
Submitted:
09 January 2024
Posted:
10 January 2024
You are already at the latest version
Abstract
Emerging zoonotic diseases are one of the main threats to human and animal health and among the agents with the potential for zoonoses, those of bacterial origin have great relevance in Public Health. Rodents are considered one of the main reservoirs of pathogens that represent a risk to human health or animal species. In this study, we used massive 16S ribosomal RNA gene amplicon sequencing and microbial community ecology analyses to survey bacteria present in the spleen of six species of rodents in Panama, in order to identify bacterial taxa with zoonotic potential in the country. We found 3,352 bacterial Amplicon Sequence Variants (ASVs i.e. phylogenetic species) in the spleen of six rodents species surveyed (Liomys adspersus, Melanomys caliginosus, Mus musculus, Proechimys semispinosus, Rattus rattus, Zygodontomys brevicauda). This bacterial community was represented by 25 phyla, 55 classes, 140 orders, 268 families, and 508 genera. The three predominant phyla were Actinobacteria, Firmicutes, and Proteobacteria and the five predominant classes were Actinobacteria, Alpha- and Gammaproteobacteria, Bacilli, and Clostridia. The high abundance genera were seven: Acinetobacter, Bartonella, Cutibacterium, Enterococcus, Sarcina, Staphylococcus, and Wolbachia. Genera found with less abundance included Bradyrhizobium, Chryseobacterium, Clostridium, Corynebacterium, Lactobacillus, Pseudonocardia, Rhodococcus, and Sphingomonas. Some of these genera (high or low abundance) have clinical importance. Overall, this study contributes to generating information on the potential of different species of rodents as reservoirs of bacterial zoonosis in Panama.
Keywords:
rodents
; Bartonella
; spleen microbiota
; zoonosis
; reservoirs
1. Introduction
Emerging zoonotic diseases are one of the main threats to human and animal health [1,2,3,4,5,6]. According to recent estimations, there are about 1407 human pathogens, of which 58 % are zoonotic and 13% are classified as emerging or reemerging [7]. The infectious agents that involve emerging and reemerging zoonotic diseases include viruses, parasites, fungi, and bacteria among others [6,8,9]. Among these etiological agents of zoonoses, those with a bacterial origin, have great relevance in public health [6,10]. In this context, a report estimated that bacteria make up 538 and 54 (10%) [7], as well as know that the 26 major emerging and reemerging infectious diseases are bacterial origin; most of them originated from an animal, or other sources (i.e., water) and are considered to be zoonoses [3]. Zoonotic bacteria can be transmitted by different animals, rodents being considered major hosts of pathogens [10,11,12], and cause risk to human health or animal species; when they act as a reservoir or amplifying hosts for these microorganisms [13,14,15,16]. This participation of rodents in the epidemiology of human pathogenic bacteria is also favored because they constitute one of the most abundant and diversified groups of mammals [12,13,17,18,19], and because of their ability to successfully colonize a wide range of habitats, where they often interact with humans, but also with other animal species [18,20]. In addition, from the ecological perspective, the transmission of diseases by rodents also involves other factors, including alterations of the ecosystem (anthropogenic or natural), and changes in the number of available hosts and vectors [21,22,23].
For instance, several authors have also pointed out that the destruction of habitats as a consequence of human expansion and land use across the globe are among the main factors that have led to a defaunation that includes the global reduced abundant of mammals [24,25,26], which in turn causes an increase in the population of rodents and their pathogens, as observed in the indirect transmission systems of Bartonella spp. from Africa [24]. Therefore, an increase in the prevalence of rodent-borne diseases occurs as a result of changes in the abundance of susceptible hosts (rodents) and by closer human-rodent contact [14,27,28]. Therefore, taxonomic surveys of microbial communities in different species of rodents can contribute to understanding the natural occurrence and dynamics of pathogenic bacteria in them and this information is valuable in the development of more precise risk models for these diseases [14,28].
A review of the diversity of rodents that make up the wild mammal fauna of Panama has shown the existence of several species of rodents that in other countries have been reported as reservoirs and hosts of zoonotic agents, which are frequently close related in the human environment (synanthropic) [14,28]. Therefore, understanding the presence in Panama of rodents with the capacity to act as a reservoir for pathogenic bacteria will provide information on the epidemiological links in the country for the circulation and transmission of bacterial zoonoses. On the other hand, the continuous deforestation, land use, and unplanned urbanization in Panama have increased human contact with rodents, which has intensified the number of infections transmitted by rodents in the human population [27].
Based on the above and considering the increase in the cases of zoonoses in various rural and suburban areas with the consequent cases of death [27,29,30], it is important to carry out studies that allow the identification of pathogenic bacteria present in different rodents, in order to derive the prevalence, co-infection, and interaction of these bacteria and their distribution in natural populations of rodents, and in this way to know the potential that these animals have to directly or indirectly transmit zoonoses.
In this study, we used massive 16S ribosomal RNA gene amplicon sequencing and microbial community ecology analyses to survey bacteria present in the spleen of six species of rodents in Panama, in order to identify and list bacteria with zoonotic potential in the country. This study contributes to generating information on the potential of different species of rodents as reservoirs of bacterial zoonosis in Panama.
2. Materials and Methods
2.1. The Materials Rodent surveys and sample collection
Six species of rodents (Liomys adspersus, Melanomys caliginosus, Mus musculus, Proechimys semispinosus, Rattus rattus, Zygodontomys brevicauda) were collected from seven sites along Panama: Cañazas Chiriquí Grande (8°54’24” N, 82°14’19” W, CCG), Comarca Ngäbe-Buglé (8°46’11” N, 81°44’02” W, CNB), Divalá (8°25’12” N, 82°43’12” W), Mercado de Abasto de Curundú (8°59’12” N, 79°32’11” W, MAC), Mercado Público de David (8°26’00” N, 82°26’00” W, MPD), Oajaca Chiguirí Arriba (8°38’12” N, 80°12’22” W, OCA), and Panama Port Balboa (8°57’27” N, 79°33’40” W, PPB), (Table 1, Figure 1). The traps were placed according to Armien’s methodology [27]. Trapping grids were separated by a minimum distance of 500 m. All trapping grids were georeferenced with a Global Positioning System (GPS) receiver (Garmin 60 CSx) using the WGS 84 / UTM zone 17 N system, and their central points (centroids) were selected. For the distribution map, we used ArcMap 10.7.1. Mammals were handled according to recommendations by Mills and others [31]. The animals were killed with inhaled isoflurane, blood and samples of the spleen, liver, kidneys, heart, and lungs were collected in separate, labeled cryovials using clean sterilized instruments for each animal. All biologic samples were immediately placed into liquid nitrogen. Data collected for all individuals captured according to Armien et al. (2009).
2.2. DNA extraction
Total DNA was extracted from the spleens using the DNeasy Blood & Tissue Kit (Qiagen, Chatsworth, CA) following the manufacturer’s protocol, and with final DNA elution in 200 µl of AE buffer. A total of 26 samples were processed.
2.3. DNA amplification
Primers 799 F and 1115R [32,33] were used to amplify a portion of the V5 and V6 region of the 16S rRNA gene. We use these primers because we can reduce the number of chloroplasts in our sequences [32,33], knowing that rodents are consumers of many plants and a wide range of crops [34,35]. These primers contained read adapters for a second PCR needed for DNA library preparation. Each sample was amplified in triplicate PCR using 2.0 µl of DNA, 2.5 µl of 10x PCR buffer, 1.5 µl 25mM MgCl2, 2.0 µl of 10 mM dNTPs, 0.75 µl of 10 µM of primers (799 F and 1115 R), 0.5 µl of Taq DNA polymerase (Taq DNA polymerase kit of Qiagen (Product catalog 201203 (Qiagen, Valencia, CA, USA) and 15 µl of molecular grade water to obtain a total volume of 25 ul. Amplifications were conducted as follows: denaturation at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 45 seconds, annealing at 50 °C for 60 seconds, and elongation of 72 °C for 90 seconds, and final extension of 72 °C for 10 minutes. We run 2 µl of PCR products on an agarose gel to verify amplifications.
2.4. DNA Library preparation
The three PCR replicates of each sample were pooled and used as a template for a second PCR conducted with primers complementary to read primer adapters and containing indexes and flow cell adapters for Illumina® DNA sequencing by synthesis technology. Reactions were conducted as follows: 14.75 µl of molecular grade water, 2 µl of 10X Buffer, 1.5 µl of 25mM MgCl2, 2 µl of 10 mM dNTPs, 1 µl of 5 µM of each index primer (forward and reverse), 0.25 µl of Taq and 2 µl of pooled DNA template. PCR reaction started with a denaturation step of 94°C for 3 min followed by six cycles of 94 °C for 45 seconds, 50 °C for 60 seconds, 72°C for 1.5 min, and a final extension of 30 seconds at 72 °C. PCR samples were combined, concentrated, and later purified using Agencourt AMPure XP following the manufacturer´s instructions (Beckman Coulter International, Nyon, Switzerland). The DNA library was quantified using a Qubit fluorometer (Invitrogen, Waltham, Massachusetts) and quality was determined on a BioAnalyzer (Agilent Technologies, Santa Clara, California). Finally, the DNA library was sequenced on an Illumina MiSeq sequencing platform following a 2 x 250 bp Paired-End sequencing (Illumina Inc., San Diego, California).
2.5. Data analysis
Using QIIME 2TM bioinformatics pipeline [36,37,38], we dereplicated and quality filtered DNA sequences using Divisive Amplicon Denoising Algorithm (DADA2), [39,40]. Read 1 (R1) was used for subsequent analyses because the sequence quality for Read 2 was low. Continuously, we trained the sequence classifier for our specific region (V5 and V6) using the SILVA database (v.138 for bacteria, www.arb-silva.de) [41,42] that was used to taxonomically annotate amplicon sequence variants (ASVs). DNA sequences of mitochondria, chloroplasts, and unassigned bacterial taxa, as well as ASVs with less than 10 counts, were excluded for further analyses. Community ecology analyses were done using QIIME 2.0 as well as the R software for subsequent plotting [43].
2.6. Bacterial diversity, and community composition
For diversity estimation analysis, sequence data from each sample was rarefied to a depth of 3000. Alpha diversity from rodent species and localities was estimated using Faith’s phylogenetic diversity (Faith’s PD), and analyzed by non-parametric Kruskal-Wallis to determine statistical differences. Faith’s PD was used to compare bacterial diversity associated with M. musculus from two sites as this was the rodent species with a comparable number of samples from two sites. Beta diversity between species was estimated based on weighted UniFrac distance using PERMANOVA and ANOSIM analyses in the vegan package [44,45] and visualized using Principle Coordinates Analysis (PCoA) phyloseq [40], and ggplot2 package [46]. We did not estimate beta diversity between localities due to two sites (Comarca Ngäbe-Buglé and Divalá) were represented by only one rodent specimen (Table 1).
3. Results
We obtained a total of 403,188 sequence reads (per sample Min=6,828; Median=12,830; Maximum=41,263; Mean=15,507) from which 3,352 (ASVs, i.e., putative bacterial species) were detected. Rarefaction curves captured the majority of the bacterial diversity dataset in this study (Figure S1A, Figure S1B).
3.1. The spleen microbiome by rodent species and locality
The spleen microbiome of six species of rodents was composed of 3,352 ASVs. This bacterial community was represented by 25 phyla, 55 classes, 140 orders, 268 families, and 508 genera. The three predominant phyla were Actinobacteria, Firmicutes, and Proteobacteria (Figure 2A, Figure 2B). The five predominant classes were Actinobacteria, Alpha- and Gammaproteobacteria, Bacilli, and Clostridia (Figure 2A, Figure 2B). The most dominant genera were seven: Acinetobacter, Bartonella, Cutibacterium, Enterococcus, Sarcina, Staphylococcus, and Wolbachia. However, there were other bacterial taxa groups in less abundance such as Bradyrhizobium, Chryseobacterium, Clostridium, Corynebacterium, Lactobacillus, Pseudonocardia, Rhodococcus and Sphingomonas. Overall, some of them could be of clinical importance (Table 2, Figure 3A, Figure 3B, Figure 3C). No statistical significance in Alpha diversity was observed by species and locality (Kruskal-Wallis: H= 9.48, p > 0.05; H= 8.36, p > 0.05). Additionally, the spleen microbiome of M. musculus did not show significant differences between sites (Kruskal-Wallis: H= 1.18, p > 0.05) (Figure 4A, Figure 4B). No significant difference was observed in Beta diversity between species (Adonis statistic: R2= 1.78, p =0.057; Anosim statistic: R= 0.09, p > 0.05) (Figure 4C).
4. Discussion
A survey of the spleen microbiome of species of rodents in Panama represents an important opportunity to explore potential zoonosis bacteria in these small mammals, which are considered one of the main hosts of pathogens. Here, we assessed the bacterial community associated with six closely related species of small mammals in Panama using a massive 16S ribosomal RNA gene amplicon sequencing for the first time.
4.1. Bacterial composition in the spleen of six species of rodents, role in rodents and implications in zoonotic diseases
Overall, we found 3,352 ASVs associated with the six species of rodents. The most common and abundant bacterial taxa included classes Actinobacteria, Alpha- and Gamma-proteobacteria following Bacilli, Clostridia, and Bacteroidia. Here, genera such as Acinetobacter, Bartonella, Cutibacterium, Enterococcus, Sarcina, Staphylococcus, and Wolbachia showed high abundance either in species of rodents or sites. Some of these bacterial taxa are pathogens responsible for several zoonotic diseases in humans and animals (i.e., domestic and wild animals)[14,47,48,49,50], and some genera or some species belong to these genera are also found in rodents, which are major reservoirs [14,49,50]. For instance, Acinetobacter which we found in all our rodents species and sites, was previously isolated from laboratory mice and rodents [51], and is associated with infections and some species are showed a high drug-resistant [52]. Bartonella, which was only found in high abundance in Z. brevicauda, is a common bacteria found in rodents worldwide [14,50,53,54,55,56], and some species are associated with many clinical manifestations including endocarditis [57,58], neurologic disorders [59], meningitis [60] among others [50]. Additionally, a study showed evidence of the transmission of Bartonella from rodents by fleas [50]. Cutibacterium, which was found in L. adspersus, contains species (i.e., Cutibacterium acnes) that are known as skin infection bacteria [61,62]. Enterococcus was found in high abundance in M. caliginosus, this bacterium is found in different animals including free-living raptors [49], and it is a commensal organism, an opportunistic pathogen associated with the mortality of humans and animals [49,63]. On the other hand, previous studies showed that species such as Enterococcus faecalis were associated with small rodents [48,64], and it has caused inflammatory disease in mice [48]. Another observation is Sarcina that was found in high abundance in M. musculus. Studies have shown some species belonging to Sarcina (i.e., Sarcina ventriculi) is a gram-positive bacterium, able to survive in extremely low pH environments [65], and it is an important pathogen that is associated with a lethal disease in sanctuary chimpanzees [66]. Staphylococcus that was found in high abundance in L. adspersus. It contains some species such as Staphylococcus aureus, this species is a commensal bacteria of the human skin, and gastrointestinal tract, which causes infections [67]. Finally, Wolbachia, interestingly was found in the spleen of rodents, and it was found in high abundance in M. caliginosus. This bacterium is associated with insects [68,69,70,71], and has implications for ecology and reproduction in various insects [68,72,73,74].
This study is a first step in screening bacterial taxa with potential for zoonosis in the rodents surveyed in Panama. Although we found several pathogenic bacteria, more studies are needed to accurately estimate their potential for zoonosis in the country. Further research is also needed to assess the core microbiome associated with different species of rodents and which one have higher potential for zoonosis.
5. Conclusions
This study resulted in the identification and relative abundance of important bacterial taxa with potential for zoonosis in six rodent species in a neotropical country. More studies are needed for determining which of the rodent species studied have higher potential for bacterial zoonosis and which environmental conditions, for example rural vs suburban or urban settings, may drive bacterial zoonosis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Rarefaction curves of bacterial phylogenetic diversity (Faith’s PD, ±SE) associated with species of rodents from Panama (A) and associated with M. musculus from two sites (B). Inner plot showed rarefaction curves from other sites collected.
Author Contributions
Conceptualization: G.G., A.M.C., B.A., and L.C.M.; methodology and data generation: G.G., P.G., B.A., and L.C.M.; formal analysis: A.M.C., B.A., and L.C.M.; writing-original draft preparation: G.G., A.M.C., P.G., B.A., and L.C.M.; writing- review & editing: G.G., A.M.C., P.G., B.A., and L.C.M.; funding acquisition B.A., and L.C.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by funds from the Ministry of Economy and Finance of Panama (Grant: FPI-MEF-056), “Epidemiología y ecología de Hantavirus, otras enfermedades zoonóticas y transmitidas por vectores (emergentes y re-emergentes) en Panamá”, Fase-I y Fase II (PhoEZyTV I-II) to B.A. 2010–2014; and the Ministry of Economy and Finance of Panama [Grant number 111130150.501.274], B.A., 2014-2019. A.M.C., B.A. and L.C.M. acknowledge support from the Sistema Nacional de Investigación (SNI-SENACYT).
Data Availability Statement
Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We thank the Panamanian Ministry of Environmental Affairs, the Gorgas Committee for Animal Care and Use, the Gorgas Memorial Institute for Health Studies (GMISH), the Ministry of Health, Ministry of Agricultural Development, and the Ministry of Environment for their support. We also thank individuals from the communities and areas surveyed, sever state organization, and the rodent ecology team of the Ministry of Health and GMISH, especially Eustiquio Broce, Omar Vargas (R.I.P), Francisco Crespo, Carlos Falcon, Ricardo Rodriguez, Juan Bosco Navarro, Santiago Bosh, Ricardo Cedeño, Daniel Gonzalez, Carlos Falconet, Jose Miguel Montenegro, Miguel Vergara, Victor Dominguez, Ariel Perez, Joel Gonzalez, Algis Vergara, Heriberto Rivera, Cesar Lombardo, Domingo Archibold, Leonel Castillo, Alexis Ortiz, Donderis Soto, Carmelo Ortiz (R.I.P), Alexis Araúz, Juán De Leon, Juán Díaz, Ramón García, Jose Santizo (R.I.P), Juan Francisco Tello, Jorge Garzón, Gilberto Niño, Ricardo Rodríguez, Dagoberto Olivares, Jony Castillo, Candelario Olivares, Emilio Salazar, Pablo Gutiérrez, Lorenzo Aldobán, Olmedo Castro, José Valencia (R.I.P), Armando Mepaquito, Silvio Bethancourt, Enrique Ramos (R.I.P), and Rafael Figueroa for support during fieldwork. We thank Claudia Dominguez, database manager. We also thank Rosa de Vargas and Iris Reyes for the significant administrative support given as part of the Department of Research in Emerging and Zoonotic Infectious Diseases, Gorgas Memorial Institute of Health Studies.
Conflicts of Interest
The authors declare no conflict of interest.
Ethics
The study was evaluated and approved by the Institutional Animal Care and Use Committee of the Gorgas Memorial Institute for Health Studies (# 001/05 CIUCAL/ICGES, July 4, 2005); using the criteria established in the “International Guiding Principles for Biomedical Research Involving Animals developed by the Council for International Organizations of Medical Sciences (CIOMIS). The study was in accordance with Law No. 23 of January 15 1997 (Animal Welfare Assurance) of Republic of Panama.
References
- Morens, D.M.; Folkers, G.K.; Anthony, S. Fauci The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Rabozzi, G.; Bonizzi, L.; Crespi, E.; Somaruga, C.; Sokooti, M.; Tabibi, R.; Vellere, F.; Brambilla, G.; Colosio, C. Emerging zoonoses: The “one health approach. ” Saf. Health Work 2012, 3, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Vouga, M.; Greub, G. Emerging bacterial pathogens: the past and beyond. Clin Microbiol Infect 2015, 1, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Barac, A.; Poljak, M.; Ong, D.S.Y. Editorial: Innovative Approaches in Diagnosis of Emerging/Re-emerging Infectious Diseases. Front. Microbiol. 2020, 11, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Magouras, I.; Brookes, V.J.; Jori, F.; Martin, A.; Pfeiffer, D.U.; Dürr, S. Emerging Zoonotic Diseases: Should We Rethink the Animal–Human Interface? Front. Vet. Sci. 2020, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; Zowalaty, M.E.E.; Rahman, A.M.M.T.; Ashour, H.M. Zoonotic diseases: Etiology, impact, and control. Microorganisms 2020, 8, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.; Kutz, S. Introduction to the Special Issue on ‘Emerging Zoonoses and Wildlife. ’ Int. J. Parasitol. Parasites Wildl. 2019, 9, 322. [Google Scholar] [CrossRef]
- Wang, W.-H.; Thitithanyanont, A.; Urbina, A.N.; Wang, S.-F. Emerging and re-emerging virus. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Cantas, L.; Suer, K. Review: The important bacterial zoonoses in “One Health” concept. Front. Public Heal. 2014, 2, 1–8. [Google Scholar] [CrossRef]
- Kruse, H.; Kirkemo, A.M.; Handeland, K. Wildlife as source of zoonotic infections. Emerg. Infect. Dis. 2004, 10, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.K.; Hitchens, P.L.; Pandit, P.S.; Rushmore, J.; Evans, T.S.; Young, C.C.W.; Doyle, M.M. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc. R. Soc. B Biol. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health Rodent-borne diseases and their risks for public health; 2009; Vol. 35; ISBN 1040841090.
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as hosts of pathogens and related zoonotic disease risk. Pathogens 2020, 9, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Mawanda, P.; Rwego, I.; Kisakye, J.J.; Sheil, D. Rodents as potential hosts and reservoirs of parasites along the edge of a central african forest: Bwindi impenetrable national park, South Western Uganda. Afr. Health Sci. 2020, 20, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.D.; Hayman, D.T.S.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.C.; Mills, J.N.; Timonin, M.E.; Willis, C.K.R.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 7039–7044. [Google Scholar] [CrossRef]
- Assefa, A.; Chelmala, S. Correction to: Comparison of rodent community between natural and modified habitats in Kafta-Sheraro National Park and its adjoining villages, Ethiopia: implication for conservation. J. Basic Appl. Zool. 2019, 80. [Google Scholar] [CrossRef]
- Williams, E.P.; Spruill-harrell, B.M.; Taylor, M.K.; Lee, J.; Nywening, A. V; Yang, Z.; Nichols, J.H.; Camp, J. V; Owen, R.D.; Jonsson, C.B. Emergence : Lessons Learned from Bat- and Rodent-Borne. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef]
- Wasserberg, G.; Abramsky, Z.; Kotler, B.P.; Ostfeld, R.S.; Yarom, I.; Warburg, A. Anthropogenic disturbances enhance occurrence of cutaneous leishmaniasis in Israel deserts: Patterns and mechanisms. Ecol. Appl. 2003, 13, 868–881. [Google Scholar] [CrossRef]
- Friggens, M.M.; Beier, P. Anthropogenic disturbance and the risk of flea-borne disease transmission. Oecologia 2010, 164, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Young, H.S.; McCauley, D.J.; Dirzo, R.; Nunn, C.L.; Campana, M.G.; Agwanda, B.; Otarola-Castillo, E.R.; Castillo, E.R.; Pringle, R.M.; Veblen, K.E.; et al. Interacting effects of land use and climate on rodent-borne pathogens in central Kenya. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Young, H.S.; Dirzo, R.; Helgen, K.M.; McCauley, D.J.; Billeter, S.A.; Kosoy, M.Y.; Osikowicz, L.M.; Salkeld, D.J.; Young, T.P.; Dittmar, K. Declines in large wildlife increase landscape-level prevalence of rodent-borne disease in Africa. Proc. Natl. Acad. Sci. U. S. A. 2014, 111, 7036–7041. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Andermann, T.; Faurby, S.; Turvey, S.T.; Antonelli, A.; Silvestro, D. The past and future human impact on mammalian diversity. Sci. Adv. 2020, 6, 1–18. [Google Scholar] [CrossRef]
- Armién, A.G.; Armién, B.; Koster, F.; Pascale, J.M.; Avila, M.; Gonzalez, P.; De La Cruz, M.; Zaldivar, Y.; Mendoza, Y.; Gracia, F.; et al. Hantavirus infection and habitat associations among rodent populations in agroecosystems of Panama: Implications for human disease risk. Am. J. Trop. Med. Hyg. 2009, 81, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Jahan, N.A.; Lindsey, L.L.; Larsen, P.A. The Role of Peridomestic Rodents as Reservoirs for Zoonotic Foodborne Pathogens. Vector-Borne Zoonotic Dis. 2021, 21, 133–148. [Google Scholar] [CrossRef]
- Gonzalez, P.; Salazar, J.R.; Salinas, T.P.; Avila, M.; Colella, J.P.; Dunnum, J.L.; Glass, G.E.; Gonzalez, G.; Juarez, E.; Lindblade, K.; et al. Two Decades of Wildlife Pathogen Surveillance: Case Study of Choclo orthohantavirus and Its Wild Reservoir Oligoryzomys costaricensis. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Armién, B.; Muñoz, C.; Cedeño, H.; Salazar, J.R.; Salinas, T.P.; González, P.; Trujillo, J.; Sánchez, D.; Mariñas, J.; Hernández, A.; et al. Hantavirus in Panama: Twenty Years of Epidemiological Surveillance Experience. Viruses 2023, 15, 1–14. [Google Scholar] [CrossRef]
- Mills, J.N.; Yates, T.L.; Childs, J.E.; Parmenter, R.R.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J. Guidelines for Working with Rodents Potentially Infected with Hantavirus. J. Mammal. 1995, 76, 716. [Google Scholar] [CrossRef]
- Hanshew, A.S.; Mason, C.J.; Raffa, K.F.; Currie, C.R. Minimization of chloroplast contamination in 16S rRNA gene pyrosequencing of insect herbivore bacterial communities. J. Microbiol. Methods 2013, 95, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. U. S. A. 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [PubMed]
- Bekele, A.; Leirs, H. Population ecology of rodents of maize fields and grassland in central Ethiopia. Belgian J. Zool. 1997, 127 Suppl, 39–48. [Google Scholar]
- Meheretu, Y.; Sluydts, V.; Welegerima, K.; Bauer, H.; Teferi, M.; Yirga, G.; Mulungu, L.; Haile, M.; Nyssen, J.; Deckers, J.; et al. Rodent abundance, stone bund density and its effects on crop damage in the Tigray highlands, Ethiopia. Crop Prot. 2014, 55, 61–67. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ Prepr. 2018, 6, e27295v1. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: from raw reads to community analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “all-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, 643–648. [Google Scholar] [CrossRef] [PubMed]
- R Development Core R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2008.
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package Version 2.0-10. 2017, 2. [Google Scholar]
- Wickham, H. ggplot2 - Elegant Graphics for Data Analysis | Hadley Wickham | Springer . 2017. [Google Scholar] [CrossRef]
- Liyai, R.; Kimita, G.; Masakhwe, C.; Abuom, D.; Mutai, B.; Onyango, D.M.; Waitumbi, J. The spleen bacteriome of wild rodents and shrews from Marigat, Baringo County, Kenya. PeerJ 2021, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Balish, E.; Warner, T. Enterococcus faecalis induces inflammatory bowel disease in interleukin-10 knockout mice. Am. J. Pathol. 2002, 160, 2253–2257. [Google Scholar] [CrossRef]
- Marrow, J.; Whittington, J.K.; Mitchell, M.; Hoyer, L.L.; Maddox, C. Prevalence and antibiotic-resistance characteristics of Enterococcus spp. isolated from free-living and captive raptors in central illinois. J. Wildl. Dis. 2009, 45, 302–313. [Google Scholar] [CrossRef]
- Gutiérrez, R.; Krasnov, B.; Morick, D.; Gottlieb, Y.; Khokhlova, I.S.; Harrus, S. Bartonella infection in rodents and their flea ectoparasites: An overview. Vector-Borne Zoonotic Dis. 2015, 15, 27–39. [Google Scholar] [CrossRef]
- Benga, L.; Feßler, A.T.; Benten, W.P.M.; Engelhardt, E.; Köhrer, K.; Schwarz, S.; Sager, M. Acinetobacter species in laboratory mice: species survey and antimicrobial resistance. Lab. Anim. 2019, 53, 470–477. [Google Scholar] [CrossRef]
- Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: Epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef]
- Pérez-Martínez, L.; Venzal, J.M.; González-Acuña, D.; Portillo, A.; Blanco, J.R.; Oteo, J.A. Bartonella rochalimae and other Bartonella spp. in fleas, Chile. Emerg Infect Dis. 2009, 1150–2. [Google Scholar] [CrossRef]
- Theonest, N.O.; Carter, R.W.; Amani, N.; Doherty, S.L.; Hugho, E.; Keyyu, J.D.; Mable, B.K.; Shirima, G.M.; Tarimo, R.; Thomas, K.M.; et al. Molecular detection and genetic characterization of Bartonella species from rodents and their associated ectoparasites from northern Tanzania. PLoS One 2019, 14, 1–17. [Google Scholar] [CrossRef]
- Landaeta-Aqueveque, C.; Moreno Salas, L.; Henríquez, A.L.; Silva-de la Fuente, M.C.; González-Acuña, D. Parasites of Native and Invasive Rodents in Chile: Ecological and Human Health Needs. Front. Vet. Sci. 2021, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Saengsawang, P.; Morand, S.; Desquesnes, M.; Yangtara, S.; Inpankaew, T. Molecular detection of Bartonella species in rodents residing in urban and suburban areas of central Thailand. Microorganisms 2021, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.S.; Worthington, M.G.; Brenner, D.J.; Moss, C.W.; Hollis, D.G.; Weyant, R.S.; Steigerwalt, A.G.; Weaver, R.E.; Daneshvar, M.I.; O’Connor, S.P. Rochalimaea elizabethae sp. nov. isolated from a patient with endocarditis. J. Clin. Microbiol. 1993, 31, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Fenollar, F.; Sire, S.; Raoult, D. Bartonella vinsonii subsp. arupensis as an agent of blood culture-negative endocarditis in a human. J. Clin. Microbiol. 2005, 43, 945–947. [Google Scholar] [CrossRef]
- Welch, D.F.; Carroll, K.C.; Hofmeister, E.K.; Persing, D.H.; Robison, D.A.; Steigerwalt, A.G.; Brenner, D.J. Isolation of a new subspecies, Bartonella vinsonii subsp. arupensis, from a cattle rancher: Identity with isolates found in conjunction with Borrelia burgdorferi and Babesia microti among naturally infected mice. J. Clin. Microbiol. 1999, 37, 2598–2601. [Google Scholar] [CrossRef] [PubMed]
- Probert, W.; Louie, J.K.; Tucker, J.R.; Longoria, R.; Hogue, R.; Moler, S.; Graves, M.; Palmer, H.J.; Cassady, J.; Fritz, C.L. Meningitis due to a “Bartonella washoensis”-like human pathogen. J. Clin. Microbiol. 2009, 47, 2332–2335. [Google Scholar] [CrossRef]
- Corvec, S. Clinical and biological features of Cutibacterium (Formerly Propionibacterium) avidum, an underrecognized microorganism. Clin. Microbiol. Rev. 2018, 31, 1–42. [Google Scholar] [CrossRef]
- Elston, M.J.; Dupaix, J.P.; Opanova, M.I.; Atkinson, R.E. Cutibacterium acnes (formerly Proprionibacterium acnes) and Shoulder Surgery. Hawai’i J. Heal. Soc. Welf. 2019, 78, 3–5. [Google Scholar]
- Ellerbroek, L.; Mac, K.N.; Peters, J.; Hultquist, L. Hazard Potential from Antibiotic-resistant Commensals like Enterococci. J. Vet. Med. 2004. [Google Scholar] [CrossRef]
- Mallon, D.J.P.; Corkill, J.E.; Hazel, S.M.; Sian Wilson, J.; French, N.P.; Bennett, M.; Hart, C.A. Excretion of vancomycin-resistant enterococci by wild mammals. Emerg. Infect. Dis. 2002, 8, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, L.P.; Valentini Junior, D.F.; Machado, S.M.D.S.; Schaefer, P.G.; Rivero, R.C.; Osvaldt, A.B. Sarcina ventriculi a rare pathogen. Autops. Case Reports 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Owens, L.A.; Colitti, B.; Hirji, I.; Pizarro, A.; Jaffe, J.E.; Moittié, S.; Bishop-Lilly, K.A.; Estrella, L.A.; Voegtly, L.J.; Kuhn, J.H.; et al. A Sarcina bacterium linked to lethal disease in sanctuary chimpanzees in Sierra Leone. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Kim, H.K.; Missiakas, D.; Schneewind, O. Mouse models for infectious diseases caused by Staphylococcus aureus. Physiol. Behav. 2018, 176, 139–148. [Google Scholar] [CrossRef]
- Poinsot, D.; Charlat, S.; Merçot, H. On the mechanism of Wolbachia-induced cytoplasmic incompatibility: Confronting the models with the facts. BioEssays 2003, 25, 259–265. [Google Scholar] [CrossRef]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and Virus Protection in Insects. Science (80-. ). 2008, 322, 702–702. [Google Scholar] [CrossRef]
- Castillo, A.M.; Saltonstall, K.; Arias, C.F.; Chavarria, K.A.; Ramírez-Camejo, L.A.; Mejía, L.C.; De León, L.F. The microbiome of neotropical water striders and its potential role in codiversification. Insects 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Castillo, A.M.; Chavarria, K.A.; Saltonstall, K.; Arias, C.F.; Mejía, L.C. Salinity effects on the microbiome of a Neotropical water strider. Hydrobiologia 2021. [Google Scholar] [CrossRef]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and Evolution of Heritable Bacterial Symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef]
- Tang, X.; Xie, G.; Shao, K.; Bayartu, S.; Chen, Y.; Gao, G. Influence of salinity on the bacterial community composition in Lake Bosten, a large oligosaline lake in arid Northwestern China. Appl. Environ. Microbiol. 2012, 78, 4748–4751. [Google Scholar] [CrossRef]
- Schuler, H.; Egan, S.P.; Hood, G.R.; Busbee, R.W.; Driscoe, A.L.; Ott, J.R. Diversity and distribution of Wolbachia in relation to geography, host plant affiliation and life cycle of a heterogonic gall wasp. BMC Evol. Biol. 2018, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sampling sites of six species of rodents in Panama. Symbols represent: L. adspersus (circle), M. caliginosus (triangle), M. musculus (filled square), P. semispinosus (plus), R. rattus (square across), Z. brevicauda (star).
Figure 1.
Sampling sites of six species of rodents in Panama. Symbols represent: L. adspersus (circle), M. caliginosus (triangle), M. musculus (filled square), P. semispinosus (plus), R. rattus (square across), Z. brevicauda (star).
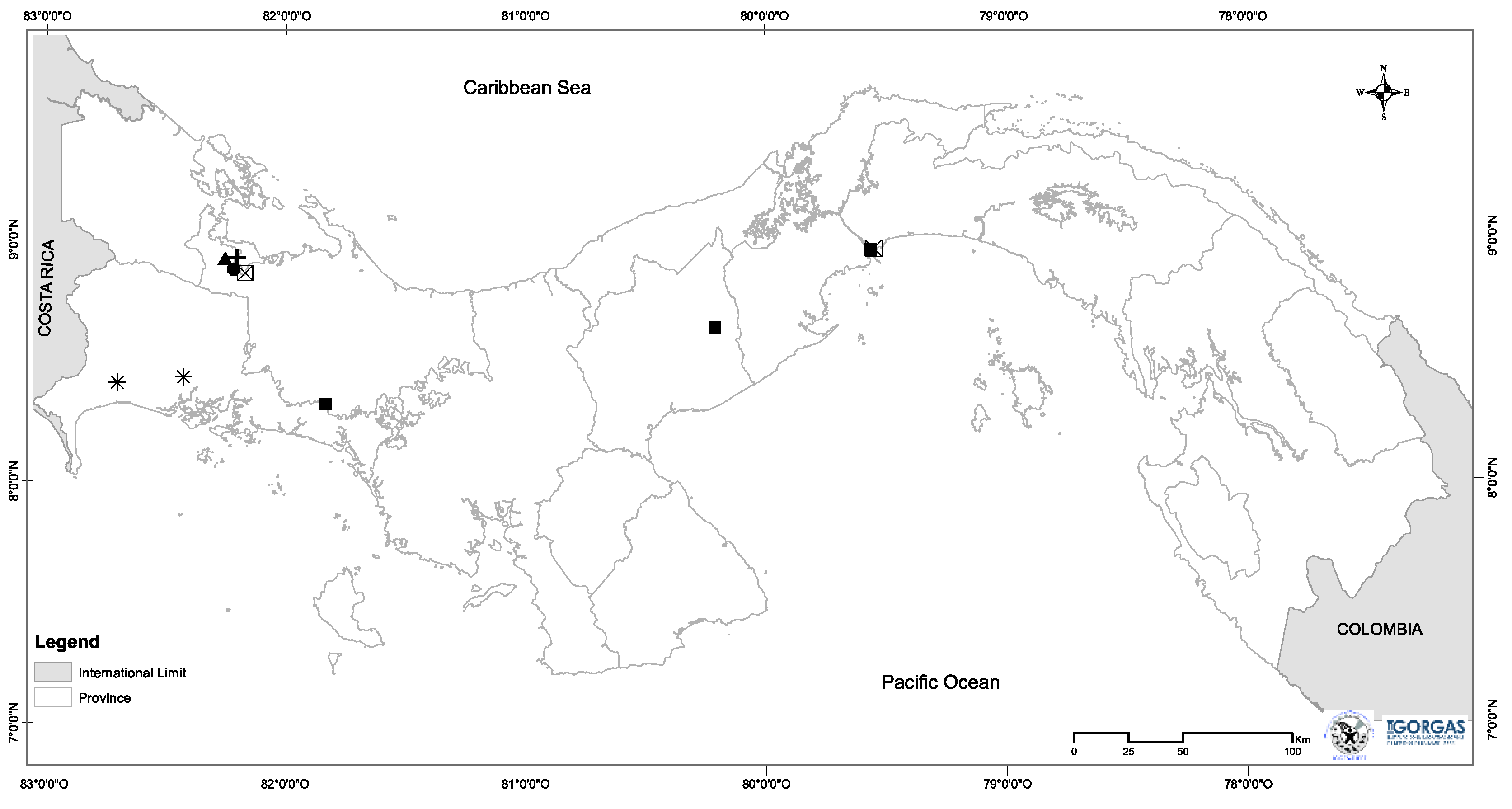
Figure 2.
Relative abundance of dominant bacteria taxa associated with species of rodents from Panama. Abundance was estimated at the level of bacterial class across species (A), bacterial class across sites (B).
Figure 2.
Relative abundance of dominant bacteria taxa associated with species of rodents from Panama. Abundance was estimated at the level of bacterial class across species (A), bacterial class across sites (B).
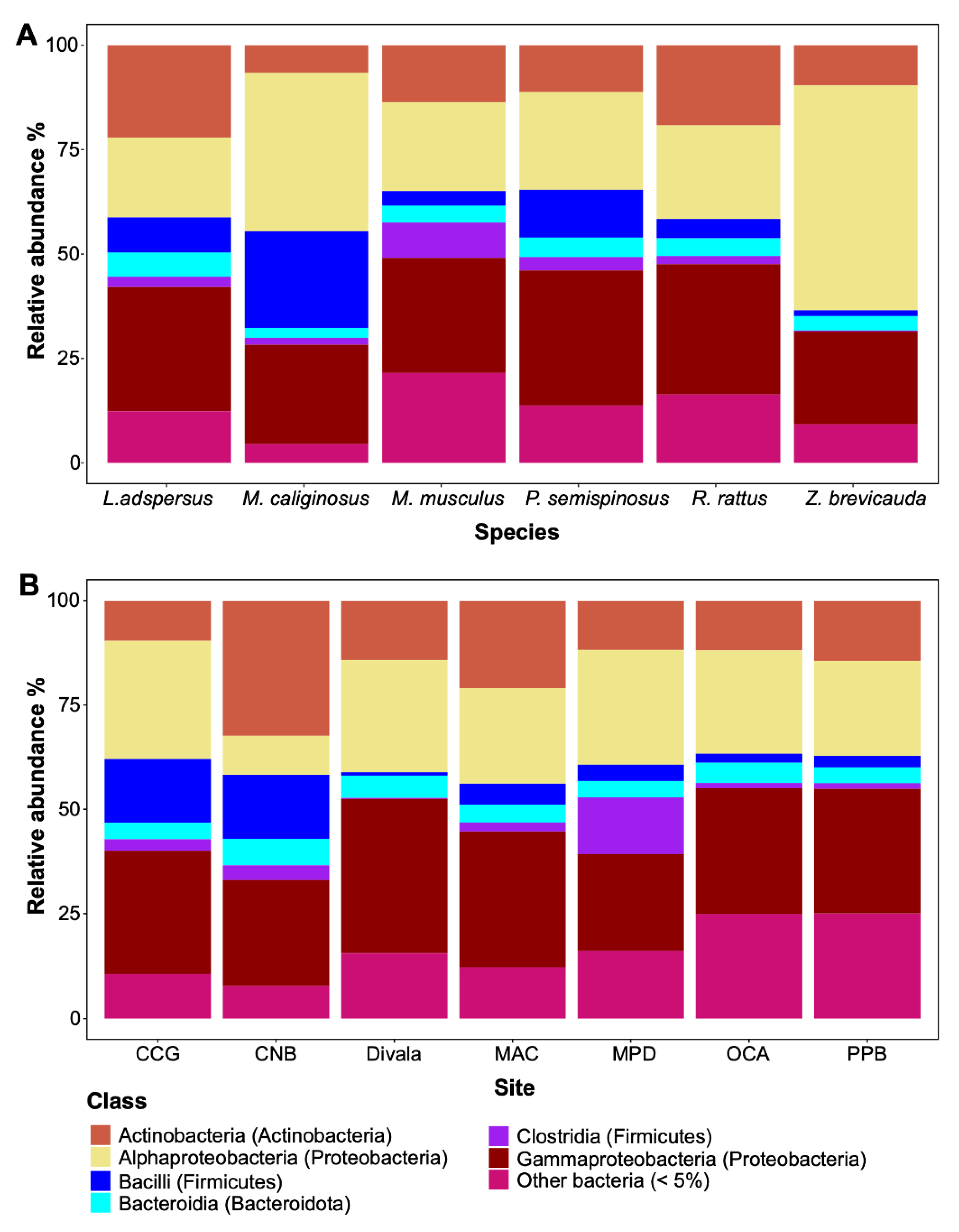
Figure 3.
Relative abundance of dominant bacterial taxa associated with species of rodents from Panama. Abundance was estimated at the genus rank across species (A), bacterial genera across M. musculus (B), and bacterial genus across sites (C).
Figure 3.
Relative abundance of dominant bacterial taxa associated with species of rodents from Panama. Abundance was estimated at the genus rank across species (A), bacterial genera across M. musculus (B), and bacterial genus across sites (C).
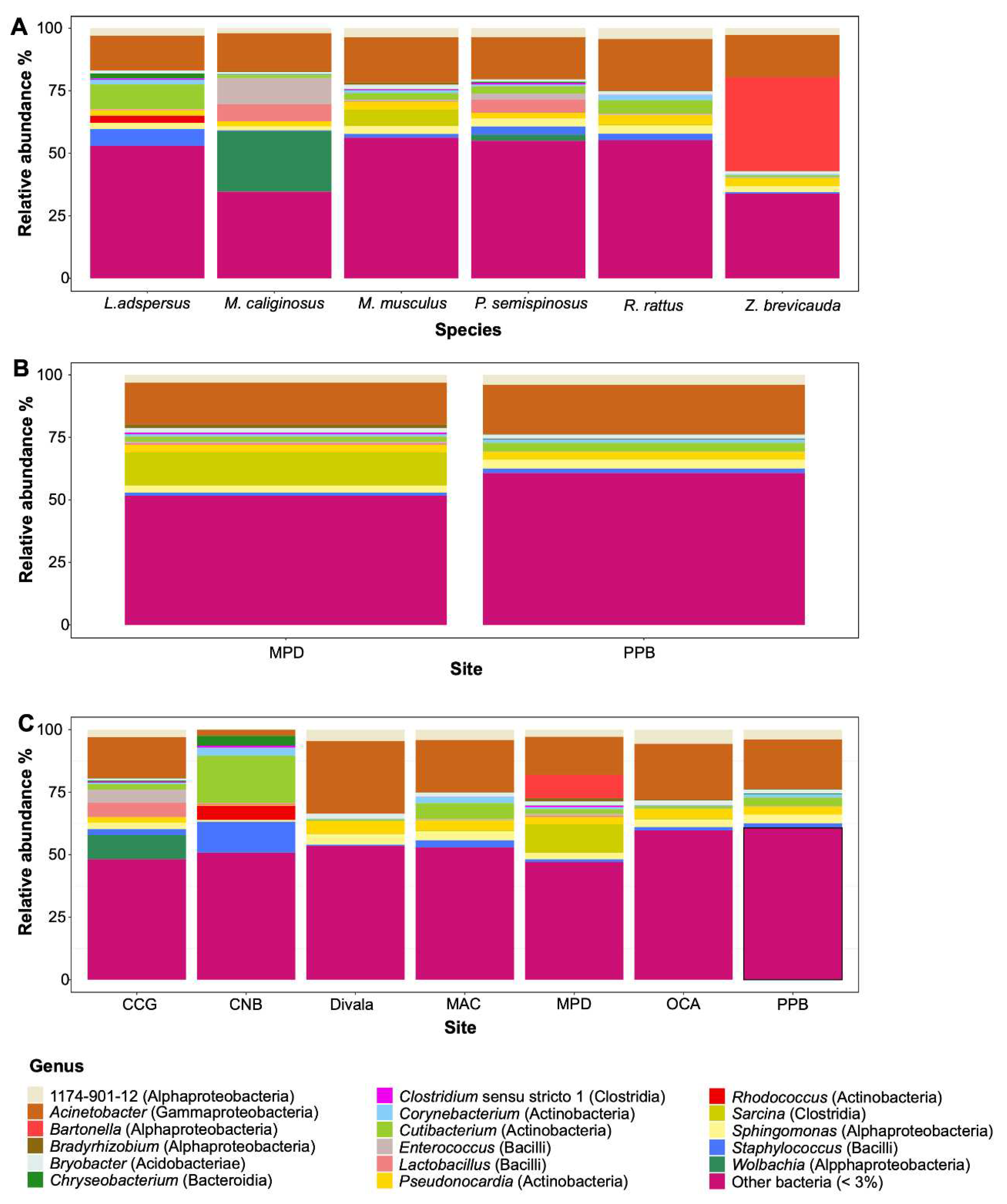
Figure 4.
Bacterial diversity associated with rodents from Panama. Graphs represent estimates of alpha diversity-based Faith’s PD for each species (A), and sites (for M. musculus) (B). Beta diversity PCoA based on weighted Unifrac distance among species (C).
Figure 4.
Bacterial diversity associated with rodents from Panama. Graphs represent estimates of alpha diversity-based Faith’s PD for each species (A), and sites (for M. musculus) (B). Beta diversity PCoA based on weighted Unifrac distance among species (C).

Table 1.
Species of rodents collected in the study, sites, and sample size (n).
| Species | Sites | n |
|---|---|---|
| L. adspersus | Comarca Ngäbe-Buglé (CNB) | 1 |
| L. adspersus | Oajaca Chiguirí Arriba (OCA) | 1 |
| M. caliginosus | Cañazas Chiriquí Grande (CCG) | 1 |
| M. musculus | Mercado Público de David (MPD) | 7 |
| M. musculus | Panama Port Balboa (PPB) | 7 |
| P. semispinosus | Cañazas Chiriquí Grande (CCG) | 2 |
| R. rattus | Oajaca Chiguirí Arriba (OCA) | 1 |
| R. rattus | Mercado de Abasto Curundú (MAC) | 4 |
| Z. brevicauda | Divalá | 1 |
| Z. brevicauda | Mercado Público de David (MPD) | 1 |
Table 2.
Relative abundance (%) bacteria at the level of genus in the six species of rodents. Sites are showed in abbreviation with exception of Divala. Number of sample size are showed in n. Only M. musculus showed relative abundance with same number of n.
Table 2.
Relative abundance (%) bacteria at the level of genus in the six species of rodents. Sites are showed in abbreviation with exception of Divala. Number of sample size are showed in n. Only M. musculus showed relative abundance with same number of n.
| Bacteria | Species of Rodents | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genus |
L. adspersus |
M. caliginosus |
M. musculus |
P. semispinosus |
R. rattus |
Z. brevicauda |
||||
| CNB | OCA | CCG | MPD | PPB | CCG | OCA | MAC | Divalá | MPD | |
| n=1 | n=1 | n=1 | n=7 | n=7 | n=2 | n=1 | n=4 | n=1 | n=1 | |
| 1174-901-12 (Alphaproteobacteria) | 0.00 | 5.97 | 1.97 | 3.16 | 3.97 | 3.54 | 5.31 | 4.11 | 4.59 | 0.66 |
| Acinetobacter (Gammaproteobacteria) | 2.46 | 25.19 | 15.24 | 16.70 | 19.62 | 16.67 | 19.36 | 20.90 | 29.01 | 5.16 |
| Bartonella (Alphaproteobacteria) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 74.87 |
| Bradyrhizobium (Alphaproteobacteria) | 0.00 | 0.36 | 0.38 | 1.45 | 0.31 | 0.25 | 0.27 | 0.22 | 0.00 | 0.22 |
| Bryobacter (Acidobacteria) | 0.00 | 2.33 | 0.71 | 1.73 | 1.60 | 0.91 | 1.55 | 1.44 | 2.03 | 0.30 |
| Chryseobacterium (Bacteroidia) | 3.83 | 0.11 | 0.31 | 0.12 | 0.29 | 0.54 | 0.00 | 0.03 | 0.03 | 0.11 |
| Clostridium sensu stricto 1 (Clostridia) | 0.90 | 0.14 | 0.00 | 0.65 | 0.09 | 0.66 | 0.00 | 0.14 | 0.00 | 0.00 |
| Corynebacterium (Actinobacteria) | 3.28 | 0.38 | 0.24 | 0.88 | 1.29 | 0.67 | 0.07 | 2.52 | 0.19 | 0.56 |
| Cutibacterium (Actinobacteria) | 18.85 | 0.92 | 1.01 | 2.05 | 3.50 | 2.87 | 1.02 | 6.41 | 0.64 | 0.93 |
| Enterococcus (Bacilli) | 0.16 | 0.00 | 10.56 | 0.64 | 0.17 | 2.50 | 0.39 | 0.46 | 0.06 | 0.63 |
| Lactobacillus (Bacilli) | 0.62 | 0.00 | 6.79 | 0.58 | 0.01 | 5.27 | 0.00 | 0.07 | 0.00 | 0.00 |
| Pseudonocardia (Actinobacteria) | 0.33 | 4.06 | 2.08 | 3.17 | 3.04 | 2.22 | 3.44 | 3.97 | 5.10 | 1.10 |
| Rhodococcus (Actinobacteria) | 5.65 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 |
| Sarcina (Clostridia) | 0.00 | 0.24 | 0.00 | 13.10 | 0.11 | 0.06 | 0.71 | 0.26 | 0.19 | 0.07 |
| Sphingomonas (Alphaproteobacteria) | 0.71 | 4.11 | 1.51 | 2.85 | 3.47 | 3.12 | 1.90 | 3.66 | 4.18 | 0.58 |
| Staphylococcus (Bacilli) | 12.36 | 1.07 | 0.31 | 1.19 | 1.82 | 3.26 | 1.58 | 2.86 | 0.49 | 0.59 |
| Wolbachia (Alphaproteobacteria) | 0.00 | 0.00 | 24.26 | 0.00 | 0.00 | 2.47 | 0.00 | 0.00 | 0.00 | 0.00 |
| Other bacteria (< 3 %) | 50.85 | 55.12 | 34.63 | 51.73 | 60.71 | 54.99 | 64.40 | 52.93 | 53.49 | 14.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



