
Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Sarcocystis sp. are coccidian protozoans belonging to the Apicomplexa phylum. As other members of this phylum, they are obligate intracellular parasites having a complex cellular machinery for the invasion of host cells. Sarcocystis spp. display dixenous life cycles, involving a predator and a prey as definitive and intermediate hosts, respectively. Specifically, these parasites develop sarcocysts in the tissues of their intermediate hosts, ranging in size from microscopic to visible to the naked eye, depending on the species. When definitive hosts consume sarcocysts, infective forms are produced in the digestive system and discharged into the environment via feces. Consumption of oocyst-contaminated water and pasture by the intermediate host completes the parasitic cycle. More than 200 Sarcocystis spp. have been described infecting wildlife, domestic animals, and humans, some of which are of economic or public health importance. Interestingly, Old World camelids (dromedary, domestic Bactrian camel, and wild Bactrian camel) and New World or South American camelids (llama, alpaca, guanaco, and vicuña) can each be infected by two different Sarcocystis species: Old World camelids by S. cameli (producing micro- and macroscopic cysts) and S. ippeni (microscopic cysts); and South American camelids by S. aucheniae (macroscopic cysts) and S. masoni (microscopic cysts). Large numbers of Old and New World camelids are bred for meat production, but the finding of macroscopic sarcocysts in carcasses significantly hampers meat commercialization. This review tries to compile the information that is currently accessible regarding the biology, epidemiology, phylogeny, and diagnosis of Sarcocystis spp. infecting Old and New World camelids. In addition, knowledge gaps will be identified to encourage research that will lead to the control of these parasites.
Keywords:
South American camelids
; Old World camels
; Sarcocystis
; sarcocysts
1. Introduction
Sarcocystosis is a parasitic infection caused by different species of protozoans belonging to the Sarcocystis genus. With over 200 described species, different Sarcocystis spp. infect mammals, including humans, as well as birds, and reptiles. Their life cycles include a predator and a prey that serve as definitive and intermediate hosts, respectively. Typical of the intermediate host phase of Sarcocystis is the formation of sarcocysts - wall-surrounded capsules in which the parasites divide asexually - which may be micro or macroscopic, depending on the species [1].
Sarcocystis complex life cycles alternate between invading, dividing and sexual stages. After a predator ingests meat of a prey containing sarcocysts, bradyzoites -the infective stage borne in these structures- invade goblet cells of the predator’s intestine. Micro and macrogametes are formed and gamete fusion leads to the formation of an oocyst which, after being excreted with the feces into the environment, sporulates to form four sporozoites. Intermediate hosts ingest oocysts when grazing or drinking contaminated pasture or water, and enzymes in their intestine lead to the liberation of sporozoites, which invade endothelial cells of mesenteric lymph node arteries. Here, they reproduce asexually by schizogony, forming schizonts with lobed nuclei, giving them the appearance of a rosette, from which merozoites eventually bud. Merozoites are released into the bloodstream, disseminating the infection in the intermediate host. They can be found free or inside mononuclear cells, where they undergo binary division by endodyogeny. When merozoites invade the endothelial cells of downstream arterioles, capillaries, and veins, a new cycle of schizogony is initiated. In turn, invasion of a myocyte or a nervous cell by a merozoite leads to the formation of sarcocysts, the final stage in the intermediate host. The parasite remains inside a parasitophorous vacuole (PV), and the PV membrane (PMV) together with parasite secretions form a wall that provides a safe microenvironment for parasite multiplication. Depending on the Sarcocystis species, final-destination cells can be myocytes of skeletal or cardiac muscles, or neural cells. In the sarcocysts, merozoites transform into metrocytes, which rapidly divide by endodyogeny and eventually turn into infective bradyzoites [1,2,3] (Figure 1).
Most Sarcocystis spp. infecting livestock have a worldwide distribution and, in general, occur with high prevalence in both industrialized and developing countries [3]. The toll of these infections varies depending on the Sarcocystis species and include mortality, morbidity, abortion, low meat yield in productive animals, and economic loss due to the presence of macroscopic sarcocysts in meat destined for human consumption. However, for most Sarcocystis species, especially those infecting wildlife, their effect on host fitness is unknown [1,4].
Camelids are found in different continental regions and, accordingly, are divided into species of the Old World and species of the New World. Both types are important for biodiversity and have a long history of association with humans. Both can also be infected by Sarcocystis spp., sometimes with relevant negative impact on local economies [5,6,7].
This review will present the available information on Sarcocystis spp. infecting camelids in the Old and the New World, draw parallels between these two scenarios and analyze the phylogenetic relationships among these parasites.
2. Main features of camelids
Camelids are Artiodactyla mammals grouped in the Camelidae family, the only extant family of the Tylopoda suborder (from the Greek: “feet with cushions”). They regurgitate and rechew food but are not ruminants and are differentiated from the latter by several anatomical features, including their stomach with three compartments, lack of horns, and the presence of real canine teeth and plantar cushions, as well as other physiological and behavioral features. Importantly, camelids differ from ruminants in their susceptibility to microbial and parasitic diseases [8].
Old World camelids (OWC) belong to the genus Camelus, which comprises three extant species: C. dromedarius (one-humped camel or dromedary), C. bactrianus (two-humped or Bactrian camel), both of which have been domesticated, and the wild and highly endangered C. ferus (two humps). The population size of OWC has been estimated to be at least 35.5 million heads, of which 95 % are dromedaries [9]. South American camelids (SAC) comprise four species: the wild Lama guanicoe (guanaco) and Vicugna vicugna (vicuña) and the domesticated Lama glama (llama) and Vicugna pacos (alpaca), with an overall estimated population of 10 million heads [10]. Notably, llamas and alpacas have been introduced to farms in some European countries, South Africa, and Australia, and thus, their geographic distribution and numbers of heads are considerably larger than those mentioned here [9,10].
Both OWC and SAC are adapted to harsh environmental conditions, including extreme temperatures, intense solar radiation, water scarcity, and poor pastures. Under such rough settings, most other livestock species are either unable to thrive or show a significant decline in production. Camelids thus constitute an attractive livestock choice in a scenario of climate change, shortage of water and reduction of agricultural areas [11].
Camelids have a long history of association with humans. Human groups who lived as gatherers and hunters must have found camels a good source of food and hides, and through their domestication some 3 to 7 thousand years ago (kya), they became important suppliers of vital goods to ancient civilizations in the Old and New Worlds. [12,13]. Currently, camelids continue to be an important asset to a considerable number of human populations, and among other benefits, their meat is a valuable source of animal protein and an attractive product for the gourmet cuisine. Meat from camelids has a higher ratio of protein to cholesterol than conventional sources of red meat. This, combined with the lower environmental impact of camelids compared to traditional cattle, meets the profile of environmentally and health-conscious consumers. Llama meat is consumed in Bolivia and northwestern Argentina, alpaca meat in Peru, and OWC meat in North Africa, the Middle East, Central Asia and China [14,15,16].
Notably, a major limitation to the marketing and consumption of SAC and OWC meat is the frequent presence of macroscopic sarcocysts, which give it an unappetizing appearance. The finding of sarcocysts can also result in confiscation of up to 60 to 100 % of carcasses by health authorities or devaluation of their commercial value, leading to financial losses. As a consequence, the number of animals slaughtered informally significantly exceeds the numbers of animals slaughtered in authorized abattoirs. This results in the informal marketing of meat obtained without the hygienic and sanitary conditions to guarantee food safety [7]. In addition, human consumption of insufficiently cooked meat contaminated with sarcocysts can lead to gastroenteritis [1,5,6].
3. Sarcocystis infecting OWC and SAC
Four Sarcocystis spp. have so far been described to infect camelids: S. cameli and S. ippeni for OWC and S. aucheniae and S. masoni for SAC [3,17].
The first observation of Sarcocystis infections in OWC was carried out in Egypt by Mason (1910) who reported macroscopic sarcocysts in muscles of camels in Egypt and used the name S. cameli to refer to the etiological agent. Later, several additional case reports of Sarcocystis species infecting OWC appeared in the literature, which were designated as S. ippeni, S. camelicanis, S. camelocanis and S. miescheri, depending on their sarcocyst wall or oocyst features. A taxonomic revision of OWC sarcocystosis led to the acceptance of S. cameli and S. ippeni as valid species, while S. camelicanis, S. camelocanis and S. miescheri were considered synonymous with S. cameli. Importantly, the vast majority of Sarcocystis reports are from dromedary camels [6,17,18].
In the case of SAC, the first description of a macroscopic sarcocyst in a llama in Australia took place in 1913, and the corresponding parasite was named S. aucheniae [19]. Later, the names S. tilopodi and guanicoecanis were used for parasites forming sarcocysts in guanaco [20,21]. A Sarcocystis species forming microscopic cysts in SACs was initially named S. lamacanis [21,22,23]. Electron microscopy and molecular studies established S. aucheniae as the only species forming macroscopic cysts in llamas, alpacas, and guanacos, while the species forming microscopic cysts was redescribed as S. masoni, in honor to Dr Eugene Mason. Thus, the names S. tilopodi, S. guanicoecanis and S. lamacanis are currently considered invalid [3,24,25].
In different Sarcocystis species, sarcocysts vary in shape (globular, filamentous, fusiform), size, and other characteristics, such as the presence or absence of internal partitions and variations in their wall ultrastructure [2,3]. In the case of Sarcocystis spp. infecting camelids, both S. aucheniae and S. cameli generate macroscopic sarcocysts that are visible to the naked eye (oval, 5-20 mm x 2 mm, and fusiform, 1.5-5 mm x 0.2-0.4 mm, respectively). Microscopic cysts of S. cameli (700 x 100 µm) have also been described. In turn, only microcysts were described for S. masoni (fusiform, 800 x 95 µm) and S. ippeni (globular, 100-120 x 50-100 µm) [3,17,25] (Table 1).
In all Sarcocystis spp., the sarcocyst wall essentially consists of the PVM covering a granular, electron-dense layer, from which septa can arise. When present, septa cross the cyst, separating its cavity into compartments, where metrocytes and bradyzoites are found. The number of parasites contained in a sarcocyst varies with the species and the stage of maturation: young cysts as small as 5 μm in diameter might contain only two parasites, while a mature macroscopic cyst can contain 20 million, as has been observed for S. aucheniae [3,24].
The sarcocyst wall can remain relatively simple in some species and, in others, form projections (villar protrusions) of different sizes and shapes, that bulge outwardly, and which can contain microfilaments, microtubules, electron-dense bodies, minute granules and small vesicles [1,3]. Dubey et al (2015) have defined at least 82 ultrastructural types of cyst walls between different Sarcocystis species. Notably, the cyst walls of both S. cameli and S. masoni have a common “9j” conformation, characterized by the presence of upright finger-like villar protrusions, with knob-like structures arising from the PVM, in which microtubules can be observed [3,25]. S. aucheniae presents a ‘type 21‘ sarcocyst wall ultrastructure, with highly branched cauliflower-like villar protrusions, similar to that of S. gigantea [1,3,25]. Finally, S. ippeni has a characteristic ‘type 32’ sarcocyst wall structure. This type of ultrastructure is characterized by thorn-like villar protrusions, with microtubules radiating into the granular layer, and has not been previously described in any other Sarcocystis species [3] (Table 1).
Figure 2 exemplifies different aspects of sarcocysts produced by S. aucheniae in the skeletal muscles of llamas and alpacas.
4. Definitive hosts
Determination of the definitive host(s) of camelid Sarcocystis spp. has so far been based on experimental infection studies. As will be briefly related in the next paragraphs, these studies have indicated that the domestic dog (Canis familiaris) can act as definitive host of S. aucheniae, S. masoni, and S. cameli. On the other hand, no reports on definitive hosts for S. ippeni, are available. In most cases, microscopic observation of oocysts or sporocysts in feces has been used as conclusive evidence. However, confirmatory studies in which the species of the excreted parasites is determined by molecular methods would be desirable.
In the case of S. aucheniae, [21] dogs, cats, rats, and mice were fed with raw guanaco meat containing macrocysts. Sporocysts were later only observed in dog feces. Excretion of Sarcocystis sp. sporocysts in feces of dogs fed with SAC meat was also confirmed by other authors [29,30,31].
For S. masoni, experimental infection has also determined that the dog can serve as a definitive host. In this case, the species of excreted oocysts was confirmed by PCR-RFLP analysis of the cytochrome c oxidase subunit I (cox-1) gene [32].
In the case of S. cameli, sporocysts were observed in feces of dogs fed with dromedary meat infested with sarcocysts of this species [33,34,35,36].
Histological studies of the intestine of dogs fed with S. masoni- or S. aucheniae-infected SAC meat showed that oocysts and sporocysts were mostly concentrated towards the tips of the vellosities, without further alterations in the mucosa [31,32]. Gamogony was observed within the lamina propria of the intestine of puppies that had been fed with S. cameli-infected meat [37]. S. cameli zygotes were first observed in the lamina propria 24 h after meat ingestion, and sporulated oocysts were evident in feces at 7-13 days post ingestion [33,34,35,36,37,38]. Prepatent periods of 8-9 days and 9-16 days were observed for S. masoni and S. aucheniae, respectively [29,30,31].
Consumption of S. aucheniae sarcocysts has been associated with toxicity and gastroenteritis signs in dogs and rabbits [39,40]. This is due to the production of a peptidic toxin, known as sarcocystin, which has been described for several other Sarcocystis spp. such as S. fayeri infecting horses, S. wapiti and S. sybillensis infecting sika deer, and S. cruzi infecting cattle [1,41,42,43]. Human consumption of raw or insufficiently cooked Sarcocystis-infected camelid meat is expected to have similar toxic effects [23]. Importantly, when infected meat is exposed to freezing, heat or alkaline conditions, the parasite is killed and the toxin inactivated [39,40,44].
5. Pathogenesis of sarcocystosis in camelids
There is a paucity of information on the pathogenic effects of sarcocystosis in camelids. Infections generally appear to be asymptomatic, although a few clinical cases have been reported. In SAC, there are two reports of acute sarcocystosis (Dalmeny’s disease) in alpacas. In one of them, a 6-year-old pregnant alpaca imported to the USA aborted and died shortly after exhibiting lethargic behavior, marked muscle tremors, and respiratory distress. Necropsy revealed numerous cysts in skeletal muscles. Inflammatory disease of the muscles was demonstrated, mostly caused by leukocyte, especially eosinophil, accumulation (eosinophilic myositis), and thought to be associated with old, degenerating S. aucheniae sarcocysts [45].
Another case of sarcocystosis-related myositis was described in an alpaca born and raised in a farm in Australia. The alpaca presented with multiple subcutaneous abscesses. Histologic examination revealed necrotizing and histiocytic myositis and cellulitis, as well as central caseation and numerous microscopic sarcocysts. However, the species of the sarcocyst-forming parasite was not identified [46].
In the case of OWC, experimental oral infections of camels with high doses of S. cameli sporocysts led to acute sarcocystosis with anorexia, lethargy, and anemia. Fatal cases were registered, and hemorrhages in several organs were observed upon necropsy [47,48]. Importantly, these are extreme cases since the numbers of sporocysts utilized (≥ 250,000 per animal) likely largely surpass the amounts ingested in a natural infection with contaminated pastures or water. In a study performed in slaughtered dromedaries of Iran, different degrees of inflammatory responses were observed in tissues surrounding old, degenerating microscopic Sarcocystis sp. sarcocysts, with infiltration of macrophages, lymphocytes, plasma cells, eosinophils, and fibroblasts [49]. In agreement with an inflammatory response, expression of the interleukin-6 gene was significantly increased in Sarcocystis sp. microcyst-infected tongue and diaphragm tissues of dromedaries from Saudi Arabia [50].
Overall, these investigations demonstrate that Sarcocystis spp. may cause considerable disease in SACs and OWCs, although infections are generally subclinical. More research is needed to understand the pathogenesis of both macro- and microscopic sarcocystosis, as well as their influence on musculoskeletal and cardiac function, immunity, well-being, and productivity of infected camelids [5,6].
6. Diagnosis
No commercial or validated diagnostic tools for sarcocystosis applicable to live camelids are so far available, and diagnosis is currently carried out post-mortem. Macroscopic sarcocysts of S. aucheniae or S. cameli can be observed upon visual inspection of skeletal muscles, which is the current procedure used in abattoirs to establish if an animal is infected [51,52]. In the case of microscopic cysts, different detection methods can be applied, including muscle squash, pepsin digestion, trypsin digestion, histopathological examination, and in some cases electron microscopic studies. However, these methods are only employed for research and not for routine examination of camelid carcasses [25,37].
PCR amplification followed by sequencing of different molecular markers such as the 18S rRNA, 28S rRNA and cox-1 genes, and the ITS region, as well as PCR-RFLP of cox-1 or 18S rRNA genes have been used for the identification and molecular characterization of SAC and OWC Sarcocystis spp. [24,25,26,32]. These methods are not practical for diagnostic purposes and are meant to be applied to tissue or cyst samples obtained after necropsy. No nucleotide sequences, on the other hand, are so far available for S. ippeni, thus molecular characterization of this parasite is still pending.
Recently, highly sensitive seminested PCR protocols for S. aucheniae were developed based on the parasite 18S rRNA gene. Using these techniques, it was possible to detect S. aucheniae DNA in the blood of live llamas from Argentina [53,54]. Primers were designed to specifically amplify S. aucheniae DNA, avoiding cross-amplification of DNA of Toxoplasma gondii or Neospora caninum, two closely related coccidians that also infect SAC [55]. In addition, the recent availability of whole-length 18S rRNA sequences of S. masoni allowed to confirm that these PCR detection protocols of S. aucheniae do not cross-react with other SAC-infecting Sarcocystis species [32].
One of these PCRs has a duplex format to simultaneously amplify a segment of the host mitochondrial 16S rRNA gene, which serves as a positive control for successful DNA extraction and amplification. To evaluate the usefulness of this method for diagnosing S. aucheniae infection, it was applied to detect parasite DNA in the blood of 80 live llamas destined for meat consumption. The presence of macroscopic cysts was analyzed postmortem by visual inspection of the carcasses. Results showed no correlation between DNA and sarcocyst detection. The observed PCR-positive/sarcocyst-negative animals could correspond to early infections in which sarcocysts have not yet been formed, while PCR-negative/sarcocyst-positive animals could correspond to older infections in which parasites are confined to muscles and do not circulate in the blood [54]. In any case, S. aucheniae parasitemia appears to be always very low, and thus, the possibility of continuous detection of parasite DNA in blood using more sensitive methods -when available- cannot be overruled.
Antibodies against Sarcocystis spp. antigens evidence the previous exposure of an animal to the parasite. This notion was used to develop a fixed indirect immunofluorescence test (IFAT) using S. aucheniae whole bradyzoites as antigen. Sera of most of the studied llamas (77 %) from the province of Jujuy, Argentina, reacted with the parasites, indicating high seroprevalence of SAC sarcocystosis in this area. The same sera also recognized bradyzoites of S. cruzi, a bovine-infecting Sarcocystis sp., with an even higher prevalence (92.5 %) indicating cross-reactivity at the genus level likely due to S. masoni infections [55].
An indirect ELISA (iELISA) was also established using a 23 kDa immunogenic protein fraction of S. aucheniae sarcocysts as antigen [56]. This assay detected seroprevalence values ranging from 23 to 50% depending on management conditions, in llamas in Argentina. This serological method is less labor-intensive than IFAT and allows a large number of samples to be processed in a time-efficient manner. However, because there is no established gold standard for serological detection of S. aucheniae, the sensitivity and specificity of this iELISA could not be yet determined. In addition, the correlation between serologic detection and the presence of sarcocysts remains to be investigated.
An important constraint of both IFAT and iELISA is the use of parasites or sarcocyst protein fractions as antigens, which might bring restrictions in reproducibility and material accessibility. Thus, identification of immunodominant conserved antigens suitable for the development of serological tests based on recombinant or chemically-synthesized peptides is highly desirable. The recent sequencing of the S. aucheniae sarcocyst transcriptome has provided a pool of attractive targets for the development of diagnostic tools. Indeed, in silico analysis of the transcriptome unraveled an array of proteins predicted to be anchored to the cell membrane through glycosylphosphatidylinositol (GPI) anchors. This type of proteins has been shown to be generally species-specific and immunodominant in other pathogenic protozoa, two requirements for serological tests [57]. These features, as well as their conservation among parasite geographical isolates needs experimental confirmation. In addition, an immunoproteomic approach was recently carried out for S. aucheniae, in which soluble immunoreactive proteins present in sarcocysts were sequenced by mass spectrometry and identified by in silico searches in the transcriptome. Highly antigenic B-cell epitopes were in silico predicted, and those that showed good water solubility and low probability of cross-reactivity with other coccidia were shortlisted for future development of peptide-based serological methods [58].
An important objective of ongoing research is the development of low-cost and reproducible diagnostic methods that can predict the presence of cysts in live camelids as this will result in a significant advance in the control of SAC sarcocystosis.
7. Epidemiology and risk factors
Macroscopic sarcocysts of S. aucheniae have been detected, mostly in intercostal and cervical skeletal muscles, in domestic SACs from Bolivia, Peru and the Northwest of Argentina [24,25,29,51,54]. Additionally, macrocysts produced by this parasite were reported in extra-Andean alpacas from the USA and Australia [45,46,59]. Guanaco and vicuña from Argentina were also shown to act as intermediate hosts of S. aucheniae with formation of macroscopic sarcocysts in their skeletal muscles [25,28,60].
Microscopic sarcocysts, referred to as Sarcocystis sp., S. lamacanis or, more recently, S. masoni, were reported in the cardiac muscle of llamas and guanacos from Argentina, alpacas from Peru and alpacas bred in the extra-Andean countries China, Iran and Australia [25,32,46,61,62,63,64,65]. Microcysts were, in some cases, also observed in other body locations, in addition to myocardium, such as liver, kidney, spleen, lung, tongue, and skeletal muscles [25,32,61,63].
Of the two reported Sarcocystis spp. infecting OWC, there is a single report on the identification of S. ippeni microscopic sarcocysts in skeletal muscles of the esophagus of two dromedaries of Egypt based on microscopy data [3,17]. On the other hand, several reports describe the presence of microscopic and macroscopic sarcocysts of S. cameli or its synonym S. camelicanis in dromedaries from Iran, Iraq, Saudi Arabia, and Egypt. The organs and tissues where cysts were found include esophagus, diaphragm, tongue, heart, and skeletal muscles [17,18,26,27,37,50]. Microcysts identified as Sarcocystis spp. were also reported in the same tissues of dromedaries from Egypt, Jordan, Saudi Arabia, Iran, Iraq, Mongolia and Ethiopia [47,49,52,66,67,68,69,70,71,72]. Unfortunately, nucleotide sequences of taxonomic relevance for S. ippeni are missing as well as species identification in the latter reports. Furthermore, there is scarce information on Sarcocystis spp. infections in Bactrian camels and no reports available for C. ferus [6,17]. Thus, knowledge on the actual distribution of Sarcocystis spp. infecting OWC is far from complete.
A few available studies on the prevalence of camelid sarcocystosis show high to very high values both in SAC and OWC [37,49,51,52,55,56,65,67].
Regarding risk factors, herd management was considered to influence Sarcocystis spp. infection in SAC. In fact, llamas raised in a fenced field, with sanitary controls, and in the absence of pastoral dogs, showed a significantly lower percentage of seropositivity to Sarcocystis spp. than those from the same region raised informally by itinerant shepherds, without sanitary controls and in the presence of shepherd dogs [56].
In addition, age was found to be a risk factor for sarcocystosis caused by S. aucheniae in llamas, S. masoni in alpacas, and Sarcocystis sp. in dromedaries. In all cases, older age was associated with higher abundance of sarcocysts, most likely due to the prolonged exposure of older animals to infective parasitic stages from the environment [49,51,63,73].
8. Parasite biology and host-pathogen interaction
As members of the Apicomplexa phylum, Sarcocystis protozoa spend most of their life cycle as obligate intracelular parasites. They have evolved sophisticated strategies to invade, live within, and egress from host cells, all of which are essential for their survival and propagation [1]. The elucidation of the molecules and mechanisms involved in the interaction of different stages of a pathogen with its host could open the way to the rational design of control tools, such as vaccines and chemotherapeutics. For Sarcocystis spp., this aspect of research lags behind compared to other pathogenic protozoa, due to the scarcity of molecular studies, with only the genome of S. neurona sequenced for this genus so far [75].
In the case of Sarcocystis spp. infecting camelids, this knowledge gap has been partially filled with the sequencing and partial analysis of the S. aucheniae sarcocyst trascriptome, which allowed the identification of molecules relevant for the parasite [57,58]. In the first place, in silico analysis of the transcriptome database allowed to identify the biosynthetic pathway of GPI, a glycolipid essential for host cell invasion. Glycosylphosphatidylinositol is highly abundant in the membranes of pathogenic protozoa, where it is present as an independent surface molecule or serving as an anchor to surface proteins, being the first form exclusive for lower eukaryotes [76]. Blockade of GPI biosynthesis or treatment of cells with phosphatidylinositol (PI)-specific phospholipase C inhibits the in vitro growth of some intracellular pathogenic protozoa, highlighting the vital role of GPIs for these microorganisms [76,77,78]. Moreover, GPIs exert strong immunomodulatory effects during the infection of Plasmodium falciparum, which are detrimental for the host [79].
As mentioned above, searches in the S. aucheniae transcriptome also identified 24 GPI-anchored proteins that, in addition to their potential usefulness as diagnostic candidates, could also be used for vaccine development [57]. Indeed, vaccine formulations based on GPI-anchored proteins of various pathogens such as Trypanosoma cruzi, Babesia canis, and Schistosoma mansoni elicited significant protection upon challenge [80,81,82].
In addition, a number of proteins expressed in S. aucheniae sarcocysts were identified by mass spectrometry and transcriptome searches. This approach provided insight into some of the processes that occur within the sarcocyst. Some of the identified proteins, predicted to be involved in chromosome separation during mitosis and protein synthesis and folding, can be assigned to the actively dividing metrocyte stage. Other proteins, associated with specialized secretory organelles of Apicomplexans (rhoptries, micronemes, and dense granules) or involved in the process of gliding motility are expected to be present in the infective stage of the bradyzoite [58]. The aforementioned studies also identified several proteases that can be hypothesized to play essential roles in S. aucheniae biology based on what is known about their homologs in other micoorganisms [58,83]. Importantly, many of these newly identified parasite proteins represent attractive chemotherapeutic and/or vaccine candidates [58].
Several enzymes involved in respiration (glycolysis, the tricarboxylic acid cycle, and the respiratory chain) and superoxide dismutase, which protects cells from oxidative damage, have been identified among S. aucheniae sarcocyst proteins. Thus, it can be concluded that this parasite undergoes aerobic respiration within the sarcocyst, which requires oxygen diffusion from host capillaries through a porous cyst wall [58].
9. Phylogeny of camelids and camelid-infecting Sarcocystis spp.
Phylogenetic relationships between S. aucheniae, S. masoni, and S. cameli and with respect to other Sarcocystis spp. were investigated by maximum likelihood, using available cytochrome c oxidase subunit I (cox-1) gene sequences (Figure 3). Camelid Sarcocystis sequences segregated into two independent and strongly supported clades: one containing exclusively S. aucheniae and the other consisting of two subclades, one of S. masoni and another of S. cameli sequences.
In the inferred tree, S. masoni and S. cameli represent closely related sister taxons that share a most recent common ancestor. Notably, as mentioned above, S. masoni and S. cameli present the same cyst structure (“9j” type of cyst wall, Table 1), further corroborating the notion of a close relationship between these species. Interestingly, S. aucheniae appears to be evolutionary more ancient than most other Sarcocystis spp. infecting mammals studied so far.
We then contrasted these observations with the evolutionary history of the Sarcocystis camelid intermediate and proposed definitive hosts (Figure 4). The ancestral camelid, Poebrotherium wilsoni, which resembles a guanaco, originated in North America in the middle Oligocene (25-30 million years ago, mya). Later, in the early Miocene, a number of changes characteristic for extant camels appeared, such as plantar cushions in each leg, the loss of upper incisive tooth and the conversion of one of them into a canine, as well as a big depression in the facial part of the maxillary bone to contain the complex lip musculature. During the Miocene, many different groups of camelids evolved, most of which became eventually extinct. One of these groups gave rise to two tribes: Camellini and Lamini, representatives of which migrated through the Bering land bridge to Eurasia and through the Panama land bridge to South America, respectively. The Camellini ancestors diverged to finally give rise to the three Camelus species that are currently living, around 4.4 mya. Lamini ancestors that arrived in SA belonged to the genus Hemiauchenia, which gave rise to the genera Paleolama, Lama and Vicugna. By the end of the Pleistocene, around 12,000 years ago, Hemiauchenia and Paleolama had become extinct, while Lama and Vicugna flourished, particularly in the arid and semiarid regions of the Andes, thanks to their adaptations to thermal stress, dehydration, and hypoxia, as found in high altitudes [86]. It is accepted that these genera gave rise to two species each: Lama to L. glama and L. guanicoe, and Vicugna to V. pacos and V. vicugna. However, the four species are interfertile and mitochondrial genome data evidence the occurrence of hybridization among them during the process of domestication [87].
Canids (Canidae family) first appeared in North America in the late Eocene (40 mya), evolving from a group of archaic carnivorous mammals, and their evolution is characterized by successive radiations and the occupation of highly diverse niches. In the late Miocene, members of the Caninae subfamily crossed to Eurasia through a land bridge at Bering strait, diversified, and finally gave rise to the modern canids of the Old World, including Canis familiaris [88]. In turn, after the Panama isthmus was formed in the Pleistocene, canids arrived in South America, around 3.9 to 3.5 mya, and diversified from a single ancestor, giving rise to numerous species, most of which but four Lycalopex spp. became extinct [89].
During the Holocene, around 10 kya, humans accompanied by dogs (Canis familaris) crossed the Bering bridge and settled first in North and then in South America. The Mexican chihuahua and the Peruvian “naked dog” derive from these pre-Columbine lineages. Finally, much later, in the XV century, European colonizers brought with them other Canis familiaris breeds that spread throughout the Americas [90].
Analyzing these data, two conclusions can be drawn. From the late Oligocene to the early Miocene periods both ancestral camelids and canids were present in North America, before the diversification into and migration of the Camellini and Lamini tribes to Eurasia and South America, respectively (Figure 4). Considering the close relation between S. masoni and S. cameli, the most recent common ancestor of these Sarcocystis spp. must have parasitized a camelid ancestor in the Miocene, using ancient canids as definitive hosts. Thus, the diversification of the ancestral camelid intermediate host into OWCs and SACs resulted in the diversification of an ancestral Sarcocystis species into S. camili and S. masoni, respectively. This coevolutionary pattern is commonly referred to as parasite-host co-speciation. Second, considering that Canis familiaris became available as definitive host for SAC Sarcocystis spp. only around 5 kya, other canids must have previously fulfilled this role, after the arrival of camelids to South America 3.9 to 3.5 mya and, later, a host shift to C. familiaris must have occurred. Extant autochthonous canids whose habitats coincide with those of SAC in the Andean regions include the Culpeo fox (Lycalopex culpaeus) and the South American gray fox (Lycalopex griseus), and with more limited distribution, the Sechuran fox or Peruvian desert fox (Lycalopex sechurae) and the Darwin´s fox (Lycalopex fulvipes) [89]. A role for these canids as definitive hosts of S. aucheniae and/or S. masoni, either by predating or -more likely scavenging on dead SAC carcasses- can be hypothesized but awaits experimental confirmation.
10. Conclusions and perspectives
Sarcocystosis is an old and well-known problem for domestic SAC and OWC meat commercialization. Prevalence in herds is high and tissues are heavily infected with cysts. Although infections are generally subclinical, ocassional pathogenicity is observed, while the effects of sarcocystosis on the well-being and fitness of domestic and wild camelids is unknown. Diagnostic methods that can be reliably applied in live animals are not available, and no tools or therapies to control these infections have so far been designed. Currently, dogs appear to be the main definitive hosts for at least three of the Sarcocystis spp. infecting camelids. However, the close relationship between camelid shepherds and dogs, resulting in their frequent feeding with raw camelid meat and the presence of feral and free-roaming dogs in camelid breeding areas make it difficult to break the parasite life cycle through parasite control in dogs.
Sarcocystis spp. have developed sophisticated adaptations to perpetuate themselves in their hosts along millions of years of evolution. Recent research has started to fill the gap of knowledge on molecules that are essential for host-pathogen interactions of camelid Sarcocystis spp. parasites and that constitute attractive targets for the development of vaccines and therapeutic interventions. These research efforts, as well as increased knowledge on the epidemiology of these parasites and novel diagnostic tests will undoubtely result in a breakthrough for camelid breeding communities.
Author Contributions
Conceptualization: MFC, SG, JRO, MACF, LS. Methodology: SNW, LOM, XBM, LS. Formal analysis: MFC, LS. Funding acquisition: MFC, JRO, MACF. Writing: original draft preparation: SNW. Writing review and editing. MFC, SG, LS. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Programa Nacional de Investigación Científica y Estudios Avanzados (proCiencia) del Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica (CONCyTEC), Peru (Grant PE501079385-2022) and the Instituto Nacional de Tecnologia Agropecuaria (INTA), Argentina (Grant PD 114).
Data Availability Statement
Publicly available datasets were analyzed in this study. Sequences included in the phylogenetic trees can be found at: https://www.ncbi.nlm.nih.gov/genbank/, using the accession numbers included in the trees.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Decker-Franco, C.; Schnittger, L.; Florin-Christensen, M., Sarcocystis, in: Parasitic Protozoa of Farm Animals and Pets. 2018a. Springer International Publishing, Cham, pp. 103–124. [CrossRef]
- Fayer, R. Sarcocystis spp. in human infections. Clin Microbiol Rev 2004, 17, 894-902. [CrossRef]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of animals and humans, second edition, 2015. [CrossRef]
- Poulsen, C.S.; Stensvold, C.R. Current status of epidemiology and diagnosis of human sarcocystosis. J Clin Microbiol. 2014, 52, 3524-30. [CrossRef]
- Saeed, M.A.; Rashid, M.H.; Vaughan, J.; Jabbar, A. Sarcocystosis in South American camelids: The state of play revisited. Parasit Vect 2018, 11, 146. [CrossRef]
- Saeed, M.A.; Vaughan, J.L.; Jabbar, A. An update on sarcocystosis in one-humped camels (Camelus dromedarius). Parasitol 2018, 145, 1367–1377. [CrossRef]
- Giuliano, S. M.; Reategui Ordoñez, J.; Barriga Marcapuda X.; Florin-Christensen M. Situación actual de la calidad de carne de camélidos sudamericanos (llama y alpaca) en Argentina y Perú y su relación con la infestación con Sarcocystis aucheniae. Proceedings of the XXIII Reunión Nacional de la Asociación Boliviana de Producción Animal – ABOPA 2023. https://www.cifumss.agro.bo/abopa/index.html.
- Fowler, M.E. Camelids are not ruminants. Zoo Wild Animal Med 2008, 375–85. [CrossRef]
- FAOSTAT. https://www.fao.org/faostat/en/#data/QCL.
- Midagri. https://www.midagri.gob.pe/portal/datero/40-sector-agrario/situacion-de-las-actividades-de-crianza-y-producci/298-camelidos-sudamericanos?start=1.
- Zarrin, M.; Riveros, J.L.; Ahmadpour, A.; De Almeida, A.M.; Konuspayeva, G.; Vargas-Bello-Pérez, E.; Faye, B.; Hernández-Castellano, L.E.. Camelids: new players in the international animal production context, Trop Anim Health Prod 2020, 52, 902-913. [CrossRef]
- Fowler, M.E. Husbandry and diseases of camelids. Rev Sci Tech 1996, 15, 155-69. [CrossRef]
- Yacobaccio, H.D. The domestication of South American camelids: a review. Anim Front 2021, 11, 43-51. [CrossRef]
- Mamani-Linares, L.W.; Gallo, C.B. Meat quality attributes of the Longissimus lumborum muscle of the Kh'ara genotype of llama (Lama glama) reared extensively in northern Chile. Meat Sci 2013, 94, 89–94.
- Mamani-Linares, L.W.; Gallo, C.B. Meat quality, proximate composition and muscle fatty acid profile of young llamas (Lama glama) supplemented with hay or concentrate during the dry season. Meat Sci 2014, 96, 394–399. [CrossRef]
- Kadim, I.T.; Al-Amri, I. S.; Alkindi, A.Y.; Quazi Haq, M. I. Nutritional values and health benefits of dromedary camel meat. Anim Front 2022, 12, 61–70. [CrossRef]
- Dubey, J.P.; Hilali, M.; Van Wilpe, E.; Calero-Bernal, R.; Verma, S.K.; Abbas, I.E. A review of sarcocystosis in camels and redescription of Sarcocystis cameli and Sarcocystis ippeni sarcocysts from the one-humped camel (Camelus dromedarius). Parasitology 2015, 142, 1481–1492. [CrossRef]
- Dubey, J.P.; A’aji, N.N.; Mowery, J.D.; Verma, S.K.; Calero-Bernal, R. Identification of Macroscopic Sarcocysts of Sarcocystis cameli from One-Humped Camel (Camelus dromedarius) in Iraq. J Parasitol 2017, 103, 168–169. [CrossRef]
- Brumpt, E. (1913). Precis de Parasitologie, 2nd Edn. Masson et cie, Paris, France.
- Quiroga, D.; Lombardero, O.; Zorrilla, R.,. Sarcocystis tilopodi N sp. en guanacos (Lama guanicoe) de la Repdblica Argentina. Gac Vet 1969, 31, 67--70.
- Gorman, T. R.; Alcaino, H. A.; Munoz, H.; Cunazza, C. Sarcocystis sp. in guanaco (Lama guanicoe) and effect of temperature on its viability. Vet Parasitol 1984, 15, 95–101.
- Leguía, G.; Casas, E. Enfermedades parasitarias y atlas parasitológico de camélidos sudamericanos 1999, Primera edn. De Mar, Lima, Perú.
- Leguía, G. The epidemiology and economic impact of llama parasites. Parasitol Today 1991, 7, 54–56. [CrossRef]
- Carletti, T.; Martin, M.; Romero, S.; Morrison, D.A.; Marcoppido, G.; Florin-Christensen, M.; Schnittger, L. Molecular identification of Sarcocystis aucheniae as the macrocyst-forming parasite of llamas. Vet Parasitol 2013, 198, 396–400. [CrossRef]
- Moré, G.; Regensburger, C.; Gos, M.L.; Pardini, L.; Verma, S.K.; Ctibor, J.; Serrano-Martínez, M.E.; Dubey, J.P.; Venturini, M.C. Sarcocystis masoni, n. sp. (Apicomplexa: Sarcocystidae), and redescription of Sarcocystis aucheniae from llama (Lama glama), guanaco (Lama guanicoe) and alpaca (Vicugna pacos). Parasitology 2016, 143, 617–626. [CrossRef]
- Metwally, D.M.; Al-Otaibi, T.T.; Al-Turaiki, I.M.; El-Khadragy, M.F.; Alajmi, R.A. Identification of Sarcocystis spp. in one-humped camels (Camelus dromedarius) from Riyadh and Dammam, Saudi Arabia, via histological and phylogenetic approaches. Animals 2020, 10, 1108. [CrossRef]
- Motamedi, G.R.; Dalimi, A.; Nouri, A.; Aghaeipour, K. Ultrastructural and molecular characterization of Sarcocystis isolated from camel (Camelus dromedarius) in Iran. Parasitol Res 2011, 108, 949-954. [CrossRef]
- Regensburger, C.; Gos, M.L.; Ctibor, J.; Moré, G. Morphological and molecular characteristics of Sarcocystis aucheniae isiolated from meat of guanaco (Lama guanicoe). J Food Qual Haz Cont 2015, 2, 118-121.
- Schnieder, T.; Kaup, F.J.; Drommer, W.; Thiel, W.; Rommel, M. [Fine structure and development of Sarcocystis aucheniae in llamas]. Z Parasitenkd 1984, 70, 451–458. [CrossRef]
- Cornejo, R.B.; Chávez, A.V.; Leyva, V.V.; Falcón, N.P.; Panez S.L.; Ticona D.S. Relationship between the size of macrocysts of Sarcocysistis aucheniae and its viability in Canis familiaris. Rev Inv Vet Perú 2007, 18, 76-83.
- Zacarías, F.S.; Sam, R. T.; Ramos, D.D.; Lucas. O.A.; Lucas, J.L. Techniques for the isolation and purification of Sarcocystis aucheniae oocysts from small intestine of experimentally infected dogs. Rev Inv Vet Perú 2013., 24, 396-403.
- Wu, Z.; Sun, J.; Hu, J.; Song, J.; Deng, S;, Zhu, N.; Yang, Y.; Tao, J. Morphological and molecular characterization, and demonstration of a definitive host, for Sarcocystis masoni from an alpaca (Vicugna pacos) in China. Biology 2022, 11, 1016. [CrossRef]
- Hilali, M.; El Sayed Imam, B.; Hassan, A. The Endogenous Stages of Sarcocystis cameli (Mason, 1910) Vet Parasitol 1982, 11, 127—129.
- Hilali, M.; Fatani, A.; Al-Atiya, S. Isolation of tissue cysts of Toxoplasma, Isospora, Hammondia and Sarcocystis from camel (Camelus dromedarius) meat in Saudi Arabia. Vet Parasitol 1995, 58, 353–356.
- Hilali, M.; Mohamed A. The dog (Canis familiaris) as the final host of Sarcocystis cameli (Mason, 1910). Tropenmed Parasitol 1982, 31, 213-214.
- Hilali, M.; Nassar, A.M.; EI-Ghaysh A.,. Camel (Camelus dromedarius) and sheep (Ovis aries) meat as a source of dog infection with some coccidian parasites. Vet Parasitol 1992, 43, 37-43.
- Abdel-Ghaffar, F.; Mehlhorn, H.; Bashtar, A.R.; Al-Rasheid, K.; Sakran, T.; El-Fayoumi, H. Life cycle of Sarcocystis camelicanis infecting the camel (Camelus dromedarius) and the dog (Canis familiaris), light and electron microscopic study. Parasitol Res 2009, 106, 189–195. [CrossRef]
- Ishag, M.Y.; Majid, A.; Magzoub, A. Isolation of a new Sarcocystis species from Sudanese camels (Camelus dromedarius). Int J Trop Med 2006, 1, 167–169.
- Godoy, Z.; Vilca, L.; Gonzáles, Z.; Leyva, V.; Sam, T. Saneamiento y detoxificación de carne de llama (Lama glama) infectada con Sarcocystis aucheniae mediante cocción, horneado, fritura y congelado. Rev Invest Vet Perú 2007, 18, 51–56.
- Vilca, M.; Durán, J.; Ramos, D.; Lucas, J. Saneamiento y eliminación de la toxicidad de carne de alpaca (Vicugna pacos) con sarcocistiosis mediante ahu mado y curado. Rev Invest Vet Perú 2013, 24, 537–54.
- Saito, M.; Taguchi, K.; Shibata, Y.; Kobayashi, T.; Shimura, K.; Itagaki, H. Toxicity and properties of the extract from Sarcocystis cruzi cysts. J Vet Med Sci 1995, 57, 1049-51. [CrossRef]
- Irikura, D.; Saito, M.; Sugita-Konishi, Y.; Ohnishi, T.; Sugiyama, K.I. et al. Characterization of Sarcocystis fayeri's actin-depolymerizing factor as a toxin that causes diarrhea. Genes Cells 2017, 22, 25-835. [CrossRef]
- Kamata Y, Saito M, Irikura D, Yahata Y, Ohnishi T, Bessho T, Inui T, Watanabe M, Sugita-Konishi Y. A toxin isolated from Sarcocystis fayeri in raw horsemeat may be responsible for food poisoning. J Food Prot 2014, 77:814-9. [CrossRef]
- Honda, M.; Sawaya; M.; Taira, K.; Yamazaki, A.; Kamata, Y.; Shimizu, H.; Kobayashi, N.; Sakata, R.; Asakura, H.; Sugita-Konishi, Y. Effects of temperature, pH and curing on the viability of Sarcocystis, a Japanese sika deer (Cervus Nippon centralis) parasite, and the inactivation of their diarrheal toxin. J Vet Med Sci 2018, 80, 1337-1344. [CrossRef]
- La Perle, K.M.D.D.; Silveria, F.; Anderson, D.E.; Blomme, E.A.G.G.. Dalmeny disease in an alpaca (Lama pacos): Sarcocystosis, eosinophilic myositis and abortion. J Comp Pathol 1999, 121, 287–293. [CrossRef]
- Gabor, M.; Gabor, L.J.; Srivastava, M.; Booth, M.; Reece, R. Chronic myositis in an australian alpaca (Lama pacos) associated with sarcocystis spp. J Vet Diagn Invest 2010, 22, 966-9 . [CrossRef]
- Fatani, A.; Elsebaie, A.; Hilali, M. Clinical and haematobiochemical changes in camels (Camelus dromedarius) experimentally inoculated with Sarcocystis cameli. J Camel Pract Res 1996, 3, 11–15.
- Manal, Y.I.; El Amin, E.; Osman, A (2001) Camels experimentally infected with Sarcocystis. Sudan J Vet Res 2014, 17, 27–33.
- Valinezhad, A.; Oryan, A.; Ahmadi, N. Sarcocystis and its complications in camels (Camelus dromedarius) of Eastern Provinces of Iran. Korean J Parasitol 2008, 46, 229–234. [CrossRef]
- Metwally, D.M.; Al-Otaibi, T.T.; Semlali, A.; Alajmi, R.A. Sarcocystis camelicanis increases interleukin (IL)-6 expression in one-humped camels (Camelus dromedarius) from Riyadh and Al Qassim, Saudi Arabia. Biosci Rep 2021 41, BSR20203140. [CrossRef]
- Rooney, A.L.; Limon, G.; Vides, H.; Cortez, A.; Guitian, J. Sarcocystis spp. in llamas (Lama glama) in Southern Bolivia: A cross sectional study of the prevalence, risk factors and loss in income caused by carcass downgrades. Prev Vet Med 2014, 116, 296-304. [CrossRef]
- Gareh, A.; Soliman, M.; Saleh, A.A.; El-Gohary, F.A.; El-Sherbiny, H.M.M.; Mohamed, R.H.; Elmahallawy, E.K.; Kotb Elmahallawy, E. Epidemiological and histopathological investigation of Sarcocystis spp. in slaughtered dromedary camels (Camelus dromedarius) in Egypt. Vet Sci 2020, 7, 162. [CrossRef]
- Martin, M.; Decker Franco, C.; Romero, S.; Carletti, T.; Schnittger, L.; Florin-Christensen, M. Molecular detection of Sarcocystis aucheniae in the blood of llamas from Argentina. Rev Argent Microbiol 2016, 48, 200–205. [CrossRef]
- Decker Franco, C.; Romero, S.; Ferrari, A.; Schnittger, L.; Florin-Christensen, M. Detection of Sarcocystis aucheniae in blood of llama using a duplex semi-nested PCR assay and its association with cyst infestation. Heliyon. 2018, 4(11), e00928. [CrossRef]
- Moré, G.; Pardini, L.; Basso, W.; Marín, R.; Bacigalupe, D.; Auad, G.; Venturini, L.; Venturini, M.C. Seroprevalence of Neospora caninum, Toxoplasma gondii and Sarcocystis sp. in llamas (Lama glama) from Jujuy, Argentina. Vet Parasitol 2008, 155, 158-60. [CrossRef]
- Romero, S.; Carletti, T.; Decker Franco, C.; Moré, G.; Schnittger, L.; Florin-Christensen, M. Seropositivity to Sarcocystis infection of llamas correlates with breeding practices. Vet Parasitol Reg Stud Reports 2017, 10, 65-70. [CrossRef]
- Decker Franco, C.; Wieser, S.N.; Soria, M.; de Alba, P.; Florin-Christensen, M.; Schnittger, L. In silico identification of immunotherapeutic and diagnostic targets in the glycosylphosphatidylinositol metabolism of the coccidian Sarcocystis aucheniae. Transbound Emerg Dis. 2020, 67 Suppl 2, 165-174. [CrossRef]
- Wieser, S.N.; Decker-Franco, C.; de Alba, P.; Romero, S.; Ferrari, A.; Schnittger, L.; Florin-Christensen, M. Discovery of antigens and cellular mechanisms in the protozoan parasite Sarcocystis aucheniae using immunoproteomics. Parasitologia 2023, 3, 349-363. [CrossRef]
- Holmdahl, O.J.M.; Morrison, D.A.; Ellis, J.T.; Huong, L.T.T. Evolution of ruminant Sarcocystis (Sporozoa) parasites based on small subunit rDNA sequences. Mol Phylogenet Evol 1999, 11, 27-37. [CrossRef]
- Wieser, S.N.; Cafrune, M.W.; Romero, S.R.; Schnittger, L.; Florin-Christensen, M. Molecular identification of Sarcocystis aucheniae infesting Vicugna vicugna,.2023, submitted.
- Beldomenico, P.M.; Uhart, M.M.; Beldomenico, P.M.; Uhart, M.; Bono, M.F.; Marull, C.; Baldi, R.; Peralta, J.L. Internal Parasites of free-ranging guanacos from Patagonia. Vet Parasitol 2003, 118, 71–77. [CrossRef]
- Lucas, J.R.; Barrios-Arpi, M.; Rodríguez, J.; Balcázarnakamatsu, S.; Zarria, J.; Namiyama, G.; Taniwaki, N.; Gonzales-Viera, O. Ultrastructural description of Sarcocystis sp. in cardiac muscle of naturally infected alpacas (Vicugna pacos). Iran J Parasitol 2019, 14, 174–179.
- Jiang, N.; Xin, S.; Zhu, N.; Yang, L.; Huang, W.; Hu, J.; Zhu, X.; Yang, Y. First report of Sarcocystis Masoni in a captive alpaca (Vicugna pacos) from China. Front Vet Sci 2021, 8, 759252. [CrossRef]
- Fernandez-F, F.; Gutiérrez-A, R.; Pacheco-S, V.; Chirinos-T, J.; Lombardo, D.M.; Olivera, L.V.M.; Bernabe-Ortiz, J.C.; López-Casaperalta, P. Determination of Sarcocystis lamacanis microcysts in the cardiac muscle of alpacas (Vicugna pacos) and their correlation with troponin CTnI. A Study performed in the High Andean region of Southern Peru. Vet Anim Sci 2022, 18, 100270. [CrossRef]
- Rodríguez, A.; Quispe-Solano, M.; Rodríguez, J.L.; Lucas, J.R. The occurrence of Sarcocystis spp. in the myocardium of alpacas (Vicugna pacos) with associated risk factors in the Peruvian Andes. Trop Anim Health Prod 2023, 55, 66. [CrossRef]
- Latif, B.M.A.; Al-Delemi, J.K.; Mohammed, B.S.; Al-Bayati, S.M.; Al-Amiry, A.M. Prevalence of Sarcocystis spp. in meat-producing animals in Iraq. Vet Parasitol 1999, 84, 85–90. [CrossRef]
- Woldemeskel, M.; Gumi, B. Prevalence of Sarcocysts in one-humped camel (Camelus dromedarius) from Southern Ethiopia. J Vet Med 2001, 48, 223–226. [CrossRef]
- Fukuyo, M.; Battsetseg, G.; Byambaa, B. Prevalence of Sarcocystis Infection in Meat-Producing Animals in Mongolia. Southeast Asian J Trop Med Pub Health 2002, 33, 490–495.
- Shekarforoush, S.S.; Shakerian, A.; Hasanpoor, M. M. Prevalence of Sarcocystis in slaughtered one-humped camels (Camelus dromedarius) in Iran. Trop Anim Health Prod 2006, 38, 301–303. [CrossRef]
- Hamidinejat, H.; Hekmatimoghaddam, S.; Jafari, H.; Sazmand, A.; Haddad Molayan, P.; Derakhshan, L.; Mirabdollahi, S. Prevalence and distribution patterns of Sarcocystis in camels (Camelus dromedarius) in Yazd Province, Iran. J Parasit Dis 2013, 37, 163–165. [CrossRef]
- Omer, S.A.; Alzuraiq, A.A.; Mohammed, O.B. Prevalence and molecular detection of Sarcocystis spp. infection in the dromedary camel (Camelus dromedarius) in Riyadh City, Saudi Arabia. Biomed Res 2017, 28, 4962–4965.
- Al-Ani, F.K.; Amr, Z. Sarcocystis Spp Prevalence in Camel Meat in Jordan . Dairy and Vet Sci J. 2017, 4, 555643. [CrossRef]
- Castro, E.; Sam, R.; López, T.; González, A.; Silva, M. Evaluaciòn de la edad como factor de riesto de seropositividad a Sarcocystis sp. en alpacas. Rev Inv Vet Perú 2004, 15, 83–86.
- Silva Rochefort, B.; Root-Bernstein, M. History of canids in Chile and impacts on prey adaptations. Ecol Evol 2021, 11, 9892-9903. [CrossRef]
- Blazejewski, T.; Nursimulu, N.; Pszenny, V.; Dangoudoubiyam, S.; Namasivayam, S.; Chiasson, M.A.; Chessman, K.; Tonkin, M.; Swapna, L.S.; Hung, S.S.; et al. Systems-Based Analysis of the Sarcocystis neurona neurona genome identifies pathways that contribute to a heteroxenous life cycle. mBio 2015, 6. [CrossRef]
- Rodríguez, A.E.; Couto, A.; Echaide, I.; Schnittger, L.; Florin-Christensen, M. Babesia bovis contains an abundant parasite-specific protein-free glycerophosphatidylinositol and the genes predicted for its assembly. Vet Parasitol 2010, 167, 227-35. [CrossRef]
- Delorenzi, M.; Sexton, A.; Shams-Eldin, H.; Schwarz, R. T.; Speed, T.; Schofield, L. Genes for glycosylphosphatidylinositol toxin biosynthesis in Plasmodium falciparum. Infect Immun 2002, 70, 4510–4522. [CrossRef]
- Rodriguez, A.E.; Florin-Christensen, M.; Flores, D.A.; Echaide, I.; Suarez, C.E.; Schnittger, L. The glycosylphosphatidylinositol-anchored protein repertoire of Babesia bovis and its significance for erythrocyte invasion. Ticks Tick Borne Dis 2014, 5, 343-8. [CrossRef]
- Deroost, K.; Pham, T. T.; Opdenakker, G.; Van den Steen, P. E. The immunological balance between host and parasite in malaria. FEMS Microbiol Rev 2016, 40, 208–257. [CrossRef]
- Martins, V.P.; Pinheiro; C.S.; Figueiredo, B.C.; Assis, N.R.; Morais, S.B.; Caliari, M.V.; Azevedo, V.; Castro-Borges, W.; Wilson, R.A.; Oliveira, S.C. Vaccination with enzymatically cleaved GPI-anchored proteins from Schistosoma mansoni induces protection against challenge infection. Clin Dev Immunol. 2012, 962538. [CrossRef]
- Hegazy-Hassan, W.; Zepeda-Escobar, J. A.; Ochoa-García, L.; et al. TcVac1 vaccine delivery by intradermal elec troporation enhances vaccine induced immune protection against Trypanosoma cruzi infection in mice. Vaccine 2018, 37, 248–257. [CrossRef]
- Moubri, K.; Kleuskens, J.; Van de Crommert, J.; et al. Discovery of a recombinant Babesia canis supernatant antigen that protects dogs against virulent challenge infection. Vet Parasitol 2018, 249, 21–29. [CrossRef]
- Florin-Christensen, M.; Sojka, D.; Ganzinelli, S.; Šnebergerová, P.; Suarez, C.E.; Schnittger, L. Degrade to survive: the intricate world of piroplasmid proteases. Trends Parasitol 2023, 39, 532-546. [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 1980, 16:111-120.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022-3027. [CrossRef]
- Wu, H.; Guang, X.; Al-Fageeh, M. B.; Cao, J.; Pan, S. Camelid genomes reveal evolution and adaptation to desert environments. Nat Commun 2014, 21, 5188. [CrossRef]
- Diaz-Maroto, P.; Rey-Iglesia, A.; Cartajena, I.; Núñez L.; et al. Ancient DNA reveals the lost domestication history of South American camelids in Northern Chile and across the Andes. Elife 2021 10, e63390. [CrossRef]
- Wang, X.; Tedford, R.H.; Van Valkenburgh, B.; Wayne, R.K. Evolutionary history, molecular systematics and evolutionary ecology of Canidae. In: The Biology of Conservation of World Canids (D.W. Macdonald and C. Sillero-Zubrin, eds.) 2004, Chapter 2, pp. 39-54. Oxford University Press, Oxford.
- Chavez, D.E.; Gronau, I.; Hains, T.; Dikow, R.B. et al. Comparative genomics uncovers the evolutionary history, demography, and molecular adaptations of South American canids. Proc Natl Acad Sci U S A. 2022, 119, e2205986119. [CrossRef]
- van Asch, B.; Zhang, A.B.; Oskarsson, M.C.; Klütsch, C.F.; Amorim, A.; Savolainen, P. Pre-Columbian origins of Native American dog breeds, with only limited replacement by European dogs, confirmed by mtDNA analysis. Proc Biol Sci 2013, 280, 20131142. [CrossRef]
Figure 1.
Life cycle of Sarcocystis. A typical life cycle of Sarcocystis species is shown, with camelids and canids as intermediate and definitive hosts, respectively.
Figure 1.
Life cycle of Sarcocystis. A typical life cycle of Sarcocystis species is shown, with camelids and canids as intermediate and definitive hosts, respectively.
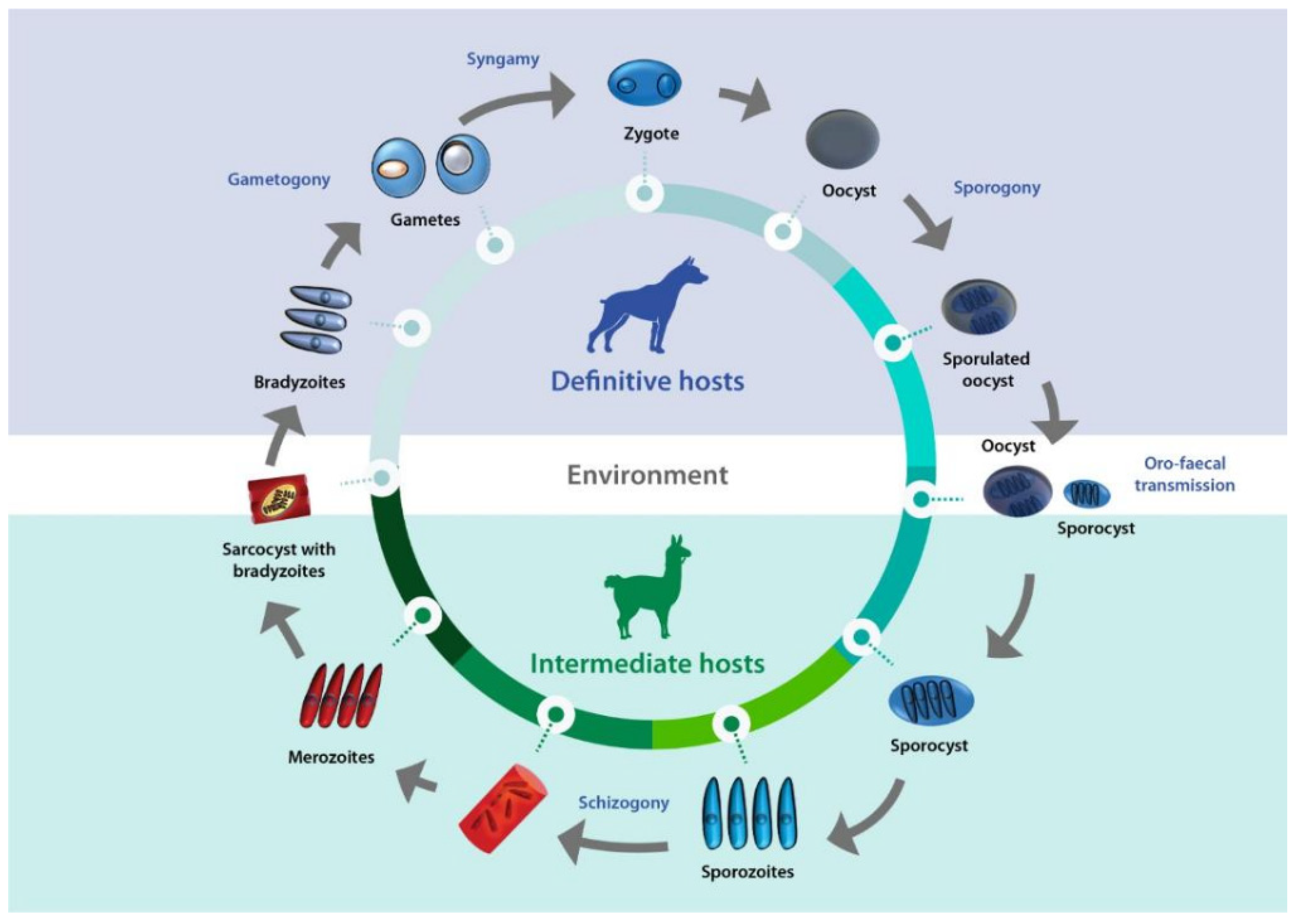
Figure 2.
Morphology of sarcocysts and bradyzoites in S. aucheniae. (a and b) macroscopic sarcocysts in llama (a) and alpaca (b) skeletal muscle; (c) hematoxylin-eosin-dyed cross-section of alpaca skeletal muscle with two macroscopic sarcocysts; in which zoites are located to the periphery and the center is empty (100x); (d and e) details of hematoxylin-eosin-dyed section of a macroscopic sarcocyst showing the morphology of the cell wall (d) and compartments with thousands of banana-shaped bradyzoites (e) (400x); (f) bradyzoites observed in the cyst stained with Giemsa (1000x).
Figure 2.
Morphology of sarcocysts and bradyzoites in S. aucheniae. (a and b) macroscopic sarcocysts in llama (a) and alpaca (b) skeletal muscle; (c) hematoxylin-eosin-dyed cross-section of alpaca skeletal muscle with two macroscopic sarcocysts; in which zoites are located to the periphery and the center is empty (100x); (d and e) details of hematoxylin-eosin-dyed section of a macroscopic sarcocyst showing the morphology of the cell wall (d) and compartments with thousands of banana-shaped bradyzoites (e) (400x); (f) bradyzoites observed in the cyst stained with Giemsa (1000x).
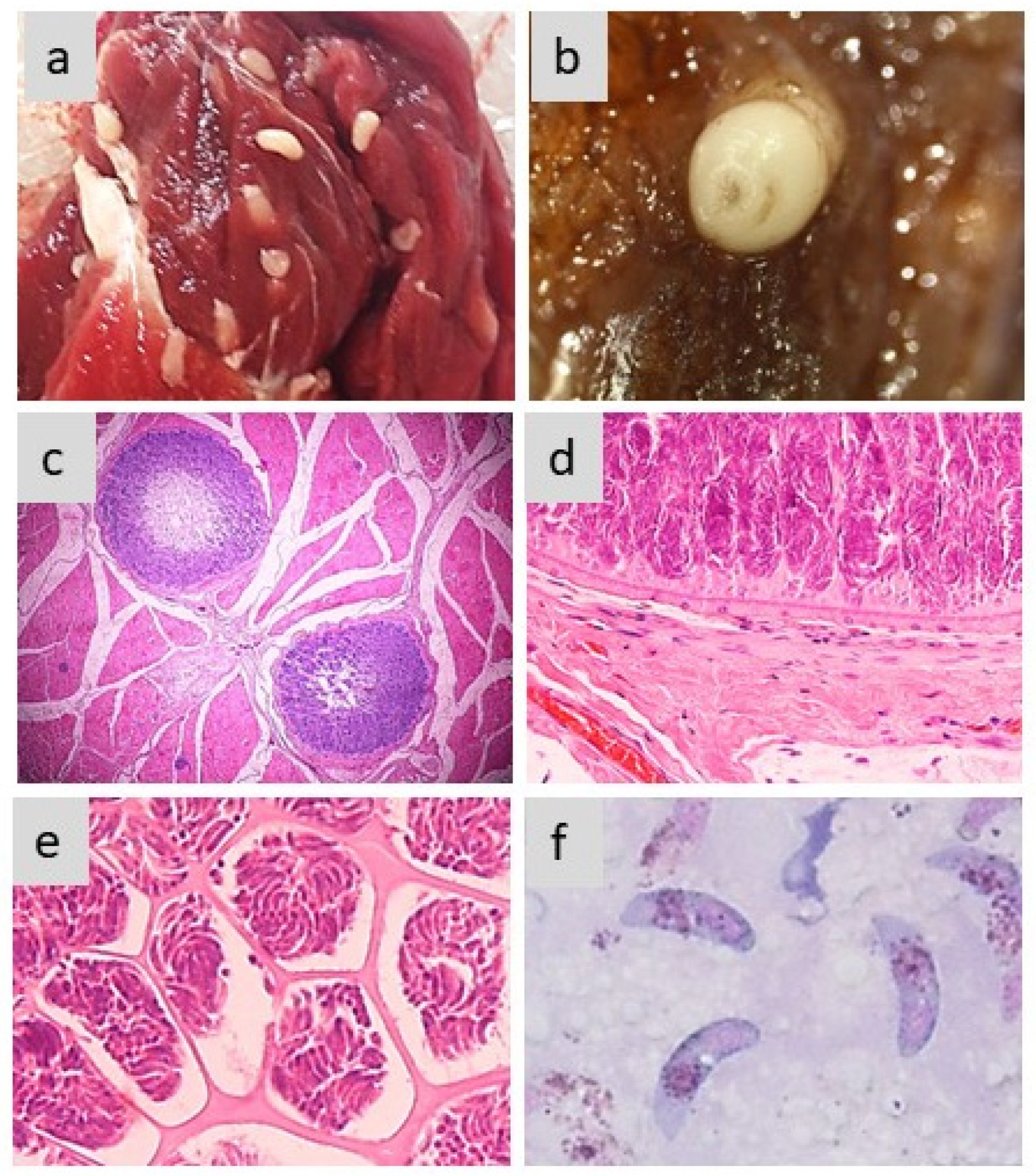
Figure 3.
Phylogenetic tree of Sarcocystis spp. cox-1 sequences. The evolutionary history was inferred by using the maximum likelihood applying the Kimura 2-parameter model [84]. The analysis involved 68 nucleotide sequences and comprised a total of 946 positions in the final dataset. Bootstrap values are shown close to the branches. Neospora caninum cox-1 gene sequence was used as outgroup. The analysis was carried out using MEGA11 [85].
Figure 3.
Phylogenetic tree of Sarcocystis spp. cox-1 sequences. The evolutionary history was inferred by using the maximum likelihood applying the Kimura 2-parameter model [84]. The analysis involved 68 nucleotide sequences and comprised a total of 946 positions in the final dataset. Bootstrap values are shown close to the branches. Neospora caninum cox-1 gene sequence was used as outgroup. The analysis was carried out using MEGA11 [85].
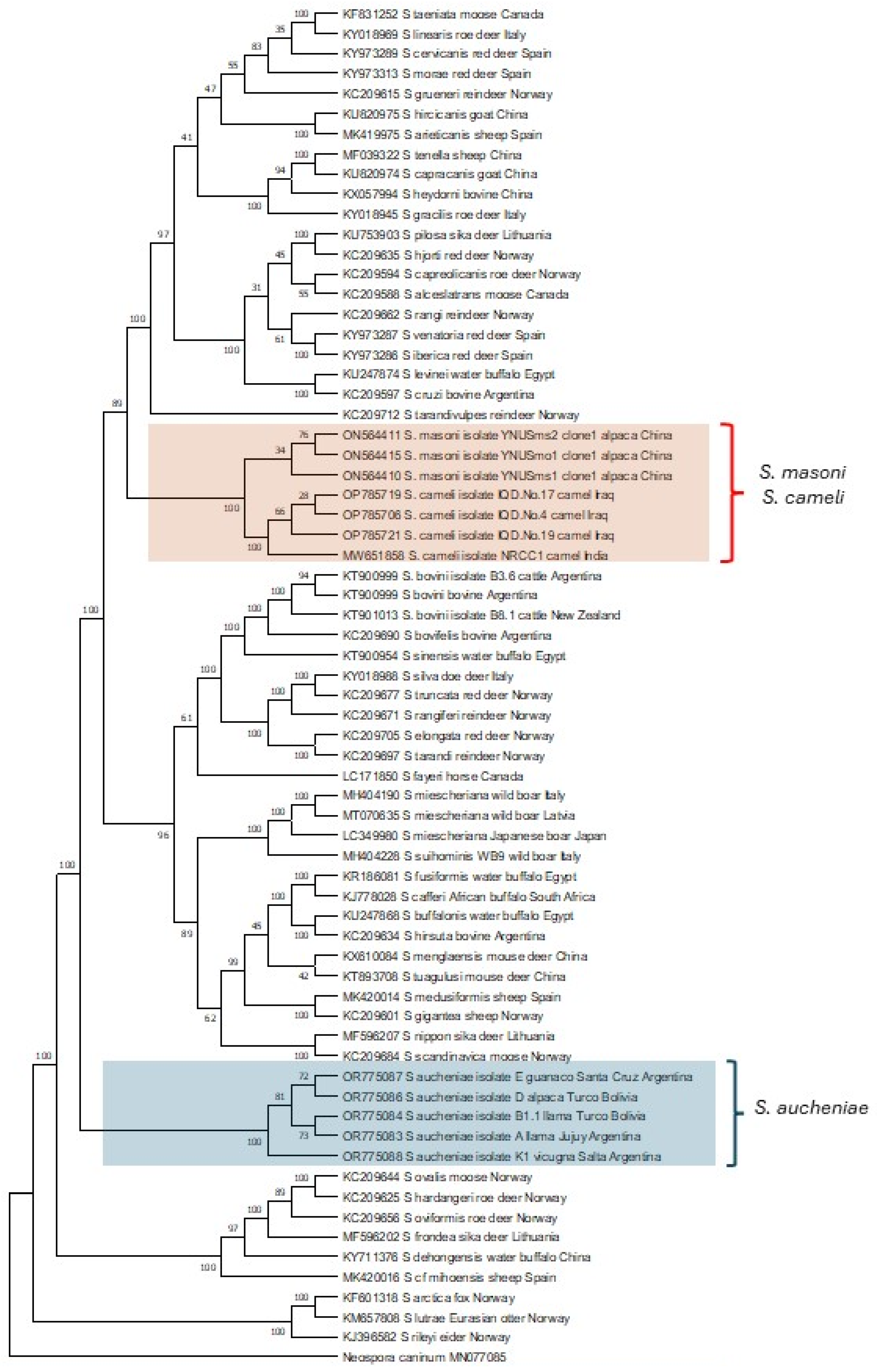
Figure 4.
Schematic representation of the evolutionary history of camelids and canids. The likely time period when the ancestor of camelid Sarcocystis spp. evolved is shown.
Figure 4.
Schematic representation of the evolutionary history of camelids and canids. The likely time period when the ancestor of camelid Sarcocystis spp. evolved is shown.
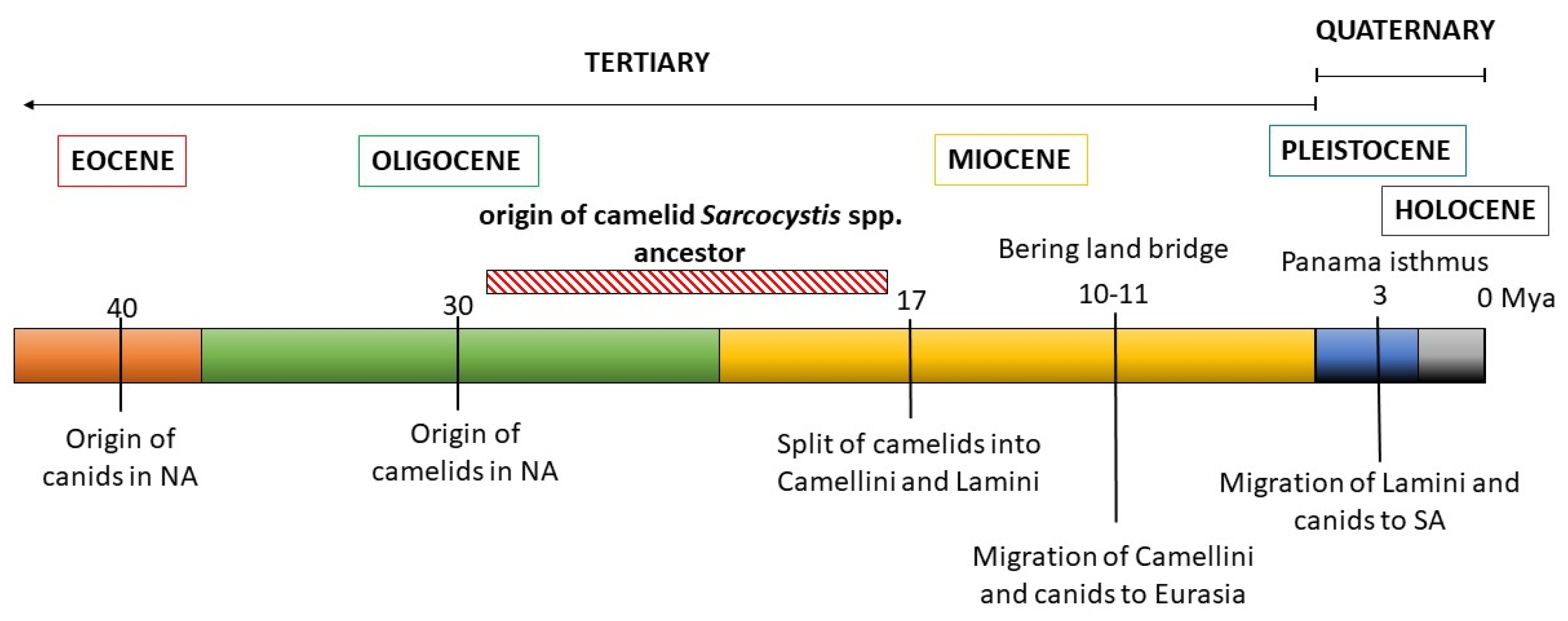

Table 1.
Characteristics of sarcocysts produced by camelid-infecting Sarcocystis sp.
| Intermediate host | Species | Sarcocyst | Ref. | ||||
|
Shape, size (length x width) |
Cyst wall | Location | |||||
| Type | Thickness (µm) |
Villar protrusions (vp) size and aspect | |||||
| OWC | S. cameli |
Fusiform, microscopic (700 x 100 µm) and macroscopic (1.5-5.0 x 0.2-0.4 mm) |
9j | < 2 | 3.0 x 0.5 µm finger-like vp |
cardiac and skeletal muscle | 3,17,18,26, 27 |
| S. ippeni | Globular, microscopic (100-120 x 50-100 µm) |
32 | 2.3-3.0 | 1.0 x 0.25 -1.2 µm thorn-like vp |
skeletal muscle | 3,17 | |
| SAC | S. aucheniae | Oval, macroscopic (0.5 -2.0 x 0.2 cm) |
21 | 6-10 |
3-4.5 x 2.5-3.5 µm branched vp, cauliflower-like wall |
skeletal muscle | 1,24,25, 28,29 |
| S. masoni | Fuisform, microscopic (800 x 95 µm) |
9j | 2.5-3.5 | 2-2.8 x 0.5-0.7 µm conical to cylindrical vp |
cardiac and skeletal muscle | 25 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



