
Submitted:
15 January 2024
Posted:
16 January 2024
You are already at the latest version
Abstract
Although crop conversion from annual to perennial crops has been considered as one path towards climate-smart and resource-efficient agriculture, the effects of this conversion on soil multifunctionality and biomass yields remain unclear. Here, we investigated the effects of one annual and three perennial crops (a grass (Lolium perenne), a legume (Medicago sativa), and their mixture) on soil multifunctionality and biomass yield on the Yellow River floodplain. Soil multifunctionality was assessed by the capacity of water regulation and the multifunctionality of carbon (C), nitrogen (N), and phosphorus (P) cycles. Results showed that perennial crops enhanced soil multifunctionality by 207% for L. perenne, 311% for M. sativa, and 438% for L. perenne + M. sativa, compared with annual winter wheat (Triticum aestivum). The effect of perennial crops on soil multifunctionality increased with infiltration rate, dissolved organic C, microbial biomass C, and extracellular enzymatic activities for both C and N acquisition. However, we observed that perennial crops had lower biomass yield than annual crop. Therefore, the transition of agricultural landscapes to perennials needs to take into account the balance between environmental protection and food security, as well as environmental heterogeneity, to promote sustainable agricultural development.
Keywords:
Land cover change
; Yellow River floodplain
; Crop type
; Soil extracellular enzymes
; Soil functions
; Annual and perennial crops
1. Introduction
Annual and perennial crops supply a multitude of functions and services in agroecosystems. Annual crop provides around 80% of the global food for humans [1], which is achieved through intensive management such as mechanized tillage, fertilization, and use of pesticides, in addition to the expansion of cropping areas [2]. However, these intensive agronomic practices are now recognized as having negative impacts on soil health such as aggravating soil erosion [3], reducing soil fertility [4,5], and impairing multiple ecosystem functions and services, that is, soil multifunctionality (SMF) [6,7]. The purpose of quantifying SMF is to assess the capacity of soil to concurrently fulfill multiple functions [8]. The initial step of the methodology involves delineating the soil function and selecting the pertinent soil parameters. Subsequently, the SMF index is computed [8]. Common methods include: a single function approach, turnover approach, averaging approach, single threshold approach and multiple thresholds approach [8]. The SMF is fundamental to soil health and represents a comprehensive manifestation of soil well-being. In agroecosystems, the understanding of soil function has evolved beyond mere maximization of biomass yield, encompassing a comprehensive pursuit of coordinated objectives such as optimizing yield, enhancing soil quality, promoting environmental sustainability, and improving resource efficiency.
Given these negative impacts of annual crops on the environment, perennial crops are increasingly suggested as an alternative option to maintain SMF and biomass yield [9]. Perennial crops can play an integral role in increasing SMF and biomass yields [10,11]. For example, decreasing tillage frequency [12] can retain biomass yield [13] and increase SMF [10]. Thus, perennial crops can provide an alternative management strategy to intensive annual crops. However, the impacts of crop conversion from annual to perennial crops on SMF and biomass yield are unclear. Furthermore, the extent to which these impacts depend on the specific perennial crops remains unknown.
Numerous studies have suggested that the conversion of annual crops to perennial crops had significant impacts on SMF [14]. Generally, grasses with fibrous and rhizomatous roots densely grow near the soil surface, forming a network that reduces soil pore space, restricts water movement, and reduces soil infiltration rate [15,16]. This phenomenon contributes to improved physical properties of the soil such as total porosity and water holding capacity [17]. Moreover, leguminous plants with robust taproots and N-fixing capabilities can enhance the physicochemical properties of the soil by increasing extracellular enzymatic activities (EEAs), thereby influencing carbon (C), nitrogen (N), and phosphorus (P) cycles [18,19]. The mixed sowing of legume-grass combinations exhibits complementary rather than competitive potential in terms of growth morphology and development rhythm, utilization of soil nutrients, as well as root distributions [20,21]. However, there are few reports on the effect of a single sowing of grass, legume or mixed legume-grass combinations on SMF.
Here, we conducted a field experiment involving crop type conversion in the Yellow River floodplain, China, to explore the influences of implementing different perennial crops on SMF and biomass yield. This region is an important agroecosystem of North China, and serves as the main region of grain production in China. The SMF was assessed by measuring soil physical, chemical, and microbial properties and soil EEAs. The Z-score average of eight soil variables related to water regulation and C, N, and P cycling was calculated (details in the Materials and methods). Our aims were to explore: (1) Does the SMF respond to annual and perennial (grasses, legumes, and mixed crops of grasses-legumes) crops differently? and (2) What are the relationships between SMF and biomass yield under crop conversion from annual to perennial crops?
2. Materials and Methods
2.1. Study site and experimental design
The field experiment was located in the Yellow River floodplain in Henan Province, China (34° 66′ N, 114° 23′ E; 76 m above sea level) (Figure 1). Based on the nearby meteorological data records at the site, the average annual temperature is 14.5 °C, and the mean annual precipitation is 627 mm. The soil was classified as Fluvisols (FAO classification). At the initial stage of the study, the surface soil at 0-20 cm depth exhibited a soil bulk density of 1.59 g cm-3, a total C content of 13.2 g kg-1, a total N content of 0.82 g kg-1, a total P content of 0.88 g kg-1, an available P content of 37.5 mg kg-1, and an available N content of 11.2 mg kg-1. The pH of the soil was 8.23. Before establishing the field experiment, there was at least a 20-years history of cultivation to produce winter wheat (Triticum aestivum) and maize (Zea mays), which were common crops for the Yellow River floodplain.
The experimental platform was established in 2020, and designed to compare biomass yield of various crops and SMF. The investigated crop types were annual winter wheat, and the perennials, Lolium perenne, Medicago sativa, and L. perenne + M. sativa. The total experimental area was about 400 m2 and arranged in a split-plot design with four blocks per crop. The factor crops were allocated to the blocks, which were each subdivided into four split-plots with the factor crop types, and there were at least one meter buffer zones between adjacent blocks. The crop types split-plots consisted of L. perenne, M. sativa, a mixture of L. perenne + M. sativa, a control treatment consisting of the annual winter wheat, and there was at least one meter buffer zones between adjacent split-plots. The size of the blocks was 4 m by 20 m. The split-plot size was 4 m by 4 m (Figure 1).
2.2. Soil sample collection
In July 2021, the samples in each plot were collected with five soil cores at 0-10 cm and 10-20 cm depth and mixed for a composite soil sample at each depth. Samples were placed on ice in a cooler and transported back to the laboratory immediately after the field sampling. All soil samples were sieved using a 2 mm mesh in the lab [22]. Each sample was separated into three portions. The first portion was air-dried to measure physical properties. The second portion was for chemical properties. The third portion was stored at 4 oC for the soil EEAs and microbial biomass.
2.3. Measurements
2.3.1. Biomass yield
The annual biomass yield of winter wheat was measured by harvesting an area of 1 m × 1 m from the middle of each plot when the crop reached physiological maturity. All perennial crops were collected in July 2021. One 1 m × 1 m quadrat was selected randomly in each plot at harvest to determine perennial biomass yields. The dry weight of the plant material was measured after oven-drying at 65 oC for 48 h [23].
2.3.2. Soil physical, chemical, and microbial properties
Soil steady infiltration rate was measured by the point source method [24]. Soil water content was determined by the drying method. Soil bulk density was measured using the metal ring (100 cm3) method. Soil total porosity and water-holding capacity were measured and calculated by the method and equations proposed by [25]. Soil pH was measured in all of the soil samples by a pH electrode in a mixture of soil and water, with a soil: water ratio of 1: 2.5 (Sartorius Basic PH Meter PB-10, Göttingen, Germany). Dissolved organic C in the extracts was detected using a multi N/ C 2100S TOC-TN analyzer (Analytik Jena AG, Jena, Germany). Soil microbial biomass C and N were measured by the chloroform fumigation extraction method with conversion factors of 0.45 and 0.54 used to calculate microbial biomass C and N [26].
2.3.3. Soil extracellular enzymatic activities
Soil EEAs including β-xylosidase (BX), β-cellobiosidase (CBH), and β-1, 4-glucosidase (BG), β-1, 4-N-acetyl-glucosaminidase (NAG), L-leucine aminopeptidase (LAP), and acid phosphatase (AP) were measured from 1 g of soil using fluorometry microplates [27]. These soil EEAs were identified using 4-methyl-umbelliferone and 7-amino-4-methylcoumarin [28]. Soil EEAs were expressed in units of nmol activity g−1 dry soil h−1 (nmol g−1 h−1).
2.3.4. Assessing soil multifunctionality
The SMF is not a one-dimensional measurable process, and includes quantifying the provision of multiple ecosystem processes and services simultaneously [31]. We evaluated multifunctionality using the SMF index, obtained as the average of several functions. The index is an averaging approach and attempts to combine a collection of soil functions into a single index, so that high values of SMF mean high values of many, but not necessarily all, of the functions included.
To obtain the SMF index for each plot, we first normalized each of the two soil function categories measured (water regulation and nutrient cycle functions) using the Z-score transformation. These standardized soil functions were then averaged to obtain an SMF index [10,32]. Water regulation was composed of total porosity and water-holding capacity [17]. C cycle multifunctionality was composed of BX, BG, and CBH activities. N cycle multifunctionality was composed of NAG and LAP activities. P cycle multifunctionality was composed of AP activity. The Z-score transformation was used to standardize soil EEAs to acquire a multifunctionality index based on the methods described by [31].
where x is the measured soil EEAs, mean is the average of enzyme i, and SD is the standard deviation of enzyme i.
2.4. Statistical analysis
Statistical analyses were performed in R (version 4.1.3) by using packages nlme [33] and ggiraphExtra. All observational data were first tested for normality using the Kolmogorov-Smirnov method, and the non-normally distributed data were logarithm-or square root-transformed. Linear mixed-effects models were performed to tested the effects of crop types and soil depth on soil properties including soil water content, bulk density, steady infiltration rate, soil pH, dissolved organic C, microbial biomass C, and microbial biomass N. The block in the experiment was considered as a random effect and crop types and soil depths as fixed effect. The effects of crop types on the eight measurable proxies, the three calculated proxies, and the five multifunctionality indexes were analyzed in linear mixed models, with crop types as a fixed factor. Between-group comparisons were performed using Tukey’s post hoc tests. Pearson correlations were used to assess the relationships between SMF, biomass, and soil properties. The comprehensive analysis between SMF and biomass yield was realized with the “ggiraphExtra” package.
3. Results
3.1. Soil physical, chemical, and microbial properties
Compared with an annual crop of winter wheat, perennial crops significantly affected soil physical, chemical, and microbial properties (Table S1; Figures 2 and 3). However, the main effects of perennial crops on soil properties were limited to the topsoil with the single exception of bulk density in both topsoil and subsoil (Table S1). Compared with an annual crop of winter wheat, perennial crops (L. perenne, M. sativa and L. perenne + M. sativa) significantly increased soil water content by 47%, 40%, and 38% (Table S1; Figure 2), steady infiltration rate by 18.4 mm h-1, 26.7 mm h-1, and 22.8 mm h-1 (Figure 3), dissolved organic C by 40.1 mg kg-1, 28.4 mg kg-1, and 52.0 mg kg-1 (Table S1; Figure 2), and microbial biomass C by 89.0 mg kg-1, 63.1 mg kg-1, and 116 mg kg-1, respectively (Table S1; Figure 2).
3.2. Soil extracellular enzymatic activities
Compared with an annual crop of winter wheat, perennial crops (L. perenne, M. sativa and L. perenne + M. sativa) significantly increased C-acq EEAs by 6.09 nmol g-1 h-1, 5.19 nmol g-1 h-1, and 10.6 nmol g-1 h-1 (Figure 4), and N-acq EEAs by 2.57 nmol g-1 h-1, 1.40 nmol g-1 h-1, and 2.72 nmol g-1 h-1, respectively (Figure 4). Perennial crops (M. sativa and L. perenne + M. sativa) also significantly increased P-acq EEAs by 61.5 nmol g-1 h-1 and 51.6 nmol g-1 h-1, respectively (Figure 4). Soil depth only significantly affected C-acq and P-acq EEAs (Table S2).
3.3. Soil multifunctionality and biomass yield
Compared with an annual crop of winter wheat, perennial crops (L. perenne, M. sativa and L. perenne + M. sativa) significantly enhanced SMF by 207%, 311%, and 438% (Figure 5), C cycle multifunctionality by 188%, 151%, and 318% (Figure 6), and N cycle multifunctionality by 170%, 100%, and 150% (Figure 6). Two perennial crops (M. sativa and L. perenne + M. sativa) significantly increased P cycle multifunctionality by 214% and 190% (Figure 6). In addition, soil depth significantly affected water regulation, C cycle multifunctionality, and P cycle multifunctionality (Table S3).
In contrast, biomass yield was significantly lower in perennial crops (L. perenne, M. sativa and L. perenne + M. sativa) than in annual crops of winter wheat, by 30.2 kg ha-1, 33.5 kg ha-1, and 28.1 kg ha-1 (Figure 3). The perennial crop combination of L. perenne + M. sativa had the highest SMF and C cycle multifunctionality, whereas perennial crops of L. perenne had the highest N cycle multifunctionality and M. sativa had the highest water regulation and P cycle multifunctionality (Figure 7).
3.4. Correlations of soil multifunctionality and biomass with soil properties
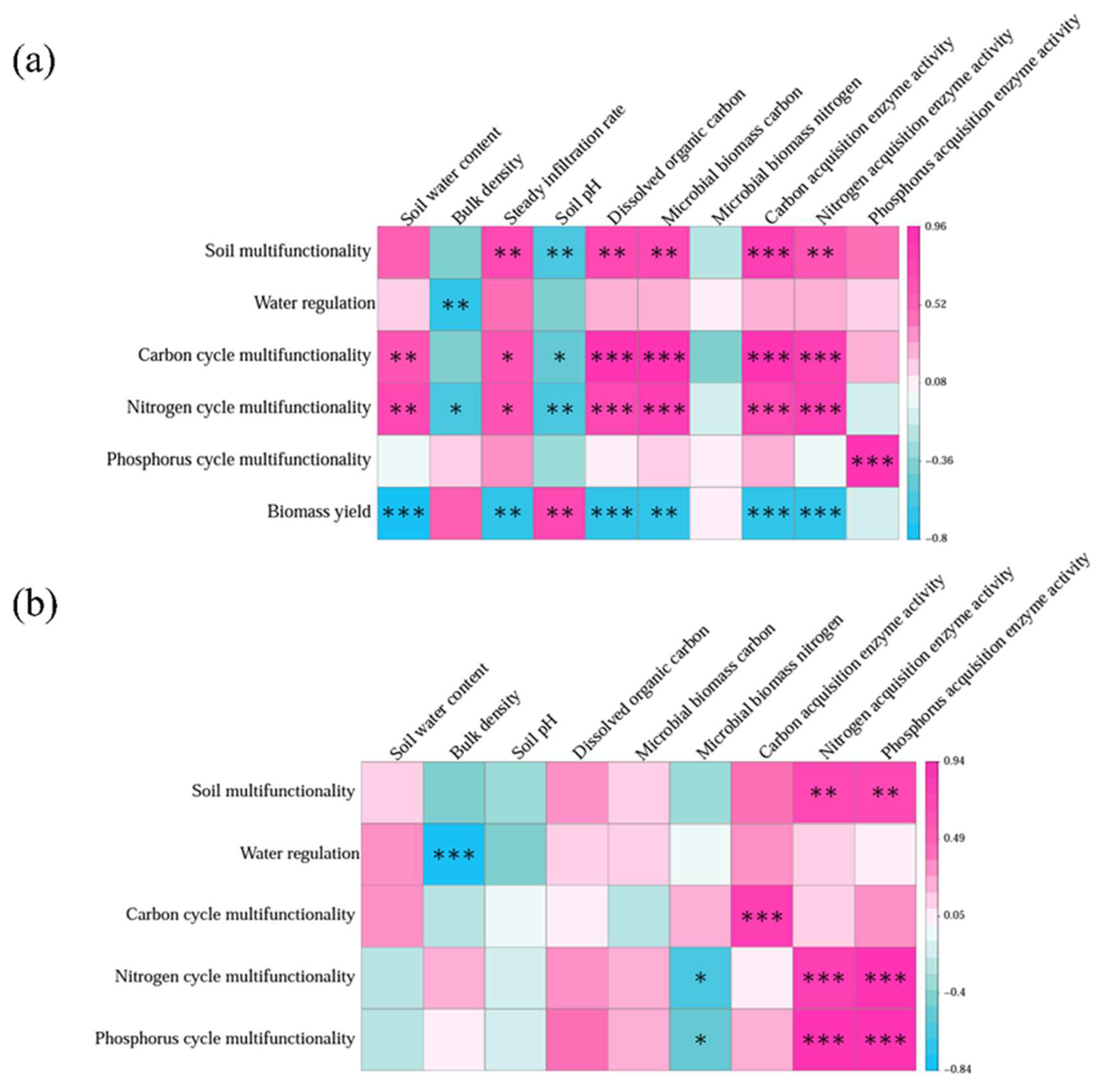
In general, SMF was positively correlated with multiple indexes related to steady infiltration rate (r = 0.71, p < 0.01), dissolved organic C (r = 0.71, p < 0.01), microbial biomass C (r = 0.71, p < 0.01), C-acq EEAs (r = 0.804, p < 0.001), and N-acq EEAs (r = 0.65, p < 0.01) in topsoil (Figure 8a). C cycle multifunctionality was positively correlated with soil water content (r = 0.63, p < 0.01), steady infiltration rate (r = 0.62, p < 0.05), dissolved organic C (r = 0.895, p < 0.001), microbial biomass C (r = 0.882, p < 0.001), and N-acq EEAs (r = 0.853, p < 0.001). Meanwhile, N cycle multifunctionality was positively correlated with soil water content (r = 0.68, p < 0.01), steady infiltration rate (r = 0.62, p < 0.05), dissolved organic C (r = 0.768, p < 0.001), microbial biomass C (r = 0.771, p < 0.001), C-acq EEAs (r = 0.747, p < 0.001). The SMF was negatively correlated with soil pH (r = -0.66, p < 0.01), which attributed to the negative correlation of C cycle multifunctionality and N cycle multifunctionality with soil pH (r = -0.56, p < 0.05, r = -0.63, p < 0.01). Water regulation was negatively correlated with bulk density (r = -0.71, p < 0.01). There was no clear relationship between SMF and soil water content, bulk density, microbial biomass N, and P-acq EEAs. Biomass yield showed significant positive correlations with soil pH (r = 0.74, p < 0.01), and negative correlations with soil water content (r = -0.804, p < 0.001), steady infiltration rate (r = -0.73, p < 0.01), dissolved organic C (r = -0.752, p < 0.001), microbial biomass C (r = -0.73, p < 0.01), C-acq EEAs (r = -0.742, p < 0.001), and N-acq EEAs (r = -0.747, p < 0.001) (Figure 8a).
In subsoil, the SMF was positively correlated with N-acq EEAs (r = 0.68, p < 0.01) and P-acq EEAs (r = 0.68, p < 0.01), which likely accounts for the observed positive correlations between N cycle multifunctionality and P-acq EEAs (r = 0.882, p < 0.001), as well as P cycle multifunctionality and N-acq EEAs (r = 0.880, p < 0.001) (Figure 8b).
3.2. Figures
Figure 1.
Location of the study area in Kaifeng, Henan Province, China. The distribution of all experiment plots. Triticum aestivum, Lolium perenne, Medicago sativa, and L. perenne + M. sativa.
Figure 1.
Location of the study area in Kaifeng, Henan Province, China. The distribution of all experiment plots. Triticum aestivum, Lolium perenne, Medicago sativa, and L. perenne + M. sativa.
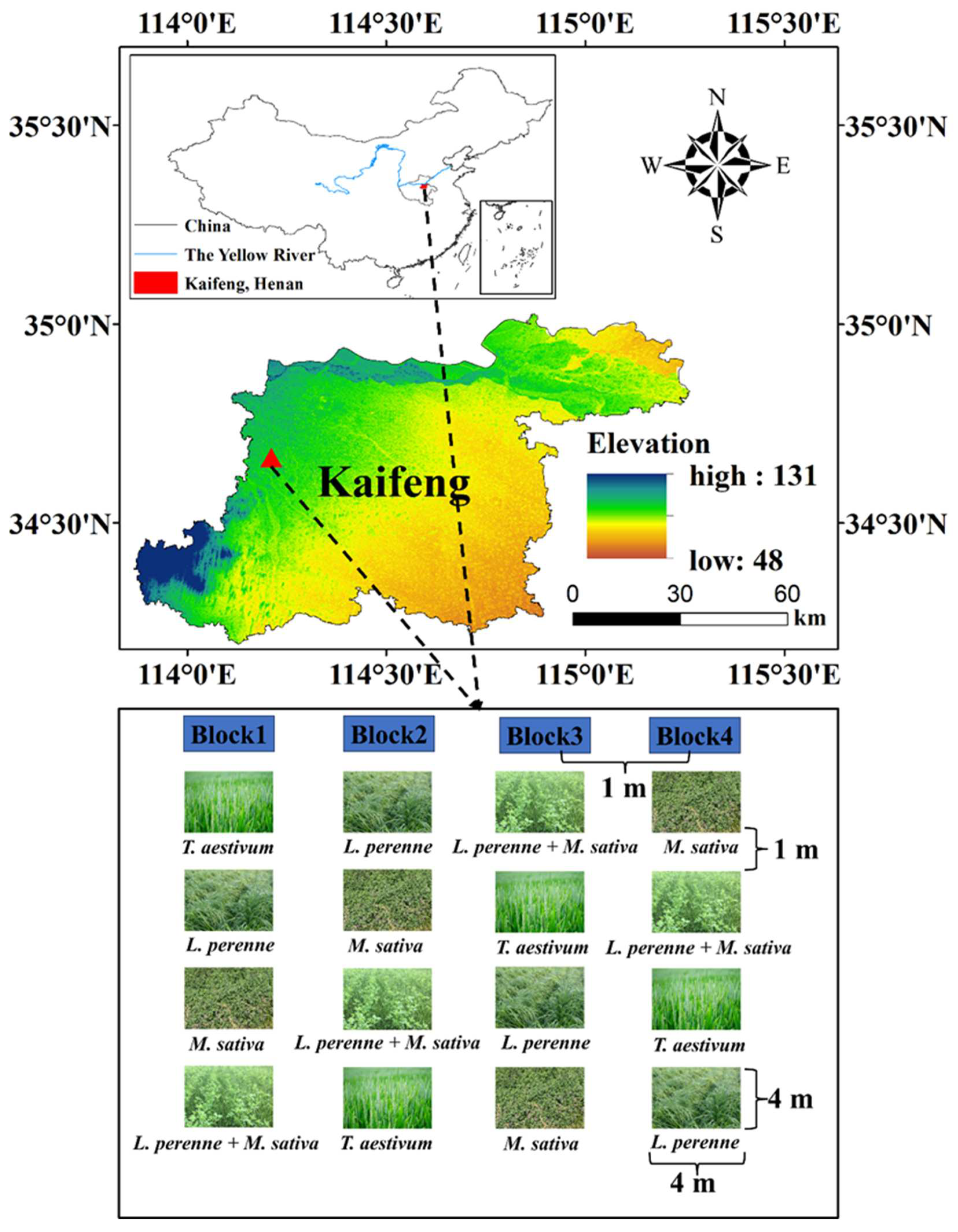
Figure 2.
Mean values are shown with standard errors for (a) soil water content, (b) bulk density, (c) soil pH, (d) dissolved organic carbon content, (e) microbial biomass carbon, and (f) microbial biomass nitrogen in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. Asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05.
Figure 2.
Mean values are shown with standard errors for (a) soil water content, (b) bulk density, (c) soil pH, (d) dissolved organic carbon content, (e) microbial biomass carbon, and (f) microbial biomass nitrogen in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. Asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05.
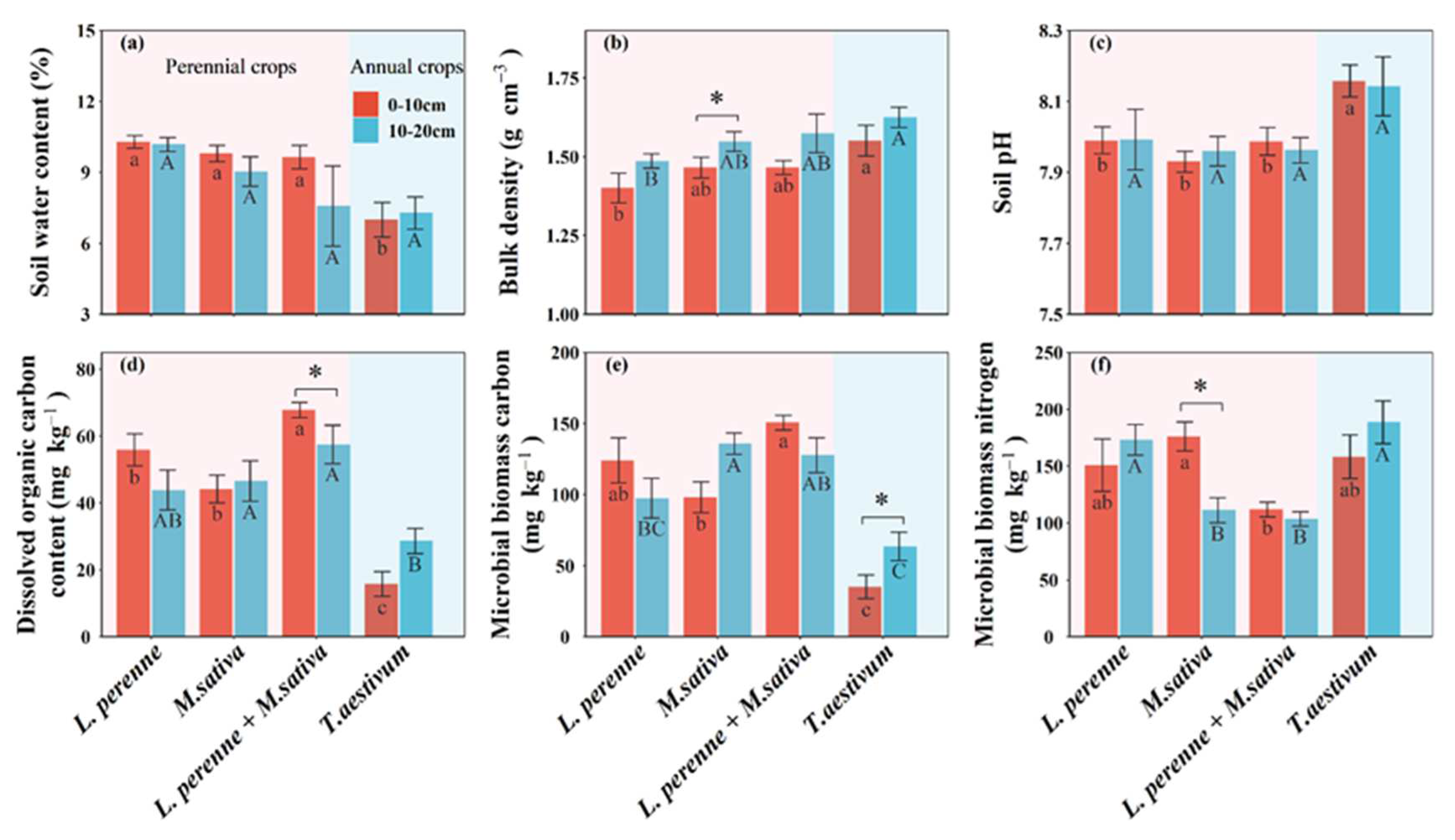
Figure 3.
Mean values are shown with standard errors for (a) biomass yield and (b) steady infiltration rate in each crop type. Note: Different letters in the bars indicate significant differences among four crop types according to Tukey's HSD tests (p < 0.05).
Figure 3.
Mean values are shown with standard errors for (a) biomass yield and (b) steady infiltration rate in each crop type. Note: Different letters in the bars indicate significant differences among four crop types according to Tukey's HSD tests (p < 0.05).
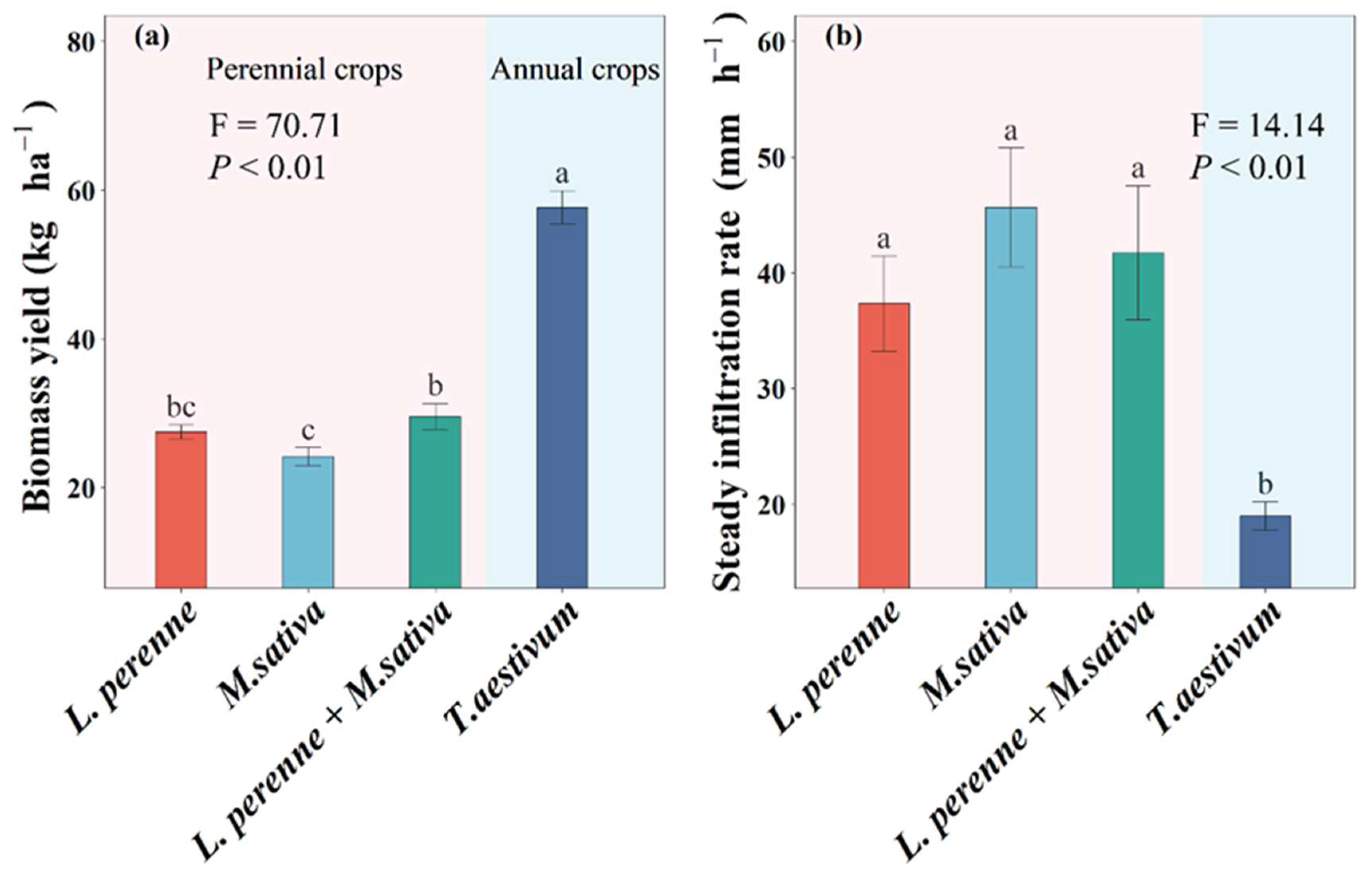
Figure 4.
Mean values are shown with standard errors for (a) carbon acquisition extracellular enzymatic activities, (b) nitrogen acquisition extracellular enzymatic activities, and (c) phosphorus acquisition extracellular enzymatic activities in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One, two, and three asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05, p < 0.01, and p < 0.001, respectively.
Figure 4.
Mean values are shown with standard errors for (a) carbon acquisition extracellular enzymatic activities, (b) nitrogen acquisition extracellular enzymatic activities, and (c) phosphorus acquisition extracellular enzymatic activities in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One, two, and three asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05, p < 0.01, and p < 0.001, respectively.
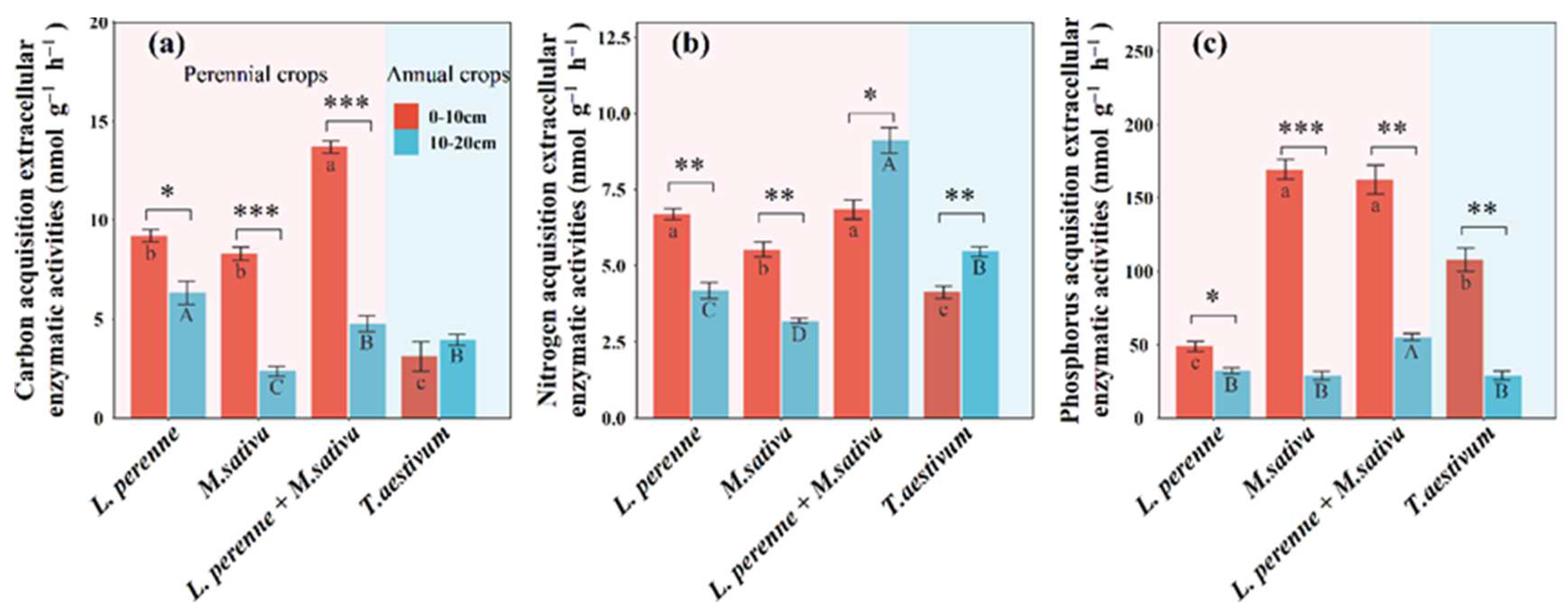
Figure 5.
Mean values are shown with standard errors for soil multifunctionality index in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One and two asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05 and p < 0.01, respectively.
Figure 5.
Mean values are shown with standard errors for soil multifunctionality index in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One and two asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05 and p < 0.01, respectively.
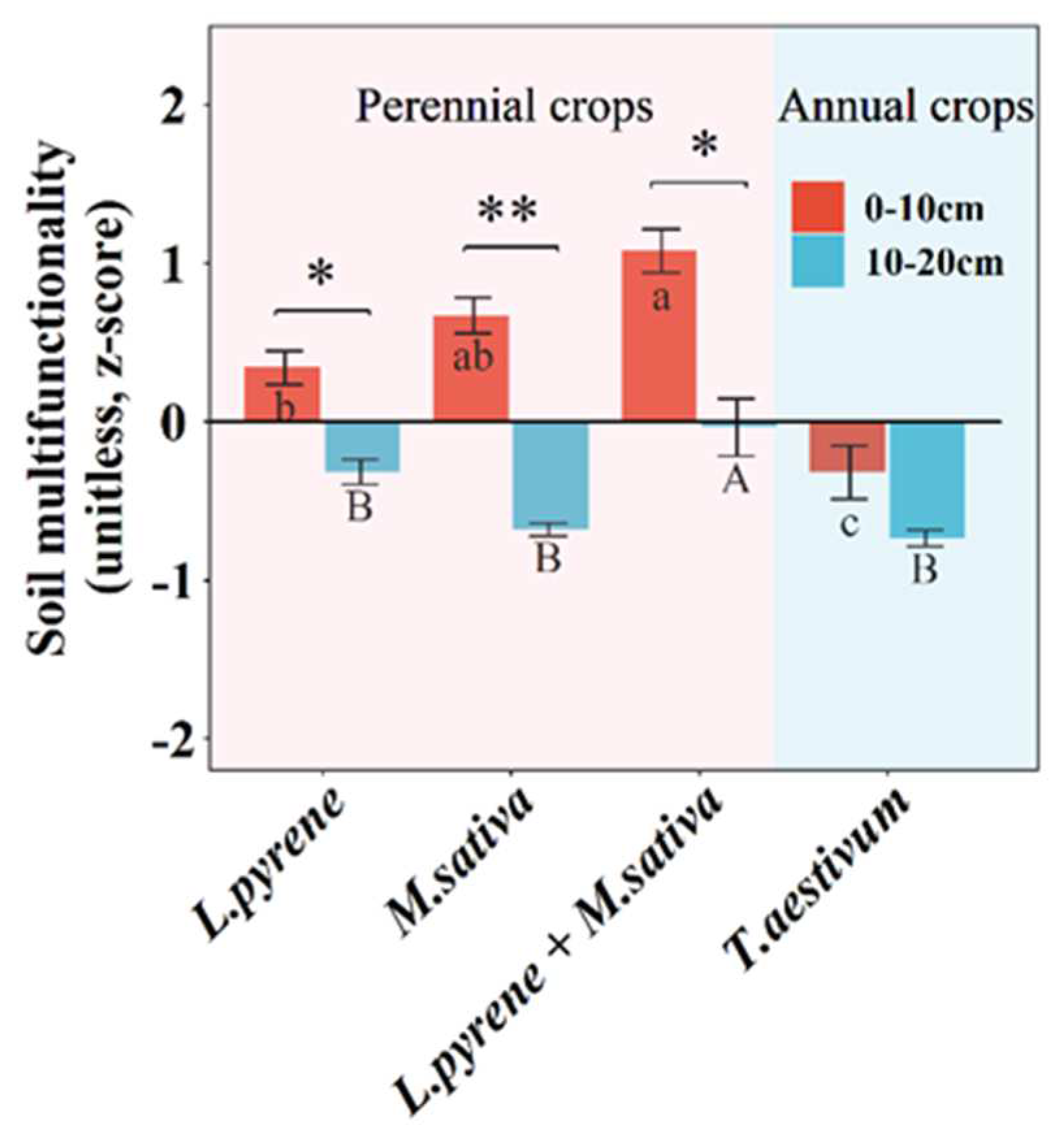
Figure 6.
Mean values are shown with standard errors for (a) soil water regulation, (b) carbon cycle multifunctionality index, (c) nitrogen cycle multifunctionality index, and (d) phosphorus cycle multifunctionality index in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One, two, and three asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05, p < 0.01, and p < 0.001, respectively.
Figure 6.
Mean values are shown with standard errors for (a) soil water regulation, (b) carbon cycle multifunctionality index, (c) nitrogen cycle multifunctionality index, and (d) phosphorus cycle multifunctionality index in each crop type and soil depth. Lowercase and upper letters denote significant differences at p < 0.05 between crop types within 0-10 cm and 10-20 cm, respectively. One, two, and three asterisks denote significant differences between 0-10 cm and 10-20 cm within each crop type at p < 0.05, p < 0.01, and p < 0.001, respectively.
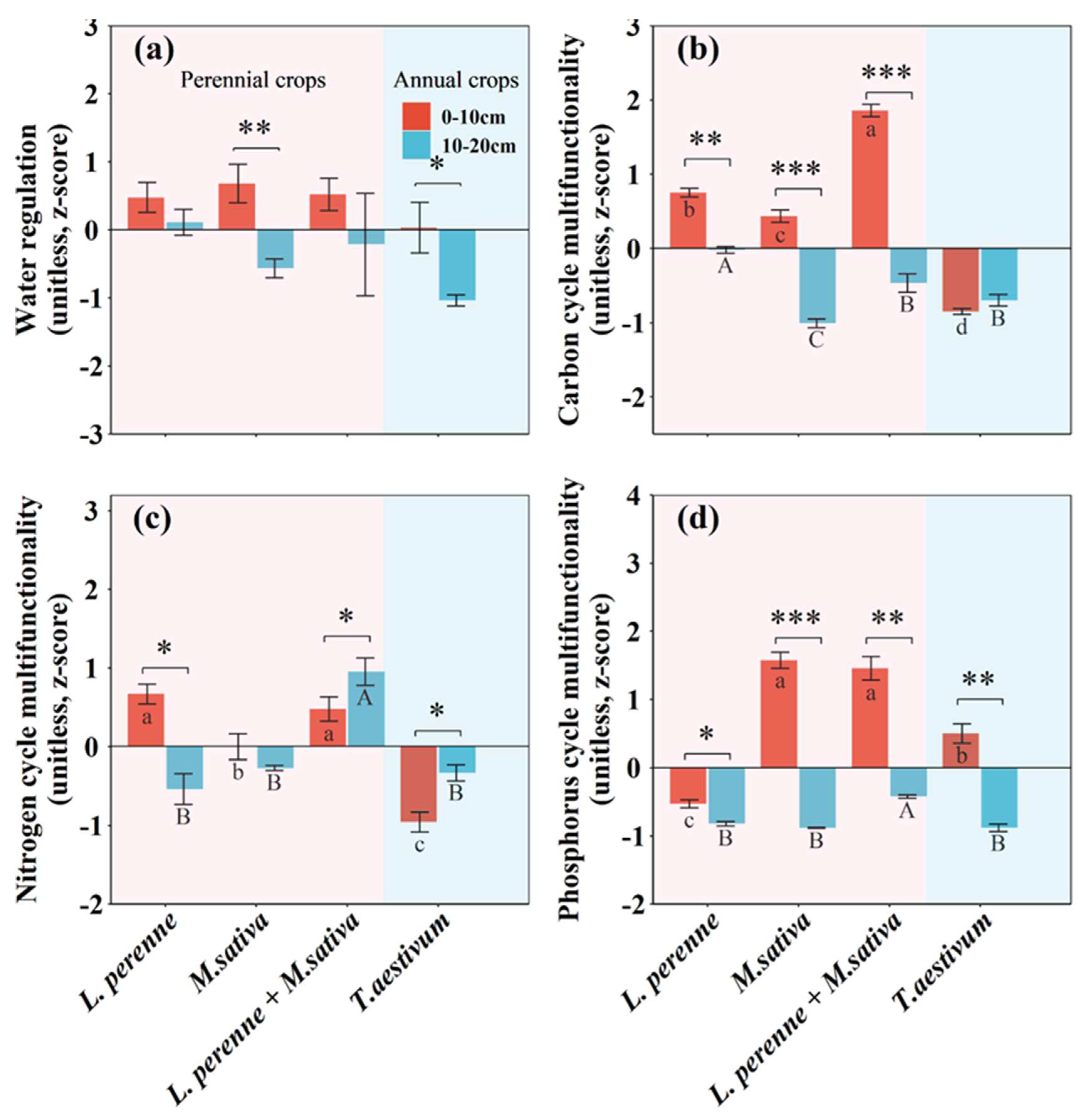
Figure 7.
Soil multifunctionality (SMF), water regulation (WR), carbon cycle multifunctionality (CCMF), nitrogen cycle multifunctionality (NCMF), phosphorus cycle multifunctionality (PCMF), and biomass yield for the T. aestivum, L. perenne, M. sativa, and L. perenne + M. sativa. To facilitate comparison among the four crop types, the data were subjected to a standardization process to allow for direct comparison.
Figure 7.
Soil multifunctionality (SMF), water regulation (WR), carbon cycle multifunctionality (CCMF), nitrogen cycle multifunctionality (NCMF), phosphorus cycle multifunctionality (PCMF), and biomass yield for the T. aestivum, L. perenne, M. sativa, and L. perenne + M. sativa. To facilitate comparison among the four crop types, the data were subjected to a standardization process to allow for direct comparison.
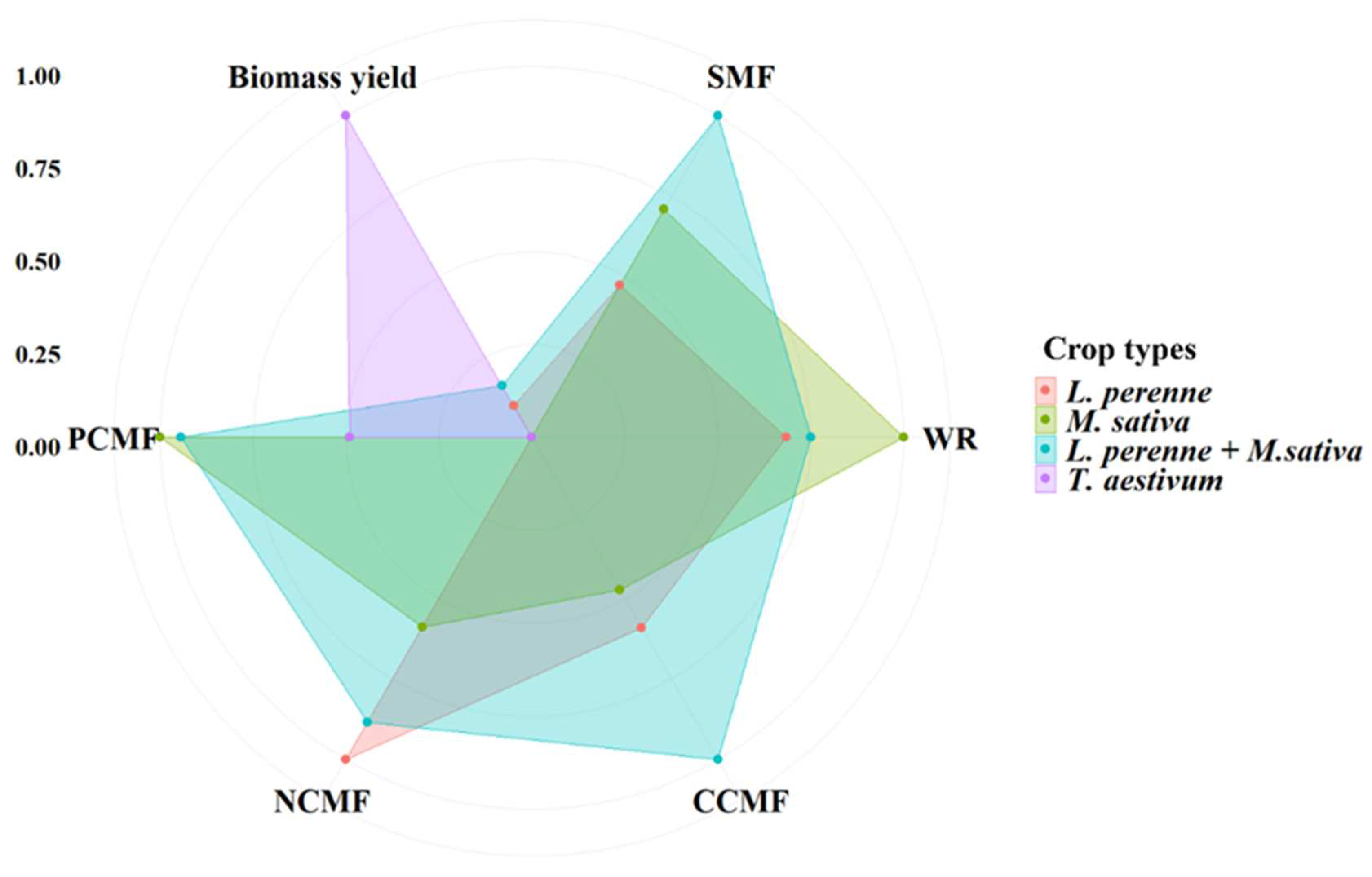
Figure 8.
Pearson’s correlation matrix among soil multifunctionality, biomass, soil properties, and soil enzyme activities at (a) 0-10 cm and (b) 10-20 cm soil depth. Blue color represents negative correlation; pink color represents positive correlation, significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
Figure 8.
Pearson’s correlation matrix among soil multifunctionality, biomass, soil properties, and soil enzyme activities at (a) 0-10 cm and (b) 10-20 cm soil depth. Blue color represents negative correlation; pink color represents positive correlation, significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
3.3. Formatting of Mathematical Components
To estimate the C-(BG, CBH and BX), N-(NAG and LAP), and P-related enzymes (AP), we used an equation to normalize the soil EEAs performing similar functions [29,30]. Taking the calculation of C acquisition (C-acq) EEAs as an example:
where x is the measured soil EEAs, mean is the average of enzyme i, and SD is the standard deviation of enzyme i.
4. Discussion
We found that perennial crops significantly increased SMF, but had varied effects on single ecosystem functions (Figures 5 and 6). [34] reported that SMF reflected a comprehensive response of all single functions, which might show positive, negative or non-significant responses. This report suggests that the effects of perennial crops on single ecosystem functions were greater in topsoil than in subsoil (Figure 6). Moreover, soil steady infiltration rate, pH, dissolved organic C, microbial biomass C, and both C-acq and N-acq EEAs were positively correlated with SMF [35], but negatively correlated with biomass yield (Figure 8a). Therefore, perennial crops, particularly mixed perennial crops, could be a promising strategy for increasing SMF in the Yellow River floodplain but at a cost to crop yield.
4.1. Perennial crops increased multiple, single functions and soil multifunctionality
Soil properties and EEAs measured in this study contributed to the effects of perennial crops on C, N, P cycling multifunctionality and SMF, but they depended strongly on the different crops. We found that the introduction of perennial crops resulted in an increase in SMF as well as C and N cycling multifunctionality (Figure 5 and Figure 6). Our results were consistent with earlier funding that complementary effects of functionally for different perennial crops promoted SMF [36]. Therefore, the incorporation of a combination of grass and legume species might represent an optimal choice for fostering sustainable development in this agroecosystem.
Compared with an annual crop of winter wheat, perennial grasses, legumes, and mixed crops of grasses-legumes increased overall SMF by enhancing C and N cycle multifunctionality. For instance, the synergistic effects of distinct single functions (such as C and N cycle multifunctionality) could mutually reinforce each other, resulting in higher SMF [37]. Indeed, perennial grasses, legumes, and mixed crops of grasses-legumes may facilitate C cycling by increasing dissolved organic C content, microbial biomass C, and C-acq EEAs, thereby increasing C cycle multifunctionality [10,30]. Moreover, perennial grasses, legumes, and mixed crops of grasses-legumes might increase microbial activity and N-acq EEAs in response to increasing dissolved organic C content, thereby increasing N cycle multifunctionality [38]. The negative correlation between water regulation and soil bulk density could counterbalance the positive effects of increased soil infiltration on water regulation (Figure 8a). These results highlight the crucial role of soil water holding capacity in water regulation, suggesting its importance over other factors [17]. In other words, regardless of whether it pertains to annual or perennial crops, the immediate impact on water regulation remains conspicuous. Our findings also suggested that P-acq EEAs play a pivotal role in distinguishing P cycle multifunctionality across different crop types [39]. The limited P-acq EEAs of perennial grasses resulted in reduced P cycle multifunctionality. Furthermore, our study underscored the importance of a neutral soil pH because it enhances SMF [40]. Compared with annual crops, perennial (grasses, legumes, and mixed crops of grasses-legumes) crops exhibited the potential to maintain a neutral soil pH. Therefore, perennial mixed crops of grasses-legumes exhibit high potential in sustaining SMF.
4.2. Relationship between soil multifunctionality and biomass
Crop types underlay the observed relationship between SMF and biomass yield. As shown in Figure 7, biomass yield was significantly higher in annual crops of winter wheat compared with perennial grasses, legumes, and mixed crops of grasses-legumes (Figure 3), although SMF was higher in perennial crops (Figure 5). We hypothesize that there are three potential factors contributing to this result. Firstly, one possible explanation is the shorter life cycle and higher rates of photosynthesis and water use efficiency observed in annual crops compared with perennial crops [41]. Furthermore, aboveground perennial mixed crops of grasses-legumes might intensify competition for light to support photosynthesis and water use efficiency [42]. Secondly due to the limited duration of planting in this study, perennial crops might exhibit underdeveloped above-ground structures; it was noteworthy that root growth exerts a substantial influence on soil dynamics, particularly in relation to C and N cycling [43]. Roots enhanced soil porosity through increased growth [44] and contributed to soil organic C content through growth and turnover [40]. Thirdly, perennial crops had been found to enhance the diversity of soil microbial communities, which had demonstrated a significant positive correlation with SMF in previous research studies [9]. Perennial crops also enhanced SMF by fostering bacterial and fungal diversity through root development [45]. Consequently, optimizing SMF and biomass yield should be further prioritized for sustainable agriculture in the future.
4.3. Implications and uncertainties
Our findings have important implications for understanding the impacts of perennial crops on SMF. For instance, perennial grasses, legumes, and mixed crops of grasses-legumes led to an increase in SMF at various levels (Figure 7). Our result is consistent with earlier funding that the maintenance of SMF is contingent upon species diversity [36]. Importantly, mixed perennial crops had the greatest effects in enhancing soil SMF and C cycle multifunctionality by significantly augmenting soil C input and EEAs (Figure 7). These findings were also consistent with previous research, indicating that mixed crops of perennials not only yield a greater quantity of matter but also provide higher energy output [46]. However, the selection of mixed perennial crops should consider not only complementary effects but also the potential for interspecific competition.
Here, we used data from one experiment as a case study to assess SMF during initial crop conversion from annual to perennial crops. The significance of our analysis lies in its demonstration of quantifiable short-term effects on SMF. It is crucial to acknowledge that we did not assign weights to individual functions, assuming equivalent importance of all functions [47]. Also, the perennial crops investigated in this study are not representative of all perennial crop types. In further research, we propose integrating additional functions, comparing a wider range of crop types, and seeking crops that could simultaneously enhance both SMF and biomass yield in both the short- and long-terms.
5. Conclusions
Our findings provide experimental evidence that the selected perennial crops can increase SMF compared to an annual crop. Perennial crops were effective for increasing SMF in an agroecological system but led to significantly lower yields than the annual crop. Mixed perennial crops have higher potential to increase SMF than single perennial crops. Finally, our results suggested that perennial crops have important implications for promoting soil functions of the agroecological system in the Yellow River floodplain. In addition, more research on potential environmental impacts is required before applying this knowledge across broad areas. Although long-term and extensive research is still required to optimize the selection of ideal annual and perennial crops, our results provide early insight by underscoring the need and the benefits of systematic research on crop-specific biomass yield and soil multifunctionality in the same agroecosystem.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Linear mixed-effects model of crop type, soil depth and their interactive effects on soil physical, chemical, and microbial properties; Table S2: Linear mixed-effects model of crop type, soil depth and their interactive effects on soil extracellular enzymatic activities; Table S3: Linear mixed-effects model of crop type, soil depth and their interactive effects on soil multifunctionality.
Author Contributions
Conceptualization, D. W. and J. C.; methodology, D. W. and J. C.; software, P.L.; validation, D. W. and J. C.; formal analysis, P. L.; investigation, P. L.; resources, D. W. and J. C.; data curation, P. L.; writing—original draft preparation, P. L.; writing—review and editing, D. W., J. C., Y. L., J. L., Y. C., G. L., C. W., C. W., and D. L M. ; visualization, P. L.; supervision, D. W. and J. C.; project administration, D. W. and J.C.; funding acquisition, D. W. and J.C. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by EU H2020 Marie Skłodowska-Curie Actions (No. 839806), Aarhus University Research Foundation (AUFF-E-2019-7-1), Danish Independent Research Foundation (1127-00015B), Nordic Committee of Agriculture and Food Research, Aarhus University iClimate, and Natural Science Basic Research Program of Shaanxi Program (No.2022JQ-897), and the National Natural Science Foundation of China (NSFC32130066, 32371672).
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
This study is granted by EU H2020 Marie Skłodowska-Curie Actions (No. 839806), Aarhus University Research Foundation (AUFF-E-2019-7-1), Danish Independent Research Foundation (1127-00015B), Nordic Committee of Agriculture and Food Research, Aarhus University iClimate, and Natural Science Basic Research Program of Shaanxi Program (No.2022JQ-897), and the National Natural Science Foundation of China (NSFC32130066, 32371672).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pimentel, D.; Cerasale, D.; Stanley, R.C.; Perlman, R.; Newman, E.M.; Brent, L.C.; Mullan, A.; Chang, D.T.I. Annual vs. perennial grain production. Agric. Ecosyst. Environ. 2012, 161, 1–9. [Google Scholar] [CrossRef]
- Malik, A.A.; Puissant, J.; Buckeridge, K.M.; Goodall, T.; Jehmlich, N.; Chowdhury, S.; Gweon, H.S.; Peyton, J.M.; Mason, K.E.; van Agtmaal, M.; Blaud, A.; Clark, I.M.; Whitaker, J.; Pywell, R.F.; Ostle, N.; Gleixner, G.; Griffiths, R.I. Land use driven change in soil pH affects microbial carbon cycling processes. Nat Commun. 2018, 9, 3591. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Geng, Y.; Fu, X.Z.; Coulter, J.A.; Chai, Q. The effects of wind erosion depending on cropping system and tillage method in a semi-arid region. Agronomy. 2020, 10(5), 732. [Google Scholar] [CrossRef]
- Singh, B.K.; Trivedi, P.; Egidi, E.; Macdonald, C.A.; Delgado-Baquerizo, M. Crop microbiome and sustainable agriculture. Nat Rev Microbiol. 2020, 18, 601–602. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Z.; Bao, X.; Sun, J.; Yang, S.; Wang, P.; Wang, C.; Wu, J.; Liu, X.; Tian, X.; Wang, Y.; Li, J.; Wang, Y.; Xia, H.; Mei, P.; Wang, X.; Zhao, J.; Yu, R.; Zhang, W.; Che, Z.; Gui, L.; Callaway, R.M.; Tilman, D.; Li, L. Long-term increased grain yields and soil fertility from intercropping. Nat Sustain. 2021, 4, 943–950. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Chu, H.; Eldridge, D.J.; Gaitan, J.J.; Liu, Y.R.; Sokoya, B.; Wang, J.T.; Hu, H.W.; He, J.Z.; Sun, W.; Cui, H.; Alfaro, F.D.; Abades, S.; Bastida, F.; Diaz-Lopez, M.; Bamigboye, A.R.; Berdugo, M.; Blanco-Pastor, J.L.; Grebenc, T.; Duran, J.; Illan, J.G.; Makhalanyane, T.P.; Mukherjee, A.; Nahberger, T.U.; Penaloza-Bojaca, G.F.; Plaza, C.; Verma, J.P.; Rey, A.; Rodriguez, A.; Siebe, C.; Teixido, A.L.; Trivedi, P.; Wang, L.; Wang, J.; Yang, T.; Zhou, X.Q.; Zhou, X.; Zaady, E.; Tedersoo, L.; Delgado-Baquerizo, M. Soil biodiversity supports the delivery of multiple ecosystem functions in urban greenspaces. Nat Ecol Evol. 2023, 7, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, J.E.K.; Gamfeldt, L.; Isbell, F.; Lefcheck, J.S.; Griffin, J.N.; Hector, A.; Cardinale, B.J.; Hooper, D.U.; Dee, L.E.; Emmett Duffy, J. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods Ecol. Evol. 2014, 5, 111–124. [Google Scholar] [CrossRef]
- Cappelli, S.L.; Domeignoz-Horta, L.A.; Loaiza, V.; Laine., A.L. Plant biodiversity promotes sustainable agriculture directly and via belowground effects. Trends Plant Sci. 2022, 27, 674–687. [Google Scholar] [CrossRef]
- Garland, G.; Edlinger, A.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Pescador, D.S.; Herzog, C.; Romdhane, S.; Saghai, A.; Spor, A.; Wagg, C.; Hallin, S.; Maestre, F.T.; Philippot, L.; Rillig, M.C.; van der Heijden, M.G.A. Crop cover is more important than rotational diversity for soil multifunctionality and cereal yieldss in European cropping systems. Nat Food. 2021, 2(1), 28–37. [Google Scholar] [CrossRef]
- Chen, J.; Manevski, K.; Lærke, P.E.; Jørgensen, U. Biomass yields, yields stability and soil carbon and nitrogen content under cropping systems destined for biorefineries. Soil Tillage Res. 2022, 221, 105397. [Google Scholar] [CrossRef]
- Lanker, M.; Bell, M.; Picasso, V.D. Farmer perspectives and experiences introducing the novel perennial grain Kernza intermediate wheatgrass in the US Midwest. Renewable Agriculture and Food Systems, 2020, 35 (6), 653-662. [CrossRef]
- Qiao, Z.; Yates, T.B.; Shrestha, H.K.; Engle, N.L.; Flanagan, A.; Morrell-Falvey, J.L.; Sun, Y.; Tschaplinski, T.J.; Abraham, P. E.; Jessy Labbé.; Wang, Z.Y.; Hettich, R.L.; Tuskan, R.A.; Wellington Muchero.; Chen, J.G. Towards engineering ectomycorrhization into switchgrass bioenergy crops via a lectin receptor-like kinase. Plant Biotechnol. J, 2021, 19, 2454-2468. [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; White, C. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst, 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Archer, N.A.L.; Quinton, J.N.; Hess, T.M. Below-ground relationships of soil texture, roots and hydraulic conductivity in two-phase mosaic vegetation in South-east Spain. J. Arid Environ, 2002, 52, 535–553. [Google Scholar] [CrossRef]
- Halli, H.M.; Govindasamy, P.; Chaudhary, M.; Srinivasan, R.; Prasad, M.; Wasnik, V.K.; Yadav, V.K.; Singh, A.K.; Kumar, S.; Vijay, D.; Pathak. H. Range grasses to improve soil properties, carbon sustainability, and fodder security in degraded lands of semi-arid regions. Sci. Total Environ, 2022, 851, 158211. [CrossRef]
- Zhang, J.; Feng, Y.; Maestre, F.T.; Berdugo, M.; Wang, J.; Coleine, C.; Saez-Sandino, T.; Garcia-Velazquez, L.; Singh, B.K.; Delgado-Baquerizo, M. Water availability creates global thresholds in multidimensional soil biodiversity and functions. Nat Ecol Evol, 2023, 7, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Luo, Y.; Li, J.; Zhou, X.; Cao, J.; Wang, R.W.; Wang, Y.; Shelton, S.; Jin, Z.; Walker, L.M.; Feng, Z.; Niu, S.; Feng, W.; Jian, Zhou, S.L. Costimulation of soil glycosidase activity and soil respiration by nitrogen addition. Glob. Chang. Biol, 2017, 23, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Sun, L.; Liu, Y.; Liu, Y.F.; López-Vicente, M.; Wei, X.H.; Wu, G.L. Alfalfa planting significantly improved alpine soil water infiltrability in the Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ, 2019, 285, 106606. [Google Scholar] [CrossRef]
- Li, L.; Li, S.M.; Sun, J.H.; Zhou, L.L.; Bao, X.G.; Zhang, H.G.; Zhang, F. S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci, 2007, 104, 11192–11196. [Google Scholar] [CrossRef]
- Cox, S.; Peel, M.D.; Creech, J.E.; Waldron, B.L.; Eun, J.; Zobell, D.R.; Miller, R.L.; Snyder, D.L. Forage production of grass–legume binary mixtures on Intermountain western USA irrigated pastures. Crop Sci, 2017, 57, 1742–1753. [Google Scholar] [CrossRef]
- Ye, J.S.; Delgado-Baquerizo, M.; Soliveres, S.; Maestre, F.T. Multifunctionality debt in global drylands linked to past biome and climate. Glob. Chang. Biol, 2019, 25 (6), 2152-2161. [CrossRef]
- Wang, D.; Liu, C.; Yang, Y.; Liu, P.; Hu, W.; Song, H.; Miao, C.; Chen, J.; Yang, Z.; Miao, Y. Clipping decreases plant cover, litter mass, and water infiltration rate in soil across six plant community sites in a semiarid grassland. Sci. Total Environ, 2023.861,160692. [CrossRef]
- Su, L.; Yang, Y.S.; Li, X.Y.; Wang, D.; Liu, Y.C.; Liu, Y.Z.; Yang, Z.L.; Li, M.M. Increasing plant diversity and forb ratio during the revegetation processes of trampled areas and trails enhances soil infiltration. Land Degrad. Dev, 2018, 29, 4025–4034. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.F.; Cui, Z.; Liu, Y.; Wang, D.; Tian, F.P.; Wu, G.L. Natural grasslands maintain soil water sustainability better than planted grasslands in arid areas. Agric. Ecosyst. Environ, 2019, 286, 106683. [Google Scholar] [CrossRef]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem, 1985, 17, 837–842. [Google Scholar] [CrossRef]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on poten tial soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA. Soil Biol. Biochem, 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Marx, M.-C.; Wood, M.; Jarvis, S.C. A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem, 2001, 33, 1633–1640. [Google Scholar] [CrossRef]
- García-Ruiz, R.; Ochoa, V.; Hinojosa, M.B.; Carreira, J.A. Suitability of enzyme activities for the monitoring of soil quality improvement in organic agricultural systems. Soil Biol. Biochem, 2008, 40 (9), 2137-2145. [CrossRef]
- Jia, R.; Zhou, J.; Chu, J.; Shahbaz, M.; Yang, Y.; Jones, D.L.; Zang, H.; Razavi, B.S.; Zeng, Z. Insights into the associations between soil quality and ecosystem multifunctionality driven by fertilization management: a case study from the North China plain. J. Clean. Prod, 2022, 362, 132265. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat Commun, 2016, 7, 10541. [Google Scholar] [CrossRef]
- Shi, X.; Wang, J.; Lucas-Borja, M.E.; Wang, Z.; Li, X.; Huang, Z. Microbial diversity regulates ecosystem multifunctionality during natural secondary succession. J. Appl. Ecol, 2021, 58 (12), 2833-2842. [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: linear and nonlinear mixed effects models. R package version, 2013, 3 (1), 111. https://cran.r-project.org/web/ packages/nlme/index.html.
- Giling, D.P.; Beaumelle, L.; Phillips, H.R. P.; Cesarz, S.; Eisenhauer, N.; Ferlian, O.; Gottschall, F.; Guerra, C.; Hines, J.; Sendek, A.; Siebert, J.; Thakur, M.P.; Barnes, A.D. Aniche for ecosystem multifunctionality in global change research. Glob. Chang. Biol, 2019, 25, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, Q.; Zhu, H.; Reich, P.B.; Banerjee, S.; van der Heijden, M.G.A.; Sadowsky, M.J.; Ishii, S.; Jia, X.; Shao, M.; Liu, B.; Jiao, H.; Li, H.; Wei, X. Erosion reduces soil microbial diversity, network complexity and multifunctionality. ISME J, 2021, 15, 2474–2489. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.M.; Bailey, R.A.; Dossena, M.; Gamfeldt, L.; Reiss, J.; Trimmer, M.; Woodward, G. Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes. Glob Chang Biol, 2015, 21, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Xiang, Y.; Seabloom, E.; Chen, H.Y. H.; Wang, J.; Yu, G.; Deng, Y.; Li, Z.; Niu, S. Ecosystem restoration and belowground multifunctionality: a network view. Ecol. Appl, 2022, 32, e2575. [Google Scholar] [CrossRef]
- Shen, H.J.; Zhang, Q.Q.; Zhang, X.; Jiang, X.Y.; Zhu, S.G.; Chen, A.F.; Wu, Z.; Xiong, Z.Q. In situ effects of biochar field-aged for six years on net N mineralization in paddy soil. Soil Tillage Res, 2021, 205, 104766. [Google Scholar] [CrossRef]
- Benavent-González, A.; Delgado-Baquerizo, M.; Fernández-Brun, L.; Singh, B.K.; Maestre, F.T.; Sancho, L.G. Identity of plant, lichen and moss species connects with microbial abundance and soil functioning in maritime Antarctica. Plant Soil, 2018, 429 (1-2), 35-52. [CrossRef]
- Zheng, J.; Zhang, F.; Zhang, B.; Chen, D.; Li, S.; Zhao, T.; Wang, Q.; Han, G.; Zhao, M. Biodiversity and soil pH regulate the recovery of ecosystem multifunctionality during secondary succession of abandoned croplands in northern China. J. Environ. Manage, 2023, 327.116882. [CrossRef]
- Albert, Á.; Kelemen, A.; Valkó, O.; Miglécz, T.; Csecserits, A.; Rédei, T.; Deák, B.; Tóthmérész, B.; Török, P. Secondary succession in sandy old fields: a promising example of spontaneous grassland recovery. Appl. Veg. Sci, 2014, 17, 214–224. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. Journal of Ecology, 2003, 91, 707–720. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, L.; Huang, Z.; Lopez-Vicente, M.; Wu, G.L. Root morphological characteristics and soil water infiltration capacity in semi-arid artificial grassland soils. Agric Water Manag, 2020, 235, 106153. [Google Scholar] [CrossRef]
- Zhao, D.; Xu, M.; Liu, G.; Ma, L.; Zhang, S.; Xiao, T.; Peng, G. Effect of vegetation type on microstructure of soil aggregates on the Loess Plateau, China. Agric. Ecosyst. Environ, 2017, 242, 1–8. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N. A.; Gallardo, A.; Garcia-Velazquez, L.; Hart, S.C.; Hayes, P.E.; He, J.Z.; Hseu, Z.Y.; Hu, H.W.; Kirchmair, M.; Neuhauser, S.; Perez, C.A.; Reed, S.C.; Santos, F.; Sullivan, B.W.; Trivedi, P.; Wang, J.T.; Weber-Grullon, L.; Williams, M.A.; Singh, B.K. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat Ecol Evol, 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Xu, R.; Shi, W.; Kamran, M.; Chang, S.; Jia, Q..; Hou, F. Grass-legume mixture and nitrogen application improve yield, quality, and water and nitrogen utilization efficiency of grazed pastures in the loess plateau. Front. Plant Sci, 2023, 14, 088849. [Google Scholar] [CrossRef]
- Wittwer, R.A.; Bender, S.F.; Hartman, K.; Hydbom, S.; Lima, R.A.A.; Loaiza, V.; Nemecek, T.; Oehl, F.; Olsson, P.A.; Petchey, O.; Prechsl, U.E.; Schlaeppi, K.; Scholten, T.; Seitz, S.; Six, J.; van der Heijden, M.G.A. Organic and conservation agriculture promote ecosystem multifunctionality. Sci. Adv, 2021, 7 (34). [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



