
Submitted:
11 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
Contamination of the environment with nano- and microplastic particles exerts a threatened impact on the aquatic ecosystems and sustainable catfish aquaculture. The presence of nanoplastics has been found to have a detrimental impact on both aquatic and terrestrial ecosystems. The present study examines the effect of polystyrene nanoplastics on DNA, erythrocytes, oxidative status and renal of catfish in addition to the potential protective effects of Chlorella vulgaris bioremeditation and selenium to hinder this effect. Six equal groups of fish were used as follows: Group 1: served as a control group and received water free from PS NPs; Group 2: was exposed to PS NPs at a concentration of 5mg/L; Group 3: was exposed to PS NPs (5mg/L) + selenium (1mg/kg diet); Group 4: was exposed to PS NPs (5mg/L) + C. vulgaris (25g/kg diet); Group 5: was supplemented with C. vulgaris (25g/kg diet); and Group 6: was supplemented with selenium (1mg/kg diet). The results indicated that PS NPs induced oxidative stress by significantly elevating MDA activities and slightly reducing antioxidant biomarkers, resulting in DNA damage, increased frequency of micronuclei, erythrocyte alterations, and numerous histopathological alterations in kidney tissue. Selenium and C. vulgaris significantly ameliorated the oxidative/antioxidant status, reduced DNA damage, micronucleus frequency, erythrocyte alterations, and morphology of kidney tissue. Nevertheless, further research is needed to evaluate in detail the mechanism behind the toxicity with nano-microplatics in aquatic system.
Keywords:
Polystyrene nanoplastics
; Oxidative stress
; Single cell gel electrophoresis
; Selenium
; Chlorella vulgaris
1. Introduction
Polystyrene nanoplastics (PS NPs) are sub-100 nm-sized plastic particles [1]. They can penetrate biological barriers and accumulate along the food chain, posing significant health risks to humans and animals [2].
Microplastics and nanoplastics (M/NPs) are present in various ecosystems such as soils, air, marine, and freshwater environments, as reported by [3]. The issue of water contamination caused by plastics has been recognized as a significant environmental peril in numerous countries worldwide, as stated by [4]. According to research conducted by [5,6], Egypt is the largest plastic consumer in Africa and ranks 7th among 192 countries in producing poorly managed plastic waste. The country generates an annual output of 0.97 million tons of such waste. plastic wastes are exposed to chemical, biological and environmental elements, and will break down into huge amounts of microplastics and nanoplastics [7].
Recent studies have reported that micro- and nanoplastics reach the human food chain in several ways for example consuming them in their natural environment, food contamination during preparation processes, or through leaching from plastic packaging of food and drinks [8,9,10].
Polystyrene nanoplastics have the ability to permeate the gut epithelium, leading to systemic exposure in humans. Several in-vivo and in-vitro studies on animals found that the probable oral bioavailability level of 50 nm polystyrene nanoparticles is ten to one hundred times greater than the level of microplastics [11,12].
There are three routes for exposure to microplastics and nanoplastics: oral, inhalation, and dermal [13,14]. After absorption, nanoplastics will be able to interact with numerous target cells and induce their toxic effects on kidney, gut, liver and other organs [15]. Extensive studies have shown that NPs can lead to different biological adverse effects, such as oxidative stress [16], cytotoxicity, genotoxicity [17], neurotoxicity [18], reproductive and developmental toxicity and immune dysfunction [19]. Furthermore, they have been observed to have detrimental effects on the survival and reproduction of aquatic organisms [20].
Clarias gariepinus inhabits various freshwater habitats, such as ponds, lakes, and pools. In addition, they are evident in rivers, rapids, and near dams. They can survive in water with a pH range of 6.5 to 8.0 and exceedingly low oxygen concentrations [21]. Their bodies produce mucous to prevent dehydration [22]. They can tolerate temperatures between 8 and 35 degrees Celsius and persist in extremely murky environments. The optimal temperature for their growth is between 28 and 30 degrees Celsius [23]. Clarias gariepinus is widely utilized in aquaculture because it is readily available throughout the year, high food conversion ratio, fast growth rate, and adapts well to laboratory conditions and stress, making it an ideal test organism [24].
Chlorella vulgaris, agreen unicellular microalgae is used in aquatic bioremediation [25]. It exhibits potent antioxidant properties due to the presence of carotenoids and phenolic bioactive compounds responsible for its antioxidant and radical-scavenging activities [26]. On the other hand selenium (Se) is necessary for fish's development and metabolic processes [27]. Selenium can be found in many essential selenoproteins and enzymes that maintain antioxidant status, DNA synthesis, fertility, reproduction, and reduce inflammation [28]. Chromosomal aberration assay, comet assay, and various types of micronucleus (MN) assays, including mammalian erythrocyte MN (EMN), are considered crucial techniques in In vivo genotoxicity research in fish [29,30,31].
The MN tests of fish erythrocytes have been commonly employed to assess genotoxicity in marine and freshwater environments [32]. These tests have also served as an indicator of the exposure of various fish species to mutagenic or genotoxic pollutants caused by numerous substances [33].
There is a dearth of research evaluating the genotoxic effect of PS NPs, the effect on erythrocyte morphology and the patholgical changes in the kidney of catfish. Also, the potential ameliorative effects of selenium and bioremediation by Chlorella vulgaris. Therefore, the present study assessed the efficacy of L-Selenomethionine and Chlorella vulgaris in mitigating the harmful effects of PS NPs on DNA, RBCs, and kidney of C. gariepinus.
2. Materials and Methods
2.1. Chemicals and materials
The Polystyrene nanoplastic particles (PS NPs) were procured from Bangs Laboratories, Inc. The procurement of Seleno-L-methionine powder was carried out from Sigma-Aldrich, USA. The powder was high purity (≥ 98%) and had a molecular weight 196.11. Additionally, the powder was identified by its CAS number, 3211-76-5. The dried green powder of Chlorella vulgaris microalgae was procured from the Institute of National Research Center, Cairo, Egypt.
Before conducting the experiments, a stock solution of PS NPs at a concentration of 5 g L−1 was prepared using deionized water. Then, the dimensions and morphology of the nanoplastics were assessed using a JEOL-JEM-100 CX II transmission electron microscope (TEM) located in the electron microscopy unit at Assiut University, Egypt.
2.2. Clarias gariepinus
For the study, a sample of 144 male juvenile catfish (C. gariepinus) was obtained from an aquaculture fish farm in Assiut, Egypt. The specimens exhibited an average weight of 160 g and a length of 25 and 30 cm. The age of the specimens was determined to be six months. The conveyance of fish to the Fish Biology Laboratory, situated within the Zoology Department of the Faculty of Science at Assiut University in Egypt, was executed. During a 35-day acclimatization period, the fish were subjected to laboratory conditions where 100 L tanks, dechlorinated tap water, and air pumps were used; this was executed before the initiation of the exposure. The piscine subjects were administered a diet comprising a commercial substrate with a crude protein content of 30%. The nutritional needs of C. gariepinus were satisfied by creating a fundamental diet comprising diverse constituents, with the estimated chemical makeup of each component outlined in Table 1.
2.3. Ethics statement
The research protocol mentioned above has received approval from the veterinary ethical committee of the Faculty of Veterinary Medicine at Assiut University, Egypt, under the reference number 06/2022/0016.
2.4. Experimental design
The fish were randomly put into six groups of 24 each, with three tanks in each group. The first group (C) was used as a control and got water that did not contain PS NPs. According to [34], the second group (NP) was exposed to 5 mg/L PS NPs in water. According to [35], the third group (NPs+Se) was given PS NPs (5 mg/L) and selenium (1 mg/kg food). According to [36], the fourth group (NPs+Ch) was exposed to PS NPs (5 mg/L) and C. vulgaris (25 g/kg food). The fifth group (Ch) got only C. vulgaris (25 g/kg food). The sixth group (Se) only got selenium (1 mg/kg food).
For 30 days, the animals were kept in tanks with 60 L of naturally dechlorinated water and air blowers for regular aeration. The experiment was done with steady water quality (temperature of 22°C, dissolved oxygen (mg/l) of 6.85, pH of 7.38, and 12:12 light: dark cycle). About 50% of the water was changed daily throughout the experiment, and PS NPs were added at the abovementioned amounts. In addition, waste products like feces were removed daily to keep the ammonia levels in the water as low as possible.
2.5. Sample collection
2.5.1. Blood samples
Following a period of 30 days of exposure, six distinct groups of fish, comprising both control and treatment groups, were captured at random using a hand net and subsequently transferred to ice in order to mitigate any potential stress, as per the study conducted by [37]. The blood collection methodology followed the approach described by [38]. First, EDTA-containing tubes were utilized to obtain blood samples from the caudal vein of C. gariepinus to detect DNA damage through the comet assay. Next, additional blood samples were collected using standard vacutainer tubes and subsequently subjected to centrifugation at 3000 revolutions per minute for 15 minutes to procure serum samples, which were then stored at a temperature of -20˚C. These samples assessed Malondialdehyde, catalase enzyme, and glutathione peroxidase enzyme levels. Finally, the blood samples were subjected to the preparation of blood smears, followed by drying at ambient temperature and fixation in absolute methanol to conduct the micronucleus assay and assess other nuclear abnormalities and erythrocyte morphological changes.
2.5.2. Tissue samples
After dissecting the fish, kidney tissues were rapidly removed, cleaned in normal saline to eliminate blood traces, and stored in 10% buffered formalin for histological investigation.
2.6. Oxidative stress biomarkers measurements
2.6.1. Determination of serum malondialdehyde (MDA) activity:
MDA was measured by spectrophotometric method (V-630, JASCO, Japan), using a commercial kit supplied by Bio Diagnostics, Egypt, with a catalog number MD 25 30 and according to the method described by [39].
2.6.2. Determination of serum catalase enzyme activity
Catalase assay was performed by spectrophotometric method (V-630, JASCO, Japan). According to [40] using test kits provided by Bio-diagnostic (Dokki, Giza, Egypt).
2.6.3. Determination of serum glutathione peroxidase (GPx) activity
Glutathione peroxidase activity was measured in serum samples using test kits provided by Bio-diagnostic (Dokki, Giza, Egypt). Providing a spectrophotometric means (V-630, JASCO, Japan) to detect the activity of GPx enzyme [41].
2.7. Genotoxicity assays
2.7.1. Detection of DNA damage by Single cell gel electrophoresis (Comet assay)
DNA damage in the erythrocyte of C. gariepinus was estimated using comet assay according to the method of [42], as follows:
- a)
- Preparation of slides
Slides were double coated with 1% standard agarose; 0.3 mL of blood was diluted in 1000 µL PBS; next, ten microliters of cell suspension was mixed with 120 µL of 2 % low melting point agarose at 37 °C, a layer of the mixture was placed on the previously prepared slides, cover-slipped and placed in the refrigerator for 5 min to solidify. Finally, 1 % agarose (100 μL) was layered on top and allowed to gel.
- b) Lysis and electrophoresis
All slides were kept in a cold lysis solution (10 mM Trizma, 100 mM Na4 EDTA, 2.5 M NaCl, 10% dimethyl sulfoxide, 0.1% sodium lauryl sulfate (SDS), and Triton X-100) at 4 °C for 60 minutes. Then transferred to the electrophoresis tank (Cleaver Scientific Ltd., UK) containing chilled alkaline solution (1 mM Na2 EDTA and 300 mM NaOH) at pH 13 for 25 min in the dark for unwinding of the DNA strands. Electrophoresis was run for 25 min at 25 V and approximately 300 mA. Next, slides were neutralized in 400 mM Tris buffer (0.4 M Tris-HCl, pH 7.5) for 7 min. Finally, the neutralized slides were dehydrated in ethanol for a few minutes and then dried at room temperature.
- c) Slide examination
The slides were examined through a fluorescence microscope (Olympus BX-43, Japan) with a green filter at a magnification of 200-fold. In addition, 50 μL of ethidium bromide at 20 μg/mL concentration was used to stain all the slides. In order to assess DNA migration, a minimum of 50 nuclei were examined per slide, with three slides per sample. In addition, the Comet Assay Software Project (CASP) was utilized to measure the diameter of the comet's head and the length of its tail.
2.7.2. Micronucleus and Erythrocyte morphological alterations
Three blood smears were prepared and dried at room temperature. For 10 minutes, the smears were fixed in methanol (100%) and stained with hematoxylin and eosin. The slides were then dehydrated with alcohol with increasing concentrations (30, 50, 70, and 90 percent, absolute) before being cleaned in xylene and permanently mounted with DPX [43]. According to [44], each nuclear abnormality observed during slide examination, including micronuclei formation, was noted. Ten photomicrographs of random fields were obtained for each slide, totaling 30 photomicrographs per experimental group, and one hundred cells were examined in each analysis field, totaling 3000 erythrocytes/group.
2.8. Histopathological examination of the kidney
Kidney specimens were fixed in neutral buffered formalin 10 %, then dehydrated in serial alcohol concentrations, cleared in xylene, and impregnated in paraffin. Five μm paraffin sections were prepared and then stained with the following stains: Hematoxylin and Eosin (H&E) according to [45], Periodic Acid Schiff (PAS) and Hematoxylin for polysaccharides (glycogen) [46] and Picrosirius red stain for collagen fibers [47].
2.9. Statistical analysis
The statistical analysis used SPSS software version 16.0 (SPSS Inc., Chicago, IL, USA). Initially, the statistical method of one-way analysis of variance (ANOVA) was employed, subsequently followed by a post hoc least significant difference (LSD) multiple range test to make comparisons between the control and exposed groups. Finally, the mean ± SE represented all outcomes for the control and experimental groups. The statistical significance level was established at a value of P < 0.05.
3. Results
3.1. Polystyrene nanoplastics characterization
The PS nanomaterials were analyzed using transmission electron microscopy (TEM), which revealed that they are made up of spherical particles with a diameter of 50.1 nm ±13.4 Figure 1.
3.2. Mortality rate
The results showed no deaths were recorded in the control group, while the NPs-exposed group showed a high mortality percentage of 36.3%. The administration of selenium reduced the rate of mortality to 14.8%. Similarly, the administration of C. vulgaris reduced the percentage of mortalities to 8.3%.
3.3. Oxidative status
3.3.1. Lipid peroxide
Serum MDA activity significantly increased (P < 0.01) in C. gariepinus exposed only to NPs compared to the control group. Conversely, treatment of fish with Se and Chlorella showed a significant (P < 0.01) decrease in the MDA activity in comparison with the NPs-exposed group in Table 2.
3.3.2. Catalase and glutathione peroxidase activities:
Catalase and glutathione peroxidase activities in the serum of C. gariepinus exposed to NPs showed a mild, non-significant (P > 0.05) decrease compared to the control group. While other groups showed no change in the activity of Catalase and GPX (Table 2).
3.4. Genotoxic biomarkers for DNA damage
3.4.1. DNA damage parameters (Comet assay)
The study's findings indicate that the administration of NPs resulted in a notable increase in DNA damage in the blood of C. gariepinus compared to the control group. In addition, the study observed a rise in damage indicators, namely tail DNA%, tail length, tail moment, and Olive tail moment, as depicted in (Figure 2 and Table 3). The different levels of DNA damage and the technique employed to evaluate DNA damage using CASP software are presented in Figure 3.
All parameters of DNA damage were significantly increased in the NPs exposed group (P < 0.01) compared with the control and other treated groups. Conversely, Chlorella (NPs+Ch) and selenium (NPs+SE)-treated groups demonstrated a significant decrease (P < 0.01) for all these parameters in comparison with the NPs-exposed group (Figure 2 and Table 3).
3.4.2. Erythrocyte micronucleus (EMN) and morphological alterations
The normal erythrocytes of C. gariepinus are rounded with a centrally located nucleus (Figure 4A). Micronucleus (MN) analysis of erythrocytes of C. gariepinus exposed to NPs indicated a significant (P < 0.01) increase in comparison with the control group (Figure 4B, 5A and Table 4). In addition, using Se and C. vulgaris significantly (P< 0.01) reduced the EMN frequency compared with the NPs-exposed group.
Erythrocyte morphological alterations and nuclear abnormalities of RBCs were significantly elevated (P < 0.01) in the NPs group compared with the control group.
Both groups exposed to NPs and treated with Se or Chlorella showed a significant decrease (P< 0.01) in the nuclear abnormalities and erythrocyte alterations compared to those only exposed to NPs.
The Predominant alterations of erythrocytes in NPs exposed group included Micro nucleated cells (Mn); one or more Mn per cell present in most observations, Vacuolated cells (Vc), Acanthocytes (Ac); Crenated cells with fewer surface projections, Eccentric nuclei (Ecn), Spindle-shaped cells (Sp), Crenated cells (echinocytes)(Cr); where the RBCs develop an irregular surface with numerous projections, Sickle cells (Sk), which vary in shape between ellipsoidal, boat-shaped, and genuine sickles; Tear-drop cells (Tr), whose body looks like a tear with pointed apices; bilobed nuclei (Biln); microcytes (Mc); enucleated cells (Enc); and notched nuclei (Non) (Figure 4B-D, 5A-L and Table 4).
3.5. Nephrotoxic effects
Histological examination of transverse kidney sections from control, selenium-treated, and C. vulgaris-treated catfish showed typical histological structures of rounded glomeruli, which are lined by the flat epithelium of Bowman's capsule, and proximal and distal convoluted tubules that are lined by simple cuboidal epithelium. The apical part of the proximal tubule cell is covered by a brush border of microvilli (Figure 6A,G,H). PAS and hematoxylin stains demonstrated increased carbohydrate substances in the brush borders of the epithelium of proximal tubules and basement membranes (Figure 7A,E,F). Transverse kidney sections of NPs-intoxicated catfish showed several pathological alterations, mainly in the renal tubules. They consisted of vacuolar degeneration, apoptosis, and coagulative necrosis of cells lining the renal tubules that later dissociated, causing detachment of the tubular epithelium (Figure 6B). The nuclei of the necrotic tubular epithelium showed necrotic changes in the form of pyknosis, karyorrhexis, or complete loss of nuclei. There was also connective tissue proliferation around the renal tubules (peritubular fibrosis), hemorrhages, and tubular dilatation (Figure 6C). Perivascular connective tissue proliferation around the renal blood vessels and thrombosis were also observed (Figure 6D). PAS and hematoxylin-stained transverse kidney sections showed a decline in the carbohydrate materials in the apical brush borders of proximal tubules, confirming the loss of brush borders (Figure 7B). Picrosirius red-stained transverse sections demonstrated the increment of the connective tissue around the renal tubules and blood vessels (Figure 8B) in comparison with the control, selenium-treated, and C. vulgaris-treated groups that showed a scarcity of connective tissue surrounding the renal tubules and blood vessels (Figure 8A, E, and F).
Administration of selenium in the NPs+Se-treated group ameliorated and inhibited the nephrotoxic effects of nanoplastics. Minimal alterative changes in the tubular epithelium were noticed. The typical histological appearance of the kidney was nearly restored with less vacuolar degeneration and necrotic changes in the tubular epithelium (Figure 6E). Hematoxylin-PAS-and Hematoxylin stained transverse sections showed increased carbohydrate materials in the apical brush borders of proximal tubules, revealing the restoration of their typical structures (Figure 7C). In addition, transverse kidney sections stained with picrosirius red stain showed a decrease in the amount of connective tissue surrounding renal tubules and blood vessels compared to that found in the NPs-intoxicated group (Figure 8C).
Administration of Chlorella in the NPs+Ch treated group restored almost all pathological alterations to near their control levels. Most renal tubules appeared nearly normal, and a few exhibited vacuolar degeneration and tubular necrosis (Figure 6F). PAS and hematoxylin-stained transverse kidney sections showed moderate carbohydrate material localization in the apical brush borders of proximal tubules (Figure 7D). The utilization of picrosirius red stain on transverse kidney sections revealed that the quantity of connective tissue surrounding the renal tubules and blood vessels in the experimental group was lower than that observed in the NPs-intoxicated group. (Figure 8D).
4. Discussion
Nanoplastics are contaminants of growing worldwide concern that could be hazardous to the aquatic ecosystem and human beings [48].
The current results showed a high mortality rate in the NPs exposed group compared to the control group which demonstrated no deaths. One study [49] demonstrated that microplastics (MPs) induced a lower survival rate in fish. The rise in the mortality rate in NPs exposed group may be attributed to intestinal blockage of the gut or tissue abrasions, which may cause lining epithelium injury, morbidity, and mortality [50,51].
The present study revealed that using selenium reduced fish mortality percentage, which is a similar result to [52], who reported that a diet supplemented with Se (0.21– 0.8 mg Se/kg) increased the survival rate of cobia (Rachycentron canadum L) juveniles. Furthermore, this result is similar to the present study as using Chlorella reduced the percentage of mortalities of the fish. Also, this result is in harmony with [53], who mentioned that adding C. vulgaris to the diet of rohu Labeo rohita enhanced the survival rate and phagocytic and antioxidant activity.
The obtained data showed that NPs enhanced oxidative stress by elevating the MDA level in catfish serum. Previous research supports that nanoplastics trigger ROS overproduction and oxidative stress, consistent with the current findings [54,55,56]. Moreover, [57] reported that exposing A. franciscana to PS NPs (50 nm) for 48 h significantly increased lipid peroxidation.
The alkaline comet assay was used in the current study for DNA damage detection in C. gariepinus blood. The results revealed that fish exposed to PS NPs showed a significant incline in DNA damage parameters (tail length, tail DNA %, tail moment, and Olive tail moment). This result is similar to [58], who reported that short-term exposure to different concentrations of 100 nm PS NPs (from 0.05 up to 50 mg/L) induced DNA damage in the hemocytes of the Mediterranean mussel (Mytilus galloprovincialis).
The findings of this study suggest that DNA damage may have an underlying cause: oxidative stress. These data agree with other studies that showed the association between increased MDA, CAT, and GPX levels and jeopardized DNA integrity [50,59,60]. Furthermore, highly reactive radicals are expected to be the principal cause of oxidative DNA damage and chain breaks of the DNA molecule [61].
Based on the actual data, it has been observed that both microplastics (MPs) and nanoparticles (NPs) can induce DNA strand breaks with the extent of the damage being influenced by their respective size and surface charge. Although the exact mechanism by which MPs/NPs induce DNA damage remains incompletely comprehended, several investigations suggest that oxidative stress and physical interaction are pivotal in instigating DNA strand breaks by NPs [62,63]. Additionally, synthetic polymers may have low-molecular-weight fragments of mono- and oligomers, catalysts, artificial stabilizers, and a variety of particular chemical additives such as (phthalates, bisphenol A, stabilizers, pigments, flame retardants, and polychlorinated biphenyls (PCBs)), each of which is substances with genotoxic properties [64].
The findings of previous studies conducted on mussels [50,65,66], and human blood cells [59,67] were in harmony with the current study findings and emphasizing the idea of PS-NPs are contaminants with genotoxic potential.
On the contrary, some studies reported that exposure to PS microparticles did not cause hemocyte genotoxic damage [68,69]. The presence of inconsistent outcomes is expected as the experiments were conducted under diverse circumstances, encompassing distinct plastic particles varieties, configurations and densities, in addition to variations in temperature and duration of the experiment. The present investigation employed the erythrocyte micronucleus (EMN) assay due to the robust correlation between the micronucleus and comet assays for various nanomaterials. [70]. The results revealed that exposure of catfish to PS NPs increased the frequency of micronucleus and several alterations in RBCs. These findings were similar to the study conducted by [71] on Sparus aurata erythrocytes. Guimarães et al. [50] concluded that PS NPs exposure caused erythrocyte morphometric changes in juveniles of Ctenopharyngodon idella.
According to Thomas et al. [72], MN happens when the structure or function of the mitotic apparatus is disrupted, failing to properly include chromosomes or fragments of chromosomes in the central nucleus.
The increase in erythrocyte nuclear abnormalities observed after the toxicity with PS NPs may be related to NPs' interaction with the chromosomal material or the oxidative stress induced by the NPs. Multiple studies have reported that oxidative stress is a consequence of exposure to NPs [17,73]. The erythrocyte vacuoles seen could come from an uneven distribution of hemoglobin, as shown by [74]. Fish need oxygen to breathe, and erythrocyte deformations lower oxygen levels, which disrupt the circulatory system and impair breathing. This respiratory stress may change the number and form of fish erythrocytes [75]. Catfish and D. rerio exposed to microplastics showed these erythrocyte changes, including a considerable increase in the frequency of nuclear abnormalities and morphological RBC changes [76,77].
The current study revealed that selenium and C. vulgaris supplementation had improved all measured parameters compared to the NPs exposed group. Where MDA activity, DNA damage, MN, and erythrocyte abnormalities were decreased. These outcomes are consistent with that obtained by [78,79], who reported that Se reduces MDA activity and increases the antioxidant levels in fish.
Bera et al. [80] explained that the protection of selenium against DNA damage might be because it is an essential constituent of selenoproteins as thioredoxin reductases and glutathione peroxidases. These proteins are crucial in antioxidant defense. Selenium therefore, may protect against DNA damage by elevating DNA glycosylase repair enzyme activity and enhancing pathways of DNA damage repair involving p53 gene.
The natural antioxidant components of C. vulgaris can scavenge free radicals that are responsible for their protective effect [81]. On the other hand, Bengwayan et., al. [82] explained that C. vulgaris has an inhibitory effect on lipid peroxidation. Also, Wu et., al. [83] stated that C. vulgaris possesses various therapeutic characteristics, such as anti-inflammatory, immunomodulatory, and antioxidant activities, which may be extremely important for animal health. They increase SOD and CAT activity, guard against DNA oxidation and lipid peroxidation, and activate cellular antioxidant enzymes.
The results revealed that C. vulgaris reduced micronucleus frequency and other morphological abnormalities in the erythrocyte. These findings were matched with [76], who demonstrated that treating microplastics-intoxicated C. gariepinus with Chlorella reduced genotoxicity and cytotoxicity in the blood.
Pathological changes observed in NPs-intoxicated C. gariepinus kidney sections were comparable to those demonstrated by [84], who studied the nephrotoxic effect of NPs and MPs in mice and observed that intoxicated kidneys showed necrosis of tubular epithelium, associated with loss of tubular brush border and tubulointerstitial fibrosis. Chen et al. [85] found that PS MPs had an oxidative nephrotoxic effect on HEK293 cells induced by inhibiting the antioxidant haem oxygenase-1. Also they observed that MPs induced apoptosis and autophagy via the depolarization of mitochondrial membranes and the formation of autophagosomes. These results support our hypothesis that PS NPs could induce nephrotoxic effects through oxidative-antioxidant homeostasis disturbance.
Administration of selenium in the NPs+Se group ameliorated and reduced the nephrotoxic effects of nanoplastics. Minimal pathological changes in the tubular epithelium were noticed, and the normal histological appearance was moderately restored. This ameliorative role of Se was also confirmed by [86], who mentions that Se has a protective role against oxidative stress, apoptosis, and kidney damage caused by zearalenone (ZEA) in mice. Zhang et al., [86] found that Se restored alterations in the biochemical and antioxidant indicators of ZEA-induced kidney damage. Moreover, Se reduced the expression of proteins and genes associated with endoplasmic reticulum stress and apoptosis caused by ZEA.
Administration of C. vulgaris in the NPs+Ch treated group almost prevents the nephrotoxic effects of nanoplastics and moderately improves the kidney's histology. Consistent with the current results, Latif et al., [87] stated that C. vulgaris reduces the adverse effects of paracetamol intoxication on hematological, biochemical, oxidative stress, and histopathological levels. In addition, they found that C. vulgaris had antioxidant, nephro-, hepato-, and cardioprotective effects against paracetamol-induced toxic effects in Wister rats.
5. Conclusion
The introduction of nano- and microplastic particles into the environment poses a significant threat to sustainable catfish aquaculture. The current research demonstrated that PS NPs have deleterious adverse effects on DNA integrity, erythrocytes, oxidative/antioxidant status, and kidneys of African catfish. Administration of selenium and C. vulgaris in NPs exposed fish ameliorated and decrease alterations and to some degree restored the normal kidney morphology. Therefore, in sustainable catfish aquaculture selenium and C. vulgaris can be used as prophylactic agents to decrease the PS NPs toxicity.
Author Contributions
Conceptualization, Zakaria M. Zaky, Doha Yahia and Alaa El-Din H. Sayed; Methodology, Shimaa A. Abdelbaky, Doha Yahia, Mohamed H. Kotob, Mohammed A. Ali, Mohammed Aufy and Alaa El-Din H. Sayed; Validation, Mohamed H. Kotob; Formal analysis, Shimaa A. Abdelbaky, Mohammed A. Ali and Mohammed Aufy; Investigation, Shimaa A. Abdelbaky and Mohamed H. Kotob; Resources, Alaa El-Din H. Sayed; Data curation, Mohammed A. Ali; Writing – original draft, Shimaa A. Abdelbaky and Mohamed H. Kotob; Writing – review & editing, Zakaria M. Zaky, Doha Yahia, Mohamed H. Kotob, Mohammed A. Ali, Mohammed Aufy and Alaa El-Din H. Sayed; Visualization, Mohammed Aufy; Supervision, Zakaria M. Zaky, Doha Yahia and Alaa El-Din H. Sayed; Project administration, Zakaria M. Zaky. All authors agree to publish the current study.
Funding
There is no fund received by the authors to conduct this experiment.
Institutional Review Board Statement
The research protocol mentioned above has received approval from the veterinary ethical committee of the Faculty of Veterinary Medicine at Assiut University, Egypt, under the reference number 06/2022/0016.
Conflict of Interest
The authors report no declaration of competing interest.
References
- Nag, A.; Baksi, A.; Ghosh, J.; Kumar, V.; Bag, S.; Mondal, B.; Ahuja, T.; Pradeep, T. Tribochemical degradation of polytetrafluoroethylene in water and generation of nanoplastics. ACS Sustain. Chem. Eng. 2019, 7, 17554–17558. [Google Scholar] [CrossRef]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain damage and behavioural disorders in fish induced by plastic nanoparticles delivered through the food chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef]
- Zhu, F.; Zhu, C.; Wang, C.; Gu, C. Occurrence and ecological impacts of microplastics in soil systems: A review. Bull. Environ. Contam. Toxicol. 2019, 102, 741–749. [Google Scholar] [CrossRef]
- Álvarez-Ruiz, R.; Picó, Y. Analysis of emerging and related pollutants in aquaticbiota. Trends Environ. Anal. Chem. 2020, 25, 00082. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A. ; Narayan, R; Law, K. L. Marine pollution. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar]
- Babayemi, J. O.; Nnorom, I. C.; Osibanjo, O.; Weber, R. Ensuring sustainability in plastics use in Africa: consumption, waste generation, and projections. Environ. Sci. Europe 2019, 31, 1–20. [Google Scholar] [CrossRef]
- Song, Y. K.; Hong, S. H.; Jang, M.; Han, G. M.; Jung, S. W.; Shim, W. J. Combined effects of UV exposure duration and mechanical abrasion on microplastic fragmentation by polymer type. Environ. Sci. Technol. 2017, 51, 4368–4376. [Google Scholar] [CrossRef]
- Santillo, D.; Miller, K.; Johnston, P. Microplastics as contaminants in commercially important seafood species. Integr. Environ. Assess. Manag. 2017, 13, 516–521. [Google Scholar] [CrossRef]
- Karami, A.; Golieskardi, A.; Keong Choo, C.; Larat, V.; Galloway, T. S.; Salamatinia, B. The presence of microplastics in commercial salts from different countries. Sci. Rep. 2017, 7, 46173. [Google Scholar] [CrossRef] [PubMed]
- Mason, S. A.; Welch, V. G.; Neratko, J. Synthetic polymer contamination in bottled water. Front. Chem. 2018, 407. [Google Scholar] [CrossRef] [PubMed]
- Jani, P.; Halbert, G. W.; Langridge, J.; Florence, A. T. Nanoparticle uptake by the rat gastrointestinal mucosa: quantitation and particle size dependency. J. Pharm. Pharmacol. 1990, 42, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Walczak, A. P.; Kramer, E.; Hendriksen, P. J.; Tromp, P.; Helsper, J. P.; van der Zande, M.; Rietjens, I.M.C.M.; Bouwmeester, H.; Bouwmeester, H. Translocation of differently sized and charged polystyrene nanoparticles in in vitro intestinal cell models of increasing complexity. Nanotoxicology. 2015, 9, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Prata, J. C.; da Costa, J. P.; Lopes, I.; Duarte, A. C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Sarkar, A.; Yadav, O. P.; Achari, G.; Slobodnik, J. Potential human health risks due to environmental exposure to nano-and microplastics and knowledge gaps: a scoping review. Sci. Total Environ. 2021, 757, 143872. [Google Scholar] [CrossRef]
- Yee, M. S. L.; Hii, L. W.; Looi, C. K.; Lim, W. M.; Wong, S. F.; Kok, Y. Y.; Tan, B.K.; Wong, C.Y.; Leong, C.O.; Leong, C. O. Impact of microplastics and nanoplastics on human health. Nanomaterials 2021, 11, 496. [Google Scholar] [CrossRef]
- Jeong, C. B.; Kang, H. M.; Lee, M. C.; Kim, D. H.; Han, J.; Hwang, D. S.; Souissi, S.; Lee, S. J.; Shin, K.H.; Park, H.G.; Lee, J. S. Adverse effects of microplastics and oxidative stress-induced MAPK/Nrf2 pathway-mediated defense mechanisms in the marine copepod Paracyclopina nana. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Brandts, I.; Cánovas, M.; Tvarijonaviciute, A.; Llorca, M.; Vega, A.; Farré, M.; Teles, M. Nanoplastics are bioaccumulated in fish liver and muscle and cause DNA damage after a chronic exposure. Environ. Res. 2022, 113433. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yin, D.; Jia, Y.; Schiwy, S.; Legradi, J.; Yang, S.; Hollert, H. Enhanced uptake of BPA in the presence of nanoplastics can lead to neurotoxic effects in adult zebrafish. Sci. Total Environ. 2017, 609, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Balbi, T.; Salis, A.; Damonte, G.; Cortese, K.; Caratto, V.; Monopoli, M.P.; Dawson, K.; Bergami, E.; Corsi, I. Interactions of cationic polystyrene nanoparticles with marine bivalve hemocytes in a physiological environment: Role of soluble hemolymph proteins. Environ. Res. 2016, 150, 73–81. [Google Scholar] [CrossRef]
- Han, Y.; Lian, F.; Xiao, Z.; Gu, S.; Cao, X.; Wang, Z.; Xing, B. Potential toxicity of nanoplastics to fish and aquatic invertebrates: Current understanding, mechanistic interpretation, and meta-analysis. J. Hazard Mater. 2021, 127870. [Google Scholar] [CrossRef]
- Pienaar, U. D. V. The freshwater fished of the Kruger National Park. Koedoe 1968, 11, 1–82. [Google Scholar] [CrossRef]
- Skelton, P. A Complete Guide to the Freshwater Fishes of Southern Africa. Halfway House: Southern Book Publishers Ltd. 1993.
- Teugels, G. A systematic revision of the African species of the genus Clarias (Pisces: Clariidae). Annales Musee Royal de l'Afrique Centrale 1986, 247, 1–199. [Google Scholar]
- Nwani, C. D.; Ifo, C. T.; Nwamba, H. O.; Ejere, V. C.; Onyishi, G. C.; Oluah, S. N.; Ikwuagwu, O.E.; Odo, G. E. Oxidative stress and biochemical responses in the tissues of African catfish Clarias gariepinus juvenile following exposure to primextra herbicide. Drug Chem. Toxicol. 2015, 38, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Brar, A.; Kumar, M.; Vivekanand, V.; Pareek, N. Photoautotrophic microorganisms and bioremediation of industrial effluents: current status and future prospects. 3 Biotech. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Safafar, H.; Wagenen, J.V.; Moller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Marine Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.J. Review of selenium toxicity in the aquatic food chain. Sci. Total Environ. 2004, 326, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M. P. The importance of selenium to human health. The lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Mohamed, S. A. K. S.; Upreti, S.; Rajendra, S. V.; Dang, R. Genotoxicity: Mechanisms, testing guidelines and methods. Int. J. Pharm. Pharm. Sci. 2017, 1, 133–138. [Google Scholar]
- Gontijo, A.M.M.C.; Barreto, R.E.; Speit, G.; Reyes, V.A.V.; Volpato, G.L.; Salvadori, D.M.F. Anesthesia of fish with benzocaine does not interfere with comet assay results. Mutat Res. 2003, 534, 165–172. [Google Scholar] [CrossRef]
- Bolognesi, C.; Hayashi, M. Micronucleus assay in aquatic animals. Mutagenesis 2011, 26, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Braham, R. P.; Blazer, V. S.; Shaw, C. H.; Mazik, P. M. Micronuclei and other erythrocyte nuclear abnormalities in fishes from the Great Lakes Basin, USA. Environ. Mol. Mutagen. 2017, 58, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.A.; Trevisan, R.; Massarsky, A.; Kozal, J.S.; Levin, E.D.; Di Giulio, R.T. Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): a case study with nanopolystyrene. Sci. Total Envi ron. 2018, 643, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Durigon, E.G.; Kunz, D.F.; Peixoto, N.C.; Uczay, J.; Lazzari, R. Diet selenium improves the antioxidant defense system of juveniles Nile tilapia (Oreochromis niloticus L.). Braz. J. Biol. 2019, 79, 527–532. [Google Scholar] [CrossRef]
- Bai, S.; Koo, J.; Kim, K.; Kim, S. Effects of Chlorella powder as a feed additive on growth performance in juvenile Korean rockfish, Sebastesschlegeli (Hilgendorf). Aquacult Res. 2001, 32, 92–98. [Google Scholar] [CrossRef]
- Wilson, J.M.; Bunte, R.M.; Carty, A.J. Evaluation of rapid cooling and tricaine methanesulfonate (ms222) as methods of euthanasia in zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 785–789. [Google Scholar]
- Bello, O. S.; Olaifa, F. E.; Emikpe, B. O. Haematological and blood biochemical changes in African catfish, Clarias gariepinus fed walnut (Tetracarpidium conophorum Mull Arg) leaf and onion (Allium cepa Linn) bulb supplemented diets. Am. J. Exp. Agric. 2014, 4, 1593–1603. [Google Scholar] [CrossRef]
- Kei, S. Serum lipid peroxide in cerebrovascular disorders determined by a new colorimetric method. Clinica. Chimica. Acta. 1978, 90, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in enzymology; Academic press, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Abd Ellah, M. R.; Niishimori, K.; Goryo, M.; Okada, K.; Yasuda, J. Glutathion peroxidase and glucose-6-phosphate dehydrogenase activities in bovine blood and liver. J. Vet. Med. Sci. 2004, 66, 1219–1221. [Google Scholar] [CrossRef]
- Villela, I.V.; Oliveira, I.M.; Silva, J.; Henriques, J.A.P. DNA damage and repair in haemolynph cells of golden mussel (Limnoperna fortunei) exposed to environmental contaminants. Mutat. Res. 2006, 605, 78–86. [Google Scholar] [CrossRef]
- Pascoe, S.; Gatehouse, D. The use of a simple haematoxylin and eosin staining procedure to demonstrate micronuclei within rodent bone marrow. Mutat. Res. 1986, 164, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, K.R.; Tilbury, K.L.; Myers, M.S. Assessment of the piscine micronucleus test as an in situ biological indicator of chemical contaminant effects. Can. Aquat. Sci. 1990, 47, 2123–2136. [Google Scholar] [CrossRef]
- Fischer, AH.; Jacobson, KA.; Rose, J.; Zeller, R. Hematoxylin and Eosin Staining of Tissue and Cell Sections. Cold Spring Harbor. Protocol, pdb. Prot. 2008, 49, 86. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.F.A. Histological demonstration of mucin after periodic acid. Nature 1946, 158, 202. [Google Scholar] [CrossRef] [PubMed]
- Bhutda, S.; Surve, M.V.; Anil., A.; Kamath, K.; Singh., N.; Modi, D; Banerjee, A. Histochemical Staining of Collagen and Identification of Its Subtypes by Picrosirius Red Dye in Mouse Reproductive Tissues. Bio-Protocol 2017, 7, 2592. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Naik, T. S. K.; Anil, A. G.; Dhiman, J.; Kumar, V.; Dhanjal, D. S.; Ramamurthy, P. C. Micro (nano) plastics in wastewater: A critical review on toxicity risk assessment, behaviour, environmental impact and challenges. Chemosphere 2022, 290, 133169. [Google Scholar] [CrossRef] [PubMed]
- Jacob, H.; Besson, M.; Oberhaensli, F.; Taylor, A.; Gillet, B.; Hughes, S.; Melvi, S.D.; Bustamante, P.; Swarzenski, P.W.; Lecchini, D.; Metian, M. A multifaceted assessment of the effects of polyethylene microplastics on juvenile gilthead seabreams (Sparus aurata). Aquat. Toxicol. 2021, 241, 106004. [Google Scholar] [CrossRef]
- Guimarães, A.T.B.; Estrela, F.N.; Pereira, P.S.; de Andrade Vieira, J.E.; de Lima Rodrigues, A.S.; Silva, F.G.; Malafaia, G. Toxicity of polystyrene nanoplastics in Ctenopharyngodon idella juveniles: a genotoxic, mutagenic and cytotoxic perspective. Science of The Total Environment 2021, 752, 141937. [Google Scholar] [CrossRef]
- Lu, Y.F.; Zhang, Y.; Deng, Y.F.; Jiang, W.; Zhao, Y.P.; Geng, J.J.; Ding, L.L.; Zhao, Y. Uptake and accumulation of polystyrene microplastics in zebrafish (Danio rerio) and toxic effects in liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Liu, K.; Wang, X.; Ai, Q.; Mai, K.; Zhang, W. Dietary selenium requirement for juvenile cobia, Rachycentron canadum L. Aquac. Res. 2010, 41, e594–e601. [Google Scholar] [CrossRef]
- Andrews, S.R.; Sahu, N.P.; Pal, A.K.; Kumar, S. Haematological modulation and growth of Labeo rohita fingerlings: effect of dietary mannan-oligosaccharide, yeast extract, protein hydrolysate and Chlorella sp. Aquacult. Res. 2009, 41, 61–69. [Google Scholar] [CrossRef]
- Eom, H.J.; Nam, S.E.; Rhee, J.S. Polystyrene Microplastics Induce Mortality through Acute Cell Stress and Inhibition of Cholinergic Activity in a Brine Shrimp. Mol. Cell. Toxicol. 2020, 16, 233–243. [Google Scholar] [CrossRef]
- Kim, J.; Rhee, J.S. Biochemospherical and Physiological Responses of the Water Flea Moina Macrocopa to Microplastics: A Multigenerational Study. Mol. Cell. Toxicol. 2021 17, 523–532.
- Lee, D.H.; Lee, S.; Rhee, J.S. Consistent exposure to microplastics induces age-specific physiological and biochemical changes in a marine mysid. Mar. Pollut. Bull. 2021, 162, 111850. [Google Scholar] [CrossRef]
- Varó, I.; Perini, A.; Torreblanca, A.; Garcia, Y.; Bergami, E.; Vannuccini, M. L.; Corsi, I. Time-dependent effects of polystyrene nanoparticles in brine shrimp Artemia franciscana at physiological, biochemical and molecular levels. Sci. Total Environ. 2019, 675, 570–580. [Google Scholar] [CrossRef]
- Brandts, I.; Teles, M.; Gonçalves, A.P.; Barreto, A.; Franco-Martinez, L.; Tvarijonaviciute, A.; Oliveira, M. Effects of nanoplastics on Mytilus galloprovincialis after individual and combined exposure with carbamazepine. Sci. Total Environ. 2018, 643, 775–784. [Google Scholar] [CrossRef]
- Gopinath, P.M.; Saranya, V.; Vijayakumar, S.; Mythili Meera, M.; Ruprekha, S.; Kunal, R.; Pranay, A.; Thomas, J.; Mukherjee, A.; Chandrasekaran, N. Assessment on interactive prospectives of nanoplastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Sci. Rep. 2019, 9, 1–15. [Google Scholar]
- Shah, N.; Khan, A.; Habib Khan, N.; Khisroon, M. Genotoxic consequences in common grass carp (Ctenopharyngodon idella Valenciennes, 1844) exposed to selected toxic metals. Biol. Trace Elem. Res. 2020, 199, 305–314. [Google Scholar] [CrossRef]
- Ribeiro, F.; Garcia, A. R.; Pereira, B. P.; Fonseca, M.; Mestre, N. C.; Fonseca, T. G.; Bebianno, M. J. Microplastics effects in Scrobicularia plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Palić, D. Micro-and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef] [PubMed]
- Mazur, A. A.; Chelomin, V. P.; Zhuravel, E. V.; Kukla, S. P.; Slobodskova, V. V.; Dovzhenko, N. V. Genotoxicity of polystyrene (PS) microspheres in short-term exposure to gametes of the sand dollar Scaphechinus mirabilis (Agassiz, 1864)(Echinodermata, Echinoidea). J. mar. sci. eng. 2021, 9, 1088. [Google Scholar] [CrossRef]
- Hamlin, H.J.; Marciano, K.; Downs, C.A. Migration of nonylphenol from food-grade plastic is toxic to the coralreef fish species Pseudochromis fridmani. Chemosphere 2015, 139, 223–228. [Google Scholar] [CrossRef]
- Gonçalves, J.M.; Sousa, V.S.; Teixeira, M.R.; Bebianno, M.J. Chronic toxicity of polystyrene nanoparticles in the marine mussel Mytilus galloprovincialis. Chemosphere 2022, 287, 132356. [Google Scholar] [CrossRef]
- Estrela, F.N.; Batista Guimarães, A.T.; Silva, F.G.; Marinho da Luz, T.; Silva, A.M.; Pereira, P.S.; Malafaia, G. Effects of polystyrene nanoplastics on Ctenopharyngodon idella (grass carp) after individual and combined exposure with zinc oxide nanoparticles. J. Hazard Mater. 2021, 403, 123879. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, S.; Domenech, J.; Barguilla, I.; Cortés, C.; Marcos, R.; Hernández, A. Genotoxic and immunomodulatory effects in human white blood cells after ex vivo exposure to polystyrene nanoplastics. Environ. Sci Nano. 2020, 7, 3431–3446. [Google Scholar] [CrossRef]
- Magni, S.; Gagné, F.; André, C.; Della Torre, C.; Auclair, J.; Hanana, H.; Parenti, C.C.; Bonasoro, F.; Binelli, A. Evaluation of uptake and chronic toxicity of virgin polystyrene microbeads in freshwater zebra mussel Dreissena polymorpha (Mollusca: Bivalvia). Sci. Total Environ. 2018, 631–632, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Zhang, J.; Zhao, X.; Feng, J.; Teng, Y.; Chen, B.; Sun, X.; Zhu, L.; Sun, X.; Qu, K. Polystyrene microplastics increase uptake, elimination and cytotoxicity of decabromodiphenyl ether (BDE-209) in the marine scallop Chlamys farreri. Environ. Pollut. 2020, 258, 113657. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.L. The comet assay in nanotoxicology research. Anal Bioanal Chemosphere 2010, 398, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Barría, C.; Martins, M.A.; Franco-Martínez, L.; Barreto, A.; Tvarijonaviciute, A.; Tort, L.; Oliveira, M.; Teles, M. Waterborne exposure of gilthead seabream (Sparus aurata) to polymethylmethacrylate nanoplastics causes effects at cellular and molecular levels. J. Hazard. Mater. 2021, 644, 123590. [Google Scholar] [CrossRef]
- Thomas, P.; Holland, N.; Bolognesi, C.; Kirsch-Volders, M.; Bonassi, S.; Zeiger, E.; Knasmueller, S.; Fenech, M. Buccal micronucleus cytome assay. Nat. Protoc. 2009, 4, 825–837. [Google Scholar] [CrossRef]
- Jacob, H.; Besson, M.; Swarzenski, P.W.; Lecchini, D.; Metian, M. Effects of virgin micro- and nanoplastics on fish: trends, meta-analysis, and perspectives. Environ. Sci. Technol. 2020, 54, 4733–4745. [Google Scholar] [CrossRef]
- Ateeq, B.; Ali, M. N.; Ahmad, W. Induction of micronuclei and erythrocyte alterations in the catfish Clarias batrachus by 2, 4-dichlorophenoxyacetic acid and butachlor. Mutat. Res. 2002, 518, 135–144. [Google Scholar] [CrossRef]
- Brown, L. Aquaculture for veterinarians: fish husbandry and medicine; Pergamon press: Oxford, North Chicago, USA, 1993; 440 pp. [Google Scholar]
- Sayed, A. H.; Hamed, M.; Badrey, A. E.A.; Soliman, H.A.M. Bioreme diation of hemotoxic and oxidative stress induced by polyeth ylene microplastic in Clarias gariepinus using lycopene, citric acid, and Chlorella. Comp. Biochem. Physiol. 2021, Part C 250, 109189. [Google Scholar]
- Da Costa Araujo, A.P.; de Andrade Vieira, J.E; Malafaia, G. Toxicity and trophic transfer of polyethylene microplastics from Poecilia reticulata to Danio rerio. Sci. Total Environ. 2020, 742, 140217. [Google Scholar] [CrossRef]
- Ashouri, S.; Keyvanshokooh, S.; Salati, A. P.; Johari, S. A.; Pasha-Zanoosi, H. Effects of different levels of dietary selenium nanoparticles on growth performance, muscle composition, blood biochemical profiles and antioxidant status of common carp (Cyprinus carpio). Aquaculture 2015, 446, 25–29. [Google Scholar] [CrossRef]
- Saffari, S.; Keyvanshokooh, S.; Zakeri, M.; Johari, S. A.; Pasha-Zanoosi, H. J. A. N. Effects of different dietary selenium sources (sodium selenite, selenomethionine and nanoselenium) on growth performance, muscle composition, blood enzymes and antioxidant status of common carp (Cyprinus carpio). Aquac. Nutr. 2017, 23, 611–617. [Google Scholar] [CrossRef]
- Bera, S.; Rosa, V. D.; Rachidi, W.; Diamond, A. M. Does a role for selenium in DNA damage repair explain apparent controversies in its use in chemoprevention? Mutagenesis 2013, 28, 127–134. [Google Scholar] [CrossRef]
- Abdelhamid, F. M.; Elshopakey, G. E.; Aziza, A. E. Ameliorative effects of dietary Chlorella vulgaris and β-glucan against diazinon-induced toxicity in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 96, 213–222. [Google Scholar] [CrossRef]
- Bengwayan, P.T.; Laygo, J.C.; Pacio, A.E.; Poyaoan, J.L.Z.; Rebugio, J.F.; Yuson, A.L.L. A comparative study on the antioxidant property of Chlorella (Chlorella sp) tablet and glutathione tablet. Int. J. Sci. Res. 2010, 2, 25–35. [Google Scholar]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: an overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, J.; Wang, W.; Gonzalez-Gil, G.; Vrouwenvelder, J. S.; Li, Z. Effects of nano-and microplastics on kidney: Physicochemospherical properties, bioaccumulation, oxidative stress and immunoreaction. Chemosphere 2022, 288, 132631. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Chen, K.F.; Lin, K.Y.A.; Chen, J.K.; Jiang, X.Y.; Lin, C.H. The nephrotoxic potential of polystyrene microplastics at realistic environmental concentrations. J. Hazard. Mat. 2022, 427, 127871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, B.; Wang, M.; Tong, J.; Pan, J.; Wang, N.; Gong, P.; Long, M. Selenium protects against zearalenone-induced oxidative stress and apoptosis in the mouse kidney by inhibiting endoplasmic reticulum stress. Oxidative Med. Cell. Longev 2020. [Google Scholar] [CrossRef]
- Latif, A. A.E.; Assar, D.H.; Elkaw, E.M.; Hamza, H.A.; Alkhalifah, D.H.M.; Hozzein, W.N.; Hamouda, R.A. Protective role of Chlorella vulgaris with Thiamine against Paracetamol induced toxic effects on haematological, biochemical, oxidative stress parameters and histopathological changes in Wistar rats. Sci. Rep. 2021, 11, 3911. [Google Scholar] [CrossRef]
Figure 1.
Transmission electron photomicrographs of the PS NPs.
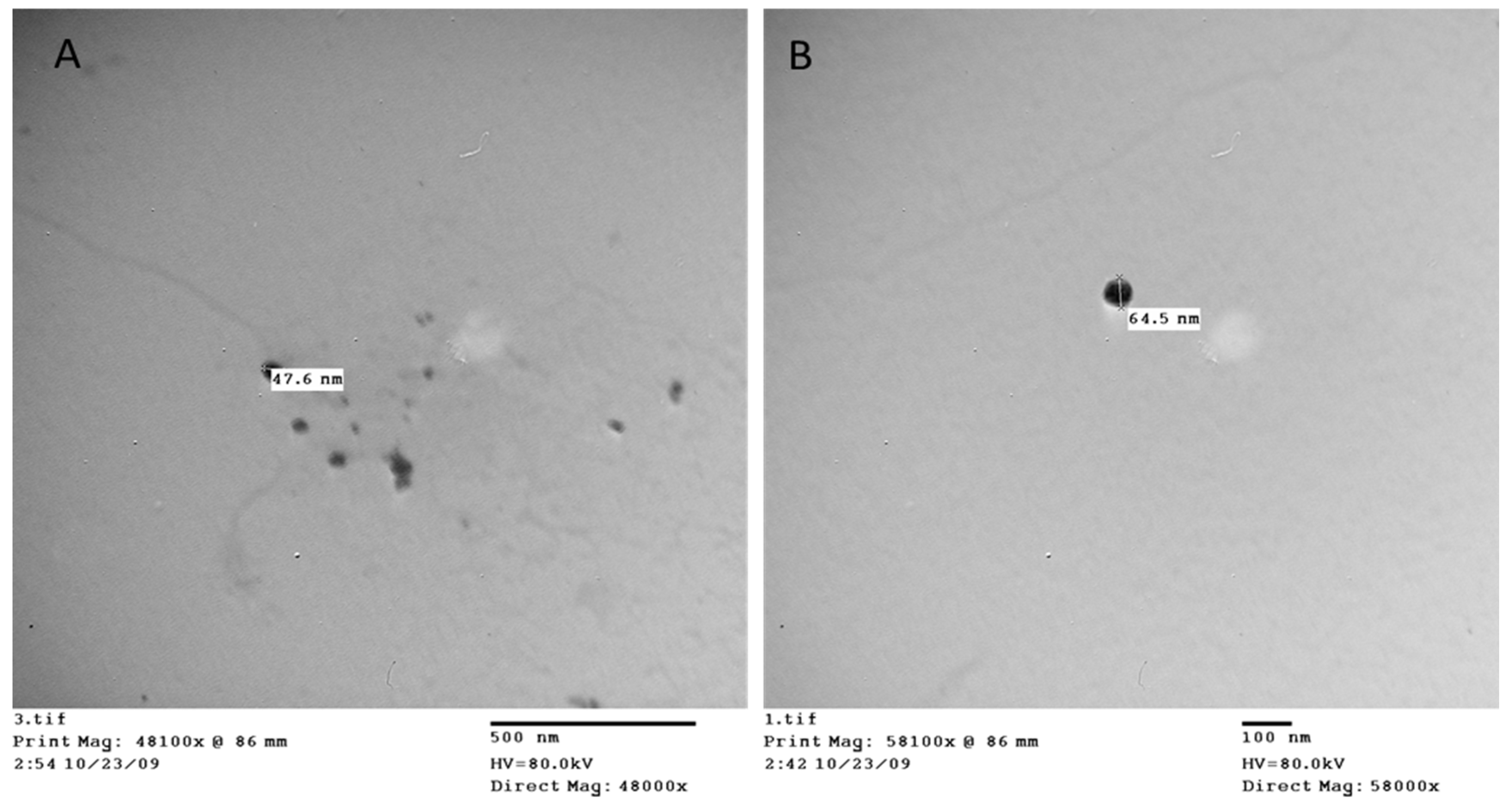
Figure 2.
DNA damage in whole blood of C. gariepinus in all the experimental groups. A: Control group; B: NPs group; C: (NPs + Se) group; D: (NPs + Ch); E: Ch; F: Se.
Figure 2.
DNA damage in whole blood of C. gariepinus in all the experimental groups. A: Control group; B: NPs group; C: (NPs + Se) group; D: (NPs + Ch); E: Ch; F: Se.
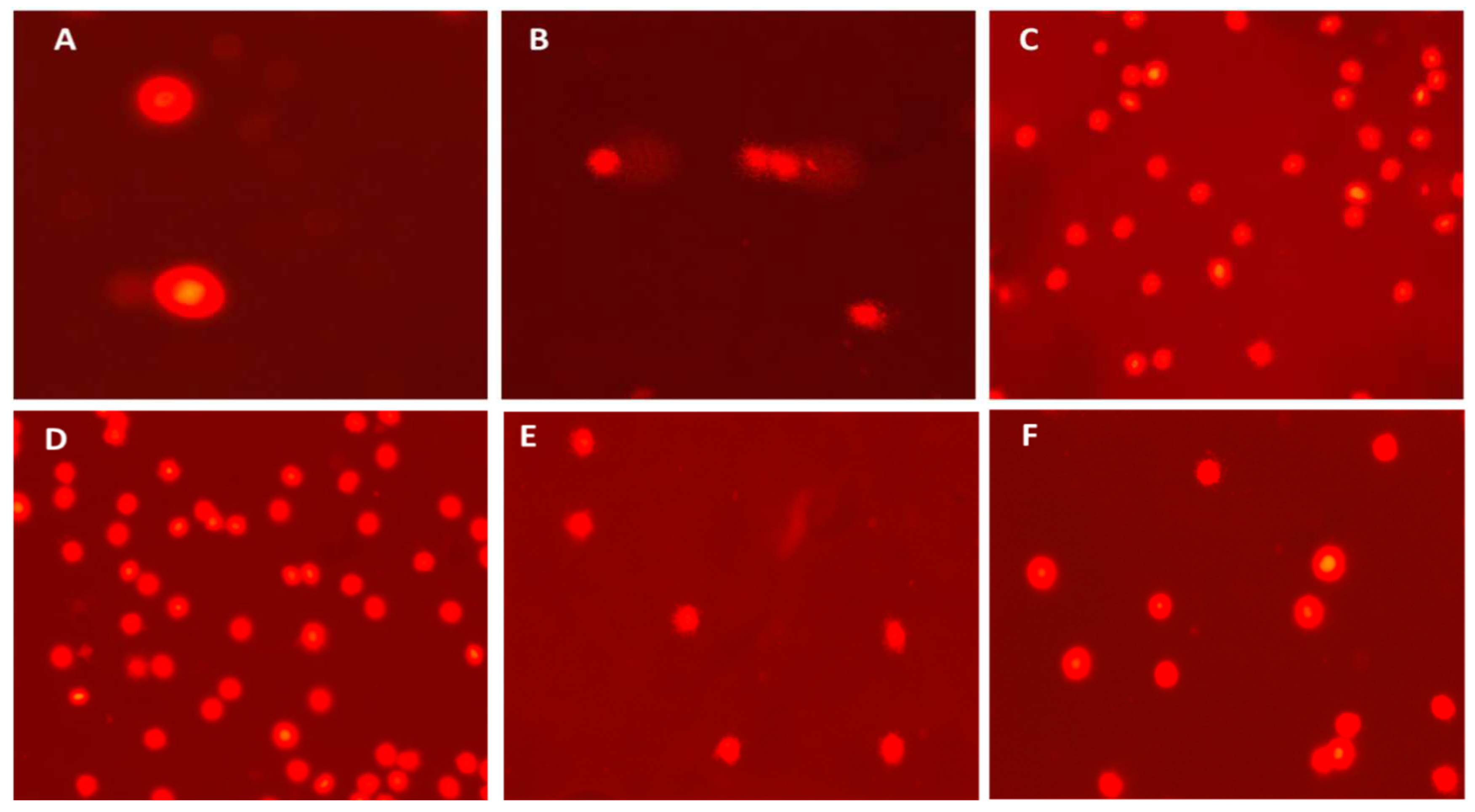
Figure 3.
Assessment of different degrees of DNA damage induced by PS NPs using CASP software showed different degrees of DNA migration. A: intact nucleus (no DNA damage). B: mild DNA damage. C: moderate degree of DNA damage. D: severe DNA damage. E&F: extensive DNA damage.
Figure 3.
Assessment of different degrees of DNA damage induced by PS NPs using CASP software showed different degrees of DNA migration. A: intact nucleus (no DNA damage). B: mild DNA damage. C: moderate degree of DNA damage. D: severe DNA damage. E&F: extensive DNA damage.
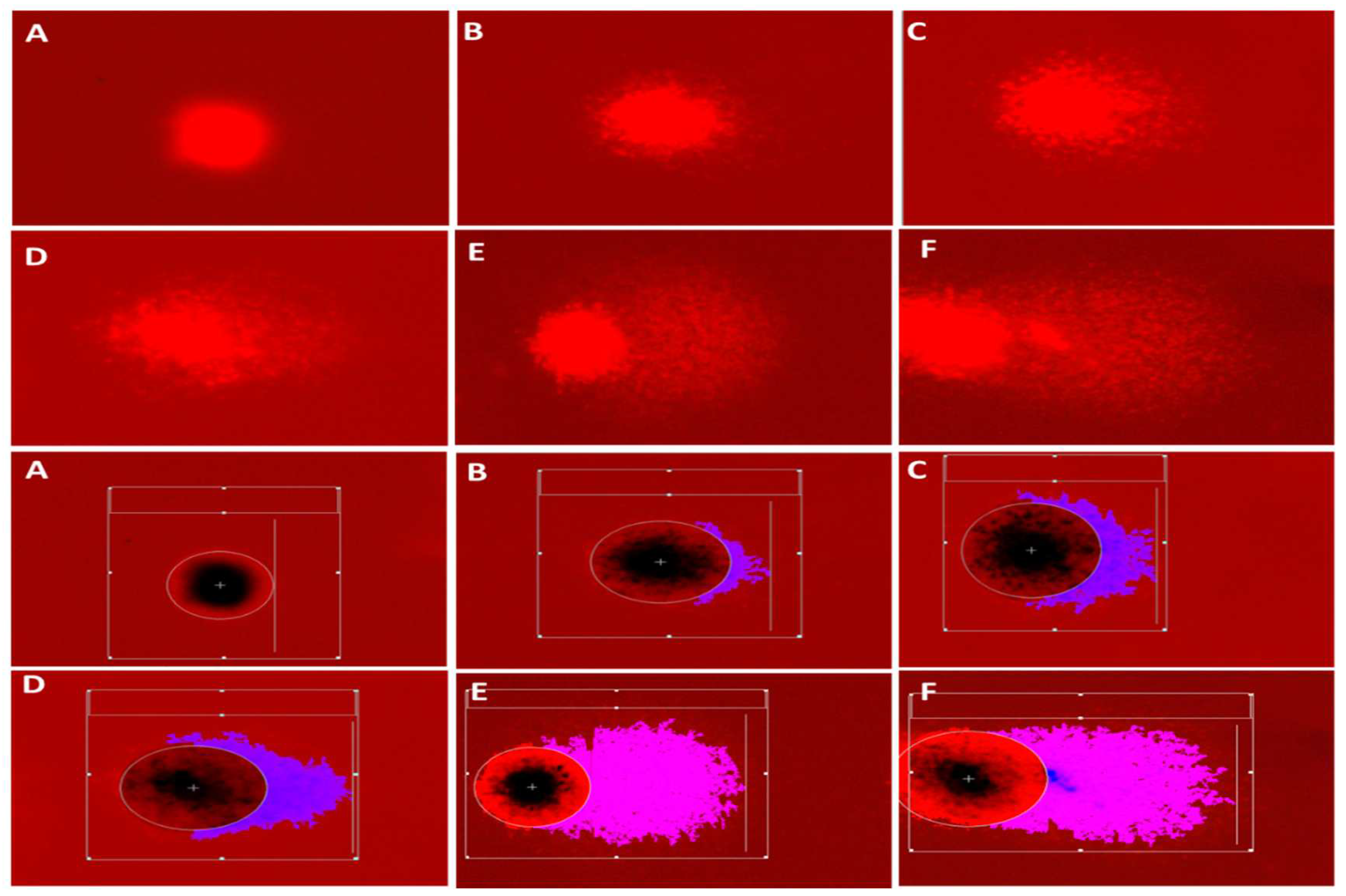
Figure 4.
Blood smears stained by H&E showed the morphological alterations and nuclear abnormalities in erythrocytes of C. gariepiuns exposed to PS NPs. (a) control fish showing normal red blood cells (RBCs), (B–D) fish exposed to PS NPs showing Mn; micronucleated cell, Vc: vacuolated cell, Bin; Bilobed nucleus cell, Ac: acanthocyte, Ecn; eccentric nucleus, Sp; spindle-shaped cell, Cr; crenated cell, Sk; sickled cell, Tr; tear-drop cell, En; enucluated cell, Mc; microcyte, Non; notched nucleated cell. Scale bar = 20 μm.
Figure 4.
Blood smears stained by H&E showed the morphological alterations and nuclear abnormalities in erythrocytes of C. gariepiuns exposed to PS NPs. (a) control fish showing normal red blood cells (RBCs), (B–D) fish exposed to PS NPs showing Mn; micronucleated cell, Vc: vacuolated cell, Bin; Bilobed nucleus cell, Ac: acanthocyte, Ecn; eccentric nucleus, Sp; spindle-shaped cell, Cr; crenated cell, Sk; sickled cell, Tr; tear-drop cell, En; enucluated cell, Mc; microcyte, Non; notched nucleated cell. Scale bar = 20 μm.

Figure 5.
Showing different erythrocyte alterations were recorded in PS NPs exposed group, A; Micronucleated cell, B; Vacuolated cell, C; Spindle cell, D; Sickle cell, E; Acanthocyte, F; crenated cell, G; notched nucleated cell, H; Bilobed nucleus, I; Microcell, J; Tear-drop like cell, K; Enucleated cell, L; Eccentric nucleus.
Figure 5.
Showing different erythrocyte alterations were recorded in PS NPs exposed group, A; Micronucleated cell, B; Vacuolated cell, C; Spindle cell, D; Sickle cell, E; Acanthocyte, F; crenated cell, G; notched nucleated cell, H; Bilobed nucleus, I; Microcell, J; Tear-drop like cell, K; Enucleated cell, L; Eccentric nucleus.
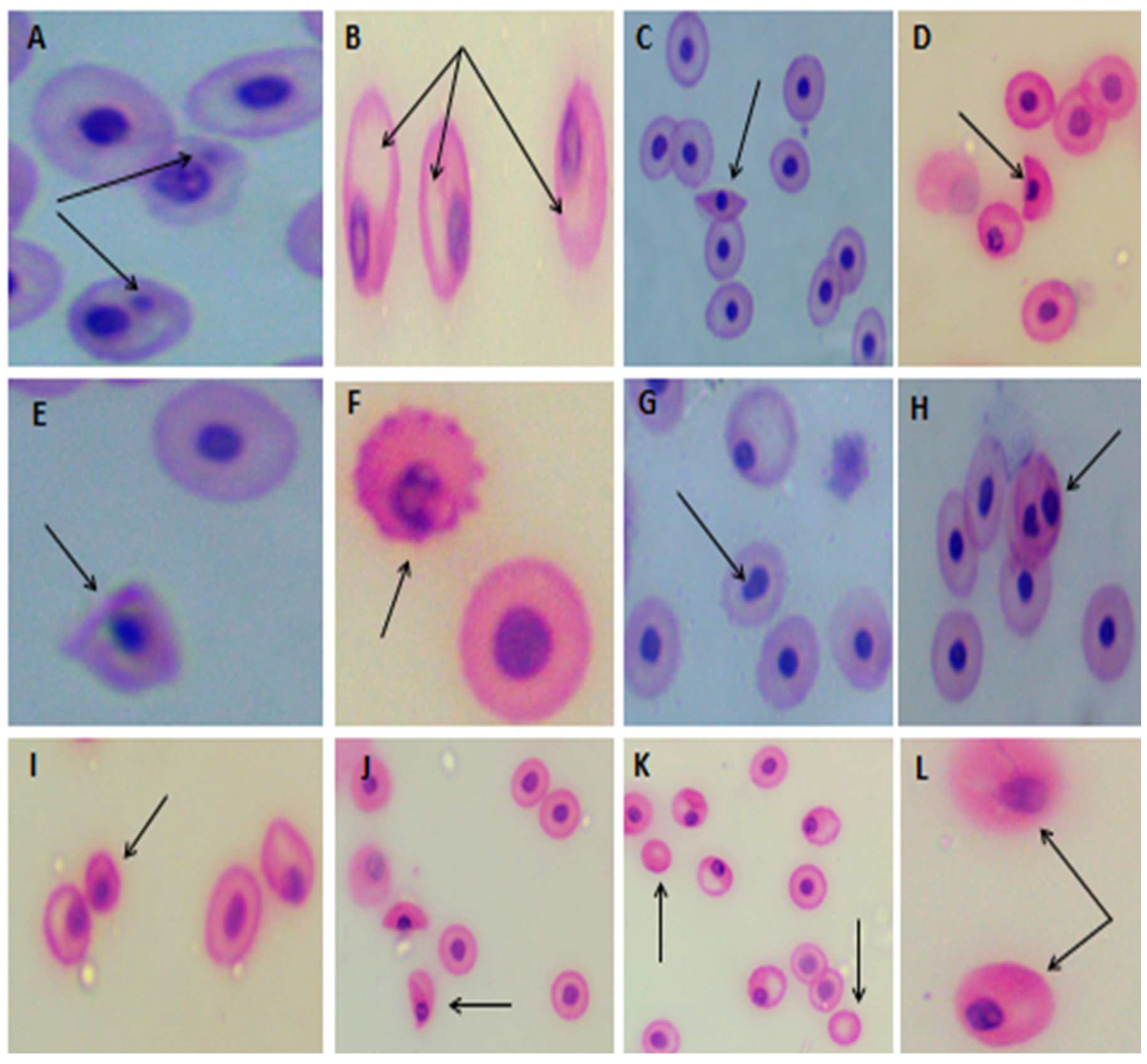
Figure 6.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus:A, G, H. Kidney sections of control, selenium and C. vulgaris treated catfish showed normal histological structures of the kidney that consisted of normally rounded glomeruli (green arrow) and renal tubules that covered by simple columnar epithelium with apical brush border (black arrow). B, C, D. Kidney transverse sections of NPs-intoxicated catfish in B showed the presence of vacuolar degeneration in the cytoplasm of tubular epithelium with peripherally located pyknotic nuclei (black arrow) and epithelial necrotic changes with nuclear alterations, dissociation of renal cells and epithelium desquamation (yellow arrow), in C showed connective tissue proliferation around the renal tubules (peritubular fibrosis) (black arrow) and necrosis and apoptosis of tubular epithelium (yellow arrow), in D showed perivascular connective tissue proliferation (black arrow), thrombosis (star) and vacuolar degeneration (yellow arrow). E. Kidney transverse section of NPs+Se treated catfish showed minimal vacuolar degenerative changes, necrotic changes of tubular epithelium, their nuclei appeared round, and vesicular centrally located (arrow). F. Kidney transverse section of NPs+Ch treated catfish showed vacuolar degenerative changes, necrotic changes of tubular epithelium and desquamation (yellow arrow), some renal tubules appeared normal with minimal cytoplasmic and nuclear alterations (black arrow). H& E. Scale bar = 20 μm.
Figure 6.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus:A, G, H. Kidney sections of control, selenium and C. vulgaris treated catfish showed normal histological structures of the kidney that consisted of normally rounded glomeruli (green arrow) and renal tubules that covered by simple columnar epithelium with apical brush border (black arrow). B, C, D. Kidney transverse sections of NPs-intoxicated catfish in B showed the presence of vacuolar degeneration in the cytoplasm of tubular epithelium with peripherally located pyknotic nuclei (black arrow) and epithelial necrotic changes with nuclear alterations, dissociation of renal cells and epithelium desquamation (yellow arrow), in C showed connective tissue proliferation around the renal tubules (peritubular fibrosis) (black arrow) and necrosis and apoptosis of tubular epithelium (yellow arrow), in D showed perivascular connective tissue proliferation (black arrow), thrombosis (star) and vacuolar degeneration (yellow arrow). E. Kidney transverse section of NPs+Se treated catfish showed minimal vacuolar degenerative changes, necrotic changes of tubular epithelium, their nuclei appeared round, and vesicular centrally located (arrow). F. Kidney transverse section of NPs+Ch treated catfish showed vacuolar degenerative changes, necrotic changes of tubular epithelium and desquamation (yellow arrow), some renal tubules appeared normal with minimal cytoplasmic and nuclear alterations (black arrow). H& E. Scale bar = 20 μm.
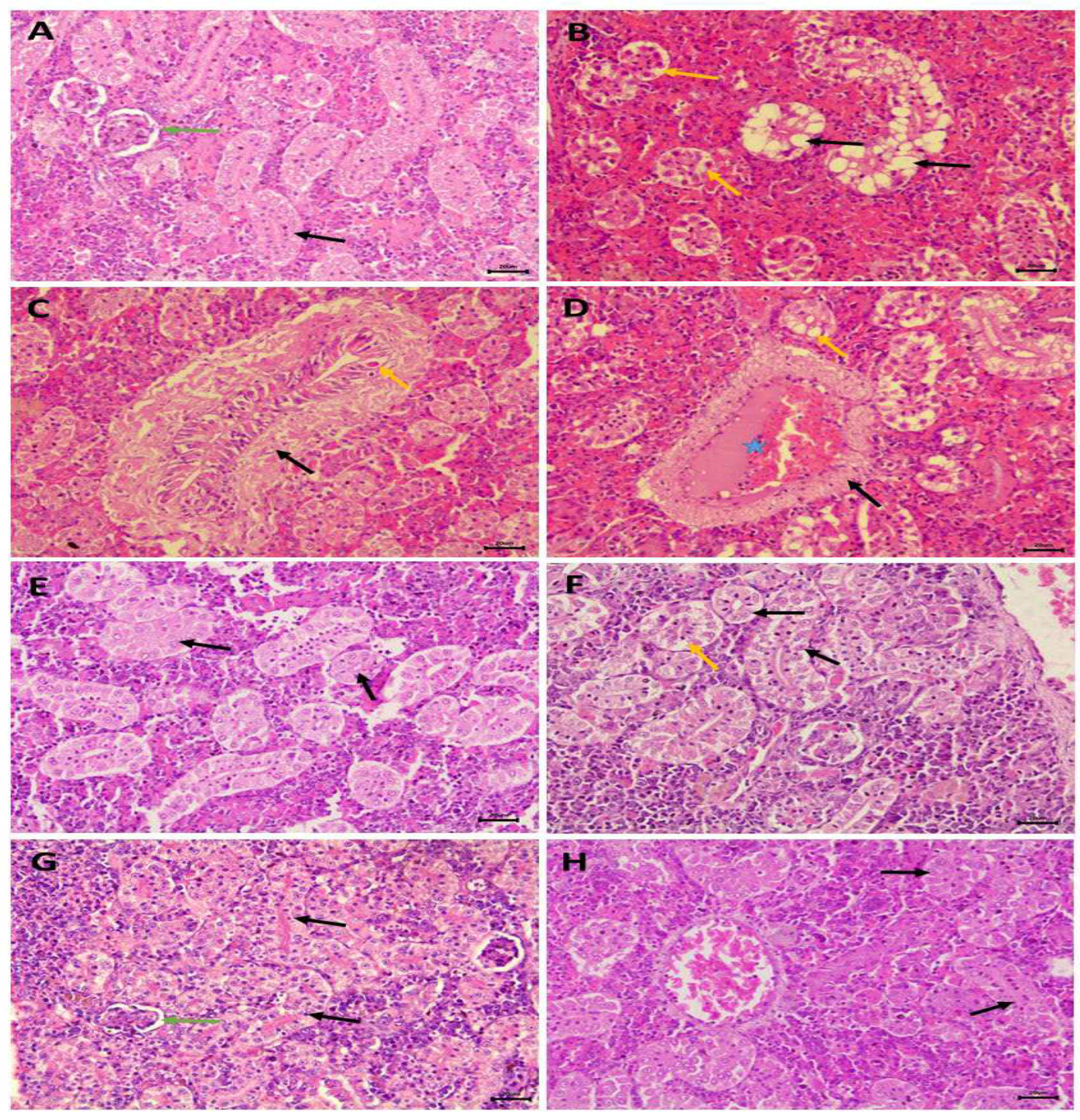
Figure 7.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus: A, E, F. Kidney transverse sections of control, selenium and C. vulgaris treated catfish showed positive PAS reactivity in the brush border (BB) and basement membrane (BM) of the renal tubules. B. Kidney transverse sections of NPs-intoxicated catfish showed a decrease in carbohydrate materials in the brush borders and the basement membranes of the renal tubules. C. Kidney transverse section of NPs+Se treated catfish showed an increase in the amount of carbohydrate observed in the basement membranes and brush borders. D. Kidney transverse section of NPs+Ch treated catfish showed moderate carbohydrate materials localization in the brush border and basement membrane of renal tissue. PAS+Hematoxylin. Scale bar = 20μm.
Figure 7.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus: A, E, F. Kidney transverse sections of control, selenium and C. vulgaris treated catfish showed positive PAS reactivity in the brush border (BB) and basement membrane (BM) of the renal tubules. B. Kidney transverse sections of NPs-intoxicated catfish showed a decrease in carbohydrate materials in the brush borders and the basement membranes of the renal tubules. C. Kidney transverse section of NPs+Se treated catfish showed an increase in the amount of carbohydrate observed in the basement membranes and brush borders. D. Kidney transverse section of NPs+Ch treated catfish showed moderate carbohydrate materials localization in the brush border and basement membrane of renal tissue. PAS+Hematoxylin. Scale bar = 20μm.
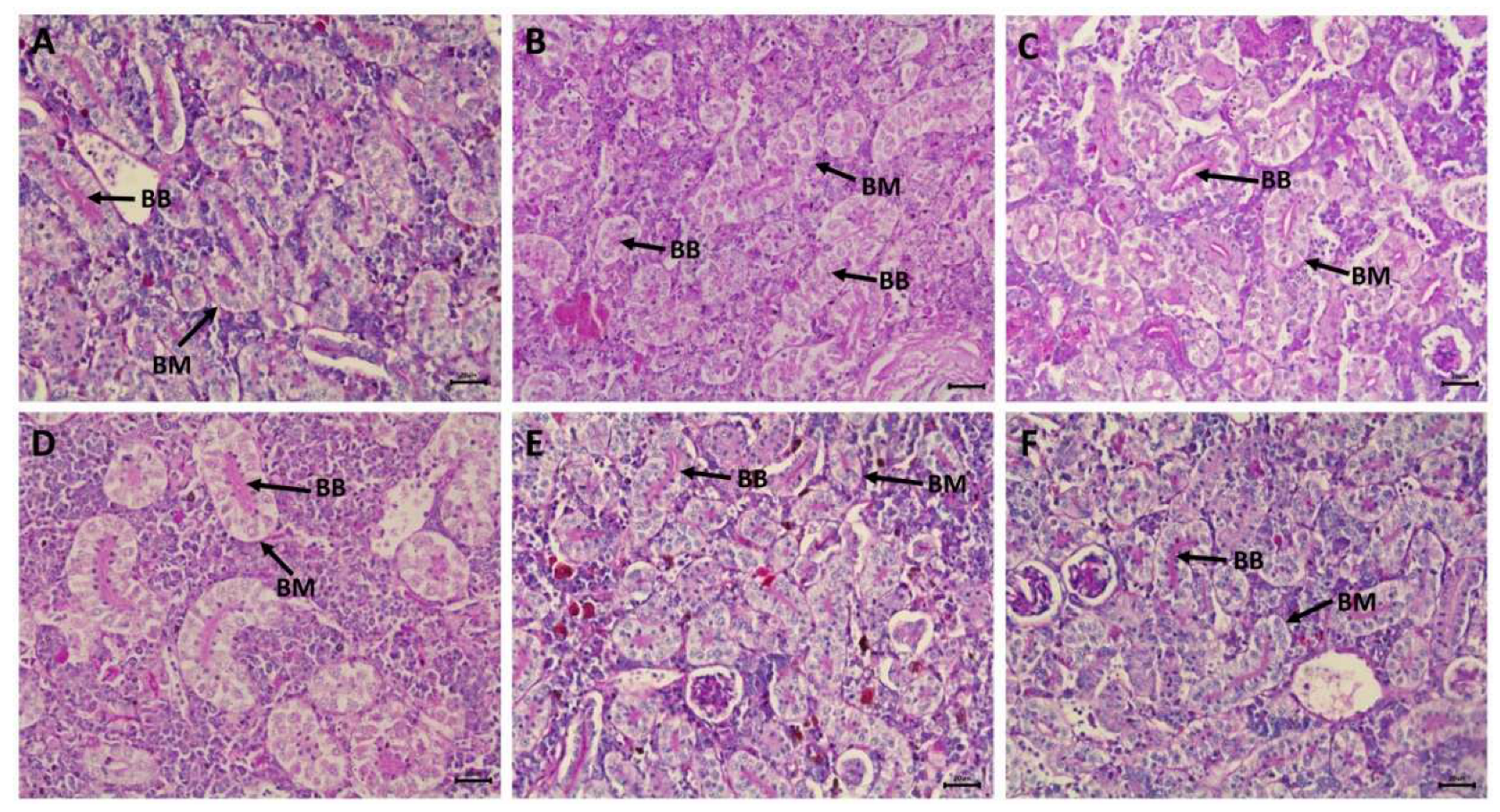
Figure 8.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus: A, E, F. Kidney transverse sections of control, selenium and C. vulgaris treated catfish showed a scanty amount of connective tissue fibers around the renal tubules and blood vessels. B. Kidney transverse sections of NPs-intoxicated catfish showed an increased amount of connective tissue fibers around the renal tubules and blood vessels with a thickened wall of blood vessels. C, D: Kidney transverse section of NPs+Se and NPs+Ch treated catfish respectively showing a decreased amount of connective tissue fibers around the renal tubules and blood vessels. Renal tubules (black arrow) and blood vessels (blue arrow). Picrosirius red stain. Scale bar = 20 μm.
Figure 8.
Photomicrographs of kidney transverse sections from control, NPs-intoxicated and treated C. gariepinus: A, E, F. Kidney transverse sections of control, selenium and C. vulgaris treated catfish showed a scanty amount of connective tissue fibers around the renal tubules and blood vessels. B. Kidney transverse sections of NPs-intoxicated catfish showed an increased amount of connective tissue fibers around the renal tubules and blood vessels with a thickened wall of blood vessels. C, D: Kidney transverse section of NPs+Se and NPs+Ch treated catfish respectively showing a decreased amount of connective tissue fibers around the renal tubules and blood vessels. Renal tubules (black arrow) and blood vessels (blue arrow). Picrosirius red stain. Scale bar = 20 μm.
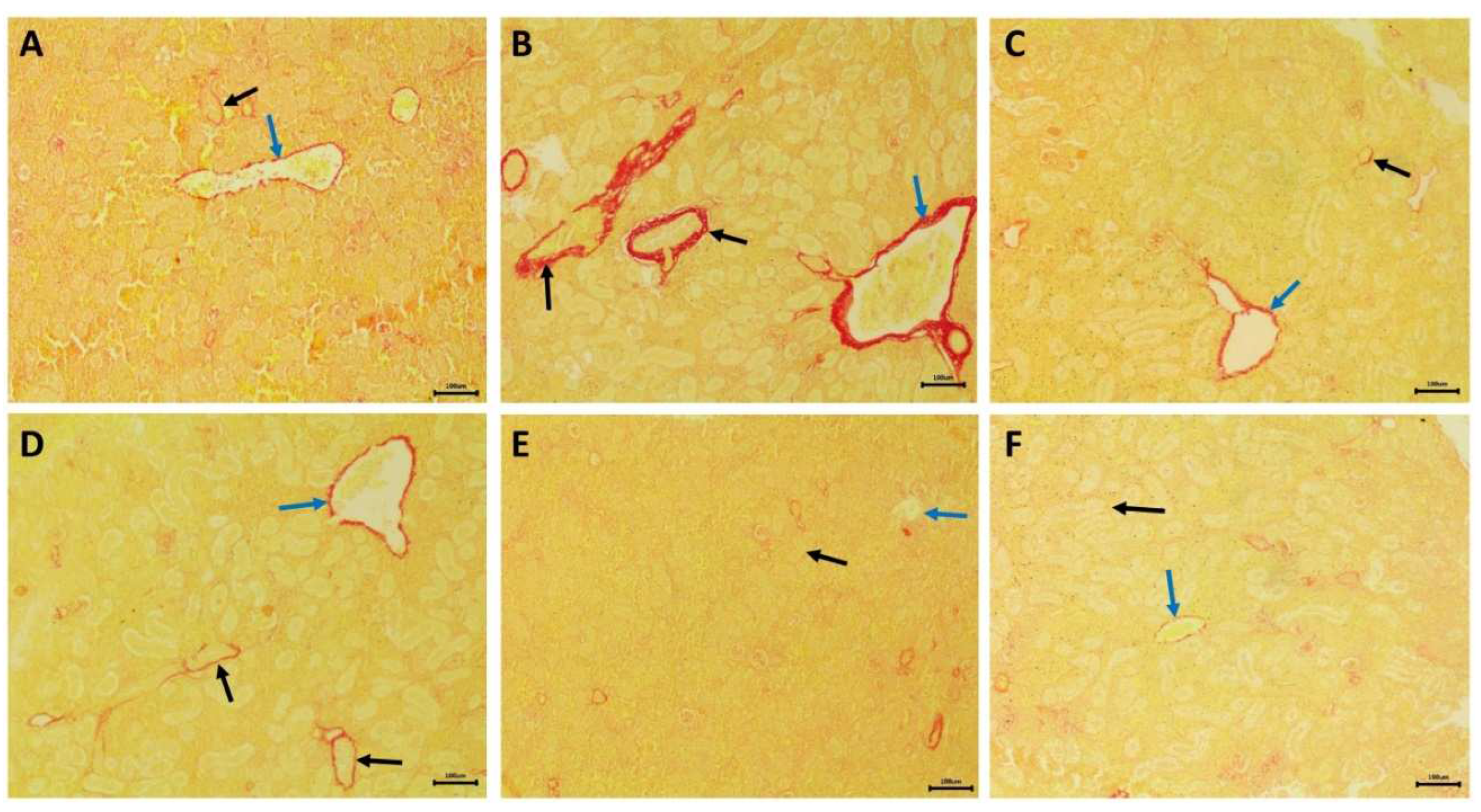

Table 1.
Ingredients and chemical composition of the basal diet (dry weight, %).
| Ingredients | % |
|---|---|
| Fish meal (65%) | 9 |
| Soybean meal (46%) | 36.85 |
| Corn gluten (60%) | 12.2 |
| Yellow corn | 19.25 |
| Wheat bran | 5.7 |
| Fish oil | 6.00 |
| Starch | 7.00 |
| Mineral premix (without se) | 2.00 |
| Vitamin premix | 2.00 |
| Total | 100 |
|
Proximate composition % dry matter (DM) |
% |
| Crude protein (CP) | 30 |
| Crude fiber (CF) | 4.8 |
| Ash | 8.2 |
| Ether extract (EE) | 6.5 |
| *Nitrogen free extract (NFE) | 50.5 |
*NFE = 100- (CP% + EE% + CF% + Ash%).
Table 2.
MDA (nmol/mL), catalase (U/L) and glutathione (U/L) activities in serum of C. gariepinus in control, NPs, (NPs+Se), (NPs+Ch), Ch and Se groups.
Table 2.
MDA (nmol/mL), catalase (U/L) and glutathione (U/L) activities in serum of C. gariepinus in control, NPs, (NPs+Se), (NPs+Ch), Ch and Se groups.
| Se | Ch | NPs+Ch | NPs+Se | NPs | Control | Groups |
|---|---|---|---|---|---|---|
| Parameters | ||||||
| 4.02±0.9b | 3.72±0.8b | 4.98± 0.7b | 5.56±0.5b | 10.22± 2.4 a | 3.71± 0.7b | MDA |
| 352.9±33.4a | 232.93±11.2b | 202.43±40.5b | 235±62.3b | 201.43±5.2b | 225.9±13.6b | CAT |
| 32.43±8.0a | 24.33±2.99a | 25.57±4.2a | 33.7± 10.4a | 19.1± 6.4a | 27.5± 5.6a | GPX |
Data are presented as means ± SE. Different letters a, b and c indicate significant changes among groups. P < 0.05. N= 6.
Table 3.
DNA damage in the blood of C. gariepinus in control, NPs, (NPs+Se), (NPs+Ch), Ch and Se groups.
Table 3.
DNA damage in the blood of C. gariepinus in control, NPs, (NPs+Se), (NPs+Ch), Ch and Se groups.
| Parameters | Control | NPs | NP+ Se | NP+ Ch | Ch | Se |
|---|---|---|---|---|---|---|
| Tail DNA% | 0.57±0.12bc | 3.43 ±0.53a | 1.26±0.19b | 0.98±0.17b | 0.56±0.16b c | 0.08±0.03c |
| Tail Length(µm) | 8.24 ±0.62c | 32.14 ±2.60a | 15.42±1.21 b | 9.77±0.60c | 7.50 ±0.45cd | 4.22 ±0.28 d |
| Tail moment | 0.19±0.05b | 3.60±0.88a | 0.44±0.07b | 0.20±0.06b | 0.15±0.03b | 0.09±0.01b |
| Olive tail moment | 0.46±0.09b | 4.58±0.66a | 1.15±0.15b | 0.59±0.11b | 0.30±0.05b | 0.11±0.02b |
Data are presented as mean ± SE. Values followed by different letters indicate significant changes among groups. P < 0.05. N= 6.
Table 4.
Erythrocytes morphological alterations and nuclear abnormalities in C.gariepinus in control, NPs, NPs+Se, NPs+Ch, Ch and Se groups.
Table 4.
Erythrocytes morphological alterations and nuclear abnormalities in C.gariepinus in control, NPs, NPs+Se, NPs+Ch, Ch and Se groups.
| Se | Ch | NPs+Ch | NPs+Se | NPs | Control | Groups |
|---|---|---|---|---|---|---|
| Parameters | ||||||
| 3.0±0.57b | 2.3±0.33b | 3.3±1.2b | 4.3±1.20b | 22.3±2.00a | 2.3±0.88b | Mn |
| 5.3±0.88b | 6.0±1.52b | 8.3±2.6b | 9.3±1.45b | 47.3±5.78a | 7.0 ±0.57b | Vc |
| 4.3±1.20b | 5.0±1.15b | 6±0.57b | 7.6±0.88b | 14.3±1.45a | 5.0±1.50b | Tc |
| 3.3±0.88b | 3.0±1.00b | 4.3±0.88b | 5.0±1.15b | 11.3±1.45a | 4.3±1.20b | Ac |
| 4.0±1.5b | 4.3±1.20b | 5.3±0.88b | 5.6±1.20b | 12.6±1.76a | 3.6±0.88b | Cr |
| 2.3±0.88b | 2.0±0.57b | 3.6±1.20b | 4.6±0.80b | 13.3±0.88a | 2.0±0.57b | Sk |
| 2.0±0.57b | 1.6±0.66b | 2.6±1.20b | 2.3±0.80b | 7.3±1.45a | 1.6±0.33b | Sp |
| 3.6±0.88b | 2.6±1.20b | 5.6±1.70b | 5.3±1.20b | 19.0±1.15a | 4.6±1.20b | Mc |
| 4.6±0.88d | 6.3±1.2bc | 7.0±2.00bc | 12.0±2.50b | 50.0 ±3.20a | 7.6±1.76bc | Ecn |
| 1.3±0.33b | 1.3±0.33b | 4.0±0.57b | 4.3±1.40b | 10±1.50a | 3.0±0.57b | Bin |
| 1.6±0.33bc | 1.0±0.33d | 2.3±0.88bc | 3.6±0.80b | 8.3±1.20a | 1.3±0.33bc | Notn |
| 2.0±0.57b | 1.3±0.33b | 2.0±0.57b | 3.0±0.57b | 9.0±1.15a | 2.6±0.88b | Enn |
Data are presented as mean ± SE. Values followed by different letters indicate significant changes among groups. P < 0.05. Mn; micronucleated cell, Vc; vacuolated cell, Tr; tear-drop cell, Ac; acanthocyte, Cr; crenated cell, Sk; sickled cell, Sp; spindle-shaped cell, Mc; microcyte, Ecn; eccentric nucleus, Bin; Bilobed nucleus, Non; notched nucleated cell, En; enucluated cell. N= 6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



