
Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
The aim of this review was to search in literature, data regarding the inflammasome, involvement in, oral pathologies (mainly periodontitis) and other diseases and the approaches to control the inflammasome-related pathogenic mechanisms. The review also investigated how phytochemicals and pharmaceutical substances could be used to inhibit the inflammasome activity. Inflammasomes are involved in inflammatory, autoimmune and autoinflammatory diseases. Pharmaceutical substances can control the inflammasome by three mechanisms: inhibiting of intracellular signaling pathways (Allopurinol and SS-31), blocking inflammasome components (VX-765, Emricasan and VX-740), and inhibiting cytokines mediated by the inflammasome (Canakinumab, Anakinra and Rilonacept). Phytochemical inhibit the inflammasome, by neutralization of reactive oxygen species. Biomaterials functionalized by the adsorption of therapeutic agents on different nanomaterials may represent a future research direction to facilitate multimodal and sequential treatment in oral pathologies.
Keywords:
inflammation
; inflammasome
; inhibitors of the inflammasome
; nanomaterials
; functionalized biomaterials
; periodontitis
; general pathology
1. Introduction
Inflammation and inflammasome activation are the main mechanism underlying the oral pathologies, particularly periodontitis, and the inter-relation between oral and systemic diseases. Periodontitis is a chronic inflammatory disease, caused by pathogenic bacterial species from the dental biofilm, which also trigger the inflammatory-immune response of the host. Among periodonto-pathogenic species, the most prominent are Porphyromonas gingivalis and Aggregatibacter Actinomycetemcomitans. The presence of inflammatory and immune cells is essential for maintaining the periodontal health. Under physiological conditions, various bacterial species that colonize the gingival sulcus live in symbiosis. Once the homeostasis is altered, the imbalance between the host`s immune response and the microbiome leads to dysbiosis, with the consequent onset of periodontal disease. The inflammatory processes is the first activated defence mechanism against periodontopathogenic bacteria. The inflammasome is part of the innate immune response that activates inflammatory caspases. Once the inflammasome is activated, pro-inflammatory cytokines, such as interleukins, are released. Recently developed therapeutic approaches for controlling the inflammation propose, inflammasome inhibition by using, phytochemicals and pharmaceuticals. Pharmaceutical substances can control the inflammasome by three mechanisms: inhibiting intracellular signaling pathways, blocking inflammasome components, and inhibiting cytokines mediated by the inflammasome. Additionally, phytochemicals could inhibit inflammasome activation by neutralization of reactive oxygen species.
Periodontal disease progresses by the periodontal tissues breakdown, affecting both the periodonatl ligament and the alveolar bone. For bone replacement, the therapeutic guidelines involve complex surgical therapies, such as bone grafts, the use of platelet-rich plasma, or guided tissue regeneration. The use of nanobiomaterials could facilitate tissue healing and improves treatment outcomes. Pharmaceutical agents adsorbed on the surface of nanobiomaterials could be released in a controlled manner at the affected periodontal site. The detailed research of the physico-chemical properties of the phytochemical and pharmaceutical substances with inhibitory effect on the inflammasome, could determine the optimal method for their adsorption on various nanostructures and in order to be used for clinical applications. These materials present numerous advantages, such as the controlled release of therapeutical agents, as well as disadvantages represented by biodegradation and cytotoxicity. Although nanobiomaterials were intensively studied in various medical and related fields, the transfer of this information to the usual practical applicability has not yet been achieved. Through future research to reduce their disadvantages, functionalized nanobiomaterials could become important adjuvants in medical practice. The aim of this review was to search the specialized literature data regarding the inflammasome, involvement in oral pathology (mainly periodontitis) and other general disease and the approaches to control the inflammasome-related pathogenic mechanisms. The review also investigated how phytochemicals and pharmaceutical substances could be used to inhibit the inflammasome activity.
2. Literature review section
- 1.
- Neutrophils – activation and involvement in periodontal disease
Neutrophils play a major role in periodontal health and disease. The absence or functional deficits of neutrophils induce the onset of severe forms of periodontitis. Thus, functional neutrophils have proven to be essential for periodontal homeostasis. However, increased number of neutrophils or their hyperreactivity, causes imbalanced interactions between the host and the microbiome, which leads to dysbiosis and periodontal breakdown [1]. Dysbiosis results from the disruption of homeostasis between the host's immune system and the oral microbiome. Thus, microbial communities develop pathogenic mechanisms to overcome the immune response of the host, causing an inflammatory reaction and the progression of periodontal disease. In the study carried out by Williams et al, a single-cell transcriptome atlas of the dental and gingival tissues was prepared to identify the cell populations that predominate in health and disease states. According to the epithelial analysis of biopsies from the oral and gingival tissue of patients with periodontitis, an important correlation was observed between inflammation and neutrophil activation [2]. Neutrophils represent the most numerous inflammatory cells, which are implicated in the innate immunity. Neutrophils exert antimicrobial responses and have the ability to release inflammatory mediators, that interact with pathogenic microorganisms. Under physiological conditions neutrophil`s responses towards the microbial communities maintain a self-limiting inflammation. In case of dysbiotic microbial attack, neutrophils show an intensified activity, leadings to dysbiotic inflammatory responses, which results in progression of the disease. The transition from the state of symbiosis, of commensal microbial communities, to the state of dysbiosis, causes tissue inflammation, by involving mechanisms of both the innate and adaptive immunity [3]. These include the complement system, CD4 T cells and neutrophils, which, through inflammation, cause the activation of osteoclasts and the resorption of alveolar bone [4] (Figure 1).
Neutrophils possess the ability to respond to the attack of microorganisms, which tend to develop survival strategies during disease progression [5]. The hyperreactive and overnumbered neutrophils discharge toxic substances in the mucosal tissues, thus contributing to tissue damage. The role of neutrophils in maintaining oral health was demonstrated by the association between neutrophilia, neutropenia and neutrophil functional deficits and periodontal disease [6]. Antimicrobial functions of neutrophils include: phagocytosis, degranulation and extrusion of chromatin DNA by forming neutrophils extracellular traps (NETs). Degranulation represents an antimicrobial attack, within the function of neutrophils, with major implications on the progression of periodontal disease [7]. Presence of myeloperoxidase in the contents of the granules determines their classification into positive and negative peroxidases. In addition to the positive peroxidases, there are other powerful anti-microbial compounds in the matrix composition, such as: serine proteases, neutrophil elastase (NE), proteinase 3 (PR3) and cathepsin G. The negative peroxidase contains antimicrobial compounds such as lactoferrin, lysozyme and metalloproteinases (MMP): gelatinase and collagenases [8]. The degree of neutrophils stimulation determines the process of hierarchical granular exocytosis. The sequence of neutrophils degranulation takes place inversely to granules formation, during maturation in the bone marrow. Literature data of the last two decades showeds that neutrophils are not limited to their antimicrobial capacity, in the context of inflammation [9]. Neutrophils possess functions with both antimicrobial and immunomodulatory effects. Neutrophils response to the microbial attack must be properly regulated, in order to minimize the negative effects on the host tissue. Priming is a reversible high alert state of neutrophils, which causes phagocytosis, degranulation, NETs formation and the release of inflammatory mediators [10]. Neutrophil priming is induced by endogenous inflammatory mediators: cytokines, growth factors, chemokines and by lipopolysaccharides (LPS). When neutrophil priming occurs during chronic systemic inflammation, it causes harmful effects [11]. Circulating neutrophils priming can occur through exposure to low systemic levels of cytokines or chemokines. When these neutrophils reach the gingival sulcus, a secondary stimulation, by local microorganisms leads to an exacerbated global response. Thus, neutrophils priming can have both beneficial and harmful effects. Reduced priming is associated with recurrent infections [12], whereas increased priming is associated with chronic inflammatory oral diseases, such as particularly periodontitis, and systemic diseases, such as rheumatoid arthritis or sepsis [13,14,15]. Circulating neutrophils isolated from patients with periodontitis showed increased production of reactive oxygen species (ROS) upon exposure to a soluble stimulus, such as phorbol myristate acetate (PMA), or to bacterial stimulation by Porphyromonas gingivalis, compared to the response elicited by cells from a healthy control group [14].
Primed cells with an altered functional state do not return to the initial state, thus improving the inflammatory response in the short term, in contrast to the trained cells returning to the initial state, which have long-term effects. The changes that persist in primed innate immune cells are called epiphenetic changes, and determine the emergence of immune memory [16] (Figure 2).
In case of dysbiosis, the pathogenic bacterial communities develop various survival mechanisms, to increase the resistance to the host's immunity, but also to benefit from the host's inflammatory response. Some pathogens can survive the immune attack, by taking advantage of molecules derived from neutrophils, while other pathogens survive by developing resistance to the antimicrobial attack [7]. In periodontitis patients, active oral lesions create an inflammatory state, which tends to provide a favorable environment for the growth and development of oral pathogens.
Porphyromonas gingivalis is a Gram-negative oral pathogen, strictly anaerobic and azacharolytic, which predominantly populates the gingival sulcus [17]. The pathogen is able to evade the antimicrobial attack of macrophages and neutrophils, but it also benefits from a nutritional substrate due to the gingival inflammatory state. Neutrophils participate in the antimicrobial attack, initially, by recognizing pathogens, due to the recognition receptors (PRR), the most important of which is the Toll-Like receptor (TRL), capable to recognize distinct microbial structures [18]. Another mechanism for maintaining symbiosis, with the involvement of the innate immune system, is through the participation of the complement system comprising approximately 50 types of proteins [19]. The complement system is activated by three pathways: the classical, the alternative and the lectin pathway. All these activation pathways converge towards the central C3 component [20] (Figure 3). Porphyromonas gingivalis first factor contacts cells of the innate immunity by binding to CD14, which is a crucial coreceptor for TLR2 activation. Thus, the family of enzymes phosphatidylinositol 3-kinases (PI3Ks) is activated, which acts on β2 integrine in monocytes and neutrophils [21]. Porphyromonas gingivalis can also bind to complement receptor 5 (C5aR1) and C5a ligand. Porphyromonas gingivalis expresses cysteine proteases, called gingipains, providing the necessary virulence to evade the antimicrobial response [22]. Gingipains can cleave the C5 component of the complement, leading to formation of the C5a, a strong chemotactic neutrophil molecule, and the C5b, which prevents terminal complement activation. According to a recent study, blockage or genetic absence of the C5aR1 receptor reduce the colonization of Porphyromonas gingivalis [23].
Aggregatibacter Actinomycetemcomitans is a Gram-negative, facultative anaerobic and non-motile coccobacillus from the Pasteurellaceae family. As an opportunistic oral pathogen, it possesses several virulence factors, responsible for its survival [24]. Aggregatibacter Actinomycetemcomitans expresses a two-component receptor called QseBC, which enables bacterium activation by the combination of iron and catecholamines (epinephrine) [25]. A recent study demonstrated that neutrophils are a source of catecholamines, due to epinephrine stored in the azurophilic granules. By increasing iron levels in the microenvironment and by using epinephrine from neutrophils, Aggregatibacter Actinomycetemcomitans growth is stimulated, which results in the induction of the QseBC operon [26].
The "neutrostat" concept defines the homeostatic mechanism of neutrophils’ recruitment and efferocytosis into the tissues. Efferocytosis of apoptotic neutrophils enables their elimination by tissue macrophages; thus, secondary necrosis is prevented and IL-23 expression is inhibited. Inhibition of IL-23 in conjunction with efferocytosis, leads to further reduction in IL-17 production by CD4 T cells, subsequently affecting the production of neutrophils in the bone marrow [27].
Hypercholesterolemia and hyperglycemia are considered favorable factors for the modification of neutrophil production and functionality [28]. Hypercholesterolemia is associated with increased production and mobilization of neutrophils, "primed" circulating capacity and exaggerated response to secondary stimulation [29]. In hyperglycemia, some neutrophil products can produce high amounts of ROS and increase NETs formation [30].
Complement represents a key element in inflammation, due to its involvement in neutrophils recruitment and their optimal functionality. Neutrophils, through specific receptors, respond to the complement activation fragments, C3a and C5a [31]. A recent study demonstrated that inhibition of the C3 fraction blocked periodontal inflammation and reduced bone loss in non-human primates (NHP) [32].
- 2.
- Activation of the inflammasome
Inflammation represents a quick and early defense response against pathogens. The inflammatory response can be physiological, as a result of the interaction between the host and different stimuli, thus offering a rapid protection against the attack [33].
The inflammasome is a pathway of the innate immune system that activates inflammatory caspases, which are a family of cysteine proteases [34]. According to a study from 2002, it was demonstrated that the inflammasome is a multiprotein complex, activated by different pathogens, which through its activation determines the activation of caspase-1. The term inflammasome has been used since 2009, after the publication of a study conducted by Bostanci et al [35]. Inflammasomes have different names according to the class of PRR sensor molecule that induces their activation [36]. NLR proteins were demonstrated to play regulatory roles in the innate immune response [37,38].
The most important inflammasomes include: NLRP2, NLRP6, NLRC4 and AIM2 [39]. The first discovered inflammasome was NLRP1, which is activated by Bacillus Anthracis and mediates the activation of caspase-1 [40]. Several studies demonstrated that the NLRP1 inflammasome is not significantly activated neither by Porphyromonas Gingivalis [41] nor by Aggregatibacter actinomycetemcomitans [42]. The NLRP2 inflammasome, inhibits NF-kB activation [43]. NLRP3 inflammasome can be activated by a number of factors, including: LPS, Staphylococcus Aureus, Klebsiella Pneumoniae and influenza virus [44], but also uric acid and ATP [45]. The NLRP6 inflammasome is involved in intestinal homeostasis [46], and through infection with Aggregatibacter Actinomycetemcomitans reduces its activity [42]. NLRP7 inflammasome is associated with Staphylococcus Aureus infection [47], but there is no scientific evidence to demonstrate its link with periodontal disease. NLRP12 inflammasome inhibits the activation of NF-kb and activates TLR and TNF-α [48]. NLRC4 inflammasome controls the release of IL-1β and intervenes on NF-kB [49], and NLRC5 inflammasome acts similarly to the NLRP3 inflammasome, decreasing the activity of caspase-1, interleukins IL-1β and IL-18 [50].
The inflammasome, a multi-protein oligomer, has been found to be involved in inflammatory processes [51]. Inflammasomes are classified into Nucleotide-binding domain leucine-rich repeat (NLR) inflammasomes and PYHIN (pyrin and hematopoietic interferon) inflammasomes [52]. Furthermore, the NLR class is divided into the NBD (nucleotide binding domain) subclass, which are involved in ribonucleotide binding, regulation of oligomerization and inflammasome assembly, and the LRR (leucine-rich proteins) subclass, which are involved in the recognition of PAMPs and in interprotein interactions [53]. According to the specialized literature, it is known that the NLRP3 inflammasome has a critical role in the activation of caspase-1 and the proteolytic processing of pro-IL-1β [39]. Caspases modulate the immune response by cleaving specific cellular substrates [34]. Inflammasome activation causes the release of pro-inflammatory cytokines and initiates a mechanism of programmed cell death called pyroptosis [54]. Apoptosis was the first type of cell death described as a “programmed cell death” in which the cell plays an active role in its own destruction [55]. Other mechanisms of cell death are represented by autophagy (the consumption of its own components in an organism subjected to starvation) [56], necroptosis (a type of programmed necrosis that occurs during caspase inhibition or insufficient caspase activation) [57], ferroptosis (programmed cell death dependent on iron and characterized by the accumulation of lipid peroxides) [58], NETosis (the process by which extracellular neutrophil traps are formed) [59] and pyroptosis (a destruction mechanism mediated by caspases) [60].
Pyroptosis was discovered in 1992 in macrophages infected with Shigella Species [61], and later in infections with Salmonella typhimurium [62]. This is a process of self-destruction mediated by caspases, which sets in quickly and causes the release of pro-inflammatory cytokines [63]. The name pyroptosis is derived from the Greek words "pyro", meaning fire, and "ptosis" which means falling [62]. Pyroptosis also induces a series of morphological and physiological changes, associated with the inflammatory reaction [60]. Due to the pores in the membrane of pyroptotic cells, the membrane permeability is altered [64]. By changing the permeability, the virus are passed from the intracellular compartment to the extracellular one, causing their increased susceptibility to immune attack [60]. The subsequent release of pro-inflammatory cytokines from the cytoplasmic granules into the extracellular environment, induces cell death, and implicitly an inflammatory response [65]. Contrarily to pyroptosis, apoptosis is a controlled cell death that does not result in inflammation or destruction of adjacent cells [66]. One of the mechanisms for activation of inflammasome and caspase-1 is triggered by staphylococcal α-hemolysin, a bacterial toxin produced by Staphylococcus aureus [67]. Another mechanism is based on the activation of caspase-1, due to lower levels of intracellular potassium (K), induced by nigericin, a toxin produced by Streptomyces hygroscopicus [68]. The nigericin-K complex, formed by nigericin and K, is transported across the cell membrane and released extracellularly [69]. Lipopolysaccharides are also able to activate caspases and induce gasdermin D (GSDMD) cleavage [33].
Pyroptosis is a process of cellular self-destruction mediated by caspases. When cells are stimulated by pathogens, the formation of inflammasomes is determined. Pyroptosis is mediated by the activation of caspase-1 by the NLRP3 inflammasome. Caspase-1 cleaves GSDMD, causing cell membrane cleavage, a process termed canonical inflammasome activation [63]. In contrast, the non-canonical activation of pyroptosis is determined by the activation of caspase-4 and 5 in humans and caspase-11 in rats, which will cause the cleavage of GSDMD. Although pyroptosis mechanisms can be activated by different signaling pathways, they all lead to the activation of GSDMD. Subsequently, cytoplasmic molecules such as interleukins 1β (IL-1β) and 18 (IL-18) are released by GSDMD and trigger an inflammatory response [70] (Figure 4). Pyroptosis is mediated by the activation of caspases [71]. Even though pyroptosis is a protective mechanism against pathogens, its overactivation causes damage to adjacent tissues [72]. The canonical pathway is represented by the detection of the pathogenic signal, followed by the formation of inflammasomes, due to the immune cells of the innate immunity, which activate capsase-1. The non-canonical inflammasome pathway requires the presence of caspases 4 and 5, which will trigger pyroptosis. Caspase-1 cleaves GSDMD into two fragments: the N-terminal fragment and the C-terminal fragment. The N-terminal fragment causes the formation of pores of small size (10-15 nm) at the level of the cell membrane, which allow the efflux of the cytoplasmic content and an increased efflux of potassium, which activates caspase-1, through the NLRP3 inflammasome [73]. Pyroptosis also induces the release of cytokines, such as IL-1β and IL-18. IL-1β stimulates vasodilatation, leukocyte migration and extravasation of immune cells, and IL-18 promotes interferon-γ production and activates T cells and macrophages [60] (Figure 4).
IL-1 consists of 2 subunits: IL-1α induces production of inflammatory mediators, differentiation of osteoclasts, and causes the destruction of alveolar bone and periodontal connective tissue, whereas the IL-1β is involved in the progression of periodontal disease [74]. IL-18 is produced in an inactive form and is activated by caspase-1, with the implication of the inflammasome. The inflammasome consists of NLR, which is a leucine-rich receptor [75]. Several types of NLRs are involved in inflammasome activation, the most common of which is NLRP3 [76]. A number of activators of the NLRP3 inflammasome have been discovered, such as: silicon particles, asbestos, cholesterol crystals, bacterial toxins, potassium efflux, cathepsin B, ribosomal mRNA, bacterial ATP, oxidized mitochondrial DNA, ROS, blocking function ribosomes, bacterial RNA, lipopeptides, guanylate-binding protein 5 and mycoplasmas [77], extracellular ATP and uric acid crystals. For inflammasome activation, a sensor (NLRP3) is required that fuses with an adapter (apoptosis-associated speck-like protein – ASC adapter), causing the formation of a filamentous structure that provides a platform for Caspase-1 cleavage [78]. The NOD3 inflammasome receptor protein (NLRP3) is activated by a series of microbial and metabolic signals. A priming step involving Toll-like receptors (TLRs) is required, causing the increase of IL-1 and the inflammasome components, Caspase-1 and NLRP3. Afterwards, the components of the inflammasome gather in the cytoplasm and Caspase-1 is cleaved [79]. Recent studies demonstrated that priming induces the synthesis of mitochondrial DNA and the mitochondrial enzyme deoxyribonucleotide kinase, which activates the NLRP3 inflammasome [80].
2.1. Implications of the inflammasome in periodontal disease
The inflammasome activation in the periodontal tissues is an important mechanism in pathogenesis of periodontal disease. Periodontal disease is an infectious chronic inflammation leading to destruction of the supporting periodontal tissues [33]. Periodontitis appears in response to microbial invasion by Gram-negative anaerobic bacteria, such as: Porphyromonas gingivalis, Aggregatibacter actinomycetemcomitans, Treponema denticola, Fusobacterium nucleatum, Tannerella forsythia and others [63]. Numerous studies reported that inflammasomes levels [81], NLRP3 expression [82], caspases and IL-18 tissue concentrations are increased in periodontal disease compared to healthy tissues [83]. There is contradictory data regarding the influence of Porphyromonas gingivalis on inflammasome, some studies indicating inflammasome activation and IL-1β production, whereas other studies reported inflammasome inhibition [77,84].
Pyroptosis represents one of the pathways leading to exacerbation of periodontal inflammation. According to current studies, the virulence factors present in the affected periodontal tissues induce inflammasome activation and tissue destruction through caspase activation, cleavage of gasdermin D and secretion of interleukins IL-1β and IL-18 [55] (Figure 4). The involvement of IL-1 in periodontal disease was mentioned for the first time in 1985 [85], and then in 1991, the role of IL-1β subunit was demonstrated [86]. According to a study carried out in 1992, IL-1 levels were increased in subjects with gingivitis [87], and IL-1 could be considered an early marker of periodontal inflammation. A study performed in 1997 demonstrated that IL-1 and TNF play an important role in pathogenesis of periodontal disease [88]. In 2009, it was demonstrated that NLRP3 and NLRP2 presented increased levels in affected periodontal tissues, compared with healthy tissues [35], confirming the involvement of the NLRP3 inflammasome in the pathogenesis of periodontitis Bostanci et al. reported biofilm-induced upregulation of inflammasome transcription causing early inflammation in periodontal tissues, whereas downregulation of transcription enabled survival of periodontal pathogens, thus demonstrating that biofilm can differentially modulate the activity of inflammasomes [89]. A study conducted in 2014 demonstrated a strong correlation between NLRP3 and advanced periodontitis [90]. According to Xue et al, the level of NRLP3 was higher in patients with chronic periodontitis or generalized aggressive periodontitis, compared to healthy subjects [91]. Porphyromonas gingivalis can activate the inflammasome through the involvement of gingipains [92].
2.2. Implications of the inflammasome in the general pathologies
The inflammasomes also have impact at systemic level. Inflammasomes represent protein complexes, located in the cytosol, which, through pathogenic stimulation, cause the appearance of inflammatory processes [93]. Inflammasomes cause the cleavage of pro-inflammatory cytokines, such as IL-1β and IL-18 [94]. The NLRP3 inflammasome reacts to a wide range of bacterial ligands, such as: LPS, bacterial RNA and peptidoglycans. They are involved in the pathogenesis of periodontal disease, rheumatoid arthritis and osteomyelitis [63]. Caspase-1 is involved in the pathogenesis of various general pathologies such as Alzheimer's disease, Crohn's disease, rheumatoid arthritis, cardiovascular diseases and endometriosis [55], thus becoming a target for pharmaceutic agents.
According to a study conducted in 2015, NLRP3 and IL-1β are increased in subjects with chronic periodontal disease associated with type 2 diabetes, compared to healthy subjects [95]. Inflammasome activation can be induced by ROS, due to mitogen-activated protein kinases (MAPK). Numerous pathologies are characterized by increased ROS production, with indirect effects on inflammasomes activation; these pathologies include: viral infections, gout and diabetes [39]. NLRP3 inflammasome activation can also be induced without ROS generation or lysosomal damage [96]. Deficiency of 25-hydroxycholesterol, which is an inhibitor of cholesterol biosynthesis, triggers the release of mitochondrial DNA and activates the inflammasome and the secretion of IL-1 and IL-18 [97]. According to a recent study, cholesterol crystals can cause the release of NETs and the activation of the macrophage inflammasome [98].
Several studies demonstrated the involvement of the NLRP3 inflammasome in the body's immune responses to viral, fungal and parasitic attacks. Dengue is a viral infection caused by the Dengue virus (DENV), transmitted to humans through the bite of infected mosquitoes, leading to inflammasome activation, caspase-1 activation and IL-1β secretion [99]. Segovia et al. demonstrated a connection between the NLRP3 inflammasome and respiratory syncytial virus [100]. Additionally, caspase-1 and IL-1β secretion was activated by infection with Mycobacterium Tuberculosis [101]. Another study demonstrated inflammasome activation by the crystalline compound alum, found in human vaccines [102]. Activation of the inflammasome was also observed in patients with diabetes [103], chronic kidney diseases [104] and autoimmune pathologies such as lupus erythematosus [105,106].
- 3.
- Production of reactive oxygen species mediated by inflammasome
The NLRP3 inflammasome senses membrane defects and induces an efflux of K+ ions, which generates mitochondrial ROS, which a major role in inflammation [96].
Oxygen is the vital element for survival, but in high concentrations, it becomes toxic, due to oxygen radicals, such as superoxide, hydroxyl, peroxyl and hydroperoxyl [107]. Through association with hydrogen peroxide and ozone, ROS are formed. The human body constantly produces ROS, but also reactive nitrogen species (RNS). According to current data, the following six reactive species are harmful to the human body: superoxide anion, hydrogen peroxide, peroxyl radicals, hydroxyl radical, singlet oxygen and peroxynitrite. Hydrogen peroxide is the reactive oxygen species, which is continuously produced in most tissues of the body, the main producer being the mitochondria [108]. The human body fights against ROS and RNS through different antioxidants, which transform the reactive species into harmless compounds. Two main subtypes of antioxidants were described: the preventive antioxidants, which are the first line of defense to stop the formation of reactive species, and the chain breakers, with secondary role. Additionally, the tertiary line of defense removes detritus and restores functionality, and the quaternary line acts through adaptation mechanisms [109]. Consumption of vegetable foods decreases oxidative stress [110], whereas foods based on animal protein increase the risk of cardiovascular diseases [111]. Polyphenols have become the most important antioxidants [112].
In neutrophils, a chain of metabolic reactions is triggered, which activates protein kinase C and induces nicotinamide adenine dinucleotide phosphate (NADPH) oxidase to decrease oxygen and form superoxide radical, which will later be metabolized in other ROS [83]. ROS are stored in phagosomes or released extracellularly, to act on pathogens that cannot be phagocytosed. Once the homeostatic balance is broken, neutrophils can induce lesions, which cause systemic inflammation [84].
ROS can directly activate caspase-1; additionally, by an indirect mechanism involving ERK1/2, ROS activate caspase-3. ERK1/2 is implicated in inflammation by guiding cells to a proapoptotic state or an antiapoptotic state [39]. Kuiper indicated a model of NLRP3 inflammasome activation, through the APANAREI axis, which includes the P2X7 receptor, NOX, ROS, ERK1/2, NLRP3, and caspase-1. Thus, the APANAREI axis involves the activation of the P2X7 receptor, mediated by ATP, which induces the production of ROS, which in turn determines the activation of ERK1/2, then the activation of caspase-1 and finally the activation of the NLRP3 inflammasome [113] (Figure 5).
- 4.
- Inflammasome inhibitors
4.1. Direct inhibitors of the inflammasome
Several pathogens can inhibit inflammasome activation, including: Mycobacterium Tuberculosis, Yersinia specia, Legionella Pneumophila and Pseudomonas Aeruginosa [77], as well as Myxoma virus and Shope fibroma virus [114]. Inflammasome inhibition can be achieved by 3 mechanisms (Figure 6): (i) inhibition of intracellular signaling pathways; (ii) blockage of inflammasome components; (iii) inhibition of cytokines mediated by inflammasome [36].
For the inhibition of signaling pathways, two pharmaceutical preparations can be mentioned: Allopurinol and SS-31. Allopurinol (Zyloprim/Aloprim®) was discovered in the 1940s in the Burroughs Wellcome program. This pharmaceutical preparation has been used for the treatment of gout and hyperuricemia, but also in type 2 diabetes [115]. SS-31 (Elamipretide, Bendavia, MTP-131) is a pharmaceutical preparation used to stabilize cardiolipin from the mitochondrial membrane; thus, the excessive production of ROS is prevented [116]. This pharmaceutical preparation has been used for the treatment of gout and hyperuricemia, but also in heart failure [117].
For blocking the inflammasome components, three pharmaceutical substances can be used: VX-765, Emricasan and VX-740. VX-765 is a reversible inhibitor of caspase-1 [118], used for the treatment of psoriasis and epilepsy [119]. Emricasan has possible indications in the treatment of diabetes and non-alcoholic hepatic steatosis [120]. The best preparation in this category is represented by VX-740 (Pralnacasan) [121]. Colchicine can also be mentioned; this pharmaceutical preparation has been used for over 3000 years for the treatment of gout [122], and is also indicated in heart diseases [123]. It has anti-inflammatory and antioxidant effects, but it is also a protector of bone structure [10]. The main mechanism of action is by blocking the assembly of microtubules [124].
Several methods have been described for inhibiting the cytokines mediated by inflammasome. Inhibition of IL-1β can be achieved using a monoclonal antibody, Canakinumab, or an antagonist of the IL-1β receptor called Anakinra, which can also be achieved through a soluble receptor, Rilonacept [36]. These pharmaceutical preparations were used for the treatment of rheumatoid arthritis and diabetes [125,126]. Canakinumab (ILARIS) is indicated for the treatment of rheumatoid arthritis and atherosclerosis [127]. Rilonacept (Arcalyst) is used to treat Mediterranean fever [128]. Anakinra (Kineret) is indicated in the treatment of type 2 diabetes and Behcet's disease [129].
4.2. Indirect inhibitors of the inflammasome by acting on reactive oxygen species
It has been shown that plant antioxidants can interfere with oxidative stress, through the redox properties of plant polyphenols acting as reducing agents and chelating metal ions [130]. Thus, it is suggested that phytochemicals may have beneficial effects [131], since ROS are neutralized by non-enzymatic antioxidants, such as vitamins C and E, but also by phytochemicals.
Aronia Melanocarpa is a plant from the rosaceae family, which originates from North America and Russia, and its fruits contain high levels of flavonoids, including: proanthocyanins, anthocyanins, flavonols and catechins, but also phenolic acids, which are hydroxylated derivatives from benzoic acid and cinnamic acid [32]. These fruits present a series of advantages due to the antidiabetic, antimutagenic and anticarcinogenic effects, as well as hepato- and cardio-protective properties [109]. In 2005, Ohgami et al. demonstrated the anti-inflammatory effect of Aronia Melanocarpa extract, suggesting that 100 mg of extract has similar effects to 10 mg of prednisolone [133].
According to Tauber et al., flavonoids controlled the production and release of ROS by directly scavenging reactive oxygen [134]. Some flavonoids can scavenge hypochlorous acid and reactive chlorinated species, whereas other flavonoids can suppress the release of arachidonic acid from cell membranes [135]. Flavonoids can also inhibit cyclooxygenase and lipoxygenase and reduce the release of hydrolytic enzymes from lysosomes, according to the study of Tordera et al [136]. In 1998, Middleton demonstrated that flavonoids can inhibit protein kinases [135]. Moreover, there may be a pathway to mitigate the activity of inflammasomes, by using antioxidants [39] (Figure 6).
Studies have shown that by preventing the production of ROS, the production of IL-1β is also inhibited, but the production of IL-18 is not [137].
- 5.
- Biomaterials functionalized with inflammasome inhibitors
Although nanobiomaterials are intensively studied in various medical and related fields, the transfer of this information to the usual practical applicability has not yet been achieved. These can present a series of benefits and advantages that improve the quality of treatments, but also a series of limitations and adverse effects that have not been sufficiently studied in the literature. By studying the physico-chemical properties of the various pharmaceutical preparations, various ways of incorporating them into specialized matrices could be discussed, for an easier therapy of the various pathologies of the oral cavity. According to the specialized literature, there are recently published studies that deal with these topics and that can be helpful in discovering various pathologies or even in therapeutic implications. The detailed study of these diagnostic or therapeutic options, even if at this moment it represents a new challenge for researchers, may be a track for future possible research, which will bring new information to the scientific community and benefits in different pathologies [138].
In periodontology, biomaterials are used to replace bone or soft tissue defects. Periodontal therapy uses allogeneic or autologous grafts, which represent a golden therapy up to this point [138]. Even if this therapeutic option is considered a "golden standard", its limitations are numerous, the most important being represented by: the need for donors, possible immunological incompatibilities, their limited quantity and especially the possibility of presenting different infectious diseases. For a material to be considered ideal, it must have a series of properties: the most important is biocompatibility, followed by: elasticity, mechanical strength, ductility and durability, malleability and good thermal properties [139].
Nanomaterials, through their optical, mechanical, electronic and magnetic properties, have gained a special place in therapeutic approaches [140]. Moreover, the possibility of using different types of nanomaterials, such as: nanoparticles, nanotubes, nanocapsules, nanocomposites and nanofibers, to restore tissue functionality has been discussed [141].
Periodontal disease therapy has two major stages, the first being represented by non-surgical periodontal therapy (professional mechanical instrumentation, bacterial plaque control, local antimicrobial adjuvant treatment, elimination of local risk factors and management of systemic risk factors), and the second, being represented by of surgical periodontal therapy. Surgical periodontal therapy is a complex approach, which encompasses three broad types of therapeutic possibilities, these being the following in the beginning: bone grafts [142], guided tissue regeneration [143] and the use of plasma rich in platelets, known as PRP [144], all with the aim of reducing inflammation and restoring optimal functionality. Nanomaterials with dimensions below 100 nm have recently been introduced, presenting a series of advantages, such as: versatility, biocompatibility, controlled release of pharmaceutical substances and mechanical resistance. According to a recent literature review, the most researched materials for bone regeneration were represented by nanohydroxyapatite and collagen. Both materials present excellent osteoconductive, mechanical and biological properties [145]. Polylactic glycolic acid and polycaprolactone are intensively studied for use in tissue engineering and regenerative medicine due to their ability to degrade into compounds that can be easily metabolized and excreted. According to studies using these biomaterials for drug delivery, they can provide long-term effects and prevent inflammation [146]. It is also known that polylactoglycolic acid must be reinforced with ceramic nanoparticles to present the adequate resistance to various therapeutic procedures [147].
The structures on which the adsorption of pharmaceutical substances that act by inhibiting the inflammasome and thus reducing the level of inflammation could be discussed, should be based on a conductive material. In order to ensure a controlled release and possibly a slow release of the adsorbed preparations, polymers can be used.
Nanofibers have also been studied for their ability to transport pharmaceutical substances, according to a study carried out in 2022, they were prepared by electrospinning, forming a polylactic acid-hydroxyapatite-doxycycline (PLA-HAP-Doxy) system. According to this study, the nanofibers were prepared in two ways: immobilization on the surface and encapsulation in the fiber structure.
It was demonstrated that the polylactic acid-hydroxyapatite-doxycycline system can be used for biomedical applications [148].
Electrospun nanofibers are indicated for the application of biomedical substances, due to their property of being flexible and high production rates [149]. Other useful properties of these fibers are represented by the large surface area, the variable degree of porosity and the controllable morphology, which gives the medicinal substance an increased bioavailability [150]. The design method of the nanofiber-drug system depends on the nature and characteristics of the pharmaceutical substance, which is desired to be adsorbed on the surface of the nanofiber. It is known that the release of the active substance depends on the interaction between the polymer and the pharmaceutical substance, the diameter of the fiber and the initial content of the substance [148]. A series of polymers and combinations of polymers can be used to control the release of the adsorbed substance, such as: polylactic acid, polyethylene-vinyl acetate and polycaprolactone [151]. Nanofibers with hydroxyapatite and polylactic acid systems have been shown to have high biocompatibility [152] and good tissue response [153,154].
The biomaterial based on polylactic acid, nanohydroxyapatite and doxycycline, according to a study published in 2023, proved to have beneficial effect for the treatment of periodontal disease, decreasing clinical parameters of inflammation, but also influenced the salivary biomarkers, by decreasing the salivary concentration of MMP-8, IL-1 and TNF-α [155]. Due to the influence on biomarkers, the biomaterial was considered to have a local and systemic biomodulatory effect in periodontal disease. Nanofibers containing polylactic acid have an increased mechanical resistance to stress, a slow degradation rate and the possibility of degradation without releasing toxic residues [156].
Pharmaceutical and phytochemical substances mentioned in the specialized literature as possible inhibitors of inflammasome, could be investigated in future studies in order to adsorb them on different nanostructures, so that they can be used for topical applications in oral therapy. In order to be able to establish a connection between the pharmaceutical substance, the nanostructure on which it will be incorporated and the ligand that will be able to control the release of the preparation, the physicochemical properties of these substances must first be studied. Biomaterials can block the inflammasome activation, through several mechanisms: (i) inhibition of intracellular signaling pathways, (ii) blocking the components of the inflammasome, (iii) inhibition of cytokines mediated by the inflammasome, (iv) inhibition of ROS.
5.1. Blocking of the inflammasome, by inhibiting intracellular signaling pathways
For the inhibition of the inflammasome by inhibiting intracellular signaling pathways, two pharmaceutical substances have been mentioned: Allopurinol (Zyloprim/Aloprim®) and SS-31 (Elamipretide, Bendavia, MTP-131). Allopurinol, also known as 1,2-dihydro-4H-pyrazolo [3,4-d] pyrimidin-4-one, is an odourless, tasteless white microcrystalline powder. Allopurinol is a bicyclic organic compound in which a pyrazole ring is linked to a hydro-substituted pyrimidine ring. It is considered to be a structural analogue of hypoxanthine, a natural purine base. In terms of its application, it is often used in the treatment of gout and hyperuricemia as it inhibits the action of the xanthine oxidase enzyme [157,158]. The main disadvantage of the drug is that it is poorly soluble in water (0.1 mg/ml) [159]. Changdeo et al prepared solid dispersions of allopurinol using different methods and hydrophilic carriers such as polyvinylpyrrolidone, polyethylene glycol 6000 to increase the water solubility. The presence of polymers had a positive effect on increasing the water solubility [159]. Kandav et al prepared chitosan nanoparticles loaded with allopurinol to improve solubility and bioavailability [160]. Varrica et al have incorporated allopurinol into nanostructured lipid carriers (NLCs) in order to improve skin penetration, solubility problems and therapeutic index of the drug. The nanostructured lipid carriers are able to make allopurinol soluble and to incorporate and deliver it into the skin [161]. Kandav et al have produced bovine serum albumin nanoparticles loaded with allopurinol in order to deliver allopurinol to the kidney level [162]. According to literature research, it can be seen that various methods have been tried to incorporate Allopurinol into various nanostructures. SS-31 (Bendavia) is a small, easily synthesised cell-permeable peptide that is used in therapy because of its water solubility. It is a variable aromatic, cationic tetrapeptide with structural properties that targets the mitochondria, penetrating its inner membrane, associating with cardiolipin and restoring membrane stability [163]. To efficiently deliver SS-31, Di Liu et al designed nanopolyplexes complexed using anionic hyaluronic acid and cationic chitosan. The electrostatic equilibrium of the nanoparticles are disrupted in the low pH environment of lysosomes, thus allowing SS-31 to be released and exert its therapeutic effect in the mitochondria. Studies have shown that the nanoparticles exhibited superior therapeutic effects compared to free SS-31 [164]. Due to the increased solubility in water, this pharmaceutical preparation is useful in therapy, and according to the study conducted by Di Liu et al, it can be affirmed its ability to be released from the structure on which it is adsorbed and the ability to exert its therapeutic effect at the site of action.
5.2. Inhibition of the inflammasome, by blocking its components
For the inhibition of the inflammasome by blocking the components of the inflammasome, three pharmaceutical substances were mentioned in the specialized literature: VX-765, Emricasanul, VX-740 (Pralnacasan). The action of colchicine can also be mentioned. Belnacasan, also known as VX-765, acts as a potent and selective inhibitor of the caspase-1 enzyme, which is being investigated for the treatment of epilepsy. It is an orally active prodrug that is converted to VRT-043198 active drug in the body. In physical appearance it is a light yellow powder, insoluble in water. Emricasan, also known as IDN-6556, is a white, solid powder that is insoluble in water but not in DMSO. Its molecular structure is C26H27F4N3O7 and its molecular weight is 569.50 g/mol [163]. Emricasan belongs to the caspase family and is an irreversible pan-caspase inhibitor. It is used in patients with non-alcoholic steatohepatitis because it reduces serum aminotransferases and caspase activity. It is the first caspase inhibitor to be tested in humans [166]. Pralnacasan also known as VX-740, is a peptidomimetic caspase-1 inhibitor targeting inflammatory components. It is used in clinical trials as a potent non-peptide inhibitor of the interleukin 1β converting enzyme [167]. Pralnacasan has a 2-naphthyl hydrophobic moiety at the P4 position and a pyridazinodiazepine- based bicyclic core that mimics the Val-Ala region. A lactone moiety is located at the C- terminal covering an aspartate residue at the P1 position. Since the discovery of the drug, researchers have sought to produce a number of inhibitors with a mono- or bicyclic backbone mimicking P3-P2 [168]. Colchicine is a light yellowish powder with molecular structure C22H25NO6 [169]. It is an alkaloid that contains an N atom in the amide form. Due to its toxic properties, its use as a medicine has been marginalized and it is often identified as a mitotic damager [170]. Colchicine is a compound containing three rings (A, B, C), in which ring A is a trimethoxy phenyl ring, ring B is a saturated seven-membered ring and ring C is a tropnol ring. Colchicine is not a basic compound and therefore, unlike other basic alkaloids, it does not form salts. It forms precipitates with various alkaloid reagents. Colchicine has a chiral centre at the C-7 position. Due to the presence of a chirality axis, Colchicine is asymmetric, with the A ring twisted by 53° relative to the B ring. Colchicine has four enantiomers, however, it should be emphasized that this requires the (aS) configuration for binding to tubulin [171]. Colchicine is an ancient drug, mentioned in the writings of several Greek physicians such as Nicader of Colphan, Discorides and Alexander of Trolles. It has also been used to relieve gout-induced pain and to treat an autoinflammatory genetic disorder, none other than Familiar Mediterranean fever. According to the specialized literature, these pharmaceutical preparations have a possible action on the inhibition of the inflammasome, and through the detailed study of their physicochemical properties, nanobiomaterials could be discovered on which they can be incorporated, to be used in the treatment of oral pathologies.
5.3. Blocking of the inflammasome, by inhibiting the cytokines
For the inhibition of the inflammasome by inhibiting the cytokines mediated by the inflammasome, three pharmaceutical substances have been mentioned in the specialized literature: Canakinumab, Anakinra and Rilonacept. Canakinumab, also known as INN, is a human anti-IL-1β monoclonal antibody with high affinity. Canakinumab consists of two light chains of 214 residues and two heavy chains of 447 or 448 residues. It has a molecular mass of 145 kDa. The molecular formula of canakinumab is based on the amino acid composition without post-translational glycosylation, but includes N-terminal pyroglutamate formation and lysine residues on the C-terminals of the heavy chains [172].
Anakinra is a 153 amino acid recombinant human interleukin-1 (IL-1) receptor antagonist. Anakinra differs from native human IL-1Ra in that it has a methionine group at the amino- terminal [173]. Rilonacept is a dimer fusion protein, an engineered compound consisting of the extracellular domains of 1L-1-RacP and 1I-1-R1. Also known as Il-1 trap, it is linked to the Fc portion of human immunoglobulin G1 (IgG1) [174].
5.4. Inhibition of the inflammasome, by using phytochemicals
Phytochemicals can also act on the inflammasome, in order to inhibit its action, through the increased content of flavonoids. Flavonoids are a family of compounds that are widely distributed in the plant kingdom and have many medicinal properties. They belong to the group of polyphenols and are found in many plants (chamomile), fruits (oranges, grapefruit) and vegetables (onions, broccoli), as well as in beverages. They are structurally characterized by a 2-phenylchroman skeleton (C6-C3-C6) with a heterocyclic pyrene ring fused to the benzene ring labelled A and linked to the phenyl ring labelled B. The flavonoid structures known to date have various substituents such as multiple hydroxyl (-OH), methoxyl (-OCH3) and glycoside groups, as well as an oxo group located at position 4 of the C ring [175,176]. Within the flavonoid family of compounds there are different subgroups. These subgroups depend on the degree of oxidation, saturation, substitution pattern, and bonding position of the C ring to the C2/C3 and C4 carbon atoms of the B and C rings. For example, in the case of isoflavones, the B ring is attached to the triple position of the C ring, whereas in the case of neoflavonoids, the B ring is attached to the quadruple position of the C ring. The metabolism, biochemical and pharmacological activity and bioavailability of flavonoids depend on the structural characteristics and configurations of the flavonoids [177]. These compounds have unique chemical and physical properties. These properties affect the solubility of flavonoids in different solvents. Their solubility in water is affected by the presence of a double bond in the C ring and the number of hydroxyl (-OH) substituents in the B ring. Solubility in water increases with the number of hydroxyl groups, whereas solubility in 1-octanol is adversely affected by the presence of hydroxyl groups. The stability of flavonoids changes when they are exposed to external stimuli such as heat and light. The scientific literature indicates that flavonoids undergo oxidative transformation when exposed to the aforementioned effects [177].
Over the past decades, the development of targeted drug delivery tools has undergone intensive development to meet stringent requirements. These requirements include the need for delivery system to increase the effectiveness of the treatment, while at the same time reducing undesirable effects. Importantly, the drug delivery system must be biocompatible and not form toxic compounds. The scientific literature shows that over the past decades, researchers have succeeded in developing drug carriers with a wide range of chemical structures and origins, ranging in molecular sizes from 1 to hundreds of nanometers. Second-generation carriers smaller than 1 μm, belonging to the group of so-called colloidal carriers, are used to deliver flavonoids. The main nanocarriers suitable for the delivery of flavonoids include: phytosomes, liposomes, solid lipid nanoparticles (SLN), nanostructured lipid carriers, nano- and microemulsions, polymer nanoparticles, chitosan based nanoparticles, gold-based nanoparticles, silver-based nanoparticles, silica nanoparticles. Zverev et al have shown that phytosomes containing flavonoids are characterized by a long-lasting tumor-targeting effect. The main property of this drug delivery system is that it does not exert toxic effects on surrounding healthy cells and tissues. The aforementioned author et al report that liposomes loaded with a flavone called luteolin and coated with tocoferyl polyethylene glycol succinate (TPGS) had no adverse effects on healthy tissues, but showed increased cytotoxicity in human lung cancer cells [178].
3. Discussion
Inflammasomes are part of the innate immune system, which normally acts beneficially on the body, but their uncontrolled activity causes harmful effects, being able to induce autoinflammatory or autoimmune diseases [179]. According to current data, the therapeutic action on inflammasomes could be considered a new treatment option for periodontal disease, as it decreases the production of cytokines [180]. Some pharmaceutical preparations and phytochemical preparations, which can act on the inhibition of the inflammasome. In order to facilitate the therapeutic effects, the adsorption or inclusion of these preparations on various structures of nanobiomaterials could be discussed. For the use on a large scale and to facilitation of various treatments, these nanobiomaterials should be involved in several studies in order to be able to exclude their limitations: inducing immunological reactions, spreading in unwanted sites, the occurrence of cytotoxicity and the control biodegradation. According to studies, the biodistribution and pharmacokinetics of these preparations depend on their shape and size [181]. Even if the incorporation of pharmaceutical preparations on various biocompatible structures could be of great help in therapy, their limits, their biocompatibility and the way they will interact with the human body must be studied. According to a literature review, it can be stated that the major problem with these materials is related to toxicity. They can enter the circulatory system and induce hematological toxicity, they can accumulate in organs (lungs, liver, spleen, kidneys) or they can produce an exaggerated quantity of ROS [182]. It is known that the size of nanomaterials is the physicochemical property that majorly influences the cellular response [183]. A nanomaterial with a reduced size presents an inversely proportional surface-volume ratio, which determines their greater reactivity [184], but also a lower accumulation in the liver and spleen [185]. However, having a small size, nanoparticle can cross the blood-brain barrier, thus presenting major health risks. The shape of the material influences the mode of absorption, biodistribution and their use [183]. It is known from the literature that a spherical shape of biomaterials has a superior capacity of pharmaceutical substance delivery than other shapes of biomaterials [186]. The asymmetric shapes of nanomaterials have an increased capacity to penetrate tissues and release pharmaceutical preparations [187]. It is also known that intelligent particles are able to change their shape depending on the need of the site [188]. Another set of properties that influence their therapeutic potential is represented by the loading capacity, binding capacity and hydrophobicity [183]. Thus, surfaces with positive charges have an increased absorption, and those with negative charges have a greater capacity to circulate for a longer time [189].
4. Conclusions
Due to the possibilities of adsorption on the surface or inclusion in nanofibres of various pharmaceutical preparations, this newly exploited branch of the universe of materials can represent a promising perspective for clinical practice, helping the practitioners to improve the quality of life of patients. Thus, these new nanobiomaterials and their combinations deserve to be studied and researched with interest, due to their capacity of action on the human body.
Author Contributions
Conceptualization, N.V., C.N.F., A.I., R.P. and B.A.B.; methodology, N.V., C.N.F., A.I., R.B., B.A.B., A.M. and D.F.; validation, N.V., C.N.F., A.I., R.B., B.A.B., A.M. and D.F.; formal analysis, N.V., A.I. and B.A.B.; investigation, N.V., A.I., R.P. and B.A.B.; resources, N.V., A.I., R.P. and B.A.B.; data curation, N.V., A.I. and B.A.B.; writing—original draft preparation, N.V., A.I. and R.P.; writing—review and editing, N.V., C.N.F., A.I., R.P. and B.A.B.; visualization, N.V., C.N.F., A.I., R.B., B.A.B., A.M. and D.F.; supervision, N.V., C.N.F., A.I., R.B., B.A.B., A.M. and D.F.; project administration, N.V and A.I. All authors have read and agreed to the published version of the manuscript.
Funding
The present study was supported by a grant from the Ministry of Research, Innovation and Digitization, CNCS—UEFISCDI, project number PN-III-P4-PCE-2021-1140.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hajishengallis, G.; Lamont, R.J. Polymicrobial communities in periodontal disease: Their quasi-organismal nature and dialogue with the host. Periodontology 2000 2021, 86, 210–230. [CrossRef]
- Williams, D.W.; Greenwell-Wild, T.; Brenchley, L.; Dutzan, N.; Overmiller, A.; Sawaya, A.P.; Webb, S.; Martin, D.; Hajishengallis, G.; Divaris, K.; et al. Human oral mucosa cell atlas reveals a stromal-neutrophil axis regulating tissue immunity. Cell 2021, 184, 4090–4104.e15. [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [CrossRef]
- Wang, H.; Ideguchi, H.; Kajikawa, T.; Mastellos, D.C.; Lambris, J.D.; Hajishengallis, G. Complement Is Required for Microbe-Driven Induction of Th17 and Periodontitis. J. Immunol. 2022, 209, 1370–1378. [CrossRef]
- Nauseef, W.M.; Borregaard, N. Neutrophils at work. Nat. Immunol. 2014, 15(7), 602-611. [CrossRef]
- Nussbaum, G.; Shapira, L. How has neutrophil research improved our understanding of periodontal pathogenesis? J. Clin. Periodontol. 2011, 38, 49-59. [CrossRef]
- Miralda, I.; Uriarte, S.M. Periodontal Pathogens’ strategies disarm neutrophils to promote dysregulated inflammation. Mol. Oral Microbiol. 2021, 36, 103–120. [CrossRef]
- Cowland, J.B.; Borregaard, N. Granulopoiesis and granules of human neutrophils. Immunol. Rev. 2016, 273, 11–28. [CrossRef]
- Cassatella, M.A.; Östberg, N.K.; Tamassia, N.; Soehnlein, O. Biological Roles of Neutrophil-Derived Granule Proteins and Cytokines. Trends Immunol. 2019, 40, 648–664. [CrossRef]
- Miralda, I.; Uriarte, S.M.; McLeish, K.R. Multiple Phenotypic Changes Define Neutrophil Priming. Front. Cell. Infect. Microbiol. 2017, 7, 217. [CrossRef]
- Irwandi, R.A.; Chiesa, S.T.; Hajishengallis, G.; Papayannopoulos, V.; Deanfield, J.E.; D’aiuto, F. The Roles of Neutrophils Linking Periodontitis and Atherosclerotic Cardiovascular Diseases. Front. Immunol. 2022, 13, 915081. [CrossRef]
- Elbim, C.; Rajagopalan-Levasseur, P.; Chollet-Martin, S.; Gaillard, J.-L.; Fay, M.; Hakim, J.; Fischer, A.; Casanova, J.-L.; Gougerot-Pocidalo, M.-A. Defective priming of the phagocyte oxidative burst in a child with recurrent intracellular infections. Microbes Infect. 1999, 1, 581–587. [CrossRef]
- Whitmore, L.C.; Hook, J.S.; Philiph, A.R.; Hilkin, B.M.; Bing, X.; Ahn, C.; Wong, H.R.; Ferguson, P.J.; Moreland, J.G. A Common Genetic Variant in TLR1 Enhances Human Neutrophil Priming and Impacts Length of Intensive Care Stay in Pediatric Sepsis. J. Immunol. 2016, 196, 1376–1386. [CrossRef]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Cooper, P.R.; Chapple, I.L.C. Hyperactivity and reactivity of peripheral blood neutrophils in chronic periodontitis. Clin. Exp. Immunol. 2007, 147, 255–264. [CrossRef]
- Eggleton, P.; Wang, L.; Penhallow, J.; Crawford, N.; A Brown, K. Differences in oxidative response of subpopulations of neutrophils from healthy subjects and patients with rheumatoid arthritis.. Ann. Rheum. Dis. 1995, 54, 916–923. [CrossRef]
- Divangahi, M.; Aaby, P.; Khader, S.A. Trained immunity, tolerance, priming and differentiation: distinct immunological processes. Nat. Immunol. 2021, 22(1), 2-6. [CrossRef]
- Hajishengallis, G.; Diaz, P.I. Porphyromonas gingivalis: Immune Subversion Activities and Role in Periodontal Dysbiosis. Curr. Oral Heal. Rep. 2020, 7, 12–21. [CrossRef]
- Prince, L.R.; Whyte, M.K.; Sabroe, I.; Parker, L.C. The role of TLRs in neutrophil activation. Curr. Opin. Pharmacol. 2011, 11, 397–403. [CrossRef]
- Hajishengallis, G.; Kajikawa, T.; Hajishengallis, E.; Maekawa, T.; Reis, E.S.; Mastellos, D.C.; Yancopoulou, D.; Hasturk, H.; Lambris, J.D. Complement-Dependent Mechanisms and Interventions in Periodontal Disease. Front. Immunol. 2019, 10, 406. [CrossRef]
- Hajishengallis, G.; Reis, E.S.; Mastellos, D.C.; Ricklin, D.; Lambris, J.D. Novel mechanisms and functions of complement. Nat. Immunol. 2017, 18, 1288–1298. [CrossRef]
- Harokopakis, E.; Hajishengallis, G. Integrin activation by bacterial fimbriae through a pathway involving CD14, Toll-like receptor 2, and phosphatidylinositol-3-kinase. Eur. J. Immunol. 2005, 35, 1201–1210. [CrossRef]
- Potempa, J.; Sroka, A.; Imamura, T.; Travis, J. Gingipains, the major cysteine proteinases and virulence factors of Porphyromonas gingivalis: structure, function and assembly of multidomain protein complexes. Curr. Protein. Pept. Sci. 2003, 4(6), 397-407. [CrossRef]
- Liang, S.; Krauss, J.L.; Domon, H.; McIntosh, M.L.; Hosur, K.B.; Qu, H.; Li, F.; Tzekou, A.; Lambris, J.D.; Hajishengallis, G. The C5a Receptor Impairs IL-12–Dependent Clearance of Porphyromonas gingivalis and Is Required for Induction of Periodontal Bone Loss. J. Immunol. 2011, 186, 869–877. [CrossRef]
- Juarez-Rodriguez, M.D.; Torres-Escobar, A.; Demuth, D.R. ygiW and qseBC are co-expressed in Aggregatibacter actinomycetemcomitans and regulate biofilm growth. Microbiology 2013, 159, 989-1001. [CrossRef]
- Weigel, W.A.; Demuth, D.R.; Torres-Escobar, A.; Rodríguez, J. Aggregatibacter actinomycetemcomitans QseBC is activated by catecholamines and iron and regulates genes encoding proteins associated with anaerobic respiration and metabolism. Mol. Oral Microbiol. 2015, 30, 384–398. [CrossRef]
- Ozuna, H.; Uriarte, S.M.; Demuth, D.R. The Hunger Games: Aggregatibacter actinomycetemcomitans Exploits Human Neutrophils As an Epinephrine Source for Survival. Front. Immunol. 2021, 12. [CrossRef]
- Stark, M.A.; Huo, Y.; Burcin, T.L.; Morris, M.A.; Olson, T.S.; Ley, K. Phagocytosis of Apoptotic Neutrophils Regulates Granulopoiesis via IL-23 and IL-17. Immunity 2005, 22, 285–294. [CrossRef]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15(2), 104. [CrossRef]
- Drechsler, M.; Megens, R.; Van Zandvoort, M.; Weber, C.; Soehnlein, O. Hyperlipidemia-Triggered Neutrophilia Promotes Early Atherosclerosis. Circulation 2010, 122, 1837–1845. [CrossRef]
- Nagareddy, P.R.; Murphy, A.J.; Stirzaker, R.A.; Hu, Y.; Yu, S.; Miller, R.G.; Ramkhelawon, B.; Distel, E.; Westerterp, M.; Huang, L.-S.; et al. Hyperglycemia Promotes Myelopoiesis and Impairs the Resolution of Atherosclerosis. Cell Metab. 2013, 17, 695–708. [CrossRef]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [CrossRef]
- Kajikawa, T.; Briones, R.A.; Resuello, R.R.; Tuplano, J.V.; Reis, E.S.; Hajishengallis, E.; Garcia, C.A.; Yancopoulou, D.; Lambris, J.D.; Hajishengallis, G. Safety and Efficacy of the Complement Inhibitor AMY-101 in a Natural Model of Periodontitis in Non-human Primates. Mol. Ther. - Methods Clin. Dev. 2017, 6, 207–215. [CrossRef]
- Dahlen, G.; Basic, A.; Bylund, J. Importance of Virulence Factors for the Persistence of Oral Bacteria in the Inflamed Gingival Crevice and in the Pathogenesis of Periodontal Disease. J. Clin. Med. 2019, 8, 1339. [CrossRef]
- Bateman, G.; Hill, B.; Knight, R.; Boucher, D. Great balls of fire: activation and signalling of inflammatory caspases. Biochem. Soc. Trans. 2021, 49, 1311–1324. [CrossRef]
- Bostanci, N.; Emingil, G.; Saygan, B.; Turkoglu, O.; Atilla, G.; A Curtis, M.; Belibasakis, G.N. Expression and regulation of the NALP3 inflammasome complex in periodontal diseases. Clin. Exp. Immunol. 2009, 157, 415–422. [CrossRef]
- Marchesan, J.T.; Girnary, M.S.; Moss, K.; Monaghan, E.T.; Egnatz, G.J.; Jiao, Y.; Zhang, S.; Beck, J.; Swanson, K.V. Role of inflammasomes in the pathogenesis of periodontal disease and therapeutics. Periodontology 2000 2020, 82, 93–114. [CrossRef]
- Harton, J.A.; Linhoff, M.W.; Zhang, J.; Ting, J.P.-Y. Cutting Edge: CATERPILLER: A Large Family of Mammalian Genes Containing CARD, Pyrin, Nucleotide-Binding, and Leucine-Rich Repeat Domains. J. Immunol. 2002, 169, 4088–4093. [CrossRef]
- Ting, J.P.; Lovering, R.C.; Alnemri, E.S.; Bertin, J.; Boss, J.M.; Davis, B.K.; Flavell, R.A.; Girardin, S.E.; Godzik, A.; Harton, J.A.; et al. The NLR Gene Family: A Standard Nomenclature. Immunity 2008, 28, 285–287. [CrossRef]
- Harijith, A.; Ebenezer, D.L.; Natarajan, V. Reactive oxygen species at the crossroads of inflammasome and inflammation. Front. Physiol. 2014, 5, 352. [CrossRef]
- Boyden, E.D.; Dietrich, W.F. Nalp1b controls mouse macrophage susceptibility to anthrax lethal toxin. Nat. Genet. 2006, 38, 240–244. [CrossRef]
- Yilmaz, .; Sater, A.A.; Yao, L.; Koutouzis, T.; Pettengill, M.; Ojcius, D.M. ATP-dependent activation of an inflammasome in primary gingival epithelial cells infected byPorphyromonas gingivalis. Cell. Microbiol. 2010, 12, 188–198. [CrossRef]
- Belibasakis, G.N.; Johansson, A. Aggregatibacter actinomycetemcomitans targets NLRP3 and NLRP6 inflammasome expression in human mononuclear leukocytes. Cytokine 2012, 59, 124–130. [CrossRef]
- Lich, J.D.; Ting, J.P. Monarch-1/PYPAF7 and other CATERPILLER (CLR, NOD, NLR) proteins with negative regulatory functions. Microbes. Infect. 2007, 9, 672-676.
- Duncan, J.A.; Gao, X.; Huang, M.T.-H.; O'Connor, B.P.; Thomas, C.E.; Willingham, S.B.; Bergstralh, D.T.; Jarvis, G.A.; Sparling, P.F.; Ting, J.P.-Y. Neisseria gonorrhoeae Activates the Proteinase Cathepsin B to Mediate the Signaling Activities of the NLRP3 and ASC-Containing Inflammasome. J. Immunol. 2009, 182, 6460–6469. [CrossRef]
- Rathinam, V.A.K.; Vanaja, S.K.; A Fitzgerald, K. Regulation of inflammasome signaling. Nat. Immunol. 2012, 13, 333–342. [CrossRef]
- Wang, P.; Zhu, S.; Yang, L.; Cui, S.; Pan, W.; Jackson, R.; Zheng, Y.; Rongvaux, A.; Sun, Q.; Yang, G.; et al. Nlrp6 regulates intestinal antiviral innate immunity. Science 2015, 350, 826–830. [CrossRef]
- Radian, A.D.; Khare, S.; Chu, L.H.; Dorfleutner, A.; Stehlik, C. ATP binding by NLRP7 is required for inflammasome activation in response to bacterial lipopeptides. Mol. Immunol. 2015, 67, 294–302. [CrossRef]
- Williams, K.L.; Lich, J.D.; Duncan, J.A.; Reed, W.; Rallabhandi, P.; Moore, C.; Kurtz, S.; Coffield, V.M.; Accavitti-Loper, M.A.; Su, L.; et al. The CATERPILLER Protein Monarch-1 Is an Antagonist of Toll-like Receptor-, Tumor Necrosis Factor α-, and Mycobacterium tuberculosis-induced Pro-inflammatory Signals. J. Biol. Chem. 2005, 280, 39914–39924. [CrossRef]
- Gong, Y.-N.; Shao, F. Sensing bacterial infections by NAIP receptors in NLRC4 inflammasome activation. Protein Cell 2012, 3, 98–105. [CrossRef]
- Davis, B.K.; Roberts, R.A.; Huang, M.T.; Willingham, S.B.; Conti, B.J.; Brickey, W.J.; Barker, B.R.; Kwan, M.; Taxman, D.J.; Accavitti-Loper, M.-A.; et al. Cutting Edge: NLRC5-Dependent Activation of the Inflammasome. J. Immunol. 2011, 186, 1333–1337. [CrossRef]
- Wu, J.; Fernandes-Alnemri, T.; Alnemri, E. S. Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by Listeria monocytogenes. J. Clin. Immunol. 2010, 30, 693–702.
- Fernandes-Alnemri, T.; Yu, J.-W.; Datta, P.; Wu, J.; Alnemri, E.S. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 2009, 458, 509–513. [CrossRef]
- Correa, R.G.; Milutinovic, S.; Reed, J.C. Roles of NOD1 (NLRC1) and NOD2 (NLRC2) in innate immunity and inflammatory diseases. Biosci. Rep. 2012, 32, 597–608. [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen Recognition by the Innate Immune System. Int. Rev. Immunol. 2011, 30, 16–34. [CrossRef]
- Sordi, M.B.; Magini, R.d.S.; Panahipour, L.; Gruber, R. Pyroptosis-Mediated Periodontal Disease. Int. J. Mol. Sci. 2022, 23, 372. [CrossRef]
- Cotranu, I.; Mareş, L. Explanatory dictionary of the Romanian language, 2nd ed. Encyclopedic Universe Gold, Romania; 2009.
- Jones, N.S.; Kshirsagar, S.; Mohanan, V.; Ramakrishnan, V.; Di Nucci, F.; Ma, L.; Mao, J.; Ding, H.; Klabunde, S.; Vucic, D.; Pan, L.; Lekkerkerker, A.N.; Chen, Y.; Rothenberg, M.E. A phase I, randomized, ascending-dose study to assess safety, pharmacokinetics, and activity of GDC-8264, a RIP1 inhibitor, in healthy volunteers. Clin. Transl. Sci. 2023, 16(10), 1997-2009. [CrossRef]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: Death by Lipid Peroxidation. Trends Cell Biol. 2016, 26, 165–176. [CrossRef]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochem. (Moscow) 2020, 85, 1178–1190. [CrossRef]
- A Miao, E.; A Leaf, I.; Treuting, P.M.; Mao, D.P.; Dors, M.; Sarkar, A.; E Warren, S.; Wewers, M.D.; Aderem, A. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 2010, 11, 1136–1142. [CrossRef]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [CrossRef]
- Cookson, B.T.; A Brennan, M. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [CrossRef]
- Yu, C.; Zhang, C.; Kuang, Z.; Zheng, Q. The Role of NLRP3 Inflammasome Activities in Bone Diseases and Vascular Calcification. Inflammation 2021, 44, 434–449. [CrossRef]
- Chen, X.; He, W.-T.; Hu, L.; Li, J.; Fang, Y.; Wang, X.; Xu, X.; Wang, Z.; Huang, K.; Han, J. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016, 26, 1007–1020. [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes and Their Roles in Health and Disease. Annu. Rev. Cell Dev. Biol. 2012, 28, 137–161. [CrossRef]
- Liu, W.; Liu, J.; Wang, W.; Wang, Y.; Ouyang, X. NLRP6 Induces Pyroptosis by Activation of Caspase-1 in Gingival Fibroblasts. J. Dent. Res. 2018, 97, 1391–1398. [CrossRef]
- Muñoz-Planillo, R.; Franchi, L.; Miller, L.S.; Núñez, G. A Critical Role for Hemolysins and Bacterial Lipoproteins inStaphylococcus aureus-Induced Activation of the Nlrp3 Inflammasome. J. Immunol. 2009, 183, 3942–3948. [CrossRef]
- Muñoz-Planillo, R.; Kuffa, P.; Martínez-Colón, G.; Smith, B.L.; Rajendiran, T.M.; Núñez, G. K+ Efflux Is the Common Trigger of NLRP3 Inflammasome Activation by Bacterial Toxins and Particulate Matter. Immunity 2013, 38, 1142–1153. [CrossRef]
- Pressman, B.C. Biological Applications of Ionophores. Annu. Rev. Biochem. 1976, 45, 501–530. [CrossRef]
- Domon, H.; Takahashi, N.; Honda, T.; Nakajima, T.; Tabeta, K.; Abiko, Y.; Yamazaki, K. Up-regulation of the endoplasmic reticulum stress-response in periodontal disease. Clin. Chim. Acta 2009, 401, 134–140. [CrossRef]
- Hörauf, J.-A.; Kany, S.; Janicova, A.; Xu, B.; Vrdoljak, T.; Sturm, R.; Dunay, I.R.; Martin, L.; Relja, B. Short Exposure to Ethanol Diminishes Caspase-1 and ASC Activation in Human HepG2 Cells In Vitro. Int. J. Mol. Sci. 2020, 21, 3196. [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [CrossRef]
- Feng, Y.; Huang, X. Methodology for Comprehensive Detection of Pyroptosis. Methods Mol. Biol. 2021, 2255, 149–157. [CrossRef]
- Graves, D.; Cochran, D. The Contribution of Interleukin-1 and Tumor Necrosis Factor to Periodontal Tissue Destruction. J. Periodontol. 2003, 74, 391–401. [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes and Their Roles in Health and Disease. Annu. Rev. Cell Dev. Biol. 2012, 28, 137–161. [CrossRef]
- Davis, B.K.; Wen, H.; Ting, J.P.-Y. The Inflammasome NLRs in Immunity, Inflammation, and Associated Diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [CrossRef]
- Ken-ichiro S. Historical aspects of studies on roles of the inflammasome in the pathogenesis of periodontal diseases. Mol. Oral Microb. 2013, 33(3), 203–211. [CrossRef]
- Schroder, K.; J. Tschopp. The inflammasomes. Cell 2010, 140, 821–832. [CrossRef]
- Martinon, F.; K. Burns; J. Tschopp. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell. 2002, 10, 417–426. [CrossRef]
- Zhong, Z.; Liang S.; Sanchez-Lopez E.; He F.; Shalapour S.; Lin X. J.; Wong J.; Ding S.; Seki E.; Schnabl B. New mitochondrial DNA synthesis enables NLRP3 inflammasome activation. Nature 2018, 560, 198–203. [CrossRef]
- Bostanci, N.; Emingil, G.; Saygan, B.; Turkoglu, O.; Atilla, G.; A Curtis, M.; Belibasakis, G.N. Expression and regulation of the NALP3 inflammasome complex in periodontal diseases. Clin. Exp. Immunol. 2009, 157, 415–422. [CrossRef]
- Xue, F.; Shu, R.; Xie, Y. The expression of NLRP3, NLRP1 and AIM2 in the gingival tissue of periodontitis patients: RT-PCR study and immunohistochemistry. Arch. Oral Biol. 2015, 60, 948–958. [CrossRef]
- Li, Y.; Li, B.; Liu, Y.; Wang, H.; He, M.; Liu, Y.; Sun, Y.; Meng, W. Porphyromonas gingivalis lipopolysaccharide affects oral epithelial connections via pyroptosis. J. Dent. Sci. 2021, 16, 1255–1263. [CrossRef]
- Shibata, K. Historical aspects of studies on roles of the inflammasome in the pathogenesis of periodontal diseases. Mol. Oral Microbiol. 2018, 33, 203–211. [CrossRef]
- Hanazawa, S.; Nakada, K.; Ohmori, Y.; Miyoshi, T.; Amano, S.; Kitano, S. Functional role of interleukin 1 in periodontal disease: induction of interleukin 1 production by Bacteroides gingivalis lipopolysaccharide in peritoneal macrophages from C3H/HeN and C3H/HeJ mice. Infect. Immun. 1985, 50, 262–270. [CrossRef]
- Jandinski, J.J.; Stashenko, P.; Feder, L.S. Localization of interleukin-1 beta in human periodontal tissue. J. Periodontol. 1991, 62(1), 36-43. [CrossRef]
- Kinane, D.; Winstanley, F.; Adonogianaki, E.; Moughal, N. Bioassay of interleukin 1 (IL-1) in human gingival crevicular fluid during experimental gingivitis. Arch. Oral Biol. 1992, 37, 153–156. [CrossRef]
- Assuma, R.; Oates, T.; Cochran, D.; Amar, S.; Graves, D.T. IL-1 and TNF Antagonists Inhibit the Inflammatory Response and Bone Loss in Experimental Periodontitis. J. Immunol. 1998, 160, 403–409. [CrossRef]
- Bostanci, N.; Meier, A.; Guggenheim, B.; Belibasakis, G.N. Regulation of NLRP3 and AIM2 inflammasome gene expression levels in gingival fibroblasts by oral biofilms. Cell. Immunol. 2011, 270, 88–93. [CrossRef]
- Guo, W.; Ye, P.; Yu, H.; Liu, Z.; Yang, P.; Hunter, N. CD24 activates the NLRP3 inflammasome through c-Src kinase activity in a model of the lining epithelium of inflamed periodontal tissues. Immun. Inflamm. Dis. 2014, 2(4), 239-253. [CrossRef]
- Xue, F.; Shu, R.; Xie, Y. The expression of NLRP3, NLRP1 and AIM2 in the gingival tissue of periodontitis patients: RT-PCR study and immunohistochemistry. Arch. Oral Biol. 2015, 60, 948–958. [CrossRef]
- Yamaguchi, Y.; Kurita-Ochiai, T.; Kobayashi, R.; Suzuki, T.; Ando, T. Activation of the NLRP3 inflammasome in Porphyromonas gingivalis-accelerated atherosclerosis. Pathog. Dis. 2015, 73. [CrossRef]
- Wang, L.; Sharif, H.; Vora, S.M.; Zheng, Y.; Wu, H. Structures and functions of the inflammasome engine. J. Allergy Clin. Immunol. 2021, 147, 2021–2029. [CrossRef]
- Bullon, P.; Navarro, J.M. Inflammasome as a Key Pathogenic Mechanism in Endometriosis. Curr. Drug Targets 2017, 18, 997–1002. [CrossRef]
- Huang, X.; Yang, X.; Ni, J.; Xie, B.; Liu, Y.; Xuan, D.; Zhang, J. Hyperglucose Contributes to Periodontitis: Involvement of the NLRP3 Pathway by Engaging the Innate Immunity of Oral Gingival Epithelium. J. Periodontol. 2015, 86, 327–335. [CrossRef]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Walle, L.V.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [CrossRef]
- Dang, E.V.; McDonald, J.G.; Russell, D.W.; Cyster, J.G. Oxysterol Restraint of Cholesterol Synthesis Prevents AIM2 Inflammasome Activation. Cell 2017, 171, 1057–1071.e11. [CrossRef]
- Warnatsch, A.; Ioannou M.; Wang Q.; Papayannopoulos V. Inflammation. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 2015, 349, 316–320. [CrossRef]
- Hottz, E.D.; Lopes, J.F.; Freitas, C.; Valls-De-Souza, R.; Oliveira, M.F.; Bozza, M.T.; Da Poian, A.T.; Weyrich, A.S.; Zimmerman, G.A.; Bozza, F.A.; et al. Platelets mediate increased endothelium permeability in dengue through NLRP3-inflammasome activation. Blood 2013, 122, 3405–3414. [CrossRef]
- Segovia, J.; Sabbah, A.; Mgbemena, V.; Tsai, S.-Y.; Chang, T.-H.; Berton, M.T.; Morris, I.R.; Allen, I.C.; Ting, J.P.-Y.; Bose, S. TLR2/MyD88/NF-κB Pathway, Reactive Oxygen Species, Potassium Efflux Activates NLRP3/ASC Inflammasome during Respiratory Syncytial Virus Infection. PLOS ONE 2012, 7, e29695. [CrossRef]
- Dorhoi, A.; Nouailles, G.; Jörg, S.; Hagens, K.; Heinemann, E.; Pradl, L.; Oberbeck-Müller, D.; Duque-Correa, M.A.; Reece, S.T.; Ruland, J.; et al. Activation of the NLRP3 inflammasome by Mycobacterium tuberculosis is uncoupled from susceptibility to active tuberculosis. Eur. J. Immunol. 2012, 42, 374–384. [CrossRef]
- Franchi, L.; Núñez, G. The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1beta secretion but dispensable for adjuvant activity. Eur. J. Immunol. 2008, 38, 2085–2089. [CrossRef]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.-H.; Brickey, W.J.; Ting, J.P.-Y. Fatty acid–induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [CrossRef]
- Vilaysane, A.; Chun, J.; Seamone, M.E.; Wang, W.; Chin, R.; Hirota, S.; Li, Y.; Clark, S.A.; Tschopp, J.; Trpkov, K.; et al. The NLRP3 Inflammasome Promotes Renal Inflammation and Contributes to CKD. J. Am. Soc. Nephrol. 2010, 21, 1732–1744. [CrossRef]
- Zhang, W.; Cai, Y.; Xu, W.; Yin, Z.; Gao, X.; Xiong, S. AIM2 Facilitates the Apoptotic DNA-induced Systemic Lupus Erythematosus via Arbitrating Macrophage Functional Maturation. J. Clin. Immunol. 2013, 33, 925–937. [CrossRef]
- Guo, H.; Callaway, J.B.; Ting, J.P.-Y. Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [CrossRef]
- Gilbert, D.L. O2 and living processes: an inter-disciplinary approach. Springer, New York, 1981.
- Halliwell, B.; Gutteridge, J. Free radicals in biology and medicine, 4th ed.; Oxford Univ. Press, Oxford, 2007.
- Denev, P.; Kratchanov, C.G.; Číž, M.; Lojek, A.; Kratchanova, M.G. Bioavailability and Antioxidant Activity of Black Chokeberry (Aronia melanocarpa) Polyphenols: in vitro and in vivo Evidences and Possible Mechanisms of Action: A Review. Compr. Rev. Food Sci. Food Saf. 2012, 11, 471–489. [CrossRef]
- Ellingsen, I.; Hjerkinn, E.M.; Seljeflot, I.; Arnesen, H.; Tonstad, S. Consumption of fruit and berries is inversely associated with carotid atherosclerosis in elderly men. Br. J. Nutr. 2008, 99, 674–681. [CrossRef]
- Rissanen, T.; Voutilainen, S.; Virtanen, J.; Venho, B.; Vanharanta, M.; Mursu, J.; Salonen, J. Low intake of fruits, berries and vegetables is associated with excess mortality in men: the Kuopio ischemic heart disease risk factor (KIHD) study. J. Nutr. 2003, 133, 199–204.
- Ross, J.A.; Kasum, C.M. DIETARY FLAVONOIDS: Bioavailability, Metabolic Effects, and Safety. Annu. Rev. Nutr. 2002, 22, 19–34. [CrossRef]
- Kuiper, J.J.W. Reactive Oxygen Species in Inflammasome Activation. MSc. I&I. thesis, University Medical Centre, Utrecht., 2009.
- Rahman, M.M.; Mohamed, M.R.; Kim, M.; Smallwood, S.; McFadden, G. Co-regulation of NF-kappaB and inflammasome-mediated inflammatory responses by myxoma virus pyrin domain-containing protein M013. PLoS Pathog. 2009, 5, e1000635. [CrossRef]
- Pacher, P.; Nivorozhkin, A.; Szabó, C. Therapeutic Effects of Xanthine Oxidase Inhibitors: Renaissance Half a Century after the Discovery of Allopurinol. Pharmacol. Rev. 2006, 58, 87–114. [CrossRef]
- Szeto, H.H. First-in-class cardiolipin-protective compound as a therapeutic agent to restore mitochondrial bioenergetics. Br. J. Pharmacol. 2014, 171(8), 2029-2050. [CrossRef]
- Daubert, M.A.; Yow, E.; Dunn, G. Novel mitochondria-targeting peptide in heart failure treatment: a randomized, placebo-controlled trial of elamipretide. Circ. Heart Fail. 2017, 10(12), e004389. [CrossRef]
- de Torre-Minguela, C.; del Castillo, P.M.; Pelegrín, P. The NLRP3 and Pyrin Inflammasomes: Implications in the Pathophysiology of Autoinflammatory Diseases. Front. Immunol. 2017, 8, 43. [CrossRef]
- MacKenzie, S.H.; Schipper, J.L.; Clark, A.C. The potential for caspases in drug discovery. Curr. Opin. Drug Discov. Devel. 2010, 13, 568-576.
- Barreyro, F.J.; Holod, S.; Finocchietto, P.V.; Camino, A.M.; Aquino, J.B.; Avagnina, A.; Carreras, M.C.; Poderoso, J.J.; Gores, G.J. The pan-caspase inhibitor Emricasan (IDN-6556) decreases liver injury and fibrosis in a murine model of non-alcoholic steatohepatitis. Liver Int. 2014, 35, 953–966. [CrossRef]
- Cornelis, S.; Kersse, K.; Festjens, N.; Lamkanfi, M.; Vandenabeele, P. Inflammatory Caspases: Targets for Novel Therapies. Curr. Pharm. Des. 2007, 13, 367–385. [CrossRef]
- Kesselheim, A.S.; Solomon, D.H. Incentives for Drug Development — The Curious Case of Colchicine. New Engl. J. Med. 2010, 362, 2045–2047. [CrossRef]
- Verma, S.; Eikelboom, J.W.; Nidorf, S.M. Colchicine in cardiac dissease: a systematic review and meta-analysis of randomized controlled trials. BMC Cardiovasc. Disord. 2015, 15, 96. [CrossRef]
- Leung, Y.Y.; Yao Hui, L.L.; Kraus, V.B. Colchicine—Update on mechanisms of action and therapeutic uses. Semin. Arthritis Rheum. 2015, 45, 341–350. [CrossRef]
- Calabrese, L.H. Anakinra treatment of patients with rheumatoid arthritis. Ann. Pharmacother. 2002, 36(7-8), 1204-1209. [CrossRef]
- Moran, A.; Bundy, B.; Becker, D.J.; A DiMeglio, L.; E Gitelman, S.; Goland, R.; Greenbaum, C.J.; Herold, K.C.; Marks, J.B.; Raskin, P.; et al. Interleukin-1 antagonism in type 1 diabetes of recent onset: two multicentre, randomised, double-blind, placebo-controlled trials. Lancet 2013, 381, 1905–1915. [CrossRef]
- Orrock, J.E.; Ilowite, N.T. Canakinumab for the treatment of active systemic juvenile idiopathic arthritis. Expert Rev. Clin. Pharmacol. 2016, 9, 1015–1024. [CrossRef]
- Ben-Zvi, I.; Kukuy, O.; Giat, E. Anakinra for colchicine-resistant familial Mediterranean fever: a randomized, double-blind, placebo-controlled trial. Arthritis Rheumatol. 2017, 69(4). [CrossRef]
- McKie, E.A.; Reid, J.L.; Mistry, P.C.; DeWall, S.L.; Abberley, L.; Ambery, P.D.; Gil-Extremera, B. A Study to Investigate the Efficacy and Safety of an Anti-Interleukin-18 Monoclonal Antibody in the Treatment of Type 2 Diabetes Mellitus. PLOS ONE 2016, 11, e0150018. [CrossRef]
- Curr. Top. Med. Chem. 2001, 1, 569–590. [CrossRef]
- Hertog, M.G.L.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S.; et al. Flavonoid Intake and Long-term Risk of Coronary Heart Disease and Cancer in the Seven Countries Study. Arch. Intern. Med. 1995, 155, 381–386. [CrossRef]
- Oszmiański, J.; Wojdylo, A. Aronia melanocarpa phenolics and their antioxidant activity. Eur. Food Res. Technol. 2005, 221, 809–813. [CrossRef]
- Ohgami, K.; Ilieva, I.; Shiratori, K.; Koyama, Y.; Jin, X.-H.; Yoshida, K.; Kase, S.; Kitaichi, N.; Suzuki, Y.; Tanaka, T.; et al. Anti-inflammatory Effects of Aronia Extract on Rat Endotoxin-Induced Uveitis. Investig. Opthalmology Vis. Sci. 2005, 46, 275–281. [CrossRef]
- Tauber, A.I.; Fay, J.R.; Marletta, M.A. Flavonoid inhibition of the human neutrophil NADPH-oxidase. Biochem. Pharmacol. 1984, 33, 1367–1369. [CrossRef]
- Middleton, E. Jr. Effect of plant flavonoids on immune and inflammatory cell function. Adv. Exp. Med. Biol. 1998, 439, 175-182.
- Tordera, M.; Ferrándiz, M.L.; Alcaraz, M.J. Influence of Anti-Inflammatory Flavonoids on Degranulation and Arachidonic Acid Release in Rat Neutrophils. 1994, 49, 235–240. [CrossRef]
- Schmidt, R.L.; Lenz, L.L. Distinct Licensing of IL-18 and IL-1β Secretion in Response to NLRP3 Inflammasome Activation. PLOS ONE 2012, 7, e45186. [CrossRef]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820.
- Nicola, C. Dental Materials. Clinical and tehnological considerations. House of science books: Cluj-Napoca, Romania, 2009; pp. 16-34.
- Bernardi, S.; Macchiarelli, G.; Bianchi, S. Autologous Materials in Regenerative Dentistry: Harvested Bone, Platelet Concentrates and Dentin Derivates. Molecules 2020, 25, 5330. [CrossRef]
- Hollý, D.; Klein, M.; Mazreku, M.; Zamborský, R.; Polák, .; Danišovič, .; Csöbönyeiová, M. Stem Cells and Their Derivatives—Implications for Alveolar Bone Regeneration: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 11746. [CrossRef]
- Vouvoudi, E.C. Overviews on the Progress of Flowable Dental Polymeric Composites: Their Composition, Polymerization Process, Flowability and Radiopacity Aspects. Polymers 2022, 14, 4182. [CrossRef]
- Valandro, L.F.; Cadore-Rodrigues, A.C.; Dapieve, K.S.; Machry, R.V.; Pereira, G.K.R. A brief review on fatigue test of ceramic and some related matters in Dentistry. J. Mech. Behav. Biomed. Mater. 2023, 138, 105607. [CrossRef]
- Albeshir, E.G.; Alsahafi, R.; Albluwi, R.; Balhaddad, A.A.; Mitwalli, H.; Oates, T.W.; Hack, G.D.; Sun, J.; Weir, M.D.; Xu, H.H.K. Low-Shrinkage Resin Matrices in Restorative Dentistry-Narrative Review. Materials 2022, 15, 2951. [CrossRef]
- Zong, C.; Bronckaers, A.; Willems, G.; He, H.; de Llano-Pérula, M.C. Nanomaterials for Periodontal Tissue Regeneration: Progress, Challenges and Future Perspectives. J. Funct. Biomater. 2023, 14, 290. [CrossRef]
- Qiu, J.; Liu, X.; You, B.; Ren, N.; Liu, H. Application of Nanomaterials in Stem Cell-Based Therapeutics for Cardiac Repair and Regeneration. Small 2023, 19, e2206487. [CrossRef]
- Hu, T.; Gu, Z.; Williams, G.R.; Strimaite, M.; Zha, J.; Zhou, Z.; Zhang, X.; Tan, C.; Liang, R. Layered double hydroxide-based nanomaterials for biomedical applications. Chem. Soc. Rev. 2022, 51, 6126–6176. [CrossRef]
- Farkas, N.-I.; Marincaș, L.; Barabás, R.; Bizo, L.; Ilea, A.; Turdean, G.L.; Toșa, M.; Cadar, O.; Barbu-Tudoran, L. Preparation and Characterization of Doxycycline-Loaded Electrospun PLA/HAP Nanofibers as a Drug Delivery System. Materials 2022, 15, 2105. [CrossRef]
- Al-Enizi, A.M.; Zagho, M.M.; Elzatahry, A.A. Polymer-Based Electrospun Nanofibers for Biomedical Applications. Nanomaterials 2018, 8, 259. [CrossRef]
- Torres-Martinez, E.J.; Cornejo-Bravo, J.M.; Serrano-Medina, A.; Perez-González, G.L.; Gómez, L.J.V. A Summary of Electrospun Nanofibers as Drug Delivery System: Drugs Loaded and Biopolymers Used as Matrices. Curr. Drug Deliv. 2018, 15, 1360–1374. [CrossRef]
- Gao, Y.; Truong, Y.B.; Zhu, Y.; Kyratzis, I.L. Electrospun antibacterial nanofibers: Production, activity, and in vivo applications. J. Appl. Polym. Sci. 2014, 131. [CrossRef]
- Zhang, H.; Fu, Q.W.; Sun, T.W.; Chen, F.; Qi, C.; Wu, J.; Cai, Z.Y.; Qian, Q.R.; Zhu, Y.J. Amorphous calcium phosphate, hydroxyapatite and poly (d,l-lactic acid) composite nanofibers: Electrospinning preparation, mineralization and in vivo bone defect repair. Colloids Surfaces B Biointerfaces 2015, 136, 27–36.
- Liu, S.; Zheng, Y.; Liu, R.; Tian, C. Preparation and characterization of a novel polylactic acid/hydroxyapatite composite scaffold with biomimetic micro-nanofibrous porous structure. J. Mater. Sci. Mater. Med. 2020, 31, 1–11. [CrossRef]
- Hartatiek; Fahmi, N.K.; Putra, A.A.D.; Yudyanto; Nasikhudin; Utomo, J.; Ahmad, N. Effect of nano-hydroxyapatite (n-HAp)/PLA scaffold composites on porosity and microstructure. AIP Conf. Proc. 2020, 2234.
- Andrei, V.; Andrei, S.; Gal, A.F.; Rus, V.; Gherman, L.-M.; Boșca, B.A.; Niculae, M.; Barabas, R.; Cadar, O.; Dinte, E.; et al. Immunomodulatory Effect of Novel Electrospun Nanofibers Loaded with Doxycycline as an Adjuvant Treatment in Periodontitis. Pharmaceutics 2023, 15, 707. [CrossRef]
- Singhvi, M.S.; Zinjarde, S.S.; Gokhale, D.V. Polylactic acid: synthesis and biomedical applications. J. Appl. Microbiol. 2019, 127, 1612–1626. [CrossRef]
- Raviolo, M.; Solana, M.; Novoa, M.; Gualdesi, M.; Alba-Soto, C.; Briñón, M. Synthesis, physicochemical properties of allopurinol derivatives and their biological activity against Trypanosoma cruzi. Eur. J. Med. Chem. 2013, 69, 455–464. [CrossRef]
- Badis, K.; Merine, H.; Ramli, Y.; Larbi, O.; Memou, C.H. Effect of Polymers nature and Stirring Speeds on Physicochemical Properties and the Controlled Release of Allopurinol-loaded Microspheres. J. Mex. Chem. Soc. 2021, 66, 17–33. [CrossRef]
- Changdeo, J.S.; Kuchekar, M.V.; Shankar, B.; Rajaram, C.A. Physicochemical characterization and solubility enhancement studies of allopurinol solid dispersions. Brazilian Journal of Pharmaceutical Sciences 2011, 47. [CrossRef]
- Kandav, G.; Bhatt, D.; Jindal, D.K. FORMULATION AND EVALUATION OF ALLOPURINOL LOADED CHITOSAN NANOPARTICLES. Int. J. Appl. Pharm. 2019, 49–52. [CrossRef]
- Varrica, C.; Carvalheiro, M.; Faria-Silva, C.; Eleutério, C.; Sandri, G.; Simões, S. Topical Allopurinol-Loaded Nanostructured Lipid Carriers: A Novel Approach for Wound Healing Management. Bioengineering 2021, 8, 192. [CrossRef]
- Kandav, G.; Bhatt, D.C.; Jindal, D.K.; Singh, S.K. Formulation, Optimization, and Evaluation of Allopurinol-Loaded Bovine Serum Albumin Nanoparticles for Targeting Kidney in Management of Hyperuricemic Nephrolithiasis: Formulation, optimization, and evaluation of ABNPs for kidney targeting, AAPS. Pharm. Sci. Tech. 2020, 21, 164. [CrossRef]
- Birk, A.V.; Liu, S.; Soong, Y.; Mills, W.; Singh, P.; Warren, J.D.; Seshan, S.V.; Pardee, J.D.; Szeto, H.H. The Mitochondrial-Targeted Compound SS-31 Re-Energizes Ischemic Mitochondria by Interacting with Cardiolipin. J. Am. Soc. Nephrol. 2013, 24, 1250–1261. [CrossRef]
- Liu, D.; Jin, F.; Shu, G.; Xu, X.; Qi, J.; Kang, X.; Yu, H.; Lu, K.; Jiang, S.; Han, F.; You, J.; Du, Y.; Ji, J. Enhanced efficiency of mitochondria-targeted peptide SS-31 for acute kidney injury by pH-responsive and AKI-kidney targeted nanopolyplexes. Biomaterials 2019, 211, 57-67. [CrossRef]
- Shan, L.; Wang, F.; Zhai, D.; Meng, X.; Liu, J.; Lv, X. New Drugs for Hepatic Fibrosis. Front. Pharmacol. 2022, 13, 874408. [CrossRef]
- Harrison, S.A.; Goodman, Z.; Jabbar, A.; Vemulapalli, R.; Younes, Z.H.; Freilich, B.; Sheikh, M.Y.; Schattenberg, J.M.; Kayali, Z.; Zivony, A.; et al. A randomized, placebo-controlled trial of emricasan in patients with NASH and F1-F3 fibrosis. J. Hepatol. 2020, 72, 816–827. [CrossRef]
- Rudolphi, K.; Gerwin, N.; Verzijl, N.; van der Kraan, P.; Berg, W.v.D. Pralnacasan, an inhibitor of interleukin-1β converting enzyme, reduces joint damage in two murine models of osteoarthritis. Osteoarthr. Cartil. 2003, 11, 738–746. [CrossRef]
- Poreba, M.; Szalek, A.; Kasperkiewicz, P.; Rut, W.; Salvesen, G.S.; Drag, M. Small Molecule Active Site Directed Tools for Studying Human Caspases. Chem. Rev. 2015, 115, 12546–12629. [CrossRef]
- Alkadi, H.; Khubeiz, M.J.; Jbeily, R. Colchicine: A Review on Chemical Structure and Clinical Usage. Infect. Disord. - Drug Targets 2018, 18, 105–121. [CrossRef]
- Sonny Larsson, N.R. Reviewing Colchicaceae Alkaloids – Perspectives of Evolution on Medicinal Chemistry. Current Topics in Med. Chem. 2014, 14, 274-289.
- Ede Bodoki, D.B.; Săndulescu, R. Ab Initio study of the Na–colchicine positively charged complex. Farmacia Journal, 2015, 64.
- Gorabi, A.M.; Kiaie, N.; Reiner, .; Carbone, F.; Montecucco, F.; Sahebkar, A. The Therapeutic Potential of Nanoparticles to Reduce Inflammation in Atherosclerosis. Biomolecules 2019, 9, 416. [CrossRef]
- Ramírez, J.; Cañete, J.D. Anakinra for the treatment of rheumatoid arthritis: a safety evaluation. Expert Opin. Drug Saf. 2018, 17, 727–732. [CrossRef]
- Dubois, E.A.; Rissmann, R.; Cohen, A.F. Rilonacept and canakinumab. Br. J. Clin. Pharmacol. 2011, 71, 639–641. [CrossRef]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [CrossRef]
- Dobrzynska, M.; Napierala, M.; Florek, E. Flavonoid Nanoparticles: A Promising Approach for Cancer Therapy. Biomolecules 2020, 10, 1268. [CrossRef]
- Zverev, Y.F.; Rykunova, A.Y. Modern Nanocarriers as a Factor in Increasing the Bioavailability and Pharmacological Activity of Flavonoids. Appl. Biochem. Microbiol. 2022, 58, 1002–1020. [CrossRef]
- Ting, J.P.-Y.; Kastner, D.L.; Hoffman, H.M. CATERPILLERs, pyrin and hereditary immunological disorders. Nat. Rev. Immunol. 2006, 6, 183–195. [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002, 10(2), 4. [CrossRef]
- Jin, S.; Xia, X.; Huang, J.; Yuan, C.; Zuo, Y.; Li, Y.; Li, J. Recent advances in PLGA-based biomaterials for bone tissue regeneration. Acta Biomater. 2021, 127, 56–79. [CrossRef]
- Armenia, I.; Ayllón, C.C.; Herrero, B.T.; Bussolari, F.; Alfranca, G.; Grazú, V.; de la Fuente, J.M. Photonic and magnetic materials for on-demand local drug delivery. Adv. Drug Deliv. Rev. 2022, 191, 114584. [CrossRef]
- Zong, C.; Van Holm, W.; Bronckaers, A.; Zhao, Z.; Čokić, S.; Aktan, M.K.; Castro, A.B.; Van Meerbeek, B.; Braem, A.; Willems, G.; et al. Biomimetic Periodontal Ligament Transplantation Activated by Gold Nanoparticles Protects Alveolar Bone. Adv. Heal. Mater. 2023, 12, e2300328. [CrossRef]
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knüchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [CrossRef]
- Ding, Q.; Cui, J.; Shen, H.; He, C.; Wang, X.; Shen, S.G.F.; Lin, K. Advances of nanomaterial applications in oral and maxillofacial tissue regeneration and disease treatment. WIREs Nanomed. Nanobiotechnology 2020, 13, e1669. [CrossRef]
- Feng, W.; Chen, Y. Chemoreactive nanomedicine. J. Mater. Chem. B 2020, 8, 6753–6764. [CrossRef]
- Lenders, V.; Koutsoumpou, X.; Sargsian, A.; Manshian, B.B. Biomedical nanomaterials for immunological applications: ongoing research and clinical trials. Nanoscale Adv. 2020, 2, 5046–5089. [CrossRef]
- Tiwari, N.; Osorio-Blanco, E.R.; Sonzogni, A.; Esporrín-Ubieto, D.; Wang, H.; Calderón, M. Nanocarriers for Skin Applications: Where Do We Stand? Angew. Chem. Int. Ed. Engl. 2022, 61, e202107960.
- Cheng, Z.; Li, M.; Dey, R.; Chen, Y. Nanomaterials for cancer therapy: current progress and perspectives. J. Hematol. Oncol. 2021, 14, 1–27. [CrossRef]
Figure 1.
The cascade underlying the transition from symbiosis, to dysbiosis, of the oral microbial communities, with the onset of inflammation, activation of adaptive immunity, followed by osteoclasts differentiation and alveolar bone resorption.
Figure 1.
The cascade underlying the transition from symbiosis, to dysbiosis, of the oral microbial communities, with the onset of inflammation, activation of adaptive immunity, followed by osteoclasts differentiation and alveolar bone resorption.
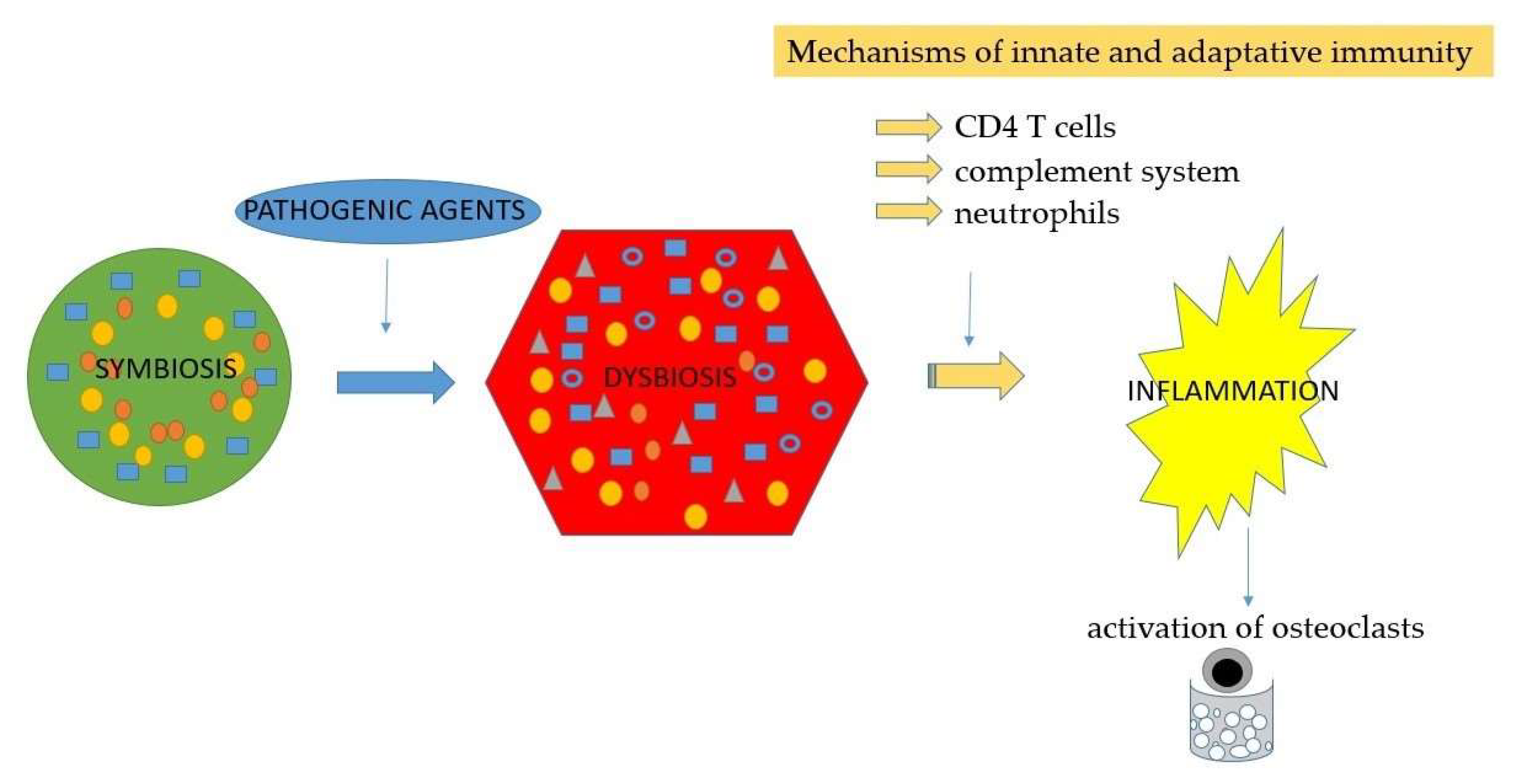
Figure 2.
The reaction to a stimulus of primed cells versus trained cells, and the induction of epiphenetic changes.
Figure 2.
The reaction to a stimulus of primed cells versus trained cells, and the induction of epiphenetic changes.
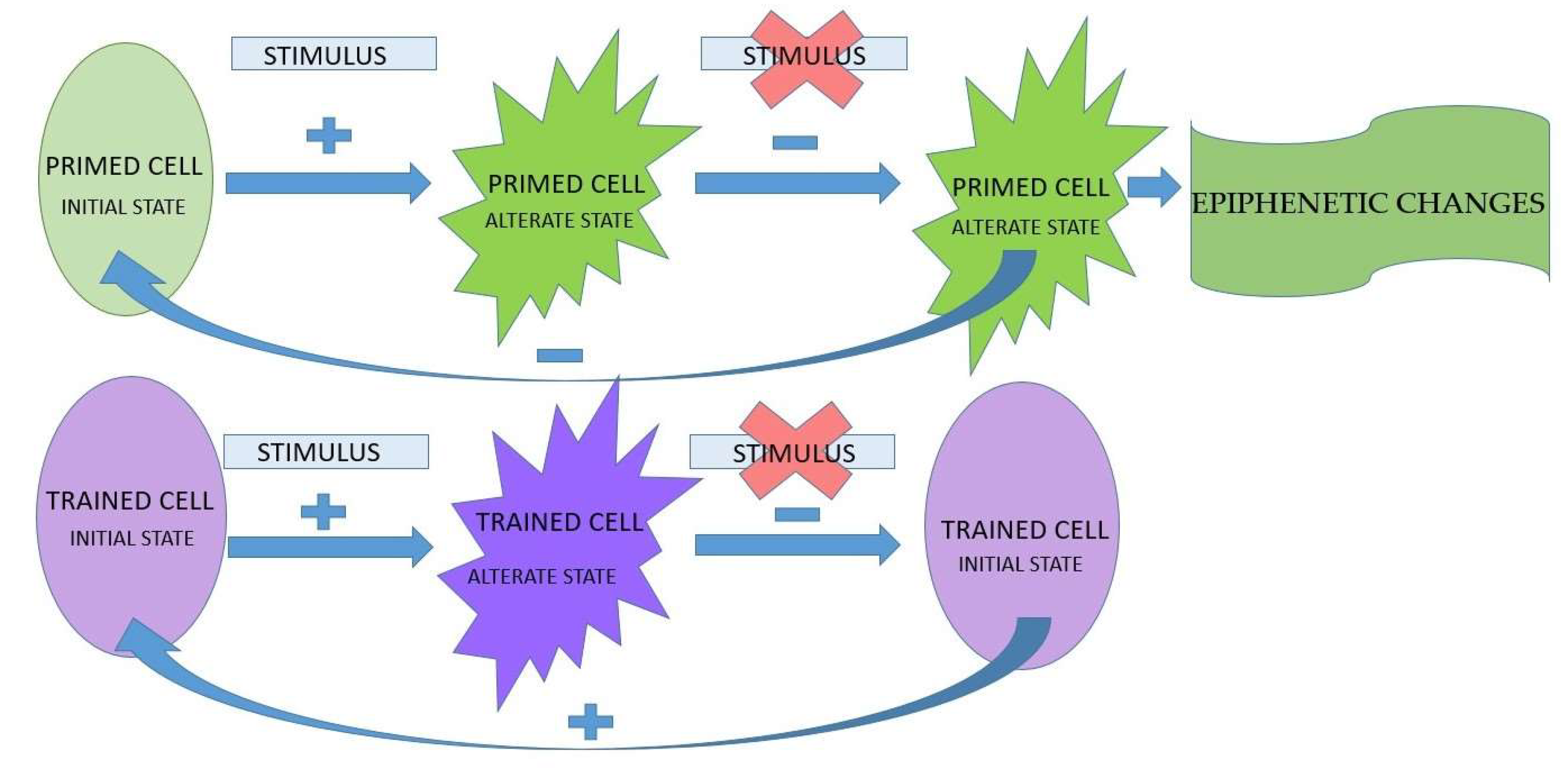
Figure 3.
Activation of the complement system through the three possible pathways, and their convergence towards the C3 central complement component.
Figure 3.
Activation of the complement system through the three possible pathways, and their convergence towards the C3 central complement component.
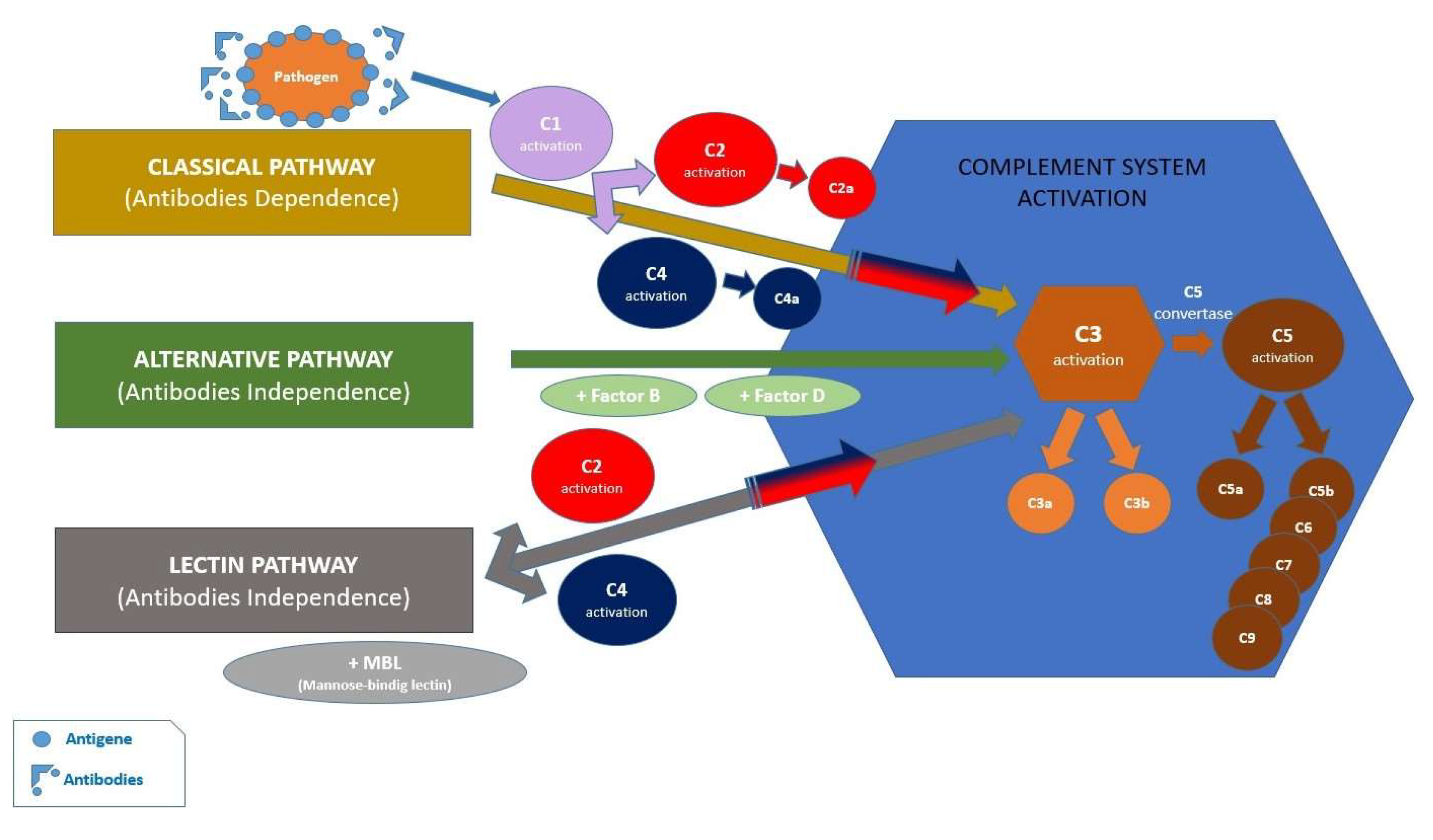
Figure 4.
Schematic representation of canonical and non-canonical pyroptosis activation. Both pathways lead GSDMD cleavage, followed by release of interleukins 1β (IL-1β) and 18 (IL-18), and the subsequent initiation of the inflammatory response.
Figure 4.
Schematic representation of canonical and non-canonical pyroptosis activation. Both pathways lead GSDMD cleavage, followed by release of interleukins 1β (IL-1β) and 18 (IL-18), and the subsequent initiation of the inflammatory response.
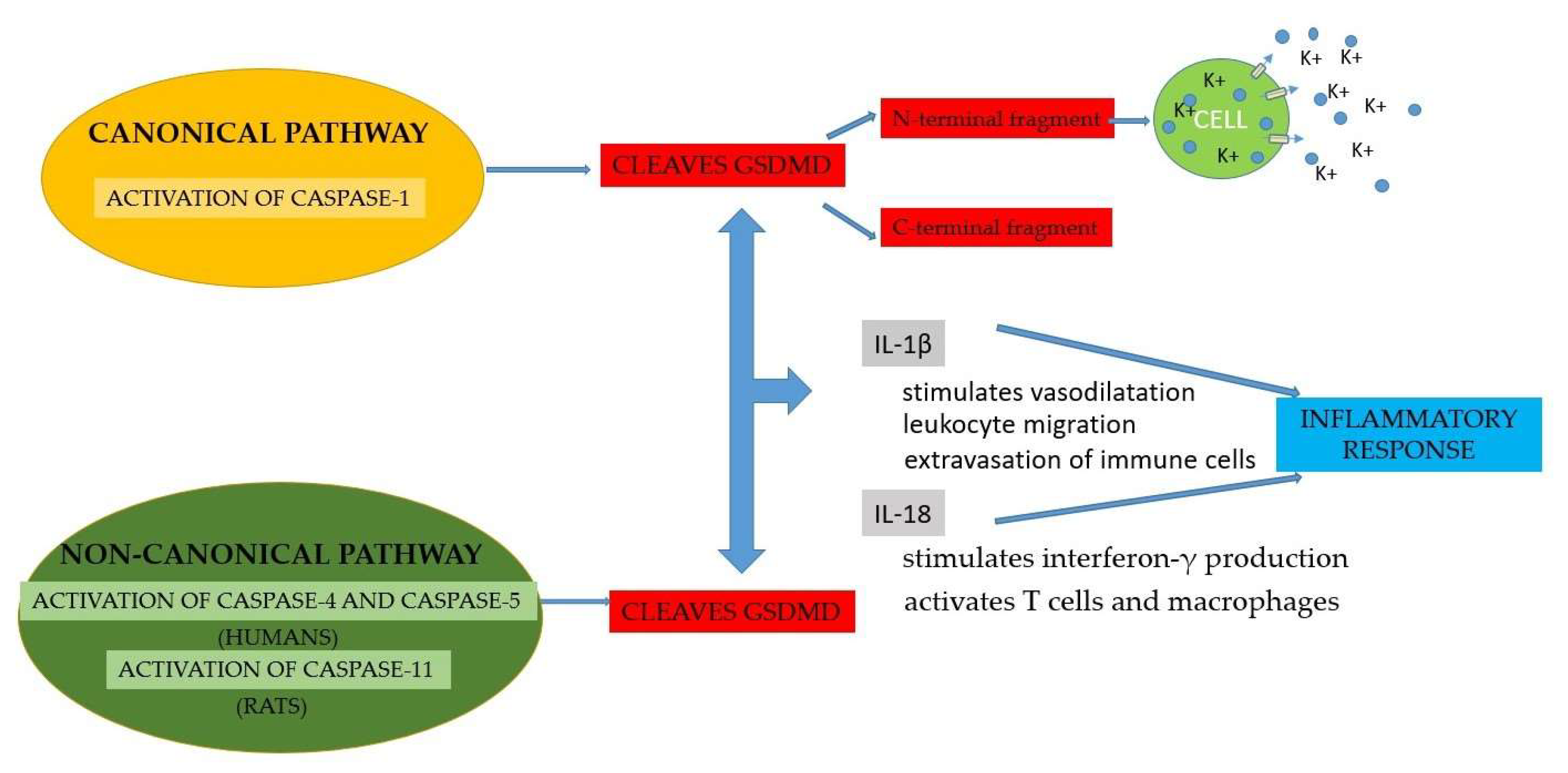
Figure 5.
Graphical representation of how ROS production can determine caspase-1 activation through a direct mechanism and caspase-3 activation through an indirect mechanism, through ERK1/2 and finally the activation of the NLRP3 inflammasome (LRR - Leucine-rich-repeat, PYD - Domain Pyrin, NBD - Nucleotide-Binding-Domain, CARD - Caspase Activation and Recruitment Domain).
Figure 5.
Graphical representation of how ROS production can determine caspase-1 activation through a direct mechanism and caspase-3 activation through an indirect mechanism, through ERK1/2 and finally the activation of the NLRP3 inflammasome (LRR - Leucine-rich-repeat, PYD - Domain Pyrin, NBD - Nucleotide-Binding-Domain, CARD - Caspase Activation and Recruitment Domain).
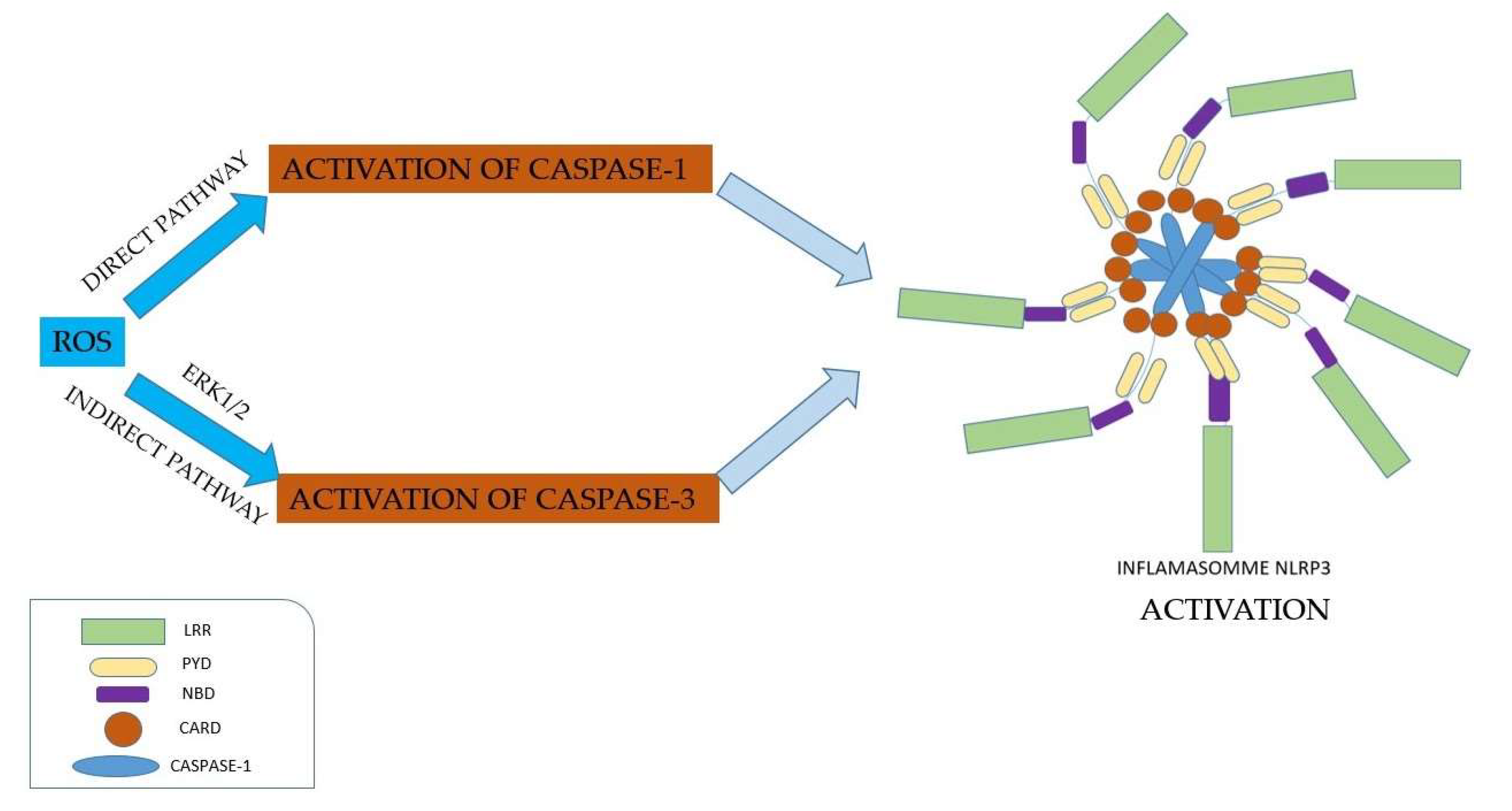
Figure 6.
Methods of inhibiting the inflammasome, through direct and indirect action. The inhibition of the inflammasome by direct mechanism can be achieved by three methods: inhibition of intracellular signaling pathways, blockage of the inflammasome components, and inhibition of cytokines mediated by the inflammasome. Several pharmaceutical preparations act on reactive oxygen species (ROS) and thus can inhibit the of the inflammasome activation, representing the indirect mechanism of inhibition (LRR - Leucine-rich-repeat, PYD - Domain Pyrin, NBD - Nucleotide-Binding-Domain, CARD - Caspase Activation and Recruitment Domain).
Figure 6.
Methods of inhibiting the inflammasome, through direct and indirect action. The inhibition of the inflammasome by direct mechanism can be achieved by three methods: inhibition of intracellular signaling pathways, blockage of the inflammasome components, and inhibition of cytokines mediated by the inflammasome. Several pharmaceutical preparations act on reactive oxygen species (ROS) and thus can inhibit the of the inflammasome activation, representing the indirect mechanism of inhibition (LRR - Leucine-rich-repeat, PYD - Domain Pyrin, NBD - Nucleotide-Binding-Domain, CARD - Caspase Activation and Recruitment Domain).
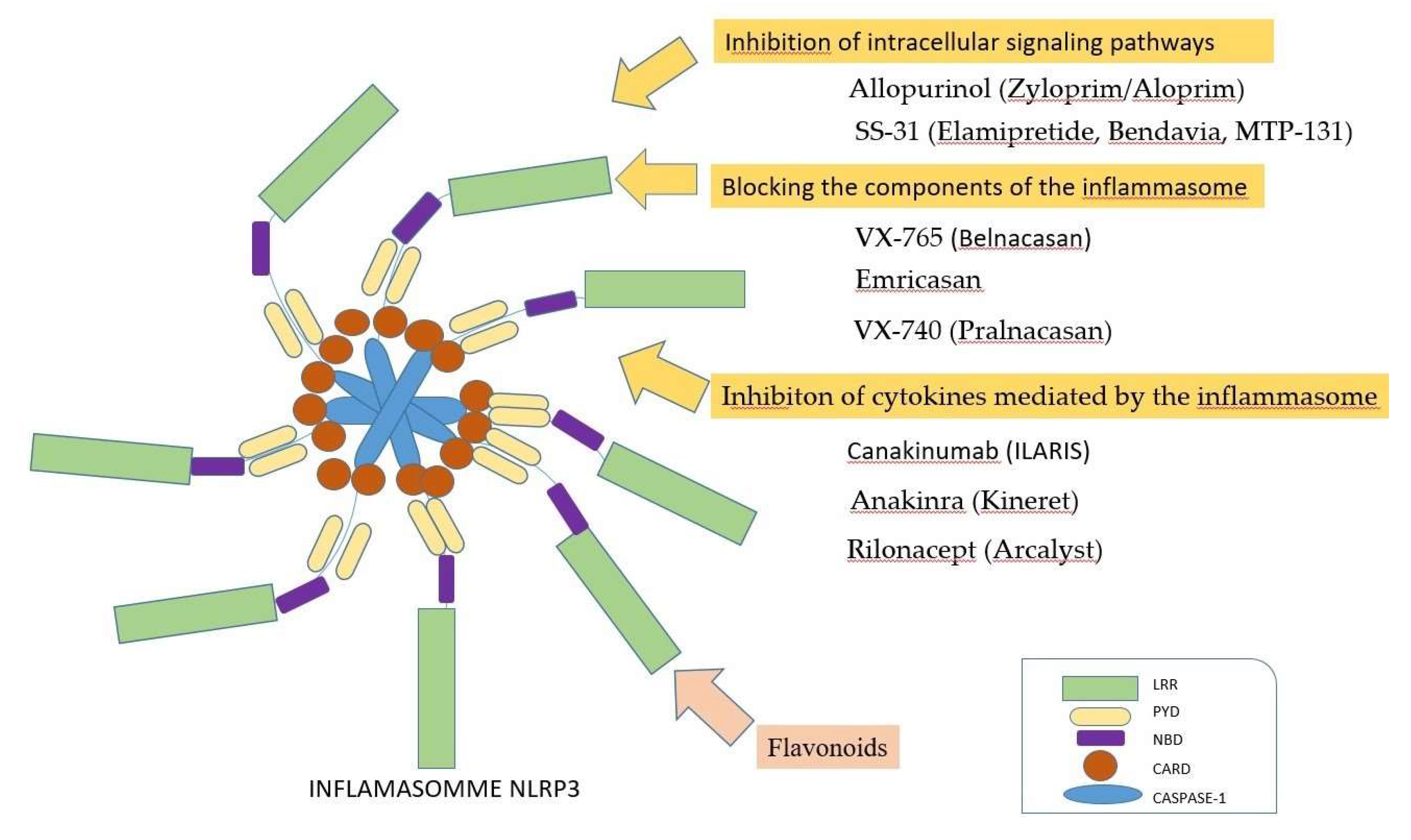
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



