
Submitted:
08 January 2024
Posted:
09 January 2024
You are already at the latest version
Abstract
The aim of the study was to determine the occurrence and characteristics of coagulase-positive Staphylococcus strains in the carcasses of wild birds and aviary capercaillies in south-eastern Poland. Samples from capercaillies and wild birds belonging to 12 orders and 26 species were tested. The resistance of the strains and the presence of selected virulence genes were examined. Similarity analysis of the isolates was performed using pulsed-field gel electrophoresis (PFGE). Coagulase-positive Staphylococcus strains accounted for 5.7% and belonged to the species: Staphylococcus aureus, Staphylococcus pseudintermedius and Staphylococcus delphini. Among isolated strains, 15.8% were multidrug resistant. The most frequently detected virulence genes were hla in 58% of isolates and hlb and hld in 47.4% of isolates., The results of multiplex PCR showed the presence of genes responsible for the production of enterotoxins C, B, E, and J, in single isolates. It can be concluded that coagulase-positive Staphylococcus strains accounted for a small percentage of staphylococci isolated from free-living birds in the study area. The occurrence of multidrug-resistant coagulase-positive Staphylococcus strains in aviary capercaillies suggests that they play a role in the transmission and spread of resistant strains into the environment. Free-living birds may be also a significant reservoir of enterotoxigenic Staphylococcus strains.
Keywords:
free-living birds
; capercaillies
; coagulase-positive Staphylococcus
; virulence genes
1. Introduction
The genus Staphylococcus includes several dozen species of immobile Gram-positive cocci that do or do not produce coagulase (CoPS, coagulase-positive staphylococci or CoNS, coagulase-negative staphylococci). Their reservoirs are people and animals, as staphylococci can only multiply in higher organisms. These microorganisms are a component of the natural microbiota of the skin and mucous membranes of animals and humans, but they can also cause endogenous or exogenous infections. Coagulase-positive staphylococci are a relatively small but fairly diverse group. Species causing infections in animals include Staphylococcus aureus, Staphylococcus intermedius, Staphylococcus delphini, Staphylococcus schleiferi subsp. coagulans, Staphylococcus pseudintermedius, Staphylococcus lutrae, Staphylococcus agnetis and coagulase-variable Staphylococcus hyicus [1]. The species S. aureus is one of the main etiological agents of nosocomial infections and is a very important economic pathogen of animals. Bacteria from the Staphylococcus intermedius group (SIG), which include S. intermedius, S. pseudintermedius and S. delphini, colonize the skin and mucous membranes of various animal species, inhabiting the mouth, nose, perineum, and groin (in dogs, horses and mink) [2]. Two new SIG species, Staphylococcus cornubiensis and Staphylococcus ursi, have also recently been described and were isolated from human skin in Cornwall, UK, and from healthy black bears (Ursus americanus) in Tennessee, USA [3,4]. The remaining species from the SIG group were also isolated from humans with endocarditis, pneumonia, brain abscess, meningitis, and even food poisoning [5,6]. Among CoPS species, those most frequently isolated from birds, especially pigeons (Columba livia), are S. aureus and species of the SIG group [7]. The numerous virulence factors they produce play an important role in the pathogenesis of infections. The intraspecific diversity of CoPS strains also translates into the variety of features presented by individual strains. Coagulase, involved in the plasma clotting process, is present in all CoPS [8]. Protein A is an important pathogenicity factor and an element of the cell wall of S. aureus, but it can also be produced by species of the SIG group [9]. Many CoPS are equipped with numerous enterotoxins and cytolysins (α-, β-, γ-, and δ-haemolysins and leukotoxins) [10]. Staphylococci have also been shown to produce Panton-Valentine Leukocidin (PVL), which is highly species-specific for S. aureus. Other leukocidins produced by staphylococci include LukAB/HG, Luk-ED, and LukM. S. aureus can also produce a number of bacterial proteins with affinity for host proteins (including fibrinogen, collagen, laminin, transferrin, and lactoferrin), which is of great importance in the initial stage of infection [10]. Like other bacteria, staphylococci may also harbour genes for many other virulence factors. The increasing resistance of staphylococci to antibiotics in recent years is a well-known phenomenon. Studies have shown that as many as 95% of S. intermedius strains are resistant to β-lactam antibiotics. An additional threat is the increase in the number of MDR (multidrug-resistant) strains, including strains resistant to fluoroquinolones and mupirocin [11,12,13]. This applies especially to animal isolates from dogs, poultry and pigeons [14]. A serious problem is the wide spread of methicillin-resistant strains (MRS). Methicillin-resistant Staphylococcus aureus (MRSA) is a significant problem in veterinary medicine and animal husbandry. It is isolated from a wide range of animals, including dogs, cats, rabbits, horses, cattle, pigs, and poultry [15,16]. The phenomenon is mainly due to the presence of the mecA gene within the staphylococcal cassette chromosome mec (SCCmec). In 2011, in Great Britain, a methicillin-resistant strain of S. aureus was isolated from the liver and intestines of a dead free-living finch (Fringilla coelebs), in which a new divergent mecA gene homologue (mecALGA251) was identified [17]. MRS strains – both coagulase-positive and negative – have also been identified among other species. Methicillin-resistant Staphylococcus pseudintermedius (MRSP) strains have been isolated from cats, horses, birds, and humans [11,13,18]. However, the first case of infection with a methicillin-resistant strain of S. pseudintermedius in humans was reported in 2006 [18]. Within the species S. delphini, there are two phenotypically and genotypically distinct groups of strains (A and B) [19]. These bacteria have been found to be the natural microflora of ferrets, badgers, horses, camels, cows, pigeons, donkeys, and foxes. The pathogenicity of this species has been demonstrated in cows and mink [20,21,22]. There are only a few reports of its isolation from humans [23]. The virulence factors of this species include Panton-Valentine leukocidin, exfoliatin, and enterotoxins [20]. Environmental transmission of CoPS strains of Staphylococcus is one of the greatest socioeconomic threats in contemporary medicine.
Due to the lack of data in the available literature on coagulase-positive Staphylococcus species in wild birds, especially staphylococci from the SIG group, the aim of the study was to analyse the occurrence of individual species and to assess the pathogenic potential of coagulase-positive Staphylococcus strains isolated from dead capercaillies kept in aviaries as well as free-living birds in south-eastern Poland.
2. Materials and Methods
2.1. Birds
Samples were collected from dead free-living birds from the Podkarpackie province and from capercaillies living in an adaptive aviary in the Wisła Forest District near Żywiec and in the Capercaillie Breeding Centre in the Leżajsk Forest District near Krosno. From November 2019 to August 2023, samples were collected from 117 dead capercaillies (adult birds, chicks, and embryos) and from wild birds belonging to 12 orders and 26 species: Ciconiiformes (white stork, Ciconia ciconia, n = 57), Galliformes (common pheasant, Phasianus colchicus, n = 50; black grouse, Lyrurus tetrix, n = 3), Strigiformes (tawny or brown owl, Strix aluco, n = 16; long-eared owl, Asio otus, n = 5; little owl, Athene noctua, n = 1), Falconiformes (common kestrel, Falco tinnunculus, n = 13), Anseriformes (mute swan, Cygnus olor, n = 6; mallard or wild duck, Anas platyrhynchos, n = 1), Passeriformes (song thrush, Turdus philomelos, n = 5; blackbird, Turdus merula, n = 10; meadow pipit, Anthus pratensis, n = 6; fieldfare, Turdus pilaris, n = 7; common grosbeak, Coccothraustes coccothraustes, n = 1; black redstart, Phoenicurus ochruros, n = 1; Savi’s warbler, Locustella luscinioides, n = 1; Eurasian blue tit, Cyanistes caeruleus, n = 1), Piciformes (great spotted woodpecker, Dendrocopos major, n = 5), Pelecaniformes (grey heron, Ardea cinerea, n = 3), Apodiformes (common swift, Apus apus, n = 4), Accipitriformes (common buzzard, Buteo buteo, n = 10; lesser spotted eagle, Clanga pomarina, n = 3; white-tailed eagle, Haliaeetus albicilla, n = 3; Eurasian sparrowhawk, Accipiter nisus, n = 2), Charadriiformes (white gull, Larus canus, n = 1), and Bucerotiformes (Eurasian hoopoe, Upupa epops, n = 1). The material for bacteriological tests consisted of swabs taken during autopsy from the internal organs of dead birds (heart, liver, and spleen), the tarsal joints, and mucous membranes (conjunctiva and palatine fissure), as well as from unhatched capercaillie embryos. Swabs were placed in transport media and transported to the laboratory under refrigerated conditions.
Consent to collect tissues for analysis from dead birds of protected species was obtained from the Regional Directorate for Environmental Protection in Rzeszów, and consent to store samples of tissues or organs of animals from protected species was obtained from the Regional Directorate for Environmental Protection in Lublin.
2.2. Identification of Staphylococcus strains
The swabs were inoculated onto blood agar medium (Blood LAB-AGAR; Biocorp, Warsaw, Poland) and Chapman’s selective medium (Mannitol Salt LAB-AGAR; Biocorp) and incubated in aerobic conditions at 37°C for 24–48 hours, depending on the bacterial growth rate. Single colonies were transferred to blood agar to isolate pure bacterial cultures, and preliminary bacteriological characterization of the isolated bacteria was performed, including Gram staining, microscopic examination of cell morphology and motility, and determination of the type of haemolysis. No quantitative colony measurement was performed. Isolated bacteria were stored at −85°C in 50% (v/v) glycerol in brain heart broth (BHI; Sigma-Aldrich, St. Louis, MO, USA) until further study.
The species of the isolates were confirmed by matrix-assisted laser desorption/ionization (MALDI)–time-of-flight mass spectrometry using the IVD MALDI Biotyper (Bruker Daltonik, Bremen, Germany), as described by Marek et al. (2016), [15]. The mass spectra of each isolate were processed with the MALDI Biotyper 3.0 software package (Bruker Daltonics, Hamburg, Germany). The results were shown as the top 10 identification matches along with confidence scores ranging from 0.000 to 3.000, according to the manufacturer’s criteria (www.bruker.com; accessed on: 19 September 2021).
2.3. Determination of bacterial sensitivity to antibiotics and chemotherapeutics based on minimum inhibitory concentration (MIC)
Selected strains of Staphylococcus spp. were analysed for sensitivity to nine antimicrobial agents: benzylpenicillin, amoxicillin, tetracycline, gentamicin, chloramphenicol, erythromycin, enrofloxacin (Roth, Zielona Góra, Poland), trimethoprim and sulfamethoxazole (Merck KGaA, Darmstadt, Germany). Broth serial microdilution panels of the antibiotics were performed in sterile 96-well spherical-bottom polystyrene titration plates (FL-MEDICAL, Italy). Bacterial suspensions were prepared from morphologically similar colonies grown overnight on nonselective blood agar medium (Blood LAB-AGAR; Biocorp, Warsaw, Poland). An inoculum of 5 × 105 CFU (colony-forming units)/mL suspended in Mueller-Hinton broth (Oxoid Ltd., Basingstoke, UK) was transferred to wells containing serial dilutions of antibiotics (50 μl of bacterial inoculum + 50 μl liquid medium with antibiotic). The plates were incubated at 35±1ºC for 18±2 hours. The MIC breakpoint was defined as the lowest concentration of the substance at which no growth of the bacterial strains could be seen. MIC values were interpreted as susceptible, intermediate or resistant using the current Clinical and Laboratory Standards Institute (CLSI) breakpoint criteria [24]. The specific breakpoints for S. aureus, S. pseudintermedius and S. delphini were as follows: for benzylpenicillin and amoxicillin, susceptibility at ≤ 0.12 and resistance at ≥ 0.250 μg/ml; for tetracycline, susceptibility at ≤ 4 μg/ml, intermediate resistance at 8 μg/ml, and resistance at ≥16 μg/ml; for gentamicin, susceptibility at ≤ 4 μg/ml, intermediate resistance at 8 μg/ml, and resistance at ≥ 16 μg/ml; for chloramphenicol, susceptibility at ≤ 8 μg/ml, intermediate resistance at 16 μg/ml, and resistance at ≥ 32 μg/ml; for erythromycin, susceptibility at ≤ 0.5 μg/ml, intermediate resistance at 1–4 μg/ml, and resistance at ≥ 8 μg/ml; for enrofloxacin, susceptibility at ≤ 0.5 μg/ml, intermediate resistance at 1–2 μg/ml, and resistance at ≥ 4 μg/ml; for trimethoprim, susceptibility at ≤ 2 μg/ml and resistance at ≥4 μg /ml; and for sulfamethoxazole, susceptibility at ≤ 38 μg/ml and resistance at ≥76 μg /ml. Isolates were tested in duplicate; each result was analysed independently. Quality control was ensured using S. aureus ATCC 29213, Enterococcus faecalis ATCC 29212, and S. pneumoniae ATCC 49619; all quality control results were within published CLSI (2020) MIC limits [24].
2.3.1. Phenotypic determination of S. aureus sensitivity to cefoxitin
Phenotypic identification of the methicillin resistance of S. aureus isolates was achieved using the cefoxitin disc diffusion test with a 30 μg disc (Oxoid), according to the Clinical and Laboratory Standards Institute [24].
2.4. Molecular analysis
2.4.1. Genomic DNA Extraction
Single bacterial colonies were inoculated into tryptic soy broth (TSB) medium (CM0129, Oxoid, Ely, UK) and incubated at 37°C for 12 hours. Cells from the overnight culture in TSB medium were harvested by centrifugation at 8000 rpm at 10°C for 15 minutes, and the supernatant was then discarded. The pellet was then washed twice with normal saline and centrifuged at 8000 rpm at 10°C for 15 minutes. The GeneMatrix Bacterial & Yeast Genomic DNA Purification Kit protocol (EURx, Gdańsk, Poland) was used to isolate bacterial DNA. The bacterial DNA was stained with ethidium bromide, agarose gel electrophoresis was performed, and finally the fluorescence of the preparations was compared with the fluorescence of the preparation of known concentration.
2.4.2. PCR for identification of Staphylococcus isolates and genotypic analysis of virulence
The identity of S. aureus isolates was confirmed using a species-specific primer encoding the thermonuclease (nuc) gene as one of the key characteristics [25]. Bacterial strains were screened for the presence of virulence-associated genes by PCR amplification with primers at a concentration of 0.04 µmol (Table 1). All isolates were tested for the presence of two resistance genes mecA and mecC; staphylococcal enterotoxin genes A to L (sea, seb, sec, sed, see, seg, sei, sej, seh, and sel); toxic shock syndrome toxin 1 (tst); exfoliative toxins A and B (eta and etb); Panton-Valentine leukocidins (pvl); two unrelated leukotoxin components lukS and lukF; and four haemolysins (hla, hlb, hlg and hld). The presence of the staphylokinase (sak) gene was analysed as well. PCR reactions were performed with a ready-to-use reaction solution (Color OptiTaq PCR Master Mix, EURx, Gdańsk, Poland) containing OptiTaq DNA polymerase, optimized reaction buffer, MgCl2, dNTP, two dyes to facilitate tracking of PCR products on the gel, and 100 ng of DNA as template. PCR conditions are detailed in Table 1. For quality control, S. aureus RF122, S. aureus ATCC43300, S. aureus ATCC25923, S. aureus ATCC13566, S. aureus NCTC13300, FRI913, FRI151m, FRI1169, CCM7056, FRI572, FRI445; S. haemolyticus ATCC 29970, S. epidermidis ATCC 12228, and S. epidermidis ATCC 12228 were used in PCR reactions. PCR products were analysed by gel electrophoresis on a 2% agarose gel and visualized using a transilluminator.
2.4.3. Pulsed-field gel electrophoresis (PFGE)
From each isolate, 250 µl of bacterial suspension from a 16-hour culture in brain heart infusion (BHI) broth (Thermo Fisher Scientific) was centrifuged for 10 minutes, 5200 x g. The cell pellet was suspended in 1 ml of TEN buffer (0.1M Tris, 0.15M NaCl, 0.1M EDTA, pH 8) and centrifuged again (5200 x g, 10 minutes). Then the cells were suspended in 0.1 ml of EC lysis buffer (6mM tris, 1M NaCl, 100mM EDTA, 0.5% Brij 58, 0.2 sodium deoxycholate, 0.5 N-lauryl sarcosine sodium, pH 8), and 5 µl of lysostaphin (1 mg/ml) (Sigma, L7386). The cell suspension was mixed with 0.1 ml of 2% LMP agarose (Sigma, A4018). The solution was placed in a cassette to form blocks, left at room temperature for 30 minutes, and then placed in 1 ml of EC buffer at 37°C for 3 hours. Then the blocks were placed in 1 ml of buffer E (9.5 M EDTA, 1% N-lauryl sarcosine sodium) with 1 mg of protease (Sigma, P6911) and incubated at 37°C for 24 hours. The blocks were washed 5 times in 1 ml of TE buffer (10mM Tris, 1mM EDTA, pH 8). Then the DNA blocks were digested in 250 µl of reaction buffer containing 30 U of SmaI restriction enzyme (Thermo Fisher Scientific) for 4 hours at 30°C. The blocks were placed in a 1% agarose gel (Sigma, A2929). Electrophoresis was performed in 0.5 x TBE buffer for 22 hours at 14°C at 6 V/cm, with a pulsing time ranging from 5 to 40 s. The fragments were separated by electrophoresis in a 1% (w/v) agarose gel (Sigma-Aldrich, Poland) using the CHEF Mapper System (BIO-RAD, Poland). Macrorestriction patterns were examined by cluster analysis using NTSYSpc ver. Software 2.02 (Exeter Software Ltd, USA). The similarity of distances between pulsotypes (PFGE patterns) was calculated using the Dice coefficient, and dendrograms were calculated based on the unweighted pair group method with arithmetic mean (UPGMA). According to the criteria proposed by Tenover et al., isolates whose PFGE pattern differed by more than six restriction fragments (bands) were genetically unrelated and were assigned to different pulsotypes, designated with capital letters. Isolates were considered related if their pulsotype differed by no more than six restriction bands [33].
3. Results
3.1. Staphylococcus strains
From samples taken from capercaillies and free-living birds, 334 strains of bacteria belonging to the genus Staphylococcus were identified. The Staphylococcus strains belonged to 24 species. Coagulase-positive strains accounted for 5.7%, while the remaining 94.3% were coagulase-negative species. The percentages of strains belonging to individual species were as follows: S. sciuri 45.2%, S. lentus 8%, S. xylosus 7.8%, S. equorum 6.6%, S. aureus 3.9%, S. kloosii 3.3%, S. saprophyticus 3%, S. chromogenes 2.7%, S. vitulinus 2.7% S. epidermidis 2.7%, S. succinus 2.4%, S. cohnii 2.1%, S. haemolyticus 2.1%, S. gallinarum 1.8%, S. pseudintermedius 1.5%, S. warneri 1.2%, S. simulans 0.6%, S. felis 0.6%, S. delphini 0.3%, S. condimenti 0.3%, S. nepalensis 0.3%, S. arlettae 0.3%, S. pasteuri 0.3% and S. stepanovicii 0.3%. Coagulase-positive Staphylococcus strains belonging to the species S. aureus, S. pseudintermedius and S. delphini were further examined. Log(score) values for these isolates obtained in MALDI-TOF mass spectrometry were higher than 2.000, which indicates a high probability of correct identification to the species level (Table 2).
3.2. Phenotypic susceptibility of isolated bacteria to selected antimicrobial agents
Analysis of antimicrobial resistance based on MIC values showed that nine of 19 coagulase-positive staphylococcal strains were fully susceptible to the antimicrobials tested. Three strains (SP3, SA9, and SA11), were resistant to three or more groups of antibiotics, while two strains (SP1 and SP4), were resistant to two groups of antibiotics. The highest number of resistant strains was observed for beta-lactam antibiotics (n = 7). Table 2 shows the lowest concentration of the antibacterial agent that completely inhibited visible growth. None of the results of the tests of quality control strains were outside the ranges defined by CLSI [24].
- Resistance of S. aureus strains to oxacillin
A surrogate test for resistance of S. aureus strains to oxacillin using the cefoxitin (30 µg) disc diffusion method showed that the growth inhibition zone for all 13 strains was equal to or greater than 22 mm, which means that all strains tested were susceptible.
3.3. Molecular analysis
- Molecular identification of Staphylococcus isolates and genotypic analysis of virulence
The presence of the thermonuclease (nuc) gene was confirmed in all 13 S. aureus isolates. The presence of the mecA and mecC genes was not detected in any of the 19 coagulase-positive Staphylococcus strains tested. Among the virulence genes analysed, the α-toxin encoding gene (hla) was found in 11/19 (58%) of the isolates and was the most frequently detected virulence gene. The presence of the hlb and hld genes was demonstrated in 9/19 isolates. Only one S. aureus strain had the hlg gene. The results of multiplex PCR for 10 enterotoxins (A–L) showed that the genome of one of the S. pseudintermedius strains contained the gene responsible for the production of enterotoxin C, and one S. aureus strain had the gene responsible for the production of enterotoxin B. Two S. aureus strains had the gene responsible for the production of enterotoxin G, and one S. aureus strain showed the presence of the two genes for enterotoxins E and J. The presence of the see gene was demonstrated in the S. delphini isolate. None of the Staphylococcus strains had the genes responsible for the production of enterotoxins A, D, H, I or L (Table 2). However, the presence of the sak gene was confirmed in one strain of S. aureus.
- Pulsed field gel electrophoresis (PFGE)
Staphylococcus isolates were digested by the SmaI enzyme. One S. aureus isolate (SA11) was not digested by this enzyme. The analysis of the phylogenetic relationship between the 13 S. aureus strains, following digestion with the SmaI endonuclease, enabled the distinction of 11 macrorestriction profiles (A–K). Two isolates (SA2 and SA7) showed the same SmaI-PFGE patterns and belonged to the same pulsotype, designated E (Figure 1(A)). The remaining S. aureus pulsotypes obtained after enzyme digestion differed by more than seven bands, which means that they were not related according to Tenover’s criteria (Figure 2(a)) [33]. The two most similar S. aureus isolates (SA6 and SA8), designated in the dendrogram as pulsotypes B and C, differed in 12 bands, indicating that they were not related (Fig 1(A)). Analysis of the phylogenetic relatedness of S. pseudintermedius strains revealed five macrorestriction profiles (A–E), indicating that all strains differed significantly and were not related (Figure 1(B), Figure 2(b)).
4. Discussion
Among the isolated Staphylococcus strains, only three coagulase-positive species were identified in our study, constituting 5.7% of all isolates obtained from dead capercaillies and free-living birds. The most numerous species was S. aureus. Reports on the occurrence of coagulase-positive staphylococci in wild birds from the last 10 years were analysed. Ruiz-Ripa et al. (2019) identified 26 (8.3%) coagulase-positive staphylococci among 324 tracheal samples collected from healthy wild birds with occasional or obligatory scavenging habits in Spain (a cinereous vulture, a red kite, and magpies). Among the isolated strains, the authors identified two CoPS species: S. aureus (n = 15 isolates) and S. delphini (n = 12 isolates) [21]. In another study, analysing samples taken from the trachea of 92 storks located in Spain, S. aureus was isolated in 23.8% of the birds. Importantly, the occurrence of S. aureus was higher in birds from landfills (55.8%) than in those from natural habitats (16.3%) [34]. Examination of samples collected from birds of prey in Portugal showed that 37.5% of them were carriers of staphylococci, of which six species were identified. Only one coagulase-positive species (S. aureus) was isolated from the common buzzard (Buteo buteo). The remaining strains belonged to coagulase-negative species [35]. Little is known about the prevalence of SIG species in free-living birds, particularly S. pseudintermedius, especially given that it is considered to be the dominant staphylococcal etiological agent of pyoderma, ear infections, and urinary tract infections in dogs [36,37]. The new coagulase-positive species S. pseudintermedius was introduced relatively recently (in 2005), based on molecular analyses of zoonotic strains phenotypically and genotypically similar to S. intermedius [37]. Literature data indicate that it occurs mainly in Canidae as an element of the physiological microbiota of the skin and mucous membranes, but it has also been isolated from cats, parrots and humans [36,37]. Most of the SIG group of staphylococci isolated from birds, especially from pigeons, have been shown to belong to the species S. intermedius and S. delphini [38]. It is very difficult to differentiate between S. intermedius, S. delphini, and S. pseudintermedius phenotypically. For a long time, S. pseudintermedius was often incorrectly identified as S. aureus or S. intermedius in many laboratories, especially since biochemical features specific to the species S. pseudintermedius have not been determined. In our study, six strains of S. aureus were isolated from capercaillies in an adaptive aviary, which constituted 5.1% of all examined capercaillies, and from seven free-living birds, which constituted 3.2% of all those examined. S. pseudintermedius was isolated from 2.6% of capercaillies and less than 1% of free-living birds, while S. delphini was isolated from one capercaillie.
Reliable identification of coagulase-positive staphylococci isolated from animals and humans has become particularly important since the occurrence of highly pathogenic strains has been confirmed in this group of microorganisms. Moreover, MIC breakpoints for some antibiotics, e.g. oxacillin, used to determine the resistance of staphylococci to methicillin, vary depending on the Staphylococcus species [24]. Rapid bacterial identification systems based on mass spectrometry, such as MALDI TOF (matrix assisted laser desorption/ionization time of flight), may be useful for identifying species belonging to the SIG group if the databases used include these bacteria. The sensitivity of the MALDI TOF method for identifying S. pseudintermedius has been estimated at 78%, and its specificity at 97% [7]. However, in recently published research by Sawhney et al. (2023), a comparative analysis of the effectiveness of whole genome sequencing (WGS) and MALDI-TOF MS in identifying staphylococci from the SIG group among 493 strains of S. pseudintermedius and seven strains of S. delphini showed that MALDI-TOF provides 99.8% and 100% precision in identifying an isolate as S. pseudintermedius and S. delphini, respectively [39]. In our study, log(score) values for S. pseudintermedius and S. delphini isolates obtained in MALDI-TOF mass spectrometry were higher than 2.000, which indicates a high probability of correct identification to the species level (Table 2).
Restriction analysis of chromosomal DNA in combination with pulse electrophoresis (PFGE) is the most frequently used method for studying the epidemiology of locally occurring Staphylococcus strains. The most commonly used restriction enzymes are SmaI and CspI. Strains that have the same restriction patterns are classified as the same macrorestriction type or PFGE type. Strains with differences in one or two bands corresponding to changes in the genome (insertions, deletions, or point mutations) are assigned to subtypes, and isolates differing in three or more bands are assigned to different PFGE types [33]. In this way, it is possible to establish genetic relationships between isolates in epidemiological studies [40]. In our study, 12 of the S. aureus isolates were typeable by SmaI enzyme digestion. One S. aureus (SA11) strain was not digested by this enzyme. It is possible that the activity of SmaI is blocked due to methylation of the restriction site [41]. Interestingly, analysis of the genetic material of 13 S. aureus strains using a rare-cutting restriction enzyme and separation of the resulting DNA fragments revealed that two isolates (SA2 and SA7) had the same SmaI-PFGE patterns and belonged to the same macrorestriction type. Isolates were obtained from birds of prey from the same area: from a wound in a common kestrel (Falco tinnunculus) and from the mucous membrane of the palatal fissure of a buzzard (Buteo buteo). It was also determined that the macrorestriction patterns of the remaining S. aureus and S. pseudintermedius isolates differed by more than seven bands, which means that they were not related according to Tenover’s criteria (Figure 1, 2(a), 2(b)), [33].
Staphylococcus aureus is a species with significant pathogenic potential, often isolated from clinical material from humans and animals. However, features previously considered typical of this species may also be present in other coagulase-positive species, which may be of importance during infections. The presence of S. aureus strains in the tissues and organs of wild birds is quite common, [17,34,35,42,43,44,45,46,47,48,49,50]. Additionally, S. aureus produces several virulence factors that allow the bacteria to survive extreme conditions in the host and to damage biological membranes, causing cell death. S. aureus maintains precise control over the expression of virulence factors, which include haemolysins, leukocidins, proteases, exfoliative toxins, enterotoxins, and immunomodulatory factors [28, 29].
In a study by Silva et al. (2022) of throat and cloacal swabs collected from 114 individuals belonging to various species of nocturnal birds of prey (Strix aluco, Tyto alba, Athene noctua and Bubo bubo) in central and northern Portugal, 25 strains of CoPS were isolated. Most of them (23 strains) were identified by MALDI-TOF as S. aureus, while two strains belonged to the species S. pseudintermedius [50]. In our study, five strains of S. pseudintermedius were isolated from samples collected from the conjunctiva, joints, and liver of three capercaillies, a meadow pipit and a white stork. One S. delphini isolate was obtained from the yolk sac of a dead capercaillie chick (Table 2). S. delphini has been isolated from many animal species in which it caused infections [21, 22]. This species can be divided into two genetically distinct clusters, S. delphini group A and group B, for which animal host specificity may exist. Some research suggests that the family Mustelidae may be the natural host of S. delphini group A, while S. delphini group B is more frequently detected in wild and domestic pigeons, although this is still under investigation [36, 38]. In our study, the group membership of the isolated S. delphini strain was not determined. The S. delphini strain was fully sensitive to nine of the antimicrobials tested. PCR testing revealed the presence of two haemolysin genes, hlb and hld, in the genome of this isolate, as well as the see gene responsible for the production of enterotoxin E.
S. pseudintermedius virulence factors, like those produced by S. aureus, promote colonization and infection [19]. Reports indicate that S. pseudintermedius is an opportunistic pathogen that can cause infections of the skin (purulent dermatitis, otitis externa, wound infections, or abscesses), and other tissues and body cavities [19,39,51,52]. The ability to haemolyse blood cells by producing cytolysins is a feature observed in various species of staphylococci. These are important factors in the pathogenicity of these bacteria [10]. The pathogenic potential of S. pseudintermedius is determined by virulence factors including coagulase, protein A, protease, enterotoxins, TSST-1 (toxic shock syndrome toxin), exfoliative toxin SIET, and cytolysins, i.e. leukotoxin Luk-I and haemolysins [19, 39]. PCR analysis for virulence determinants of strains isolated from capercaillies and free-living birds showed that genes involved in avoidance and invasion (hla and hlb) were the most common in S. aureus isolates. The α-toxin encoding gene (hla) was found in 10/13 (77%) of the S. aureus isolates, and it was the most frequently detected virulence gene. The presence of the hlb and hld genes was found in 9/13 and 7/13 S. aureus strains, respectively. In the case of other Staphylococcus strains, the presence of hla and hld genes was confirmed in one strain of S. pseudintermedius isolated from the joint of a capercaillie, and the presence of hlb and hld genes in an isolate of S. delphini from the yolk sac of a capercaillie. Silva et al. (2022), analysing strains of S. aureus and S. pseudintermedius isolated from nocturnal birds of prey in Portugal, also detected genes hla and hlb encoding virulence factors. All isolates carried at least one virulence gene, with the hla gene present in all isolates and the hlb gene in 15 isolates. Four S. aureus isolates contained the sak gene [50]. As some authors point out, in the case of S. pseudintermedius strains, β-haemolysin is produced constitutively, while δ- and α-toxin are rarely produced [20, 51]. The results of research by Afzal et al. 2022 on staphylococci isolated from humans indicate that the hlg gene is more common in infectious strains of S. aureus than in non-infectious strains [53]. Alpha-haemolysin is by far the most commonly studied of the S. aureus cytotoxins, because it is produced by many strains and is toxic to a broad spectrum of mammalian cells. In our study, the presence of the hlg gene was detected in only one of the S. aureus strains, which was isolated from the palatal fissure of a capercaillie.
Carriers of virulence factors may be genes located on mobile genetic elements called pathogenicity islands, e.g. TSST and some enterotoxins, or lysogenic bacteriophages, e.g. Panton-Valentine leukocidin (PVL), while others are integrated into the bacterial chromosome, such as staphylokinase (SAK) [54,55]. The most important virulence factors for diagnostics include enzymes: staphylokinase (plasminogen activator) and haemolysins, primarily β-toxin, also called sphingomyelinase, which destroys the cell membrane of macrophages, leukocytes and erythrocytes [31, 53]. Staphylokinase, encoded by the sak gene, can be produced by lysogenic strains of both S. aureus and non-aureus staphylococci [54]. PCR testing for the staphylokinase (sak) gene in CoPS strains from capercaillies and free-living birds showed its presence in one strain of S. aureus isolated from the song thrush in the palatal fissure. The role of this protein as a virulence factor of staphylococci is assumed to stem from its interaction with plasminogen and defensins. Binding of staphylokinase to plasminogen may affect bacterial invasion into the host tissues. The activity of virulence factors encoded by bacteriophages, such as staphylokinase, is assumed to be specific to human cells, which indicates a close relationship between the host and the pathogen [56]. Some authors suggest that Staphylococcus strains of animal origin carry sak-containing phages less frequently than human isolates [57].
PVL is a two-component, β-barrel pore-forming toxin [58]. PVL and γ-haemolysin are toxins that act on cell membranes through the synergy of two pore-forming proteins. A small percentage of S. aureus isolates produce PVL [55]. PVL is also the most leukocytolytic toxin and causes skin necrosis [59]. To date, PVL-positive S. aureus strains have been documented in companion animals but have rarely been found in other animal species [16]. The presence of genes responsible for the production of toxic shock syndrome toxin-1 (TSST-1), Panton-Valentine leukocidin (PVL), or exfoliative toxins was not confirmed in any CoPS strains isolated from capercaillies and free-living birds.
Staphylococcal enterotoxins (SE) constitute a family of structurally related toxins. Thus far, 33 enterotoxins (SEs) and SE-like toxins (SEls) have been described. The most frequently identified toxins (>75%) worldwide are SEA to SEE enterotoxins, referred to as ‘classic SE’ [60]. They are mainly produced by coagulase-positive staphylococci, but the genes responsible for the production of enterotoxins have also been identified in coagulase-negative strains isolated from birds (poultry) [61]. Genes encoding enterotoxins (se) are most often located on mobile genetic elements, such as plasmids, prophages, or pathogenicity islands [28]. Screening of CoPS strains isolated from capercaillies and free-living birds for 10 enterotoxin genes (classic A–E and new G–L) confirmed the presence of enterotoxin genes in four of 13 S. aureus isolates. However, PFGE analysis showed that two S. aureus strains (SA2 and SA7) carrying the seg gene were closely related, but isolated from two different bird species. Moreover, one strain of S. aureus, isolated from the spleen of a common buzzard, had the gene responsible for the production of enterotoxin B, and the presence of two genes, see and sej, was confirmed in one strain isolated from a song thrush. In the genome of one strain of S. pseudintermedius, isolated from a capercaillie, the gene responsible for the production of classical enterotoxin C was found. The S. delphini strain isolated from a capercaillie also had the gene for a classical enterotoxin (see). Staphylococcal strains isolated from wild animals are rarely characterized in terms of potential enterotoxicity. Moreover, unlike S. aureus, no enterotoxin screening has been performed at the genome level for SIG species. The few studies of veterinary isolates of S. intermedius from horses, dogs, and cats and of human strains showed that just over 10% of the strains were positive for the enterotoxin genes [60].
An additional threat is the increase in the number of drug-resistant strains recorded in recent years, including methicillin-resistant and multi-drug-resistant (MDR) strains of Staphylococcus [52], especially those resistant to fluoroquinolones, isolated from companion and farm animals [62]. Resistance genes are most often located on mobile genetic elements (MGEs), such as the staphylococcal cassette chromosome (SCCmec), plasmids, and transposons [58]. Methicillin resistance in various staphylococcal species isolated from animals has proven to be a serious therapeutic challenge over the last decade. S. pseudintermedius strains carrying the mecA gene are referred to as MRSP (methicillin-resistant S. pseudintermedius) and, like MRSA (methicillin-resistant Staphylococcus aureus), are usually multi-drug resistant, because SCCmec often contains resistance genes to other antibiotics and chemotherapeutics as well. Cases of MRSP infections have been reported, especially in dogs, cats, birds, horses and humans [11]. However, studies of the role of wildlife as a reservoir of MRSA and other methicillin-resistant CoPS are limited, in comparison with humans and domestic animals. In a study by Silva et al. (2022), on strains isolated from nocturnal birds of prey, only one of 23 S. aureus isolates was resistant to cefoxitin and contained the mecC gene. Two isolates showed phenotypic resistance to macrolides and lincosamides. However, two S. pseudintermedius isolates were sensitive to all antibiotics tested, but the presence of the mecA gene was identified in one of them [50]. In our study, the analysis of the presence of the mecA gene and a novel mecA homologue, mecC, did not show the presence of these genes in any of the 19 isolates from capercaillies and free-living birds in south-eastern Poland. The study of phenotypic resistance to antimicrobials using the broth microdilution method showed that 36.8% of the coagulase-positive strains were resistant to beta-lactam antibiotics (penicillin and amoxicillin), 26.3% were resistant to chloramphenicol, and 15.8% to tetracycline, erythromycin, and trimethoprim. However, three of the 19 strains were multidrug-resistant (MDR). MDR strains were isolated from capercaillies kept in adaptive aviaries. This phenomenon should probably be linked to the fact that capercaillies kept in aviaries have closer contact with the human environment. None of the CoPS strains showed phenotypic resistance to gentamicin, enrofloxacin, or sulfamethoxazole. The pathogenic potential of the bacterial strains isolated from wild birds obviously varies depending on their environment (urban or farmland), which is undoubtedly linked to the presence of people, farm animals, hospitals and production plants. In the latest study of samples from the breeding sites of wild birds living in parks and near hospital waste dumps in Islamabad (Pakistan), 174 isolates were biochemically identified as S. aureus. Antimicrobial sensitivity testing showed that nearly 29% of them were multidrug-resistant, and half of them were designated as MRSA. The majority of MDR isolates came from faecal samples, collected from the vicinity of hospitals [63].The available literature contains reports of the occurrence of MRSA strains isolated from bird species such as white stork, cinereous vulture, Eurasian griffon vulture, and magpie in Spain; lesser yellowlegs in the United States; and common chaffinch in Scotland, and the mecA homologue (mecA LGA251) has been found in strains isolated from Canada goose faecal samples [17, 21, 34, 45, 47, 49 ]. According to studies on S. aureus, the population structure may vary regionally [15, 34 43]. Analysis of coagulase-positive strains from wild birds in Spain showed that among the collection of 15 S. aureus isolates, 13 were MRSA. Moreover, the MRSA isolate was mecA-positive, and 12 MRSA isolates harboured the mecC gene [21].
Due to the progress of civilization, contact between wildlife and the human environment is becoming more frequent, increasing the possibility of exchange of microbial virulence factors in various ecosystems. There are many reports of the pathogenic potential of staphylococci isolated from humans, companion animals, and farm animals. However, populations of coagulase-positive staphylococci found in wild animals, especially birds, in various regions of the world are still poorly characterized.
5. Conclusions
Coagulase-positive Staphylococcus strains constituted only a small percentage (5.7%) of staphylococcal species isolated from dead capercaillies and from various species of free-living birds in south-eastern Poland.
Although staphylococci from the SIG group are rarely isolated from free-living birds, our results indicate that S. pseudintermedius and S. delphini may be present on the mucous membranes and internal organs of capercaillies and other species of wild birds.
The occurrence of multidrug-resistant CoPS in aviary capercaillies suggests their role in the transmission and spread of resistant strains into the environment. The largest number of resistant strains isolated from capercaillies and free-living birds was found for beta-lactam antibiotics. The study results clearly indicate the need for monitoring of resistant bacteria in wild birds and other wildlife.
The analysis of virulence determinants of strains isolated from capercaillies and free-living birds showed that genes involved in avoidance and invasion (hla and hlb) were the most common in the isolates. Enterotoxin production by CoPS isolates from free-living birds has not previously been assessed. We identified S. aureus and SIG group isolates carrying genes for both classical and novel enterotoxins. Our results suggest that free-living birds may be a significant reservoir of enterotoxigenic Staphylococcus strains.
Author Contributions
Conceptualization, M.S. and A.M.; methodology, M.S. and A.M.; software, Ł.S.J; validation, Ł.S.J. and T.H.; formal analysis, all authors.; investigation, all authors; M.S. and A.M.; data curation, all authors; writing—original draft preparation, M.S. and A.M.; writing—review and editing, A.M. and M.S.; visualization, A.M. and M.S.; supervision, A.M. and M.S.; project administration, M.S;. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parte, A.C. LPSN – list of prokaryotic names with standing in nomenclature. Nucleic Acids Res 2014, 42, D613–D616. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Karasawa, T.; Piao, C.; Itoda, I.; Hidai, H.; Yamaura, H.; Totsuka, K.; Morikawa, T.; Takayama, M. Molecular confirmation of transmission route of Staphylococcus intermedius in mastoid cavity infection from dog saliva. J. Infect. Chemother. 2004, 10, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.K.; Lee, J.; Bendall, R.; Zhang, L.; Sunde, M.; Slettemeås, J.S.; Gaze, W.; Page, A.J.; Vos, M. Staphylococcus cornubiensis sp. nov., a member of the Staphylococcus intermedius Group (SIG). Int J Syst Evol Microbiol 2018, 68, 3404–3408. [Google Scholar] [CrossRef] [PubMed]
- Perreten, V.; Kania, S.A; Bemis, D. Staphylococcus ursi sp. nov., a new member of the ‘ Staphylococcus intermedius group’ isolated from healthy black bears. Int J Syst Evol Microbiol. 2020, 70, 4637–4645. [Google Scholar] [CrossRef]
- Gerstadt, K.; Daly, J.S.; Mitchell, M.; Wessolossky, M.; Cheeseman, S.H. Methicillin-resistant Staphylococcus intermedius pneumonia following coronary artery bypass grafting. Clin. Infect. Dis. 1999, 29, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Mahoudeau, I.; Delabranche, X.; Prevost, G.; Monteil, H.; Piemont, Y. Frequency of isolation of Staphylococcus intermedius from humans. J. Clin. Microbiol. 1997, 35, 2153–2155. [Google Scholar] [CrossRef]
- Decristophoris, P.; Fasola, A.; Benagli, C.; Tonolla, M.; Petrini, O. Identification of Staphylococcus intermedius Group by MALDI--TOF MS. Syst. Appl. Microbiol. 2011, 34, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Hale, J.H.; Smith, M.M. The role of coagulase in staphylococcal infections. Brit J Exp Pathol 1947, 28, 57–67. [Google Scholar]
- Fehrer, S.L.; Boyle, M.D.; Halliwell, R.E. Identification of protein A from Staphylococcus intermedius isolated from canine skin. Am J Vet Res. 1988, 49, 697–701. [Google Scholar]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus Hemolysins, bi-component Leukocidins, and Cytolytic Peptides: A Redundant Arsenal of Membrane-Damaging Virulence Factors? Front Cell Infect Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef]
- Ruscher, C.; Lübke-Becker, A.; Semmler, T.; Wleklinski, C.G.; Paasch, A.; Soba, A.; Stamm, I.; Kopp, P.; Wieler, L.H.; Walther, B. Widespread rapid emergence of a distinct methicillin- and Multidrug-resistant Staphylococcus pseudintermedius (MRSP) genetic lineage in Europe. Vet. Microbiol. 2010, 144, 340–346. [Google Scholar] [CrossRef]
- Ünal, N.; Çinar, O.D. Detection of staphylococcal enterotoxin, methicillin-resistant and Panton–Valentine leukocidin genes in coagulase-negative staphylococci isolated from cows and ewes with subclinical mastitis. Trop Anim Health Prod 2012, 44, 369–375. [Google Scholar] [CrossRef]
- Weese, J.S.; van Duijkeren, E. Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary medicine. Vet. Microbiol. 2010, 140, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Bean, D.C.; Wigmore, S.M. Carriage rate and antibiotic susceptibility of coagulase-positive staphylococci isolated from healthy dogs in Victoria, Australia. Australia. Aust Vet J. 2016, 94, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Stępień-Pyśniak, D.; Pyzik, E.; Adaszek, Ł.; Wilczyński, J.; Winiarczyk, S. Occurrence and characterization of Staphylococcus bacteria isolated from poultry in Western Poland. Berl Munch Tierarztl Wochenschr 2016, 129, 147–152. [Google Scholar] [CrossRef]
- Morgan, M. Methicillin-resistant Staphylococcus aureus and animals: zoonosis or humanosis? J Antimicrob Chemother. 2008, 62, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Robb, A.; Pennycott, T.; Duncan, G.; Foster, G. Staphylococcus aureus carrying divergent mecA homologue (mecA LGA251) isolated from a free-ranging wild bird. Vet. Microbiol. 2013, 162, 300–301. [Google Scholar] [CrossRef]
- Van Hoovels, L.; Vankeerberghen, A.; Boel, A.; Van Vaerenbergh, K.; De Beenhouwer, H. First case of Staphylococcus pseudintermedius infection in a human. J Clin Microbiol. 2006, 44, 4609–12. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, D.B. The Staphylococcus intermedius group of bacterial pathogens: species re-classification, pathogenesis and the emergence of methicillin resistance. Vet. Dermatol. 2009, 20, 490–495. [Google Scholar] [CrossRef]
- Gharsa, H.; Slama, K.B.; Gómez-Sanz, E.; Gómez, P.; Klibi, N.; Zarazaga, M.; Boudabous, A.; Torres, C. Characterisation of nasal Staphylococcus delphini and Staphylococcus pesudintermedius isolates from healthy donkeys in Tunisia. Equine Vet. J. 2014, 47, 463–466. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Gomez, P.; Alonso, C.A.; Camacho, M.C.; de la Puente, J.; Fernandez-Fernandez, R.; Ramiro, Y.; Quivedo, M.A.; Blanco, J.M.; Zarazaga, M.; Höfle, U.; Torres, C. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu(A) in Magpies and Cinereous Vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Gary, J.M.; Langohr, I.M.; Lim, A.; Bolin, S.; Bolin, C.; Moore, I.; Kiupel, M. Enteric colonization by Staphylococcus delphini in four ferret kits with diarrhoea. J. Comp. Pathol. 2014, 151, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Magleby, R.; Bemis; D.A.; Kim, D.; Carroll, K.C.; Castanheira, M.; Kania, S.A.; Jenkins, S.G.; Westblade, L.F. First reported human isolation of Staphylococcus delphini. Diagn Microbiol Infect Dis. 2019, 94, 274–276. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Othman, E.H.; Merza, S.N.; Jubrael, S.M. Nucleotide sequence analysis of methicillin-resistant Staphylococcus aureus in Kurtdistan Region-Iraq. J. Univ. Zakho. 2014, 2, 1–13. [Google Scholar]
- Murakami, K.; Minamide, W.; Wada, K.; Nakamura, E.; Teraoka, H.; Watanabe, S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. J Clin Microbiol 1991, 29, 2240–2244. [Google Scholar] [CrossRef] [PubMed]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecA(LGA251). Clin Microbiol Infect 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance. J Clin Microbiol 2000, 38, 1032–1035. [Google Scholar] [CrossRef] [PubMed]
- Cremonesi, P.; Perez, G.; Pisoni, G.; Moroni, P.; Morandi, S.; Luzzana, M.; Brasca, M.; Castiglioni, B. Detection of enterotoxigenic Staphylococcus aureus isolates in raw milk cheese. Lett Appl Microbiol 2007, 45, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Futagawa-Saito, K.; Sugiyama, T.; Karube, S.; Sakurai, N.; Ba-Thein, W.; Fukuyasu, T. Prevalence and characterization of leukotoxin-producing Staphylococcus intermedius in isolates from dogs and pigeons. J. Clin. Microbiol. 2004, 42, 5324–5326. [Google Scholar] [CrossRef]
- Sung, J. M.; Lloyd, D.H.; Lindsay, J.A. Staphylococcus aureus host specificity: Comparative genomics of human versus animal isolates by multi-strain microarray. Microbiology, 2008; 154, 1949–1959. [Google Scholar]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Gomez, P.; Lozano, C.; Camacho, M.C.; Lima-Barbero, J.F.; Hernandez, J.M.; Zarazaga, M.; Hofle, U.; Torres, C. Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 2016, 71, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.; Silva, N.; Igrejas, G.; Silva, F.; Sargo, R.; Alegria, N.; Benito, D.; Gomez, P.; Lozano, C.; Gomez-Sanz, E.; Torres, C.; Caniça, M.; Poeta, P. Antimicrobial resistance determinants in Staphylococcus spp. recovered from birds of prey in Portugal. Vet. Microbiol. 2014, 171, 436–440. [Google Scholar] [PubMed]
- Guardabassi, L.; Schmidt, K.R.; Petersen, T.S.; Espinosa-Gongora, C.; Moodley, A.; Agerso, Y.; Olsen, J.E. Mustelidae are natural hosts of Staphylococcus delphini group A. Vet Microbiol 2012, 159, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.; Vancanneyt, M.; Baele, M.; Vaneechoutte, M.; De Graef, E.; Snauwaert, C.; Cleenwerck, I.; Dawyndt, P.; Swings, J.; Decostere, A.; Haesebrouck, F. Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. Int. J. Syst. Evol. Microbiol. 2005, 55, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kikuchi, K.; Tanaka, Y.; Takahashi, N.; Kamata, S.; Hiramatsu, K. Reclassification of phenotypically identified Staphylococcus intermedius strains. J. Clin. Microbiol. 2007, 45, 2770–2778. [Google Scholar] [CrossRef] [PubMed]
- Sawhney, S.S.; Vargas, R.C.; Wallace, M.A.; Muenks, C.E.; Lubbers, B.V.; Stephanie, A.; Fritz, S.A.; Burnham, C.D.; Dantas, G. Diagnostic and commensal Staphylococcus pseudintermedius genomes reveal niche adaptation through parallel selection of defense mechanisms. Nature Communications 2023, 14, 7065. [Google Scholar] [CrossRef] [PubMed]
- Doškař, J.; Pantůček, R.; Růžičková, V.; Sedláček, I. Molecular diagnostics of Staphylococcus aureus. W: Detection of bacteria, viruses, parasites and fungi. Bioterrorism prevention, red. M.V. Magni, Springer, Dordrecht 2010, 139-184.
- Bens, C.P.M.; Voss, A.; Klaassen, C.H.W. Presence of a novel DNA methylation enzyme in methicillin-resistant Staphylococcus aureus isolates associated with pig farming leads to uninterprettable results in standard pulsed-field gel electrophoresis analysis. J Clin Microbiol 2006, 44, 1875–1876. [Google Scholar] [CrossRef] [PubMed]
- Awad-Alla, M.E.; Abdien, H.M.; Dessouki, A.A. Prevalence of bacteria and parasites in White Ibis in Egypt. Vet. Ital. 2010, 46, 277–286. [Google Scholar]
- Monecke, S.; Gavier-Widen, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Muller, E.; Reissig, A.; Ruppelt-Lorz, A.; Walter, B.; Coleman, D.C.; Ehricht, R. Diversity of Staphylococcus aureus Isolates in European Wildlife. PLoS ONE 2016, 11, e0168433. [Google Scholar] [CrossRef]
- Porrero, M.C.; Mentaberre, G.; Sanchez, S.; Fernandez-Llario, P.; Casas-Diaz, E.; Mateos, A.; Vidal, D.; Lavin, S.; Fernández-Garayzábal, J.-F.; Dominguez, L. Carriage of Staphylococcus aureus by free-living wild animals in Spain. Appl. Environ. Microbiol. 2014, 80, 4865–4870. [Google Scholar] [CrossRef] [PubMed]
- Porrero, M.C.; Mentaberre, G.; Sanchez, S.; Fernandez-Llario, P.; Gomez-Barrero, S.; Navarro-Gonzalez, N.; Serrano, E.; Casas-Diaz, E.; Marco, I.; Fernández-Garayzabal, J.F.; Mateos, A.; Vidal, D.; Lavín, S.; Domínguez, L. Methicillin resistant Staphylococcus aureus (MRSA) carriage in different free-living wild animal species in Spain. Vet. J. 2013, 198, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Silvanose, C.D.; Bailey, T.A.; Naldo, J.L.; Howlett, J.C. Bacterial flora of the conjunctiva and nasal cavity in normal and diseased captive bustards. Avian Dis. 2001, 45, 447–451. [Google Scholar] [CrossRef]
- Thapaliya, D.; Dalman, M.; Kadariya, J.; Little, K.; Mansell, V.; Taha, M.Y.; Grenier, D.; Smith, T.C. Characterization of Staphylococcus aureus in Goose Feces from State Parks in Northeast Ohio. Ecohealth 2017, 14, 303–309. [Google Scholar] [CrossRef]
- Vidal, A.; Baldoma, L.; Molina-Lopez, R.A.; Martin, M.; Darwich, L. Microbiological diagnosis and antimicrobial sensitivity profiles in diseased free-living raptors. Avian Pathol. 2017, 46, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, S.E.; Kauffman, L.K.; Smith, T.C. Methicillin-resistant Staphylococcus aureus in central Iowa wildlife. J. Wildl. Dis. 2012, 48, 1069–1073. [Google Scholar] [CrossRef]
- Silva, V.; Lopes, A.F.; Soeiro, V.; Caniça, M.; Manageiro, V.; Pereira, J.E.; Maltez, L.; Capelo, J.L.; Igrejas, G.; Poeta, P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics 2022, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Chrobak, D.; Kizerwetter-Świda, M.; Rzewuska, M.; Moodley, A.; Guardabassi, L.; Binek, M. Molecular characterization of Staphylococcus pseudintermedius strains isolated from clinical samples of animal origin. Folia Microbiol. 2011, 56, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Chae, M.; Yoon, J.; Kim, S.; Lee, S.; Yoo, J.; Park, H.M. Antibiotic resistance and molecular characterization of ophthalmic Staphylococcus pseudintermedius isolates from dogs. J. Vet. Sci. 2014, 15, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Vijay, A.K.; Stapleton, F.; Willcox, M. Virulence genes of Staphylococcus aureus associated with keratitis, conjunctivitis and contact lens-assocaied inflammation. Transl. Vis. Sci. Technol. 2022, 11, 5. [Google Scholar] [CrossRef]
- Novick, R. P. Pathogenicity factors and their regulation. In V. A. Fuschetti, R. P. Novick, J. J. Ferretti, D. A. Portnoy, & J. I. Rood (Eds.), Gram-positive pathogens, 2000, 392–407. Washington, DC: AMS Press.
- Prévost, G.; Cribier, B.; Couppié, P.; Petiau, P.; Supersac, G.; Finck-Barbançon, V.; Monteil, H.; & Piémont, Y. Panton-Valentine leucocidin and gamma-hemolysin from Staphylococcus aureus ATCC 49775 are encoded by distinct genetic loci and have different biological activities. Infection and Immunity 1995, 63, 4121–4129. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Wirtz, C.; Flückiger, U.; Wolz, C. Extensive phage dynamics in Staphylococcus aureus contributes to adaptation to the human host during infection. Mol Microbiol 2006, 61, 1673–1685. [Google Scholar] [CrossRef]
- Matthews, A.M.; Novick, R.P. Staphylococcal phages in Phages. Their role in bacterial pathogenesis and biotechnology. Waldor MK, Friedman DI, Adhya SL (eds). Washington, DC: American Society for Microbiology Press,2005,297–318.
- Humphreys, H.; Coleman, D.C. Contribution of whole-genome sequencing to understanding of the epidemiology and control of meticillin-resistant Staphylococcus aureus. J. Hosp. Infect. 2019, 102, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; Lian, J.; Ito, T.; Kanamori, M.; Matsumaru, H.; Maruyama, A.; Murakami, H.; Hosoyama, A.; Mizutani-Ui, Y.; Takahashi, N.K.; Sawano, T.; Hiramatsu, K. Whole genome sequencing of methicillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Keller, B.; von Eiff, C.; Brück, M.; Lubritz, G.; Etienne, J.; Peters, G. Enterotoxigenic potential of Staphylococcus intermedius. Appl Environ Microbiol. 2001, 67, 5551–7. [Google Scholar] [CrossRef] [PubMed]
- Pyzik, E.; Marek, A.; Stępień-Pyśniak, D.; Urban-Chmiel, R.; Jarosz, Ł.S.; Jagiełło-Podębska, I. Detection of Antibiotic Resistance and Classical Enterotoxin Genes in Coagulase -negative Staphylococci Isolated from Poultry in Poland. J Vet Res. 2019, 12;63, 183-190. [CrossRef]
- Kang, M.; Chae, M.; Yoon, J.; Lee, S.; Yoo, J.; Park, H.M. Resistance to fluoroquinolones and methicillin in ophthalmic isolates of Staphylococcus pseudintermedius from companion animals. Vet. Rec. 2014, 55, 678–682. [Google Scholar]
- Tareen, A.R.; Zahra, R. Community acquired methicillin resistant Staphylococci (CA-MRS) in fecal matter of wild birds – A ‘one health’ point of concern. J Infect Public Health. 2023, 16, 877–883. [Google Scholar] [CrossRef]
Figure 1.
Dendrogram showing the level of similarity of SmaI restriction PFGE patterns of the genomic DNA of S. aureus (A) and S. pseudintermedius (B) isolates.
Figure 1.
Dendrogram showing the level of similarity of SmaI restriction PFGE patterns of the genomic DNA of S. aureus (A) and S. pseudintermedius (B) isolates.
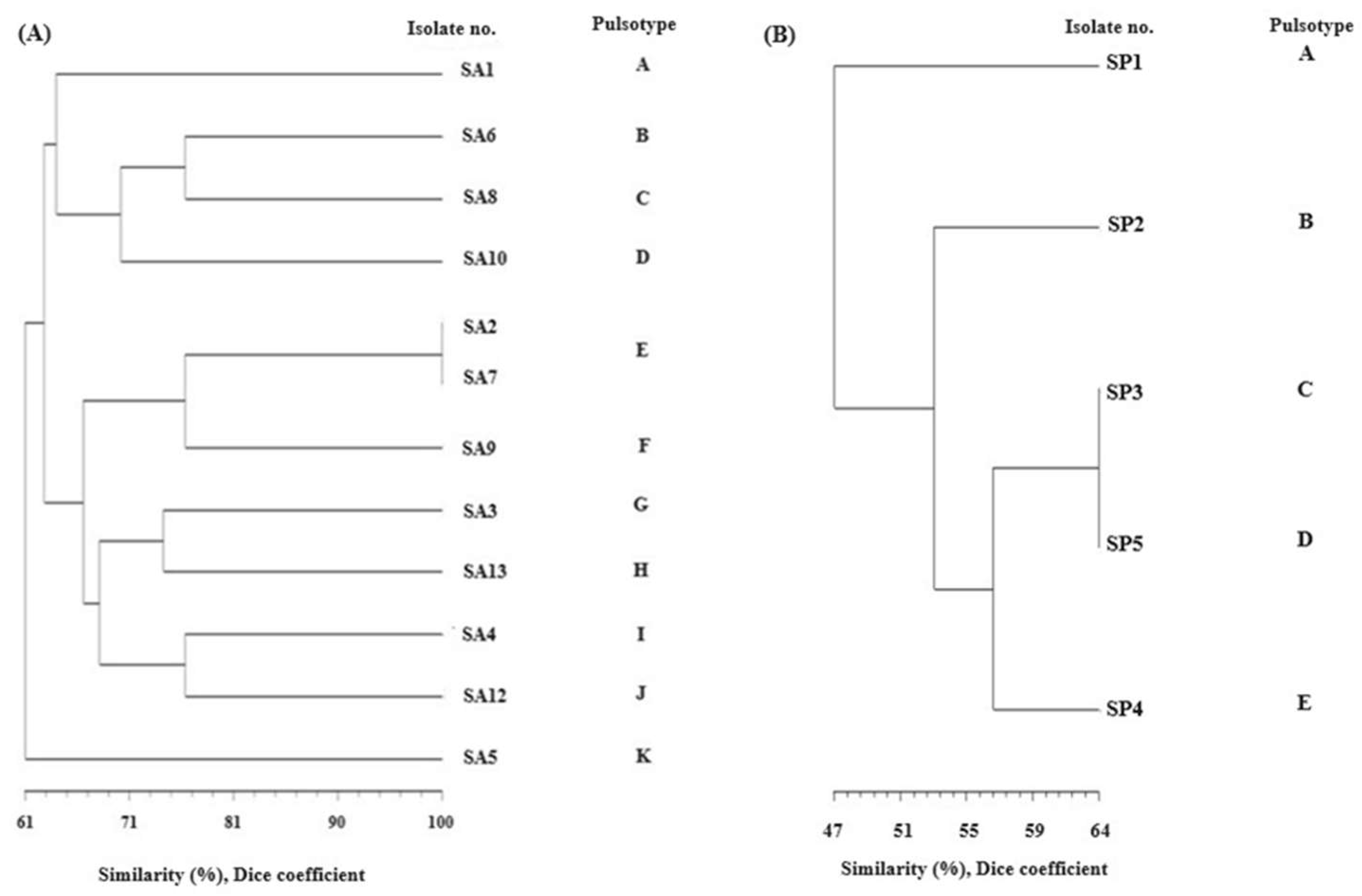
Figure 2.
PFGE patterns of S. aureus (a) and S. pseudointermedius (b) isolates digested with SmaI restriction enzyme.
Figure 2.
PFGE patterns of S. aureus (a) and S. pseudointermedius (b) isolates digested with SmaI restriction enzyme.


Table 1.
Nucleotide sequences and sizes of PCR products of amplified genes.
| Primer name | Oligonucleotide sequence (5′-3′)* | Amplicon size (bp) | Target gene | PCR conditions | Reference |
|---|---|---|---|---|---|
| nuc | GCGATTGATGGTGATACGGTTAGCCAAGCCTTGACGAACTAAAGC | 270 | nuc | 94°C, 5 min, 37 cycles of 94°C for 1 min, 55°C for 30 s, 72°C for 1 min, final extension 72°C for 7 min | [25] |
| mecA | AAAATCGATGGTAAAGGTTGGCAGTTCTGGCACTACCGGATTTGC | 533 | mecA | 94℃, 5 min, 40 cycles of 94℃ for 1 min, 58℃ for 1 min, 72℃ for 2 min, final extension 72℃ for 5 min | [26] |
| mecC | GAA AAA AAG GCT TAG AAC GCC TCGAA GAT CTT TTC CGT TTT CAG C | 138 | mecC | 94℃, 15 min, 30 cycles of 94℃ for 30 s, 59℃ for 1 min, 72℃ for 1 min, final extension for 10 min | [27] |
| sea | ACGATCAATTTTTACAGCTGCATGTTTTCAGAGTTAATC | 544 | enterotoxins | 94℃, 5 min, 35 cycles of 94℃ for 2 min, 57℃ for 2 min, 72℃ for 1 min, final extension 72℃ for 7 min | [28] |
| seb | GAATGATATTAATTCGCATCTCTTTGTCGTAAGATAAACTTC | 416 | |||
| sec | GACATAAAAGCTAGGAATTTAAATCGGATTAACATTATCCA | 257 | |||
| sed | TTACTAGTTTGGTAATATCTCCTTCCACCATAACAATTAATGC | 334 | |||
| see | ATAGATAAAGTTAAAACAAGCAATAACTTACCGTGGACCC | 170 | |||
| seg | CCACCTGTTGAAGGAAGAGGTGCAGAACCATCAAACTCGT | 432 | 94°C for 2 min, 30 cycles of 94°C for 25 s, 50°C for 20 s, 72°C for 40 s, final extension 72°C for 6 min | [29] | |
| seh | TCACATCATATGCGAAAGCAGTCGGACAATATTTTTCTGATCTTT | 463 | |||
| sei | CTCAAGGTGATATTGGTGTAGGCAGGCAGTCCATCTCCTGTA | 529 | |||
| sej | GGTTTTCAATGTTCTGGTGGTAACCAACGGTTCTTTTGAGG | 306 | |||
| sel | CACCAGAATCACACCGCTTACTGTTTGATGCTTGCCATTG | 240 | |||
| tst | ACCCCTGTTCCCTTATCATCTTTTCAGTATTTGTAACGCC | 326 | tsst-1 | 95℃, 5 min, 30 cycles of 94℃ for 1 min, 55℃ for 30 s, 72℃ for 1 min, final extension at 72℃ for 5 min | [12] |
| pvl | ATCATTAGGTAAAATGTCTGGACATGATCCAGCATCAASTGTATTGGATAGCAAAAGC | 433 | pvl | ||
| eta | GCAGGTGTTGATTTAGCATTAGATGTCCCTATTTTTGCTG | 93 | etaetb | ||
| etb | ACAAGCAAAAGAATACAGCGGTTTTTGGCTGCTTCTCTTG | 226 | |||
| lukS | TGTAAGCAGCAGAAAATGGGGGCCCGATAGGACTTCTTACAA | 503 | lukS | 94°C, 3 min, 35 cycles of 94°C for 1 min, 57°C for1 min, 72°C for1Min, final extension 72°C for7 min | [30] |
| lukF | CCTGTCTATGCCGCTAATCAAAGGTCATGGAAGCTATCTCGA | 572 | lukF | ||
| sak | TGAGGTAAGTGCATCAAGTTCACCTTTGTAATTAAGTTGAATCCAGG | 403 | sak | 94°C, 15 min, 35 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min | [31] |
| hla | CTGATTACTATCCAAGAAATTCGATTGCTTTCCAGCCTACTTTTTTATCAGT | 209 | haemolysins | 95 °C for 5 min, 30 cycles of 95 °C for 1 min, 58 °C for 1 min and 72 °C for 2 min, final extension of 72 °C for 10 min | [32] |
| hlb | GTGCACTTACTGACAATAGTGCGTTGATGAGTAGCTACCTTCAG | 309 | |||
| hld | AAGAATTTTTATCTTAATTAAGGAAGGAGTGTTAGTGAATTTGTTCACTGTGTCGA | 111 | |||
| hlg | GTCAYAGAGTCCATAATGCATTTAACACCAAATGTATAGCCTAAAGTG | 535 |
*F – forward primer; R – reverse primer. The sets of primers were synthesized by Genomed S.A, Poland. The concentration of primers was 0.04 µmol.
Table 2.
Susceptibility to antibiotics and chemotherapeutics (MIC) and the presence of virulence genes in CoPS strains.
Table 2.
Susceptibility to antibiotics and chemotherapeutics (MIC) and the presence of virulence genes in CoPS strains.
| Staphylococcus species | bird species | organ | MIC breakpointsμg/ml | Presence of virulence genes | ||||||||||
| MALDI-TOF MSBiotyperLog (Score) | Penicillin | Amoxicillin | Gentamicin | Chloramphenicol | Tetracycline | Enrofloxacin | Erythromycin | Trimethoprim | Sulfamethoxazole | |||||
| S. pseudintermedius | SP1 | 2.100 | Capercaillie | conjunctiva | ≤ 0.062 | ≤ 0.062 | ≤ 0.5 | ≥32 | ≤ 0.125 | ≤ 0.125 | ≤ 0.125 | ≥ 16 | ≤ 19 | - |
| SP2 | 2.096 | Capercaillie | joint | ≤ 0.062 | ≤ 0.062 | ≤ 0.5 | ≤ 4 | ≤ 0.125 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 38 | sec, hla, hld | |
| SP3 | 2.367 | Capercaillie | liver | ≥ 64 | ≥ 1 | ≤ 0.250 | ≥ 64 | ≥ 32 | ≤ 0.5 | ≥ 256 | ≤ 1 | ≤ 38 | - | |
| SP4 | 2.168 | Meadow pipit | joint | ≤ 0.125 | ≤ 0.125 | ≤ 0.250 | ≥ 32 | ≤ 0.250 | ≤ 0.125 | ≥ 16 | ≤ 0.5 | ≤ 38 | - | |
| SP5 | 2.008 | White stork | conjunctiva | ≥16 | ≥ 0.250 | ≤ 0.250 | ≤ 4 | ≤ 0.125 | ≤ 0.5 | ≤ 0.250 | ≤ 1 | ≤ 19 | - | |
| S. aureus | SA1 | 2.205 | Capercaillie | embryo | ≥ 32 | ≥ 64 | ≤ 1 | ≤ 4 | ≤ 0.125 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 19 | nuc, hla, hld |
| SA2 | 2.260 | Common kestrel | wound | ≤ 0.062 | ≤ 0.062 | ≤ 1 | ≤ 4 | ≤ 0.125 | ≤ 1 | ≤ 0.5 | ≤ 2 | ≤ 38 | nuc, seg, hla, hlb | |
| SA3 | 2.249 | Song thrush | palatal fissure | ≥ 8 | ≥ 4 | ≤ 1 | ≤ 8 | ≤ 0.250 | ≤ 0.250 | ≤ 0.5 | ≤ 0.250 | ≤ 19 | nuc, see, sej, sak | |
| SA4 | 2.314 | Capercaillie | conjunctiva | ≤ 0.062 | ≤ 0.062 | ≤ 1 | ≤ 4 | ≤ 0.250 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 38 | nuc, hla, hlb, hld | |
| SA5 | 2.419 | Capercaillie | embryo | ≤ 0.125 | ≤ 0.125 | ≤ 0.5 | ≤ 4 | ≤ 0.125 | ≤ 0.125 | ≤ 0.250 | ≤ 2 | ≤ 19 | nuc, hla, hlb | |
| SA6 | 2.363 | Common kestrel | palatal fissure | ≤ 0.062 | ≤ 0.062 | ≤ 0.5 | ≤ 8 | ≤ 0.250 | ≤ 0.125 | ≤ 0.5 | ≤ 1 | ≤ 19 | nuc, hla, hlb, hld | |
| SA7 | 2.054 | Common buzzard | palatal fissure | ≤ 0.062 | ≤ 0.062 | ≤ 1 | ≤ 4 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 2 | ≤ 9,5 | nuc, seg, hla, hlb | |
| SA8 | 2.146 | Capercaillie | liver | ≤ 0.062 | ≤ 0.062 | ≤ 0.250 | ≤ 4 | ≤ 0.5 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 19 | nuc, hla, hlb,hld | |
| SA9 | 2.326 | Capercaillie | embryo | ≤0.062 | ≤ 0.062 | ≤ 1 | 16 | ≥ 16 | ≤ 0.250 | ≤ 0.5 | ≥ 16 | ≤ 38 | nuc, hla, hlb | |
| SA10 | 2.359 | Common kestrel | liver | ≥ 0.5 | ≥ 0.5 | ≤ 1 | ≤ 4 | ≤ 0.5 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 19 | nuc, hla, hlb, hld | |
| SA11 | 2.230 | Capercaillie | palatal fissure | ≥ 4 | ≥ 2 | ≤ 1 | ≥ 256 | ≥ 256 | ≤ 0.125 | ≥ 256 | ≥ 8 | ≤ 38 | nuc, hlg | |
| SA12 | 2.030 | Common buzzard | spleen | ≤ 0.125 | ≤ 0.125 | ≤ 1 | ≤ 8 | ≤ 1 | ≤ 0.125 | ≤ 0.5 | ≤ 2 | ≤ 38 | nuc, seb, hld | |
| SA13 | 2.342 | Common buzzard | joint | ≥ 0.250 | ≥ 0.250 | ≤ 1 | ≤ 8 | ≤ 0.250 | ≤ 0.125 | ≤ 0.5 | ≤ 0.5 | ≤ 19 | nuc, hla, hld | |
| S. delphini | SD1 | 2.028 | Capercaillie | yolk sac | ≤ 0.062 | ≤ 0.062 | ≤ 0.250 | ≤ 4 | ≤ 0.125 | ≤ 0.125 | ≤ 0.5 | ≤ 2 | ≤ 38 | see, hlb, hld |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



