
Submitted:
05 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Research studies related to neuroplastic changes that occur secondary to a bout of cystitis at a critical neonatal period of development in female rats (P14-16) are reviewed. They reveal that neonatal bladder inflammation (NBI) leads to an adult phenotype with many of the features of the human disorder, interstitial cystitis. Neurochemical, behavioral and neurophysiological differences between rats which experienced NBI and control animals are discussed and new data presented.
Keywords:
interstitial cystitis
; visceral pain
; neonatal
; urinary bladder
Introduction
Events that occur during childhood can lead to profound and permanent changes that become manifest later in life103. In animal models, neonatal pain experiences have been demonstrated to “prime” adult pain responses. This phenomenon was first reported by Ruda et al.100 using hindpaw inflammation, an observation that has been replicatede.g.,115. It has also been demonstrated following neonatal hindpaw incision8,25,53, facial incision82, neonatal systemic LPS exposure33, neonatal nerve injury116, neonatal muscle injection51, neonatal gastric suctioning109, neonatal colorectal distension35,122,123 and neonatal colonic inflammation4. Consistent with these multiple reports of adult hypersensitivity induced by nociceptive neonatal events are alterations that occur secondary to an episode of cystitis experienced at a critical period of development in rodents. First described by Randich et al.95 these changes mirror many of the phenotypic features of the disorder interstitial cystitis (IC) in humans61. As a consequence, this neonatal bladder inflammation (NBI) paradigm is able to serve as a model of the human disorder. In this review we propose to support the veracity of this statement by presenting a short summary of information related to IC in humans followed by a review of studies that have used NBI as an experimental manipulation. This review has identified some gaps in knowledge and when possible, we will present novel data to help fill those gaps. To optimize the ease of reading this information, the Materials and Methods section related to the new data is placed at the end of the document.
The disease to be modelled: Interstitial Cystitis (IC). IC is a chronic bladder disorder characterized by recurring pelvic discomfort or pain associated with altered urological organ function which is not due to readily defined causes of such sensations such as active urinary tract infections, stones or tumors. Dependent upon the precise definition of the disorder3,14,15, IC is present in either 1 in 10,000 females (1 in 100,000 males) when utilizing stringent criteria or in up to 2% of the population when utilizing purely symptom-based criteria. The term Bladder Pain Syndrome has often been used synonymously with IC. For purposes of this literature review, we will use the term “IC” and assign the diagnosis based on the following criteria: Inclusionary criteria: discomfort (pain and/or urgency) with low volume bladder filling or emptying and findings on cystoscopic examination that may include either a mucosal ulcer (Hunner’s patch) or submucosal petechial hemorrhages following hydrodistension (glomerulations indicative of vascular fragility); Exclusionary criteria: active urinary tract infection using standard criteria114; neoplasm on cystoscopy; anatomic abnormalities such as bladder diverticuli; current bladder or kidney stones; and neurological disease with identifiable pathology (e.g. spinal cord injury).
Common signs and symptoms. In addition to pain, subjects with the diagnosis of IC report frequent urination during the daytime (up to 30 times/day), persistent urgency, dyspareunia and nocturia13. Pain and urological symptoms are obviously related, but are viewed by many as distinct sensory entities29,45. Pain is commonly localized to the pelvis, but may extend into the abdomen and the perineum. Studies by the MAPP (Multidisciplinary Approach to Chronic Pelvic Pain of Urological Origin) Research Network, which is sponsored by the U.S. National Institute of Diabetes, Digestive and Kidney Diseases have characterized sites of pain in subjects with IC and identified that pain may be “global” in some subjects with evidence of diffuse widespread pain in some subjects44 and segmental pain in others2,13. Co-morbidities are common, especially “functional” disorders such as irritable bowel syndrome and fibromyalgia10,40. Some have suggested that the presence of such chronic overlapping pain conditions should be the basis for separation of the IC population into subgroups which are bladder-centric (pain more localized to regions in or near the bladder)50 and non-bladder-centric (or bladder-plus other) groups in which pain may be widespread44,46 including a group with multiple morbidities who are polysymptomatic and polysyndromic47. Brain imaging studies, psychophysical testing and cystoscopic findings support the stratification of the IC population into subgroups24,41,83,90,101. Flares in pain are the rule, rather than the exception, in the IC patient population and have been attributed to many causes with the most frequent being recurrent UTI, acutely stressful events and neighboring pathology associated with co-existing disease111,112. A common historical finding associated with the IC patient population is a history of childhood bladder infection88 and/or other adverse childhood events both psychological and physical30,49,89,102. Pain and urinary symptoms (e.g., urgency, frequency) occur commonly together, but clinically important differences in the severity of each, as part of flare symptomatology, have been noted110.
Physical exam findings. On examination, IC patients commonly report lower abdominal and pelvic floor tenderness to palpation31,42. Formal psychophysical testing has demonstrated statistically significant, hypersensitivity associated with deep tissue tests such as the ischemic forearm task, the cold pressor test, thumbnail bed pressure or muscle pressure thresholds23,24,32,42. In general, subjects with IC have not proven to be hypersensitive to cutaneous testing at either segmental or non-segmental levels using heat or superficial mechanical stimuli, but appear to require activation of deep (e.g. muscular) structures to demonstrate tenderness. Neuroimaging of the IC population has been extensive and differences in gray matter structure, bloodflow and connectivity have been demonstrated23,24,26,34,36,39,119. Studies have observed subgroup differences when other co-morbidities such as fibromyalgia are taken into account24.
Evocative Tests. At present, the gold standards for assessing pain are patient-based metrics. To refine localization of pain generation in IC, manipulations are performed to evoke bladder pain. For IC, evocative tests include cystometrograms (CMGs), the intravesical application of potassium solutions and the ice water test. CMGs are standard clinical tests used to diagnose altered bladder sensation. The test consists of catheterization of the bladder through the urethra followed by a slow infusion of a fluid (typically normal saline). A pressure transducer measures the intravesical pressure and the volume of fluid infused can be calculated. Subjects are instructed to report the point at which they have various sensations such as “First Pain” and “Maximal Toleration.” The volumes and pressures within the bladder at the time of the sensory reports are recorded, as are any contractions of the bladder. In experimental studies, numeric sensory intensity ratings are also reported during bladder filling. Studies in healthy females have demonstrated that repeated, sequential CMGs result in sensitization to bladder distension74 and there is a tight correlation between increases in intravesical pressure and sensory reports. Subjects with the diagnosis of IC report pain with lower intravesical volumes and pressures than healthy controls and tolerate much lower volumes and pressures of bladder filling75. The “Potassium Test” is uncommonly employed and consists of placement of a urinary catheter through the urethra and infusion of solutions of either normal saline or potassium chloride which are then allowed to dwell in the bladder85. If the patient reports an evocation of pain, the test is “positive.” It is intended to identify a “leaky bladder” in which potassium solutions are able to cross the urothelial barrier of the bladder and directly activate submucosal nerves. Parsons85 reported high sensitivity of the test in the IC population which he tested. A similar test is the “Ice Water Test” which is occasionally used to assess for neurogenic contractions of the bladder. It consists of placement of a urinary catheter and infusion of an ice water solution. An Ice Water Test is positive for central nervous system (CNS) injury if the infusion evokes a bladder contraction. Notably, subjects with IC given the Ice Water Test report bladder pain secondary to ice water infusion whereas healthy subjects do not. Hence, the diagnostic criteria of the test when looking for IC relates to a “negative” test for CNS injury (no bladder contraction), but a positive report of pain55. At present, there are no formal lab tests that have obtained acceptance as diagnostic for IC. Many substances including urinary fungi81, growth factors1 and neuropeptides in the bladder84 have been identified as being altered in subjects with IC when compared with controls, but at this point these do not serve as biomarkers but as topics for research.
Lack of Conditioned Pain Modulation (CPM) in IC subjects. A notable phenotypic finding related to subjects with the diagnosis of IC is that they, on formal testing, have deficient CPM mechanisms. In healthy humans, application of a painful stimulus to one part of the body as a conditioning stimulus reliably inhibits pain sensation in other body parts. This is typically quantified as an increase in thermal pain threshold or thermal pain tolerance following application of a painful stimulus to another body site. Ness et al.72 demonstrated that healthy subjects had robust CPM such that an increase in thermal pain thresholds/tolerance measured at the ankle was apparent after immersing a hand in ice water but subjects with IC had no such increase (no inhibitory CPM) and in some cases had a decrease in pain threshold/tolerance. This indicated a blunting or absence of this endogenous pain inhibitory system.
Summary of findings in humans with IC that can find correlate in rats. For a model of IC to have face validity it needs to have correlates to the human condition43. For this reason, we proposed that for a model of IC to be valid, it would need to demonstrate the following: hypersensitivity to bladder distension consistent with nociception (correlate to discomfort/pain); sensitivity to intravesical fluids such as in CMGs, ice water or potassium solutions (correlate to evocative tests); increases in symptoms with insults such as bladder inflammation, nearby pathology or stress (correlate to flares); altered central nervous system processing (correlate to altered fMRI); and altered endogenous antinociceptive systems (correlate to lack of CPM). We propose that, in the female rat, neuroplastic changes following NBI result in the fulfillment of these criteria.
Whole animal reflex responses that change following NBI. Models of visceral pain differ from those that use stimuli applied to the skin. Characterized by Sherrington at the beginning of the 20th century, pseudaffective reflexes are responses by animals that occur in response to noxious stimuli which are suggestive of responses to pain, with all of its emotional and physiological components, but which does not require a cortical level of processing of information67. Common measures include flexion-withdrawal reflexes evoked by noxious stimuli (e.g. heat, pinch) applied to superficial layers of the body. These reflexes are experienced by humans and their vigor correlates with human reports of pain. Visceral sensations have proved to be harder to model, but the distension of hollow organs produces pain in humans and robust visceromotor (abdominal contractions) and cardiovascular (typically pressor) reflex responses in both humans and non-humans9,12,67,71,74. These responses occur when the bladder is distended in rats and have correlates in humans74,75. The effects of NBI on these varied responses to UBD will be discussed individually below.
Effect of NBI on Reflex Responses to Bladder Filling. In a primary characterization paper, Randich et al.95 identified that there was a critical period of development during postnatal days 14 to 16 (P14-16; the end of the neonatal period) in which bladder inflammation induced by the transurethral intravesical administration of the yeast cell wall derivative zymosan (1% in saline, 30 minutes) resulted in an altered adult phenotype that had increased micturition episodes (measured using a Void Spot Assay) and increased vigor of cardiovascular and visceromotor (VMR; abdominal contractile) responses to UBD following acute bladder re-inflammation61,95. Neonatal control treatments for those experiments were intravesical saline and/or anesthesia/antibiotic treatments identical to what was received by the rats receiving intravesical treatments. Zymosan is a Toll-like Receptor 2 agonist (TLR2) and 1% solutions were utilized since previous experiments in adult rats had identified that concentration as optimal for the production of a time-limited bladder inflammation94. Similar intravesical administration of zymosan at P28-30 (equivalent of rat adolescence) did not produce the same effects in adults95. Subsequent reports refined timing and other methodology. Bladder inflammation on days P7-9 was not effective at producing adult effects61. Bladder inflammation on P21-23 led to increased micturition episodes as an adult, but not an augmentation of VMRs61. All subsequent timepoints had no effects on adult measures7,95. It is notable that bulbospinal inhibitory systems are forming in rats during the third week of life27. A single day of intravesical zymosan treatment on P14 was not sufficient to produce robust adult VMR effects but three daily treatments were sufficient61. Anecdotally, more than three days of NBI was associated with an undesirable level of pup mortality whereas three days of treatment had no effect on survival or weight gain after the period of treatment. Use of lipopolysaccharide (LPS; 100 mg/ml concentration) administered intravesically on P14-16 failed to produce adult effects61 suggesting subtleties related to innate inflammatory processes since LPS works via a TLR4 mechanism. The primary site for recording VMRs, quantified as electromyograms, has been the left external oblique musculature immediately superior to the inguinal ligament61. Similar (but less robust) visceromotor responses were also present in the pelvic floor musculature (pubococcygeus muscle). Both basal tone and evoked responsiveness of the pubococcygeus muscle was more robust in rats which experienced zymosan-induced bladder inflammation on P14-1661. Other nociceptive endpoints such as facial grimacing, vocalizations, locomotion and similar spontaneous behaviors have not been utilized in studies of NBI.
Protocols for Neonatal and Adult Bladder Inflammation. Based on these characterization studies a “standard” protocol for the production of NBI was established which is described more completely in the Materials and Methods section which consists of intravesical treatment with a 1% zymosan solution on days P14-16. Unless otherwise stated, the term “NBI” will indicate use of this standard protocol. Adult bladder inflammation (ABI) also followed a standard protocol in these studies which is also described in the Materials and Methods section and consisted of a single intravesical treatment with a 1% zymosan solution the day before testing. Unless otherwise stated, the term “ABI” will indicate use of this standard protocol.
NBI Increases Responsiveness to Infusion of Intravesical Solutions. Cystometrograms (CMGs) are the most common clinical measures of urinary bladder sensation. NBI results in reduced volume and pressure thresholds for the evocation of micturition responses20 when rats are tested as adults and compared with those that received neonatal control treatments. VMRs were also observed to be evoked by bladder contractions during the CMG testing when of sufficient intensity20. Adult rats which had experienced NBI also demonstrated VMRs following the infusion of a 0.4 mM solution of potassium chloride into their bladders (same as the human Potassium Test), whereas control rats do not61. Similarly, infusion of ice cold water (a correlate to the Ice Water Test) evokes VMRs but not bladder contractions in rats which experienced NBI, an apparent TRPA1-related phenomenon93, but not in control rats.
NBI Does Not Alter Hindpaw Somatic Nociception Measures. Both Randich et al.95 and DeBerry et al.20 examined hindpaw flexion withdrawal responses to thermal and mechanical stimuli and did not observe any quantitative differences between rats which had experienced NBI and their controls.
NBI Alters Elevated Plus Maze Measures. Elevated plus maze measures differ between rats which experienced NBI and their controls. NBI-treated rats spend significantly less time in the open arm of an Elevated Plus Maze compared to controls suggesting they have higher baseline levels of anxiety61.
Secondary Insults and Nociceptive Reflexes. As will be noted in following sections, NBI results in numerous biochemical and physiological alterations in adult rats without any additional manipulation. However, the evocation of augmented pseudaffective reflex responses (both cardiovascular and visceromotor) has required the administration of a secondary insult (NBI being the primary insult). The most common secondary insult which has been employed is ABI as described above using zymosan. However, infusion of a solution of 100 mg/ml LPS into adult rats which experienced NBI also produced augmented VMRs61. Notably, recent studies have identified that NBI not only increases the vigor of VMRs to UBD when treated as adults with these previously tested concentrations of zymosan and LPS, but also reduced the concentration of these TLR2/TLR4 agonists which was necessary to result in an augmention of VMRs to UBD when compared with controls6. Other secondary insults which have been demonstrated to produce an augmentation of VMRs to UBD in rats which experienced NBI include hindpaw inflammation (produced by injection of Complete Freund’s Adjuvant 3 days prior to testing)61,65, chronic constriction of the infraorbital nerve61 and acute footshock stress61,66. These secondary insults and their production of bladder hypersensitivity serve as correlates to flares in the symptomatology of IC subjects. Interestingly, others38,52 have noted differential effects on the vigor of VMRs evoked by non-bladder stimuli that occur between NBI-treated rats and their controls. In these studies, NBI-treated rats demonstrated significantly more robust colorectal distension-evoked VMRs than those of control rats.
Absence of Stress-Induced Somatic Analgesia. One of the robust endogenous inhibitory systems that can be evoked in animal models without pharmacological manipulation is the phenomenon of Stress-Induced Analgesia11. Whereas normal rats will demonstrate a stress-induced elevation in their cutaneous thermal and mechanical thresholds for evocation of a withdrawal reflex (i.e., it takes a higher temperature to evoke a flexion reflex following a stressor), rats which experienced NBI were demonstrated to have no effect of an episode of acute footshock stress on reflex responses to somatic stimuli66.
Summary of Changes in Whole Animal Measures Following NBI. Using whole animal reflexes and responses as primary endpoints, these studies have characterized hypersensitivity to bladder distension that is consistent with increased nociception. It is notable that the augmentation of pseudaffective reflex responses such as VMRs and cardiovascular responses that define hypersensitivity, require an accomplice: a second, adult insult. The present rodent model utilizing NBI is therefore presented as a “two-hit” model of pathology requiring a developmental insult (NBI) as a necessary but not sufficient event which must be followed by a secondary insult in order to become “clinically” manifest. The first insult appears to “prime” the system and the second insult makes the pain component of the disorder become manifest. Secondary insults which have been reported to increase the vigor or pseudaffective responses include segmental inflammatory events (e.g., bladder inflammation, hindpaw inflammation), distant neuropathic pain and acute stress61. These increases in responses also have correlates to the flares in symptoms that occur in subjects with IC. Other features of the NBI model include sensitivity to intravesical fluids such as occurs with ice water infusion or potassium chloride solution infusions, both of which activate VMRs in NBI-treated rats but not controls. Cystometrograms in NBI-treated rats also demonstrate hypersensitivity to fluid distension with lower pressure/volume thresholds needed to evoke micturition. Last, but not least, whole animal studies have demonstrated alterations in endogenous antinociceptive systems by demonstrated an absence of stress-induced analgesic mechanisms. Additional findings such as pelvic floor hypersensitivity and increased measures of anxiety round out positively comparative results between rats and humans. Taken together, these findings indicate that rats which experienced NBI have many of the behavioral phenotypic features of IC in humans. We would argue that the NBI model in female rats has good face value as a model of IC. Notably, a similar, but less extensive set of studies has also been performed examining the effects of NBI in female mice18 with similar, but not identical findings.
Theories related to etiologies. Ultimately, the goal of any model is to identify mechanisms related to the disease of interest so that preventative or interventional therapeutics may be devised in relation to the disorder. The main theories related to the etiology of IC include the following: Subjects with IC have a urothelial abnormality such that primary afferents in the bladder become exposed to “toxic urine”; IC is an autoimmune disease; IC is a variant of neuropathic pain; IC is a variant of myofascial pain; IC is a local manifestation of a total body pain mechanism; IC is due to genetic factors; IC is a psychiatric disorder; and IC is due to infection by a fastidious organism. What most proponents of these various theories fail to recognize is that they are not mutually exclusive as etiologies of IC. As noted above, a “two hit” model requires an initial developmental insult (in this case NBI) followed by an adult insult/susceptibility (one of the above). For purposes of face value of the NBI model, one also needs to understand specific neuroplastic changes that are due to NBI that occur in elements of sensation associated with the bladder. We will describe the ones that are known below, but first we will make an assessment of some current deficits of knowledge related to whole animal measures and attempt to fill some of the identified gaps with new data.
Gap #1 – what are effective second insults in the NBI model? This is obviously an open-ended question which will hopefully be addressed by individuals who ascribe to the different theories related to the etiology of IC since the theories are not mutually exclusive when a two-hit mechanism is hypothesized. A simpler, testable question is whether all nociceptive events in the body are sufficient to result in bladder hypersensitivity in the NBI model. To address this question we asked an even more mechanistic question: Is inflammatory pain anywhere always an effective “second insult”? Studies published-to-date which used inflammation as an adult pretreatment in the NBI model chose to inflame nearby tissues which had afferent information processed in the same, or immediately adjacent, spinal segments as conditioning stimuli (i.e. bladder re-inflammation, hindpaw inflammation). Therefore, studies identical to those described in Ness et al.61,65 were performed, the only exception in methodology being that instead of just inflaming the left hindpaw of the rat by injecting CFA three days prior to subsequent measurement of VMRs to UBD, the right hindpaw, cheek or forepaw were injected with the same amount of CFA (100 ml, or saline for controls) three days prior to testing. Measured endpoints were the VMRs evoked by graded UBD using electrodes placed into the left superior oblique musculature. Mechanical hypersensitivity at the sites of injection were confirmed by probing the sites with a cotton swab which activated a withdrawal response. Redness and swelling were also present at the sites of injection. A more complete description of these methods is presented in the Materials and Methods section.
Results of these studies reproduced earlier reports indicating that CFA-induced inflammation of the left hindpaw on the side ipsilateral to the EMG recording sites (a perisegmental inflammatory stimulus) produced an augmentation of VMRs to UBD in NBI-treated, but not in neonatal control-treated (NoNBI) rats which actually were slightly reduced [Figure 1A]. A similar finding was observed when the site of CFA-induced inflammation was in the hindpaw contralateral to the EMG recording site (also a perisegmental inflammatory stimulus) [Figure 1B]. However, when the site of CFA-induced inflammation was the contralateral forepaw or contralateral cheek region, there was no augmentation of VMRs to UBD in either NBI-treated or control rats (heterosegmental inflammatory stimuli) [Figure 1C,D]. These findings suggest that any form of nociception experienced at another site of the body is not sufficient as a secondary insult in the NBI model. Other secondary insults are yet to be investigated.
Bladder Tissue Measures That Change With NBI and/or ABI. The following sections will examine changes that occur as a consequence of NBI at different sites along the sensory pathway associated with the urinary bladder. ABI was used as a second insult in these studies.
Gross Histological Findings. When using Hematoxylin & Eosin, Gomori’s Trichrome or Toluidine Blue stains on thin sections of bladder tissue, there were no quantitative histological differences noted between rats which had experienced NBI and their controls20 when no second insult was presented. It was therefore notable that following “hydrodistension” (30 minutes of sustained intravesical pressure produced by a column of normal saline which is similar to what is done clinically in patients with suspected IC), the rats which had experienced NBI had a threefold increase in the number of hemorrhages upon release of the pressure than rats which had received neonatal control treatments61. These hemorrhages are a correlate to glomerulations that are noted following hydrodistension in humans with IC and suggest a vascular fragility within the bladder wall of rats which experienced NBI. Some human studies1 have suggested that Vascular Endothelial Growth Factor (VEGF) is increased in subjects with the diagnosis of IC. Since additional vascularity could explain the noted hemorrhages, we briefly investigated whether VEGF was changed in the NBI model.
Gap #2 – Are there differences in VEGF content of the bladder secondary to NBI?
More fully described in the Materials and Methods section, the experimental process was to form four groups of rats similar to previous studies which underwent combinations of the NBI and ABI protocols using intravesical zymosan as the inflammogen: NBI+ABI, NBI Only, Control Only and ABI Only groups. As adults, bladders were harvested and using ELISA measures, VEGF levels measured.
As apparent from the data presented in Table 1, ABI led to statistically significant increases in VEGF levels, but NBI had no statistically significant effect. This indicates that VEGF levels in the NBI model are elevated, consistent with human studies, but only when local inflammation is present.
Other Endogenous Agonists and Receptors. As indicated in Table 1 bladder content of the toll-like receptors TLR2 and TLR4 have been reported to be greater in rats which experienced NBI when compared with those which did not experience NBI6. Interestingly, the bladder content of the endogenous kappa opioid agonist, dynorphin, was reduced in rats which had experienced both NBI and ABI when compared to controls but NBI by itself was not sufficient to cause this reduction108. In association with bladder sensation, the neuropeptides Substance P and Calcitonin Gene-Related Peptide (CGRP) have particular relevance. Mustard oil-induced neurogenic inflammation, measured as Evan’s Blue extravasation has been demonstrated to be greater in rats which experienced NBI when compared with their controls20. Thought to be an indirect measure of primary afferent neuropeptide content, this observation prompted more extensive studies related to Substance P and CGRP total content within the bladder both of which were demonstrated to be greater in rats which experienced NBI20. Using ELISA measures, Shaffer et al.106 examined the whole bladder neuropeptide content of rats which were in four groups: those which experienced NBI and ABI; those which experienced NBI Only; those which received ABI Only treatments and those which received Control Only treatments. Results for these studies are given in Table 2. NBI led to both increased Substance P and CGRP content in rats with the greatest increases in rats which also experienced ABI. An additional study of note used a similar model of neonatal bladder irritation as described by Park et al.86 in which 0.2% acetic acid solutions were infused into the bladders of female rats on P10. These investigators observed increased expression of TRPV1 in both bladder tissues and in lumbosacral dorsal root ganglia of the same rats when 8 weeks old. There are still a few lingering questions related to these studies. For example, the precise localization of the increased content of sensory neuropeptides in the bladder tissues due to NBI is unknown. To address this gap in knowledge, the following studies were performed.
Gap #3 – Does NBI change the location of Substance P and CGRP in the bladder? Studies published-to-date which demonstrated NBI-induced increases in Substance P and GCRP content (which are noted in Table 2) only measured total bladder content of the neuropeptides. Electrophysiologic study of primary afferents associated with the urinary bladder indicate that there are subpopulations of these neurons with differential distribution in the bladder tissues120. Recent studies have identified a particular uniqueness to the “urothelial” bladder primary afferents which have bare nerve endings immediately adjacent to and within the urothelium16,17,37 when compared with “non-urothelial” bladder primary afferents (all the rest). Using standard immunohistochemical techniques for Substance P and CGRP labelling of bladder tissues, it was possible to take photomicrographs that could be analyzed quantitatively for the intensity of neuropeptide labelling in the urothelial/immediate suburothelial area and to compare it with the labelling of the total bladder, generating a ratio indicating the relative labelling of the urothelial area. This data is presented in Table 2. Specific methods for this study are given in the Materials and Methods section.
In these studies, NBI, by itself, produced a statistically significant increase in the Urothelial versus Non-Urothelial labelling of CGRP and a statistically significant decrease in the Urothelial versus Non-Urothelial distribution of Substance P labelling when compared with control animals. ABI produced a statistically significant increase in the Urothelial to Non-Urothelial ratio of both CGRP and Substance P labelling in rats which had not experienced NBI, but had no effect on relative distribution in rats which had experienced NBI (Table 2).
Other Effects of NBI on Bladder Primary Afferent Neurons. Apart from the study by Park et al86, noted above, there is limited information related to changes induced by NBI measured in dorsal root ganglion neurons. There are also very few studies which have electrophysiologically assessed whether there are differences in the activity of bladder primary afferents that occurs secondary to NBI treatment. Kannampalli et al.38 examined the effects of NBI on a small sample of pelvic nerve afferents and observed a slightly lower rate of spontaneous activity in NBI-treated rats in comparison with control rats. However, this difference was not statistically significant due to low sample size. The level of excitation of these neurons evoked by bladder filling was similarly less in NBI-treated rats but interpretation was also limited by small sample size. Afferent pelvic nerve fibers in both treated and untreated rats appeared to encode for UBD in a similar accelerating monotonic fashion. Application of a selective GABAAa-2 receptor agonist increased the spontaneous activity of NBI-treated pelvic nerve afferents but had no statistical effect on control afferents. Zhang et al122 also identified that NBI led to downregulation of lumbosacral spinal tissue measures of potassium chloride co-transporters and vesicular GABA transport protein expression, but it not known whether this is primary afferent or second order neuron effect.
More deficits of knowledge. Any list of what we don’t know about changes in bladder primary afferent neurons as well as the bladder itself which are induced by NBI could be endless. Obvious candidates for investigation include other growth factors, other neuropeptides, measures related to the muscular, vasculature and immune systems, autonomic function measures and more. Particularly notable, our neurophysiological understanding of primary afferents associated with the bladder and how they may change secondary to NBI is almost non-existent. Obviously, neuropeptide content and distribution changes are relevant to our understanding. This is particularly true for CGRP changes since this peptide is only contained in primary afferents. It is known that bladder primary afferent neurons travel to both thoracolumbar and lumbosacral spinal segments5 and that characteristics of the different subpopulations which have different spinal destinations are different16,28,120 but at present, these spinal destination differences have only been characterized in the healthy or acutely inflamed states and NBI as a manipulation is still to be tested. Clearly, there is fertile ground for more studies, particularly in the realm of bladder primary afferent neurons. At present, we have to extrapolate NBI-induced changes observed in spinal processing and following spinal drug administration as indirect measures of their function.
What we know about “normal” spinal neurons excited by urinary bladder distension (UBD). Quantitative characterization studies of spinal dorsal horn neurons responsive to UBD in the rat have identified at least two populations of neurons that can be differentiated from each other by the existence versus absence of heterosegmental inhibitory influences56. Excitatory responses to visceral and cutaneous inputs are important defining characteristics, but have less predictive value in response to subsequent manipulations. Approximately half of spinal neurons that are excited by UBD demonstrate spontaneous activity that is inhibited >20% by the application of a noxious stimulus to a heterosegmental site (e.g., pinch of forepaw, neck skin, nose). These neurons are designated as Type I neurons. The other half of the UBD-excited neuronal sample (Type II neurons) are not inhibited by this stimulus demonstrating either no response to heterosegmental noxious stimuli or, in some cases, excitation. Both types of neurons demonstrate convergent excitatory cutaneous receptive fields, but the size of these receptive fields is smaller in Type I neurons (Type II neurons frequently have whole body noxious cutaneous excitatory inputs) and demonstrate an “inhibitory surround” characteristic where stimulation of cutaneous structures immediately outside of the excitatory field produce inhibition of the neuron56. A roughly 50%-50% Type I-Type II distribution has also been noted in spinal neurons excited by colorectal distension68. This Type I-Type II classification system has proven predictive of responsiveness to local inflammation and various pharmacological agents56,57,68-70. Notably, when examining spinal dorsal horn neurons in the lumbar enlargement which are excited by noxious somatic stimuli (e.g., heat, pinch) the Type I-Type II distribution is roughly 95%-5%76, but still predictive of response to pharmacological agents76. Categorization according to neurons’ excitatory cutaneous inputs (as Class 2 or Class 3 neurons, excited by both noxious and noxious inputs versus only noxious inputs respectively) was less predictive of the effects of pharmacological agents. Other characteristics of spinal neurons excited by UBD is that they are located in the superficial and deep laminae (including lamina X in the dorsal commissure region of the spinal cord) of the L6-S2 spinal segments, as well as the T10-L2 spinal segments and are generally absent from the lumbar segments exclusively associated with the hindlimb (L3-4). Both Type I and Type II neurons encode for UBD in a monotonic, accelerating fashion. Following acute inflammation, Type I neurons become less active and Type II neurons become more active57,69.
NBI and the activity of individual spinal dorsal horn neurons. In the absence of other manipulations, Type I and Type II neurons demonstrate similar levels of spontaneous activity as well as activity evoked by UBD56. This changes following NBI64,65,77. If no secondary pretreatments occur as adults, then both the spontaneous and UBD-evoked activity of Type I neurons in NBI-treated rats becomes reduced in comparison with their controls. However, if ABI occurs as an adult in NBI-treated rats, then the spontaneous and evoked activities of both Type I and Type II neurons are more robust when compared to the NBI-treated rats which did not get their bladders re-inflamed64,77. In contrast, rats which did not experience NBI but instead received neonatal control treatments, Type I neurons become less active following ABI and Type II neurons become more active in a fashion identical to that noted above in “naïve” (otherwise untreated) rats64,77. A similar phenomenon occurs with acute hindpaw inflammation induced by injection of Complete Freund’s Adjuvant: Type I neurons, which are inhibited by this treatment in control rats become more active in NBI-treated rats; Type II neurons become more active in both control and NBI-treated rats65. The simplest mechanism to describe these effects is that inhibitory influences on Type I neurons which are activated by noxious stimuli are reduced or absent in NBI-treated rats. As was noted above, human subjects with the diagnosis of IC also have reduced or absent Conditioned Pain Modulation, the human equivalent of noxious stimulus-activated inhibition72. As previously noted, in rats, stress-induced analgesia (another endogenous antinociceptive system) also appears to be reduced or absent in NBI-treated rats66. Kannampalli et al.38 also examined the effects of NBI on spinal dorsal horn neurons in a limited sample of neurons. In their hands, the spontaneous activity of spinal dorsal horn neurons in NBI-treated rats was similar to that of controls, but it was notable that the neurons in control rats appeared to be excited by a slow infusion of saline into the bladder whereas the same was not true for the NBI-treated rats. The findings of this study are difficult to compare with other studies as other characteristics of the neurons (e.g., cutaneous receptive fields, effect of heterosegmental noxious stimuli) were not described nor was the effect of segmental somatic stimuli. What was notable in these studies was that inhibition of dorsal horn neurons which was produced by the direct spinal application of a selective GABAAa-2 receptor agonist in control rats was absent in NBI-treated rats giving additional evidence of missing inhibitory mechanisms in NBI-treated rats. These findings were consistent with studies by these same investigators which identified deficits in other GABAA subunit receptor expression in the spinal cord of rats which had experienced NBI104. It is not known whether NBI alters excitability of different neuronal groups located within different laminae of the spinal cord (Type I and Type II neurons appear intermixed in both superficial and deep laminae) and so a set of studies was performed to answer the following question:
Gap #4. Does the location of dorsal horn neurons excited by UBD change with NBI?
Using nuclear c-Fos protein induction as an indirect measure of neuronal excitation, the location of spinal neurons excited by UBD was mapped to four locations: the lateral dorsal horn, the medial dorsal horn, the dorsal commissure region and the spinal parasympathetic nucleus (intermediolateal gray matter) in a fashion previously described by Vizzard117 (more complete methods in the Material and Methods section). The data from these studies is indicated in Table 3. Whereas UBD resulted in a robust, statistically significant increase in the number of c-Fos positive nuclei, particularly in the dorsal commissure area of the spinal cord, ANOVA analysis demonstrated no statistical effect of either NBI or ABI. This would suggest that the location of neurons which are excited by UBD is not grossly changed by NBI but this does not rule out changes in responsiveness. Namely, neurons do not appear to change from an excited to inhibited condition, but rather, may be less excited, such as was suggested by electrophysiological studies77.
NBI-Induced Changes Following Intrathecal Receptor Antagonists. Using the effect of spinal intrathecal drug administration to localize effects of pharmacological antagonists, it is possible to imply tonic mechanisms that may be altering spinal neuronal processing in rats which have experienced NBI. These studies suggest there are tonic effects of alpha adrenergic63, serotonergic63, GABAergic63, glycinergic63 and NMDA glutaminergic66 receptor activation. The caveat associated with these studies is that other manipulations, such as peripheral nerve stimulation, were also being studied and so results could have be affected by the experimental preparation.
NBI Alters the Role of Endogenous Opioids. Numerous changes occur in the spinal opioid systems of rats when they experience neonatal injuriese.g.53 including NBI. As noted in Table 1, both bladder and spinal cord contents of opioidergic peptides are altered in rats which experienced NBI108. Particularly notable is that the endogenous kappa opioid receptor agonist, dynorphin, was increased in the lumbosacral spinal cord of rats which had experienced NBI108. In a series of studies, the nonselective opioid receptor antagonist, naloxone, was used to probe for a role of endogenous opioids in the modulation of sensory processing in rats which had experienced NBI. Spinal intrathecal administration of the naloxone allowed for localization of drug effect to spinal levels. Previous studies by Ball et al.7 observed that intrathecal naloxone had minimal effects on rats which had not experienced any form of bladder inflammation. However, following ABI, otherwise naive-to-treatment rats demonstrated augmented VMRs to UBD which became even more magnified if pretreated with intrathecal naloxone. This indicated that in healthy rats, ABI results in the activation of a reactive, inhibitory, spinal opioidergic system which suppresses the vigor of VMRs. Deberry et al.19 extended this observation to rats which had experienced NBI by demonstrating that this opioidergic system was absent in NBI-treated rats. In other words, rats which had received neonatal control treatments, when treated as adults with ABI had augmented VMRs that augmented further with systemic naloxone treatment (just like the rats in Ball et al.’s study). However, rats which had experienced NBI when further treated with ABI had augmented VMRs, but these VMRs were not altered by naloxone treatment.
A similar absence of an endogenous inhibitory opioidergic effect was noted in studies examining acute footshock (AFS) -induced bladder hyperalgesia. In these studies, naloxone, administered intrathecally, demonstrated that endogenous spinal opioids appear to switch from producing inhibition of VMRs following AFS in control rats to producing a pronociceptive effect in rats which experienced NBI and then AFS as an adult60,66 . In these studies, intrathecal naloxone led to more robust VMRs in rats which experienced AFS but no NBI. In contrast, the same dose of intrathecal naloxone reduced the robustness of VMRs in rats which had experienced both NBI and AFS66. These observations are consistent with a failed opioid inhibitory system, as noted above, but also suggests that an excitatory opioidergic system becomes manifest following NBI. As noted above, spinal cord dynorphin levels are increased in rats which experienced NBI and dynorphin is often a candidate for opioid-induced excitation. Other common candidates for pronociceptive processes include NMDA-glutamate receptor activation which was also demonstrated to increase VMR vigor in rats following AFS whether they had experienced NBI or control treatments66.
Effects of NBI on Responses to Exogenously Administered Opioids. Shaffer et al.107 also examined opioid effects in a functional fashion by administering the mu opioid receptor agonist, morphine, and the kappa opioid receptor agonist, U50,488H, systemically to rats which had experienced NBI and/or ABI and then compared the effect of these drugs on the VMRs to UBD with those of control rats. Whereas Control Only rats (no pretreatments) demonstrated dose dependent inhibition of VMRs to UBD for both drugs at all doses tested, rats which experienced both NBI and ABI demonstrated augmentation of their VMRs by a 1 mg/kg dose and a significantly diminished inhibitory effect of a 4 mg/kg dose of U50,488H. Rats which experienced NBI Only or ABI Only did not differ in their responses from rats which experienced neither NBI nor ABI. This differential effect of U50,488H was not observed when morphine was used as an analgesic as all groups demonstrated similar inhibitory effects of drug independent of previous treatment with NBI and/or ABI59,107. Subsequent studies using the kappa opioid antagonist, nor-BNI, administered into the spinal intrathecal space 24 hours prior to testing, gave good evidence that the sites of action of the U50,488H were at a spinal level, but methodological issues related to effects of the drug and spinal catheter placement on baseline measures of VMRs precluded a definitive demonstration of effect105.
Response to Baclofen. The analgesic/antispastic GABAB receptor agonist baclofen was also studied in the NBI model79. In rats which had not experienced NBI, baclofen was demonstrated to produce robust, dose-dependent inhibition of UBD-evoked VMRs and cardiovascular responses to UBD (pressor responses) when it was administered systemically or intrathecally. Receptor specificity was demonstrated using pretreatment with selective GABAB antagonists and a spinal action of effect was verified by topical application to the spinal cord which produced inhibition of spinal dorsal horn neurons excited by UBD. In rats which had experienced NBI and ABI baclofen was also demonstrated to produce a robust, dose-dependent inhibition of UBD-evoked VMRs which was statistically indistinguishable in magnitude from that measured in rats which had not experienced NBI. This suggests that GABAB-related spinal mechanisms are not altered by NBI.
CRFR1/CRFR2. Receptors important in stress-related hypersensitivity phenomena are those that are responsive to corticotropin releasing factor (CRF) and urocortins: CRF receptor 1 (CRFR1 – endogenous agonists are CRF and Urocortin 1) and CRF receptor 2 (CRFR2 – endogenous agonists are Urocortins 1-3). Studies by Robbins et al. observed that chronic stress produces an augmentation of VMRs to UBD96,97. Spinal intrathecal administration of CRFR2 but not CRFR1 antagonists blocked this VMR augmentation when produced by chronic footshock stress97. Chronic footshock was also demonstrated to alter CRFR1, CRFR2, CRF and Urocortins 1-3 content96. Due to these observations of augmentation in relation to bladder sensation, these agents were investigated in association with NBI-induced augmentation of UBD-evoked VMRs and dorsal horn neuronal responses.
When ABI is used as an adult pretreatment in rats which experienced NBI, there are multiple changes in CRFR1, CRFR2, CRF peptide and Urocortin 2 content is the lumbosacral spinal cord64 (Table 1). Most important of these is an increase in both the CRFR2 content and the associated content of its agonist Urocortin 2. CRFR1 content and the content of its agonist CRF are also increased in this group, but it appears to be more related to the acute inflammatory state than the neonatal treatment (Table 1). VMRs in rats which experienced NBI and ABI were reduced when the CRFR2 antagonists aSVG30 and Astressin 2B were administered intrathecally. In rats which experienced NBI but no ABI, the aSVG30 paradoxically increased responses. Similar things happened in association with Type I and Type II spinal dorsal horn neurons following topical application of aSVG30: in rats which had experienced NBI and ABI, the aSVG30 was inhibitory; but in rats which experienced NBI but no ABI, application of the aSVG30 results in more vigorous responses, at least within the Type II neurons.
When CFA-induced hindpaw inflammation was used as a second insult in adult rats which had experienced NBI or control treatments, it was observed that this manipulation increased spinal CRFR2 content in rats which had experienced NBI independent of adult treatments but also increased CRFR2 content in all rats which received CFA in their paws65 (Table 1). Notably, spinal CRFR1 content also increased in all NBI-treated rats whether they experienced CFA injections or not, but did not increase secondary to the hindpaw inflammation treatment by itself (Table 1). The administration of spinal intrathecal CRFR2 antagonists blunted the augmentation of UBD-evoked VMRs produced by hindpaw inflammation in rats which had experienced NBI but produced an augmentation of VMRs in rats which experienced NBI, but no hindpaw inflammation65. In a parallel set of studies, topical administration of the same antagonist to the exposed lumbosacral spinal cord led to an inhibition of spinal dorsal neurons in rats which had experienced NBI and hindpaw inflammation but an augmentation of responses in rats which experienced NBI, but no hindpaw inflammation65. These findings were similar to those noted in NBI-treated rats which experienced ABI as a second insult. These findings which were consistent between VMR measures and dorsal horn neuronal measures and suggest that there is a tonic inhibitory effect of CRFR2 in rats which have experienced NBI but no second insult. This inhibition appears to convert to an excitatory effect of CRFR2 activation following the secondary insult. To test whether this was the case, we performed a brief set of experiments using the intrathecal administration of the CRFR2 agonist, Urocortin 2 to modify VMRs in rats which had experienced NBI and/or ABI in order to answer the following question:
Gap #5. Does the effect of Urocortin 2 change with a second insult in rats which had experienced NBI?
To answer this, rats which had experienced NBI also received either ABI or an adult control treatment. 24 hours later they were anesthetized, an intrathecal catheter placed and a 15 mg dose of Urocortin 2 administered to the lumbosacral cord (more complete methods are given in the Materials and Methods section). 15 minutes later VMRs to UBD were obtained. The data (Figure 2) demonstrated that administration of Urocortin 2 produced analgesia in NBI-treated rats if their bladders were not re-inflamed when compared with similar rats given intrathecal normal saline. However, if the bladders were re-inflamed, augmented VMRs were observed when compared with those rats with uninflamed bladders. No additional augmentation of VMRs was noted in those rats with the administration of Urocortin 2. This result is consistent with the presence of an inhibitory mechanism that uses CRFR2s in NBI-treated rats which did not experience ABI. Following ABI there is either a loss of these inhibitory influences and/or they are overwhelmed by excitatory influences also occurring via CRFR2 mechanisms. The precise neurophysiological mechanism will take single cell neuronal studies to identify. At this point we can say that these studies have confirmed that an alteration in the overall effects of CRFR2s has occurred due to NBI. A similar, but not identical, switch in receptor effect in the kappa opioid receptor experiments was described above. In those studies, a 1 mg/kg IV dose of U50,488H which was inhibitory in control rats, became excitatory in NBI-treated rats107.
An additional unanswered question raised by these experiments is whether all effective secondary insults in the NBI model utilize CRFR2 mechanisms to produce their augmentation of responses to UBD. To answer this question we examined the role of CRFRs following a different secondary insult, namely acute footshock stress (AFS), in NBI-treated rats and answered the more precise question:
Gap #6 – What is the role of CRFRs in the augmentation of VMRs produced by AFS?
As noted, ABI and CFA-induced hindpaw inflammation were both demonstrated to augment VMRs to UBD in NBI-treated rats through CRFR2-related mechanisms. Using similar methods (described more completely in the Materials and Methods section), rats were assigned to four groups: those that experienced NBI and then AFS as adults (NBI+AFS), those that experienced NBI but no AFS (NBI Only), those that received neonatal control treatments and AFS as adults (AFS Only) and those that received both neonatal and adult control treatments (Control Only). Immediately following their adult treatments rats were either sacrificed for harvest of their L6-S2 spinal cord segments for ELISA studies or underwent testing for VMRs to UBD. The spinal cord content of CRFR1s and CRFR2s was found to be statistically lower in rats which experienced NBI when compared to rats which did not experience NBI (two way ANOVA for NBI effect for CRFR1: F1,1,16 = 11.657, p=0.004; two way ANOVA for NBI effect for CRFR2: F1,1,16 = 12.457, p=0.003). The spinal cord content of CRFR1s and CRFR2s was statistically unaffected by AFS and there were no interaction effects in the data. These data are displayed in Table 1. An augmentation effect related to AFS61 was confirmed by comparing data from different pretreatment groups subsequently treated with IT normal saline (filled circles in Figure 3A–D). Using only that subset of data, it is apparent that graded VMRs evoked by graded distensions were statistically more robust in rats that received AFS pretreatment whether they had been pretreated with both NBI+AFS (Figure 3A) or received AFS Only pretreatment (Figure 3B) when compared with rats that had been pretreated with NBI Only (Figure 3C) or Control Only Pretreatments (Figure 3D). A repeated-measures two-way ANOVA of the IT saline-treated data from all four pretreatment groups demonstrated statistical significance of the AFS pretreatment effect (F1,1,27 = 7.711; p=0.010) and the NBI+AFS group had the most robust responses. To examine the role of CRFR1s and CRFR2s on the increased vigor of rats which experienced AFS, the CRFR1 antagonist antalarmin and the CRFR2 antagonist aSVG30 were administered intrathecally prior to the measure of VMRs to UBD. As apparent in Figure 3, neither the CRFR1 nor CRFR2 antagonists at the doses employed were effective at reducing the vigor of the VMRs to UBD when compared with VMRs in rats administered intrathecal saline in any of the pretreatment groups.
These findings demonstrated that the augmenting effects of the adult exposure to AFS was unaffected by CRFR1 or CRFR2 antagonists at the doses tested and the lumbosacral spinal cord content of both CRFR1s and CRFR2s were reduced (rather than increased) in those rats which had experienced NBI. These findings are diametrically opposite to the effects of inflammatory treatments (to bladder or hind limb) which appear to activate CRFR1 and CRFR2 related mechanisms in rats which experienced NBI. As a consequence, we can answer the question of whether all effective secondary insults in the NBI model act via CRFR2-related mechanisms in the negative. Other mechanisms appeared to be in play when AFS is used as a pretreatment. These have been studied and reported60,66 subsequent to the present studies. As noted, NBI-treated rats appear to lack a stress-activated, opioidergic inhibitory system that is present in rats which have not experienced NBI. At the same time, in these rats, there is an intact excitatory glutamatergic system present which is activated by AFS. The result is an unopposed facilitatory effect with resultant increased vigor in VMRs. Therefore, our best explanation for second insult-related augmentation of VMRs is that the lack of an opioidergic inhibitory system in NBI-treated rats leads to an unopposed pro-nociceptive mechanism which in some paradigms are CRFR2-related and in others are NMDA-related. The end result of both of these alterations in pain modulatory systems is the presence of augmented VMRs to UBD. An important caveat in the AFS studies is that there was only limited time between AFS and the subsequent measures reported such that there may have been insufficient time for the induction of protein synthesis important to CRFR2-related mechanisms.
Obvious areas of ignorance related to spinal processing. Clearly, there are many more areas to define when it comes to the neuroplastic effects of NBI on spinal sensory processing. Intracellular recordings may be able to answer specific questions related to excitatory and inhibitory inputs that cannot be addressed by extracellular recordings which only have the quantal action potential response as readout. Lamina specific phenomena still need to be defined as do potential segmental phenomena and other measures of local connectivity. The neurochemistry of inputs, outputs and modulations of spinal neurons that may be altered by NBI is poorly understood. That said, there are even greater areas of ignorance in our understanding of areas lying cephalad to the spinal cord.
SupraSpinal Processing. Similar to our limited understanding of bladder primary afferents and how they are changed by NBI and our limited understanding of spinal processing, our neurophysiological understanding of supraspinal mechanisms related to bladder sensation and the effect of NBI is close to non-existent. Although supraspinal neurons responsive to bladder distension have been extensively studied, as have their spinal pathways for inpute.g. 98,99, study of the effects of NBI on specific supraspinal sites, to the best of our knowledge, is minimal. Talluri et al.113 characterized the effect of NBI on neurons located in the rostral ventromedial medulla (RVM) which received bladder inputs and/or colon inputs as identified by pseudorabies virus tracing. Following NBI there was an increase in the number of RVM neurons receiving both bladder and colon inputs. There was also a downregulation of vesicular GABA transporter and an increase in tryptophan hydroxylase 2 in bladder-linked RVM neurons.
Other studies have given indirect information related to supraspinal sites of bladder sensory processes by examining the effects of electrolytic lesions of the amygdala. Previous studies of AFS-induced hyperalgesia in the absence of NBI, demonstrated that structures in the amygdala were required for the induction of bladder hypersensitivity21,62 since an electrolytic lesion of the central nucleus of the amygdala appeared to abolish this phenomenon. As noted above, AFS has also been demonstrated to augment VMRs in rats which experienced NBI to a greater magnitude than the same manipulation augments VMRs in rats which did not experience NBI61. Interestingly, in rats which experienced NBI, AFS continues to augment VMRs despite the presence of lesions of the amygdala that were sufficient to abolish augmentation in rats which did not experience NBI62. This indicates that the amygdala is not needed for the “extra” augmenting effects observed to be induced by NBI in rats experiencing AFS. Clearly, there is plenty we do not know about the effect of NBI on supraspinal processing related to bladder sensation that are in need of investigation.
Novel Systemic Therapeutics Tested in the NBI Model. Neuroplastic changes that occur secondary to NBI is the focus of this review, but we would be remiss if we did not also mention the use of the NBI model to test potential therapeutics. These have included pharmacological agents administered systemically (as opposed to intrathecally, that was previously discussed) that have found a role in the treatment of other bladder disorders or in the treatment of pain. Mirabegron and CL316243, b3 adrenoceptor agonists, both inhibited VMRs to UBD in rats which had experienced both NBI and ABI as did the a1 adrenoceptor antagonist, tamsulocin73. The muscarinic antagonist oxybutynin and the antidepressant amitriptyline had no effect on the VMRs of these rats73. In contrast, the benzodiazepines diazepam and midazolam both produced dose-dependent inhibition of VMRs of these rats58 as did the intravenous infusion of the general anesthetic propofol73. Notably, differential effects of these drug treatments in NBI-treated versus controls was not performed.
Other novel therapeutics which have been studied are those which use electrical neuromodulation. In rodent models, such treatments produce antinociceptive effects when rats have been treated with NBI and ABI, but not in control animals. Spinal cord stimulation80 and bilateral Pudendal Nerve Stimulation78 (bPNS) have both found efficacy in reducing VMRs to UBD in NBI-treated rats, but not in their controls. In contrast, bilateral T13 or L6 spinal root stimulation were not similarly effective78. As in any investigation, not all options are effective for all types of pain, so it is particularly notable that bladder hypersensitivity induced by either ABI without previous NBI or by chronic footshock stress were not affected by bPNS. The inhibitory neuromodulatory effects were exclusive to the NBI model. The limited number of studies of therapeutics done-to-date suggest there is fertile ground available for research using the NBI model to study new drugs or other interventions.
Conclusions
Bladder inflammation at a critical period of development in female rats has been demonstrated to produce multiple neuroplastic changes that persist into adulthood. The critical period of development is the time period where bulbospinal inhibitory influences are being established103. The present article has reviewed the literature related to NBI. The changes that have been observed are summarized in Figure 4. These changes have many correlates in humans with the disorder IC which are compared in Table 4. Given the many similarities, this supports the assertion that the NBI model in female rats is a valid model of IC. This NBI model requires a sustained bout of bladder inflammation in the third week of life as a necessary but not sufficient event, which when coupled with a second pro-nociceptive insult, as an adult, results in phenotypic features of IC. Bladder hypersensitivity appears to occur due to a combination of deficits in inhibitory mechanisms and an augmentation of facilitatory mechanisms which include mechanisms using glutamatergic and CRFR2-related receptors. Human epidemiological evidence related to the frequency of infant urinary tract infections and their increased history in subjects who develop IC as adults88 gives additional face value to the model and support that it has utility for testing new therapeutics for IC.
Material and Methods
Common methodology
General. All studies were approved by the University of Alabama at Birmingham’s Institutional Animal Care and Use Committee and followed the ethical guidelines of the International Association for the Study of Pain124. To obtain female rat pups, timed pregnant females were obtained from Envigo Laboratories (Formerly Harland, Indianapolis, IN) and date of birth verified by daily observation of cages. All rats were raised to 12-15 weeks of age. Female rats were utilized for both methodological reasons48 including the relative ease of bladder cannulation in females, and due to the greater prevalence of IC in females versus males13.
Neonatal Bladder Inflammation (NBI). The standard protocol for NBI consisted of anesth-etizing female rat pups on days P14-16 using a potent inhalational agent (halothane or isoflurane, 2-3% in oxygen), injecting ampicillin (50-100 mg/kg s.c.), swabbing the urethral orifice with an iodine/povidone solution, placing a transurethral intravesical catheter (24 gauge angiocath), infusing 0.1 ml of a 1% solution of zymosan A (Sigma Aldrich, St. Louis, MO) in saline and allowing the solution to dwell for 30 minutes while keeping the rat warm on a heating blanket; followed by recovery with return to their home cage. Zymosan was used as an intravesical inflammatory agent due to our extensive characterization of responses to this substance94 and our previous demonstration that NBI induced by other substances such as bacterial LPS do not produce the same long-lasting effects on reflex responses to UBD61. Primary characterization studies did not observe any statistical differences between rats which received intravesical saline as step above95 and in an attempt to avoid any non-specific inflammatory events, the neonatal control treatments for all studies described here consisted of steps [1]-above, followed by maintenance of anesthesia for 30 minutes and return to home cages.
Protocol for Adult Bladder Inflammation (ABI). Adult bladder inflammation (ABI) also followed a standard protocol in these studies which consisted of anesthetizing female rats approximately 24 hours prior to subsequent testing using a potent inhalational agent (halothane or isoflurane, 2-3% in oxygen), injecting ampicillin (50-100 mg/kg s.c.), swabbing the urethral orifice with an iodine/povidone solution, placing a transurethral intravesical catheter (22 gauge angiocath), infusing 0.5 ml of a 1% solution of zymosan A (in saline; Sigma Aldrich, St. Louis, MO) or other inflammatory compound and allowing the solution to dwell for 30 minutes while keeping the rat warm on a heating blanket; followed by recovery with return to their home cage. Unless otherwise stated, the term “ABI” indicates use of this standard protocol. Similar to NBI, characterization studies did not observe any statistical differences between rats which received intravesical saline as step and in an attempt to avoid any non-specific inflammatory events, the adult control treatments for these studies consisted of steps [1]-above, followed by maintenance of anesthesia for 30 minutes and return to home cages.
Visceromotor Response (VMR) Measures. Adult rats were initially anesthetized with 2-5% isoflurane followed by injection of urethane (1.2-1.4 gm/kg IP), and a 22 gauge angiocatheter was placed transurethrally into the bladder and held in place by a tight suture around the distal urethral orifice. A 7.8 cm catheter made of PE10 tubing was inserted via an incision in the atlanto-occipital membrane following surgical exposure and threaded down through the subarachnoid space to the lumbosacral region. Silver wire electrodes were placed in the left external oblique musculature immediately superior to the inguinal ligament. Isoflurane was then lowered until flexion reflexes were present (<1%). UBDs (20 s, 10-60 mm Hg) were produced using compressed air, and intravesical pressure was monitored using an in-line pressure transducer. Contraction of the abdominal musculature in response to UBD has been well characterized8,32 and was quantified as electromyographic (EMG) activity measured via the external oblique electrodes using standard differential amplification (Grass, Inc. P511 AC amplifiers; 50 x amplification, 60 Hz clipping, low filter setting 10 Hz – high filter setting 3 kHz). The analog EMG signal was digitized with a sampling rate of 10 kHz and saved on computer with digital rectification via software (Spike 4 software; Cambridge Electronic Design Limited, Cambridge, UK) allowing for calculation of mean EMG activity (in mV) during any defined time period. Notably, for each data set the same amplifier and filter settings were used for all rats. After a 15 minute waiting period, three repeated 20 second duration UBDs at 60 mm Hg intensity were presented at 3 minute intervals and then followed by the measurement of responses to graded constant pressure stimuli (10-60 mmHg, 20 seconds, 1-minute intertrial intervals).
Quantitative Analysis of VMRs. The vigor of VMRs is expressed as “Visceromotor Response” which is defined as increases in mean EMG activity during the 20 s period of UBD over the immediate pre-stimulus level of activity. Logistically, during each experimental run, the differential amplifier output is digitized and rectified by computer software (Spike 2, Cambridge Electronic Design, Inc., Cambridge UK) to positive values, which are then averaged over a period of time to give a mean voltage measure of amplified EMG activity. Measures of the Visceromotor Response for each distending pressure were then treated as discrete data points and analyzed using a repeated measures ANOVA for each pretreatment group. We have previously demonstrated that discrete Visceromotor Response measures are normally distributed61. In addition, when utilizing multiple different intensities of UBD, a stimulus-response function can be generated which can then be associated with an Area-Under-the-Curve statistic (AUC measure). A more extensive demonstration of this analysis, as well as a graphical description of the experimental apparatus related to VMRs is given in Ness et al. 61 AUC measures have also been demonstrated to be normally distributed61.
Statistics. Data are reported as means + SEM unless otherwise stated. For statistical analyses, the SYSTAT 12 (SPSS, Inc. San Jose, CA, United States) software package was utilized. One-way or two-way ANOVAs were used to identify treatment effects with Tukey’s HSD for post hoc pairwise comparisons unless otherwise stated.
Gap #1 Specific Study Methods. In order to allow a direct comparison, much of the methodology of these studies is identical to our previous reports which used either bladder re-inflammation as an adult insult or hindlimb inflammation64,65. Anesthetics, drug doses, timing and other variables were therefore chosen to be consistent with that other report. Studies were performed in 171 female Sprague-Dawley rats. All rats underwent NBI or control pretreatments on days P14, P15 and P16 using the standard NBI protocol. As adults (12-15 weeks of age) female rats were anesthetized with 2-5% isoflurane in oxygen, injected with ampicillin (50-100 mg/kg IP) and the skin at the site of injection was painted with an iodine-povidone solution. Complete Freund’s Adjuvant (CFA) pretreatment consisted of 100 ml of CFA (Sigma Aldrich, St. Louis, MO) injected into the subcutaneous tissues of one of four sites using a 30-gauge needle: the left or right hindpaw’s dorsolateral surface; the right forepaw’s dorsolateral surface, or; the right cheek subcutaneous area. Control treatments for CFA consisted of similar anesthetic, iodine swabbing and antibiotic treatment and in most cases 100 ml of normal saline was injected. After pretreatment, rats were kept warm on a heating blanket, allowed to recover and returned to their home cages. CFA/Control treatments occurred only once and were performed approximately 72 hours prior to additional testing described below. Gross examination of the CFA-injected sites demonstrated them all to be swollen and red and sensitive to probing with a cotton-tipped applicator. On the day of the experiment, VMRs to UBD were obtained as described above.
Gap #2 Specific Study Methods. Thirty-five female rats (half NBI-treated, half neonatal control treated), as adults underwent induction of ABI or control pretreatment according to the protocols described above. 24 hours later they were deeply anesthetized with 5% isoflurane and then euthanized via decapitation. Bladders were removed and processed according to ELISA kit instructions (LSBio kits: LS-F542; Lifespan Biosciences. Seattle, WA, USA). Protein concentrations were determined using the Pierce BCA Assay Reagent Kit (Thermo Fisher Scientific, Rockford, IL USA). Samples and serial dilutions of standards were processed according to kit protocols. A one way ANOVA was used to identify treatment effects.
Gap #3 Specific Study Methods. 21 rats underwent NBI and/or ABI pretreatments according to the standard protocols described above. Bladders were removed and drop-fixed in 4% paraformaldehyde overnight and cryoprotected in 30% sucrose. They were then processed for histology and 40 mm thick frozen sections were obtained as “rings” from the base of the bladder using a Leica CM1850 UV cryostat. Adjacent sections were processed for immunocytochemical labeling for SP or CGRP. Primary antisera were either rabbit anti-SubP antibody (1:1000; ImmunoStar cat. #20064) or rabbit anti-CGRP antibody (1:2000; ImmunoStar cat. #24112). Secondary antibodies used were Alexa 568-conjugated goat anti-rabbit antibody (1:1000; Jackson Immunoresearch code #111-005-003). Sections were mounted on slides, incubated in the primary antiserum for overnight at 4oC, and incubated with the secondary antibody. The specificity of the immunocytochemical staining was verified in control experiments that excluded the primary antiserum for the processing and using adsorption control experiments. Slides were then examined with a MicroBrightField system interfaced with an upright Olympus BX-51 microscope using narrow band cubes for Alexa 568 (DM 568, excitation filter 540–560, barrier filter 575–645 nm). Images were captured with a high resolution color digital camera. The same time of exposure, gain and gamma adjustment for the control and the experimental samples for each fluorescence staining was used. Quantification of the labeling used a procedure similar to our previous studies16,17,37. Tissue was analyzed by an observer blinded to pretreatment group using Image J software. Areas consisting of the urothelium and immediately adjacent submucosal tissues were outlined using a selection tool, as was an adjacent full thickness sample of deeper bladder tissues out from the urothelial sample site to the serosa. The density of fluorescent activity (pixels/area) was measured for both sites and a ratio comparing the urothelial versus non-urothelial fluorescent density determined. Data was subsequently unblinded and quantitative comparisons performed with ANOVAs and post hoc tests as appropriate.
Gap #4 – Specific Study Methods. 32 female rats underwent NBI and/or ABI protocols or control treatments as described above and were subsequently placed into either a “Distended” or “Not Distended” group such that there were 4 rats in each of 8 groups. As adults, these rats were anesthetized with inhaled isoflurane (5% for induction, 4% for maintenance throughout surgery). The trachea was cannulated for artificial respiration. In animals in the Distended group, the urinary bladder was catheterized with a 22-gauge angiocatheter via the urethra and held in place with a tight suture tied around the distal urethra orifice. The rat was moved to a recording area where it was artificially respirated and body temperature was maintained at approximately 37.0˚C with a heating pad. The anesthesia level was lowered to 1% isoflurane and maintained for 10 minutes. The bladder was then distended via air pressure according to the following sequence: 3 x 20 s, 60 mmHg distensions, followed by graded distensions of 10, 20, 30, 40, 50, 60, 70, and 80 mmHg, respectively. The intertrial interval was 3 minutes. The rats were maintained at 2% isoflurane for 90 minutes to allow for c-Fos expression. Rats in the Not Distended group were not cannulated and otherwise maintained in a warm, anesthetized state for the same period of time. At the end of experimental procedure, the anesthesia level was increased to 4% for 5 minutes, after which time the animal was killed by transcardial perfusion with saline followed by 4% formalin. The spinal cord was removed by hydraulic extrusion. Tissue was postfixed in 4% formalin and cryoprotected in 4% formalin and 12% sucrose followed by 4% formalin and 25% sucrose, each for 24 hours, after which time 40 µm sections of T12-S2 were cut using a Leica CM1850 UV cryostat. To avoid double counting cells, nonconsecutive sections were selected. Tissue was processed using an avidin-biotinylated enzyme complex (Vectastain® ABC Kit, Rabbit IgG) and visualized using the Vector® VIP Peroxidase Kit. Both kits were acquired from Vector Laboratories (Burlingame, CA). The protocol was as follows: Tissue was washed three times with phosphate-buffered saline (PBS) for a total time of at least 5 minutes, incubated in a solution of 0.3% hydrogen peroxide and 0.3% normal sera for 5 minutes, washed twice with PBS for a total of 5 minutes, incubated in a solution of 1.5% normal serum 0.2% triton for 20 minutes, and incubated in a solution of polyclonal rabbit anti-c-fos primary antibody (1:20,000; Ab-5, Calbiochem, La Jolla, CA USA ) and 0.2% triton overnight at room temperature. The next day, two washes in PBS for a total of 5 minutes and incubation in anti-goat biotinylated secondary antibody (1:200) with 1.5% normal serum for 30 minutes were performed, followed by 2 washes in PBS for a total of 5 minutes, incubation in reagent for 30 minutes, and incubation in chromagen for 6 minutes for visualization. Finally, sections were transferred to tap water, mounted, and cover slipped using Fisher Scientific Permount Mounting Medium (Fisher Scientific, Pittsburg, PA USA).
3 slices from each section, L5, L6 and S1, out of an average of 10 slices per section were randomly selected from each animal. Pictures were taken using a Leica microscope and a Hamasura camera at 10x magnification and examined using ImagePro-Plus (Media Cybernetics, Silver Springs, MD, USA). Analysis was carried out using ImageJ software (U.S. National Institutes of Health, Bethesda, MD, USA). The number of c-Fos positive cells was counted in four sections for each slice: the dorsal commissure (DCM), lateral lamina V-VII including the sacral parasympathetic nucleus in L6 and S1 sections and the lateral spinal nucleus in L5 sections, the medial dorsal horn (MDH), and the lateral dorsal horn (LDH), as described by Vizzard117. Cells with a mean coloration 2 times darker than background were counted as c-Fos positive by two observers blind to the experimental conditions. The average of the counts of the two observers was taken and considered as the number of c-Fos labelled neurons in each section for each slice. The average and standard error of the four animals in each condition was calculated and a three-way analysis of variance was conducted to test for differences between groups and interactions between neonatal treatment, adult treatment, and distension. To assess the effect of neonatal insult, adult inflammation, and distension separately, separate one-way ANOVAs were conducted for each manipulation. Post hoc analyses were conducted as appropriate.
Gap #5 Study Specific Methods. For these studies, all 32 rats (8/group x 4 groups) experienced the standard NBI protocol. Half of those experienced the standard ABI protocol while the other half received the adult control treatment 24 hours prior to testing. On the day of the experiment, they were anesthetized and prepared for VMR measures per the above protocol. Prior to the presentation of any UBDs, the intrathecal catheter was dosed with one of the following in a 10 l volume followed by a 10 l normal saline flush: normal saline; or 15 mg of Urocortin 2 peptide (Sigma-Aldrich, St. Louis, MO, USA). A 15 minute waiting period occurred and then five 20 s, 60 mm Hg UBDs were administered at 3 minute intervals followed by graded 20 s UBDs administered in ascending order of 10, 20, 30, 40, 50 and 60 mm Hg. VMRs were measured and quantified. Statistical analysis consisted of two-way, repeated measures ANOVAs.
Gap #6 Specific Study Methods. The methods used in the present study mirror those of our published study66 that examined the interaction between acute footshock (AFS) and NBI so that a comparison could be made to that study. In all, a total of 121 adult, female Sprague-Dawley rats were studied which were raised from birth from timed-pregnant dams (Harlan/Envigo Laboratories, Sprattville, AL) and received either NBI or neonatal control treatments. They were then tested at 12-15 weeks of age. As adults they received AFS or control treatments. AFS was administered in an operant conditioning chamber similar to our previous studies21,61. All rats were placed in the chamber for a 15-min accommodation session for a total of six sessions in the days prior to final testing. On the day of testing, rats to be pretreated with AFS were placed in the chamber and received intermittent footshock (1.0 mA, 1 sec duration, total of 30 shocks in 15 min) administered via a parallel rod floor on a variable-interval schedule. Control pretreatments consisted of placement in the chamber for 15 min without shocks. Rats were then immediately anesthetized for either VMR measures or tissue harvest. Those for the latter procedure were deeply anesthetized with 5% isoflurane and then euthanized via decapitation. Spinal cords were hydraulically extracted and lumbosacral (L6-S2) segments were isolated and processed according to ELISA kit instructions. Spinal tissues from 5 rats/group were used for the neurochemical study. Protein concentrations were determined using the Pierce BCA Assay Reagent Kit (Thermo Fisher Scientific, Rockford, IL). CRFR1 and CRFR2 contents were quantified using LSBio kits [Lifespan Biosciences, Seattle, WA]. Samples and serial dilutions of standards were processed according to kit protocols. The other 101 rats underwent VMR testing per the protocol described above. Four rats which demonstrated neurological deficits after intrathecal catheter insertion were discarded from additional study. The following drugs were administered IT in a 10 l volume followed by a 10 l normal saline flush: aSVG30 (12 g IT; Phoenix Pharmaceuticals, Inc., Burlingame, CA USA), antalarmin (24 g IT in water/9% DMSO vehicle; Sigma-Aldrich, St. Louis, MO USA) or normal saline. 15 minutes later VMRs to five repeated distensions (60 mmHg, 20 s, 3 min interval) followed by graded distensions (10-60 mm Hg, 20 s, 1 min intervals) were measured. A two-way ANOVA was utilized to analyze ELISA data. A repeated-measures two-way ANOVA was used to compare graded VMR data. Tukey’s HSD was used for post hoc tests.
Funding Information
These studies were supported by DK51413.
Acknowledgements
The technical efforts and expertise of Dr. Alan Randich, Dr. Amber Shaffer and Dr. Chelsea Ball in blinded histological analyses are gratefully acknowledged.
Author Contributions
TJN was involved in data collection, data analysis, drafting of the manuscript and has given final approval of its form. CD was involved in data collection, data analysis, editing of the manuscript and has given final approval of its form
Conflict of Interest
The authors declare no conflicts of interest.
Ethics Statements
All studies were approved by the University of Alabama at Birmingham’s Institutional Animal Care and Use Committee and followed the ethical guidelines of the International Association for the Study of Pain124.
References
- Abreu-Mendes P, Costa A, Charrua A, Almeida Pinto R, Cruz F. The role of urinary VEGF in observational studies of BPS/IC patients: a systematic review. Diagnostics (Basel) 2022; 12:1037. [CrossRef]
- Afari N, Buchwald D, Clauw D, Hong B, Hou X, Krieger JN, Mullins C, Stephens-Shields AJ, Gasperi M, Williams DA; MAPP Research Network. A MAPP network case-control study of urological chronic pelvic pain compared with nonurological pain conditions. Clin J Pain 2020; 36:8-15. [CrossRef]
- Akiyama Y and Hanno P. Phenotyping of interstitial cystitis/bladder pain syndrome. Int J Urol 2019; 1: 17-19. [CrossRef]
- Al-Chaer ED, Kawasaki M, Pasricha TJ. A new model of chronic visceral hypersensitivity in adult rats induced by colon irritation during postnatal development. Gastroenterology 2000; 119: 1276-128. [CrossRef]
- Applebaum AE, Vance WH, Coggeshall RE. Segmental localization of sensory cells that innervate the bladder. J Comp Neurol 1980; 192:203-209. [CrossRef]
- Archer AC, DeBerry JJ, DeWitte C, Ness TJ. Neonatal cystitis makes adult female rat urinary bladders more sensitive to low concentration microbial antigens. Res Rep Urol 2023; 15: 531-539. [CrossRef]
- Ball CL, Ness TJ and Randich A. Opioid blockade and inflammation reveal estrous cycle effects on visceromotor reflexes evoked by bladder distension. J Urol 2010; 184: 1529-1535. [CrossRef]
- Beggs S, Currie G, Salter MW, Fitzgerald M, Walker SM. Priming of adult pain responses by neonatal pain experience: maintenance by central neuroimmune activity. Brain 2012; 135:404-417. [CrossRef]
- Blatt LK, Lashinger ES, Laping NJ and Su X. Evaluation of pressor and visceromotor reflex responses to bladder distension in urethane anesthetized rats. Neurourol Urodyn 2009; 28: 442-446. [CrossRef]
- Bullones Rodríguez MÁ, Afari N, Buchwald DS; National Institute of Diabetes and Digestive and Kidney Diseases Working Group on Urological Chronic Pelvic Pain. Evidence for overlap between urological and nonurological unexplained clinical conditions. J Urol 2013; 189(1 Suppl): S66-74.
- Butler RK, Finn DP. Stress-induced analgesia. Prog Neurobiol 2009; 88: 184-202. [CrossRef]
- Castroman P, Ness TJ. Vigor of visceromotor responses to urinary bladder distension in rats increases with repeated trials and stimulus intensity. Neurosci Letts 2001; 306:97-100. [CrossRef]
- Clemens JQ, Clauw DJ, Kreder K, Krieger JN, Kusek JW, Lai HH, Rodriguez L, Williams DA, Hou X, Stephens A, Landis JR; MAPP Research Network. Comparison of baseline urological symptoms in men and women in the MAPP research cohort. J Urol 2015; 193:1554-1558. [CrossRef]
- Clemens JQ, Meenan RT, O’Keeffe Rosetti MC, Gao SY, Calhoun EA. Prevalence and incidence of interstitial cystitis in a managed care population. J Urol 2005; 173: 98-102. [CrossRef]
- Clemens JQ, Mullins C, Ackerman AL, Bavendam T, van Bokhoven A, Ellingson BM, Harte SE, Kutch JJ, Lai HH, Martucci KT, Moldwin R, Naliboff BD, Pontari MA, Sutcliffe S, Landis JR; MAPP Research Network Study Group. Urologic chronic pelvic pain syndrome: insights from the MAPP Research Network. Nat Rev Urol. 2019; 16:187-200. [CrossRef]
- Clodfelder-Miller B, DeBerry JJ, Ness TJ. Urothelial bladder afferents selectively project to L6/S1 levels and are more peptidergic than those projecting to the T13/L1 levels in female rats. Heliyon 2023; 2023: e18495. [CrossRef]
- Clodfelder-Miller BJ, Kanda H, Gu J, Creighton JR, Ness TJ and Deberry JJ. Urothelial bladder afferent neurons in the rat are anatomically and neurochemically distinct from non-urothelial afferents. Brain Res 2018; 1689: 45-53. [CrossRef]
- Clodfelder-Miller B, Ness TJ and DeBerry JJ. Neonatal bladder inflammation results in adult female mouse phenotype with increased frequency and nociceptive responses to bladder filling. Front Syst Neurosci. 2022; 16: 858220. [CrossRef]
- Deberry JJ, Ness TJ, Robbins MT, Randich A. Inflammation-induced enhancement of the visceromotor reflex to urinary bladder distention: Modulation by endogenous opioids and the effects of early-in-life experience with bladder inflammation. J Pain 2007; 8:914-23. [CrossRef]
- Deberry JJ, Randich A., Shaffer A., Robbins M, and Ness TJ. Neonatal bladder inflammation produces functional changes and alters neuropeptide content in bladders of adult female rats. J Pain 2010; 11:247-255. [CrossRef]
- DeBerry JJ, Robbins MT, Ness TJ The amygdala central nucleus is required for acute stress-induced bladder hyperalgesia in a rat visceral pain model. Brain Res 2015; 1606:77-85. [CrossRef]
- DeGroat WC, Araki I, Vizzard MA. Developmental and injury induced plasticity in the micturition reflex pathway. Behav Brain Res 1998: 92: 127-140. [CrossRef]
- Deutsch G, Deshpande H, Frölich MA, Lai HH, Ness TJ. Bladder distension increases blood flow in pain related brain structures in subjects with interstitial cystitis. J Urol 2016; 196: 902-10. [CrossRef]
- Deutsch G, Deshpande H, Lai HH, Kutch JJ, Ness TJ. Cerebral perfusion and sensory testing results differ in interstitial cystitis/bladder pain syndrome patients with and without fibromyalgia: a site-specific MAPP Network study. J Pain Res 2021; 14: 3887-3895. [CrossRef]
- Ding X, Liang Y-J, Su L, Liao F-Ff, ang D, Tai J, Xing G-G. BDNF contributes to the neonatal incision-induced facilitation of spinal long-term potentiation and the exacerbation of incisional pain in adult rats. Neuropharm 2018; 137: 114-132. [CrossRef]
- Farmer MA, Huang L, Martucci K, Yang CC, Maravilla KR, Harris RE, Clauw DJ, Mackey S, Ellingson BM, Mayer EA, Schaeffer AJ, Apkarian AV; MAPP Research Network. Brain white matter abnormalities in female interstitial cystitis/bladder pain syndrome: A MAPP Network Neuroimaging Study. J Urol.2015; 194:118-26. [CrossRef]
- Fitzgerald M, Koltzenberg M. The functional development of descending inhibitory pathways in the dorsolateral funiculus of the newborn rat spinal cord. Brain Res. 1986; 389: 261-270. [CrossRef]
- Gold MS, Zhang L, Wrigley DL, Traub RJ. Prostaglandin E(2) modulates TTX-RI(Na) in rat colonic sensory neurons. J Neurophysiol 2002; 88:1512-1522. [CrossRef]
- Griffith JW, Stephens-Shields AJ, Hou X, Naliboff BD, Pontari M, Edwards TC, Williams DA, Clemens JQ, Afari N, Tu F, Lloyd RB, Patrick DL, Mullins C, Kusek JW, Sutcliffe S, Hong BA, Lai HH, Krieger JN, Bradley CS, Kim J, Landis JR. Pain and urinary symptoms should not be combined into a single score: psychometric findings from the MAPP Research Network. J Urol 2016; 195: 949-54. [CrossRef]
- Gupta A, Bhatt RR, Naliboff BD, Kutch JJ, Labus JS, Vora PP, Alaverdyan M, Schrepf A, Lutgendorf S, Mayer EA; MAPP Research Network. Impact of early adverse life events and sex on functional brain networks in patients with urological chronic pelvic pain syndrome (UCPPS): A MAPP Research Network study. PLoS One 2019;14: e0217610. [CrossRef]
- Gupta P, Gallop R, Spitznagle T, Lai H, Tu F, Krieger JN, Clemens JQ, Bradley CS, Yang C, Sutcliffe S, Moldwin R, Kreder K, Kutch J, Rodriguez LV. Is pelvic floor muscle tenderness a distinct urologic chronic pelvic pain syndrome phenotype? Findings from the MAPP Research Network symptom pattern study. J Urol 2022; 208: 341-349. [CrossRef]
- Harte SE, Schrepf A, Gallop R, Kruger GH, Lai HHH, Sutcliffe S, Halvorson M, Ichesco E, Naliboff BD, Afari N, Harris RE, Farrar JT, Tu F, Landis JR, Clauw DJ; MAPP Research Network. Quantitative assessment of nonpelvic pressure pain sensitivity in urologic chronic pelvic pain syndrome: a MAPP Research Network study. Pain 2019; 160: 1270-1280. [CrossRef]
- Hsieh C-T, Lee Y-J, Dai X, Ojeda NB, Lee HJ, Tien L-T, Fan L-W. Systemic lipopolysaccharide-induced pain sensitivity and spinal inflammation were reduced by minocycline in neonatal rats. Int J Mol Sci 2018; 19:2947. [CrossRef]
- Huang L, Kutch JJ, Ellingson BM, Martucci KT, Harris RE, Clauw DJ, Mackey S, Mayer EA, Schaeffer AJ, Apkarian AV, Farmer MA; MAPP Research Network. Brain white matter changes associated with urological chronic pelvic pain syndrome: multisite neuroimaging from a MAPP case-control study. Pain 2016; 157: 2782-2791. [CrossRef]
- Ji N-N, Meng Q-X, Wang Y, Zhou Z-M, Song Y, Hua R, Zhang Y-M. Microglia-derived TNF-a inhibiting GABAergic neurons in the anterior lateral bed nucleus of the stria terminalis precipitates visceral hypersensitivity induced by colorectal distension in rats. Neurobio Stress 2022; 18: 100449. [CrossRef]
- Kairys AE, Schmidt-Wilcke T, Pulu T, Ichesco E, Labus JS, Martucci K, Farmer MA, Ness TJ, Deutsch G, Mayer EA, Mackey S, Apkarian AV, Maravilla K, Clauw DJ, Harris RE. Increased brain gray matter in the primary somatosensory cortex is associated with increased pain and mood disturbance in patients with interstitial cystitis/painful bladder syndrome. J Urol. 2015; 193:131-7. [CrossRef]
- Kanda H, Clodfelder-Miller BJ, Gu JG, Ness TJ, DeBerry JJ. Electrophysiological properties of lumbosacral primary afferent neurons innervating urothelial and non-urothelial layers of mouse urinary bladder. Brain Res 2016; 1648:81-9. [CrossRef]
- Kannampalli P, Babygirija R, Zhang J, Poe MM et al. Neonatal bladder inflammation induces long-term visceral pain and altered responses of spinal neurons in adult rats. Neuroscience 346: 349-364, 2017. [CrossRef]
- Kilpatrick LA, Kutch JJ, Tillisch K, Naliboff BD, Labus JS, Jiang Z, Farmer MA, Apkarian AV, Mackey S, Martucci KT, Clauw DJ, Harris RE, Deutsch G, Ness TJ, Yang CC, Maravilla K, Mullins C, Mayer EA. Alterations in resting state oscillations and connectivity in sensory and motor networks in women with interstitial cystitis/painful bladder syndrome. J Urol 2014; 192: 947-55. [CrossRef]
- Krieger JN, Stephens AJ, Landis JR, Clemens JQ, Kreder K, Lai HH, Afari N, Rodríguez L, Schaeffer A, Mackey S, Andriole GL, Williams DA; MAPP Research Network. Relationship between chronic nonurological associated somatic syndromes and symptom severity in urological chronic pelvic pain syndromes: baseline evaluation of the MAPP study. J Urol 2015;193: 1254-62. [CrossRef]
- Kutch JJ, Ichesco E, Hampson JP, Labus JS, Farmer MA, Martucci KT, Ness TJ, Deutsch G, Apkarian AV, Mackey SC, Klumpp DJ, Schaeffer AJ, Rodriguez LV, Kreder KJ, Buchwald D, Andriole GL, Lai HH, Mullins C, Kusek JW, Landis JR, Mayer EA, Clemens JQ, Clauw D, Harris RE, MAPP Research Network. Brain signature and functional impact of centralized pain: a multidisciplinary approach to the study of chronic pelvic pain (MAPP) network study. Pain 2017; 158:1979-1991. [CrossRef]
- Lai HH, Gardner V, Ness TJ, Gereau RW 4th. Segmental hyperalgesia to mechanical stimulus in interstitial cystitis/bladder pain syndrome - evidence of central sensitization. J Urol 2014; 191:1294-9. [CrossRef]
- Lai H, Gereau RW 4th, Luo Y, O'Donnell M, Rudick CN, Pontari M, Mullins C, Klumpp DJ. Animal models of urologic chronic pelvic pain syndromes: findings from the multidisciplinary approach to the study of chronic pelvic pain research network. Urology 2015; 85: 1454-65. [CrossRef]
- Lai HH, Jemielita T, Sutcliffe S, Bradley CS, Naliboff B, Williams DA, Gereau RW 4th, Kreder K, Clemens JQ, Rodriguez LV, Krieger JN, Farrar JT, Robinson N, Landis JR; MAPP Research Network. Characterization of whole-body pain in urological chronic pelvic pain syndrome at baseline: A MAPP Research Network Study. J Urol 2017; 198: 622-631. [CrossRef]
- Lai HH, Krieger JN, Pontari MA, Buchwald D, Hou X, Landis JR; MAPP Research Network. Painful bladder filling and painful urgency are distinct characteristics in men and women with urological chronic pelvic pain syndromes: A MAPP Research Network Study. J Urol 2015; 194: 1634-41. [CrossRef]
- Lai HH, Newcomb C, Harte S, Appleby D, Ackerman AL, Anger JT, Nickel JC, Gupta P, Rodriguez LV, Landis JR, Clemens JQ; MAPP Research Network. Comparison of deep phenotyping features of UCPPS with and without Hunner lesion: A MAPP-II Research Network Study. Neurourol Urodyn 2021; 40: 810-818. [CrossRef]
- Lai HH, North CS, Andriole GL, Cupps L, Song D, Ness TJ, Hong BA. Urological symptoms in a subset of patients with urological chronic pelvic pain syndrome and a polysymptomatic, polysyndromic pattern of presentation. J Urol 2014; 191: 1802-7. [CrossRef]
- LaPrairie JL, Murphy AZ. Female rats are more vulnerable to the long-term consequences of neonatal inflammatory injury. Pain 2007; 132: S124-S133. [CrossRef]
- Lutgendorf SK, Zia S, Luo Y, O'Donnell M, van Bokhoven A, Bradley CS, Gallup R, Pierce J, Taple BJ, Naliboff BD, Quentin Clemens J, Kreder KJ, Schrepf A. Early and recent exposure to adversity, TLR-4 stimulated inflammation, and diurnal cortisol in women with interstitial cystitis/bladder pain syndrome: A MAPP research network study. Brain Behav Immun 2023; 111: 116-123. [CrossRef]
- Mazeaud C, Rigaud J, Levesque A et al. Stratification of patients with interstitial cystitis-bladder pain syndrome according to the anatomical bladder capacity. Urology 2019; 123: 87-92. [CrossRef]
- Miranda A, Mickle A, Bruckert M et al. NMDA receptor mediates chronic visceral pain induced by neonatal noxious somatic stimulation. Eur J Pharmacol 2014; 744: 28-35. [CrossRef]
- Miranda A, Mickle A, Schmidt J, Zhang Z et al. Neonatal cystitis-induced colonic hypersensitivity in adult rats: a model of viscero-visceral convergence. Neurogastroenterol Motil 2011; 23: 683-e281. [CrossRef]
- Moriarty O, Harrington L, Beggs S, Walker SM. Opioid analgesia and the somatosensory memory of neonatal surgical injury in the adult rat. Br J Anaesth 2018; 121: 314-324. [CrossRef]
- Moriarity O, Tu Y, Sengar AS, Salter MW, Beggs S, Walker SM. Priming of adult incision response by early-life injury: neonatal microglial inhibition has persistent but sexually dimorphic effects in adult rats. J Neurosci 2019; 39: 3081-3093. [CrossRef]
- Mukerji G, Waters J, Chessell IP, Bountra C, Agarwal SK, Anand P. Pain during ice water test distinguishes clinical bladder hypersensitivity from overactivity disorders. BMC Urol 2006; 6: 31. [CrossRef]
- Ness TJ, Castroman P. Evidence for two populations of rat spinal nociceptive neurons excited by urinary bladder distension. Brain Res 2001; 923:147-156. [CrossRef]
- Ness TJ, Castroman PJ, Randich A. Acute bladder inflammation differentially affects spinal visceral nociceptive neurons. Neurosci Lett 2009; 467:150-4. [CrossRef]
- Ness TJ, Clodfelder-Miller B, McNaught J, Miller DE and Su X. Benzodiazepines suppress neuromodulatory effects of pudendal nerve stimulation on rat bladder nociception. Anesth Analg, 2020; 130:1077-1084. [CrossRef]
- Ness TJ, Clodfelder-Miller B, McNaught J, Miller DE and Su X. Neuromodulatory effects of pudendal nerve stimulation on bladder hypersensitivity are present in opioid-pretreated rats" Reg Anesth Pain Med 2019; 44: 1015-1020. [CrossRef]
- Ness TJ, DeWitte C, DeBerry JJ. Spinal neurochemical mechanisms of acute stress-induced visceral hypersensitivity in healthy rats. Neurosci Letts 2022; 770: 136401. [CrossRef]
- Ness TJ, DeWitte C, DeBerry JJ, Hart MP, Clodfelder-Miller B, Gu JG, Ling J, Randich A. A model in female rats with phenotypic features similar to interstitial cystitis/bladder pain syndrome. Front Pain Res 2021; 7: 791045. [CrossRef]
- Ness TJ, DeWitte C, DeBerry JJ, Randich A. Neonatal bladder inflammation alters the role of the central amygdala in hypersensitivity produced by acute footshock stress in adult female rats. Brain Res. 2018; 1698: 99-105. [CrossRef]
- Ness TJ, DeWitte C, McNaught J, Clodfelder-Miller B and Su X. Spinal mechanisms of pudendal nerve stimulation-induced inhibition of bladder hypersensitivity in rats. Neurosci Letts 2018; 686: 181-185. [CrossRef]
- Ness TJ, DeWitte C, Randich A. Neonatal cystitis leads to alterations in spinal corticotropin releasing factor receptor-type 2 content and function in adult rats following bladder re-inflammation. Brain Res 2022; 1788: 147927. [CrossRef]
- Ness TJ, DeWitte C, Randich A. The double insult of neonatal cystitis plus adult somatic inflammation results in corticotropin releasing factor Type II receptor-dependent bladder hypersensitivity in female rats. J Pain 2022; 23: 2167-2178. [CrossRef]
- Ness TJ, DeWitte C, Robbins MT and DeBerry JJ. Neonatal cystitis alters mechanisms of stress-induced visceral hypersensitivity in rats. Neurosci Letts 2022; 778: 136617. [CrossRef]
- Ness TJ, Gebhart GF. Colorectal distension as a noxious visceral stimulus: physiologic and pharmacologic characterization of pseudaffective reflexes in the rat. Brain Res 1988; 450:153-169. [CrossRef]
- Ness TJ, Gebhart GF. Interactions between visceral and cutaneous nociception. I. Noxious cutaneous stimuli inhibit visceral nociceptive neurons and reflexes. J Neurophysiol 1991; 66:20-28. https://doi.org/10.1152/jn.1991.66.1.20. [CrossRef]
- Ness TJ, Gebhart GF. Acute inflammation differentially alters the activity of two classes of rat spinal visceral nociceptive neurons. Neurosci Lett 2000; 281:131-134. [CrossRef]
- Ness TJ, Gebhart GF. Inflammation enhances reflex and spinal neuron responses to noxious visceral stimulation in rats. Am J Physiol 2001; 280: G649-G657. [CrossRef]
- Ness TJ, Lewis-Sides A, Castroman P. Characterization of pressor and visceromotor reflex responses to bladder distension in rats: sources of variability and effect of analgesics. J Urol 2001; 165: 968-974.
- Ness TJ, Lloyd LK, Fillingim RB. An endogenous pain control system is altered in subjects with interstitial cystitis. J Urol 2014; 191: 364-70. [CrossRef]
- Ness TJ, McNaught J, Clodfelder-Miller B, Su X. Medications used to treat bladder disorders may alter effects of neuromodulation. Neurourol Urodyn 2020; 39:1313-1320. [CrossRef]
- Ness TJ, Richter HE, Varner RE, Fillingim RB. A psychophysical study of discomfort produced by repeated filling of the urinary bladder. Pain 1998; 76: 61-69. [CrossRef]
- Ness TJ, Powell-Boone T, Cannon R, Lloyd LK, Fillingiim RB. Psychophysical evidence of hypersensitivity in subjects with interstitial cystitis. J Urol 2005; 173: 1983-1987. [CrossRef]
- Ness TJ, Randich A. Which spinal cutaneous nociceptive neurons are inhibited by intravenous lidocaine in the rat? Reg Anesth Pain Med 2006; 31: 248-253. [CrossRef]
- Ness TJ, Randich A. Neonatal bladder inflammation alters activity of adult rat spinal visceral nociceptive neurons. Neurosci Lett 2010; 472: 210-214. [CrossRef]
- Ness TJ, Randich A, Nelson DE, Su X. Screening and optimization of nerve targets and parameters reveals inhibitory effect of pudendal stimulation on rat bladder hypersensitivity. Reg Anesth Pain Med. 2016; 41: 737-743. [CrossRef]
- Ness TJ, Randich A, Su X, DeWitte C and Hildebrand K. Systemic and intrathecal baclofen produce bladder antinociception in rats. BMC Urology 2021; 21: 139. [CrossRef]
- Ness TJ and Su X. Parametric assessment of spinal cord stimulation on bladder pain-like responses in rats. Neuromodulation 2022; 25: 1134-1140.
- Nickel JC, Stephens A, Landis JR, Mullins C, van Bokhoven A, Anger JT, Ackerman AL, Kim J, Sutcliffe S, Krol JE, Sen B, Hammond J, Ehrlich GD; Multidisciplinary Approach to the Study of Chronic Pelvic Pain (MAPP) Research Network. Urinary fungi associated with urinary symptom severity among women with interstitial cystitis/bladder pain syndrome (IC/BPS). World J Urol 2020; 38: 433-446.
- Otsuji J, Hayashi Y, Hitomi S, Soma C, Soma K, Shibuta I, Iwata K, Shirakawa T, Shinoda M. Decreased PPARgamma in the trigeminal spinal subnucleus caudalis due to neonatal injury contributes to incision-induced mechanical allodynia in female rats. Sci Reps 2022; 12: 19314. [CrossRef]
- Overholt, TL, Matthews CA, Evans RJ, BaDLANI g, Ahn C, Simon T, Walker SJ. Small fiber polyneuropathy is associated with non-bladder-centric interstitial cystitis/bladder pain syndrome patients. Female Pelvic Med Reconstr Surg 2021; 27: 581-585. [CrossRef]
- Pang X, Marchand J, Sant GR, Kream RM, Theoharides TC. Increased number of substance P positive nerve fibers in interstitial cystitis. J Urol 1995; 75: 744-750.
- Parsons CL. Potassium sensitivity test. Tech Urol 1996. 2:171-173.
- Park JS, Jung HD, Cho YS, Jin MH, Hong CH. Neonatal bladder irritation is associated with vanilloid receptor TRPV1 expression in adult rats. Int Neurourol J 2018; 22: 169-176. [CrossRef]
- Pattinson D and Fitzgerald M. The neurobiology of infant pain: development of excitatory and inhibitory neurotransmission in the spinal dorsal horn. Reg Anesth Pain Med 2004; 29: 36-44. [CrossRef]
- Peters KM, Killinger KA and Ibrahim IA. Childhood symptoms and events in women with interstitial cystitis/painful bladder syndrome. Urology 2009; 73: 258-62. [CrossRef]
- Pierce J, Harte SE, Afari N, Bradley CS, Griffith JW, Kim J, Lutgendorf S, Naliboff BD, Rodriguez LV, Taple BJ, Williams D, Harris RE, Schrepf A. Mediators of the association between childhood trauma and pain sensitivity in adulthood: a MAPP Research Network analysis. Pain 2023; 164: 1995-2008.
- Plair A, Evans RJ, Langefeld CD, Matthews CA, Badlani G, Walker SJ. Anesthetic bladder capacity is a clinical biomarker for interstitial cystitis/bladder pain syndrome subtypes. J Urol 2021 158: 74-80. [CrossRef]
- Randich A, DeWitte C, DeBerry JJ, Robbins MT, and Ness TJ. Lesions of the central amygdala and ventromedial medulla reduce bladder hypersensitivity produced by acute but not chronic foot shock. Brain Res 2017; 1675: 1-7. [CrossRef]
- Randich A, Mebane H, Deberry JJ, Ness TJ. Rostral ventral medulla modulation of the visceromotor reflex evoked by urinary bladder distension in female rats. J Pain 2008; 9: 920-926. [CrossRef]
- Randich A, Mebane H, Ness TJ. Ice water testing reveals hypersensitivity in adult rats that experienced neonatal bladder inflammation: Implications for painful bladder syndrome/interstitial cystitis. J Urol 2009; 182: 337-42. [CrossRef]
- Randich A, Uzzell TW, Cannon RS and Ness TJ. Inflammation and enhanced nociceptive responses to bladder distension produced by intravesical zymosan in the rat. BMC Urol 2006; 6:2. [CrossRef]
- Randich A, Uzzell TW, DeBerry JJ and Ness TJ. Neonatal urinary bladder inflammation produces adult bladder hypersensitivity. J Pain 2006; 7: 469-79. [CrossRef]
- Robbins MT, DeWitte C, Ness TJ. Stress-induced bladder hypersensitivity: effect of corticotropin releasing factor receptors assessed by spinal neurophysiology and neurochemistry. Neuropharmacology 2023; 224: 109369. [CrossRef]
- Robbins MT, Ness TJ. Footshock-induced urinary bladder hypersensitivity: role of spinal corticotrophin-releasing factor receptors. J Pain 2008; 9: 991-998.
- Robbins MT, Uzzell TW, Aly S and Ness TJ. Visceral nociceptive input to the area of the medullary lateral reticular nucleus ascends in the lateral spinal cord. Neurosci Lett 2005; 381: 329-333. [CrossRef]
- Robbins MT, Uzzell TW, Aly S, and Ness TJ. Characterization of thalamic neuronal responses to urinary bladder distension, including the effect of acute spinal lesions in the rat. J Pain 2006; 7: 218-224.
- Ruda MA, Ling QD, Hohmann AG, Peng YB, Tachibana T. Altered nociceptive neuronal circuits after neonatal peripheral inflammation. Science. 2000; 289:628-31. [CrossRef]
- Schachar JS, Evans RJ, Parks GE, Zambon J, Badlani G, Walker SJ. Histological evidence supports low anesthetic bladder capacity as a marker of a bladder-centric disease subtype ininterstitial cystitis/bladder pain syndrome. Int Urogynecol J 2019; 30: 1863-1870.
- Schrepf A, Naliboff B, Williams DA, Stephens-Shields AJ, Landis JR, Gupta A, Mayer E, Rodriguez LV, Lai H, Luo Y, Bradley C, Kreder K, Lutgendorf SK; MAPP Research Network. Adverse childhood experiences and symptoms of urologic chronic pelvic pain syndrome: A Multidisciplinary Approach to the Study of Chronic Pelvic Pain Research Network Study. Ann Behav Med 2018; 52: 865-877.
- Schwaller F and Fitzgerald M. The consequences of pain in early life: injury-induced plasticity in developing pain pathways. Eur J Neurosci 2014; 39: 344-352. [CrossRef]
- Sengupta JN, Pochiraju S, Kannampalli P, Bruckert M, Addya S, Yadav P, Miranda AM, Shaker R, Banerjee B. MicroRNA-mediated GABA Aa-1 receptor subunit down-regulation in adult spinal cord following neonatal cystitis-induced chronic visceral pain in rats. Pain 2013; 154: 59-70.
- Shaffer AD. “Early-in-Life Bladder Inflammation Alters Cappa-Opioid Modulation of Inflammatory Bladder Pain” All ETDs from UAB. 2012; 2938. Available online: https://digitalcommons.library.uab.edu/etd-collection/2938.
- Shaffer AD, Ball CL, Robbins MT, Ness TJ and Randich A. Effects of acute adult and early-in-life bladder inflammation on bladder neuropeptides in adult female rats. BMC Urol 2011; 11: 18. [CrossRef]
- Shaffer AD, Ness TJ, Randich A. Early-in-life bladder inflammation alters U50,488H but not morphine-induced inhibition of visceromotor responses to urinary bladder distension. Neurosci Lett 2013; 534:150-154. [CrossRef]
- Shaffer AD, Ness TJ, Robbins MT, Randich A. Early in life bladder inflammation alters opioid peptide content in the spinal cord and bladder of adult female rats. J Urol 2013; 189:352-358. [CrossRef]
- Smith C, Nordstrom E, Sengupta JN, Miranda A. Neonatal gastric suctioning results in chronic visceral and somatic hyperalgesia: role of corticotropin releasing factor. Neurogastroenterol Motil 2007; 19: 692-699. [CrossRef]
- Stephens-Shields AJ, Lai HH, Landis JR, Kreder K, Rodriguez LV, Naliboff BD, Afari N, Sutcliffe S, Moldwin R, Griffith JW, Clemens JQ, Bradley CS, Quallich S, Gupta P, Harte SE, Farrar JT. Clinically important differences for pain and urinary symptoms in urological chronic pelvic pain syndrome: a MAPP Network study. J Urol 2023; 209: 1132-1140.
- Sutcliffe S, Colditz GA, Goodman MS, Pakpahan R, Vetter J, Ness TJ, Andriole GL, Lai HH. Urological chronic pelvic pain syndrome symptom flares: characterisation of the full range of flares at two sites in the Multidisciplinary Approach to the Study of Chronic Pelvic Pain (MAPP) Research Network. BJU Int 2014;114: 916-25. [CrossRef]
- Sutcliffe S, Colditz GA, Pakpahan R, Bradley CS, Goodman MS, Andriole GL, Lai HH. Changes in symptoms during urologic chronic pelvic pain syndrome symptom flares: findings from one site of the MAPP Research Network. Neurourol Urodyn 2015; 34: 188-95.
- Talluri B, Hoelzel F, Medda BK, Terashvili, Sanvanson P, Shaker R, Banerjee A, Sengupta JN, Banerjee B. Identification and characterization of rostral ventromedial medulla neurons synaptically connected to the urinary bladder afferents in female rats with or without neonatal cystitis. J Comp Neurol 2022; 530: 1129-1147.
- Tullus K, Shaikh N. Urinary tract infections in children. Lancet 2020; 395:1659–1668.
- Vega-Avelaira D, Ballesteros JJ, Lopez-Garcia JA. Inflammation-induced hyperalgesia and spinal microglia reactivity in neonatal rats. Eur J Pain 2013; 17: 1180-1188. [CrossRef]
- Vega-Avelaira D, McKelvey R, Hathway G, Fitzgerald M. The emergence of adolescent onset pain hypersensitivity following neonatal nerve injury. Mol Pain 2012; 8: 30. [CrossRef]
- Vizzard MA. Alterations in spinal cord Fos protein expression induced by bladder stimulation following cystitis. Am J Physiol 2000; 278: R1027-R1039. [CrossRef]
- Wilson ML and Gaido L. Laboratory diagnosis of urinary tract infections in adult patients. Med Micro 2004; 38: 1150-1158. [CrossRef]
- Woodworth D, Mayer E, Leu K, Ashe-McNalley C, Naliboff BD, Labus JS, Tillisch K, Kutch JJ, Farmer MA, Apkarian AV, Johnson KA, Mackey SC, Ness TJ, Landis JR, Deutsch G, Harris RE, Clauw DJ, Mullins C, Ellingson BM; MAPP Research Network. Unique microstructural changes in the brain associated with urological chronic pelvic pain syndrome (UCPPS) revealed by diffusion tensor MRI, super-resolution track density imaging, and statistical parameter mapping: A MAPP Network Neuroimaging Study. PLoS One 2015;10: e0140250. [CrossRef]
- Xu L, Gebhart GF. Characterization of mouse lumbar splanchnic and pelvic nerve urinary bladder mechanosensory afferents. J Neurophysiol 2008; 99: 244-253. [CrossRef]
- Zhang G, Yu L, Chen Z-Y, Zhu J-S, Hua R, Qin X, Cao J-L, Zhang Y-M. Activation of corticotropin-releasing factor neurons and microglia in paraventricular nucleus precipitates visceral hypersensitivity induced by colorectal distension in rats. Brain Behav Immun 2016; 55: 93-104. [CrossRef]
- Zhang J, Yu J, Kannampalli P, Nie L et al. MicroRNA-mediated downregulation of potassium-chloride-cotransporter and vesicular GABA transporter expression in spinal cord contributes to neonatal cystitis-induced visceral pain in rats. Pain 2017; 158: 2461-2474. [CrossRef]
- Zhang G, Zhao B-X, Hua R, Kang J, Shao B-M, Carbonaro TM, Zhang Y-M. Hippocampal microglial activation and glucocorticoid receptor down-regulation precipitate visceral hypersensitivity induced by colorectal distension in rats. Neuropharm 2016; 102: 295-303. [CrossRef]
- Zimmermann M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983; 16: 109-110. [CrossRef]
Figure 1.
Site-dependent effect of CFA-induced inflammation. Visceromotor responses (VMRs) to graded urinary bladder distension were measured in rats which had experienced neonatal bladder inflammation (NBI) or neonatal control treatments (NoNBI) and, as adults, subsequent injection of Complete Freund’s Adjuvant (CFA) or Saline into one of four body sites: A, the left hindpaw which was ipsilateral to the site of VMR electrode placement (n=7-22/group); B, the right hindpaw (n=6-16/group); C, the right forepaw (n=5/group), or: D, the right cheek subcutaneous tissues (n=6-16/group). Injection of CFA into either hindpaw resulted in augmented VMRs but injection into the forepaw or cheek were ineffective. * indicates significantly greater VMR measures when compared with controls with p<0.05. .
Figure 1.
Site-dependent effect of CFA-induced inflammation. Visceromotor responses (VMRs) to graded urinary bladder distension were measured in rats which had experienced neonatal bladder inflammation (NBI) or neonatal control treatments (NoNBI) and, as adults, subsequent injection of Complete Freund’s Adjuvant (CFA) or Saline into one of four body sites: A, the left hindpaw which was ipsilateral to the site of VMR electrode placement (n=7-22/group); B, the right hindpaw (n=6-16/group); C, the right forepaw (n=5/group), or: D, the right cheek subcutaneous tissues (n=6-16/group). Injection of CFA into either hindpaw resulted in augmented VMRs but injection into the forepaw or cheek were ineffective. * indicates significantly greater VMR measures when compared with controls with p<0.05. .
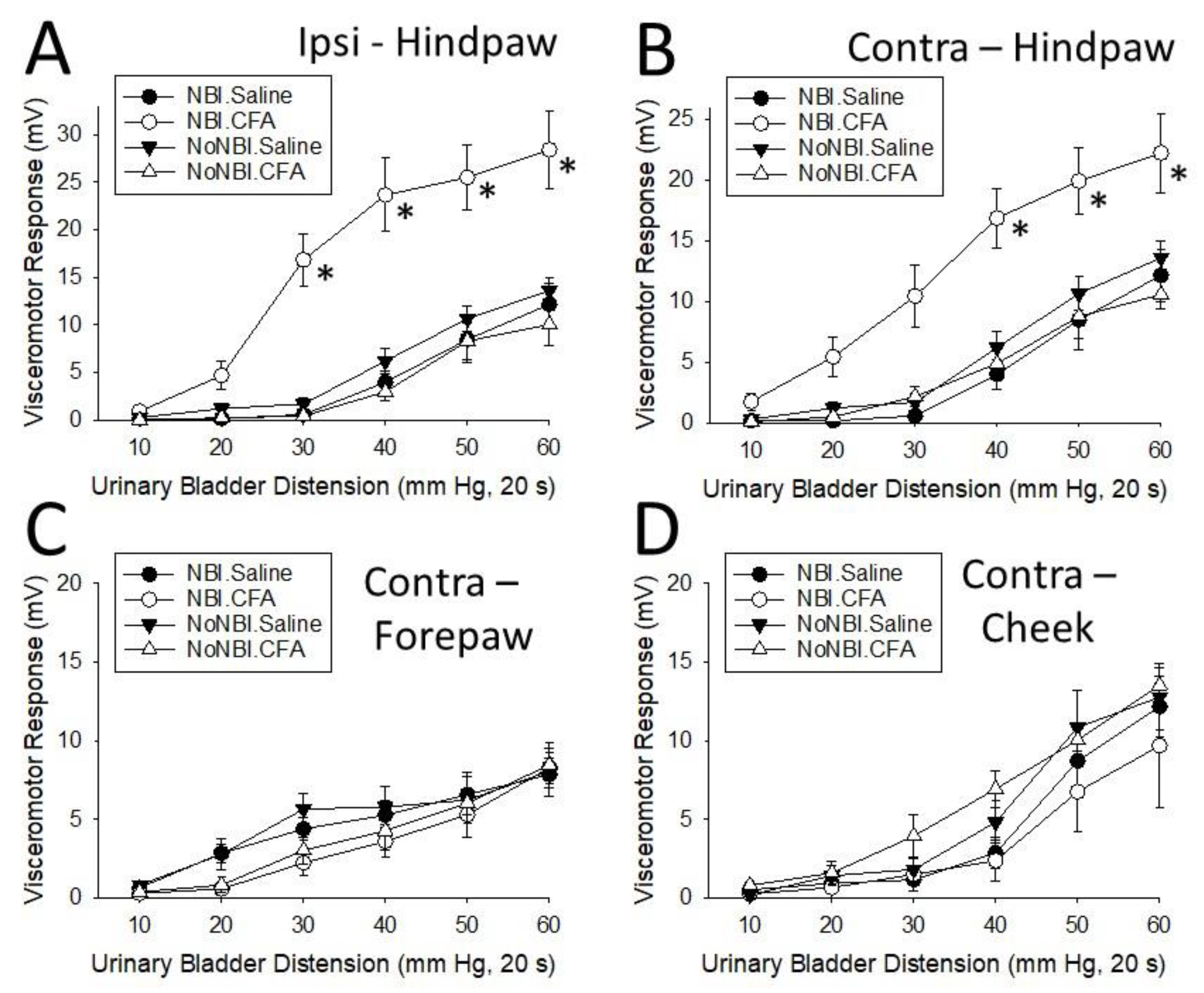
Figure 2.
ABI Alters Response to Spinal Urocortin in Rats Which Experienced NBI. Visceromotor responses (VMRs) to graded urinary bladder distension were measured in rats which had experienced neonatal bladder inflammation (NBI) and, as adults, subsequent acute bladder inflammation (ABI) or control treatments (NoABI) followed by the intrathecal injection of 15 mg of Urocortin 2 or saline. In rats which experienced NBI, but NoABI, the effect of Urocortin was profoundly inhibitory. * indicates significantly greater VMR measures when compared with saline treated controls with p<0.05. N=8 in all groups.
Figure 2.
ABI Alters Response to Spinal Urocortin in Rats Which Experienced NBI. Visceromotor responses (VMRs) to graded urinary bladder distension were measured in rats which had experienced neonatal bladder inflammation (NBI) and, as adults, subsequent acute bladder inflammation (ABI) or control treatments (NoABI) followed by the intrathecal injection of 15 mg of Urocortin 2 or saline. In rats which experienced NBI, but NoABI, the effect of Urocortin was profoundly inhibitory. * indicates significantly greater VMR measures when compared with saline treated controls with p<0.05. N=8 in all groups.
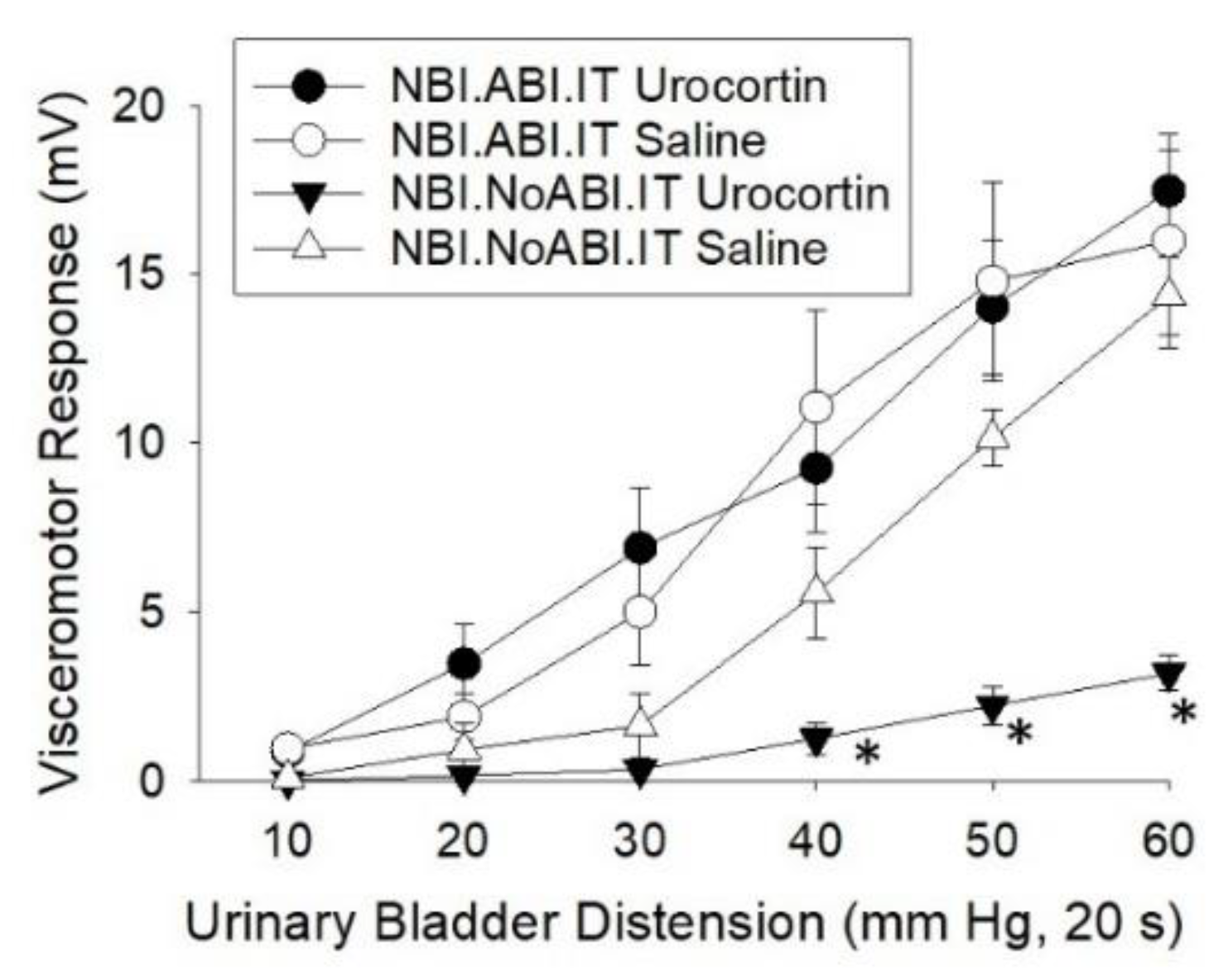
Figure 3.
Effect of intrathecal CRFR antagonists on visceromotor responses (VMRs) to urinary bladder distension (UBD). Data represent mean + SEM. NBI indicates rats experienced Neonatal Bladder Inflammation. AFS indicates rats experienced Acute Footshock treatments. See text for more description. Rats received 10 l IT injections of either Normal Saline, Antalarmin (24 g; a CRFR1 antagonist) or aSVG30 (12 g; a CRFR2 antagonist). None of the drugs utilized had statistically significant effects on VMRs to UBD in any of the pretreatment groups. The same doses of IT drugs have been effective following other secondary insults64,65.
Figure 3.
Effect of intrathecal CRFR antagonists on visceromotor responses (VMRs) to urinary bladder distension (UBD). Data represent mean + SEM. NBI indicates rats experienced Neonatal Bladder Inflammation. AFS indicates rats experienced Acute Footshock treatments. See text for more description. Rats received 10 l IT injections of either Normal Saline, Antalarmin (24 g; a CRFR1 antagonist) or aSVG30 (12 g; a CRFR2 antagonist). None of the drugs utilized had statistically significant effects on VMRs to UBD in any of the pretreatment groups. The same doses of IT drugs have been effective following other secondary insults64,65.
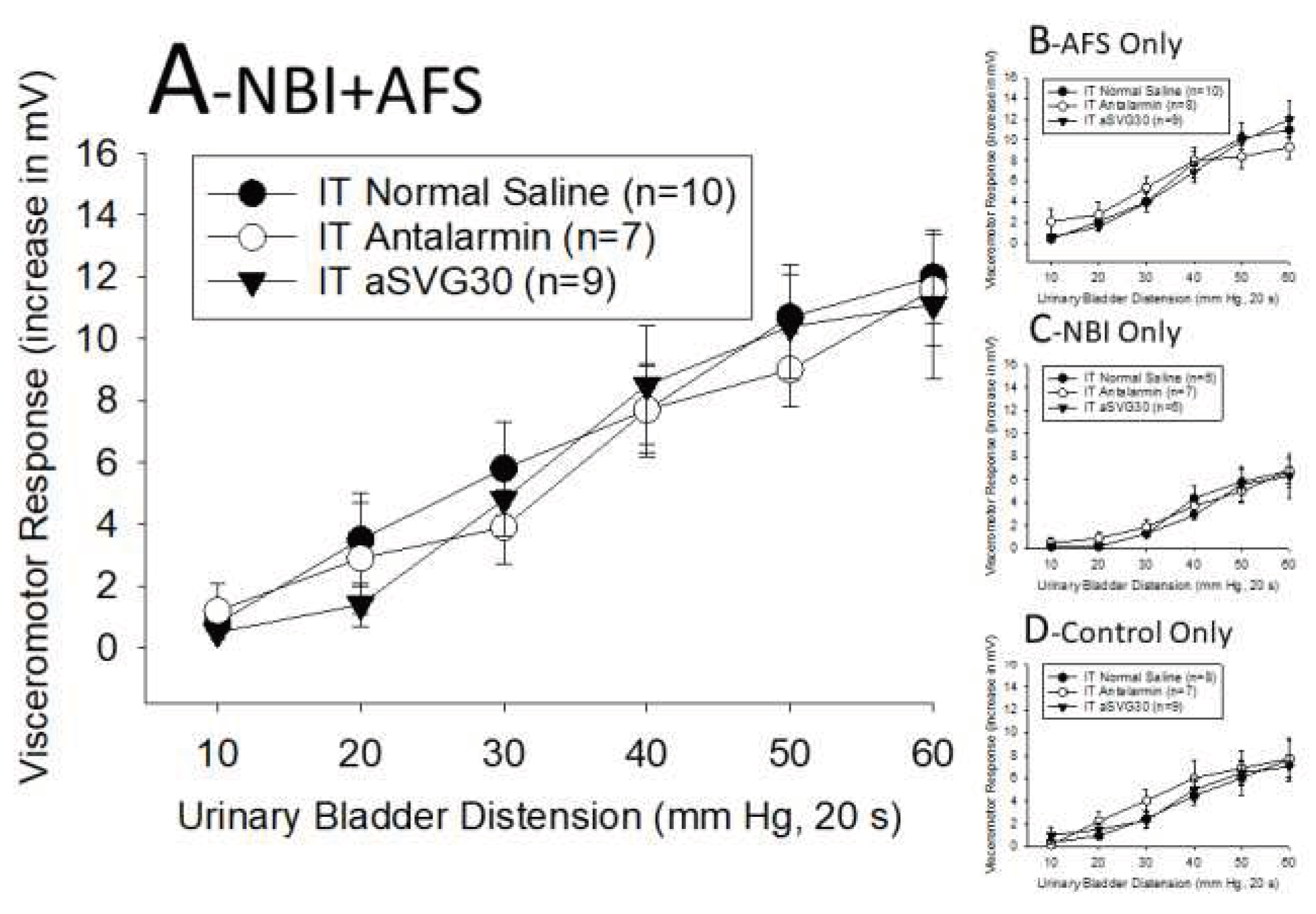
Figure 4.
Summary of changes that occur secondary to NBI.
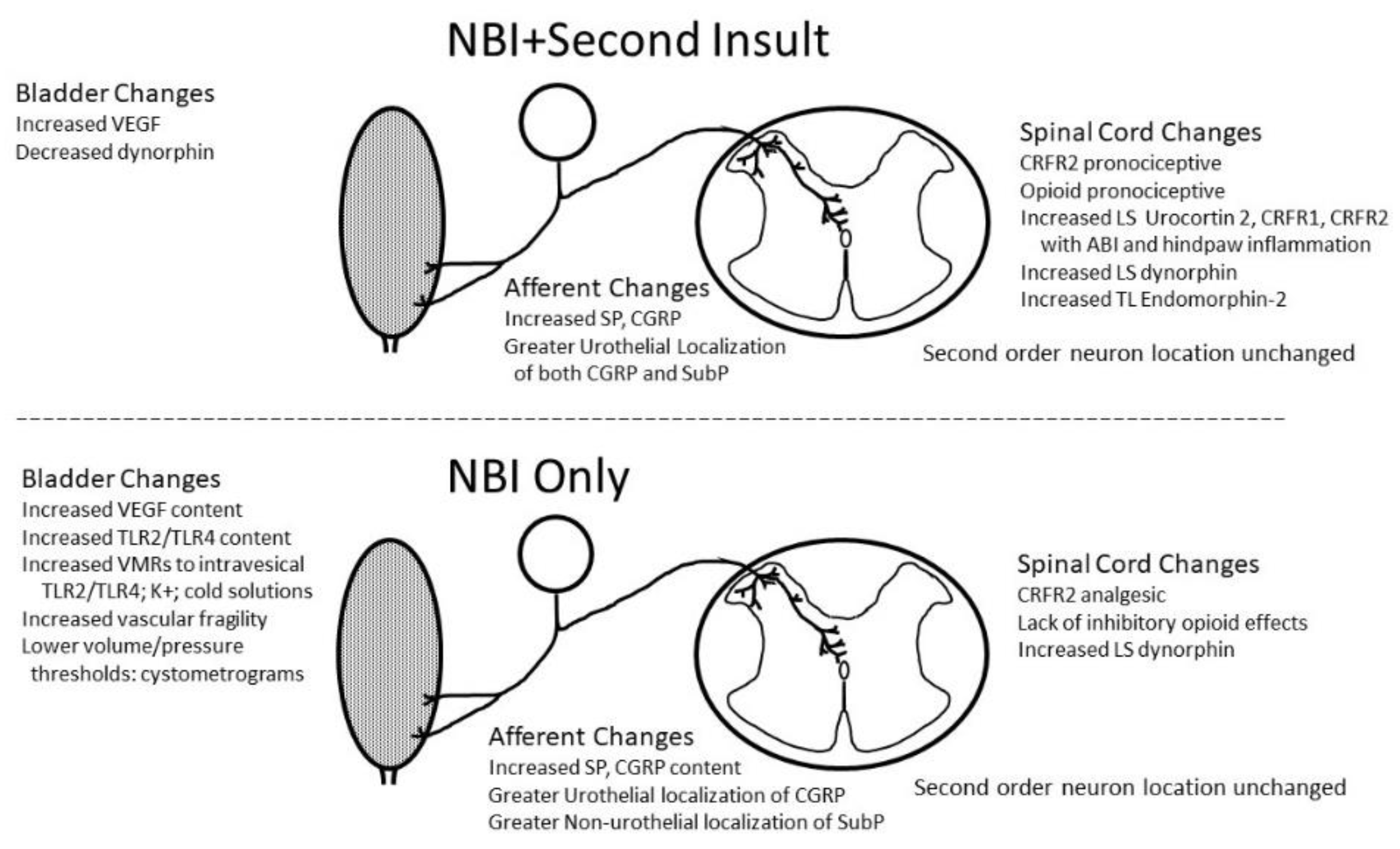

Table 1.
Neurochemical Changes Due to NBI.
| SERUM/PLASMA | Control Only | NBI Only | NBI+ABI | ABI only | |
| Dynorphin108 [% Control Only mean] (n) | 100 + 5 (14) | 104 + 6 (16) | 107 + 6 (15) | 100 + 6 (16) | |
| Beta Endorphin108 [% Control Only mean] (n) | 100 + 7 (14) | 90 + 6 (16) | 98 + 5 (15) | 85 + 5 (16) | |
| WHOLE BLADDER | Control Only | NBI Only | NBI+ABI | ABI only | |
| TLR26 [ng/mg protein] (n) | 163 + 18 (8) | 289 + 51* (8) | n.m. | n.m. | |
| TLR46 [ng/mg protein] (n) | 8 + 1 (8) | 17 + 3** (8) | n.m. | n.m. | |
| Dynorphin108 [ng/mg protein] (n) | 0.39 + 0.03 (4) | 0.46 + 0.04 (5) | 0.32 + 0.02* (7) | 0.42+0.02 (7) | |
| VEGF [pg/mg protein] (n) | 51.4 + 3.3 (9) | 52.0 + 5.7 (9) | 62.3 + 2.3* (9) | 62.0 + 3.5* (8) | |
| SPINAL CORD | Level | Control Only | NBI Only | NBI+ABI | ABI only |
| TLR26 [ng/mg protein] (n) | TL | 669 + 121 (6) | 869 + 67 (5) | n.m. | n.m. |
| LS | 508 + 80 (6) | 511 + 104 (6) | n.m. | n.m. | |
| TLR46 [ng/mg protein] (n) | TL | 4.2 + 0.7 (6) | 4.4 + 0.7 (5) | n.m. | n.m. |
| LS | 1.7 + 0.3 (6) | 2.5 + 0.8 (6) | n.m. | n.m. | |
| Dynorphin108 [ng/mg protein] (n) | TL | 0.28+0.03 (6) | 0.30 + 0.01 (6) | 0.28 + 0.01 (8) | 0.24 + 0.02 (7) |
| LS | 0.17 + 0.02 (5) | 0.27 + 0.01* (6) | 0.26 + 0.02* (8) | 0.18 + 0.01 (7) | |
| Endomorphin-2108 [ng/mg protein] (n) | TL | 1.31 + 0.03 (6) | 1.18 + 0.06 (6) | 0.92 + 0.10* (8) | 1.32 + 0.11 (7) |
| LS | 1.61 + 0.16 (5) | 1.79 + 0.18 (6) | 1.76 + 0.07 (8) | 1.73 + 0.04 (7) | |
| CRFR164 [ng/mg protein] (n) | LS | 1.40 + 0.17 (8) | 1.56 + 0.07 (8) | 2.39 + 0.23** (8) | 1.72 + 0.13 (8) |
| CRFR264 [ng/mg protein] (n) | LS | 1.42 + 0.21 (8) | 1.83 + 0.07 (8) | 3.37 + 0.62** (8) | 1.83 + 0.31 (8) |
| CRF Peptide64 [ng/mg protein] (n) | LS | 27 + 12 (8) | 11 + 4 (8) | 99 + 41 | 291 + 114* (8) |
| Urocortin64 [ng/mg protein] (n) | LS | 5.1 + 0.5 | 6.5 + 0.9 | 7.5 + 0.8* | 5.5 + 0.2 |
| SPINAL CORD | Level | Control Only | NBI Only | NBI+CFA | CFA Only |
| CRFR165 [ng/mg protein] (n) | LS | 2.23 + 0.10 (5) | 2.53 + 0.18 (5) | 2.87 + 0.14* (5) | 2.50 + 0.04 95) |
| CRFR265 [ng/mg protein] (n) | LS | 1.53 + 0.08 (5) | 2.15 + 0.09** (5) | 2.37 + 0.10** (5) | 1.93 + 0.07* (5) |
| SPINAL CORD | Level | Control Only | NBI Only | NBI+AFS | AFS Only |
| CRFR1 [ng/mg protein] (n) | LS | 2.53 + 0.14 (5) | 1.90 + 0.05 (5) | 2.13 + 0.16 (5) | 2.41 + 0.15 (5) |
| CRFR2 [ng/mg protein] (n) | LS | 1.99 + 0.13 (5) | 1.58 + 0.04* (5) | 1.62 + 0.06* (5) | 1.91 + 0.13 (5) |
Data represents Mean + SEM. New data from this manuscript and indicated by italic script. TL indicates thoracolumbar spinal cord; LS indicates lumbosacral spinal cord. * and bolded data indicates values statistically different from corresponding Control Only group, p<0.05. “n.m.” indicates not measured.
Table 2.
Effect of NBI on Neuropeptide Content of the Bladder.
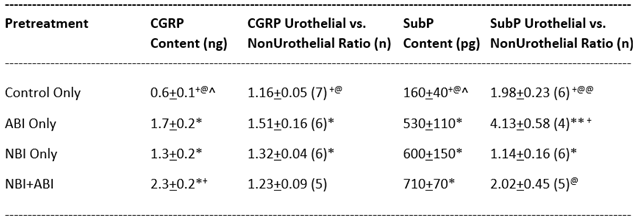
All values are Mean + SEM. NBI indicates Neonatal Bladder Inflammation; ABI indicates Adult Bladder Inflammation; CGRP indicates calcitonin gene-related peptide; SubP indicates Substance P *,** indicate significant difference from Control Only value at P<0.05 and 0.01 values +indicates significant difference from NBI Only value at P<0.05 value. @,@@indicate significant difference from ABI Only value at P<0.05 and 0.01 values ^ indicates significant difference from NBI+ABI value at P<0.05 value. Content data from Shaffer et al106. Ratio data is novel and methods described in text.
Table 3.
Spinal c-Fos expression: effects of NBI & ABI.
| L5 Lateral Dorsal Horn | Not Distended | Distended | L6 Lateral Dorsal Horn | Not Distended | Distended | S1 Lateral Dorsal Horn | Not Distended | Distended |
| Control Only | 1.60 + 0.39 | 3.63 + 0.59* | Control Only | 2.71 + 0.46 | 4.88 + 0.65* | Control Only | 2.21 + 0.33 | 2.17 + 0.63 |
| ABI Only | 2.25 + 0.68 | 3.77 + 0.89 | ABI Only | 3.54 + 1.33 | 4.19 + 1.30 | ABI Only | 1.21 + 0.42 | 1.54 + 0.28 |
| NBI Only | 3.69 + 0.99 | 4.44 + 0.42 | NBI Only | 3.13 + 0.30 | 7.36 + 1.03* | NBI Only | 2.09 + 0.92 | 2.86 + 0.95 |
| NBI+ABI | 2.69 + 0.62 | 4.34 + 0.14* | NBI+ABI | 4.00 + 1.95 | 4.83 + 0.40 | NBI+ABI | 2.23 + 1.29 | 1.75 + 0.34 |
| L5 Medial Dorsal Horn | Not Distended | Distended | L6 Medial Dorsal Horn | Not Distended | Distended | S1 Medial Dorsal Horn | Not Distended | Distended |
| Control Only | 9.11 + 0.83 | 13.06 + 4.22 | Control Only | 8.42 + 1.30 | 17.73 + 2.78* | Control Only | 4.88 + 1.11 | 4.17 + 2.28 |
| ABI Only | 13.61 + 3.66 | 13.50 + 3.03 | ABI Only | 11.88 + 4.34 | 11.29 + 3.36 | ABI Only | 3.56 + 1.43 | 2.12 + 0.84 |
| NBI Only | 9.92 + 3.75 | 20.25 + 0.69* | NBI Only | 10.42 + 3.42 | 18.98 + 4.21 | NBI Only | 4.42 + 1.25 | 6.44 + 3.00 |
| NBI+ABI | 10.79 + 1.52 | 12.46 + 1.46 | NBI+ABI | 10.86 + 3.28 | 12.83 + 1.20 | NBI+ABI | 5.81 + 4.45 | 3.38 + 1.06 |
| L5 Spinal Nucleus | Not Distended | Distended | L6 Parasympathetic Nucleus | Not Distended | Distended | S1 Parasympathetic Nucleus | Not Distended | Distended |
| Control Only | 0.75 + 0.14 | 6.46 + 1.97* | Control Only | 3.78 + 1.48 | 12.42 + 1.40* | Control Only | 1.44 + 0.25 | 4.81 + 1.64 |
| ABI Only | 1.04 + 0.25 | 4.46 + 0.74* | ABI Only | 1.96 + 0.59 | 8.40 + 2.67 | ABI Only | 0.90 + 0.20 | 2.40 + 0.47* |
| NBI Only | 0.46 + 0.38 | 5.98 + 1.41* | NBI Only | 1.42 + 0.92 | 6.96 + 1.93* | NBI Only | 0.79 + 0.29 | 3.77 + 1.28 |
| NBI+ABI | 0.94 + 0.17 | 6.34 + 1.64* | NBI+ABI | 2.29 + 0.78 | 6.75 + 1.63* | NBI+ABI | 1.11 + 0.52 | 3.46 + 1.49 |
| L5 Dorsal Commissure | Not Distended | Distended | L6 Dorsal Commissure | Not Distended | Distended | S1 Dorsal Commissure | Not Distended | Distended |
| Control Only | 4.73 + 1.40 | 23.40 + 4.99* | Control Only | 4.82 + 0.88 | 23.00 + 3.87* | Control Only | 3.50 + 0.68 | 6.00 + 1.96 |
| ABI Only | 7.92 + 2.16 | 17.82 + 3.52* | ABI Only | 5.23 + 2.09 | 13.79 + 3.21* | ABI Only | 2.35 + 0.98 | 3.13 + 1.27 |
| NBI Only | 4.13 + 0.13 | 24.71 + 4.99* | NBI Only | 2.29 + 0.79 | 16.21 + 1.78* | NBI Only | 1.46 + 0.21 | 8.02 + 1.90* |
| NBI+ABI | 5.73 + 1.19 | 17.02 + 3.82* | NBI+ABI | 4.65 + 1.12 | 14.21 + 4.68* | NBI+ABI | 2.55 + 1.73 | 5.23 + 2.72 |
|
*effect of distension; p<0.05 Bold indicates significant difference from “Not Distended” measure |
Data represents Mean + SEM Fos+ nuclei | |||||||
|
Repeated Final Columns for Reviewers – formatting issues with conversion S1 Lateral Dorsal Horn |
Not Distended | Distended | ||||||
| Control Only | 2.21 + 0.33 | 2.17 + 0.63 | ||||||
| ABI Only | 1.21 + 0.42 | 1.54 + 0.28 | ||||||
| NBI Only | 2.09 + 0.92 | 2.86 + 0.95 | ||||||
| NBI+ABI | 2.23 + 1.29 | 1.75 + 0.34 | ||||||
| S1 Medial Dorsal Horn | Not Distended | Distended | ||||||
| Control Only | 4.88 + 1.11 | 4.17 + 2.28 | ||||||
| ABI Only | 3.56 + 1.43 | 2.12 + 0.84 | ||||||
| NBI Only | 4.42 + 1.25 | 6.44 + 3.00 | ||||||
| NBI+ABI | 5.81 + 4.45 | 3.38 + 1.06 | ||||||
| S1 Parasympathetic Nucleus | Not Distended | Distended | ||||||
| Control Only | 1.44 + 0.25 | 4.81 + 1.64 | ||||||
| ABI Only | 0.90 + 0.20 | 2.40 + 0.47* | ||||||
| NBI Only | 0.79 + 0.29 | 3.77 + 1.28 | ||||||
| NBI+ABI | 1.11 + 0.52 | 3.46 + 1.49 | ||||||
| S1 Dorsal Commissure | Not Distended | Distended | ||||||
| Control Only | 3.50 + 0.68 | 6.00 + 1.96 | ||||||
| ABI Only | 2.35 + 0.98 | 3.13 + 1.27 | ||||||
| NBI Only | 1.46 + 0.21 | 8.02 + 1.90* | ||||||
| NBI+ABI | 2.55 + 1.73 | 5.23 + 2.72 | ||||||
Table 4.
Comparison of Human and Rodent Features.
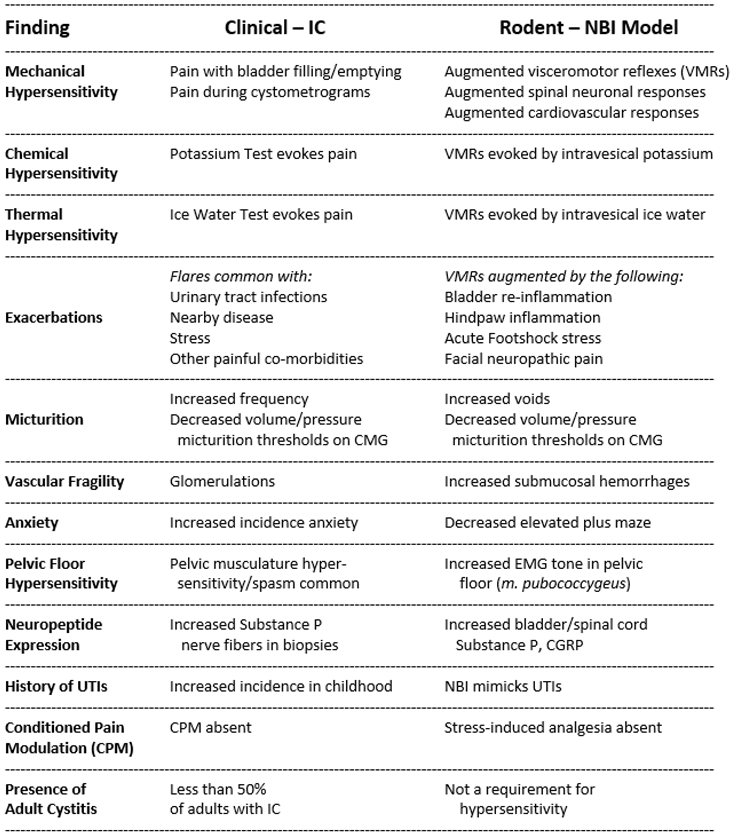
Abbreviations: NBI – neonatal bladder inflammation; UTI – urinary tract infection; VMRs – visceromotor responses; IC – interstitial cystitis; CPM – Conditioned Pain Modulation; EMG – electromyogram.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



