
Submitted:
11 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
Rice is an important cereal crop worldwide, the growth of which is affected by rice blast disease, caused by the fungal pathogen Magnaporthe oryzae. As climate change increases the diversity of pathogens, the disease-resistance genes (R genes) in plants must be identified. The major blast-resistance genes have been identified in indica rice varieties; therefore, japonica rice varieties with R genes now need to be identified. Because leucine-rich repeat (LRR) domain proteins possess R-gene properties, we used bioinformatics analysis to identify the rice candidate LRR domain receptor-like proteins (OsLRR-RLPs). OsLRR-RLP2, which contains six LRR domains, showed differences in the DNA sequence, containing 43 single-nucleotide polymorphisms (SNPs) in indica and japonica subpopulations. The results of M. oryzae inoculation analysis indicated that indica varieties with partial deletion of OsLRR-RLP2 showed susceptibility, whereas japonica varieties with intact OsLRR-RLP2 showed resistance. oslrr-rlp2 mutant generated using clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) showed increased pathogen susceptibility, whereas plants overexpressing this gene showed pathogen resistance. These results indicate that OsLRR-RLP2 confers resistance to rice, and OsLRR-RLP2 may be useful for breeding resistant cultivars.
Keywords:
LRR-RLP
; Magnaporthe oryzae
; rice
; japonica resistance
1. Introduction
Rice (Oryza sativa L.) accounts for approximately half of the total food crop consumption. However, global rice production is being seriously threatened by rice blast disease, which is caused by the fungus Magnaporthe oryzae [1]. The annual yield loss due to rice blast disease is approximately 10–30% [2]. Therefore, cultivating resistant varieties can reduce rice yield losses. Plants possess immune recognition pathways for pests and pathogens. Resistance genes (R genes) provide disease resistance to plants. Many R genes are being studied; however, because of the increasing variety of pathogens and their resistance to R genes, additional research on R genes is important for rice breeding.
Plants defend themselves against pathogens through pattern-triggered immunity (PTI) and effector-triggered immunity (ETI) activation. The first plant defense mechanism is PTI, which is mediated by pattern recognition receptors (PRRs), which are activated upon direct recognition of pathogen-associated molecular patterns (PAMPs) by PRRs [3]. The second plant defense mechanism is ETI, which is mediated by the direct or indirect recognition of effectors by nucleotide-binding and leucine-rich repeat receptors (NLRs) [4]. The innate immune receptors can be initially recognized using NLR and PRRs. PRRs are located at the plasma membrane and recognize extracellular signals, termed PAMPs [5]. Receptor-like kinases (RLKs) are members of the PRR family. Receptor kinases perform recognition and signal transduction [6]. The intracellular kinase domains encoded by PRRs perform immune-sensing functions. In contrast to RLKs, receptor-like proteins (RLPs) contain an ectodomain, a transmembrane domain, and a short cytoplasmic tail [7]. As RLKs, RLPs are involved in plant defense and development [8,9]. Both use extracellular leucine-rich repeat (LRR) domains to perceive PAMPs; however, unlike RLKs, RLPs lack an intracellular kinase domain. The molecular mechanisms underlying the recognition and signaling of RLKs and RLPs are still poorly understood. Given the limited research available on this topic, further studies on receptor identification are required.
The LRR is involved in protein –protein recognition. The LRRs are generally 20–29 residues long and contain 11 conserved residues with the consensus sequence LxxLxLxxN/CxL, in which any amino acid is “x”; “L” is Val, Leu, or Ile, “N” is Asn, Thr, Ser, or Cys; and “C” is Cys or Ser [10]. Leucine-rich repeat receptor-like proteins (LRR-RLPs) are also involved in the activation of the immune response [11]. LRR-RLPs have been discovered in various plant species since they were first characterized in tomatoes (Lycopersicon esculentum) [8]. A total of 57 genes encoding LRR-RLPs have been identified in Arabidopsis [11], some of which have been functionally characterized [7]. For example, the receptor of the enigmatic MAMP of Xanthomonas (ReMAX) is responsible for the recognition of eMAX by Xanthomonas strains [12]. Solanum lycopersicum LRR-RLPs confer resistance against pathogens by recognizing avirulence proteins [13]. Ninety genes encoding LRR-RLPs have been identified [10]. OsRLP1 is involved in resistance to rice black-streaked dwarf virus [14]. RLPs have rarely been studied; research particularly in rice is scarce.
Cultivated rice varieties belong to the indica and japonica subspecies, which have substantially changed, including their stress resistance, during domestication [15,16]. However, most blast genes have been identified as indica accessions and are less well-known in japonica cultivars [17]. In this study, we identified another RLP, named OsLRR-RLP2, in a japonica cultivar and evaluated its response to M. oryzae. To identify the R gene related to rice blast disease in the rice variety, we systematically analyzed single-nucleotide polymorphisms (SNPs) with Indels and screened T-DNA insertional lines screening of the LRR domain protein genes. We obtained accessions possessing mutations within the LRR and subsequently performed a series of experimental investigations under blast treatment. Identification of the disease resistance genes related to PRRs provides genetic resources that can be used to improve our understanding of rice immunity. These results provide a useful reference for the development of rice plant with enhanced resistance to blast R gene.
2. Results
2.1. Selection of LRR Candidates and Phylogenetic Analysis
We first downloaded 3240 genes encoding LRR proteins from a rice genome database (Rice Genome Annotation Project, http://rice.uga.edu/, accessed on 21 October 2021). After removing duplicate values and changing them to the RAP-DB ID (https://rapdb. dna. affrc. go. jp/, accessed on 28 October 2021), 857 genes were analyzed for indels (≥ 3bp) of 453 high quality rice varieties compared to Nipponbare and the frequency of indels was presented as a heatmap (yellow boxes indicate the high frequency and the blue ones indicates low frequency) (Figure 1A). We then identified 316 genes that showed high genetic diversity among the rice varieties compared to the reference (Nipponbare) genome. We further analyzed the microarray data for 277 genes with probes in rice Affymetrix gene chip (Figure 1B), and 16 genes of them showed significant changes in both the BLAST or bacterial blight disease treatments (Figure 1C). Through the analysis of the T-DNA mutant lines, one LRR domain gene was selected for further analysis due to its response to rice blast disease. This LRR domain gene, named OsLRR-RLP2, contains LRR N-terminus domain (LRRNT) and six LRR domains (Figure 2A).
To determine phylogenetic relationships with other species, the protein sequences of all species were searched using BLASTx in NCBI GenBank. We found high amino acid sequence identity with Oryza species: 93% similarity with O. sativa indica sequence (EAY86640.1) and 83.1% similarity with Oryza glaberrima (XP_052143053.1) (Figure 2B). Other species, including monocot species, have low amino acid sequence identities ranging from 39% to 55%. To find homologs in the model plant Arabidopsis, we also searched for homologs in the TAIR database (TAIR https://www.arabidopsis.org/, accessed on 13 June 2022). Although their similarity was very low (39.4%), the homolog with the highest similarity, AtRLP19 (OAP10192.1), has 27 LRR domains. There was no T-DNA insertion in AtRLP19 because mutation in this gene causes lethality [7]. All of the LRR domains in one protein form a single continuous structure and adopt an arc or horseshoe shape, a bowed tube on the concave face, a parallel β sheet, and a convex face with a variety of secondary structures, such as α helices, 310 helices, pII, and β turns [18]. LRR domains forms a stack of the parallel β strands on the inner concave face in both OsLRR-RLP2 and AtRLP19 (Figure 2C). A secondary helical structure is located on the outer convex face. In contrast to OsLRR-RLP2, AtRLP19 has 27 LRR domains, resulting in an expanded structural profile that includes a larger stack of parallel strands and transmembrane (TM) domains. To date, no functional studies have been conducted on this gene in any species.
2.2. Expression and Localization Analysis of OsLRR-RLP
Transcriptome data from Nipponbare (CAFRI-Rice database, https://cafri-rice.khu.ac.kr/, accessed on 21 February 2023) showed OsLRR-RLP2 is expressed at very low levels most of tissues/organs except the leaves with high expression (Figure 2D). To detect the gene transcripts in japonica cultivar Dongjin (cv. DJ), we performed quantitative RT-PCR (qRT-PCR) analysis using the total RNA prepared from vegetative and reproductive organs. Similar to the RNA-seq data, expression of OsLRR-RLP2 is overall low but preferential in the leaves (Figure 2E).
Protein localization was examined to validate the presence of the gene in the cellular level. Using online tools, the predominant site of OsLRR-RLP protein localization was predicted to be plasma membrane. Subcellular localization was observed through transient expression of the C-terminal GFP-tagged protein in tobacco epidermal cells. Control (p35S:GFP) signals were distributed in the nucleus, cytoplasm, and plasma membrane of epidermal tobacco cells (Figure 3A). OsLRR-RLP2 signals, which express the p35S:OsLRR-RLP2-GFP construct, were observed in the cytoplasm and plasma membrane (Figure 3B). To distinguish the signals between the plasma membrane and the cell wall, the leaves were treated with 1 N NaCl. After plasmolysis, both control and OsLRR-RLP2 GFP signals were detected in the cytoplasm and plasma membrane, but not in the cell wall, when observed via merging with FM4-64, a plasma membrane marker (Figure 3C,D). The protein localization is similar to that of OsRLP1, which localizes to the cytoplasm and plasma membrane [14].
2.3. Haplotype Analysis of OsLRR-RLP2 Sequence and Validation of M. oryzae Response
Using the rice SNP database, we observed that OsLRR-RLP2 was a subspecies-unbalanced gene whose frequency in japonica is 5% greater than that in indica subgroups, indicating a japonica-dominant gene (Figure 4A). The allele frequency of OsLRR-RLP2 in different rice subpopulations showed sequence variation compared to reference-type alleles, particularly in indica and australia (aus) (Figure 4B). The tropical and temperate rice types mostly had reference-type alleles, whereas the indica and aus groups possessed alternate alleles and belonged to another haplotypic group compared with the japonica type. In the OsLRR-RLP2 gene, a total of 43 SNPs were identified. Among these, 28 were identified as nonsynonymous SNPs (Figure 4C).
To confirm the defense responses based on SNP differences, accessions from the Korean rice collection (RWG) were acquired [19]. The RWGs were categorized as temperate japonica and indica. Among these, we obtained 9 RWG lines with indels in OsLRR-RLP2 (Table S1). To investigate the correlation between sequence variation and M. oryzae responses, we inoculated 9 RWG lines using punch inoculation. Following inoculation, compared with the DJ cultivar, susceptibility was observed in indica accessions, RWG-006 (CT9993-5-10-1-M, Colombia introduction rice), RWG-034 (BALA, India introduction rice), RWG-123 (Chungdo Hwayang 14, Korean weedy rice), and RWG-165 (Milyang 23, Korean breeding line), whereas resistance was observed in japonica accessions RWG-084 (Syalebyeo-163-1-B, Korean weedy rice), RWG-138 (Jinbu Byeo, Korean breeding line), RWG-140 (Hopyung, Korean breeding line), RWG-231 (Gangchan, Korean breeding line), and RWG-281 (Jinbaek, Korean breeding line) (Figure 5A,B). Fungal biomass was higher in susceptible lines than in resistant lines (Figure 5C). A subsequent PCR-based sequencing analysis was conducted to examine the OsLRR-RLP2 sequences of the RWG lines. Different sequence sizes of RWG lines were observed using PCR of the 1.5 kb full-length gene in combination with primers F1 and R2 (Figure S1A,B). Further sequencing of the PCR product using primers F2 and R2 showed deletion of the fourth LRR domain in indica RWG-006, RWG-034, RWG-123, and RWG-165 compared with intact LRR domains in japonica accessions (Figure S1C). Although other possibilities of M. oryzae response to RWG lines exist, japonica varieties were resistant and indica varieties were susceptible to rice blast disease, to which the sequence variation in OsLRR-RLP2 could be a contributing factor.
2.4. Functional Analysis of OsLRR-RLP2 Involving Rice Blast Disease
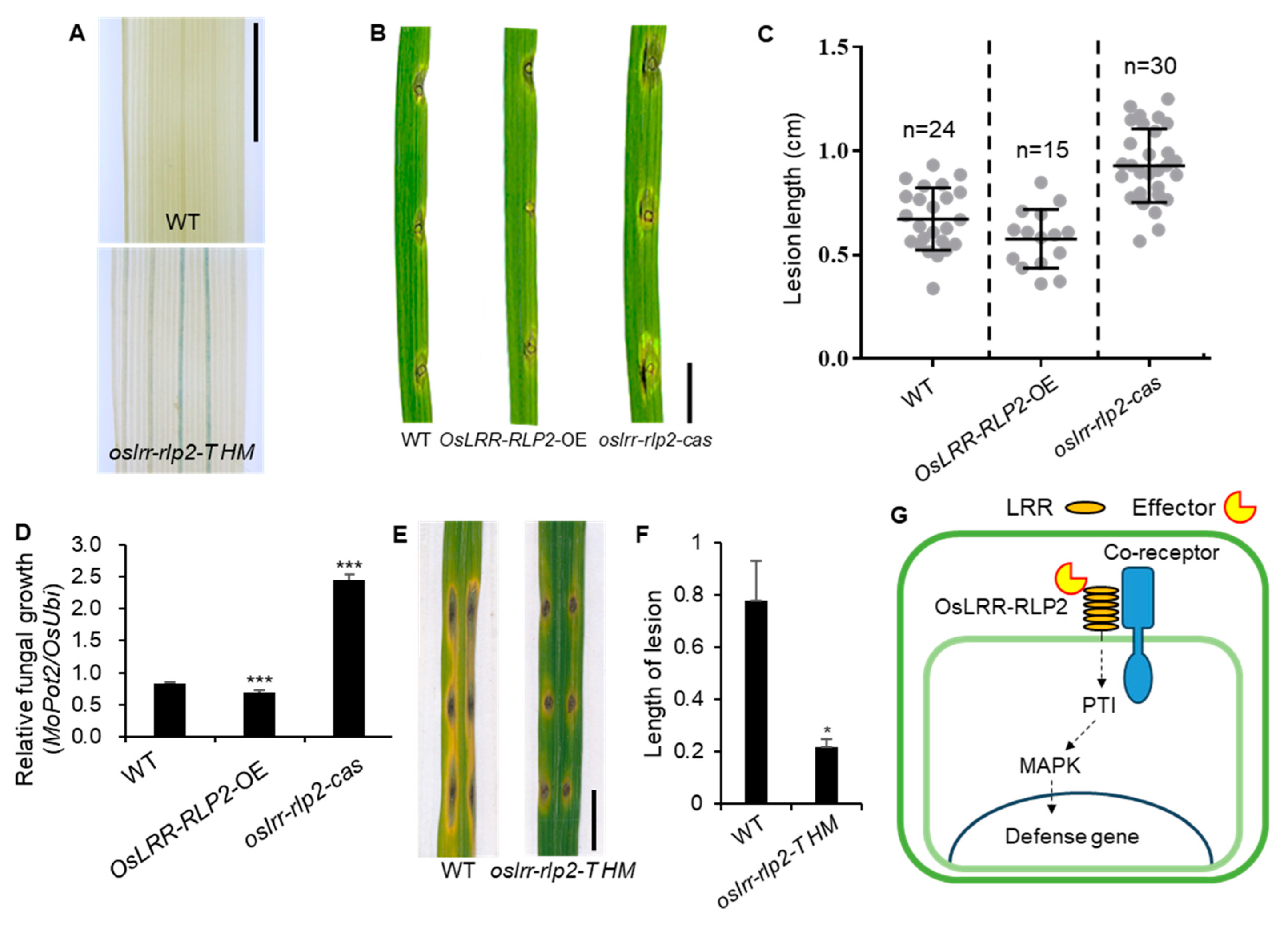
To examine the role of OsLRR-RLP2 in rice immunity against blast disease, we isolated a homozygous (HM) T-DNA-tagged mutant, 3D-01328 (Figures S2A), and generated overexpression and knockout lines (Figure S2B). To verify T-DNA insertion, we analyzed beta-glucuronidase (GUS) activity. Histochemical staining for GUS activity in transgenic rice plants showed that OsLRR-RLP2 was constitutively expressed, and GUS staining was observed in the main vein tissue of rice leaves (Figure 6A). We used PCR-based genotyping to confirm T-DNA insertion in the rice genome; the amplified PCR products showed the presence of an approximately 500 bp band for OsLRR-RLP2 genes from transgenic plants, and no amplified DNA fragment from wild-type (WT) plants confirmed the insertion of T-DNA fragments into the rice genome (Figure S2C). Inserted T-DNA vector is pGA2772, which contains enhancer element, therefore we examined expression value. T-DNA transgenic plants exhibited higher OsLRR-RLP2 expression levels than the WT plants (Figure S2D), indicating it is activation tagging line.
To confirm overexpressed gene effect in activation T-DNA tagging line, we constructed overexpressing plants using the ubiquitin promoter and OsLRR-RLP2 transcription levels in OsLRR-RLP2-OE plants were ten times higher than in WT plants (Figure S2E). Moreover, we generated knockout plants using the clustered regularly interspaced palindromic repeat (CRISPR)/CRISPR-associated protein 9 (Cas9) system. Through genotyping, we selected knockout oslrr-rlp2-cas1 and oslrr-rlp2-cas2 for further analysis, which are homozygous mutants with 7 bp deletions and 1 bp insertions, respectively, for further analysis (Figure S2F).
We performed a blast disease assay on leaves after punch inoculation with M. oryzae. WT, OsLRR-RLP2-OE, and oslrr-rlp2-cas plants were inoculated with M. oryzae, and disease symptoms were quantified at 7 days post inoculation (dpi). The lesion size on oslrr-rlp2-cas leaves was substantially larger than that on WT leaves, whereas that on OsLRR-RLP2-OE leaves was considerably smaller (Figure 6B,C). The propagation of M. oryzae on infected leaves was quantified via qRT-PCR using primers specific for MoPot2, an M. oryzae housekeeping gene. In agreement with the resistant phenotype, the relative fungal biomass was notably smaller in the rice plants inoculated with OsLRR-RLP2-OE than in the WT plants, whereas the expression was higher in oslrr-rlp2-cas (Figure 6D). oslrr-rlp2-T HM lines exhibited enhanced resistance to M. oryzae compared with the WT (Figure 6E,F). oslrr-rlp2-cas showed susceptibility to M. oryzae compared with the WT, whereas OsLRR-RLP2-OE exhibited resistance compared with the WT. These results indicate that OsLRR-RLP2 positively regulates the blast resistance response in rice.
3. Discussion
Global rice production must double by 2050 to meet the demands of the growing population [20]; however, rice yield is affected by biotic and abiotic stresses. Rice blast, caused by M. oryzae, is one of the most serious diseases of the crop [1]. The plant immune system is initially mediated by receptors; PRRs, the receptor proteins, include RLKs or RLPs, and typically have LRR domains [21]. In this study, we identified LRR candidates that seem specific to Oryza spp. and predominantly exist in japonica rice. The gene has only LRR domains, so we named it OsLRR-RLP2.
We identified OsLRR-RLP2 as a positive regulator of rice blast resistance in rice. Overexpression of OsLRR-RLP2 increased rice defenses, whereas the OsLRR-RLP2 mutant was more susceptible to rice blast infection than WT plants. RLPs are plasma membrane-localized receptors that lack intracellular signaling domains [22]. The RLP genes in several plant species have been implicated in disease resistance. RXEG1, an RLP from N. benthamiana, is associated with XEG1 via the LRR domain in the apoplast and forms a complex with the LRR-receptor-like kinases BAK1 and SOBIR1 to transduce XEG1-induced defense signals [23]. Arabidopsis RLP1 and AtRLP1 have also been implicated in PAMP recognition and PTI activation [12]. Despite AtRLP1 lacking glycine residues in its TM, AtRLP1 is capable of binding to the coreceptor RLK, named AtSOBIR1 [24]. Another RLP, AtRLP23, mediates the recognition of a peptide motif (nlp20) found in numerous bacterial, fungal, and oomycete necrosis and ethylene-inducing peptide 1-like proteins (NLPs) [24]. AtRLP30 plays a role in resistance against necrotrophic fungi in Arabidopsis [25]. Further investigations to identify the adaptor kinases associated with OsLRR-RLP2 and the recognition of M. oryzae PAMP, such as MSP1 [26], will reveal the mechanism of RLP in the rice immune response.
Understanding the subcellular localization of resistance genes is important because of the site of interaction between R proteins and cognate pathogen effector proteins. Protein localization and molecular functions can be determined through transient expression in transgenic plants. R proteins are typically localized in the cytoplasm, nucleus, or cell membrane [27,28]. The cytoplasmic localization of resistance proteins contributes to the hypersensitive response (HR) [29]. In the future, whether OsLRR-RLP2 is required to induce HR will need to be determined. We also performed a GUS assay to determine the insertion of T-DNA and expression pattern of OsLRR-RLP2. The expression of this gene in the leaves was high in the main vein tissue. LRR-RLKs are predominantly expressed in leaf vascular tissues [30]. Plant vascular bundles are responsible for the transport of water and nutrients and are colonized by M. oryzae [31]. Therefore, expression in the vasculature indicates that OsLRR-RLP2 functions there to provide resistance against M. oryzae. This suggests that OsLRR-RLP2 plays a role in the immune response in leaf veins.
Genome-wide analyses of the LRR-RLP family have been conducted in Arabidopsis, rice, Brassica napus, and poplar [7,11,32,33]. Multicopy tandem repeats support the diversifying selection of R genes available for adaptation to pathogen challenges [11]. The R genes Pid3, Pi5, Pb1, and Pik are specifically distributed in the genomes of rice subspecies, japonica-type accessions, and indica-type accessions [34]. These results are possible because R genes have experienced substantial differentiation owing to reproductive isolation during the evolutionary process. This distribution specificity of R genes implies that the R genes conferring resistance to M. oryzae differ between the indica and japonica subspecies. The SNP-seek database revealed that OsLRR-RLP2 has 43 SNPs, 8 deletions, and 3 insertions in exons. However, OsLRR-RLP2 has more deletions in the fourth LRR domain. This variation can be attributed to distinctions arising from the Korean landrace compared to others. The results of M. oryzae inoculation analysis indicated that the indica group with partial deletion of the OsLRR-RLP2 showed susceptibility, and the japonica group with an intact OsLRR-RLP2 showed resistance. However, in some lines, mutations in the OsLRR-RLP2 domain did not completely match the phenotypic characteristics of rice blast disease. Other mechanisms may also occur in RWG lines. Use of the OsLRR-RLP2 gene shows considerable potential for advancing research on blast disease resistance in japonica rice.
4. Materials and Methods
4.1. Plant Materials and Blast Inoculation
All the rice plants including WT, transgenic plants, and RWG lines were grown in a growth chamber at 28 ℃ with a 12/12h (light/dark) photoperiod. To obtain seeds, rice plants were grown in a greenhouse at 28 ℃ (16 h light) and 22 ℃ (8 h dark), or in a living modified organism (LMO)-regulated paddy field at Kyung Hee University in Suwon, or at Pusan National University in Miryang, South Korea. The seeds were germinated on MSO medium for approximately 10 days and then transferred to soil. Approximately 4- to 5-week-old seedlings were used for M. oryzae PO6-6 inoculation. The uppermost 2 to 4 expanded rice leaves were inoculated with M. oryzae. The disease response was recorded 7 dpi by measuring the lesion length of the infected leaves, based on which the mean and standard deviation were calculated.
The RWG line was selected from Oryza sativa varieties with the LRR-RLP gene with many mutations through insertions/deletions (indels) analysis. Consequently, 9 RWG lines were selected, which are listed in Table S1. Line 3D-01328 carrying a T-DNA-GUS insertion in Os02g39660 (cv. Dongjin) was detected in a T-DNA insertion mutant population [35].
4.2. Gene Identification and Sequence Analysis
We identified OsLRR-RLP2 proteins from the Rice Genome Annotation Project (RGAP, http://rice.uga.edu/, accessed on 21 February 2022) using a Pfam domain search. We researched OsLRR-RLP2 using CAFRI-Rice (https://cafri-rice.khu.ac.kr/, accessed on 21 February 2022). To construct the phylogenetic tree, we collected protein sequences using NCBI blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 13 June 2022) and Phytozome (https://phytozome-next.jgi.doe.gov/, accessed on 13 June 2022). Phylogenetic analysis was performed using the neighbor-joining method with 500 bootstrap repeats in MEGA4. Three-dimensional (3D) models were predicted using OsLRR-RLP2 and AtRLP19 as templates in AlphaFold (https://alphafold.ebi.ac.uk/, accessed on 13 July 2022).
SNP and haplotype variations were confirmed using the SNP-Seek database (https://snp-seek.irri.org/, accessed on 26 November 2022). An allele frequency chart was retrieved from Rice Pan-genome Browser (https://cgm.sjtu.edu.cn/3kricedb/index.php, accessed on 26 November 2022).
4.3. Construction of Vectors and Plant Transformation
To construct the plasmids, a guide RNA for the CRISPR/Cas9 vector was designed, and two target regions were selected using CRISPRdirect software (http://crispr.dbcls.jp/, accessed on 1 December 2021). The gRNA fragment with annealed primers was ligated into the cloning site of the pRGEB32 binary vector [36]. In addition, the overexpression lines were cloned using OsLRR-RLP2 CDS and Ubiquitin promoter into pGA3426 fusion vector. The vectors were transformed into Escherichia coli TOP10, and the verified plasmid was transformed into Agrobacterium tumefaciens LBA4404 for rice transformation. Rice calli were cocultured with Agrobacterium to generate transgenic rice [37].
4.4. DNA Isolation and Genotypic Analysis
Rice leaves were sampled for genomic DNA extraction using the classical cetyltrimethylammonium bromide (CTAB) extraction buffer. gDNA was used to sequence the CRISPR/cas9 mutants and RWG lines. A PCR-based genotyping assay was performed to screen for homozygous transgenic lines and genotype SNPs. The primers used are listed in Supplementary Table S2. In addition, fusion transcript T-DNA-tagged genes were detected via amplified PCR using the synthesized DNA as the template, GUS primers, and gene-specific primers (Table S2).
4.5. RNA Isolation, cDNA Synthesis, and qRT-PCR
Total RNA was extracted using the leaf tissue infected with M. oryzae strains with an RNeasy Plant Mini kit (Qiagen, Hilden, Germany). cDNA was synthesized from RNA (Takara, Shiga, Japan). qRT-PCR analysis was performed using a Roter-Gene Q system (Qiagen, Hilden, Germany) and 2X SYBR Master mix. Relative expression was calculated with qRT-PCR using the threshold cycle value (CT) of each target gene against the CT value of the rice genomic ubiquitin (OsUbi) gene according the 2−ΔΔCT method [38]. qRT-PCR was conducted with three replications. The primer sequences used for qRT-PCR analysis are listed in Table S2.
4.6. Generation of Fusion Proteins and Subcellular Localization Analysis
For in silico analysis, online software, Plant-mSubP (https://bioinfo.usu.edu/Plant-mSubP/, accessed on 6 January 2023) and LocTree3 (https://rostlab.org/services/loctree3/, accessed on 6 January 2023), was used to determine the subcellular localization of OsLRR-RLP2 protein.
Nicotiana benthamiana localization assays were performed as described by Sparkes et al. [39]. The entire coding region of OsLRR-RLP2 was amplified using primers LRR-RLP_F and LRR-RLP_R (Table S2). Validated cDNA inserts were cloned into the p35S:OsLRR-RLP2-GFP fusion vector at the 5’-end of the GFP gene via digestion with HindIII and EcoRI. The resulting fusion construct and control vector (p35S:GFP) were transformed into A. tumefaciens strain GV3101. Transformed GV3101 was infiltrated into N. benthamiana leaves. Transient expression in N. benthamiana leaves was observed using a K1-Fluo confocal microscope (Nanoscope System, Daejeon, South Korea) after 72 h of infiltration and observed with green fluorescent protein (GFP) at 480 nm excitation and 550 nm emission wavelengths.
To detect the membrane localization of OsLRR-RLP2, FM4-64 staining of N. benthamiana leaves was performed and observed with red fluorescent protein (RFP) at 400 nm excitation and 500 nm emission wavelengths.
4.7. Histochemical GUS Assay
Leaves were immersed in a GUS staining solution [40]. After the tissues had been incubated at 37 °C for 2 h, the chlorophyll was removed in 70% ethanol. The samples were photographed using a BX21 microscope (Olympus, Tokyo, Japan).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
H.J.K., J.W.J., V.T., J.H.K., T.P., C.W.P., Y.S.G., and E.J.K. performed the experiments; S.W.K. provided resources; J.S.J. and S.T.K. reviewed and edited the manuscript; H.J.K. and Y.J.K. analyzed the data and wrote the paper; K.H.J. and Y.J.K. supervised the experiments. All the authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the National Research Foundation, Ministry of Education, Science, and Technology (NRF-2021R1A4A2001968 to YJK and NRF-2021K1A3A1A61002988 to KHJ).
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dean, R.; Van Kan, J. A.; Pretorius, Z. A.; Hammond-Kosack, K. E.; Di Pietro, A.; Spanu, P. D.; Rudd, J. J.; Dickman, M.; Kahmann, R.; Ellis, J., The Top 10 fungal pathogens in molecular plant pathology. Molecular plant pathology 2012, 13, 414-430. [CrossRef]
- Skamnioti, P.; Gurr, S. J., Against the grain: safeguarding rice from rice blast disease. Trends in biotechnology 2009, 27, 141-150. [CrossRef]
- Couto, D.; Zipfel, C., Regulation of pattern recognition receptor signalling in plants. Nature Reviews Immunology 2016, 16, 537-552. [CrossRef]
- Cui, H.; Tsuda, K.; Parker, J. E., Effector-triggered immunity: from pathogen perception to robust defense. Annual review of plant biology 2015, 66, 487-511. [CrossRef]
- Li, D.; Wu, M., Pattern recognition receptors in health and diseases. Signal transduction and targeted therapy 2021, 6, 291. [CrossRef]
- Boutrot, F.; Zipfel, C., Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Annual review of phytopathology 2017, 55, 257-286. [CrossRef]
- Wang, G.; Ellendorff, U.; Kemp, B.; Mansfield, J. W.; Forsyth, A.; Mitchell, K.; Bastas, K.; Liu, C.-M.; Woods-Tör, A.; Zipfel, C., A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant physiology 2008, 147, 503-517. [CrossRef]
- Jones, D. A.; Thomas, C. M.; Hammond-Kosack, K. E.; Balint-Kurti, P. J.; Jones, J. D., Isolation of the tomato Cf-9 gene for resistance to Cladosporium fulvum by transposon tagging. Science 1994, 266, 789-793. [CrossRef]
- Nadeau, J. A.; Sack, F. D., Control of stomatal distribution on the Arabidopsis leaf surface. Science 2002, 296, 1697-1700. [CrossRef]
- Zipfel, C., Plant pattern-recognition receptors. Trends in immunology 2014, 35, 345-351. [CrossRef]
- Fritz-Laylin, L. K.; Krishnamurthy, N.; Tör, M.; Sjölander, K. V.; Jones, J. D., Phylogenomic analysis of the receptor-like proteins of rice and Arabidopsis. Plant physiology 2005, 138, 611-623. [CrossRef]
- Jehle, A. K.; Lipschis, M.; Albert, M.; Fallahzadeh-Mamaghani, V.; Fürst, U.; Mueller, K.; Felix, G., The receptor-like protein ReMAX of Arabidopsis detects the microbe-associated molecular pattern eMax from Xanthomonas. The Plant Cell 2013, 25, 2330-2340. [CrossRef]
- Thomas, C. M.; Jones, D. A.; Parniske, M.; Harrison, K.; Balint-Kurti, P. J.; Hatzixanthis, K.; Jones, J. D., Characterization of the tomato Cf-4 gene for resistance to Cladosporium fulvum identifies sequences that determine recognitional specificity in Cf-4 and Cf-9. The Plant Cell 1997, 9, 2209-2224. [CrossRef]
- Zhang, H.; Chen, C.; Li, L.; Tan, X.; Wei, Z.; Li, Y.; Li, J.; Yan, F.; Chen, J.; Sun, Z., A rice LRR receptor-like protein associates with its adaptor kinase OsSOBIR1 to mediate plant immunity against viral infection. Plant Biotechnology Journal 2021, 19, 2319-2332. [CrossRef]
- Chen, E.; Huang, X.; Tian, Z.; Wing, R. A.; Han, B., The genomics of Oryza species provides insights into rice domestication and heterosis. Annual review of plant biology 2019, 70, 639-665. [CrossRef]
- Garris, A. J.; Tai, T. H.; Coburn, J.; Kresovich, S.; McCouch, S., Genetic structure and diversity in Oryza sativa L. Genetics 2005, 169, 1631-1638. [CrossRef]
- Escolà, G.; González-Miguel, V. M.; Campo, S.; Catala-Forner, M.; Domingo, C.; Marqués, L.; San Segundo, B., Development and Genome-Wide Analysis of a Blast-Resistant japonica Rice Variety. Plants 2023, 12, 3536. [CrossRef]
- Enkhbayar, P.; Kamiya, M.; Osaki, M.; Matsumoto, T.; Matsushima, N., Structural principles of leucine-rich repeat (LRR) proteins. Proteins: Structure, Function, and Bioinformatics 2004, 54, 394-403. [CrossRef]
- Lee, D.; Zhang, H.; Zeng, Y.; Kim, B.; Kwon, S.-W., Discovery of Genomic Regions and Candidate Genes for Awn Length Using QTL-seq in Rice (Oryza sativa L.). 2023. [CrossRef]
- Ray, D. K.; Mueller, N. D.; West, P. C.; Foley, J. A., Yield trends are insufficient to double global crop production by 2050. PloS one 2013, 8, e66428. [CrossRef]
- van Ooijen, G.; van den Burg, H. A.; Cornelissen, B. J.; Takken, F. L., Structure and function of resistance proteins in solanaceous plants. Annu. Rev. Phytopathol. 2007, 45, 43-72. [CrossRef]
- Gust, A. A.; Felix, G., Receptor like proteins associate with SOBIR1-type of adaptors to form bimolecular receptor kinases. Current opinion in plant biology 2014, 21, 104-111. [CrossRef]
- Sun, Y.; Wang, Y.; Zhang, X.; Chen, Z.; Xia, Y.; Wang, L.; Sun, Y.; Zhang, M.; Xiao, Y.; Han, Z., Plant receptor-like protein activation by a microbial glycoside hydrolase. Nature 2022, 610, 335-342. [CrossRef]
- Albert, I.; Zhang, L.; Bemm, H.; Nürnberger, T., Structure-function analysis of immune receptor at RLP23 with its ligand nlp20 and coreceptors at SOBIR1 and At BAK1. Molecular Plant-Microbe Interactions 2019, 32, 1038-1046. [CrossRef]
- Zhang, W.; Fraiture, M.; Kolb, D.; Löffelhardt, B.; Desaki, Y.; Boutrot, F. F.; Tör, M.; Zipfel, C.; Gust, A. A.; Brunner, F., Arabidopsis receptor-like protein30 and receptor-like kinase suppressor of BIR1-1/EVERSHED mediate innate immunity to necrotrophic fungi. The Plant Cell 2013, 25, 4227-4241. [CrossRef]
- Min, C. W.; Jang, J. W.; Lee, G. H.; Gupta, R.; Yoon, J.; Park, H. J.; Cho, H. S.; Park, S. R.; Kwon, S.-W.; Cho, L.-H., TMT-based quantitative membrane proteomics identified PRRs potentially involved in the perception of MSP1 in rice leaves. Journal of Proteomics 2022, 267, 104687. [CrossRef]
- Singh, J.; Gupta, S. K.; Devanna, B.; Singh, S.; Upadhyay, A.; Sharma, T. R., Blast resistance gene Pi54 over-expressed in rice to understand its cellular and sub-cellular localization and response to different pathogens. Scientific reports 2020, 10, 5243. [CrossRef]
- Wang, G.-F.; Balint-Kurti, P. J., Cytoplasmic and nuclear localizations are important for the hypersensitive response conferred by maize autoactive Rp1-D21 protein. Molecular Plant-Microbe Interactions 2015, 28, 1023-1031. [CrossRef]
- Bai, S.; Liu, J.; Chang, C.; Zhang, L.; Maekawa, T.; Wang, Q.; Xiao, W.; Liu, Y.; Chai, J.; Takken, F. L., Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS pathogens 2012, 8, e1002752. [CrossRef]
- Wu, Y.; Xun, Q.; Guo, Y.; Zhang, J.; Cheng, K.; Shi, T.; He, K.; Hou, S.; Gou, X.; Li, J., Genome-wide expression pattern analyses of the Arabidopsis leucine-rich repeat receptor-like kinases. Molecular plant 2016, 9, 289-300. [CrossRef]
- Rodrigues, F. Á.; Benhamou, N.; Datnoff, L. E.; Jones, J. B.; Bélanger, R. R., Ultrastructural and cytochemical aspects of silicon-mediated rice blast resistance. Phytopathology 2003, 93, 535-546. [CrossRef]
- Dolatabadian, A.; Bayer, P. E.; Tirnaz, S.; Hurgobin, B.; Edwards, D.; Batley, J., Characterization of disease resistance genes in the Brassica napus pangenome reveals significant structural variation. Plant biotechnology journal 2020, 18, 969-982. [CrossRef]
- Petre, B.; Hacquard, S.; Duplessis, S.; Rouhier, N., Genome analysis of poplar LRR-RLP gene clusters reveals RISP, a defense-related gene coding a candidate endogenous peptide elicitor. Frontiers in plant science 2014, 5, 111. [CrossRef]
- Wu, Y.; Xiao, N.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Liu, G.; Dai, Z.; Pan, X.; Li, A., Combination patterns of major R genes determine the level of resistance to the M. oryzae in rice (Oryza sativa L.). PLoS One 2015, 10, e0126130. [CrossRef]
- An, S.; Park, S.; Jeong, D.-H.; Lee, D.-Y.; Kang, H.-G.; Yu, J.-H.; Hur, J.; Kim, S.-R.; Kim, Y.-H.; Lee, M., Generation and analysis of end sequence database for T-DNA tagging lines in rice. Plant physiology 2003, 133, 2040-2047. [CrossRef]
- Xie, K.; Minkenberg, B.; Yang, Y., Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proceedings of the National Academy of Sciences 2015, 112, 3570-3575. [CrossRef]
- Lee, S.; Jeon, J.-S.; Jung, K.-H.; An, G., Binary vectors for efficient transformation of rice. Journal of Plant Biology 1999, 42, 310-316. [CrossRef]
- Livak, K. J.; Schmittgen, T. D., Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods 2001, 25, 402-408. [CrossRef]
- Sparkes, I. A.; Runions, J.; Kearns, A.; Hawes, C., Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nature protocols 2006, 1, 2019-2025. [CrossRef]
- Moon, S.; Chandran, A. K. N.; Kim, Y.-J.; Gho, Y.; Hong, W.-J.; An, G.; Lee, C.; Jung, K.-H., Rice RHC encoding a putative cellulase is essential for normal root hair elongation. Journal of Plant Biology 2019, 62, 82-91. [CrossRef]
Figure 1.
Heatmap analysis for indel (≥3 bp) frequency of 453 high-quality accessions compared to Nipponbare of 857 leucine-rich repeat domain (LRR) genes in rice. (A) Heatmap representing genes with LRR domains between difference species, (B) 277 genes significanly differentially expressed under biotic stress, and (C) 16 genes upregulated under blast and white bacterial blight disease treatments.
Figure 1.
Heatmap analysis for indel (≥3 bp) frequency of 453 high-quality accessions compared to Nipponbare of 857 leucine-rich repeat domain (LRR) genes in rice. (A) Heatmap representing genes with LRR domains between difference species, (B) 277 genes significanly differentially expressed under biotic stress, and (C) 16 genes upregulated under blast and white bacterial blight disease treatments.
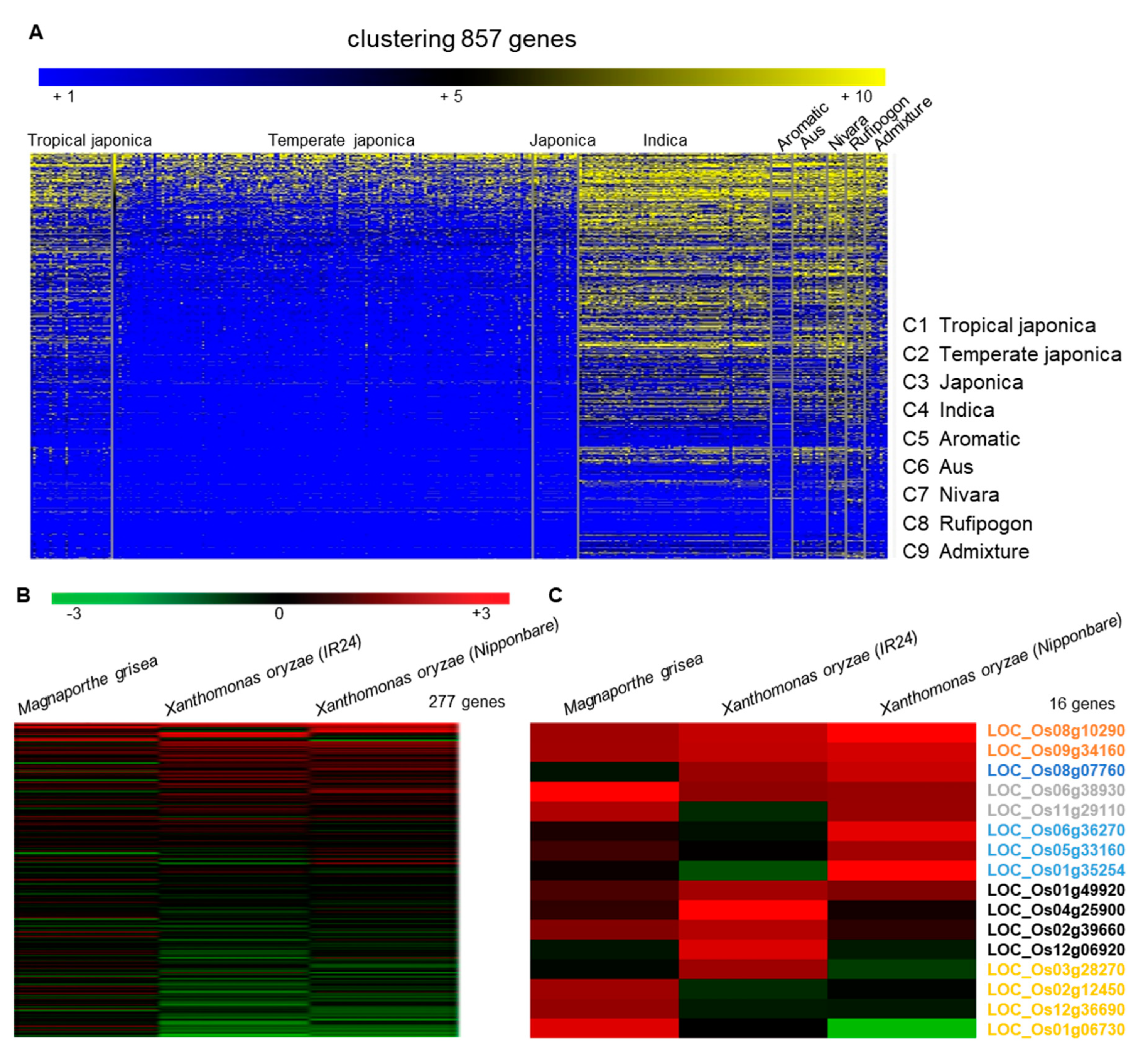
Figure 2.
Results of protein domain, phylogenetic tree, and gene expressions analyses. (A) Schematic representation of OsLRR-RLP2 using SMART (http://smart.embl-heidelberg.de/, accessed on 21 February 2022). LRR, leucine-rich repeat domain; LRRNT, leucine-rich repeat N-terminus. (B) Phylogenetic tree of OsLRR-RLP2. The protein sequences were identified using NCBI BLASTX. Phylogenetic tree based on the neighbor-joining method was constructed using MEGA 4 with 500 bootstrap replications. (C) Three-dimensional modeling of OsLRR-RLP2 and AtRLP19 protein. (D) CAFRI-RICE analysis of OsLRR-RLP2. OsLRR-RLP2 showed high expression in leaves. (E) Validation of OsLRR-RLP2 expression using qRT-PCR analysis. The roots and shoots of 10-day-old seedlings, flowers, and 3-month-old leaves were analyzed. Three technical replicates were performed, and the results were normalized using rice ubiquitin (OsUbi5, Os01g22490).
Figure 2.
Results of protein domain, phylogenetic tree, and gene expressions analyses. (A) Schematic representation of OsLRR-RLP2 using SMART (http://smart.embl-heidelberg.de/, accessed on 21 February 2022). LRR, leucine-rich repeat domain; LRRNT, leucine-rich repeat N-terminus. (B) Phylogenetic tree of OsLRR-RLP2. The protein sequences were identified using NCBI BLASTX. Phylogenetic tree based on the neighbor-joining method was constructed using MEGA 4 with 500 bootstrap replications. (C) Three-dimensional modeling of OsLRR-RLP2 and AtRLP19 protein. (D) CAFRI-RICE analysis of OsLRR-RLP2. OsLRR-RLP2 showed high expression in leaves. (E) Validation of OsLRR-RLP2 expression using qRT-PCR analysis. The roots and shoots of 10-day-old seedlings, flowers, and 3-month-old leaves were analyzed. Three technical replicates were performed, and the results were normalized using rice ubiquitin (OsUbi5, Os01g22490).
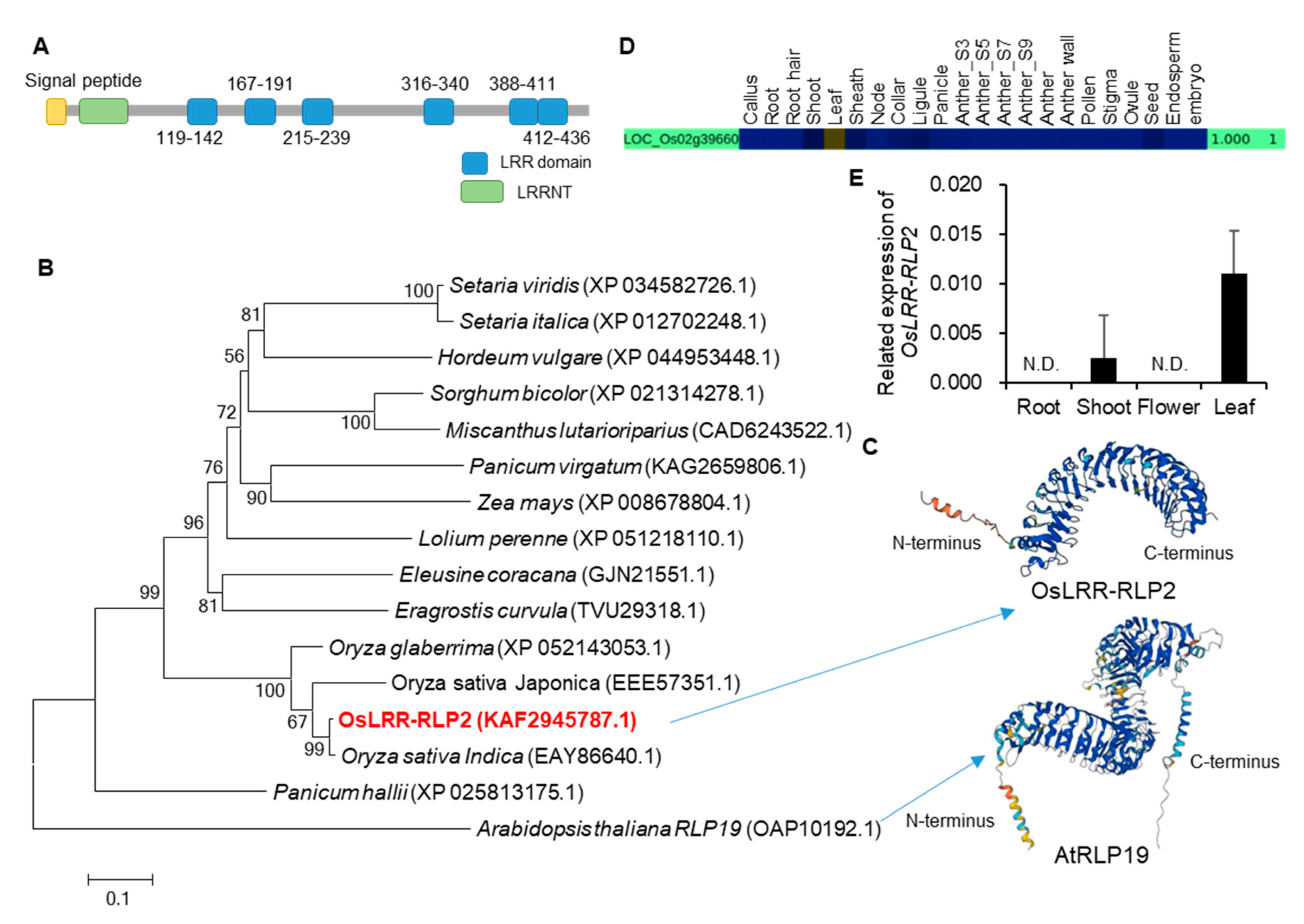
Figure 3.
Subcellular localization of OsLRR-RLP2. GFP signals of OsLRR-RLP2 in tobacco epidermal cells stained with the membrane marker FM4-64. The tobacco leaves were treated with 1 N NaCl to induce plasmolysis. (A,C) Control vector (p35S:GFP) expressed in epidermal cells of tobacco leaves. (B,D) Recombinant vector (p35S:OsLRR-RLP2-GFP) expressed in epidermal cells of tobacco leaves. GFP, green fluorescent protein; BF, bright-field; scale bar: 30 μm.
Figure 3.
Subcellular localization of OsLRR-RLP2. GFP signals of OsLRR-RLP2 in tobacco epidermal cells stained with the membrane marker FM4-64. The tobacco leaves were treated with 1 N NaCl to induce plasmolysis. (A,C) Control vector (p35S:GFP) expressed in epidermal cells of tobacco leaves. (B,D) Recombinant vector (p35S:OsLRR-RLP2-GFP) expressed in epidermal cells of tobacco leaves. GFP, green fluorescent protein; BF, bright-field; scale bar: 30 μm.
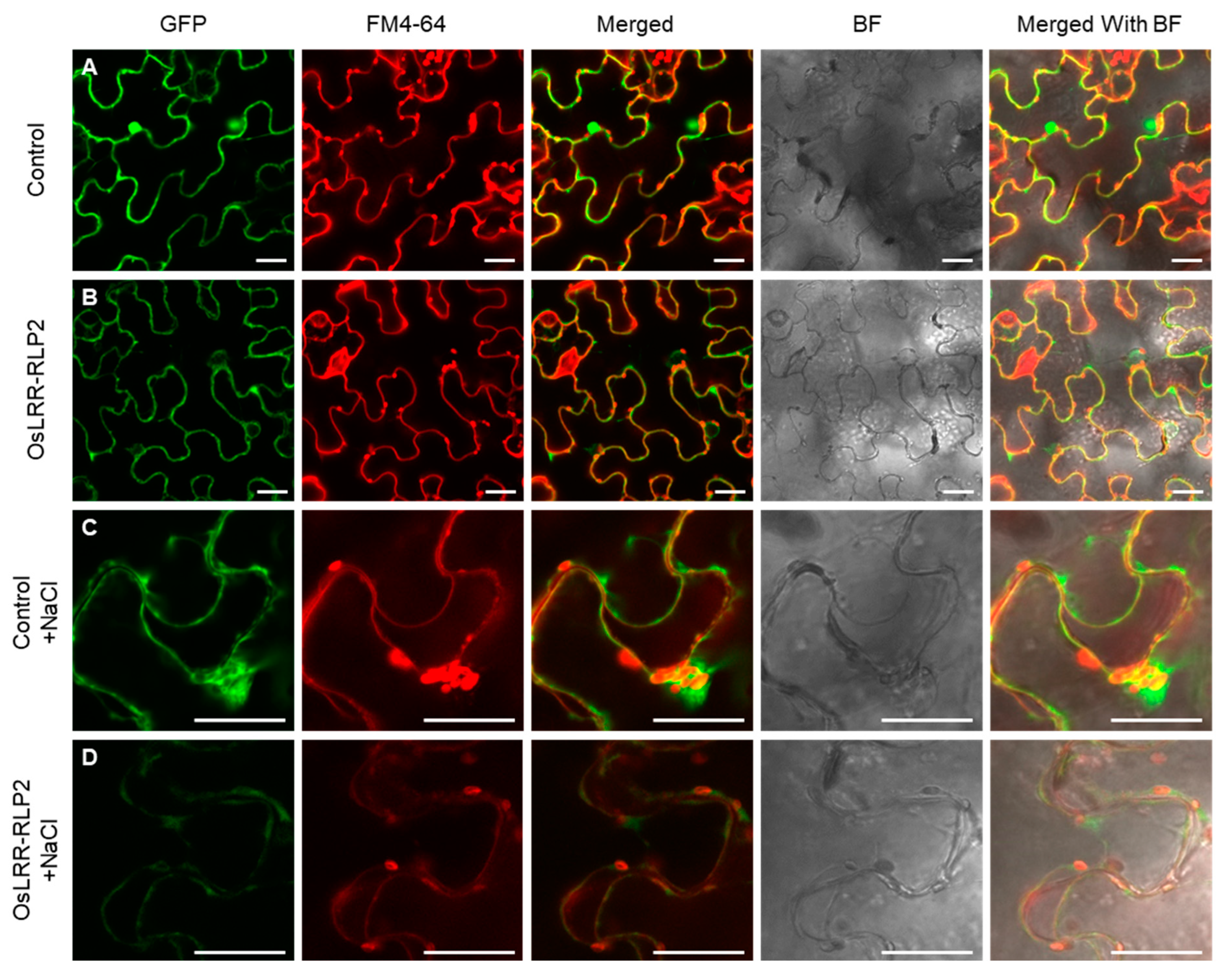
Figure 4.
Single-nucelotide polymorphisms (SNPs) and synteny of the genomic sequences of OsLRR-RLP2. (A) Phylogenetic tree constructed based on presence/absence variations (PAVs) among 453 high-quality accessions. OsLRR-RLP2 can be classified into japonica-dominant genes and Indica-subgroup-unbalanced genes. (B) Allele frequencies of OsLRR-RLP2 in 3024 accessions available from SNP-Seek database. Numbers on X-axis represent positions of indels. Numbers on Y-axis indicate allele frequency in percentage. (C) SNPs of OsLRR-RLP2 gene across the ~3024 rice accessions depicted based on the sequence variation data retrieved from the SNP-Seek database.
Figure 4.
Single-nucelotide polymorphisms (SNPs) and synteny of the genomic sequences of OsLRR-RLP2. (A) Phylogenetic tree constructed based on presence/absence variations (PAVs) among 453 high-quality accessions. OsLRR-RLP2 can be classified into japonica-dominant genes and Indica-subgroup-unbalanced genes. (B) Allele frequencies of OsLRR-RLP2 in 3024 accessions available from SNP-Seek database. Numbers on X-axis represent positions of indels. Numbers on Y-axis indicate allele frequency in percentage. (C) SNPs of OsLRR-RLP2 gene across the ~3024 rice accessions depicted based on the sequence variation data retrieved from the SNP-Seek database.
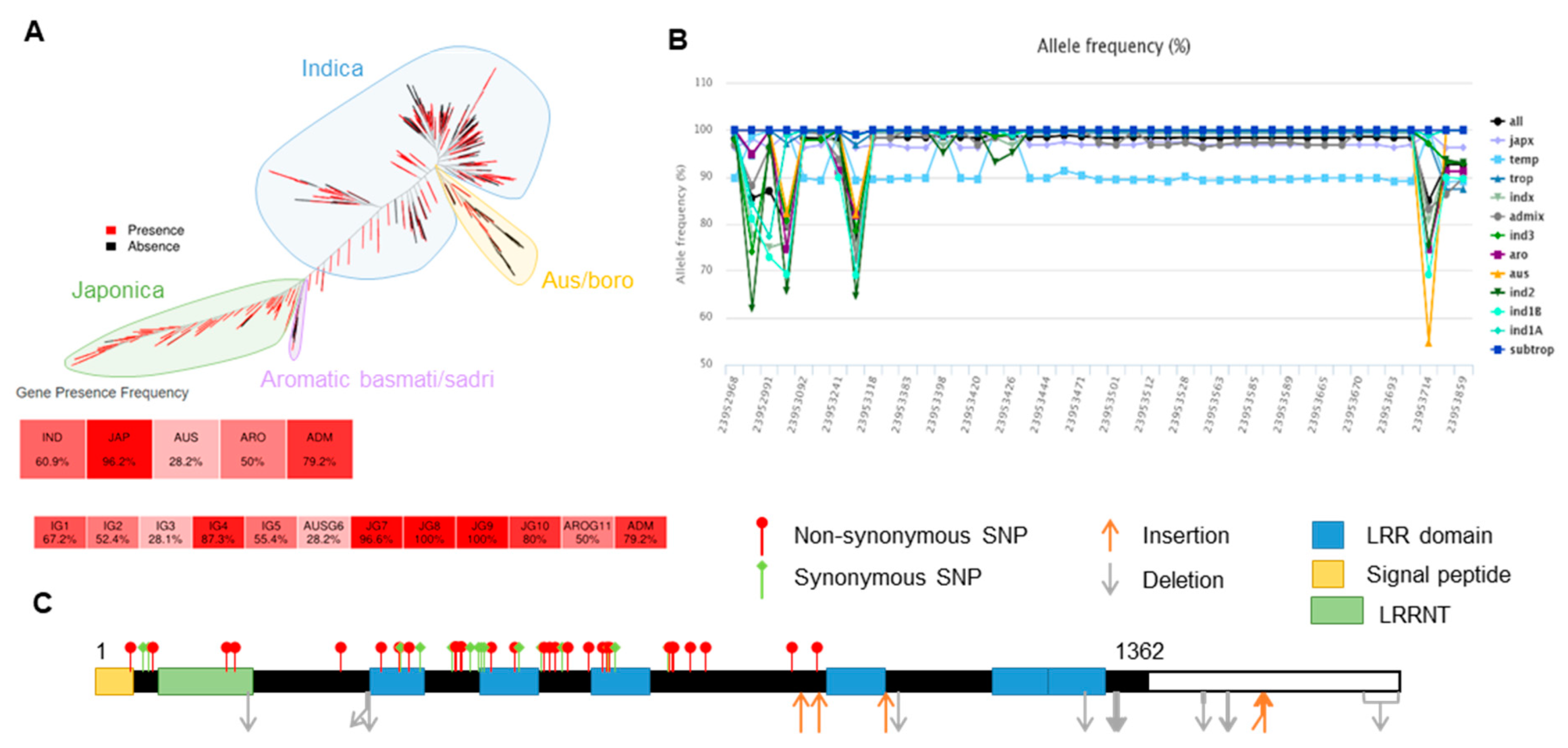
Figure 5.
Inoculation of DJ and 9 RWG lines in rice blast disease. (A) Representative leaves showing disease lesions in five-week-old from RWG lines inoculated with M. oryzae (PO6-6). The leaves were photographed 7 days postinoculation (dpi). Scale bar =1 cm. (B) Disease lesion lengths in leaves of five-week-old RWG lines were measured 7 dpi. (C) Relative fungal biomass w determined by examining the expression level of M. oryzae Pot2 against OsUbiquitin DNA level. Three technical replicates were performed. Error bars indicate ± SD.
Figure 5.
Inoculation of DJ and 9 RWG lines in rice blast disease. (A) Representative leaves showing disease lesions in five-week-old from RWG lines inoculated with M. oryzae (PO6-6). The leaves were photographed 7 days postinoculation (dpi). Scale bar =1 cm. (B) Disease lesion lengths in leaves of five-week-old RWG lines were measured 7 dpi. (C) Relative fungal biomass w determined by examining the expression level of M. oryzae Pot2 against OsUbiquitin DNA level. Three technical replicates were performed. Error bars indicate ± SD.
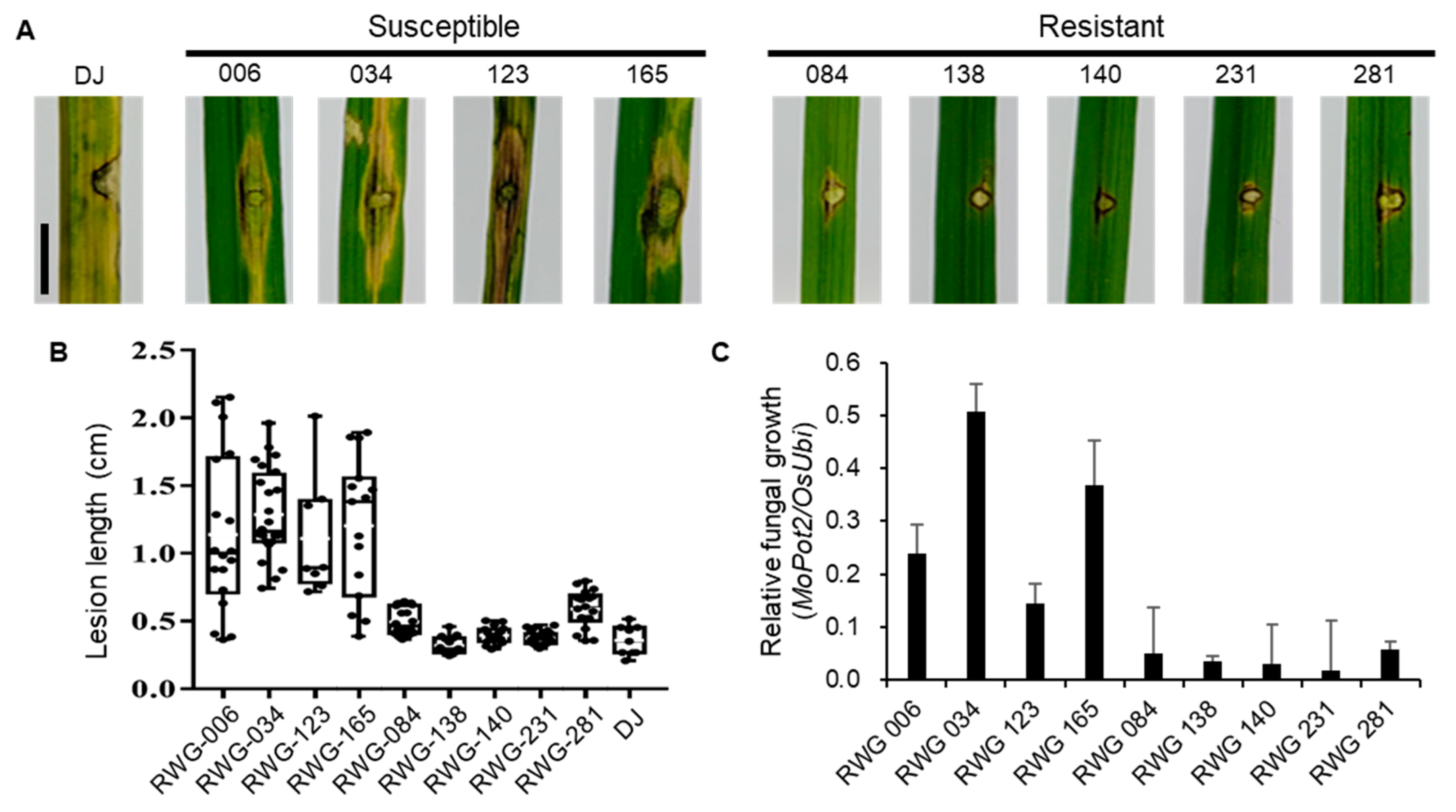
Figure 6.
Functional analysis of OsLRR-RLP2. (A) GUS staining of T-DNA plants. Expression appears in main vein tissue of the leaf. Scale bar = 1 cm. (B) Representative leaves from WT, OsLRR-RLP2-OE, and oslrr-rlp2-cas inoculated with M. oryzae (PO6-6). The leaves were photographed 7 dpi. Scale bar = 1 cm. WT, wild-type; OE, overexpression. (C) Disease lesion lengths in five-week-old leaves from WT and transgenic plants were measured 7 dpi with M. oryzae (PO6-6). (D) Relative fungal biomass was determined by examining the expression levels of M. oryzae Pot2 and OsUbiquitin DNA. Three technical replicates were performed. Error bars indicate ± SD. Asterisks indicate significant differences between samples according to the t-test (***P < 0.001). (E) Representative leaves from WT and oslrr-rlp2 T-DNA HM inoculated with M. oryzae (PO6-6). The leaves were photographed 7 dpi. (F) Disease lesion lengths in five-week-old leaves from WT and T-DNA HM were measured 7 dpi with M. oryzae (PO6-6). Asterisks indicate significant differences between samples according to the t-test (*P < 0.05). (G) Model for defense activation by OsLRR-RLP2.
Figure 6.
Functional analysis of OsLRR-RLP2. (A) GUS staining of T-DNA plants. Expression appears in main vein tissue of the leaf. Scale bar = 1 cm. (B) Representative leaves from WT, OsLRR-RLP2-OE, and oslrr-rlp2-cas inoculated with M. oryzae (PO6-6). The leaves were photographed 7 dpi. Scale bar = 1 cm. WT, wild-type; OE, overexpression. (C) Disease lesion lengths in five-week-old leaves from WT and transgenic plants were measured 7 dpi with M. oryzae (PO6-6). (D) Relative fungal biomass was determined by examining the expression levels of M. oryzae Pot2 and OsUbiquitin DNA. Three technical replicates were performed. Error bars indicate ± SD. Asterisks indicate significant differences between samples according to the t-test (***P < 0.001). (E) Representative leaves from WT and oslrr-rlp2 T-DNA HM inoculated with M. oryzae (PO6-6). The leaves were photographed 7 dpi. (F) Disease lesion lengths in five-week-old leaves from WT and T-DNA HM were measured 7 dpi with M. oryzae (PO6-6). Asterisks indicate significant differences between samples according to the t-test (*P < 0.05). (G) Model for defense activation by OsLRR-RLP2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



