Submitted:
12 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
Recent studies suggest that the interaction of dietary constituents with taste and olfactory receptors and nociceptor expressed in the oral cavity, nasal cavity and gastrointestinal tract may regulate homeostasis through activation of the neuroendocrine system. Polyphenols, of which 8,000 have been identified to date, represent the greatest diversity of secondary metabolites in plants, most of which are bitter and some of them astringent. Epidemiological studies have shown that polyphenol intake contributes to maintaining and improving cardiovascular, cognitive and sensory health. However, because polyphenols have very low bioavailability, the mechanisms of their beneficial effects are unknown. In this review, we focus on the taste of polyphenols from the perspective of sensory nutrition, summarize the results of previous studies on their relationship with bioregulation, and discuss their future potential.
Keywords:
Polyphenol
; sensory nutrition
; bitter
; astringency
; central nervous system
1. Introduction
In recent years, attention has focused on sensory nutrition, a field of study that examines how the senses conveyed by what people eat and drink act on the brain and how these signals affect human behavior and homeostasis [1]. In 2019, the National Institutes of Health (NIH) hosted a workshop on ’Sensory Nutrition and Disease’. The workshop invited a diverse group of researchers from neuroscience, food science, psychology, nutrition, and health sciences to understand how chemosensory influences affect eating. The workshop encouraged people to explore how these influences impact their choices and health [2]. Three topics were discussed at the conference: a) the need to optimize chemosensory testing and assessment in humans, b) the plasticity of the chemosensory system, and c) the interplay between chemosensory signals, cognitive signals, dietary intake, and metabolism, providing some guidance in advancing sensory nutrition research.
Chemosensory receptors expressed in the oral-nasal trigeminal system have been well studied to transmit signals from food to the brain and determine feeding behavior, such as ingestion or rejection [3,4]. Olfactory receptors respond to thousands of different types of volatiles [5,6,7]. Taste receptors respond to salts, sugars, amino acids, alkaloids, acids and fats [8,9]. In addition, stimuli such as temperature, osmotic pressure, pH, pungency and menthol are recognized by transient receptor potential (TRP) [10,11,12]. Flavor perception is formed by integrating signals from various chemosensory systems in the oral and nasal cavities within the brain [13,14]. Flavor perception is thought to be shaped by neural processes occurring in chemosensory regions of the brain, including the anterior insula, frontal operculum, orbitofrontal cortex and anterior cingulate cortex, and by interactions with other heteromodal areas including the posterior parietal cortex and ventral lateral prefrontal cortex. In recent years, it has been discovered that chemoreceptors responsible for flavor formation are expressed outside of the oral-nasal trigeminal system [4,15,16]. For instance, it is known that olfactory receptors are in sperm regulated their motility [17], in skeletal muscle controlled differentiation and regeneration [18], in adipocytes [19] and liver [20] regulated energy metabolism, and involved in gastrointestinal hormone secretion in the intestinal tract [21]. The sweet taste receptor 2/3 (T1R2/3) is expressed in various parts of the body, including the gut, pancreas, brain, bladder, bone, and adipose tissue and plays a role in energy metabolism [22]. In addition, it has been shown to induce strong relaxation in the bladder [23]. The bitter taste receptor, the taste receptor 2 (T2R) receptor, is expressed in several tissues, including the lungs, gastrointestinal tract, kidneys, genitals and brain. Xie et al. described the identification of numerous T2Rs in the gastrointestinal tract [24], and they are also expressed in the brain [25], heart [26], respiratory tract [27], reproductive system [28], bone marrow stroma and the vascular wall [29]. T1R2/3 and T2R are believed to act antagonistically in regulating innate immunity in the respiratory tract [30]. Heterodimers of the taste receptors Taste receptor 1 and Taste receptor 3 (T1R1/3) regulate gastrointestinal hormones in the gastrointestinal tract [31]. It has also been reported that it controls the fertilization ability of sperm [32]. Salt-taste channels, epithelial sodium channels (ENaCs), are known to be expressed in the kidney, distal colon, bladder, stomach and lungs [33]. Almost every cell in the body expresses at least one member of the TRP channel family, which has distinct biophysical, mechanical, and pharmacological properties [34]. In particular, TRP channels involved in sensory signaling are the best characterized and have been used as experimental models to understand functional aspects.
Sensory receptors are located in different organs and are thought to have different functions, but many details remain unknown. In particular, it is not well understood how these sensory receptors in the gut react to the nutrients and phytochemicals in food, and how the subsequent signals from these food components are transmitted to the central nervous system (CNS). Recent reports suggest that food signals may contribute to homeostasis via sensory receptors in the gastrointestinal tract. For example, in the gastrointestinal tract, by the activation of GPCRs, olfactory receptors and sweet/umami/bitter taste receptors, secrete gastrointestinal hormones. These gastrointestinal (GI) hormones have been reported to activate the vagus nerve, and the stimulation is transmitted to the CNS, affecting eating and cognitive function [35,36]. As such reports accumulate, there is a growing need to investigate the importance of sensory nutrition, that is, how chemosensory signals from food contribute to maintaining homeostasis and reducing disease risk via the gastrointestinal-brain axis.
Polyphenols are the largest and most diverse group of plant secondary metabolites, with 8000 identified to date [37]. Polyphenols are known to be coloring [38], bitter [39,40]and astringent [41,42] and have a significant impact on food palatability. In addition, daily consumption of these beverages can transform astringency and bitterness, which are inherently aversive perceptions, into pleasurable stimuli [43,44,45]..These results suggest that changes in taste preference and avoidance due to long-term taste exposure can be considered as adaptations accompanied by neuroplasticity. However, the mechanisms by which taste preferences change over time are not yet understood. Epidemiological studies have reported a negative correlation between polyphenol intake and cardiovascular disease [46], neurodegenerative diseases [47,48], age-related deterioration of sensory organs [49]. There are reports that people who are regular consumers of astringent and bitter drinks may be less at risk of type 2 diabetes and cardiovascular diseases, such as coffee and tea [50,51]. Large-scale intake studies of polyphenols from cocoa have also shown reduced cardiovascular deaths [52] and hippocampus-dependent cognitive improvements in the elderly [53]. In general, polyphenols are rarely absorbed from the upper gastrointestinal tract and move to the lower gastrointestinal tract, where they are partly broken down by intestinal bacteria, but mostly excreted in the feces [54]. Their beneficial mechanism of action is not known because it is very unlikely to be distributed in the blood or organs, including the brain. Recent studies have reported that polyphenols ingested from the diet can alter the composition of the gut flora [55]. It has been reported that the composition of secondary metabolites in the colon changes accordingly and that these may be absorbed and affect cardiovascular disease and cognitive function [56]. On the other hand, it is extremely difficult to elucidate these causal relationships because the type and quantity of polyphenols ingested vary greatly with diet, and the quality and quantity of polyphenol metabolites produced by the gut microbiota and themselves vary widely between individuals.
This review therefore focuses on the interaction of bitter and astringent tastes of polyphenols with sensory receptors, particularly with those that are expressed extra-oral cavity. In addition, we summarized the results of previous studies on how signals produced by polyphenols affect the CNS via gastrointestinal sensory receptors and how such signals affect peripheral organs, and explored the mechanisms involved.
2. Bitter Taste Receptors and Polyphenols
2.1. Bitter Taste Receptors Expressed Extra-Oral Cavity and GI Hormones.
Most pharmaceuticals are considered to present a bitter taste, and it is also known that bitter substances such as alkaloids and humulones are known to be present in plant food [57]. It has been well studied that these bitter substances are recognized by T2Rs, one of the G protein-coupled receptors (GPCRs), which are seven transmembrane receptors expressed on taste buds [58,59,60]. In human, 25 members of the 291-334 amino acid long T2R superfamily are involved in the perception of bitter taste [61,62]. As mentioned above, T2Rs are known to be distributed in various organs outside the oral cavity [24,25,26,27,28,29]. In recent years, research has increasingly focused on the relationship between bitter taste receptors expressed in the gastrointestinal tract and gastrointestinal hormones released by endocrine cell [63,64]. Typical gastrointestinal hormones involved in these are glucose-dependent insulinotropic polypeptide (GIP), which is derived from K cells located in the upper small intestine, and glucagon-like peptides, which are secreted mainly by L cells located in the distal small intestine. Incretins such as -1 (GLP-1) [65,66,67]. Another hormone of particular interest is cholecystokinin (CCK), which is produced by I-cells in the upper part of the small intestine [68]. These hormones have been reported to be motility modulators of the gastropyloric duodenum that delay gastric content evacuation [68,69]. The peptides secreted by neuropodal cells inhibit the gastric inhibitory vagal motor circuit (GIVMC) via the vagus nerve and nucleus tractus solitarius (NTS), resulting in delayed gastric emptying. This mechanism is suggested to reduce appetite and thus energy intake [69,70].
In addition, the secreted incretin stimulates insulin secretion, which has been extensively studied for its ability to reduce the increase in blood glucose levels after a meal [70,71]. GLP-1 has glucagonostatic effects, and its analogue and receptor agonists are commonly used to treat type 2 diabetes [72,73,74]. GLP-1 receptor agonists in particular are widely used in clinical practice but are known to cause gastrointestinal complications. Therefore, it can be difficult to use this medicine as a preventative measure. On the other hand, GLP-1 secretion has been reported to be induced by oral administration of typical bitter substances such as quinine [75,76] and denatonium benzoate [77]. This has led to increased interest in drug discovery based on the interaction between bitter substances and T2Rs [78,79].
2.2. Interactions between Bitter Taste Receptors and Polyphenols
Polyphenols are chemically classified as flavonoids (Figure 1a) including flavanols, flavanones, flavones, flavonols, flavanols, isoflavones, anthocyanins), chalcones, tannins (proanthocyanidins, hydrolysable tannins, fluorotannins, complex tannins) and phenols [55,80]. Polyphenol compounds are commonly associated with a bitter taste. However, there is limited practical evidence on the degree of bitterness associated with specific compounds as determined by human sensory evaluation. The specific sub-chemical structure of polyphenols that contributes to the bitter taste remains unclear. The interaction between polyphenols and T2R has been validated by in vitro experiments [81]. Most of the experiments have involved the expression of human T2R in HEK293 cells, the addition of polyphenols and the detection of calcium influx into the cells using fluorescent probes. A comprehensive survey of these previous studies was conducted in the following method.
The databases BitterDB (https://bitterdb.agri.huji.ac.il/dbbitter.php), Phenol explore (http://phenol-explorer.eu/) and the database of natural TAS2R agonists (https://github.com/dipizio/Natural_TAS2R_agonists) were used. Search formulae were "polyphenols" "flavonoids" "flavanols" " flavanones" "flavones" "flavonols" "flavanols" "isoflavones" "anthocyanins" "chalcones" "tannins " "pro(antho)cyanidins" "hydrolysable tannins" "fluorotannins "complex tannins" "phenols" and "taste receptor 2" "T2R" "TAS2R”. The survey was conducted in June 2023.
The results showed that 102 polyphenolic compounds have been tested to present using the methods described above. The findings are summarised in Table 1. Most of the compounds interacted with T2R14 and T2R39. However, (-)-epicatechin (Figure 1b), epigallocatechin gallate (EGCG, Figure 1c) and procyanidins (Figure 1d), oligomers of epicatechin, specifically interacted with T2R5 [81]. Additionally, procyanidins were found to interact with T2R7 [81]. Castalagin, grandinin, and vescalagin are hydrolyzable tannins that interact with T2R7. These tannins have a strong bitter taste similar to procyanidin, a condensed tannin [81]. The results indicate that T2R7 may have a significant impact on the bitterness experienced by humans. Among these polyphenols, green tea’s EGCG and amarogentin (Figure 1e), a type of secoiridoid glycoside, were found to interact with multiple T2Rs. EGCG was found to interact with T2R4, T2R5, T2R30 and T2R43 [39,81]. Amarogentin is a constituent of Gentian root, which has traditionally been used as a bitter stomachic. It has been reported to interact with eight T2Rs: T2R1, T2R4, T2R10, T2R30, T2R39, T2R43, T2R46, and T2R50 [82]. In these in vitro experiments, attempts have been made to calculate the EC50 of polyphenols that activate the T2R. The concentration range varied from a few micromoles to several tens of micromoles [39,40,82,83,84,85,86,87,88,89]. The EC50 values were similar to those of well-known bitter substances, such as absinthin, acetylthiourea, chloroquine, denatonium benzoate, ethylpyrazine, and methylthiourea [59]. The above suggests that polyphenols have high potential as a seed for drug design using bitterness.
Furthermore, it should be noted that the in vitro experiments only targeted a limited number of the 25 T2Rs, therefore the interactions with all T2Rs remain unknown. Also, due to the different experimental conditions in each report, it was unclear to what extent the calculated EC50 was correct. The specific sub-chemical structure of polyphenols that contributes to the bitter taste remains unclear. The unclear steric structure of T2R was partly responsible for the uncertainty involved. In 2022, Xu et al. revealed the conformation of T2R46 for the first time using cryo-EM [90]. This paper showed that when the ligand strychnine binds to T2R46 in the apo state and activates the receptor, Y241 acts as a ’toggle switch’. Meanwhile, DeepMind has developed an AI system so called “AlphaFold” predicts the 3D structure of proteins, including an estimated structure of 25 T2Rs in humans “https://www.proteinatlas.org/search/tas2r”. Docking simulation methods and/or molecular dynamics simulation methods, etc. can be used to calculate the binding mode and binding energy of T2R and the ligand polyphenol based on available information. These methods would reveal the partial structure of polyphenols and their respective affinities for bitter taste receptors. Subsequent results could be applied to the discovery of bitter-based medicines.
3. Astringent Taste Receptors and Polyphenols
3.1. Mechanisms of Astringent Taste Perception
Astringency is a taste that is specific to polyphenols, and bitter compounds can be either synthetic chemicals, such as pharmaceuticals, or natural products. Astringency is a sensory attribute that is often described as a drying, roughening, and puckering sensation in the mouth. Astringency is a common characteristic of anthocyanins, flavanols (including catechins, gallated catechins, and procyanidins), and hydrolyzed tannins [41,91]. Furthermore, it is a crucial factor in determining the quality of foods and drinks such as berries, red wine and chocolate. However, the mechanisms by which astringent polyphenol stimuli are recognized remain unclear. Taste refers to the sensation resulting from direct stimulation of taste receptors in the taste buds. The taste sensation is transmitted centrally via the sensory branches of the facial nerve (VII), the glossopharyngeal nerve (IX) or the vagus nerve (X) [92]. In contrast, somatosensation is transmitted by the general sensory branches of the trigeminal (V), glossopharyngeal (IX) or vagus (X) nerves [93]. Astringency, rather than taste, has been reported to be transmitted as somatosensory information via the trigeminal nerve [94]. The mucosa surrounding taste buds contains various somatosensory receptors, such as mechanoreceptors, temperature receptors, and nociceptors [95]. It is thought that humans perceive “flavor” through complex multisensory inputs in the oral cavity and olfactory inputs from the back of the nose that are transmitted to the CNS. Previous fMRI studies in humans have reported that the primary taste cortex, the insular cortex, is activated not only by sweet taste stimuli but also by somatosensory capsaicin and astringent taste stimuli, and that these three types of stimuli activate overlapping subregions [96]. Astringency has been reported to activate not only the insular cortex, a transient taste area, but also the superior orbitofrontal cortex, the cingulate cortex and frontal inferior triangularis with the intensity of activation being greater than that of sweet taste or capsaicin [96]. On the other hand, astringency was attributed to oral friction caused by the interaction of polyphenols with proline-rich proteins in saliva, which are recognized by mechanotropic receptors [41,42]. This theory is explained by a three-stage model in which a reversible bond between the hydrophobic face of the aromatic ring of the polyphenol and the pyrrolidine ring of the proline residue of the protein first forms a soluble complex, additional polyphenols cross-link these peptides to form a larger insoluble complex, which then aggregates further [97]. However, it is not known whether such complex reactions take place in the oral cavity in the short time between food ingestion and swallowing. It has also recently been reported that interactions with mucins in saliva may also be involved in the formation of astringent taste sensations [98]. But the target molecules that recognize this phenomenon are not at all clear. According to this theory, salivary proteins or mucins are essential for mammals to recognise astringent tastes. However, recent reports have shown that astringent polyphenols can activate GPCRs [94] or TRPs [99,100,101] and trigger the activity of the trigeminal nerve independently of saliva and mucins. In summary, the perception of astringency does not necessarily require the presence of saliva or mucin, suggesting that it is not a taste, but rather a type of somatosensory perception.
3.2. Astringent Polyphenols and Stress Response
Consumption of cocoa extracts, which are rich in flavanols, a type of astringent polyphenol, can significantly reduce risk of cardiovascular diseases [52]. A significant reduction in the number of cardiovascular events and deaths from cardiovascular disease was observed when elderly people were given an average of 500 mg of flavanols per day for 3.6 years. Repeated flavanols are also known to improve blood pressure a risk factors for cardiovascular disease [102,103]. Furthermore, a lot of intervention trials have demonstrated that two hours following a single intake of flavanol, there was a significant increase in blood flow-dependent vasorelaxation (FMD) levels [104]. There are also reports of an optimal dosage for these effects, known as hormesis, where the effects are attenuated at low or high doses [105,106,107]. The hormetic concept is based on the idea that low levels of stress up-regulate adaptive responses that not only precondition, repair and restore normal function to damaged tissues/organs, but also modestly over-compensate, reducing ongoing background damage [108,109,110]. We have focused on the results of this study and developed an evaluation system to reproduce this effect in experimental animals [111]. A marked increase in blood flow in skeletal muscle arterioles was observed immediately after ingestion of the cocoa-derived flavanol fraction in rats [112]. In addition, it was found that at doses generally taken by humans from food (10 mg/kg), this blood flow-increasing effect was observed, but at doses 10 times higher, the effect disappeared [113]. In experimental animals, hormesis was also observed, similar to that found in humans regarding hemodynamic alterations. It is widely recognized that exposure to stress in mammals leads to an increase in sympathetic nerve activity, causing alterations in the circulatory system [114,115]. There were several experiments conducted using adrenergic receptor inhibitors to investigate the relationship between these effects and the sympathetic nervous system. The results suggests that the rapid changes in hemodynamics after a single dose of flavanols were mediated by adrenergic receptors [113,116]. Furthermore, it has been suggested that the haemodynamic absence seen with high doses of flavanols compared to low doses was due to activation of α2 receptors, the central autoreceptor [113,116]. Similarly, single doses of anthocyanins, polyphenols with an astringent taste, showed circulatory changes via sympathoadrenergic receptors [117]. Increased sympathetic activity is also known to promote white to beige fat cell conversion and skeletal muscle protein formation [118]. After being administered a flavanol fraction from cocoa for two weeks, browning was shown in mice inguinal adipose tissue. This was characterized by polycytosis of fat, a marked reduction in adipocyte size, and increased expression of the heat-producing protein, uncoupling protein 1 (UCP-1) [119]. Repeated administration of the flavanol cinnamomycin A2 (A2) for two weeks also resulted in significant activation of the Akt/mTOR pathway and a marked increase in mean muscle cross-sectional area in the soleus muscle [120]. It has also been observed that after a single dose of flavanol, blood catecholamine levels were significantly increased [121] and urinary catecholamine excretion was also elevated [119]. Sympathetic nerve activity is known to be increased by stress. Examples include spicy substances such as capsaicin [122], temperature changes [123] and exercise [124]. It is known that when mammals are exposed to stress, activation of the hypothalamic-pituitary-adrenal axis (HPA) occurs along with hyperactivation of sympathetic nervous [125]. To confirm that the effects of astringent polyphenols on circulatory dynamics are a stress response response, we administered a single dose of flavanols and observed their effects on the HPA axis. The results showed a significant increase in the expression of the stress hormone corticotropin-releasing hormone (CRH) mRNA in the paraventricular nucleus of the mouse hypothalamus [126,127]. It was found that the duration of CRH mRNA expression onset shortened as the doses increased. A subsequent increase in blood cortisol (or corticosterone in rodents) levels was confirmed. The administration of astringent polyphenols leads to hyperactivation of sympathetic nervous and activation of the HPA axis in mammals, indicating that astringency is perceived as a stressor.
3.3. Astringent Polyphenols and TRP Channels
According to the research results to date, it seems that astringency is likely to be a somatic sensation [94,95]. Schöbel et al. reported that the sensation of astringency is not impaired in human subjects when the taste nerve is denervated or blocked by local anaesthesia. In addition, they showed that salty, sweet, sour and bitter tastes were almost completely lost in these subjects, and that subjects lost the sensation of astringency only when both trigeminal and gustatory nerves were blocked [94]. In 1997, it was demonstrated that the somatic sensation caused by capsaicin stimulation is recognized by Transient Receptor Potential V1 (TRPV1), which is one of the nociceptors [128]. In mammals, there are 28 different TRP channel proteins expressed, which are classified into seven subfamilies based on amino acid sequence homology [34]. TRP channels are involved in a variety of sensory responses, including heat, cold, pain, stress, vision and taste, and are activated by a number of stimuli. These channels are composed of six transmembrane polypeptide subunits that assemble as a tetramer to form a cation-permeable pore [129]. TRP vanilloid 1 and ankyrin 1 (TRPV1 and TRPA1) are expressed mainly in sensory neurons and are also distributed in the oral and gastrointestinal tracts [130,131]. It is widely recognized that capsaicin activates sympathoexcitation via the CNS by activating TRPV1 expressed on sensory nerves, thereby promoting heat production [132] and altering hemodynamics [133]. The astringent polyphenol flavanols have been observed to have the same effect described as above. Interactions between TRP channels and astringent polyphenols have been reported in several cellular experiments [99,100,101]. In Ca2+ imaging experiments, it has been reported that EGCG, an astringent and bitter polyphenol, activates the mouse enteroendocrine cell line STC-1. TRPA1 receptor inhibitors canceled this effect [100]. We investigated whether astringent polyphenols activate TRP channels and investigated the involvement of TRP channels in the effect of increasing blood flow in rats. The rats were administered flavanol, A2, and TRPV1 or TRPA1 channel inhibitors simultaneously through single oral doses. The marked increase in blood flow observed with A2 alone was eliminated by the combined use of TRP channel inhibitors [134]. These findings suggest that TRP channels contribute to the sympathetic hyperactivity of flavanols as a stress response. The interaction between A2 and each TRP channel was then observed using modeling simulation methods. The binding energies of the A2 and ligand binding sites for each TRP channel were relatively high. These results suggested that there was no direct interaction between them [134].
Polyphenols are stored in vacuoles within plants, which have a weakly acidic pH of 4-5, and are stable [135]. However, when consumed by mammals, they are exposed to neutral pH conditions in gut such as the mouth, small intestine, and large intestine. It is widely recognized that flavanols [136] and anthocyanins [137], which are astringent polyphenols, undergo rapid oxidation and decomposition or condensation under neutral pH. In our study, (-)-epicatechin (EC) and its tetramer, A2, also showed superoxide radical (O2・- ) scavenging activity at acidic pH but significantly enhanced O2・- production under neutral pH [134]. Furthermore, an in vitro study conducted on TRP channel-expressing HEK293 cells has demonstrated that oxidized EGCG, which was incubatied at neutral pH for several hours, significantly enhances calcium influx [101]. In contrast, freshly dissolved, non-oxidized EGCG showed no such effect. In measuring peripheral vascular blood flow in rats after co-administration of O2・- scavengers and A2, the blood flow increasing effect observed with A2 alone was nullified [134].
It was reported that TRPV1 and TRPA1 can be activated by reactive oxygen species (ROS) through cytoplasmic side cysteine residues [138]. In hTRPV1, Cys-258 is highly oxidizable, leading to the formation of a disulfide bond between Cys-258 and Cys-742, which opens the channel [139]. In TRPA1, mutational analysis has reported the presence of multiple reactive oxygen species domains, including Cys421, Cys621, Cys641 and Cys665 [140]. These findings suggest that orally ingested flavanols and anthocyanins produce ROS in the neutral pH environment of the gastrointestinal tract, which are oxidatively degraded by successive oxidative reactions in the molecule, and the produced ROS may interact with the ROS recognition domain of the TRP channel and become activated (Figure 2). Furthermore, the responses of ROS-TRP channels on sensory nerves could be perceived in the CNS as an astringent taste. However, further research is needed on how TRP channels are involved in the mechanism of astringent taste perception.
4. Bioavailability of Polyphenols
Numerous studies have been conducted on the bioavailability of polyphenols [141,142,143]. Although a proportion of ingested polyphenols are absorbed in the small intestine, the rate and total amount of absorption of polyphenols are known to be strongly influenced by their chemical structure. Compared to their bioavailability in humans, the urinary excretion of most polyphenols is a few percent, whereas the excretion of astringent polyphenols such as anthocyanins and flavanols is quite low [144]. Flavonoids as one of the main polyphenols are compounds with a 3,4-dihydro-2-phenyl-2H-1-benzopyran structure and there might be an inverse correlation between the number of hydroxyl groups in the B-ring and bioavailability. For example, in an intervention study, the urinary excretion rate of pelargonidin, an anthocyanin aglycon with a single hydroxyl group on the B-ring, was reported to be 1.8% [145]. In contrast, cyanidin, which has two hydroxyl groups, is around 0.1% [146]. Methylated flavonoids, which have methoxy groups on the B ring, are also known to be highly absorbable [147]. These differences in absorption may depend on the chemical stability of polyphenols in the neutral to slightly alkaline conditions of the small intestine. For example, cyanididins are readily degraded under neutral pH conditions to protocatechuic acid (PCA) and phloroglucinaldehyde (PGA), and it has been reported that blood concentrations of the degradation product PGA were four times higher than those of cyanidin [148]. Most polyphenolic compounds are known to be glucuronidated or sulfated by the small intestine or liver and exist as metabolites in the blood [54]. These metabolites have difficulty penetrating cell membranes because they are more water soluble. Ingested polyphenols pass through the upper gastrointestinal tract and reach the large intestine. Ingested polyphenols pass through the upper gastrointestinal tract and reach the large intestine. A part of ingested polyphenols is known to be degraded by intestinal bacteria in the colon. Several comprehensive reviews of these have been reported so far [149,150,151]. However, the amount and type of metabolic degradation of polyphenols by gut bacteria relative to the total polyphenols consumed, and the kinetics of this degradation, is not well understood. Unlike nutrients and easily absorbed chemicals, such as fat-soluble substances, ingested polyphenols are present in high concentrations in the oral cavity, stomach, small and large intestines. Therefore, until they are excreted from the body as feces, they keep in contact with I-, K- and L-cells that express bitter taste receptors, or with gastrointestinal sensory nerves and epithelial cells that express TRP channels for an extended period. Considering these behaviours in the gastrointestinal tract, it is likely that polyphenols exert their various physiological effects via sensory receptors expressed in the gastrointestinal tract.
5. Biological Regulation through Bitter and Astringency of Polyphenols
Polyphenols have been shown in epidemiological studies to have the potential to prevent cardiovascular disease, neurodegenerative diseases, and age-related sensory organ deterioration [46,47,48,49]. Large-scale intervention trial of flavanols have also reported reduced cardiovascular mortality [52] and hippocampus-dependent improvements in cognitive function [53]. As previously discussed, the nervous system including CNS plays a significant role in controlling obese, hyperglycaemia, dyslipidaemia, and hypertension, all of which are risk factors for cardiovascular disease [152]. However, it is important to note that polyphenols are poorly absorbed and, even if absorbed, are only present in the blood as metabolites. Therefore, it is unlikely that they can pass through the blood-brain barrier (BBB) and be distributed in the brain. There is no discussion of how bitterness and astringency contribute to these effects. In considering the mechanisms of these beneficial effects of polyphenols, we proposed to discuss the involvement of afferent and efferent nerve. The following is a review of the various physiological effects of polyphenols reported so far and how they contribute to bitter and astringent tastes.
5.1. Circulation and Polyphenol
The effects of cocoa astringent flavanol on the circulatory system are well studied: a meta-analysis of RCTs of adult participants at cardiometabolic risk reported between 2000 and 2021 observed a marked increase in FMD [153]. These activities on FMD of astringent anthocyanin [154] was also confirmed by the meta-analyses. On the other hand, for compounds without a pronounced taste, such as resveratrol [155] or isoflavones [156], the onset of action has been reported to depend on the study design. There are few reports of other polyphenol enhancement in FMD, it is because many experiments have been conducted with mixtures, therefore the main compounds cannot be limited. FMD-enhancing effect of a single dose of flavanols shows a non-linear hormesis response [106,107]. Anthocyanins, [105,157] or flavanol [105] have been reported to be more effective on FMD in single oral doses compared to repeated doses. In addition, repeated intake of these polyphenols is also well known to affect blood pressure [158]. In particular, meta-analyses have shown that repeated intake of flavanols significantly reduces systolic and diastolic blood pressure [54,159].
In experimental animal studies, a single oral administration of flavanols or anthocyanins also results in a significant increase in blood flow of skeletal muscle arteriole [112,113,117]. Furthermore, repeated administration of these chemicals considerably decreased the mean blood pressure in rodent [160,161]. These modifications are consistent with the changes that occur in the circulatory system as a result of exercise (Figure 3).
In brief, exercise induces acute changes in the microcirculation, which consequently leads to changes in the systemic circulation [159]. Exercise increases skeletal muscle blood flow through hyperactivation of the sympathetic nervous system, subsequently inducing shear stress on vascular endothelial cells [162]. Mechanosensors consisting of vascular endothelial growth factor (VEGF) receptor 2, vascular endothelial (VE) cadherin and CD31 (platelet endothelial cell adhesion molecule-1, PECAM-1) expressed on endothelial cells respond to shear stress and release vasorelaxing factor NO via the Akt /eNOS pathway, causing the vessel to relax [163]. (Figure 3a). The rise in FMD resulting from astringent polyphenols observed in intervention trials may be attributed to these alterations in hemodynamics. In addition, these alteration leads to the differentiation of vascular endothelial cells via mechanosensors, With the repetition of the exercise, these changes in hemodynamics are also repeated and are known to trigger the formation of new capillaries [164]. Continuous fluid shear stress angiogenesis through the cellular production of proangiogenic growth factors, such as VEGF. Resting endothelial cells are transformed into tip cells by the secretion of angiogenesis-promoting factors from a hypoxic or metabolically active microenvironment [165]. These tip cells proliferate to become stalk cells, which extend a growing sprout and form a lumen. Once the two spraut fuse, they form a capillary tube, which lowers the resistance to blood flow, and the endothelial cells return to quiescence. In this quiescent state, endothelial cells produce mechano-sensing complexes and form tight junctions [166] (Figure 3b). These factors lead to angiogenesis, which is an increase in blood vessel volume and a decrease in blood pressure.
In rodent studies, a single administration of astringent polyphenols induced phosphorylation of eNOS in the aorta [112,113,117,161,167]. Moreover, repeated administration of astringent polyphenols to rodents has been shown to lower blood pressure. Furthermore, repeated administration reduced blood pressure and significantly increased CD31 expression, a marker of neovascularization, in the soleus [161]. This change suggested the promotion of angiogenesis. This process of angiogenesis increases vascular capacity, reducing the risk of exposure to shear stress. Therefore, repeated ingestion of astringent polyphenols might result in a decrease in blood pressure along with a reduction in the FMD response.
5.2. Blood Glucose and Polyphenols
Interest in using bitter compounds to improve glucose tolerance by stimulating gastrointestinal hormone secretion has recently increased [78,79]. Most polyphenols have a bitter taste and low bioavailability, which makes them potentially useful for this objective. For example, interventional studies have been conducted on changes in glucose tolerance and blood levels of incretin after single doses of green tea flavanol [168], coffee chlorogenic acid [169,170] or combinations thereof [168], olive oleuropein [171] and anthocyanin fraction from blackcurrant [172] in healthy subjects. Intake of 540 mg of green tea flavanol reduced the increase in blood glucose levels caused by a high-fat, high-carbohydrate diet, increased blood levels of GLP-1 and reduced GIP levels [168]. In addition, when 355 mg of chlorogenic acid was given to healthy subjects, an increase in blood levels of GLP-1 was only observed in subjects with a low insulinogenic index [170]. These combinations (beverages containing 540 mg of green tea flavanols and 150 mg of chlorogenic acid) have also been reported to reduce the rise in blood glucose, increase blood GLP-1 levels and reduce GIP levels after consumption of high-fat, high-carbohydrate cookies [168]. A single oral dose of oleuropein (20 mg) decreased blood glucose and dipeptidyl peptidase (DPP)-4 levels, which inactivates active GLP-1, and increased blood levels of GLP-1 and insulin 2 hours after consumption of the test meal [171]. These findings suggest that a single ingestion of bitter polyphenols may increase incretin secretion from gastrointestinal secretory cells, promote active incretin retention, induce insulin secretion, and suppress meal-derived blood glucose levels after a few hours. Improvements in glucose tolerance as measured by the homeostatic model assessment for insulin resistance (HOMA-IR) have been reported in studies investigating repeated administration of green tea flavanol, either alone [173] or in combination with chlorogenic acid [174], pine bark flavanol [175], acacia flavanols [176], red wine polyphenol [177], eriocitrin [178], and curcumin [179]. However, reports were suggesting that repeated intake of resveratrol can improve insulin resistance [180], or that no benefit is observed [181]. Further research is needed to determine the specific types and amounts of polyphenols that are effective in improving glucose tolerance. Yusoff et al. showed that the antidiabetic effect can depend on the chemical structure of the flavonoid [182]. It was reported that flavonoids with antidiabetic activity should have a C2-C3 double bond (C ring) in both the A and B rings of the flavonoid skeleton and hydroxyl groups in the C3’, C4’, C5 and C7 positions (Figure 1a). The study results demonstrate that the beneficial activity of polyphenols is highly variable, depending on the chemical structure. Therefore, for drug discovery, studies that elucidate the subchemical structures of polyphenols essential for T2R activation are crucial.
5.3. Obesity and Polyphenols
It has been suggested that consuming polyphenols may help to control obesity [183,184]. A meta-analysis of clinical trials published between 2010 and 2021 showed that polyphenols produced statistically significant reductions in body weight, BMI and abdominal circumference, but no significant reduction in body fat. Subgroup analyses showed that polyphenol intake had significant effects in subjects under 50 years of age, in the Asian population and in patients with obesity-related health problems, for more than three months and at doses of around 220 mg d-[185]. To date, the anti-obesity effects of polyphenols have been attributed to (1) appetite suppression, (2) reduced digestion and absorption of lipids and carbohydrates through inhibition of digestive enzymes, (3) adipocyte differentiation, (4) regulation of lipid metabolism, (5) stimulation of energy expenditure, (6) improvement of intestinal microflora, (7) amelioration of obesity-related mild inflammation; and (8) reduction of oxidative stress [186,187,188]. However, it should be noted that the bioavailability of polyphenols is very low. While they may inhibit digestive enzymes and improve gut microbiota, other effects are likely to be secondary. As mentioned above, bitter substances such as polyphenols may reduce energy intake due to suppressed appetite via the secretion of gastrointestinal hormones, resulting in vagus nervous stimulation [24,69]. Polyphenols appear to have a similar effect to GLP-1 analogues in weight loss by suppressing appetite and reducing gastric emptying [189,190]. Increased sympathetic activity after ingestion of astringent polyphenols is thought to cause fat browning as confirmed in the study using rodent [119]. In addition, this promotion of lipid oxidation induced by the consumption of astringent polyphenols has also been shown to increase HDL cholesterol levels in the blood [191,192]. Many of the hypotheses proposed as mechanisms for the anti-obesity effects of polyphenols can be explained by their bitter or astringent taste.
5.4. Brain Function and Polyphenol
Gastrointestinal hormones secreted by the ingestion of polyphenols have been reported to activate the vagus nerve as a neurotransmitter [193]. Gastrointestinal hormones are secreted by neuropodal cells in response to bitter, sweet and umami taste stimuli, and this sensory information is transmitted directly to the mNTS within milliseconds [194,195,196]. Upstream of the NTS, it projects to multiple brain regions, as well as transsynaptically to the dorsal hippocampus [197] and frontal cortex [198]. These sensory transmissions have been shown to improve mood and memory in studies using severed gastrointestinal vagus nerves [194]. Recently, results from intervention studies have shown that stimulating the vagus nerve can help depression recovery [199]. The bitterness of polyphenols may play a role in the homeostasis of the brain.
Polyphenols with a markedly astringent taste have been shown to trigger the stress response and to activate the hypothalamic-pituitary-adrenal (HPA) axis with an increase in sympathetic activity [126,127]. Stress causes CRH to be secreted from the paraventricular nucleus of the hypothalamus into the pituitary portal system, stimulating the anterior pituitary to release adrenocorticotropic hormone (ACTH) [200]. ACTH subsequently circulates to the adrenal glands and stimulates the release of cortisol into the blood [201]. Cortisol readily crosses BBB and binds to receptors in the amygdala, prefrontal cortex and hippocampus [202]. Chronic stress is an allosteric load and excess cortisol is released by the adrenal glands [203]. The resulting prolonged activation of neuroendocrine, cardiovascular and emotional responses can be damaging to health with increased cardiovascular risk, cognitive dysfunction and depressive mood [204]. On the other hand, exercise has also been reported to activate the HPA axis and increase blood cortisol levels, despite its benefits for cognition, and mood and promoting adult neurogenesis [205,206]. A cortisol paradox between chronic stress and exercise on brain function has been observed [207,208,209]. The reason for this paradox is unclear, but one possibility is the involvement of cortisol-glucocorticoid receptor (GR)-dopamine (DA)-dopamine D2 receptors [210]; Cortisol secreted into the blood activates GR in medial prefrontal cortex and enhances glutamatergic input from mPFC to ventral tegmental area (VTA). This input to the VTA enhances the projection of DA neurons from the VTA to the mPFC. This increase in DA in the mPFC activates GABAergic neurons in the nucleus of the anterior nucleus of the terminal line (aBNST), which project to the paraventricular nucleus (PVH) of the hypothalamus. It provides negative feedback control and prevents over-reaction of the HPA axis [210]. These mechanisms suggest that exercise may have the effect of reducing excessive HPA activation and improving mood and memory function, despite increasing basal levels of glucocorticoids. In a large intervention study in elderly subjects, 3.6 years of cocoa extract, mainly composed of astringent polyphenols, was reported to restore hippocampus-dependent memory in participants in the lower tertile of habitual diet quality or flavanol intake [53]. In rodents, repeated administration of highly astringent catechin tetramers A2 has also been shown to improve spatial memory with enhanced adult neurogenesis [211]. In summary, astringent stimulation may benefit brain function via the GR-DA-DAR pathway as well as exercise, but much more research is needed to elucidate the mechanism.
6. Conclusion
The consumption of polyphenols id beneficial for the maintenance and promotion of human health. However, the mechanisms remain unclear. As most polyphenols have a bitter taste, migrate from the oral cavity to the gastrointestinal tract and interact with intestinal secretory cells, they likely regulate sugar metabolism or feeding via the T2R. On the other hand, if we consider the effects of astringent polyphenols on the circulatory system, metabolism and brain function, their effects have a great deal in common with the benefits of exercise. Astringency, which is considered a stressor, is believed to have beneficial effects due to hormesis. Research on the bio-modulation of polyphenols with taste, which has not received much attention to date, may provide a solution to the polyphenol paradox, in which polyphenols exert bio-regulatory effects despite their extremely low bioavailability.
Author Contributions
Conceptualization, N.O. and V.C.; Drafting of the manuscript. N.O., T.S F.T. F.Y..; Review and editing, N.O. and V.C.;Funding acquisition, N.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by JSPS KAKENHI Grant Number JP 23H02166.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reed, D.R.; Mainland, J.D.; Arayata, C.J. Sensory nutrition: The role of taste in the reviews of commercial food products. Physiology & behavior 2019, 209, 112579. [Google Scholar]
- Reed, D.R.; Alhadeff, A.L.; Beauchamp, G.K.; Chaudhari, N.; Duffy, V.B.; Dus, M.; Fontanini, A.; Glendinning, J.I.; Green, B.G.; Joseph, P.V.; et al. NIH Workshop Report: sensory nutrition and disease. The American journal of clinical nutrition 2021, 113, 232–245. [Google Scholar] [CrossRef]
- Lindemann, B. Receptors and transduction in taste. Nature 2001, 413, 219–225. [Google Scholar] [CrossRef]
- Lee, S.J.; Depoortere, I.; Hatt, H. Therapeutic potential of ectopic olfactory and taste receptors. Nat Rev Drug Discov 2019, 18, 116–138. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, R.; Aier, I.; Semwal, R.; Tyagi, P.; Varadwaj, P. Sense of Smell: Structural, Functional, Mechanistic Advancements and Challenges in Human Olfactory Research. Current neuropharmacology 2019, 17, 891–911. [Google Scholar] [CrossRef]
- Kato, A.; Touhara, K. Mammalian olfactory receptors: pharmacology, G protein coupling and desensitization. Cell Mol Life Sci 2009, 66, 3743–3753. [Google Scholar] [CrossRef]
- Spehr, M.; Munger, S.D. Olfactory receptors: G protein-coupled receptors and beyond. J Neurochem 2009, 109, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Dubovski, N.; Fierro, F.; Margulis, E.; Ben Shoshan-Galeczki, Y.; Peri, L.; Niv, M.Y. Taste GPCRs and their ligands. Prog Mol Biol Transl Sci 2022, 193, 177–193. [Google Scholar]
- Chaudhari, N.; Roper, S.D. The cell biology of taste. J Cell Biol 2010, 190, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Roper, S.D. TRPs in taste and chemesthesis. Handbook of experimental pharmacology 2014, 223, 827–871. [Google Scholar]
- Zufall, F. TRPs in olfaction. Handbook of experimental pharmacology 2014, 223, 917–933. [Google Scholar] [PubMed]
- Green, B.G. Chemesthesis and the chemical senses as components of a "chemofensor complex". Chem Senses 2012, 37, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Guichard, E.; Barba, C.; Thomas-Danguin, T.; Tromelin, A. Multivariate Statistical Analysis and Odor-Taste Network To Reveal Odor-Taste Associations. Journal of agricultural and food chemistry 2020, 68, 10318–10328. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M.; Prescott, J. Odor/taste integration and the perception of flavor. Exp Brain Res 2005, 166, 345–357. [Google Scholar] [CrossRef]
- Tong, T.; Wang, Y.; Kang, S.G.; Huang, K. Ectopic Odorant Receptor Responding to Flavor Compounds: Versatile Roles in Health and Disease. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, O.; Drago, F. Pharmacological significance of extra-oral taste receptors. Eur J Pharmacol 2021, 910, 174480. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, M.; Libert, F.; Schurmans, S.; Schiffmann, S.; Lefort, A.; Eggerickx, D.; Ledent, C.; Mollereau, C.; Gérard, C.; Perret, J.; et al. Expression of members of the putative olfactory receptor gene family in mammalian germ cells. Nature 1992, 355, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Griffin, C.A.; Kafadar, K.A.; Pavlath, G.K. MOR23 promotes muscle regeneration and regulates cell adhesion and migration. Dev Cell 2009, 17, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Park, J.; Moon, C.; Park, T. Regulation of Adipogenesis and Thermogenesis through Mouse Olfactory Receptor 23 Stimulated by α-Cedrene in 3T3-L1 Cells. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Giusepponi, M.E.; Kern, M.; Chakaroun, R.; Wohland, T.; Kovacs, P.; Dietrich, A.; Schön, M.R.; Krohn, K.; Pucci, M.; Polidori, C.; et al. Gene expression profiling in adipose tissue of Sprague Dawley rats identifies olfactory receptor 984 as a potential obesity treatment target. Biochem Biophys Res Commun 2018, 505, 801–806. [Google Scholar] [CrossRef]
- Bellono, N.W.; Bayrer, J.R.; Leitch, D.B.; Castro, J.; Zhang, C.; O’Donnell, T.A.; Brierley, S.M.; Ingraham, H.A.; Julius, D. Enterochromaffin Cells Are Gut Chemosensors that Couple to Sensory Neural Pathways. Cell 2017, 170, 185–198.e16. [Google Scholar] [CrossRef] [PubMed]
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral and extraoral tissues. Current opinion in clinical nutrition and metabolic care 2014, 17, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Zhai, K.; Yang, Z.; Zhu, X.; Nyirimigabo, E.; Mi, Y.; Wang, Y.; Liu, Q.; Man, L.; Wu, S.; Jin, J.; et al. Activation of bitter taste receptors (tas2rs) relaxes detrusor smooth muscle and suppresses overactive bladder symptoms. Oncotarget 2016, 7, 21156–21167. [Google Scholar] [CrossRef] [PubMed]
- Xie, C. Role of Intestinal Bitter Sensing in Enteroendocrine Hormone Secretion and Metabolic Control. Frontiers in endocrinology (Lausanne) 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ansoleaga, B.; Garcia-Esparcia, P.; Pinacho, R.; Haro, J.M.; Ramos, B.; Ferrer, I. Decrease in olfactory and taste receptor expression in the dorsolateral prefrontal cortex in chronic schizophrenia. J Psychiatr Res 2015, 60, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Blank, K.; See Hoe, L.E.; Behrens, M.; Meyerhof, W.; Peart, J.N.; Thomas, W.G. Bitter taste receptor agonists elicit G-protein-dependent negative inotropy in the murine heart. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 2014, 28, 4497–4508. [Google Scholar] [CrossRef] [PubMed]
- Tizzano, M.; Finger, T.E. Chemosensors in the nose: guardians of the airways. Physiology (Bethesda) 2013, 28, 51–60. [Google Scholar] [CrossRef]
- Xu, J.; Cao, J.; Iguchi, N.; Riethmacher, D.; Huang, L. Functional characterization of bitter-taste receptors expressed in mammalian testis. Mol Hum Reprod 2013, 19, 17–28. [Google Scholar] [CrossRef]
- Lund, T.C.; Kobs, A.J.; Kramer, A.; Nyquist, M.; Kuroki, M.T.; Osborn, J.; Lidke, D.S.; Low-Nam, S.T.; Blazar, B.R.; Tolar, J. Bone Marrow Stromal and Vascular Smooth Muscle Cells Have Chemosensory Capacity via Bitter Taste Receptor Expression. PloS one 2013, 8, e58945. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: chemical properties, biological activities, and synthesis. Angew Chem Int Ed Engl 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Vancleef, L.; Van Den Broeck, T.; Thijs, T.; Steensels, S.; Briand, L.; Tack, J.; Depoortere, I. Chemosensory signalling pathways involved in sensing of amino acids by the ghrelin cell. Sci Rep 2015, 5, 15725. [Google Scholar] [CrossRef]
- Li, F. Taste perception: from the tongue to the testis. Mol Hum Reprod 2013, 19, 349–360. [Google Scholar] [CrossRef]
- Hill, M.A.; Sowers, J.R. Mineralocorticoid antagonists and ENaC inhibitors in hyperaldosteronism. Journal of clinical hypertension (Greenwich, Conn.) 2019, 21, 929–931. [Google Scholar] [CrossRef]
- Fallah, H.P.; Ahuja, E.; Lin, H.; Qi, J.; He, Q.; Gao, S.; An, H.; Zhang, J.; Xie, Y.; Liang, D. A Review on the Role of TRP Channels and Their Potential as Drug Targets_An Insight Into the TRP Channel Drug Discovery Methodologies. Frontiers in pharmacology 2022, 13, 914499. [Google Scholar] [CrossRef]
- Emery, E.C.; Diakogiannaki, E.; Gentry, C.; Psichas, A.; Habib, A.M.; Bevan, S.; Fischer, M.J.; Reimann, F.; Gribble, F.M. Stimulation of GLP-1 secretion downstream of the ligand-gated ion channel TRPA1. Diabetes 2015, 64, 1202–1210. [Google Scholar] [CrossRef]
- Mayer, F.; Gunawan, A.L.; Tso, P.; Aponte, G.W. Glucagon-like peptide 1 and glucose-dependent insulinotropic polypeptide stimulate release of substance P from TRPV1- and TRPA1-expressing sensory nerves. Am J Physiol Gastrointest Liver Physiol 2020, 319, G23–g35. [Google Scholar] [CrossRef]
- Arts, I.C.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. The American journal of clinical nutrition 2005, 81, 317s–325s. [Google Scholar] [CrossRef]
- Dabas, D. Polyphenols as Colorants. Advances in Food Technology and Nutritional Sciences - Open Journal 2018, SE, S1–S6. [Google Scholar]
- Soares, S.; Kohl, S.; Thalmann, S.; Mateus, N.; Meyerhof, W.; De Freitas, V. Different phenolic compounds activate distinct human bitter taste receptors. Journal of agricultural and food chemistry 2013, 61, 1525–1533. [Google Scholar] [CrossRef]
- Soares, S.; Silva, M.S.; García-Estevez, I.; Groβmann, P.; Brás, N.; Brandão, E.; Mateus, N.; de Freitas, V.; Behrens, M.; Meyerhof, W. Human Bitter Taste Receptors Are Activated by Different Classes of Polyphenols. Journal of agricultural and food chemistry 2018, 66, 8814–8823. [Google Scholar] [CrossRef]
- Soares, S.; Brandão, E.; Guerreiro, C.; Soares, S.; Mateus, N.; de Freitas, V. Tannins in Food: Insights into the Molecular Perception of Astringency and Bitter Taste. Molecules (Basel, Switzerland) 2020, 25. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Quijada-Morín, N.; Brás, N.F.; Gomes, P.; de Freitas, V.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Characterization of Sensory Properties of Flavanols - A Molecular Dynamic Approach. Chem Senses 2015, 40, 381–390. [Google Scholar] [CrossRef]
- Labbe, D.; Damevin, L.; Vaccher, C.; Morgenegg, C.; Martin, N. Modulation of perceived taste by olfaction in familiar and unfamiliar beverages. Food Quality and Preference 2006, 17, 582–589. [Google Scholar] [CrossRef]
- Mattes, R.D. Influences on acceptance of bitter foods and beverages. Physiology & behavior 1994, 56, 1229–1236. [Google Scholar]
- Stein, L.J.; Nagai, H.; Nakagawa, M.; Beauchamp, G.K. Effects of repeated exposure and health-related information on hedonic evaluation and acceptance of a bitter beverage. Appetite 2003, 40, 119–129. [Google Scholar] [CrossRef]
- Dugo, L.; Tripodo, G.; Santi, L.; Fanali, C. Cocoa Polyphenols: Chemistry, Bioavailability and Effects on Cardiovascular Performance. Curr Med Chem 2018, 25, 4903–4917. [Google Scholar] [CrossRef]
- Zhang, X.; Molsberry, S.A.; Yeh, T.S.; Cassidy, A.; Schwarzschild, M.A.; Ascherio, A.; Gao, X. Intake of Flavonoids and Flavonoid-Rich Foods and Mortality Risk Among Individuals With Parkinson Disease: A Prospective Cohort Study. Neurology 2022, 98, e1064–e1076. [Google Scholar] [CrossRef]
- Holland, T.M.; Agarwal, P.; Wang, Y.; Leurgans, S.E.; Bennett, D.A.; Booth, S.L.; Morris, M.C. Dietary flavonols and risk of Alzheimer dementia. Neurology 2020, 94, e1749–e1756. [Google Scholar] [CrossRef]
- Tang, D.; Tran, Y.; Shekhawat, G.S.; Gopinath, B. Dietary Flavonoid Intake and Chronic Sensory Conditions: A Scoping Review. Antioxidants (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef]
- Imamura, F.; Schulze, M.B.; Sharp, S.J.; Guevara, M.; Romaguera, D.; Bendinelli, B.; Salamanca-Fernández, E.; Ardanaz, E.; Arriola, L.; Aune, D.; et al. Estimated Substitution of Tea or Coffee for Sugar-Sweetened Beverages Was Associated with Lower Type 2 Diabetes Incidence in Case-Cohort Analysis across 8 European Countries in the EPIC-InterAct Study. The Journal of nutrition 2019, 149, 1985–1993. [Google Scholar] [CrossRef]
- Ma, L.; Hu, Y.; Alperet, D.J.; Liu, G.; Malik, V.; Manson, J.E.; Rimm, E.B.; Hu, F.B.; Sun, Q. Beverage consumption and mortality among adults with type 2 diabetes: prospective cohort study. BMJ (Clinical research ed.) 2023, 381, e073406. [Google Scholar] [CrossRef]
- Sesso, H.D.; Manson, J.E.; Aragaki, A.K.; Rist, P.M.; Johnson, L.G.; Friedenberg, G.; Copeland, T.; Clar, A.; Mora, S.; Moorthy, M.V.; et al. Effect of cocoa flavanol supplementation for prevention of cardiovascular disease events: The COSMOS randomized clinical trial. The American journal of clinical nutrition 2022. [Google Scholar]
- Brickman, A.M.; Yeung, L.K.; Alschuler, D.M.; Ottaviani, J.I.; Kuhnle, G.G.C.; Sloan, R.P.; Luttmann-Gibson, H.; Copeland, T.; Schroeter, H.; Sesso, H.D.; et al. Dietary flavanols restore hippocampal-dependent memory in older adults with lower diet quality and lower habitual flavanol consumption. Proceedings of the National Academy of Sciences of the United States of America 2023, 120, e2216932120. [Google Scholar] [CrossRef]
- Osakabe, N.; Terao, J. Possible mechanisms of postprandial physiological alterations following flavan 3-ol ingestion. Nutr Rev 2018, 76, 174–186. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary Polyphenol, Gut Microbiota, and Health Benefits. Antioxidants (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Frontiers in nutrition 2019, 6, 188. [Google Scholar] [CrossRef]
- Yan, J.; Tong, H. An overview of bitter compounds in foodstuffs: Classifications, evaluation methods for sensory contribution, separation and identification techniques, and mechanism of bitter taste transduction. Compr Rev Food Sci Food Saf 2023, 22, 187–232. [Google Scholar] [CrossRef]
- Brockhoff, A.; Behrens, M.; Niv, M.Y.; Meyerhof, W. Structural requirements of bitter taste receptor activation. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 11110–11115. [Google Scholar] [CrossRef]
- Meyerhof, W. Elucidation of mammalian bitter taste. Reviews of physiology, biochemistry and pharmacology 2005, 154, 37–72. [Google Scholar]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Frontiers in pharmacology 2020, 11, 587664. [Google Scholar] [CrossRef]
- Lossow, K.; Hübner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive Analysis of Mouse Bitter Taste Receptors Reveals Different Molecular Receptive Ranges for Orthologous Receptors in Mice and Humans. J Biol Chem 2016, 291, 15358–15377. [Google Scholar] [CrossRef]
- Wooding, S.P.; Ramirez, V.A.; Behrens, M. Bitter taste receptors: Genes, evolution and health. Evol Med Public Health 2021, 9, 431–447. [Google Scholar] [CrossRef]
- Ye, L.; Liddle, R.A. Gastrointestinal hormones and the gut connectome. Curr Opin Endocrinol Diabetes Obes 2017, 24, 9–14. [Google Scholar] [CrossRef]
- Clark, A.A.; Liggett, S.B.; Munger, S.D. Extraoral bitter taste receptors as mediators of off-target drug effects. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 2012, 26, 4827–4831. [Google Scholar] [CrossRef]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes Metab 2018, 20 Suppl 1, 5–21. [Google Scholar] [CrossRef]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Pfeiffer, A.F.H. The evolving story of incretins (GIP and GLP-1) in metabolic and cardiovascular disease: A pathophysiological update. Diabetes Obes Metab 2021, 23 Suppl 3, 5–29. [Google Scholar] [CrossRef]
- Xie, C.; Wang, X.; Young, R.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Role of Intestinal Bitter Sensing in Enteroendocrine Hormone Secretion and Metabolic Control. Frontiers in endocrinology 2018, 9, 576. [Google Scholar] [CrossRef]
- Miller, L.J.; Harikumar, K.G.; Wootten, D.; Sexton, P.M. Roles of Cholecystokinin in the Nutritional Continuum. Physiology and Potential Therapeutics. Frontiers in endocrinology 2021, 12, 684656. [Google Scholar] [CrossRef]
- Tack, J.; Verbeure, W.; Mori, H.; Schol, J.; Van den Houte, K.; Huang, I.H.; Balsiger, L.; Broeders, B.; Colomier, E.; Scarpellini, E.; et al. The gastrointestinal tract in hunger and satiety signalling. United European Gastroenterol J 2021, 9, 727–734. [Google Scholar] [CrossRef]
- Kim, W.; Egan, J.M. The role of incretins in glucose homeostasis and diabetes treatment. Pharmacological reviews 2008, 60, 470–512. [Google Scholar] [CrossRef]
- Holst, J.J.; Gasbjerg, L.S.; Rosenkilde, M.M. The Role of Incretins on Insulin Function and Glucose Homeostasis. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- Zhao, T.C. Glucagon-like peptide-1 (GLP-1) and protective effects in cardiovascular disease: a new therapeutic approach for myocardial protection. Cardiovasc Diabetol 2013, 12, 90. [Google Scholar] [CrossRef]
- Christensen, M.; Bagger, J.I.; Vilsbøll, T.; Knop, F.K. The alpha-cell as target for type 2 diabetes therapy. Rev Diabet Stud 2011, 8, 369–381. [Google Scholar] [CrossRef]
- Deacon, C.F.; Ahrén, B. Physiology of incretins in health and disease. Rev Diabet Stud 2011, 8, 293–306. [Google Scholar] [CrossRef]
- Rezaie, P.; Bitarafan, V.; Rose, B.D.; Lange, K.; Mohammadpour, Z.; Rehfeld, J.F.; Horowitz, M.; Feinle-Bisset, C. Effects of Quinine on the Glycaemic Response to, and Gastric Emptying of, a Mixed-Nutrient Drink in Females and Males. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Rose, B.D.; Bitarafan, V.; Rezaie, P.; Fitzgerald, P.C.E.; Horowitz, M.; Feinle-Bisset, C. Comparative Effects of Intragastric and Intraduodenal Administration of Quinine on the Plasma Glucose Response to a Mixed-Nutrient Drink in Healthy Men: Relations with Glucoregulatory Hormones and Gastric Emptying. The Journal of nutrition 2021, 151, 1453–1461. [Google Scholar] [CrossRef]
- Verbeure, W.; Deloose, E.; Tóth, J.; Rehfeld, J.F.; Van Oudenhove, L.; Depoortere, I.; Tack, J. The endocrine effects of bitter tastant administration in the gastrointestinal system: intragastric versus intraduodenal administration. American journal of physiology. Endocrinology and metabolism 2021, 321, E1–e10. [Google Scholar] [CrossRef]
- Koh, G.Y.; Rowling, M.J.; Pritchard, S.K. Possible role of type 1 and type 2 taste receptors on obesity-induced inflammation. Nutr Rev 2022, 80, 1919–1926. [Google Scholar] [CrossRef]
- Medapati, M.R.; Bhagirath, A.Y.; Singh, N.; Chelikani, P. Pharmacology of T2R Mediated Host-Microbe Interactions. Handbook of experimental pharmacology 2022, 275, 177–202. [Google Scholar]
- Li, W.; Chen, H.; Xu, B.; Wang, Y.; Zhang, C.; Cao, Y.; Xing, X. Research progress on classification, sources and functions of dietary polyphenols for prevention and treatment of chronic diseases. Journal of Future Foods 2023, 3, 289–305. [Google Scholar] [CrossRef]
- Tarragon, E.; Moreno, J.J. Polyphenols and taste 2 receptors. Physiological, pathophysiological and pharmacological implications. Biochemical pharmacology 2020, 178, 114086. [Google Scholar] [CrossRef]
- Behrens, M.; Brockhoff, A.; Batram, C.; Kuhn, C.; Appendino, G.; Meyerhof, W. The human bitter taste receptor hTAS2R50 is activated by the two natural bitter terpenoids andrographolide and amarogentin. Journal of agricultural and food chemistry 2009, 57, 9860–9866. [Google Scholar] [CrossRef] [PubMed]
- Roland, W.S.; Vincken, J.P.; Gouka, R.J.; van Buren, L.; Gruppen, H.; Smit, G. Soy isoflavones and other isoflavonoids activate the human bitter taste receptors hTAS2R14 and hTAS2R39. Journal of agricultural and food chemistry 2011, 59, 11764–11771. [Google Scholar] [CrossRef]
- Narukawa, M.; Noga, C.; Ueno, Y.; Sato, T.; Misaka, T.; Watanabe, T. Evaluation of the bitterness of green tea catechins by a cell-based assay with the human bitter taste receptor hTAS2R39. Biochem Biophys Res Commun 2011, 405, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Narukawa, M.; Mochizuki, M.; Misaka, T.; Watanabe, T. Activation of the hTAS2R14 human bitter-taste receptor by (-)-epigallocatechin gallate and (-)-epicatechin gallate. Bioscience, biotechnology, and biochemistry 2013, 77, 1981–1983. [Google Scholar] [CrossRef] [PubMed]
- Intelmann, D.; Batram, C.; Kuhn, C.; Haseleu, G.; Meyerhof, W.; Hofmann, T. Three TAS2R Bitter Taste Receptors Mediate the Psychophysical Responses to Bitter Compounds of Hops (Humulus lupulus L.) and Beer. Chemosensory Perception 2009, 2, 118–132. [Google Scholar] [CrossRef]
- Roland, W.S.; van Buren, L.; Gruppen, H.; Driesse, M.; Gouka, R.J.; Smit, G.; Vincken, J.P. Bitter taste receptor activation by flavonoids and isoflavonoids: modeled structural requirements for activation of hTAS2R14 and hTAS2R39. Journal of agricultural and food chemistry 2013, 61, 10454–10466. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Misaka, T.; Ishiguro, M.; Masuda, K.; Sugawara, T.; Ito, K.; Kobayashi, T.; Matsuo, S.; Ishimaru, Y.; Asakura, T.; et al. Characterization of the beta-D-glucopyranoside binding site of the human bitter taste receptor hTAS2R16. J Biol Chem 2010, 285, 28373–28378. [Google Scholar] [CrossRef]
- Xu, W.; Wu, L.; Liu, S.; Liu, X.; Cao, X.; Zhou, C.; Zhang, J.; Fu, Y.; Guo, Y.; Wu, Y.; et al. Structural basis for strychnine activation of human bitter taste receptor TAS2R46. Science 2022, 377, 1298–1304. [Google Scholar] [CrossRef]
- Cheynier, V. Polyphenols in foods are more complex than often thought. The American journal of clinical nutrition 2005, 81, 223s–229s. [Google Scholar] [CrossRef]
- Gibbons, J.R.; Sadiq, N.M. Neuroanatomy, Neural Taste Pathway. In StatPearls, StatPearls Publishing Copyright © 2023; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Huang, R.; Xu, C. An overview of the perception and mitigation of astringency associated with phenolic compounds. Compr Rev Food Sci Food Saf 2021, 20, 1036–1074. [Google Scholar] [CrossRef] [PubMed]
- Schöbel, N.; Radtke, D.; Kyereme, J.; Wollmann, N.; Cichy, A.; Obst, K.; Kallweit, K.; Kletke, O.; Minovi, A.; Dazert, S.; et al. Astringency is a trigeminal sensation that involves the activation of G protein-coupled signaling by phenolic compounds. Chem Senses 2014, 39, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, O.G. Gastrophysics of the Oral Cavity. Curr Pharm Des 2016, 22, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Kishi, M.; Sadachi, H.; Nakamura, J.; Tonoike, M. Functional magnetic resonance imaging investigation of brain regions associated with astringency. Neurosci Res 2017, 122, 9–16. [Google Scholar] [CrossRef] [PubMed]
- García-Estévez, I.; Ramos-Pineda, A.M.; Escribano-Bailón, M.T. Interactions between wine phenolic compounds and human saliva in astringency perception. Food Funct 2018, 9, 1294–1309. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rochera, B.; Manjón, E.; Escribano-Bailón, M.T.; García-Estévez, I. Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Kurogi, M.; Saitoh, O. The diversity in sensitivity of TRPA1 and TRPV1 of various animals to polyphenols. Biomed Res 2021, 42, 43–51. [Google Scholar] [CrossRef]
- Kurogi, M.; Miyashita, M.; Emoto, Y.; Kubo, Y.; Saitoh, O. Green tea polyphenol epigallocatechin gallate activates TRPA1 in an intestinal enteroendocrine cell line, STC-1. Chem Senses 2012, 37, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Kurogi, M.; Kawai, Y.; Nagatomo, K.; Tateyama, M.; Kubo, Y.; Saitoh, O. Auto-oxidation products of epigallocatechin gallate activate TRPA1 and TRPV1 in sensory neurons. Chem Senses 2015, 40, 27–46. [Google Scholar] [CrossRef]
- Amoah, I.; Lim, J.J.; Osei, E.O.; Arthur, M.; Tawiah, P.; Oduro, I.N.; Aduama-Larbi, M.S.; Lowor, S.T.; Rush, E. Effect of Cocoa Beverage and Dark Chocolate Consumption on Blood Pressure in Those with Normal and Elevated Blood Pressure: A Systematic Review and Meta-Analysis. Foods 2022, 11. [Google Scholar] [CrossRef]
- Ried, K.; Sullivan, T.R.; Fakler, P.; Frank, O.R.; Stocks, N.P. Effect of cocoa on blood pressure. The Cochrane database of systematic reviews 2012, 8, Cd008893. [Google Scholar]
- Hooper, L.; Kay, C.; Abdelhamid, A.; Kroon, P.A.; Cohn, J.S.; Rimm, E.B.; Cassidy, A. Effects of chocolate, cocoa, and flavan-3-ols on cardiovascular health: a systematic review and meta-analysis of randomized trials. The American journal of clinical nutrition 2012, 95, 740–751. [Google Scholar] [CrossRef]
- Ebaditabar, M.; Djafarian, K.; Saeidifard, N.; Shab-Bidar, S. Effect of dark chocolate on flow-mediated dilatation: Systematic review, meta-analysis, and dose-response analysis of randomized controlled trials. Clin Nutr ESPEN 2020, 36, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, N.; Fushimi, T.; Fujii, Y. Hormetic response to B-type procyanidin ingestion involves stress-related neuromodulation via the gut-brain axis: Preclinical and clinical observations. Frontiers in nutrition 2022, 9, 969823. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zimmermann, D.; De Castro, C.A.; Actis-Goretta, L. Dose-response relationship between cocoa flavanols and human endothelial function: a systematic review and meta-analysis of randomized trials. Food Funct 2019, 10, 6322–6330. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Osakabe, N.; Di Paola, R.; Siracusa, R.; Fusco, R.; D’Amico, R.; Impellizzeri, D.; Cuzzocrea, S.; Fritsch, T.; Abdelhameed, A.S.; et al. Hormesis defines the limits of lifespan. Ageing research reviews 2023, 91, 102074. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Cornelius, C.; Cuzzocrea, S.; Iavicoli, I.; Rizzarelli, E.; Calabrese, E.J. Hormesis, cellular stress response and vitagenes as critical determinants in aging and longevity. Mol Aspects Med 2011, 32, 279–304. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, C.; Graziano, A.; Calabrese, E.J.; Calabrese, V. Hormesis and vitagenes in aging and longevity: mitochondrial control and hormonal regulation. Horm Mol Biol Clin Investig 2013, 16, 73–89. [Google Scholar] [CrossRef]
- Fushimi, T.; Fujii, Y.; Koshino, H.; Inagawa, K.; Saito, A.; Koizumi, R.; Shibata, M.; Osakabe, N. Method for detecting hemodynamic alterations following a single gavage in rats. Exp Anim 2021, 70, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Ingawa, K.; Aruga, N.; Matsumura, Y.; Shibata, M.; Osakabe, N. Alteration of the systemic and microcirculation by a single oral dose of flavan-3-ols. PloS one 2014, 9, e94853. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Inagawa, K.; Ebe, R.; Fukase, S.; Horikoshi, Y.; Shibata, M.; Osakabe, N. Onset of a hypotensive effect following ingestion of flavan 3-ols involved in the activation of adrenergic receptors. Free radical biology & medicine 2016, 99, 584–592. [Google Scholar]
- Charkoudian, N.; Rabbitts, J.A. Sympathetic neural mechanisms in human cardiovascular health and disease. Mayo Clinic proceedings 2009, 84, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Charkoudian, N.; Wallin, B.G. Sympathetic neural activity to the cardiovascular system: integrator of systemic physiology and interindividual characteristics. Comprehensive Physiology 2014, 4, 825–850. [Google Scholar] [PubMed]
- Koizumi, R.; Fushimi, T.; Sato, Y.; Fujii, Y.; Sato, H.; Osakabe, N. Relationship between hemodynamic alteration and sympathetic nerve activation following a single oral dose of cinnamtannin A2. Free radical research 2020, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, A.; Horikoshi, Y.; Fushimi, T.; Saito, A.; Koizumi, R.; Fujii, Y.; Hu, Q.Q.; Hirota, Y.; Aizawa, K.; Osakabe, N. Acylated anthocyanins derived from purple carrot (Daucus carota L.) induce elevation of blood flow in rat cremaster arteriole. Food Funct 2019, 10, 1726–1735. [Google Scholar] [CrossRef] [PubMed]
- Tentolouris, N.; Liatis, S.; Katsilambros, N. Sympathetic system activity in obesity and metabolic syndrome. Annals of the New York Academy of Sciences 2006, 1083, 129–152. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Muta, O.; Teshima, T.; Hirasima, N.; Odaka, M.; Fushimi, T.; Fujii, Y.; Osakabe, N. Repeated Oral Administration of Flavan-3-ols Induces Browning in Mice Adipose Tissues through Sympathetic Nerve Activation. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Muta, O.; Oyama, S.; Odaka, M.; Shimizu, K.; Katsuragawa, S.; Suzuki, K.; Fushimi, T.; Fujii, Y.; Akagi, R.; Osakabe, N. Cinnamtannin A2, (-)-epicatechin tetramer, attenuates skeletal muscle wasting in disuse atrophy model mice induced by hindlimb suspension. J Clin Biochem Nutr 2023, 73, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Ishimura, K.; Oya, S.; Kamino, M.; Fujii, Y.; Nanba, F.; Toda, T.; Ishii, T.; Adachi, T.; Suhara, Y.; et al. Comparison of the sympathetic stimulatory abilities of B-type procyanidins based on induction of uncoupling protein-1 in brown adipose tissue (BAT) and increased plasma catecholamine (CA) in mice. PloS one 2018, 13, e0201203. [Google Scholar] [CrossRef]
- Ogawa, N.; Kurokawa, T.; Mori, Y. Sensing of redox status by TRP channels. Cell Calcium 2016, 60, 115–122. [Google Scholar] [CrossRef]
- Machado, S.A.; Pasquarelli-do-Nascimento, G.; da Silva, D.S.; Farias, G.R.; de Oliveira Santos, I.; Baptista, L.B.; Magalhães, K.G. Browning of the white adipose tissue regulation: new insights into nutritional and metabolic relevance in health and diseases. Nutrition & metabolism 2022, 19, 61. [Google Scholar]
- Molina-Hidalgo, C.; Stillman, C.M.; Collins, A.M.; Velazquez-Diaz, D.; Ripperger, H.S.; Drake, J.A.; Gianaros, P.J.; Marsland, A.L.; Erickson, K.I. Changes in stress pathways as a possible mechanism of aerobic exercise training on brain health: a scoping review of existing studies. Frontiers in physiology 2023, 14, 1273981. [Google Scholar] [CrossRef] [PubMed]
- Wadsworth, M.E.; Broderick, A.V.; Loughlin-Presnal, J.E.; Bendezu, J.J.; Joos, C.M.; Ahlkvist, J.A.; Perzow, S.E.D.; McDonald, A. Co-activation of SAM and HPA responses to acute stress: A review of the literature and test of differential associations with preadolescents’ internalizing and externalizing. Dev Psychobiol 2019, 61, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Suzuki, K.; Adachi, T.; Taira, S.; Osakabe, N. Corticotropin-releasing hormone is significantly upregulated in the mouse paraventricular nucleus following a single oral dose of cinnamtannin A2 as an (-)-epicatechin tetramer. J Clin Biochem Nutr 2019, 65, 29–33. [Google Scholar] [CrossRef]
- Fujii, Y.; Suzuki, K.; Hasegawa, Y.; Nanba, F.; Toda, T.; Adachi, T.; Taira, S.; Osakabe, N. Single oral administration of flavan 3-ols induces stress responses monitored with stress hormone elevations in the plasma and paraventricular nucleus. Neuroscience letters 2018, 682, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: a clinical perspective. British journal of pharmacology 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yu, M.; Liu, Y.; Yu, S. TRP channel functions in the gastrointestinal tract. Seminars in immunopathology 2016, 38, 385–396. [Google Scholar] [CrossRef]
- Uchida, K.; Dezaki, K.; Yoneshiro, T.; Watanabe, T.; Yamazaki, J.; Saito, M.; Yada, T.; Tominaga, M.; Iwasaki, Y. Involvement of thermosensitive TRP channels in energy metabolism. J Physiol Sci 2017, 67, 549–560. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.H. Neural control of blood pressure: focusing on capsaicin-sensitive sensory nerves. Cardiovascular & hematological disorders drug targets 2007, 7, 37–46. [Google Scholar]
- Fushimi, T.; Hirahata, C.; Hiroki, K.; Fujii, Y.; Calabrese, V.; Suhara, Y.; Osakabe, N. Activation of transient receptor potential channels is involved in reactive oxygen species (ROS)-dependent regulation of blood flow by (-)-epicatechin tetramer cinnamtannin A2. Biochemical pharmacology 2023, 214, 115682. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, K.C.; Duke, S.O. Function of polyphenol oxidase in higher plants. Physiologia Plantarum 1984, 60, 106–112. [Google Scholar] [CrossRef]
- Tan, J.; de Bruijn, W.J.C.; van Zadelhoff, A.; Lin, Z.; Vincken, J.-P. Browning of Epicatechin (EC) and Epigallocatechin (EGC) by Auto-Oxidation. Journal of agricultural and food chemistry 2020, 68, 13879–13887. [Google Scholar] [CrossRef]
- Fang, J. Bioavailability of anthocyanins. Drug metabolism reviews 2014, 46, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Kozai, D.; Ogawa, N.; Mori, Y. Redox regulation of transient receptor potential channels. Antioxidants & redox signaling 2014, 21, 971–986. [Google Scholar]
- Ogawa, N.; Kurokawa, T.; Fujiwara, K.; Polat, O.K.; Badr, H.; Takahashi, N.; Mori, Y. Functional and Structural Divergence in Human TRPV1 Channel Subunits by Oxidative Cysteine Modification*. Journal of Biological Chemistry 2016, 291, 4197–4210. [Google Scholar] [CrossRef] [PubMed]
- Kawase, M.; Chen, W.; Kawaguchi, K.; Nyasha, M.R.; Sasaki, S.; Hatakeyama, H.; Kaneko, T.; Kanzaki, M. TRPA1 and TRPV1 channels participate in atmospheric-pressure plasma-induced [Ca(2+)](i) response. Sci Rep 2020, 10, 9687. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Chen, L. Polyphenols and bioavailability: an update. Crit Rev Food Sci Nutr 2019, 59, 2040–2051. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Xing, X.; Wang, S. Health benefits of dietary polyphenols: insight into interindividual variability in absorption and metabolism. Current Opinion in Food Science 2022, 48, 100941. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. The American journal of clinical nutrition 2005, 81, 230s–242s. [Google Scholar] [CrossRef]
- Felgines, C.; Talavéra, S.; Gonthier, M.P.; Texier, O.; Scalbert, A.; Lamaison, J.L.; Rémésy, C. Strawberry anthocyanins are recovered in urine as glucuro- and sulfoconjugates in humans. The Journal of nutrition 2003, 133, 1296–1301. [Google Scholar] [CrossRef] [PubMed]
- Bitsch, R.; Netzel, M.; Sonntag, S.; Strass, G.; Frank, T.; Bitsch, I. Urinary Excretion of Cyanidin Glucosides and Glucuronides in Healthy Humans After Elderberry Juice Ingestion. J Biomed Biotechnol 2004, 2004, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Koirala, N.; Thuan, N.H.; Ghimire, G.P.; Thang, D.V.; Sohng, J.K. Methylation of flavonoids: Chemical structures, bioactivities, progress and perspectives for biotechnological production. Enzyme and Microbial Technology 2016, 86, 103–116. [Google Scholar] [CrossRef] [PubMed]
- de Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. British journal of pharmacology 2014, 171, 3268–3282. [Google Scholar] [CrossRef]
- Luo, C.; Wei, X.; Song, J.; Xu, X.; Huang, H.; Fan, S.; Zhang, D.; Han, L.; Lin, J. Interactions between Gut Microbiota and Polyphenols: New Insights into the Treatment of Fatigue. Molecules (Basel, Switzerland) 2022, 27. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Frontiers in nutrition 2019, 6. [Google Scholar] [CrossRef]
- Wang, X.; Yu, J.; Zhang, X. Dietary Polyphenols as Prospective Natural-Compound Depression Treatment from the Perspective of Intestinal Microbiota Regulation. Molecules (Basel, Switzerland) 2022, 27. [Google Scholar] [CrossRef]
- Rojas, M.; Chávez-Castillo, M.; Pirela, D.; Parra, H.; Nava, M.; Chacín, M.; Angarita, L.; Añez, R.; Salazar, J.; Ortiz, R.; et al. Metabolic Syndrome: Is It Time to Add the Central Nervous System? Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Kiyimba, T.; Yiga, P.; Bamuwamye, M.; Ogwok, P.; Van der Schueren, B.; Matthys, C. Efficacy of Dietary Polyphenols from Whole Foods and Purified Food Polyphenol Extracts in Optimizing Cardiometabolic Health: A Meta-Analysis of Randomized Controlled Trials. Advances in Nutrition 2023, 14, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Fairlie-Jones, L.; Davison, K.; Fromentin, E.; Hill, A.M. The Effect of Anthocyanin-Rich Foods or Extracts on Vascular Function in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2017, 9, 908. [Google Scholar] [CrossRef] [PubMed]
- Mohammadipoor, N.; Shafiee, F.; Rostami, A.; Kahrizi, M.S.; Soleimanpour, H.; Ghodsi, M.; Ansari, M.J.; Bokov, D.O.; Jannat, B.; Mosharkesh, E.; et al. Resveratrol supplementation efficiently improves endothelial health: A systematic review and meta-analysis of randomized controlled trials. Phytother Res 2022, 36, 3529–3539. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Liu, X.X.; Bai, Y.Y.; Wang, X.J.; Sun, K.; Chen, J.Z.; Hui, R.T. Effect of oral isoflavone supplementation on vascular endothelial function in postmenopausal women: a meta-analysis of randomized placebo-controlled trials. The American journal of clinical nutrition 2010, 91, 480–486. [Google Scholar] [CrossRef]
- Fairlie-Jones, L.; Davison, K.; Fromentin, E.; Hill, A.M. The Effect of Anthocyanin-Rich Foods or Extracts on Vascular Function in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Godos, J.; Currenti, W.; Micek, A.; Falzone, L.; Libra, M.; Giampieri, F.; Forbes-Hernández, T.Y.; Quiles, J.L.; Battino, M.; et al. The Effect of Dietary Polyphenols on Vascular Health and Hypertension: Current Evidence and Mechanisms of Action. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.D.; Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cassidy, A. Relative impact of flavonoid composition, dose and structure on vascular function: a systematic review of randomised controlled trials of flavonoid-rich food products. Mol Nutr Food Res 2012, 56, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Inagawa, K.; Shibata, M.; Osakabe, N. Flavan-3-ol fraction from cocoa powder promotes mitochondrial biogenesis in skeletal muscle in mice. Lipids in health and disease 2014, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, T.; Oyama, S.; Koizumi, R.; Fujii, Y.; Osakabe, N. Impact of cyanidin 3-O-glucoside on rat micro-and systemic circulation, possibly thorough angiogenesis. J Clin Biochem Nutr 2023, 72, 132–138. [Google Scholar] [CrossRef]
- Resnick, N.; Yahav, H.; Shay-Salit, A.; Shushy, M.; Schubert, S.; Zilberman, L.C.; Wofovitz, E. Fluid shear stress and the vascular endothelium: for better and for worse. Progress in biophysics and molecular biology 2003, 81, 177–199. [Google Scholar] [CrossRef]
- Li, Y.S.; Haga, J.H.; Chien, S. Molecular basis of the effects of shear stress on vascular endothelial cells. J Biomech 2005, 38, 1949–1971. [Google Scholar] [CrossRef] [PubMed]
- Gorski, T.; De Bock, K. Metabolic regulation of exercise-induced angiogenesis. Vasc Biol 2019, 1, H1–h8. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J Cell Biol 2003, 161, 1163–1177. [Google Scholar] [CrossRef]
- Galie, P.A.; Nguyen, D.H.; Choi, C.K.; Cohen, D.M.; Janmey, P.A.; Chen, C.S. Fluid shear stress threshold regulates angiogenic sprouting. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 7968–7973. [Google Scholar] [CrossRef]
- Koizumi, R.; Fushimi, T.; Sato, Y.; Fujii, Y.; Sato, H.; Osakabe, N. Relationship between hemodynamic alteration and sympathetic nerve activation following a single oral dose of cinnamtannin A2. Free radical research 2021, 55, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Yanagimoto, A.; Matsui, Y.; Yamaguchi, T.; Saito, S.; Hanada, R.; Hibi, M. Acute Dose-Response Effectiveness of Combined Catechins and Chlorogenic Acids on Postprandial Glycemic Responses in Healthy Men: Results from Two Randomized Studies. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.E.G.; Paiva, C.; Amato, A.A.; Lofrano-Porto, A.; Wassell, S.; Bluck, L.J.C.; Dórea, J.G.; da Costa, T.H.M. Decaffeinated coffee improves insulin sensitivity in healthy men. The British journal of nutrition 2018, 119, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Jokura, H.; Watanabe, I.; Umeda, M.; Hase, T.; Shimotoyodome, A. Coffee polyphenol consumption improves postprandial hyperglycemia associated with impaired vascular endothelial function in healthy male adults. Nutrition research (New York, N.Y.) 2015, 35, 873–881. [Google Scholar] [CrossRef]
- Carnevale, R.; Silvestri, R.; Loffredo, L.; Novo, M.; Cammisotto, V.; Castellani, V.; Bartimoccia, S.; Nocella, C.; Violi, F. Oleuropein, a component of extra virgin olive oil, lowers postprandial glycaemia in healthy subjects. British journal of clinical pharmacology 2018, 84, 1566–1574. [Google Scholar] [CrossRef]
- Castro-Acosta, M.L.; Smith, L.; Miller, R.J.; McCarthy, D.I.; Farrimond, J.A.; Hall, W.L. Drinks containing anthocyanin-rich blackcurrant extract decrease postprandial blood glucose, insulin and incretin concentrations. J Nutr Biochem 2016, 38, 154–161. [Google Scholar] [CrossRef]
- Liu, C.Y.; Huang, C.J.; Huang, L.H.; Chen, I.J.; Chiu, J.P.; Hsu, C.H. Effects of green tea extract on insulin resistance and glucagon-like peptide 1 in patients with type 2 diabetes and lipid abnormalities: a randomized, double-blinded, and placebo-controlled trial. PloS one 2014, 9, e91163. [Google Scholar] [CrossRef] [PubMed]
- Yanagimoto, A.; Matsui, Y.; Yamaguchi, T.; Hibi, M.; Kobayashi, S.; Osaki, N. Effects of Ingesting Both Catechins and Chlorogenic Acids on Glucose, Incretin, and Insulin Sensitivity in Healthy Men: A Randomized, Double-Blinded, Placebo-Controlled Crossover Trial. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zibadi, S.; Rohdewald, P.J.; Park, D.; Watson, R.R. Reduction of cardiovascular risk factors in subjects with type 2 diabetes by Pycnogenol supplementation. Nutrition research (New York, N.Y.) 2008, 28, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Matsumae, T.; Kataoka, T.; Yazaki, Y.; Yamaguchi, H. Effect of acacia polyphenol on glucose homeostasis in subjects with impaired glucose tolerance: A randomized multicenter feeding trial. Exp Ther Med 2013, 5, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Ros, E.; Valderas-Martinez, P.; Casas, R.; Arranz, S.; Guillén, M.; Lamuela-Raventós, R.M.; Llorach, R.; Andres-Lacueva, C.; et al. Effects of red wine polyphenols and alcohol on glucose metabolism and the lipid profile: a randomized clinical trial. Clin Nutr 2013, 32, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Cesar, T.B.; Ramos, F.M.M.; Ribeiro, C.B. Nutraceutical Eriocitrin (Eriomin) Reduces Hyperglycemia by Increasing Glucagon-Like Peptide 1 and Downregulates Systemic Inflammation: A Crossover-Randomized Clinical Trial. Journal of medicinal food 2022, 25, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Luechapudiporn, R.; Phisalaphong, C.; Jirawatnotai, S. Curcumin extract for prevention of type 2 diabetes. Diabetes care 2012, 35, 2121–2127. [Google Scholar] [CrossRef] [PubMed]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. The British journal of nutrition 2011, 106, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Thazhath, S.S.; Wu, T.; Bound, M.J.; Checklin, H.L.; Standfield, S.; Jones, K.L.; Horowitz, M.; Rayner, C.K. Administration of resveratrol for 5 wk has no effect on glucagon-like peptide 1 secretion, gastric emptying, or glycemic control in type 2 diabetes: a randomized controlled trial. The American journal of clinical nutrition 2016, 103, 66–70. [Google Scholar] [CrossRef]
- Shamsudin, N.F.; Ahmed, Q.U.; Mahmood, S.; Shah, S.A.A.; Sarian, M.N.; Khattak, M.; Khatib, A.; Sabere, A.S.M.; Yusoff, Y.M.; Latip, J. Flavonoids as Antidiabetic and Anti-Inflammatory Agents: A Review on Structural Activity Relationship-Based Studies and Meta-Analysis. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Kim, N.H.; Kilonzi, S.M.; Oh, D.H. Insights on Dietary Polyphenols as Agents against Metabolic Disorders: Obesity as a Target Disease. Antioxidants (Basel, Switzerland) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. International journal of molecular sciences 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Balasooriya, H.; Sirisena, S.; Ng, K. The effectiveness of dietary polyphenols in obesity management: A systematic review and meta-analysis of human clinical trials. Food Chem 2023, 404, 134668. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules (Basel, Switzerland) 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Wang, H.N.; Xiang, J.Z.; Qi, Z.; Du, M. Plant extracts in prevention of obesity. Crit Rev Food Sci Nutr 2022, 62, 2221–2234. [Google Scholar] [CrossRef] [PubMed]
- Kawser Hossain, M.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Kumar Saha, S.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Molecular Mechanisms of the Anti-Obesity and Anti-Diabetic Properties of Flavonoids. International journal of molecular sciences 2016, 17, 569. [Google Scholar] [CrossRef] [PubMed]
- Raven, L.M.; Stoita, A.; Feller, R.B.; Brown, C.; Greenfield, J.R. Delayed Gastric Emptying with Perioperative Use of Glucagon-like Peptide-1 Receptor Agonists. Am J Med 2023. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes - state-of-the-art. Mol Metab 2021, 46, 101102. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.; Natsume, M.; Yasuda, A.; Nakamura, Y.; Tamura, T.; Osakabe, N.; Kanegae, M.; Kondo, K. Plasma LDL and HDL cholesterol and oxidized LDL concentrations are altered in normo- and hypercholesterolemic humans after intake of different levels of cocoa powder. The Journal of nutrition 2007, 137, 1436–1441. [Google Scholar] [CrossRef]
- Baba, S.; Osakabe, N.; Kato, Y.; Natsume, M.; Yasuda, A.; Kido, T.; Fukuda, K.; Muto, Y.; Kondo, K. Continuous intake of polyphenolic compounds containing cocoa powder reduces LDL oxidative susceptibility and has beneficial effects on plasma HDL-cholesterol concentrations in humans. The American journal of clinical nutrition 2007, 85, 709–717. [Google Scholar] [CrossRef]
- Kaelberer, M.M.; Buchanan, K.L.; Klein, M.E.; Barth, B.B.; Montoya, M.M.; Shen, X.; Bohórquez, D.V. A gut-brain neural circuit for nutrient sensory transduction. Science 2018, 361. [Google Scholar] [CrossRef] [PubMed]
- Kaelberer, M.M.; Rupprecht, L.E.; Liu, W.W.; Weng, P.; Bohórquez, D.V. Neuropod Cells: The Emerging Biology of Gut-Brain Sensory Transduction. Annu Rev Neurosci 2020, 43, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Fraser, K.A.; Davison, J.S. Cholecystokinin-induced c-fos expression in the rat brain stem is influenced by vagal nerve integrity. Experimental physiology 1992, 77, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Rinaman, L.; Baker, E.A.; Hoffman, G.E.; Stricker, E.M.; Verbalis, J.G. Medullary c-Fos activation in rats after ingestion of a satiating meal. Am J Physiol 1998, 275, R262–R268. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.N.; Hsu, T.M.; Liu, C.M.; Noble, E.E.; Cortella, A.M.; Nakamoto, E.M.; Hahn, J.D.; de Lartigue, G.; Kanoski, S.E. Gut vagal sensory signaling regulates hippocampus function through multi-order pathways. Nature communications 2018, 9, 2181. [Google Scholar] [CrossRef] [PubMed]
- Klarer, M.; Arnold, M.; Günther, L.; Winter, C.; Langhans, W.; Meyer, U. Gut vagal afferents differentially modulate innate anxiety and learned fear. J Neurosci 2014, 34, 7067–7076. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Yan, Q.; Ma, Y.; Fang, J.; Yang, Y. Recognizing the role of the vagus nerve in depression from microbiota-gut brain axis. Front Neurol 2022, 13, 1015175. [Google Scholar] [CrossRef]
- Lightman, S.L.; Birnie, M.T.; Conway-Campbell, B.L. Dynamics of ACTH and Cortisol Secretion and Implications for Disease. Endocr Rev 2020, 41. [Google Scholar] [CrossRef]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Frontiers in immunology 2019, 10, 1545. [Google Scholar] [CrossRef]
- Bermejo, J.L.; Valldecabres, R.; Villarrasa-Sapiña, I.; Monfort-Torres, G.; Marco-Ahulló, A.; Ribeiro Do Couto, B. Increased cortisol levels caused by acute resistance physical exercise impair memory and learning ability. PeerJ 2022, 10, e13000. [Google Scholar] [CrossRef]
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The Role of Cortisol in Chronic Stress, Neurodegenerative Diseases, and Psychological Disorders. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; McEwen, B.S.; Friston, K. Uncertainty and stress: Why it causes diseases and how it is mastered by the brain. Prog Neurobiol 2017, 156, 164–188. [Google Scholar] [CrossRef] [PubMed]
- Kanaley, J.A.; Weltman, J.Y.; Pieper, K.S.; Weltman, A.; Hartman, M.L. Cortisol and growth hormone responses to exercise at different times of day. J Clin Endocrinol Metab 2001, 86, 2881–2889. [Google Scholar] [CrossRef] [PubMed]
- Stranahan, A.M.; Lee, K.; Mattson, M.P. Central mechanisms of HPA axis regulation by voluntary exercise. Neuromolecular Med 2008, 10, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Cotman, C.W. Voluntary exercise protects against stress-induced decreases in brain-derived neurotrophic factor protein expression. Neuroscience 2004, 124, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, T.J.; Gould, E. Stress, stress hormones, and adult neurogenesis. Exp Neurol 2012, 233, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Nakagawa, S.; Kitaichi, Y.; An, Y.; Omiya, Y.; Song, N.; Koga, M.; Kato, A.; Inoue, T.; Kusumi, I. The role of medial prefrontal corticosterone and dopamine in the antidepressant-like effect of exercise. Psychoneuroendocrinology 2016, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Nakagawa, S.; An, Y.; Ito, K.; Kitaichi, Y.; Kusumi, I. The exercise-glucocorticoid paradox: How exercise is beneficial to cognition, mood, and the brain while increasing glucocorticoid levels. Frontiers in neuroendocrinology 2017, 44, 83–102. [Google Scholar] [CrossRef]
- Fujii, Y.; Sakata, J.; Sato, F.; Onishi, K.; Yamato, Y.; Sakata, K.; Taira, S.; Sato, H.; Osakabe, N. Impact of short-term oral dose of cinnamtannin A2, an (-)-epicatechin tetramer, on spatial memory and adult hippocampal neurogenesis in mouse. Biochem Biophys Res Commun 2021, 585, 1–7. [Google Scholar] [CrossRef]
Figure 1.
Chemical structure of flavonoid (a), (-)-epicatechin(b), epigallocatechin gallate (c), procyanidin(d), amarogentin (e).
Figure 1.
Chemical structure of flavonoid (a), (-)-epicatechin(b), epigallocatechin gallate (c), procyanidin(d), amarogentin (e).
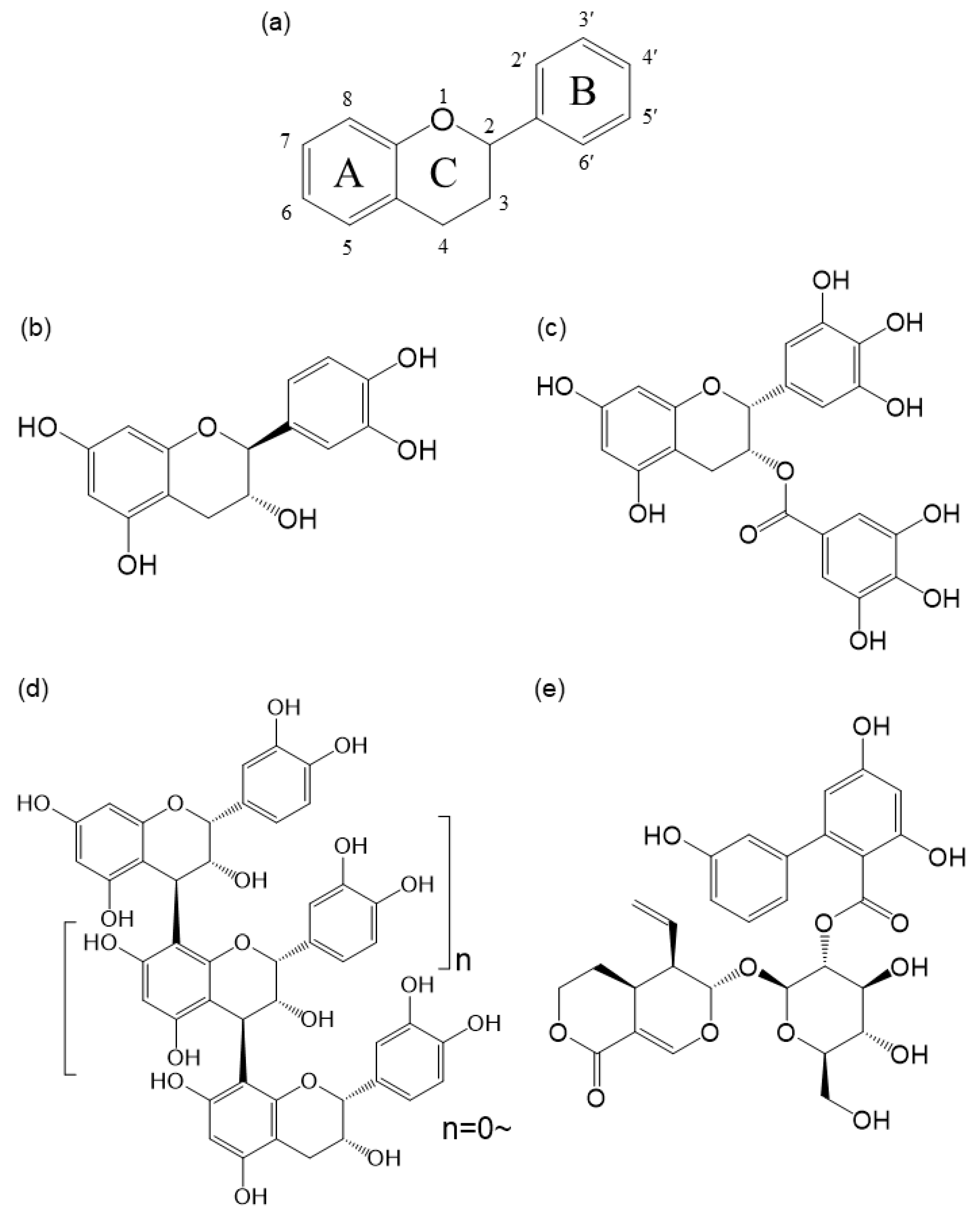
Figure 2.
Hypothetical recognition mechanism of astringent polyphenols by TRP channel.
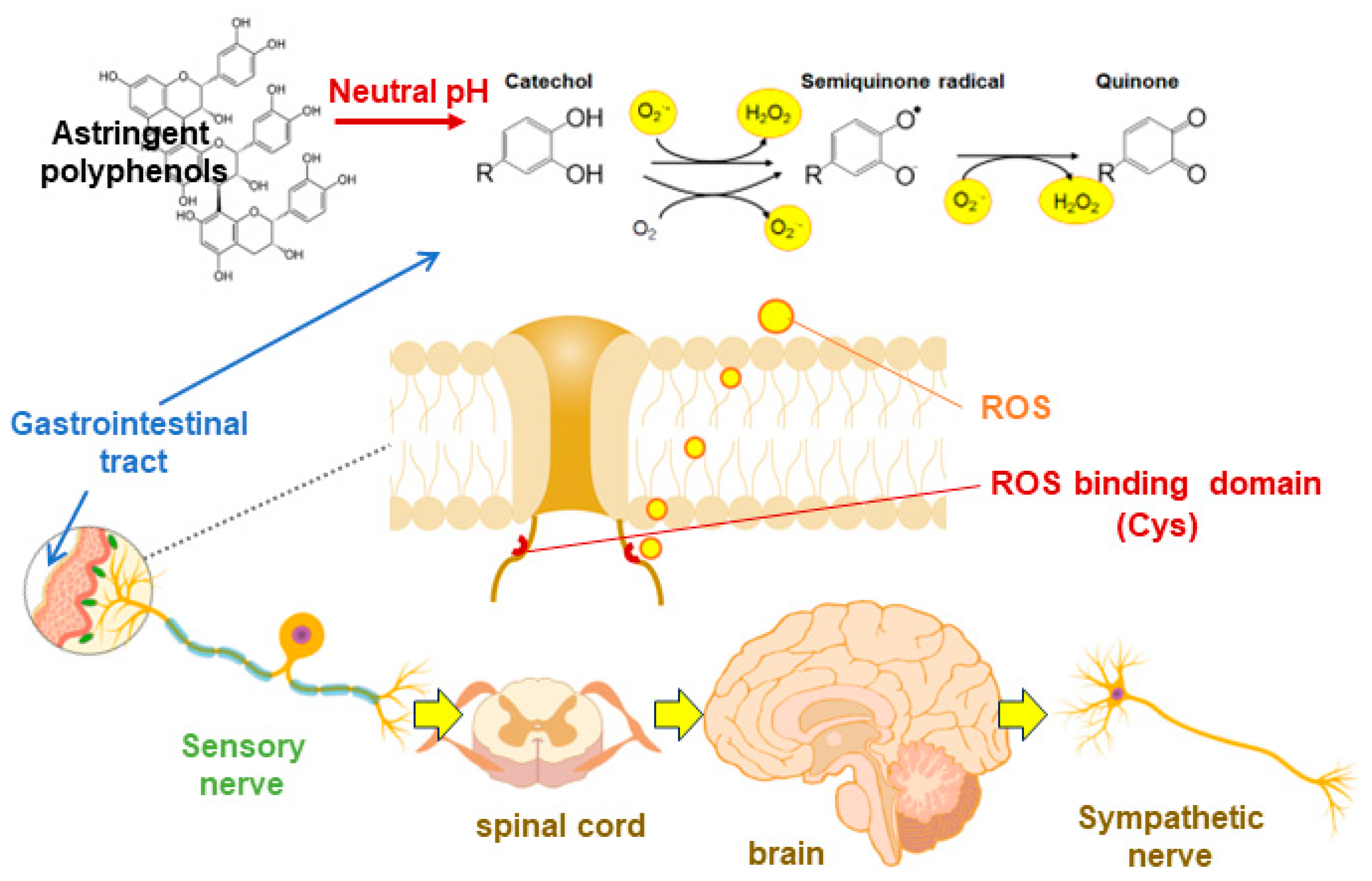
Figure 3.
Illustrations of two hypotheses for the mechanisms underlying the observed effects of a single (a) and repeated oral administration (b) of astringent polyphenols on micro-and systemic circulation.
Figure 3.
Illustrations of two hypotheses for the mechanisms underlying the observed effects of a single (a) and repeated oral administration (b) of astringent polyphenols on micro-and systemic circulation.
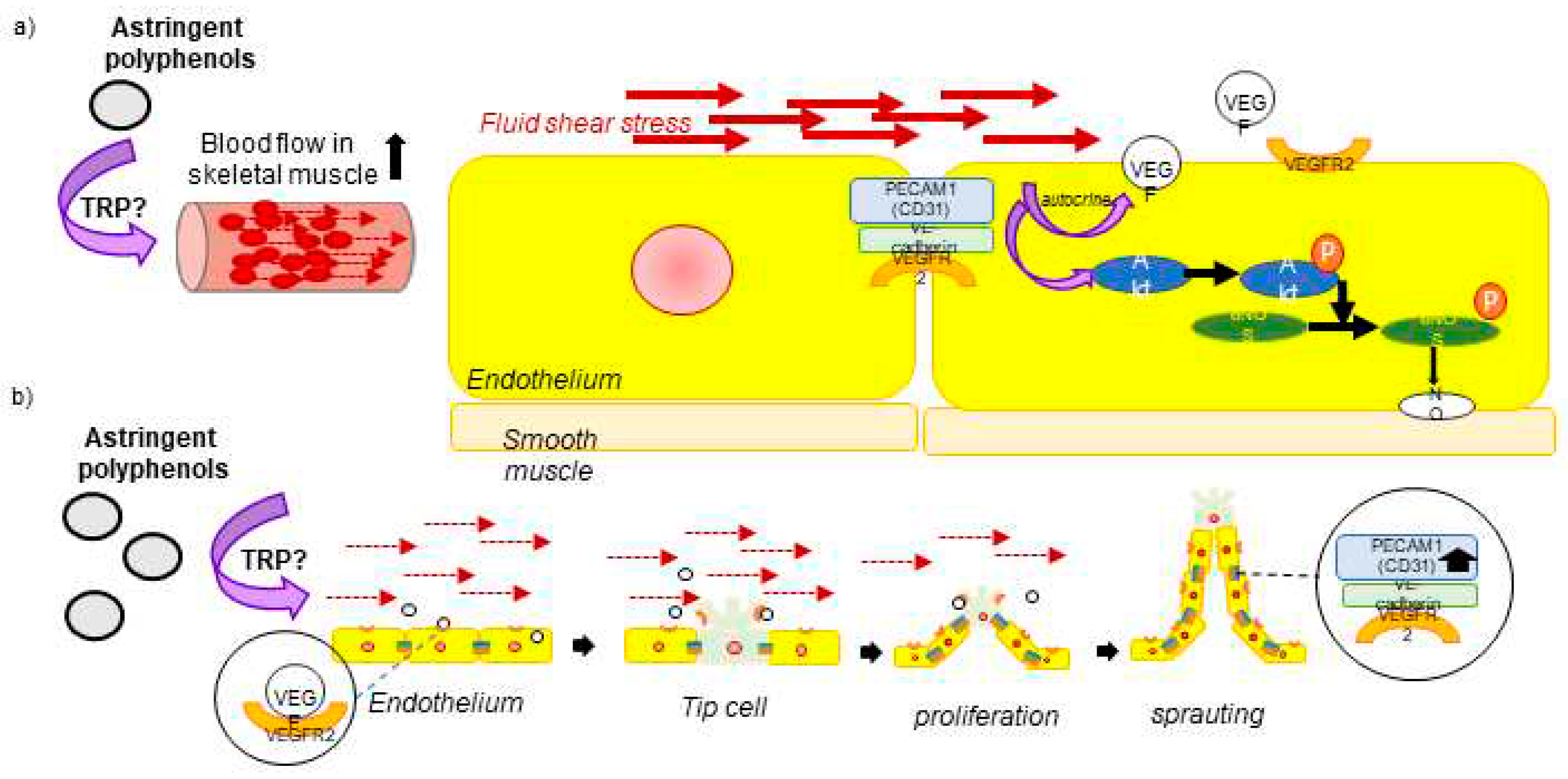

Table 1.
The interaction of polyphenols and T2R in vitro studies.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



