef]
- Chang, J. , Wang, Z., Tang, E., Fan, Z., McCauley, L., Franceschi, R., Guan, K., Krebsbach, P. H., & Wang, C. Y. (2009). Inhibition of osteoblastic bone formation by nuclear factor-kappaB. Nature medicine, 15(6), 682–689. [CrossRef]
- Boyce, B. F. , Li, J., Yao, Z., & Xing, L. (2023). Nuclear Factor-Kappa B Regulation of Osteoclastogenesis and Osteoblastogenesis. Endocrinology and metabolism (Seoul, Korea), 38(5), 504–521. [CrossRef]
- Cong, Q. , Jia, H., Li, P., Qiu, S., Yeh, J., Wang, Y., Zhang, Z. L., Ao, J., Li, B., & Liu, H. (2017). p38α MAPK regulates proliferation and differentiation of osteoclast progenitors and bone remodeling in an aging-dependent manner. Scientific reports, 7, 45964. [CrossRef]
- Yousefzadeh, N. , Kashfi, K., Jeddi, S., & Ghasemi, A. (2020). Ovariectomized rat model of osteoporosis: a practical guide. EXCLI journal, 19, 89–107. [CrossRef]
- van Straalen, J. P. , Sanders, E., Prummel, M. F., & Sanders, G. T. (1991). Bone-alkaline phosphatase as indicator of bone formation. Clinica chimica acta; international journal of clinical chemistry, 201(1-2), 27–33. [CrossRef]
- Nakamura, A., Dohi, Y., Akahane, M., Ohgushi, H., Nakajima, H., Funaoka, H., & Takakura, Y. (2009). Osteocalcin secretion as an early marker of in vitro osteogenic differentiation of rat mesenchymal stem cells. Tissue engineering. Part C, Methods, 15(2), 169–180. 2. [CrossRef]
- Zhang, W. , Yu, L., Wang, F., Chen, M., & Li, H. (2023). Rosavin regulates bone homeostasis through HDAC1-induced epigenetic regulation of EEF2. Chemico-biological interactions, 384, 110696. [CrossRef]
- Shakespear, M. R., Halili, M. A., Irvine, K. M., Fairlie, D. P., & Sweet, M. J. (2011). Histone deacetylases as regulators of inflammation and immunity. Trends in immunology, 32(7), 335–343. 7. [CrossRef]
- Cantley, M. D. , Zannettino, A. C. W., Bartold, P. M., Fairlie, D. P., & Haynes, D. R. (2017). Histone deacetylases (HDAC) in physiological and pathological bone remodelling. Bone, 95, 162–174. [CrossRef]
- Parveen, R., Harihar, D., & Chatterji, B. P. (2023). Recent histone deacetylase inhibitors in cancer therapy. Cancer, 129(21), 3372–3380. 21. [CrossRef]
- Chen, J. , Wang, J., Wu, X., Simon, N., Svensson, C. I., Yuan, J., Hart, D. A., Ahmed, A. S., & Ackermann, P. W. (2023). eEF2 improves dense connective tissue repair and healing outcome by regulating cellular death, autophagy, apoptosis, proliferation and migration. Cellular and molecular life sciences : CMLS, 80(5), 128. [CrossRef]
- Kwon, J. Y., Lee, S. H., Jhun, J., Choi, J., Jung, K., Cho, K. H., Kim, S. J., Yang, C. W., Park, S. H., & Cho, M. L. (2018). The Combination of Probiotic Complex, Rosavin, and Zinc Improves Pain and Cartilage Destruction in an Osteoarthritis Rat Model. Journal of medicinal food, 21(4), 364–371. 4. [CrossRef]
- Im, G. I., & Kim, M. K. (2014). The relationship between osteoarthritis and osteoporosis. Journal of bone and mineral metabolism, 32(2), 101–109. 2. [CrossRef]
- Kaspiris, A. , Hadjimichael, A. C., Lianou, I., Iliopoulos, I. D., Ntourantonis, D., Melissaridou, D., Savvidou, O. D., Papadimitriou, E., & Chronopoulos, E. (2023). Subchondral Bone Cyst Development in Osteoarthritis: From Pathophysiology to Bone Microarchitecture Changes and Clinical Implementations. Journal of clinical medicine, 12(3), 815. [CrossRef]
- McLean R., R. (2009). Proinflammatory cytokines and osteoporosis. Current osteoporosis reports, 7(4), 134–139. [CrossRef]
- Jehan, F. , Zarka, M., de la Houssaye, G., Veziers, J., Ostertag, A., Cohen-Solal, M., & Geoffroy, V. (2022). New insights into the role of matrix metalloproteinase 3 (MMP3) in bone. FASEB bioAdvances, 4(8), 524–538. [CrossRef]
- Zhang, Q. , Chen, B., Yan, F., Guo, J., Zhu, X., Ma, S., & Yang, W. (2014). Interleukin-10 inhibits bone resorption: a potential therapeutic strategy in periodontitis and other bone loss diseases. BioMed research international, 2014, 284836. [CrossRef]
- Shen, Y. , Winkler, I. G., Barbier, V., Sims, N. A., Hendy, J., & Lévesque, J. P. (2010). Tissue inhibitor of metalloproteinase-3 (TIMP-3) regulates hematopoiesis and bone formation in vivo. PloS one, 5(9), e13086. [CrossRef]
- Eek, D. , Krohe, M., Mazar, I., Horsfield, A., Pompilus, F., Friebe, R., & Shields, A. L. (2016). Patient-reported preferences for oral versus intravenous administration for the treatment of cancer: a review of the literature. Patient preference and adherence, 10, 1609–1621. [CrossRef]
- Zhang, J. K. , Yang, L., Meng, G. L., Yuan, Z., Fan, J., Li, D., Chen, J. Z., Shi, T. Y., Hu, H. M., Wei, B. Y., Luo, Z. J., & Liu, J. (2013). Protection by salidroside against bone loss via inhibition of oxidative stress and bone-resorbing mediators. PloS one, 8(2), e57251. [CrossRef]
- Fu, S. , Yan, M., Fan, Q., & Xu, J. (2022). Salidroside promotes osteoblast proliferation and differentiation via the activation of AMPK to inhibit bone resorption of knee osteoarthritis mice. Tissue & cell, 79, 101917. [CrossRef]
- Zheng, H. , Qi, S., & Chen, C. (2018). Salidroside Improves Bone Histomorphology and Prevents Bone Loss in Ovariectomized Diabetic Rats by Upregulating the OPG/RANKL Ratio. Molecules (Basel, Switzerland), 23(9), 2398. [CrossRef]
Figure 1.
The two-dimensional chemical structure of rosavin (C20H28O10). Oxygen (O) atoms are denoted in red, and hydrogen (H) atoms are indicated in blue.
Figure 1.
The two-dimensional chemical structure of rosavin (C20H28O10). Oxygen (O) atoms are denoted in red, and hydrogen (H) atoms are indicated in blue.
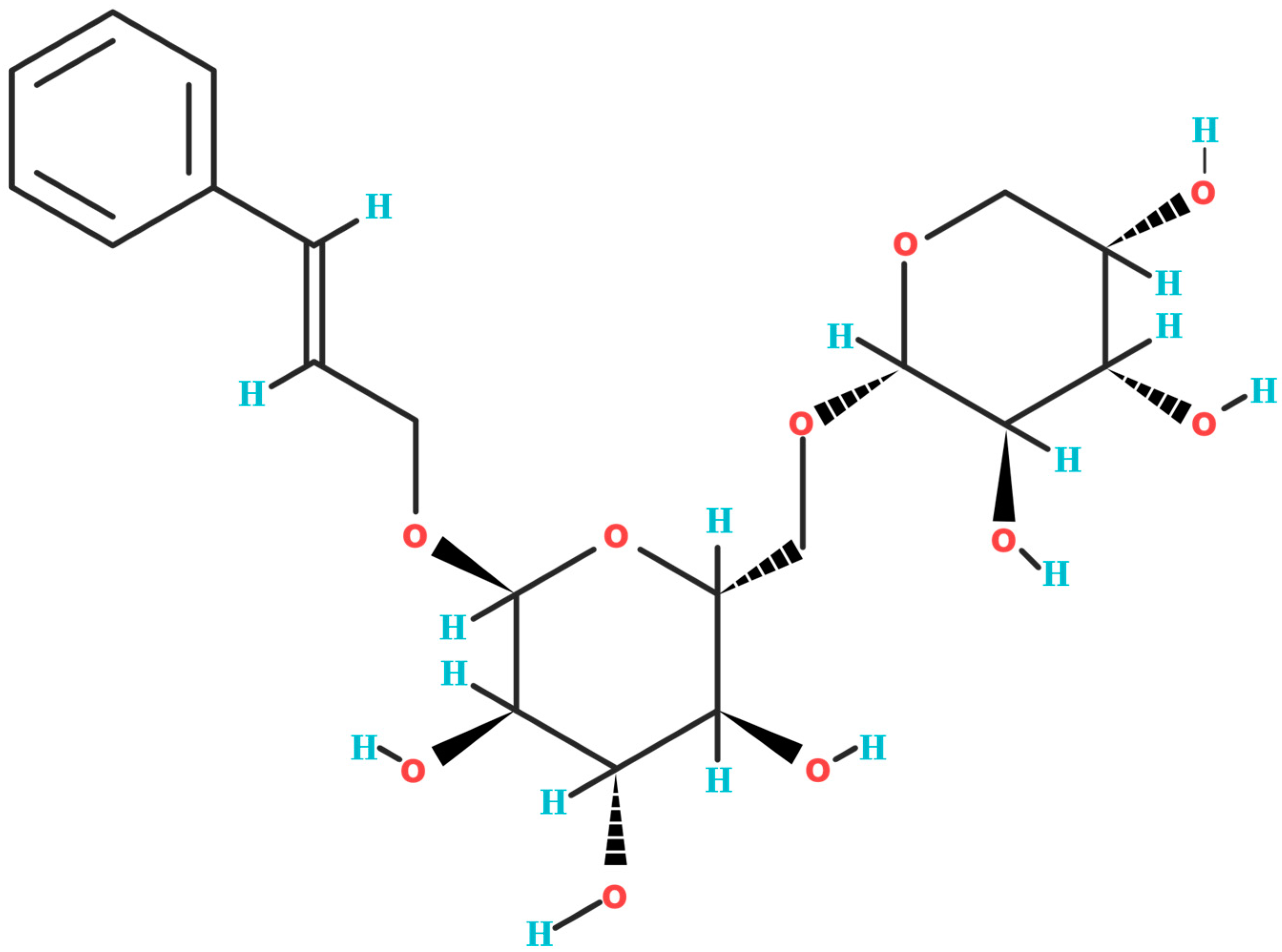
Figure 2.
A graphic diagram illustrating the impact of rosavin on the metabolism of murine bone tissue through the HDAC1/EEF2 signaling axis (postmenopausal osteoporosis model, PMOP). The illustration includes three-dimensional models of the murine femoral bone, presenting both macroscopic and microscopic perspectives (trabecular bone tissue). ↑ - increase, ↓ - decrease, HDAC1 - histone deacetylase 1, EEF2 - eukaryotic translation elongation factor 2, MAPK - mitogen-activated protein kinase, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, TRAP - tartrate-resistant acid phosphatase, NFATc1 - nuclear factor of activated T-cells cytoplasmic 1.
Figure 2.
A graphic diagram illustrating the impact of rosavin on the metabolism of murine bone tissue through the HDAC1/EEF2 signaling axis (postmenopausal osteoporosis model, PMOP). The illustration includes three-dimensional models of the murine femoral bone, presenting both macroscopic and microscopic perspectives (trabecular bone tissue). ↑ - increase, ↓ - decrease, HDAC1 - histone deacetylase 1, EEF2 - eukaryotic translation elongation factor 2, MAPK - mitogen-activated protein kinase, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, TRAP - tartrate-resistant acid phosphatase, NFATc1 - nuclear factor of activated T-cells cytoplasmic 1.
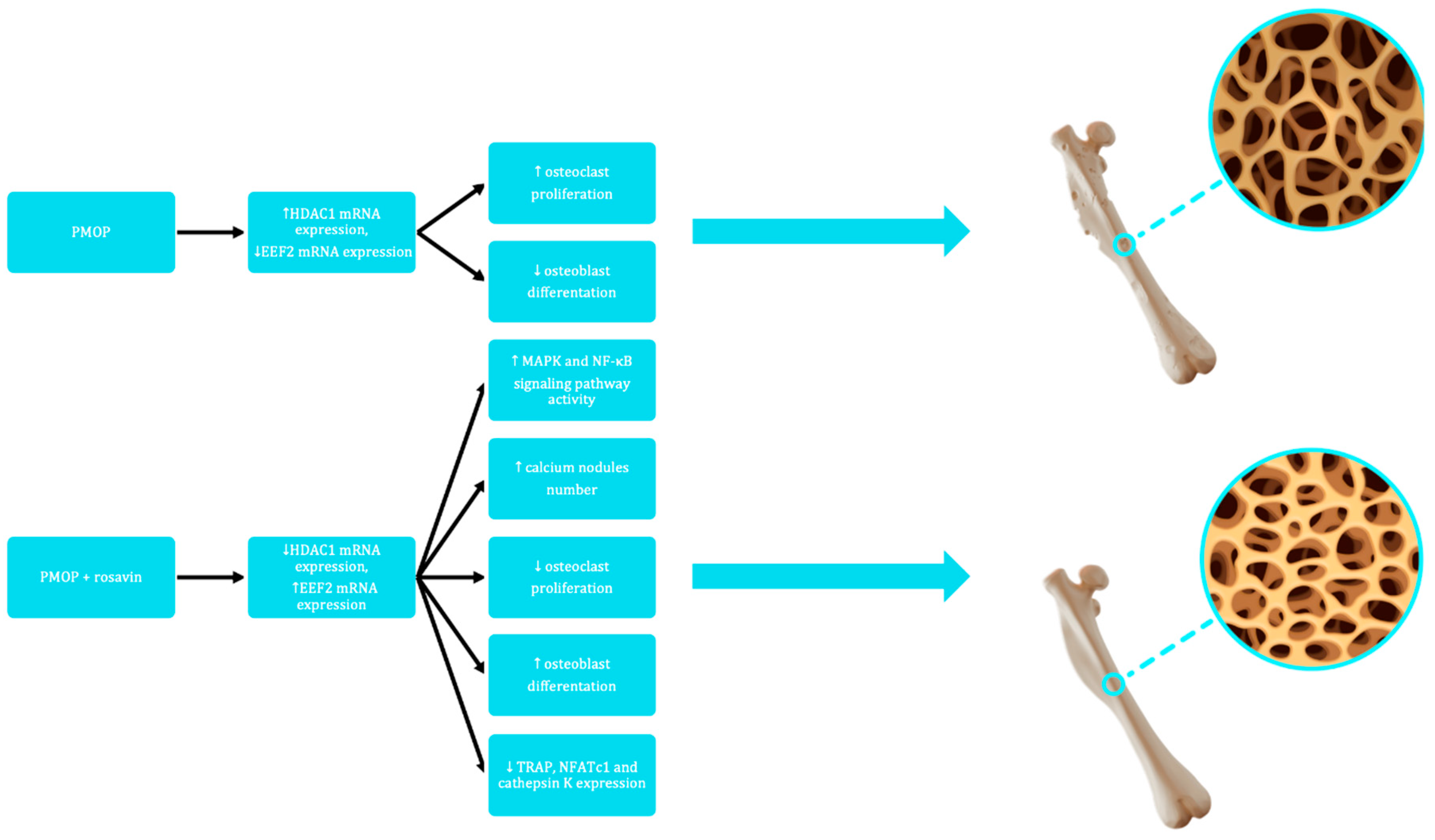
Table 1.
Biological manifestation of rosavin's impact in in vitro studies on specific cell cultures. (+) - stimulating/activating effect, (–) - inhibitory effect, (↔) - no effect, n/a - no data available, BMMSCs - bone marrow mesenchymal stem cells, RAW264.7 - murine macrophage cells, MC3T3-E - a mouse clonal, osteoblast-like cell line. TRAP - tartrate-resistant acid phosphatase, OCN – osteocalcin, Runx2 - mouse runt-related transcription factor 2, RANK - receptor activator of nuclear factor-κB, c-fms - colony-stimulating factor-1 receptor, CTR - calcitonin receptor, TRAF6 - tumor necrosis factor receptor-associated factor 6, MMP-9 - matrix metallopeptidase 9, NFATc1 - nuclear factor of activated T-cells cytoplasmic 1, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, MAPK - mitogen-activated protein kinase, ERK - extracellular signal-regulated kinase, p38 - human protein 38, p56 - human protein 56, JNK - c-Jun N-terminal kinase, IκBα - inhibitory subunit of NF-κB α, EEF2 - eukaryotic translation elongation factor 2, HDAC1 - histone deacetylase 1.
Table 1.
Biological manifestation of rosavin's impact in in vitro studies on specific cell cultures. (+) - stimulating/activating effect, (–) - inhibitory effect, (↔) - no effect, n/a - no data available, BMMSCs - bone marrow mesenchymal stem cells, RAW264.7 - murine macrophage cells, MC3T3-E - a mouse clonal, osteoblast-like cell line. TRAP - tartrate-resistant acid phosphatase, OCN – osteocalcin, Runx2 - mouse runt-related transcription factor 2, RANK - receptor activator of nuclear factor-κB, c-fms - colony-stimulating factor-1 receptor, CTR - calcitonin receptor, TRAF6 - tumor necrosis factor receptor-associated factor 6, MMP-9 - matrix metallopeptidase 9, NFATc1 - nuclear factor of activated T-cells cytoplasmic 1, NF-κB - nuclear factor kappa-light-chain-enhancer of activated B cells, MAPK - mitogen-activated protein kinase, ERK - extracellular signal-regulated kinase, p38 - human protein 38, p56 - human protein 56, JNK - c-Jun N-terminal kinase, IκBα - inhibitory subunit of NF-κB α, EEF2 - eukaryotic translation elongation factor 2, HDAC1 - histone deacetylase 1.
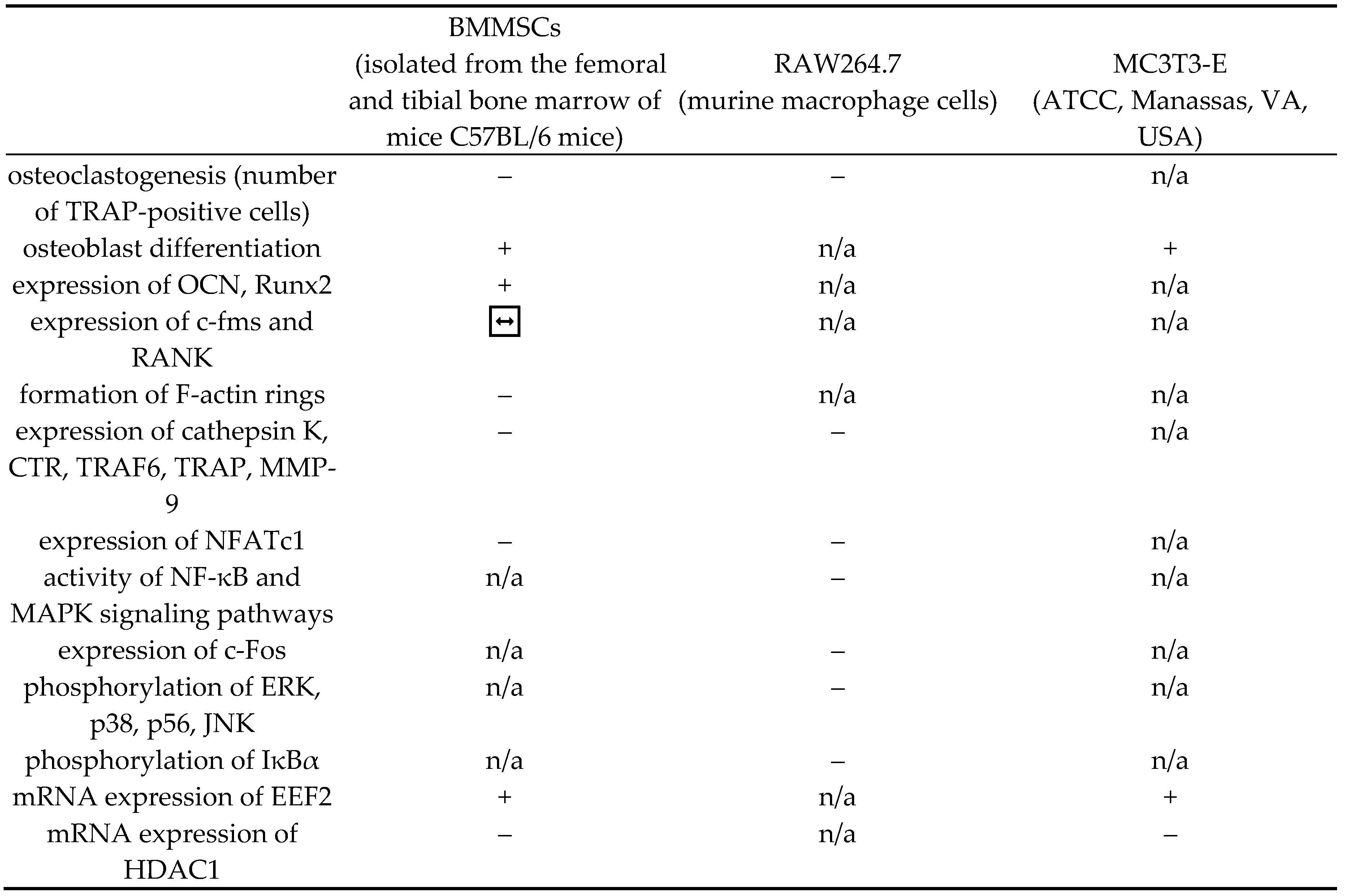
Table 2.
Table 2. Biological manifestation of rosavin's action in in vivo studies on blood serum, bone tissue and synovial membrane depending on the animal species and form of rosavin administration. ↑ - increase, ↓ - decrease, n/a - no data available, CTX-1 - cross linked C-telopeptide of type I collagen, TRAcp5b - tartrate-resistant acid phosphatase 5b, RANKL - receptor activator for nuclear factor κ B ligand, M-CSF - macrophage colony-stimulating factor, TRAP - tartrate-resistant acid phosphatase, ALP - alkaline phosphatase, OCN – osteocalcin, EEF2 - eukaryotic translation elongation factor 2, HDAC1 - histone deacetylase 1, MMP-3 - matrix metallopeptidase 3, IL-6 – interleukin 6, TNF-α – tumor necrosis factor α, IL-10 – interleukin 10, TIMP3 - tissue inhibitor of metalloproteinase 3
Table 2.
Table 2. Biological manifestation of rosavin's action in in vivo studies on blood serum, bone tissue and synovial membrane depending on the animal species and form of rosavin administration. ↑ - increase, ↓ - decrease, n/a - no data available, CTX-1 - cross linked C-telopeptide of type I collagen, TRAcp5b - tartrate-resistant acid phosphatase 5b, RANKL - receptor activator for nuclear factor κ B ligand, M-CSF - macrophage colony-stimulating factor, TRAP - tartrate-resistant acid phosphatase, ALP - alkaline phosphatase, OCN – osteocalcin, EEF2 - eukaryotic translation elongation factor 2, HDAC1 - histone deacetylase 1, MMP-3 - matrix metallopeptidase 3, IL-6 – interleukin 6, TNF-α – tumor necrosis factor α, IL-10 – interleukin 10, TIMP3 - tissue inhibitor of metalloproteinase 3
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



