Submitted:
12 January 2024
Posted:
15 January 2024
You are already at the latest version
Abstract
Emotional experiences are a part of our lives. The maladaptive functioning of an individual's emotional field can lead to emotional disturbances of various kinds, such as anxiety and depression. Currently, there is an increasing prevalence of emotional disorders that cause great human suffering and high socio-economic costs. Emotional processing has a biological basis. The major neuroscientific theories of emotion are based on biological functioning, and all of them take into account the anatomy and function of the tenth cranial nerve: the vagus nerve. The vagus nerve connects the subdiaphragmatic and supradiaphragmatic areas and modulates emotional processing as the basis of interoceptive functioning. Auricular vagus nerve stimulation is a new and innovative neuromodulation technique based on the function of the vagus nerve. Several interventions have shown that this new neurostimulation technique is a very promising resource for treating emotional disorders. In this paper, we summarise three neuroscientific theories of emotion, explain what transauricular vagus nerve stimulation is, and present arguments for its use and continued research.
Keywords:
emotion
; emotional disorders
; neuromodulation
; transauricular vagus nerve stimulation
Emotional life is a constitutive human reality. As a biological system, emotions take place within our biology; an adaptive emotional life is an indicator of mental health. According to recent data, between 1990 and 2019, the global number of disability-adjusted life years (DALYS) attributable to mental disorders increased from 80.8 million in 1990 to 125.3 million in 2019 [1]; the economic value associated with this burden is estimated to be in the order of five million US dollars [2]. Within mental disorders, depressive and anxiety disorders are among the leading causes of DALYS worldwide (ranked 13th and 24th, respectively). In 2019, 301 million people had an anxiety disorder and 280 million depression. The continued high prevalence of these disorders, along with bipolar and eating disorders, is of particular concern because they also increase the risk of suicide (ranked 18th among the leading causes of death in the 2019 GBD).
These disorders occur in specific neural circuits involved in cortico-amygdalar connections [3,4,5] and are intricately involved in human emotional experience [6,7,8]. Emotional experience is the result of the dynamics generated as a consequence of emotional reactions and the capacity, ability, and willingness to regulate such reactions and the resulting experiences [9]. Emotional response is one of the dimensions that shape temperament, understanding personality as the result of the interaction between temperament and character, where character is the volitional part of personality and temperament; from an epigenetic perspective, it is the part of personality most closely related to our genetic configuration [10].
Therefore, based on these potentials of emotional reactivity in temperament and the capacity for control found in the character dimension of personality, it would be possible to identify, based on scientific knowledge and current technological resources, how to improve emotional states.
To this end, solid explanatory theories based on the current scientific corpus, such as that proposed by LeDoux [11]; the Theory of constructed emotion [12]; and the Polyvagal theory [13,14], will be presented below. Additionally, based on the foundations of these theories, together with the description of the technique, arguments will be made for the use of transauricular vagus nerve stimulation (tVNS) to improve the emotional state.
1. The LeDoux Proposal (2021)
LeDoux [11] considers emotions to be conscious experiences that arise in biologically or psychologically significant situations. Concerning consciousness, and based on Endel Tulving’s theory, three states are distinguished: autonoetic, noetic, and anoetic. Each depends on a different kind of memory: autonoetic consciousness depends on episodic memory, noetic depends on semantic memory, and anoetic depends on procedural memory. So, for example, if you are confronted with the proximity of a snake using semantic memory, the stimulus is recognised as a snake and is likely to be conceptualised as a potentially dangerous stimulus and situation. The result is a case of noetic awareness of danger, which includes awareness of the relationship between danger and the experience of fear. With the addition of episodic memory, memories of your ’I’ are added and ’you’ become part of the experience: you conceptualise that the snake can harm you. The experience has become an autonoetic state of reflexive self-consciousness in which you are afraid of what might happen to you. Not all autonoetic states are emotional states, however, all emotional states are autonoetic states.
As for the process of emotion generation (see Figure 1), which is a multi-stage process that develops in three phases of the nervous system (unconscious, pre-conscious, and conscious), it begins with a stimulus from the world (internal-external) that is meaningful to the subject. Significance is related to risk, which is seen as a reality that threatens the equilibrium of the organism. This significant stimulus is thus processed at an unconscious level by the senses in interaction with the memory and the body’s risk detection mechanisms, and the resulting physiological and behavioural responses. The risk detection mechanisms generate automatic physiological and behavioural responses that, together with the processing of memory and information from the senses, are the inputs for the processing that will take place at the preconscious level. Here, with the information from the non-conscious level, schemas of the situation, of the self, and of the emotion will emerge; these, in turn, will form a mental model that will be the input received by the conscious level. At the conscious level, the conscious emotional experience is generated through the self-reported narrative of the mental model. The narrative can be about immediate experiences or memories. The conscious emotional experience resulting from the narrative is of a particular categorical type of emotion (e.g. fear, joy, anger, jealousy, envy, pride) or an undefined state of distress or well-being. Because emotional situations can change from one moment to another, so can schemas, mental models, and narratives. Consequently, emotional experience is dynamic in real time: distress can turn into fear or anger, or fear can turn into anger, jealousy, or shame.
For LeDoux [11,15,16], the key is threat detection, which is a survival mechanism common to all forms of life. In mammals, in particular, a central component of this mechanism is a set of nuclei in the temporal lobe known as the amygdalar complex [17]. The amygdalar complex contains at least 13 separate, distinct nuclei, each with unique patterns of connectivity and functionality [18,19]. These subnuclei are typically grouped into three main divisions: a) the deep basolateral complex (BLA), consisting of the lateral, basal, and accessory basal nuclei; b) the superficial cortical nuclei and the nuclei of the lateral olfactory tract; and c) the centromedial group, consisting of the medial and central nuclei (CeA) [20]. In general, multisensory information from the environment is first received by the amygdala via projections from the nuclei of the thalamus and sensory cortex, and transmitted to the BLA and adjacent CeA. The BLA transmits information to cortical regions; this transmission is regulated by excitatory projections from the cortex [21] and by associated intercalated cells that can shut down the transmission of nerve impulses between the BLA and CeA [22]. The CeA plays a key role in modulating autonomic and endocrine responses for a range of visceral functions [23]. The cortico-medial region is considered “evolutionarily primitive” due to its connections with the olfactory system. In contrast, the BLA is considered "evolutionarily more recent" due to its predominant coupling with the neocortex, mainly through the medial prefrontal cortex and sensory association areas, as well as other subcortical structures such as the hippocampus [17]. In this line, Barger et al. [24] carried out a comparative work on the number of neurons in humans and non-human primates, the result of which showed a higher cell density in humans concentrated in the lateral nuclei of the amygdala, which have dense connections with different regions of the cortex [17]. It is the different areas in the cortex that are responsible for generating the self-reported narrative of one’s own self-reported emotional experience per se [11]. Concerning emotional experiences, recent studies show that a deficient functional interaction between areas of the prefrontal cortex and amygdaloid nuclei is clearly related to major depressive disorder [5,25]; post-traumatic stress disorder (PTSD) [26,27]; and various anxiety-related disorders [4,28].
2. Theory of Constructed Emotion
The Theory of constructed emotion is based on the bayesian model of brain function. According to this model, as the brain processes sensory inputs, it searches within itself for previous models associated with these incoming inputs, activating those that show the greatest similarity. Based on this information, it makes predictions about what will happen. Thus, when predictions and reality resemble one another, certainty is generated; when they don’t resemble one another, uncertainty is generated [7]. While certainty is considered a success, uncertainty is equated with prediction errors (surprises), which the brain uses to revise its models of the world. In all this processing, the brain tries to reduce entropy—prediction errors—by continuously actively adjusting the predictions it makes, i.e., by allostasis. This is because the attempt to reduce surprise is always a consequence of an inherently self-evident mechanism [29]. At the brain level, prediction and prediction error occupy the deep and superficial pyramidal layers of the brain, respectively [30], and from the cortical hierarchy down to the lower levels of the hierarchy, i.e. to the sensory stimuli. At all levels, the goal is to use as few resources as possible to make the connection between sensory input and one’s own schema of the world.
In this conception, the theory of constructed emotion [11,12,31] designates as "predictive coding" the process that involves both encoding sensory inputs and the subsequent activation of prior mental models related to the new inputs, and the subsequent process of predicting, with its certainties and surprises. This process also involves the reduction of entropy through allostasis. This predictive coding occurs both concerning the internal environment of the subject’s own body and as a result of the subject’s interaction with the external environment [32,33].
On the one hand, as regards the internal environment, the brain models the world from the perspective of the body’s physiological needs through interoceptive processing [34,35]. In such processing, ascending interoceptive signals carry sensory prediction errors from the internal environment to the brain via lamina I and vagal afferent pathways, and are anatomically positioned to be modulated by descending visceromotor predictions that control the internal environment [12]. Recent research suggests that interoception is at the core of the brain’s internal model [12,36,37]. Interoceptive sensations are often experienced as low-dimensional affective feelings [12,38]. As such, they possess the properties of affect, i.e., their own valence and arousal [39].
Furthermore, on the other hand, concerning the external world, during the process of socialisation, the subject will integrate, concerning emotional experiences, the properties of the different emotional states granted by the social group to which they belong, integrating within mental schemas the different emotional categories, which will be activated by predictive coding [32]. Specifically, category construction occurs automatically and continuously throughout the lifespan via predictive processing, that is, by predicting, selecting, and processing correctly [12,36,40,41]. Each emotional event begins as a category, constructed as a set of interrelated physical signals that evolve in the brain. The incoming sensory signals, together with the information contained in mental models based on integrated emotional categories, will be the main agents in the allostatic processing aimed at reducing entropy; they will provide the selection of a similar category, whose activation and assembly with the incoming inputs will coordinate motor actions and conscious experience. When unexpected signals arrive from sensory surfaces or expected signals fail to materialise, i.e., when ’prediction errors’ are made as entropy increases, the brain will attempt to correct its predictions (known as ’learning’) [32]. Prediction errors are associated with negative valence of emotional experiences, whereas positive valence indicates emotional experiences that resolve uncertainty and provide a sense of control [42]. This links emotional states to uncertainty resolution, neuromodulation, and cortical control of gain [43], giving positive valence to prediction accuracy and negative valence to prediction error.
Based on the interaction between one’s body and the environment, this theory suggests that emotions are created from a dimensional and categorical perspective. The dimensional perspective refers to the dimensions of valence and activation resulting from interoceptive processing. The categorical perspective refers to the process by which the subject, in the face of certain changes in the internal and external world, effortlessly attempts to make sense of the emotional experience based on emotional categories acquired in the process of socialisation [32]. Multiple areas of the brain are involved in the categorical elaboration of emotional experience, and biological circuits are not differentiated for specific emotions [44]. A review by Wager et al. [45] analysed patterns of human brain activity from 148 studies on emotional categories (2159 participants in total). They concluded that emotional categories correspond to predictive patterns in cortical and subcortical areas across multiple brain networks. This means that even category-based emotional experience is rooted in predictive hits and misses, as observed in patterns of brain activity.
3. Polyvagal Theory
The Polyvagal theory [13,14,46] suggests that the origin of emotional responses resides in automatic and unconscious perceptions made by the autonomic nervous system (ANS) based on perceived safety, risk, or extreme risk. Based on this theory, the ANS is hierarchically structured around the configuration of the vagus nerve. The vagus nerve has multiple innervations and connections to much of the body [47,48]. Structurally, the vagus nerve consists of the ventral and dorsal vagal branches; the dorsal branch lacks myelin and is phylogenetically the oldest, whereas the ventral branch has myelin and is phylogenetically the most recent [49,50]. Together with the sympathetic-adrenal axis, which is specific to the sympathetic nervous system, these two complexes form the three neurobiological circuits of the ANS [13].
The ventral-vagal complex (VVC), or myelinated vagus, is activated when the organism perceives safety. It is located in the nucleus ambiguus (NA) and its innervations are directed to supradiaphragmatic areas. From the ventral part of the NA, it exchanges information with the nucleus of the solitary tract (NTS), with some cranial nerves, and with the sinoatrial node of the heart. As regards the NTS, it makes connections with the hypothalamus, the limbic system, the periaqueductal grey matter, the amygdala, and various parts of the cortex [47]. In terms of cranial nerves, various innervations of the glossopharyngeal and facial nerves are also found in the NA. As a consequence of the automatic activation of the VVC, the face and voice show prosocial patterns such as smiling and a pleasant tone of voice [13]. Also, concerning the sinoatrial node, at the level of the heart, the myelinated vagus is an inhibitor of the sympathetic system, which acts as a brake, allowing a rapid slowing of the heart rate [50]. The emotional response generated in the organism is typical of a state of uncertainty.
When the body automatically detects a risk, the influence of the ventral vagus disappears and the sympathetic-adrenal system is activated. The sympathetic-adrenal system is part of the sympathetic nervous system and is considered an adaptive activation system that supports fight-flight behaviour, which is associated with an almost complete withdrawal of the parasympathetic influence of the VVC [13]. The autonomic functions of such a complex are increased heart rate, reduced heart rate variability, secretion of glucocorticoids and catecholamines, energy production including glucose, and conversion of norepinephrine to epinephrine [46]. As the neurobehavioural goal is fight or flight, cognitive function is subordinated to amygdalar function [51]. The emotional response generated is typical of a state of stress.
Finally, when extreme risk is perceived, the dorsal-vagal complex (DVC) or unmyelinated vagus system is activated, mainly innervating subdiaphragmatic areas. The neurobehavioural functions of this complex are immobilisation or passive adaptation, including apparent death and loss of consciousness [46]. Thus, the emotional response generated is related to the characteristics of emotional shock.
Therefore, according to the Polyvagal theory, it is assumed that the genesis of the emotional experience is found in that automatic perceptual stage and outside the will that the organism performs based on the safety, risk, or extreme risk detected, which generates a primary emotional reaction of well-being, stress, or emotional shock in the organism. The emotional experience is created based on this automatic ANS response.
Having summarised the three theories, the following recapitulation can be made. In the LeDoux model, at the unconscious level of the ANS, the interoceptive state together with risk detection are key; both processes involve the vagus nerve. From the Theory of constructed emotion, the affective state of the body (valence and arousal) depends on interoceptive processing and predictive coding, suggesting that the sensory prediction errors of the internal environment are transported by vagal afferents, demonstrating the importance of the vagus nerve. According to Polyvagal theory, the vagus nerve is responsible for orchestrating the physiological state of the emotional response based on the vagus nerve’s detection of safety, risk, or extreme risk.
The vagus nerve, the tenth pair of cranial nerves, is the largest component of the parasympathetic nervous system and links the peripheral organs to the brain [52,53], providing information to the brainstem about the state of the sensory organs [54], i.e., the interoceptive state. It also contains efferents originating in the medulla nuclei that regulate cardiac, pulmonary, and gastrointestinal activity [55]. Optimal regulation of this bidirectional centre-periphery pathway (involving interactions between brainstem structures, elements of the central autonomic network, and the prefrontal cortex) is thought to be associated with mental well-being and to influence emotional regulatory functions [56].
The question now is, can we improve emotional state via the vagus nerve? One possible answer seems to lie in an emerging neurostimulation technique, which we will discuss below.
4. Transauricular Vagus Nerve Stimulation
tVNS is a non-invasive brain stimulation technique [57] where electrodes apply electrical impulses to the skin of specific areas of the ear [58,59] (see Figure 2) to stimulate the auricular branch of the vagus nerve, also known as Alderman’s or Arnold’s nerve [60,61]. The location of the auricular vagus nerve was revealed by dissecting the ear of human cadavers [62,63] and mapping the areas of the ear. The outer ear is the only site to which the vagus nerve sends its peripheral branch, the auricular vagus nerve (aVN) (see Figure 3). The aVN is mainly an afferent fibre, innervating the ear and connecting it to the main branch of the vagus nerve [62,63]. Approximately 80% of vagus nerve fibres are considered to be afferent, while 20% are efferent [54]. Like the vagus nerve, the aVN is composed of myelinated A and B fibres and unmyelinated C fibres [58]. Thus, upon application of tVNS, the thick myelinated Ab fibres of Arnold’s nerve are excited and the afferent signal propagates from the peripheral nerves to the brainstem [65,66,67]and various subdiaphragmatic areas [58,68].
Auricular stimulation is not a new procedure [69]. In one of the earliest Chinese medical treatises written around 2,500 years ago—"Huang Di Nei Jing" - The Yellow Emperor’s Classic of Internal Medicine—auricular stimulation using acupuncture is considered a healing practice [70]. Therefore, the proposal for the application of tVNS is novel, nevertheless, its use and development are very old [71]. Until 2000, vagus nerve stimulation could only be performed invasively, via surgical procedures inserting implants in the body that directly stimulate the vagus nerve [55,72]. tVNS was an alternative developed by Ventureyra [73] for epilepsy in 2000, inspired by vagus nerve stimulation (VNS), auricular acupuncture, and the distribution of the aVN in the auricular concha [74]. Several studies conclude that tVNS generally shows similar benefits to internal VNS [69,75]. In addition, unlike internal stimulation, tVNS does not require medical intervention for placement, making it less invasive and less costly [76,77,78]. Therefore, it has received particular interest in basic and clinical studies since its introduction [59,79,80].
As an emerging neuromodulation therapy, tVNS has been shown to be safe [77,80,81]. In a recent meta-analysis and systematic review on the risks of tVNS published by Kim et al. [82], they show that there are no significant differences in terms of risks between the experimental and control groups analysed. Regarding tVNS stimulation parameters, they can vary in current intensity (mA), pulse width (μs), frequency (Hz), duty cycle (s), and session duration (min) [81]. For the mA, μs, and Hz parameters, devices marketed for tVNS are configured with appropriate safety values, although there is still no clear consensus on the optimal parameters [64,83,84].
tVNS stimulation devices have currently acquired CE-marked approval for epilepsy, depression, anxiety, pain, and migraine [64]. They are used for epilepsy, major depressive disorder, insomnia, glucose metabolic disorders, pain, stroke, stroke rehabilitation, anxiety, fear, cognitive impairment, cardiovascular disorders, tinnitus, Prader-Willi syndrome, and COVID-19 [69]. The Food and Drug Association (FDA) first approved VNS via a cervical implantable device for the treatment of depression in 1997 [85]. It is currently FDA-approved for the treatment of depression and PTSD [58]. Therefore, the last two decades have seen the rapid development of tVNS devices [55], which, as shown in Figure 4 (see Figure 4), stimulate various supradiaphragmatic and subdiaphragmatic areas innervated by the vagus nerve by stimulating the tragus or cymba conchae.
Specifically, tVNS projects the electrical signal to the NTS and parabrachial nucleus (PBN), from where it projects to cortical and subcortical structures, such as the locus coeruleus (LC), amygdala (A), anterior cingulate gyrus (ACC), and medial prefrontal cortex (mPFC) [81,87,88]. These regions are important nodes of the emotional regulation network, which is associated with the appropriate interpretation of emotions [45,89,90,91,92] and with emotional regulation strategies [93,94].
Regarding the projection of the NTS to the LC, it is worth mentioning that the LC is the largest noradrenergic nucleus [95]. Brainstem norepinephrine networks (e.g., LC and NTS) are the main recipients of vagal afferent fibres [53], from where they project to limbic and hypothalamic regions where emotion and motivation appear to be controlled [86].Two key noradrenergic regions that receive these projections are the paraventricular nucleus (PVN) and the supraoptic nucleus (SON), which contain neurons that synthesise the neuropeptide oxytocin and regulate its secretion [96]. tVNS stimulation has been shown to directly modulate the brainstem activity of the locus coeruleus-norepinephrine network (LC-NE) [77,87,94,97,98]. Indirect noradrenergic physiological markers have been proposed, such as P300, salivary alpha-amylase (SAA), and pupil dilation. Results regarding the effect of tVNS on these physiological markers are inconclusive. On the one hand, laboratory studies show that tVNS affects these non-invasive physiological markers of noradrenergic activity [100,101,102]; on the other hand, others show no such effect [97,103,104]. However, the studies showing no effects all use long on/off cycles or continuous stimulation at relatively low stimulation intensities (e.g.,0.5 mA).
As for the subdiaphragmatic projections of the vagus nerve [71], tVNS influences gastrointestinal activity by stimulating the vagus nerve [55,86]; this bidirectional brain-microbiota communication influences cognition, social behaviour, anxiety expression, and stress responses [105]. The vagus nerve plays a key role in such communication [106], influencing central nervous system (CNS) reward neurons [107] based on the predictive processing mentioned above [32]. For example, glutamate released from enteroendocrine cells can activate these vagal afferents [107], as can serotonin [108]. Concerning the gut microbiota, a growing number of studies show that it is associated with anxiety disorders and depression [109,110]. The gut microbiota is a key regulator within the brain-gut axis: bacterial species regulate the production of neurotransmitters and their precursors (e.g., serotonin, GABA, and tryptophan) and can secrete and upregulate essential proteins and metabolites involved in the release of neuropeptides and gut hormones [111,112]. In addition, vagal and spinal afferent pathways mediate neural communication between the gut microbiota and the CNS. Also, the gut microbiota modulates immune signalling from the gut to the brain through cytokine induction [113,114]. Simpson et al. [115]conducted a systematic review of the relationship between microbiota and anxiety disorders and depression, concluding that such disorders may be characterised by an increased abundance of pro-inflammatory species (e.g. Enterobacteriaceae and Desulfovibrionaceae) and decreased production of short-chain fatty acids (e.g., Faecalibacterium). It is also suggested that several taxa and their mechanisms of action may be related to the pathophysiology of anxiety and depression by the transmission of peripheral inflammation to the brain.
5. Discussion and Future Research
To summarise the impact of the vagus nerve on the brain-microbiota axis and its relationship with emotional experiences, it is worth mentioning that the first tVNS interventions associated with the emotional domain were aimed at patients resistant to treatment for depression, who, after tVNS, presented improvements in depression-associated symptomatology [116]. The study by Fang et al. [117] found that after one month of treatment with tVNS, functional connectivity improved in depressed patients compared with the control/placebo group. Another study in patients diagnosed with depression [118] found that, after two weeks of tVNS, the severity of their depression decreased. This finding was later replicated in a larger sample of patients [119].
Regarding anxiety, Lamb et al. [120] conducted a pilot intervention in 22 combat veterans with PTSD, concluding that tVNS affected the systems underlying emotional dysregulation and improved their emotional state. Bottari et al. [121] also conducted a pilot study using tVNS to improve sleep in 13 veterans, with results showing that tVNS can improve sleep depth and stability and increase parasympathetically mediated nocturnal autonomic activity in veterans with PTSD. Schwartz et al. [122] also conducted a pilot intervention with tVNS in 10 World Trade Center survivors, who still had PTSD 20 years afterwards, with the intervention reducing their stress responses and hyperarousal. Sanchez-Perez et al. [123] assessed the efficacy of three nerve stimulation modalities in reducing physiological manifestations of stress: tVNS, cervical vagus nerve stimulation (cVNS), and median nerve stimulation (tMNS) in 19 healthy young subjects exposed to three different acute mental and physiological stressors while receiving the different stimulations. The results indicated that tVNS and cVNS produced significant changes in the reduction of sympathetic outflow.
The number of both single-session (e.g.,[103,124,125,126,127]) and multi-session (e.g.[121,128,129]) tVNS interventions are increasing. There is no international consensus on the ideal parameters for tVNS, however, the increasing number of interventions and their positive effects indicate that tVNS is a promising technique for improving emotional states.
Thus, in the future, different tVNS interventions could be developed for specific emotional dysfunction profiles, e.g., transient anxiety states, PTSD, anxiety disorders, etc. Additionally, given that the vagus nerve supports the brain-microbiota axis, we also believe that future interventions should analyse both the aforementioned noradrenergic and adrenergic indicators, as well as markers of the human gut microbiota, to obtain more evidence of the effects of tVNS.
6. Conclusions
Given the complexity of the human emotional experience and its multiple intervening variables, tVNS is presented as a modest neuromodulatory technique for the dimensional physiological improvement of the interoceptive state of the subject’s emotional state. In other words, via tVNS we can influence and improve the interoceptive state. This more appropriate interoceptive state that, in terms of LeDoux’s model [11], involves a more adaptive reduction of the activation of mechanisms associated with risk detection, i.e., the amygdalar nuclei and the adrenal sympathetic axis, and, thus, would contribute positively to the construction of a more adaptive narrative of emotional experience. While, in terms of the Theory of constructed emotion [11,12,31], at the dimensional level, this more adequate interoceptive state generates an affective state that is as close as possible to a pleasant valence and an adequate activation of the organism, a consequence of the reduction of surprise and the increase of certainty in the organism’s predictive processing. This would allow a possible better execution of the categorical processing of the emotional experience. Finally, from the Polyvagal theory [13,46], stimulation of the ventral vagus involves the activation of the vagal brake, which inhibits and prevents the activation of the sympathetic adrenal axis, generating an interoceptive state of maximum well-being in the organism, which can be achieved in the absence of risk, thus allowing better cognitive processing as a result of the cerebral state of activation of the vagal brake [14].
That is to say, together with the improvement of the interoceptive state—a consequence of tVNS—which is fundamental for the adequate management of emotional processing, for subjects to have the most appropriate emotional experiences at any given moment, it will be necessary to develop, in parallel and in the most adaptive way possible, the capacity to generate coherent and healthy narratives; these, from an ethical viewpoint, will allow a non-damaging categorisation. In this way, the scientific knowledge addressed in this work will contribute to reducing the negative impact of realities generated as a consequence of inadequate management of emotional experiences, both at the individual and social levels.
Author Contributions
Complete.
Acknowledgments
No.
Conflicts of Interest
No.
References
- GBD 2019, Disease and Injuries Collaborators (2022). Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019, The Lancet Psychiatry, 9(2), 137-150. 1016. [CrossRef]
- Arias, D., Saxena, S., & Verguet, S. (2022). Quantifying the global burden of mental disorders and their economic value. Eclinicalmedicine, 54. [CrossRef]
- Hagihara, K. M. , Bukalo, O., Zeller, M., Aksoy-Aksel, A., Karalis, N., Limoges, A., Rigg, T., Campbell, T., Mendez, A., Weinholtz, C., Mahn, M., Zweifel, L. S., Palmiter, R. D., Ehrlich, I., Lüthi Andreas, & Holmes, A. (2021). Intercalated amygdala clusters orchestrate a switch in fear state. Nature: International Weekly Journal of Science, 594(7863), 403–407. [CrossRef]
- Kenwood, M. M. , Kalin, N. H., & Barbas, H. (2021). The prefrontal cortex, pathological anxiety, and anxiety disorders. Neuropsychopharmacology : At the Intersection of Brain, Behavior, and Therapeutics, 47(1), 260–275. [CrossRef]
- Zhang, A., Yang, C., Li, G., Wang, Y., Liu, P. H., Liu, Z., Sun, N., & Zhang, K. (2020). Functional connectivity of the prefrontal cortex and amygdala is related to depression status in major depressive disorder. Journal of Affective Disorders, 274, 897–902. 274, 897–902. [CrossRef]
- Bhatnagar, A., Murray, G., & Ray, S. (2023). Circadian biology to advance therapeutics for mood disorders. Trends in Pharmacological Sciences, 44(10), 689–704. [CrossRef]
- Clark, J. E. , Watson, S., & Friston, K. J. (2018). What is mood? a computational perspective. Psychological Medicine, 48(14), 2277–2284. [CrossRef]
- Panchal, P., Kaltenboeck, A., & Harmer, C. J. (2019). Cognitive emotional processing across mood disorders. Cns Spectrums, 24(1), 54–63. [CrossRef]
- Aranberri Ruiz, A. (2023). Emotional experience and its biological underpinnings: improving emotional well-being through vagal tone. Papeles Del Psicólogo - Psychologist Papers, 44(2), 95–95. [CrossRef]
- Cloninger, C. R., & Zwir, I.(2022) Genetics of human character and temperament. eLS, 3, 1-20. [CrossRef]
- LeDoux, J. E. (2021). What emotions might be like in other animals. Current Biology : Cb, 31(13), 829. [CrossRef]
- Barrett, L. F. (2017). The theory of constructed emotion: an active inference account of interoception and categorization. Social Cognitive and Affective Neuroscience, 12(1), 1–23. [CrossRef]
- Porges, S.W. (2001). The polyvagal theory: phylogenetic substrates of a social nervous system. Int J Psychophysiol.;42:123–146.
- Porges, S. W. (2023). The vagal paradox: a polyvagal solution. Comprehensive Psychoneuroendocrinology, 16. [CrossRef]
- LeDoux, J. (2012). Rethinking the emotional brain. Neuron, 73(4), 653–676. [CrossRef]
- LeDoux, J. E. (2000). Emotion circuits in the brain. Annual Review of Neuroscience, 23, 155–184. [CrossRef]
- Chin, R. , Chang, S. W. C., & Holmes, A. J. (2023). cortex: The evolution of the human brain. Psychological Review, 130(2), 285–307. [CrossRef]
- Brabec, J. , Rulseh, A., Hoyt, B., Vizek, M., Horinek, D., Hort, J., & Petrovicky, P. (2010). Volumetry of the human amygdala—An anatomical study. Psychiatry Research: Neuroimaging, 182(1), 67–72. 1016. [Google Scholar] [CrossRef]
- Pape, H.-C. , & Pare, D. (2010). Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiological Reviews, 90(2), 419–419. [CrossRef]
- Tyszka, J. M. , & Pauli, W. M. (2016). In vivo delineation of subdivisions of the human amygdaloid complex in a high-resolution group template. Human Brain Mapping, 37(11), 3979–3998. [CrossRef]
- Janak, P. H., & Tye, K. M. (2015). From circuits to behaviour in the amygdala. Nature, 517(7534), 284–292. [CrossRef]
- Milad, M. R. , & Quirk, G. J. (2012). Fear extinction as a model for translational neuroscience: Ten years of progress. Annual Review of Psychology, 63, 129–151. [CrossRef]
- LeDoux, J. E. (2014). Coming to terms with fear. Proceedings of the National Academy of Sciences, 111(8), 2871–2878. [CrossRef]
- Barger, N. , Hanson, K. L., Teffer, K., Schenker-Ahmed, N. M., & Semendeferi, K. (2014). Evidence for evolutionary specialization in human limbic structures. Frontiers in Human Neuroscience, 8, Article 277. [CrossRef]
- Yang, Z., Jian, L., Qiu, H., Zhang, C., Cheng, S., Ji, J., Li, T., Wang, Y., Li, J., & Li, K. (2021). Understanding complex functional wiring patterns in major depressive disorder through brain functional connectome. Translational Psychiatry, 11(1), 526–526. [CrossRef]
- Haris, E. M. , Bryant, R. A., Williamson, T., & Korgaonkar, M. S. (2023). Functional connectivity of amygdala subnuclei in ptsd: a narrative review. Molecular Psychiatry, 28(9), 3581–3594. [CrossRef]
- Henigsberg, N. , Kalember, P., Petrović, Z. K., & Šečić, A. (2019). Neuroimaging research in posttraumatic stress disorder - focus on amygdala, hippocampus and prefrontal cortex. Progress in Neuropsychopharmacology & Biological Psychiatry, 90, 37–42. [CrossRef]
- Millan, M. J. (2022). Agomelatine for the treatment of generalized anxiety disorder: focus on its distinctive mechanism of action. Therapeutic Advances in Psychopharmacology, 12. [CrossRef]
- Hohwy, J. (2020). New directions in predictive processing. Mind & Language, 35(2), 209–223. [CrossRef]
- Shipp, S. , Adams, R. A., & Friston, K. J. (2013). Reflections on agranular architecture: predictive coding in the motor cortex. Trends in Neurosciences, 36(12), 706–706. [CrossRef]
- Barrett, L. F. (2019). In search of emotions. Current Biology, 29(5), 142. [CrossRef]
- Barrett, L. F. (2022). Context reconsidered: complex signal ensembles, relational meaning, and population thinking in psychological science. The American Psychologist, 77(8), 894–894. [CrossRef]
- Barrett, L.F. , & Finlay, B. L. (2018). Concepts, goals and the control of survival-related behaviors. Current Opinion in Behavioral Sciences, 24, 172–179. [CrossRef]
- Craig, A.D. (2015). How Do You Feel?: an Interoceptive Moment with Your Neurobiological Self. Princeton University Press.
- Kurtin, D. L. , Giunchiglia, V., Vohryzek, J., Cabral, J., Skeldon, A. C., & Violante, I. R. (2023). Moving from phenomenological to predictive modelling: progress and pitfalls of modelling brain stimulation in-silico. Neuroimage, 272. [CrossRef]
- Barrett, L. F., & Simmons, W. K. (2015). Interoceptive predictions in the brain. Nature Reviews. Neuroscience, 16(7), 419–29. [CrossRef]
- Kleckner, I. R. , Zhang, J., Touroutoglou, A., Chanes, L., Xia, C., Simmons, W. K.,... & Feldman Barrett, L. (2017). Evidence for a large-scale brain system supporting allostasis and interoception in humans. Nature human behaviour, 1(5), 0069.
- Ventura-Bort, C. , Wendt, J., & Weymar, M. (2021). The role of interoceptive sensibility and emotional conceptualization for the experience of emotions. Frontiers in Psychology, 12. [CrossRef]
- Kuppens, P., Tuerlinckx, F., Russell, J. A., & Barrett, L. F. (2013). The relation between valence and arousal in subjective experience. Psychological Bulletin, 139(4), 917–940. [CrossRef]
- Katsumi, Y. , Kamona, N., Zhang, J., Bunce, J. G., Hutchinson, J. B., Yarossi, M., & Barrett, L. F. (2021). Functional connectivity gradients as a common neural architecture for predictive processing in the human brain. BioRxiv, 2021-09. [CrossRef]
- Katsumi, Y., Theriault, J. E., Quigley, K. S., & Barrett, L. F. (2022). Allostasis as a core feature of hierarchical gradients in the human brain. Network Neuroscience, 6(4), 1010–1031. [CrossRef]
- Barrett, L. F. , & Satpute, A. B. (2013). Large-scale brain networks in affective and social neuroscience: towards an integrative functional architecture of the brain. Current Opinion in Neurobiology, 23(3), 361–72. [CrossRef]
- Press, C., Kok, P., & Yon, D. (2020). The perceptual prediction paradox. Trends in Cognitive Sciences, 24(1), 13–24. [CrossRef]
- Touroutoglou, A., Bliss-Moreau, E., Zhang, J., Mantini, D., Vanduffel, W., Dickerson, B. C., & Barrett, L. F. (2016). A ventral salience network in the macaque brain. Neuroimage, 132, 190–197. [CrossRef]
- Wager, T. D. , Kang, J., Johnson, T. D., Nichols, T. E., Satpute, A. B., Barrett, L. F., & Diedrichsen, J. (2015). A bayesian model of category-specific emotional brain responses. Plos Computational Biology, 11(4). [CrossRef]
- Porges, S. W. (2007). The polyvagal perspective. Biological Psychology, 74(2), 116–143. [CrossRef]
- Berthoud, H. R. , & Neuhuber, W. L. (2000). Functional and chemical anatomy of the afferent vagal system. Autonomic Neuroscience: Basic & Clinicall, 85(1–3), 1–17.
- Neuhuber, W. L. , & Berthoud, H.-R. (2021). Functional anatomy of the vagus system - emphasis on the somato-visceral interface. Autonomic Neuroscience: Basic and Clinical, 236. [CrossRef]
- Gourine, A. V. , Machhada, A., Trapp, S., & Spyer, K. M. (2016). Cardiac vagal preganglionic neurones: an update. Autonomic Neuroscience : Basic & Clinical, 199, 24–8. [CrossRef]
- Porges, S. W. (1997). Emotion: an evolutionary by-product of the neural regulation of the autonomic nervous system. Annals of the New York Academy of Sciences, 807, 62–77.
- Arnsten, A. F. T. (2015). Stress weakens prefrontal networks: molecular insults to higher cognition. Nature Neuroscience, 18(10), 1376–1385. [CrossRef]
- Ruffoli, R., Giorgi, F. S., Pizzanelli, C., Murri, L., Paparelli, A., & Fornai, F. (2011). The chemical neuroanatomy of vagus nerve stimulation. Journal of Chemical Neuroanatomy, 42(4), 288–296. [CrossRef]
- Yuan, H., & Silberstein, S. D. (2018). Histamine and migraine. Headache, 58(1), 184–193. [CrossRef]
- Butt, M.F. , Albusoda, A., Farmer, A.D. & Aziz, Q. (2020), The anatomical basis for transcutaneous auricular vagus nerve stimulation. J. Anat, 236: 588-611. [CrossRef]
- Stegeman, I. , Velde, H. M., Robe, P. A. J. T., Stokroos, R. J., & Smit, A. L. (2021). Tinnitus treatment by vagus nerve stimulation: a systematic review. Plos One 16(3), 0247221. [CrossRef]
- Mather, M., & Thayer, J. F. (2018). How heart rate variability affects emotion regulation brain networks. Current Opinion in Behavioral Sciences, 19, 98–104. [CrossRef]
- De Smet, S. , Ottaviani, C., Verkuil, B., Kappen, M., & Baeken, C. (2023). Effects of non-invasive vagus nerve stimulation on cognitive and autonomic correlates of perseverative cognition. Psychophysiology, 60(6). [CrossRef]
- Goggins, E. , Mitani, S., & Tanaka, S. (2022). Clinical perspectives on vagus nerve stimulation: present and future. Clinical Science, 136(9), 695–709. [CrossRef]
- Warren, C.M. , Tona, K.D., Ouwerkerk, L., van Paridon, J., Poletiek, F., van Steenbergen, H., Bosch, J.A., Nieuwenhuis, S., (2019). The neuromodulatory and hormonal effects of transcutaneous vagus nerve stimulation as evidenced by salivary alpha amylase, salivary cortisol, pupil diameter, and the P3 event-related potential. Brain Stimul 12, 635–642. [CrossRef]
- Assenza, G. , Campana, C., Colicchio, G., Tombini, M., Assenza, F., Di Pino, G., & Di Lazzaro, V. (2017). Transcutaneous and invasive vagal nerve stimulations engage the same neural pathways: In-vivo human evidence. Brain Stimulation, 10(4), 853–854. [CrossRef]
- Peuker, E. T., & Filler, T. J. (2002). The nerve supply of the human auricle. Clinical Anatomy, 15(1), 35–37. [CrossRef]
- Kiyokawa, J. , Yamaguchi, K., Okada, R., Maehara, T. & Akita, K. (2014) Origin, course and distribution of the nerves to the posterosuperior wall of the external acoustic meatus. Anat Sci Int. Sep;89(4):238-45. [CrossRef] [PubMed]
- Watanabe, K. , Tubbs, R. S., Satoh, S., Zomorodi, A. R., Liedtke, W., Labidi, M., Friedman, A. H., & Fukushima, T. (2016). Isolated Deep Ear Canal Pain: Possible Role of Auricular Branch of Vagus Nerve-Case Illustrations with Cadaveric Correlation. World neurosurgery, 96, 293–301. [CrossRef]
- Farmer, A. D., Strzelczyk, A., Finisguerra, A., Gourine, A. V., Gharabaghi, A., Hasan, A., Burger, A. M., Jaramillo, A. M., Mertens, A., Majid, A., Verkuil, B., Badran, B. W., Ventura-Bort, C., Gaul, C., Beste, C., Warren, C. M., Quintana, D. S., Hämmerer, D., Freri, E., … Weymar, M. (2021). International consensus based review and recommendations for minimum reporting standards in research on transcutaneous vagus nerve stimulation (version 2020). Frontiers in Human Neuroscience, 14. [CrossRef]
- Shiozawa, P. , Silva, M. E. D., Carvalho, T. C. D., Cordeiro, Q., Brunoni, A. R., & Fregni, F. (2014). Transcutaneous vagus and trigeminal nerve stimulation for neuropsychiatric disorders: A systematic review. Arquivos de Neuro-psiquiatria, 72(7), 542e547.
- Sellaro, R., de Gelder, B., Finisguerra, A., & Colzato, L. S. (2018). Transcutaneous vagus nerve stimulation (tvns) enhances recognition of emotions in faces but not bodies. Cortex, 99, 213–223. [CrossRef]
- Vonck, K. , Raedt, R., Naulaerts, J., De, V. F., Thiery, E., Van, R. D., Miatton, M., Boon, P., & Aldenkamp, B. (2014). Vagus nerve stimulation...25 years later! what do we know about the effects on cognition?. Neuroscience and Biobehavioral Reviews, 45, 63–71. [CrossRef]
- Tan, C. , Yan, Q., Ma, Y., Fang, J., & Yang, Y. (2022). Recognizing the role of the vagus nerve in depression from microbiota-gut brain axis. Frontiers in Neurology, 13, 1015175–1015175. [CrossRef]
- Wang, Y. , Li, S.-Y., Wang, D., Wu, M.-Z., He, J.-K., Zhang, J.-L., Zhao, B., Hou, L.-W., Wang, J.-Y., Wang, L., Wang, Y.-F., Zhang, Y., Zhang, Z.-X., & Rong, P.-J. (2020). Transcutaneous auricular vagus nerve stimulation: from concept to application. Neuroscience Bulletin, 37(6), 853–862. [CrossRef]
- Pu-Wei, H. , Hsin-Cheng, H., Yi-Wen, L., Nou-Ying, T., Chin-Yi, C., & Ching-Liang, H. (2015). The history, mechanism, and clinical application of auricular therapy in traditional chinese medicine. Evidence-Based Complementary and Alternative Medicine, 2015. [CrossRef]
- Kong, J. , Fang, J., Park, J., Li, S., & Rong, P. (2018). Treating depression with transcutaneous auricular vagus nerve stimulation: state of the art and future perspectives. Frontiers in Psychiatry, 9. [CrossRef]
- Ferstl, M. , Teckentrup, V., Lin, W. M., Kräutlein, F., Kühnel, A., Klaus, J., Martin, W., & Kroemer, N. B. (2022). Non-invasive vagus nerve stimulation boosts mood recovery after effort exertion. Psychological Medicine, 52(14), 3029–3039. [CrossRef]
- Ventureyra, E. C. G. (2000). Transcutaneous vagus nerve stimulation for partial onset seizure therapy. Childs Nerv. Syst. 16, 101–102. [CrossRef]
- Yap, J. Y. Y. , Keatch, C., Lambert, E., Woods, W., Stoddart, P. R., & Kameneva, T. (2020). Critical review of transcutaneous vagus nerve stimulation: challenges for translation to clinical practice. Frontiers in Neuroscience, N/a. [CrossRef]
- Carreno, F. R., & Frazer, A. (2016). The allure of transcutaneous vagus nerve stimulation as a novel therapeutic modality. Biological Psychiatry, 79(4), 260–1. [CrossRef]
- Daban, C. , Martinez-Aran, A., Cruz, N., & Vieta, E. (2008). Safety and efficacy of vagus nerve stimulation in treatment-resistant depression. a systematic review. Journal of Affective Disorders, 110(1–2), 1–15. [CrossRef]
- Diedrich, A. , Urechie, V., Shiffer, D., Rigo, S., Minonzio, M., Cairo, B., Smith, E. C., Okamoto, L. E., Barbic, F., Bisoglio, A., Porta, A., Biaggioni, I., & Furlan, R. (2021). Transdermal auricular vagus stimulation for the treatment of postural tachycardia syndrome. Autonomic Neuroscience: Basic and Clinical, 236. [CrossRef]
- Nicholson, W. C. , Kempf, M.-C., Moneyham, L., & Vance, D. E. (2017). The potential role of vagus-nerve stimulation in the treatment of hiv-associated depression: a review of literature. Neuropsychiatric Disease and Treatment, 13, 1677–1689. [CrossRef]
- Barbella, G. , Cocco, I., Freri, E., Marotta, G., Visani, E., Franceschetti, S., & Casazza, M. (2018). Transcutaneous vagal nerve stimulatio (t-vns): an adjunctive treatment option for refractory epilepsy. Seizure: European Journal of Epilepsy, 60, 115–119. [CrossRef]
- Bauer, S. , Baier, H., Baumgartner, C., Bohlmann, K., Fauser, S., Graf, W., Hillenbrand, B., Hirsch, M., Last, C., Lerche, H., Mayer, T., Schulze-Bonhage, A., Steinhoff, B. J., Weber, Y., Hartlep, A., Rosenow, F., & Hamer, H. M. (2016). Transcutaneous vagus nerve stimulation (tvns) for treatment of drug-resistant epilepsy: a randomized, double-blind clinical trial (cmpse02). Brain Stimulation, 9(3), 356–363. [CrossRef]
- Badran, B. W. , Mithoefer, O. J., Summer, C. E., LaBate, N. T., Glusman, C. E., Badran, A. W., DeVries, W. H., Summers, P. M., Austelle, C. W., McTeague, L. M., Borckardt, J. J., & George, M. S. (2018). Short trains of transcutaneous auricular vagus nerve stimulation (tavns) have parameter-specific effects on heart rate. Brain Stimulation, 11(4), 699–708. [CrossRef]
- Kim, A. Y. , Marduy, A., de Melo, P. S., Gianlorenco, A. C., Kim, C. K., Choi, H., Song, J.-J., & Fregni, F. (2022). Safety of transcutaneous auricular vagus nerve stimulation (tavns): a systematic review and meta-analysis. Scientific Reports, 12(1), 22055–22055. 2205. [Google Scholar] [CrossRef]
- Burger, A. M. , D’Agostini, M., Verkuil, B., & Van Diest, I. (2020). Moving beyond belief: a narrative review of potential biomarkers for transcutaneous vagus nerve stimulation. Psychophysiology, 57(6). [CrossRef]
- Redgrave, J. , Day, D., Leung, H., Laud, P. J., Ali, A., Lindert, R., & Majid, A. (2018). Safety and tolerability of transcutaneous vagus nerve stimulation in humans; a systematic review. Brain Stimulation, 11(6), 1225–105 1238. [CrossRef]
- Bonaz, B. , Picq, C., Sinniger, V., Mayol, J. F. & Clarençon, D. (2013).Vagus nerve stimulation: From epilepsy to the cholinergic anti-inflammatory pathway. Neurogastroenterol. Motil. 25, 208–221. [CrossRef]
- Zhu, S., Zhang, X., Zhou, M., Kendrick, K. M., & Zhao, W. (2022). Therapeutic applications of transcutaneous auricular vagus nerve stimulation with potential for application in neurodevelopmental or other pediatric disorders. Frontiers in Endocrinology, 13, 1000758–1000758. [CrossRef]
- Frangos, E. , Ellrich, J., & Komisaruk, B. R. (2015). Non-invasive access to the vagus nerve central projections via electrical stimulation of the external ear: fmri evidence in humans. Brain Stimulation, 8(3), 624–36. [CrossRef]
- Komisaruk, B. R. , & Frangos, E. (2022). Vagus nerve afferent stimulation: projection into the brain, reflexive physiological, perceptual, and behavioral responses, and clinical relevance. Autonomic Neuroscience: Basic and Clinical, 237. [CrossRef]
- Banks, S. J. , Eddy, K. T., Angstadt, M., Nathan, P. J., & Phan, K. L. (2007). Amygdala-frontal connectivity during emotion regulation. Social Cognitive and Affective Neuroscience, 2(4), 303–12. [CrossRef]
- Berboth, S. , & Morawetz, C. (2021). Amygdala-prefrontal connectivity during emotion regulation: a meta-analysis of psychophysiological interactions. Neuropsychologia, 153. [CrossRef]
- Kohn, N. , Eickhoff, S. B., Scheller, M., Laird, A. R., Fox, P. T., & Habel, U. (2014). Neural network of cognitive emotion regulation — an ale meta-analysis and macm analysis. Neuroimage, 87, 345–355. [CrossRef]
- Vanderhasselt, M.-A. , Baeken, C., Van Schuerbeek, P., Luypaert, R., & De Raedt, R. (2013). Inter-individual differences in the habitual use of cognitive reappraisal and expressive suppression are associated with variations in prefrontal cognitive control for emotional information: an event related fmri study. Biological Psychology, 92(3), 433–439. [CrossRef]
- Messina, I. , Grecucci, A., & Viviani, R. (2021). Neurobiological models of emotion regulation: a meta-analysis of neuroimaging studies of acceptance as an emotion regulation strategy. Social Cognitive and Affective Neuroscience, 16(3), 257–267. [CrossRef]
- Zhou, X. , & Zhen, R. (2022). How do physical and emotional abuse affect depression and problematic behaviors in adolescents? the roles of emotional regulation and anger. Child Abuse & Neglect, 129. [CrossRef]
- Fornai, F., Ruffoli, R., Giorgi, F. S., & Paparelli, A. (2011). The role of locus coeruleus in the antiepileptic activity induced by vagus nerve stimulation. European Journal of Neuroscience, 33(12), 2169–2178. [CrossRef]
- Panaro, M. A. , Benameur, T., & Porro, C. (2020). Hypothalamic neuropeptide brain protection: focus on oxytocin. Journal of Clinical Medicine, 9(5). [CrossRef]
- D’Agostini, M., Burger, A. M., Franssen, M., Perkovic, A., Claes, S., von Leupoldt, A., Murphy, P. R., & Van Diest, I. (2023). Short bursts of transcutaneous auricular vagus nerve stimulation enhance evoked pupil dilation as a function of stimulation parameters. Cortex, 159, 233–253. [CrossRef]
- Manta, S. , Dong, J., Debonnel, G., & Blier, P. (2009). Enhancement of the function of rat serotonin and norepinephrine neurons by sustained vagus nerve stimulation. Journal of Psychiatry & Neuroscience : Jpn, 34(4), 272–80.
- Wienke, C. , Grueschow, M., Haghikia, A., & Zaehle, T. (2023). Phasic, Event-Related Transcutaneous Auricular Vagus Nerve Stimulation Modifies Behavioral, Pupillary, and Low-Frequency Oscillatory Power Responses. Journal of Neuroscience. 6319. [Google Scholar]
- Giraudier, M. , Ventura-Bort, C., Burger, A. M., Claes, N., D’Agostini, M., Fischer, R., Franssen, M., Kaess, M., Koenig, J., Liepelt, R., Nieuwenhuis, S., Sommer, A., Usichenko, T., Van Diest, I., von Leupoldt, A., Warren, C. M., & Weymar, M. (2022). Evidence for a modulating effect of transcutaneous auricular vagus nerve stimulation (tavns) on salivary alpha-amylase as indirect noradrenergic marker: a pooled mega-analysis. Brain Stimulation, 15(6), 1378–1388. [CrossRef]
- Lloyd, B., Wurm, F., de Kleijn, R., &; Nieuwenhuis, S. (2023). Short-term transcutaneous vagus nerve stimulation increases pupil size but does not affect eeg alpha power: a replication of sharon et al. (2021, journal of neuroscience). Brain Stimulation, 16(4), 1001–1008. [CrossRef]
- Ventura-Bort, C. , Wirkner, J., Genheimer, H., Wendt, J., Hamm, A. O., & Weymar, M. (2018). Effects of transcutaneous vagus nerve stimulation (tvns) on the p300 and alpha-amylase level: a pilot study. Frontiers in Human Neuroscience, 12. [CrossRef]
- Burger, A. M. , Van der Does, W., Brosschot, J. F., & Verkuil, B. (2020). From ear to eye? No effect of transcutaneous vagus nerve stimulation on human pupil dilation: a report of three studies. Biological psychology, 152, 107863. [CrossRef]
- Keute, M. , Demirezen, M., Graf, A., Mueller, N. G., & Zaehle, T. (2019). No modulation of pupil size and event-related pupil response by transcutaneous auricular vagus nerve stimulation (tavns). Scientific Reports, 9(1), 1–10. [CrossRef]
- Long-Smith, C. , O’Riordan, K. J., Clarke, G., Stanton, C., Dinan, T. G., & Cryan, J. F. (2020). Microbiota-gut-brain axis: new therapeutic opportunities. Annual Review of Pharmacology and Toxicology, 60, 477–502. [CrossRef]
- Fülling, C. , Dinan, T. G., & Cryan, J. F. (2019). Gut microbe to brain signaling: what happens in vagus…. Neuron, 101(6), 998–1002. [CrossRef]
- Han, W. , Tellez, L. A., Perkins, M. H., Perez, I. O., Qu, T., Ferreira, J., Ferreira, T. L., Quinn, D., Liu, Z.-W., Gao, X.-B., Kaelberer, M. M., Bohórquez, D. V., Shammah-Lagnado, S. J., de Lartigue, G., & de Araujo, I. E. (2018). A neural circuit for gut-induced reward. Cell, 175(3), 665–678. [CrossRef]
- Buchanan, K. L. , Klein, M. E., Barth, B. B., Montoya, M. M., & Bohórquez, D. V. (2018). A gut-brain neural circuit for nutrient sensory transduction. Science, 361(6408). [CrossRef]
- Bellono, N. W., Bayrer, J. R., Leitch, D. B., Castro, J., Zhang, C., O’Donnell, T. A., Brierley, S. M., Ingraham, H. A., & Julius, D. (2017). Enterochromaffin cells are gut chemosensors that couple to sensory neural pathways. Cell, 170(1), 185–198. [CrossRef]
- Bermúdez-Humarán, L.G. , Salinas, E., Ortiz, G. G., Ramirez-Jirano, L., Morales, J. A., & Bitzer-Quintero, O. (2019). From Probiotics to Psychobiotics: Live Beneficial Bacteria Which Act on the Brain-Gut Axis. Nutrients, 11(4), 890. [CrossRef]
- Morais, L. H. , Schreiber, Henry L., & Mazmanian, S. K. (2021). The gut microbiota–brain axis in behaviour and brain disorders. Nature Reviews.Microbiology, 19(4), 241-255. [CrossRef]
- Bany Bakar, R. , Reimann, F., & Gribble, F. M. (2023). The intestine as an endocrine organ and the role of gut hormones in metabolic regulation. Nature Reviews. Gastroenterology & Hepatology, 20(12), 784–796. [CrossRef]
- Venegas, D. P. , De, L. F. M. K., Landskron, G., Hermoso, M. A., Dijkstra, G., Faber, K. N., Gonzalez, M. J., Quera, R., & Harmsen, H. J. M. (2019). Short chain fatty acids (scfas)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Frontiers in Immunology, 10(Mar). [CrossRef]
- Agirman, G. , Yu, K. B., & Hsiao, E. Y. (2021). Signaling inflammation across the gut-brain axis. Science, 374(6571), 1087–1092. 1087. [Google Scholar] [CrossRef]
- Foster, J. A. , Rinaman, L., & Cryan, J. F. (2017). Stress & the gut-brain axis: regulation by the microbiome. Neurobiology of Stress, 7, 124–136. [CrossRef]
- Simpson, C. A. , Diaz-Arteche, C., Eliby, D., Schwartz, O. S., Simmons, J. G., & Cowan, C. S. M. (2021). The gut microbiota in anxiety and depression - a systematic review. Clinical Psychology Review, 83, 101943–101943. 1019. [Google Scholar] [CrossRef]
- Wang, Y. , Zhan, G., Cai, Z., Jiao, B., Zhao, Y., Li, S., & Luo, A. (2021). Vagus nerve stimulation in brain diseases: therapeutic applications and biological mechanisms. Neuroscience and Biobehavioral Reviews, 127, 37-53. [CrossRef]
- Fang, J., Egorova, N., Rong, P., Liu, J., Hong, Y., Fan, Y., Wang, X., Wang, H., Yu, Y., Ma, Y., Xu, C., Li, S., Zhao, J., Luo, M., Zhu, B., & Kong, J. (2017). Early cortical biomarkers of longitudinal transcutaneous vagus nerve stimulation treatment success in depression. Neuroimage. Clinical, 14, 105–111. [CrossRef]
- Hein, E. , Nowak, M., Kiess, O., Biermann, T., Bayerlein, K., Kornhuber, J., & Kraus, T. (2013). Auricular transcutaneous electrical nerve stimulation in depressed patients: a randomized controlled pilot study. Journal of Neural Transmission, 120(5), 821–7. [CrossRef]
- Rong, P.-J. , Fang, J.-L., Wang, L.-P., Meng, H., Liu, J., Ma, Y., Ben, H., Li, L., Liu, R.-P., Huang, Z.-X., Zhao, Y.-F., Li, X., Zhu, B., & Kong, J. (2012). Transcutaneous vagus nerve stimulation for the treatment of depression: a study protocol for a double blinded randomized clinical trial. Bmc Complementary and Alternative Medicine : The Official Journal of the International Society for Complementary Medicine Research(Iscmr), 12(1), 1–6. [CrossRef]
- Lamb, D. G. , Porges, E. C., Lewis, G. F., & Williamson, J. B. (2017). Non-invasive vagal nerve stimulation effects on hyperarousal and autonomic state in patients with posttraumatic stress disorder and history of mild traumatic brain injury: preliminary evidence. Frontiers in Medicine, 4. [CrossRef]
- Bottari, S. A. , Lamb, D. G., Porges, E. C., Murphy, A. J., Tran, A. B., Ferri, R., Jaffee, M. S., Davila, M. I., Hartmann, S., Baumert, M., & Williamson, J. B. (2023). Preliminary evidence of transcutaneous vagus nerve stimulation effects on sleep in veterans with post-traumatic stress disorder. Journal of Sleep Research, E13891, 13891. [CrossRef]
- Schwartz, R. M. , Shaam, P., Williams, M. S., McCann-Pineo, M., Ryniker, L., Debnath, S., & Zanos, T. P. (2022). Understanding mental health needs and gathering feedback on transcutaneous auricular vagus nerve stimulation as a potential PTSD treatment among 9/11 responders living with PTSD symptoms 20 years later: A qualitative approach. International Journal of Environmental Research and Public Health, 19(8), 4847. [CrossRef]
- Sanchez-Perez, J. A. , Gazi, A. H., Rahman, F. N., Seith, A., Saks, G., Sundararaj, S., Erbrick, R., Harrison, A. B., Nichols, C. J., Modak, M., Chalumuri, Y. R., Snow, T. K., Hahn, J.-O., & Inan, O. T. (2023). Transcutaneous auricular vagus nerve stimulation and median nerve stimulation reduce acute stress in young healthy adults: a single-blind sham-controlled crossover study. Frontiers in Neuroscience, 17. [CrossRef]
- Borgmann, D. , Rigoux, L., Kuzmanovic, B., Edwin Thanarajah, S., Münte, T. F., Fenselau, H., & Tittgemeyer, M. (2021). Technical note: modulation of fmri brainstem responses by transcutaneous vagus nerve stimulation. Neuroimage, 244. [CrossRef]
- Colzato, L. S., Ritter, S. M., & Steenbergen, L. (2018). Transcutaneous vagus nerve stimulation (tvns) enhances divergent thinking. Neuropsychologia, 111, 72–76. [CrossRef]
- Jongkees, B. J., Immink, M. A., Finisguerra, A., & Colzato, L. S. (2018). Transcutaneous vagus nerve stimulation (tvns) enhances response selection during sequential action. Frontiers in Psychology, 9, 1159–1159. [CrossRef]
- Szeska, C. , Richter, J., Wendt, J., Weymar, M., & Hamm, A. O. (2020). Promoting long-term inhibition of human fear responses by non-invasive transcutaneous vagus nerve stimulation during extinction training. Scientific Reports, 10(1). [CrossRef]
- Liu, J., Fang, J., Wang, Z., Rong, P., Hong, Y., Fan, Y., & Kong, J. (2016). Transcutaneous vagus nerve stimulation modulates amygdala functional connectivity in patients with depression. Journal of Affective Disorders, 205, 319–326. [CrossRef]
- Wu, C. , Liu, P., Fu, H., Chen, W., Cui, S., Lu, L., & Tang, C. (2018). Transcutaneous auricular vagus nerve stimulation in treating major depressive disorder: A systematic review and meta-analysis. Medicine, 97(52), e13845. [CrossRef]
Figure 1.
Generation of the emotional experience. Generating emotional experiences. The key to emotional experiences is the integration of perceptual and memory signals with signals related to brain and body states to form situational, self, and emotion schemas. These merge into an unconscious, or more accurately, a pre-conscious mental model of the emotional situation. The result of the model is a narrative that constitutes the penultimate preconscious antecedent of conscious emotional experience and is also the antecedent of verbal self-report. Adapted from LeDoux, [11].
Figure 1.
Generation of the emotional experience. Generating emotional experiences. The key to emotional experiences is the integration of perceptual and memory signals with signals related to brain and body states to form situational, self, and emotion schemas. These merge into an unconscious, or more accurately, a pre-conscious mental model of the emotional situation. The result of the model is a narrative that constitutes the penultimate preconscious antecedent of conscious emotional experience and is also the antecedent of verbal self-report. Adapted from LeDoux, [11].
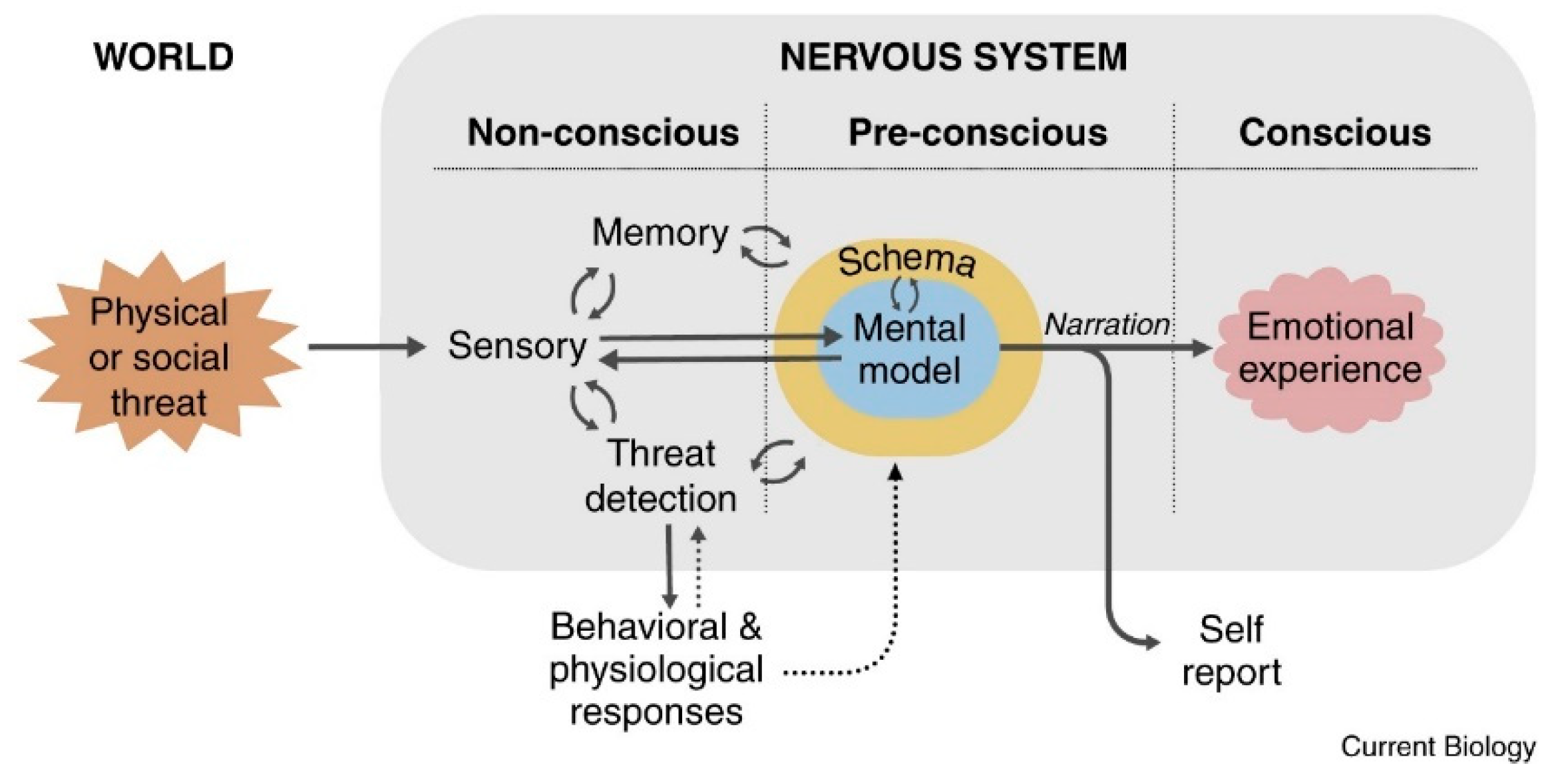
Figure 2.
tVNS device. The image shows the electrodes located in the areas innervated by the auricular vagus nerve. Adapted from Warren et al. [59].
Figure 2.
tVNS device. The image shows the electrodes located in the areas innervated by the auricular vagus nerve. Adapted from Warren et al. [59].
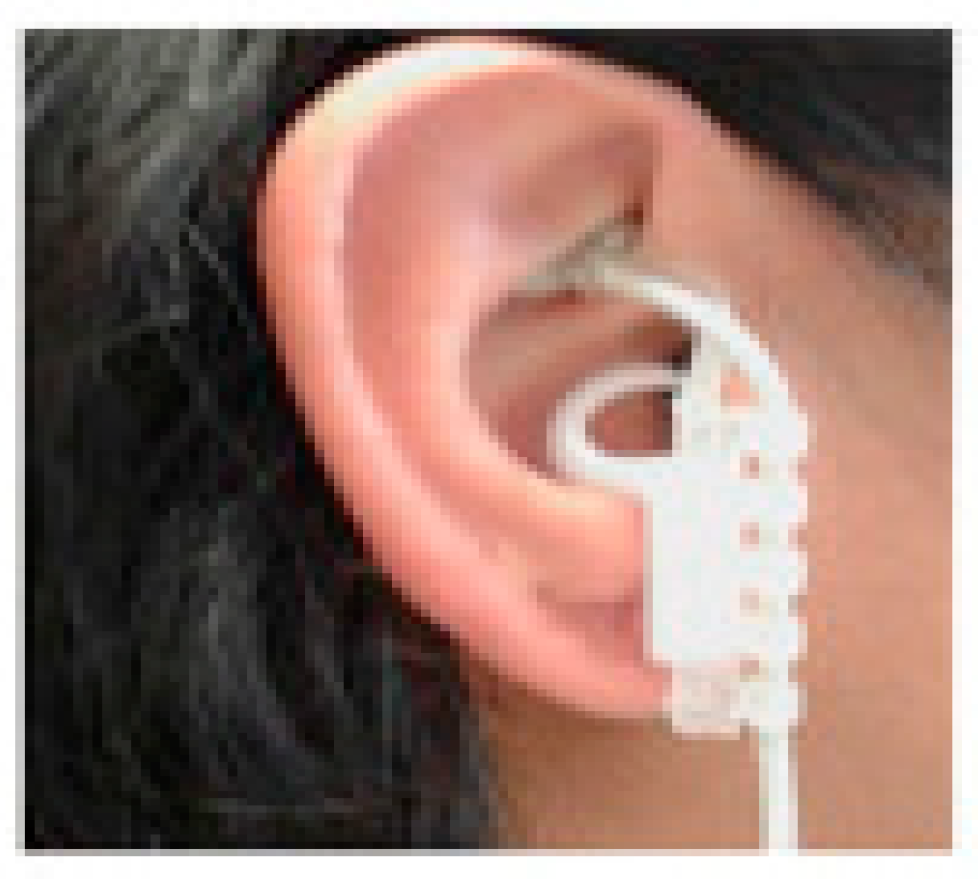
Figure 3.
Auricular branch of the vagus nerve. ABVN: Auricular Branch of Vagus Nerve. ATN: Auricular Temporal Nerve. GAN: Great Auricular Nerve. The purple areas correspond with those of the auricular vagus nerve. Adapted from Butt et al.[54].
Figure 3.
Auricular branch of the vagus nerve. ABVN: Auricular Branch of Vagus Nerve. ATN: Auricular Temporal Nerve. GAN: Great Auricular Nerve. The purple areas correspond with those of the auricular vagus nerve. Adapted from Butt et al.[54].

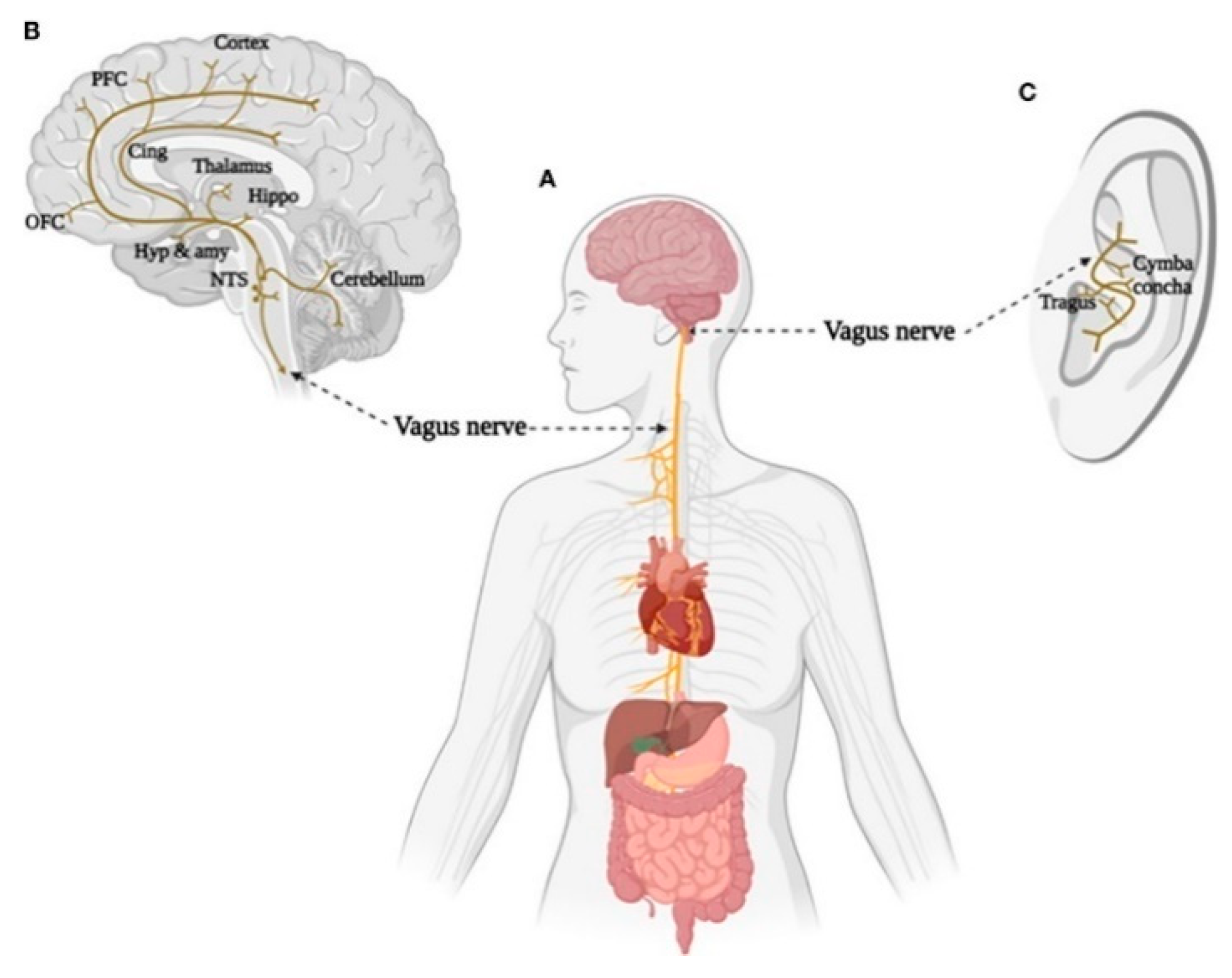
Figure 4.
Brain and body projections of the vagus nerve. (A) Illustration of the connection between the brain and major body organs via the vagus nerve. (B) Brain areas involved in the vagal afferent pathway. Nucleus of the solitary tract (NTS), hypothalamus (Hyp), amygdala (amy), hippocampus (Hippo), cingulate cortex (Cing), orbital frontal cortex (OFC), and prefrontal cortex (PFC). (C) Distribution of the vagus nerve in the external ear. Adapted from Zhu et al. [86].
Figure 4.
Brain and body projections of the vagus nerve. (A) Illustration of the connection between the brain and major body organs via the vagus nerve. (B) Brain areas involved in the vagal afferent pathway. Nucleus of the solitary tract (NTS), hypothalamus (Hyp), amygdala (amy), hippocampus (Hippo), cingulate cortex (Cing), orbital frontal cortex (OFC), and prefrontal cortex (PFC). (C) Distribution of the vagus nerve in the external ear. Adapted from Zhu et al. [86].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



