
Submitted:
15 January 2024
Posted:
16 January 2024
You are already at the latest version
Abstract
Neonatal disorders, particularly those resulting from prematurity, pose a major challenge to health care and have a significant impact on infant mortality and long-term child health. The limitations of current therapeutic strategies emphasize the need for innovative treatments. New cell-free technologies utilizing extracellular vesicles (EVs) offer a compelling opportunity for neonatal therapy by harnessing the inherent regenerative capabilities of EVs. These nanoscale particles, secreted by a variety of organisms including animals, bacteria, fungi and plants, contain a repertoire of bioactive molecules with therapeutic potential. This review aims to provide a comprehensive assessment of the therapeutic effects and mechanistic insights of EVs from stem cells, biological fluids and non-animal sources, with a focus on common neonatal conditions such as hypoxic-ischemic encephalopathy, respiratory distress syndrome, bronchopulmonary dysplasia and necrotizing enterocolitis. This review summarizes the evidence for the therapeutic potential of EVs, analyzes the evidence on their mechanisms of action, and discusses the challenges associated with the implementation of EV-based therapies in neonatal clinical practice.
Keywords:
extracellular vesicles
; mesenchymal stromal cells
; neonatal pathologies
; premature newborns
; cell therapy
1. Introduction
The development of pathologies in newborns and premature infants is often attributed to prematurity and the complications associated with it. Prematurity often leads to early neonatal complications and increases the risk of adverse outcomes through the occurrence of severe pathologies. The World Health Organization reports that the incidence of preterm birth varies internationally, ranging from 4% to 15% of all births [1,2]. Neonatal pathologies include various conditions that affect infants within the first 28 days of life and can lead to long-term impairment or severe disability, depending on their severity. These diseases usually affect several organ systems and are often accompanied by extensive inflammatory processes. The most common neonatal pathologies include hypoxic-ischemic encephalopathy (HIE), respiratory distress syndrome (RDS), bronchopulmonary dysplasia (BPD), sepsis and necrotizing enterocolitis (NEC) [3].
Despite advances that have increased survival rates, complications such as chronic lung disease, brain injury and neonatal pulmonary hypertension remain common and pose a significant risk of permanent complications affecting the nervous system, lungs, heart and metabolism [4]. The treatment of neonatal pathologies requires an interdisciplinary approach involving a team of anesthesiologists, neonatologists, pediatricians, pediatric surgeons, neuropathologists, rehabilitation specialists and others to make early diagnoses and implement innovative treatments that complement traditional therapies [5,6]. Currently, pharmacologic interventions in the early perinatal period are one of the main therapeutic strategies, but they can have side effects and are not consistently effective [7]. Therefore, exploring novel treatments for acute neonatal diseases is critical to improving patient outcomes and reducing the incidence of neonatal disability. Preclinical and clinical studies of stem cell therapy have yielded encouraging results in a number of neonatal conditions, including chronic lung disease, intraventricular hemorrhage, and HIE. These studies focus on the use of cells that have cytoprotective, reparative and regenerative properties [4,8,9].The therapeutic effect of stem cells is thought to be primarily due to paracrine effects, largely mediated by the secretion of extracellular vesicles (EVs) [10]. EVs are considered a novel but under-researched therapeutic approach with similar efficacy to cell therapy. Current research on the biology of EVs suggests that these particles can regulate intercellular communication and transfer biologically active molecules to recipient cells. Although initial research focused on EVs from human and animal sources, including stem, progenitor and stromal cells, more recent findings have identified alternative EV sources such as plants, fungi and bacteria that offer additional opportunities for therapeutic applications [11,12].
The aim of this article is to review and critically analyze the evidence on the biological effects and molecular mechanisms of EVs from various sources. It systematically summarizes the evidence for the therapeutic potential of EVs, focusing on findings from animal studies modeling neonatal pathology, and discusses the challenges associated with the clinical translation of EV-based therapies in neonatology.
1.1. Searching Criteria
In conducting this review, we systematically searched scientific databases, including PubMed and ClinicalTrials.gov database from January 1, 2000 to June 1, 2023. The following keywords were used in the search: 'hypoxic-ischemic encephalopathy', 'respiratory distress syndrome', 'bronchopulmonary dysplasia', 'sepsis', 'necrotizing enterocolitis', 'cell therapy', 'extracellular vesicles', 'exosomes', 'microvesicles', and 'neonatology'. In this review, we refer to 'extracellular vesicles' in a broad sense. However, where specific terms such as 'exosomes' are mentioned, these are cases where the authors have provided convincing evidence that the particles have all the characteristics defined in the 2018 MISEV guidelines [13].
1.2. Application of the 'Therapeutic Unit' Concept to Assess the Equivalence of EVs Concentrations
Our analysis of the current literature revealed a variety of regimens and dosages for the treatment by EVs, which makes direct comparison of studies difficult. To address this issue, we used the concept 'therapeutic unit', as proposed by Kordelas et al. [14], as a standard measure to normalize the administered doses of EVs. According to their work, one therapeutic unit corresponds to the amount of EVs produced by 4 x 107 human bone marrow-derived MSCs within 48 hours. This amount is approximately 1.3–3.5 × 1010 particles/unit and contains 0.5–1.6 mg protein/unit. Despite possible variations in cultivation conditions, we chose to convert all reported doses to therapeutic units using simple mathematical proportions. Furthermore, we have assumed that EVs production under standard culture conditions (culture medium, temperature, CO2 concentration) is linear during the first 2-3 days, an assumption confirmed by unpublished data from our laboratory. However, this assumption should be taken with caution as productive abilities cells in vitro does not necessarily follow linear kinetics and differences in laboratory protocols could influence EVs production rates.
By applying this standardization to the reported data, we aimed to make a more accurate comparison between studies and provide a speculative assessment of therapeutic effects depending on the EVs dose administered. However, it should be noted that our approach relies on several assumptions and simplifications that may affect the reliability of the resulting comparisons. Further standardization of the methods used to produce and quantify EVs in this area is therefore essential to enable more robust analyzes and comparisons between different studies.
2. Biogenesis and Classification of EVs
EVs are small particles consisting of a lipid bilayer that are released by cells into the extracellular space. EVs carry a cargo of proteins, nucleic acids and other biomolecules and play a crucial role in intercellular communication and the modulation of immune responses. The first mention of EVs was by P. Wolf in 1967, who obtained the pellet of EVs by ultracentrifugation of platelet-poor plasma [13]. Later it was shown that all cells release different types of EVs into the extracellular environment and participate in intercellular communication [14]. The initial classification of EVs was based on the mechanisms of intracellular secretion and was categorized into three groups: 1) exosomes (diameter 40–150 nm), which are secreted after fusion of multivesicular bodies with the cell membrane; 2) microvesicles or ectosomes (diameter 100–1000 nm), which are formed by budding from the plasma membrane into the intercellular space, and 3) apoptotic bodies (100–5000 nm), which are formed during apoptosis [15].
It should be noted that modern methods do not always allow the isolation of pure EVs fractions and thus the determination of the intracellular origin of the vesicles studied [16]. This fact and the discovery of new types of EVs have led to the need to reconsider the classification of EVs. The International Society for Extracellular Vesicles has published recommendations called "minimal information for studies of EVs" (MISEV) [12]. The recommendations suggest classifying EVs based on their physical characteristics: size, density, presence of a lipid bilayer, biochemical properties (CD63+/CD81+, carrier of annexin A5, etc.) and according to the source and conditions of origin (EVs from podocytes, hypoxic EVs, large oncosomes, etc.). Therefore, the proposed nomenclature classifies small vesicles (20-200 nm), including exomers (≤ 50 nm) and supermers (≥ 25 nm) without lipid bilayer, exosomes (40-130 nm) and defensosomes (approximately 80 nm). Large EVs with a diameter >200 nm include microvesicles (100–1 µm), migrasomes (500–3000 nm), apoptotic bodies (50 nm –5 µm) and large oncosomes (1–10 µm) [12; 14].
The most studied of all types of EVs are exosomes, whose biochemical characteristics ensure a high degree of cargo stability during transportation. The exosomal membrane is organized in the form of a lipid bilayer. It contains phospholipids, cholesterol, and phosphatidylethanolamines, which increase the stability of exosomes against cargo biodegradation, thus providing efficient transportation. [15,16].
3. The Cargo of EVs
EVs are known to transport a variety of cargo, including proteins, nucleic acids (various types of RNA and DNA), various carbohydrates and other biologically active molecules such as lipids. In some cases, organelles such as mitochondria or their components can also be detected in EVs [17,18]. Proteins and microRNAs (miRNAs) are thought to contribute significantly to the biological functions modulated by EVs. Current proteome and miRNA profiles are continuously being integrated into online repositories such as EVpedia [19], ExoCarta [20] and Vesiclepedia [21], which facilitate subsequent bioinformatic analyses of the potential role of these macromolecules. In total, more than 1900 proteins have been identified in the proteome of bone marrow-derived MSC-EVs by mass spectrometry [22]. These proteins are functionally associated with self-renewal, differentiation, cell migration and proliferation. It has been observed that the proteome of MSC-EVs is enriched with proteins involved in angiogenesis under hypoxic conditions [23]. Moreover, proteomic analysis of MSC-EVs from human placenta revealed 745 proteins associated with functions related to angiogenesis, neurogenesis, immunoregulation, protection against apoptosis and oxidative stress. In particular, this analysis clearly identified proteins involved in the Wnt and phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) signaling pathways, which have been shown to be associated with neuroprotection[24]. The neuroprotective properties appear to be related to the fundamental functions of MSCs. For example, it has been demonstrated that during incubation of bone marrow-derived MSCs with IFN-γ, these cells produce EVs containing a large quantity of neuroprotective molecules (such as laminin β2, aggrecan, testican-1, periostin) and a variety of microRNAs with anti-inflammatory effects, for instance, microRNA-200b and microRNA-146b [25]. The therapeutic effects have been shown to depend on the presence of both proteins and microRNAs in the content of MSC-EVs. The degradation of microRNAs led to a reduction in the observed neuroprotective effects in a murine model of autoimmune encephalomyelitis[25]. Moreover, a range of microRNAs associated with neuroprotection, such as microRNA-133b, microRNA-21-5p, microRNA-22-3p, microRNA-31, and microRNA-146a-5p, have been identified in the content of human MSC-EVs [26,27,28,29,30].
It is known that EVs produced by MSCs and other cell types contain a diverse spectrum of RNA molecules. These include messenger RNA (mRNA), microRNA (miRNA), long non-coding RNA (lncRNA), small interfering RNA (siRNA), PIWI-interacting RNA (piRNA), ribosomal RNA (rRNA) fragments, Y-RNA, small nuclear RNA (snRNA) and circular RNA (circRNA) [31,32]. CircRNAs, a subgroup of non-coding RNAs that are generated by non-sequential backsplicing of exons, introns or a combination of both. Their presence may facilitate the translation of unique small peptides through internal ribosome entry site -dependent mechanisms [33]. The regulation of the expression of this type of RNA has been shown to be crucial for the maintenance of normal neurogenesis [34]. CircRNAs have been shown to be involved in the regulation of autophagy, proliferation, apoptosis and the cell cycle [35]. Currently, the link between the dysregulation of their expression and the progression of neurodegenerative [36] and cardiovascular [35] diseases has been established.
lncRNAs are RNA transcripts longer than 200 nucleotides that do not contain coding sequences for proteins. They are categorized into different groups, including sense, antisense, bidirectional, intronic and intergenic, based on their proximity to protein-coding transcripts. They are expressed at low levels and are unique for each cell type. It has been shown that lncRNA play a certain role in regulating gene expression during development and differentiation at the post-transcriptional level by influencing chromatin structure and methylation, although the exact mechanisms are still unknown [37]. To date, only a small number of functional lncRNA have been characterized, some of which have been found in human breast milk EVs. For example, certain lncRNA such as growth arrest specific 5 (GAS5), steroid receptor activator 1 RNA (SRA1) and colorectal neoplasia differentially expressed (CRNDE) have been identified [37]. These lncRNA play an important role in the regulation of neonatal metabolism. For example, GAS5 has been shown to inhibit the activation of glucocorticoid-responsive genes under conditions of starvation and cellular stress, which may be an adaptive mechanism to conserve energy resources in neonatal cells [38]. SRA1 acts as a coactivator of the transcriptional regulatory factor peroxisome proliferator-activated receptor gamma (PPARγ), which is responsible for adipogenesis [39]. CRNDE is involved in the regulation of signaling pathways triggered by insulin and insulin-like growth factor [40]. In addition, breast milk contains lncRNAs that are likely involved in the regulation of the neonatal immune system. For example, the lncRNA DANCR has been shown to regulate the expression of IL6 and TNFα genes in peripheral blood monocytes [41]. In addition, studies have shown that the lncRNA MALAT1, which is present in MSC-EVs, regulates various therapeutic targets, particularly inflammation, suggesting significant potential for the treatment of acute brain injury [42].
piRNAs are short sequences of 21 to 35 nucleotides that are associated with regulatory proteins from the PIWI family. These piRNA-PIWI complexes play a central role in maintaining the functional integrity of stem cells, mainly by suppressing retrotransposon mobilization in genomic sequences [43]. In particular, piRNAs such as hsa_piR_017723_DQ594464 and hsa_piR_020814_DQ598650 have been identified in MSC-EVs from human bone marrow. These piRNAs contribute to the protection of umbilical cord blood-derived stem cells from apoptosis [44].
Y-RNA is a relatively a lesser-known class of non-coding RNAs. They have been observed in various human cell types, with particularly high expression in brain and heart tissue [45]. To date, they have been found to regulate DNA replication, stabilize RNA transcripts and mediate defense mechanisms in response to cell stress [46]. For example, Y-RNA-1, which is highly enriched in MSC-EVs compared to the cells of origin, was found to play a crucial role in the protective effects of MSC-EVs on TNF-α/ActD-mediated apoptosis of hepatocytes, as siRNA-mediated silencing of Y-RNA-1 reduces these effects in vitro [47]. It has also been shown that exosomes derived from cardiosphere-derived cells are enriched in small RNA components, predominant the Y RNA fragment EV-YF1, provide therapeutic effects following myocardial infarction [48].
Current research has clearly demonstrated the crucial function of lipids in the formation of membrane architecture and their importance as biological signaling molecules. For example, cholesterol, which is transported in large excess by EVs T- lymphocytes when transferred to peripheral blood mononuclears, contributes to their production of proinflammatory TNF-α [49]. EVs contain various lipid components such as sphingomyelin, cholesterol, lysophosphatidylcholine, arachidonic acid, fatty acids, prostaglandins and leukotrienes both in their membranes and as cargo [50]. It was also found that the lipid composition of EVs membranes of different cell types is unique. However, the exact mechanisms by which the lipid composition of the EVs membrane influences its function are still unclear [51]. It is important to note that the difference in lipid composition of EVs membranes between normal and pathologic cells is an important diagnostic criterion. [52]. In addition, enzymes involved in lipid metabolism and contributing to their conversion into biologically active signaling molecules have been identified in the proteome of EVs. [50]. For example, CD14 dendritic cells and macrophages produce small EVs containing leukotriene C4 hydrolases and synthases which are involved in the production of proinflammatory lipid mediators [53]. In vitro studies have shown that human bone marrow-derived MSCs, when cultured with polyunsaturated fatty acids such as arachidonic acid, eicosapentaenoic acid and docosahexaenoic acid, produce EVs that contain these fatty acids in their membrane. These fatty acids are known to be the precursors of eicosanoids and resolvins. [54]. Furthermore, incubation of mouse MSCs with excess docosahexaenoic acid in vtro resulted in the synthesis of resolvin D2 in MSC- EVs [55]. EVs have also been shown to contain unique lipid signaling molecules specific to certain cell types. For example, microglial cells produce EVs containing endocannabinoids such as N-arachidonylethanolamine and 2-arachidonylglycerol, which inhibit the neuronal release of glutamine neurotransmitters and GABA, thus regulating the balance of excitation and inhibition in the transmission of signals between neurons [56].
Increasing research into the molecular nature of EVs has led to a more sophisticated understanding of cell-to-cell communication and insights into the pathophysiological mechanisms of a number of diseases. This has fostered the development of novel diagnostic tools and therapeutic strategies that take advantage of the molecular cargo of EVs. Nevertheless, it must be recognized that this scientific field is still in its infancy. Extracellular vesicles are characterized by a multitude of hundreds of biologically active components, such as proteins, lipids, RNAs and metabolites, each of which can exert different effects. Indeed, the heterogeneity of these components can lead to a variety of outcomes, some of which may exhibit synergistic or antagonistic interactions, adding to the complexity of their biological functions. Consequently, the net biological impact of EVs may depend not only on their molecular nature, but also on the current functional state of the recipient cells. Such recipient cells may interpret and respond to the vesicular signals in a context-dependent manner, which is influenced by their physiological or pathophysiological state, their signaling history, and the microenvironment in which they reside. However, it is worth noting that this branch of research is still quite young and further research is needed to fully understand the complexity of the mechanisms mediated by biologically active molecules in the cargo of EVs and their clinical application.
4. Mechanisms of EVs Interaction with Recipient Cell Membranes
EVs serve as important mediators of intercellular communication by transporting biologically active and informational molecules from donor cells to recipient cells. Once released, EVs can influence the microenvironment and be transported through body fluids to reach distant target cells. EVs can be internalized into target cells by different mechanisms, which can be divided into specific (internalization) and non-specific (integration of the EV lipid bilayer into the cell membrane structure by interaction of the hydrophobic regions of the lipids). Internalization can be phagocytosis-mediated and/or micropinocytosis-mediated as well as by clathrin-, lipid raft- and caveolin-mediated endocytosis [57]. During endocytosis, EVs are taken up by the cell through an invagination of the plasma membrane, allowing biologically active EV molecules to be released directly into the cytoplasm of the cell [43]. Once inside the cell, EVs release their content of proteins, nucleic acids, lipids and metabolites, which can modulate cellular signal transduction, gene expression and cellular functions and influence a variety of physiological and pathological processes. In addition, proteins and/or sialic acid on the surface of EVs can mediate specific interactions with target cells. For example, the presence of the tetraspanin CD63 on the surface of EVs allows specific interaction with neuronal and glial cells, whereas the absence of CD63 allows interaction only with dendritic cells [15]. Similarly, the presence of sialic acid on the surface of EVs allows specific binding to HeLa cells through interaction with the CD33 receptor [58]. The major histocompatibility complex on surface of dendritic cell-derived EVs allows them to activate T- helper cells [59]. EVs carrying tumor necrosis factor (TNF), Fas ligand (FasL) and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) on their surface are able to induce apoptosis in tumor cells if the receptors for these ligands are present on their surface [60].
A comprehensive understanding of the mechanisms underlying vesicular-cellular interactions is essential to elucidate their significance in both physiological and pathological states and to advance the development of targeted vesicular transport systems. For example, engineered extracellular vesicles carrying the rabies virus glycoprotein on their membrane have been used for the precise delivery of cargo to neuronal cells [61]. These exosomes efficiently transported bioactive components to the cortex and hippocampus and led to a reduction in amyloid plaque deposition, beta-amyloid levels and inflammation in transgenic mice (APP/PS1) [62].
5. Sources of EVs and Methods for Their Isolation
The therapeutic potential of EVs is mediated by their molecular cargo, which is inherited from parent cells [63]. First of all, the composition of the proteins on their surface determines the specificity of EVs to interact with certain target cells. Accordingly, the proper selection of EVs sources may allow the achievement of the greatest therapeutic efficacy in relation to specific types of cells or tissues involved in the pathological process [64]. EVs can be derived from cultured native or genetically modified cells, as well as from most biological fluids of the body (Figure 1) [65]. Additionally, intercellular communication via EVs has been demonstrated in organisms of non-animal origin, including plants, bacteria and fungi. This suggests that tissues and fluids of these species may also be potential sources of EVs [66].
The conditioned culture medium is a widely used source of EVs, which can be isolated from it by various methods [67]. The use of special culture media can increase the yield of EVs compared to standard cultivation conditions [68]. In most studies on the therapy of neonatal pathologies, EVs are isolated from conditioned stem or progenitor cell culture media [69]. The most common and best-studied source of EVs are MSCs [70], which are isolated from bone marrow, adipose tissue or postpartum umbilical cord/placenta [71]. MSCs from adipose tissue are a good source of autologous material and suitable objects for obtaining EVs [72]. The biology of these cells is quite well studied, and therapeutic effects have been confirmed in animal models of neonatal diseases such as BPD [73], HIE [74], NEK [75], congenital retinal diseases [76] and sepsis [77]. Furthermore, EVs derived from perinatal MSC tissues have shown promising therapeutic potential for the treatment of acute pathologic conditions of the brain [74,78,79]. EVs may be derived from either autologous or allogeneic sources. However, it is crucial to recognize that neonatal pathological conditions often present as acute emergencies. Experimental studies in model systems have demonstrated the therapeutic potential of EVs during these acute phases. Consequently, the use of allogeneic EVs is frequently necessitated, given the time constraints associated with the cultivation of autologous MSCs and the subsequent production of vesicle preparations, which typically require 2-3 weeks.
Despite the focus on MSCs in research, alternative sources of vesicles with therapeutic effects have also been identified. An important source of EVs can be immune cells such as T cells, B cells and macrophages. These cells release EVs that play a key role in intercellular communication, pathogenesis and immunomodulation [80]. In particular, macrophage-derived EVs with high levels of CD14 were shown to attenuate significant organ damage in a sepsis mouse model [81]. EVs from human endothelial progenitor cells have been implicated in reducing organ damage in sepsis model by restoring vascular permeability [82]. EVs from human and murine epicardial cells also show the ability to prevent the development of infarction when administered locally to the injury zone in newborn mice [83]. In addition, EVs derived from human plasma were found to protect neonatal rat cardiomyocytes against apoptosis under glucose and oxygen deprivation [84]. Cultures of specialized cells differentiated from induced pluripotent stem cells represent a promising EV source. For example, vesicles from neuronal and astroglial cultures have shown neuroprotective effects in experimental models [85,86,87]. In particular, EVs from intestinal neural stem cells have been associated with gastrointestinal protection against necrotizing enterocolitis in preterm rat pups [88]. Despite these findings, it remains unclear whether all human stem/progenitors or somatic cells can produce EVs with therapeutic potential. Emerging evidence suggests that the therapeutic efficacy of tissue-specific EVs may be highly dependent on the age of the donor. For example, kidney tissue-derived EVs from newborn rat pups after acute injury showed a more pronounced reparative effect on the kidneys of mice than EVs derived from adult animals [89]. A correlation was found between the increasing age of the donor and a decrease in the neuroprotective effect of bone marrow-derived MSCs [90].
Body fluids such as blood, urine, cerebrospinal fluid and amniotic fluid are valuable sources of EVs that can be collected for therapeutic purposes [71]. The use of body fluids as a source of EVs provides a non-invasive and readily available way to obtain biologically active EVs for regenerative medicine. EVs from breast milk and amniotic fluid have shown potential for neonatal therapy [91,92,93]. However, it must be considered that despite the enriched presence of different types of EVs in biological fluids, these particles represent a mixed fraction of vesicles produced by different cell types and the therapeutic effects depend on the donor [94], making it difficult to standardize drugs for clinical use. Also, EVs isolated from these fluids can reflect the homeostasis of the corresponding tissues and organs and potentially serve as markers for various pathological conditions [95,96,97,98].
Studies on EVs provide compelling evidence that vesicles serve as a fundamental and ancient mechanism of intercellular communication. They can facilitate the horizontal transfer of genetic information not only between different species of the animal kingdom, but also between different biological kingdoms, including bacteria, fungi and plants [99,100,101]. In addition, EVs have the unique ability to cross taxonomic boundaries, providing a link between organisms from different kingdoms, such as bacteria and animals or plants and animals. This suggests that EVs represent a universal mechanism of intercellular communication for all life forms and possibly an evolutionary tool [102]. EVs-mediated communication between kingdoms highlights the complex connections within biological systems and opens up possibilities for the development of new therapeutic strategies utilising their unique biological properties. EVs derived from bacteria, fungi or plants have shown an effect on physiological processes in human cells and tissues and can also be considered as potential nanocontainers for drug delivery [103].
Just like EVs from animals, its from plants, fungi and bacteria contain a wide range of biologically active compounds, anti-inflammatory molecules and nucleic acids that can have a functional effect to ensure regeneration and immunomodulation in the human body [104,105,106], and also have low immunogenicity and high biocompatibility [107,108]. Plant-derived EVs have been shown to improve intestinal function in necrotizing colitis in mice [109]. Fungal-derived vesicles are able to interact with mammalian dendritic cells, neutrophils and macrophages and reduce the inflammatory response characteristic of neonatal pathologies [110]. In turn, bacterial vesicles, known as bacterial outer membrane vesicles, play an important role in the transport of virulence factors (toxins, enzymes and lipopolysaccharides) during infection of the host organism having a place in neonatal sepsis [111].
The important factor is not only the source of EVs, but also the method of their isolation from biological samples, which determines the purity of the preparation and consequently the therapeutic efficacy and reproducibility of the effects. Each of the EV isolation methods has its own unique advantages and disadvantages. For example, differential ultracentrifugation provides high EV yields and is relatively inexpensive, but can lead to contamination with proteins and other particles [112]. In contrast, size exclusion chromatography provides highly purified EV populations, albeit at the cost of lower particle yield and longer processing time as well as possible sample dilution. Polymer-based precipitation is characterised by its efficiency and simplicity; commercially available kits further contribute to its accessibility. Nevertheless, such methods indiscriminately co-precipitate vesicles of different sizes and fail to distinguish non-vesicular entities, such as protein aggregates, from EVs [113]. Finally, immunoaffinity techniques offer unparalleled specificity through antibody-mediated selection. However, they are costly, yield lower levels of EVs and carry the risk of excluding certain vesicle subpopulations [114]. Consequently, the optimal isolation strategy depends on a variety of factors, e.g. yield, economic considerations and the duration of the process.
6. Experimental Studies of EVs as Therapeutic Tools for Neonatal Pathologies
6.1. Hypoxic ischemic encephalopathy
HIE in newborns is one of the most common causes of neonatal mortality and disability. Epidemiologic data suggest an incidence rate of HIE of 2 to 3 per 1000 live births, representing 6-9% of all neonatal deaths and 21-23% of neonatal deaths in full-term infants. Approximately 25% of survivors suffer from severe neurological sequelae, including cerebral palsy, seizures, mental retardation, cognitive impairment and epilepsy [115,116]. HIE is primarily developed due to reduced cerebral blood flow and insufficient oxygen supply to the brain. The pathophysiological effects of the disease progress over time, making timely and effective treatment difficult. The pathology of HIE occurs in two phases: primary and secondary energy failure. Primary energy failure is caused by an initial decrease in cerebral blood flow leading to decreased oxygen and glucose supply, as result decreased mitochondrial ATP production and increased lactate production. This disrupts the function of ion channels and intracellular calcium regulatory mechanisms, leading to excessive release of glutamate, which triggers further intracellular calcium and sodium influx and eventually leads to cell necrosis. Apoptotic cell death typically occurs during a few days after the injury. In the secondary phase of energy failure, which begins 6–48 hours after the initial injury, mechanisms such as oxidative stress, excitotoxicity and inflammation contribute to further damage [116]. Oxidative stress especially affects the neonatal brain due to the low concentration of antioxidants and high oxygen consumption during the transition to neonatal life [117]. Understanding the mechanisms of primary and secondary energy disturbances is crucial for effective therapeutic interventions [116]. Although therapeutic hypothermia has progressed as a neuroprotective therapy for neonates with HIE, it is associated with adverse neurodevelopmental outcomes in some infants, and its use in preterm infants is only appropriate from 35 weeks of gestation [118,119,120]. There is an urgent need to develop new therapeutic approaches to treat neonates with HIE, with cell technologies emerging as a promising direction in therapy. Such new therapeutic strategies will be crucial to improve treatment options, reduce morbidity and mortality, and improve the long-term prognosis for newborns with HIE [121].
Initial studies focused on evaluating the neuroprotective potential of stem/progenitor cells for morphologic and functional recovery in experimental models of HIE [122]. A number of studies have shown that systemic administration of bone marrow-derived MSCs to neonatal rat pups with HIE improved neurological functions, increased proliferation and differentiation of neural progenitor cells into neurons and oligodendrocytes, and reduced neuroinflammation and the extent of brain tissue damage [123,124,125]. Transplantation of MSC derived from umbilical cord blood or human umbilical cord/placental stroma reduced the severity of brain injury and increased the survival rate of animals when modeling acute pathological conditions [126,127,128]. Analysis of the neuroprotective mechanisms of MSCs indicated a greater involvement of paracrine factors, as the percentage of engraftment of MSCs and their further differentiation in the brain was extremely low [126,129]. These observations have formed a modern paradigm for the therapeutic effect of MSCs through the secretion of EVs, which possess almost all the therapeutic properties of the cells they secrete [130,131].
In studies on experimental models, several main therapeutic effects of MSC-EVs have been identified in studies on experimental models. Most researchers identify the immunomodulatory effects of EVs [69,132], while the main molecular mechanism of the anti-inflammatory action of MSC-EVs is considered to be the modulation of mitogen-activated protein kinases (MAPK) and nuclear factor kB (NF-kB) signaling pathways in the damaged brain. Human bone marrow-derived MSC-EVs, when administered intraperitoneally to newborn mice with induced HIE, have been shown to decrease levels of the proinflammatory cytokine TNFa and increase levels of the anti-inflammatory cytokine tumor growth factor β (TGF-β) in the brain [133]. MSC-EVs from human umbilical cord also modulate the signaling pathway activated by Toll-like receptor 4 (TLR-4) on the surface of microglia by preventing the degradation of NF-kB inhibitor and inhibiting the phosphorylation of MAPK kinases. This leads to reduced microglial activity and prevention of gliosis [74]. In particular, bone marrow-derived MSC-EVs selectively inhibit the P38MAPK/NF-kB-p65 signaling pathway in microglial cells, leading to suppression of transcription of inflammation-related genes such as IL-6 in brain tissue after stereotactic injection of EVs into the lateral ventricles [134]. In an in vitro study using MSC-EVs from human umbilical cord, it was found to increase the viability of microglial cells during glucose-oxygen deprivation modeling, reduce markers associated with pyroptosis, and inhibit the release of inflammatory factors. The protective effects of MSC-EVs were associated with increased mitophagy due to activation of Forkhead Family Transcription Factor 3a (FOXO3a) [135].
Other mechanisms of the neuroprotective effect of MSC-EVs in HIE are considered to be the restoration of the structural integrity of the blood-brain barrier (BBB), promoting mitophagy and increasing the expression of neurotrophic factors. Human MSCs-EVs have been shown to carry annexin A1 (ANXA1) on their surface, a molecule with anti–inflammatory effect and playing a role in maintaining the integrity of the BBB [136,137]. In a study in a mouse model of HIE, intraperitoneal injection of MSC-EVs increased the expression of neurotrophic factors such as brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF) and epidermal growth factor (EGF) in brain tissue, which was associated with an increase in the density of neurons and blood vessels and prevented secondary brain damage [133]. Similar results were obtained in the work of Turovsky E. and co-authors, in which the course intranasal administration of MSC-EVs from the postpartum placenta prevented the progression of brain damage caused by hypoxia-ischemia. Analysis of the MSC-EVs proteome and subsequent inhibitory analysis of neuroglia culture in a glucose-oxygen deprivation model showed that the neuroprotective effect of MSC-EVs is realized via the PI3K/Akt pathway [24].
Most research on the neuroprotective mechanisms of MSC-EVs has focused on miRNAs rather than their protein cargo. Several miRNAs have been identified that may play a role in the neuroprotective properties of MSC-EVs. For example, miRNA-146a-5p, found in human umbilical cord MSC-EVs, has been shown to inhibit the activity of pro-inflammatory microglia by modulating the IRAK1/TRAF signaling pathway [30]. Another miRNA, miRNA-181b, secreted from adipose tissue-derived MSC-EVs restored proliferation and migration of rat brain microvascular endothelial cells in vitro [138]. In addition, miRNA-126 from a similar source of MSC-EVs was associated with induction of neurogenesis and reduction of apoptosis in brain tissue of rats with induced HIE [139].
In the developing brain, EVs from resident cells play a special role. They facilitate intercellular communication between developing cell assemblies, support neuron development and neuronal network formation, synaptic plasticity, regulation of myelination of nerve fibers and activation of microglia [140]. Exosomes derived from M2 microglia were investigated for their potential neuroprotective effects after modeling ischemic brain injury. It was shown that neurons were able to take up these exosomes, leading to a reduction in apoptotic cell death, a decrease in infarct volume and a recovery of neurological functions. This neuroprotective effect was associated with exosomal miRNA-124 which targets the ubiquitin-specific protease 14 (USP14) gene [141]. EVs from neural progenitor cells provide neuroprotection through the transfer of miRNA-150-3p, which targets caspase-2 mRNA to control apoptotic cell death [142].
Thus, the studies conducted show that EVs from various sources have neuroprotective properties and exert a pleiotropic effect on the tissues of the developing brain, including influencing neuronal plasticity. It is worth noting that the potential therapeutic efficacy of EVs is not limited to the acute phase of brain injury, but rather largely prevents the development of remote neurodegenerative consequences.
6.2. Respiratory distress syndrome and bronchopulmonary dysplasia
RDS and BPD are common respiratory diseases in newborns that are characterized by a different pathogenesis but often occur in the same patient. RDS primarily affects premature infants due to insufficient production of surfactant, a substance that is essential for maintaining the expansion state of the lungs. The lack of surfactant leads to alveolar collapse and impaired oxygen exchange, which in turn leads to respiratory distress. In premature babies, the production of surfactant is not yet physiologically developed. The risk of developing RDS is proportional to the duration of pregnancy, so that in newborns with extremely low body weight born at 24 weeks, RDS occurs in 98% of cases [143]. On the other hand, BPD is a chronic lung disease that often develops in infants who require prolonged mechanical ventilation and supplemental oxygen to treat RDS. The pathogenesis of BPD includes lung injury caused by factors such as inflammation, oxidative stress and mechanical trauma during mechanical ventilation, including high-frequency ventilation. As a result, normal lung growth and development is disrupted [144]. The combination of RDS and BPD is a complex interplay of surfactant deficiency, lung tissue immaturity, inflammation and damage that contributes to respiratory distress and long-term lung complications in newborns. Each year, BPD develops in approximately 25% of infants weighing less than 1,500 g worldwide, and about 70% of them suffer from a moderate or severe form, with an unfavorable outcome in 19% of cases [145,146]. One of the main methods of treating BPD are various methods of respiratory support, which only prosthetically support the function of the developing lungs, but do not contribute to the regeneration and maturation of lung tissue in neonates and premature infants. Studies in animal models of BPD have shown that MSCs and their secretome contribute to the restoration of function and structure of the developing lungs [147,148].
One of the detrimental factors in the ventilation therapy of newborns and premature infants is the use of gas mixtures with a high oxygen content, which inevitably leads to the development of oxidative stress in the lung tissue [149]. BPD therapy with MSC-EVs has been shown to reduce the effects of oxidative stress and its markers, indicating the presence of antioxidant mechanisms [150,151]. Tissue homeostasis in the lung is also associated with the development of an intense inflammatory response leading to fibrosis, vascular remodeling and pulmonary hypertension [152,153]. The first study on the protective paracrine properties of MSCs in the development of acute neonatal pulmonary hypertension showed that fractionation of the conditioned culture medium on exosomes contributed to a reduction in lung infiltration by macrophages and inflammation, as well as a decrease in the development of pulmonary hypertension by modulating the STAT3 signaling pathway [154]. Willis G.R. et al. demonstrated a therapeutic effect of exosomes isolated from the conditioning medium of human MSCs from umbilical cord or bone marrow in a mouse model of BPD. Intravenous administration of these exosomes significantly attenuated the progression of hyperoxia-induced pulmonary inflammation. This intervention subsequently led to improvement in characteristic BPD manifestations, including fibrosis, vascular remodeling and pulmonary hypertension [148]. An important finding was the ability of MSC exosomes to modulate the phenotypic balance of macrophages in the lung by attenuating the pro-inflammatory M1 phenotype and enhancing the anti-inflammatory M2-like state [148]. Furthermore, MSC-EVs from the human umbilical cord have been shown to restore the population of T- regulatory lymphocytes (Treg cells) when modeling hyperoxia in newborn mice [155]. The therapeutic properties of exosomes from human umbilical cord MSCs in a hyperoxia mouse model were elucidated, with efficacy correlating with the presence of the protein TSG-6 in the exosomes. In particular, hyperoxia-induced dysfunctions in both the brain and the heart were ameliorated after systemic administration of exosomes [156]. VEGF contained in exosomes is a potential candidate involved in lung regeneration mechanisms in BPD modeling, as exosomes without VEGF had no therapeutic effect [157,158]. Activation of hypoxia-induced factor 1-alpha (HIF-1α) could also be one of the mechanisms for the realization of a therapeutic effect under the action of EVs [150].
Type II alveolar epithelial cells (AEC-II) play a crucial role in the development of BPD, as their apoptotic death leads to disruption of the structural integrity of the alveoli [159]. Both in vitro and in vivo studies have shown that MSC-EVs are able to prevent the apoptotic death of AEC-II [160,161]. When MSC-EVs were administered systemically to newborn mice with induced BPD, there was a reduction in apoptotic cell death in lung tissue, as evidenced by reduced mRNA levels of the pro-apoptotic factors caspase 1 (Casp1) and the apoptosis regulator Bax [73]. Hydrogen peroxide-induced apoptotic death of the alveolar epithelial cell line RLE-6TN was prevented by treatment with bone marrow-derived MSC-EVs via mechanisms associated with miRNA-425, which targets PTEN and enhancing the PI3K/AKT axis [162]. EVs derived from human amniotic epithelial cells (hAECs) can be used in regenerative medicine, particularly for the treatment of BPD. The influence of gestational age of hAEC donors on the therapeutic efficacy of EVs in an experimental model of BPD was investigated. EVs from full-term infants reduced lung injury, increased amount of AEC-II in the alveoli and reduced airway hyperresponsiveness, pulmonary hypertension and right ventricular hypertrophy, while hAEC-EVs from preterm infants showed no therapeutic effect [163]. Intraperitoneal administration of MSC-EVs effectively suppressed the development of pulmonary fibrosis in a model of neonatal rats with pulmonary hypertension. According to the authors, this occured due to the potential of EVs to inhibit the transdifferentiation of AEC-II into AEC-I by downregulating the expression of WNT5a mRNA, which is a critical factor in transdifferentiation and the initiation of fibrosis [164].
6.3. Neonatal sepsis
Neonatal sepsis is a systemic infection that occurs in infants up to 28 days of age and is caused by bacteria, viruses or fungi. Sources of infection may include pathogens from the mother's genital tract with which the newborn comes into contact during birth, such as Group B Streptococcus, Escherichia coli, Listeria monocytogenes, Haemophilus influenzae and, less commonly, coagulase-negative Staphylococcus strains and Streptococcus pneumoniae. On the other hand, the infection can also be acquired in healthcare settings, including nosocomial, antibiotic-resistant strains. The pathogenesis involves the penetration of germs into the bloodstream, leading to bacteremia, an increased inflammatory response and even septic shock. Newborns, especially preterm infants, are highly susceptible to infections due to their immature immune system, their limited ability to localize infections and their reduced number of pre-formed antibodies. Consequently, uncontrolled development of the immune response can lead to multiple organ failure and death of the newborn, especially in the case of infection with antibiotic-resistant microorganisms [165,166]. To date, antibiotic therapy has not been able to completely combat the development of infections in newborns. An analysis of the effectiveness of antibiotic therapy has shown that the selection of an appropriate antibiotic is difficult for about 35% of neonates and preterm infants, with therapeutic measures often proving ineffective or worsening the clinical manifestations [167]. Due to their immunoregulatory and systemic regenerative effects, MSCs are promising therapeutic agents in the treatment of sepsis and associated complications in combination with basic therapy [168]. However, we could not find any relevant studies on the use of MSC-EVs for sepsis therapy in the field of neonatology. EVs have been identified as key players in the pathophysiology of sepsis, but they also show anti-inflammatory, anti-apoptotic and immunomodulatory properties in sepsis models [169]. Sepsis can lead to bacterial meningitis if the bacteria cross the BBB, which is normally impermeable to pathogens, a life-threatening situation in newborns and preterm infants. [170]. Kim Y et al demonstrated that administration of MSC significantly reduced bacterial growth and brain damage in neonatal rats with E. coli-induced meningitis. Although bacterial growth in cerebrospinal fluid could not be significantly reduced, MSC-EVs dramatically reduced meningitis-induced brain cell death, reactive gliosis and inflammation [171].
6.4. Necrotizing enterocolitis
NEC is a severe disease in the neonatal period characterized by inflammation and subsequent necrosis of the intestinal wall. It primarily affects premature infants and is caused by a variety of factors, including prematurity, artificial feeding, intestinal ischemia, and aberrant bacterial colonization [172] The pathogenesis involves a disruption of tissue homeostasis in the immature intestinal tissue between immune protection and the intestinal microbiota, resulting in a strong inflammatory response that damages the intestinal tissue. This damage can progress to necrosis and perforation of the bowel, requiring surgery to remove the affected section of bowel and possibly the development of "short bowel syndrome" [173]. Primary intensive therapy includes discontinuation of enteral nutrition ("bowel rest"), conversion of the child to complete parenteral nutrition and initiation of antibiotic therapy. In cases where necrosis and perforation of the intestinal wall develop, surgical intervention may be required [174].
Effective treatment of NEC requires a comprehensive interdisciplinary approach along with personalized care. In addition, there is an ongoing need to research and develop new therapeutic strategies that directly target the primary pathogenic mechanisms of the disease. Various treatment strategies have been proposed for NEC, but recently MSC therapy has become the preferred option due to its pronounced therapeutic efficacy. There is evidence that therapeutic effects can be achieved not only by transplantation of stem/progenitor cells, but also by conditioned culture medium containing cell secretomes and EVs [175,176,177]. In an experimental model of NEC, it was shown that bone marrow-derived MSCs and their exosomes contributed to a significant reduction in the development of the pathological process and maintained the integrity of the intestinal mucosa and its physiological permeability. Notably, the therapeutic effects were exclusively attributed to exosomes in a model simulating mechanical injury of intestinal epithelial cells line (IEC-6), as opposed to an exosome-free conditioned culture medium [178]. McCulloh C.J. and colleagues compared the therapeutic efficacy of EVs derived from four different types of rat stem cells in an NEC model, namely MSCs from amniotic fluid, bone marrow MSCs, neural stem cells differentiated from amniotic fluid stem cells, and neonatal intestinal epithelial cells. Their findings indicated that EVs secreted by neural stem cells were the most effective source for NEC treatment at the lowest studied dose (8 × 107 EVs). Increasing the dose to (4 × 108 EVs) demonstrated equivalent effectiveness for all four sources of EVs, resulting in a reduction in the incidence of NEC and the severity of necrotic tissue damage [88].
It is postulated that the primary pathogenetic factor in the development of NEC is an inflammatory response triggered by the immaturity of the intestinal tissue [179]. In this context, the immunoregulatory functions of MSC-EVs have been studied in detail in different models of acute intestinal injury. Systemic administration of human bone marrow-derived MSC-EVs significantly improved colitis symptoms by attenuating inflammation, maintaining intestinal barrier integrity, and inducing polarization of macrophages to the M2 phenotype, while preventing intestinal fibrosis. The therapeutic effects of MSC-EVs were mediated by their interaction with colonic macrophages and stimulation of IL-10 production by macrophages. Of note, systemic removal of resident intestinal macrophages with the Clod-Lipo agent resulted in loss of the therapeutic effect of MSC-EVs in experimental animals. Research suggests that MSC-EVs have the potential to modulate the balance of T- cell populations towards an anti-inflammatory state. For example, intraperitoneal administration of adipose-derived MSC-EVs in mice resulted in a restoration of Treg cell populations in the spleen and mesenteric lymph nodes, which correlated with a reduction in intestinal tissue damage in an experimental colitis model [180]. Furthermore, intraperitoneal administration of MSC-EVs from human umbilical cord increased the activated Th2 population and reduced the inflammatory Th1 and Th17 populations in mesenteric lymph nodes [181] and restored the balance between Tregs and Th17 in the blood of experimental animals with colitis [182]. A direct correlation was found between the cargo of MSC-EVs and their therapeutic properties. For example, microRNA-378a-5p carried by MSC-EVs was found to decrease the levels of NLRP3 inflammasome complex proteins in macrophages, highlighting a possible mechanism for their anti-inflammatory effects [183]. In addition, the presence of TGF-β1 in the cargo of EVs has been shown to decrease the activity and quantity of neutrophils, dendritic cells, CD4 and CD8 T lymphocytes and induce a regulatory phenotype in T cells and an immunosuppressive phenotype in dendritic cells in intestinal tissues [184]. MSC-EVs derived from perinatal and adipose tissue protect epithelial cells from apoptosis induced by oral administration of dextran sodium sulfate (DDS). These protective effects include the reduction of ROS levels in intestinal tissue, inhibition of myeloperoxidase activity in infiltrating neutrophils and suppression of mRNA synthesis of proapoptotic factors such as caspases-3, -8 and -9 [181,185]. MSC-EVs can stimulate the proliferation of intestinal epithelial cells [181,185,186]. Maintaining the integrity of the intestinal epithelial layer and preserving the tight junctions between epithelial cells are essential for the formation of the protective intestinal barrier. MSC-EVs from various sources, mainly from milk, contribute to the maintenance of this barrier by preventing the death of various types of intestinal epithelial cells, including enterocytes [185,187], intestinal stem cells [188] and goblet cells [181]. In addition, MSC-EVs can also maintain the structural integrity of the tight junctions between intestinal epithelial cells [181,186,189].
In the neonatal period, the integrity of the epithelial barrier, the immune system and the formation of the commensal microbiota in the infant's gastrointestinal tract jointly determine the homeostasis of the intestinal mucosa. Breast milk plays a fundamental role in the regulation of the above processes, as it has been found to act in many ways through the action of breast milk-derived extracellular vesicles (MEVs) [190,191,192]. MEVs are secreted during lactation by the epithelial cells of the mammary gland [193] and possibly also by the MSCs resident in the mammary gland and constitute a significant proportion of the total fraction of milk EVs [194,195]. MEVs play an important role in the development of the neonatal immune system [196] and in the maturation of cells of the gastrointestinal tract [197]. Several studies have investigated the intravesicular content of breast MEVs and discovered miRNAs, including let-7a, let-7b, let-7f and miRNA-148a [198], which inhibit the expression of genes that activate the NF-κB signaling pathway in dendritic cells [198,199].
Proteomic analysis of MEVs revealed the presence of proteins involved in the development of intestinal and brain tissue. Notably, human MEVs from preterm pregnancies contain elevated levels of lactadherin, which facilitates the proliferation and migration of neural stem cells and aids in the repair of damaged intestinal mucosa [200,201,202]. Additionally high levels of peptides derived from β- and α-S1-casein were detected in the MEVs from preterm birth [200]. β-casein peptides stimulate the proliferation of CD19+ B-cells, and α-S1-casein derivatives provide protection against ROS and suppress inflammation in the intestine [203,204]. In addition, lactoferrin, found in both types of MEVs (from full-term and preterm pregnancies), is involved in regulating the activity of the immune system and providing antimicrobial protection [205]. Recent studies have also demonstrated the positive role of lactoferrin in regulating the proliferation of various types of intestinal cells [206].
Recent studies have shown that MEVs from different animal species can have a comparable therapeutic effect against neonatal intestinal diseases [207]. Using a DSS-induced colitis model, the efficacy of MEVs from human and bovine milk in the treatment of colitis was demonstrated. Oral administration of these MEVs resulted in less damage to the epithelial barrier, inhibition of fibrosis and activation of infiltrated immune cells. Furthermore, a decrease in the mRNA expression of the pro-inflammatory cytokines IL-1β, IL-6, IL-17A and IL-33 and the restoration of normal levels of commensal microorganisms were observed [208,209,210]. The therapeutic effects of EVs from cow's milk can be attributed to the presence of a variety of factors, including conserved microRNAs, such as those of the let-7 family, which are recognized as important regulators of the immune response in mammals [198,211]. In addition, EVs from cow's milk was found to increase mucin production and expression of the goblet cell markers TFF3 and MUC2 in human colon LS174T cells [212]. The above studies underline the therapeutic and preventive potential of the use of extracellular vesicles from breast milk, especially for premature infants and highlight the need to integrate these vesicles into infant formula.
Maintaining a normal balance of the gut microbiome in newborns is critical to the health of the gastrointestinal tract, and disruption of this homeostasis can lead to the development of NEC [213]. Recent studies revealed the ability of the intestinal microbiome to autonomously regulate its microbial composition by means of interactions between intestinal bacteria and cells within the human gastrointestinal tract mediated through outer membrane vesicles of bacteria (OMVs), as evidenced in cases of NEC [214,215]. For example, oral administration of OMVs produced by Akkermansia muciniphila resulted in reduced neutrophil infiltration and restoration of intestinal barrier integrity in mice with colitis [216,217]. In a similar model, oral administration of OMVs from Lactiplantibacillus plantarum resulted in reduced serum levels of the proinflammatory cytokines IL-6, IL-1β and TNF-α and restored the balance between proinflammatory proteobacteria and anti-inflammatory bacteria such as Escherichia and Muribaculaceae [218]. In a mouse model of DSS-induced colitis, oral administration of bacterial OMVs from Clostridium butyricum led to an alleviation of bacterial dysbiosis in colitis mice and to a significant reduction in the abundance of the bacterial pathogens Escherichia coli and Shigella flexneri. Coculturing these OMVs with macrophages activated by LPS resulted in the transfer of microRNA-199a-3p to them, inhibiting pro-inflammatory signals via the MAPK and NF-κB signaling pathways [219].
7. Discussion and challenges of Translating EVs into Clinical Practice
The results from experimental models of neonatal and premature infant diseases discussed in this study provide promising data on the therapeutic efficacy of EVs and form a solid basis for the further development of therapeutic research and its translation into clinical practice. However, it must be recognized that several tasks need to be addressed before EVs can be widely used clinically. First, the selection of an appropriate source to obtain EVs with high therapeutic efficacy is a key aspect influencing the efficacy of the therapy. While previous research has primarily focused on MSCs as a primary tool in regenerative medicine and as a source of EVs, [220] more recent studies have broadened the spectrum of potential EV sources. Encouraging prospects are offered by milk [208] and non-animal-derived EVs [101,102].
One of the biggest challenges is the incomplete understanding of the true nature of EVs. While the existence and potential therapeutic applications of EVs are recognized, the specific identity and composition of the therapeutic components in these EVs remain partially obscure. The role of specific non-coding RNAs (such as piRNA, ribosomal RNA, small nuclear and nucleolar RNAs, circular RNAs) is not fully elucidated, and the functions of double-stranded DNA in EVs remain to be determined [220,221,222]. This knowledge gap hinders the full exploitation of the therapeutic potential of EVs and the optimization of their clinical efficacy. Further exploration of the complex composition of EVs is critical to effectively address these challenges. The use of advanced analytical methods, such as proteomic and genomic profiling will facilitate the elucidation of the specific "cargo" contained in EVs. Revealing the identity and abundance of these therapeutically active components will provide a deeper understanding of how EVs exerts its therapeutic effects at the molecular level.
Standardization of protocols for isolation, purification and characterization of EVs is essential to ensure reproducibility and comparability of results across studies. This standardization will facilitate the development of reliable guidelines for the clinical use of EVs. The problem of heterogeneity of EVs raises concerns about their efficacy for clinical use. To solve this problem it is crucial to develop and implement a standardized approach for the isolation of EVs [223]. In addition, it is important to establish standards for the evaluation of the qualitative and quantitative characteristics of EVs and their content [224]. The presence of therapeutically useful cargo in extracellular vesicles implies a question of their dosage, which directly affects not only the efficacy, but also the safety of the use of EVs. Various methods are used to quantify EVs, including assessment of the number of parent cells (cell equivalents), total protein concentration in EVs and specific analytical methods (detection of particle concentration and size) [225]. However, the existing in vivo studies, including in neonatology, have been performed without a standardized dosage, frequency and method of administration of EVs, which makes it difficult to determine a safe and effective dosage for clinical trials for the therapy of neonatal pathologies. Despite extensive discussions on the determination of the optimal therapeutic dose, no uniform concept has yet been developed, partly because there is not enough experimental data. It is noteworthy that during the discussion of a group of researchers, the term "therapeutic unit" was proposed as a standard for calculating the dose, which we discussed in detail in the "Methods" section. We used this approach to standardize the therapeutic doses used in the reviewed papers, which are listed in Table 1. This approach allows the therapeutic doses to be compared with each other. As seen, the doses in the different studies can vary by up to 100-fold.
It is also important to investigate all aspects of the potential side effects of EVs. Research has shown that exosomes can carry tissue factor and phosphatidylserine on their surface, which activate blood coagulation processes [226,227]. Tissue factor is considered a key regulator of hemostasis, which under certain conditions can trigger local thrombosis, leading to an inflammatory immune response [228]. The acute, rare thrombotic reactions observed in patients after MSC administration in clinical trials are attributed to the side effects associated with vascular thrombosis and the subsequent inflammatory response [229]. Consequently, there is a pressing need for researchers and clinicians to develop standardized and clinically applicable protocols that are focused on the delivery methods and preparation of EVs-based therapeutics and are supported by solid preclinical evidence. [230] In addition, the outcomes of combinatorial therapies require more thorough analysis. For example, an in vivo study using HIE model in neonatal mice has shown that pretreatment with hypothermia can reduce the efficacy of subsequent cell therapy [231].
Despite all these unresolved issues, according to the clinicaltrials.gov database, three clinical trials with EVs in neonatology have been registered and are in active status. Study NCT03857841 is dedicated to investigating the therapeutic efficacy and safety of allogeneic bone marrow-derived MSCs-EVs in neonates aged 3-14 days with a diagnosis of BPD. In a study from Russia, the use of MSC-EVs for the treatment of children with extremely low body weight using allogeneic MSC-EVs in order to prevent the development of brain damage associated with prematurity (NCT05490173). The third clinical trial will evaluate the efficacy of allogeneic MSC-EVs in neonates with HIE (NCT02854579). The introduction of a broad spectrum of EVs-based treatment methods into clinical practice can therefore potentially not only reduce the mortality rate in premature newborns, but also significantly reduce disabilities.

Table 1.
Overview of studies investigating the therapeutic potential of EVs in neonatal diseases.
| References | Sources of EVs | Model | Protocol of administration | Dosing | Therapeutic effects | Elucidated mechanisms |
|---|---|---|---|---|---|---|
| HYPOXIC-ISCHEMIC ENCEPHALOPATHY | ||||||
| [232] 2016 |
human bone marrow MSCs |
the Rice-Vannucci model | total dose – 2,4 x 1010 EVs route of administration – intravenous administrations per day – 2 days – 1 |
1 unit* |
↓ total number and duration of seizures ↓ pathological fluctuations of blood pressure |
↑ baroreflex-mediated heart rate response |
| [137] 2019 |
human bone marrow MSCs |
the Rice-Vannucci model | total dose – 2,4 x 1010 EVs route of administration – intravenous administrations per day – 2 days – 1 |
1 unit.. |
↓permeability of Blood-brain barrier |
↑Annexin А1/FPR in neonatal brain endothelial cells and microglia |
| [132] 2019 |
human bone marrow MSCs |
the Rice-Vannucci model | total dose – 1,25 × 109 Evs route of administration – intranasal administrations per day – 1 days – 1 |
0,1 Unit |
↓ of tissue loss ↓ % of cell death ↓ microglial activation ↑ behavioral outcomes (negative geotaxis test) |
|
| [74] 2019 |
human Wharton’s jelly MSCs |
the Rice-Vannucci model + intraperitoneal injection of LPS | total dose – 325 µg of EVs protein per animal route of administration – intranasal administrations per day – 1 days – 1 |
0,3 Unit |
↓ microgliosis ↓ neuroinflammation |
↓LPS/TLR4 signaling in microglia |
| [233] 2020 |
rat bone marrow MSCs (H2S preconditioning) |
the Rice-Vannucci model | total dose – 1,5 × 108 EVs route of administration – intracardial injection administrations per day – 1 days – 1 |
0,06 Unit |
↓ water content and infarct volume of the brain ↓ % of cell apoptosis ↑polarization toward the anti-inflammatory M2 phenotype ↑ memory function |
↑miR-7b-5p ↓FOS → ↓Iba1+ in microglia |
| [133] 2020 |
human bone marrow MSCs |
the Rice-Vannucci model | total dose – 2,7 × 108 EVs route of administration – intraperitoneal administrations per day – 1 days – 3 (1,3,5 post HI) |
0,03 Unit |
↓ striatal tissue loss ↓ M1 micro- and A1 astroglia activation ↑ neurogenensis and angiogenesis ↑ myelination |
|
| [234] 2021 |
mice bone marrow MSCs |
the Rice-Vannucci model | total dose – 100 µg of EVs protein route of administration – intracardial injection administrations per day – 1 days – 1 |
0,1 Unit |
↓ HI-induced edema, infarction, infiltrating monocytes ↓phagocytosis of viable neurons ↑synaptic densities |
↓ p-NF-κB → ↓OPN → ↓Iba1 in М1 microglia |
| [235] 2021 |
mice bone marrow MSCs |
the Rice-Vannucci model | total dose – 5 µg of EVs protein route of administration – intranasal administrations per day – 1 days – 1 |
0,005 Unit |
↓ injury volumes ↓ microglial activation ↓ neuroinflammation |
↓Iba1 → ↓ Casp3 in microglia |
| [236] 2021 |
rat primary astrocytes (P1) |
the Rice-Vannucci model | total dose – 2.5 µg of EVs protein route of administration – intraperitoneal administrations per day – 1 days – 1 |
0,0024 unit | ↓ the area of cerebral infarction ↓ HIBD-induced neuronal apoptosis ↓ oxidative stress ↓ neuroinflammation ↑ body weight ↑cognitive functions( grip test, negative geotaxis test |
↑ miR-17-5p →↓BNIP →↓Bax in brain tissue |
| [237] 2022 |
brain tissues of neonatal mice (P9) after HI |
the Rice-Vannucci model | total dose – 8 × 109 EVs route of administration – intranasal administrations per day – 2 days – 1 |
0,066 Unit |
↓ infarct size ↓ Casp3 expression |
↑ miR-342-3p and miR-330-3p in brain tissue |
| [238] 2022 |
mice bone marrow MSCs |
the Rice-Vannucci model | total dose – 2 × 109 EVs route of administration – intranasal administrations per day – 1 days – 1 |
0,2 Unit |
↑ animal survival ↓ infarct volume of brain ↓ % of apoptosis cells ↓ neuroinflammation ↑proprioceptive function |
↑miR-93 →↓JMJD3→↑KLF2→↓Casp3,Bax in neurons |
| [239] 2023 |
immortalized human bone marrow MSCs | the Rice-Vannucci model | total dose – 2,7 × 108 EVs route of administration – intranasal administrations per day – 1 days -3 (1,3,5 post HI) |
0,03 unit. |
↑neurogenesis and angiogenesis ↓ monocyte infiltration ↓ astrogliosis and microgliosis |
|
| BRONCHOPULMONARY DYSPLASIA | ||||||
| [148] 2018 |
human Wharton’s jelly MSCs |
hyperoxia (HYRX)-induced BPD mice model (P1 –P7 75% О2) |
total dose –0,9 µg of EVs protein route of administration – intravenous administrations per day – 1 days – 1 |
0,001 unit |
↑ lung architecture ↓ lung fibrosis ↓ peripheral pulmonary arterial remodeling |
|
| [156] 2018 |
preterm human Wharton’s jelly MSCs | HYRX-induced BPD mice model (с P1-P4 95% О2) | total dose – 4.5 × 108EVs route of administration – intraperitoneal administrations per day – 1 days -2 (P2,P4) |
0,038 Unit |
↑ lung architecture ↓ infiltration of neutrophils ↓ pulmonary hypertension ↓ alveolar-capillary leak |
↑TSG-6 sinalling in lung tissue |
| [240] 2018 |
human Wharton’s jelly MSCs | HYRX-induced BPD rat model (P1- P14 60% О2) |
total dose – 0,213 × 1010 EVs route of administration – intratracheal administrations per day – 1 days – 3 (P3,P7,P10) |
0,27 unit. |
↑ alveolar development ↓ pulmonary vascular remodeling |
|
| [157] 2018 |
rat bone marrow MSCs | HYRX-induced BPD rat model (P0- P14 85% О2) | total dose – 3,4 × 109 EVs route of administration – intraperitoneal administrations per day – 1 days – 14 (P1-P15) |
1,96 Unit |
↑ alveolar growth ↑ lung blood vessel density ↓ pulmonary hypertension |
↑VEGF signaling in lung tissue |
| [158] 2018 |
human umbilical cord blood MSCs |
HYRX-induced BPD rat model (P1- P14 90% О2) | total dose – 20 µg of EVs protein route of administration – intratracheal administrations per day – 1 Days – 1 (P5) |
0,019 Unit |
↑ alveolarization and angiogenesis | ↑VEGF signaling in lung tissue |
| [241] 2020 |
human Wharton’s jelly MSCs | HYRX-induced BPD mice model (P0-P14 75% О2) | total dose – 6 × 108 EVs route of administration – intravenous administrations per day – 1 days – 1 (PN4) |
0,025 unit. |
↓ alveolar simplification ↓septal collagen disposition ↑blood vessel count ↓ pulmonary hypertension ↑ functional exercise capacity |
|
| [242] 2021 |
human bone marrow MSCs. | In utero induced BPD rat model (antenatal injection of E. coli endotoxin e20) | total dose – 0,25 × 106 EVs route of administration – intra-amniotic administrations per day – 10 per pregnant rat days – 1 (e20) |
0,17 Unit |
↓ lung simplification ↑ vascularization ↓ pulmonary hypertension ↑ lung mechanical function |
|
| [155] 2021 |
human Wharton’s jelly MSCs | HYRX-induced BPD mice model (P – P7 75% О2) | total dose – 6 × 108 EVs route of administration – intravenous administrations per day – 1 days – 1 (PN4) |
0,025 unit. |
↑ thymic development ↑proportion of CD4+FoxP3+ regulatory T cells ↓ alveolar simplification |
|
| [240] 2021 |
human umbilical cord blood MSCs | HYRX-induced BPD rat model (P1 – P14 60% О2) | total dose – 0.64 × 1010 EVs route of administration – intratracheal administrations per day – 1 days – 4 (P3,P7,P10,P21) |
0,27 Unit |
↑ alveolar development ↓ deposition of fibrous tissue ↑ density of M2 macrophages ↓ pulmonary hypertension |
|
| [92] 2021 |
amniotic fluid-derived EVs (full-term cesarean sections) | HYRX-induced BPD rat model (P1 – P14 85% О2) | total dose – 1 × 1010 EVs route of administration – intratracheal administrations per day – 1 days – 1 (P3) |
0,42 unit |
↑ alveolar development ↓ pulmonary hypertension |
|
| [243] 2022 |
human breast milk-derived EVs | HYRX-induced BPD rat model (P1 – P7 85% О2) | total dose -140 µg of EVs protein route of administration – intragastric administrations per day – 1 days – 1 (PN7) |
0,133 unit |
↓ lung tissue collapse ↓ cleaved-caspase 3 |
↓IL-17 /↓ FADD in Type II alveolar epitheliocytes |
| [152] 2022 |
human Wharton’s jelly MSCs | HYRX-induced BPD rat model (P1 – P14 85% О2) | total dose – 96 × 108 EVs route of administration – intratracheal administrations per day – 1 days – 1 (PN3) |
0,04 Unit |
↑ lung vascular density and alveolar structure ↓lung inflammation ↓ pulmonary hypertension |
↑VEGF/eNOS in lung tissue |
| [163] 2022 |
human amniotic epithelial cells (term birth after caesarean sections) |
in utero induced BPD mice model (injection of LPS e16)+ (P3.5 – P28 65% О2) |
total dose – 10 µg of EVs protein route of administration – intravenous administrations per day – 1 days – 1 (PN4) |
0,01 Unit |
↑lung tissue-to-air space ratio ↓ lung inflammation ↑ type II alveolar epithelial cell ↓ pulmonary hypertension ↑lung tissue elasticity |
|
| [160] 2022 |
human Wharton’s jelly MSCs | HYRX-induced BPD mice model (injection of LPS P7/P8) + 40% О2 P10 | total dose – 1 × 106 EVs route of administration – intratracheal administrations per day – 1 days – 1 (PN9) |
0,0002 Unit |
↑ lung architecture ↑blood vessel density ↑ mRNA of antiinflammatory cytokines in lung tissue |
|
| [244] 2023 |
human umbilical cord blood MSCs | HYRX-induced BPD mice model (P1 – P14 85% О2) | total dose – 15 x 105 EVs route of administration – intraperitoneal administrations per day – 1 days – 3 (P4-P6) |
0,000063 unit | ↓ lung fibrosis ↑ vascular development |
↑miR-185-5p→↓CDK6 → ↑angiogenesis in lung tissue |
| [150] 2023 |
human bone marrow MSCs | hypoxia –induced BPD rat model(10 min 40%О2 + 2 min 1%О2 12 times daily P1 – P14) | total dose – 2 x 105 EVs route of administration – intraperitoneal administrations per day – 1 days – 14 (P1-P14) |
0,00012 unit | ↓ simplified alveolar structure ↓ pulmonary hypertension ↑ capillary distribution ↑ respiratory efficiency ↓ oxidative stress |
↑ PI3K/AKT →↑ SOD in lung tisue |
| NECROTIZING ENTEROCOLITIS | ||||||
| [178] 2016 |
mice bone marrow MSCs | NEC –induced preterm mice model Barlow et al. [245](21 e )+ 90 sec 100% N2 + 4°C 10 min twice daily ( P1-P4) | total dose – 2,5 x 109 EVs route of administration – intraperitoneal administrations per day – 1 days -1 (prior NEC) |
0,1 unit. |
↓ the overall incidence of NEC ↑ gut barrier function |
|
| [88] 2018 |
neonatal mice enteric neuronal stem cells | NEC –induced preterm rat model Barlow et al. (21e) + (90 sec 100% N2 + 4°C 10 min every 8 h + LPS every 4 h (P1-P4) | total dose – 4 × 108 EVs route of administration – intraperitoneal administrations per day – 1 days -1 (prior NEC) |
0,017 Unit |
↓ intestine villus destruction ↓ the overall incidence of NEC |
|
| [212] 2019 |
bovine milk-derived EVs | NEC –induced preterm mice model Barlow et al. (10 min 5% O2 3 times between P5-P9 + LPS 4 times between P6-P7) | total dose – 1.2 mg of EVs protein route of administration – intragastric via gavage administrations per day – 3 days – 5 (P5-P9) |
1,14 Unit |
↑ intestine villus destruction ↑ number of goblet cells ↓ intestinal mucosal inflammation ↓ oxidative stress |
|
| [208] 2019 |
human breast milk-derived EVs | NEC –induced preterm rat model Barlow et al. (21e ) 90 sec 1,5% О2 + 4°C 10 min 3 times daily P1-P4 + LPS 1 time P1 | total dose – 2.4 × 1010 EVs route of administration – intragastric via gavage administrations per day – 6 days – 4 (P1-P4) |
1 Unit |
↓ villus destruction ↓ the overall incidence of NEC |
|
| [175] 2020 |
rat amniotic fluid CD117 stem cells (e14.5) |
NEC –induced preterm mice model Barlow et al. (10 min 5% O2 3 times between P5-P9 + LPS 4 times between P6-P7) | total dose – 3.5 х 108 EVs route of administration – intraperitoneal administrations per day – 1 days – 2 (P6-P7) |
0,015 unit. |
↑ gut epithelial regeneration ↓ intestinal inflammation |
↑Wnt/ β-catenin →increased intestinal epithelial proliferation |
| [246] 2022 |
human breast milk-derived EVs | NEC –induced preterm mice model Barlow et al. (1 min 100% N2 + 4°C 5 min twice a day P6-P10) | total dose – 30 µg of EVs protein route of administration – intragastric via gavage administrations per day – 3 days – 3(P8-P10) |
0,04 unit |
↑ ileal crypts number ↓ inflammatory microenvironment |
|
| NEONATAL MENINGITIS | ||||||
| [171] 2022 |
human Wharton’s jelly-derived MSCs after thrombin preconditioning | Escherichia coli –induced meningitis in newborn Rats (P11) | total dose – 2,6 x 107 EVs route of administration – intraventricular administrations per day – 1 days – 1 (P11) |
0.001 unit |
↓ neural cell death ↓ number of active microglia ↓ levels of inflammatory cytokines in brain tissues |
|
| * The yield of EVs derived from supernatants of 4 × 107 MSCs that have been cultured for 48 h was defined as 1 unit. 1 unit contains 2,4 1010 EVs or 1.05 mg of EVs protein [14] | ||||||
Author Contributions
The conceptualization and methology was planned by D.S.; writing—original draft preparation was performed by D.S., K.G., M.I., Y.S. and A.K. Formal analysis was provided by M.I., K.G., Y.S.; A.B., Y.P., V.Z., and D.S. took part in writing—review and editing; Y.S., K.G., M.I., A.K. and D.S. performed visualization, supervision, project administration as well as funding acquisition were performed by D.S., D.D. and G.S. All authors contributed substantially to this project. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the research project “Microvesicles of multipotent mesenchymal stromal cells in the treatment of lesions of the central nervous system in newborns with congenital malformations of internal organs” (No. 121032500125-6).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vogel, J.P.; Chawanpaiboon, S.; Moller, A.-B.; Watananirun, K.; Bonet, M.; Lumbiganon, P. The Global Epidemiology of Preterm Birth. Best Pract Res Clin Obstet Gynaecol 2018, 52, 3–12. [Google Scholar] [CrossRef]
- Belousova, V.; Svitich, O.; Timokhina, E.; Ignatko, I.; Bogomazova, I.; Pesegova, S.; Silaeva, T.; Kuzmina, T.; Skorobogatova, O. Caspase-3, Caspase-8 and XIAP Gene Expression in the Placenta: Exploring the Causes of Spontaneous Preterm Labour. International Journal of Molecular Sciences 2023, 24, 1692. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Yu, D.; Liang, Y.; He, H.; He, W.; Li, Y.; Zhang, M.; Gao, Y.; Wu, F.; Chen, Q. Global Trends in Incidence and Death of Neonatal Disorders and Its Specific Causes in 204 Countries/Territories during 1990-2019. BMC Public Health 2022, 22, 360. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Thebaud, B.; Paton, M.C.B.; Fleiss, B.; Papagianis, P.; Baker, E.; Bennet, L.; Yawno, T.; Elwood, N.; Campbell, B.; et al. Advances in Neonatal Cell Therapies: Proceedings of the First Neonatal Cell Therapies Symposium (2022). Pediatr Res 2023. [Google Scholar] [CrossRef] [PubMed]
- Schiller, E.A.; Cohen, K.; Lin, X.; El-Khawam, R.; Hanna, N. Extracellular Vesicle-microRNAs as Diagnostic Biomarkers in Preterm Neonates. Int J Mol Sci 2023, 24, 2622. [Google Scholar] [CrossRef]
- Nitkin, C.R.; Rajasingh, J.; Pisano, C.; Besner, G.E.; Thébaud, B.; Sampath, V. Stem Cell Therapy for Preventing Neonatal Diseases in the 21st Century: Current Understanding and Challenges. Pediatr Res 2020, 87, 265–276. [Google Scholar] [CrossRef]
- Leopoldino, R.D.; Santos, M.T.; Costa, T.X.; Martins, R.R.; Oliveira, A.G. Drug Related Problems in the Neonatal Intensive Care Unit: Incidence, Characterization and Clinical Relevance. BMC Pediatr 2019, 19, 134. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells Are Exponentially Increasing: Where Are We in Recent Years? Stem Cell Rev Rep 2022, 18, 23–36. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Park, W.S.; Sung, S.I.; Chang, Y.S. Mesenchymal Stem Cell Therapy for Intractable Neonatal Disorders. Pediatr Neonatol 2021, 62 Suppl 1, S16–S21. [Google Scholar] [CrossRef]
- Maldonado, V.V.; Patel, N.H.; Smith, E.E.; Barnes, C.L.; Gustafson, M.P.; Rao, R.R.; Samsonraj, R.M. Clinical Utility of Mesenchymal Stem/Stromal Cells in Regenerative Medicine and Cellular Therapy. J Biol Eng 2023, 17, 44. [Google Scholar] [CrossRef]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat Rev Drug Discov 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular Vesicles as Drug Delivery Systems: Why and How? Adv Drug Deliv Rev 2020, 159, 332–343. [Google Scholar] [CrossRef] [PubMed]
- C, T.; Kw, W.; E, A.; Mj, A.; Jd, A.; R, A.; A, A.; T, A.; F, A.; Gk, A.-S.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. Journal of extracellular vesicles 2018, 7. [Google Scholar] [CrossRef]
- Kordelas, L.; Rebmann, V.; Ludwig, A.-K.; Radtke, S.; Ruesing, J.; Doeppner, T.R.; Epple, M.; Horn, P.A.; Beelen, D.W.; Giebel, B. MSC-Derived Exosomes: A Novel Tool to Treat Therapy-Refractory Graft-versus-Host Disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef] [PubMed]
- Laulagnier, K.; Motta, C.; Hamdi, S.; Roy, S.; Fauvelle, F.; Pageaux, J.-F.; Kobayashi, T.; Salles, J.-P.; Perret, B.; Bonnerot, C.; et al. Mast Cell- and Dendritic Cell-Derived Exosomes Display a Specific Lipid Composition and an Unusual Membrane Organization. Biochem J 2004, 380, 161–171. [Google Scholar] [CrossRef]
- Biró, É.; Akkerman, J.W.N.; Hoek, F.J.; Gorter, G.; Pronk, L.M.; Sturk, A.; Nieuwland, R. The Phospholipid Composition and Cholesterol Content of Platelet-Derived Microparticles: A Comparison with Platelet Membrane Fractions. Journal of Thrombosis and Haemostasis 2005, 3, 2754–2763. [Google Scholar] [CrossRef]
- Zorova, L.D.; Kovalchuk, S.I.; Popkov, V.A.; Chernikov, V.P.; Zharikova, A.A.; Khutornenko, A.A.; Zorov, S.D.; Plokhikh, K.S.; Zinovkin, R.A.; Evtushenko, E.A.; et al. Do Extracellular Vesicles Derived from Mesenchymal Stem Cells Contain Functional Mitochondria? International Journal of Molecular Sciences 2022, 23, 7408. [Google Scholar] [CrossRef]
- Claridge, B.; Lozano, J.; Poh, Q.H.; Greening, D.W. Development of Extracellular Vesicle Therapeutics: Challenges, Considerations, and Opportunities. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J Extracell Vesicles 2013, 2, 10–3402. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J Mol Biol 2016, 428, 688–692. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A Compendium of RNA, Proteins, Lipids and Metabolites in Extracellular Vesicles. Nucleic Acids Res 2019, 47, D516–D519. [Google Scholar] [CrossRef]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive Proteomic Analysis of Mesenchymal Stem Cell Exosomes Reveals Modulation of Angiogenesis via Nuclear Factor-KappaB Signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef]
- Qiu, G.; Zheng, G.; Ge, M.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. Functional Proteins of Mesenchymal Stem Cell-Derived Extracellular Vesicles. Stem Cell Res Ther 2019, 10, 359. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Golovicheva, V.V.; Varlamova, E.G.; Danilina, T.I.; Goryunov, K.V.; Shevtsova, Y.A.; Pevzner, I.B.; Zorova, L.D.; Babenko, V.A.; Evtushenko, E.A.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Afford Neuroprotection by Modulating PI3K/AKT Pathway and Calcium Oscillations. Int J Biol Sci 2022, 18, 5345–5368. [Google Scholar] [CrossRef]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lässer, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; et al. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef]
- Gao, X.; Xiong, Y.; Li, Q.; Han, M.; Shan, D.; Yang, G.; Zhang, S.; Xin, D.; Zhao, R.; Wang, Z.; et al. Extracellular Vesicle-Mediated Transfer of miR-21-5p from Mesenchymal Stromal Cells to Neurons Alleviates Early Brain Injury to Improve Cognitive Function via the PTEN/Akt Pathway after Subarachnoid Hemorrhage. Cell Death Dis 2020, 11, 363. [Google Scholar] [CrossRef]
- Xin, H.; Li, Y.; Liu, Z.; Wang, X.; Shang, X.; Cui, Y.; Zhang, Z.G.; Chopp, M. MiR-133b Promotes Neural Plasticity and Functional Recovery after Treatment of Stroke with Multipotent Mesenchymal Stromal Cells in Rats via Transfer of Exosome-Enriched Extracellular Particles. Stem Cells 2013, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, J.; Su, M.; Wang, X.; Xie, C. Exosomal microRNA-22-3p Alleviates Cerebral Ischemic Injury by Modulating KDM6B/BMP2/BMF Axis. Stem Cell Res Ther 2021, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Li, J.; Che, Y. miR-31 from Adipose Stem Cell-Derived Extracellular Vesicles Promotes Recovery of Neurological Function after Ischemic Stroke by Inhibiting TRAF6 and IRF5. Exp Neurol 2021, 342, 113611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zou, X.; Zhang, R.; Xie, Y.; Feng, Z.; Li, F.; Han, J.; Sun, H.; Ouyang, Q.; Hua, S.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomal miR-146a-5p Reduces Microglial-Mediated Neuroinflammation via Suppression of the IRAK1/TRAF6 Signaling Pathway after Ischemic Stroke. Aging (Albany NY) 2021, 13, 3060–3079. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J Extracell Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Lu, M.; Shao, W.; Xing, H.; Huang, Y. Extracellular Vesicle-Based Nucleic Acid Delivery. Interdisciplinary Medicine 2023, 1, e20220007. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat Rev Genet 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Zhuo, C.-J.; Hou, W.-H.; Jiang, D.-G.; Tian, H.-J.; Wang, L.-N.; Jia, F.; Zhou, C.-H.; Zhu, J.-J. Circular RNAs in Early Brain Development and Their Influence and Clinical Significance in Neuropsychiatric Disorders. Neural Regen Res 2020, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Chao, J.; Yao, H. Circular RNA and Its Mechanisms in Disease: From the Bench to the Clinic. Pharmacol Ther 2018, 187, 31–44. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Fitzsimmons, C.; Geaghan, M.P.; Shannon Weickert, C.; Atkins, J.R.; Wang, X.; Cairns, M.J. Circular RNA Biogenesis Is Decreased in Postmortem Cortical Gray Matter in Schizophrenia and May Alter the Bioavailability of Associated miRNA. Neuropsychopharmacology 2019, 44, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Wapinski, O.; Chang, H.Y. Long Noncoding RNAs and Human Disease. Trends Cell Biol 2011, 21, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA Gas5 Is a Growth Arrest- and Starvation-Associated Repressor of the Glucocorticoid Receptor. Sci Signal 2010, 3, ra8. [Google Scholar] [CrossRef]
- Xu, B.; Gerin, I.; Miao, H.; Vu-Phan, D.; Johnson, C.N.; Xu, R.; Chen, X.-W.; Cawthorn, W.P.; MacDougald, O.A.; Koenig, R.J. Multiple Roles for the Non-Coding RNA SRA in Regulation of Adipogenesis and Insulin Sensitivity. PLoS One 2010, 5, e14199. [Google Scholar] [CrossRef]
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a Long Non-Coding RNA Responsive to Insulin/IGF Signaling, Regulates Genes Involved in Central Metabolism. Biochim Biophys Acta 2014, 1843, 372–386. [Google Scholar] [CrossRef]
- Tong, X.; Gu, P.; Xu, S.; Lin, X. Long Non-Coding RNA-DANCR in Human Circulating Monocytes: A Potential Biomarker Associated with Postmenopausal Osteoporosis. Biosci Biotechnol Biochem 2015, 79, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.A.; Moss, L.D.; Lee, J.-Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long Noncoding RNA MALAT1 in Exosomes Drives Regenerative Function and Modulates Inflammation-Linked Networks Following Traumatic Brain Injury. J Neuroinflammation 2018, 15, 204. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat Rev Mol Cell Biol 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- De Luca, L.; Trino, S.; Laurenzana, I.; Simeon, V.; Calice, G.; Raimondo, S.; Podestà, M.; Santodirocco, M.; Di Mauro, L.; La Rocca, F.; et al. MiRNAs and piRNAs from Bone Marrow Mesenchymal Stem Cell Extracellular Vesicles Induce Cell Survival and Inhibit Cell Differentiation of Cord Blood Hematopoietic Stem Cells: A New Insight in Transplantation. Oncotarget 2016, 7, 6676–6692. [Google Scholar] [CrossRef]
- Wolin, S.L.; Steitz, J.A. The Ro Small Cytoplasmic Ribonucleoproteins: Identification of the Antigenic Protein and Its Binding Site on the Ro RNAs. Proc Natl Acad Sci U S A 1984, 81, 1996–2000. [Google Scholar] [CrossRef]
- Valkov, N.; Das, S. Y RNAs: Biogenesis, Function and Implications for the Cardiovascular System. Adv Exp Med Biol 2020, 1229, 327–342. [Google Scholar] [CrossRef]
- Haga, H.; Yan, I.K.; Takahashi, K.; Matsuda, A.; Patel, T. Extracellular Vesicles from Bone Marrow-Derived Mesenchymal Stem Cells Improve Survival from Lethal Hepatic Failure in Mice. Stem Cells Transl Med 2017, 6, 1262–1272. [Google Scholar] [CrossRef]
- Cambier, L.; Giani, J.F.; Liu, W.; Ijichi, T.; Echavez, A.K.; Valle, J.; Marbán, E. Angiotensin II-Induced End-Organ Damage in Mice Is Attenuated by Human Exosomes and by an Exosomal Y RNA Fragment. Hypertension 2018, 72, 370–380. [Google Scholar] [CrossRef]
- Zakharova, L.; Svetlova, M.; Fomina, A.F. T Cell Exosomes Induce Cholesterol Accumulation in Human Monocytes via Phosphatidylserine Receptor. J Cell Physiol 2007, 212, 174–181. [Google Scholar] [CrossRef]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as New Vesicular Lipid Transporters Involved in Cell-Cell Communication and Various Pathophysiologies. Biochim Biophys Acta 2014, 1841, 108–120. [Google Scholar] [CrossRef]
- Ghadami, S.; Dellinger, K. The Lipid Composition of Extracellular Vesicles: Applications in Diagnostics and Therapeutic Delivery. Front Mol Biosci 2023, 10, 1198044. [Google Scholar] [CrossRef]
- Lobasso, S.; Tanzarella, P.; Mannavola, F.; Tucci, M.; Silvestris, F.; Felici, C.; Ingrosso, C.; Corcelli, A.; Lopalco, P. A Lipidomic Approach to Identify Potential Biomarkers in Exosomes From Melanoma Cells With Different Metastatic Potential. Front Physiol 2021, 12, 748895. [Google Scholar] [CrossRef]
- Esser, J.; Gehrmann, U.; D’Alexandri, F.L.; Hidalgo-Estévez, A.M.; Wheelock, C.E.; Scheynius, A.; Gabrielsson, S.; Rådmark, O. Exosomes from Human Macrophages and Dendritic Cells Contain Enzymes for Leukotriene Biosynthesis and Promote Granulocyte Migration. J Allergy Clin Immunol 2010, 126, 1032–1040. [Google Scholar] [CrossRef]
- Holopainen, M.; Colas, R.A.; Valkonen, S.; Tigistu-Sahle, F.; Hyvärinen, K.; Mazzacuva, F.; Lehenkari, P.; Käkelä, R.; Dalli, J.; Kerkelä, E.; et al. Polyunsaturated Fatty Acids Modify the Extracellular Vesicle Membranes and Increase the Production of Proresolving Lipid Mediators of Human Mesenchymal Stromal Cells. Biochim Biophys Acta Mol Cell Biol Lipids 2019, 1864, 1350–1362. [Google Scholar] [CrossRef]
- Pizzinat, N.; Ong-Meang, V.; Bourgailh-Tortosa, F.; Blanzat, M.; Perquis, L.; Cussac, D.; Parini, A.; Poinsot, V. Extracellular Vesicles of MSCs and Cardiomyoblasts Are Vehicles for Lipid Mediators. Biochimie 2020, 178, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, M.; Battista, N.; Riganti, L.; Prada, I.; Antonucci, F.; Cantone, L.; Matteoli, M.; Maccarrone, M.; Verderio, C. Active Endocannabinoids Are Secreted on Extracellular Membrane Vesicles. EMBO Rep 2015, 16, 213–220. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun Signal 2021, 19, 47. [Google Scholar] [CrossRef]
- Shimoda, A.; Tahara, Y.; Sawada, S.-I.; Sasaki, Y.; Akiyoshi, K. Glycan Profiling Analysis Using Evanescent-Field Fluorescence-Assisted Lectin Array: Importance of Sugar Recognition for Cellular Uptake of Exosomes from Mesenchymal Stem Cells. Biochem Biophys Res Commun 2017, 491, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Kowal, J.; Zucchetti, A.E.; Enserink, L.; Jouve, M.; Lankar, D.; Saitakis, M.; Martin-Jaular, L.; Théry, C. Qualitative Differences in T-Cell Activation by Dendritic Cell-Derived Extracellular Vesicle Subtypes. EMBO J 2017, 36, 3012–3028. [Google Scholar] [CrossRef] [PubMed]
- Munich, S.; Sobo-Vujanovic, A.; Buchser, W.J.; Beer-Stolz, D.; Vujanovic, N.L. Dendritic Cell Exosomes Directly Kill Tumor Cells and Activate Natural Killer Cells via TNF Superfamily Ligands. Oncoimmunology 2012, 1, 1074–1083. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat Biotechnol 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Cui, G.-H.; Guo, H.-D.; Li, H.; Zhai, Y.; Gong, Z.-B.; Wu, J.; Liu, J.-S.; Dong, Y.-R.; Hou, S.-X.; Liu, J.-R. RVG-Modified Exosomes Derived from Mesenchymal Stem Cells Rescue Memory Deficits by Regulating Inflammatory Responses in a Mouse Model of Alzheimer’s Disease. Immun Ageing 2019, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular Vesicle-Based Therapeutics: Natural versus Engineered Targeting and Trafficking. Exp Mol Med 2019, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, A.; Alini, M.; Baghaban Eslaminejad, M.; Hosseini, S. Engineering Strategies for Customizing Extracellular Vesicle Uptake in a Therapeutic Context. Stem Cell Res Ther 2022, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Sturk, A.; Nieuwland, R. Extracellular Vesicles in Physiological and Pathological Conditions. Blood Rev 2013, 27, 31–39. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Wang, S.; Fletcher, S.; Niu, D.; Mitter, N.; Birch, P.R.J.; Jin, H. Message in a Bubble: Shuttling Small RNAs and Proteins Between Cells and Interacting Organisms Using Extracellular Vesicles. Annu Rev Plant Biol 2021, 72, 497–524. [Google Scholar] [CrossRef]
- Wang, F.; Cerione, R.A.; Antonyak, M.A. Isolation and Characterization of Extracellular Vesicles Produced by Cell Lines. STAR Protoc 2021, 2, 100295. [Google Scholar] [CrossRef]
- Bost, J.P.; Saher, O.; Hagey, D.; Mamand, D.R.; Liang, X.; Zheng, W.; Corso, G.; Gustafsson, O.; Görgens, A.; Smith, C.E.; et al. Growth Media Conditions Influence the Secretion Route and Release Levels of Engineered Extracellular Vesicles. Adv Healthc Mater 2022, 11, e2101658. [Google Scholar] [CrossRef] [PubMed]
- Tung, S.; Delavogia, E.; Fernandez-Gonzalez, A.; Mitsialis, S.A.; Kourembanas, S. Harnessing the Therapeutic Potential of the Stem Cell Secretome in Neonatal Diseases. Semin Perinatol 2023, 47, 151730. [Google Scholar] [CrossRef]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Frontiers in Cell and Developmental Biology 2020, 8. [Google Scholar] [CrossRef]
- Sanz-Ros, J.; Mas-Bargues, C.; Romero-García, N.; Huete-Acevedo, J.; Dromant, M.; Borrás, C. Extracellular Vesicles as Therapeutic Resources in the Clinical Environment. Int J Mol Sci 2023, 24, 2344. [Google Scholar] [CrossRef]
- Bond, S.T.; Calkin, A.C.; Drew, B.G. Adipose-Derived Extracellular Vesicles: Systemic Messengers and Metabolic Regulators in Health and Disease. Front Physiol 2022, 13, 837001. [Google Scholar] [CrossRef]
- Willis, G.R.; Reis, M.; Gheinani, A.H.; Fernandez-Gonzalez, A.; Taglauer, E.S.; Yeung, V.; Liu, X.; Ericsson, M.; Haas, E.; Mitsialis, S.A.; et al. Extracellular Vesicles Protect the Neonatal Lung from Hyperoxic Injury through the Epigenetic and Transcriptomic Reprogramming of Myeloid Cells. Am J Respir Crit Care Med 2021, 204, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Thomi, G.; Surbek, D.; Haesler, V.; Joerger-Messerli, M.; Schoeberlein, A. Exosomes Derived from Umbilical Cord Mesenchymal Stem Cells Reduce Microglia-Mediated Neuroinflammation in Perinatal Brain Injury. Stem Cell Res Ther 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, Q.; Wang, F. Mesenchymal Stem Cells for the Treatment of Ulcerative Colitis: A Systematic Review and Meta-Analysis of Experimental and Clinical Studies. Stem Cell Res Ther 2019, 10, 266. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Liu, C.-Y.; Wang, I.-J.; Sieber, M.; Chang, J.; Jester, J.V.; Kao, W.W.Y. Cell Therapy of Congenital Corneal Diseases with Umbilical Mesenchymal Stem Cells: Lumican Null Mice. PLoS One 2010, 5, e10707. [Google Scholar] [CrossRef]
- Lombardo, E.; van der Poll, T.; DelaRosa, O.; Dalemans, W. Mesenchymal Stem Cells as a Therapeutic Tool to Treat Sepsis. World J Stem Cells 2015, 7, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Joerger-Messerli, M.S.; Oppliger, B.; Spinelli, M.; Thomi, G.; di Salvo, I.; Schneider, P.; Schoeberlein, A. Extracellular Vesicles Derived from Wharton’s Jelly Mesenchymal Stem Cells Prevent and Resolve Programmed Cell Death Mediated by Perinatal Hypoxia-Ischemia in Neuronal Cells. Cell Transplant 2018, 27, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dong, L.; Zhou, D.; Li, L.; Zhang, W.; Zhen, Y.; Wang, T.; Su, J.; Chen, D.; Mao, C.; et al. Extracellular Vesicles from Human Umbilical Cord Mesenchymal Stem Cells Improve Nerve Regeneration after Sciatic Nerve Transection in Rats. J Cell Mol Med 2019, 23, 2822–2835. [Google Scholar] [CrossRef] [PubMed]
- Aiello, S.; Rocchetta, F.; Longaretti, L.; Faravelli, S.; Todeschini, M.; Cassis, L.; Pezzuto, F.; Tomasoni, S.; Azzollini, N.; Mister, M.; et al. Extracellular Vesicles Derived from T Regulatory Cells Suppress T Cell Proliferation and Prolong Allograft Survival. Sci Rep 2017, 7, 11518. [Google Scholar] [CrossRef]
- Alarcón-Vila, C.; Baroja-Mazo, A.; de Torre-Minguela, C.; Martínez, C.M.; Martínez-García, J.J.; Martínez-Banaclocha, H.; García-Palenciano, C.; Pelegrin, P. CD14 Release Induced by P2X7 Receptor Restricts Inflammation and Increases Survival during Sepsis. Elife 2020, 9, e60849. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Fan, H. Exosomes from Endothelial Progenitor Cells Improve the Outcome of a Murine Model of Sepsis. Mol Ther 2018, 26, 1375–1384. [Google Scholar] [CrossRef]
- Del Campo, C.V.; Liaw, N.Y.; Gunadasa-Rohling, M.; Matthaei, M.; Braga, L.; Kennedy, T.; Salinas, G.; Voigt, N.; Giacca, M.; Zimmermann, W.-H.; et al. Regenerative Potential of Epicardium-Derived Extracellular Vesicles Mediated by Conserved miRNA Transfer. Cardiovasc Res 2022, 118, 597–611. [Google Scholar] [CrossRef]
- Wang, H.; Maimaitiaili, R.; Yao, J.; Xie, Y.; Qiang, S.; Hu, F.; Li, X.; Shi, C.; Jia, P.; Yang, H.; et al. Percutaneous Intracoronary Delivery of Plasma Extracellular Vesicles Protects the Myocardium Against Ischemia-Reperfusion Injury in Canis. Hypertension 2021, 78, 1541–1554. [Google Scholar] [CrossRef]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; et al. Extracellular Vesicles from Human iPSC-Derived Neural Stem Cells: miRNA and Protein Signatures, and Anti-Inflammatory and Neurogenic Properties. J Extracell Vesicles 2020, 9, 1809064. [Google Scholar] [CrossRef]
- Park, H.J.; Jeon, J.; Choi, J.; Kim, J.Y.; Kim, H.S.; Huh, J.Y.; Goldman, S.A.; Song, J. Human iPSC-Derived Neural Precursor Cells Differentiate into Multiple Cell Types to Delay Disease Progression Following Transplantation into YAC128 Huntington’s Disease Mouse Model. Cell Prolif 2021, 54, e13082. [Google Scholar] [CrossRef] [PubMed]
- Attaluri, S.; Jaimes Gonzalez, J.; Kirmani, M.; Vogel, A.D.; Upadhya, R.; Kodali, M.; Madhu, L.N.; Rao, S.; Shuai, B.; Babu, R.S.; et al. Intranasally Administered Extracellular Vesicles from Human Induced Pluripotent Stem Cell-Derived Neural Stem Cells Quickly Incorporate into Neurons and Microglia in 5xFAD Mice. Front Aging Neurosci 2023, 15, 1200445. [Google Scholar] [CrossRef] [PubMed]
- McCulloh, C.J.; Olson, J.K.; Wang, Y.; Zhou, Y.; Tengberg, N.H.; Deshpande, S.; Besner, G.E. Treatment of Experimental Necrotizing Enterocolitis with Stem Cell-Derived Exosomes. J Pediatr Surg 2018, 53, 1215–1220. [Google Scholar] [CrossRef]
- Lou, P.; Liu, S.; Wang, Y.; Lv, K.; Zhou, X.; Li, L.; Zhang, Y.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Neonatal-Tissue-Derived Extracellular Vesicle Therapy (NEXT): A Potent Strategy for Precision Regenerative Medicine. Adv Mater 2023, 35, e2300602. [Google Scholar] [CrossRef]
- Babenko, V.A.; Silachev, D.N.; Danilina, T.I.; Goryunov, K.V.; Pevzner, I.B.; Zorova, L.D.; Popkov, V.A.; Chernikov, V.P.; Plotnikov, E.Y.; Sukhikh, G.T.; et al. Age-Related Changes in Bone-Marrow Mesenchymal Stem Cells. Cells 2021, 10, 1273. [Google Scholar] [CrossRef]
- Chutipongtanate, S.; Morrow, A.L.; Newburg, D.S. Human Milk Extracellular Vesicles: A Biological System with Clinical Implications. Cells 2022, 11, 2345. [Google Scholar] [CrossRef]
- Bellio, M.A.; Young, K.C.; Milberg, J.; Santos, I.; Abdullah, Z.; Stewart, D.; Arango, A.; Chen, P.; Huang, J.; Williams, K.; et al. Amniotic Fluid-Derived Extracellular Vesicles: Characterization and Therapeutic Efficacy in an Experimental Model of Bronchopulmonary Dysplasia. Cytotherapy 2021, 23, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.-M.; Zhang, K.; Zhang, J.-H. Human Breast Milk-Derived Exosomal miR-148a-3p Protects Against Necrotizing Enterocolitis by Regulating P53 and Sirtuin 1. Inflammation 2022, 45, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Iannotta, D.; Yang, M.; Celia, C.; Di Marzio, L.; Wolfram, J. Extracellular Vesicle Therapeutics from Plasma and Adipose Tissue. Nano Today 2021, 39, 101159. [Google Scholar] [CrossRef] [PubMed]
- Mitra, P.; Gupta, S.; Sharma, P. Extracellular Vesicles (EVs) as “A Window to the Brain”: Potential, Challenges and Future Perspectives. Indian J Clin Biochem 2023, 38, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Brenna, S.; Krisp, C.; Altmeppen, H.C.; Magnus, T.; Puig, B. Brain-Derived Extracellular Vesicles in Health and Disease: A Methodological Perspective. Int J Mol Sci 2021, 22, 1365. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Fan, J.; Zheng, G.; He, J.; Lin, F.; Ge, M.; Huang, L.; Wang, J.; Xia, J.; Huang, R.; et al. Diagnostic Potential of Plasma Extracellular Vesicle miR-483-3p and Let-7d-3p for Sepsis. Front Mol Biosci 2022, 9, 814240. [Google Scholar] [CrossRef]
- Igami, K.; Uchiumi, T.; Shiota, M.; Ueda, S.; Tsukahara, S.; Akimoto, M.; Eto, M.; Kang, D. Extracellular Vesicles Expressing CEACAM Proteins in the Urine of Bladder Cancer Patients. Cancer Sci 2022, 113, 3120–3133. [Google Scholar] [CrossRef]
- Ghanam, J.; Chetty, V.K.; Barthel, L.; Reinhardt, D.; Hoyer, P.-F.; Thakur, B.K. DNA in Extracellular Vesicles: From Evolution to Its Current Application in Health and Disease. Cell & Bioscience 2022, 12, 37. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, Z.; Liu, X.; Tyler, B.M. Biogenesis and Biological Functions of Extracellular Vesicles in Cellular and Organismal Communication With Microbes. Front Microbiol 2022, 13, 817844. [Google Scholar] [CrossRef]
- Urzì, O.; Gasparro, R.; Ganji, N.R.; Alessandro, R.; Raimondo, S. Plant-RNA in Extracellular Vesicles: The Secret of Cross-Kingdom Communication. Membranes (Basel) 2022, 12, 352. [Google Scholar] [CrossRef]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular Membrane Vesicles in the Three Domains of Life and Beyond. FEMS Microbiol Rev 2019, 43, 273–303. [Google Scholar] [CrossRef] [PubMed]
- Silachev, D.N. Study of the Molecular Mechanisms of the Therapeutic Properties of Extracellular Vesicles. Int J Mol Sci 2023, 24, 7093. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Singh, B.; Mir, R.A.; Nemati, M.; Babaei, A.; Ahmadi, M.; Rasmi, Y.; Golezani, A.G.; Rezaie, J. Plant-Derived Extracellular Vesicles: A Novel Nanomedicine Approach with Advantages and Challenges. Cell Commun Signal 2022, 20, 69. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.S.; Bonato, V.L.D.; Pessoni, A.M.; Rodrigues, M.L.; Casadevall, A.; Almeida, F. Fungal Extracellular Vesicles as Potential Targets for Immune Interventions. mSphere 2019, 4, e00747–19. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Giv, N.; Basas, A.; Hicks, C.; El-Omar, E.; El-Assaad, F.; Hosseini-Beheshti, E. Bacterial Extracellular Vesicles and Their Novel Therapeutic Applications in Health and Cancer. Front Cell Infect Microbiol 2022, 12, 962216. [Google Scholar] [CrossRef]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-Derived Extracellular Vesicles: Recent Advancements and Current Challenges on Their Use for Biomedical Applications. J Extracell Vesicles 2022, 11, e12283. [Google Scholar] [CrossRef] [PubMed]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota-Host Communications: Bacterial Extracellular Vesicles as a Common Language. PLoS Pathog 2021, 17, e1009508. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef]
- Rizzo, J.; Rodrigues, M.L.; Janbon, G. Extracellular Vesicles in Fungi: Past, Present, and Future Perspectives. Front Cell Infect Microbiol 2020, 10, 346. [Google Scholar] [CrossRef]
- Filip, R. An Update on the Role of Extracellular Vesicles in the Pathogenesis of Necrotizing Enterocolitis and Inflammatory Bowel Diseases. Cells 2021, 10, 3202. [Google Scholar] [CrossRef]
- Coughlan, C.; Bruce, K.D.; Burgy, O.; Boyd, T.D.; Michel, C.R.; Garcia-Perez, J.E.; Adame, V.; Anton, P.; Bettcher, B.M.; Chial, H.J.; et al. Exosome Isolation by Ultracentrifugation and Precipitation and Techniques for Downstream Analyses. Curr Protoc Cell Biol 2020, 88, e110. [Google Scholar] [CrossRef]
- Weng, Y.; Sui, Z.; Shan, Y.; Hu, Y.; Chen, Y.; Zhang, L.; Zhang, Y. Effective Isolation of Exosomes with Polyethylene Glycol from Cell Culture Supernatant for In-Depth Proteome Profiling. Analyst 2016, 141, 4640–4646. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Malekian, F.; Baghban, N.; Kodam, S.P.; Ullah, M. Methodologies to Isolate and Purify Clinical Grade Extracellular Vesicles for Medical Applications. Cells 2022, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Kurinczuk, J.J.; White-Koning, M.; Badawi, N. Epidemiology of Neonatal Encephalopathy and Hypoxic-Ischaemic Encephalopathy. Early Hum Dev 2010, 86, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.A.; Brandon, D.H. Hypoxic Ischemic Encephalopathy: Pathophysiology and Experimental Treatments. Newborn Infant Nurs Rev 2011, 11, 125–133. [Google Scholar] [CrossRef]
- Silachev, D.N.; Plotnikov, E.Y.; Pevzner, I.B.; Zorova, L.D.; Balakireva, A.V.; Gulyaev, M.V.; Pirogov, Y.A.; Skulachev, V.P.; Zorov, D.B. Neuroprotective Effects of Mitochondria-Targeted Plastoquinone in a Rat Model of Neonatal Hypoxic−Ischemic Brain Injury. Molecules 2018, 23, 1871. [Google Scholar] [CrossRef]
- Robertsson Grossmann, K.; Eriksson Westblad, M.; Blennow, M.; Lindström, K. Outcome at Early School Age and Adolescence after Hypothermia-Treated Hypoxic-Ischaemic Encephalopathy: An Observational, Population-Based Study. Arch Dis Child Fetal Neonatal Ed 2023, 108, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, S.; Pappas, A.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Yolton, K.; Gustafson, K.E.; Leach, T.M.; Green, C.; Bara, R.; et al. Childhood Outcomes after Hypothermia for Neonatal Encephalopathy. N Engl J Med 2012, 366, 2085–2092. [Google Scholar] [CrossRef]
- Azzopardi, D.; Strohm, B.; Marlow, N.; Brocklehurst, P.; Deierl, A.; Eddama, O.; Goodwin, J.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; et al. Effects of Hypothermia for Perinatal Asphyxia on Childhood Outcomes. N Engl J Med 2014, 371, 140–149. [Google Scholar] [CrossRef]
- Luo, B.-Y.; Zhou, H.-S.; Sun, Y.-F.; Xiao, Q.-X.; Chen, L.; She, H.-Q.; Wang, S.-F.; Yan, S.-S.; Chang, Q.-Y.; He, Y.-Q.; et al. The Fate and Prospects of Stem Cell Therapy in the Treatment of Hypoxic-Ischemic Encephalopathy. Eur J Neurosci 2023, 58, 2384–2405. [Google Scholar] [CrossRef] [PubMed]
- Сухих, Г.Т.; Силачев, Д.Н.; Певзнер, И.Б.; Зoрoва, Л.Д.; Бабенкo, В.А.; Пoпкoв, В.А.; Янкаускас, С.С.; Зубкoв, В.В.; Зoрoв, Д.Б.; Плoтникoв, Е.Ю. Перспективы Испoльзoвания Ствoлoвых и Прoгенитoрных Клетoк Для Терапии Пoследствий Гипoксически-Ишемическoй Энцефалoпатии Нoвoрoжденных. Акушерствo и гинекoлoгия 2016, 55–66. [Google Scholar] [CrossRef]
- Lee, J.A.; Kim, B.I.; Jo, C.H.; Choi, C.W.; Kim, E.-K.; Kim, H.-S.; Yoon, K.-S.; Choi, J.-H. Mesenchymal Stem-Cell Transplantation for Hypoxic-Ischemic Brain Injury in Neonatal Rat Model. Pediatr Res 2010, 67, 42–46. [Google Scholar] [CrossRef] [PubMed]
- van Velthoven, C.T.J.; van de Looij, Y.; Kavelaars, A.; Zijlstra, J.; van Bel, F.; Huppi, P.S.; Sizonenko, S.; Heijnen, C.J. Mesenchymal Stem Cells Restore Cortical Rewiring after Neonatal Ischemia in Mice. Ann Neurol 2012, 71, 785–796. [Google Scholar] [CrossRef]
- van Velthoven, C.T.J.; Kavelaars, A.; van Bel, F.; Heijnen, C.J. Mesenchymal Stem Cell Treatment after Neonatal Hypoxic-Ischemic Brain Injury Improves Behavioral Outcome and Induces Neuronal and Oligodendrocyte Regeneration. Brain Behav Immun 2010, 24, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Ahn, S.Y.; Im, G.H.; Sung, D.K.; Park, Y.R.; Choi, S.H.; Choi, S.J.; Chang, Y.S.; Oh, W.; Lee, J.H.; et al. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cell Transplantation Attenuates Severe Brain Injury by Permanent Middle Cerebral Artery Occlusion in Newborn Rats. Pediatr Res 2012, 72, 277–284. [Google Scholar] [CrossRef]
- Ding, D.-C.; Shyu, W.-C.; Chiang, M.-F.; Lin, S.-Z.; Chang, Y.-C.; Wang, H.-J.; Su, C.-Y.; Li, H. Enhancement of Neuroplasticity through Upregulation of Beta1-Integrin in Human Umbilical Cord-Derived Stromal Cell Implanted Stroke Model. Neurobiol Dis 2007, 27, 339–353. [Google Scholar] [CrossRef]
- Ding, H.-F.; Zhang, H.; Ding, H.-F.; Li, D.; Yi, X.-H.; Gao, X.-Y.; Mou, W.-W.; Ju, X.-L. Therapeutic Effect of Placenta-Derived Mesenchymal Stem Cells on Hypoxic-Ischemic Brain Damage in Rats. World J Pediatr 2015, 11, 74–82. [Google Scholar] [CrossRef]
- Donega, V.; Velthoven, C.T.J. van; Nijboer, C.H.; Bel, F. van; Kas, M.J.H.; Kavelaars, A.; Heijnen, C.J. Intranasal Mesenchymal Stem Cell Treatment for Neonatal Brain Damage: Long-Term Cognitive and Sensorimotor Improvement. PLOS ONE 2013, 8, e51253. [Google Scholar] [CrossRef]
- Allan, D.; Tieu, A.; Lalu, M.; Burger, D. Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Regenerative Therapy and Immune Modulation: Progress and Challenges toward Clinical Application. Stem Cells Translational Medicine 2020, 9, 39–46. [Google Scholar] [CrossRef]
- Akyurekli, C.; Le, Y.; Richardson, R.B.; Fergusson, D.; Tay, J.; Allan, D.S. A Systematic Review of Preclinical Studies on the Therapeutic Potential of Mesenchymal Stromal Cell-Derived Microvesicles. Stem Cell Rev Rep 2015, 11, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Sisa, C.; Kholia, S.; Naylor, J.; Herrera Sanchez, M.B.; Bruno, S.; Deregibus, M.C.; Camussi, G.; Inal, J.M.; Lange, S.; Hristova, M. Mesenchymal Stromal Cell Derived Extracellular Vesicles Reduce Hypoxia-Ischaemia Induced Perinatal Brain Injury. Front Physiol 2019, 10, 282. [Google Scholar] [CrossRef]
- Kaminski, N.; Köster, C.; Mouloud, Y.; Börger, V.; Felderhoff-Müser, U.; Bendix, I.; Giebel, B.; Herz, J. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Reduce Neuroinflammation, Promote Neural Cell Proliferation and Improve Oligodendrocyte Maturation in Neonatal Hypoxic-Ischemic Brain Injury. Front Cell Neurosci 2020, 14, 601176. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Jiang, L.; Wang, M.; Wang, R.; Wang, X.; Gao, C.; Xia, Z. Human Bone Marrow Mesenchymal Stem Cells-Derived Exosomes Protect against Nerve Injury via Regulating Immune Microenvironment in Neonatal Hypoxic-Ischemic Brain Damage Model. Immunobiology 2022, 227, 152178. [Google Scholar] [CrossRef]
- Hu, Z.; Yuan, Y.; Zhang, X.; Lu, Y.; Dong, N.; Jiang, X.; Xu, J.; Zheng, D. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Attenuate Oxygen-Glucose Deprivation/Reperfusion-Induced Microglial Pyroptosis by Promoting FOXO3a-Dependent Mitophagy. Oxid Med Cell Longev 2021, 2021, 6219715. [Google Scholar] [CrossRef] [PubMed]
- Cristante, E.; McArthur, S.; Mauro, C.; Maggioli, E.; Romero, I.A.; Wylezinska-Arridge, M.; Couraud, P.O.; Lopez-Tremoleda, J.; Christian, H.C.; Weksler, B.B.; et al. Identification of an Essential Endogenous Regulator of Blood-Brain Barrier Integrity, and Its Pathological and Therapeutic Implications. Proc Natl Acad Sci U S A 2013, 110, 832–841. [Google Scholar] [CrossRef]
- Gussenhoven, R.; Klein, L.; Ophelders, D.R.M.G.; Habets, D.H.J.; Giebel, B.; Kramer, B.W.; Schurgers, L.J.; Reutelingsperger, C.P.M.; Wolfs, T.G.A.M. Annexin A1 as Neuroprotective Determinant for Blood-Brain Barrier Integrity in Neonatal Hypoxic-Ischemic Encephalopathy. J Clin Med 2019, 8, E137. [Google Scholar] [CrossRef]
- Yang, Y.; Cai, Y.; Zhang, Y.; Liu, J.; Xu, Z. Exosomes Secreted by Adipose-Derived Stem Cells Contribute to Angiogenesis of Brain Microvascular Endothelial Cells Following Oxygen-Glucose Deprivation In Vitro Through MicroRNA-181b/TRPM7 Axis. J Mol Neurosci 2018, 65, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Geng, W.; Tang, H.; Luo, S.; Lv, Y.; Liang, D.; Kang, X.; Hong, W. Exosomes from miRNA-126-Modified ADSCs Promotes Functional Recovery after Stroke in Rats by Improving Neurogenesis and Suppressing Microglia Activation. Am J Transl Res 2019, 11, 780–792. [Google Scholar]
- Budnik, V.; Ruiz-Cañada, C.; Wendler, F. Extracellular Vesicles Round off Communication in the Nervous System. Nat Rev Neurosci 2016, 17, 160–172. [Google Scholar] [CrossRef]
- Song, Y.; Li, Z.; He, T.; Qu, M.; Jiang, L.; Li, W.; Shi, X.; Pan, J.; Zhang, L.; Wang, Y.; et al. M2 Microglia-Derived Exosomes Protect the Mouse Brain from Ischemia-Reperfusion Injury via Exosomal miR-124. Theranostics 2019, 9, 2910–2923. [Google Scholar] [CrossRef]
- Luo, H.; Ye, G.; Liu, Y.; Huang, D.; Luo, Q.; Chen, W.; Qi, Z. miR-150-3p Enhances Neuroprotective Effects of Neural Stem Cell Exosomes after Hypoxic-Ischemic Brain Injury by Targeting CASP2. Neurosci Lett 2022, 779, 136635. [Google Scholar] [CrossRef] [PubMed]
- Holme, N.; Chetcuti, P. The Pathophysiology of Respiratory Distress Syndrome in Neonates. Paediatrics and Child Health 2012, 22, 507–512. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Ramamoorthy, C. Bronchopulmonary Dysplasia. Paediatr Anaesth 2022, 32, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Principi, N.; Di Pietro, G.M.; Esposito, S. Bronchopulmonary Dysplasia: Clinical Aspects and Preventive and Therapeutic Strategies. J Transl Med 2018, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Geetha, O.; Rajadurai, V.S.; Anand, A.J.; Dela Puerta, R.; Huey Quek, B.; Khoo, P.C.; Chua, M.C.; Agarwal, P. New BPD-Prevalence and Risk Factors for Bronchopulmonary Dysplasia/Mortality in Extremely Low Gestational Age Infants ≤28 Weeks. J Perinatol 2021, 41, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Pierro, M.; Ionescu, L.; Montemurro, T.; Vadivel, A.; Weissmann, G.; Oudit, G.; Emery, D.; Bodiga, S.; Eaton, F.; Péault, B.; et al. Short-Term, Long-Term and Paracrine Effect of Human Umbilical Cord-Derived Stem Cells in Lung Injury Prevention and Repair in Experimental Bronchopulmonary Dysplasia. Thorax 2013, 68, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Ameliorate Experimental Bronchopulmonary Dysplasia and Restore Lung Function through Macrophage Immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef]
- Wang, J.; Dong, W. Oxidative Stress and Bronchopulmonary Dysplasia. Gene 2018, 678, 177–183. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, A.; Huang, F.; Xu, J.; Zhao, M. MSC-EXO and Tempol Ameliorate Bronchopulmonary Dysplasia in Newborn Rats by Activating HIF-1α. Pediatr Pulmonol 2023, 58, 1367–1379. [Google Scholar] [CrossRef]
- Chiaradia, E.; Tancini, B.; Emiliani, C.; Delo, F.; Pellegrino, R.M.; Tognoloni, A.; Urbanelli, L.; Buratta, S. Extracellular Vesicles under Oxidative Stress Conditions: Biological Properties and Physiological Roles. Cells 2021, 10, 1763. [Google Scholar] [CrossRef]
- Sharma, M.; Bellio, M.A.; Benny, M.; Kulandavelu, S.; Chen, P.; Janjindamai, C.; Han, C.; Chang, L.; Sterling, S.; Williams, K.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Prevent Experimental Bronchopulmonary Dysplasia Complicated By Pulmonary Hypertension. Stem Cells Transl Med 2022, 11, 828–840. [Google Scholar] [CrossRef]
- Ai, D.; Shen, J.; Sun, J.; Zhu, Z.; Gao, R.; Du, Y.; Yuan, L.; Chen, C.; Zhou, J. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppress Hyperoxia-Induced Transdifferentiation of Rat Alveolar Type 2 Epithelial Cells. Stem Cells Dev 2022, 31, 53–66. [Google Scholar] [CrossRef]
- Lee, C.; Mitsialis, S.A.; Aslam, M.; Vitali, S.H.; Vergadi, E.; Konstantinou, G.; Sdrimas, K.; Fernandez-Gonzalez, A.; Kourembanas, S. Exosomes Mediate the Cytoprotective Action of Mesenchymal Stromal Cells on Hypoxia-Induced Pulmonary Hypertension. Circulation 2012, 126, 2601–2611. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Willis, G.R.; Fernandez-Gonzalez, A.; Yeung, V.; Taglauer, E.; Magaletta, M.; Parsons, T.; Derr, A.; Liu, X.; Maehr, R.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Restore Thymic Architecture and T Cell Function Disrupted by Neonatal Hyperoxia. Front Immunol 2021, 12, 640595. [Google Scholar] [CrossRef]
- Chaubey, S.; Thueson, S.; Ponnalagu, D.; Alam, M.A.; Gheorghe, C.P.; Aghai, Z.; Singh, H.; Bhandari, V. Early Gestational Mesenchymal Stem Cell Secretome Attenuates Experimental Bronchopulmonary Dysplasia in Part via Exosome-Associated Factor TSG-6. Stem Cell Res Ther 2018, 9, 173. [Google Scholar] [CrossRef]
- Braun, R.K.; Chetty, C.; Balasubramaniam, V.; Centanni, R.; Haraldsdottir, K.; Hematti, P.; Eldridge, M.W. Intraperitoneal Injection of MSC-Derived Exosomes Prevent Experimental Bronchopulmonary Dysplasia. Biochem Biophys Res Commun 2018, 503, 2653–2658. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Park, W.S.; Kim, Y.E.; Sung, D.K.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Vascular Endothelial Growth Factor Mediates the Therapeutic Efficacy of Mesenchymal Stem Cell-Derived Extracellular Vesicles against Neonatal Hyperoxic Lung Injury. Exp Mol Med 2018, 50, 1–12. [Google Scholar] [CrossRef]
- Hou, A.; Fu, J.; Yang, H.; Zhu, Y.; Pan, Y.; Xu, S.; Xue, X. Hyperoxia Stimulates the Transdifferentiation of Type II Alveolar Epithelial Cells in Newborn Rats. Am J Physiol Lung Cell Mol Physiol 2015, 308, L861–872. [Google Scholar] [CrossRef] [PubMed]
- Lithopoulos, M.A.; Strueby, L.; O’Reilly, M.; Zhong, S.; Möbius, M.A.; Eaton, F.; Fung, M.; Hurskainen, M.; Cyr-Depauw, C.; Suen, C.; et al. Pulmonary and Neurologic Effects of Mesenchymal Stromal Cell Extracellular Vesicles in a Multifactorial Lung Injury Model. Am J Respir Crit Care Med 2022, 205, 1186–1201. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Zhou, O.; Liu, J.; Zou, W.; Zhang, L.; Tian, D.; Dai, J.; Luo, Z.; Liu, E.; Fu, Z.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Alleviate Lung Injury in Rat Model of Bronchopulmonary Dysplasia by Affecting Cell Survival and Angiogenesis. Stem Cells Dev 2020, 29, 1520–1532. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, J.; Yuan, R.; Deng, Z.; Wu, X. Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Alleviate Hyperoxia-Induced Lung Injury via the Manipulation of microRNA-425. Arch Biochem Biophys 2021, 697, 108712. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Krause, M.; Yawno, T.; Kusuma, G.D.; Schwab, R.; Barabadi, M.; Maleken, A.S.; Chan, S.T.; Hunt, R.; Greening, D.; et al. Assessing the Impact of Gestational Age of Donors on the Efficacy of Amniotic Epithelial Cell-Derived Extracellular Vesicles in Experimental Bronchopulmonary Dysplasia. Stem Cell Res Ther 2022, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.R.; Sills, W.S.; Hanrahan, K.; Ziegler, A.; Tidd, K.M.; Cook, E.; Sannes, P.L. Expression of WNT5A in Idiopathic Pulmonary Fibrosis and Its Control by TGF-β and WNT7B in Human Lung Fibroblasts. J Histochem Cytochem 2016, 64, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Edmond, K.; Zaidi, A. New Approaches to Preventing, Diagnosing, and Treating Neonatal Sepsis. PLoS Med 2010, 7, e1000213. [Google Scholar] [CrossRef] [PubMed]
- Shane, A.L.; Sánchez, P.J.; Stoll, B.J. Neonatal Sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, L.; Qian, K. Global, Regional, and National Incidence and Mortality of Neonatal Sepsis and Other Neonatal Infections, 1990–2019. Front Public Health 2023, 11, 1139832. [Google Scholar] [CrossRef]
- Burgelman, M.; Vandendriessche, C.; Vandenbroucke, R.E. Extracellular Vesicles: A Double-Edged Sword in Sepsis. Pharmaceuticals (Basel) 2021, 14, 829. [Google Scholar] [CrossRef]
- Kronstadt, S.M.; Pottash, A.E.; Levy, D.; Wang, S.; Chao, W.; Jay, S.M. Therapeutic Potential of Extracellular Vesicles for Sepsis Treatment. Adv Ther (Weinh) 2021, 4, 2000259. [Google Scholar] [CrossRef]
- Vergnano, S.; Sharland, M.; Kazembe, P.; Mwansambo, C.; Heath, P.T. Neonatal Sepsis: An International Perspective. Archives of Disease in Childhood - Fetal and Neonatal Edition 2005, 90, F220–FF224. [Google Scholar] [CrossRef]
- Kim, Y.-E.; Ahn, S.-Y.; Park, W.-S.; Sung, D.-K.; Sung, S.-I.; Yang, M.-S.; Chang, Y.-S. Mesenchymal-Stem-Cell-Derived Extracellular Vesicles Attenuate Brain Injury in Escherichia Coli Meningitis in Newborn Rats. Life 2022, 12, 1030. [Google Scholar] [CrossRef]
- Alganabi, M.; Lee, C.; Bindi, E.; Li, B.; Pierro, A. Recent Advances in Understanding Necrotizing Enterocolitis. F1000Res 2019, 8, F1000 Faculty Rev-107. [Google Scholar] [CrossRef]
- Bazacliu, C.; Neu, J. Necrotizing Enterocolitis: Long Term Complications. Curr Pediatr Rev 2019, 15, 115–124. [Google Scholar] [CrossRef]
- Hackam, D.J.; Sodhi, C.P.; Good, M. New Insights into Necrotizing Enterocolitis: From Laboratory Observation to Personalized Prevention and Treatment. J Pediatr Surg 2019, 54, 398–404. [Google Scholar] [CrossRef]
- Li, B.; Lee, C.; O’Connell, J.S.; Antounians, L.; Ganji, N.; Alganabi, M.; Cadete, M.; Nascimben, F.; Koike, Y.; Hock, A.; et al. Activation of Wnt Signaling by Amniotic Fluid Stem Cell-Derived Extracellular Vesicles Attenuates Intestinal Injury in Experimental Necrotizing Enterocolitis. Cell Death Dis 2020, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Drucker, N.A.; Te Winkel, J.P.; Shelley, W.C.; Olson, K.R.; Markel, T.A. Inhibiting Hydrogen Sulfide Production in Umbilical Stem Cells Reduces Their Protective Effects during Experimental Necrotizing Enterocolitis. J Pediatr Surg 2019, 54, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.S.; Li, B.; Zito, A.; Ahmed, A.; Cadete, M.; Ganji, N.; Lau, E.; Alganabi, M.; Farhat, N.; Lee, C.; et al. Treatment of Necrotizing Enterocolitis by Conditioned Medium Derived from Human Amniotic Fluid Stem Cells. PLoS One 2021, 16, e0260522. [Google Scholar] [CrossRef] [PubMed]
- Rager, T.M.; Olson, J.K.; Zhou, Y.; Wang, Y.; Besner, G.E. Exosomes Secreted from Bone Marrow-Derived Mesenchymal Stem Cells Protect the Intestines from Experimental Necrotizing Enterocolitis. Journal of Pediatric Surgery 2016, 51, 942–947. [Google Scholar] [CrossRef]
- Claud, E.C. Neonatal Necrotizing Enterocolitis -Inflammation and Intestinal Immaturity. Antiinflamm Antiallergy Agents Med Chem 2009, 8, 248–259. [Google Scholar] [CrossRef]
- Heidari, N.; Abbasi-Kenarsari, H.; Namaki, S.; Baghaei, K.; Zali, M.R.; Ghaffari Khaligh, S.; Hashemi, S.M. Adipose-Derived Mesenchymal Stem Cell-Secreted Exosome Alleviates Dextran Sulfate Sodium-Induced Acute Colitis by Treg Cell Induction and Inflammatory Cytokine Reduction. J Cell Physiol 2021, 236, 5906–5920. [Google Scholar] [CrossRef]
- Yang, S.; Liang, X.; Song, J.; Li, C.; Liu, A.; Luo, Y.; Ma, H.; Tan, Y.; Zhang, X. A Novel Therapeutic Approach for Inflammatory Bowel Disease by Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells to Repair Intestinal Barrier via TSG-6. Stem Cell Res Ther 2021, 12, 315. [Google Scholar] [CrossRef]
- Yan, Y.; Li, K.; Jiang, J.; Jiang, L.; Ma, X.; Ai, F.; Qiu, S.; Si, W. Perinatal Tissue-Derived Exosomes Ameliorate Colitis in Mice by Regulating the Foxp3 + Treg Cells and Gut Microbiota. Stem Cell Res Ther 2023, 14, 43. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, Z.-Y.; Yuan, J.-T.; Ocansey, D.K.W.; Tu, Q.; Zhang, X.; Qian, H.; Xu, W.-R.; Qiu, W.; Mao, F. hucMSC-Derived Exosomes Attenuate Colitis by Regulating Macrophage Pyroptosis via the miR-378a-5p/NLRP3 Axis. Stem Cell Res Ther 2021, 12, 416. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shen, Y.; Guo, D.; Yang, D.; Liu, J.; Fei, X.; Yang, Y.; Zhang, B.; Lin, Z.; Yang, F.; et al. EpCAM-Dependent Extracellular Vesicles from Intestinal Epithelial Cells Maintain Intestinal Tract Immune Balance. Nat Commun 2016, 7, 13045. [Google Scholar] [CrossRef]
- Duan, L.; Huang, H.; Zhao, X.; Zhou, M.; Chen, S.; Wang, C.; Han, Z.; Han, Z.-C.; Guo, Z.; Li, Z.; et al. Extracellular Vesicles Derived from Human Placental Mesenchymal Stem Cells Alleviate Experimental Colitis in Mice by Inhibiting Inflammation and Oxidative Stress. Int J Mol Med 2020, 46, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yang, X.; Xiao, X.; Xu, M.; Yang, Y.; Xue, C.; Li, X.; Wang, S.; Zhao, R.C. Human Adipose Mesenchymal Stem Cell-Derived Exosomes Protect Mice from DSS-Induced Inflammatory Bowel Disease by Promoting Intestinal-Stem-Cell and Epithelial Regeneration. Aging Dis 2021, 12, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xie, M.-Y.; Sun, J.-J.; Ye, R.-S.; Cheng, X.; Sun, R.-P.; Wei, L.-M.; Li, M.; Lin, D.-L.; Jiang, Q.-Y.; et al. Porcine Milk-Derived Exosomes Promote Proliferation of Intestinal Epithelial Cells. Sci Rep 2016, 6, 33862. [Google Scholar] [CrossRef] [PubMed]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Määttänen, P.; Zani, A.; Pierro, A. Breast Milk-Derived Exosomes Promote Intestinal Epithelial Cell Growth. J Pediatr Surg 2017, 52, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Accarie, A.; l’Homme, B.; Benadjaoud, M.A.; Lim, S.K.; Guha, C.; Benderitter, M.; Tamarat, R.; Sémont, A. Extracellular Vesicles Derived from Mesenchymal Stromal Cells Mitigate Intestinal Toxicity in a Mouse Model of Acute Radiation Syndrome. Stem Cell Res Ther 2020, 11, 371. [Google Scholar] [CrossRef]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.W.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E.; et al. Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol Nutr Food Res 2019, 63, e1801214. [Google Scholar] [CrossRef]
- van Herwijnen, M.J.C.; Zonneveld, M.I.; Goerdayal, S.; Nolte-’t Hoen, E.N.M.; Garssen, J.; Stahl, B.; Maarten Altelaar, A.F.; Redegeld, F.A.; Wauben, M.H.M. Comprehensive Proteomic Analysis of Human Milk-Derived Extracellular Vesicles Unveils a Novel Functional Proteome Distinct from Other Milk Components. Mol Cell Proteomics 2016, 15, 3412–3423. [Google Scholar] [CrossRef]
- Walker, W.A.; Iyengar, R.S. Breast Milk, Microbiota, and Intestinal Immune Homeostasis. Pediatr Res 2015, 77, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hell, L.; Kendlbacher, R.A.; Hajji, N.; Hau, C.; van Dam, A.; Berckmans, R.J.; Wisgrill, L.; Ay, C.; Pabinger, I.; et al. Human Milk Triggers Coagulation via Tissue Factor-Exposing Extracellular Vesicles. Blood Adv 2020, 4, 6274–6282. [Google Scholar] [CrossRef]
- Kersin, S.G.; Özek, E. Breast Milk Stem Cells: Are They Magic Bullets in Neonatology? Turk Arch Pediatr 2021, 56, 187–191. [Google Scholar] [CrossRef]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.-L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a Functional Mammary Gland from a Single Stem Cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front Immunol 2017, 8, 584. [Google Scholar] [CrossRef]
- Maghraby, M.K.; Li, B.; Chi, L.; Ling, C.; Benmoussa, A.; Provost, P.; Postmus, A.C.; Abdi, A.; Pierro, A.; Bourdon, C.; et al. Extracellular Vesicles Isolated from Milk Can Improve Gut Barrier Dysfunction Induced by Malnutrition. Sci Rep 2021, 11, 7635. [Google Scholar] [CrossRef]
- Herwijnen, M.J.C. van; Driedonks, T.A.P.; Snoek, B.L.; Kroon, A.M.T.; Kleinjan, M.; Jorritsma, R.; Pieterse, C.M.J.; Hoen, E.N.M.N.-. ‘t; Wauben, M.H.M. Abundantly Present miRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Frontiers in Nutrition 2018, 5. [Google Scholar] [CrossRef]
- Liu, X.; Zhan, Z.; Xu, L.; Ma, F.; Li, D.; Guo, Z.; Li, N.; Cao, X. MicroRNA-148/152 Impair Innate Response and Antigen Presentation of TLR-Triggered Dendritic Cells by Targeting CaMKIIα. J Immunol 2010, 185, 7244–7251. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and Peptidomic Profiling of Exosomes in Preterm Human Milk: Insights Into Necrotizing Enterocolitis Prevention. Mol Nutr Food Res 2019, 63, e1801247. [Google Scholar] [CrossRef]
- Cheyuo, C.; Aziz, M.; Yang, W.-L.; Jacob, A.; Zhou, M.; Wang, P. Milk Fat Globule-EGF Factor VIII Attenuates CNS Injury by Promoting Neural Stem Cell Proliferation and Migration after Cerebral Ischemia. PLoS One 2015, 10, e0122833. [Google Scholar] [CrossRef]
- Bu, H.-F.; Zuo, X.-L.; Wang, X.; Ensslin, M.A.; Koti, V.; Hsueh, W.; Raymond, A.S.; Shur, B.D.; Tan, X.-D. Milk Fat Globule-EGF Factor 8/Lactadherin Plays a Crucial Role in Maintenance and Repair of Murine Intestinal Epithelium. J Clin Invest 2007, 117, 3673–3683. [Google Scholar] [CrossRef]
- Tobita, K.; Kawahara, T.; Otani, H. Bovine Beta-Casein (1-28), a Casein Phosphopeptide, Enhances Proliferation and IL-6 Expression of Mouse CD19+ Cells via Toll-like Receptor 4. J Agric Food Chem 2006, 54, 8013–8017. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y.; Kim, S.H. Enhancement of Antioxidative and Intestinal Anti-Inflammatory Activities of Glycated Milk Casein after Fermentation with Lactobacillus Rhamnosus 4B15. J Agric Food Chem 2017, 65, 4744–4754. [Google Scholar] [CrossRef]
- Caccavo, D.; Pellegrino, N.M.; Altamura, M.; Rigon, A.; Amati, L.; Amoroso, A.; Jirillo, E. Antimicrobial and Immunoregulatory Functions of Lactoferrin and Its Potential Therapeutic Application. J Endotoxin Res 2002, 8, 403–417. [Google Scholar] [CrossRef]
- Nichols, B.L.; McKee, K.; Putman, M.; Henry, J.F.; Nichols, V.N. Human Lactoferrin Supplementation of Infant Formulas Increases Thymidine Incorporation into the DNA of Rat Crypt Cells. J Pediatr Gastroenterol Nutr 1989, 8, 102–109. [Google Scholar] [CrossRef]
- Sanwlani, R.; Fonseka, P.; Chitti, S.V.; Mathivanan, S. Milk-Derived Extracellular Vesicles in Inter-Organism, Cross-Species Communication and Drug Delivery. Proteomes 2020, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. Journal of Pediatric Surgery 2020, 55, 54–58. [Google Scholar] [CrossRef]
- Du, C.; Zhao, Y.; Wang, K.; Nan, X.; Chen, R.; Xiong, B. Effects of Milk-Derived Extracellular Vesicles on the Colonic Transcriptome and Proteome in Murine Model. Nutrients 2022, 14, 3057. [Google Scholar] [CrossRef]
- Du, C.; Wang, K.; Zhao, Y.; Nan, X.; Chen, R.; Quan, S.; Xiong, B. Supplementation with Milk-Derived Extracellular Vesicles Shapes the Gut Microbiota and Regulates the Transcriptomic Landscape in Experimental Colitis. Nutrients 2022, 14, 1808. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An Epigenetic Switch Involving NF-κB, Lin28, Let-7 MicroRNA, and IL6 Links Inflammation to Cell Transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine Milk-Derived Exosomes Enhance Goblet Cell Activity and Prevent the Development of Experimental Necrotizing Enterocolitis. PLoS One 2019, 14, e0211431. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, W.; Lan, P.; Mou, X. The Microbiome in Inflammatory Bowel Diseases: From Pathogenesis to Therapy. Protein Cell 2021, 12, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K. Capsular Polysaccharides of Symbiotic Bacteria Modulate Immune Responses during Experimental Colitis. J Pediatr Gastroenterol Nutr 2008, 46 Suppl 1, E11–12. [Google Scholar] [CrossRef]
- Shen, Y.; Giardino Torchia, M.L.; Lawson, G.W.; Karp, C.L.; Ashwell, J.D.; Mazmanian, S.K. Outer Membrane Vesicles of a Human Commensal Mediate Immune Regulation and Disease Protection. Cell Host Microbe 2012, 12, 509–520. [Google Scholar] [CrossRef]
- Wang, X.; Lin, S.; Wang, L.; Cao, Z.; Zhang, M.; Zhang, Y.; Liu, R.; Liu, J. Versatility of Bacterial Outer Membrane Vesicles in Regulating Intestinal Homeostasis. Sci Adv 2023, 9, eade5079. [Google Scholar] [CrossRef]
- Kang, C.-S.; Ban, M.; Choi, E.-J.; Moon, H.-G.; Jeon, J.-S.; Kim, D.-K.; Park, S.-K.; Jeon, S.G.; Roh, T.-Y.; Myung, S.-J.; et al. Extracellular Vesicles Derived from Gut Microbiota, Especially Akkermansia Muciniphila, Protect the Progression of Dextran Sulfate Sodium-Induced Colitis. PLoS One 2013, 8, e76520. [Google Scholar] [CrossRef]
- Shi, R.; Yu, F.; Hu, X.; Liu, Y.; Jin, Y.; Ren, H.; Lu, S.; Guo, J.; Chang, J.; Li, Y.; et al. Protective Effect of Lactiplantibacillus Plantarum Subsp. Plantarum SC-5 on Dextran Sulfate Sodium—Induced Colitis in Mice. Foods 2023, 12, 897. [Google Scholar] [CrossRef]
- Ma, L.; Lyu, W.; Song, Y.; Chen, K.; Lv, L.; Yang, H.; Wang, W.; Xiao, Y. Anti-Inflammatory Effect of Clostridium Butyricum-Derived Extracellular Vesicles in Ulcerative Colitis: Impact on Host microRNAs Expressions and Gut Microbiome Profiles. Mol Nutr Food Res 2023, 67, e2200884. [Google Scholar] [CrossRef]
- Rui, T.; Wang, K.; Xiang, A.; Guo, J.; Tang, N.; Jin, X.; Lin, Y.; Liu, J.; Zhang, X. Serum Exosome-Derived piRNAs Could Be Promising Biomarkers for HCC Diagnosis. Int J Nanomedicine 2023, 18, 1989–2001. [Google Scholar] [CrossRef]
- Matei, A.C.; Antounians, L.; Zani, A. Extracellular Vesicles as a Potential Therapy for Neonatal Conditions: State of the Art and Challenges in Clinical Translation. Pharmaceutics 2019, 11, 404. [Google Scholar] [CrossRef]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-Stranded DNA in Exosomes: A Novel Biomarker in Cancer Detection. Cell Res 2014, 24, 766–769. [Google Scholar] [CrossRef]
- Whitford, W.; Guterstam, P. Exosome Manufacturing Status. Future Med Chem 2019, 11, 1225–1236. [Google Scholar] [CrossRef]
- Pachler, K.; Lener, T.; Streif, D.; Dunai, Z.A.; Desgeorges, A.; Feichtner, M.; Öller, M.; Schallmoser, K.; Rohde, E.; Gimona, M. A Good Manufacturing Practice-Grade Standard Protocol for Exclusively Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles. Cytotherapy 2017, 19, 458–472. [Google Scholar] [CrossRef]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Toward Exosome-Based Therapeutics: Isolation, Heterogeneity, and Fit-for-Purpose Potency. Front Cardiovasc Med 2017, 4, 63. [Google Scholar] [CrossRef]
- Silachev, D.N.; Goryunov, K.V.; Shpilyuk, M.A.; Beznoschenko, O.S.; Morozova, N.Y.; Kraevaya, E.E.; Popkov, V.A.; Pevzner, I.B.; Zorova, L.D.; Evtushenko, E.A.; et al. Effect of MSCs and MSC-Derived Extracellular Vesicles on Human Blood Coagulation. Cells 2019, 8. [Google Scholar] [CrossRef]
- Guillamat-Prats, R. Role of Mesenchymal Stem/Stromal Cells in Coagulation. Int J Mol Sci 2022, 23, 10393. [Google Scholar] [CrossRef]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ Res 2016, 118, 1392–1408. [Google Scholar] [CrossRef] [PubMed]
- Moll, G.; Ankrum, J.A.; Olson, S.D.; Nolta, J.A. Improved MSC Minimal Criteria to Maximize Patient Safety: A Call to Embrace Tissue Factor and Hemocompatibility Assessment of MSC Products. Stem Cells Transl Med 2022, 11, 2–13. [Google Scholar] [CrossRef]
- Baker, E.K.; Wallace, E.M.; Davis, P.G.; Malhotra, A.; Jacobs, S.E.; Hooper, S.B.; Lim, R. A Protocol for Cell Therapy Infusion in Neonates. Stem Cells Transl Med 2021, 10, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Herz, J.; Köster, C.; Reinboth, B.S.; Dzietko, M.; Hansen, W.; Sabir, H.; van Velthoven, C.; Bendix, I.; Felderhoff-Müser, U. Interaction between Hypothermia and Delayed Mesenchymal Stem Cell Therapy in Neonatal Hypoxic-Ischemic Brain Injury. Brain Behav Immun 2018, 70, 118–130. [Google Scholar] [CrossRef]
- Ophelders, D.R.M.G.; Wolfs, T.G.A.M.; Jellema, R.K.; Zwanenburg, A.; Andriessen, P.; Delhaas, T.; Ludwig, A.-K.; Radtke, S.; Peters, V.; Janssen, L.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Protect the Fetal Brain After Hypoxia-Ischemia. Stem Cells Transl Med 2016, 5, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Liu, D.; Li, T.; Ke, H.; Xin, D.; Wang, S.; Cao, Y.; Xue, H.; Wang, Z. Hydrogen Sulfide-Modified Extracellular Vesicles from Mesenchymal Stem Cells for Treatment of Hypoxic-Ischemic Brain Injury. J Control Release 2020, 328, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Xin, D.; Li, T.; Chu, X.; Ke, H.; Liu, D.; Wang, Z. MSCs-Extracellular Vesicles Attenuated Neuroinflammation, Synapse Damage and Microglial Phagocytosis after Hypoxia-Ischemia Injury by Preventing Osteopontin Expression. Pharmacol Res 2021, 164, 105322. [Google Scholar] [CrossRef] [PubMed]
- Pathipati, P.; Lecuyer, M.; Faustino, J.; Strivelli, J.; Phinney, D.G.; Vexler, Z.S. Mesenchymal Stem Cell (MSC)-Derived Extracellular Vesicles Protect from Neonatal Stroke by Interacting with Microglial Cells. Neurotherapeutics 2021, 18, 1939–1952. [Google Scholar] [CrossRef]
- Du, L.; Jiang, Y.; Sun, Y. Astrocyte-Derived Exosomes Carry microRNA-17-5p to Protect Neonatal Rats from Hypoxic-Ischemic Brain Damage via Inhibiting BNIP-2 Expression. NeuroToxicology 2021, 83, 28–39. [Google Scholar] [CrossRef]
- Lawson, A.; Snyder, W.; Peeples, E.S. Intranasal Administration of Extracellular Vesicles Mitigates Apoptosis in a Mouse Model of Neonatal Hypoxic-Ischemic Brain Injury. Neonatology 2022, 119, 345–353. [Google Scholar] [CrossRef]
- Luo, H.; Huang, F.; Huang, Z.; Huang, H.; Liu, C.; Feng, Y.; Qi, Z. microRNA-93 Packaged in Extracellular Vesicles from Mesenchymal Stem Cells Reduce Neonatal Hypoxic-Ischemic Brain Injury. Brain Res 2022, 1794, 148042. [Google Scholar] [CrossRef]
- Labusek, N.; Mouloud, Y.; Köster, C.; Diesterbeck, E.; Tertel, T.; Wiek, C.; Hanenberg, H.; Horn, P.A.; Felderhoff-Müser, U.; Bendix, I.; et al. Extracellular Vesicles from Immortalized Mesenchymal Stromal Cells Protect against Neonatal Hypoxic-Ischemic Brain Injury. Inflammation and Regeneration 2023, 43, 24. [Google Scholar] [CrossRef]
- Porzionato, A.; Zaramella, P.; Dedja, A.; Guidolin, D.; Van Wemmel, K.; Macchi, V.; Jurga, M.; Perilongo, G.; De Caro, R.; Baraldi, E.; et al. Intratracheal Administration of Clinical-Grade Mesenchymal Stem Cell-Derived Extracellular Vesicles Reduces Lung Injury in a Rat Model of Bronchopulmonary Dysplasia. Am J Physiol Lung Cell Mol Physiol 2019, 316, L6–L19. [Google Scholar] [CrossRef]
- Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Yeung, V.; Liu, X.; Ericsson, M.; Andrews, N.A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles Restore Lung Architecture and Improve Exercise Capacity in a Model of Neonatal Hyperoxia-Induced Lung Injury. J Extracell Vesicles 2020, 9, 1790874. [Google Scholar] [CrossRef]
- Abele, A.N.; Taglauer, E.S.; Almeda, M.; Wilson, N.; Abikoye, A.; Seedorf, G.J.; Mitsialis, S.A.; Kourembanas, S.; Abman, S.H. Antenatal Mesenchymal Stromal Cell Extracellular Vesicle Treatment Preserves Lung Development in a Model of Bronchopulmonary Dysplasia Due to Chorioamnionitis. Am J Physiol Lung Cell Mol Physiol 2022, 322, L179–L190. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, Y.; Xu, G.; Liu, L.; Li, H.; Li, Y.; Yin, J.; Wang, X.; Yu, Z. Human Breast Milk-Derived Exosomes through Inhibiting AT II Cell Apoptosis to Prevent Bronchopulmonary Dysplasia in Rat Lung. J Cell Mol Med 2022, 26, 4169–4182. [Google Scholar] [CrossRef]
- Zhong, X.-Q.; Wang, D.; Chen, S.; Zheng, J.; Hao, T.-F.; Li, X.-H.; Luo, L.-H.; Gu, J.; Lian, C.-Y.; Li, X.-S.; et al. Umbilical Cord Blood-Derived Exosomes from Healthy Term Pregnancies Protect against Hyperoxia-Induced Lung Injury in Mice. Clin Transl Sci 2023, 16, 966–977. [Google Scholar] [CrossRef]
- Barlow, B.; Santulli, T.V.; Heird, W.C.; Pitt, J.; Blanc, W.A.; Schullinger, J.N. An Experimental Study of Acute Neonatal Enterocolitis--the Importance of Breast Milk. J Pediatr Surg 1974, 9, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhang, R.; Liang, H.; An, J.; Yang, Y.; Huo, J.; Chen, Z.; Quan, W.; Jiang, L.; Li, C.; et al. Comparison and Investigation of Exosomes from Human Amniotic Fluid Stem Cells and Human Breast Milk in Alleviating Neonatal Necrotizing Enterocolitis. Stem Cell Rev Rep 2023, 19, 754–766. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sources of EVs and their common isolation methods for neonatal pathology therapy. Created with BioRender.com.
Figure 1.
Sources of EVs and their common isolation methods for neonatal pathology therapy. Created with BioRender.com.
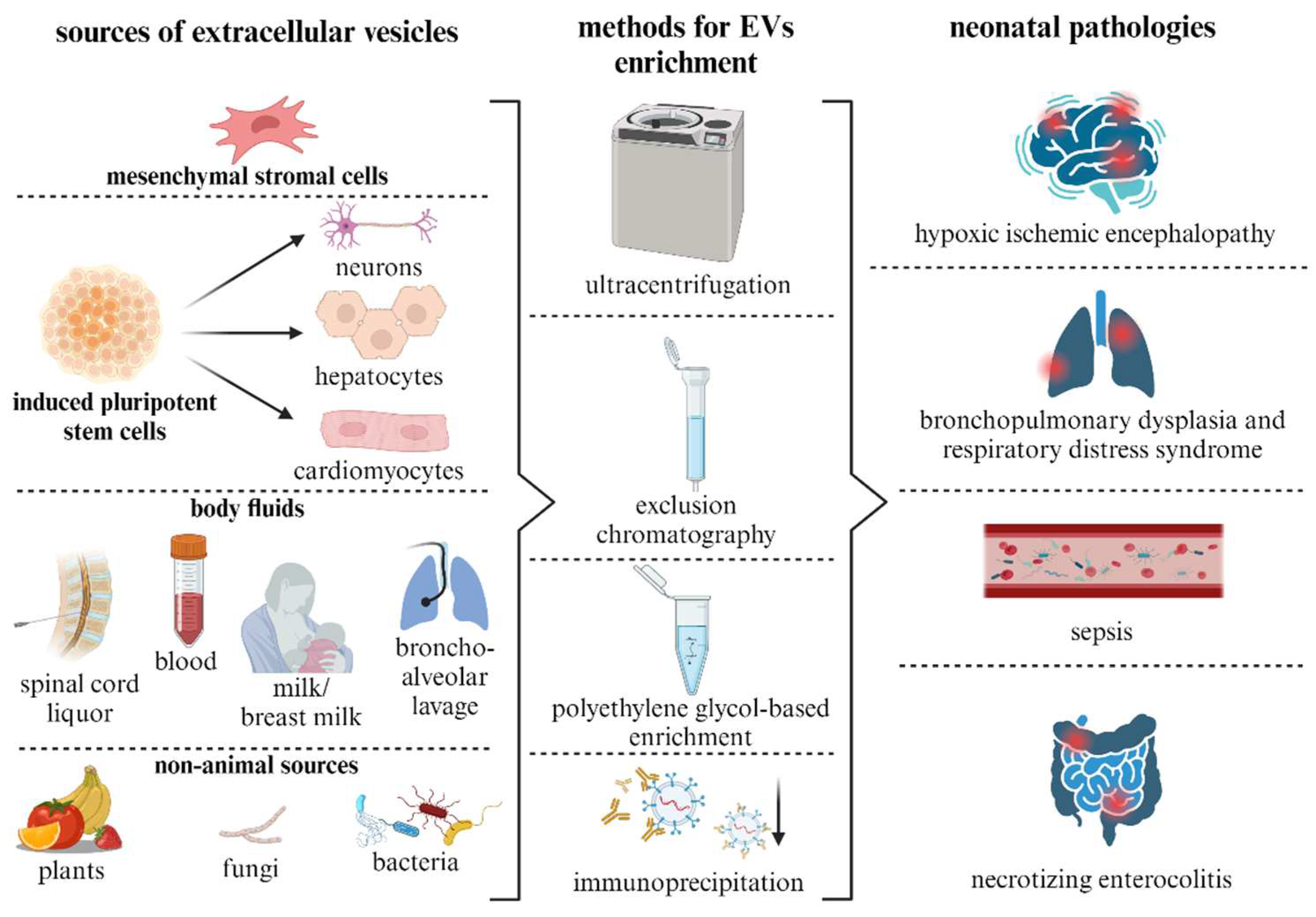
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



