
Submitted:
08 January 2024
Posted:
09 January 2024
You are already at the latest version
Abstract
This comprehensive review scrutinizes tissue culture and micropropagation methodologies in geophytes, focusing on bulbous plants. The examination encompasses key stages, including somatic embryogenesis, bulb growth, dormancy breaking, and planting. Studies underscore the pivotal role of plant growth regulators in plant regeneration and bulb growth. Bioreactor systems for healthy plant regeneration, rooting methods, acclimatization strategies, and considerations for ex vitro survival are elucidated. The review also delves into somaclonal variation dynamics and acknowledges the burgeoning field of genetic editing, particularly CRISPR studies, as a promising avenue for enhancing valuable compound content in geophytes. The conclusion emphasizes the necessity of a multifaceted approach integrating biochemistry, physiology, and molecular biology to address existing challenges and propel tissue culture protocols for diverse geophyte species.
Keywords:
flower bulbs
; organogenesis
; somaclonal variations
; somatic embryogenesis.
1. Ornamental Geophytes History
For centuries, flower bulbs have captivated societies worldwide, drawing attention for their enticing fragrance, vibrant colors, and diverse blooming periods. These bulbs have held a significant place in various forms of art [1-3]. Greek philosophy dating back to 300 BC mentions the medicinal and consumable properties of flower bulbs like Crocus, Colchicum, and Gladiolus. Greek mythology also references bulbs such as Lilium, Hyacinthus, Crocus, Iris, and Narcissus [4,5]. Japan and China are known to have numerous wild Lilium species endemic to their regions. The United Kingdom is the origin of Narcissus pseudonarcissus, commonly known as the lenty lily, which serves as the ancestor for many cultivated daffodil varieties. The Persian-style gardens of the Taj Mahal, an iconic 17th-century monument, feature essential flower bulbs like Anemone, Ranunculus, Fritillaria, Iris, Lilium, Pancratium, and Gloriosa [6]. Flower bulbs such as Aztec lily, canna, dahlias, and tuberose are reported to have originated from the Americas [7]. In the 1180s, flower bulbs, particularly those with iris motifs, adorned the military uniforms of Italians from Florence-Tuscany and the French [8].
The worldwide spread of tulips has sparked numerous hypotheses. Originally found in the western regions of China, the Caucasus, Turkey, Iran, and Central Asia, tulips first appeared in Anatolia in 12th-century Konya motifs. The breeding of tulips began during the Ottoman Empire in the 16th century [9]. Tulip mania reached its peak between 1673-1736 during the Ottoman era, earning that period its name. The arrival of tulips in Europe has trigged offseveral hypotheses. One suggests that Busbeck, an ambassador representing the Austro-Hungarian Empire in 1554, brought tulip bulbs from Istanbul to his friend Carolus Clusius in Austria. Another possibility involves a French physician named Belon, who traveled to the Near East in 1549 and visited Istanbul. Belon recorded in his memoirs that numerous foreigners journeyed to Istanbul by ship to acquire tulip bulbs, which he referred to as the "red lily." These accounts shed light on the early introduction of tulips to Europe and the growing fascination with these captivating flowers. In the 17th century, tulip mania took hold in the Netherlands. The tulip, originally imported from Ottoman lands in the 16th century, became a highly sought-after commodity in the 18th century, primarily sourced from the Netherlands [9-12].
As mentioned by Rees [13], ancestor’s habitats of some flower bulbs was reported by Bailey [14] such as Tulipa: Mediterranean and across Asia to Japan; Hyacinthus: Greece, Syria, and Asia Minor; Narcissus pseudonarcissus L.: Sweden to Spain and Romania; Narcissus tazetta L.: Canary Isles to Japan; Narcissus poeticus L.: France to Greece; Iris tingitana Boiss. & Reut.: Morocco; Iris reticulata Bieb.: Caucasus; Iris xiphium L.: Spain and North Africa; Freesia South Africa; Lilium longiflorum Thunb.: Japan; Lilium regale Wils.: West China.
Over time, flower bulbs have been recognized for their unique characteristics and have been cultivated with consideration for their potential uses and economic and social values. They have transitioned from being sold as individual bulbs to being traded in bulk quantities and have found commercial applications as cut flowers and potted plants. By the conclusion of the twentieth century, the Netherlands emerged as the leading global player in the flower bulb trade, asserting its dominance in this industry [8].
2. World Ornamental Plants Sector – Flower Bulbs situation in the sector
The global production of ornamental plants has witnessed a widespread increase, playing a crucial role in boosting the economies of more than 50 countries. According to Gabellini and Scaramuzzi [15], world cut flower and potted plants production counts for a value of about 35.5 billion with a cultivation area of 745,000 ha. Asia-Pacific is the largest region covering about 79% of the total surface area dedicated to ornamentals. The European Union occupies about 10% of the world flower production surface [16] but thanks to the high productivity per hectare, the ornamental industry represents about 40% of the worldwide production (value of more than 11 billion EUR). The biggest producers are the Netherlands (32%), France (12%), Germany (12%), Spain (12%) and Italy (11%). Outside EU, Mexico, Colombia, Ecuador are important producer countries in Central America. Thanks to favorable climatic conditions, foreign investments, and relatively cheap labor, Latin American countries have become an important export region for both the American and European markets. In South America, Brazilian floriculture has developed in the past five years with a cultivated surface of about 15,600 ha and a production value of 1.7 billion EUR; the internal market absorbs most of this production [17].
Countries such as Kenya, Tanzania, Ethiopia, Uganda, and Zambia in Africa, especially in the equatorial belt, have boosted the production and export of cut flowers. Advantages such as the economy being based on agriculture, suitable climatic conditions and cheap labor in African countries have led to the development of ornamental plant production. Production areas are generally processed by professional companies and consist of large-scale nurseries owned by foreign investors.
The global trade volume of ornamental plants is expected to surpass 60 billion USD (about 54.6 billion EUR) in 2027 at CAGR of 8.8% (source: Flower and Ornamental Plant Global Market Report; https://www.thebusinessresearchcompany.com/report/flower-and-ornamental-plant-global-market-report), with a particular emphasis on markets with strong purchasing power, including Europe, USA, and Japan. The Netherlands confirms its central and dominant role in the international trade of flowers and ornamental plants thanks to the combination of national production and re-exported products. Other important exporting countries in the world are Colombia, Kenya, Ecuador, and Israel. The EU is a net exporter of pot plants, conifers and hardy perennial plants, bulbs and corms, and a net importer of cut flowers and cut foliage. The most important European importing countries for flowers and potted plants are Germany, France, and UK
Ornamental geophytes, also called flower bulbs, contribute significantly to the global ornamental industry, and are used for commercial bulbs, cut flowers or potted flowering plant production, landscaping, and private gardening. Moreover, it is well recognized their utility as food or medicinal and aromatic plants [16,18-21].
In a report published in 2010, Benschop, et al. [1] estimated the global value of the flower bulb industry at more than 1 billion USD with perspectives of expansion all over the world. In 2022, the product category “Bulbs and Roots” including dormant and in growth bulbs, tubers, roots, corms, crowns and rhizomes registered a global value of export of over 2 billion USD (about 1.82 billion EUR) with a slight decrease with respect to the year 2021 counterbalanced by a 4% increase in the period 2018-20 (source: Trade statistics for international business development; https://www.trademap.org/). The leading producer is the Netherlands, but significant producers of ornamental geophytes emerged in Chile, New Zealand, and China. The production is also present in other Latin American and Asian countries and in Southern Africa. Although ornamental geophytes consist of more than 800 different genera, the industry is still dominated by a few genera. Beside genera that comprise most of the worldwide bulb production (Tulipa, Lilium, Narcissus, Gladiolus, Hyacinthus, Crocus and Iris), there are other ornamental geophytes of economic importance in the cut flower industry (e.g. Freesia, Altroemeria, Hippeatrum and Zantedeschia). Anemone and Ranunculus are two important genera for cut flower production under Mediterranean conditions. In 2022, the EU exported a total of over 100 million EUR worth of orchid, hyacinth, narcissi, and tulip bulbs in growth or in flower; the imports were of 30.9 million EUR. Of all EU countries, the Netherlands was responsible for 81% of all exports of these bulbs and other exporter countries were Lithuana (7%), Poland (5%), Denmark (2%) and Latvia (2%). The most part of these exported bulbs are addressed to the European area (Switzerland, UK, Norway, Russia and Ukraine; Eurostat database, 2023; https://ec.europa.eu/eurostat/web/products-eurostat-news/-/edn-20230406-1).
As the global economic situation improves and people want to spend more, the demand for high-quality products increases. This trend also applies to the global market of flowers and ornamental plants. Consequently, there is a growing demand for high-quality bouquets from different flower collections. Thanks to the huge diversity of the genera, flower bulb can satisfy the need of new products; in addition, the increasing research progresses enable to set a productive chain also enhancing the local flower industry. This can be of particular interest for countries which are aimed at diversifying their production from the major popular floriculture products. An interesting example is provided by Turkey which is very rich in natural resources of ornamental geophytes and quite recently started to develop a commercial production for flower bulbs with 42 ha dedicated (Turkish Statistical Institute-2023).
In the following paragraphs, we want to review the most salient and recent findings for the micropropagation of flower bulb crops by providing examples of successful application of tissue culture techniques and outlining concluding remarks on the state of the art of bulbs in vitro culture and future perspectives.
3. Flower bulbs propagation and challenges
Horticulturally, flower bulbs, a broad synonym of ornamental geophytes, encompass many categories of plants used for flowering at different times of the year, used for landscaping, amenity, cut- flower production or grown for their foliage as indoor or outdoor plants. Flower bulbs exhibit remarkable diversity in their developmental biology and physiological responses to environmental cues, and several genera hold significant economic value [1,22,23]. Ornamental geophytes can be evergreen or deciduous, annual, or perennial and can change their growth and flowering characteristics in cultivation or in a different environment [24]. The majority of bulbous plant species, such as tulip and lily, belong to the monocot group, which includes important ornamental plants [25].
Geophytes are plants with storage organs which are of several kinds, morphologically and physiologically (Figure 1). These storage organs can allow the plant to survive the unfavorable periods (whether too dry, too hot, or too cold) [13,24,26]. Thanks to the nutrient reserves within these modified structures the plant viability and the further plant development is ensured despite the severe external conditions [27,28]. Prior to the onset of favorable environmental conditions, the differentiation of new buds occurs, allowing for shoot development through the ongoing physiological processes in the underground organs. Additionally, the storage organs enable vegetative reproduction of flower bulbs, offering a viable alternative when seed propagation is hindered by adverse environmental factors [16,22,29]. Geophytes can be propagated through generative (seed production) and vegetative methods [16]. The commercial production of flower bulbs through seed propagation is not widely preferred due to several reasons [29,30]. Firstly, seed production could result in offspring that are not identical to the parent plant, leading to increased diversity. This lack of uniformity makes this kind of propagation unsuitable for commercial production, except for specific seed-raised crop. Furthermore, the seed of some bulb plants has limited viability and precise requirements of germination could be necessary to produce seedlings. Another difficulty in applying seed multiplication is the extended juvenile phase, lasting up to 5-7 years before flowering, which could be faced for certain species [30]. Nevertheless, seed production is highly valuable in breeding programs, allowing for the development of new cultivars.
Seed propagation is particularly advantageous if a high number of seeds can be obtained, the juvenile phase is relatively short, and variability is not a concern compared to vegetative propagation [24]. Another advantage in seed propagation is that in almost all cases, plants produced are free of virus infe ction; vice versa, through vegetative propagation methods there is the risk of spreading pathogenic microorganism and compromising the production. Examples of geophytes that can be commercially propagated through seed production include Anemone, Allium, Begonia, Cyclamen, Freesia, Fritillaria and Ranunculus [16,22,29-31].
Bulbs go through distinct developmental phases: juvenile vegetative, adult vegetative, and reproductive. The transition to the adult vegetative phase is necessary for flower initiation, which occurs after several years in the juvenile phase for certain species like Tulipa and Narcissus. The shift to the reproductive phase is triggered by high temperatures, leading to flower bud formation. Dormancy follows, requiring a prolonged period of cold for dormancy release and preparation for spring growth. This life cycle is common among bulbous plants, including Tulipa, Crocus, and Hyacinthus [13,22,25].
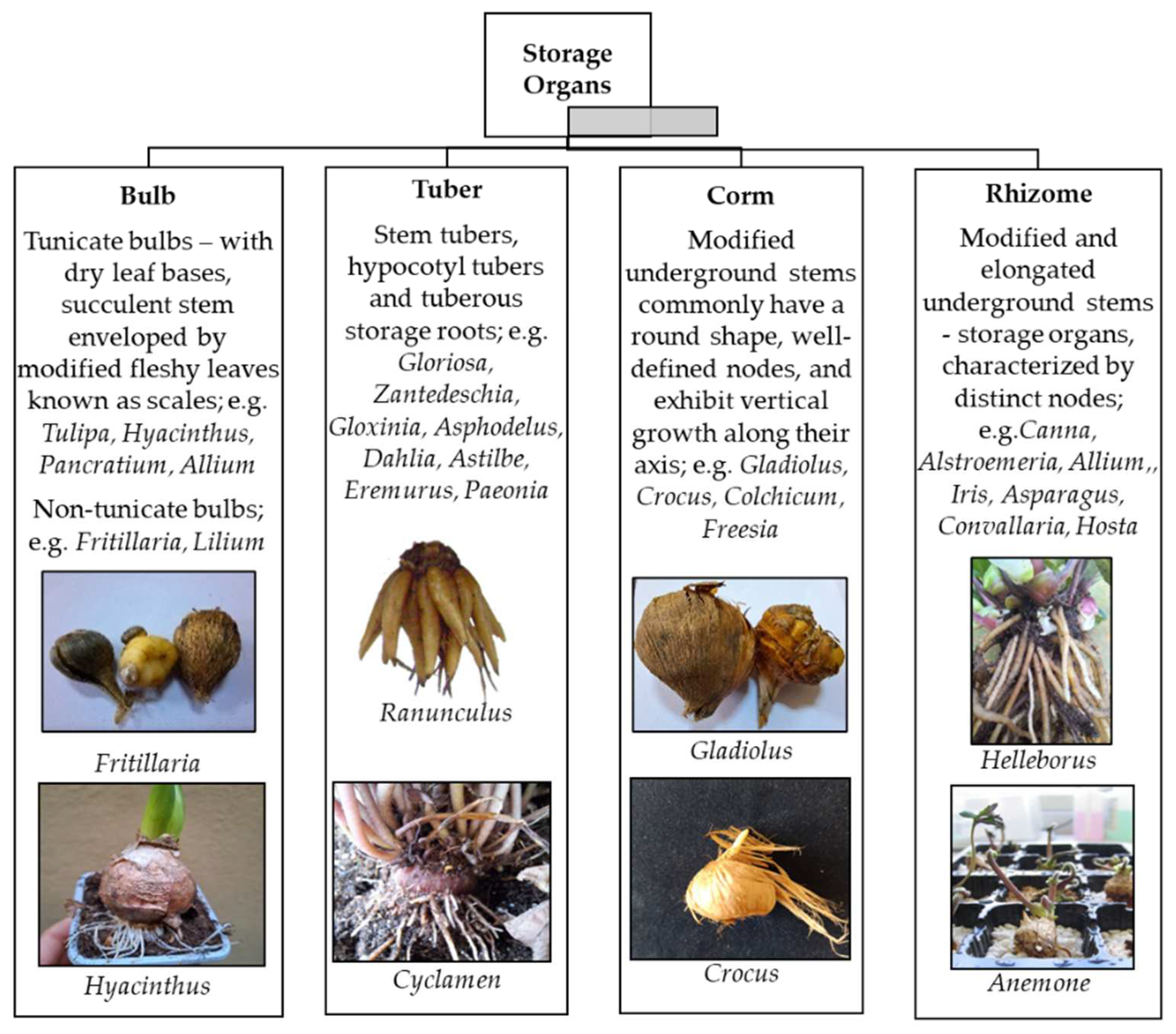
Flower bulbs propagate in nature with natural division. Natural division can be with daughter bulbs (tulip, crocus), offsets (hyacinths, muscari) bulblets (lily), bulbils (lily), cormels, rhizomes [32], branched rhizomes and stolons (allium,oxalis) [24,30,33]. Multiplication rates of the natural division are quite low [13,16,24]. For this reason, it has been envisaged that flower bulbs need alternative propagation methods. Traditional propagation methods include scaling, twin scaling, chipping, scooping, scoring, as well as stem and leaf cuttings [33]. Scaling is the method where individual scales are removed from the bulb to produce bulblets. Twin scaling, a variation of scaling, involves attaching two scales to a piece of the basal plate. Chipping, a mechanical technique similar to twin scaling, is employed to separate scales. Scooping involves removing the basal plate from the scales, while scoring entails cutting the basal plate. Additionally, stem and leaf cuttings can be used as propagation methods for certain bulbous plants. These techniques have been widely practiced by horticulturists and gardeners to propagate and expand their bulb collections, ensuring the continued availability and diversity of these plants [16,30,31]. It can also be considered the bulbous structure and propagation method. Tunicate bulbs can be multiplied using techniques such as offsets, scoring, scooping, coring, sectioning, and cottage. Non-tunicate bulbs, on the other hand, can be propagated through scaling after they have bloomed. Corms can be multiplied using cormels, which are small corms that develop between the old and new corms. It typically takes one to two years of growth for cormels to reach the size when they can produce flowers. Tubers can be propagated by either planting the entire tuber or dividing it into sections, ensuring each section has at least one eye or bud. Rhizomes are divided into sections, each containing at least one shoot bud or active shoot. Some rhizomes have roots attached to their bottoms and can be individually planted in containers. Rhizomes can also be planted in nursery beds and used as a source of bare-root stock for planting or for cultural purposes such as basketry. Crown division is another method of propagation that differs slightly from dividing rhizomes [24,34]. However, these methods also present notable limitations. Firstly, generating a substantial number of plants that are true-to-type within a practical time frame can be challenging. Secondly, the propagation process carries a significant risk of disease transmission. Tissue culture techniques offer effective solutions to conventional challenges in plant propagation with advantageous for geophytes such as expediting the multiplication of species and cultivars, generating plants free from viruses, contributing to advancements in crop quality, and allowing the 0synthesis of secondary metabolites. [16].
4. Micropropagation
Micropropagation is a widely recognized method of vegetative propagation that allows to produce numerous offspring plants. This process involves multiplying plants under sterile conditions on a nutrient culture medium with known composition and optimal artificial culture conditions [35]. Micropropagation represents an intricate plant multiplication technique involving substantial capital investment, significant operational costs, and a need for specialized expertise [36-38]. However, tissue culture methods offer various benefits, particularly in micropropagation which is employed to multiply diverse plants, including genetically modified or conventionally bred ones. It proves useful for producing plantlets from seedless or challenging-to-reproduce plants economically and quickly, significantly reducing the time needed for abundant plantlet production [29]. Podwyszyńska, et al. [39] reported that the global annual production of ornamental plants through in vitro cultures has surged from 800 million to 2 billion in the last decade. According to a recent research report, the micropropagation market size is expected to rise by 2030 reaching the value of 2.8 billion USD (about 2.5 billion EUR; report "Micropropagation Market- Global Outlook and Forecast 2022-2027"- https://www.reportlinker.com/p06313600/?utm_source=GNW). Europe is one of the most prominent geographical segments in the global micropropagation market with 140 commercial in vitro laboratories for which the production of ornamental plants is pivotal in their production [39].
4.1. In vitro regeneration pathways
In vitro plant regeneration is a process where explants, through cell division and differentiation, give rise to the formation of organs and tissues [40]. This comprehensive process involves the regeneration of an entire plant from various sources, including adult tissues, organs, unorganized calli, or even a single cell [41,42].
Somatic embryogenesis (SE) is a remarkable process in which plant somatic cells undergo dedifferentiation, acquiring embryonic cell characteristics, and subsequently through embryonic development form complete plants [43-45]. This process highlights the totipotency of plant cells, demonstrated by the formation of embryogenic callus [46,47]. It involves the development of structures resembling zygotic embryos from cells that are not associated with zygotes or the original tissue, closely mirroring the characteristic stages observed during zygotic embryogenesis [45]. Somatic embryos can differentiate through two pathways: direct somatic embryogenesis, where they differentiate directly from explants without the formation of callus, and indirect somatic embryogenesis, where differentiation occurs after passing through a callus stage [48]. Somatic embryogenesis is favored for mass propagation due to its higher proliferation rate, ease of liquid culture medium utilization, efficient handling of numerous embryos, and greater adaptability for bioreactors [29,40]. However, various factors such as explant source, genotype, nutrient media, stress conditions, hormone induction, subculture, electrical excitation or gene expression modification can influence somatic embryogenesis [43,44,46,49,50].
Freshly formed structures like shoots, roots, or embryos can emerge on plant tissues without preexisting meristems, and these recently developed organs are termed adventive or adventitious and this initiation of a novel structure and organization is termed organogenesis [51,52]. Organogenesis involves perception of phytohormones, differentiation of specialized cells to gain organogenic competence, cell division regulation, cell expansion, and the overall patterning of the organ [32,53,54]. The presence or absence of the callus stage results in two distinct developmental stages. The developmental stage that includes the callus phase is called indirect organogenesis. In indirect organogenesis, the likelihood of observing somaclonal variation is higher. The developmental stage that occurs without the callus phase is termed as direct organogenesis [55,56]. The first step in the propagation of plant material using organogenesis technique is the selection of the explant source from the donor plant. Usually, embryos, seedling parts, apical meristems (root or shoot tips), primordial organs, protoplasts, young flower buds are used [54,55].
4.2. The key factors affecting micropropagation of flower bulbs
Establishing a successful micropropagation protocol is a complex task, influenced by various interacting factors, including plant materials, culture conditions, and culture media ingredients. Despite the development of micropropagation protocols for many flower bulb crops, challenges hinder commercial viability. Key limitations include the high cost of tissue-culture plantlets, inefficiencies in the initiation phase, low propagation rates for numerous genotypes, challenges during the acclimatization of ex vitro plantlets, and the occurrence of off-types.
4.2.1. Explant choice
The selection and preparation of suitable explants play a crucial role in the successful culture of ornamental bulbous plants. Factors such as the regeneration capacity, physiological state and hormonal balance of the donor plant, the origin, age and size of the explants, their polarity, and pre-treatment methods are key considerations [57]. The induction of bulblets in vitro is primarily influenced by the temperatures and storage durations of the mother plant. These situations will be explained in the part 5.1. A variety of tissue sources, including shoot tips (apical and axillary buds), bulb scales, leaves, stems, and different parts of the inflorescence, are utilized as explants in the initial stages of culturing these plants [30,58]. Yasemin, et al. [59] emphasized the significance of explant types regarding factors such as callus color, callus hardness, callus fragility, callus formation rate, embryogenic callus rate, and callus growth rate of Pancratium maritimum.
4.2.2. Culture medium
The composition of the culture medium is a critical factor influencing the growth and morphogenesis of plant tissues in culture. The Murashige and Skoog (1962; MS) formulation is widely used, initially designed to support optimal growth of tobacco callus through extensive dose-response curve studies for essential minerals. Furthermore, N6 Chu [60], Woody Plant Medium (WPM)[61], and B5 [62] are also used as culture media in tissue culture [46]. Plant tissue and cell culture media are generally made up of some or all the following components: macronutrients, micronutrients, sugar(s), vitamins, amino acids or other nitrogen supplements, other undefined supplements, solidifying agents or support systems, and growth regulators [63]. Undefined elements can be coconut milk, meat, malt, yeast extracts, potato extracts, juices, pulps, fruit extracts, plant/plant pars/seedling extracts. Osmotic balance, pH and buffers of the culture medium also effect the target in the tissue culture [64]. Elicitors such as chitosan, aminolevulinic acid (ALA), alginate (Ag), N-acetylglucosamine (NAG), salicylic acid (SA), hyaluronic acid (HA), silver nitrate (AgNO3), jasmonic acid (JA), methyl jasmonate (MeJA), phloroglucinol (PG), and pectin, are extensively employed to induce secondary metabolites in plant tissue culture [65]. Methyl-Jasmonate (MeJA) and various polyamines, also prove advantageous in tulip tissue culture by promoting efficient bulb formation during micropropagation [66]. Moreover, the medium's physico-chemical properties, consistency can alter the nutrient status of microshoots. The growth and multiplication of Ranunculus asiaticus shoots were influenced by the agar brand used for culture medium solidification, resulting in varied water and nutrient availability; in addition, the agar impurities affected the biological performance of microshoots [67] (Figure 3). The use of solidified media in micropropagation offers advantages, including easy visibility and recovery of small explants, maintained explant orientation, improved aeration without special measures, and orderly growth of shoots and roots. In contrast, liquid media can cause disoriented growth and difficulties in shoot separation. However, there are drawbacks to semi-solid media, such as the potential presence of inhibitory substances in agars, slow growth rates, and limited diffusion of toxic exudates. Poor oxygen diffusion to developing roots is a concern, even in aerated liquid media. Adherence of gel to roots can pose issues during plantlet transfer to soil, and cleaning containers for re-use becomes time-consuming with semi-solid media [68]. Different plants have varying nutritional requirements, affecting optimal growth and morphogenesis. Tissues from different plant parts may have unique needs for satisfactory growth [46].
4.2.3. Environment conditions
Temperature and light have the key roles in micropropagation. Under in vitro conditions, younger tissues typically import sucrose and have limited photosynthesis. Despite this, low light intensities are essential for organ formation [69]. In vitro morphogenesis or organogenesis can be greatly affected by altering the light color and intensity of incubation [30,65]. White fluorescent light (350~750 nm) is conventional in in vitro culture but has drawbacks like high electricity consumption and uneven radiation. Monochromatic LEDs with specific wavelengths are now widely used for more energy-efficient and controlled in vitro plant propagation [65,70]. Light is crucial for regenerating storage organs in the in vitro cultures of geophytes. Phytochrome governs the plant's light response, influencing organogenesis induction and direction. Blue light and its receptor cryptochrome play a role in storage organ formation [71]. Examining light quality effects during adventitious organogenesis in bulbous plants, researchers utilized monochromatic fluorescent lamps. The results revealed that in in vitro cultures of Hyacinthus orientalis, blue light stimulates adventitious shoot development, while red light encourages the formation of adventitious bulbs [72]. Studies have explored the impact of light on plant growth and development parameters in various lily species and cultivars during in vitro organogenesis. [73,74,75 2023]. According to Bach and Sochacki [30], the choice of light may interact with tissue cytokinin metabolism, potentially influencing in vitro tissue dormancy and subsequent development. While light is crucial for typical green shoot and plantlet growth, unorganized cell and tissue cultures can thrive without it. Surprisingly, darkness may enhance growth and morphogenesis, with brief dark periods observed to promote shoot morphogenesis in certain cases [68]. Embryo explants from P. maritimum were cultivated under both photoperiod and dark conditions. The results, based on unpublished data from Yasemin S., indicate that the dark condition was more successful in generating callus and Figure 4 illustrates the outcomes after one month of incubation.
Temperature influences both growth rates and the transition between vegetative and reproductive phases in plant development. In flower bulbs undergoing in vitro culture, temperature is a vital factor influencing regeneration, bulbing, and dormancy. For temperate ornamental geophytes, lower temperatures mimic winter, prompting dormancy. Under in vitro conditions, reduced temperatures induce storage organs to withstand unfavorable growth conditions [76]. Temperature is another key factor able to influence regeneration, bulbing and dormancy of flower bulbs during the in vitro culture [77]. In vitro regenerated bulblets necessitate dormancy breaking to facilitate ongoing development. A cold treatment was proved to be effective for bulblet initiation and forbreaking dormancy in the regenerated bulblets [30].
5. Micropropagation of flower bulbs
Micropropagation of ornamental plants has been studying since the 1950s [76,78]. The impact of this technology on flower bulbs is also testified by the literature cited in this review. Micropropagation involves enhancing axillary bud development through organogenesis, adventitious shoot formation, and somatic embryogenesis [79].
Plant biotechnology significantly contributes to the production of ornamental geophytes through various approaches. These methods focus on propagating specific genotypes, acquiring virus-free plant material, and supporting breeding and crop improvement programs [16,29,69,79]. Techniques such as callus culture, embryo rescue, in vitro pollination, somatic hybridization, induction of somaclonal variation, protoplast culture, synthetic seed production, and in vitro ploidy manipulation, genetic transformation, gene mapping, and DNA fingerprinting play essential roles [19,65,79]. In vitro pollination and fertilization, embryo, ovary, ovule, anther and pollen culture have been reported for some geophytes as Lilium, Cyclamen, Anemone, Ranunculus and Amaryllis [80-87]. Dhooghe, et al. [80], investigated pollen-stigma interactions in crosses between A. coronaria and R. asiaticus which are important ornamental species from Ranunculaceae. In their study, they confirmed the pre-fertilization barriers with pollen growth obstacles. They showed with some stigma treatments that these obstacles can be overcome. Treating the stigma with the combination of auxin 2,4 D and cytokinin kinetin 24 hours after pollination, regardless of the cross-direction, led to a significantly higher ratio of pollen tube length to total style length and an improved seed set. Tütüncü and Mendi [83], evaluated the effects of pollination with gamma-irradiated pollen on in vitro ovule cultured in Cyclamen persicum L. They ϒ-irraditated (0-control-, 50, 100, 150, 200, 300, and 450 Gy doses) flower buds which were collected before anthesis. The fruits were gathered 30 days post-pollination, and ovule explants were subsequently cultured. They also investigated different MS media combinations with different plant growth regulators. The effects of nutrient media and irradiation dose on plant formation were important. The lowest plantlet regeneration (0.33%) was obtained from ovule explants which pollinated with the pollen exposed to 50 Gy gamma rays and cultured on M3 (full-strength MS salts and vitamins, 30 g/L sucrose, 2 g/L glucose, 2.0 mg/L 2,4-D and 0.8 mg/L 6-( γ, γ-dimethylallylamino) purine-2iP-) media while the highest plantlet regeneration (2.66%) was obtained from ovule explants cultured on M1 media (full-strength MS salts and vitamin, 30 g/L sucrose, 2 g/L glucose, 10 g/L maltose, 1.0 g/L proline, 2.0 g/L peptone, 200.0 mg/L spermidine and 0.5 mg/L kinetin) 30 days after pollination with non-irradiated pollen grains. In their study in situ parthenogenesis was not obtained. They supposed that the lack of in situ parthenogenesis can be explained by the factors such as plant genotype, type of applied radiation, and amount of radiation dose. Tütüncü and Mendi [88], evaluated the pollen viability, fertilization, pollen grain germination and zygote formation via histological analysis in wild and commercial C. persicum. According to their results, pollen viability was between 82.1-84.3%, and in vitro germination rates were between 63.8-65.2%. Morever, they found the fertilization in the first two days after pollination and as a result of histologic analysis, zygote formation was detected at the 20th day after pollination in wild cyclamen. In the commercial cultivar pollen tube arrived the ovary between 5-7 days after pollination and zygote formation occurred 30th and 40th day. Sevindik, et al. [81] aimed to devise a proficient method for haploid plant regeneration in both wild Cyclamen persicum and the commercial F1 Melody cultivar. This was achieved through anther and ovule culture techniques. Microspore uninuclear stages were discerned using DAPI dye after subjecting buds to a two-day cold pre-treatment at 4 °C. Successful haploid embryo development was observed in C. persicum on B5 medium with 1.0 mg/L NAA (100% success rate). An efficient ovule culture protocol was established for C. persicum using 2.0 mg/L 2,4-D and 0.8 mg/L 2iP, and for Melody F1 with 2.0 mg/L 2,4-D and 0.5 mg/L 2iP, both achieving a 100% success rate. Spontaneous double haploidization was verified in C. persicum through flow cytometric analysis. Further scientific inquiry is warranted to evaluate the feasibility and potential applicability of protoplast culture and somatic cell hybridization techniques in the context of geophytes. Karamian and Ebrahimzadeh [89] investigated protoplast isolation and regeneration in Crocus cancellatus using embryogenic calli. They obtained calli from shoot meristem culture and isolated protoplasts, finding optimal growth in Ca-alginate beads with nurse cells. Microcalli appeared after 4–5 weeks and transferring them to half-strength MS medium enhanced embryogenic calli growth. Somatic embryos developed on growth regulator-free or abscisic acid-supplemented medium. Matured embryos germinated with gibberellic acid, and plantlet formation occurred with 6-benzyladenine and α-naphthaleneacetic acid. Recently, Koetle, et al. [90] reported about the procedures using Agrobacterium strains to transform to the geophytes such as Crocus, Allium, Agapanthus, Lilium, Tulip, Gladioulus, Hyacinthus, Narcissus [91-100].
Micropropagation systems were developed for major bulbous plants but commercially propagation was achieved just for few bulbous plants such as Lily, Zantadeschia [29]. For other ornamental geophytes, there is a need for more efforts due to the initial limited success in micropropagation. Indeed, many scientific papers have been published on flower bulbs highlighting the discrepancy between the potentiality of this technique and the practical application. Following, we would like to analyze the most important findings related to the major steps in micropropagation of flower bulbs crops which are: preparation of mother plants, initiation, multiplication, bulbing/bulb growth, dormancy breaking, planting. Dormancy breaking is the different step from the other herbaceous and woody plants [16,29].
5.1. Stage 0: Preparation of mother stock plant material
Happy plants make happy plantlets. Stage 0 in ornamental geophyte propagation represents the initial and crucial phase dedicated to ensuring the selection and cultivation of healthy plants [37]. This pivotal stage aims to pre-process the chosen specimens to prevent contamination, ultimately fostering the development of thriving plantlets [33]. Key pre-treatments, such as regulating humidity, irrigation methods, temperature, and light exposure, are implemented to curtail the potential source of contamination and maintain the genetic integrity of the desired traits. Additionally, the duration of storage impacts the formation of bulb-like structures in geophytes, further emphasizing the significance of meticulous handling during this phase [16,36,69,101-104].
To initiate the in vitro culture of tulip, Podwyszyńska and Sochacki [105] selected healthy, true to type, and virus-free plants during their flowering phase in spring. After having carried out ELISA tests to verify the virus-free status of their plants, they harvested the bulbs in June and stored them at 17–20°C until October, followed by dry cooling at 5°C. The selected healthy bulbs (the recommended bulb size is 10 cm diameter) were then planted in pots with a fungicide (Captan)-soaked perlite substrate and forced in darkness at 9°C for 5–14 days prior to initiate the in vitro culture. The effect of a bulb pre-treatment at 5±2 °C for a period of 3–4 weeks was positively referred also by Muraseva and Novikova [106] who developed an efficient protocol for in vitro propagation from bulb scale explants for Fritillaria ruthenica Wikstr., an endangered and rare species. In other cases, natural conditions were advisable to prepare the mother plants. Kumar, et al. [107] successfully established an efficient plant regeneration system for Lachenalia. viridiflora via somatic embryogenesis by growing the mother stock plants in the greenhouses under controlled conditions that closely resembled the natural habitat of this plant species which is collected from a specific sub-population in St. Helena Bay, Western Cape, South Africa, credited to Prof. Graham Duncan. Around 10 mature bulbs of L. viridiflora were carefully selected and planted in terracotta pots with a standardized diameter of 200 mm. The potted plants were then nurtured under ambient temperature conditions and exposed to the natural photoperiod under the controlled environment of the University of KwaZulu-Natal Botanical Gardens' greenhouse. The researchers took special care to ensure regular watering and maintained a weed-free environment to promote optimal growth and development, mimicking the plants' natural conditions as closely as possible. Mirici, et al. [108] studied the in vitro bulblet regeneration from fresh bulb scale and immature embryos of the endangered geophytes Sternbergia fischeriana. Before explant sterilization, the collected bulbs were subjected to a drying process in the dark at room temperature for a duration of 6 weeks.
In conclusion, it could be argued that closed environments, such as glasshouses and tunnels are high-quality places for plant cultivation with a reduced risk of contamination. Treating plants with fungicides and insecticides before obtaining plant material helps to decrease the risk of contamination. Various practices are available to enhance in vitro development, altering the physiological state of the parent plant from which explants will be taken. These practices include implementing long-day treatments, cultivating under red light, maintaining bulbous plants at low temperatures, and applying plant growth regulators to the leaves [38]. By focusing on the careful selection and pre-processing of mother plants, stage 0 lays the foundation for a successful propagation process, leading to the cultivation of healthy ornamental geophytes [37].
5.2. Stage 1: Establishment of aseptic culture
In this stage, the selected explants are excised from the stock plant material and disinfected with certain chemicals before they are inoculated in the culture medium. Disinfecting plant explants for successful establishment in in vitro conditions, particularly in the context of plant tissue culture is very important. The primary goal is to eliminate microbial contamination, both exogenous and endogenous, without causing harm to the plant tissues. The success of the disinfection process is crucial for the subsequent stages of tissue culture [52,109-112]. Various chemicals such as antibiotics, fungicides, sodium hypochlorite, mercuric chloride, ethanol, hydrogen peroxide, calcium hypochlorite and silver nitrate are commonly used for surface sterilization. The concentration and duration of the disinfection process need to be carefully considered to achieve effective sterilization without compromising the viability and regeneration ability of the explants. The choice of disinfectants depends on the specific plant species and type of explant. The goal is to create aseptic conditions for plant tissue culture by addressing the challenges associated with microbial contamination and maintaining the health of the explants [110,113]. The use of seeds as an initial material in in vitro studies is advantageous because it decreases the risk of contamination. After germination of the seeds, in vitro seedling parts can be used as explant [52,112,114,115]. Yasemin et al. [112] conducted the disinfection experiments for the seeds of P. maritimum. They used the 70% ethanol and washed the seed with the distilled water following by treatment with different concentrations of NaOCl (1.6% and 3.2%) at different durations (15, 20 and 25 min). High NaOCl and long durations had negative effects on the seeds viability, germination and plant formation. Kumar, et al. [107] described the successful surface sterilization process for the Lachenalia viridiflora leaves. The leaves were treated in a laminar flow bench using 70% ethanol (v/v) for 60 seconds, followed by immersion in 2% sodium hypochlorite (NaOCl) for 10 minutes. To aid in the process, a few drops of Tween 20 were added as a surfactant. After sterilization, the leaves were rinsed three times with sterile distilled water.
Unfortunately, the contamination issue represents a substantial concern for the flower bulb tissue culture, particularly when underground plant tissues are used to initiate the culture. Although surface sterilization is successful, the plant tissues can be contaminated due to endogenous microorganisms in the bulbs and this could appear also in the further culture cycles [29]. Therefore, a thorough sterilization process is indispensable to eliminate contaminants from the geophytic storage organ explants. In the study conducted by Lagram, et al. [116], a protocol was employed for the preparation of saffron daughter corms (Crocus sativus L.). The corms were subjected to a series of treatments, including a 30-minute immersion in tap water, gentle brushing with Tween-20, a 1-minute exposure to 80% ethanol, a 20-minute treatment with freshly prepared HgCl2 at a concentration of 0.2%, and finally, rinsing four times for 4 minutes each with sterile distilled water. Sochacki and Orlikowska [117] in their studies aimed at improving the micropropagation protocol for two Narcissus cultivars, tested several pre-treatments of bulbs including the use of fungicides and hot water treatment (44.4°C for 3h) followed by a disinfection procedure with mercuric chloride and chloramine T. By these methods, they were successful in reducing the initial contamination to 14-17% and they showed that different genotypes could differently react to the same treatment.
Seeds of P. maritimum were used as explants in some studies and disinfection has been successful [59,112,114,118]. However disinfection of the flower bulbs are not quite easy. According to studies conducted by Yasemin S (unpublished data), Gladioulus corms and Fritillaria bulbs were washed under tap water then applied 70% ethanol and 2.5 % NaOCl and washed by sterile distilled water (3 times). All explants were contaminated. Because the mother plant was already not healthy. This could show the importance of the mother plant material selection. Some explant photos are shown in Figure 5.
Evaluating the negative impacts of sterilizing agents on plant tissues is crucial, with explant survival data serving as a key parameter. Rafiq, et al. [119] and Rather, et al. [120] indicate that combined chemicals, was more effective on disinfection achievement but the use of combined sterilant leads to a reduction in explant survival. Rafiq, et al. [119] experimented diverse sterilization approaches, incorporating sodium hypochlorite (1%) and mercuric chloride (0.1%) at varying durations (10 and 20 min), along with combinations involving 70% ethyl alcohol and carbendazim (200 ppm). Their chosen explants were bulb scale-base and bulb scale-tip from oriental hybrid Lilium cv. Ravenna. The most effective surface sterilization method entailed treating healthy bulb scales with carbendazim (200 ppm) for 30 min, followed by 0.1% mercuric chloride for 10 min, and then 70% ethyl alcohol for 30 s. It was obtained a good achievement with the highest culture asepsis (77.08%) and greater explant survival (86.12%). The survival rate was more than 90% whereas surface sterilization was less than 55% in the alone sterilant usage. Farooq, et al. [121] used bulb scales and young leaves to initiate the culture of Lilium LA hybrids ‘Indian Summerset’ and ‘Nashville’. A pre-treatment of both explants has been made by washing the tissues in a water solution containing Tween-20 and the fungicide carbendezim. Afterwards, a surface-sterilization of the explants was applied using carbendenzim, mercuric chloride and etyl-alcohol for different concentrations and duration. The highest disinfection success for bulb scales (Indian Summerset: 85.41%, Nashwille: 89.58%) was higher in the combinational sterilant. Chib, et al. [92] also detailed an optimized two-step surface sterilization method for Crocus sativus L. This involved separately employing 0.1% mercuric chloride and 4% sodium hypochlorite, leading to effective asepsis and a noteworthy 86% survival rate of explants. The disinfection of corms was successfully achieved through this process. Application of antioxidants, such as ascorbic acid, citric acid can effectively prevent browning caused by oxidation. Furthermore, some additive such as polyvinylpyrrolidone (PVP), charcoal, fungicide can also prevent browning of explants and endogenous contamination risks. Appleton, et al. [122] performed in vitro regeneration of the Hypoxis colchicifolia. When establishing in vitro cultures, they have experimented PVP, activated charcoal, ascorbic acid, citric acid to inhibit browning and benomyl solutions as fungicide to inhibit contamination. They have solved the browning problem with PVP and partially contamination problem with benomyl. However, these applications are not creating the same response in every plant or plant part. It should be analyzed and optimized for each plants. Devi, et al. [123] have used charcoal and ascorbic acid to reduce the phenolics exudates, but charcoal inhibited the tissue growth and ascorbic acid caused the somatic embyros death in saffron (Crocus sativus L.).
In general, young tissues and organs have a high regeneration capacity than the older ones. Apical and axillary buds from tubers or bulbs can be used to initiate the in vitro culture. Furthermore, adventitious buds generally are induced from bulb scale or flower stems. Twin scales are useful explants although the use of underground storage organs leads to addressing serious contamination problems during the initiation and the further multiplication phase. Flower stems has advantage with low endogenous contamination rates compared to other tissues [16,21,29]. Vegetative segments of plants often regenerate more easily in vitro than generative ones. Rafiq, et al. [119] used basal and tip bulb scales to micropropagate the oriental hybrid Lilium cv. Ravenna. They also found that the explant survival rate of basal scales was higher than the tip bulb scales. Lapiz-Culqui, et al. [124] and Patil, et al. [125] used the bulb scales to cultivate different Lilium cultivars. Youssef, et al. [126] used the leaf as explant in their study to increase Lilium bulb number and size. Ozel, et al. [127] used twin scale bulb explant to micropropagate the endemic Muscari muscarimi. Twin scales were also used by Santos, et al. [128] to propagate Narcissus asturiensis and Kukulczanka, et al. [129] to propagate Frittilaria melagris through the same system. Kumar, et al. [130] used the bulb scales of the critically endangered Frittillaria roylei for in vitro culture. Sevindik and Mendi [131], Taheri-Dehkordi, et al. [132] used the corms as explants to propagate the Crocus sativus L. Furthermore, Slimani, et al. [133], informed that the vegetative apices, apical and axillary buds, meristematic zone, segments leave, ovaries, protoplasts, corm, root are useful to induce the somatic embryogenesis of Crocus sativus L.
Success in this initiation stage can be influenced by environmental factors encompassing nutrient composition, plant growth regulators (PGRs), light exposure, temperature, atmospheric composition, and various culture methodologies [16]. Devi, et al. [123] reported that the dark conditions were effective for somatic embryos proliferation of the saffron.
5.3. Stage 2: Multiplication
The process of multiplication, which is a continuation of in vitro initiation, constitutes one of the most critical stages in micropropagation. As mentioned in previous stages, leaves, inflorescences, and bulbous structures (such as bulbs, corms, tubers, etc.) are utilized for multiplication [44,79]. The selection of the medium to be used (solid, liquid, semi-solid, or media type- MS, B5, WPM etc) is crucial for achieving successful responses, in addition to the use of different explants. The optimization of factors such as salt mixtures (essential micro-macro elements, carbon sources, vitamins), sugar derivatives, ratios, types, and concentrations of plant growth regulators, light, temperature, and inductive agents is necessary [69,79]. The concentration of the hormones (e.g., cytokinin) is critical, as it can simultaneously promote multiplication and development while causing adverse effects [134-137]. Each step implemented here will influence both multiplication and subsequent planting stages; hence, optimal requirements need to be determined. Unfortunately, due to the variability in results obtained for each species and even within varieties, specific optimization efforts are required for each plant. This situation poses one of the challenges of tissue culture. Additionally, high production costs add to these challenges [138]. The commercial viability of geophyte propagation through tissue culture has also been questioned [79].
To achieve effective results at this stage, understanding the plant's physiology and mimicking its natural requirements based on the choices made during this phase can be impactful. The selections made during this stage lead to different differentiations according to the meristematic zones that will be formed as a result of stimulations in explants (somatic tissues) [79]. In the regeneration stage, unipolar meristems give rise to shoots or roots, which is organogenesis. Bipolar meristems, on the other hand, lead to the development of a complete plant or microcorm, representing somatic embryogenesis [139].
Plant growth regulators can stimulate or inhibit the in vitro development of geophytes. Sochacki, et al. [140] aimed at evaluating the influence of growth regulators (PGRs) on in vitro shoots multiplication of Tulipa L 'Heart of Warsaw' and they yielded the best performance (9.14 shoots/clump) when MS medium was supplemented with 2iP 0.1 mg/L, NAA 0.1 mg/L, and mT 5.0 mg/L. Additionally, this study showed that the type of carbohydrate added to the nutrient solution had a significant effect on the shoot proliferation of the tulip cultivar; the highest multiplication of the shoots (number of shoots for one starting clump) was achieved with sucrose (34.33 shoots/per clump) and glucose (35.88 shoots/per clump) as carbohydrates. Lagram, et al. [116], used excised mother corm buds under different BAP, 2.4-D and NAA concentrations in MS medium. The highest bud sprouting (96.67%) and shoot growth (8.87 cm) were obtained from 1 mg/L BAP and 1 mg/L NAA. They also evaluated adventitious shoot formations of the explants. The highest adventitious shoot (80%) was obtained from 0.5 mg/L NAA and 2.75 mg/L BAP. Recently micropropagation via organogenesis has been in increasing trend. Succesful adventitious bud regenerations were obtained in Lilium, Narcissus, Sternbergia, Hippeastrum, Firttilaria, Muscari, Tulip, Iris, Lachenelia, Hyacinthus [75,77,106,108,127,141-152]. As seen in the Figure 6, organogenic structures were obtained from bulb scales in P. maritimum.
Slimani, et al. [133] explained the importance of somatic embyrogenesis for the Crocus sativus L.to get healthy corms propagation. MS, LS, B5 media were supplemented with the growth regulators BA, NAA, Kn, TDZ 2.4D tested at different concentrations. Ebrahimzadeh, et al. [153], obtained somatic embryos by using meristem explants in LS culture medium included 2 µM BA and 2 µM NAA in Crocus sativus L. Sheibani, et al. [154], used corm explants in MS medium included TDZ (0, 0.1, 0.25 and 0.5 mg/L), and they obtained somatic embryos from 0.5 mg/L TDZ. Marković, et al. [155] delved into the intricate world of somatic embryogenesis in F. meleagris, emphasizing the crucial role of plant growth regulators (PGRs) in this process. Their study illuminated that the medium without PGRs exhibited exceptional efficiency throughout the experiment, showcasing a robust somatic embryogenic response. Moreover, the medium with lower concentrations of PGRs also proved to be conducive to somatic embryogenesis. The pinnacle of morphogenetic success was achieved in a BAP/2,4-D-containing medium with the lowest PGR concentrations. This insight into optimal conditions for somatic embryogenesis opens avenues for enhanced propagation strategies, potentially revolutionizing large-scale production of F. meleagris. Kocak, et al. [156] investigated somatic embryogenesis potential in various explants (ovules, divided ovary parts, leaves, and petiole segments) from 15 different genotypes of the wild species Cyclamen persicum Mill. The explants were cultured on a medium with specific plant growth regulators to induce embryogenic callus. The study found significant variations in embryogenic potential among explants and genotypes. While petiole explants produced the most callus, ovary explants were most efficient in forming somatic embryos. On average across genotypes, petiole explants had a 34.3% callus formation rate, followed by ovary (30.16%), ovule (26.6%), and leaf (15.6%). The percentages of somatic embryos formed were highest in ovary explants (11.3%), followed by petiole (8.00%), leaf (4.16%), and ovule (2.83%) explants. Recently an efficient method for tulip regeneration via SE was developed [157,158]. Some reports on SE are also available by using leaf, petiole, ovary, anther, roots and aseptic seedling tissues as explants in cyclamen [156,159-164]. Aseptic seedlings tissues (cotyledons, petioles, tubers, and roots) have been also used as explants to initiate the SE culture in many medicinal and endemic geophytes (e.g. Crocus, Iris, Hypoxis,Colchicum etc.) [123,131,153,154,165-168]. Some embryo- like structures from P maritimum bulb explants are shown in Figure 7A, 7B, and 7C. Morever, embryogenic callus and regeneration in R. asiaticus were shown in Figure 7D, 7E. Thalamus derived callus in R. asiaticus L. calli have been initiated on MS medium containing the growth regulator 2,4-D and cytokinins (BA and kinetin) according to Beruto, et al. [169].
5.3. Stage 3: Bulb growth
Following the multiplication phase, in non-bulbous plants, the optimization of media plays a crucial role in enhancing the successful rooting of developed shoots. However, this phenomenon is not uniformly applicable to geophytes, where the acquisition of storage organs proves to be challenging and time intensive [140]. This phase in geophytes is directed towards the acquisition of shoots, primordia, and bulbs. In some plant species, the sequence involves the initial rooting of shoots followed by subsequent bulb formation, while in others, bulbs are generated directly [16]. The induction of bulb formation occurs through diverse mechanisms. Investigations into bulb formation have explored the influence of various sugar types and concentrations, plant growth regulators, light quality, temperature, distinct tissue culture conditions (solid, liquid medium, bioreactor utilization). Sochacki, et al. [140] conducted an investigation on the bulb formation of previously multiplied Tulipa L 'Heart of Warsaw' plantlets, examining various carbohydrate types, different phase media, and plant growth regulators. The highest bulb formation (28.00 bulbs) and the most mature bulbs (14.50 bulbs) were achieved in a two-phase (liquid) system with 1 mg/L PBZ and glucose. Pałka, et al. [75] utilized bulb scales of Lilium candidum as explants in an in vitro MS media and assessed the impact of different light qualities (spectral compositions of red (100%), blue (100%), and red and blue light (RB ratio 7:3), RB light was mixed in equal proportions (50%) with green (RBG), yellow (RBY), UV (RBUV), and far-red (RBfR) light, white LED (Wled), Fluorescent lamp light (Fl), and darkness (D)) on bulb formation without the use of plant growth regulators. The study resulted in a bulb formation rate of 79-100% across all explants. The best results in terms of bulb number were obtained from Fl (16.3), RBY (13.33), and RBfR (13.33), B (12.69), RB (12.63), and Wled (14.36) treatments. The lowest rate (3.00) was observed in plants kept in darkness. The largest bulb diameter (5.41 mm) and the highest photosynthetic pigment content were obtained from the RBG treatment. Darkness and red light induced etiolation. Additionally, soluble sugars in bulbs were stimulated by darkness and blue light. Lagram et al. [116] used different sucrose, IBA and NAA concentrations on corm production and root regeneration in ½ MS. Furthermore, they considered the photoperiod effect (16h/8h and dark conditions) on the parameters. They obtained the best results in ½ MS, 6% sucrose, 1 mg/L NAA, and dark conditions with 100% corm production, 7.9 g mini-corm weight, 93.8 % root formation, 14.9 root number per mini-corm. In the realm of bulb growth in F. meleagris, Marković et al. [155] unraveled key factors influencing the development of bulbs in vitro. Their investigation revealed that bulbing ability remained relatively high in the absence of PGRs, and for the most part, was unaffected by varying concentrations of the tested PGRs. Notably, the study identified that the lowest cytokinin concentration, in combination with low auxin, significantly enhanced bulb formation when the cultures were transferred to a higher temperature. This finding underscores the importance of temperature modulation and specific PGR combinations for optimizing bulb growth in F. meleagris. Understanding these dynamics can contribute to refining propagation methods and accelerating the scale-up of bulb production for this species. Azeri and Öztürk [170] investigated the most effective hormone treatment to induce and produce Lilium monodelphum M. Bieb, var. Armenum in tissue culture rapidly and efficiently. Optimal bud regeneration (11.67) was achieved in a medium containing 3.0 mg/L TDZ, 0.25 mg/L NAA, and 0.1 mg/L GA3. Additionally, the highest microbulb formation (15.83) was observed in a medium comprising 2.0 mg/L PAC, 0.2 mg/L NAA, and 0.1 mg/L GA3.

Figure 8.
Bulbs formation from seeds and callus derived from bulb scales in Pancratium maritimum.
In recent years, the use of bioreactor systems has shown an increase for healthy plant regeneration. Studies on the regeneration of bulbs, corms, rhizomes, microtubers, shoots, and subsequent rooting, as well as somatic embryogenesis, have been on the rise [171]. In their published article, Murthy, et al. [171] indicated the proliferation of Lilium hybrids and Allium sativum bulbs, Crocus sativus and Alocasia amazonica corms, microtubers of Solanum tuberosum varieties, and rhizomes of Cymbidium sinense using various bioreactor systems [74,172-184]. According to Murthy, et al. [171] the selection of an appropriate bioreactor system is crucial due to various factors (design, principle, inoculation density, aeration, temperature, light intensity, etc.) that influence the regeneration of propagules. To minimize losses of small bulblets and prevent dormancy, direct transplantation of plantlets is often preferred.
Rooting can be improved through various methods, such as adding auxin and/or activated charcoal (AC) to the culture medium, adjusting the auxin to cytokinin ratio, and using half-strength salts and sucrose. In the study by Azeri and Öztürk [170], microbulbs were subjected to rooting media with IBA (0.5 and 1.0 mg/L). Successful plant development, with root numbers of 6.4 and 5.9, respectively, was observed in media containing 0.5 and 1 mg/L IBA.
For successful acclimatization hardening is essential. This process enhances tolerance to moisture stress and prevents hyperhydricity [16]. In Rafiq, et al. [119] study on Oriental Lilium hybrid cv. Ravenna, varying combinations of IBA and NAA significantly influenced microshoot rooting behavior. IBA outperformed NAA, with the highest rooting (92.71%) and primary root characteristics observed in a medium supplemented with 1.50 mg/L IBA. Explants showed differences, with basal scale segments exhibiting the highest rooting (92.71%), percentage root number/shoot (10.02), length of primary roots (2.17 cm) while tip scale segments had the lowest (77.55%, 9.08, 1.65 cm). Notably, primary hardening influenced plantlet survival, with the highest ex vitro survival (98.96%) in plantlets from IBA (1.5 mg/L)-fortified media, particularly in basal scale segments. Rooted plantlets were hardened in media containing perlite and vermiculite (1:1). This suggests that IBA-treated cultures resulted in superior ex vitro survival, possibly due to enhanced rooting characteristics such as root number and length.
Direct bulblet induction in certain genotypes can provide several advantages, including the elimination of in vitro rooting, prevention of hyperhydricity, avoidance of the need for hardening, increased survival rates, and a shortened bulb production period. Conditions conducive to bulb formation involve high sucrose concentrations, the application of plant growth retardants, exposure to low temperatures, and the utilization of aged shoots. It's worth noting that in vitro formed bulblets may experience spontaneous dormancy, as reported by Kim and De Hertogh [16]. In the research conducted by Chib, et al. [92], they experimented with a high sucrose concentration combined with plant growth regulators (PGRs) to enhance efficiency over a 90-day timeframe. The most favorable outcomes were observed when using a combination of MS medium, TDZ, IAA, activated charcoal, and 4% sucrose, resulting in a notable 68% efficiency. Lower sucrose concentrations led to sluggish growth, while concentrations exceeding 4% resulted in cell death, as evidenced by the blackening of the callus. Different sucrose, BA and 2,4D concentrations in MS media affected the growing bulbs in in vitro conditions for P. maritimum [59].
5.3. Stage 4: Dormancy breaking
Geophytes produce new buds that enable vegetative propagation through their bulbs. After an active growth and flowering period in spring, senescence of aboveground tissues is followed by root senescence, leading the plant into a dormant phase with no visible organogenesis. To survive in unfavorable environmental conditions that are not conducive to their development and proliferation, these organs enter a state of dormancy. Dormancy is characterized by the inability to initiate growth from meristems under favorable conditions. In bulbs, dormancy can manifest in three different types: endodormancy (internal inability to grow), ecodormancy (environmental conditions), and paradormancy (apical dominance, hormonal status, metabolite-sugar levels) [185-187]. When evaluating the concepts of dormancy and true dormancy, it is important to note that true dormancy persists until dormancy is completely terminated, even if favorable environmental conditions are present. Unlike many other plants, geophytes do not exhibit true dormancy, as activities continue even during dormant phases. Most bulbous structures obtained through tissue culture display dormancy. During in vitro regeneration, bulbs and other storage organs such as tubers and corms typically undergo dormancy, similar to their behavior in natural conditions. The level of dormancy may vary depending on factors such as sucrose concentration, age of the bulb, and environmental conditions [16,104,188]. The growth and sprouting of geophytes in vitro are influenced by dormancy, bulb size, and maturity [189]. Therefore, an efficient in vitro protocol, including dormancy release, is crucial for the rapid, efficient, and valid commercial exploitation of all horticultural geophytes. To overcome this constraint, in some Fritillaria species, different temperature regimes were tested under in vitro conditions. Kizil and Khawar [146] treated F. persica seeds at 4°C for 75 days, 10°C for 15 days, and 4°C for 30 days in a MS medium with different plant growth regulators (BAP, IBA) and the study successfully developed a protocol for breaking seed dormancy. Optimal results were achieved with 80% dormancy break using 2.0 mg/L BAP + 1.0 mg/L IBA and 40% bulblet induction with 1.0 mg/L BAP + 1.0 mg/L IBA. Alternating temperatures of 4°C and 10°C for specific durations influenced seed germination and bulblet induction. A minimum incubation period of 30 days at 4°C, followed by 60 days at 10°C, was needed to break seed dormancy. Bulblet diameter increased on MS medium with 50 mg/L sucrose after 30 days at 4°C. Successful rooting of Fritillaria bulblets was achieved on MS medium with 0.5 mg/L NAA. Marković, et al. [155], carried out bulb scale culture with different plant growth regulators combinations for four weeks at 7°C in Frittilaria meleagris. They found that in the control medium (PGR-free medium), shoots per explants were higher and maximum at the end of the chilling. In the study conducted by Carasso and Mucciarelli [190], seeds of Fritillaria tubiformis Gren. & Godr were chilled in 4°C 30, 60 and 90 days in 1% agar medium. At the end of cold stratification, seeds again disinfected and immature zygotic embryos were rescued from seed coats. The highest number of somatic embryos occurred in zygotic embryos subjected to 30 days of cold stratification on MS2 (BA 2 mg/L and NAA 0.5 mg/L) medium. Successful conversion of somatic embryos into bulblets necessitated transfer to a maturation medium supplemented with 4% (w/v) sucrose. In the presence of IBA 1 mg/L, bulblets sprouted and developed roots, leading to the establishment of newly formed plants suitable for pot cultivation.
5.3. Stage 5: Planting
Transplanting shoots or bulblets to sterile rooting media is successful for various plant species. However, moving plantlets from in vitro to greenhouse conditions requires specific care. Initially, it is crucial to maintain high humidity and low light for a few days until the plantlets become autotrophic. Subsequently, they can be cultivated under lower humidity and higher light levels in the greenhouse [16]. In the study by Azeri and Öztürk [170], microbulbs were subjected to rooting media with IBA (0.5 and 1.0 mg/L). Successful plant development, with root lengths of 6.4 and 5.9, respectively, was observed in media containing 0.5 and 1 mg/L IBA. All plantlets obtained through tissue culture demonstrated survival when transferred to soil. Yasemin, et al. [59], transferred the P. maritimum plantlets into sand, sand:peat (1:1), and peat and the survival rates were more than 90%. Rafiq, et al. [119] transferred the rooted plantlets in media containing perlite and vermiculite (1:1). The survival rates were more than 80%. Frittiaria ruthenica was acclimatized in coconut fiber and sand (3:1) mixture in greenhouse. The survival rate was found as 72% [106]. In the study by İzgü, et al. [160], separated the plantlets from embryogenic-like structures (ELSs) and further germinated on plant growth regulator (PGR)-free culture initiation medium (CIM). Developed plantlets were transplanted to pots with peat and sand. Plantlet formations were found between 15% and 38 % in Cyclamen species. Subsequently, successful acclimatization in a greenhouse was achieved, with survival rates of 70%, 63%, 54%, and 25% for C. mirabile, C. pseudibericum, C. cilicium, and C. parviflorum, respectively.
6. Somaclonal Variation
In vitro isolation and culture of explants often lead to callus formation, influenced by species and a high auxin-to-cytokinin ratio. Continuous callus culture may result in genetic variation and loss of regeneration potential over generations. Callus cultures in geophytes have been used for plant regeneration, cell suspension production, and isolating somaclonal variants [16,191]. The evaluation of somaclonal variation is crucial for ensuring the success of in vitro propagation to achieve true-to-type clones. While somaclonal variations may pose challenges in clonal propagation, they can be advantageous in breeding programs [192]. Van Harmelen, et al. [193] determined somaclonal variations from callus derived from bulb scales of L. longliflorum. The callus was kept 3 years at 20°C in the dark and after this period, regeneration was detected; their findings showed mutations in the regenerated plants such as dwarf plants, malformation of the leaves, and male sterility. The use of molecular markers is one of the most effective strategies for monitoring somaclonal variations and nowadays ISSR markers have been used in many studies [194-196]. Memon, et al. [197], detected somaclonal variation among in vitro propagated cormels of gladiolus using RAPD and ISSR molecular markers. The observed variations had varying degrees from the mother cormels and were evident across different varieties of gladiolus. Asadi, et al. [198] found that indirect embryogenesis resulted in somaclonal variation, while direct embryogenesis produced uniform plants in Galanthus transcaucasicus. The researchers utilized twin-scaling explants and exposed them to varying concentrations of naphthaleneacetic acid (NAA) and benzyladenine (BA), including 0.5, 1, 2, 3, 4, 6, 8, and 10 mg/L for NAA, and 0.5, 1, 2, 3, and 4 mg/L for BA. Subsequently, bulblets were transferred to a medium containing 0.5, 1, 2, 3, and 4 mg/L NAA, along with 0.5, 1, 2, 3, and 4 mg/L BA. The most pronounced callus formation occurred at either 4 mg/L NAA or 8 mg/L NAA in combination with 1 mg/L BA. Optimal regeneration of bulblets was observed at 6 mg/L NAA and 2 mg/L BA, yielding the highest number of regenerated bulblets. Higher NAA concentrations increased somaclonal variation, with the highest observed at the maximum NAA concentration. ISSR analysis showed no somaclonal variation in plants from low BA and NAA concentrations, but significant variation occurred at high NAA concentrations. Kritskaya, et al. [134] investigated the somaclonal variations in Tulipa suaveolens with ISSR markers. They obtained bulblets through direct organogenesis, showing morphological similarities with intact plants. However, ISSR analysis detected a notable level of somaclonal variability, ranging from 13.9% to 15.8%.
7. Conclusions and future perspectives
Tissue culture techniques can reply to the requirements of the present-day floriculture asking for novelties and good quality of the starting plant material to introduce in the productive flow. The tissue culture found a great application in the flower bulb industry and many new methods of micropropagation have been developed over these last decades. Many research papers aimed at describing micropropagation protocols for specific species but, as deduced from our review, there is a need of a multifunctional approach able to face the important challenges which are still present. Beside the insights on biochemistry and physiology, the molecular biology could be of great help in advancing the establishment of valuable tissue culture protocols for new genera/species.
Like in other plants, gen editing studies are conducted in geophytes as well. Particularly, in geophytes of economic importance with high medicinal and aromatic values, we can come across genetic editing studies aimed at increasing the content of important compounds in storage organs or plant parts. However, Kamo, et al. [191] emphasized that the application of modern biotechnological approaches to geophytes, facing challenges in developing transgenic varieties. Issues include the lack of efficient transformation systems and difficulties in gene integration into specific genome regions. Ornamental geophytes, with large genomes, pose research complexities. Despite solvable challenges, uncertainties exist regarding sufficient funding, given their relatively small production value compared to major crops. Nevertheless, biotechnological research on geophytes contributes to expanding our understanding of plant biology. Indeed, genome editing technology is fastly improving and precisely genome editing studies like CRISPR are importantly increasing. This approach is touching also geophytes even though the geophytes have a lot challenges. For example, Crocus sativus contains numerous aromatic and volatile compounds. Genetic editing has been targeted to enhance the content of crocin, picrocrocin, and safranal apocarotenoids in this plant, as they are responsible for color, bitter taste, and aroma. Preliminary CRISPR studies have been conducted for this purpose [92]. Using the CRISPR method, it is possible to enhance and replicate many important medicinal and aromatical contents of geophytes, overcome difficulties in their multiplication, and increase tolerance to biotic and abiotic stresses.
References
- Benschop, M.; Kamenetsky, R.; Le Nard, M.; Okubo, H.; De Hertogh, A. 1 The global flower bulb industry: Production, utilization, research. Horticultural reviews 2010, 36, 1–115. [Google Scholar]
- Correvon, H.; Massé, H. Les iris dans les jardins; Aux jardins Correvon: 1907.
- Reynolds, M.; Meachem, W. garden bulbs of spring. 1967.
- Doerflinger, F. The bulb book. (No Title) 1973.
- Margaris, N. Flowers in Greek mythology. In Proceedings of the IV International Symposium on New Floricultural Crops 541; 1999; pp. 23–29. [Google Scholar]
- Janick, J.; Kamenetsky, R.; Puttaswamy, S.H. Horticulture of the Taj Mahal: Gardens of the imagination. Chronica Horticulturae 2010, 50, 30–33. [Google Scholar]
- Bryan, J.E. Bulbs. (No Title) 1989.
- De Hertogh, A.; Schepeen, J.; Kamenetsky, R.; Le Nard, M.; Okubo, H. The Globalization of Flower Bulb Industry. Ornamental geophytes: from basic science to sustainable production 2013, 1. [Google Scholar]
- Pavord, A. The Tulip: The Story ofa Flower That Has Made Men Mad. 1999.
- Marasek-Ciolakowska, A.; Sochacki, D.; Marciniak, P. Breeding Aspects of Selected Ornamental Bulbous Crops. Agronomy 2021, 11. [Google Scholar] [CrossRef]
- Orlikowska, T.; Podwyszyńska, M.; Marasek-Ciołakowska, A.; Sochacki, D.; Szymański, R. Tulip. Ornamental crops 2018, 769–802. [Google Scholar]
- Sarac, Y.I.B., Ahmet; Deligoz, Ilyas. Lale Gece Publishing: 2021.
- Rees, A. The physiology of ornamental bulbous plants. The Botanical Review 1966, 32, 1–23. [Google Scholar] [CrossRef]
- Bailey, L.H. Manual of cultivated plants. Manual of Cultivated Plants. 1949. [Google Scholar]
- Gabellini, S.; Scaramuzzi, S. Evolving consumption trends, marketing strategies, and governance settings in ornamental horticulture: A grey literature review. Horticulturae 2022, 8, 234. [Google Scholar] [CrossRef]
- Kim, K.-W.; De Hertogh, A. Flowering Bulbs (Geophytes). Horticultural Reviews, Volume 18 1997, 18, 87. [Google Scholar]
- Pizano, M. Innovation and sustainability in South American floriculture. In Proceedings of the XXXI International Horticultural Congress (IHC2022): International Symposium on Innovations in Ornamentals: from Breeding to 1368; 2022; pp. 77–84. [Google Scholar]
- Janowska, B.; Andrzejak, R. Plant growth regulators for the cultivation and vase life of geophyte flowers and leaves. Agriculture 2023, 13, 855. [Google Scholar] [CrossRef]
- Kocak, M.; Sevindik, B.; Izgu, T.; Tutuncu, M.; Mendi, Y.Y. Synthetic seed production of flower bulbs. Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects 2019, 283–299. [Google Scholar]
- Seyidoglu N., Z. M., Ayasligil Y. Position and application areas of geophytes within landscape design. African Journal of Agricultural Research 2009, 4, 1351–1357. [Google Scholar]
- Ziv, M.; Lilien-Kipnis, H. Bud regeneration from inflorescence explants for rapid propagation of geophytes in vitro. Plant Cell Reports 2000, 19, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Kamenetsky, R.; Okubo, H. Ornamental geophytes: from basic science to sustainable production; CRC press: 2012.
- Marasek-Ciolakowska, A.; Sochacki, D.; Marciniak, P. Breeding aspects of selected ornamental bulbous crops. Agronomy 2021, 11, 1709. [Google Scholar] [CrossRef]
- Rees, A.R. Ornamental bulbs, corms and tubers; CAB international: 1992.
- Leeggangers, H.C.F.; Moreno-Pachon, N.; Gude, H.; Immink, R.H. Transfer of knowledge about flowering and vegetative propagation from model species to bulbous plants. Int. J. Dev. Biol. 2013, 57, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Kamenetsky, R. Biodiversity of Geophytes: Phytogeography, Morphology, and Survival Strategies. In Ornamental geophytes: from basic science to sustainable production, Kamenetsky, R., Okubo, H., Eds.; CRC press: 2012; pp. 57-76.
- Dafni, A.; Cohen, D.; Noy-Mier, I. Life-cycle variation in geophytes. Annals of the Missouri Botanical Garden 1981, 652–660. [Google Scholar] [CrossRef]
- Woodhead, T.W. Ecology of woodland plants in the neighbourhood of Huddersfield. Botanical Journal of the Linnean Society 1906, 37, 333–406. [Google Scholar] [CrossRef]
- De Klerk, G. Micropropagation of bulbous crops: technology and present state. Floriculture and Ornamental Biotechnology 2012, 6, 1–8. [Google Scholar]
- Bach, A.; Sochacki, D. Ornamental Geophytes from Basic Science to Sustainable Production; Phsiology and Menagement Systems. Kamenetsky, R and Okubo, H. 2013.
- Ulus, A.; Seyidoğlu, N. Bazi Doğal Geofitlerin Doku Kültürü İle Üretimi. Journal of the Faculty of Forestry Istanbul University 2006, 56, 71–80. [Google Scholar]
- Irish, V.F. The Arabidopsis petal: a model for plant organogenesis. Trends in Plant Science 2008, 13, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Langens-Gerrits, M.M.; De Klerk, G.-J.M. Micropropagation of flower bulbs: Lily and Narcissus. Plant Cell Culture Protocols 1999, 141–147. [Google Scholar]
- Luna, T. Vegetative propagation. Nursery manual for native plants: A guide for tribal nurseries 2009, 1, 153–175. [Google Scholar]
- Hartmann, H.T.; Kester, D.E. Plant propagation: principles and practice. Soil Science 1963, 95, 89. [Google Scholar] [CrossRef]
- Debergh, P.; Maene, L. A scheme for commercial propagation of ornamental plants by tissue culture. Scientia horticulturae 1981, 14, 335–345. [Google Scholar] [CrossRef]
- Debergh, P.C.; Read, P. Micropropagation. In Micropropagation: technology and application; Springer: 1991; pp. 1-13.
- Ozzambak, M.E.; Senel, U.; Gulden;, H.; Zeybekoglu, E.; Baser, S. Mikroçoğaltım teknikleri. In Süs Bitkileri Islahı (Klasik ve Biyoteknolojik Yöntemler), Yalcin Mendi, Y.K.S., Ed.; Gece Publishing: 2021; Volume 341-360.
- Podwyszyńska, M.; Orlikowska, T.; Trojak-Goluch, A.; Wojtania, A. Application and Improvement of In Vitro Culture Systems for Commercial Production of Ornamental, Fruit, and Industrial Plants in Poland. Acta Societatis Botanicorum Poloniae 2022, 91. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Jain, S.M. Cellular, molecular, and physiological aspects of in vitro plant regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New insights into tissue culture plant-regeneration mechanisms. Frontiers in plant science 2022, 13, 926752. [Google Scholar] [CrossRef] [PubMed]
- Von Arnold, S. Somatic embryogenesis. Plant propagation by tissue culture. Volume 1: the background 2008, 335-354.
- Williams, E.; Maheswaran, G. Somatic embryogenesis: factors influencing coordinated behaviour of cells as an embryogenic group. Annals of botany 1986, 57, 443–462. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X. Regulation of Somatic Embryogenesis in Higher Plants. Critical Reviews in Plant Sciences 2010, 29, 36–57. [Google Scholar] [CrossRef]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New Insights Into Tissue Culture Plant-Regeneration Mechanisms. Front Plant Sci 2022, 13, 926752. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.L. Somatic embryogenesis: a model for early development in higher plants. The plant cell 1993, 5, 1411. [Google Scholar] [CrossRef] [PubMed]
- Sharp, W.; Sondahl, M.; Caldas, L.; Maraffa, S. The physiology of in vitro asexual embryogenesis. Horticultural reviews 1980, 2, 268–310. [Google Scholar]
- Sevindik, B.; Tutuncu, M.; P;, E.C.; Y, Y.M. Somatik embryogenesis. In Süs Bitkileri Islahı (Klasik ve Biyoteknolojik Yöntemler), Yalcin Mendi, Y.K.S., Ed.; Gece Publishing: 2021; pp. 413-440.
- Von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant cell, Tissue and Organ culture 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Gahan, P.; George, E. Adventitious regeneration. Plant propagation by tissue culture. Volume 1: the background 2008, 355-401.
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Plant propagation by tissue culture 3rd Edition. The Netherland, The Back Ground Springer 2008.
- Sugiyama, M. Organogenesis in vitro. Current opinion in plant biology 1999, 2, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Tutuncu, M.; Basar;, S.; Yalcin Mendi, N.Y. Organogenesis. In Süs Bitkileri Islahı (Klasik ve Biyoteknolojik Yöntemler), Yalcin Mendi, Y.K.S., Ed.; Gece Publishing: 2021; pp. 361-392.
- Hicks, G.S. Patterns of organ development in plant tissue culture and the problem of organ determination. The Botanical Review 1980, 46, 1–23. [Google Scholar] [CrossRef]
- Schwarz, O.J.; Beaty, R.M. Organogenesis. In Plant Tissue Culture Concepts and Laboratory Exercises; Routledge: 2018; pp. 125-138.
- Preece, J. Stock plant physiological factors affecting growth and morphogenesis. Plant propagation by tissue culture. Volume 1: the background 2008, 403-422.
- Van Aartrijk, J.; Van der Linde, P. In vitro propagation of flower-bulb crops. In Proceedings of the Tissue culture as a plant production system for horticultural crops: Conference on Tissue Culture as a Plant Production System for Horticultural Crops, Beltsville, MD, October 20–23, 1985, 1986; pp. 317-331.
- Yasemin, S.; Koksal, N.; Buyukalaca, S. Indirect organogenesis and in vitro bulb formation of Pancratium maritimum. Plant Cell, Tissue and Organ Culture (PCTOC) 2023, 1-15. [Google Scholar] [CrossRef]
- Chu, C. Establishment of an efficient medium for another culture of rice through comparative experiments on the nitrogen sources. Scientia Sinica 1975, 18, 223–231. [Google Scholar]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. 1980, 30, 421–427. [Google Scholar]
- Gamborg, O.L.c.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Experimental cell research 1968, 50, 151–158. [Google Scholar] [CrossRef]
- George, E.; de Klerk, G. The components of plant tissue culture media I: Macro-and micro-nutrients. Plant propagation by tissue culture. Volume 1: the background 2008, 65-113.
- Thorpe, T.; Stasolla, C.; Yeung, E.; de Klerk, G.; Roberts, A.; George, E. The components of plant tissue culture media II: Organic additions, osmotic and pH effects, and support systems. Plant propagation by tissue culture. Volume 1: the background 2008, 115-173.
- Mehbub, H.; Akter, A.; Akter, M.A.; Mandal, M.S.H.; Hoque, M.A.; Tuleja, M.; Mehraj, H. Tissue Culture in Ornamentals: Cultivation Factors, Propagation Techniques, and Its Application. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Podwyszyńska, M.; Kosson, R.; Treder, J. Polyamines and methyl jasmonate in bulb formation of in vitro propagated tulips. Plant Cell, Tissue and Organ Culture (PCTOC) 2015, 123, 591–605. [Google Scholar] [CrossRef]
- Beruto, M.; Curir, P. Effects of agar and gel characteristics on micropropagation: Ranunculus asiaticus, a case study. Floriculture, ornamental and plant biotechnology 2006, 277-284. [Google Scholar]
- George, E.; Davies, W. Effects of the physical environment. Plant propagation by tissue culture. Volume 1: the background 2008, 423-464.
- Hussey, G. The application of tissue culture to the vegetative propagation of plants. Science Progress (1933-) 1978, 185-208. [Google Scholar]
- Bornwaßer, T.; Tantau, H.-J. Evaluation of LED lighting systems in in vitro cultures. In Proceedings of the VII International Symposium on Light in Horticultural Systems 956; 2012; pp. 555–562. [Google Scholar]
- Goeden, K.; Tong, C. The effects of blue light on potato tuberisation. In Proceedings of the Abstr, 543. Poster presented for the annual meeting of the American Society Plant Biologists, 2003.
- Bach, A.; Świderski, A. The effect of light quality on organogenesis of Hyacinthus orientalis L. in vitro. Acta Biologica Cracoviensia. Series Botanica 2000, 42, 115–120. [Google Scholar]
- Bakhshaie, M.; Khosravi, S.; Azadi, P.; Bagheri, H.; van Tuyl, J.M. Biotechnological advances in Lilium. Plant cell reports 2016, 35, 1799–1826. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.; Chakrabarty, D.; Paek, K. Growth and uptake of sucrose and mineral ions by bulblets of Lilium oriental hybrid ‘Casablanca’during bioreactor culture. The Journal of Horticultural Science and Biotechnology 2002, 77, 253–257. [Google Scholar] [CrossRef]
- Pałka, P.; Cioć, M.; Hura, K.; Szewczyk-Taranek, B.; Pawłowska, B. Adventitious organogenesis and phytochemical composition of Madonna lily (Lilium candidum L.) in vitro modeled by different light quality. Plant Cell, Tissue and Organ Culture (PCTOC) 2023, 152, 99–114. [Google Scholar] [CrossRef]
- Ascough, G.D.; van Staden, J.; Erwin, J.E. In Vitro Storage Organ Formation of Ornamental Geophytes. In Horticultural Reviews; 2008; pp. 417-445.
- Van Aartrijk, J.; Blom-Barnhoorn, G.J. Adventitious Bud Formation from Bulb-scale Explants of Lilium speciosum Thunb. in vitro Interacting effects of NAA, TIBA, wounding, and temperature. J Plant Physiol 1984, 116, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Robb, S.M. The Culture of Excised Tissue 3 Lilium speciosum Thun. Journal of Experimental Botany 1957, 8, 348–352. [Google Scholar] [CrossRef]
- Ziv, M. THE CONTRIBUTION OF BIOTECHNOLOGY TO BREEDING, PROPAGATION AND DISEASE RESISTANCE IN GEOPHYTES. In Proceedings of the VII International Symposium on Flowerbulbs, Herzliya, Israel 1 December 1997 1997.
- Dhooghe, E.; Reheul, D.; Van Labeke, M.-C. Overcoming Pre-Fertilization Barriers in Intertribal Crosses between Anemone coronaria L. and Ranunculus asiaticus L. Horticulturae 2021, 7, 529. [Google Scholar] [CrossRef]
- Sevindik, B.; İzgü, T.; Tütüncü, M.; Çürük, P.; Sarı, N.; Mendi, Y.Y. Double-haploid plant production through anther and ovule culture of wild Cyclamen persicum Mill. and Melody F1 cyclamen cultivar. In Vitro Cellular & Developmental Biology - Plant 2023. [Google Scholar] [CrossRef]
- Sevindik, B.; İzgü, T.; Tütüncü, M.; Çürük, P.; Söğüt, Z.; Mendİ, Y.Y. Effects of different plant growth regulators on ovule culture of Turkish Cyclamen persicum Mill. and commercial variety" Melody F1". alatarım 2017, 16, 1–9. [Google Scholar]
- Tütüncü, M.; Mendi, Y.Y. Effect of pollination with gamma irradiated pollen on in vitro regeneration of ovule culture in Cyclamen. Turkish Journal of Agriculture-Food Science and Technology 2022, 10, 2415–2420. [Google Scholar] [CrossRef]
- Van Creij, M.; Kerckhoffs, D.; De Bruijn, S.; Vreugdenhil, D.; Van Tuyl, J. The effect of medium composition on ovary-slice culture and ovule culture in intraspecific Tulipa gesneriana crosses. Plant cell, tissue and organ culture 2000, 60, 61–67. [Google Scholar] [CrossRef]
- Van Tuyl, J.; Van Creij, M.; Van Dien, M. In vitro pollination and ovary culture as a breeding tool in wide hybridization of Lilium and Nerine. In Proceedings of the VI International Symposium on Flower Bulbs 325; 1992; pp. 461–466. [Google Scholar]
- Van Tuyl, J.M.; Van Diën, M.P.; Van Creij, M.G.M.; Van Kleinwee, T.C.M.; Franken, J.; Bino, R.J. Application of in vitro pollination, ovary culture, ovule culture and embryo rescue for overcoming incongruity barriers in interspecific Lilium crosses. Plant Science 1991, 74, 115–126. [Google Scholar] [CrossRef]
- Wang, J.; Huang, L.; Bao, M.-z.; Liu, G.-f. Production of interspecific hybrids between Lilium longiflorum and L. lophophorum var. linearifolium via ovule culture at early stage. Euphytica 2009, 167, 45–55. [Google Scholar] [CrossRef]
- Tütüncü, M.; Mendi, Y. Evaluation of pollen tube growth and fertilization via histological analysis in Cyclamen persicum. In Proceedings of the XXVI International Eucarpia Symposium Section Ornamentals: Editing Novelty 1283; 2020; pp. 21–26. [Google Scholar]
- Karamian, R.; Ebrahimzadeh, H. Plantlet regeneration from protoplast-derived embryogenic calli of Crocus cancellatus. Plant Cell, Tissue and Organ Culture 2001, 65, 115–121. [Google Scholar] [CrossRef]
- Koetle, M.; Finnie, J.; Balázs, E.; Van Staden, J. A review on factors affecting the Agrobacterium-mediated genetic transformation in ornamental monocotyledonous geophytes. South African Journal of Botany 2015, 98, 37–44. [Google Scholar] [CrossRef]
- Babu, P.; Chawla, H. In vitro regeneration and Agrobacterium mediated transformation in gladiolus. The Journal of Horticultural Science and Biotechnology 2000, 75, 400–404. [Google Scholar] [CrossRef]
- Chib, S.; Thangaraj, A.; Kaul, S.; Dhar, M.K.; Kaul, T. Development of a system for efficient callus production, somatic embryogenesis and gene editing using CRISPR/Cas9 in Saffron (Crocus sativus L.). Plant methods 2020, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Meredith, C.P. Agrobacterium-mediated transformation of Lilium. In Proceedings of the VI International Symposium on Flower Bulbs 325; 1992; pp. 611–618. [Google Scholar]
- Hoshi, Y.; Kondo, M.; Mori, S.; Adachi, Y.; Nakano, M.; Kobayashi, H. Production of transgenic lily plants by Agrobacterium-mediated transformation. Plant Cell Reports 2004, 22, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Kamo, K.; Jordan, R.; Guaragna, M.A.; Hsu, H.-t.; Ueng, P. Resistance to Cucumber mosaic virus in Gladiolus plants transformed with either a defective replicase or coat protein subgroup II gene from Cucumber mosaic virus. Plant cell reports 2010, 29, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Hasegawa, H.; Suzuki, M. Transformation and regeneration of garlic (Allium sativum L.) by Agrobacterium-mediated gene transfer. Plant cell reports 2000, 19, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Zou, Q.; Guo, D.; Zhuang, X.; Yu, X.; Xiang, X.; Cao, J. Agrobacterium tumefaciens-mediated transformation of Narcissus tazzeta var. chinensis. Plant cell reports 2007, 26, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Popowich, E.; Firsov, A.; Mitiouchkina, T.; Filipenya, V.; Dolgov, S.; Reshetnikov, V. Agrobacterium-mediated transformation of Hyacinthus orientalis with thaumatin II gene to control fungal diseases. Plant cell, tissue and organ culture 2007, 90, 237–244. [Google Scholar] [CrossRef]
- Suzuki, S.; Supaibulwatana, K.; Mii, M.; Nakano, M. Production of transgenic plants of the Liliaceous ornamental plant Agapanthus praecox ssp. orientalis (Leighton) Leighton via Agrobacterium-mediated transformation of embryogenic calli. Plant Science 2001, 161, 89–97. [Google Scholar] [CrossRef]
- Wilmink, A.; Van de Ven, B.; Dons, J. Expression of the GUS-gene in the monocot tulip after introduction by particle bombardment and Agrobacterium. Plant Cell Reports 1992, 11, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Amaki, W.; Shinohara, Y.; Hayata, Y.; Sano, H.; Suzuki, Y. Effects of bulb desiccation and storage on the in vitro propagation of hyacinth. Scientia horticulturae 1984, 23, 353–360. [Google Scholar] [CrossRef]
- Gavinlertvatana, P.; Read, P.E.; Wilkins, H.; Heins, R. Influence of Photoperiod and Daminozide Stock Plant Pretreatments on Ethylene and CO2 Levels and Callus Formation from Dahlia Leaf Segment Cultures1. Journal of the American Society for Horticultural Science 1979, 104, 849–852. [Google Scholar] [CrossRef]
- Hosoki, T.; Sagawa, Y. Clonal Propagation of Ginger (Zingiber officinale Roscoe) through Tissue Culture1. HortScience 1977, 12, 451–452. [Google Scholar] [CrossRef]
- Stimart, D.P.; Ascher, P.D. Developmental Responses of Lilium longiflorum Bulblets to Constant or Alternating Temperatures in Vitro1. Journal of the American Society for Horticultural Science 1981, 106, 450–454. [Google Scholar] [CrossRef]
- Podwyszyńska, M.; Sochacki, D. Micropropagation of tulip: production of virus-free stock plants. Protocols for in vitro propagation of ornamental plants 2010, 243–256. [Google Scholar]
- Muraseva, D.S.; Novikova, T.I. Efficient protocol for in vitro propagation from bulb scale explants of Fritillaria ruthenica Wikstr. (Liliaceae), a rare ornamental species. Rendiconti Lincei. Scienze Fisiche e Naturali 2018, 29, 491–497. [Google Scholar] [CrossRef]
- Kumar, V.; Moyo, M.; Van Staden, J. Enhancing plant regeneration of Lachenalia viridiflora, a critically endangered ornamental geophyte with high floricultural potential. Scientia Horticulturae 2016, 211, 263–268. [Google Scholar] [CrossRef]
- Mirici, S.; Parmaksız, İ.; Özcan, S.; Sancak, C.; Uranbey, S.; Sarıhan, E.O.; Gümüşcü, A.; Gürbüz, B.; Arslan, N. Efficient in vitro bulblet regeneration from immature embryos of endangered Sternbergia fischeriana. Plant Cell, Tissue and Organ Culture 2005, 80, 239–246. [Google Scholar] [CrossRef]
- Buckley, P.M.; Reed, B.M. 045 ANTIBIOTIC SUSCEPTIBILITY OF PLANT-ASSOCIATED BACTERIA. HortScience 1994, 29, 434c–434. [Google Scholar] [CrossRef]
- George, E.F. Plant propagation by tissue culture part 1. The technology 1993. [Google Scholar]
- Silva, J.A.T.d.; Winarto, B.; Dobránszki, J.; Zeng, S. Disinfection procedures for propagation of. Folia Horticulturae 2015, 27, 3–14. [Google Scholar] [CrossRef]
- Yasemin, S.; Köksal, N.; Büyükalaca, S. Effects of disinfection conditions and culture media on in vitro germination of sea daffodil (Pancratium maritimum). J. BIOL. ENVIRON. SCI. 2018, 34, 13–22. [Google Scholar]
- Barrett, C.; Cassells, A.C. An evaluation of antibiotics for the elimination of Xanthomonas campestris pv. pelargonii (Brown) from Pelargonium x domesticum cv.‘Grand Slam’explants in vitro. Plant Cell, Tissue and Organ Culture 1994, 36, 169–175. [Google Scholar] [CrossRef]
- Alexopoulos, A.A.; Mavrommati, E.; Kartsonas, E.; Petropoulos, S.A. Effect of Temperature and Sucrose on In Vitro Seed Germination and Bulblet Production of Pancratium maritimum L. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Nikopoulos, D.; Alexopoulos, A.A. In vitro propagation of an endangered medicinal plant: Pancratium maritimum L. Journal of food, agriculture & environment 2008. [Google Scholar]
- Lagram, K.; El Merzougui, S.; Boudadi, I.; Ben El Caid, M.; El Boullani, R.; El Mousadik, A.; Serghini, M.A. In vitro shoot formation and enrooted mini-corm production by direct organogenesis in saffron (crocus sativus L.). Vegetos 2023. [Google Scholar] [CrossRef]
- Sochacki, D.; Orlikowska, T. The obtaining of narcissus plants free from potyviruses via adventitious shoot regeneration in vitro from infected bulbs. Scientia horticulturae 2005, 103, 219–225. [Google Scholar] [CrossRef]
- Redhwan, A.; Acemi, A.; Özen, F. Effects of plant growth regulators on in vitro seed germination, organ development and callogenesis in Pancratium maritimum L. Plant Cell, Tissue and Organ Culture (PCTOC) 2023, 1-14. [Google Scholar] [CrossRef]
- Rafiq, S.; Rather, Z.; Bhat, R.A.; Nazki, I.; Al-Harbi, M.S.; Banday, N.; Farooq, I.; Samra, B.N.; Khan, M.; Ahmed, A.F. Standardization of in vitro micropropagation procedure of Oriental Lilium Hybrid Cv.‘Ravenna’. Saudi journal of biological sciences 2021, 28, 7581–7587. [Google Scholar] [CrossRef] [PubMed]
- Rather, Z.; Nazki, I.; Qadri, Z.; Mir, M.; Bhat, K.; Hussain, G. In vitro propagation of herbaceous peony (Paeonia lactiflora Pall.) cv. Sara Bernhardt using shoot tips. Indian Journal of Horticulture 2014, 71, 385–389. [Google Scholar]
- Farooq, I.; Banday, N.; Nazki, I.; Malik, A.A.; Khan, F.; Mir, M.; Yaseen, T. In vitro sterilization of Lilium LA Hybrids “Indian Summerset” and “Nashville” as influenced by different sterilant combinations. The Pharma Innovation Journal 2022, 11, 1795–1798. [Google Scholar]
- Appleton, M.; Ascough, G.; Van Staden, J. In vitro regeneration of Hypoxis colchicifolia plantlets. South African Journal of Botany 2012, 80, 25–35. [Google Scholar] [CrossRef]
- Devi, K.; Sharma, M.; Ahuja, P. Direct somatic embryogenesis with high frequency plantlet regeneration and successive cormlet production in saffron (Crocus sativus L.). South African Journal of Botany 2014, 93, 207–216. [Google Scholar] [CrossRef]
- Lapiz-Culqui, Y.K.; Meléndez-Mori, J.B.; Mállap-Detquizán, G.; Tejada-Alvarado, J.J.; Vilca-Valqui, N.C.; Huaman-Human, E.; Oliva, M.; Goñas, M. In Vitro Bulbification of Five Lily Varieties: An Effective Method to Produce Quality Seeds and Flowers. International Journal of Agronomy 2022, 2022. [Google Scholar] [CrossRef]
- Patil, A.M.; Gunjal, P.P.; Das, S. In vitro micropropagation of Lilium candidum bulb by application of multiple hormone concentrations using plant tissue culture technique. International Journal for Research in Applied Sciences and Biotechnology 2021, 8, 244–253. [Google Scholar] [CrossRef]
- Youssef, N.M.; Shaaban, S.A.; Ghareeb, Z.F.; Taha, L.S. In vitro bulb formation of direct and indirect regeneration of Lilium orientalis cv.“Starfighter” plants. Bulletin of the National Research Centre 2019, 43, 1–9. [Google Scholar] [CrossRef]
- Ozel, C.A.; Khawar, K.M.; Unal, F. Factors affecting efficient in vitro micropropagation of Muscari muscarimi Medikus using twin bulb scale. Saudi Journal of Biological Sciences 2015, 22, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Fidalgo, F.; Santos, I.; Salema, R. In vitro bulb formation of Narcissus asturiensis, a threatened species of the Amaryllidaceae. The Journal of Horticultural Science and Biotechnology 2002, 77, 149–152. [Google Scholar] [CrossRef]
- Kukulczanka, K.; Kromer, K.; Czastka, B. Propagation of Fritillaria meleagris L. through tissue culture. In Proceedings of the III International Symposium on Growth Regulators in Ornamental Horticulture 251, 1988; pp. 147-154.
- Kumar, P.; Partap, M.; Ashrita; Rana, D.; Kumar, P.; Warghat, A.R. Metabolite and expression profiling of steroidal alkaloids in wild tissues compared to bulb derived in vitro cultures of Fritillaria roylei – High value critically endangered Himalayan medicinal herb. Industrial Crops and Products 2020, 145. [CrossRef]
- Sevindik, B.; Mendi, Y.Y. Somatic embryogenesis in Crocus sativus L. In Vitro Embryogenesis in Higher Plants 2016, 351–357. [Google Scholar]
- Taheri-Dehkordi, A.; Naderi, R.; Martinelli, F.; Salami, S.A. A robust workflow for indirect somatic embryogenesis and cormlet production in saffron (Crocus sativus L.) and its wild allies; C. caspius and C. speciosus. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Slimani, C.; El Goumi, Y.; Chaimae, R.; El Ghadraoui, L.; Benjelloun, M.; Lazraq, A. Micropropagation and potential of bioactive compounds of saffron (Crocus sativus L.) for nutrition and health. Notulae Scientia Biologicae 2022, 14, 11278–11278. [Google Scholar] [CrossRef]
- Kritskaya, T.; Kashin, A.; Kasatkin, M.Y. Micropropagation and Somaclonal Variation of Tulipa suaveolens (Liliaceae) in vitro. Russian Journal of Developmental Biology 2019, 50, 209–215. [Google Scholar] [CrossRef]
- Pierik, R.; Sprenkels, P.; Van Der Harst, B.; Van Der Meys, Q. Seed germination and further development of plantlets of Paphiopedilum ciliolare Pfitz. in vitro. Scientia Horticulturae 1988, 34, 139–153. [Google Scholar] [CrossRef]
- Tubić, L.; Savić, J.; Mitić, N.; Milojević, J.; Janošević, D.; Budimir, S.; Zdravković-Korać, S. Cytokinins differentially affect regeneration, plant growth and antioxidative enzymes activity in chive (Allium schoenoprasum L.). Plant Cell, Tissue and Organ Culture (PCTOC) 2015, 124, 1–14. [Google Scholar] [CrossRef]
- Ziv, M. Morphogenesis of gladiolus buds in bioreactors—implication for scaled-up propagation of geophytes. In Proceedings of the Progress in Plant Cellular and Molecular Biology: Proceedings of the VIIth International Congress on Plant Tissue and Cell Culture, Amsterdam, The Netherlands, 24–29 June 1990, 1990; pp. 119-124.
- Pence, V.C. Evaluating costs for the in vitro propagation and preservation of endangered plants. In Vitro Cellular & Developmental Biology-Plant 2011, 47, 176–187. [Google Scholar]
- Backs-Hüsemann, D.; Reinert, J. Embryobildung durch isolierte Einzelzellen aus Gewebekulturen vonDaucus carota. Protoplasma 1970, 70, 49–60. [Google Scholar] [CrossRef]
- Sochacki, D.; Marciniak, P.; Ciesielska, M.; Zaród, J.; Sutrisno. The Influence of Selected Plant Growth Regulators and Carbohydrates on In Vitro Shoot Multiplication and Bulbing of the Tulip (Tulipa L.). Plants 2023, 12, 1134. [CrossRef] [PubMed]
- Plessner, O.; Ziv, M.; Negbi, M. In vitro corm production in the saffron crocus (Crocus sativus L.). Plant Cell, Tissue and Organ Culture 1990, 20, 89–94. [Google Scholar] [CrossRef]
- Santos, J.; Santos, I.; Salema, R. In vitro production of bulbs of Narcissus bulbocodium flowering in the first season of growth. Scientia Horticulturae 1998, 76, 205–217. [Google Scholar] [CrossRef]
- Santos, A.; Fidalgo, F.; Santos, I.; Salema, R. In vitrobulb formation ofNarcissus asturiensis, a threatened species of theAmaryllidaceae. The Journal of Horticultural Science and Biotechnology 2015, 77, 149–152. [Google Scholar] [CrossRef]
- Bahr, L.R.; Compton, M.E. Competence for in vitro bulblet regeneration among eight Lilium genotypes. HortScience 2004, 39, 127–129. [Google Scholar] [CrossRef]
- Sultana, J.; Sutlana, N.; Siddique, M.; Islam, A.; Hossain, M.; Hossain, T. In vitro bulb production in Hippeastrum (Hippeastrum hybridum). Journal of Central European Agriculture 2010. [Google Scholar]
- Kizil, S.; Khawar, K.M. The effects of plant growth regulators and incubation temperatures on germination and bulb formation of Fritillaria persica L. Propag Ornam Plants 2014, 14, 133–138. [Google Scholar]
- Podwyszyńska, M.; Kosson, R.; Treder, J. Polyamines and methyl jasmonate in bulb formation of in vitro propagated tulips. Plant Cell, Tissue and Organ Culture (PCTOC) 2015, 123, 591–605. [Google Scholar] [CrossRef]
- Stanišić, M.; Raspor, M.; Ninković, S.; Milošević, S.; Ćalić, D.; Bohanec, B.; Trifunović, M.; Petrić, M.; Subotić, A.; Jevremović, S. Clonal fidelity of Iris sibirica plants regenerated by somatic embryogenesis and organogenesis in leaf-base culture—RAPD and flow cytometer analyses. South African Journal of Botany 2015, 96, 42–52. [Google Scholar] [CrossRef]
- ÇAkmak, D.; KaraoĞLu, C.; Aasim, M.; Sancak, C.; ÖZcan, S. Advancement in protocol for in vitro seed germination, regeneration, bulblet maturation, and acclimatization of Fritillaria persica. Turkish Journal of Biology 2016, 40, 878–888. [Google Scholar] [CrossRef]
- Youssef, N.M.; Shaaban, S.A.; Ghareeb, Z.F.; Taha, L.S. In vitro bulb formation of direct and indirect regeneration of Lilium orientalis cv. “Starfighter” plants. Bulletin of the National Research Centre 2019, 43, 211. [Google Scholar] [CrossRef]
- Maślanka, M.; Mazur, J.; Kapczyńska, A. In Vitro Organogenesis of Critically Endangered Lachenalia viridiflora. Agronomy 2022, 12, 475. [Google Scholar] [CrossRef]
- Doğan, S.; Çağlar, G.; Palaz, E.B. The effect of different applications on in vitro bulb development of an endemic hyacinth plant (Hyacinthus orientalis L. subsp. chionophyllus Wendelbo) grown in Turkey. Turkish Journal of Agriculture-Food science and technology 2020, 8, 1713–1719. [Google Scholar] [CrossRef]
- Ebrahimzadeh, H.; Karamian, R.; NOURI, D.M. Somatic Embryogenesis and Regeneration of Plantlet in Saffron, Crocus Sativus L. 2000.
- Sheibani, M.; Nemati, S.H.; Davarinejad, G.H.; Azghandi, A.V.; Habashi, A.A. INDUCTION OF SOMATIC EMBRYOGENESIS IN SAFFRON USING THIDIAZURON (TDZ). 2007; pp. 259-267.
- Marković, M.; Trifunović-Momčilov, M.; Radulović, O.; Paunović, D.M.; Antonić Reljin, D.D.; Uzelac, B.; Subotić, A. The Effects of Different Auxin–Cytokinin Combinations on Morphogenesis of Fritillaria meleagris Using Bulb Scale Sections In Vitro. Horticulturae 2023, 9. [Google Scholar] [CrossRef]
- Kocak, M.; Izgu, T.; Sevindik, B.; Tutuncu, M.; Curuk, P.; Simsek, O.; Aka Kacar, Y.; Teixeira da Silva, J.A.; Yalcin Mendi, Y. Somatic embryogenesis of Turkish Cyclamen persicum Mill. Scientia Horticulturae 2014, 172, 26–33. [Google Scholar] [CrossRef]
- Bach, A.; Ptak, A. Somatic embryogenesis and plant regeneration from ovaries of Tulipa gesneriana L. in in vitro cultures. Acta Horticulturae 2001, 391–394. [Google Scholar] [CrossRef]
- Podwyszyńska, M.; Marasek-Ciolakowska, A. Micropropagation of tulip via somatic embryogenesis. Agronomy 2020, 10, 1857. [Google Scholar] [CrossRef]
- Fersing, J.; Mouras, A.; Lutz, A. First stage in the industrial vegetative propagation of Cyclamen by in vitro culture. Pepinieristes, horticulteurs, maraichers 1982. [Google Scholar]
- İzgü, T.; Sevindik, B.; Çürük, P.; Şimşek, Ö.; Aka Kaçar, Y.; Teixeira da Silva, J.A.; Yalçın Mendi, Y. Development of an efficient regeneration protocol for four Cyclamen species endemic to Turkey. Plant Cell, Tissue and Organ Culture (PCTOC) 2016, 127, 95–113. [Google Scholar] [CrossRef]
- Kiviharju, E.; Tuominen, U.; Törmälä, T. The effect of explant material on somatic embryogenesis of Cyclamen persicum Mill. Plant Cell, Tissue and Organ Culture 1992, 28, 187–194. [Google Scholar] [CrossRef]
- Otani, M.; Shimada, T. Somatic embryogenesis and plant regeneration from Cyclamen persicum Mill. leaf cultures. Plant tissue culture letters 1991, 8, 121–123. [Google Scholar] [CrossRef]
- Takamura, T.; Miyajima, I.; Matsuo, E. Somatic embryogenesis of Cyclamen persicum Mill.‘Anneke’from aseptic seedlings. Plant Cell Reports 1995, 15, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Wicart, G.; Mouras, A.; Lutz, A. Histological study of organogenesis and embryogenesis in Cyclamen persicum Mill. tissue cultures: evidence for a single organogenetic pattern. Protoplasma 1984, 119, 159–167. [Google Scholar] [CrossRef]
- Kumari, A.; Baskaran, P.; Van Staden, J. In vitro propagation via organogenesis and embryogenesis of Cyrtanthus mackenii: a valuable threatened medicinal plant. Plant Cell, Tissue and Organ Culture (PCTOC) 2017, 131, 407–415. [Google Scholar] [CrossRef]
- Nautiyal, A.; Ramlal, A.; Agnihotri, A.; Rashid, A. Stress-induced somatic embryogenesis on seedlings of Azadirachta indica A. Juss. by thidiazuron and its inhibition by ethylene modulators. Plant Cell, Tissue and Organ Culture (PCTOC) 2023, 153, 357–366. [Google Scholar] [CrossRef]
- Yu, Y.; Qin, W.; Li, Y.; Zhang, C.; Wang, Y.; Yang, Z.; Ge, X.; Li, F. Red light promotes cotton embryogenic callus formation by influencing endogenous hormones, polyamines and antioxidative enzyme activities. Plant Growth Regulation 2019, 87, 187–199. [Google Scholar] [CrossRef]
- MENDİ, N.Y.Y.; ÖZDEMİR, Ş.; UĞUR, S.; SEVİNDİK, B.; İZGÜ, T.; KAYA, E.; TÜTÜNCÜ, M.; ÇÜRÜK, P.; ŞİMŞEK, Ö.; KOYUNCU, O. In vitro Regeneration and Synthetic Seed Production of Colchicum cilicicum Grown Naturally in Turkey. alatarım, 18.
- Beruto, M.; Curir, P.; Debergh, P. Callus growth and somatic embryogenesis in thalamus tissue of Ranunculus asiaticus L. cultivated in vitro: Cytokinin effect and phenol metabolism. In Vitro Cellular & Developmental Biology-Plant 1996, 32, 154–160. [Google Scholar]
- Azeri, F.N.; Öztürk, G. Microbulb and plantlet formation of a native bulbous flower, Lilium monodelphum M. Bieb, var. Armenum, through tissue culture propagation. Biotechnology Reports 2021, 32, e00665. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Bioreactor systems for micropropagation of plants: present scenario and future prospects. Front Plant Sci 2023, 14, 1159588. [Google Scholar] [CrossRef] [PubMed]
- Akita, M.; Takayama, S. Induction and development of potato tubers in a jar fermentor. Plant Cell, Tissue and Organ Culture 1994, 36, 177–182. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Alsadon, A.; Al-Aizari, A.A.; Al-Mohidib, M. In vitro floral emergence and improved formation of saffron daughter corms. Horticulturae 2022, 8, 973. [Google Scholar] [CrossRef]
- Gao, R.; Wu, S.-Q.; Piao, X.-C.; Park, S.-Y.; Lian, M.-L. Micropropagation of Cymbidium sinense using continuous and temporary airlift bioreactor systems. Acta physiologiae plantarum 2014, 36, 117–124. [Google Scholar] [CrossRef]
- Hao, Z.; Ouyang, F.; Geng, Y.; Deng, X.; Hu, Z.; Chen, Z. Propagation of potato tubers in a nutrient mist bioreactor. Biotechnology techniques 1998, 12, 641–644. [Google Scholar] [CrossRef]
- Jiménez, E.; Pérez, N.; de Feria, M.; Barbón, R.; Capote, A.; Chávez, M.; Quiala, E.; Pérez, J.C. Improved production of potato microtubers using a temporary immersion system. Plant Cell, Tissue and Organ Culture 1999, 59, 19–23. [Google Scholar] [CrossRef]
- Jo, U.; Murthy, H.; Hahn, E.; Paek, K. Micropropagation of Alocasia amazonica using semisolid and liquid cultures. In Vitro Cellular & Developmental Biology-Plant 2008, 44, 26–32. [Google Scholar]
- Kim, E.; Hahn, E.; Murthy, H.; Paek, K. Enhanced shoot and bulblet proliferation of garlic (Allium sativum L.) in bioreactor systems. The Journal of Horticultural Science and Biotechnology 2004, 79, 818–822. [Google Scholar] [CrossRef]
- Lian, M.-L.; Piao, X.-C.; Park, S.-Y. Mass production of lilium bulblets in bioreactors. Production of biomass and bioactive compounds using bioreactor technology 2014, 389–415. [Google Scholar]
- Lian, M.; Chakrabarty, D.; Paek, K. Bulblet formation from bulbscale segments of Lilium using bioreactor system. Biologia Plantarum 2003, 46, 199–203. [Google Scholar] [CrossRef]
- Lim, S.; Seon, J.; Paek, K.; Han, B.; Son, S. Development of pilot scale process for mass production of Lilium bulblets in vitro. In Proceedings of the International Symposium on Biotechnology of Tropical and Subtropical Species Part 2 461; 1997; pp. 237–242. [Google Scholar]
- Piao, X.C.; Chakrabarty, D.; Hahn, E.J.; Paek, K.Y. A simple method for mass production of potato microtubers using a bioreactor system. Current Science 2003, 1129–1132. [Google Scholar]
- Teisson, C.; Alvard, D. In vitro production of potato microtubers in liquid medium using temporary immersion. Potato research 1999, 42, 499–504. [Google Scholar] [CrossRef]
- Yu, W.-C.; Joyce, P.; Cameron, D.; McCown, B. Sucrose utilization during potato microtuber growth in bioreactors. Plant Cell Reports 2000, 19, 407–413. [Google Scholar] [CrossRef]
- Marković, M.; Trifunović Momčilov, M.; Uzelac, B.; Jevremović, S.; Subotić, A. Bulb Dormancy In Vitro—Fritillaria meleagris: Initiation, Release and Physiological Parameters. Plants 2021, 10, 902. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, F.R.; Jose-Santhi, J.; Kalia, D.; Singh, K.; Singh, R.K. Sugars as the regulators of dormancy and sprouting in geophytes. Industrial Crops and Products 2022, 189, 115817. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, C.; Sui, J.; Liang, J.; Ge, J.; Li, J.; Pan, W.; Yi, M.; Du, Y.; Wu, J. A wake-up call: signaling in regulating ornamental geophytes dormancy. Ornamental Plant Research 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Takayama, S.; Misawa, M. Differentiation in Lilium bulbscales grown in vitro. Effects of activated charcoal, physiological age of bulbs and sucrose concentration on differentiation and scale leaf formation in vitro. Physiologia Plantarum 1980, 48, 121–125. [Google Scholar] [CrossRef]
- Langens-Gerrits, M.M.; Miller, W.B.; Croes, A.F.; De Klerk, G.-J. Effect of low temperature on dormancy breaking and growth after planting in lily bulblets regenerated in vitro. Plant growth regulation 2003, 40, 267–275. [Google Scholar] [CrossRef]
- Carasso, V.; Mucciarelli, M. In vitro bulblet production and plant regeneration from immature embryos of Fritillaria tubiformis Gren. & Godr. Propagation of Ornamental Plants 2014, 14, 101–111. [Google Scholar]
- Kamo, K.; Roh, M.; Blowers, A.; Smith, F.; Eck, J. I. 12 Transgenic Gladiolus. Transgenic Crops III 2013, 48, 155. [Google Scholar]
- Van Tuyl, J.M.; Arens, P.; Marasek-Ciolakowska, A. Breeding and genetics of ornamental geophytes. Ornamental Geophytes: From Basic Science to Sustainable Horticultural Production, 1st ed.; Kamenetsky, R., Okubo, H., Eds 2012, 131-158.
- Van Harmelen, M.; Löffler, H.; Van Tuyl, J. Somaclonal variation in lily after in vitro cultivation. In Proceedings of the VII International Symposium on Flowerbulbs 430; 1996; pp. 347–350. [Google Scholar]
- Abdolinejad, R.; Shekafandeh, A.; Jowkar, A.; Gharaghani, A.; Alemzadeh, A. Indirect regeneration of Ficus carica by the TCL technique and genetic fidelity evaluation of the regenerated plants using flow cytometry and ISSR. Plant Cell, Tissue and Organ Culture (PCTOC) 2020, 143, 131–144. [Google Scholar] [CrossRef]
- Konar, S.; Adhikari, S.; Karmakar, J.; Ray, A.; Bandyopadhyay, T.K. Evaluation of subculture ages on organogenic response from root callus and SPAR based genetic fidelity assessment in the regenerants of Hibiscus sabdariffa L. Industrial Crops and Products 2019, 135, 321–329. [Google Scholar] [CrossRef]
- Vitamvas, J.; Viehmannova, I.; Cepkova, P.H.; Mrhalova, H.; Eliasova, K. Assessment of somaclonal variation in indirect morphogenesis-derived plants of Arracacia xanthorrhiza. Pesquisa Agropecuária Brasileira 2019, 54. [Google Scholar] [CrossRef]
- Memon, N.; Yasmin, A.; Pahoja, V.; Hussain, Z.; Ahmad, I. In vitro regeneration of gladiolus propagules. J. Agr. Technol 2012, 8, 2331–2351. [Google Scholar]
- Asadi, N.; Zarei, H.; Hashemi-Petroudi, S.H.; Mousavizadeh, S.J. Micropropagation and assessment of somaclonal variation in Galanthus transcaucasicus in vitro plantlets. Ornamental Horticulture 2021, 27, 505–515. [Google Scholar] [CrossRef]
Figure 2.
In vitro regeneration pathways.
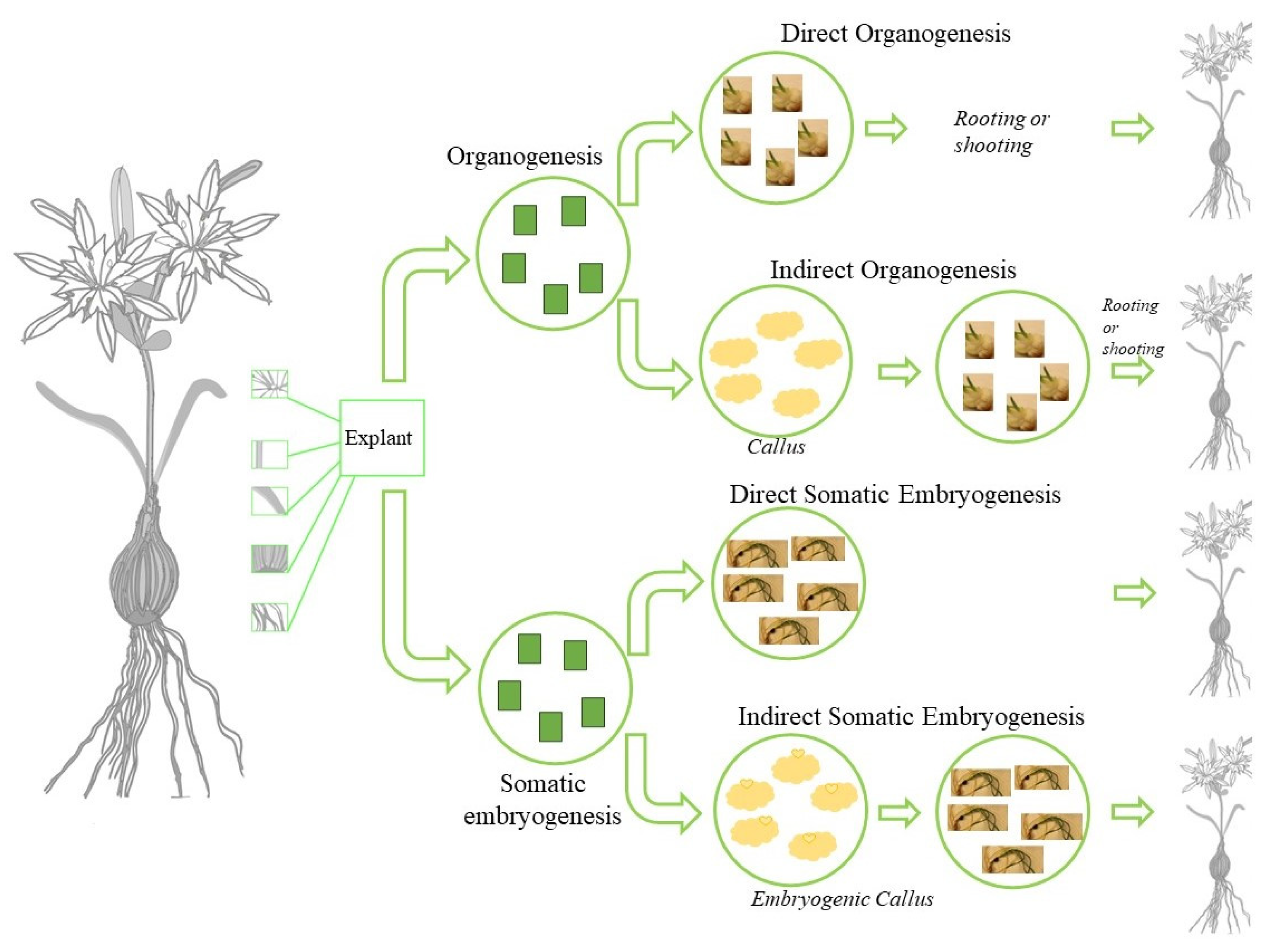
Figure 3.
Biological performance of Ranunculus shoots in vitro cultured on basal MS medium gelled with different agar brands. A= agar Oxoid (OX; cd L13(3)), 8 g/L; B= agar Roth (RT; cd 4508), 8 g/L; C= agar OX (8 g/L) supplemented with the impurities collected from dialysis of 8 g of RT agar powder; D= washed agar RT, 18 g/L.
Figure 3.
Biological performance of Ranunculus shoots in vitro cultured on basal MS medium gelled with different agar brands. A= agar Oxoid (OX; cd L13(3)), 8 g/L; B= agar Roth (RT; cd 4508), 8 g/L; C= agar OX (8 g/L) supplemented with the impurities collected from dialysis of 8 g of RT agar powder; D= washed agar RT, 18 g/L.

Figure 4.
One month after the embryos of Pancratium maritimum inoculated under 16/8 h photoperiod (A) and dark (B) conditions.
Figure 4.
One month after the embryos of Pancratium maritimum inoculated under 16/8 h photoperiod (A) and dark (B) conditions.
Figure 5.
Different explants examples from in vitro propagation of flower bulbs; Sea daffodil seeds (A), Gladioulus corm (B) and Fritillaria bulbs (C).
Figure 5.
Different explants examples from in vitro propagation of flower bulbs; Sea daffodil seeds (A), Gladioulus corm (B) and Fritillaria bulbs (C).
Figure 6.
Organogenic structure, leaf and bulbs growth from P. maritimum callus derived from bulb scales.
Figure 6.
Organogenic structure, leaf and bulbs growth from P. maritimum callus derived from bulb scales.

Figure 7.
Embryo-like structures (ELS) in callus of P. maritimum (A, B, C) and embryogenic callus and regeneration in Ranunculus asiaticus (D, E).
Figure 7.
Embryo-like structures (ELS) in callus of P. maritimum (A, B, C) and embryogenic callus and regeneration in Ranunculus asiaticus (D, E).
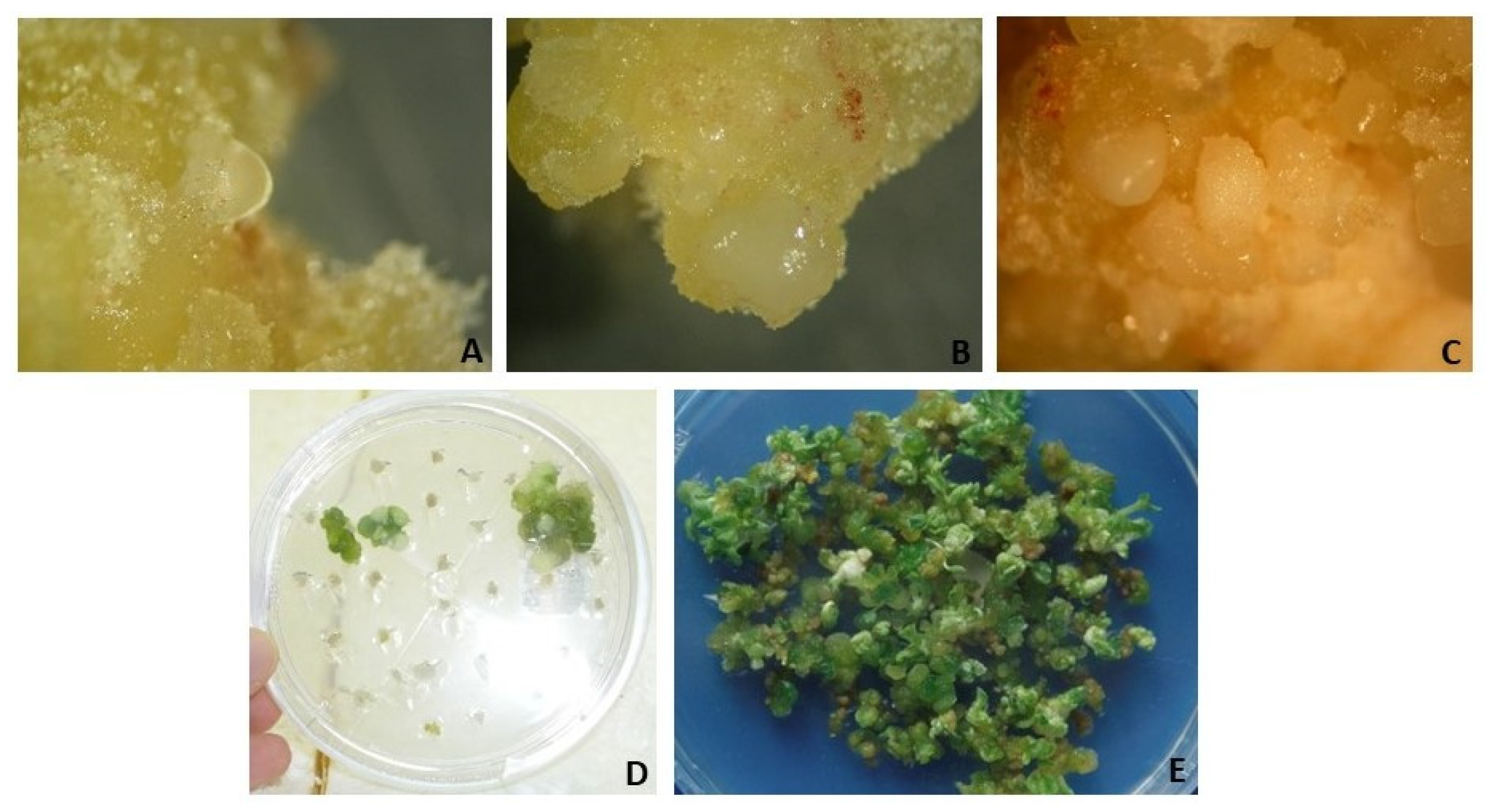
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



