
Submitted:
07 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Metformin is an oral antidiabetic agent with antimicrobial properties, excreted in an active and unmetabolized form. Like some non-antibiotic drugs, subinhibitory concentrations of metformin may contribute to the expression of microbial resistance in wild-type strains. This work aimed to evaluate the biofilm formation of Pseudomonas aeruginosa TGC04 exposed to 100 and 200 µg/mL of metformin, varying the Chemical Oxygen Demand (COD) and flow regime. Biofilms were quantified after 2 and 24 hours, observing reductions and increases in biofilm growth around 13 and 64%, respectively. The greatest biofilm growth occurred in cells exposed to 200 µg/mL of metformin. Both the flow regime and COD concentration were important for the development of the biofilm, whose cells were mostly weakly to moderately adhered. The study provides evidence that in polluted aquatic systems with turbulent regimes, the inhibitory concentration of metformin promoted the least disturbances to the biofilm.
Keywords:
Emerging contaminants
; Non-antibiotic drugs
; Antimicrobial activity.
1. Introduction
With the evolution of the pharmaceutical industry, the consumption of medicines by society has become a common practice, contributing to the increase in the gram ratio of drug intake per person [1]. As a result, active quantities of drugs and their metabolites are discarded into the environment in various ways. Their presence in water bodies represents a threat to health, as it can alter the trophic chain [2], promoting endocrine disruption in animals [3] and inducing microbial tolerance [4].
The United Nations Environment Program recently published a report which makes a grim prediction about future pandemics caused by multidrug-resistant bacteria as spread through water. This is because a great amount of still-active antibiotic and non-antibiotic drugs is excreted through feces and urine. These reach water bodies through water and may pass through sewage treatment plants. The excretion of drugs in active form depends on both their pharmacological and pharmacokinetic properties, as well as the health status of the human or veterinary patient, to whom these drugs have been administered [5].
Metformin is a biguanide derived from galegine, a metabolite produced by Galega officinalis L., and presents fruitful results in relation to glucose metabolism [6]. Metformin is the most prescribed oral antidiabetic medication worldwide, making up 45 to 50% of all prescriptions, used by more than 150 million people annually [7]. In Brazil, it was among the three most sold medicines in the last five years, grossing more than US$ 100 million a year [8]. Metformin is also worrying because its undiscriminating automedical use by Brazilians wanting to lose weight has grown enormously [9].
Metformin has negative effects, however, on ecosystem services when released into the environment. Little is known about its effect on the metabolism of many aquatic microorganisms [10]. Metformin is thought to be the most prevalent drug discarded into water bodies [11]. The drug is eliminated unchanged through urine and its fate in the environment is a transformation into guanylurea (GGU), followed by its mineralization [12]. In addition, both molecules are highly hydrophilic, which makes rapid and passive diffusion across the membrane unlikely [13]. They also do not bioaccumulate [14] and can be metabolized by animals and vegetables.
Like other non-antibiotic drugs, metformin demonstrates a certain degree of antimicrobial activity against Pseudomonas aeruginosa, but also other Gram-negative pathogens, Gram-positive bacteria and yeasts may exhibit sensitivity to the drug [15]. The proposed mechanism of action indicates the potentiation of antibiotics because it promotes changes in the outer membrane and favors the intracellular accumulation of antimicrobials [16]. However, metformin can promote multidrug resistance, as has already been reported for other Gram-negative cases, via chromosome mutagenesis [17].
Biofilm acts as a physical barrier in hostile environments and enables cells to become resistant. Contact with subinhibitory levels of bioactive compounds favors the development of persistent subpopulations, resulting from the horizontal transfer of resistance factors within the cell community [18]. Since P. aeruginosa is believed to be an opportunistic microorganism, the development of antibiotic resistance due to contact with non-antibiotic drugs represents a risk that needs to be studied. Thus, this work aimed to test the hypothesis that in polluted waters, exposure to inhibitory concentrations of metformin positively affects the growth of P. aeruginosa biofilm.
2. Materials and Methods
2.1. Strain
The study used the wild strain P. aeruginosa TGC04 (Brazilian SisGen #A40AD65; UFPEDA 1063B), recovered from oil-contaminated soil and maintained at the Environmental Microbiology Laboratory (CBiotec/UFPB).
2.2. Metformin
Metformin hydrochloride was purchased (Sigma-Aldrich) and the standard solution was prepared in sterilized distilled water (50,000 µg/mL).
2.3. In vitro planktonic cell perturbation assay
The Minimum Inhibitory Concentration (MIC) verification test was carried out in microdilution plates containing 100 µL Müeller-Hinton broth, 100 µL of metformin solution; and 10 µL of the bacterial suspension, prepared in NaCl 0.85%, from freshly cultured cells (turbidity = tube 0.5 on the MacFarland scale). Metformin concentrations ranging from 25,000 up to 50 µg/mL diluted at a 1:5 ratio). The system was incubated at 29±1 ºC for 24h. Afterwards, the wells were examined at λ= 630 nm (BioteK, EL800) and the MIC was interpreted as the concentration at which the presence of viable cells was no longer observed [19]. The qualification of the inhibitory action was interpreted according to the scale proposed by Sartorato et al. [20]. The assay was performed in triplicate.
2.4. In vitro biofilm formation assay
Nine glass microcosms were prepared, measuring 110 mm (height) x 97 mm (width) x 97 mm (length), filled with 500 mL of synthetic wastewater with Chemical Oxygen Demand (COD) at 10,000 and 1,000 mg/L, respectively. the composition of the synthetic wastewater with COD at 10,000 mg/L was, as follows (in mg/L): sucrose (8.5); yeast extract (1.96); urea (0.62); CaCl2 (0.03); MgSO4 (0.02); K2HPO4 (0.03); KH2PO4 (0.003) e FeCl3 (0.003) [21]. The metformin solution was then mixed with the medium to make concentrations of 200 (MIC) and 100 µg/mL (Sub-MIC). Two incubation conditions were tested, with and without aeration as provided by a submerged aquarium pump (JAD, S-510), 220 V, with a flow rate of 4 L/min and pressure of 0.01 MPa. At the top of the microcosms, a support was built for polyamide thread to suspend 6 equidistant glass slides, with a submerged area of 875 mm2 per face (Figure 1). The microcosms were aseptically capped with plastic film and left to incubate at 25 ºC for 24h. Samples were taken in triplicate during the first two hours of incubation and the rest were collected after 24 hours.
Biofilm was quantified by the crystal violet test [22]. The glass slides were removed and subsequently washed with tap water to eliminate cells not adhered to the surface. After drying, the glass slides were treated with 10 mL of 1% crystal violet for 20 minutes. Afterwards, the excess dye was removed with tap water and the glass slide was washed again with an equal volume of absolute ethanol. The absorbance of the second wash solution, containing the crystal violet-ethanol mixture, was determined at λ= 590 nm (Kasvi, K37-VIS). Furthermore, the amount of biofilm remaining after exposure to metformin was expressed using the formula:
% = [(C–A) – (T–B) / (C–A)] x 100
Where, C = control absorbance, T = treatment absorbance; B = absorbance of the medium and A, the absorbance of the metformin solutions.
Any reduction in the adhesion of P. aeruginosa TGC04 was understood as antimicrobial activity. The antibiofilm effect of metformin was only considered, however, when the percentage of reduction was equal to or greater than 80%, compared to the control. Likewise, the adherence percentage was considered weak if < 40%, moderate if between 40-80%, and high if > 80% [23]. The assay was performed in triplicate randomly. The test control was performed without the addition of metformin.
3. Results
Table 1 summarizes the results of the in vitro biofilm formation assay with addition of metformin at MIC (200 µg/mL) and sub-MIC (100 µg/mL). Under some of the tested conditions, the values obtained at 2 h of testing were very different from those observed at 24 h. This was because under certain conditions a greater percentage of reduction in adhesion was observed at 2 h; but later decreased as the biofilm continued to form. To give more understanding to the results, and considering biofilm formation as a dynamic, cumulative, or dispersive process, the results were expressed as the variation between the two times (Δ24h-2h). Negative numbers represent an increase in biofilm growth, while the reduction was represented by positive numbers. Subtracting 100% from the 24-hour reduction percentage in the test served to classify the type of cell adhesion.
The greatest increase and reduction in biofilm growth occurred under exposure to sub-MIC of metformin (100 µg/mL), ≈64% and ≈13%, respectively. The COD content and flow rate were important variables for the fate of the P. aeruginosa TGC04 biofilm. Under the stagnant regime and exposure to sub-MIC, the higher the COD, the greater the adhesion of cells to the glass surface; while under turbulent regime, the opposite was observed (inhibition of the biofilm). On the other hand, metformin exposure to the MIC (200 µg/mL) resulted in a pattern more dependent on the flow rate, i.e., the turbulent regime favored biofilm growth while the stagnant regime resulted in a reduction in the number of adhered cells with COD at 1000 mg/L.
4. Discussion
In this work, the effect of metformin on the growth of the P. aeruginosa TGC04 biofilm was studied in a model simulating a polluted aquatic environment, under both a turbulent and a stagnant regime. The results suggest that biofilm growth was disturbed due to shear stress on the surface where the biofilm formed; and the COD content resulted in no significant antibiofilm effect. These findings agreed with previously reported results where low nutrient content favored biofilm growth, and where high nutrient concentrations did not promote an inhibitory effect [24].
This present study revealed that in a stagnant regime, a lower COD provided conditions that culminated in a reduction in biofilm growth. Likewise, under conditions with higher COD, both sub-MIC and MIC concentrations of metformin have not exhibited antimicrobial activity in the stagnant and turbulent regimes. This can be explained from two distinct points of view: subinhibitory concentrations of metformin stimulate the formation of cell aggregation; or, the higher the nutrient level, the less biomass loss there is, even though metformin is found at inhibitory concentrations.
A higher concentration of organic matter in a body of water reflects an increase in COD and this is an important indicator of anthropogenic pollution [25]. In addition, biofilm growth demonstrates a positive correlation with organic matter content and P. aeruginosa is able to grow with both low or high COD [26]. Furthermore, hydrodynamic forces are one of the most important factors that favor the development of biofilms, whether in coherent or dispersed. These forces enable the mass transfer of nutrients, metabolites, oxygen, and cells from the medium to the biofilm [27,28].
Under a stagnant regime, it is not possible to have a significant growth of the biofilm when compared to that formed in a turbulent regime [29]. At faster flow rates, biofilms develop in a monolayer, increasing their resistance [30]. A previous study observed that the biofilm formed by a strain of Escherichia coli exhibited doubled and tripled growth when the flow rate was 2.13 and 4.25 L/min, respectively [31]. In addition, according to Mello [32], when cells were exposed to flow rates < 9.4 L/min (or 2 m/s) there was no impediment to biofilm formation. Growth may be more evident at flow rates < 4 .7 L/min (or < 1 m/s). In this study, a flow rate of 4 L/min was used, which may have contributed to the increase in cell adhesion, even in test 4 (Table 1), where there was only a little reduction < 0.5%. Furthermore, the adhesive glass surface favored cell aggregation. This was contrary to what was described by a previous work that used steel surfaces exposed to a flow of 1 m/s and it was observed no growth [33].
Various studies have reported that nutrient content is crucial for biofilm growth of different microorganisms [34,35]. Nutritional stress conditions can stimulate the growth of biofilms [36] and their growth rate increases with increased organic carbon assimilation. In a non-turbulent regime, nutrient assimilation is disadvantaged and consequently cell multiplication can be disturbed [37].
Oligotrophic and/or nitrogen-deficient environments lead to a reduction in planktonic life and stimulate the growth of biofilms [38]. Biofilm forms a protective structure in a nutrient-poor environment; in high-nutrient sites, there is greater conservation of planktonic forms [39]. Additionally, in the presence of inhibitors, planktonic cells may be more susceptible to the inhibitory effects exerted by antimicrobials. However, this change in lifestyle functions is an adaptive strategy of P. aeruginosa in response to the environmental stresses to which it is subjected, such as exposure to ultraviolet radiation [40], temperature variations [41], different pH [42] and exposure to antimicrobials [43]. P. aeruginosa biofilm can tolerate between 100 and 1000 times the concentrations of these compounds [44].
A specific choice of P. aeruginosa used in this study was made; this biofilm is characterized by its rapid adaptive response, as well as ecological flexibility when subjected to environmental stress. As noted by Elabed et al. [45], P. aeruginosa can adapt to stress caused by various environmental factors. Even if the exposure time to stressors is long, it responds quickly, exhibiting phenotypes with losses or gains in physiological and biochemical abilities, as well as presenting changes in morphology. In addition, P. aeruginosa demonstrates a high capacity to change lifestyle from free-swimming cells to organized aggregates, given its affinity for a variety of surfaces [46].
P. aeruginosa can adhere to both hydrophobic and hydrophilic surfaces because it can alter the structure of the O-antigen of its LPS, the largest constituent of the Gram-negative wall. It is known that the bacteria express two glycoforms of O-antigen. The first, highly conserved and more hydrophobic, is called the Common Polysaccharide Antigen (CPA) and is composed of homopolymer of D-rhamnose in trisaccharide repeating units. The second, O-Specific Antigen (OSA) is a heteropolymer composed of variable structures [47]. This results in a differentiation between strains and a change in the hydrophobicity profile of the cell wall, resulting in an increase in the types of surfaces available for adhesion [48].
In terms of adhesion, P. aeruginosa commonly exhibits moderate to weak and occasionally high adhesion. This characteristic represents one of the bacteria's evolutionary strategies to enable detachment and migration to colonize new surfaces when it is exposed to environmental stresses or when it is in competition with other microbes [49]. Additionally, a recent study observed in the behavior of Burkholderia contaminans (Pseudomonadota) that the fact of moderate adherence is an element that may help to understand the microbial response to nutrient limitation in the environment, i.e., adherence in order to obtain a more immediate response and to provide means for colonizing sites richer in nutrients [50].
Environmental stresses cause microorganisms to exhibit various genotypic and phenotypic responses and the growth of biofilms is one of them; the mechanisms this response, however, has not yet been fully elucidated [51]. Based on our results, the inhibitory potential of metformin was notable against planktonic cells of P. aeruginosa. These findings agree with what has been previously reported [52]. The expected antibiofilm activity was not observed. Biofilm growth, however, is subject to other factors which are listed in the literature [53]. Specifically, as found in our study, those behaviors that seemed to have contributed most were the bioavailability of nutrients, the flow rate and the time of exposure to metformin.
The subinhibitory concentration of metformin inhibited P. aeruginosa TGC04, especially under a stagnant regime. This may have been a result of the amount of assay time. On the other hand, there were two conditions under which the cells were highly adhered to the glass surface (Table 1). It is not uncommon to find material in the literature that describes antimicrobials stimulating the formation of microbial biofilms after exposure to subinhibitory concentrations [54]. In our study, this increase in biofilm concentration was up to around 64% in comparison to the control. This value was also greater than found in a previous investigation where subinhibitory concentrations of different drugs increased the Enterococcus faecalis biofilm by around 50% [55]. In contrast, another study, carried out with E. coli, found that subinhibitory concentrations of several antimicrobials exhibited antibiofilm activity through a mechanism of inhibition of pili formation, important in the adhesion process. It should be further noted that the response is dependent on the type of drug, the time of exposure to the drug and the form of the bacteria, i.e., planktonic or sessile [56]. The inhibition of growth at approximately 72% in our study, however, was possibly related to motility.
The mechanism that affects P. aeruginosa motility involves changes in quorum sensing systems. The P. aeruginosa PAO1 strain in one study was investigated in terms of sensitivity to metformin. The MIC observed was excessively high, 100 mg/mL, five hundred times higher than that observed in our study. When applied at a concentration of MIC÷10, however, the metformin demonstrated significant inhibition in the expression of important virulence factors such as pyocyanin, hemolysin, protease and elastase. The bacterial sensitivity to oxidative stress was also increased. These phenotypes are linked to quorum sensing systems [57].
Jing Zuo et al. [58] analyzed the effects of metformin on the formation of Streptococcus suis biofilm, and found results contradictory to those of this research, as they did not observe inhibitory activity in planktonic cells. However, the authors reported significant biofilm reduction by inhibiting the synthesis of the autoinducer AI-2. This reinforces the assumption that biofilm reduction in P. aeruginosa may strongly involve anti-quorum sensing mechanisms. Future research may elucidate this hypothesis, based on the premise that inhibition of cell-cell communication may be a way to control pathogen growth.
The Covid-19 pandemic was a wake-up call to turn our attention to future pandemics caused by multidrug-resistant pathogens. There is clear evidence that the disposal of non-antibiotic drugs and their metabolites into the environment can trigger mechanisms that end up involving both tolerance and resistance to antimicrobials in wild microbiota. Thus, the transfer of resistance genes is guaranteed. Given this, there is an urgent need to provide new perspectives to manage urban effluents to reflect this new reality.
5. Conclusions
This study provided evidence that in turbulent and polluted aquatic systems, the response of P. aeruginosa TGC04 exposed to metformin is to form biofilm, with nutrient availability as a relevant factor. Exposure of inhibitory and subinhibitory concentrations of metformin caused stress and led to adhesion and biofilm formation by P. aeruginosa TGC04.
Acknowledgments
The authors would like to express their gratitude to Universidade Federal da Paraíba (UFPB) for supporting the research by grants to students affiliated with the PVI14908-2022 Project (Call #01/2022 – PROPESQ/CGPAIC/UFPB). The English text of this paper has been revised by Sidney Pratt, Canadian, MAT (The Johns Hopkins University), RSAdip - TESL (Cambridge University).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Morais, V.C.; Soeder, J.; Özcan, E.; Gomes, V.T.; Lima, K.Y.G.; Vasconcelos, U. 2022. Different solutions prepared with over-the-counter and under prescription medicines alters germination and seedling growth of linseed (Linaceae). World J Pharm Pharm Sci. 2022, 11, 1-11.
- Valdéz-Carrillo, M.; Abrell, L.; Ramírez-Hernández, J.; Reyes-López, J.; Corréon-Diazconti, C. Pharmaceuticals as emerging contaminants in the aquatic environment of Latin America: a review. Environ Sci Pollut Res. 2020, 27, 44683–44891. [Google Scholar] [CrossRef] [PubMed]
- Kasonga, T.K.; Coetzee, M.A.A.; Kamika, I.; Ngole-Jeme, V.M.; Momba, M.N.B. Endocrine-disruptive chemicals as contaminants of emerging concern in wastewater and surface water: A review. J Environ Manage. 2011, 277, 111485. [Google Scholar] [CrossRef] [PubMed]
- Kock, N.; Islam, N.F.; Sonowal, S.; Prasad, R.; Sarma, H. Environmental antibiotics and resistance genes as emerging contaminants: Methods of detection and bioremediation. Curr Res Microb Sci. 2020, 2: 100027. [CrossRef]
- United Nations Environment Programme. Environmental dimensions of antimicrobial resistance: Summary for policymakers. United Nations Environment Programme: Nairobi, Kenya, 2022.
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia. 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Drzewoski, J.; Hanefeld, M. The current and potential therapeutic use of metformin-The good old drug. Pharmaceuticals, 2021, 14, 122. [CrossRef]
- Executive Secretariat of the Drugs Market Regulation Chamber. Statistical yearbook of the pharmaceutical market. CMED: Brasília, Brazil, 2022.
- Dantas, P.; Azevedo, T.R.; Costa, H.P.; Rios, E.R.V.; Cavalcanti, M.G.; Oliveira, E.G. Off-Label use of metformin for the treatment of obesity: a risk or success in clinical practice? Biomed J Sci Tech Res. 2023, 48, 39158–39161. [Google Scholar]
- Cummings, B.M.; Needoba, J.A.; Peterson, T.D. Effect of metformin exposure on growth and photosynthetic performance in the unicellular freshwater chlorophyte, Chlorella vulgaris. PlosOne. 2018, 13, e0207041. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, C.; Berset, D.; Wolschke, H. Occurrence of the antidiabetic drug Metformin and its ultimate transformation product guanylurea in several compartments of the aquatic cycle. Environ Int. 2014, 70, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Straub, J.O.; Caldwell, D.J.; Davidson, T.; D'Aco, V.; Kappler, K.; Robinson, P.F.; Simon-Hettich, B.; Tell, J. Environmental risk assessment of metformin and its transformation product guanylurea. I. Environmental fate. Chemosphere. 2019, 216, 844–854. [Google Scholar] [CrossRef]
- Foretz, M.; Guigas, B.; Bertrand, L.; Pollak, M.; Viollet, B. Metformin: From mechanisms of action to therapies. Cell Metab. 2014, 20, 953–966. [Google Scholar] [CrossRef]
- Ussery, E.; Nielsen, K.; Pandelides, Z.; Kirkwood, A.; Bonetta, D.; Venables, B.; Guchardi, J.; Holdway, D. Effects of environmentally relevant metformin exposure on Japanese medaka (Oryzias latipes). Aquat Toxicol. 2018, 205, 58–65. [Google Scholar] [CrossRef]
- Meherunisa; Saptna, J.; Vikas, S. Study of metformin effect on antimicrobial property. Int Arch Biomed Clin Res. 2018, 4, 85–87. [Google Scholar]
- Masadeh, M.M.; Alzoubi, K.H.; Masadeh, M.M.; Aburashed, Z.O. Metformin as a potential adjuvant antimicrobial agent against multidrug resistant bacteria. Clin Pharmacol. 2021, 13: 83-90.
- Wei, Z.; Wei, Y.; Li, H.; Shi, D.; Yang, D.; Yin, J.; Zhou, S.; Chen, T.; Li, J.; Jin, M. Emerging pollutant metformin in water promotes the development of multiple-antibiotic resistance in Escherichia coli via chromosome mutagenesis. J Hazard Mat. 2022, 430, 128474. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci Biotechnol Biochem. 2015, 80, 7–12. [Google Scholar] [CrossRef]
- Olajuyigbe, O.O; Afolayan, A.J. In vitro antibacterial and time-kill assessment of crude methanolic stem bark extract of Acacia mearnsii De Wild against bacteria in shigellosis. Molecules. 2012, 17, 2103–2118. [Google Scholar] [CrossRef] [PubMed]
- Sartoratto, A.; Machado, A.L.M.; Delarmelina, C.; Figueira, G.M.; Duarte, M.C.T.; Rehder, V.L.G. Composition and antimicrobial activity of essential oils from aromatic plants used in Brazil. Braz J Microbiol. 2004, 35, 275–280. [Google Scholar] [CrossRef]
- Reyes-Lara, S.; Reyes-Mazzoco, R. Effect of hydraulic and organic loads on the mass removal of a structured packing in a trickling filter. Rev Mex Ing Quim. 2009, 8, 101–109. [Google Scholar]
- Ommen, P.; Zobek, N.; Meyer, R. Quantification of biofilm biomass by staining: Nontoxic safranin can replace the popular crystal violet. J Microbiol Methods. 2017, 141, 87–89. [Google Scholar] [CrossRef]
- Rossi, E.; Paroni, M.; Landini, P. Biofilm and motility in response to environmental and host-related signals in Gram negative opportunistic pathogens. Appl Microbiol. 2018, 125, 1587–1602. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm formation and control strategies of foodborne pathogens: food safety perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef]
- Han, X.; Chen, X.; Ma, J.; Chen, J.; Xie, B.; Yin, W.; Yang, Y.; Jia, W.; Xie, D.; Huang, F. Discrimination of Chemical Oxygen Demand pollution in surface water based on visible near-infrared spectroscopy. Water. 2022, 14, 3003. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, X.; Bian, W. Impact of organic matter on biofilm growth and microbial community diversity. Desalination Water Treat. 2017, 66, 10–16. [Google Scholar] [CrossRef]
- Ahmerkamp, S.; Jajaluddin, F.M.; Cui, Y.; Brumley, D.R.; Pacherres, C.O.; Berg, J.S.; Stocker, R.; Kuypers, M.M.M.; Koren, K.; Behrendt, L. Simultaneous visualization off low field sand oxygen concentrations to unravel transport and metabolic processes in biological systems. Cell Rep Method. 2022, 2, 100216. [Google Scholar] [CrossRef] [PubMed]
- Tsagkari, E.; Connelly, S.; Liu, Z.; McBride, A.; Sloan, W.T. . The role of shear dynamics in biofilm formation. NPJ Biofilms Microbiomes. 2022, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Manoel, C.M.; Nunes, O.C.; Melo, L.F. Dynamics of drinking water biofilm in flow/non-flow conditions. Water Res. 2007, 41: 551-562.
- Laspidou, C.S.; Rittmann, B.E.; Karamanos, S.A. Finite element modeling to expand the UMCCA model to describe biofilm mechanical behavior. Water Sci Technol. 2005, 52, 161e166. [Google Scholar] [CrossRef]
- Simunič, U.; Pipp, P.; Dular, M.; Stopar, D. The limitations of hydrodynamic removal of biofilms from the dead-ends in a model drinking water distribution system. Water Res. 2020, 178, 115838. [Google Scholar] [CrossRef] [PubMed]
- Mello, F.L. Biofilm formation and its role in fixed film processes. In Handbook of Water and Wastewater Microbiology, 1st ed.; Mara, D., Horan, N., Eds.; Academic Press: London, United Kingdom, 2003; pp. 337–349. [Google Scholar]
- Liu, T.; Cheng, Y.F.; Sharma, M.; Voordouw, G. Effect of fluid flow on biofilm formation and microbiologically influenced corrosion of pipelines in oilfield produced water. J Petroleum Sci Eng. 2017, 156, 451–459. [Google Scholar] [CrossRef]
- Salgar-Chaparroa, S.J.; Lepkova, K.; Pojtanabuntoeng, T.; Darwin, A.; Machuca, L.L. Nutrient level determines biofilm characteristics and subsequent impact on microbial corrosion and biocide effectiveness. Environ Microbiol. 2020, 86, e02885-19. [Google Scholar] [CrossRef] [PubMed]
- Legner, M.; McMillen, D.R.; Cvitkovitch, D.G. Role of dilution rate and nutrient availability in the formation of microbial biofilms. Front Microbiol. 2019, 10, 916. [Google Scholar] [CrossRef]
- Landini, P. Cross-talk mechanisms in biofilm formation and responses to environmental and physiological stress in Escherichia coli. Res Microbiol. 2009, 160, 259–266. [Google Scholar] [CrossRef]
- Tsai, Y-P. Simulation of biofilm formation at different assimilable organic carbon concentrations under lower flow velocity condition. J Basic Microbiol. 2005, 45, 475–485. [Google Scholar] [CrossRef]
- Shaikh, S.; Rashid, N.; Onwusogh, U.; McKay, G.; Mackey, H.R. Effect of nutrients deficiency on biofilm formation and single cell protein production with a purple non-sulphur bacteria enriched culture. Biofilms. 2023, 5, 100098. [Google Scholar] [CrossRef]
- Shao, D.; Li, J.; Li, J.; Tang, R.; Liu, L.; Shi, J.; Huang, Q.; Yang, H. Inhibition of gallic acid on the growth and biofilm formation of Escherichia coli and Streptococcus mutans. J Food Sci. 2015, 80, M1299–M1305. [Google Scholar] [CrossRef] [PubMed]
- Elasri, M.O.; Miller, R.V. Study of the response of a biofilm bacterial community to UV radiation. Appl Environ Microbiol. 1999, 65, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Norwood, D.E.; Gilmour, A. The differential adherence capabilities of two Listeria monocytogenes strains in monoculture and multispecies biofilms as a function of temperature. Lett Appl Microbiol. 2001, 33, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Hostacká, A.; Ciznár, I.; Stefkovicová, M. Temperature and pH affect the production of bacterial biofilm. Folia Microbiol. 2010, 55, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival and persistence. Front Cell Infect Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Macia, M.D.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin Microbiol Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Elabed, H.; Bakhrouf, A.; Hamza, R.; Gaddour, K. Evidence of the adaptive response in Pseudomonas aeruginosa to 14 years of incubation in seawater. Ann Microbiol. 2012, 62, 1385–1394. [Google Scholar] [CrossRef]
- O’Neal, L.; Baraquet, C.; Suo, Z.; Parsek, M.R. The Wsp system of Pseudomonas aeruginosa links surface sensing and cell envelope stress. PNAS. 2022, 119, e2117633119. [Google Scholar] [CrossRef]
- Melamed, J.; Kocev, A.; Torgov, V.; Veselovsky, V.; Brockhausen, I. Biosynthesis of the Pseudomonas aeruginosa common polysaccharide antigen by D-Rhamnosyltransferases WbpX and WbpY. Glycoconj J. 2022, 39, 393–411. [Google Scholar] [CrossRef]
- Azimi, S.; Thomas, J.; Cleland, S.E.; Curtis, J.E.; Goldberg, J.B.; Diggle, S.P. O-Specific Antigen-Dependent surface hydrophobicity mediates aggregate assembly type in Pseudomonas aeruginosa. mBio. 2021, 12, e0086021. [Google Scholar] [CrossRef]
- Silva, E.C.; Gomes, V.T.; Pragana, L.G.; Bandeira, J.A.C.; Santos, L.F.A.; Travassos, R.A.; Vasconcelos, U. Adherence reduction of Pseudomonas aeruginosa UFPEDA 416 under blue led light irradiation and curcumin exposure. Contemporary J. 2023, 3, 4437–4454. [Google Scholar] [CrossRef]
- Diaz, A.; Dixit, A.R.; Khodadad, CLM, Hummerick ME, Justiano-Velez Y-A, Li W, O’Rurke A. Biofilm formation is correlated with low nutrients and simulated microgravidity conditions in a Bulkholderia isolate from the ISS water processor assembly. Biofilm. 2023, 5, 100110. [Google Scholar] [CrossRef]
- Bernardi, S.; Anderson, A.; Macchiarelli, G.; Hellwig, E.; Cieplik, F.; Vach K, Al-Ahmad A. Subinhibitory antibiotic concentrations enhance biofilm formation of clinical Enterococcus faecalis isolates. Antibiotics. 2021, 10, 874. [Google Scholar] [CrossRef]
- Chada, J.; Khullar, L.; Gulati, P.; Chhibber, S.; Harjai, K. Anti-virulence prospects of Metformin against Pseudomonas aeruginosa: A new dimension to a multifaceted drug. Microb Pathog. 2013, 183, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.N.; Silveira, W.C.D.; Teixeira, L.E.M.; Araújo, I.D. Mechanisms of bacterial adhesion to biomaterials. Rev Med Minas Gerais. 2013, 23, 96–101. [Google Scholar] [CrossRef]
- Rosman, C.W.K.; van der Mei, H.C.; Sjollema, J. Influence of sub-inhibitory concentrations of antimicrobials on micrococcal nuclease and biofilm formation in Staphylococcus aureus. Sci Rep. 2021, 11, 13241. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Mekni, M.A.; Domenico, P.; Nguyen, D.; Jacques, M. Biofilms: Microbial shelters against antibiotics. Microb Drug Resist. 2017, 23, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Wojnicz, D.; Tichaczek-Goska, D. Effect of sub-minimum inhibitory concentrations of ciprofloxacin, amikacin and colistin on biofilm formation and virulence factors of Escherichia coli planktonic and biofilm forms isolated from human urine. Braz J Microbiol. 2013, 44, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.A.; Elsherbini, A.M.; Shaldam, M.A. Repurposing metformin as a quorum sensing inhibitor in Pseudomonas aeruginosa. Afr Health Sci. 2017, 17, 808–819. [Google Scholar] [CrossRef]
- Zuo, J.; Shen, Y.; Wang, H.; Gao, S.; Yuan, S.; Song, D.; Wang, Y.; Wang, Y. Effects of metformin on Streptococcus suis LuxS/AI-2 quorum sensing system and biofilm formation. Microb Pathog. 2023, 181, 106183. [Google Scholar] [CrossRef]
Figure 1.
Simplified diagram of microcosms without aeration (A) and with aeration (B).
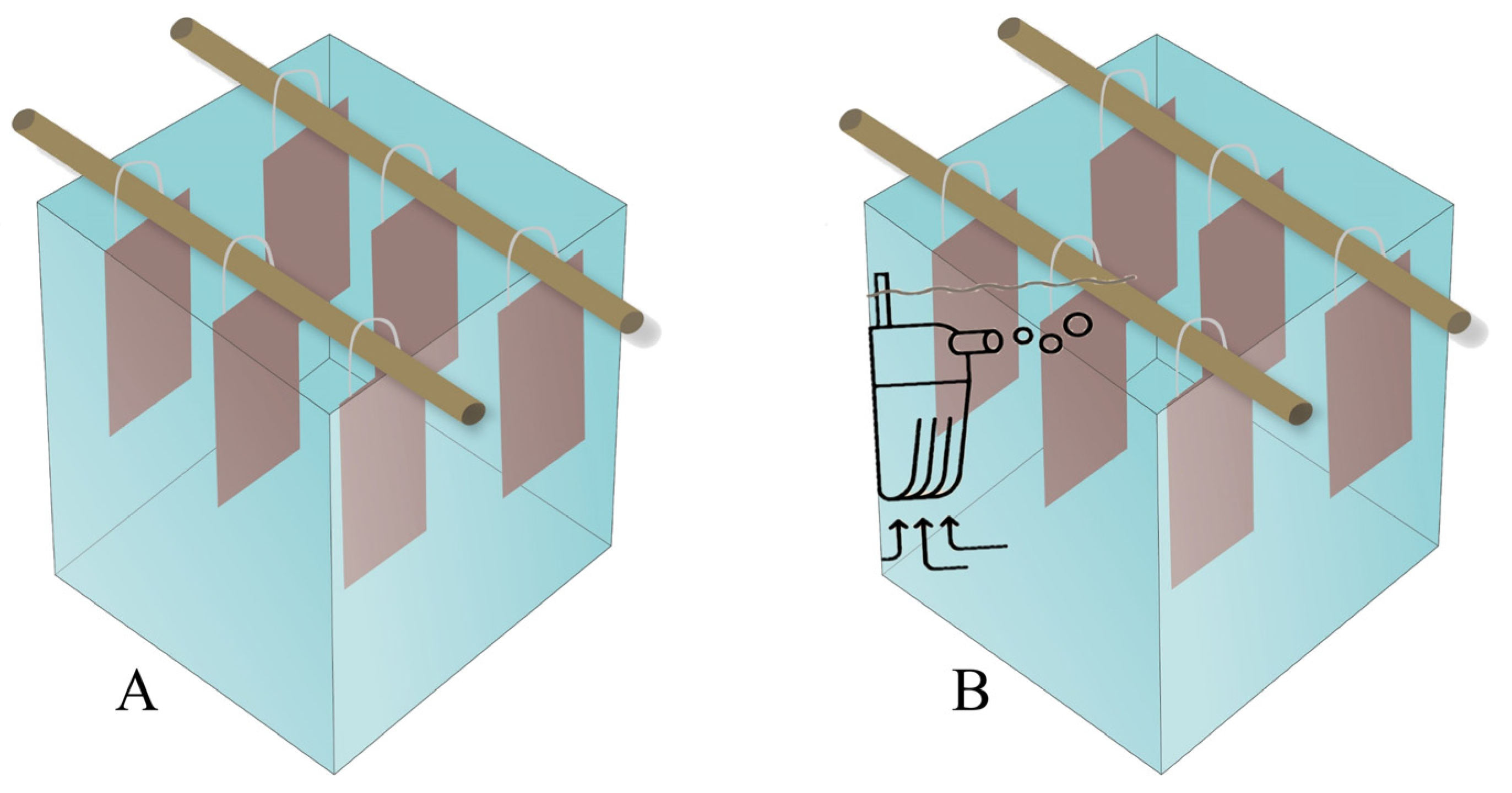

Table 1.
Percentages of reduction or increase in Pseudomonas aeruginosa TGC04 biofilm, compared to the control. The asterisk indicates antibiofilm activity of metformin.
Table 1.
Percentages of reduction or increase in Pseudomonas aeruginosa TGC04 biofilm, compared to the control. The asterisk indicates antibiofilm activity of metformin.
| Test | Metformin (µg/mL) | COD (mg/L) | Flow rate (L/min) | Reduction (%) | Δ24-2h 1 | Type of adhesion (24h) | |
| 2h | 24h | ||||||
| 1 | 100 | 1000 | 0 | 60.00±0.02 | 72.64±0.02 | + 12.64 | Weakly |
| 2 | 100 | 1000 | 4 | 13.85±0.03 | 8.22±0.04 | – 5.63 | Highly |
| 3 | 100 | 10000 | 0 | 72.48±0.02 | 8.33±0.02 | – 64.15 | Highly |
| 4 | 100 | 10000 | 4 | 32.10±0.03 | 32.47±0.03 | + 0.37 | Moderately |
| 5 | 200 | 1000 | 0 | 65.88±0.02 | 67.92±0.02 | + 2.04 | Moderately |
| 6 | 200 | 1000 | 4 | 84.40±0.07* | 44.44±0.02 | – 39.96 | Moderately |
| 7 | 200 | 10000 | 0 | 75.38±0.01 | 49.32±0.03 | – 26.06 | Moderately |
| 8 | 200 | 10000 | 4 | 75.31±0.01 | 24.68±0.03 | – 50.63 | Weakly |
1 Positive numbers indicate reduction and negative numbers indicate an increase in biofilm growth, compared to the control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



