
Submitted:
09 January 2024
Posted:
09 January 2024
You are already at the latest version
Abstract
Orthobunyaviruses (order Bunyavirales, family Peribunyaviridae) in the Simbu serogroup have been responsible for widespread epidemics of congenital disease in ruminants. Australia has a national program to monitor arboviruses of veterinary importance. While monitoring for Akabane virus, a novel orthobunyavirus was detected. To inform the priority that should be given to this detection, a scoping review was undertaken to 1) Characterise the associated disease presentations and establish which of the Simbu group viruses are of veterinary importance; 2) Examine the diagnostic assays that have undergone development and validation for this group of viruses, and 3) Describe the methods used to monitor the distribution of these viruses. Two search strategies identified 224 peer-reviewed publications for 33 viruses in the serogroup. Viruses in this group may cause severe animal health impacts, but only those phylogenetically arranged in clade B are associated with animal disease. Six viruses (Akabane, Schmallenberg, Aino, Shuni, Peaton, and Shamonda) were associated with congenital malformations, neurological signs, and reproductive disease. Diagnostic test interpretation is complicated by cross-reactivity, the timing of foetal immunocompetence and sample type. Serological testing in surveys remains a mainstay of the methods used to monitor the distribution of SGVs. Due to significant differences in survey designs, only broad mean seroprevalence estimates could be provided. Further research is required to characterise the SGV detected and determine the disease risk posed by novel orthobunyaviruses and how they could challenge current diagnostic and surveillance capabilities.
Keywords:
Arboviruses
; Animal Health
; Orthobunyavirus
; Simbu serogroup
* Correspondence email: tiffany.o'connor@dpi.nsw.gov.au
1. Introduction
Arboviruses within the Orthobunyavirus genus (order Bunyavirales, family Peribunyaviridae,) pose a substantial animal health threat as the geographical spread of insect-borne viruses expands, and the frequency and magnitude of their epidemics increase [1,2]. Orthobunyaviruses are approximately 90-100nm in diameter, spherical, and double-membraned with a fringe of protein spikes [2]. The glycoprotein spikes are arranged in an ordered lattice and protrude 18nm from the surface [2]. The genome consists of three separate, single-stranded, negative-sense RNA segments that are named according to their size: small (S, ~1kb), medium (M, ~4.5kb) and large (L, ~6kb) [3]. Several orthobunyaviruses, including Schmallenberg virus (Orthobunyavirus schmallenbergense) and Akabane virus (Orthobunyavirus akabaneense), have been responsible for widespread epidemics of congenital disease in ruminants [2].
Taxonomic classification of orthobunyaviruses is challenging due to the genetic diversity and the large number of viruses in the genus. Historically, many orthobunyaviruses were isolated through international discovery programs and named based on their antigenic similarity [4,5]. The antigenic properties are related to the two surface glycoproteins (Gn and Gc) and the nucleoprotein (N) protein encoded by the M and S segments, respectively [6]. The genus was organised into 18 serogroups based on antigenic similarities between viruses [5,7]. The current taxonomic classification system is based on the evolutionary history of the viruses [8,9]. Orthobunyaviruses are grouped into species complexes when there is >96% identity in the complete amino acid sequence of the L protein, an RNA-dependent RNA polymerase, translated from the L segment [8,10]. As a result, species complexes have grouped together multiple viruses with different antigenic characteristics [11] (Supplementary Table S1). This genus currently comprises over 189 named viruses assigned into 103 species complexes [12].
The Simbu group is one of the larger serogroups comprising 33 named viruses assigned to 19 species complexes with a global distribution (Supplementary Table S1) [5,8]. The Simbu group viruses (SGVs) may be further genetically divided into two monophyletic clades, Clade A and Clade B. Clade designation is based on phylogenetic relationships between SGVs and reflects an evolutionary divide that is consistent with previously established serogroups [13]. Serogroup and clade assignment allow for virus classification by both their antigenic and phylogenetic properties [11,13]. The vertebrate hosts, vectors, and/or pathogenicity of over 60% of the SGVs within Clade A remain unknown (Supplementary Table S1). Several Clade A viruses are classified in the Oropouche species complex (Orthobunyavirus oropoucheense) and notably two are associated with fever and encephalitis in humans. Clade B comprises viruses that are important animal pathogens, including Akabane virus (Orthobunyavirus akabaneense), Schmallenberg virus (Orthobunyavirus schmallenbergense), and Shuni virus (Orthobunyavirus shuniense) [14,15,16].
For SGVs in Clade B, virus transmission to mammalian hosts occurs via hematophagous arthropods such as biting midges of the genus Culicoides (Supplementary Table S1). These Clade B viruses, several of which are recognised teratogens, have been found worldwide, with the exception of North America (Figure 1, Supplementary Table S1). In endemic regions, virus infection typically results in minimal disease in adult animals during transmission seasons of high vector activity. However, epidemics occur when the distribution of the insect vector intermittently overlaps with that of a naïve host population [17,18]. This occurs when environmental conditions allow the insect vectors to move into new populations of susceptible livestock, or when these insect vectors are absent from part of their normal distribution range for some time, giving rise to susceptible mammalian hosts [18]. The most severe disease syndrome is observed when naïve pregnant animals are infected at critical stages of gestation resulting in foetal pathology following in-utero infection.
In Australia, the distribution of selected insect-borne viruses of veterinary importance are monitored by the National Arbovirus Monitoring Program. This active surveillance system involves the longitudinal sampling of sentinel herds across Australia and trapping Culicoides midges [19]. Data on both population immunity and vector activity can be used to identify areas free from arbovirus transmission, where naïve animals may be at risk of disease and to support trade. In January 2017, an unknown orthobunyavirus was detected in a sentinel cattle herd during surveillance activities in New South Wales (NSW), Australia. Preliminary analyses suggest that this isolate may be most similar to Shamonda virus (Orthobunyavirus schmallenbergense). To inform the priority that should be given to the detection of a Shamonda-like virus in Australia, a scoping review was undertaken to 1) Characterise the associated disease presentations and establish which of the Simbu group viruses (SGVs) are of veterinary importance; 2) Examine the diagnostic assays that have undergone development and validation for this group of viruses, and 3) Describe the methods used to monitor the distribution of the SGVs.
2. Materials and Methods
2.1. Protocol
The review protocol was completed in accordance with the Preferred Reporting Items for Systematic reviews and Meta-Analyses extension for Scoping Reviews guidelines [20]. The protocol was agreed by all authors on 28 April 2022.
2.2. Search strategies
Several combinations of search terms were attempted to identify eligible publications with no restrictions applied on search fields. Initially, given the detection of a Shamonda-like virus, a search to identify publications describing SGVs and specifically for this virus was used to frame the study objectives. This first strategy used the genus name but restricted publications to those including the serogroup name or the virus name. However, to encompass the whole Simbu group, this strategy was expanded to include varying combinations of genus and individual virus names. This second search strategy listing the genus name and all 33 virus names returned 276,211 publications in Scopus. This search strategy was refined further by applying restriction to the domestic animal species of interest only (bovine, ovine, caprine, equine and swine). This refined search strategy returned 8,677 publications in Scopus before it was used across other scientific databases. Both search strategies were combined to identify publications in five databases: Scopus, Web of Science, CAB Abstracts, BIOSIS and Medline. The first search was performed on 26 May 2022. The second search was performed on 11 August 2022.
- (“Bunyavir*” OR “orthobunyavirus*”) AND (“Shamonda*” OR “Simbu*”)
- (“Bunyavir*” OR “orthobunyavirus” OR “Akabane” OR “Tinaroo” OR “Yaba-7” OR “Jatobal” OR “Buttonwillow” OR “Cat Que” OR “Ingwavuma” OR “Inini” OR “Manzanilla” OR “Mermet” OR “Facey's paddock” OR “Iquitos” OR “Madre de Dios” OR “Oropouche” OR “Perdoes” OR “Pintupo” OR “Utinga” OR “Utive” OR “Douglas” OR “Sango” OR “Sathuperi” OR “Peaton” OR “Schmallenberg” OR “Shamonda” OR “Sabo” OR “Aino” OR “Kaikalur” OR “Shuni” OR “Oya” OR “Para” OR “Simbu” OR “Thimiri”) AND (“bovine” OR “cattle” OR “cow” OR “calf” OR “calves” OR “dairy” OR “beef” OR “ovine” OR “sheep” OR “caprine” OR “goat” OR “equine” OR “horse” OR “porcine” OR “pig” or “pork”)
The search was applied across all databases and the publications retrieved from both strategies were exported into the citation manager software Endnote.
2.3. Eligibility criteria
A systematic approach was applied to find eligible publications using three levels of screening (Figure 2): A) Screening by publication type for original research publications in English, with no restrictions on publication date and type. Duplicates and non-peer-reviewed publications were excluded; B) Screening by title and keywords for eligible publications relevant to the 33 viruses of the Simbu group (Supplementary Table S1), and by title and abstract for domestic ruminant, equid or porcine species; and C) Screening by abstract for eligible publications providing information relevant to at least one of the three themes of interest: 1) Animal disease presentation, which recorded observations of clinical signs associated with SGVs; 2) Diagnostic tests, which described the development or validation of tests for detecting SGVs; and 3) Monitoring, which determined the distribution of SGVs. Allocation to a theme was based on the aims and objectives of the research publication. Where the publication could be relevant to more than one theme, allocation was made following evaluation of the text in full.
2.4. Data collection and analysis
Data were extracted for all three themes according to the details listed in Table 1. The data were recorded in Excel (Microsoft, 2023) before data summaries were prepared in R (R Core Team, 2022). For publications where multiple categorical levels of one variable were applicable (e.g., host species had >1 species listed if an eligible publication described disease presentation in multiple species), the details were recorded as a comma-separated list. Vector operations, grouped into the ‘RepRows’ function, in R were used to extract the recorded individual items in a free text list and match these variables to the details of each publication. A record was generated for each extracted item in the free text list from each publication. This allowed for multiple recorded items for the same variable to be counted and analysed from the one publication (i.e., for multiple host species, a separate record for bovine and ovine would be generated for the same publication). To avoid double counting, multiple variables were not extracted together for analysis.

Table 1.
Data extracted from eligible publications in a scoping review to identify veterinary-important Simbu group viruses. This information was used to produce a summary of the associated disease presentations, monitoring methods, and diagnostic assays of the SGVs with veterinary importance.
Table 1.
Data extracted from eligible publications in a scoping review to identify veterinary-important Simbu group viruses. This information was used to produce a summary of the associated disease presentations, monitoring methods, and diagnostic assays of the SGVs with veterinary importance.
| Variable | Description | Unit/Levels |
|---|---|---|
| Authors | The names of the authors listed on the publication | Free text list |
| Publication Date | The date for which the publication was published | Date/Month/Year format |
| Publication Year | The year for which the publication was published | Date/Month/Year format |
| Country | A list of the country affiliations of the authors | Free text list |
| Continent | The continent in which the study was conducted | Americas Africa Asia Australia/Oceania Europe |
| Title | The title of the publication | Free text list |
| Journal | The name of the journal of publication | Free text list |
| Virus name | The Simbu group virus/es investigated | see Supplementary Table S1 for the 33 viruses included in the serogroup |
| Theme: The veterinary important SGVs and the associated animal disease presentation | ||
| Study type | Study type/design | Descriptive Observational Experimental Theoretical |
| Infection type | Natural exposure or experimental challenge | Natural Experimental |
| Case definition1 | Diagnostic criterion applied to recruit cases into this review to provide a systematic description of disease presentations associated with SGV infection |
Clinical presentation only Antibody detection only Agent detection only Consistent clinical presentation with confirmation of viral infection with a SGV |
| Diagnostic test | Test type(s) utilized for confirmation of disease due to the virus of interest | Free text list |
| Start | The start date of the period of observation | Date/Month/Year format |
| End | The end date of the period of observation | Date/Month/Year format |
| Season | The season in which the observations of animal disease occurred. The date of disease report was used to determine the season based on the period and location of the observation. |
Summer Autumn Spring Winter |
| Host species | Species of animal host with disease | Bovine Ovine Caprine Porcine Equine |
| Host age group2 | Age group of the animal host with disease |
Foetal: unborn Juvenile: born to <12 months of age Adult: 12 months of age |
| Disease system2 | The organ system involved | Circulatory Respiratory Gastrointestinal Musculoskeletal Nervous Urinary Reproductive Congenital Non-specific |
| Clinical signs2 | A list of all clinical signs reported | Free text list |
| Frequency of clinical signs listed | The count of the observations of clinical signs by the number of cases | Numeric |
| Differentials considered | What other diseases were considered | Free text list |
| Diagnostic tests | Assay types used to diagnose the virus of interest | Free text list |
| Histopathological description | The inclusion of histopathological description of disease | Yes No |
| Economic losses estimated | The inclusion of an estimate or economic assessment of the impact of associated disease | Yes, with information collected in a free text list No |
|
1 This assessment was completed to ensure that cases of clinical disease described was confirmed to be a result of infection with the virus of interest. The four diagnostic criteria are based on the unified case definitions proposed by European agencies during data collection for the Schmallenberg virus epidemic [21]. The diagnostic challenge in identifying a SGV as a cause of disease were exemplified by the different and varying case definitions used by affected countries during the outbreak in Europe. In January 2012, different countries had varying criteria for what constituted a confirmed case following Schmallenberg virus infection. These criteria depended on factors such as the age of the host (offspring or adult) or the location of the infection (within or outside the known range). Additionally, laboratory confirmation of the viral infection was achieved through a variety of agent and/or antibody tests. 2 To provide a distinction between maternal and foetal outcomes of infection, clinical signs relating to reproductive and neurological disease were categorised given the context provided from the host age group. This allowed for congenital disease to be considered as a foetal outcome and reproductive disease to be measured as a maternal effect. Theme: Diagnostic tests – development and validation studies | ||
| Test scenario | Reason for testing | Monitoring populations for infection/exposure Diagnosis in animals where disease is suspected |
| Sample types | List of the samples utilized in test development or validation | Free text list |
| Test types | List of the test types investigated | Free text list |
| Test level | Test is designed to detect SGVs either at a group level or to differentiate a specific named virus | Group Virus |
| Test target | Test designed for serology or agent detection | Serology Agent |
| Test details | Details such as the test manufacturer, reagents, primer, and probe sequences | Free text list |
| Test validation | If the assay has undergone validation and if so, to which stage along the validation pathway as defined by WOAH guidelines [22] | None: No validation data provided Stage 1: Analytical characteristics Stage 2: Diagnostic characteristics and cut-off values Stage 3: Reproducibility Stage 4: Implementation |
|
Theme: Monitoring methods | ||
| Virus | Simbu group virus of interest for the activity | Free text list |
| Survey Area | Area of interest for the study | Local region (intra-country) Country Multinational Continent |
| Survey location | Details of the country and region where the activity occurred | Free text list |
| Survey period | Length of time over the course of the study | Numeric (months) |
| Sample collection | The type sample collection used in the study: Active if the samples were primarily collected for the surveillance activity. Passive if the samples were collected for another purpose (diagnostic investigation or a non-SGV surveillance program) |
Active Passive |
| Survey test | Test type(s) utilized in the monitoring activity | Free text list |
| Animal | Animal species from where samples were collected from | Free text list |
| Sample frequency | Frequency of sampling used for the study | Single Serial – repeated/ daily/ weekly/ monthly/ quarterly/ Yearly Paired |
| Sample size calculation | - If sample size calculations were considered and the details of the:
|
Yes/No Numeric for the information on the sample size calculation |
| Sample size | Sample size | Numeric |
| Seroprevalence3 | Reported seroprevalence in each study was charted by:
|
Numeric |
| 3 A quantitative summary of seroprevalence by country, animal species, and Simbu group virus was prepared using R with data extracted (Supplementary Figure S1). | ||
3. Results
3.1. Selection of sources of evidence
Implementation of the protocol and screening processes identified 87 eligible publications from the first search strategy and 217 publications from the second search strategy (Figure 2). Eight publications were incorporated through the first strategy, which were not identified via the second approach. Most publications (n = 46,609) were removed in the second search strategy after Title and Keyword screening for relevance (Figure 1). A total of 224 publications were combined from both search strategies, with 95 publications allocated to the animal disease presentation, 38 publications for diagnostic test development and validation, and 91 publications for monitoring methods to determine SGV distribution.
3.2. Characteristics of sources of evidence
The 224 eligible peer-reviewed publications were retrieved from 70 different journals. The five journals with the most publications were: Transboundary and Emerging Diseases (n = 23), Veterinary Microbiology (n = 19), Emerging Infectious Diseases (n = 12), Australian Veterinary Journal (n = 11) and BMC Veterinary Research (n = 10). Authors of the publications represented 50 countries, with most from Japan (n = 45), Germany (n = 30), Australia (n = 28), Belgium (n = 23), and France (n = 18).
A total of 321 records were extracted by virus name from all 224 eligible publications. Overall, Schmallenberg virus was the most frequently investigated (n = 108), followed by Akabane virus (n = 95), Aino virus (n = 37) and Peaton virus (n = 17). Following these, four viruses: Shuni virus, Shamonda virus and Douglas virus were investigated with equal frequency (n = 10). Records encompassed 17 of the 33 named viruses within the Simbu group. This included 10 viruses investigated for animal health impacts, 13 viruses for monitoring distribution, and 16 viruses for diagnostic tests (Table 2). The publications spanned from 1973 to 2022. Records for both Akabane virus and Aino virus were consistently published over this 49-year period (Figure 3).
3.3. Animal health impacts associated with Simbu group viruses
3.3.1. Clinical presentation of livestock disease
Overall, congenital malformations in ruminants were the most frequently observed disease syndrome that was attributed to SGV infection. Disease observations were most frequently recorded for bovine and most often with Akabane virus. Six SGVs resulted in congenital malformations in cattle: Akabane virus, Schmallenberg virus, Aino virus, Shuni virus, Peaton virus and Shamonda virus. In contrast, only three SGVs (Akabane virus, Schmallenberg virus and Shuni virus) were attributed with congenital malformations in sheep and goats. The type of congenital malformations depended on the gestational age at the time of infection (Table 3).
Most reports of severe neurological disease involved calves following in-utero infection with Akabane virus at one of two distinct periods of gestation (Table 4). Neurological signs were also reported in calves born alive following in-utero infection with Shuni virus [35,36], Aino virus [37,38], Peaton virus [39] and Schmallenberg virus [40]. Neurological signs were also reported in lambs following in-utero infection with Akabane virus [41,42]. The neurological signs included: Dysstasia/astasia, ataxia, circling, dull/slow to respond to surroundings, proprioceptive deficits, nystagmus, hypersensitivity, paralysis, tremors, abnormal gait, recumbency, unconscious paddling, loss of proprioception, swallowing difficulties and tongue paralysis. The musculoskeletal signs (stiff joints, lameness) were only reported in conjunction with neurological disease in calves and lambs following in-utero infection from Schmallenberg virus or Akabane virus [40,43,44].
Reproductive disease in adult animals was also observed across ruminant species and included reports of early embryonic death, repeated oestrus, and increased return to inseminations. Again, most disease was reported for cattle, with reproductive disease also associated with five SGVs: Schmallenberg virus, Akabane virus, Aino virus, Shuni virus and Peaton virus. For sheep and goats, the same three SGVs (Schmallenberg virus, Akabane virus and Shuni virus) were reported in association with both congenital malformations and reproductive signs. In comparison to congenital malformations, reproductive disease observations were most often associated with Schmallenberg virus across all three ruminant species. This strong association between Schmallenberg virus and reproductive disease is most obvious in sheep.
Non-reproductive disease in horses and adult ruminants was uncommon. There were five publications describing disease in adult animals attributed to infection with an SGV. Although neurological disease in horses following Shuni virus infection is the most common disease presentation in an adult animal [16,45], there were three case reports of neurological disease in adult cattle following infection with Shuni virus [35] and Akabane virus [46,47]. The neurological signs were comparable between juvenile and adult animals (Table 4).
Other disease observations in horses and adult ruminants included non-specific signs including pyrexia, reduced appetite with subsequent weight loss, temporarily reduced milk yield and excess salivation. Diarrhoea, the only gastrointestinal clinical sign observed, was reported in adult cattle and sheep following natural and experimental infection with Schmallenberg virus [48,49,50]. Nasal discharge and tachypnoea were the only two respiratory signs observed. These subtle signs were observed only during experimental infection with Schmallenberg virus in sheep [50] or Akabane virus in cattle [51]. There were no cases of clinical disease for swine examined in this review. This is because the two records retrieved following infection with Cát Quế virus and Ingwavuma virus retrieved in this systematic search did not fulfill the case definition required (Figure 4).
Disease was reported more often in winter (n = 26), spring (n = 22), and autumn (n = 20) than in summer (n = 12). This seasonal trend was consistent across all countries and viruses investigated, apart from Shuni virus. For Shuni virus, disease was more often reported in summer (n = 3) and autumn (n = 3) than in winter (n = 1) and spring (n = 1). The case numbers and frequency of clinical signs were extracted from 37 publications to provide the descriptive summary below (Table 4). A histopathological description was provided in some publications describing disease in ruminants (29/95, Supplementary Table S2).
In addition to the loss of replacement animals, economic impacts associated with disease from SGVs are due to costs associated with veterinary treatment and reduced production. Replacement animal costs represent a substantial economic loss. The Schmallenberg virus outbreak in Ireland in 2012 resulted in an estimated 10% decrease in weaning rates for affected sheep farms [52]. Veterinary costs were attributed to dystocia, which is common for ruminants following abortion, stillbirth, or attempted birth of a malformed foetus. Direct veterinary costs were estimated to be €65-107 per animal affected in Belgium in 2011 [53]. In dairy cows experiencing dystocia, milk yield decreased by 11.4-24.7% compared to normal parturition [54]. Adding to these production losses, acute infection in adult ruminants may result in reduced milk yield (Table 4). Overall, the economic impact of disease associated with Schmallenberg virus in cattle and sheep was best summarised in two European studies. In 2015, estimated disease costs per cow per year were £9.7-48.6 in France and £8.2-51.4 in the UK for cattle [55]. In 2017, estimated disease cost per cow per year was €23-43 in France [56]. This 2017 study also estimated the disease costs per ewe per year to be €19- 37 in France [56].
3.3.2. The Simbu group viruses associated with livestock disease
Most publications were observational studies (66/95) encompassing descriptive case series (n = 25), cross-sectional studies (n = 16), single case reports (n = 15), case-control studies (n = 4), and cohort studies (n = 2). Just over two-thirds of the publications fulfilled at least one of the four diagnostic criteria for Simbu group virus infection as listed in Table 1 (61/95, Figure 4). Akabane virus was extensively studied as a causative agent of animal disease. There are 31 records where clinical disease caused by Akabane virus infection was diagnosed. Five other Simbu group viruses met the required case definition, including Schmallenberg virus (n = 19) and then Aino virus (n = 6), Shuni virus (n = 5), Peaton virus (n = 2), and Shamonda virus (n = 1). While Ingwavuma virus, Douglas virus, Tinaroo virus and Sango virus were also investigated, these viruses were not diagnosed as the cause of clinical disease in livestock (Figure 4).
Laboratory tests for infections with SGVs in the 95 publications describing livestock disease presentation involved a range of different assay types. The earliest diagnostic tests used were virus neutralisation tests (VNT), virus isolation, immunofluorescence testing (IFAT), and histopathological examination. The earliest ELISA tests for SGVs were used in a publication from 1988 [42], and PCR diagnostics tests became available from 2002 [47]. Supportive evidence for viral infection was provided through a range of assays: PCR (n = 11), VNT (n = 11), ELISA (n = 4), histopathological examination (n = 4), or virus isolation (n = 2). In over half the publications (n = 50), a combination of assay types was utilised to provide confirmation of viral infection. This included a combination of the aforementioned assays and/or the following – IFAT, immunohistochemistry, agar gel immunodiffusion test (AGID), complement fixation test (CFT), in-situ hybridisation, nucleic acid sequencing and foetal IgG, IgA, or IgM testing. The most frequent combinations included two test types – VNT with virus isolation (n = 7), ELISA with PCR (n = 5), and VNT with histopathological examination (n = 4). Thirteen publications described the clinical presentation only and did not provide details of laboratory testing (Figure 4).
Disease was observed following experimental infection for four SGVs from 29 publications: Akabane virus (n = 16), Schmallenberg virus (n = 10), Aino virus (n = 2) and Shuni virus (n = 1). Inoculation was most often by subcutaneous (n = 11) or intravenous (n = 11) injection in pregnant animals resulting in congenital malformations of the developing foetus. Other successful routes include intramuscular injection (n = 2), oronasal administration (n = 1) and intrauterine inoculation (n = 1) including intra-peritoneal injection of the foetus (n = 1). Intracerebral injection (n = 2) of juvenile calves resulted in the development of neurological disease with Akabane virus [43] and non-specific signs such as lethargy and pyrexia with Aino virus [44]. No clinical disease was reported following experimental challenge (n = 21) of non-pregnant adult ruminants with Shamonda virus, Douglas virus, Sango virus, or Peaton virus despite laboratory confirmation of viral infection by agent or antibody detection (Figure 4).
3.4. Test development and validation studies for assays to detect viruses in the Simbu group
Out of the 38 publications for review, 23 described assay development and the remaining 15 included descriptions of test validation. Validation, according to the World Organisation for Animal Health (WOAH) guidelines, was completed to Stage 1 for 8/15 publications, where the study determined analytical sensitivity and specificity and demonstrated repeatability and reproducibility compared to standard test methods [22]. Validation was completed to Stage 2 for 7/15 publications, where the study established the diagnostic sensitivity and specificity of the candidate test on a reference population, and determined cut-off values [22].
Most publications described an assay designed for the purpose of diagnosis (33/38), where disease was suspected to be a consequence of SGV infection. Diagnostic tests were most often developed for Schmallenberg virus (n = 21) and Akabane virus (n = 20) (Table 2). Assays in development were more often designed to detect a specific named virus (31/38), rather than a pan-reactive group test (7/38). Only a few described an assay that was used for surveillance purposes (5/38), where testing is completed in situations where there was no suspicion of disease. Given this limited validation of diagnostic tests for surveillance purposes, most of the assays were designed for use with individual animal samples (36/38). Only 2 out of 38 assays were designed for pooled samples for herd or population testing. In these instances, bulk milk was used as a population sample for developing serological tests.
Blood, whether in the form of serum (n = 22), whole blood (n = 9), or substitutes like thoracic fluid (n = 1), was the most used sample type for developing and validating assays. Although blood is an obvious choice for serological testing, publications were almost evenly split between tests targeting antibody (18/38) or agent (17/38) detection. Agent detection assays also used a range of tissues as sample types in test development and validation (n = 14) with the brain the most frequently used organ (n = 5). Semen (n = 2), Culicoides homogenates (n = 2) and milk (n =1) were less often utilised.
The assay types that were most often evaluated were: PCR (n = 18), ELISA (n = 18), and VNT (n = 17). Less commonly evaluated assay types included haemagglutination inhibition test (n = 2), electron microscopy (n = 1), IFAT (n = 1). Novel technologies were also investigated such as loop-mediated isothermal amplification (LAMP) (n = 2), recombinase polymerase amplification (n = 1) and antibody detection with the Luminex MAGPIX® system (n = 1). Further details on the test components were available for 30/38 publications allowing for the details on test design to be presented below.
3.4.1. Agent detection
Molecular assays for agent detection were most often designed with amplification targets in the S segment (n = 17) compared to M (n = 2) and L segments (n = 3). The full details on the primers and probe sequences are provided in Supplementary Table S3. Specific PCR assays for named viruses within the serogroup were mostly developed for Akabane virus (n = 8), Schmallenberg virus (n = 5), and Aino virus (n = 5). The S segment was the target for most of these specific virus assays (n = 15). The M segment is a highly variable genomic region and this segment was used in the development of a virus-specific PCR for Peaton virus [57] and Schmallenberg virus [58]. Two virus species specific PCRs based on the L segment were described for Schmallenberg virus [58,59,60]. To develop a group reactive PCR, primer and probes sequences were designed for the S [57,61,62] and L segments [58,61,63,64].
To achieve WOAH Stage 1 validation, known virus isolates were utilised in the PCR development to demonstrate analytical specificity against the different SGVs and analytical sensitivity with dilution series conducted to determine the limit of detection (Table 2). Only two PCR assays were validated with reference samples to establish diagnostic specificity and sensitivity [62,65]. Reference samples in both publications were limited to materials with Schmallenberg virus with known positive samples sourced from experimentally infected animals and/or field samples. However, the diagnostic specificity could not be estimated for the Schmallenberg virus real-time PCR assay (SBV-S) because no known negative samples were tested in this publication. During testing to determine the analytical specificity of the SBV-S assay, false positive results were observed for five out of nine virus isolates, which included Sabo virus, Shamonda virus, Simbu virus, Peaton virus, and Douglas virus. Nonetheless, the SBV-S PCR test has a diagnostic sensitivity of 100% when assessed using reference samples that are positive for the Schmallenberg virus (91.8-100%, n = 43)[65]. The universal S-segment-based real-time PCR assay (Uni-S) has a diagnostic specificity of 100% (89.1-100%, n = 32), and a diagnostic sensitivity of 83.7% (69.3-93.2%, n = 43) [62] when assessed using reference samples for the Schmallenberg virus, Akabane virus and Shuni virus.
3.4.2. Antibody detection
For ELISAs, detection of orthobunyaviruses within the same serogroup is achieved when the test is designed against the N protein. However, the detection of a specific virus within the serogroup can be achieved when the Gc protein is used as the test target, such as for Akabane virus [66]. Details were available across 11 publications, extracted into 15 records, for ELISAs designed to detect antibodies for Schmallenberg virus or Akabane virus. Six commercially available ELISAs were described (Table 5). General designs for commercial antibody tests included competition and indirect ELISA formats utilising N protein (n = 3), Gc protein (n = 1) or whole virus antigens (n = 2) to detect antibodies. An in-house ELISA was described in four publications (n = 1 for each) [67,68,69,70].
To achieve WOAH Stage 1 validation, hyperimmune serum against known viruses were utilised in the ELISA development to demonstrate analytical specificity. For Stage 2 validation, most publications, utilised reference samples taken from field submissions that were tested using a VNT for Schmallenberg virus, Akabane virus or Shuni virus (Table 2). In addition to the validation testing completed for the commercial assays (Table 5), an ELISA test to detect specific viruses in the Simbu serogroup has shown overall diagnostic specificities of 84.56%, 94.68%, and 89.39% and sensitivities of 89.08%, 69.44%, and 84.91% for Schmallenberg virus, Akabane virus, or Shuni virus, respectively [66]. The test was conducted on a sample of 477 known negative, 238 known Schmallenberg virus antibody positive, 36 known Akabane virus antibody positive, and 53 known Schmallenberg virus antibody positive cases.
3.5. Monitoring the distribution of viruses in the Simbu group
Across the 91 publications describing the distribution of SGVs, Schmallenberg virus (n = 51) was the most studied, followed by Akabane virus (n = 32) and Aino virus (n = 21) (Table 2). Most studies were conducted within a country (n = 59) or local region (n = 24) rather than across multiple countries (n = 6) or continents (n = 2). Overall, the monitoring methods described were conducted across 56 countries and covered all continents except North America. The countries most frequently studied were Australia (n = 9), Korea (n = 9), Belgium (n = 8), Japan (n = 8), Germany (n = 5), Netherlands (n = 5), Türkiye (n = 5), China (n = 4), France (n = 4) and Ireland (n = 4).
Most publications described active sample collection (n = 70) rather than the opportunistic use of existing samples (n = 20) collected in surveillance for other viral pathogens. One publication described a mixed approach to SGV surveillance. Most surveys (n = 66) aimed to detect only one virus. Some investigated a combination of SGVs including other animal pathogens (n = 20). In some cases, the publication described a general arbovirus survey where a SGV was detected, without specifying the virus of interest (n = 5).
In over half the publications, samples were collected from a single mammalian host species: bovine (n = 35), ovine (n = 5), caprine (n = 3), equine (n = 2), and porcine (n = 1) or samples of vectors, Culicoides spp. (n = 2). In 43 publications, samples were collected from a combination of animals and insect species. This included a combination of two or more ruminant species (n = 27), or a combination of a ruminant and other host species such as camelids, swine or equine (n = 6), or finally a combination of a mammalian species and Culicoides (n = 10).
Overall, a single sample type was also most often collected: serum (n = 67), milk (n = 6) or Culicoides (n = 2). Occasionally, a combination of these sample types (n = 8) including, aborted foetal tissues (n = 4), EDTA treated blood (n = 3) and vaginal swabs (n = 1) were utilised. Milk yield data from dairy cattle was used in a novel approach to provide an early-warning system for Schmallenberg virus transmission [76].
Both agent and antibody assays were utilised as part of the efforts to detect SGVs and determine the distribution of these viruses. Most publications described the use of only one type of assay in their testing method. In these circumstances, antibody tests such as ELISAs (n = 32) and VNTs (n = 23) were more often used rather than agent-specific PCRs (n = 2). Less than a third of publications (n = 24) utilised a combination of two assay types to both detect and distinguish the SGV surveyed, including: (A) Multiple antibody tests – ELISAs with VNT (7/24) or haemagglutination inhibition tests (HIT) with VNT (3/24); or (B), a combination of both antibody and agent test types – ELISAs with PCR (9/24) or VNTs with PCR (2/24); or (C), a specific antibody test and general assay such as a VNT with virus isolation (3/24). More than three tests were combined with varying permutations of assay types in 10 publications. These combinations utilised an agent specific test, such as PCR (7/10), with antibody assays, including ELISA (6/10), VNT (9/10), HIT (1/10), IFAT (2/10), AGID (2/10) and CFT (1/10), and general test types such as virus isolation (4/10) and electron microscopy (1/10).
Only one publication conducted a survey with the purpose to demonstrate proof of freedom. Instead, almost all publications were conducted with the intent of virus detection. As such, most samples were collected at a single time point (62) rather than repeated (27). For repeated sampling, a monthly period was the most frequent interval (16), followed by quarterly (5) and yearly (2). Daily (1) and weekly (1) sampling was uncommon. Sampling was also repeated following specific circumstances such as calving (2). The information on sampling interval was not available for 2 publications. Autumn was most often the season in which sampling commenced (15/48), closely followed by spring (13/48), summer (12/48) and then winter (8/48). The median sampling period was 14.38 months (minimum 1 month, mean 6 months, maximum 108 months) described from 32/91 publications.
Overall, the median total number of animals sampled for over a third of all publications (71/91) was 507 (min. 9, Q1 239, mean 2285, Q3 1936, max. 21833). Sample size calculations were reported in almost one third of the publications (25/91). Where sample size calculations were made, the median sample size was 1301 (min. 42, Q1 361, mean 3647, Q3 4670, max. 21833). The assumptions in survey design were most varied based on the anticipated prevalence (median 20%, range 1-98%), whereas error (median 5%, range 1-15%), and confidence (median 95%, range 95-99%) were within a narrow range. Based on the seroprevalence measure reported across 57 publications, overall, the median seroprevalence was 18.8 % (min. 0, Q1 2.9%, mean 28.9%, Q3 48%, max. 100%). The mean seroprevalence was highly varied and has been presented by country, animal species and virus in Supplementary Figure S1.
4. Discussion
Orthobunyaviruses in the Simbu group can pose a significant threat to livestock production. Emergence of these viruses, in particular Akabane virus and Schmallenberg virus, in naïve populations has led to epidemics with profound animal health and welfare impacts. This scoping review covers almost five decades of research on SGVs and presents the anticipated disease presentations of the veterinary-important orthobunyaviruses in livestock, with considerations to the published diagnostic assays and surveillance strategies used to monitor the distribution of Simbu group viruses. The findings justify the need to revisit these viruses as re-emerging pathogens with significant animal health impacts.
This review examined literature from 1973 to 2022. The peak output years of publications were associated with the emergence of epidemics caused by SGVs (Figure 3). This includes the publications from the 1970-80s, associated with the epidemics of Akabane virus in Australia, Japan and Israel, and from 2011 in the decade following the emergence of Schmallenberg virus across Europe [15]. The disease epidemics associated with these SGVs are not the only explanation for the expansion of publications from 2011 (Figure 3). More viruses in this serogroup are being recognised as causes of disease in livestock. For example, Shuni virus was first isolated from a healthy cow in Nigeria in 1966 [77]. Reports of neurological disease in horses associated with Shuni virus were much later investigated in South Africa in 2009 [16]. Shuni virus has been linked to congenital malformations and reproductive disease in cattle, sheep, and goats in Israel between 2014-15 [36] and also caused neurological disease in a 16-month-old heifer in Israel in 2016 [35]. Peaton virus was first isolated from a healthy cow in Australia in 1979 [78] and was recently recognised as a cause of congenital malformations in calves in both Japan and Israel in 2018-2019 [39,79]. Finally, Shamonda virus was first isolated from cattle in Nigerian markets in 1965 [77]. Serological evidence suggests that Shamonda virus is a cause of congenital malformations in calves in Japan in the early 2000’s [80,81]. More recently, Shamonda virus has been detected in cases of febrile illness associated with abortions in goats in South Africa in 2023 [82]. The detection of a Shamonda-like isolate during surveillance activities in NSW provides the justification to revisit the SGVs and investigate its potential impacts on livestock health in Australia.
Congenital malformations in ruminants, particularly in cattle, was identified as the most frequent disease manifestation of viruses within the Simbu group. Gestational age determines the form of congenital malformations observed (Table 4). The pathogenesis of congenital disease was determined during the outbreak investigations of Akabane virus in Australia and Japan [83,84] and is well characterised by experimental infection studies [23,24,28,29,30,32]. This observation was further confirmed during the Schmallenberg virus epidemic that occurred across Europe [85]. Infection before embryonic attachment, may also cause embryo loss resulting in observations of reproductive disease for the adult animal [86,87]. Following foetal infection, neuroglial cells appear to be the sites of virus predilection and neuronal cells in the spinal ventral horn [88]. If the developing foetus survives, this results in pathological changes in the brain and spinal cord. If infected in mid-gestation, the lesions in central nervous system also result in musculoskeletal changes due to the lack of movement in the developing foetus. The developing skeletal muscle may also be infected and is often atrophied and replaced by adipose tissue [23]. In the late gestation and perinatal period, following the development of immunocompetence, infection may lead to inflammatory changes in the central nervous system. If the neonate is not aborted, survives to term and is not stillborn, encephalomyelitis may be observed with neurological signs.
While reviewing the frequency of clinical signs and necropsy findings for each virus within the Simbu group, it became evident that orthobunyaviruses in this serogroup exhibit varying propensities for foetal pathology (Table 4). Akabane virus is more often associated with congenital malformations, while Schmallenberg virus is found to frequently affect reproductive outcomes. In previous experimental infection challenges that utilised different strains of the same named virus within this serogroup, it was shown that there could be varying propensities for disease expression. The differences in pathogenicity of strains are most often reported for Akabane virus isolates. A mouse model experiment has demonstrated the differences in the neuroinvasiveness and neurovirulence of Akabane virus isolates [89]. These strain differences were also observed in a comparison of teratogenicity observed in ovine foetuses following separate experimental inoculations conducted in Japan and Australia [29,32]. The degree of foetal pathology observed was influenced by the number of passages in cell culture and the cell line used [38], and this method has been utilized in the production of a vaccine for Akabane virus [90,91]. The determinants of pathogenicity for SGVs have not been identified, although some elements of pathogenesis have been characterised, such as the ability to induce viraemia in a mammalian host [92,93]. Further investigation into the molecular relationships between the pathogenic and non-pathogenic viruses could help define the genetic determinants of disease. Knowledge of the drivers for pathogenicity would be especially useful for selecting safe vaccines and also allows for virulence to be monitored during production [94].
Out of the 33 named viruses in this serogroup, only six were identified as a cause of disease in livestock: Akabane virus (Orthobunyavirus akabaneense), Schmallenberg virus and Shamonda virus (Orthobunyavirus schmallenbergense), Aino virus (Orthobunyavirus ainoense), Shuni virus (Orthobunyavirus shuniense), and Peaton virus (Orthobunyavirus peachesterense). All are phylogenetically classified into Clade B. However, there are other named virus entities within this phylogenetic structure that are closely related but have no recognised disease manifestations in livestock (Table 3). This includes Tinaroo virus and Yaba-7 virus (both belonging to the species Orthobunyavirus akabaneense), Sathuperi virus and Douglas virus (both Orthobunyavirus schmallenbergense), and Kaikalur virus (Orthobunyavirus shuniense). To gain a better understanding of the risks posed by orthobunyaviruses, it is necessary to establish a standardized approach to determine their molecular, antigenic, and pathogenic characteristics. Investigating the molecular and antigenic relationships between the pathogenic and non-pathogenic viruses could help identify the determinants of disease. The hypothesised relationships could be investigated in experimental infection trials. Pathogenicity trials are possible for SGVs because a laboratory infection model has already been established in chicken embryos [95,96,97,98]. The suitability of the chicken embryo model is best demonstrated by the embryo pathology described in a comparison between Schmallenberg and Akabane [99]. Embryo mortality and pathological findings were akin to natural Schmallenberg virus and Akabane virus infection in ruminants [99]. Techniques in molecular epidemiology could complement traditional virological methods to help determine the threat posed by the Shamonda-like isolate detected during surveillance in the Australian orthobunyavirus ecosystem.
The type of diagnostic tests utilised also evolved with the advent of molecular technology and this resulted in some challenges when comparing early literature with the more recent Schmallenberg virus epidemic. A range of agent and antibody tests and sample types have been utilised to diagnose a SGV as the cause of congenital disease. The detection of a SGV in the foetus with congenital disease provides a strong association between virus and its role as the aetiological agent. However, as foetal immunocompetence develops, the virus may be cleared as the foetus becomes able to recognise and respond to pathogens [26]. As a result, the causative virus may not be isolated or detectable within foetal tissues [60]. Consequently, a negative virus culture or PCR result does not rule out an SGV infection as the cause of disease. This is especially the case for cattle due to the longer gestation period in comparison to the smaller ruminant species (Table 3). As the ruminant placenta prevents the transfer of maternal antibodies during gestation [26], antibodies detected in a foetus are presumed to be the outcome of in-utero infection [100]. Serological tests on the blood of a stillborn or aborted neonate can support a definitive diagnosis for congenital disease caused by SGVs that are known foetal pathogens. In the absence of foetal samples, blood from the associated dam should be submitted for testing to provide supportive evidence of previous viral infection.
The usefulness of blood is reflected in this review as it is the most common sample type utilised in both diagnostic test evaluation and monitoring surveys. Blood is necessary to detect SGVs in transmission, but the window to detect viraemia is short-lived [92,93]. Virus may be isolated from blood for a window of 1-4 days in the 1-6 days following experimental infection [93,101,102]. By PCR testing, this window of detection or viraemia may be up to 8 days post infection [103]. Meanwhile, following viraemia, neutralizing antibodies develop within a few days and may be detected for up to three years or longer [41,97,104]. The prolonged presence of antibodies following infection complicates the interpretation of serological tests based on a single sample for adult animals. This is because the antibodies detected may not indicate recent infection but rather exposure during previous transmission seasons. This is also complicated when animals may be infected with more than one orthobunyavirus in the Simbu group. It is difficult to distinguish the different viruses within the serogroup using serological tests because SGVs share antigenic similarities [105]. Further, the false positive results for the virus-specific SBV-S PCR demonstrates that agent detection tests also struggle to distinguish between SGVs [65]. However, using a broadly, or cross-reactive test, allows for the detection of a virus at the serogroup level, including potentially previously unrecognised SGVs.
The commercially available ELISAs (Table 5) and the virus-specific SBV-S PCR are cross-reactive tests, but it is important to note that this detection does not necessarily reflect the risks or likelihood of disease. Currently, additional test methods are utilised following the detection of a SGV to identify the specific viruses of veterinary importance. Novel reagents and technologies could be investigated to develop more efficient test methods. This includes utilising the Gc protein in ELISAs and investigating tools that allow for the simultaneous testing of antibodies against multiple proteins, such as the Luminex MAGPIX® system [66,106]. Assays that can distinguish the individual SGVs are necessary in outbreak investigations to correctly identify the aetiological agent. These new diagnostic tests must be validated appropriately, using known positive and negative reference samples that encompass all SGVs.
Surveillance programs monitoring SGV transmission can also help provide supportive evidence in diagnostic investigations. Seroprevalence surveys were identified as the main method to determine the geographical distribution of SGVs in livestock. The distribution of SGVs may be monitored by collecting a serum sample at a single time point from wildlife [107,108,109], animals held at a saleyard [42], or even opportunistically using samples collected during control programs for other animal pathogens [110]. However, due to the persistence of antibodies, serosurveys sampled at a single time point, without regard to the age or location of the sampled animal, are not a real-time representation of virus transmission and limit interpretation of the data over a defined time interval. This is particularly evident in the high level of variability observed in the reported seroprevalence by location, host species sampled and virus of interest (Supplementary Figure S1). As a result, the mean seroprevalence (Table 2) is presented to help provide rough estimates for sample size calculations for future surveys. The seasonal transmission of SGVs can be monitored through active surveillance programs that use a network of sentinel herds [102,111,112,113,114]. To effectively monitor SGV distribution using a sentinel network, it is critical to consider seasonal variations and geoclimatic regions when selecting locations for herds [19]. For a robust surveillance system, these active programs should be supplemented with strategic testing of diagnostic submissions.
This review provides the justification to include strategic testing for cases of neurological disease in livestock. If a pathological SGV was to emerge in a naïve animal population, it could be anticipated that this disease presentation could precede reports of congenital malformations. This is exemplified by the difference in seasonal observations with Shuni virus. The acute onset of neurological disease in horses follows infection during times of high vector activity resulting in disease primarily being reported in summer and autumn [16]. In contrast, for the SGVs causing congenital malformations in ruminants, there is an interval between infection and parturition, with disease more likely to be observed in autumn or winter. Infections are more likely to occur in young animals because of a lack of acquired immunity in previous transmission seasons. Neurological disease in the post-natal period was observed preceding the observations of congenitally malformed calves and lambs for Akabane virus [35,36,41,42]. Young horses were also more often reported with neurological signs following Shuni virus infection [45]. All animals showing consistent clinical neurological signs, during favourable seasonal conditions for virus transmission, should be investigated to ensure SGV-associated disease is not missed.
While rare, horses and adult cattle can develop a fatal neurological disease following infection with Shuni virus or Akabane virus [16,35,45,46,47]. Diagnoses of neurological signs associated with SGVs could be missed because disease would occur following acute infection. This means that serological tests may not be able to diagnose disease because antibodies may not yet be present. Given the viral tropism for nervous tissue, the brain is the obvious organ of choice for PCR testing, but the collection of brain requires necropsy. This sample type would also be problematic to collect in regions with other viruses of high zoonotic consequences, such as for horses in Australia with concerns for Hendra virus (Henipavirus hendraense). In these circumstances, agent detection on blood or cerebral spinal fluid may present an alternative diagnostic approach. This approach was successful in retrospectively confirming viral infection with Shuni virus in horses with a consistent clinical presentation [16]. Alternatively, demonstration of seroconversion would confirm recent infection. This strategic testing of appropriate diagnostic cases aids passive surveillance to help strengthen existing monitoring of SGVs.
Publication bias and/or the lack of surveillance activity could result in gaps in the distribution and diversity of SGVs. This could explain the apparent absence of Clade B SGVs in southeast Asia and North America (Figure 1). The absence of a Clade B SGV in southeast Asia is unlikely. Recently, antibodies against Schmallenberg virus were detected in serum collected from small ruminants in Malaysia [115]. However, because the commercial ELISA utilised in this study was designed on the N protein, the antibodies cannot be distinguished from other SGVs. Nonetheless, the absence of a Clade B virus does not appear likely in southeast Asia given the neighbouring regions have reported SGVs from both Clades. Given the geographical isolation, the observations of an absence of a Clade B SGV in North America may be valid. However, this difficult to evaluate because no publications of surveillance strategies from North America were assessed in this review. Additionally, the presence of a distantly related orthobunyavirus, Cache Valley virus (Orthobunyavirus cacheense), that also causes congenital defects in ruminants, could mask the emergence of a teratogenic SGV [116].
Restrictions applied during the systematic screening process of this scoping review has led to several limitations. Publications were screened to include only original research, in English language with relevance to livestock species of interest. The restriction to English-language publications meant that the experimental infection of pregnant pigs with Akabane virus (Supplementary Table S1), that were written in Japanese and the investigation of swine as a reservoir for Schmallenberg virus in French were not included [117,118]. In applying restrictions to original research, this review only evaluated assays with established development or validation pathways. This meant that diagnostic assays that have no evaluation pathways, like virus isolation and genomic sequencing were not included. These methods are critical virological techniques and are essential when the virus of interest may be unknown. Finally, the focus on livestock species resulted in publications describing vector distribution to be excluded. As arboviruses, vectors play an important role in the epidemiology of the diseases caused by SGVs. While reviewing the identified literature Culidoides spp. midges were identified as a sample type for diagnostic test and surveillance programs and their relevance and significance in these areas cannot be ignored.
It was also surprising that the field of molecular epidemiology and its associated tools were not identified during the review of surveillance methods despite its tremendous growth in the last decade. This could be because allocation to a theme within this review was based on the study objectives of animal health impacts, diagnostic tests or surveillance strategies. This focus meant that the details on the molecular characterisation of SGVs in the publications identified by our search strategy, were not extracted for analysis. This is a significant limitation in this review as molecular epidemiology allows for a better understanding of the distribution and regionalization of SGVs and how new viruses may have evolved in a particular region.
5. Conclusions
The Simbu group of viruses may cause severe livestock disease, which usually manifest as congenital malformations, reproductive loss and neurological disease. Although there can be a range of clinical presentations, clusters of congenital malformations, characterised by arthrogryposis and hydranencephaly, should raise a high level of suspicion of the involvement of a SGV. Among the 33 named orthobunyaviruses in the SGV subgroup, six have been identified to cause animal diseases: Akabane virus, Schmallenberg virus, Aino virus, Shuni virus, Peaton virus, and Shamonda virus. Disease diagnosis is complicated by the timing of foetal immunocompetence and test cross-reactivity. To demonstrate assay specificity, tests for both agent and antibody should be validated with reference samples that encompass all the SGVs. While broadly reactive tests are available to detect SGVs, novel diagnostic tools should be investigated so that individual viruses may be identified in a rapid and cost-effective manner. Tests must also be validated for surveillance purposes as serosurveys remain the main stay of monitoring methods, and an understanding of the spatial-temporal transmission must rely heavily on longitudinal sampling of sentinel herds. Finally, further research is required to help determine the disease risk of novel orthobunyaviruses and how they could challenge current diagnostic and surveillance capabilities.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1,
- Supplementary Table S1: The distribution, insect vector, host range, and disease observations for the 33 viruses within the Simbu serogroup as arranged by their species designation;
- Supplementary Table S2: Histopathological details extracted from publications describing the animal health outcomes following natural or experimental infection of a Simbu group virus in livestock;
- Supplementary Table S3: PCR details extracted from publications describing the molecular assays developed and validated for the detection of Simbu group viruses
- Supplementary Figure S1: Mean seroprevalence extracted from the seroprevalence studies describing the distribution and diversity of SGVs by A) country, B) host species , and C) virus.
- RepRows function to match variables entered as a comma-separated list and extract the details into multiple records for the same publication.
Author Contributions
Conceptualization, T.O.C. and J.A.L.M.L.T.; methodology, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T.; software, T.O.C.; validation, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T; formal analysis, T.O.C., and J.A.L.M.L.T.; investigation, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T.; resources, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T.; data curation, T.O.C.; writing—original draft preparation, T.O.C.; writing—review and editing, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T.; visualization, T.O.C., P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T.; supervision, P.M.H., D.S.F., P.D.K., and J.A.L.M.L.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to express their gratitude to: Thomas Goodfellow, academic liaison librarian at the University of Sydney, for his assistance in refining our search strategy; and, Dr. Damian Collins, biometrician at the NSW Department of Primary Industries, for developing the 'repRows' function, which efficiently matches variables with record details in R. The function has been submitted with this manuscript with his permission.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Girard M, Nelson CB, Picot V, Gubler DJ. Arboviruses: A global public health threat. Vaccine. 2020;38(24):3989-94. Epub 2020/04/24. [CrossRef] [PubMed]
- Elliott, RM. Orthobunyaviruses: recent genetic and structural insights. Nature Reviews Microbiology. 2014;12(10):673-85. [CrossRef]
- Hughes HR, Adkins S, Alkhovskiy S, Beer M, Blair C, Calisher CH, et al. ICTV Virus Taxonomy Profile: Peribunyaviridae. Journal of General Virology. 2020;101(1):1-2. [CrossRef]
- Vasilakis N, Tesh RB, Popov VL, Widen SG, Wood TG, Forrester NL, et al. Exploiting the Legacy of the Arbovirus Hunters. Viruses. 2019;11(5). Epub 2019/05/28. [CrossRef] [PubMed]
- Calisher, CH. History, Classification, and Taxonomy of Viruses in the Family Bunyaviridae. In: Elliott RM, editor. The Bunyaviridae. Boston, MA: Springer US; 1996. p. 1-17.
- Walter CT, Barr JN. Recent advances in the molecular and cellular biology of bunyaviruses. Journal of General Virology. 2011;92(11):2467-84. [CrossRef]
- Briese T, Calisher CH, Higgs S. Viruses of the family Bunyaviridae: Are all available isolates reassortants? Virology. 2013;446(1-2):207-16. [CrossRef] [PubMed]
- Abudurexiti A, Adkins S, Alioto D, Alkhovsky SV, Avšič-Županc T, Ballinger MJ, et al. Taxonomy of the order Bunyavirales: update 2019. Archives of Virology. 2019;164(7):1949-65. Epub 2019/05/09. [CrossRef] [PubMed]
- Simmonds P, Adriaenssens EM, Zerbini FM, Abrescia NGA, Aiewsakun P, Alfenas-Zerbini P, et al. Four principles to establish a universal virus taxonomy. PLOS Biology. 2023;21(2):e3001922. [CrossRef]
- Jin H, Elliott RM. Mutagenesis of the L protein encoded by Bunyamwera virus and production of monospecific antibodies. Journal of General Virology. 1992;73(9):2235-44. [CrossRef]
- Kinney RM, Calisher CH. Antigenic Relationships among Simbu Serogroup (Bunyaviridae) Viruses. The American Journal of Tropical Medicine and Hygiene. 1981;30(6):1307-18. [CrossRef]
- Yanase T, Murota K, Suda Y. Whole-genome sequence analysis of a novel orthobunyavirus isolated in Japan in the 1980s. Archives of Virology. 2023;168(2):67. [CrossRef]
- Ladner JT, Savji N, Lofts L, Travassos da Rosa A, Wiley MR, Gestole MC, et al. Genomic and phylogenetic characterization of viruses included in the Manzanilla and Oropouche species complexes of the genus Orthobunyavirus, family Bunyaviridae. Journal of General Virology. 2014;95(5):1055-66. [CrossRef]
- Miura Y, Hayashi S, Ishihara T, Inaba Y, Omori T. Neutralizing antibody against akabane virus in precolostral sera from calves with congenital arthrogryposis hydranencephaly syndrome. Arch Gesamte Virusforsch. 1974;46(3-4):377-80. [CrossRef]
- Hoffmann B, Scheuch M, Höper D, Jungblut R, Holsteg M, Schirrmeier H, et al. Novel orthobunyavirus in cattle, Europe, 2011. Emerging Infectious Diseases. 2012;18(3):469-72. [CrossRef]
- van Eeden C, Williams JH, Gerdes TGH, van Wilpe E, Viljoen A, Swanepoel R, et al. Shuni virus as cause of neurologic disease in horses. Emerging Infectious Diseases. 2012;18(2):318-21. [CrossRef]
- De Regge, N. Akabane, Aino and Schmallenberg virus—where do we stand and what do we know about the role of domestic ruminant hosts and Culicoides vectors in virus transmission and overwintering? Current Opinion in Virology. 2017;27:15-30. [CrossRef]
- Murray, MD. Akabane epizootics in New South Wales: evidence for long-distance dispersal of the biting midge Culicoides brevitarsis. Australian Veterinary Journal. 1987;64(10):305-8. [CrossRef]
- Geoghegan JL, Walker PJ, Duchemin J-B, Jeanne I, Holmes EC. Seasonal Drivers of the Epidemiology of Arthropod-Borne Viruses in Australia. PLOS Neglected Tropical Diseases. 2014;8(11):e3325. [CrossRef]
- Tricco AC, Lillie E, Zarin W, O'Brien KK, Colquhoun H, Levac D, et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Annals of internal medicine. 2018;169(7):467-73. [CrossRef]
- Afonso A, Abrahantes JC, Conraths F, Veldhuis A, Elbers A, Roberts H, et al. The Schmallenberg virus epidemic in Europe-2011-2013. Preventive Veterinary Medicine. 2014;116(4):391-403. [CrossRef]
- World Organisation for Animal Health. Chapter 1.1.6. Principles and Methods of Validation of Diagnostic Assays for Infectious Diseases. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 20222018.
- Kurogi H, Inaba Y, Takahashi E, Sato K, Omori T, Miura Y, et al. Epizootic congenital arthrogryposis-hydranencephaly syndrome in cattle: Isolation of Akabane virus from affected fetuses. Archives of Virology. 1976;51(1-2):67-74. [CrossRef]
- Kirkland PD, Barry RD, Harper PA, Zelski RZ. The development of Akabane virus-induced congenital abnormalities in cattle. The Veterinary Record. 1988;122(24):582-6. [CrossRef]
- Gibney EH, Kipar A, Rosbottom A, Guy CS, Smith RF, Hetzel U, et al. The extent of parasite-associated necrosis in the placenta and foetal tissues of cattle following Neospora caninum infection in early and late gestation correlates with foetal death. International Journal for Parasitology. 2008;38(5):579-88. [CrossRef]
- Osburn, B. Ontogeny of immune responses in cattle. In: Morrison WI, editor. Ruminant immune system in health and disease. 1986.
- Bayrou C, Garigliany M-M, Sarlet M, Sartelet A, Cassart D, Desmecht D. Natural Intrauterine Infection with Schmallenberg Virus in Malformed Newborn Calves. Emerging Infectious Diseases. 2014;20(8):1327-30. [CrossRef]
- Hirashima Y, Kitahara S, Kato T, Shirafuji H, Tanaka S, Yanase T. Congenital malformations of calves infected with Shamonda virus, southern Japan. Emerging Infectious Diseases. 2017;23(6):993-6. [CrossRef]
- Parsonson IM, Della-Porta AJ, Snowdon WA. Congenital abnormalities in newborn lambs after infection of pregnant sheep with Akabane virus. Infection and Immunity. 1977;15(1):254-62. [CrossRef]
- Parsonson IM, Della-Porta AJ, Snowdon WA. Akabane virus infection in the pregnant ewe. 2. Pathology of the foetus. Veterinary Microbiology. 1981;6(3):209-24. [CrossRef]
- McCullagh, P. Interception of the development of self tolerance in fetal lambs. European Journal of Immunology. 1989;19(8):1387-92. Epub 1989/08/01. [CrossRef] [PubMed]
- Hashiguchi Y, Nanba K, Kumagai T. Congenital abnormalities in newborn lambs following Akabane virus infection in pregnant ewes. National Institute of Animal Health Quarterly. 1979;19(1-2):1-11. PubMed PMID: WOS:A1979HN75400001.
- Laloy E, Bréard E, Trapp S, Pozzi N, Riou M, Barc C, et al. Fetopathic effects of experimental Schmallenberg virus infection in pregnant goats. Veterinary Microbiology. 2017;211:141-9. [CrossRef]
- Porto WJN, Horcajo P, Kim PdCP, Regidor-Cerrillo J, Romão EA, Álvarez-García G, et al. Peripheral and placental immune responses in goats after primoinfection with Neospora caninum at early, mid and late gestation. Veterinary Parasitology. 2017;242:38-43. [CrossRef]
- Golender N, Bumbarov V, Assis I, Beer M, Khinich Y, Koren O, et al. Shuni virus in Israel: Neurological disease and fatalities in cattle. Transboundary and Emerging Diseases. 2019;66(3):1126-31. [CrossRef]
- Golender N, Brenner J, Valdman M, Khinich Y, Bumbarov V, Panshin A, et al. Malformations Caused by Shuni Virus in Ruminants, Israel, 2014-2015. Emerging Infectious Diseases. 2015;21(12):2267-8. [CrossRef]
- Moriwaki M, Miura Y, Hayashi S, Ishitani R. Histopathological findings of calves infected experimentally with Aino virus. National Institute of Animal Health quarterly. 1977;17(3):95-106. PubMed Central PMCID: PMC917129.
- Coverdale OR, Cybinski DH, St George TD. Congenital abnormalites in calves associated with Akabane virus and Aino virus. Australian Veterinary Journal. 1978;54(3):151-2. [CrossRef]
- Behar A, Leibovich BB, Edery N, Yanase T, Brenner J. First genomic detection of Peaton virus in a calf with hydranencephaly in Israel. Veterinary Medicine and Science. 2019;5(1):87-92. [CrossRef]
- Garigliany M-M, Hoffmann B, Dive M, Sartelet A, Bayrou C, Cassart D, et al. Schmallenberg Virus in Calf Born at Term with Porencephaly, Belgium. Emerging Infectious Diseases. 2012;18(6):1005-6. [CrossRef]
- Parsonson IM, McPhee DA, Della-Porta AJ, McClure S, McCullagh P. Transmission of Akabane virus from the ewe to the early fetus (32 to 53 days). Journal of Comparative Pathology. 1988;99(2):215-27. [CrossRef]
- Haughey KG, Hartley WJ, Della-Porta AJ, Murray MD. Akabane disease in sheep. Australian Veterinary Journal. 1988;65(5):136-40. [CrossRef]
- Saegerman C, Martinelle L, Dal Pozzo F, Kirschvink N. Preliminary survey on the impact of Schmallenberg virus on sheep flocks in South of Belgium. Transboundary and Emerging Diseases. 2014;61(5):469-72. [CrossRef]
- Liao YK, Lu YS, Goto Y, Inaba Y. The isolation of Akabane virus (Iriki strain) from calves in Taiwan. Journal of Basic Microbiology. 1996;36(1):33-9. [CrossRef]
- Motlou TP, Williams J, Venter M. Epidemiology of Shuni Virus in Horses in South Africa. Viruses. 2021;13(5). [CrossRef]
- Oem JK, Lee KH, Kim HR, Bae YC, Chung JY, Lee OS, et al. Bovine Epizootic Encephalomyelitis caused by Akabane Virus Infection in Korea. Journal of Comparative Pathology. 2012;147(2-3):101-5. [CrossRef]
- Lee JK, Park JS, Choi JH, Park BK, Lee BC, Hwang WS, et al. Encephalomyelitis Associated with Akabane Virus Infection in Adult Cows. Veterinary Pathology. 2002;39(2):269-73. [CrossRef]
- Lechner I, Wüthrich M, Meylan M, van den Borne BHP, Schüpbach-Regula G. Association of clinical signs after acute Schmallenberg virus infection with milk production and fertility in Swiss dairy cows. Preventive Veterinary Medicine. 2017;146:121-9. [CrossRef]
- Shaw AE, Mellor DJ, Purse BV, Shaw PE, McCorkell BF, Palmarini M. Transmission of Schmallenberg virus in a housed dairy herd in the UK. Veterinary Record. 2013;173(24):609. [CrossRef]
- Laloy E, Riou M, Barc C, Belbis G, Breard E, Breton S, et al. Schmallenberg virus: experimental infection in goats and bucks. BMC Veterinary Research. 2015;11. [CrossRef]
- Miyazato S, Miura Y, Hase M, Kubo M, Goto Y, Kono Y. Encephalitis of cattle caused by Iriki isolate, a new strain belonging to Akabane virus. Nippon juigaku zasshi The Japanese Journal of Veterinary Science. 1989;51(1):128-36. [CrossRef]
- Barrett D, O'Neill R, Sammin D, Clegg TA, More SJ. The impact of infection with Schmallenberg virus on weaning rate in Irish sheep flocks. Preventive Veterinary Medicine. 2015;122(3):332-8. [CrossRef]
- Martinelle L, Dal Pozzo F, Gauthier B, Kirschvink N, Saegerman C. Field veterinary survey on clinical and economic impact of schmallenberg virus in Belgium. Transboundary and Emerging Diseases. 2014;61(3):285-8. [CrossRef]
- Horikita T, Yoshinaga S, Okatani AT, Yamane I, Honda E, Hayashidani H. Loss of milk yield due to Akabane disease in dairy cows. Journal of Veterinary Medical Science. 2005;67(3):287-90. [CrossRef]
- Häsler B, Alarcon P, Raboisson D, Waret-Szkuta A, Rushton J. Integration of production and financial models to analyse the financial impact of livestock diseases: A case study of Schmallenberg virus disease on British and French dairy farms. Veterinary Record. 2015;2(1). [CrossRef]
- Waret-Szkuta A, Alarconl P, Häsler B, Rushton J, Corbière F, Raboisson D. Economic assessment of an emerging disease: The case of Schmallenberg virus in France. OIE Revue Scientifique et Technique. 2017;36(1):265-77. [CrossRef]
- Shirafuji H, Yazaki R, Shuto Y, Yanase T, Kato T, Ishikura Y, et al. Broad-range detection of arboviruses belonging to Simbu serogroup lineage 1 and specific detection of Akabane, Aino and Peaton viruses by newly developed multiple TaqMan assays. Journal of Virological Methods. 2015;225:9-15. [CrossRef]
- Fischer M, Schirrmeier H, Wernike K, Wegelt A, Beer M, Hoffmann B. Development of a pan-Simbu real-time reverse transcriptase PCR for the detection of Simbu serogroup viruses and comparison with SBV diagnostic PCR systems. Virology Journal. 2013;10. [CrossRef]
- De Regge N, Madder M, Deblauwe I, Losson B, Fassotte C, Demeulemeester J, et al. Schmallenberg virus circulation in Culicoides in Belgium in 2012: Field validation of a real time RT-PCR approach to assess virus replication and dissemination in midges. PLoS ONE. 2014;9(1). [CrossRef]
- De Regge N, Van Den Berg T, Georges L, Cay B. Diagnosis of Schmallenberg virus infection in malformed lambs and calves and first indications for virus clearance in the fetus. Veterinary Microbiology. 2013;162(2-4):595-600. [CrossRef]
- Camarão AAR, Swanepoel R, Boinas F, Quan M. Development and analytical validation of a group-specific RT-qPCR assay for the detection of the Simbu serogroup orthobunyaviruses. Journal of Virological Methods. 2019;271. [CrossRef]
- Golender N, Bumbarov VY, Erster O, Beer M, Khinich Y, Wernike K. Development and validation of a universal S-segment-based real-time RT-PCR assay for the detection of Simbu serogroup viruses. Journal of Virological Methods. 2018;261:80-5. [CrossRef]
- Gurau MR, Baraitareanu S, Manescu MA, Popp MC, Danes D. Studies concerning the optimisation of reverse transcription polymerase chain reaction technique of pan-Simbu virus group. Scientific Bulletin Series F Biotechnologies. 2016;20:329-32. PubMed PMID: CABI:20173026101.
- Wernike K, Beer M. International proficiency trial demonstrates reliable Schmallenberg virus infection diagnosis in endemic and non-affected countries. PLoS ONE. 2019;14(6). [CrossRef]
- Aebischer A, Wernike K, Hoffmann B, Beer M. Rapid genome detection of Schmallenberg virus and bovine viral diarrhea virus by use of isothermal amplification methods and high-speed real-time reverse transcriptase PCR. Journal of Clinical Microbiology. 2014;52(6):1883-92. [CrossRef]
- Wernike K, Aebischer A, Sick F, Szillat KP, Beer M. Differentiation of antibodies against selected Simbu serogroup viruses by a glycoprotein Gc-based triplex ELISA. Veterinary Sciences. 2021;8(1):1-14. [CrossRef]
- Tsuda T, Yoshida K, Yanase T, Ohashi S, Yamakawa M. Competitive enzyme-linked immunosorbent assay for the detection of the antibodies specific to Akabane virus. Journal of Veterinary Diagnostic Investigation. 2004;16(6):571-6. [CrossRef]
- Blacksell SD, Lunt RA, White JR. Rapid identification of Australian bunyavirus isolates belonging to the Simbu serogroup using indirect ELISA formats. Journal of Virological Methods. 1997;66(1):123-33. [CrossRef]
- Näslund K, Blomqvist G, Vernersson C, Zientara S, Bréard E, Valarcher JF. Development and evaluation of an indirect enzyme-linked immunosorbent assay for serological detection of Schmallenberg virus antibodies in ruminants using whole virus antigen. Acta veterinaria Scandinavica. 2014;56:71. [CrossRef]
- Ungar-Waron H, Gluckman A, Trainin Z. ELISA test for the serodiagnosis of Akabane virus infection in cattle. Tropical Animal Health and Production. 1989;21(3):205-10. [CrossRef]
- Kittelberger R, McFadden AMJ, Kirkland PD, Hannah MJ, Orr D, Bueno R, et al. Evaluation of two commercial enzyme-linked immunosorbent assay kits for the detection of serum antibodies against Akabane virus in cattle. Journal of Veterinary Diagnostic Investigation. 2013;25(5):645-8. [CrossRef]
- Li X, Jing H, Liu X, Wang Q, Qiu S, Liu D, et al. Comparative evaluation of two commercial ELISA kits for detection of antibodies against Akabane virus in cattle serum. BMC Veterinary Research. 2019;15(1). [CrossRef]
- Pejaković S, Wiggers L, Coupeau D, Kirschvink N, Mason J, Muylkens B. Test selection for antibody detection according to the seroprevalence level of Schmallenberg virus in sheep. PLoS ONE. 2018;13(4). [CrossRef]
- Bréard E, Lara E, Comtet L, Viarouge C, Doceul V, Desprat A, et al. Validation of a Commercially Available Indirect Elisa Using a Nucleocapside Recombinant Protein for Detection of Schmallenberg Virus Antibodies. PLoS ONE. 2013;8(1). [CrossRef]
- Daly JM, King B, Tarlinton R, Gough KC, Maddison BC, Blowey R. Comparison of Schmallenberg virus antibody levels detected in milk and serum from individual cows. BMC Veterinary Research. 2015;11(56):(11 March 2015)-(11 March ). PubMed PMID: CABI:20153115888.
- Douglass AP, O'Grady L, McGrath G, Tratalos J, Mee JF, Barrett D, et al. Development of a syndromic surveillance system for Irish dairy cattle using milk recording data. Preventive Veterinary Medicine. 2022;204. [CrossRef]
- Causey OR, Kemp GE, Causey CE, Lee VH. Isolations of Simbu-group viruses in Ibadan, Nigeria 1964–69, including the new types Sango, Shamonda, Sabo and Shuni. Annals of Tropical Medicine and Parasitology. 1972;66(3):357-62. [CrossRef]
- St George TD, Cybinski DH, Filippich C, Carley JG. Isolation of three Simbu group viruses new to Australia. Australian Journal of Experimental Biology and Medical Science. 1979;57(6):581-2. PubMed PMID: BIOSIS:PREV198019052059.
- Matsumori Y, Aizawa M, Sakai Y, Inoue D, Kodani M, Tsuha O, et al. Congenital abnormalities in calves associated with Peaton virus infection in Japan. Journal of Veterinary Diagnostic Investigation. 2018;30(6):855-61. Epub 2018/09/12. [CrossRef] [PubMed]
- Yanase T, Maeda K, Kato T, Nyuta S, Kamata H, Yamakawa M, et al. The resurgence of Shamonda virus, an African Simbu group virus of the genus Orthobunyavirus, in Japan. Archives of Virology. 2005;150(2):361-9. [CrossRef]
- Hirashima Y, Sakaguchi Z, Okada D, Fujioka M, Kitahara S, Iwamoto J, et al. Retrospective survey of shamonda virus infection in sentinel and breeding cattle and in malformed calves in Kagoshima Prefecture. Journal of the Japan Veterinary Medical Association. 2017;70(11):729-34. [CrossRef]
- Walt Mvd, Rakaki ME, MacIntyre C, Mendes A, Junglen S, Theron C, et al. Identification and Molecular Characterization of Shamonda Virus in an Aborted Goat Fetus in South Africa. Pathogens. 2023;12(9):1100. PubMed PMID:. [CrossRef]
- Konno S, Moriwaki M, Nakagawa M. Akabane Disease in Cattle: Congenital Abnormalities caused by Viral Infection. Spontaneous Disease. Veterinary Pathology. 1982;19(3):246-66. [CrossRef]
- Hartley WJ, Desaram WG, Della-Porta AJ, Snowdon WA, Shepherd NC. Pathology of congenital bovine episootic arthrogryposis and hydranenecphaly and its relationship to Akabane virus. Australian Veterinary Journal. 1977;53(7):319-25. [CrossRef]
- Herder V, Wohlsein P, Peters M, Hansmann F, Baumgaertner W. Salient Lesions in Domestic Ruminants Infected With the Emerging So-called Schmallenberg Virus in Germany. Veterinary Pathology. 2012;49(4):588-91. [CrossRef]
- König P, Wernike K, Hechinger S, Tauscher K, Breithaupt A, Beer M. Fetal infection with Schmallenberg virus — An experimental pathogenesis study in pregnant cows. Transboundary and Emerging Diseases. 2019;66(1):454-62. [CrossRef]
- Steinrigl A, Schiefer P, Schleicher C, Peinhopf W, Wodak E, Bago Z, et al. Rapid spread and association of Schmallenberg virus with ruminant abortions and foetal death in Austria in 2012/2013. Preventive Veterinary Medicine. 2014;116(4):350-9. [CrossRef]
- Noda Y, Yokoyama H, Katsuki T, Kurashige S, Uchinuno Y, Narita M. Demonstration of Akabane virus antigen using immunohistochemistry in naturally infected newborn calves. Veterinary Pathology. 2001;38(2):216-8. [CrossRef]
- Murata Y, Uchida K, Shioda C, Uema A, Bangphoomi N, Chambers JK, et al. Histopathological Studies on the Neuropathogenicity of the Iriki and OBE-1 Strains of Akabane Virus in BALB/cAJcl Mice. Journal of Comparative Pathology. 2015;153(2-3):140-9. [CrossRef]
- Park B-K, Chang C-H, Lyoo Y-S, Son S-W, Lee P-S, Rhee J-C, et al. Studies on an attenuated live Akabane virus vaccine against Akabane disease. Research Reports of the Rural Development Administration (Suweon). 1992;34(2 VET.):20-6. PubMed PMID: BIOSIS:PREV199395108926.
- Kurogi H, Inaba Y, Takahashi E, Sato K, Akashi H, Satoda K, et al. An attenuated strain of Akabane virus: a candidate for live virus vaccine. National Institute of Animal Health Quarterly. 1979;19(1-2):12-22.
- Kraatz F, Wernike K, Hechinger S, Koenig P, Granzow H, Reimann I, et al. Deletion Mutants of Schmallenberg Virus Are Avirulent and Protect from Virus Challenge. Journal of Virology. 2015;89(3):1825-37. [CrossRef]
- Sick F, Breithaupt A, Golender N, Bumbarov V, Beer M, Wernike K. Shuni virus-induced meningoencephalitis after experimental infection of cattle. Transboundary and Emerging Diseases. 2021;68(3):1531-40. [CrossRef]
- Yang D-K, Kim H-H, Nah J-J, Choi S-S, Seok K-O, Kim S-Y, et al. Evaluation of Akabane vaccine strains based on molecular characterization. Journal of Preventive Veterinary Medicine. 2012;36(4):180-5. PubMed PMID: KJD:ART001754575.
- McPhee DA, Parsonson IM, Della Porta AJ, Jarrett RG. Teratogenicity of Australian Simbu serogroup and some other Bunyaviridae viruses: The embryonated chicken egg as a model. Infection and Immunity. 1984;43(1):413-20. [CrossRef]
- Kurogi H, Inaba Y, Akashi H, Takahashi E, Sato K, Satoda K, et al. Immune response of various animals to Akabane disease live virus vaccine. National Institute of Animal Health Quarterly. 1979;19(1-2):23-31. PubMed Central PMCID: PMC537650.
- Poskin A, Verite S, Comtet L, Van Der Stede Y, Cay B, De Regge N. Persistence of the protective immunity and kinetics of the isotype specific antibody response against the viral nucleocapsid protein after experimental Schmallenberg virus infection of sheep. Veterinary Research. 2015;46(1). [CrossRef]
- McClure S, McCullagh P, Parsonson IM, McPhee DA, Della-Porta AJ, Orsini A. Maturation of immunological reactivity in the fetal lamb infected with Akabane virus. Journal of Comparative Pathology. 1988;99(2):133-43. [CrossRef]
- Collins ÁB, Mee JF, Kirkland PD. Pathogenicity and teratogenicity of Schmallenberg virus and Akabane virus in experimentally infected chicken embryos. Veterinary Microbiology. 2018;216:31-7. [CrossRef]
- Parsonson IM, Della-Porta AJ, Snowdon WA. Developmental disorders of the fetus in some arthropod-borne virus infections. American Journal of Tropical Medicine and Hygiene. 1981;30(3):660-73. [CrossRef]
- Kurogi H, Inaba Y, Takahashi E, Sato K, Satoda K, Goto Y, et al. Congenital abnormalities in newborn calves after inoculation of pregnant cows with Akabane virus. Infection and Immunity. 1977;17(2):338-43. [CrossRef]
- St George TD, Standfast HA, Cybinski DH. Isolations of Akabane virus from sentinel cattle and Culicoides brevitarsis. Australian Veterinary Journal. 1978;54(12):558-61. [CrossRef]
- Wernike K, Eschbaumer M, Breithaupt A, Hoffmann B, Beer M. Schmallenberg virus challenge models in cattle: infectious serum or culture-grown virus? Veterinary Research. 2012;43(1):84. [CrossRef]
- Wernike K, Holsteg M, Sasserath M, Beer M. Schmallenberg virus antibody development and decline in a naturally infected dairy cattle herd in Germany, 2011-2014. Veterinary Microbiology. 2015;181(3-4):294-7. Epub 2015/11/01. [CrossRef] [PubMed]
- Wernike K, Aebischer A, Sick F, Szillat KP, Beer M. Differentiation of Antibodies against Selected Simbu Serogroup Viruses by a Glycoprotein Gc-Based Triplex ELISA. Veterinary Sciences. 2021;8(1):12. PubMed PMID:. [CrossRef]
- Hoste ACR, Ruiz T, Fernández-Pacheco P, Jiménez-Clavero MÁ, Djadjovski I, Moreno S, et al. Development of a multiplex assay for antibody detection in serum against pathogens affecting ruminants. Transboundary and Emerging Diseases. 2021;68(3):1229-39. [CrossRef]
- Varol, K. Investigation of Schmallenberg virus seroprevalence in Honamli goats from Burdur region. Kocatepe Veterinary Journal. 2022;15(1):115-9. [CrossRef]
- Jiménez-Ruiz S, Risalde MA, Acevedo P, Arnal MC, Gómez-Guillamón F, Prieto P, et al. Serosurveillance of Schmallenberg virus in wild ruminants in Spain. Transboundary and Emerging Diseases. 2021;68(2):347-54. [CrossRef]
- Yang D-K, Hwang I-J, Kim B-H, Kweon C-H, Lee K-W, Kang M-I, et al. Serosurveillance of Viral Diseases in Korean Native Goats (Capra hircus). Journal of Veterinary Medical Science. 2008;70(9):977-9. [CrossRef]
- de Souza Nunes Martins M, Pituco EM, Taniwaki SA, Okuda LH, Richtzenhain LJ. Schmallenberg virus: research on viral circulation in Brazil. Brazilian Journal of Microbiology. 2022;53(1):377-83. [CrossRef]
- St. George TD. A sentinel herd system for the study of arbovirus infections in Australia and Papua-New Guinea. Veterinary Science Communications. 1980;4(1):39-51. PubMed PMID: CABI:19800577999.
- Behar A, Friedgut O, Rotenberg D, Zalesky O, Izhaki O, Yulzary A, et al. Insights on Transmission, Spread, and Possible Endemization of Selected Arboviruses in Israel—Interim Results from Five-Year Surveillance. Veterinary Sciences. 2022;9(2). [CrossRef]
- Shin YK, Oem JK, Yoon S, Hyun BH, Cho IS, Yoon SS, et al. Monitoring of five bovine arboviral diseases transmitted by arthropod vectors in Korea. Journal of Bacteriology and Virology. 2009;39(4):353-62. [CrossRef]
- Abu Elzein EME, Al-Afaleq AI, Mellor PS, El-Bashir AM, Hassanein MM. Study of Akabane infection in Saudi Arabia by the use of sentinel ruminants. Journal of Comparative Pathology. 1998;119(4):473-8. [CrossRef]
- Jesse FFA, Paul BT, Hashi HA, Chung ELT, Sedek NIHBM, Lila MAM. Schmallenberg Virus (SBV) Infection Among Small Ruminants in Selected States of Peninsular Malaysia. Journal of Advanced Veterinary Research. 2022;12(1):62-7.
- Waddell L, Pachal N, Mascarenhas M, Greig J, Harding S, Young I, et al. Cache Valley virus: A scoping review of the global evidence. Zoonoses Public Health. 2019;66(7):739-58. Epub 2019/06/30. [CrossRef] [PubMed]
- Maruyama S, Maruyama F, Tanaka Y, Itoh H, Uematsu N. Experimental infection of pigs with Akabane virus. Uirusu Journal of Virology. 1983;33(2):131-3. [CrossRef]
- Zimmer J-Y, Saegerman C, Martinelle L, Losson B, Leroy P, Haubruge E, et al. Pig farms: reservoirs of vectors of Bluetongue and Schmallenberg viruses? Biotechnologie Agronomie Societe Et Environnement. 2014;18(4):480-7. PubMed PMID: WOS:000346244100002.
Figure 1.
The distribution of the orthobunyaviruses within the Simbu group; Clade A viruses were reported within the countries in yellow, Clade B viruses in countries in blue, and viruses from both clades were reported within countries in green.
Figure 1.
The distribution of the orthobunyaviruses within the Simbu group; Clade A viruses were reported within the countries in yellow, Clade B viruses in countries in blue, and viruses from both clades were reported within countries in green.
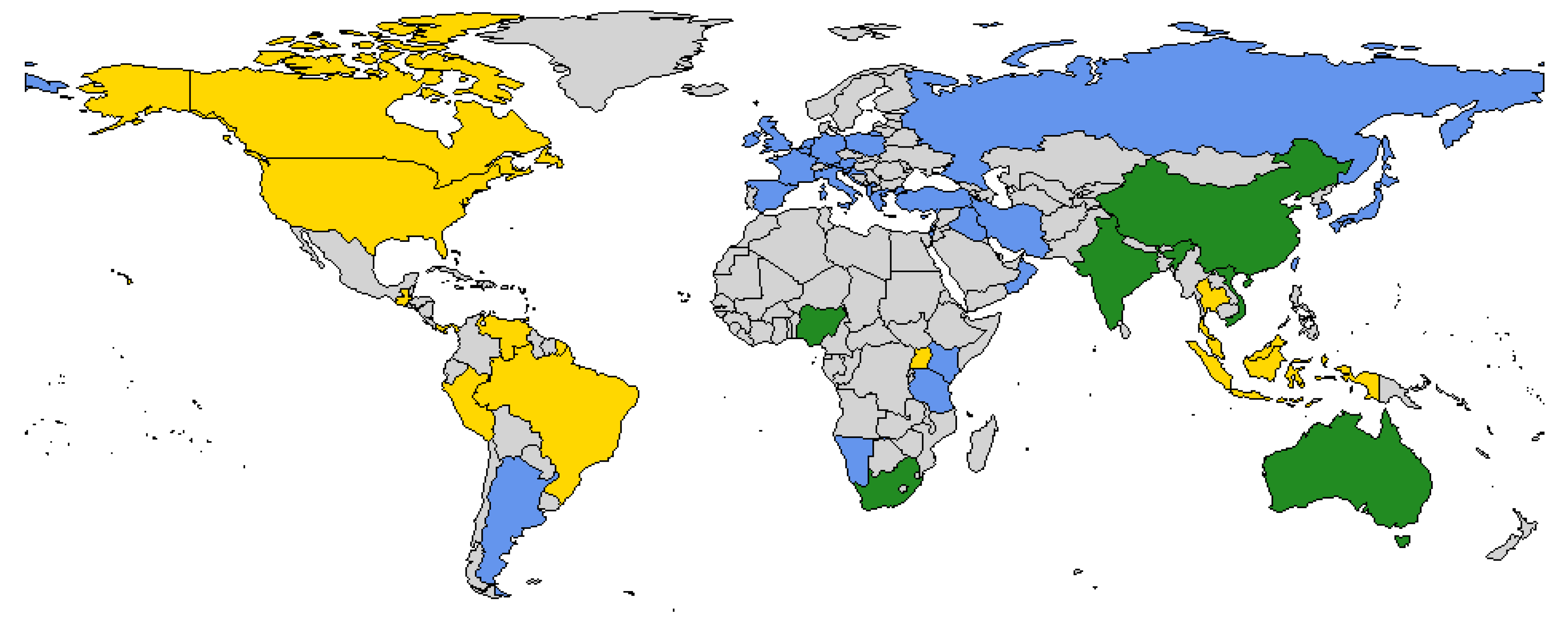
Figure 2.
Workflow used to screen for eligible literature in this scoping review to determine the animal disease, diagnostic tests, and monitoring methods for orthobunyaviruses in the Simbu group with veterinary importance. 1 Eight publications were incorporated from the first search strategy, which were not identified via the second approach.
Figure 2.
Workflow used to screen for eligible literature in this scoping review to determine the animal disease, diagnostic tests, and monitoring methods for orthobunyaviruses in the Simbu group with veterinary importance. 1 Eight publications were incorporated from the first search strategy, which were not identified via the second approach.
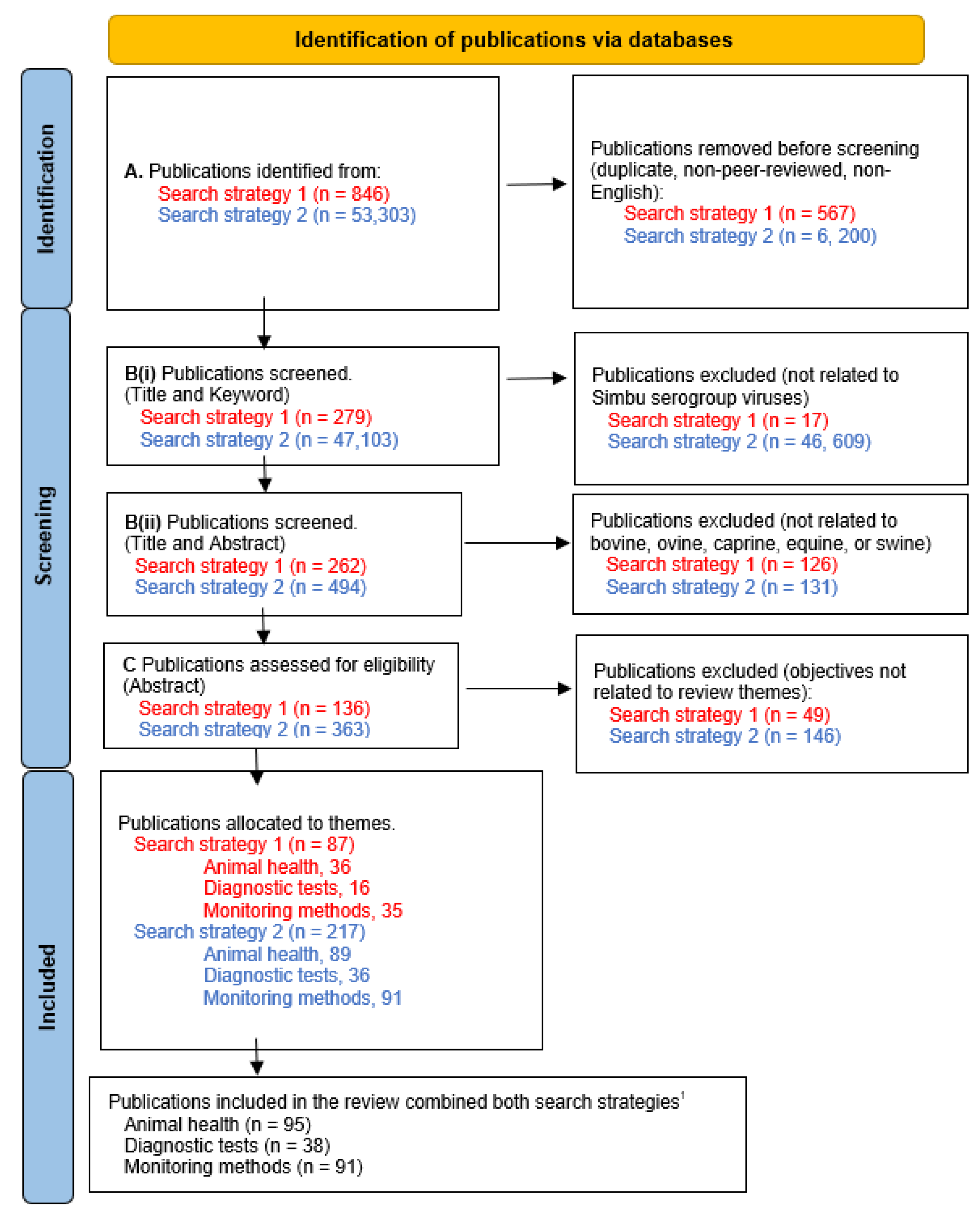
Figure 3.
The publication year and number of records for viruses in the Simbu group in the 321 records extracted from the 224 eligible publications retrieved from the scoping review protocol.
Figure 3.
The publication year and number of records for viruses in the Simbu group in the 321 records extracted from the 224 eligible publications retrieved from the scoping review protocol.
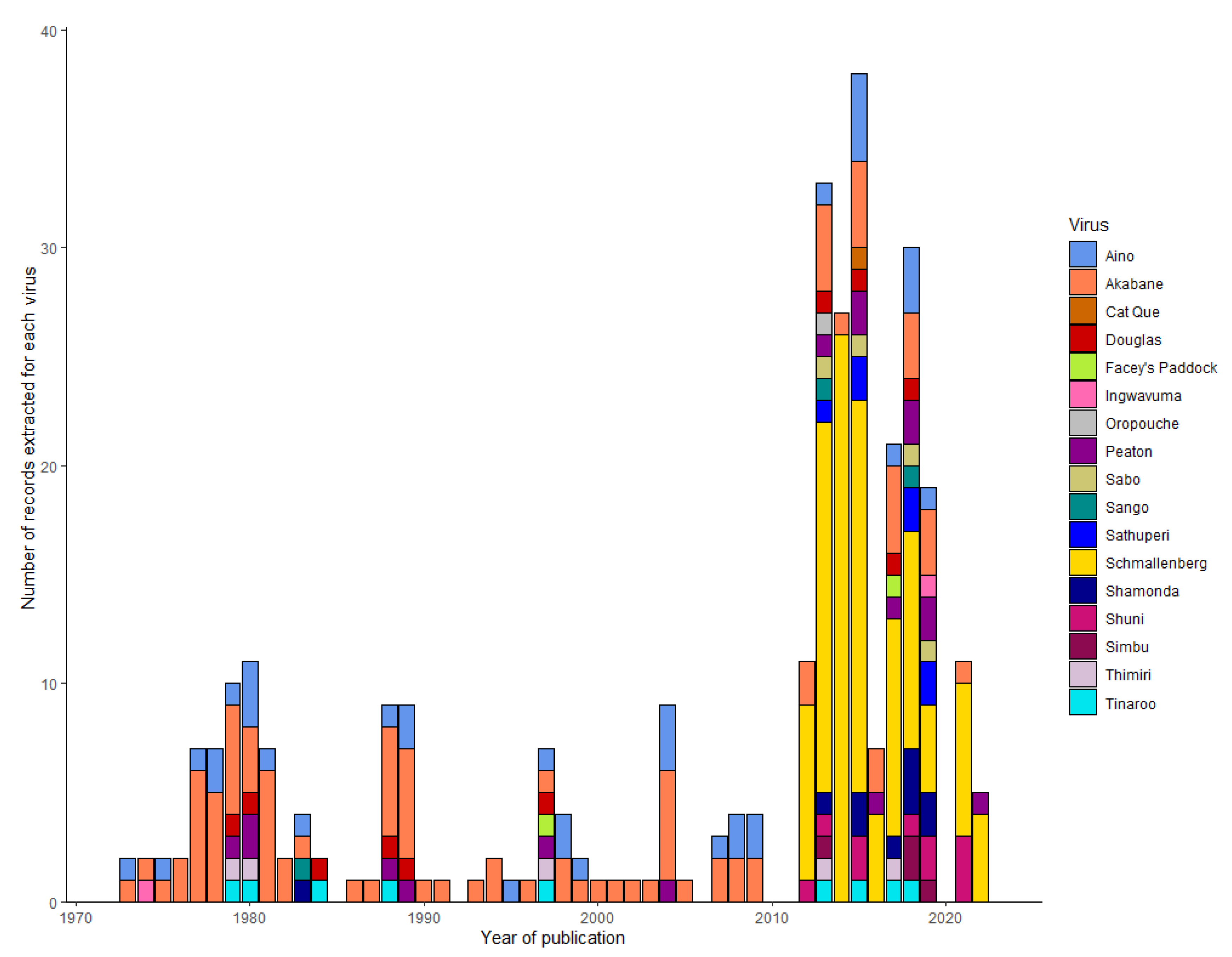
Figure 4.
The Simbu group viruses investigated in disease reports in different livestock species. The four diagnostic criteria were based on case definitions provided by European agencies during data collection for the Schmallenberg virus epidemic [21] with antibody detection only and agent detection only categories combined as ‘No clinical signs following viral infection’ for this figure.
Figure 4.
The Simbu group viruses investigated in disease reports in different livestock species. The four diagnostic criteria were based on case definitions provided by European agencies during data collection for the Schmallenberg virus epidemic [21] with antibody detection only and agent detection only categories combined as ‘No clinical signs following viral infection’ for this figure.
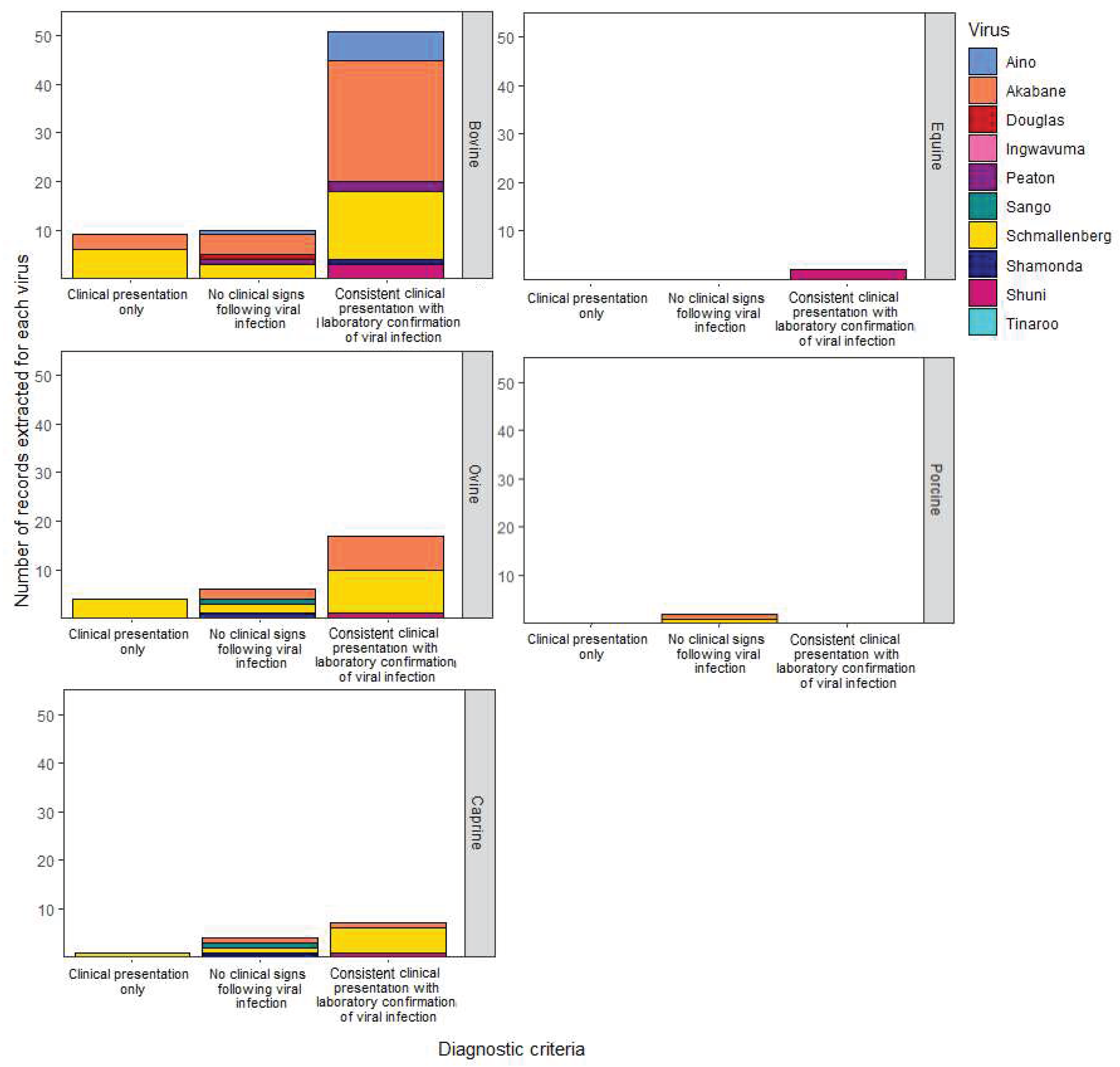
Table 2.
The number of records for viruses in the Simbu group extracted from the 224 publications identified for the scoping review protocol. A summary of the key animal health impacts, diagnostic assays and monitoring methods is also provided.
Table 2.
The number of records for viruses in the Simbu group extracted from the 224 publications identified for the scoping review protocol. A summary of the key animal health impacts, diagnostic assays and monitoring methods is also provided.
| Clade | Virus species | Virus names | Animal Health | Diagnostic Tests | Distribution | Total records by virus | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of records | Species | Key disease syndrome | No. of records | Highest level of WOAH validation | Validated assay types | No. of records | Mean seroprevalence | 95% CI | ||||
| A | Orthobunyavirus buttonwillowense | Buttonwillow virus | ||||||||||
| A | Orthobunyavirus catqueense | Cat Que virus | 1 | 60 | 50-70 | 1 | ||||||
| Oya virus | ||||||||||||
| A | Orthobunyavirus faceyense | Facey's paddock virus | 1 | No validation | 1 | - | - | 2 | ||||
| A | Orthobunyavirus ingwavumaense | Ingwavuma virus | 1 | Porcine | No disease | 1 | Stage 1 | PCR | 2 | |||
| A | Orthobunyavirus jatobalense | Jatobal virus | ||||||||||
| A | Orthobunyavirus manzanillaense | Manzanilla virus | ||||||||||
| Inini virus | ||||||||||||
| A | Orthobunyavirus mermetense | Mermet virus | ||||||||||
| A | Orthobunyavirus oropoucheense | Oropouche virus | 1 | Stage 1 | PCR | 1 | ||||||
| Iquitos virus | ||||||||||||
| Madre de Dios virus | ||||||||||||
| Perdoes virus | ||||||||||||
| Pintupo virus | ||||||||||||
| A | Orthobunyavirus thimiriense | Thimiri virus | 2 | Stage 1 | PCR | 3 | 0 | 0 | 5 | |||
| A | Orthobunyavirus utingaense | Utinga virus | ||||||||||
| Utive virus | ||||||||||||
| N/A | Orthobunyavirus oyoense | Oyo virus | ||||||||||
| B | Orthobunyavirus ainoense | Aino virus | 8 | Bovine | Congenital malformations, Neurological syndrome, Reproductive disease |
8 | Stage 1 | ELISA, PCR | 21 | 27.9 | 20.4-35.5 | 37 |
| B | Orthobunyavirus akabaneense | Akabane virus | 43 | Bovine, Ovine, Caprine Porcine |
Congenital malformations, Neurological syndrome, Reproductive disease No disease |
20 | Stage 2 | ELISA, PCR | 32 | 23.5 | 14.6-32.4 | 95 |
| Tinaroo virus | 1 | Bovine | No disease | 3 | Stage 1 | PCR | 5 | 24.6 | 20.9-28.3 | 9 | ||
| Yaba-7 virus | ||||||||||||
| B |
Orthobunyavirus schmallenbergense | Schmallenberg virus | 36 | Bovine, Ovine, Caprine Porcine |
Reproductive disease, Congenital malformations, Non-specific & Gastrointestinal No disease |
21 | Stage 2 | ELISA, PCR, VNT, LAMP, RPA, IFA | 51 | 34.9 | 27.0-42.8 | 108 |
| Shamonda virus | 2 | Bovine, Ovine, Caprine | Congenital malformations | 4 | Stage 1 | PCR | 4 | 34.1 | 3.1-65.1 | 10 | ||
| Douglas virus | 2 | Bovine | No disease | 3 | Stage 1 | PCR | 5 | 14.0 | 8.8-19.3 | 10 | ||
| Sathuperi virus | 4 | 3 | 34.9 | 8.2-61.5 | 7 | |||||||
| B | Orthobunyavirus peachesterense | Peaton virus | 3 | Bovine | Congenital malformations, Reproductive disease |
5 | Stage 1 | PCR | 9 | 21.6 | 15.7-27.5 | 17 |
| B | Orthobunyavirus saboense | Sabo virus | 3 | Stage 1 | PCR | 1 | 6.6 | 0-13.9 | 4 | |||
| B | Orthobunyavirus saboense | Sango virus | 1 | Ovine, Caprine | No disease | 2 | Stage 1 | PCR | 3 | |||
| B | Orthobunyavirus shuniense | Shuni virus | 6 | Bovine, Equine | Neurological, Congenital malformations, Reproductive |
4 | Stage 2 | ELISA, PCR | 10 | |||
| Kaikalur virus | ||||||||||||
| B | Orthobunyavirus simbuense | Simbu virus | 3 | Stage 1 | PCR | 1 | 9.5 | 1.9-17.1 | 4 | |||
| Para virus | ||||||||||||
Table 3.
Congenital malformations and foetal pathology caused by Simbu group viruses during in-utero development.
Table 3.
Congenital malformations and foetal pathology caused by Simbu group viruses during in-utero development.
| Gestational age at infection (days) | Species | ||
|
Bovine Endometrium attachment: 35 days Foetal immunocompetence: ≥120-150days Full term: 279–292days |
Ovine Endometrium attachment: 21 days Foetal immunocompetence: ≥64-87days Full term: 144-151days |
Caprine Endometrium attachment: 19 days Foetal immunocompetence: ≥40days Full term: 145-155 days |
|
| 0-28 | Embryonic death (day 28) | ||
| 29-59 |
At gestational age 30-36 days: Arthrogryposis Brachygnathism Brain agenesis or hydranencephaly Lung hypoplasia Microencephaly Muscle neuronal atrophy and degeneration Porencephaly Scoliosis Spinal cord agenesis or hypoplasia |
Foetal death (day 42) | |
| 60-90 | Hydranencephaly Porencephaly Myositis of skeletal muscles |
||
| 91-120 | Arthrogryposis Hydranencephaly Porencephaly Stillborn |
||
| 121-150 | Arthrogryposis | ||
| 151-180 | Arthrogryposis Stillborn |
||
| 181-210 | |||
| 211-240 | Degenerative encephalopathy | ||
| 241-270 | Clinical encephalopathy Stillborn |
||
| References | [23,24,25,26,27,28] | [29,30,31,32] | [33,34] |
Table 4.
The frequency of clinical signs and necropsy findings for each virus in the Simbu group viruses recognised as a cause of animal disease.
Table 4.
The frequency of clinical signs and necropsy findings for each virus in the Simbu group viruses recognised as a cause of animal disease.
| Akabane virus | Schmallenberg virus | Shuni virus | Aino virus | Shamonda virus | Peaton virus | |
|---|---|---|---|---|---|---|
| Total cases | 1675 | 553 | 48 | 17 | 15 | 2 |
| Clinical signs |
Congenital malformations: Unable to suckle, 140 Arthrogryposis, 118 Blind, 109 Scoliosis/ Vertebral deformities, 51 Brachgnathia inferior/ prognathism, 45 Torticollis, 37 Skeletal muscle atrophy, 19 Lameness, 13 Leg extension, 11 Weak, 9 Deaf, 7 Cleft palate, 3 Dwarf, 2 Cryptorchid, 1 Neurological Astasia/Dysstasia, 90 Dull, 47 Recumbency, 31 Convulsions/tremors, 24 Circling, 15 Nystagmus, 15 Hypersensitivity, 14 Collison with barrier, 9 Paralysis. 8 Pupillary contraction, 4 Altered consciousness, 2 Reproductive: Abortion, 76 Dystocia, 33 Perinatal mortality, 4 Mummified/autolyzed foetus, 2 Stillborn, 2 Non-specific: Tachypnoea,47 Pyreixa,46 Ataxia, 44 Tachycardia, 42 Salivation, 25 Reduced appetite, 13 Hypothermia, 4 Jaundice, 1 Gastrointestinal: Diarrhea, 3 |
Non-specific: Temporary decrease in milk yield, 350 Pyrexia, 227 Gastrointestinal: Diarrhea, 323 Congenital malformations: Arthrogryposis, 109 Scoliosis/ Vertebral deformities, 82 Brachgnathia inferior/ prognathism, 63 Torticollis, 46 Skeletal muscle atrophy, 1 Blind, 1 Cleft palate, 1 Neurological (only in animals surviving in-utero infection): Dull, 1 Proprioception deficits, 1, Strabismus, 1 Reproductive: Abortion, 50 Mummified/autolyzed foetus, 16 Perinatal mortality, 10 Stillborn, 2 |
Non-specific: Found dead, 12 Pyrexia, 9 Reduced appetite, 5 Anemia, 2 Leukopenia, 2 Neurological: Ataxia, 10 Recumbency, 7 Paralysis, 5 Convulsions/tremors, 2 Hypersensitivity, 1 Circling, 1 Weak, 1 Reproductive: Abortion, 3 Congenital malformations: Arthrogryposis, 1 Torticollis, 1 |
Congenital malformations: Arthrogryposis, 13 Torticollis, 1 Leg extension, 7 Neurological (only in animals surviving in-utero infection): Ataxia, 8 Astasia/Dysstasia, 8 Recumbency, 7 Hypersensitivity, 7 Circling, 7 Nystagmus, 1 Non-specific: Pyrexia, 7 Reduced appetite, 7 Reproductive Mummified/autolyzed foetus, 7 Stillborn, 4 Abortion, 1 |
Congenital malformations: Arthrogryposis, 12 Scoliosis/ Vertebral deformities, 11 Torticollis, 10 |
Congenital malformations: Arthrogryposis, 1 Scoliosis/ Vertebral deformities, 1 Blind, 1 Reproductive: Stillborn, 1 |
| Necropsy findings |
Congenital malformations: Micro-/hydranencephaly, 190 Hydrocephalus/ porencephaly, 90 Hypoplasia of the cerebellum, 56 Hypoplasia of the spinal cord, 55 Vestigial lungs/pulmonary hypoplasia, 25 Hypoplasia of the cerebrum, 3 |
Congenital malformations: Hypoplasia of the cerebellum, 86 Hypoplasia of the spinal cord, 67 Hydrocephalus/ porencephaly, 45 Micro-/hydranencephaly, 41 Hypoplasia of the cerebrum, 12 Cardiac ventricular septal defect, 3 Vestigial lungs/pulmonary hypoplasia, 1 Colonic atresia, 1 Ectopic cordis, 1 Unilateral hydronephrosis, 1 |
No reports |
Congenital malformations: Micro-/hydranencephaly, 6 Hypoplasia of the cerebellum, 3 |
Congenital malformations: Micro-/hydranencephaly, 2 Hypoplasia of the cerebrum, 1 |
Congenital malformations: Hydrocephalus/ porencephaly, 2 Hypoplasia of the cerebellum, 1 |
Table 5.
Details of six commercially available ELISAs for Akabane virus or Schmallenberg virus. The positive and negative agreement percentages were obtained with samples of a known status by comparison studies using a virus neutralisation test as the relative gold standard.
Table 5.
Details of six commercially available ELISAs for Akabane virus or Schmallenberg virus. The positive and negative agreement percentages were obtained with samples of a known status by comparison studies using a virus neutralisation test as the relative gold standard.
| ELISA | Format | Target (virus) | Sample | Sample status and size | Positive agreement (%) | Negative agreement (%) | Reference |
|---|---|---|---|---|---|---|---|
| ID-Screen Akabane ELISA (Innovative Diagnostics, Grabels, France) |
Competition | Whole virus (Akabane virus) | Serum | Positive (n = 378) Negative (n=334) |
96.0-98.9 | 99.7 | [71] |
| Serum | Positive (n = 153) Negative (n = 537) |
93.5 | 82.3 | [72] | |||
| Akabane ELISA kit (Chisso Corp, Yokohama) |
Competition | Gc glycoprotein (Akabane virus) | Serum | Positive (n = 378) Negative (n=334) |
78.0 | 100 | [71] |
| ID-Screen Schmallenberg virus Multi-species ELISA (Innovative Diagnostics, Grabels, France) | Competition | Nucleoprotein (Schmallenberg virus) | Serum | Positive (n = 45) Negative (n = 45) |
96-100 | 100 | [73] |
| ID-Screen Schmallenberg virus Indirect Multi-species ELISA (Innovative Diagnostics, Grabels, France) |
Indirect | Nucleoprotein (Schmallenberg virus) | Serum | Positive (n = 180) Negative (n = 1364) |
97.2 | 99.8 | [74] |
| IDEXX Schmallenberg virus Antibody Test Kit (IDEXX, Hoofddorp, Netherlands) |
Indirect | Nucleoprotein (Schmallenberg virus) | Serum | Positive (n = 45) Negative (n = 45) |
78-93 | 93-98 | [73] |
| Serum | Positive (n = 153) Negative (n = 537) |
80.4 | 93.5 | [72] | |||
| Svanovir Schmallenberg antibody ELISA (Svanova, Uppsala, Sweden) |
Indirect | Whole virus (Schmallenberg virus) |
Serum Bulk milk |
Positive (n = 82) Negative (n = 6) Positive (n = 82) Negative ( n = 6) |
94 98 |
50 100 |
[75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



