
Submitted:
11 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
The intricate functionality of the vertebrate retina relies on the interplay between neurotransmitter activity and calcium dynamics, offering important insights into de-velopmental processes, physiological functioning, and disease progression. Neuro-transmitters orchestrate cellular processes to shape the behavior of the retina under diverse circumstances. Despite research to elucidate the roles of individual neuro-transmitters in the visual system, there remains a gap in our understanding of the holistic integration of their interplay with calcium dynamics in the broader context of neuronal development, health, and disease. To address this gap, the present review explores the mechanisms used by the neurotransmitters glutamate, GABA, glycine, dopamine, and acetylcholine and their interplay with calcium dynamics. This con-ceptual outline is intended to inform and guide future research, underpinning novel therapeutic avenues for retinal-associated disorders.
Keywords:
Retina
; neurotransmitters
; calcium
; neural processes
; vision
; retinal development
; synaptic transmission
; retinal diseases
; ocular disorders
; therapeutic advancements.
1. Introduction
An integral component of the central nervous system (CNS), the retina provides crucial insights into brain development, function, and disease progression. Spanning an approximate thickness of 250 µm in humans, 170 µm in rats, 165 µm in rabbits, and 160 µm in zebrafish, the retina is composed of rod and cone photoreceptors, horizontal cells, bipolar cells (BPCs), amacrine cells, glial cells, microglia, astrocytes, Müller cells, and ganglion cells [1,2,3,4,5]. Within this compact, elegant structure, these components perform visual functions, such as light reception, phototransduction, and signal processing [1]. The cellular components communicate with one another via multiple neurotransmitters, each of which is governed by calcium (Ca2+) dynamics in a distinct way to ensure proper retinal development and healthy physiological functioning. Thus, comprehension of retinal structure, its components, and its neuronal mechanisms is crucial for the advancement of research in the field.
Neurotransmission requires interaction between Ca2+, various neurotransmitters including glutamate, GABA, glycine, dopamine, and acetylcholine (ACh), and their respective receptors that enable precise and rapid communication between retinal cells [6,7]. The heteromeric, multi-subunit neurotransmitter receptors facilitate increased diversity and specificity of their synapses [8]. In the retina, optimal neurotransmission relies on the precise interplay between neurotransmitters and Ca2+ dynamics, since Ca2+ ions act as second messengers that influence numerous cellular functions and facilitate neurotransmitter release, which is essential for maintaining visual sensitivity, perception, and signal processing [9,10,11,12]. Any disruptions of this interplay between neurotransmitter release and Ca2+ dynamics can lead to visual impairments and eye-related disorders, as discussed in detail for each neurotransmitter [13,14,15,16,17,18,19,20,21].
Due to its accessibility and direct connection to the central nervous system, the study of the retina in different animal models can offer insight into the detection of neurodegenerative diseases, including Huntington’s disease (HD), Parkinson’s disease (PD), and Alzheimer’s disease (AD), and ocular disorders, including glaucoma, age-related macular degeneration, and diabetic retinopathy in humans [22,23,24,25]. For each of these, the pathogenicity involves changes in neurotransmission and Ca2+ dynamics within the retina, highlighting this unique structure as the center point of a crucial area of research for potential therapeutic intervention [13,14,15,16,17,18,19,20,21]. The current review will explore important aspects of the retina, including its structure, synapses, visual processes, and development, and describe the roles of the retinal neurotransmitters glutamate, GABA, glycine, dopamine, and acetylcholine and their interplay with Ca2+ ions in retinal development, normal physiological functioning, and disease.
1.1. The retina
1.1. Retinal structure and information processing
The complex, well-defined circuitry of the retina relies on the arrangement of key components across various cellular layers that facilitate vision in various organisms. Therefore, an understanding of retinal structure is fundamental to understanding its function and mechanisms. The general structure of the human retina is shown schematically in Figure 1. Starting from the innermost layer that is closer to the anterior part of the face and the vitreous humor that fills the eye, the retina is comprised of the inner limiting membrane (ILM), retinal nerve fiber layer (RNFL), retinal ganglion cells (RGCs) layer, inner plexiform layer (IPL), inner nuclear layer (INL), outer plexiform layer (OPL), outer nuclear layer (ONL), the ellipsoid zone (EZ), the photoreceptor outer segments (POS), and the retinal pigment epithelium (RPE) [26]. As described by Masland et al. (2012), each layer is comprised of different cell types with unique functions relevant to different aspects of vision [1]. The ILM, composed of astrocytes and the end feet of Müller cells, creates a barrier between the vitreous humor and the retina itself. The RNFL is formed by the axons of RGCs, the RGCs layer by the cell bodies of RGCs and by displaced amacrine cells, the IPL by amacrine cells and synapses between the dendrites of RGCs and the axons of BPCs, and the INL is comprised of horizontal, bipolar, amacrine, displaced ganglion and the cell bodies of Müller cells. The OPL is formed of synapses between photoreceptors, horizontal cells, and BPCs, and the ONL by the nuclei of the photoreceptors. The EZ, also known as the inner segment (IS) area, contains the inner segments of photoreceptor cells, while their outer segments reside in the POS. Finally, the RPE right behind the photoreceptors contains a simple layer of cuboidal cells and is connected to Bruch’s membrane and the choroid [1], as shown in Figure 1.
The visual process involves a complex interplay among various retinal cell types, each holding unique functions to contribute to the transmission of visual information. First-order neurons, the photoreceptors, are divided into two types: rods and cones. Rod photoreceptors specialize in detecting low-light levels, while cone photoreceptors enable photopic vision with high spatial acuity and color differentiation [27]. Horizontal cells, which are second-order neurons, modulate information transfer between BPCs and photoreceptors and provide lateral inhibition to enhance contrast and contribute to spatial receptive fields [28]. BPCs, another type of excitatory second-order neurons, receive input from photoreceptors and provide a link between them and the different types of RGCs. The various types of BPCs play distinct roles in collecting and modulating signals from photoreceptors, initiating the processing of visual stimuli within the inner retina [29,30]. Although amacrine cells typically release inhibitory neurotransmitters, the presence of gap junctions allows them to play both inhibitory and excitatory roles, acting as interneurons that interact with cells of the vertical retinal pathway – the pathway that involves the transmission of visual information from photoreceptors to BPCs to ganglion cells – and modulate signal transmission between neurons [31]. RGCs are the main output neurons for the retina and play dual roles in image-forming and non-image-forming processes [32]. Müller glial cells are involved in various metabolic processes and provide support to retinal neurons [33].
Each neuronal class exists in specialized types to perform unique functions, with three cone photoreceptor types, approximately three different types of horizontal cells, twelve types of BPCs, approximately twenty types of amacrine cells, and up to 45 types of RGCs [34]. Although the specific classifications are out of the scope of this review, we will mention necessary classifications in different contexts, and they will be further explained within each section when needed. Here, we briefly describe the major retinal neuronal classifications. Cone photoreceptors are composed of visual pigments, called opsins, and based on the structure of their opsins, cones are sensitive to either short- (or blue), medium- (or green), or long-wavelengths (or red wavelengths) of light [35]. In this way, these neurons can be classified as short-wavelength or blue cones, medium-wavelength or green cones, and long-wavelength or red cones according to their relative spectral sensitivity [35]. There is only one type of rod photoreceptors, which contains rhodopsin, the light receptor that initiates scotopic vision [36]. Horizontal cells have three types that have been identified: HA-1, HA-2, and HB [37]. Although more research is required to determine the selectivity of horizontal cells towards the different photoreceptors, HA-1 and HA-2 cells associate with cones without specificity, whereas HB cells seem to associate specifically with blue-sensitive cones given their large terminals organized in a rectilinear configuration [38]. There are, as previously mentioned, at least twelve types of BPCs, with one type connecting with rod photoreceptors (rod bipolar cell) and the remaining types connecting with cone photoreceptors (cone BPCs) [39]. These neurons are mainly divided into ON-type and OFF-type BPCs. OFF-type BPCs are hyperpolarized in the light and depolarized in the dark [40]. ON-type BPCs, on the other hand, are hyperpolarized in the dark and depolarized in the light [40]. Amacrine cell types can be broadly divided into ON-type, OFF-type, and ON-OFF amacrine cells [40]. ON-type amacrine cells receive information exclusively from ON-type BPCs, depolarizing in the presence of light. OFF-type amacrine cells receive information exclusively from OFF-type BPCs, therefore depolarizing in the absence of light. ON-OFF amacrine cells receive information from both types of BPCs and depolarize in both the presence or absence of illumination [40]. Amacrine cells may also be divided into subtypes based on the neurotransmitter they release, being mainly classified into either GABAergic amacrine cells or glycinergic amacrine cells, although a small population of these neurons has been shown to be neither GABAergic nor glycinergic (nGnG) [41]. Finally, RGCs can be classified as ON-type, OFF-type, and ON-OFF-type RGCs. ON-type RGCs are excited by illuminating their receptive field center and inhibited by illuminating their receptive field surround. OFF-type RGCs work oppositely, excited in their receptive field surroundings and inhibited by light in their receptive field center. ON-OFF RGCs can respond to both increase and decrease in light intensity within their receptive fields [40].
These different neuronal classifications play specialized functions in visual processing. Light first enters the eye through the cornea and passes through the pupil, iris, and lens before reaching retinal photoreceptor, where phototransduction, the conversion of photons of light into electrical signals, takes place [40]. In the vertebrate retina, both rod and cones respond to light with hyperpolarizing receptor potentials [42]. In the dark, the plasma membranes of rod and cone photoreceptors in the outer segment are extremely permeable to cations, as evidenced by the so-called dark current, which is a continuous influx of sodium ions (Na+) into the outer segments [42]. To achieve an electrical balance, efflux of potassium ions (K+) into the inner segments occurs via the activity of a Na+/K+ ATPase [43]. Photoreceptors are depolarized by the dark current, resulting in a tonic release of glutamate in the synaptic cleft forms a signal to the postsynaptic neurons [43].
In the presence of light, absorption of photons by photo-pigments in the outer segments reduces Na+ conductance and decreases the dark current, resulting in hyperpolarization of the membrane potential and a decrease in the release of tonic glutamate [44]. The sign-inversing and sign-conserving responses of bipolar neurons to glutamate are discussed in section 2.1.2. Briefly, under light stimulation ON-center BPCs become depolarized, while the OFF-center BPCs become hyperpolarized. BPCs release glutamate into the synaptic cleft to communicate with RGCs and amacrine cells in the IPL [45]. Depending on whether the signals are from ON or OFF BPCs, RGCs convey action potentials to the visual cortex for further processing [46]. Horizontal cells also hyperpolarize in response to light, thereby reducing the release of the neurotransmitter gamma-aminobutyric acid (GABA) in synapses with cone photoreceptors and BPCs [45]. Signals from neighboring rod and cone photoreceptors converge onto the same RGC. Such convergence allows rod photoreceptors to combine smaller signals and and produce more substantial responses in BPCs [47]. Finally, the resulting signals are transmitted to the brain through the RGCs, which form the optic nerve through the combination of their axons [40].
This review will focus mainly on information from literature on mouse and primate retina. The Mouse retina is rod-dominated with a rod:cone ratio of 35:1.[48,49] Only 5% of cones express purely a short (S) wavelength sensitive opsin, while the remaining co-express S-opsin and a middle (M) wavelength sensitive opsin [50,51]. On the other hand, the primate retina is enriched in cone photoreceptors with a rod:cone ratio of 20:1 [52]. Three opsins are expressed by the correspondent types of cones: S (blue), M (green) and L (red), allowing for fine discrimination between colors in the visible spectrum. The central retina of primates also contains a region specialized for high acuity called fovea. Within the cone-abundant fovea, a single cone photoreceptor contracts synapse with a single (midget) bipolar cell, which in turn contacts a single (midget) ganglion cell [53], ensuring the highest acuity vision by providing each cone a private line of communication to the brain [54]. The arrangement of photoreceptors in primate retina is matched to the function of foveal vs peripheral retina. The foveal region is crucial for color vision, with a ratio of cone types of L:M:S cones of 11:5:1 [55], whereas peripheral retina is more important for luminance coding and therefore abundant in rod photoreceptors.
1.2. Retinal Synapses
As we introduced in the previous section, neurons must communicate with each other precisely to transmit signals from photoreceptors to BPCs via synaptic transmission. Retinal synapses are comprised of two different types: conventional synapses, and specialized ribbon synapses. Within the central nervous system (CNS), the most common type of synapse is the conventional (also called central) synapse, which consists of an active zone containing synaptic vesicles in the pre-synaptic terminal and a post-synaptic density in the post-synaptic terminal. Conventional synapses are formed by all retinal cell types, except for photoreceptors and BPCs which form ribbon synapses, and vary in their functional properties based on the cell type [56].
Systems such as vision and hearing present ribbon synapses, specialized to respond to sensory stimuli and transmit these signals to appropriate cells to encode diverse information via neurotransmission [57]. The active zone of these synapses contains dense ribbon-like specialized organelles, called synaptic ribbons, that are tethered to the cell membrane [58]. In the retina, photoreceptors and BPCs contain such ribbon synapses, signaling graded changes in membrane potential [11,58,59,60]. Ribbons tether synaptic vesicles and calcium channels and have been hypothesized to play important roles in the facilitation of synaptic vesicle release within the synaptic vesicle cycle in a calcium-dependent manner, with vesicles being released when calcium channels are opened [59,61,62,63,64,65,66,67]. In ribbon synapses, Ca2+ influx has been shown to occur in hotspots located near the ribbon through voltage gated Ca2+ (CaV) channels [59,67].
Cav channels are grouped into CaV1, CaV2, or CaV3 subtypes based on the properties of the currents the channels mediate [68]. CaV1 channels mediate L-type currents, which are long-lasting currents that require strong polarization for their activation [69]. CaV2 channels mediate P/Q-, N-, and R-type currents, which are activated by high voltage and classified based on their sensitivity to different blockers and toxins [70,71]. CaV3 channels mediate T-type currents, which are transient currents that are activated by low voltage [69,70]. Ribbon synapses are driven by L-type Ca channels, or CaV1 channels, which are further classified into CaV1.1, CaV1.2, CaV1.3, and CaV1.4 [68]. These channels are tightly coupled to “big-potassium,” or BK, channels at presynaptic active zones in order to regulate the entry of calcium and the release of neurotransmitters [72]. BK channels are voltage-gated potassium channels that have a large potassium ion (K+) conductance across the cell membrane when activated by membrane depolarization and Ca2+ entry through CaV channels [73].
In conventional synapses, BK channels have been described as negative regulators of neurotransmitter release in frogs and lizards [74,75]. Similarly, in conventional synapses in the retina of rats, the contribution of CaV1 channels of amacrine cells to GABAergic release was reduced by BK channels as they suppressed postsynaptic depolarization of the neurons and limited the activation of CaV channels [76]. In ribbon synapses, on the other hand, results have shown mixed and dual roles of BK channels in neurotransmission. In the rod photoreceptors of salamanders, BK channels were found to facilitate transmitter release, with BK channel activation increasing extracellular levels of K+, therefore enhancing the Ca2+ channel currents and amplifying synaptic transmission [77]. In the mouse retina, BK channels did not seem to modulate the activity of cone BPCs and photoreceptors, but modulate the activity of rod BPCs instead, where the absence of BK channels under scotopic light conditions resultes in a significant reduction in electroretinogram signals [78]. Therefore, the function of BK channels and their interactions with CaV channels seems to vary based on experimental and physiological conditions, requiring a clearer understanding of the variability between studies and differences between the activity of Ca2+ and K+ channels in different retinal neuronal types.
1.3. Retinal Development
Retinal development is a complex and tightly regulated process that occurs during embryonic and early postnatal time. Across species, the basic components of the eyes and retina retain similar functions, although the timing of critical developmental events varies from one species to another [79]. This review will focus on vertebrate development, with a focus on mice, as retinal development in this species has been well-detailed by Hoon et al., 2015 and Cepko, 2014. Figure 2 illustrates the timescale of embryonic and postnatal development for various retinal neuronal types in mice. This figure, adapted from Hoon et al., 2015 and Cepko, 2014, will be recurrent in the paper as we detail the activity and presence of different neurotransmitters during retinal development.
Like other regions of the CNS, the developing retina is derived from the neural tube and is formed by the invagination of the optic vesicle to form the optic cup [80]. The external part of the optic cup becomes the RPE, while the internal part develops into the multi-layered retina [81]. The RGCs are the first cells to differentiate, at approximately E10 in mice, and extend their axons to form the optic nerve [82]. As development progresses, other cell types differentiate and establish connections within the retina in the following order: horizontal cells, cone photoreceptors, amacrine cells, rod photoreceptors, BPCs, and finally Müller glial cells [82,83].
Throughout the intricate development of the retina, events like cell migration, fate determination, and synaptogenesis ensure proper retinal organization and function [84,85,86,87]. After birth, the retina continues to refine and mature via visual experience and synaptic plasticity, with synaptic connections being finely tuned for proper visual pathways and receptive fields [88]. During retinal development, neurotransmitters play vital roles in shaping neural circuits and establishing functional connections, thereby contributing to the precise processing and transmission of visual information in the mature retina. The refinement of retinal connections and receptive fields relies on Ca2+-dependent and Ca2+-independent processes together with a fine balance between excitatory and inhibitory neurotransmitters, ensuring the development of a proper visual pathway. The culmination of retinal development is a well-organized and functional neural network that facilitates the detection and transmission of visual information to the brain.
Neurotransmission
This section will provide an overview of the current understanding of the retinal neurotransmitters glutamate, gamma-aminobutyric acid (GABA), glycine, dopamine, and acetylcholine (ACh) and of their interplay with Ca2+ in multiple retinal mechanisms. Table 1 summarizes the roles of these neurotransmitters in retinal development and normal retinal physiology, Table 2 summarizes their involvement in different retinal diseases, and Table 3 outlines their interplay with Ca2+-dependent processes discussed herein.
1.4. Glutamate
The primary excitatory neurotransmitter in the nervous system is the amino acid glutamic acid (glutamate), which plays a key role in human retinal function. Glutamate is released by photoreceptors, bipolar neurons, vglut3 amacrine cells, and RGCs. It is primarily involved in the vertical pathways of the retina [89,90], where it stimulates postsynaptic membranes via rapid depolarization and Ca2+ ion influx [91,92]. Glutamate receptors are classified into ionotropic receptors (iGluRs), which consist of NMDA receptors, AMPA receptors, and kainite receptors (KAR), and metabotropic receptors (mGluRs). Ionotropic receptors are usually ligand-gated ion channels, whereas metabotropic receptors activate signaling cascades through G-proteins and second messengers [93].
2.1.1. The Role of Glutamate in Retinal Development
Glutamate plays a significant role in the development and maturation of RGCs by regulating spontaneous neural activity during retinal waves and by guiding activity-dependent processing of the visual field map [94,95,96]. Retinal waves are comprised of spontaneous neural activity that occurs during RGC development and serve as a foundation for the activity-dependent refinement of retinotopic maps in the brain [97]. While the establishment of glutamatergic retinal waves was initially attributed to amacrine cells and BPCs, more recent reports reveal the involvement of Müller glial cells in the process [98]. During the occurrence of retinal waves, the ionotropic α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) glutamate receptors regulate Ca2+ activity in Müller glial cells, while the glutamate transporters, including the excitatory amino acid transporters EAAT1 and EAAT2, play a modulatory role, allowing the cells to sense and control the retinal waves by adjusting the concentration of extracellular glutamate [98,99]. In mice, glutamatergic retinal waves occur between postnatal days 10 and 14 [100,101], as shown in Figure 3, gray shaded bar.
The retina also expresses Ca2+-permeable N-methyl-D-aspartate (NMDA) receptors for glutamate that, together with non-NMDA receptors, regulate the outgrowth and stabilization of RGC dendrites during pre-embryonic development and their remodeling during postnatal further development [96,102,103]. In many experimental animal models, neurons in the IPL and GCL express high levels of NMDA receptors, beginning during the early phases of retinal development and increasing after birth. These results suggest that they are likely involved in early retinal development and the establishment of synapses within the inner retina, primarily in amacrine cells and RGCs [103,104,105,106]. Thus, NMDA receptors are likely to regulate the differentiation of different cell types during pre-embryonic development [103].
After birth, glutamate-mediated neuronal activity may be more prominent in defining the structure of the inner retina, primarily the remodeling of the dendrites and stratification of RGCs [95,103]. NMDA conductance determines the amount of current that can pass through NMDA receptor channels when open. Interestingly, NMDA conductance decreases as retinal development progresses, meaning that the receptor channels become less effective [102]. In contrast, the rate of desensitization of ionotropic glutamate receptors increases as development progresses [102]. It is hypothesized that such change in behavior throughout the developmental process likely enables RGCs to integrate synaptic information, facilitating the transmission of electrical signals to the brain. These observations reinforce the idea of the various roles played by glutamate throughout retinal development, although more research will be required to form a more complete understanding of the functions of glutamate and its receptors during development.
2.1.2. The Role of Glutamate in Normal Retinal Physiology
In the mature retina, glutamate acts on vertical pathway neurons and is used primarily by photoreceptors as an excitatory neurotransmitter for the relay of signals to the second-order neurons [45]. While photoreceptors release glutamate upon depolarization, providing the basis for signal transmission and subsequent visual processing [107], the mechanism of release is distinct for cone and rod photoreceptors. Cones release glutamate solely through synaptic ribbon sites, whereas similarly to BPCs, rods exhibit a broader release pattern from both ribbon and non-ribbon sites [108,109,110]. Glutamate undergoes modifications that are mediated by horizontal cells, different bipolar cell types, and is further refined by amacrine cells [40]. ON BPCs express metabotropic glutamate receptors and exhibit sign-inversing responses by coupling to G-proteins, in which glutamate hyperpolarizes the membrane of ON BPCs, inhibiting synaptic transduction currents. Glutamate receptors of OFF BPCs, on the other hand, display sign-conserving behavior, in which an increase in glutamate signaling results in an increased electrical response of the OFF BPCs [111,112,113].
The intricate distribution and activity of glutamate receptors further contribute to their role in normal retinal functioning. Glutamate receptor subtypes, including AMPA, NMDA, and KA receptors, are differentially located across different neurons (Figure 4), where they play distinct roles. The AMPA and KA receptors are located at postsynaptic density sites, where they allow passage of primarily Na+ and K+ ions in OFF BPCs, horizontal cells, and RGCs, while the peri-synaptic NMDA receptors are present in some ON BPCs, amacrine cells and RGCs, and permeable to Na+, K+, and Ca2+ ions [107,114,115]. NMDA receptors, alongside AMPA receptors, are capable of modulating glutamate release in the retina without significant limitation of the temporal tuning of synaptic responses in RGCs [114].
Alongside with iGluRs, the retina expresses mGluRs, which are present in eight types across the different retinal neurons (mGluR1-8). In rodents, mGluR1 has been shown to be present in bipolar, amacrine, and RGCs; mGluR2 in amacrine and RGCs; mGluR3 in RGCs; mGluR4 in amacrine and RGCs; mGluR5 in horizontal, bipolar, amacrine, and RGCs; mGluR6 in bipolar and RGCs; mGluR7 in bipolar, amacrine, and RGCs; and mGluR8 in photoreceptors, horizontal, amacrine, and RGCs (Figure 4) [116]. The sign-inversing response of the metabotropic glutamate receptors described earlier is possible especially because of mGluR6 on the dendrites of ON BPCs. mGluR6 binds the glutamate from photoreceptors and activates the G protein Go, which leads to the closing of the cation channel TRPM1 (Transient Receptor Potential Cation Channel Subfamily M Member 1), therefore hyperpolarizing the neuron [117,118,119].
2.1.3. The Role of Glutamate in Retinal Diseases
Dysregulation in the balance of glutamate can severely impact retinal function. The metabolism of glutamate is governed by its uptake and conversion by the adenosine triphosphate (ATP)-dependent glutamine synthetase (GS) to non-toxic glutamine in Müller glial cells, a highly energy-dependent process [120]. Thus, a lack of energy can disrupt the cell’s glutamate equilibrium, resulting in oxidative stress, mitochondrial impairment, and upregulation of NMDA receptors, ultimately inducing excitotoxicity [121,122]. This process has been implicated in the degeneration of RGCs, which vary in sensitivity by cell type, with α-type RGCs, which are RGCs with large somata and dendritic field [123], displaying more resistance and J-type RGCs, which are direction selective cells that respond specifically to upward motion [124], being the most sensitive [34]. Excitotoxicity can also result from excessive stimulation of glutamate receptors, underlying a pivotal mechanism that contributes to retinal pathology [125,126]. Interestingly, this process can be mitigated by glutamate-release inhibitors, which may serve as the basis for potential therapeutic interventions that aim to reduce glutamate neurotransmitter activity [127].
Dysfunctional glutamate metabolism is linked to retinopathies such as retinal ischemia, which is induced by the occlusion of retinal or choroidal blood vessels. Acute retinal ischemia sparks a critical depletion of ATP and glutamine synthetase [128,129,130], resulting in elevated levels of extracellular glutamate that trigger a destructive excitotoxicity cascade characterized by neuron depolarization, the influx of Ca2+ ions, oxidative stress, and robust glutamatergic stimulation, which contributes to the degenerative repercussions of ischemia and reperfusion scenarios [131]. Notably, the function of the key Müller cell glutamate aspartate transporter (GLAST) decreases significantly after retinal ischemia, resulting in an elevation of glutamate levels within the aqueous humor and the inner retina [132,133,134]. However, this impairment is not permanent, since the function of GLAST is recovered on reperfusion [135].
The pathogenesis of the ocular disorder glaucoma, characterized by the apoptotic loss of RGCs, remains poorly understood but appears to involve glutamate excitotoxicity [136]. A reduction in the levels of the glutamate transporter GLAST is associated with increased intraocular pressure (IOP) poses a significant risk for the development and progression of glaucoma, causes damage to the optic nerve, and ultimately leads to vision loss [137,138]. Interestingly, in animal models of glaucoma, the absence or downregulation of different glutamate transporters resulted in the death of RGCs, even in the absence of high IOP [138]. In normotensive glaucoma, deletion of the apoptosis signal-regulating kinase 1 (ASK1) gene suppresses the oxidative stress that is linked to glutamate-mediated excitotoxicity. increases survival of RGCs and reduces the production of the pro-inflammatory cytokine tumor necrosis factor-alpha (TNF-α) by macrophages, microglia, and astrocytes [139,140]. Thus, a potential therapeutic approach to reverse the effects of glutamate-related toxicity might include ASK1 inhibitors, such as the highly selective inhibitor selonsertib.
The progression of retinal degeneration in diabetic retinopathy, once thought to result from a microcirculatory disorder, is now known to involve glutamate and Ca2+ signaling pathways. The early stages of diabetes mellitus are associated with heightened levels of glutamate and oxidative stress [141,142]. In rat Müller cells, NMDA receptors inhibit the secretion of vascular endothelial growth factor (VEGF), thereby countering blood-retinal barrier breakdown, indicating the protective effect of a well-maintained glutamatergic system [143,144]. In rat models of diabetes, the NMDA receptor inhibitor memantine blocks elevated levels of VEGF [144]. However, as noted above, the protective role of glutamate is compromised in diabetic retinopathy, since Müller cells in the inner retina of diabetic individuals exhibit decreases in GLAST activity and Müller cell immunoreactivity [145,146]. To mitigate the detrimental impact of dysregulated glutamate and Ca2+ signaling in diabetic retinopathy, novel therapeutic targets might include the mitochondrial membrane lipid cardiolipin, which is linked to apoptosis cascades, and antioxidant factors such as lipoic acid, carotenoids, and nuclear factor (erythroid-derived 2)-like 2 (Nrf2), which plays a crucial protective role against oxidative stress [147,148,149,150,151].
2.1.4. The Interplay of Glutamate and Calcium
As described above, retinal waves generate spontaneous bursts of Ca2+ that occur during retinal development [101]. During retinal waves, depolarization of ON cone BPCs leads to a transient increase in intracellular Ca2+ levels and a release of glutamate to the next neuronal cells, the ON RGCs, which generate bursts of action potentials in the form of retinal waves [152]. In the presence of glutamate, RGCs and amacrine cells display a significant increase in Ca2+ levels [153]. These responses are initially mediated by AMPA and KA receptors, but the NMDA receptors fulfill this role as development progresses, underlining the hypothesis that the different types of receptors play different roles during development. The interplay between glutamate activity and changes in intracellular Ca2+ levels is crucial for the outgrowth, stabilization, and differentiation of various cell types, especially amacrine cells, and RGCs, during development [96,102,104,153].
In normal physiological conditions, glutamate is present in vast amounts, especially at retinal ribbon synapses where synaptic vesicles filled with glutamate are attached [11]. Ribbons are crucial in the pre-synaptic release of glutamate, in which they assist with the replenishment of synaptic vesicles and organize the proteins necessary for glutamate release, vesicular clustering, and positioning of voltage-gated Ca2+ channels at synaptic active sites [154,155]. Non-ribbon glutamate release also occurs in the retina, involving Ca2+-mediated processes, such as Ca2+-induced calcium release (CICR) from intracellular endoplasmic reticulum (ER) stores [156,157].
In retinal ischemia, the rise of intracellular Ca2+ levels determines the beginning of excitotoxic glutamate-related neuronal death [158] that occurs in RGCs of rats due to the activation of AMPA/kainite receptors during visual processing, which facilitates the influx of Ca2+ through NMDA receptors [92]. In living animals, such depolarization of RGCs is mediated by glutamate release during visual stimuli in the retina [92]. One possible mechanism of cell death in diabetic retinopathy involves a step-wise increase in retinal glutamate levels, activation of NMDA receptors, increased levels of intracellular Ca2+, induction of RGC apoptosis, and further activation of NMDA receptors, a cycle that contributes to the death of numerous retinal cells [159].
1.5. Gamma-aminobutyric acid (GABA)
The principal inhibitory neurotransmitter in the CNS and retina is gamma-aminobutyric acid (GABA), which plays a critical role in regulating visual processing. GABA is released, primarily via Ca2+-dependent conventional exocytosis and Ca2+-independent mechanisms via GABA transporters [160], from GABAergic horizontal cells, amacrine cells, to BPCs and RGCs, where GABA is crucial for the processing of visual information and the development of retinal circuitry in the inner retina [161].
2.2.1. The Role of GABA in Retinal Development
As noted above, GABA released by amacrine cells and RGCs is an important regulator of inner retinal development [7,162,163,164,165,166,167]. GABAB receptors are metabotropic G protein-coupled receptors on amacrine cells and RGCs, where they are capable of inducing neuronal hyperpolarization through the opening of G protein-gated inwardly rectifying K+ channels and of inhibiting neurotransmitter release by inhibiting voltage-gated Ca2+ channels both pre- and post-synaptically [168]. During early neurogenesis in the retina of embryonic chickens, an agonist of GABAB blocked Ca2+ signaling during retinal Ca2+ waves, while the GABAB antagonist p-3-aminopropyl-p-dietoxymethyl phosphoric acid increased the duration of the transient, illustrating that the GABA receptor subtype plays a crucial role in regulating Ca2+ waves prior to synapse formation [169]. In the mouse retina, expression of the enzyme L-glutamate decarboxylase (GAD) was used to track the developmental pattern of GABAergic neurons (Figure 5). GAD expression was robust during the period that spanned embryonic day 17 to postnatal day 3, then significantly declined after postnatal day 12 [170].
GABA is also implicated in postnatal stages of retinal development. During the early postnatal weeks in the ferret retina, GABA potentiates activity on all RGCs through the action of GABAA receptors [171]. As development progresses and ON-OFF cell segregation occurs, the role of GABA shifts to suppression of ganglion cell burst activity [171]. This developmental transition underscores the multifaceted roles of GABA in shaping distinct patterns of presynaptic activity, extending its functions its inhibitory characteristics.
2.2.2. The Role of GABA in Normal Retinal Physiology
GABA plays an important role in shaping the retinal response to visual stimuli. Its physiological impact is mediated via three types of membrane receptors: ionotropic GABAA and GABAC receptors and metabotropic GABAB receptors [172,173,174]. The GABAA receptor is comprised of five subunits that encircle a chloride channel and is highly sensitive to the receptor antagonist bicuculline and the chloride channel blocker picrotoxin [175,176]. GABAA receptors have been shown to be present in photoreceptors, horizontal cells, bipolar cell axons and dendrites, amacrine cell processes, RGCs, and Müller glial cells in different species [162,163]. As demonstrated in mice, GABAA receptor activation inhibits the proliferation and self-renewal of retinal progenitor cells [177]. Metabotropic G protein-coupled GABAB receptors in the retina are present in horizontal neurons, amacrine neurons, and RGCs [178], playing crucial roles in signal modulation, inhibiting neurotransmitter release from the presynaptic terminal or modulating the balance of brief and sustained signals in the retinal circuitry [168]. GABAC receptors are present at different synaptic sites in cone photoreceptors, horizontal cells, BPCs, amacrine cells, and RGCs, and gate chloride ion (Cl-) channels [107]. They function as auto-receptors, limiting the release of GABA itself [179]. GABAC receptors have been identified as GABA receptors that are resistant to bicuculline, and baclofen, a GABAB receptor agonist [180]. However, given that the retinal bipolar neurons of goldfish contain two distinct bicuculline/baclofen-resistant receptors, one coupled to Cl- channels (a GABAA-type receptor) and another coupled to Ca2+ channels through a GTP-dependent process (a GABAB-type receptor), Matthews et al. proposed that receptors that were previously classified as GABAC receptors belong in the category of GABAA and GABAB receptors instead [180].
The release of GABA is tightly regulated by both Ca2+-dependent and Ca2+-independent mechanisms [181]. In the catfish retina, an increase in the concentration of intracellular Na+ triggers GABA transporters to release GABA via a non-vesicular, Ca2+-independent mechanism [181]. Ca2+-dependent mechanisms occur upon the influx of Ca2+ ions during depolarization, ultimately leading to the release of different neurotransmitters. In such mechanisms, for example, during brief depolarization in retinal amacrine cells, the intracellular release of Ca2+ from internal stores further elevates its concentration, resulting in GABA release [182]. Following the release of GABA, its concentration is maintained collectively by the GABA transporters (GAT1, GAT2, GAT3, and GAT4) and the vesicular transporter VGAT [183,184,185], while the termination of the signal is achieved via uptake mechanisms that involve both presynaptic terminals and glial cells [186]. In the inner retina, amacrine cells and Müller glial cells participate in GABA uptake, while Müller glial cells are the neurons primarily responsible for this function in the outer retina [187]. Upon entry into Müller cells, GABA is metabolized by the mitochondrial enzyme GABA transaminase to yield succinate semialdehyde [187]. Figure 6 illustrates the localization of the different types of GABA receptors in the retina.
2.2.3. The Role of GABA in Retinal Diseases
GABA dysregulation is associated with diabetic retinopathy and retinal ischemia. Diabetic retinopathy involves alterations in GABA metabolism in Müller cells whereby heightened activity of GAD leads to an increase in accumulation of GABA while the [15] GABA transaminase (GABA-T) that metabolizes GABA exhibits reduced activity [15]. In rats, this shift is accompanied by the detection of reduced oscillatory potentials (OPs) using an electroretinogram (ERG), highlighting the functional impairment associated with diabetic retinopathy [15]. Retinal ischemia, a pathological condition previously discussed in terms of glutamate activity, leads to the rapid accumulation of GABA in a manner similar to that observed for glutamate [16]. Interestingly, in glial cells, the accumulation of GABA is more rapid and reaches higher peak levels due to a decrease in GABA transaminase activity in these cells [188]. Thus, glial cells will be relevant for the development of novel targeted treatments for retinal ischemia.
Therapeutics that target GABA pathways have the potential for the treatment of various diseases that affect retinal health. For example, anticonvulsants that influence GABAergic signaling also exert neuroprotection of RGCs against excitotoxicity, thereby representing promising treatments for retinal ischemia [18]. The GABA-transaminase inhibitor vigabatrin (VGB) shows efficacy against retinal ischemia, although with side effects that include possible irreversible visual field defects [189]. Similarly, an inhibitor of the high-affinity GABA transporter GAT1 tiagabine (TGB) offers reversible protection of the retina but may cause altered color perception [190]. Recent investigations involve the use of chloride derivatives as alternative therapeutics that maximize positive outcomes while minimizing irreversible side effects, thereby bypassing retinal risks [191].
2.2.4. The Interplay of GABA and Calcium
The interplay of GABA and Ca2+ during retinal development occurs in various animal models. In the turtle retina, GABA is responsible for changes in the dynamics of retinal Ca2+ waves throughout development [192]. Similarly, GABAB receptors in chicken retinas regulate the retinal Ca2+ waves that appear early in development [169]. GABAB receptors appear to modulate the rate and duration of Ca2+ waves in the retina by inhibiting Ca2+ channels, resulting in a decrease in the influx of Ca2+ in RGCs and amacrine cells [169]. The release of GABA from amacrine cells is dependent on a slow, asynchronous release of Ca2+ resulting from its accumulation in the cell terminals [193]. Horizontal cells also release GABA to photoreceptors, where it stimulates the opening of Ca2+ channels, leading to depolarization of photoreceptors to balance the hyperpolarization caused by light [194,195].
Levels of Ca2+ are also related to diseases in which GABA is involved. For example, in diabetic retinopathy, reduced Ca2+ levels in pre-synaptic GABAergic amacrine cells are associated with a reduced release of GABA [196]. While little is known about the effects on Ca2+-dependent mechanisms of the antiepileptic drugs used to treat retinal ischemia, other drugs typically used to treat epilepsy such as ethosuximide are associated with reduced Ca2+ currents through T-type channels [197]. Additionally, intraocular pressure (IOP) in the eye can damage the optic nerve, contributing to the risk of developing glaucoma. Chintalapudi et al. (2017) used mice to test the effects on IOP of the gabapentinoid drug pregabalin which is specific for the Ca2+ voltage-gated channel auxiliary subunit alpha2delta1 (Cacn2d1) gene [198]. These authors found that pregabalin reduces the flow of Ca2+ through its selective channels, thereby reducing intracellular Ca2+ levels and lowering the impact of IOP [198].
1.6. Glycine
The amino acid neurotransmitter glycine is found in the synapses between AII amacrine cells and OFF RGCs, where it plays an integral part in scotopic (low-light) vision [199]. As one of the main inhibitory amino acids in the CNS, glycine helps to shape neuronal pathways and facilitates visual processing in the dark [200].
2.3.1. The Role of Glycine in Retinal Development
Glycine receptors exhibit both excitatory and modulatory properties that are crucial for the development of retinal synapses. Neonatal glycine receptors (GlyRs), comprised of α2 subunit homopentamers, contribute to the establishment of a depolarizing glycine-gated Cl- flux that subsequently triggers Ca2+ influx. This process is integral to neuronal specialization, including the development of glycinergic synapses [201,202]. GlyR α2 subunits are expressed at very high levels in retinal progenitor cells of both the developing and the adult retina (Figure 7), starting at birth (postnatal day 0) in mice [203]. The accumulation of GlyR α1, α3, α4, and β subunits also exhibits a steady increase during postnatal retinal development [204]. However, there is a paucity of studies regarding the role of glycine in retinal development and further research will be necessary to uncover its mechanisms and influences during development.
2.3.2. The Role of Glycine in Normal Retinal Physiology
In the retina, glycine is found in some cone BPCs and most amacrine cells, the latter of which regulate inhibitory neurotransmission [205], as follows. The retina efficiently segregates visual signals into ON and OFF pathways before they ascend to the visual cortex. Notably, the AII (rod) amacrine cells release glycine at their synapses with OFF BPCs, other amacrine cells, and RGCs, parsing out signals from the rod photoreceptors to both the ON and OFF pathways in the INL – with the transmission of signals from AII to the ON pathway primarily relying on gap junctions –, each with tailored glycinergic responses to distinct types of BPCs [206]. Glycine and glutamate play vital roles in modulating crossover pathways that lead to the dominant ON crossover inhibition to OFF BPCs and RGCs, shown in Figure 8, which was adapted from Hsueh et al., 2008. OFF RGCs receive glutamatergic OFF excitation from OFF BPCs, which is compensated by crossover inhibition of OFF BPCs and RGCs from ON amacrine cells [207]. This crossover inhibition is compensated by glutamatergic excitation of ON amacrine cells from ON BPCs [207]. Finally, crossover inhibition from OFF amacrine cells onto ON amacrine cells compensates for the previously mentioned glutamatergic excitation [207].
Post-development, the functional roles of glycine are facilitated by glycine receptors (GlyRs) comprised of α and β subunit heterooligomers [208]. The GlyRs are ligand-gated ion channels that are sensitive to strychnine, which increases the permeability of the postsynaptic membrane to chloride ions [209]. Their distribution spans the IPL, INL, and RGC layer, with different kinetic speeds that are tailored to their functions. For example, in BPCs and A-type RGCs, α1β-containing GlyRs facilitate rapid signal transmission, with spontaneous inhibitory synaptic currents (sIPSCs) displaying medium fast kinetics with a lower decay time constant, while in AII amacrine cells, α3β-containing GlyRs relay rod light signals with reduced temporal resolution [205]. In narrow-field amacrine cells, α2β- and α4β-containing GlyRs contribute to modulatory functions where temporal precision is less important [205]. Notably, the α1 subunits of GlyRs play a critical role in scotopic vision, particularly at the synapses of AII amacrine cells and OFF RGCs, where they modulate the transmission of visual signals under low-light conditions [200].
2.3.3. The Role of Glycine in Retinal Diseases
Since diabetes mellitus is characterized by a significant reduction in levels of the hormone insulin, which is crucial for amino acid metabolism, it has been hypothesized that the pathogenesis of the disease is related to a deficiency of amino acids, including glycine [19]. For this reason, the therapeutic potential of glycine supplementation has been explored for the treatment of diabetic retinopathy. In diabetic rats, treatment with glycine improved the chromatin distribution and nuclear profiles of RGCs facing apoptosis [21]. Glycine possesses antioxidant, anti-inflammatory, and anti-glycation properties [210]. On glycine treatment, Müller cells exhibit increased expression of glial fibrillary acidic protein (GFAP) and other intermediate filament proteins, resulting in the accumulation of advanced glycation end products (AGEs) and inflammatory mediators and an increase in levels of oxidative stress [210]. Glycine also prevents Müller cell reactivation and improves disorganized arrangements of the outer segments of cone and rod photoreceptors [210]. Finally, glycine supplementation attenuates retinal neuronal damage in rat models of diabetes, ameliorating its detrimental effects on the INL, ONL, and RGCL [211].
2.3.4. The Interplay of Glycine and Calcium
While the interplay between glycine and Ca2+ during development or in retinal diseases remains largely unknown, it is observed during normal physiological function. Glycine signaling via a G-protein-associated receptor pathway inhibits voltage-gated Ca2+ currents, thereby reducing signal transmission in the retina from BPCs to RGCs [212]. Furthermore, ATP suppresses the Ca2+ currents mediated by glycine receptors in OFF RGCs via the mediation of metabotropic P2Y(1) and P2Y(11) purinergic receptors, thus altering the pattern of action potentials in the retina [213]. Additionally, glycine release has been shown to be potentiated by cyclic AMP (cAMP) through exchange protein activated by cAMP2 (EPAC2) and Ca2+ stores in AII-type amacrine cells [214]. By using agonists and antagonists for EPAC2, Marc et al. found that the cAMP-induced increase of glycinergic release requires the activation of EPAC2 and intact internal Ca2+ stores [214]. cAMP level dynamics are an important part of glycine release within the retina and it has been proposed that the activation of EPAC2 results in Ca2+ release from Ca2+ stores, which in turn increase the release of glycine onto OFF-cone BPCs terminals [214]. Such release modulation is important because it ensures proper synaptic transmission in the retina, allowing for sharper vision with reduced synaptic noise [215].
1.7. Dopamine
Dopamine, being produced and released by dopaminergic amacrine cells in the retina, plays a fundamental role in the functioning of human vision, being largely responsible for retinal physiology and visual perception [216]. The neurotransmitter influences spatiotemporal vision, brightness, and the circadian clock, all of which contribute to the refinement and enhancement of our interpretation of the world [217].
2.4.1. The Role of Dopamine in Retinal Development
Although retinal cells that will develop into dopaminergic neurons are detected early in retinal development, dopamine does not appear to contribute to the development of the retina or its synapses [216]. In mice, dopamine levels become detectable starting on postnatal day 6 and increase post-birth (Figure 9), when its function in relation to light-dark cycles is first observed [218,219].
2.4.2. The Role of Dopamine in Normal Retinal Physiology
Dopamine plays a vital role in the normal functioning of the retina that is mediated primarily via the dopamine G protein-coupled receptors of types D1R and D2R, which influence cellular activity via the cAMP/PKA transduction pathway [220]. D1R enhances voltage-gated Ca2+ channels in neurons via stimulatory G proteins (Gs), stimulating the release of dopamine from synaptic vesicles [216,220], while D2R utilizes inhibitory G proteins (Gi) receptors to reduce dopamine release [220].
The synthesis and release of dopamine by amacrine cells are closely tied to the intensity of incident light on the retina, with dopamine levels increasing significantly during the day in response to light. In an opposing balance with melatonin-secreting cells, dopamine uses a feedback mechanism to modulate rod photoreceptor signaling, thereby aiding adaptation to changing light levels [216,221]. Interestingly, the response of dopamine to non-Circadian changes in levels of light and darkness influences the receptive field properties of certain cells, enhancing their ability to detect specific visual features (Figure 10) [222]. With bright illumination (e.g., during daytime), dopamine release activates low-affinity D1 receptors on cone-BPCs, strengthening their receptive field, which is hypothesized to enhance the detection of edges and fine details in the animal retina [223,224].
2.4.3. The Role of Dopamine in Retinal Diseases
Visual impairments can be associated with diseases such as Parkinson’s disease (PD) that involve an imbalance in dopamine levels; in PD, there is a significant decrease in dopamine levels, including in the retina [225]. Individuals with PD can exhibit deficits in contrast sensitivity, loss of color vision, and reduced photopic and scotopic vision [226]. These symptoms are associated with the loss of dopaminergic amacrine and RGCs in the retina and impairment of electroretinographic response, suggesting that dopamine deficiency plays at least a partial role in the visual symptoms of PD [14,17].
Low levels of dopamine are also associated with form-deprivation myopia, a type of nearsightedness that occurs due to a developmental elongation of the eye [13,20], and in animal models, the administration of dopamine prevents the development of form-deprivation myopia [227,228]. Because light stimulates the release of dopamine, bright light exposure has been explored as a possible alternative treatment for form-deprivation myopia [229,230,231]. Thus, given the important interplay between light exposure and dopamine, it will be important to explore treatments and preventative methods to reduce visual impairment that combine dopamine and light.
2.4.4. The Interplay of Dopamine and Calcium
While little is known about the roles of the interplay between Ca2+ and dopamine in development and retinal disease, dopamine receptors and Ca2+ work together in neurotransmission in the healthy retina. When activated, D4 receptors in photoreceptors induce changes in the elicited Ca2+ current that can increase or reduce neurotransmission in response to light, depending on the cell subtype [232,233]. The balance between melatonin and dopamine release also seems to be dependent on Ca2+. In the rabbit retina, Dubocovich (1983) reported that melatonin inhibits the Ca2+-dependent release of dopamine in a dose-dependent manner [234]. Similarly, Nowak et al. (1989) reported that melatonin blocks electrically-evoked Ca2+-dependent dopamine release in the retina, suggesting that melatonin and dopamine, combined with Ca2+ dynamics, may regulate retinal mechanisms related to the Circadian rhythm [235].
1.8. Acetylcholine
The activity of the neurotransmitter acetylcholine (ACh) in the CNS is critical for such physiological activities as direction selectivity, regulation of retinal waves, and providing input to RGCs [161].
2.5.1. The Role of Acetylcholine in Retinal Development
ACh plays a crucial role in retinal development, particularly in the context of the stage II cholinergic retinal waves that are crucial for the establishment of the retinal circuitry and precede the glutamatergic waves in mice, occurring during postnatal days 1 – 10 [100,101], as shown in Figure 11. Just before the eyes of neonatal mice open, there is a shift from cholinergic connections to glutamatergic circuit activity, and retinal waves become regulated by glutamate, as described earlier in this review [98,99,236].
In RGCs, retinal waves occur in synchronized Ca2+ bursts that seem to coincide with the release of ACh [237,238]. The effects of ACh are closely linked to the activity of starburst amacrine cells (SACs), which use transient action potentials during development and may function as a part of a network that aids in the establishment of neural connectivity [239]. In various animals, blockage of cholinergic waves is associated with changes in retinal development, such as the reduction of dendritic motility, stratification and growth in RGCs, and reduction of the receptive field [240,241,242]. Arroyo et al. (2016) showed that the blockade of cholinergic retinal waves results in an increase of intrinsically photosensitive RGC gap junction networks with other neurons. This suggests that cholinergic waves normally suppress the formation of communication networks between these cells, supporting the interpretation that such network modulation is regulated through dopaminergic release of these waves during retinal development [243]. Combined, such studies led to the hypothesis that ACh plays an important role in the development of the intricate retinal network, perhaps by contributing to the initiation and propagation of retinal waves, while also contributing to the growth and connectivity of retinal cells.
2.5.2. The Role of Acetylcholine in Normal Retinal Physiology
ACh is produced from choline in starburst amacrine cells (SACs) and released. The effects of ACh in various cell types, including other amacrine cells, BPCs, and RGCs, are regulated by muscarinic acetylcholine receptors (mAChRs) and nicotinic acetylcholine receptors (nAChRs) [244,245,246,247]. Both mAChRs and nAChRs are present in BPCs, amacrine cells, and RGCs [247,248,249,250,251,252,253]. There are five subtypes of mAChRs, M1, M2, M3, M4, and M5 and five subtypes of nAChRs, α3, α4, α6, α7, β2 and β4 [253]. While mAChRs can enhance or diminish RGC responsiveness, nAChRs usually increase their responses by allowing the entry of Na+ and Ca2+ ions [247]. The activation of these receptors thus mediates the strength of the peak and inward current response of these cells to light stimuli (Figure 12), although the role of such changes is not clearly understood [247]. Additionally, the role of the retinal mAChRs in regulating functions such as pupil constriction, accommodation, and tear production has been the focal point of several studies [254]. Within the retina, the M1 subtype of mAchRs is especially important for the survival of RGCs [255].
Importantly, ACh plays a crucial role in direction selectivity, an important aspect of vision where direction-selective cells increase their activity in favor of specific orientations presented in the field of view [256]. The release of ACh from SACs occurs in a direction- and motion-sensitive manner and is transmitted to ON-OFF direction-selective RGCs [256,257,258,259]. These cells also secrete GABA, which is only released in response to null directions, thereby demonstrating an important interplay between GABA and ACh in determining direction selectivity [257]. Type 7 retinal BPCs normally contain nAChRs, which led to the investigation of the neuron’s role in the direction selectivity observed in SACs [260]. Type 7 BPCs isolated from mice with a genetic nAChR knockout were found to contribute to the ability of the SACs to respond to stimuli in a direction- and motion-dependent manner via cholinergic feedback [260]. This suggests that the cholinergic feedback to BPCs amplifies direction-selective signaling in SACs, highlighting the important role of ACh in augmenting motion detection in the retina [260].
2.5.3. The Role of Acetylcholine in Retinal Diseases
ACh is not particularly associated with the development of retinal diseases, ACh receptors provide a promising avenue for neuroprotection from glaucoma and delaying the death of retinal cells. ACh can control neuroprotective pathways in both the developing and mature retina, particularly the mAchRs that are vital for neuronal survival [254]. This is supported by the observation that in cases of acute closed-angle glaucoma, the ACh agonist pilocarpine constricts the iris via the M3 receptors, thereby shielding the retina from long-term damage via vascular mechanisms [244]. However, it is important to mention that non-subtype-selective mAChR agonists can result in unwanted side effects. For example, pilocarpine has been used for control of IOP, but its long-term use is associated with ciliary spasms, blurred vision, miosis, and retinal detachment [261], highlighting the need for the future development of treatments that maximize the benefits and minimize side effects.
2.5.4. The Interplay of Acetylcholine and Calcium
As previously discussed, the Ca2+ bursts that occur during development propagate across the retina, where they coincide with the extracellular release of ACh, thereby contributing to the establishment of neuronal networks [257,260]. Thus, the interplay between Ca2+ and ACh is crucial in shaping retinal development. G protein-coupled mAChRs in the retina (including subtypes M1, M3, and M5) activate the alpha subunits of Gq/11 proteins, raising the levels of intracellular Ca2+ [253]. This surge in Ca2+ can enhance PKC activity and potentially stimulate the production of nitric oxide (NO) via Ca2+-dependent neuronal nitric oxide synthase [253]. The M2 and M4 receptors are coupled to Gi and Go proteins, respectively, and inhibit cAMP-dependent pathways, thereby indirectly influencing Ca2+ dynamics [253]. This intricate interplay between ACh and Ca2+ is a crucial regulatory mechanism in retinal function and signaling. The direction-selective release of ACh and GABA is a Ca2+-specific process and these neurotransmitters are likely released from different synaptic vesicle pools [257].
As discussed, although Ach is not associated with the development of many retinal diseases, some ACh agonists are neuroprotective. Drugs such as pilocarpine which are effective against glaucoma play neuroprotective roles by preserving the Ca2+ balance and the integrity of the mitochondrial membranes [262].
5. Conclusion and Future Perspectives
In summary, the retina plays a vital role in processing visual information, with neurotransmitters and calcium dynamics being key factors in its function, development, and susceptibility to disease. This review highlighted the roles of the major retinal neurotransmitters, glutamate, GABA, glycine, dopamine, and ACH, shedding light on their functions and interactions and offering valuable insights for future research directions and the development of novel therapeutic agents to treat retinal diseases. Nevertheless, there remain major gaps in our understanding of the precise mechanisms and interactions in the retina, particularly concerning the interplay between neurotransmitters and Ca2+. For instance, anti-epileptic drugs used to treat GABA-related retinal ischemia are often associated with reduced Ca2+ currents through T-type channels, little is known about the Ca2+-dependent mechanisms of these drugs in relation to GABA. Also, although glycine, dopamine, and Ca2+ are both important for different aspects of development and retinal disease, little research has addressed the dynamics of the interplay between each individual neurotransmitter and Ca2+. Another unclear understanding of the interplay between neurotransmitters and Ca2+ is observed with acetylcholine. nAChRs allow for the entry of Na+ and Ca2+, and the activation of nAChRs mediates the strength of the peak and inward current responses of retinal cells, but the purpose of these interactions remains poorly understood. Addressing such knowledge gaps will be crucial for future investigations. Exploring the early patterns and interactions of the neurotransmitters discussed in this review with Ca2+ during development and early life stages will contribute to an enhanced understanding of normal retinal functioning. Such knowledge will allow for the deeper exploration of early disease detection and advancement of various visual impairments.
Author Contributions
Conceptualization, J.M.B., A.P.S., S.M., N.V.., L.D.S. and T.V.; writing—original draft preparation, T.V., J.M.B.; writing—review and editing, J.M.B., A.P.S., S.M., N.V.., L.D.S. and T.V.; visualization, J.M.B., and T.V; supervision, J.M.B., A.P.S., S.M., N.V.., L.D.S. and T.V.; project administration, L.D.S., and T.V.; funding acquisition, L.D.S., and T.V.; All authors have read and agreed to the published version of the manuscript.
Funding
Research reported in this publication was supported by the National Eye Institute of the National Institutes of Health under Award Number R01EY030863 and UTHSC College of Medicine Faculty Research Growth Award to T.V and R21-EY032567 to LDS and P30-EY007551 to University of Houston College of Optometry.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
The authors would like to thank Dr. Kyle Johnson Moore in the UTHSC Office of Scientific Writing for the critical review and editing of the manuscript.
Conflicts of Interest
“The authors declare no conflicts of interest.” Authors must identify and declare any personal circumstances or interest that may be perceived as inappropriately influencing the representation or interpretation of reported research results. Any role of the funders in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results must be declared in this section. If there is no role, please state “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Masland Richard, H. The Neuronal Organization of the Retina. Neuron. 2012, 76, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Myers CE, Klein BE, Meuer SM. Retinal thickness measured by spectral-domain optical coherence tomography in eyes without retinal abnormalities: the Beaver Dam Eye Study. Am J Ophthalmol. 2015, 159, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Muraoka Y, Ikeda HO, Nakano N, Hangai M, Toda Y, Okamoto-Furuta K, Kohda H, Kondo M, Terasaki H, Kakizuka A, et al. Real-Time Imaging of Rabbit Retina with Retinal Degeneration by Using Spectral-Domain Optical Coherence Tomography. PLOS ONE. 2012, 7, e36135. [Google Scholar] [CrossRef]
- Quint WH, Tadema KCD, Crins JHC, Kokke N, Meester-Smoor MA, Willemsen R, Klaver CCW, Iglesias AI. Zebrafish: An In Vivo Screening Model to Study Ocular Phenotypes. Transl Vis Sci Technol. 2022, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Guo L, Normando E, Nizari S, Lara D, Cordeiro MF. Tracking Longitudinal Retinal Changes in Experimental Ocular Hypertension Using the cSLO and Spectral Domain-OCT. Investigative ophthalmology & visual science. 2010, 51, 6504–6513. [Google Scholar] [CrossRef]
- Pycock, CJ. Retinal neurotransmission. Survey of Ophthalmology. 1985, 29, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Wu SM, Maple BR. Amino acid neurotransmitters in the retina: a functional overview. Vision Research. 1998, 38, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Brockie PJ, Madsen DM, Zheng Y, Mellem J, Maricq AV. Differential expression of glutamate receptor subunits in the nervous system of Caenorhabditis elegans and their regulation by the homeodomain protein UNC-42. J Neurosci. 2001, 21, 1510–1522. [Google Scholar] [CrossRef]
- Fedorovich SV, Waseem TV, Lavrukevich TV, Konev SV. Role of Calcium in Exocytosis Induced by Hypotonic Swelling. Annals of the New York Academy of Sciences. 2005, 1048, 337–340. [Google Scholar] [CrossRef]
- Cerella C, Diederich M, Ghibelli L. The dual role of calcium as messenger and stressor in cell damage, death, and survival. Int J Cell Biol. 2010, 2010, 546163. [Google Scholar] [CrossRef]
- Vaithianathan T, Henry D, Akmentin W, Matthews G. Nanoscale dynamics of synaptic vesicle trafficking and fusion at the presynaptic active zone. eLife. 2016, 5, e13245. [Google Scholar] [CrossRef]
- Mennerick S, Matthews G. Ultrafast exocytosis elicited by calcium current in synaptic terminals of retinal bipolar neurons. Neuron. 1996, 17, 1241–1249. [Google Scholar] [CrossRef]
- Dong F, Zhi Z, Pan M, Xie R, Qin X, Lu R, Mao X, Chen JF, Willcox MD, Qu J, et al. Inhibition of experimental myopia by a dopamine agonist: different effectiveness between form deprivation and hyperopic defocus in guinea pigs. Mol Vis. 2011, 17, 2824–2834. [Google Scholar]
- Esteve-Rudd J, Fernández-Sánchez L, Lax P, De Juan E, Martín-Nieto J, Cuenca N. Rotenone induces degeneration of photoreceptors and impairs the dopaminergic system in the rat retina. Neurobiol Dis. 2011, 44, 102–115. [Google Scholar] [CrossRef]
- Ishikawa A, Ishiguro S, Tamai M. Changes in GABA metabolism in streptozotocin-induced diabetic rat retinas. Curr Eye Res. 1996, 15, 63–71. [Google Scholar] [CrossRef]
- Napper GA, Pianta MJ, Kalloniatis M. Localization of amino acid neurotransmitters following in vitro ischemia and anoxia in the rat retina. Vis Neurosci. 2001, 18, 413–427. [Google Scholar] [CrossRef]
- Ortuño-Lizarán I, Sánchez-Sáez X, Lax P, Serrano GE, Beach TG, Adler CH, Cuenca N. Dopaminergic Retinal Cell Loss and Visual Dysfunction in Parkinson Disease. Ann Neurol. 2020, 88, 893–906. [Google Scholar] [CrossRef]
- Pisani F, Costa C, Caccamo D, Mazzon E, Gorgone G, Oteri G, Calabresi P. Tiagabine and vigabatrin reduce the severity of NMDA-induced excitotoxicity in chick retina. Experimental brain research. 2006, 171, 511–515. [Google Scholar] [CrossRef]
- Rosenlund, BL. Effects of insulin on free amino acids in plasma and the role of the amino acid metabolism in the etiology of diabetic microangiopathy. Biochem Med Metab Biol. 1993, 49, 375–391. [Google Scholar] [CrossRef]
- Stone RA, Lin T, Laties AM, Iuvone PM. Retinal dopamine and form-deprivation myopia. Proc Natl Acad Sci U S A. 1989, 86, 704–706. [Google Scholar] [CrossRef]
- van Dijk HW, Verbraak FD, Kok PH, Stehouwer M, Garvin MK, Sonka M, DeVries JH, Schlingemann RO, Abràmoff MD. Early neurodegeneration in the retina of type 2 diabetic patients. Invest Ophthalmol Vis Sci. 2012, 53, 2715–2719. [Google Scholar] [CrossRef]
- Amini E, Moghaddasi M, Habibi SAH, Azad Z, Miri S, Nilforushan N, Mirshahi R, Cubo E, Mohammadzadeh N, Rohani M. Huntington's disease and neurovascular structure of retina. Neurol Sci. 2022, 43, 5933–5941. [Google Scholar] [CrossRef]
- Zhang Y, Zhang X, Yue Y, Tian T. Retinal Degeneration: A Window to Understand the Origin and Progression of Parkinson's Disease? Front Neurosci. 2021, 15, 799526. [Google Scholar] [CrossRef]
- Liao C, Xu J, Chen Y. Retinal Dysfunction in Alzheimer's Disease and Implications for Biomarkers. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Marchesi N, Fahmideh F, Boschi F. Ocular Neurodegenerative Diseases: Interconnection between Retina and Cortical Areas. Cells 2021, 10. [Google Scholar] [CrossRef]
- Wang YX, Pan Z, Xue CC, Xie H, Wu X, Jonas JB. Macular outer nuclear layer, ellipsoid zone and outer photoreceptor segment band thickness, axial length and other determinants. Scientific Reports. 2023, 13, 5386. [Google Scholar] [CrossRef]
- Kawamura S, Tachibanaki S. Rod and cone photoreceptors: Molecular basis of the difference in their physiology. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology. 2008, 150, 369–377. [Google Scholar] [CrossRef]
- Thoreson WB, Mangel SC. Lateral interactions in the outer retina. Progress in Retinal and Eye Research. 2012, 31, 407–441. [Google Scholar] [CrossRef]
- Martemyanov KA, Sampath AP. The Transduction Cascade in Retinal ON-Bipolar Cells: Signal Processing and Disease. Annu Rev Vis Sci. 2017, 3, 25–51. [Google Scholar] [CrossRef]
- Euler T, Haverkamp S, Schubert T, Baden T. Retinal bipolar cells: elementary building blocks of vision. Nature Reviews Neuroscience. 2014, 15, 507–519. [Google Scholar] [CrossRef]
- Grimes WN, Aytürk DG, Hoon M, Yoshimatsu T, Gamlin C, Carrera D, Nath A, Nadal-Nicolás FM, Ahlquist RM, Sabnis A, et al. A high-density narrow-field inhibitory retinal interneuron with direct coupling to Müller glia. J Neurosci. 2021, 41, 6018–6037. [Google Scholar] [CrossRef]
- Yu DY, Cringle SJ, Balaratnasingam C, Morgan WH, Yu PK, Su EN. Retinal ganglion cells: Energetics, compartmentation, axonal transport, cytoskeletons and vulnerability. Prog Retin Eye Res. 2013, 36, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach A, Bringmann A. New functions of Müller cells. Glia. 2013, 61, 651–678. [Google Scholar] [CrossRef] [PubMed]
- Christensen I, Lu B, Yang N, Huang K, Wang P, Tian N. The Susceptibility of Retinal Ganglion Cells to Glutamatergic Excitotoxicity Is Type-Specific. Front Neurosci. 2019, 13, 219. [Google Scholar] [CrossRef] [PubMed]
- Zhang F, Kurokawa K, Lassoued A, Crowell JA, Miller DT. Cone photoreceptor classification in the living human eye from photostimulation-induced phase dynamics. Proceedings of the National Academy of Sciences. 2019, 116, 7951–7956. [Google Scholar] [CrossRef] [PubMed]
- Park, PS. Constitutively active rhodopsin and retinal disease. Adv Pharmacol. 2014, 70, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Connaughton VP, Graham D, Nelson R. Identification and morphological classification of horizontal, bipolar, and amacrine cells within the zebrafish retina. J Comp Neurol. 2004, 477, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Song PI, Matsui JI, Dowling JE. Morphological types and connectivity of horizontal cells found in the adult zebrafish (Danio rerio) retina. J Comp Neurol. 2008, 506, 328–338. [Google Scholar] [CrossRef]
- Bertalmío, M. Chapter 2 - The biological basis of vision: the retina. In Vision Models for High Dynamic Range and Wide Colour Gamut Imaging; Bertalmío M, ed, Ed.; Academic Press, 2020; pp. 11–46. [Google Scholar]
- Matthews, GG. Neurobiology: Molecules, Cells and Systems; Wiley, 2000. [Google Scholar]
- Balasubramanian R, Gan L. Development of Retinal Amacrine Cells and Their Dendritic Stratification. Curr Ophthalmol Rep. 2014, 2, 100–106. [Google Scholar] [CrossRef]
- Werblin, FS. Regenerative hyperpolarization in rods. The Journal of Physiology. 1975, 244, 53–81. [Google Scholar] [CrossRef]
- Oakley B, 2nd, Flaming DG, Brown KT. Effects of the rod receptor potential upon retinal extracellular potassium concentration. Journal of General Physiology. 1979, 74, 713–737. [Google Scholar] [CrossRef]
- Nakatani K, Yau KW. Calcium and magnesium fluxes across the plasma membrane of the toad rod outer segment. The Journal of Physiology. 1988, 395, 695–729. [Google Scholar] [CrossRef] [PubMed]
- Euler T, Masland RH. Light-Evoked Responses of Bipolar Cells in a Mammalian Retina. Journal of Neurophysiology. 2000, 83, 1817–1829. [Google Scholar] [CrossRef]
- Baden T, Berens P, Franke K, Román Rosón M, Bethge M, Euler T. The functional diversity of retinal ganglion cells in the mouse. Nature. 2016, 529, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Field GD, Gauthier JL, Sher A, Greschner M, Machado TA, Jepson LH, Shlens J, Gunning DE, Mathieson K, Dabrowski W, et al. Functional connectivity in the retina at the resolution of photoreceptors. Nature. 2010, 467, 673–677. [Google Scholar] [CrossRef]
- La Vail MM, Rapaport DH, Rakic P. Cytogenesis in the monkey retina. J Comp Neurol. 1991, 309, 86–114. [Google Scholar] [CrossRef] [PubMed]
- Carter-Dawson LD, LaVail MM. Rods and cones in the mouse retina. I. Structural analysis using light and electron microscopy. J Comp Neurol. 1979, 188, 245–262. [Google Scholar] [CrossRef]
- Haverkamp S, Wässle H, Duebel J, Kuner T, Augustine GJ, Feng G, Euler T. The primordial, blue-cone color system of the mouse retina. J Neurosci. 2005, 25, 5438–5445. [Google Scholar] [CrossRef]
- Nikonov SS, Kholodenko R, Lem J, Pugh EN, Jr. Physiological features of the S- and M-cone photoreceptors of wild-type mice from single-cell recordings. J Gen Physiol. 2006, 127, 359–374. [Google Scholar] [CrossRef]
- Wikler KC, Williams RW, Rakic P. Photoreceptor mosaic: number and distribution of rods and cones in the rhesus monkey retina. J Comp Neurol. 1990, 297, 499–508. [Google Scholar] [CrossRef]
- Wässle H, Boycott BB. Functional architecture of the mammalian retina. Physiol Rev. 1991, 71, 447–480. [Google Scholar] [CrossRef] [PubMed]
- Wässle, H. Parallel processing in the mammalian retina. Nat Rev Neurosci. 2004, 5, 747–757. [Google Scholar] [CrossRef]
- Roorda A, Metha AB, Lennie P, Williams DR. Packing arrangement of the three cone classes in primate retina. Vision Res. 2001, 41, 1291–1306. [Google Scholar] [CrossRef] [PubMed]
- Dowling JE, Wells GP. Synaptic organization of the frog retina: an electron microscopic analysis comparing the retinas of frogs and primates. Proceedings of the Royal Society of London Series B Biological Sciences. 1997, 170, 205–228. [Google Scholar] [CrossRef]
- Sterling P, Matthews G. Structure and function of ribbon synapses. Trends Neurosci. 2005, 28, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zenisek D, Horst NK, Merrifield C, Sterling P, Matthews G. Visualizing synaptic ribbons in the living cell. J Neurosci. 2004, 24, 9752–9759. [Google Scholar] [CrossRef]
- Vaithianathan T, Matthews G. Visualizing synaptic vesicle turnover and pool refilling driven by calcium nanodomains at presynaptic active zones of ribbon synapses. Proceedings of the National Academy of Sciences. 2014, 111, 8655–8660. [Google Scholar] [CrossRef]
- Heidelberger R, Wang MM, Sherry DM. Differential distribution of synaptotagmin immunoreactivity among synapses in the goldfish, salamander, and mouse retina. Vis Neurosci. 2003, 20, 37–49. [Google Scholar] [CrossRef]
- Henrique von G, Gary M. Depletion and Replenishment of Vesicle Pools at a Ribbon-Type Synaptic Terminal. The Journal of Neuroscience. 1997, 17, 1919. [Google Scholar] [CrossRef]
- von Gersdorff H, Vardi E, Matthews G, Sterling P. Evidence That Vesicles on the Synaptic Ribbon of Retinal Bipolar Neurons Can Be Rapidly Released. Neuron. 1996, 16, 1221–1227. [Google Scholar] [CrossRef]
- Gary M, Peter S. Evidence That Vesicles Undergo Compound Fusion on the Synaptic Ribbon. The Journal of Neuroscience. 2008, 28, 5403. [Google Scholar] [CrossRef]
- Ling-Gang W, Timothy AR, Leon L. Modes of Vesicle Retrieval at Ribbon Synapses, Calyx-Type Synapses, and Small Central Synapses. The Journal of Neuroscience. 2007, 27, 11793. [Google Scholar] [CrossRef]
- Cho S, von Gersdorff H. Ca(2+) influx and neurotransmitter release at ribbon synapses. Cell Calcium. 2012, 52, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Coggins M, Zenisek D. Evidence that exocytosis is driven by calcium entry through multiple calcium channels in goldfish retinal bipolar cells. J Neurophysiol. 2009, 101, 2601–2619. [Google Scholar] [CrossRef] [PubMed]
- Zenisek D, Davila V, Wan L, Almers W. Imaging calcium entry sites and ribbon structures in two presynaptic cells. J Neurosci. 2003, 23, 2538–2548. [Google Scholar] [CrossRef] [PubMed]
- Pangrsic T, Singer JH, Koschak A. Voltage-Gated Calcium Channels: Key Players in Sensory Coding in the Retina and the Inner Ear. Physiological Reviews. 2018, 98, 2063–2096. [Google Scholar] [CrossRef] [PubMed]
- Kushner J, Ferrer X, Marx SO. Roles and Regulation of Voltage-gated Calcium Channels in Arrhythmias. J Innov Card Rhythm Manag. 2019, 10, 3874–3880. [Google Scholar] [CrossRef]
- Fox AP, Nowycky MC, Tsien RW. Kinetic and pharmacological properties distinguishing three types of calcium currents in chick sensory neurones. The Journal of Physiology. 1987, 394, 149–172. [Google Scholar] [CrossRef] [PubMed]
- Neelands TR, King APJ, Macdonald RL. Functional Expression of L-, N-, P/Q-, and R-Type Calcium Channels in the Human NT2-N Cell Line. Journal of Neurophysiology. 2000, 84, 2933–2944. [Google Scholar] [CrossRef] [PubMed]
- Griguoli M, Sgritta M, Cherubini E. Presynaptic BK channels control transmitter release: physiological relevance and potential therapeutic implications. J Physiol. 2016, 594, 3489–3500. [Google Scholar] [CrossRef]
- Wang, ZW. Regulation of synaptic transmission by presynaptic CaMKII and BK channels. Mol Neurobiol. 2008, 38, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Robitaille R, Charlton MP. Presynaptic calcium signals and transmitter release are modulated by calcium-activated potassium channels. J Neurosci. 1992, 12, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Morita K, Barrett EF. Evidence for two calcium-dependent potassium conductances in lizard motor nerve terminals. J Neurosci. 1990, 10, 2614–2625. [Google Scholar] [CrossRef] [PubMed]
- Grimes WN, Li W, Chávez AE, Diamond JS. BK channels modulate pre- and postsynaptic signaling at reciprocal synapses in retina. Nat Neurosci. 2009, 12, 585–592. [Google Scholar] [CrossRef]
- Jian Wei X, Malcolm MS. Large-Conductance Calcium-Activated Potassium Channels Facilitate Transmitter Release in Salamander Rod Synapse. The Journal of Neuroscience. 2005, 25, 7660. [Google Scholar] [CrossRef]
- Naoyuki T, Vithiyanjali S, Thomas E, Peter R, Mathias WS, Timm S. BK Channels Mediate Pathway-Specific Modulation of Visual Signals in the <em>In Vivo</em> Mouse Retina. The Journal of Neuroscience. 2012, 32, 4861. [Google Scholar] [CrossRef]
- Van Cruchten S, Vrolyk V, Perron Lepage M-F, Baudon M, Voute H, Schoofs S, Haruna J, Benoit-Biancamano M-O, Ruot B, Allegaert K. Pre- and Postnatal Development of the Eye: A Species Comparison. Birth Defects Research. 2017, 109, 1540–1567. [Google Scholar] [CrossRef]
- Graw, J. Eye development. Curr Top Dev Biol. 2010, 90, 343–386. [Google Scholar] [CrossRef]
- Harada T, Harada C, Parada LF. Molecular regulation of visual system development: more than meets the eye. Genes Dev. 2007, 21, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Marquardt T, Gruss P. Generating neuronal diversity in the retina: one for nearly all. Trends Neurosci. 2002, 25, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Cantrup R, Kaushik G, Schuurmans C. Control of Retinal Development by Tumor Suppressor Genes. In Tumor Suppressor Genes; Cheng Y, ed, Ed.; IntechOpen: London, U.K., 2012. [Google Scholar]
- Turner DL, Snyder EY, Cepko CL. Lineage-independent determination of cell type in the embryonic mouse retina. Neuron. 1990, 4, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Wetts R, Fraser SE. Multipotent Precursors Can Give Rise to All Major Cell Types of the Frog Retina. Science. 1988, 239, 1142–1145. [Google Scholar] [CrossRef]
- Holt CE, Bertsch TW, Ellis HM, Harris WA. Cellular determination in the Xenopus retina is independent of lineage and birth date. Neuron. 1988, 1, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Turner DL, Cepko CL. A common progenitor for neurons and glia persists in rat retina late in development. Nature. 1987, 328, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Hernan G, de Mera-Rodríguez JA, Gañán Y, Solana-Fajardo J, Martín-Partido G, Rodríguez-León J, Francisco-Morcillo J. Development and postnatal neurogenesis in the retina: a comparison between altricial and precocial bird species. Neural Regen Res. 2021, 16, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Finlayson PG, Iezzi R. Glutamate stimulation of retinal ganglion cells in normal and s334ter-4 rat retinas: a candidate for a neurotransmitter-based retinal prosthesis. Invest Ophthalmol Vis Sci. 2010, 51, 3619–3628. [Google Scholar] [CrossRef] [PubMed]
- Lee S, Chen L, Chen M, Ye M, Seal RP, Zhou ZJ. An unconventional glutamatergic circuit in the retina formed by vGluT3 amacrine cells. Neuron. 2014, 84, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Ruan G-X, Allen GC, Yamazaki S, McMahon DG. An Autonomous Circadian Clock in the Inner Mouse Retina Regulated by Dopamine and GABA. PLOS Biology. 2008, 6, e249. [Google Scholar] [CrossRef]
- Hartwick AT, Hamilton CM, Baldridge WH. Glutamatergic calcium dynamics and deregulation of rat retinal ganglion cells. J Physiol. 2008, 586, 3425–3446. [Google Scholar] [CrossRef]
- Roth, BL. Molecular pharmacology of metabotropic receptors targeted by neuropsychiatric drugs. Nat Struct Mol Biol. 2019, 26, 535–544. [Google Scholar] [CrossRef]
- Wong, RO. Retinal waves and visual system development. Annu Rev Neurosci. 1999, 22, 29–47. [Google Scholar] [CrossRef]
- Bodnarenko SR, Chalupa LM. Stratification of ON and OFF ganglion cell dendrites depends on glutamate-mediated afferent activity in the developing retina. Nature. 1993, 364, 144–146. [Google Scholar] [CrossRef]
- Bodnarenko SR, Jeyarasasingam G, Chalupa LM. Development and regulation of dendritic stratification in retinal ganglion cells by glutamate-mediated afferent activity. J Neurosci. 1995, 15, 7037–7045. [Google Scholar] [CrossRef] [PubMed]
- Galli L, Maffei L. Spontaneous impulse activity of rat retinal ganglion cells in prenatal life. Science. 1988, 242, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang RW, Du WJ, Prober DA, Du JL. Müller Glial Cells Participate in Retinal Waves via Glutamate Transporters and AMPA Receptors. Cell Rep. 2019, 27, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki T, Yamasaki M, Hashimoto K, Kohda K, Yuzaki M, Shimamoto K, Tanaka K, Kano M, Watanabe M. Glutamate transporter GLAST controls synaptic wrapping by Bergmann glia and ensures proper wiring of Purkinje cells. Proc Natl Acad Sci U S A. 2017, 114, 7438–7443. [Google Scholar] [CrossRef]
- Blankenship AG, Ford KJ, Johnson J, Seal RP, Edwards RH, Copenhagen DR, Feller MB. Synaptic and extrasynaptic factors governing glutamatergic retinal waves. Neuron. 2009, 62, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Xu H-P, Burbridge TJ, Ye M, Chen M, Ge X, Zhou ZJ, Crair MC. Retinal Wave Patterns Are Governed by Mutual Excitation among Starburst Amacrine Cells and Drive the Refinement and Maintenance of Visual Circuits. The Journal of Neuroscience. 2016, 36, 3871–3886. [Google Scholar] [CrossRef] [PubMed]
- Liets LC, Chalupa LM. Glutamate-mediated responses in developing retinal ganglion cells. Prog Brain Res. 2001, 134, 1–16. [Google Scholar] [CrossRef]
- Chang Y-C, Chen C-Y, Chiao C-C. Visual Experience–Independent Functional Expression of NMDA Receptors in the Developing Rabbit Retina. Investigative Ophthalmology & Visual Science. 2010, 51, 2744–2754. [Google Scholar] [CrossRef]
- Gründer T, Kohler K, Kaletta A, Guenther E. The distribution and developmental regulation of NMDA receptor subunit proteins in the outer and inner retina of the rat. J Neurobiol. 2000, 44, 333–342. [Google Scholar] [CrossRef]
- Hartveit E, Brandstätter JH, Sassoè-Pognetto M, Laurie DJ, Seeburg PH, Wässle H. Localization and developmental expression of the NMDA receptor subunit NR2A in the mammalian retina. J Comp Neurol. 1994, 348, 570–582. [Google Scholar] [CrossRef]
- Pourcho RG, Qin P, Goebel DJ. Cellular and subcellular distribution of NMDA receptor subunit NR2B in the retina. J Comp Neurol. 2001, 433, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Popova, E. ON-OFF Interactions in the Retina: Role of Glycine and GABA. Curr Neuropharmacol. 2014, 12, 509–526. [Google Scholar] [CrossRef]
- Mehta B, Ke J-B, Zhang L, Baden AD, Markowitz AL, Nayak S, Briggman KL, Zenisek D, Singer JH. Global Ca2+ Signaling Drives Ribbon-Independent Synaptic Transmission at Rod Bipolar Cell Synapses. The Journal of Neuroscience. 2014, 34, 6233–6244. [Google Scholar] [CrossRef] [PubMed]
- Mehta B, Snellman J, Chen S, Li W, Zenisek D. Synaptic ribbons influence the size and frequency of miniature-like evoked postsynaptic currents. Neuron. 2013, 77, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Thoreson, WB. Transmission at rod and cone ribbon synapses in the retina. Pflugers Arch. 2021, 473, 1469–1491. [Google Scholar] [CrossRef]
- Werblin FS, Dowling JE. Organization of the retina of the mudpuppy, Necturus maculosus. II. Intracellular recording. J Neurophysiol. 1969, 32, 339–355. [Google Scholar] [CrossRef]
- Kaneko, A. Physiological and morphological identification of horizontal, bipolar and amacrine cells in goldfish retina. J Physiol. 1970, 207, 623–633. [Google Scholar] [CrossRef]
- DeVries SH, Schwartz EA. Kainate receptors mediate synaptic transmission between cones and 'Off' bipolar cells in a mammalian retina. Nature. 1999, 397, 157–160. [Google Scholar] [CrossRef]
- Zhang J, Diamond JS. Distinct perisynaptic and synaptic localization of NMDA and AMPA receptors on ganglion cells in rat retina. J Comp Neurol. 2006, 498, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Gustafson EC, Morgans CW, Tekmen M, Sullivan SJ, Esguerra M, Konno R, Miller RF. Retinal NMDA receptor function and expression are altered in a mouse lacking D-amino acid oxidase. J Neurophysiol. 2013, 110, 2718–2726. [Google Scholar] [CrossRef] [PubMed]
- Dhingra A, Vardi N. "mGlu Receptors in the Retina" - WIREs Membrane Transport and Signaling. Wiley Interdiscip Rev Membr Transp Signal. 2012, 1, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Nawy, S. The metabotropic receptor mGluR6 may signal through G(o), but not phosphodiesterase, in retinal bipolar cells. J Neurosci. 1999, 19, 2938–2944. [Google Scholar] [CrossRef]
- Shen Y, Heimel JA, Kamermans M, Peachey NS, Gregg RG, Nawy S. A transient receptor potential-like channel mediates synaptic transmission in rod bipolar cells. J Neurosci. 2009, 29, 6088–6093. [Google Scholar] [CrossRef]
- Morgans CW, Brown RL, Duvoisin RM. TRPM1, the endpoint of the mGluR6 signal transduction cascade in retinal ON-bipolar cells. Bioessays. 2010, 32, 609–614. [Google Scholar] [CrossRef]
- Lie-Venema H, Hakvoort TB, van Hemert FJ, Moorman AF, Lamers WH. Regulation of the spatiotemporal pattern of expression of the glutamine synthetase gene. Prog Nucleic Acid Res Mol Biol. 1998, 61, 243–308. [Google Scholar] [CrossRef]
- Novelli A, Reilly JA, Lysko PG, Henneberry RC. Glutamate becomes neurotoxic via the N-methyl-D-aspartate receptor when intracellular energy levels are reduced. Brain Res. 1988, 451, 205–212. [Google Scholar] [CrossRef]
- Nguyen D, Alavi MV, Kim KY, Kang T, Scott RT, Noh YH, Lindsey JD, Wissinger B, Ellisman MH, Weinreb RN, et al. A new vicious cycle involving glutamate excitotoxicity, oxidative stress and mitochondrial dynamics. Cell Death Dis. 2011, 2, e240. [Google Scholar] [CrossRef]
- Peichl, L. Alpha ganglion cells in mammalian retinae: common properties, species differences, and some comments on other ganglion cells. Vis Neurosci. 1991, 7, 155–169. [Google Scholar] [CrossRef]
- Sanes JR, Masland RH. The Types of Retinal Ganglion Cells: Current Status and Implications for Neuronal Classification. Annual Review of Neuroscience. 2015, 38, 221–246. [Google Scholar] [CrossRef]
- Lucas DR, Newhouse JP. The toxic effect of sodium L-glutamate on the inner layers of the retina. AMA Arch Ophthalmol. 1957, 58, 193–201. [Google Scholar] [CrossRef]
- Izumi Y, Shimamoto K, Benz AM, Hammerman SB, Olney JW, Zorumski CF. Glutamate transporters and retinal excitotoxicity. Glia. 2002, 39, 58–68. [Google Scholar] [CrossRef]
- Shimamoto K, Lebrun B, Yasuda-Kamatani Y, Sakaitani M, Shigeri Y, Yumoto N, Nakajima T. DL-threo-beta-benzyloxyaspartate, a potent blocker of excitatory amino acid transporters. Mol Pharmacol. 1998, 53, 195–201. [Google Scholar] [CrossRef]
- Wasilewa P, Hockwin O, Korte I. Glycogen concentration changes in retina, vitreous body and other eye tissues caused by disturbances of blood circulation. Albrecht Von Graefes Arch Klin Exp Ophthalmol. 1976, 199, 115–120. [Google Scholar] [CrossRef]
- Kosenko E, Llansola M, Montoliu C, Monfort P, Rodrigo R, Hernandez-Viadel M, Erceg S, Sánchez-Perez AM, Felipo V. Glutamine synthetase activity and glutamine content in brain: modulation by NMDA receptors and nitric oxide. Neurochem Int. 2003, 43, 493–499. [Google Scholar] [CrossRef]
- Fernandez DC, Chianelli MS, Rosenstein RE. Involvement of glutamate in retinal protection against ischemia/reperfusion damage induced by post-conditioning. Journal of Neurochemistry. 2009, 111, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Osborne NN, Casson RJ, Wood JP, Chidlow G, Graham M, Melena J. Retinal ischemia: mechanisms of damage and potential therapeutic strategies. Prog Retin Eye Res. 2004, 23, 91–147. [Google Scholar] [CrossRef] [PubMed]
- Barnett NL, Pow DV, Bull ND. Differential perturbation of neuronal and glial glutamate transport systems in retinal ischaemia. Neurochem Int. 2001, 39, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Liu Y, Wang C, Su G. Cellular Signaling in Müller Glia: Progenitor Cells for Regenerative and Neuroprotective Responses in Pharmacological Models of Retinal Degeneration. J Ophthalmol. 2019, 2019, 5743109. [Google Scholar] [CrossRef]
- Bringmann A, Uckermann O, Pannicke T, Iandiev I, Reichenbach A, Wiedemann P. Neuronal versus glial cell swelling in the ischaemic retina. Acta Ophthalmol Scand. 2005, 83, 528–538. [Google Scholar] [CrossRef]
- Barnett NL, Grozdanic SD. Glutamate transporter localization does not correspond to the temporary functional recovery and late degeneration after acute ocular ischemia in rats. Exp Eye Res. 2004, 79, 513–524. [Google Scholar] [CrossRef]
- Casson, RJ. Possible role of excitotoxicity in the pathogenesis of glaucoma. Clin Exp Ophthalmol. 2006, 34, 54–63. [Google Scholar] [CrossRef]
- Naskar R, Vorwerk CK, Dreyer EB. Concurrent downregulation of a glutamate transporter and receptor in glaucoma. Invest Ophthalmol Vis Sci. 2000, 41, 1940–1944. [Google Scholar]
- Harada T, Harada C, Nakamura K, Quah HM, Okumura A, Namekata K, Saeki T, Aihara M, Yoshida H, Mitani A, et al. The potential role of glutamate transporters in the pathogenesis of normal tension glaucoma. J Clin Invest. 2007, 117, 1763–1770. [Google Scholar] [CrossRef]
- Harada C, Nakamura K, Namekata K, Okumura A, Mitamura Y, Iizuka Y, Kashiwagi K, Yoshida K, Ohno S, Matsuzawa A, et al. Role of apoptosis signal-regulating kinase 1 in stress-induced neural cell apoptosis in vivo. Am J Pathol. 2006, 168, 261–269. [Google Scholar] [CrossRef]
- Osaka N, Takahashi T, Murakami S, Matsuzawa A, Noguchi T, Fujiwara T, Aburatani H, Moriyama K, Takeda K, Ichijo H. ASK1-dependent recruitment and activation of macrophages induce hair growth in skin wounds. J Cell Biol. 2007, 176, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Ambati J, Chalam KV, Chawla DK, D'Angio CT, Guillet EG, Rose SJ, Vanderlinde RE, Ambati BK. Elevated gamma-aminobutyric acid, glutamate, and vascular endothelial growth factor levels in the vitreous of patients with proliferative diabetic retinopathy. Arch Ophthalmol. 1997, 115, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Hernández C, Simó R. Neuroprotection in diabetic retinopathy. Curr Diab Rep. 2012, 12, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Villagrana AR, Garcia-Román J, González-Espinosa C, Lamas M. Pharmacological inhibition of N-methyl d-aspartate receptor promotes secretion of vascular endothelial growth factor in müller cells: effects of hyperglycemia and hypoxia. Curr Eye Res. 2010, 35, 733–741. [Google Scholar] [CrossRef]
- Kusari J, Zhou SX, Padillo E, Clarke KG, Gil DW. Inhibition of Vitreoretinal VEGF Elevation and Blood–Retinal Barrier Breakdown in Streptozotocin-Induced Diabetic Rats by Brimonidine. Investigative Ophthalmology & Visual Science. 2010, 51, 1044–1051. [Google Scholar] [CrossRef]
- Li Q, Puro DG. Diabetes-induced dysfunction of the glutamate transporter in retinal Müller cells. Invest Ophthalmol Vis Sci. 2002, 43, 3109–3116. [Google Scholar]
- Zeng K, Xu H, Mi M, Zhang Q, Zhang Y, Chen K, Chen F, Zhu J, Yu X. Dietary taurine supplementation prevents glial alterations in retina of diabetic rats. Neurochem Res. 2009, 34, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Paradies G, Petrosillo G, Paradies V, Ruggiero FM. Role of cardiolipin peroxidation and Ca2+ in mitochondrial dysfunction and disease. Cell Calcium. 2009, 45, 643–650. [Google Scholar] [CrossRef]
- Aydın A, Orhan H, Sayal A, Özata M, Şahin G, Işımer A. Oxidative stress and nitric oxide related parameters in type II diabetes mellitus: effects of glycemic control. Clinical Biochemistry. 2001, 34, 65–70. [Google Scholar] [CrossRef]
- Dong LY, Jin J, Lu G, Kang XL. Astaxanthin attenuates the apoptosis of retinal ganglion cells in db/db mice by inhibition of oxidative stress. Mar Drugs. 2013, 11, 960–974. [Google Scholar] [CrossRef]
- Mittal R, Kumar A, Singh DP, Bishnoi M, Nag TC. Ameliorative potential of rutin in combination with nimesulide in STZ model of diabetic neuropathy: targeting Nrf2/HO-1/NF-kB and COX signalling pathway. Inflammopharmacology. 2018, 26, 755–768. [Google Scholar] [CrossRef]
- Xu Z, Wei Y, Gong J, Cho H, Park JK, Sung ER, Huang H, Wu L, Eberhart C, Handa JT, et al. NRF2 plays a protective role in diabetic retinopathy in mice. Diabetologia. 2014, 57, 204–213. [Google Scholar] [CrossRef]
- Firl A, Sack GS, Newman ZL, Tani H, Feller MB. Extrasynaptic glutamate and inhibitory neurotransmission modulate ganglion cell participation during glutamatergic retinal waves. Journal of Neurophysiology. 2013, 109, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Wong, RO. Effects of glutamate and its analogs on intracellular calcium levels in the developing retina. Vis Neurosci. 1995, 12, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Mesnard CS, Barta CL, Sladek AL, Zenisek D, Thoreson WB. Eliminating Synaptic Ribbons from Rods and Cones Halves the Releasable Vesicle Pool and Slows Down Replenishment. Eliminating Synaptic Ribbons from Rods and Cones Halves the Releasable Vesicle Pool and Slows Down Replenishment. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef]
- Frank T, Rutherford MA, Strenzke N, Neef A, Pangršič T, Khimich D, Fejtova A, Gundelfinger ED, Liberman MC, Harke B, et al. Bassoon and the synaptic ribbon organize Ca²+ channels and vesicles to add release sites and promote refilling. Neuron. 2010, 68, 724–738. [Google Scholar] [CrossRef] [PubMed]
- Chen M, Van Hook MJ, Zenisek D, Thoreson WB. Properties of ribbon and non-ribbon release from rod photoreceptors revealed by visualizing individual synaptic vesicles. J Neurosci. 2013, 33, 2071–2086. [Google Scholar] [CrossRef] [PubMed]
- Krizaj D, Lai FA, Copenhagen DR. Ryanodine stores and calcium regulation in the inner segments of salamander rods and cones. J Physiol. 2003, 547, 761–774. [Google Scholar] [CrossRef]
- Sattler R, Tymianski M. Molecular mechanisms of calcium-dependent excitotoxicity. Journal of Molecular Medicine. 2000, 78, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Smith, SB. Diabetic Retinopathy and the NMDA Receptor. Drug News Perspect. 2002, 15, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Wu X-S, Wu L-G. The Yin and Yang of Calcium Effects on Synaptic Vesicle Endocytosis. The Journal of Neuroscience. 2014, 34, 2652–2659. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp S, Wässle H. Immunocytochemical analysis of the mouse retina. J Comp Neurol. 2000, 424, 1–23. [Google Scholar] [CrossRef]
- Vaughn JE, Famiglietti EV, Barber Jr. RP, Saito K, Roberts E, Ribak CE. GABAergic amacrine cells in rat retina: Immunocytochemical identification and synaptic connectivity. Journal of Comparative Neurology. 1981, 197, 113–127. [Google Scholar] [CrossRef]
- Mosinger JL, Yazulla S, Studholme KM. GABA-like immunoreactivity in the vertebrate retina: a species comparison. Exp Eye Res. 1986, 42, 631–644. [Google Scholar] [CrossRef]
- Davanger S, Ottersen OP, Storm-Mathisen J. Glutamate, GABA, and glycine in the human retina: an immunocytochemical investigation. J Comp Neurol. 1991, 311, 483–494. [Google Scholar] [CrossRef]
- Agardh E, Ehinger B, Wu JY. GABA and GAD-like immunoreactivity in the primate retina. Histochemistry. 1987, 86, 485–490. [Google Scholar] [CrossRef]
- Yu BCY, Watt CB, Lam DMK, Fry KR. GABAergic ganglion cells in the rabbit retina. Brain Research. 1988, 439, 376–382. [Google Scholar] [CrossRef]
- Sandell, JH. GABA as a developmental signal in the inner retina and optic nerve. Perspect Dev Neurobiol. 1998, 5, 269–278. [Google Scholar] [PubMed]
- Shaye H, Stauch B, Gati C. Molecular mechanisms of metabotropic GABA(B) receptor function. Sci Adv 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Catsicas M, Mobbs P. GABAb receptors regulate chick retinal calcium waves. J Neurosci. 2001, 21, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer J, Rusoff AC. Horizontal cells of the mouse retina contain glutamic acid decarboxylase-like immunoreactivity during early developmental stages. J Neurosci. 1984, 4, 2948–2955. [Google Scholar] [CrossRef] [PubMed]
- Fischer KF, Lukasiewicz PD, Wong RO. Age-dependent and cell class-specific modulation of retinal ganglion cell bursting activity by GABA. J Neurosci. 1998, 18, 3767–3778. [Google Scholar] [CrossRef]
- Buddhala C, Hsu CC, Wu JY. A novel mechanism for GABA synthesis and packaging into synaptic vesicles. Neurochem Int. 2009, 55, 9–12. [Google Scholar] [CrossRef]
- Mikkelsen M, Harris AD, Edden RAE, Puts NAJ. Macromolecule-suppressed GABA measurements correlate more strongly with behavior than macromolecule-contaminated GABA+ measurements. Brain Res. 2018, 1701, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Hyland N, Cryan J. A Gut Feeling about GABA: Focus on GABAB Receptors. Frontiers in Pharmacology 2010, 1. [Google Scholar] [CrossRef] [PubMed]
- Macdonald RL, Olsen RW. GABAA receptor channels. Annu Rev Neurosci. 1994, 17, 569–602. [Google Scholar] [CrossRef] [PubMed]
- Bormann, J. Electrophysiology of GABAA and GABAB receptor subtypes. Trends in Neurosciences. 1988, 11, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Wang S, Du L, Peng G, Li W. GABA inhibits proliferation and self-renewal of mouse retinal progenitor cell. Cell Death Discovery. 2019, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Koulen P, Malitschek B, Kuhn R, Bettler B, Wässle H, Brandstätter JH. Presynaptic and postsynaptic localization of GABAB receptors in neurons of the rat retina. European Journal of Neuroscience. 1998, 10, 1446–1456. [Google Scholar] [CrossRef]
- Kamermans M, Werblin F. GABA-mediated positive autofeedback loop controls horizontal cell kinetics in tiger salamander retina. The Journal of Neuroscience. 1992, 12, 2451–2463. [Google Scholar] [CrossRef]
- Matthews G, Ayoub GS, Heidelberger R. Presynaptic inhibition by GABA is mediated via two distinct GABA receptors with novel pharmacology. J Neurosci. 1994, 14, 1079–1090. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, EA. Depolarization without calcium can release gamma-aminobutyric acid from a retinal neuron. Science. 1987, 238, 350–355. [Google Scholar] [CrossRef]
- Warrier A, Borges S, Dalcino D, Walters C, Wilson M. Calcium from internal stores triggers GABA release from retinal amacrine cells. J Neurophysiol. 2005, 94, 4196–4208. [Google Scholar] [CrossRef]
- Brecha NC, Weigmann C. Expression of GAT-1, a high-affinity gamma-aminobutyric acid plasma membrane transporter in the rat retina. J Comp Neurol. 1994, 345, 602–611. [Google Scholar] [CrossRef]
- Honda S, Yamamoto M, Saito N. Immunocytochemical localization of three subtypes of GABA transporter in rat retina. Molecular Brain Research. 1995, 33, 319–325. [Google Scholar] [CrossRef]
- Schousboe, A. Role of astrocytes in the maintenance and modulation of glutamatergic and GABAergic neurotransmission. Neurochem Res. 2003, 28, 347–352. [Google Scholar] [CrossRef]
- Moran J, Pasantes-Morales H, Redburn DA. Glutamate receptor agonists release [3H]GABA preferentially from horizontal cells. Brain Research. 1986, 398, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Marc, RE. Structural organization of GABAergic circuitry in ectotherm retinas. Prog Brain Res. 1992, 90, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Barnett NL, Osborne NN. Redistribution of GABA immunoreactivity following central retinal artery occlusion. Brain Research. 1995, 677, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Malmgren K, Ben-Menachem E, Frisén L. Vigabatrin Visual Toxicity: Evolution and Dose Dependence. Epilepsia. 2001, 42, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Biermann J, Grieshaber P, Goebel U, Martin G, Thanos S, Di Giovanni S, Lagrèze WA. Valproic acid-mediated neuroprotection and regeneration in injured retinal ganglion cells. Invest Ophthalmol Vis Sci. 2010, 51, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Schur RM, Gao S, Yu G, Chen Y, Maeda A, Palczewski K, Lu ZR. New GABA modulators protect photoreceptor cells from light-induced degeneration in mouse models. FASEB J. 2018, 32, 3289–3300. [Google Scholar] [CrossRef] [PubMed]
- Sernagor E, Young C, Eglen SJ. Developmental modulation of retinal wave dynamics: shedding light on the GABA saga. J Neurosci. 2003, 23, 7621–7629. [Google Scholar] [CrossRef]
- Eggers ED, Klein JS, Moore-Dotson JM. Slow changes in Ca2(+) cause prolonged release from GABAergic retinal amacrine cells. J Neurophysiol. 2013, 110, 709–719. [Google Scholar] [CrossRef]
- Hirano AA, Vuong HE, Kornmann HL, Schietroma C, Stella SL, Barnes S, Brecha NC. Vesicular Release of GABA by Mammalian Horizontal Cells Mediates Inhibitory Output to Photoreceptors. Vesicular Release of GABA by Mammalian Horizontal Cells Mediates Inhibitory Output to Photoreceptors. Frontiers in Cellular Neuroscience. 2020, 14. [Google Scholar] [CrossRef]
- Verweij J, Kamermans M, Spekreijse H. Horizontal cells feed back to cones by shifting the cone calcium-current activation range. Vision Res. 1996, 36, 3943–3953. [Google Scholar] [CrossRef]
- Moore-Dotson JM, Eggers ED. Reductions in Calcium Signaling Limit Inhibition to Diabetic Retinal Rod Bipolar Cells. Invest Ophthalmol Vis Sci. 2019, 60, 4063–4073. [Google Scholar] [CrossRef]
- Czapiński P, Blaszczyk B, Czuczwar SJ. Mechanisms of action of antiepileptic drugs. Curr Top Med Chem. 2005, 5, 3–14. [Google Scholar] [CrossRef]
- Chintalapudi SR, Maria D, Di Wang X, Bailey JNC, Allingham R, Brilliant M, Budenz D, Fingert J, Gaasterland D, Gaasterland T, et al. Systems genetics identifies a role for Cacna2d1 regulation in elevated intraocular pressure and glaucoma susceptibility. Nature Communications. 2017, 8, 1755. [Google Scholar] [CrossRef] [PubMed]
- Pow DV, Barnett NL. Changing patterns of spatial buffering of glutamate in developing rat retinae are mediated by the Müller cell glutamate transporter GLAST. Cell Tissue Res. 1999, 297, 57–66. [Google Scholar] [CrossRef]
- Sassoe-Pognetto M, Wassle H, Grunert U. Glycinergic synapses in the rod pathway of the rat retina: cone bipolar cells express the alpha 1 subunit of the glycine receptor. The Journal of Neuroscience. 1994, 14, 5131–5146. [Google Scholar] [CrossRef]
- Lynch, JW. Native glycine receptor subtypes and their physiological roles. Neuropharmacology. 2009, 56, 303–309. [Google Scholar] [CrossRef]
- Kirsch J, Betz H. Glycine-receptor activation is required for receptor clustering in spinal neurons. Nature. 1998, 392, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chávez G, Velázquez-Flores MÁ, Ruiz Esparza-Garrido R, Salceda R. Glycine receptor subunits expression in the developing rat retina. Neurochemistry International. 2017, 108, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Young TL, Cepko CL. A Role for Ligand-Gated Ion Channels in Rod Photoreceptor Development. Neuron. 2004, 41, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, S. Glycine Receptor Diversity in the Mammalian Retina. In Webvision: The Organization of the Retina and Visual System; Kolb H, Nelson R, Fernandez E, Ed.; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 1995. [Google Scholar]
- Famiglietti EV, Jr. , Kolb H. A bistratified amacrine cell and synaptic cirucitry in the inner plexiform layer of the retina. Brain Res. 1975, 84, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Hsueh HA, Molnar A, Werblin FS. Amacrine-to-amacrine cell inhibition in the rabbit retina. J Neurophysiol. 2008, 100, 2077–2088. [Google Scholar] [CrossRef]
- Bormann J, Rundström N, Betz H, Langosch D. Residues within transmembrane segment M2 determine chloride conductance of glycine receptor homo- and hetero-oligomers. Embo j. 1993, 12, 3729–3737. [Google Scholar] [CrossRef]
- Betz, H. Structure and function of inhibitory glycine receptors. Q Rev Biophys. 1992, 25, 381–394. [Google Scholar] [CrossRef]
- Guidry, C. The role of Müller cells in fibrocontractive retinal disorders. Prog Retin Eye Res. 2005, 24, 75–86. [Google Scholar] [CrossRef]
- Gholami S, Kamali Y, Reza Rostamzad M. Glycine Supplementation Ameliorates Retinal Neuronal Damage in an Experimental Model of Diabetes in Rats: A Light and Electron Microscopic Study. J Ophthalmic Vis Res. 2019, 14, 448–456. [Google Scholar] [CrossRef]
- Hou M, Duan L, Slaughter MM. Synaptic inhibition by glycine acting at a metabotropic receptor in tiger salamander retina. J Physiol. 2008, 586, 2913–2926. [Google Scholar] [CrossRef] [PubMed]
- Zhang P-P, Zhang G, Zhou W, Weng S-J, Yang X-L, Zhong Y-M. Signaling mechanism for modulation by ATP of glycine receptors on rat retinal ganglion cells. Scientific Reports. 2016, 6, 28938. [Google Scholar] [CrossRef]
- Marc AM, Veeramuthu B, Xiaohan W, Henrique von G. Glycine Release Is Potentiated by cAMP via EPAC2 and Ca<sup>2+</sup> Stores in a Retinal Interneuron. The Journal of Neuroscience. 2021, 41, 9503. [Google Scholar] [CrossRef]
- Freed MA, Liang Z. Synaptic noise is an information bottleneck in the inner retina during dynamic visual stimulation. The Journal of Physiology. 2014, 592, 635–651. [Google Scholar] [CrossRef]
- Witkovsky, P. Dopamine and retinal function. Documenta Ophthalmologica. 2004, 108, 17–39. [Google Scholar] [CrossRef]
- Hattar S, Liao HW, Takao M, Berson DM, Yau KW. Melanopsin-Containing Retinal Ganglion Cells: Architecture, Projections, and Intrinsic Photosensitivity. Science. 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Versaux-Botteri C, Verney C, Zecevic N, Nguyen-Legros J. Early appearance of tyrosine hydroxylase immunoreactivity in the retina of human embryos. Developmental Brain Research. 1992, 69, 283–287. [Google Scholar] [CrossRef]
- Wulle I, Schnitzer J. Distribution and morphology of tyrosine hydroxylase-immunoreactive neurons in the developing mouse retina. Developmental Brain Research. 1989, 48, 59–72. [Google Scholar] [CrossRef]
- Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. Dopamine Receptors: From Structure to Function. Physiological Reviews. 1998, 78, 189–225. [Google Scholar] [CrossRef] [PubMed]
- Iuvone PM, Tosini G, Pozdeyev N, Haque R, Klein DC, Chaurasia SS. Circadian clocks, clock networks, arylalkylamine N-acetyltransferase, and melatonin in the retina. Prog Retin Eye Res. 2005, 24, 433–456. [Google Scholar] [CrossRef]
- Goel M, Mangel SC. Dopamine-Mediated Circadian and Light/Dark-Adaptive Modulation of Chemical and Electrical Synapses in the Outer Retina. Frontiers in Cellular Neuroscience 2021, 15. [Google Scholar] [CrossRef]
- Troy JB, Shou T. The receptive fields of cat retinal ganglion cells in physiological and pathological states: where we are after half a century of research. Prog Retin Eye Res. 2002, 21, 263–302. [Google Scholar] [CrossRef] [PubMed]
- Hammond, P. Receptive field mechanisms of sustained and transient retinal ganglion cells in the cat. Exp Brain Res. 1975, 23, 113–128. [Google Scholar] [CrossRef]
- Harnois C, Di Paolo T. Decreased dopamine in the retinas of patients with Parkinson's disease. Invest Ophthalmol Vis Sci. 1990, 31, 2473–2475. [Google Scholar]
- Moschos MM, Tagaris G, Markopoulos L, Margetis L, Tsapakis S, Kanakis M, Koutsandrea C. Morphologic Changes and Functional Retinal Impairment in Patients with Parkinson Disease without Visual Loss. European Journal of Ophthalmology. 2011, 21, 24–29. [Google Scholar] [CrossRef]
- Mao J, Liu S, Qin W, Li F, Wu X, Tan Q. Levodopa inhibits the development of form-deprivation myopia in guinea pigs. Optom Vis Sci. 2010, 87, 53–60. [Google Scholar] [CrossRef]
- Gao Q, Liu Q, Ma P, Zhong X, Wu J, Ge J. Effects of direct intravitreal dopamine injections on the development of lid-suture induced myopia in rabbits. Graefes Arch Clin Exp Ophthalmol. 2006, 244, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Ashby RS, Schaeffel F. The effect of bright light on lens compensation in chicks. Invest Ophthalmol Vis Sci. 2010, 51, 5247–5253. [Google Scholar] [CrossRef]
- Backhouse S, Collins AV, Phillips JR. Influence of periodic vs continuous daily bright light exposure on development of experimental myopia in the chick. Ophthalmic Physiol Opt. 2013, 33, 563–572. [Google Scholar] [CrossRef]
- Lan W, Feldkaemper M, Schaeffel F. Intermittent episodes of bright light suppress myopia in the chicken more than continuous bright light. PLoS One. 2014, 9, e110906. [Google Scholar] [CrossRef]
- Ivanova TN, Alonso-Gomez AL, Iuvone PM. Dopamine D4 receptors regulate intracellular calcium concentration in cultured chicken cone photoreceptor cells: Relationship to dopamine receptor-mediated inhibition of cAMP formation. Brain Research. 2008, 1207, 111–119. [Google Scholar] [CrossRef]
- Jackson CR, Chaurasia SS, Zhou H, Haque R, Storm DR, Iuvone PM. Essential roles of dopamine D4 receptors and the type 1 adenylyl cyclase in photic control of cyclic AMP in photoreceptor cells. J Neurochem. 2009, 109, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, ML. Melatonin is a potent modulator of dopamine release in the retina. Nature. 1983, 306, 782–784. [Google Scholar] [CrossRef]
- Nowak JZ, Z̵urawska E, Zawilska J. Melatonin and its generating system in vertebrate retina: circadian rhythm, effect of environmental lighting and interaction with dopamine. Neurochemistry International. 1989, 14, 397–406. [Google Scholar] [CrossRef]
- Ford KJ, Feller MB. Assembly and disassembly of a retinal cholinergic network. Vis Neurosci. 2012, 29, 61–71. [Google Scholar] [CrossRef]
- Feller MB, Wellis DP, Stellwagen D, Werblin FS, Shatz CJ. Requirement for Cholinergic Synaptic Transmission in the Propagation of Spontaneous Retinal Waves. Science. 1996, 272, 1182–1187. [Google Scholar] [CrossRef]
- Burbridge Timothy J, Xu H-P, Ackman James B, Ge X, Zhang Y, Ye M-J, Zhou ZJ, Xu J, Contractor A, Crair Michael C. Visual Circuit Development Requires Patterned Activity Mediated by Retinal Acetylcholine Receptors. Neuron. 2014, 84, 1049–1064. [Google Scholar] [CrossRef]
- Zhou ZJ, Fain GL. Starburst amacrine cells change from spiking to nonspiking neurons during retinal development. Proc Natl Acad Sci U S A. 1996, 93, 8057–8062. [Google Scholar] [CrossRef]
- Sernagor E, Grzywacz NM. Influence of spontaneous activity and visual experience on developing retinal receptive fields. Curr Biol. 1996, 6, 1503–1508. [Google Scholar] [CrossRef]
- Wong WT, Wong RO. Changing specificity of neurotransmitter regulation of rapid dendritic remodeling during synaptogenesis. Nat Neurosci. 2001, 4, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Mehta V, Sernagor E. Early neural activity and dendritic growth in turtle retinal ganglion cells. Eur J Neurosci. 2006, 24, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Arroyo DA, Kirkby LA, Feller MB. Retinal Waves Modulate an Intraretinal Circuit of Intrinsically Photosensitive Retinal Ganglion Cells. J Neurosci. 2016, 36, 6892–6905. [Google Scholar] [CrossRef] [PubMed]
- Zeisel SH, Da Costa K-A, Franklin PD, Alexander EA, Lamont JT, Sheard NF, Beiser A. Choline, an essential nutrient for humans. The FASEB Journal. 1991, 5, 2093–2098. [Google Scholar] [CrossRef]
- Voigt, T. Cholinergic amacrine cells in the rat retina. Journal of Comparative Neurology. 1986, 248, 19–35. [Google Scholar] [CrossRef]
- Townes-Anderson E, Vogt BA. Distribution of muscarinic acetylcholine receptors on processes of isolated retinal cells. J Comp Neurol. 1989, 290, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Strang CE, Long Y, Gavrikov KE, Amthor FR, Keyser KT. Nicotinic and muscarinic acetylcholine receptors shape ganglion cell response properties. Journal of Neurophysiology. 2014, 113, 203–217. [Google Scholar] [CrossRef]
- Hall LM, Hellmer CB, Koehler CC, Ichinose T. Bipolar Cell Type-Specific Expression and Conductance of Alpha-7 Nicotinic Acetylcholine Receptors in the Mouse Retina. Invest Ophthalmol Vis Sci. 2019, 60, 1353–1361. [Google Scholar] [CrossRef]
- Strang CE, Renna JM, Amthor FR, Keyser KT. Nicotinic acetylcholine receptor expression by directionally selective ganglion cells. Vis Neurosci. 2007, 24, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Keyser KT, MacNeil MA, Dmitrieva N, Wang F, Masland RH, Lindstrom JM. Amacrine, ganglion, and displaced amacrine cells in the rabbit retina express nicotinic acetylcholine receptors. Vis Neurosci. 2000, 17, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva NA, Strang CE, Keyser KT. Expression of alpha 7 nicotinic acetylcholine receptors by bipolar, amacrine, and ganglion cells of the rabbit retina. J Histochem Cytochem. 2007, 55, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Fischer AJ, McKinnon LA, Nathanson NM, Stell WK. Identification and localization of muscarinic acetylcholine receptors in the ocular tissues of the chick. J Comp Neurol. 1998, 392, 273–284. [Google Scholar] [CrossRef]
- Ruan Y, Patzak A, Pfeiffer N. Muscarinic Acetylcholine Receptors in the Retina-Therapeutic Implications. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Bernard V, Normand E, Bloch B. Phenotypical characterization of the rat striatal neurons expressing muscarinic receptor genes. The Journal of Neuroscience. 1992, 12, 3591–3600. [Google Scholar] [CrossRef]
- Laspas P, Zhutdieva MB, Brochhausen C, Musayeva A, Zadeh JK, Pfeiffer N, Xia N, Li H, Wess J, Gericke A. The M1 muscarinic acetylcholine receptor subtype is important for retinal neuron survival in aging mice. Scientific Reports. 2019, 9, 5222. [Google Scholar] [CrossRef]
- Elstrott J, Anishchenko A, Greschner M, Sher A, Litke AM, Chichilnisky EJ, Feller MB. Direction selectivity in the retina is established independent of visual experience and cholinergic retinal waves. Neuron. 2008, 58, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Lee S, Kim K, Zhou ZJ. Role of ACh-GABA Cotransmission in Detecting Image Motion and Motion Direction. Neuron. 2010, 68, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Vaney DI, Sivyer B, Taylor WR. Direction selectivity in the retina: symmetry and asymmetry in structure and function. Nature Reviews Neuroscience. 2012, 13, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Tiriac A, Bistrong K, Pitcher MN, Tworig JM, Feller MB. The influence of spontaneous and visual activity on the development of direction selectivity maps in mouse retina. Cell Rep. 2022, 38, 110225. [Google Scholar] [CrossRef] [PubMed]
- Hellmer CB, Hall LM, Bohl JM, Sharpe ZJ, Smith RG, Ichinose T. Cholinergic feedback to bipolar cells contributes to motion detection in the mouse retina. Cell Rep. 2021, 37, 110106. [Google Scholar] [CrossRef] [PubMed]
- Lusthaus J, Goldberg I. Current management of glaucoma. Med J Aust. 2019, 210, 180–187. [Google Scholar] [CrossRef]
- Shen J, Wang Y, Yao K. Protection of retinal ganglion cells in glaucoma: Current status and future. Exp Eye Res. 2021, 205, 108506. [Google Scholar] [CrossRef]
Figure 1.
Left: Schematic diagram of the eye where a section of the retina is highlighted by the orange circle, which is expanded in the right figure. Right: Schematic diagram of retinal layers, retinal cells, and neurotransmitters. Abbreviations used: ACh, acetylcholine; EZ, ellipsoid zone; GCL, ganglion cell layer; Glu, glutamate; Gly, glycine; ILM, inner limiting membrane; INL, inner nuclear layer; IPL, inner plexiform layer; OPL, outer plexiform layer; ONL, outer nuclear layer; POS, photoreceptor outer segments; RNFL, retinal nerve fiber layer; RPE, retinal pigment epithelium.
Figure 1.
Left: Schematic diagram of the eye where a section of the retina is highlighted by the orange circle, which is expanded in the right figure. Right: Schematic diagram of retinal layers, retinal cells, and neurotransmitters. Abbreviations used: ACh, acetylcholine; EZ, ellipsoid zone; GCL, ganglion cell layer; Glu, glutamate; Gly, glycine; ILM, inner limiting membrane; INL, inner nuclear layer; IPL, inner plexiform layer; OPL, outer plexiform layer; ONL, outer nuclear layer; POS, photoreceptor outer segments; RNFL, retinal nerve fiber layer; RPE, retinal pigment epithelium.
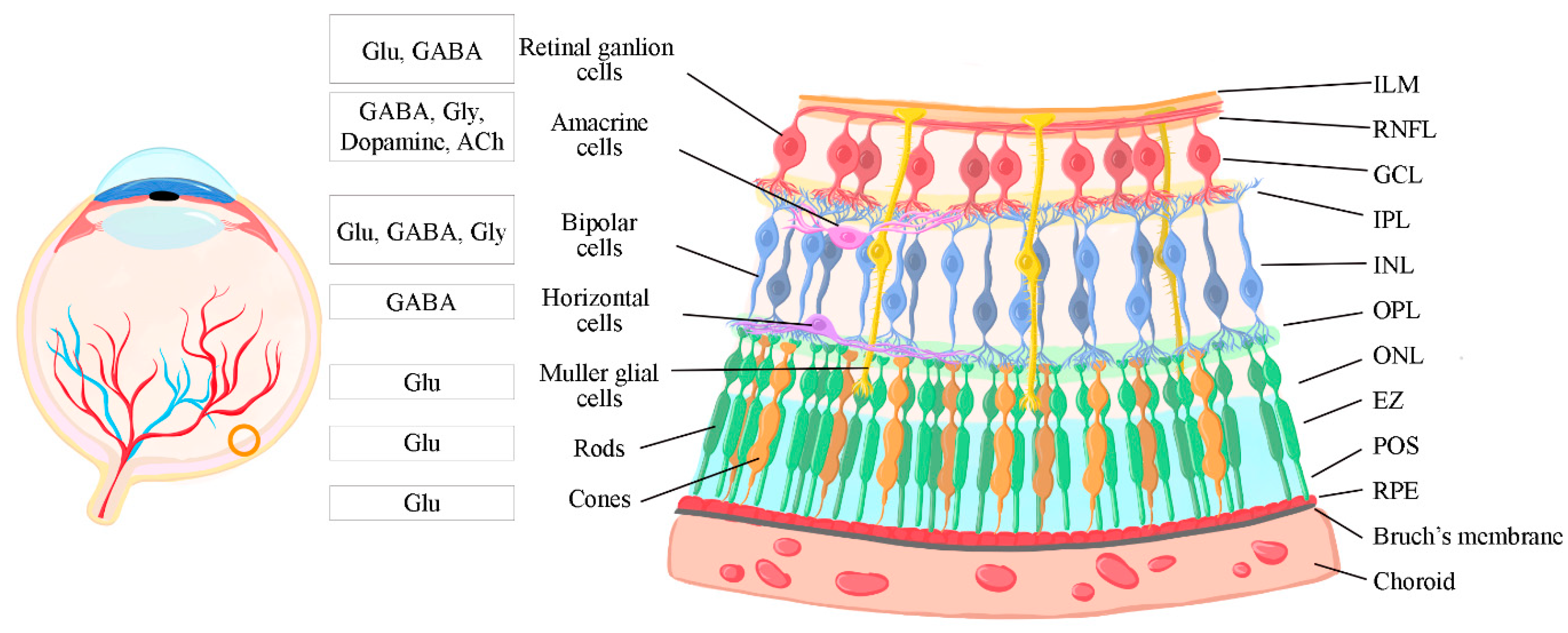
Figure 2.
Timeline of retinal development in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day.
Figure 2.
Timeline of retinal development in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day.
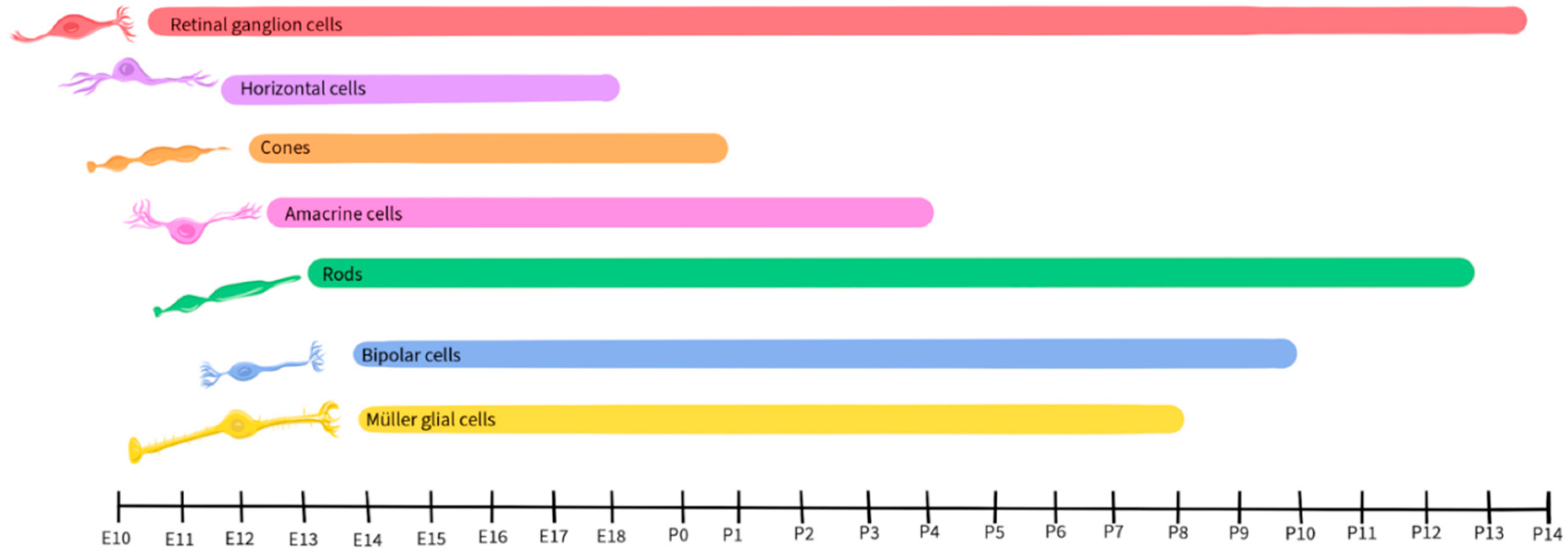
Figure 3.
Timeline of retinal development and glutamate activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. The gray line below the timescale shows glutamatergic activity.
Figure 3.
Timeline of retinal development and glutamate activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. The gray line below the timescale shows glutamatergic activity.

Figure 4.
Localization of glutamate receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different glutamate receptors. Right: color legend indicating the types of glutamate receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; AC, amacrine cell; RGC, retinal ganglion cell.
Figure 4.
Localization of glutamate receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different glutamate receptors. Right: color legend indicating the types of glutamate receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; AC, amacrine cell; RGC, retinal ganglion cell.
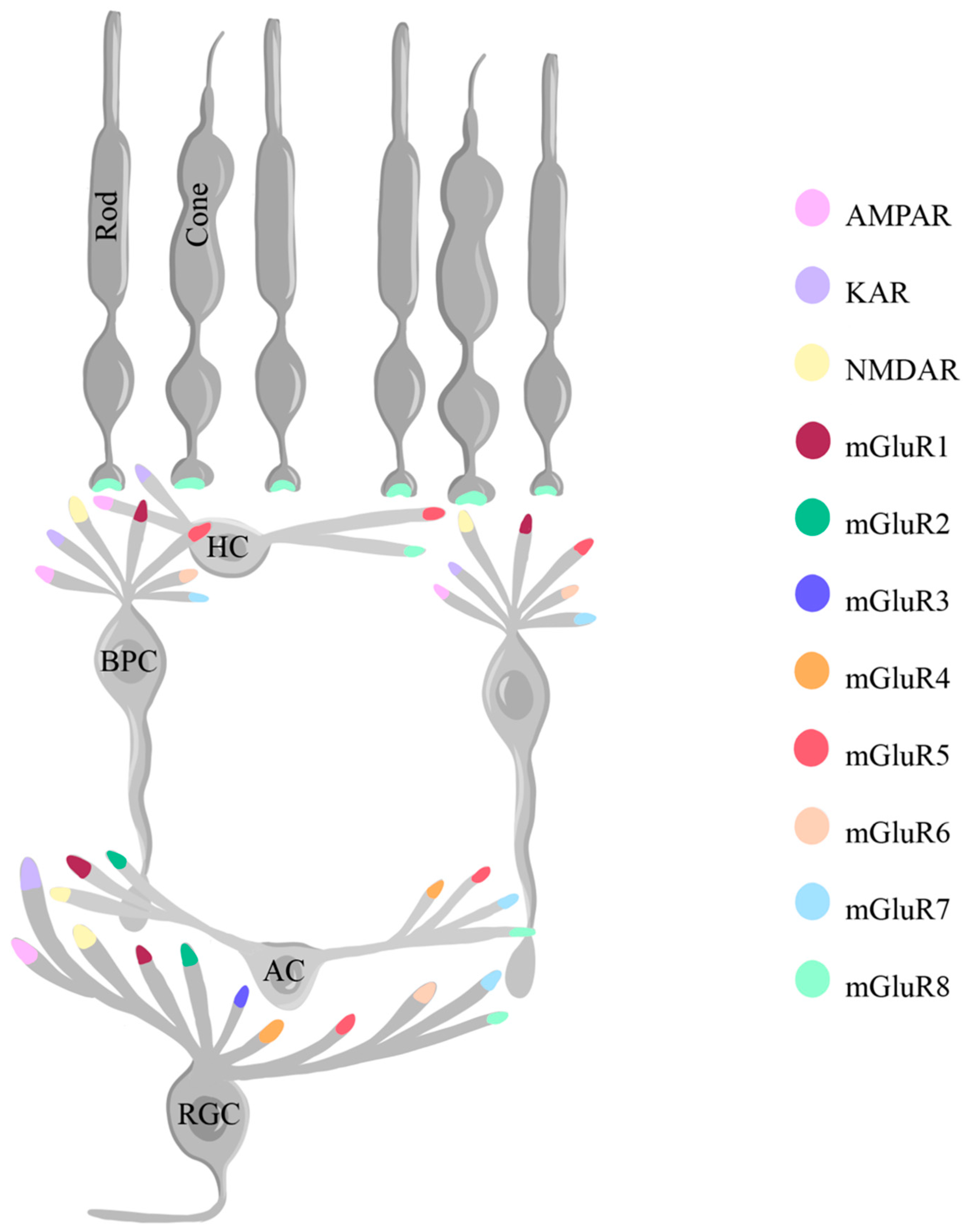
Figure 5.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic and GABAergic activity.
Figure 5.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic and GABAergic activity.

Figure 6.
Localization of GABA receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different GABA receptors. Right: color legend indicating the types of GABA receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; MGC, Müller glial cell AC, amacrine cell; RGC, retinal ganglion cell.
Figure 6.
Localization of GABA receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different GABA receptors. Right: color legend indicating the types of GABA receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; MGC, Müller glial cell AC, amacrine cell; RGC, retinal ganglion cell.
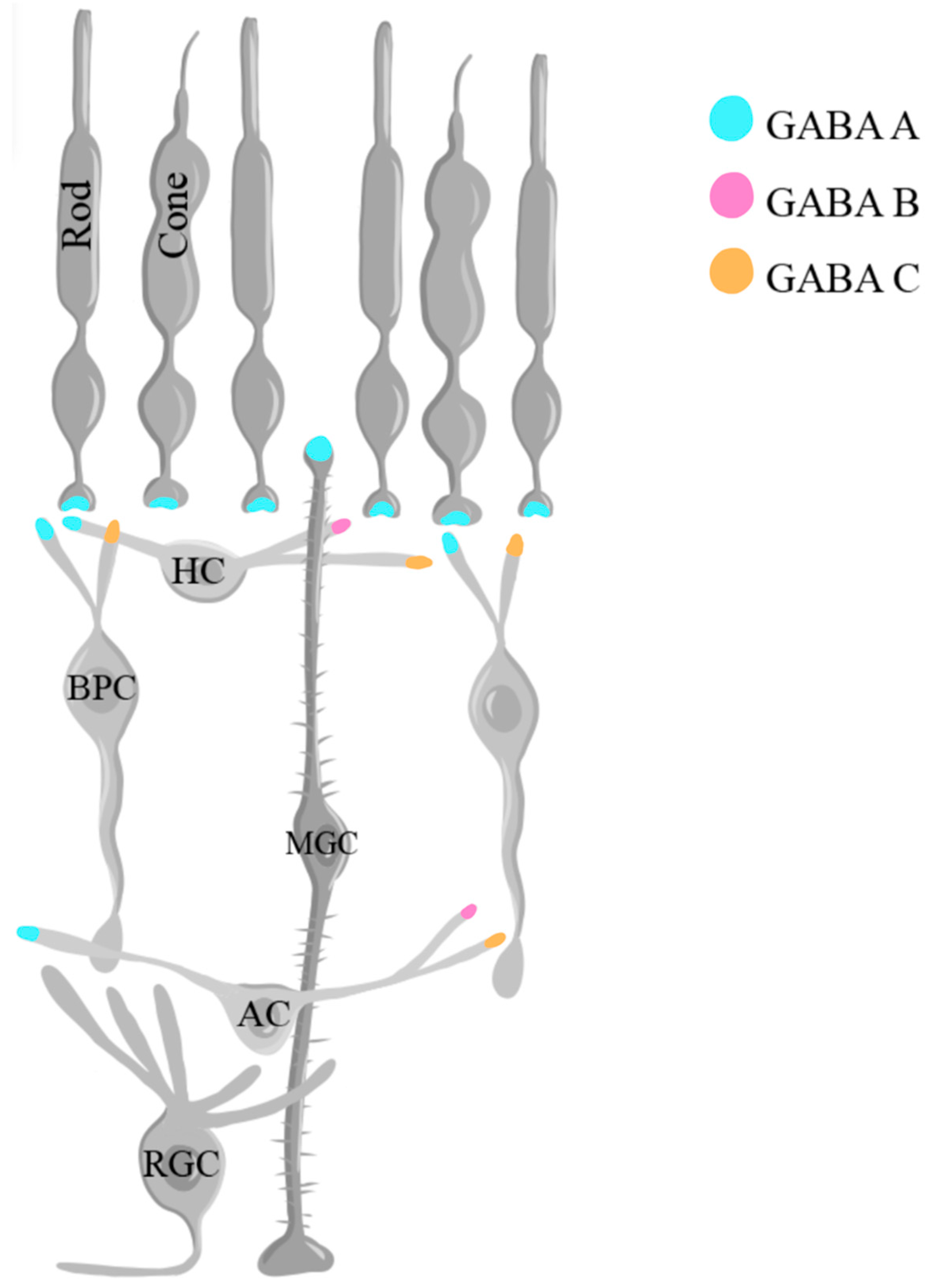
Figure 7.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, and glycinergic activity.
Figure 7.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, and glycinergic activity.
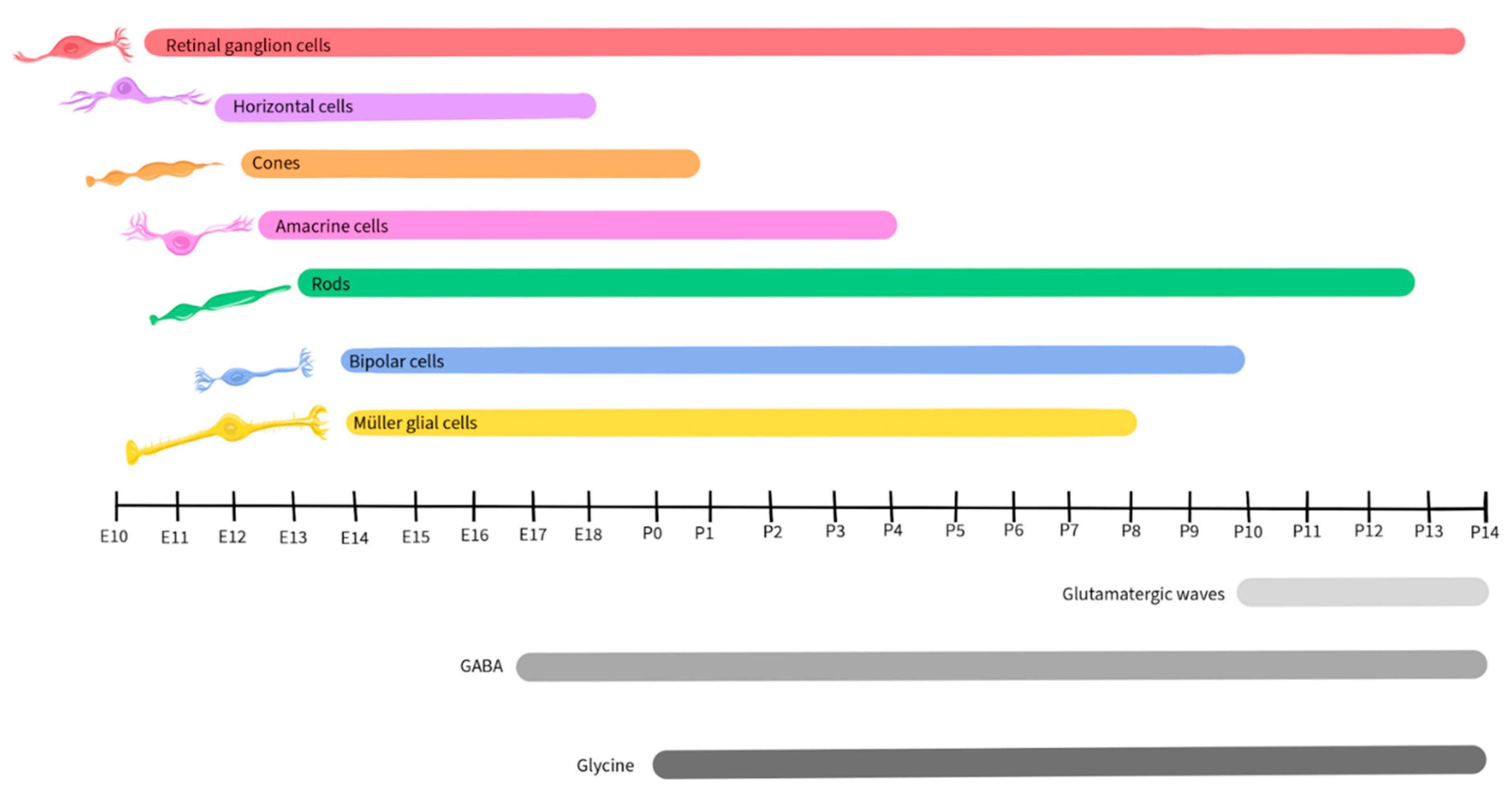
Figure 8.
Diagram of glycine crossover inhibition in the retina adapted from Hsueh et al., 2008. Grey illustrations depict the different retinal neurons. Blue arrows represent glutamate excitation. Orange lines represent glycinergic inhibition. Abbreviations: BPC, bipolar cell; RGC, retinal ganglion cell.
Figure 8.
Diagram of glycine crossover inhibition in the retina adapted from Hsueh et al., 2008. Grey illustrations depict the different retinal neurons. Blue arrows represent glutamate excitation. Orange lines represent glycinergic inhibition. Abbreviations: BPC, bipolar cell; RGC, retinal ganglion cell.
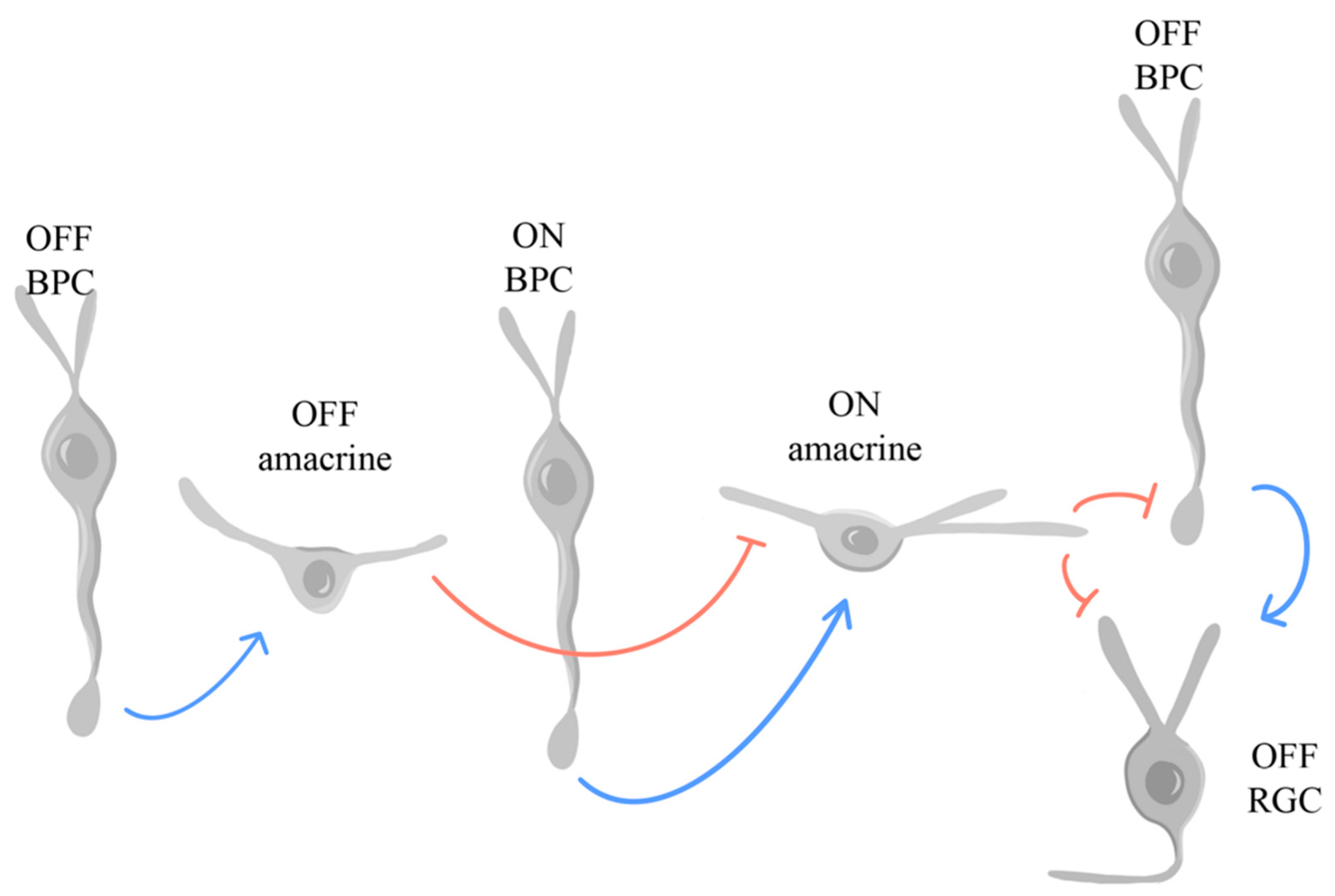
Figure 9.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, glycinergic, and dopaminergic activity.
Figure 9.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, glycinergic, and dopaminergic activity.
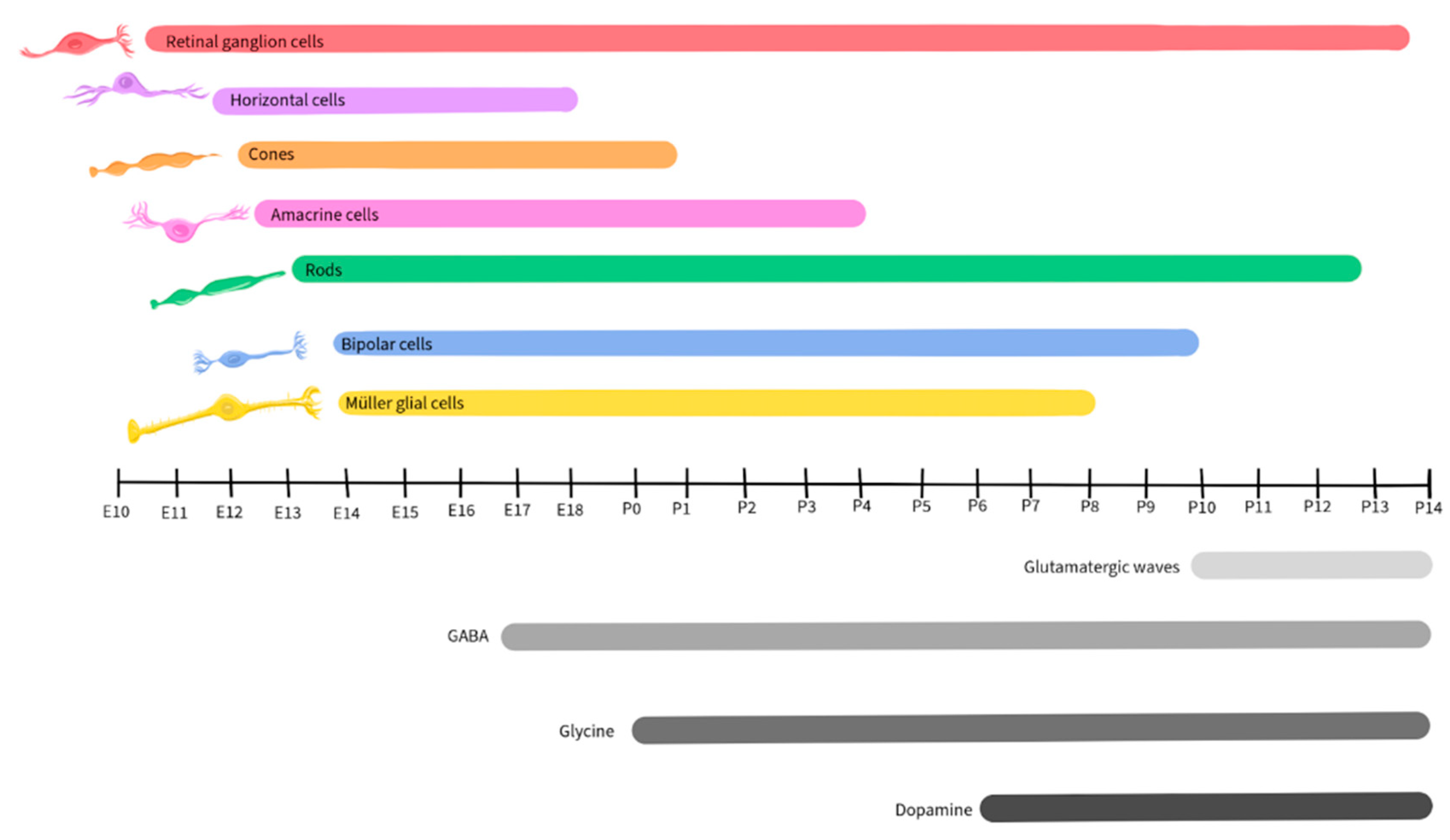
Figure 10.
Flow diagram of dopamine transmission across the retina.
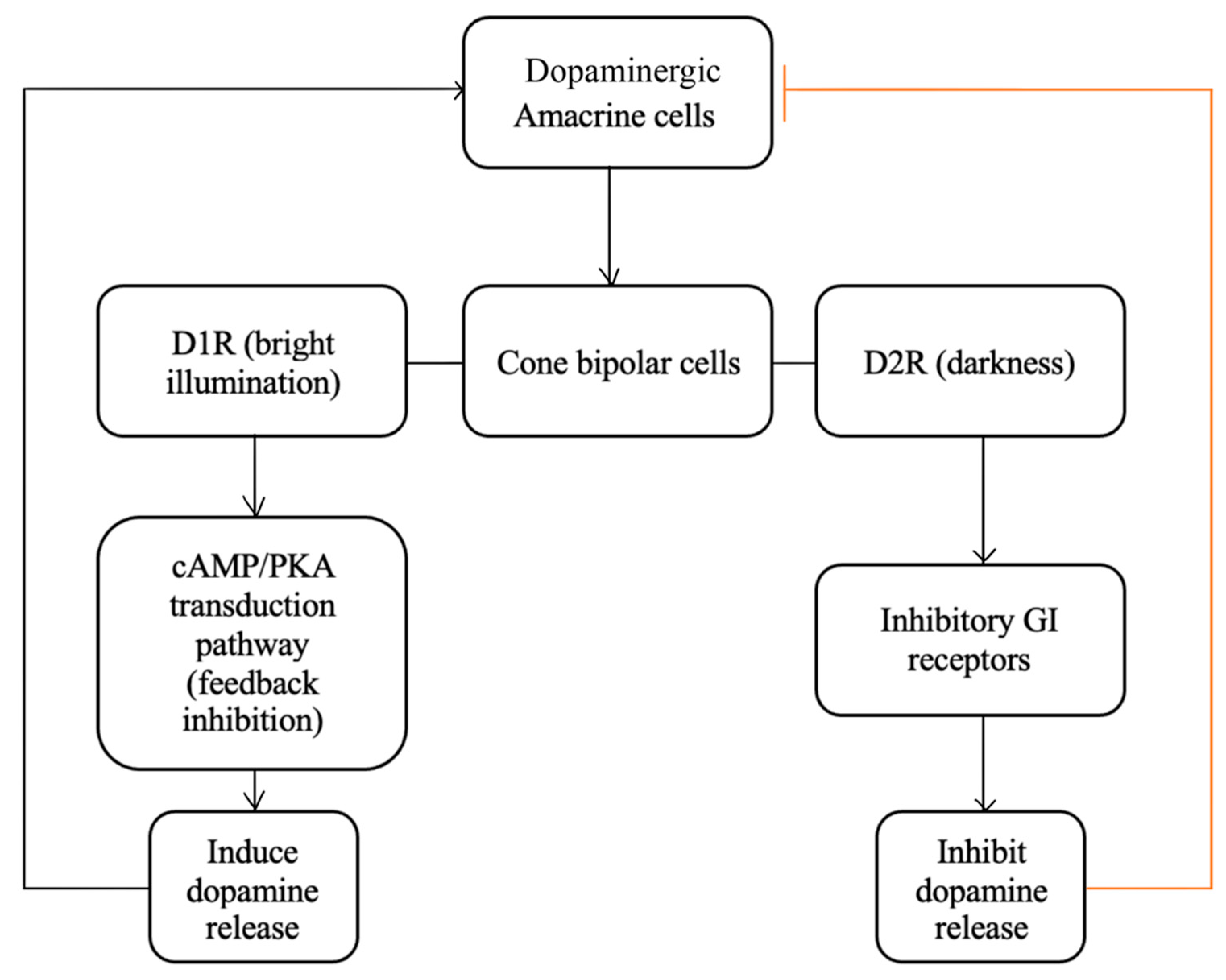
Figure 11.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, glycinergic, dopaminergic, and cholinergic activity.
Figure 11.
Timeline of retinal development and neurotransmitter activity in mice. Colored lines above the timescale show the time during embryonic and postnatal development when each retinal cell type arises. Abbreviations used: E, embryonic day; P, postnatal day. Gray lines below the timescale show glutamatergic, GABAergic, glycinergic, dopaminergic, and cholinergic activity.
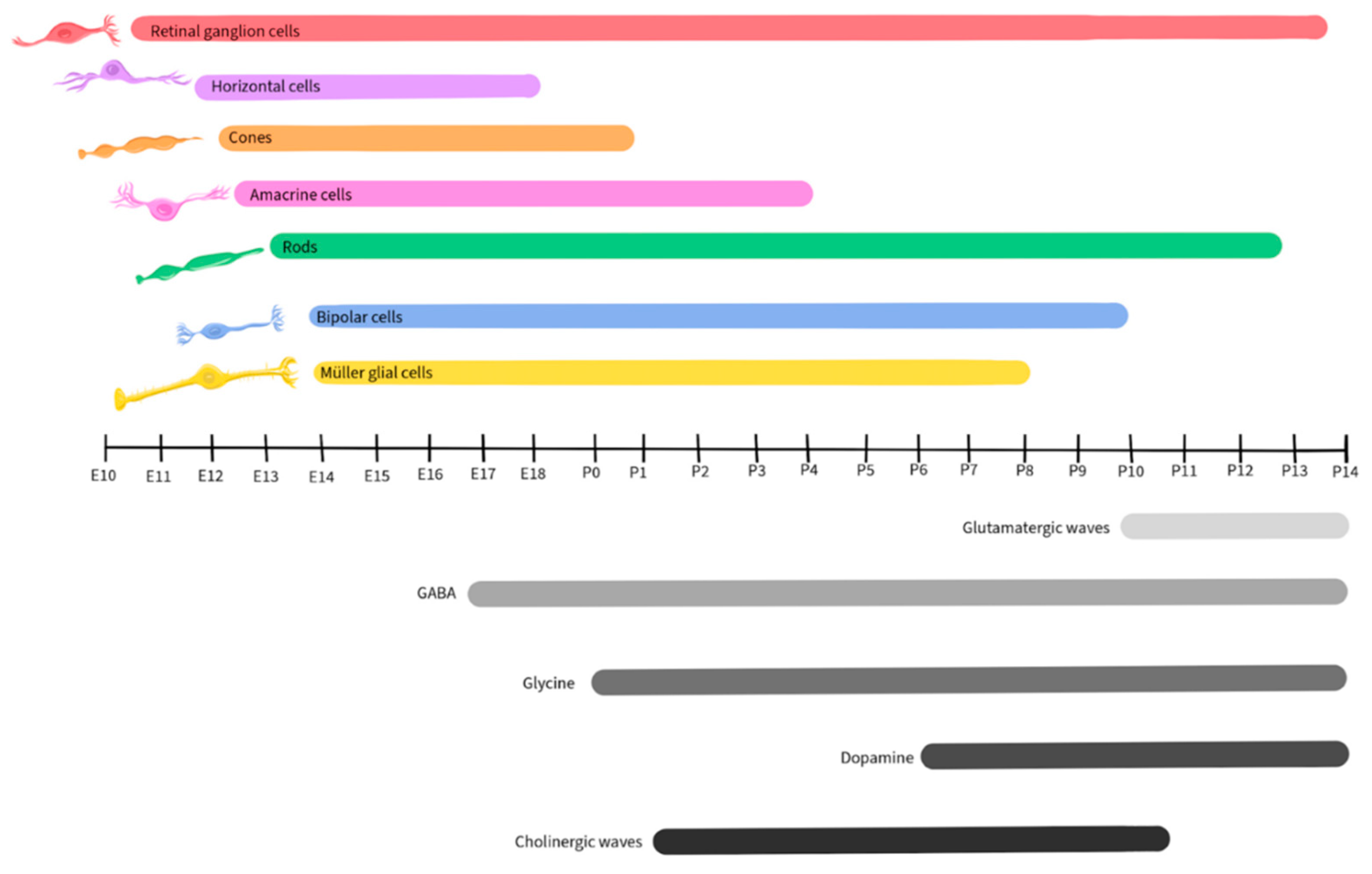
Figure 12.
Localization of acetylcholine receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different ACh receptors. Right: color legend indicating the types of ACh receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; SAC, starburst amacrine cell; AC, amacrine cell; RGC, retinal ganglion cell.
Figure 12.
Localization of acetylcholine receptors in the retina. Left: grey illustrations depict the different retinal neurons and colored portions depict the expression of different ACh receptors. Right: color legend indicating the types of ACh receptors represented in the image on the left. Abbreviations: HC, horizontal cell; BPC, bipolar cell; SAC, starburst amacrine cell; AC, amacrine cell; RGC, retinal ganglion cell.
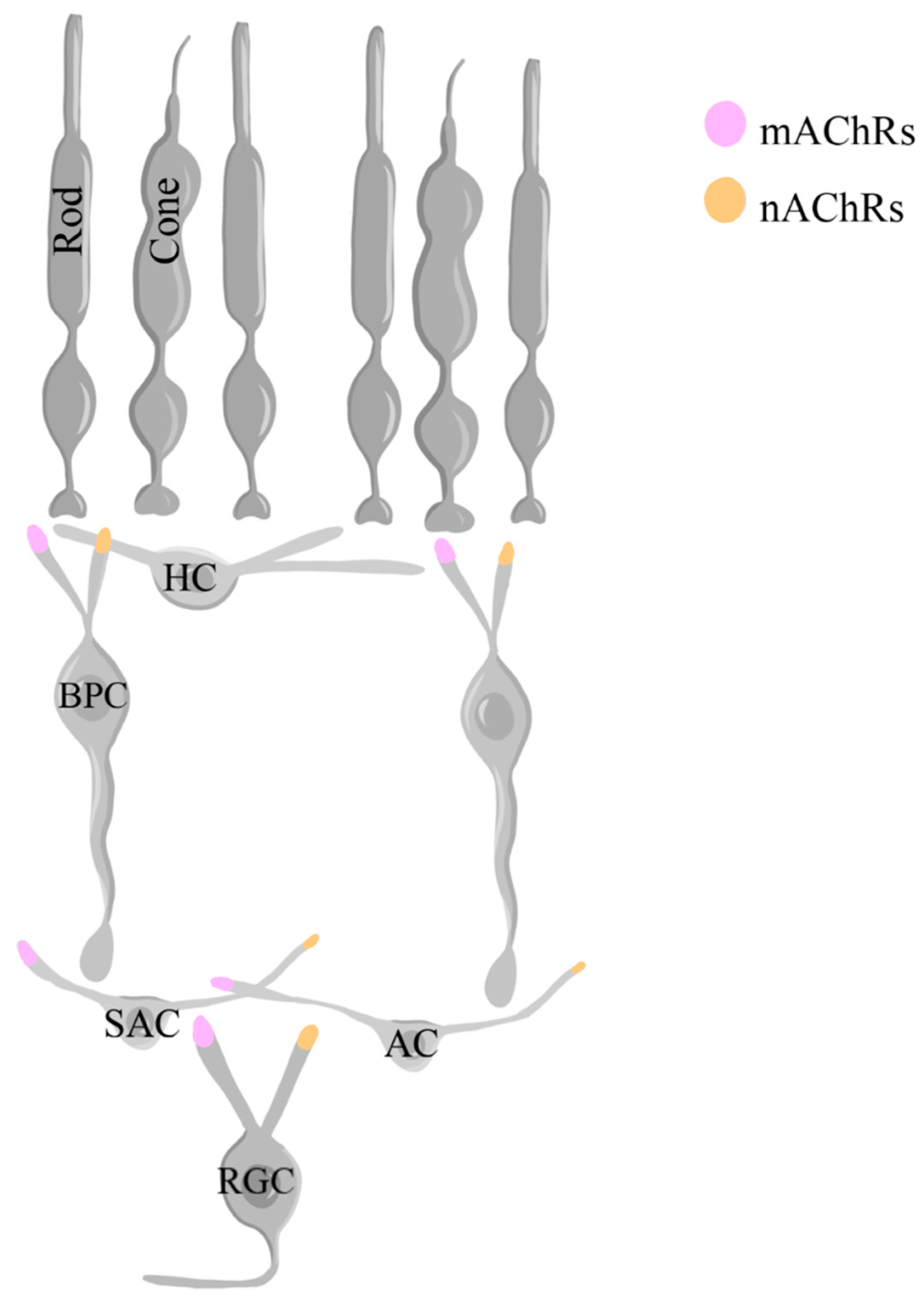

Table 1.
Summary of neurotransmitter involvement in retinal development and healthy function.
| Neurotransmitter | Retinal Development | Normal Retinal Physiology |
|---|---|---|
| Glutamate |
|
|
| GABA |
|
|
| Glycine |
|
|
| Dopamine |
|
|
| Acetylcholine (ACh) |
|
|
Table 2.
Summary of neurotransmitter involvement in retinal disease.
| Glaucoma | Retinal Ischemia | Diabetic Retinopathy | Parkinson’s Disease Visual Impairment | Form-Depravation Myopia | |
|---|---|---|---|---|---|
| Glutamate |
|
|
|
Unknown. | Unknown. |
| GABA | Unknown. |
|
|
Unknown. | Unknown. |
| Glycine | Unknown. | Unknown. |
|
Unknown. | Unknown. |
| Dopamine | Unknown. | Unknown. | Unknown. |
|
|
| Acetylcholine (ACh) |
|
Unknown. | Unknown. | Unknown. | Unknown. |
Table 3.
Summary of calcium interplay with different neurotransmitters in retinal development, health, and disease as discussed in the present review.
Table 3.
Summary of calcium interplay with different neurotransmitters in retinal development, health, and disease as discussed in the present review.
| Neurotransmitter | Role of Calcium Interplay |
|---|---|
| Glutamate |
|
| GABA |
|
| Glycine |
|
| Dopamine |
|
| Acetylcholine (ACh) |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



