Submitted:
06 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
The goal of this study was to characterize the initial growth trajectory of Epicrates assisi, 1 Epicrates cenchria, and Epicrates crassus snakes raised in captivity. An exponential model with 2 parameters for birth weight (A) and growth rate (k) was fitted for each animal. There was a significant 3 difference A and k among the three species, the means (standard error) for A and k parameters were 4 31 (0.3) g and 0.0027 (0.00006) for E. assisi, 27.6 (0.6) g and 0.0046 (0.00013) for E. cenchria and 25.9 5 (1.4) g and 0.0033 (0.00027) for E. crassus. We showed a significative variability between species for 6 initial growth trajectory. In addition, we also demonstrated the existence of variability within species, 7 which can be used to change the growth trajectory of snakes, being a tool for selection and genetic 8 improvement.9
Keywords:
growth curve
; performance
; ex situ
; rainbow boa
; reproduction
‡ These authors contributed equally to this work.
1. Introduction
The Epicrates genus is composed of five species which are endemic at the neotropical region, occurring from Nicaragua to northwestern Argentina. In Brazil they are distributed across almost all biomes [15]. They are popularly known as “salamanta” or “rainbow boa” due to the iridescent effect of the dorsal scales under the incidence of light. All species of Epicrates are carnivorous constrictors with aglyphous dentition.
Snakes have increasingly been bred as pets around the world. Their diet in the wild consists of small mammals, amphibians, lizards, and birds. In captivity, their diet consists of mice for the smaller and younger snakes and rats for the larger ones [17]. Pet rainbow boas are usually sold once they are large enough to receive a transponder-type microchip with a universal identification number, which occurs between the third and fifth month of life.
The use of growth curves is important to understand the development of an animal both in captivity and in the wild. Fitting of nonlinear growth curves models allows to summarize a massive age-weight databases into a few parameters for studying and characterize the growth of the population; identify the superior animals at younger ages; evaluate the development of animals over time and study the interactions of responses of treatments with time [25]. Once genetic variability allows individuals to respond in different ways to changes that occur in the environment and are therefore, of great importance for the evolution of species.
Knowing the growth trajectory of these snakes, as well as the factors that influence their parameters, is important to predict the growth of animals in a population and explore the best management strategies. As sexual maturity is directly related to body size, low growth rate may result in reproductive delay of the animals. Therefore, the objective of this study was to analyze the variability and characterize the growth trajectory in the early phase of captive bred Epicrates assisi, Epicrates cenchria, and Epicrates crassus.
2. Materials and Methods
A database with repeated measurements of age-weight records of Epicrates assisi, Epicrates cenchria and Epicrates crassus snakes, born in captivity between 2017 and 2021, was provided by a commercial snake farm located in Betim, Minas Gerais (Brazil).
Animals were identified and weighed after birth and housed individually in clear plastic boxes measuring 22.2 x 15 x 6 cm with paper towel substrate. A bowl type water trough and an artificial heating source per box were used to provide a thermal gradient with a temperature range between 33°C to 35°C. The room had also had lighting (photoperiod 12:12 hours), temperature and humidity controlled. The animals were weighed every fifteen days on electronic scales accurate to one gram.
The first feeding of the offspring occurred after skin moulting, it was always used mouse (Mus musculus) neonates. Neonates of mice (Rattus norvegicus) could be offered at subsequent feedings. All animals were fed weekly at the rate of 5 to 10% of body weight and the water was provided ad libitum.
As this is an observational study with a database obtained under commercial conditions and uncontrolled variations, there was neither a planned experimental design nor the need for protocol to be approved by the Ethics Committee (CEUA).
This is an example of a quote.
2.1. Data Editing
Only data from animals (females and males) E. assisi, E. cenchria, and E. crassus species born between the years 2017 and 2021, with at least five valid weightings, the first weighing being performed until the 30th day of life and the last weighing after 180 days of life were considered. After editing, the database was composed of 7210 weight data from six hundred and twenty animals, as presented in Table 1. For the individual trajectories of the animals see Figure 1.
There were variations in relation to age at first weighing. From the years 2017 and 2019, the animals were weighed earlier while the animals born in 2020 and 2021 were weighed later, especially for E. assisi in 2021. Because the data was obtained from a commercial breeding farm, the variations were not controlled as well as management strategies varied over the seasons of birth which explains the differences in relation to age at first weighing of animals.
No specific pattern was observed for maximum age at weighing across years. However, the maximum age was higher for animals of the E. crassus species, intermediate for E. assisi and lower for E cenchria, specifically in the years 2020 and 2021. As these animals are destined for sale, weight taking is closed on the eve of the animal leaving the farm. This sales order is also related to the popularization of each species. E. cenchria are more valuable and desired because they exhibit more intense colors and because of the number of rare color pattern variations obtained by the hatchery. Thus, animals of this species are sold faster than others. Estimates of the A, and k parameters of an exponential model were obtained for each animal using the nls_table function of the forestmangr package [6] of the R software [23]. The exponential model used was as follows:
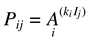
where:
- Pij represents the weight of animal i, at age j – expressed in grams;
- Ai represents the weight of animal i, at age j – expressed in grams;
- ki represents the weight of animal i, at age j – expressed in grams;
- Ij represents the weight of animal i, at age j – expressed in grams;
- represents the weight of animal i, at age j – expressed in grams;
The estimates of parameters A(Aˆi ) and k(kˆi ) of each animal were used to predict their weights at the same ages they were measured P(Pˆi j)). Next, the following adjustment criteria were calculated for each animal:
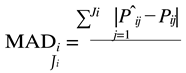
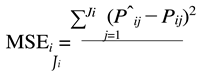
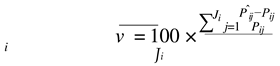
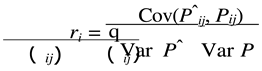
where:
- MADi i represents the mean of the absolute values of the deviations of animal i – expressed in grams;
- Jirepresents the total number of measurements of animal i;
- MSEirepresents the mean squared residues of animal i – expressed in grams2;
- vi represents the average of the adjustment bias for animal i – expressed in percentage;
- rirepresents the correlation between predicted and observed values of animal i;
- Cov(Pˆij, Pij) represents the covariance between predicted and observed values for the weight of animal i at age j;
- Var(Pˆij ) represents the variance of the predicted weights of animal i at age j;
- Var(Pij) represents the variance of the observed weights of animal i at age j; and the other terms as defined above.
2.2. Analysis of Variance and Contrasts
The four fitting statistics, described above, and the estimates of the A and k parameters were considered as variables response and subjected to analyses of variance considering the following statistical model:
yijkl = µ + Ei + Sj + Ak + ESij + EAik + SAjk + eijkl
where:
yijkl represents the value of the response variable in species i, sex j, born in year k, and repetition l;
µ represents a general constant present in all observations;
Ei represents the effect of species i;
Sjrepresents the effect of gender j;
Ak represents the effect of birth year k;
ESij, EAik, SAjk represent the effects of the double interactions of the factors described above; and eijkl represents the random error associated with each observation.
The null hypotheses for differences between means (for some factor of the statistical model, for example: species, sex, and year of birth) adjusted (for the other factors of the model) were tested by means of contrasts, using Tukey’s method for p-value adjustment in multiple comparisons, using the options of the emmeans package [12] of the R software [23].
3. Results
Estimates of birth weight and relative growth rate (Table 2) along with analyzes of the adjusted individual growth trajectories for variations between and within species (Figure 2).
The adjustment criteria indicated that the exponential model fit well with the snake 111 growth trajectory. It was not possible to adjust other non-linear models, such as Gompertz, 112 Richards, Von Bertalanffy among others, since the data analyzed contemplated only the 113 exponential growth phase, being necessary to measure the weight for a longer period 114 until a phase in which there was a decrease in growth rate and weight stabilization was 115 identified.
The parameters indicating birth weight (A) and relative growth rate (k) were significantly influenced by species, year of birth, and the interaction of these factors (Table 3). The relative growth rate was also influenced by the interaction of sex and year of birth (Table 3).
E. assisi was born heavier than E. cenchria and E. crassus (p<0.05). There was no 120 difference (p>0.05) between birth weight of E. cenchria and E. crassus. The relative growth 121 rate of E. cenchria was higher than the other two species (p<0.05) (Table 4).
There was no significant difference for adjusted A in relation to the year of birth. In general, it is noticeable that the mean growth rates (k) decreased over time, however the only significant statistical differences (p<0.05) were in the years 2018 x 2019 and 2018 x 2021 (Table 5). The species x year of birth interaction was significant because there was a change in the magnitude of the year effect within species.
4. Discussion
The parameters indicating birth weight (A) and relative growth rate (k) were signifi- cantly influenced by species, year of birth, and the interaction of these factors (Table 3). The relative growth rate was also influenced by the interaction of sex and year of birth (Table 3).
E. assisi was born heavier than E. cenchria and E. crassus (p<0.05). There was no difference (p>0.05) between birth weight of E. cenchria and E. crassus. The relative growth rate of E. cenchria was higher than the other two species (p<0.05) (Table 4).
There was no significant difference for adjusted A in relation to the year of birth. In general, it is noticeable that the mean growth rates (k) decreased over time, however the only significant statistical differences (p<0.05) were in the years 2018 x 2019 and 2018 x 2021 (Table 5). The species x year of birth interaction was significant because there was a change in the magnitude of the year effect within species.
The exponential model yi = A eki although not indicated for a long period of growth, is relatively common because of its simplicity. Growth can be easily described with only the initial and final weights [8]. Where A is the estimated initial weight; k is the relative growth rate and i is the age. Thus, following the development of Epicrates snakes after sexual maturity, after two to or three years, would be an excellent complement to this work.
Although E. assisi is endemic to the Caatinga biome (from the states of Piauí to southern Bahia and northern Minas Gerais in Brazil) it may occur in syntopy with E. crassus in southwestern Bahia state to northern Minas Gerais in transition areas between Caatinga and Cerrado [15]. Such characteristics may explain the similarity in growth profile between the species.
E. assisi had a higher birth weight in relation to the other species. This fact may be an adaptation to the conditions of their natural habitat, which is characterized by extreme droughts and dry periods typical of the semi-arid climate. Heavier offspring may have more chances of survival even with the lower availability of food.
E. cenchria inhabits the Amazon rainforest, considered to have the greatest biodiversity on the planet. The abundance of food and the favorable climate optimizes the development of this species, which making it one of the largest and heaviest snakes of the Epicrates family, possibly exceeding 2.20 meters.
The Cerrado biome presents an abundance of prey intermediate between Caatinga and Amazon rainforest. This fact may bring the ‘Cerrado salamanta’ closer to both the Caatinga and Amazon ‘salamanta’ being found similarities in birth weight between E. crassus and E. cenchria. Variations were detected regarding the individual growth trajectory and relative growth rate between and within species, these differences may have occurred because growth patterns vary, and the model used was not robust enough to fit all growth patterns in the same way. However, variability can be caused by some not identified difference in management or due to genetic differences between animals of the same species, which can be a meaningful material for the evolution of species. To better understand the variability, it would be necessary to have proper design in which the family of the parents of these animals or the molecular markers are known.
In commercial breeding, knowing the growth behavior of animals from birth is fundamental to the success of the activity. Animals that grow faster may be sold sooner, contributing to the reduction of operational and production costs. In the reproductive bias, fertility tends to increase as alongside the growth of the body of female snakes. Size at birth can be influenced by female size and litter size [11]. Sexual maturity in these species is more related to body weight and size than age. Thus, selecting snakes with higher growth rates until puberty becomes an important tool for genetic selection and herd improvement.
Growth rates among species can be totally different even among species whose adults are similar in size. This difference occurs due to physiological reasons that allow a higher rate of food conversion into mass [24].
Snakes continues to growth even after reaching reproductive status, females accumulate body reserves for long periods before reproduction to effect vitellogenesis. Reproductive performance and body weight may interact and vary according to environmental conditions and food availability [1]. In addition, larger females tend to give birth to larger offspring, which is desirable for the production system.
Some snake taxa exhibit sexual dimorphism by size. Snakes that exhibit male-male combat behavior have larger male individuals than females, while females of highly fertile species are larger than males [23]; [20]; [9]).[21] maintained a litter of Bothrops fonsecai under a controlled feeding regime and showed that females, even though they received the same amount of food as males, showed higher growth rates in all measured parameters.
The authors discuss that sexual dimorphism is independent of the amount of food ingested, but rather connected to the different food conversion between the sexes and suggest that this difference may be linked to the efficiency of energy assimilation by the female. A population of Agkistrodon piscivorus (Lacépède, 1789) was studied for 14 years by Ford (2002) and similar growth rates were found between the sexes at the juvenile stage. However, males grew more after reaching sexual maturity.
5. Conclusions
The present study provided evidence of the existence of significant variability for the initial growth trajectory. E. assissi is heavier than E. cenchria and E. crassus at birth, but the growth rate is higher in E. cenchria than in the other two species. The variability within species can be used to alter the growth trajectory of snakes, being a tool for selection and genetic improvement, since it is to elucidate growth patterns, the relationship between size and age, or even possible to identify the existence of differences between individuals of the same species.
6. Patents
This section is not mandatory, but may be added if there are patents resulting from the work reported in this manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
“Not applicable”.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We thank the students Hebiene Laiane da Silva Lobo, Nathalia Teixeira Garcia, and Maria Luiza da Cunha Cabral for their help in compiling the raw data. Thanks to Nicolas Eleuterio Fontana for his help and comments on the manuscript. This work was supported by Capes -Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior, CNPq- Conselho Nacional de Desenvolvimento Científico e Tecnológicico end Fapemig- Fundação de Amparo à pesquisa de Minas Gerais.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds ... are available from the authors.
References
- Andrews, R.M.T. Patterns of growth in Reptiles. Biologyof the Reptilia 1982, 13, 273–320. [Google Scholar]
- Beaupre, S.J.; Duvall, D.; O’Leile, J. Ontogenetic variation in growth and sexual size dimorphism in a central Arizona population of the western diamondback rattlesnake (Crotalus atrox). Copeia 1988, 40–47. [Google Scholar] [CrossRef]
- Bertona, M.; Chiaraviglio, M. Reproductive biology, mating aggregations, and sexual dimorphism of the Argentine Boa constrictor (Boa constrictor occidentalis). Journal of Herpetology 2003, 37, 510–516. [Google Scholar] [CrossRef]
- Bonnet, X.; Naulleau, G.; Mauget, R. The influence of body condition on 17- estradiol levels in relation to vitellogenesis in female Vipera aspis (Reptilia, Viperidae). General and comparative endocrinology 1994, 93, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, X.; Shine, R.; Naulleau, G.; Thiburce, C. The influence of body condition on 17- estradiol levels in relation to vitellogenesis in female Vipera aspis (Reptilia, Viperidae). Journal of Zoology 2001, 255, 341–351. [Google Scholar] [CrossRef]
- Braga, S.R.; de Oliveira, M.L.R.; Gorgens, E.B. forestmangr: Forest mensuration and management. 2021. R package version 0.9.
- Ford, N.B. Ecology of the Western cottonmouth (Agkistrodon piscivorus leucostoma) in Northeastern Texas. Biology of the Vipers. 2002, USA, 167–177. [Google Scholar]
- Gamito, S. Growth models and their use in ecological modelling: an application to a fish population. Ecological modelling 1988, 113, 83–94. [Google Scholar] [CrossRef]
- Hendry, C.R.; Guiher, T.J.; Pyron, R.A. Ecological divergence and sexual selection drive sexual size dimorphism in new world pitvipers (Serpentes: Viperidae). Journal Of Evolutionary Biology 2014, 27, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Hill III, J.G.; Beaupre, S.J. Body size, growth, and reproduction in a population of Western Cottonmouths (Agkistrodon piscivorus leucostoma) in the Ozark Mountains of northwest Arkansas. Copeia 2008, 1, 105–114. [Google Scholar] [CrossRef]
- King, R.B.; Bittner, T.D.; Queral-Regil, A.; Cline, J.H. Sexual dimorphism in neonate and adult snakes. Journal of Zoology 1999, 247, 19–28. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: estimated marginal means. R package version 1.4. 4. Am. Stat 2020, 34, 216–221. [Google Scholar]
- Lourdais, O.; Shine, R.; Bonnet, X.; Brischoux, F. Sex differences in body composition, performance and behaviour in the Colombian rainbow boa (Epicrates cenchria maurus, Boidae). Journal of Zoology 2006, 269, 175–182. [Google Scholar] [CrossRef]
- Naulleau, G.; Bonnet, X. Body condition threshold for breeding in a viviparous snake. Oecologia 1996, 107, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Passos, P.; Fernandes, R. Revision of the Epicrates cenchria complex (Serpentes: Boidae). Herpetological Monographs 2008, 22, 1–30. [Google Scholar] [CrossRef]
- Pearson, D.; Shine, R.; Williams, A. Geographic variation in sexual size dimorphism within a single snake species (Morelia spilota, Pythonidae). Oecologia 2002, 131, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S.; Wissink-Argilaga, N. Nutrition—lizards and snakes. Companion Animal 2015, 20, 362–366. [Google Scholar] [CrossRef]
- Reynolds, R.G.; Henderson, R.W. Boas of the world (Superfamily Booidae): a checklist with systematic, taxonomic, and conservation assessments. Bulletin of the Museum of Comparative Zoology 2018, 162, 1–58. [Google Scholar] [CrossRef]
- Shine, R. Intersexual dietary divergence and the evolution of sexual dimorphism in snakes. The American Naturalist 1991, 138, 103–122. [Google Scholar] [CrossRef]
- Stahlschmidt, Z.R.; Davis, J.R.; DeNardo, D.F. Sexual variation in assimilation efficiency: its link to phenotype and potential role in sexual dimorphism. Journal of Comparative Physiology 2011, 181, 383–389. [Google Scholar] [CrossRef]
- Stuginski, D.R.; Mendes, G.F.; Sant’Anna, S.S.; Rubio, D.T.; Vieira, S.E.M.; Oitaven, L.P.C.; Grego, K.F. Sexual differences in growth rates of juveniles from a litter of Bothrops fonsecai: the role of feeding conversion in a female-biased SSD species. South American Journal of Herpetology 2017, 12, 193–199. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. Published online 2021. [Google Scholar]
- Weatherhead, P.J.; Barry, F.E.; Brown, G.P.; Forbes, M.R. Sex ratios, mating behavior and sexual size dimorphism of the northern water snake, Nerodia sipedon. Behavioral Ecology and Sociobiology 1995, 36, 301–311. [Google Scholar] [CrossRef]
- Case, T.J. On the evolution and adaptive significance of postnatal growth rates in the terrestrial vertebrates. The Quarterly review of biology 1978, 53, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Davidian, M.; Giltinan, D.M. Nonlinear models for repeated measurement data: an overview and update. Journal of agricultural, biological, and environmental statistics 2003, 8, 387–419. [Google Scholar] [CrossRef]
- eigel, Richard A. ; Huggins, M.M.; Ford, Neil B. Reduction in locomotor ability as a cost of reproduction in gravid snakes. Oecologia 1987, 73, 481–485. [Google Scholar] [CrossRef]
Figure 1.
Figure 1. Individual growth trajectories observed for female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria, and E. crassus.
Figure 1.
Figure 1. Individual growth trajectories observed for female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria, and E. crassus.
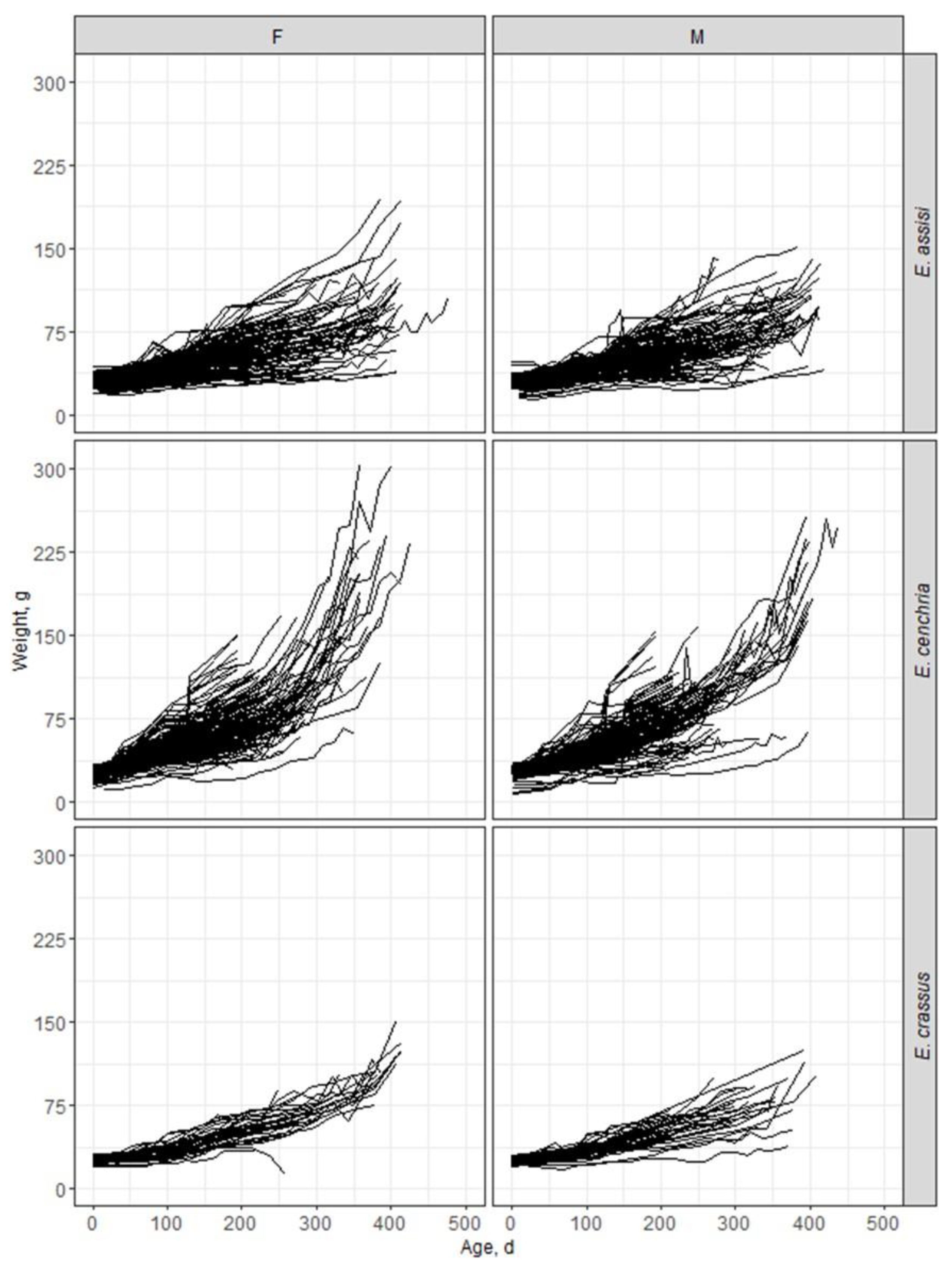
Figure 2.
Individual growth trajectories fitted by an exponential model for female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria, and E. crassus.
Figure 2.
Individual growth trajectories fitted by an exponential model for female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria, and E. crassus.
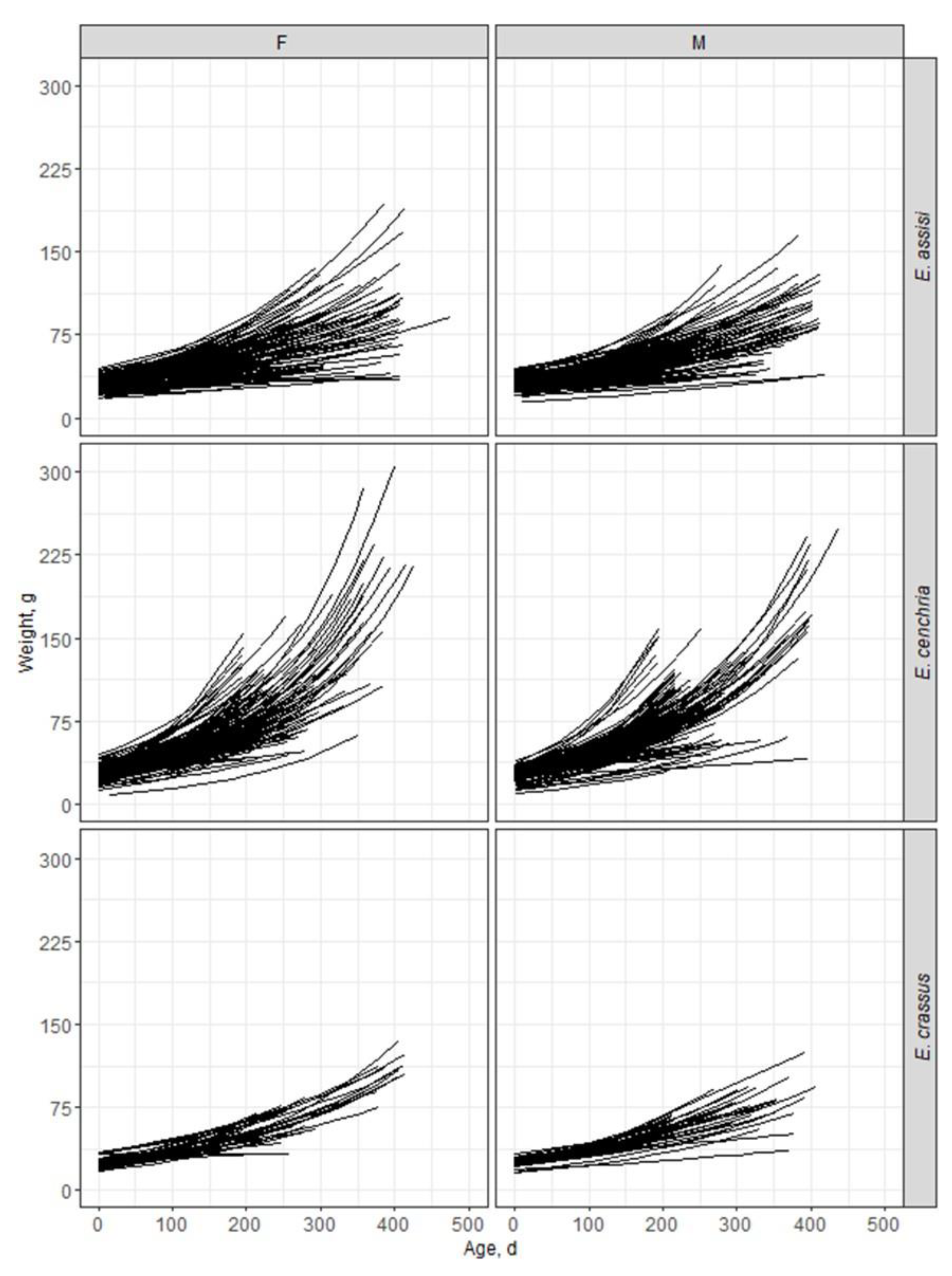

Table 1.
Descriptive statistics regarding the number of animals, weighings, and initial and final ages, according to species and sex.
Table 1.
Descriptive statistics regarding the number of animals, weighings, and initial and final ages, according to species and sex.
| Starting age (days) Final age (days) Number of weighings | ||||||||||||
| Species | Sex | Animals | Med | Min | Max | Med | Min | Max | Med | Min | Max | Total |
| E. assisi | F | 129 | 2.3 | 0 | 25 | 289.6 | 184 | 475 | 12 | 5 | 36 | 1555 |
| E. assisi | M | 138 | 1.9 | 0 | 18 | 281.1 | 184 | 419 | 12.5 | 5 | 36 | 1721 |
| E. cenchria | F | 141 | 1.9 | 0 | 21 | 255.9 | 182 | 425 | 11.72 | 5 | 30 | 1653 |
| E. cenchria | M | 131 | 1.5 | 0 | 15 | 255.7 | 190 | 436 | 10.9 | 6 | 44 | 1426 |
| E. crassus | F | 38 | 0.7 | 0 | 6 | 302.1 | 195 | 415 | 11.2 | 6 | 21 | 425 |
| E. crassus | M | 43 | 0.8 | 0 | 8 | 284.2 | 186 | 406 | 10 | 6 | 21 | 430 |
1 Tables may have a footer.
Table 2.
Means (standard error in parentheses) of fit criteria and estimates of birth weight and relative growth rate of female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria and E. crassus.
Table 2.
Means (standard error in parentheses) of fit criteria and estimates of birth weight and relative growth rate of female (F) and male (M) snakes of the captive-bred species Epicrates assisi, E. cenchria and E. crassus.
| Species | Sex | N | MAD (g) | MSE (g2) | Bias (%) | Corr | A (g) | k (g) |
| E. assisi | F | 129 | 3.175 | 18.940 | 0.857 | 0.918 | 30.974 | 0.00281 |
| E. assisi | M | 138 | (0.134) | (1.711) | (0.085) | (0.009) | (0.488) | (0.00008) |
| E. cenchria | F | 141 | 3.069 | 19.083 | 0.879 | 0.926 | 30.697 | 0.00286 |
| E. cenchria | M | 131 | (0.141) | (2.164) | (0.095) | (0.008) | (0.459) | (0.00008) |
| E. crassus | F | 38 | 4.785 | 41.461 | 0.974 | 0.950 | 27.855 | 0.00486 |
| E. crassus | M | 43 | (0.203) | (3.290) | (0.208) | (0.006) | (0.504) | (0.00012) |
1MAD- mean of the absolute values of the deviations; MSE- mean of the squared residuals; Corr- correlation; A- birth weight; K- relative growth rate.
Table 3.
P-values associated with each source of variation of the statistical model used for the analysis of variance of the fit criteria1 and the estimates of birth weight (A) and relative growth rate (k) of captive bred snakes of the species Epicrates assisi, E. cenchria and E. crassus.
Table 3.
P-values associated with each source of variation of the statistical model used for the analysis of variance of the fit criteria1 and the estimates of birth weight (A) and relative growth rate (k) of captive bred snakes of the species Epicrates assisi, E. cenchria and E. crassus.
| Source of variation | MAD (g) | MSE (g2) | Bias (%) | Correlation | A (g) | k (g) |
| Species (E) | 0.0000 | 0.0000 | 0.0000 | 0.0006 | 0.0000 | 0.0000 |
| Sex (S) | 0.4487 | 0.5853 | 0.4321 | 0.4914 | 0.2816 | 0.4669 |
| Year of birth (A) | 0.0296 | 0.1281 | 0.7897 | 0.0000 | 0.0000 | 0.0000 |
| E x S | 0.3955 | 0.5999 | 0.2920 | 0.1940 | 0.7429 | 0.7746 |
| E x A | 0.0000 | 0.0000 | 0.0000 | 0.0145 | 0.0000 | 0.0000 |
| S x A | 0.5320 | 0.7264 | 0.2951 | 0.0253 | 0.2136 | 0.0283 |
1MAD- mean of the absolute values of the deviations; MSE- mean of the squared residuals; Corr- correlation; A- birth weight; K- relative growth rate.
Table 4.
Means (standard error in parentheses) of birth weight (A) and relative growth rate (k) for snakes of the species Epicrates assisi, E. cenchria and E. crassus, adjusted for the other effects included in the statistical model of the analysis of variance1.
Table 4.
Means (standard error in parentheses) of birth weight (A) and relative growth rate (k) for snakes of the species Epicrates assisi, E. cenchria and E. crassus, adjusted for the other effects included in the statistical model of the analysis of variance1.
| Species | A (g) | k (g) |
| E. assisi | 31.0 (0.3) a | 0.00277 (0.00006) a |
| E. cenchria | 27.6 (0.6) b | 0.00460 (0.00013) b |
| E. crassus | 25.9 (1.4) b | 0.00330 (0.00027) a |
1Means adjusted for the effects of sex, year of birth and by the double interactions involving the effects of species, sex and year of birth. Different letters in the same column indicate that the contrast between means is statistically significant (p<0.05), using Tukey’s method for p-value adjustment in multiple comparisons.
Table 5.
Means (standard error in parentheses) of birth weight (A) and relative growth rate (k) for the year of birth for snakes of the species Epicrates assisi, E. cenchria and E. crassus, adjusted for the other effects included in the statistical model of the analysis variance1.
Table 5.
Means (standard error in parentheses) of birth weight (A) and relative growth rate (k) for the year of birth for snakes of the species Epicrates assisi, E. cenchria and E. crassus, adjusted for the other effects included in the statistical model of the analysis variance1.
| Year of birth | A (g) | k (g) |
| 2017 | 29.0 (1.3) a | 0.00399 (0.00025) ab |
| 2018 | 28.2 (0.4) a | 0.00392 (0.00008) a |
| 2019 | 28.2 (0.4) a | 0.00356 (0.00008) b |
| 2020 | 28.7 (1.8) a | 0.00316 (0.00024) ab |
| 2021 | 26.8 (1.2) a | 0.00316 (0.00024) b |
11Means adjusted for the effects of sex, year of birth and by the double interactions involving the effects of species, sex, and year of birth. Different letters in the same column indicate that the contrast between means is statistically significant (p<0.05), using Tukey’s method for p-value adjustment in multiple comparisons.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



