
Submitted:
05 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Several traps are recommended for monitoring corn earworm, Helicoverpa zea (Boddie), an important pest of field, vegetable and fruit crops in the U.S. These traps, which are meant to capture H. zea in many cases end up capturing other insects. Helicoverpa zea has recently been found feeding on different seeds and dual-type (seed and fiber) hemp, Cannabis sativa L. (Rosales: Cannabaceae) causing serious damage. Limited work has been done on developing integrated pest management (IPM) for H. zea industrial hemp in Missouri. We evaluated the attractiveness of different traps with the aim of developing a monitoring system for the adult male H. zea in industrial hemp fields in two Missouri locations In addition, we recorded other non-target insects in the traps to determine trap selectivity. Commercially available green, clear, tricolor bucket traps Heliothis Scentry and Scentry Delta 1X traps baited with H. zea sex pheromones were evaluated in 2021 and 2022. Tricolor traps captured significantly more adult male H. zea than the other traps in both years. Non-target insects, mainly Xylocopa virginica (L.) (Hymenoptera: Apidae), Bombus spp (Hymenoptera: Apidae), and Apis mellifera L. (Hymenoptera: Apidae) were also captured. The tricolor trap captured the most bees. The presence of non-target species highlights the risk of using tricolor traps as a monitoring tool. This data provides information for planning the monitoring of corn earworm in industrial hemp farms in Missouri.
Keywords:
Cannabis sativa
; Helicoverpa zea
; non-target species
; pest
; pheromone-baited traps
1. Introduction
Production of industrial hemp, C. sativa has increased steadily in the U.S. since the introduction of the 2014 Farm Bill and the Agricultural Improvement Act of the 2018 Farm Bill [1,2]. Hemp has been grown in the U.S. since the early 1600s, reaching peak production in the mid-1800s [3], with Missouri being second in hemp production in the U.S. [4]. Hemp production was, however, restricted following the passage of the Marijuana Tax Act in 1937 [5]. As a result, between 1958, when the last hemp crop was planted in Wisconsin, and 1999, when a permit was issued for experimental test plots in Hawaii, no hemp was legally grown in the U.S. until the 2014 Farm Bill was amended [2].
Recognizing the potential benefits of hemp in the U.S. economy, Congress approved the legalization of hemp in the U.S. Currently, all U.S. states have federally legalized industrial hemp (herein referred to as “hemp”) production [1]. The value of hemp production in the open and under protection for the United States totaled $238 million in 2022 [6]. This includes food and body products, clothing, auto parts, building materials, and other products [7]. Since hemp has huge potential and has been approved for cultivation, it may become a field crop in the U.S., which may be grown in large acreages. As the production of this crop continues and expands, change in the arthropod complex and their abundance and damage is expected.
Like most field crops, hemp is infested by several arthropod species [8,9], and the importance of these arthropod complexes may be significant as production of this crop continues to expand. Approximately 272 insects and mites are associated with Cannabis spp. globally [10]. In the U.S., about 150 insects and several arthropod pests are associated with hemp production and the pest management needs of some of the pests have been described [8,9]. However, little to no information exists on the taxa and diversity of insects that attack hemp in Missouri. Based on our preliminary work in the 2020 hemp growing season, we found that corn earworm, Helicoverpa zea (Boddie, 1850), yellow-striped armyworm, Spodoptera ornithogalli (Guenée), brown stink bug, Euschistus spp. (Hemiptera: Pentatomidae), Southern green stink bug, Nezara viridula L. (Hemiptera: Pentatomidae), and green stink bug, Chinavia halaris (Say) (Hemiptera: Pentatomidae), and several other arthropods have established themselves as key pests of seed, dual (hybrid) and cannabinoid/floral varieties of hemp. Among these pests, H. zea [8], has demonstrated the greatest potential for crop injury in Missouri hemp. This report confirms previous observations by other researchers across several states [9,11,12,13]. The larvae of H. zea cause significant damage to a diverse array of host plants, including some of the most cultivated crops like corn, tomato, pepper, soybean, etc [14]. Helicoverpa zea preferentially feeds on the floral regions of CBD and seeds of grain variety hemp [13].
Although, corn earworms, and the associated plants they infests, have been around for over a century [13], several studies have shown the population dynamics of this insect in many host plant plants except in industrial hemp. Limited knowledge exists on their monitoring except for work done using black light and various pheromone baited traps mainly in corn (Zea mays L.) [17,18,19]. In addition, some studies have identified that the type of trap is critical when monitoring lepidopteran pests because of their potential to attract other non-targets like bees. One major trap design used in many monitoring studies is bucket traps with different colors and shapes. Also, the type of agricultural practices and landscape composition at various scales are known to shape the composition of bee and other arthropod communities [15,16,20,21].
Hemp is a prolific pollen producer, and most males produce an abundance of pollen that serves as vital subsistence resources to bees at a point in the season when they are resource-limited [22]. Hemp flowers attract a diverse range of bee species, and taller varieties of hemp attract a greater diversity of bees, making hemp a valuable resource for bees during floral scarcity and potentially benefitting other crops in the agricultural landscape [56]. Therefore, any pest monitoring in hemp should consider the effect that will have on bee communities.
Since hemp is a new crop in Missouri, it is important to develop an IPM program that will permit better decision-making on how to suppress pest populations below the economic injury level. A critical step in developing an IPM program for any key pest depends on a good monitoring program that provides vital information on pests’ abundance and seasonality. Various methods have been used to monitor pest moths, including the use of pheromone-baited sticky traps [19,23,24], black light traps [25,26,27,28], drop cloths [29,30] and pheromone-baited traps [19,31,32]. The performance of these traps in attracting corn earworms has not been well studied, particularly in hemp. In addition to these traps attracting lepidopteran pests, they have also been shown to attract other nontarget species (“bycatch”), such as bees. A good monitoring trap should, however, attract more of the target pest but few other nontargets.
In this study, we evaluated three trap types, namely the bucket, Heliothis Scentry, and Scentry Delta 1X traps that are frequently used to monitor several species of adult male moths in corn and other crops. These traps have produced different results in different agricultural production systems, particularly on bycatches of bees. Therefore, our objective was to evaluate and determine the effectiveness of some commercially available traps and sex pheromones for monitoring H. zea and their bycatch of bees in Missouri hemp fields. Ultimately, we aim to identify and recommend a good trap for effectively monitoring corn earworm adults while reducing bycatch of other nontarget insects in industrial hemp.
2. Materials and Methods
2.1. Study Area and Crop Management
The experiments were conducted in two Missouri locations, Lincoln University George Washington Carver, Jefferson City, MO (38o31’33.202”N 92o8’26.606”W) (herein referred to as Carver Farm), and Mid-West Natural Fiber AGRIPAK farm site located in Sikeston, MO (36o 55’ 28.51”N 89o 37’37.128”W) (herein referred to as Sikeston Farm) in 2021 and 2022 growing seasons (Figure 1). These two locations represent the state’s Central and Southeast regions. Each field consisted of ~2 ha plots planted with different hemp varieties and types (dual, seed, and fiber). In both years, 32 varieties of 7 dual types, and 25 fiber types were planted in a variety trial. The dual varieties were planted at seeding rates of ~ 22.68 kg/ha and 18.14 kg/ha for the seed and fiber varieties respectively. Seeds of all types and varieties were planted directly in the soil using a seeder mounted on a tractor. Seeding was done between May 15-22 each year and at each location. The plots were not managed in terms of weed control and pesticide applications. Agricultural weeds were abundant during the late seasons at all sites, which is more common in hemp than many other crops because no herbicides are registered for use on hemp in Missouri. The plots were irrigated when needed using overheard sprinkler irrigation systems.
2.2. Description of experimental plots
At both locations, hemp seeds were planted on rows of 3 m by 30 m plots (Figure 1 A and B) and inter-row spacing of about 2.5 m at Carver Farm and 6 m at Sikeston Farm, depending on the tractor used.
Figure 1.
Layout of plots in A) 2021 and B) 2022. Traps were randomized in each row. A row was considered a block within which traps were randomized. In both years, each trap received the same corn earworm lure. Because the lures were the same, the maximum of 30 m (A) and minimum of 15 (B) distances between the traps in a row and between rows or block (20 m) was deemed adequate for insects to respond to the trap since this work did not test different lures. The only factor was the type of trap.
Figure 1.
Layout of plots in A) 2021 and B) 2022. Traps were randomized in each row. A row was considered a block within which traps were randomized. In both years, each trap received the same corn earworm lure. Because the lures were the same, the maximum of 30 m (A) and minimum of 15 (B) distances between the traps in a row and between rows or block (20 m) was deemed adequate for insects to respond to the trap since this work did not test different lures. The only factor was the type of trap.
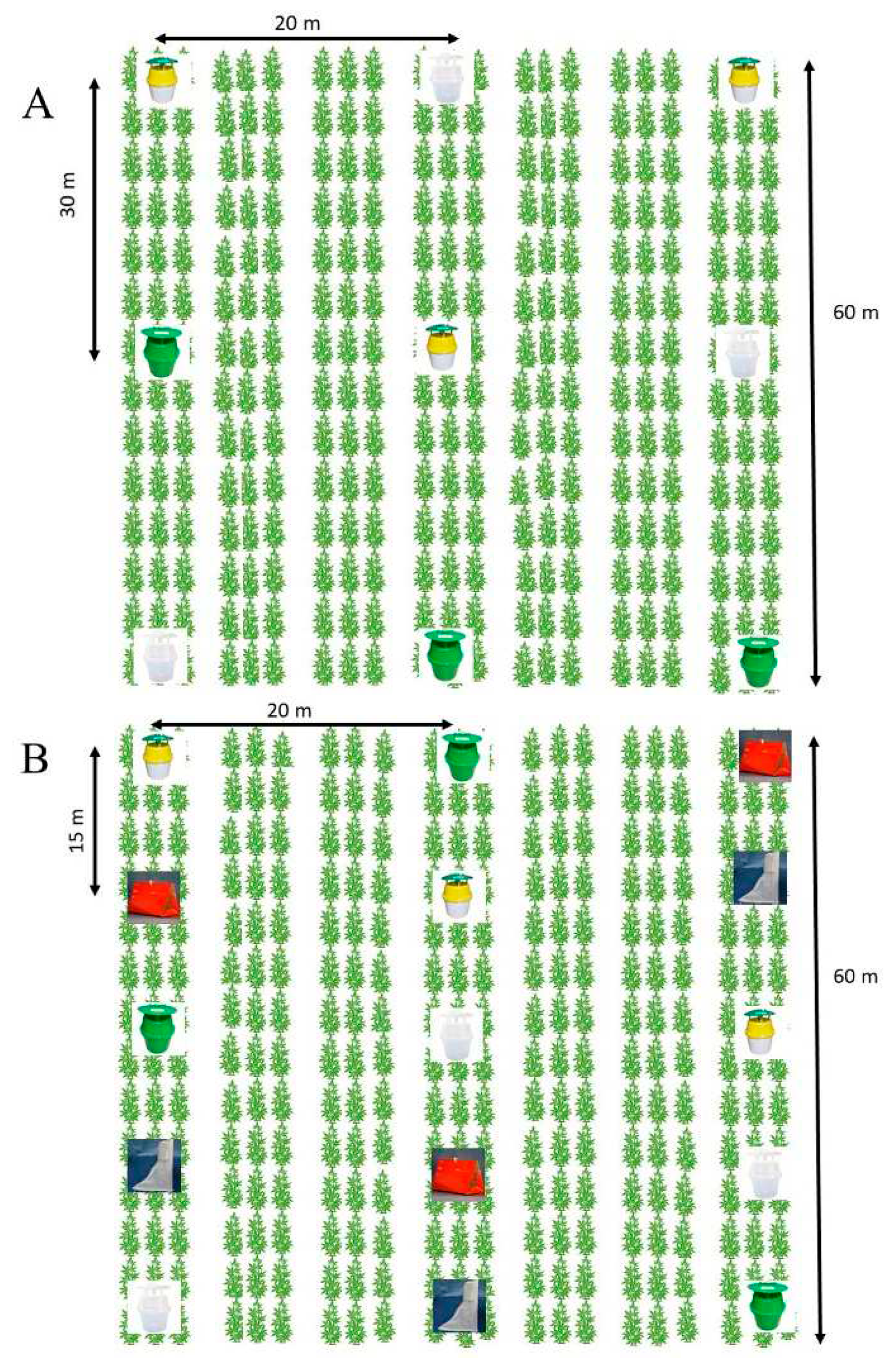
2.2. Sampling of Moths Using Pheromone-baited Trap
In 2021, three Trécé bucket traps of different colors (tricolor, green, and clear) baited with Trécé™ H. zea pheromone lure were evaluated to determine their performance in attracting male H. zea (Figure 1 A). The traps and lures were purchased from Great Lakes IPM (Vestaburg, MI) and deployed in each site from August through September. The tricolor trap consisted of a white bucket (0.13 m tall and 0.16 m in diam.), a yellow funnel (bottom opening 0.03 m) on top of the bucket, and a dark green cover (0.16 m diam.) attached to the funnel by four posts that allowed a 0.03-m circular space in between cover and funnel (Figure 2-A). The pheromone lure was placed in a green basket (0.05 m long) that hung in the middle of the green cover. The green (Figure 2-A) and the clear (Figure 2-B) bucket traps were similar in shape and size to the tricolor trap (Figure 2-C), except that the entire unit consisted of green and clear colors, respectively. The pheromone lures for the green and clear bucket trap were placed in a green and clear basket-top cover. Traps were hung on 1.52 m wooden stakes using office clips and metal strings, so the trap height was about 1.45 m above ground. Helicoverpa zea, rubber septa lures were replaced every six weeks. Traps within each block were rotated monthly. The mechanism of moth capture was by direct flight into the bucket through the entrance in the trap top. Moths attracted to the pheromone “fell” downwards (usually after flying inside the posts and bouncing off the top cover) through the funnel into the bucket where they were killed through starvation. Traps within blocks and the separation among blocks were at least 15.00 m and 20.00 m apart respectively. Traps were inspected weekly unless conditions did not permit. Adults of male H. zea from the traps were removed, held in Ziploc® bags, counted, and recorded during each visit. Extreme care was taken to avoid the escape of live adults. Sampling was performed once every week. Although our focus was on H. zea, we also collected data on non-targets mainly bees (Hymenoptera: Apidae): Eastern carpenter bee Xylocopa virginica (L.), bumble bee Bombus spp. and honeybee Apis mellifera L. Each trap block was replicated three times and included one of each H. zea pheromone-baited bucket trap. In 2022, two additional traps, Heliothis Scentry [(HS), Figure 2-E] and Scentry Delta 1X [(SD-1X), Figure 2-D] traps, and Trécé™ were included except at the Sikeston farm where the HS trap was not included due to availability (Figure 3). For convenience, these traps are hereafter coded HS and SD-1X, respectively. Traps and lures were also purchased from Great Lakes IPM and deployed in each trapping site from May to September. The SD-1X trap measured 0.18 m in length by 0.10 m in width by 0.13 m height, with a sticky inner liner (0.17 m length by 0.10 m width by 0.09 m height). Moths captured in the SD-1X trap generally died overnight. Heliothis Scentry trap was composed of two cones. The base cone measured 0.80 m long, with a bottom opening of 0.34 m that narrowed to 0.15 m at the top; the apex cone measured 0.27 m long, with a bottom opening of 0.15 m that narrowed to 0.06 m at the top. The bottom portion of the apex cone was secured to the top portion of the base cone with Velcro material. The lure was placed along a cord stretching across the bottom of the base cone. Moths captured in the HS traps died over a couple of days. Traps were also hung on wooden stakes as previously described in 2021. Helicoverpa zea rubber septa lures were replaced every six weeks. Traps within each trap block were rotated monthly. Similarly, traps were replicated three times but included one of each H. zea pheromone-baited bucket traps (clear, green, and tricolor), HS, and SD-1X traps (Figure 2). Traps within blocks and the separation among blocks were at least 15 m apart. Traps were inspected weekly unless conditions did not permit. Adults of male H. zea captured were removed from the traps, held in Ziploc® bags, counted, destroyed, and removed from the field during each visit. Data on non-targets, mainly Eastern carpenter, bumble, and honeybees, were also recorded.
2.3. Statistical Analysis
Data for each year and location were first analyzed for ANOVA's normality and heteroscedasticity assumptions. Data on H. zea larvae and adults that did not meet the assumptions of ANOVA were square root transformed. Where both actual and transformed data did not meet the assumptions of ANOVA, a generalized linear model was performed assuming Poisson distribution and log-link function to determine the effects of trap type, sampling date, and trap type * sampling date interaction. Where the trap type and sampling date interaction was significant, a one-way ANOVA was used to determine the performance of the trap types in each sampling date. A nonparametric Kruskal Wallis rank sum test was performed, and a Chi-square approximation (χ2) was used to determine the significant effect of trap type in each sampling date where applicable. All data were analyzed using JMP (JMPIN version 19.0, SAS Institute 2019) and the results were considered statistically significant at P < 0.05.
3. Results
3.1. Trap Performance in 2021
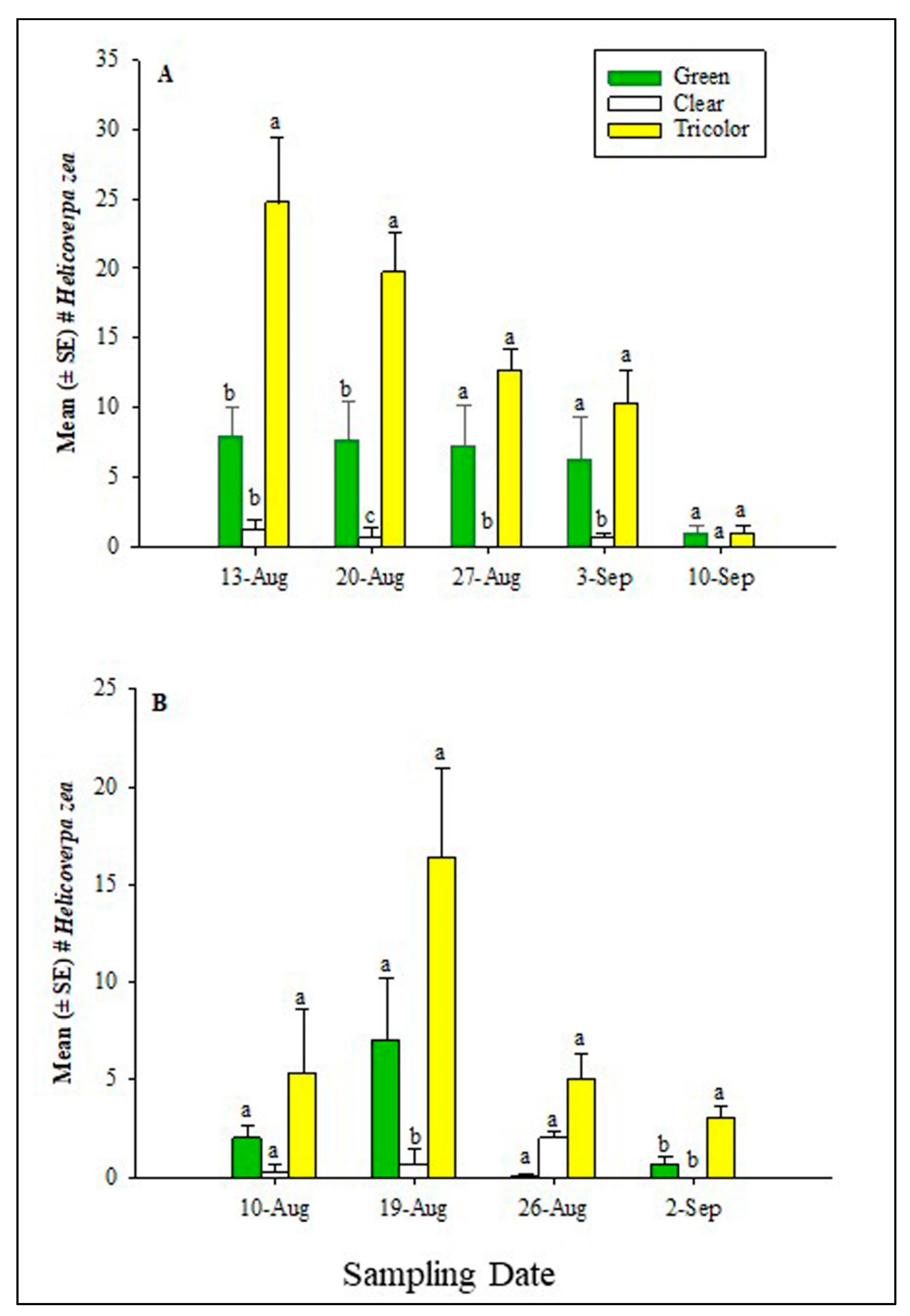
In 2021, at the Carver Farm, the GLM showed significant differences among sampling date (χ2 = 41.7, df = 4, P < 0.0001), trap type (χ2 = 86.8, df = 2, P < 0.0001), but sampling date * trap type interaction was not significant (χ2 = 9.6, df = 2, P = 0.2901) for the adult H. zea captured in the traps. Analysis based on sampling date, showed significant differences on August 13 (χ2 = 7.3, df = 2; P = 0.0257), August 20 (χ2 = 7.3, df = 2, P = 0.0265), August 27 (χ2 = 6.7, df = 2, P = 0.0349), and September 3 (χ2 = 7.3, df = 2, P = 0.0496). However, no significant difference occurred in trap captures on September 10 (χ2 = 2.9, df = 2, P = 0.2301). In most of the sampling dates, tricolor traps captured significantly or numerically more H. zea than clear and green traps (Figure 2a). At the Sikeston Farm, the GLM showed a significant difference for only the sampling date (χ2 = 33.4, df = 3, P < 0.0001), but trap type (χ2 = 3.5, df = 2, P = 1.0000) and sampling date * trap type interaction (χ2 = 1.5, df = 6; P = 0.9586) were not significant. When the data were analyzed in terms of sampling date, the traps showed a marginally significant difference on August 19 (χ2 = 6.2, df = 2; P = 0.0457) and September 2 (χ2= 6.0, df = 2, P = 0.0495) but not on August 10 (χ2 = 2.9, df = 2, P = 0.2301).and August 26 (χ2 = 0.00, df = 2; P = 1.0000) (Figure 2b). The tricolor trap captured significantly and numerically more H. zea male adults compared to clear and green traps.
Figure 3.
Mean (± SE) total number of adult male H. zea in traps at (A) Lincoln University Washington Carver Farm, Jefferson City, MO, and (B) Sikeston Farm during the 2021 hemp season. Different letters above bars indicate a significant difference (P > 0.05) among treatments within each sampling date. Mean separation was done using the Wilcoxon Chi-square approximation test.
Figure 3.
Mean (± SE) total number of adult male H. zea in traps at (A) Lincoln University Washington Carver Farm, Jefferson City, MO, and (B) Sikeston Farm during the 2021 hemp season. Different letters above bars indicate a significant difference (P > 0.05) among treatments within each sampling date. Mean separation was done using the Wilcoxon Chi-square approximation test.
3.3. Trap Performance in 2022
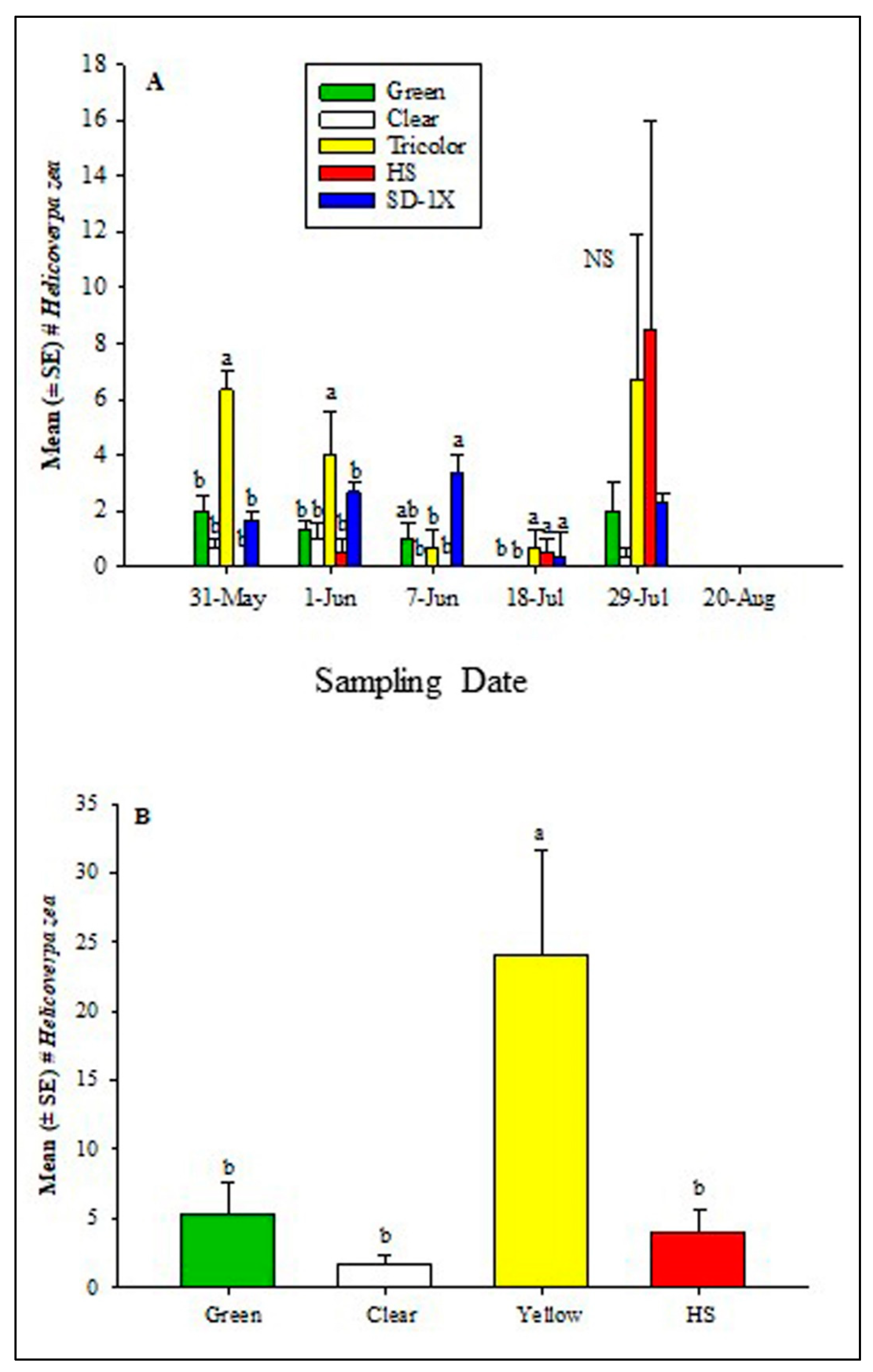
In 2022, at the Carver Farm, the GLM showed significant differences among sampling dates (χ2 = 21.6, df = 4, P = 0.0002), trap type (χ2 = 48.1, df = 4, P < 0.0001), and sampling date * trap type interaction (χ2 = 85.7, df = 16, P < 0.0001). When the data were analyzed in terms of sampling date, the traps differed significantly on May 31 (χ2 = 10.6, df = 4, P = 0.0309), June 1 (χ2 = 10.9, df = 4; P = 0.0281), June 7 (χ2 = 8.9, df = 4, P = 0.0542), July 18 (χ2 = 9.3, df = 4, P = 0.0548) and September 3 (χ2 = 7.3, df = 2; P = 0.0496). However, no significant difference was found on July 29 (χ2 = 4.3, df = 2, P = 0.3613) (Figure 4a). In all the sampling dates, tricolor traps captured significantly or numerically more H. zea than green, clear, HS, and SD- 1X traps except on July 29 when HS recorded numerically higher H. zea males (Figure 4a). At Sikeston, only one sampling date was recorded due to distance. The GLM showed a marginally significant difference for only the trap type (χ2 = 7.6, df = 3, P = 0.0547) (Figure 4b). Similarly, the tricolor trap captured significantly more H. zea than green, clear, or HS traps.
Figure 4.
Mean (± SE) the total number of adult male H. zea traps at (A) Lincoln University Washington Carver Farm, Jefferson City, MO, and (B) Sikeston Farm, MO, during the 2022 hemp season. There was only one sampling date at the Sikeston location. Different letters above the bars indicate a significant difference (P > 0.05) among treatments within each sampling date. Mean separation was done using the Wilcoxon Chi-square approximation test.
Figure 4.
Mean (± SE) the total number of adult male H. zea traps at (A) Lincoln University Washington Carver Farm, Jefferson City, MO, and (B) Sikeston Farm, MO, during the 2022 hemp season. There was only one sampling date at the Sikeston location. Different letters above the bars indicate a significant difference (P > 0.05) among treatments within each sampling date. Mean separation was done using the Wilcoxon Chi-square approximation test.
3.4. Captures of Common Pollinators in 2021
In 2021, at the Carver Farm, a total of 204 bees were captured in the traps. Among the bees captured, 45.1% were carpenter bees, 28.3% were bumble bees and 26.5% were honeybees. The tricolor trap captured significantly more bees compared with clear traps, while no significant difference occurred between tricolor and green traps for all bee species (Table 1). At the Sikeston Farm, a total of 352 bees were captured in the traps. Among the bees captured, 19.6% were carpenters, 65.3% were bumble and 15.5% were honeybees. In both locations, the tricolor traps had significantly more bees compared with clear and green traps (Figure 5; Table 1).
3.5. Captures of Common Pollinators in 2022
In 2022, at the Carver Farm, the traps baited with H. zea lures captured a total of 553 (N = 75) bees. Among the bees captured, 15.9% were carpenter bees, 66.0% were bumble bees and 17.2% were honeybees. At the Sikeston Farm where only one sampling date was performed, the traps that contained H. zea, captured a total of 117 bees (N =12). Among the bees captured, 15.9% were carpenter bees, 66.0% were bumble bees and 17.2% were honeybees. In both locations, the tricolor trap had significantly and numerically more bees than green, clear, HS and SD-1X traps (Figure 4; Table 2).
4. Discussion
In this study, we sought to dtermine the capture of different traps for corn earworm monitoring and identify the by-catch of the bee community captured in the corn earworm pheromone-baited traps in hemp. In 2021, we found significant differences in captures among the different bucket traps with the tricolor bucket trap recording significantly more adult H. zea males than green and clear bucket traps in most of the sampling dates. In 2022, the tricolor trap was still better at capturing more H. zea even when HS and SD-1X traps were added. We attribute the capture in the traps as due to the trap design and color because the same pheromone dispensers, pheromone load, and release rates, lures longevity, and trap height were used. Insects use olfactory and visual stimuli to locate resources that have been used to monitor many insects successfully [33,34], particularly moths, flies, and beetles. We consider that both visual stimuli provided by the tricolor trap and olfactory stimuli provided by the H. zea pheromone contributed to H. zea being attracted to the traps.
Although both olfactory and visual stimuli acted to attract H. zea, color played a major role in the performance of the traps since there was a significant difference between the traps with the same design but different colors and those with different designs. The response of insects to color varies, but color can enhance trap numbers in many situations. In a similar study which evaluated different traps in capturing wild male velvet bean caterpillar moths (VBC), Anticarsia gemmatalis Hübner (Lepidoptera: Erebidae), and fall armyworm moths (FAW), Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), significantly fewer VBC and FAW males were captured in pheromone-baited mono-colored (forest green) bucket traps than standard multicolored bucket traps [35]. Our inability to measure the reflectance and hue of the different trap colors used in this study is a major setback. However, several authors have shown that the compound eyes of some noctuid moths have a bimodal sensitivity to light with peaks in the UV (350 – 370 nm) and green (500 – 575 nm) regions [35,36]. Yet, other noctuid species respond more strongly to traps emitting low spectral reflectance in these regions [37,38]. Although green and tricolor are ubiquitous colors in the landscape, it is reasonable to expect that combining these colors will enhance trap capture. However, show that not all of these colors can enhance trap captures [39]. Adding white with 100% reflectance to the tricolor trap may have contributed to the enhanced attractiveness of this trap.
The capture of different bee species in the traps was not surprising because several studies investigating pheromone traps for moth monitoring resulted in the capture of non-targets including bees [40,41,42]. Due to its temporally unique floral phenology, hemp has the potential to provide an important food resource to diverse bee communities during periods of flower scarcity, thereby providing support for other crops in landscapes across agroecosystems [56].
The main non-target species found in the traps were carpenter, bumble, and honeybees. These were particularly high in numbers in the tricolor bucket trap, contrary to the report that bees usually respond to blue color. Studies on a different project using blue vane traps support the hypothesis that bees respond to blue color [Akotsen-Mensah unpublished]. In 2021 and 2022, a total of 757 bees were captured in the traps. Bycatch of non-target insects in monitoring traps has been shown to increase trap processing time, decrease trap effectiveness for the target species, and can include beneficial arthropods that provide important ecosystem services [40,41]. Beneficial species frequently captured in monitoring traps might negatively impact their populations and the services they provide.
Hymenopteran pollinators comprise much of the insect bycatch in moth pheromone-baited traps in various agroecosystems [43,44,45,46,47]. Although the impact of bee removal from agroecosystems in monitoring traps that target moth pests is unknown [44], it may negatively influence local bee abundance and species diversity, which could alter pollination services for both wild plants and managed crops and could reduce crop yields [48,49]. As there is already widespread recognition of global declines of wild bee populations [48,49,50], practices that systematically remove bees from agricultural landscapes should be avoided. Although the tricolor traps did significantly better than green and clear, their ability to also attract bees may not make this trap a good monitoring tool. The reason is that four out of five North American bumble bee species, one of which was captured in our study have been earmarked on the list of Endangered Species Act [51]. Since pollinator declines can have cascading effects on associated ecosystems [52,53,54], any trap that will attract pollinators may not be accepted by growers [55].
If the tricolor bucket trap should be used to monitor H. zea in the hemp field, it will be appropriate to find a way to reduce its ability to capture bees and other pollinators. At this point, we can recommend the green bucket trap as an alternative since it was also able to attract H. zea but had fewer bees. The high number of bees in the field may be due to the search for pollen from the male hemp plant, which was in abundance in the field. Since the plot we used was a variety trial with many male flowers that shed pollen, it is possible many bees came to the field in search of pollen and eventually ended up being captured by the traps.
5. Conclusions
This study has confirmed that H. zea (both adults and larvae) can be present in industrial hemp fields in Missouri from the time of planting (usually mid-May) until harvest (end-September). The presence of these insects may pose significant threats to hemp seed production. Tricolor bucket traps should be installed early in the season to detect these insects. However, due to concern about the tricolor trap capturing bees, particularly carpenter and bumble bees, this trap can be substituted with the green color bucket trap, if the monitoring goal is to detect the insects. Our results have provided information on monitoring these insects, particularly H. zea. Although our research aims to use the trap numbers to make management decisions, it is too early to use the trap numbers to guide insecticide applications. We suggest further studies to determine the economic threshold of insect damage and landscape and weather factors to better understand the population dynamics and economic damage of these insects. The information obtained can be useful for effective insecticide applications for production profitability and environmental sustainability of industrial hemp in Missouri.
Data Availability Statement
Data are available upon request.
Acknowledgments
The authors thank the farm managers of Lincoln University Carver Farm, Eric Groose and Troy Wieberg. We also thank Jeff Limbaugh and Patrick Van Meter of Mid-West Natural Fiber AGRIPAK Farm at Sikeston, MO for technical assistance. Garrett Avery helped to transport students to the sites to establish plots and collect data. The authors acknowledge Drs. Henry Ofosuhene-Sintim and Kamalendu Paul for reviewing the draft manuscript. This research was supported by funding from the USDA Capacity Building Grant awarded to Lincoln University Hemp Institute (Award # 2021-38821-34726).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adesina, I.; Bhowmik, A.; Sharma, H.; Shahbazi, A. A review on the current state of knowledge of growing conditions, agronomic soil health practices and utilities of hemp in the United States. 2020, Agric. 10. [CrossRef]
- Mark, T.; Shepherd, J.; Olson, D.; Snell, W.; Proper, S.; Thornsbury, S. 2020. Economic Viability of Industrial Hemp in the United States: A review of state pilot programs. United States Dep. Agric. Econ. Res. Serv. 2020, 1–77. [CrossRef]
- Tancig, M.; Kelly-Begazo, C.; Kaur, N.; Sharma, L.; Brym, Z. Industrial hemp in the United States: Definition and History. Edis. 2021, 2021: 2–4. [CrossRef]
- Horner, J.; Ohmes, A.; Massey, R.; Luce, G.; Bissonnette, K.; Milhollin, R.; Lim, T.; Roach, A.; Morrison, C.; Schneider. R. Missouri Industrial Hemp Production. 2019, 1–45.
- Johnson, N. American Weed: A History of cannabis cultivation in the United States. EchoGéo. 2019, 0–22. [CrossRef]
- NASS. National Hemp Report (April 2023) 29 USDA, National Agricultural Statistics Service. 2023, National Hemp Report 04/19/2023 (cornell.edu).
- Agricultural Marketing Resource Center. A national information resource for value- added agriculture. 2023, (https://www.agmrc.org/commodities-products/fiber/industrial-hemp).
- McPartland, J.; Clarke, R.; Watson, D. Hemp diseases and pests management and biological control. 2000, CABI Publisher.
- Cranshaw, W.; Schreiner, M.; Britt, K.; Kuhar, T. P.; McPartland, J.; Grant, J.; Bogran, C. Developing Insect Pest Management Systems for Hemp in the United States: A Work in Progress. J. Integr. Pest Manag. 2019, 10. [CrossRef]
- Mostafa, A. R. ; Messenger. P. S. lnsects and mites associated with plants of the 45 genera Argemone, Cannabis, Glaucium, Erythroxylum, Eschscholtzia, Humulus, and Papaver. Unpubl. manuscript, Univ. California, Berkeley, 1972.
- Schreiner, M. A survey of the arthropod fauna associated with hemp (Cannabis sativa L.) Grown in Eastern Colorado (Doctoral dissertation, Colorado State University), 2019. [CrossRef]
- Ajayi, O. S.; Samuel-Foo, M. Hemp pest spectrum and potential relationship between Helicoverpa zea station and hemp production in the United States in the face of climate change. Insects. 2021, 12: 1–11. [CrossRef]
- Britt, K. E.; Kuhar, T. P.; Cranshaw, W.; McCullough, C. T.; Taylor, S. V.; Arends, B. R.; Burrack, H.; Pulkoski, M.; Owens, D.; Tolosa, T. A.; Zebelo, S.; Kesheimer, K. A.; Ajayi, O. S.; Samuel-Foo, M.; Davis, J. A.; Arey, N.; Doughty, H.; Jones, J.; Bolt, M.; Fritz, B. J.; Grant, J. F.; Cosner, J.; Schreiner, M. Pest management needs and limitations for corn earworm (Lepidoptera: Noctuidae), an emergent key pest of hemp in the United States. J. Integr. Pest Manag. 2021, 12: 1–11. [CrossRef]
- Arey, N. C.; Lord, N. P. ; Davis. J. A. Evaluation of Hemp (Cannabis sativa) (Rosales: Cannabaceae) as an alternative host plant for polyphagous Noctuid Pests. J. Econ. Entomol. 2022, 115: 1947–1955. [CrossRef]
- Le Feon, V. , Schermann-Legionnet, A., Delettre, Y., Aviron, S., Billeter, R., Bugter, R., Hendrickx, F., Burel, F. Intensification of agriculture, landscape composition and wild bee communities: a large scale study in four European countries. Agric. Ecosyst. Environ. 2010, 137: 143–150. [CrossRef]
- Senapathi, D. , Biesmeijer, J.C., Breeze, T.D., Kleijn, D., Potts, S.G., Carvalheiro, L.G. Pollinator conservation — the difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12: 93–101. [CrossRef]
- Weber, D. C.; Ferro, D. N. Nontarget noctuids complicate integrated pest management monitoring of sweet corn with pheromone traps in Massachusetts. J. Econ. Entomol. 1991, 84: 1364–1369. [CrossRef]
- Coop, L. B.; Drapek, R. J.; Croft, B. A.; Fisher, G. C. Relationship of corn earworm (Lepidoptera: Noctuidae) pheromone catch and silking to infestation levels in Oregon sweet corn. J. Econ. Entomol.1992, 85: 240–245. [CrossRef]
- Guerrero, S.; Brambila, J.; Meagher, R. L. Efficacies of four pheromone-baited traps in capturing male Helicoverpa (Lepidoptera: Noctuidae) Moths in northern Florida. Florida Entomol. 2014, 97: 1671–1678. [CrossRef]
- Lichtenberg, E. M. , Mendenhall, C. D., Brosi, B. Foraging traits modulate stingless bee community disassembly under forest loss. J. Anim. Ecol. 2017, 86(6): 1404-1416. [CrossRef]
- Grab, H., E. J. Blitzer, B. Danforth, G. Loeb, and K. Poveda. Temporally dependent pollinator competition and facilitation with mass flowering crops affects yield in co-blooming crops. Sci. Rep. 2017, 7: 45296. [CrossRef]
- Dalio, J. S. Cannabis sativa – an important subsistence pollen source for Apis. J. Pharm. Biol. Sci. 2012, 1: 1–3.
- Byers, J. A.; Naranjo, S. E. Detection and monitoring of pink bollworm moths and invasive insects using pheromone traps and encounter rate models. J. Appl. Ecol. 2014, 51: 1041– 1049. [CrossRef]
- Ferracini, C.; Pogolotti, C.; Lentini, G.; Saitta, V.; Busato, E.; Rama, F.; Alma, A. Performance of pheromone-baited traps to monitor the seasonal abundance of tortrix moths in chestnut groves. Insects. 2020, 11: 1–15. [CrossRef]
- Falcon, L.; Van Den Bosch, R.; Etzel, L.; Ferris, C.; Stromberg, L. Light traps as detection devices for moths of cabbage looper and bollworm. Calif. Agric. 1967, 21: 12–14. [CrossRef]
- Hayes, J. L. Dynamics of nocturnal activity of moths in the Heliothis complex (Lepidoptera: Noctuidae) in cotton. J. Econ. Entomol.1991, 84: 855–865. [CrossRef]
- Gregg, P. C.; Fitt, G. P.; Coombs, M.; Henderson, G. S. Migrating moths (Lepidoptera) collected in tower-mounted light traps in northern New South Wales, Australia: species composition and seasonal abundance. Bull. Entomol. Res. 1993, 83: 563–578. [CrossRef]
- Harrison, T.; Berenbaum, M. R. Moth diversity in three biofuel crops and native prairie in Illinois. Insect Sci. 2013, 20: 407–419. [CrossRef]
- Studebaker, G. E.; Spurgeon, D. W.; Mueller, A. J. Calibration of ground cloth and sweep net sampling methods for larvae of corn ear worm and soybean looper (Lepidoptera: Noctuidae) in soybean. J. Econ. Entomol.1991, 84: 1625–1629. [CrossRef]
- Swenson, S. J.; Prischmann-Voldseth, D. A.; Musser, F. R. Corn Earworms (Lepidoptera: Noctuidae) as pests of soybean. J. Integr. Pest Manag. 2013, 4: 1–8. [CrossRef]
- Pearson, A. C.; Sevacherian, V.; Ballmer, G. R.; Vail, P. V.; Henneberry, T. J. Population dynamics of Heliothis virescens and H. zea (Lepidoptera: Noctuidae) in the Imperial Valley of California. Environ. Entomol. 1989, 18: 970–979. [CrossRef]
- Parajulee, M. N.; Rummel, D. R.; Arnold, M. D.; Carroll, S. C. Long-term seasonal abundance patterns of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) in the Texas High Plains. J. Econ. Entomol. 2004, 97: 668–677. [CrossRef]
- Piñero, J. C.; Jácome, I.; Vargas, R.; Prokopy, R. J. 2006. Response of female melon fly, Bactrocera cucurbitae, to host-associated visual and olfactory stimuli. Entomol. Exp. et Appl. 2008, 121 (3), 261-269. [CrossRef]
- Pinero, J. C.; Souder, S. K.; Vargas, R. I. Vision-mediated exploitation of a novel host plant by a tephritid fruit fly. PLoS One, 2017, 12 (4), e0174636. [CrossRef]
- Mitchell, E. R.; Agee, H. R.; Heath, R. R. Influence of pheromone trap color and design on capture of male velvetbean caterpillar and fall armyworm moths (Lepidoptera: Noctuidae). J. Chem. Ecol. 1989, 15: 1775–1784. [CrossRef]
- Agee, H. R. Spectral sensitivity of the compound eyes of field-collected adult bollworm and tobacco budworms. Ann. Entomol. Soc. Am. 1973, 66: 613–615. [CrossRef]
- McLaughlin, J. R.; Brogdon, J. E.; Agee, H. R.; Mitchell, E. R. Effect of trap color on captures of male cabbage looper and soybean looper in double-cone pheromone traps. J. Georg. Entomol. Soc. 1975, 10: 174–179. [CrossRef]
- Méndez, A.; Martín, L.; Arines, J.; Carballeira, R.; Sanmartín, P. Attraction of insects to ornamental lighting used on cultural heritage buildings: A case study in an urban area. Insects. 2022, 13(12): 1153. [CrossRef]
- Knodel, J. J.; Agnello, A. M. Field comparison of nonsticky and sticky pheromone traps for monitoring fruit pests in western New York. J. Econ. Entomol. 1990, 83: 197–204. [CrossRef]
- Cha, D. H.; Hesler, S. P.; Park, S.; Adams, T. B.; Zack, R. S.; Rogg, H.; Loeb, G. M.; Landolt, P. J. Simpler is better: fewer non-target insects trapped with a four-component chemical lure vs. a chemically more complex food-type bait for Drosophila suzukii. Entomol. Exp. Appl. 2015,154: 251–260. [CrossRef]
- Landolt, P.; Zhang, Q.-H. Discovery and development of chemical attractants used to trap pestiferous social wasps (Hymenoptera: Vespidae). J. Chem. Ecol. 2016, 42: 655–665. [CrossRef]
- Grocock, N. L.; Batallas, R. E.; McNamara, E. A.; Sturm, A. B.; Manson, J. S.; Evenden, M. L. Bumble bees (Hymenoptera: Apidae) respond to moth (Lepidoptera: Noctuidae) pheromone components, leading to bee bycatch in monitoring traps targeting moth Pests. Front. Ecol. Evol. 2020, 8: 576692. [CrossRef]
- Adams, R. G.; Murray, K. D.; Los, L. M. Effectiveness and Selectivity of Sex Pheromone Lures and Traps for Monitoring Fall Armyworm (Lepidoptera: Noctuidae) Adults in Connecticut Sweet Corn. J. Econ. Entomol. 1989, 82: 285–290. [CrossRef]
- Meagher, R. L.; Mitchell, E. R. Nontarget hymenoptera collected in pheromone- and synthetic floral volatile-baited traps. Environ. Entomol. 1999, 28: 367–371. [CrossRef]
- Knight, A. L.; Miliczky, E. Influence of trap colour on the capture of codling moth (Lepidoptera: Tortricidae), honeybees, and non-target flies. J. Entomol. Soc. Brit. Columbia. 2003, 100: 65–70.
- Aurelian, V. M.; Evenden, M. L.; Judd, G. J. R. Diversity and abundance of arthropod by-catch in semiochemical-baited traps targeting apple clearwing moth (Lepidoptera: Sesiidae) in organic and conventional apple orchards in British Columbia, Canada. Can. Entomol. 2015, 147: 227–243. [CrossRef]
- Spears, L. R.; Looney, C.; Ikerd, H.; Koch, J. B.; Griswold, T.; Strange, J. P.; Ramirez, R. A. Pheromone Lure and Trap Color A. Environ. Entomol. 2016, 45: 1009–1016. [CrossRef]
- Goulson, D. Conserving wild bees for crop pollination. Food, Agric. Environ. 2003, 1: 142–144.
- Potts, S. G.; Biesmeijer, J. C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W. E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25: 345– 353. [CrossRef]
- Zattara, E. E.; Aizen, M. A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth. 2021, 4:114–123. [CrossRef]
- Graves, T. A.; Janousek, W. M.; Gaulke, S. M.; Nicholas, A. C.; Keinath, D. A.; Bell, C. M. , Cannings, S.; Hatfield, R. G.; Heron, J. M.; Koch, J. B.; Loffland, H. L.; Richardson, L. L.; Rohde, A. T.; Rykken, J.; Strange, J. P.; Tronstad, L. M.; Sheffield, C. S. 2020. Western bumble bee: declines in the continental United States and range-wide information gaps. Ecosphere. 2020, 11. [CrossRef]
- Biesmeijer, J. C.; Roberts, S. P.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A. P.; Potts, S. G.; Kleukers, R. J.; Thomas, C. D.; Settele, J.; Kunin, W. E. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, (80-.). 313: 351–354. [CrossRef]
- Cameron, S. A.; Lozier, J. D.; Strange, J. P.; Koch, J. B.; Cordes, N.; Solter, L. F.; Griswold, T. L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. U. S. A. 2011, 108: 662–667. [CrossRef]
- Powney, G. D.; Carvell, C.; Edwards, M.; Morris, R. K. A.; Roy, H. E.; Woodcock, B. A.; Isaac, N. J. B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10: 1–6. [CrossRef]
- Gross, H. R.; Carpenter, J. E. Role of the fall armyworm (Lepidoptera: Noctuidae) pheromone and other factors in the capture of bumblebees (Hymenoptera: Aphidae) by universal moth traps. Environ. Entomol. 1991, 20: 377–381. [CrossRef]
- Flicker, N. R. , Poveda, K., Grab, H. The Bee Community of Cannabis sativa and Corresponding Effects of Landscape Composition. Environ. Entomol. 2019, 49(1): 197–202. [CrossRef]

Table 1.
Mean (± SE) number of common bee species captured per sampling week in traps in 2021.
| Location | Trap Type | Carpenter Bee Mean ± SE |
Bumble Bee Mean ± SE |
Honey Bee Mean ± SE |
| Carver Farm | Green | 1.40 ± 0.53 ab | 1.27 ± 0.51 a | 1.07 ± 0.73 a |
| Clear | 0.47 ± 0.24 b | 0.33 ± 0.19 a | 0.60 ± 0.47 a | |
| Tricolor | 4.27 ± 1.39 a | 2.27 ± 0.88 a | 1.93 ± 1.47 a | |
| χ2 | 9.12 | 5.38 | 0.33 | |
| df | 2, 44 | 2, 44 | 2, 44 | |
| p | 0.0104 | 0.0679 | 0.8473 | |
| Sikeston | Green | 0.58 ± 0.26 b | 1.33 ± 0.61 b | 1.33 ± 0.45 a |
| Clear | 1.08 ± 0.34 b | 2.92 ± 0.94 b | 1.25 ± 0.49 a | |
| Tricolor | 4.08 ± 1.08 a | 14.92 ± 4.67 a | 1.83 ± 0.58 a | |
| χ2 | 9.15 | 9.75 | 0.69 | |
| df | 2, 33 | 2, 33 | 2, 33 | |
| p | 0.0103 | 0.0076 | 0.7091 |
Means followed by different letters in the same column are significantly different from each other at p = 0.05. Means followed by the same letter within each column are not significantly different (P > 0.05, Chi-square approximation).
Table 2.
Mean total (± SE) number of bees captured in traps baited with H. zea lures in two locations in 2022.
Table 2.
Mean total (± SE) number of bees captured in traps baited with H. zea lures in two locations in 2022.
| Trap Type | H. zea | |
|---|---|---|
| Carver Farm | Sikeston Farm | |
| Green | 11.00 ± 2.08 b | 10.67 ± 2.33 b |
| Clear | 8.3 ± 2.70 b | 2.67 ± 0.89 b |
| Tricolor | 134.6 ± 56.2 a | 31.67 ± 1.8 a |
| HS | 14.0 ± 2.73 b | NA |
| SD-1X | 16.33 ± 4.9 b | 22.00 ± 6.5 ab |
| χ2 | 8.8 | 8.00 |
| df | 4 | 3 |
| P | 0.064 | 0.0459 |
Means followed by the same letter within each column are not significantly different (P > 0.05, Chi-square approximation). HS is the Heliothis Scentry trap and SD-1X is Scentry Delta trap.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



