
Submitted:
16 January 2024
Posted:
16 January 2024
You are already at the latest version
Abstract
The impacts of spice (sumac, cumin, black pepper, and red pepper) extract on Gram-positive (S. aureus and E. faecalis) and Gram-negative (K. pneumonia, P. aureginosa, C. jejuni, A. hydrophila, S. Paratyphi A, and Y. enterocolitica) food-borne pathogen bacterial strains (FBP) for their antibacterial activity and accumulation of ammonia (AMN), trimethylamine (TMA), and biogenic amines (BAs) in tyrosine decarboxylase broth (TDB) were investigated. Sumac extract exhibited the highest an-tibacterial potential against all FBP followed by cumin and peppers. Ammonia (AMN) production (844.24-543 mg/L) was strongly inhibited by sumac extract (55.10 mg/L), whilst suppression of trimethylamine (TMA) production (53.66-3.21 mg/L) was observed by sumac (1.28 mg/L), followed by red pepper (0.91 mg/L) extract. Cumin extracts stimulated the production of cadaverine, spermidine, tryptamine, and 2-phenylethylamine. Furthermore, sumac extracts inhibited hista-mine and tyramine production in the majority of FBP. This research suggests the application of sumac extracts as natural preservatives for inhibiting the growth of FBP and inhibiting the pro-duction of AMN, TMA, and BAs.
Keywords:
biogenic amine
; food pathogenic bacteria
; inhibition
; spice
; food safety
1. Introduction
Due to a rise in the cases of food poisoning-related mortality, there has been a growing global demand for safe food. The food-borne pathogens (FBP) are known to spread disease through infection, and they can also produce toxins that result in food poisoning. Gram-positive and Gram-negative bacteria are the primary cause of most illnesses and fatalities [1]. Most foodborne illness outbreaks that have been documented are linked to well-known organisms, including Salmonella, Campylobacter, Norovirus, Listeria monocytogenes, and Escherichia coli that produces Shiga toxin. Staphylococcus aureus, Clostridium species, Bacillus cereus, Yersinia enterocolitica, parasites, and other pathogens have also been shown to cause diseases on occasion [2]. Biogenic amine-related toxins have become a significant concern due to their potential to be poisonous and carcinogenic, as well as to trigger headaches, dizziness, and heart palpitations [3]. As a result of the activities of microbes during processes of decarboxylation, transamination, reducing amination, and compound degradation, poisonous nitrogenous chemicals called biological amines are produced [4]. Consequently, biogenic amines (BAs) are frequently used as a sign of the quality and safety of food.
Histamine and tyramine, the two main BAs found in foods, are among the most hazardous and extensively studied amines [4]. A strict monitoring system and regulatory limits have been established at various levels for histamine and tyramine BAs, taking into account the variations in foods and processing methods. It is known that the other BAs have a synergistic effect on raising the harmfulness of histamine and tyramine [2]. Therefore, different approaches to prevent or manage the concentration of BAs are required to enhance food safety and quality as well as human health [5]. Many practices have been assessed for lower BAs, including: using food additives or bioactive compounds (phenolic or terpenoids); using multiple starter cultures during the fermentation process; gamma irradiation; cold storage temperatures; high-hydrostatic pressure processing (HHP); food packaging procedures; and so on [6,7]. There has been a growing interest among consumers in clean label foods, which are the foods preserved using natural antimicrobials [8]. Due to their high antioxidative and antibacterial activity, spices are among the most commonly used natural antimicrobials for food preservation.
Spices are made from various plant parts, including roots, rhizomes, stem bark, leaves, fruits, flowers, and seeds [9,10]. Spices are frequently added to foods in order to improve their flavour and colour [11]. Spices tend to be subject to food fraud because they are most frequently offered in powdered form [12]. It has been reported that many unbranded spices readily available in the markets contain synthetic dyes; therefore, pose a health risk to humans [13]. As an alternative to their powdered form, extracts of spices have often been used to formulate foods. In this respect, various extraction solvents are used; however they have an impact on the bioactivity of the extracts. Spice extracts have gained a lot of attention due to their wide range of bioactivities and are generally recognized as safe (GRAS). However, there is currently little research on how spice extracts affect the synthesis of bacterial biogenic amines in various mediums. The presence of biogenic amines can be detected by using a variety of media. Using different types of media for detecting biogenic amines has its advantages and disadvantages. For example, agar plates provide a simple and cost-effective method, but they may have limited sensitivity. On the other hand, liquid media offer higher sensitivity, but they can be more time-consuming and require specialized equipment for analysis. In this respect, different broths have been used to count bacteria that produce amines to determine the presence of BAs. Furthermore, an accumulation of amines is found to be greater in Tyrosine decarboxylase broth (TDB) [14,15,16]. Therefore, the present study aims at assessing the effects of diethyl ether -extracted spice extract on the growth and generation of biogenic amines by Gram-positive (Staphylococcus aureus ATCC 29213 and Enterococcus faecalis ATCC 29212) and Gram-negative (Klebsiella pneumoniae ATCC 700603, Pseudomonas aeruginosa ATCC 27853, Campylobacter jejuni ATCC 33560, Aeromonas hydrophila NCIMB1135, Salmonella Paratyphi A NCTC13, and Yersinia enterocolitica NCTC 11175) food-borne bacteria.
2. Materials and Methods
2.1. Spices, Chemicals, and Cultural Media
A total of four different dried spices identified based on their botanical names were used in this study: sumac (Rhus coriaria L.), cumin (Cuminum cyminum L.), black pepper (Piper nigrum), and red pepper (Capsicum annuum). All these spices were acquired from a local market in Adana, Turkey. The spices were ground and dried. The compounds of diethyl ether, active carbon, tyrosine, peptone, Lab-Lemco powder, NaCl, pyridoxal–HCl, trimethylamine hydrochloride, ammonium chloride were obtained from Merck (Darmstadt, Germany) and Sigma-Aldrich (Seelze, Germany). Biogenic amines standards: e.g., histamine dihydrochloride, tyramine hydrochloride, tryptamine hydrochloride, putrescine dihydrochloride, 2-phenylethylamine hydrochloride, cadaverine dihydrochloride, spermidine trihydrochloride, spermine tetrahydrochloride, 5-hydroxytryptamine (serotonin), 3-hydroxytyramine hydrochloride (dopamine), agmatine sulphate, trichloroacetic acid, benzoyl chloride, acetonitrile NaOH, were acquired from Merck (Darmstadt, Germany) and Sigma-Aldrich (Seelze, Germany). All of them were of analytical reagent quality. Culture media; e.g., nutrient broth and plate count agar (PCA) were purchased from Merck (Darmstadt, Germany), and Biokar (Beauvais, France) Difco, respectively.
2.2. Bacterial Strains
Reference bacterial strains, e.g., Staphylococcus aureus (ATCC 29213), Klebsiella pneumoniae (ATCC 700603), Enterococcus faecalis (ATCC 29212), Pseudomonas aeruginosa (ATCC 27853), and Campylobacter jejuni (ATCC 33560) used in this study were obtained from the American Type Culture Collection (Rockville, MD, USA). Aeromonas hydrophila (NCIMB1135), Salmonella Paratyphi A (NCTC13), and Yersinia enterocolitica (NCTC 11175) were obtained from the National Collection of Industrial Food and Marine Bacteria (Aberdeen, UK) and the National Collection of Type Cultures (London, UK).
2.3. Spice Extraction
The solvent extraction technique was used to extract the spices. An extraction thimble (30 x 80 mm, Whatman 2810-338, UK) made from cellulose was used to combine 200 g of powdered spice with 1 L of diethyl ether and carried out in a reflux extractor. The mixture was then extracted for 4 hours at 60°C. Extraction process was carried out twice for each spice. To remove the color of the extracts, 40 g of activated carbon (Merck, Darmstadt, Germany) was used to bleach them for 30 minutes at 60°C after extraction. After the extracts had been filtered through Whatman No. 1 filter paper (Maidstone, UK), the impurities were eliminated from the extracts. A rotary evaporator (Heidolph WB 2000, Heidolph Instruments, Schwabach, Germany) was used to extract the organic solvent. Before further use, the dried extracts were stored at -20°C and protected from light. In order to carry out the antibacterial and biogenic amine analyses, the spice extracts were sterilized for 15 min. at room temperature (22°C) in a Telstar Bio IIA biological cabinet (Telstar, Madrid, Spain) using UV radiation (30 W, 253.7 nm wavelength, 50 cm away from the light source).
2.4. Culture Media and Biogenic Amines (BAs) Extraction
The method outlined by Klausen and Huss [17] was used to measure the synthesis of ammonia (AMN), trimethylamine (TMA), and BAs by reference to FBP staining, in tyrosine decarboxylase broth (TDB). Food-borne pathogens were cultured for two or three days at their ideal growth temperature in nutrient broth. After that, 0.5 mL of each bacterial culture was added to the TDB for tyrosine decarboxylation over the course of 72 hours, yielding 106 colony-forming units per mL (106 cfu/mL) as measured by the McFarland cell densitometer (Biosan DEN 1, Latvia). Spice extracts were added to the TDB at a rate of 1% (w/v), following bacterial inoculation. All extracts were tested in triplicate on the same day for all groups. As a part of the extraction process, five milliliters of TDB containing food-borne pathogens were divided into separate bottles and then added with two milliliters of trichloroacetic acid (6%, w/v) in order to extract biogenic amines. A filter paper with a pore size of 11 m (Schleicher and Schuell, Dassel, Germany) was then used to filter the extracts, then they were centrifuged for 10 minutes at 3000 g. A total of four milliliters of each bacterial supernatant was collected for the analysis, and the procedure was carried out in three duplicates.
2.5. Analysis of BA by HPLC after Derivatization
The method outlined by Özogul [18] was followed in order to prepare a standard amine-mixed aqueous solution containing ammonium chloride, trimethylamine hydrochloride and twelve amine. Derivatization of a 100 microliter standard amine solution containing 10 mg of each amine per microliter was accomplished by adding 40 mL of 2% (v/v) benzoyl chloride in acetonitrile and 1 mL of aqueous 2M NaOH solution. After shaking the solution for one minute in a vortex mixer, it was allowed to stand at room temperature and shielded from light for 20 min. Afterwards, The derivatization was stopped by adding 2 mL of saturated aqueous NaCl solution. The resultant solution was extracted twice with two milliliters of diethyl ether. The top layer was then separated, put into sterile sample tubes, dried with a nitrogen stream, and combined with one milliliter of acetonitrile. The BAs were separated and quantified by performing triplicate injections of 10 µL of the produced solution into Shimadzu HPLC equipment (Kyoto, Japan), following the HPLC approach previously described by Özogul [19]. The samples of extracted bacterial cultures were prepared in the same manner as those of the standard mixed amine solution, with the exception that 4 mL of each extracted bacterial culture was replaced with 100 mL of the standard mixed amine solution during the derivatization procedure.
The method outlined by Özogul [19] was used to determine the concentrations of BAs, TMA, and AMN. The results were expressed as milligrams of BAs (or TMA and ammonia) per litter of TDB (mg/L). There was an HPLC apparatus used in this study, which was a Shimadzu Prominence HPLC unit (Shimadzu, Kyoto, Japan), equipped with an HPLC ODS Hypersil column, 5 μm (250 × 4.6) mm (Phenomenex, Macclesfield, Cheshire, U.K.), an autosampler (SIL 20AC), a column oven (CTO-20AC), a communication bus module (CBM-20A) featuring a valve unit FCV-11AL, and two binary gradient pumps (Shimadzu LC-10AT).
2.6. Chromatographic Separation
In order to conduct the chromatographic separation, gradient elutions were performed using acetonitrile (eluant A) and HPLC grade water (eluant B) at a flow rate of 1.2 mL/min. The injection volume was 10 µL, and the overall separation time was less than 20 minutes. Detection was monitored at 254 nm. Standard curves were created for each amine ranging from 0 to 50 mg/mL. A correlation coefficient of peak area versus amine standard concentrations was computed for each compound following the injection of five duplicates of each standard solution of amine. The curves for each benzoylated amine showed a correlation coefficient (r) greater than 0.99.
2.7. Determination of Different Bacterial Growths in Tyrosine Decarboxylase Broth (TDB)
After appropriate dilutions (10–10 CFU/mL) were made of each bacterial culture in the TDB, 0.1 mL was inoculated in triplicate onto plate count agar (PCA, Merck, Darmstadt, Germany) plates using a spread plate approach. Following 72 hours of incubation at 30°C, the results were obtained as the logarithm of total viable colony-forming units per milliliter of broth, log (average standard deviation), and log (CFU/mL).
2.8. Statistical Analysis
The results were calculated using triplicate samples for each spice (per treatment). An analysis of variance (ANOVA) was performed and Duncan's multiple range tests were run on the data when there were significant differences at P<0.05. Statistical differences between the control and spice extracts were determined based on pathogen concentrations and BA contents. All statistical analyses were conducted using SPSS version 19 for Windows (SPSS Inc.; Chicago, IL, USA).
3. Results and Discussions
3.1. Bacterial Growth in Tyrosine Decarboxylase Broth
The results of different food-borne pathogen bacterial growths in TDB are shown in Figure 1. Due to sumac's higher antimicrobial activity, sumac extract demonstrated significant inhibition of both Gram-positive (S. aureus and E. faecalis) and Gram-negative (K. pneumoniae, P. aeruginosa, C. jejuni, A. hydrophila, S. Paratyphi A, and Y. enterocolitica) bacteria. The highest inhibition levels were observed for Y. enterocolitica, ranging from 8.61 to 5.05 log (CFU/ml). P. aureginosa was inhibited at similar levels by cumin extract and sumac extract. Cumin, red pepper, and black pepper extracts also inhibited K. pneumoniae and E. faecalis at similar levels. For all microorganisms tested, cumin, black pepper, and red pepper spice extracts inhibited bacteria below 1.0 log (CFU/ml). Sumac extract was shown to be effective against several Gram-positive and Gram-negative bacteria in previous studies [20,21]. Presence of several polyphenolic compounds in sumac were linked to antibacterial activity [22]. On the other hand, a previous study reported that cumin extract exhibited lower bactericidal activities [23]. Impacts of drying technique and extraction solvents on antibacterial activity were earlier discussed, highlighting the role of different drying techniques (degrading the bioactive compound) and solvents (poor solubility of the bioactive compound) on the extraction of bioactive compounds responsible for activity [24,25].
3.2. Ammonia, Trimethylamine and BAs Production in Tyrosine Decarboxylase Broth
There are three types of BAs that are present in food. Heterocyclic BAs (histamine and tryptamine), aliphatic BAs (putrescine and cadaverine), and aromatic BAs (tyramine and phenylethylamine). A further categorization is based on the quantity of amine groups, which include polyamines (spermidine and spermine), diamines (histamine, putrescine, and cadaverine), and monoamines (tyramine and phenylethylamine) [26]. BAs such as diamines, polyamines, and TMA are detected to monitor the freshness or spoilage rate of food. The most dangerous amines are histamine and tyramine, which are the two primary BAs present in food [4]. Inhibitory effects of four spice extracts, e.g., sumac, black pepper, red pepper, and cumin on the production of ammonia (AMN), trimethylamine (TMA) and the formed BAs (putrescine, cadaverine, spermidine, tryptamine, phenylethylamine, spermine, serotonin, dopamine, and agmatine) produced by eight food-borne bacteria using TDB are presented in Table 1. AMN production was between 543 mg/L by A. hydrophila and 844 mg/L by K. pneumoniae. A significant inhibition of ammonia production was observed with all spice extracts (>75 %), particularly sumac, which inhibited five microbial strains, e.g., S. aureus (90%), S. Paratyphi A (91%), K. pneumoniae (80%), E. faecalis (87%), and Y. enterocolitica (92%). There was an 80% inhibition of P. aeruginosa and a 75% inhibition of C. jejuni by black pepper whereas 89% of inhibition of A. hydrophila by cumin was observed. The maximum production of cadaverine and putrescin was recorded by S. Paratyphi A (4.39 mg/L) and C. jejuni (35.49 mg/L). The control sample (without spice extract) had a generally high level of BAs formation (except for tryptamine and phenylethylamine). Extracts of cumin exhibited higher values than control samples. A similar pattern was observed with putrescine, where sumac extract inhibited five species (>50%) other than K. pneumoniae (50%), whereas black pepper extract inhibited stronger inhibition for A. hydrophila (47%) and Y. enterocolitica (90%). The production of cadaverine in all evaluated microorganisms was most resistant to spice extracts, with the exception of S. Paratyphi A, for which an inhibition of 60% was observed in the presence of sumac extract. However, among extracts tested, there was a higher increase in cadaverine production with cumin extract over control. Sumac extract was the most effective inhibitor of the four spice extracts evaluated, followed by black pepper and red pepper extracts. Spermidine production peaked at 90.22 mg/L, primarily generated by E. faecalis. Black pepper extract was found to be the most effective against spermidine production, inhibiting over 70% of all evaluated microorganisms. Similarly, sumac inhibited spermine production in all samples. In comparison to other extracts, sumac extract were found to promote serotonin generation in bacteria (S. aureus, S. Paratyphi A, K. pneumoniae, E. faecalis, and P. aeruginosa). Additionally, cumin extract promoted serotonin production for all three remaining microorganisms. The results showed that red pepper extract was effective on inhibition the production of trimethylamine by A. hydrophila, while black pepper inhibited the formation of trimethylamine by P. aeruginosa, E. faecalis, and S. Paratyphi A. Among all tested microorganisms, pepper-based extracts significantly retarded dopamine production. Sumac extract inhibited the production of agmatine by all tested microorganisms with the exception of S. aureus.
Microorganisms secrete endogenous enzymes (amino acid decarboxylase) and exogenous enzymes for decarboxylation of proteins and amino acids [4]. Cumin extract intensifies the production of tryptamine, phenylethylamine, and spermidine BAs due to its synergistic effect with TDB broth in decarboxylating phenylalanine and tryptophan, thereby producing phenylethylamine and tryptamine. This is the first study to suggest that spice extract increases BAs production, which could be explained by the abundance of alkaloid in cumin, because alkaloid content has been associated with increasing BAs production. [4,27,28,29]. The sumac extract was the most effective inhibitor, followed by the black pepper, red pepper, and the cumin extracts. In this study, sumac extract was observed to suppress BAS production, which could be ascribed to the fact that sumac extract contained 211 different kinds of phytochemicals, such as polyphenols, organic acids (mallic and tannic acid), and flavonoids [30,31,32,33,34]. Peppers (black and red) were reported to be the sources of bioactive capsicin, piperin, flavonoid, amide, and organic acid constituents, confirming their ability to inhibit biogenic amine production. Several studies show that the bioactive components in spice extracts are antibacterial, inhibiting the actions of endogenous enzymes, and also targeting Gram-positive and Gram-negative bacteria [8,32,35,36,37,38,39]. However, the antibacterial action, however, led to the inactivation of microorganisms that triggered BA production. In addition, bioactive constituents present in spices were reported to inhibit enzymatic activities due to their high antioxidant potential, which is mainly responsible for decarboxylation of amino acids [40,41,42].
3.2.1. Histamine Production by Food-Borne Pathogen Bacteria in Tyrosine Decarboxylase Broth
Histamine is regarded as BAs that is dangerous when taken into the human body at high proportions-(100 mg/kg) [32]. Histamine is usually produced from histidine converted by microorganisms or their enzymes [43]. Histamine is generally used for the estimation of quality and freshness indexes for meat-based foods [3]. Among all of the microorganisms evaluated, sumac extract had the strongest inhibitory effect (< 10 mg/L) on histamine production (Figure 2). On the other hand, Y. enterocolitica was completely inhibited. This was followed by red pepper, which exhibited significant inhibition (< 10 mg/L) for S. Paratyphi A, K. pneumoniae, Y. enterocolitica, P. aeruginosa, and A. hydrophila. As for the rest of the samples, production was above 10 mg/L but below the maximum allowable level. On the other hand, black and red pepper extracts were observed to promote the production of histamine by S. aureus, S. Paratyphi A, and Y. enterocolitica. Cumin extracts increased production by all FBP except for E. faecalis and A. hydrophila. Therefore, we can speculate that spice extracts can be used against the production of histidine due to their ability to inhibit bacterial growth, arrest biogenic amine synthesis, and inhibit amino acid decarboxylation, specifically enzyme (histidine decarboxylase) activity [44,45].
Similar results were demonstrated by Shakila, Vasundhara [46] who detected the efficacy of spice extracts (cinnamon, clove, turmeric, and cardamom) on in vitro histamine production by Morganella morganii. Based on the proposed inhibition of histamine decarboxylation activity exhibited by spices, the aforementioned results were obtained. Some extracts, however, were also found to promote amine production due to their lower activity in inhibiting histamine decarboxylation, corresponding to a delay in amine production [42].
3.2.2. Tyramine Production by Food-Borne Pathogen Bacteria in Tyrosine Decarboxylase Broth
The tyramine production profile of microorganisms in TBD is shown in Figure 3. The sumac extract significantly increased the formation of tyramine by S. aureus, S. Paratyphi A, and P. aeruginosa. The black and red peppers were found to significantly inhibit tyramine production by S. aureus, S. Paratyphi A, E. faecalis, Y. enterocolitica, and A. hydrophila. Also, cumin extracts were observed to promote the production of tyramine at levels higher than those produced in the control sample, with the exception of the strain C. jejuni, A. hydrophila, and S. aureus. Tyrosine was observed to be produced by the tyrosine amino acid by the action of a microbial enzyme [3]. Tyramine is associated with several disorders in humans, and in some cases its toxicity was reported to be higher as compared to histamine [47]. There have been similar results regarding the impacts of spice extracts on BAs (tyramine) production in minced meat [48]. It was concluded that spice extract inhibited microorganism growth and amino acid decarboxylase activity, lowering the production of amine. A recent molecular docking study has shown the ability of spices to bind with the amino acid decarboxylase active site and inhibit the enzyme, resulting in less BAs production and accumulation [49].
5. Conclusions
Sumac extract exhibited significant inhibition of Gram-positive and Gram-negative bacteria, while cumin, black pepper, and red pepper spice extracts had lower bactericidal activities. This study also demonstrated the inhibitory effects of four spice extracts (sumac, black pepper, red pepper, and cumin) on the production of ammonia and BAs (histamine, tyramine, putrescine, cadaverine, spermidine, tryptamine, phenylethylamine, spermine, trimethylamine, serotonin, dopamine, and agmatine) by eight food-borne pathogen bacteria using tyrosine decarboxylase broth. Results showed that sumac was the most effective inhibitor, followed by black pepper, red pepper, and cumin extracts. All evaluated microorganisms produced less histamine when sumac extract was used, while cumin extract induced histamine production. In order to ensure food safety, sumac extract is recommended as a food preservative for controlling biogenic amine production. Further research should be conducted on the various methods of extracting these materials, particularly sumac extract. A variety of foods, their antioxidant and antibacterial properties, and their safety aspects should be discussed in addition to their integrated or combined use with other technologies. Integrating sumac extract with other technologies has the potential to enhance its effectiveness and expand its applications. By combining it with innovative delivery systems or processing techniques, we can unlock new possibilities for preserving food, improving health, and combating bacterial infections. This integration could lead to synergistic effects and create unique solutions in various fields such as food science, medicine, and environmental sustainability.
Author Contributions
F.K., investigation, methodology, software (microbiological and statistical analyses); E.K., software (microbiological and chemical analysis), review & editing; N.B.R., original draft preparation, writing; M.T.Y., original draft preparation, writing, manuscript submission and editing; F.Ö., Project administration, conceptualization, review & editing.
Funding
The project was funded by the Scientific Research Projects Unit of Cukurova University (Project No: FYL-2015-4813). The authors are grateful for their financial support.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ramachandran, G. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence 2014, 5, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Gourama, H. Foodborne pathogens. Food Safety Engineering 2020, 25–49. [Google Scholar]
- Özogul, Y.; Özogul, F. Biogenic amines formation, toxicity, regulations in food. Biogenic amines in food: Analysis, occurrence and toxicity 2019, 1–17. [Google Scholar]
- Gardini, F.; et al. Technological factors affecting biogenic amine content in foods: A review. Frontiers in microbiology 2016, 7, 1218. [Google Scholar] [CrossRef] [PubMed]
- Tsafack, P.B.; Tsopmo, A. Effects of bioactive molecules on the concentration of biogenic amines in foods and biological systems. Heliyon 2022, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.R.; Anwar, M.M.; Sallam, E.M.; Emam, W.H. Quality and safety of irradiated food regarding biogenic amines: Ras cheese. International Journal of Food Science & Technology 2016, 51, 1048–1054. [Google Scholar]
- Jaguey-Hernandez, Y.; Aguilar-Arteaga, K.; Ojeda-Ramirez, D.; Anorve-Morga, J.; González-Olivares, L.G.; Castaneda-Ovando, A. Biogenic amines levels in food processing: Efforts for their control in foodstuffs. Food Research International 2021, 144, 110341. [Google Scholar] [CrossRef]
- Rathod, N.B.; et al. Recent developments of natural antimicrobials and antioxidants on fish and fishery food products. Comprehensive Reviews in Food Science and Food Safety 2021, 20, 4182–4210. [Google Scholar] [CrossRef]
- El-Sayed, S.M.; Youssef, A.M. Potential application of herbs and spices and their effects in functional dairy products. Heliyon 2019, 5, 1–7. [Google Scholar] [CrossRef]
- Batiha, G.E.S.; Ogunyemi, O.M.; Shaheen, H.M.; Kutu, F.R.; Olaiya, C.O.; Sabatier, J.M.; De Waard, M. Rhus coriaria L.(Sumac), a versatile and resourceful food spice with cornucopia of polyphenols. Molecules 2022, 27, 5179. [Google Scholar] [PubMed]
- Pervez, M.K.; Ahmed, F.; Dewani, R.; Ayaz, T.; Mehboob, S.J.; Soomro, S.A. Qualitative investigation of prohibited food colors in red hot chilli & curry collected from Karachi City. Pakistan Journal of Pharmacology 2017, 34, 17–22. [Google Scholar]
- Modupalli, N.; Naik, M.; Sunil, C.K.; Natarajan, V. Emerging non-destructive methods for quality and safety monitoring of spices. Trends in Food Science & Technology 2021, 108, 133–147. [Google Scholar]
- Ullah, A.; Chan, M.W.H.; Aslam, S.; Khan, A.; Abbas, Q.; Ali, S.; et al. Banned Sudan dyes in spices available at markets in Karachi, Pakistan. Food Additives & Contaminants: Part B 2023, 16, 69–76. [Google Scholar]
- Daud, N.M.; et al. Valorisation of plant seed as natural bioactive compounds by various extraction methods: A review. Trends in Food Science & Technology 2022, 119, 201–214. [Google Scholar]
- Kuley, E.; Özogul, F. Synergistic and antagonistic effect of lactic acid bacteria on tyramine production by food-borne pathogenic bacteria in tyrosine decarboxylase broth. Food chemistry 2011, 127, 1163–1168. [Google Scholar] [CrossRef]
- Yazgan, H.; et al. The antimicrobial properties and biogenic amine production of lactic acid bacteria isolated from various fermented food products. Journal of Food Processing and Preservation 2021, 45, e15085. [Google Scholar] [CrossRef]
- Klausen, N.K.; Huss, H.H. A rapid method for detection of histamine-producing bacteria. International Journal of Food Microbiology 1987, 5, 137–146. [Google Scholar] [CrossRef]
- Özogul, F. Effects of specific lactic acid bacteria species on biogenic amine production by foodborne pathogen. International journal of food science & technology 2011, 46, 478–484. [Google Scholar]
- Özoğul, F. Production of biogenic amines by Morganella morganii, Klebsiella pneumoniae and Hafnia alvei using a rapid HPLC method. European Food Research and Technology 2004, 219, 465–469. [Google Scholar] [CrossRef]
- Fazeli, M.R.; et al. Antimicrobial activities of Iranian sumac and avishan-e shirazi (Zataria multiflora) against some food-borne bacteria. Food control 2007, 18, 646–649. [Google Scholar] [CrossRef]
- Nasar-Abbas, S.; Halkman, A.K. Antimicrobial effect of water extract of sumac (Rhus coriaria L.) on the growth of some food borne bacteria including pathogens. International journal of food microbiology 2004, 97, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Rayne, S.; Mazza, G. Biological activities of extracts from sumac (Rhus spp.): A review. Nature precedings 2017, 1–1. [Google Scholar]
- Mostafa, A.A.; et al. Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi journal of biological sciences 2018, 25, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Mohammad Salamatullah, A.; et al. Effects of different solvents extractions on total polyphenol content, HPLC analysis, antioxidant capacity, and antimicrobial properties of peppers (red, yellow, and green (Capsicum annum L.). Evidence-Based Complementary and Alternative Medicine 2022, 2022.
- Pundir, R.K.; Jain, P. Comparative studies on the antimicrobial activity of black pepper (Piper nigrum) and turmeric (Curcuma longa) extracts. International Journal of Applied Biology and Pharmaceutical Technology 2010, 1, 492–500. [Google Scholar]
- Tırıs, G.; Yanıkoğlu, R.S.; Ceylan, B.; Egeli, D.; Tekkeli, E.K.; Önal, A. A review of the currently developed analytical methods for the determination of biogenic amines in food products. Food Chemistry 2023, 398, 133919. [Google Scholar] [CrossRef]
- Mallik, S.; Sharangi, A.; Sarkar, T. Phytochemicals of coriander, cumin, fenugreek, fennel and black cumin: A preliminary study. National Academy Science Letters 2020, 43, 477–480. [Google Scholar] [CrossRef]
- Merah, O.; et al. Biochemical composition of cumin seeds, and biorefining study. Biomolecules 2020, 10, 1054. [Google Scholar] [CrossRef]
- Sharma, A.; et al. Genetic engineering approach using early Vinca alkaloid biosynthesis genes led to increased tryptamine and terpenoid indole alkaloids biosynthesis in differentiating cultures of Catharanthus roseus. Protoplasma 2018, 255, 425–435. [Google Scholar]
- Abu-Reidah, I.M.; et al. HPLC–DAD–ESI-MS/MS screening of bioactive components from Rhus coriaria L.(Sumac) fruits. Food chemistry 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Burgut, A.; et al. Suppression effects of aqueous and ethanolic extracts of propolis on biogenic amine production by Morganella psychrotolerans. LWT 2020, 131, 109771. [Google Scholar] [CrossRef]
- Houicher, A.; et al. Control of biogenic amine production and bacterial growth in fish and seafood products using phytochemicals as biopreservatives: A review. Food Bioscience 2021, 39, 100807. [Google Scholar] [CrossRef]
- Rathod, N.B.; et al. Recent developments in polyphenol applications on human health: A review with current knowledge. Plants 2023, 12, 1217. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, F. Chemical composition and biological activity of staghorn sumac (Rhus typhina). Food Chemistry 2017, 237, 431–443. [Google Scholar] [CrossRef]
- Ghorbani, P.; et al. Sumac silver novel biodegradable nano composite for bio-medical application: Antibacterial activity. Molecules 2015, 20, 12946–12958. [Google Scholar] [CrossRef] [PubMed]
- Harich, M.; et al. Evaluation of antibacterial activity of two natural bio-preservatives formulations on freshness and sensory quality of ready to eat (RTE) foods. Food Control 2018, 85, 29–41. [Google Scholar] [CrossRef]
- Karsha, P.V.; Lakshmi, O.B. Antibacterial activity of black pepper (Piper nigrum Linn.) with special reference to its mode of action on bacteria. 2010.
- Milenković, A.N.; Stanojević, L.P. Black pepper: Chemical composition and biological activities. Advanced Technologies 2021, 10, 40–50. [Google Scholar] [CrossRef]
- Zou, L.; Hu, Y.-Y.; Chen, W.-X. Antibacterial mechanism and activities of black pepper chloroform extract. Journal of Food Science and Technology 2015, 52, 8196–8203. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; et al. Effects of spices on the formation of biogenic amines during the fermentation of dry fermented mutton sausage. Food Chemistry 2020, 321, 126723. [Google Scholar] [CrossRef] [PubMed]
- Trifan, A.; et al. Phytochemical characterization and evaluation of the antioxidant and anti-enzymatic activity of five common spices: Focus on their essential oils and spent material extractives. Plants 2021, 10, 2692. [Google Scholar] [CrossRef] [PubMed]
- Wendakoon, C.N.; Sakaguchi, M. Inhibition of amino acid decarboxylase activity of Enterobacter aerogenes by active components in spices. Journal of food protection 1995, 58, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Oktariani, A.F.; Ramona, Y.; Sudaryatma, P.E.; Dewi, I.A.M.M.; Shetty, K. Role of marine bacterial contaminants in histamine formation in seafood products: A review. Microorganisms 2022, 10, 1197. [Google Scholar] [CrossRef] [PubMed]
- Jeon, A.R.; Lee, J.H.; Mah, J.-H. Biogenic amine formation and bacterial contribution in Cheonggukjang, a Korean traditional fermented soybean food. LWT 2018, 92, 282–289. [Google Scholar] [CrossRef]
- Kim, D.H.; et al. Inhibitory effects of brown algae extracts on histamine production in mackerel muscle via inhibition of growth and histidine decarboxylase activity of Morganella morganii. Journal of Microbiology and Biotechnology 2014, 24, 465–474. [Google Scholar] [CrossRef]
- Shakila, R.J.; Vasundhara, T.; Vijaya Rao, D. Inhibitory effect of spices on in vitro histamine production and histidine decarboxylase activity of Morganella morganii and on the biogenic amine formation in mackerel stored at 30 C. Zeitschrift für Lebensmittel-Untersuchung und Forschung 1996, 203, 71–76. [Google Scholar] [CrossRef]
- Linares, D.M.; et al. Comparative analysis of the in vitro cytotoxicity of the dietary biogenic amines tyramine and histamine. Food chemistry 2016, 197, 658–663. [Google Scholar] [CrossRef]
- Mahmoud, A.F.A.; et al. Reduction of biogenic amines production in chilled minced meat using antimicrobial seasonings. Journal of microbiology, biotechnology and food sciences 2021, 10, e3663–e3663. [Google Scholar] [CrossRef]
- Lin, Z.; Wu, Z.-y.; Zhang, W.-x. Bioinformatics analysis of amino acid decarboxylases related to four major biogenic amines in pickles. Food Chemistry 2022, 393, 133339. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Food-borne pathogen growth in tyrosine decarboxylase broth. a–c indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
Figure 1.
Food-borne pathogen growth in tyrosine decarboxylase broth. a–c indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
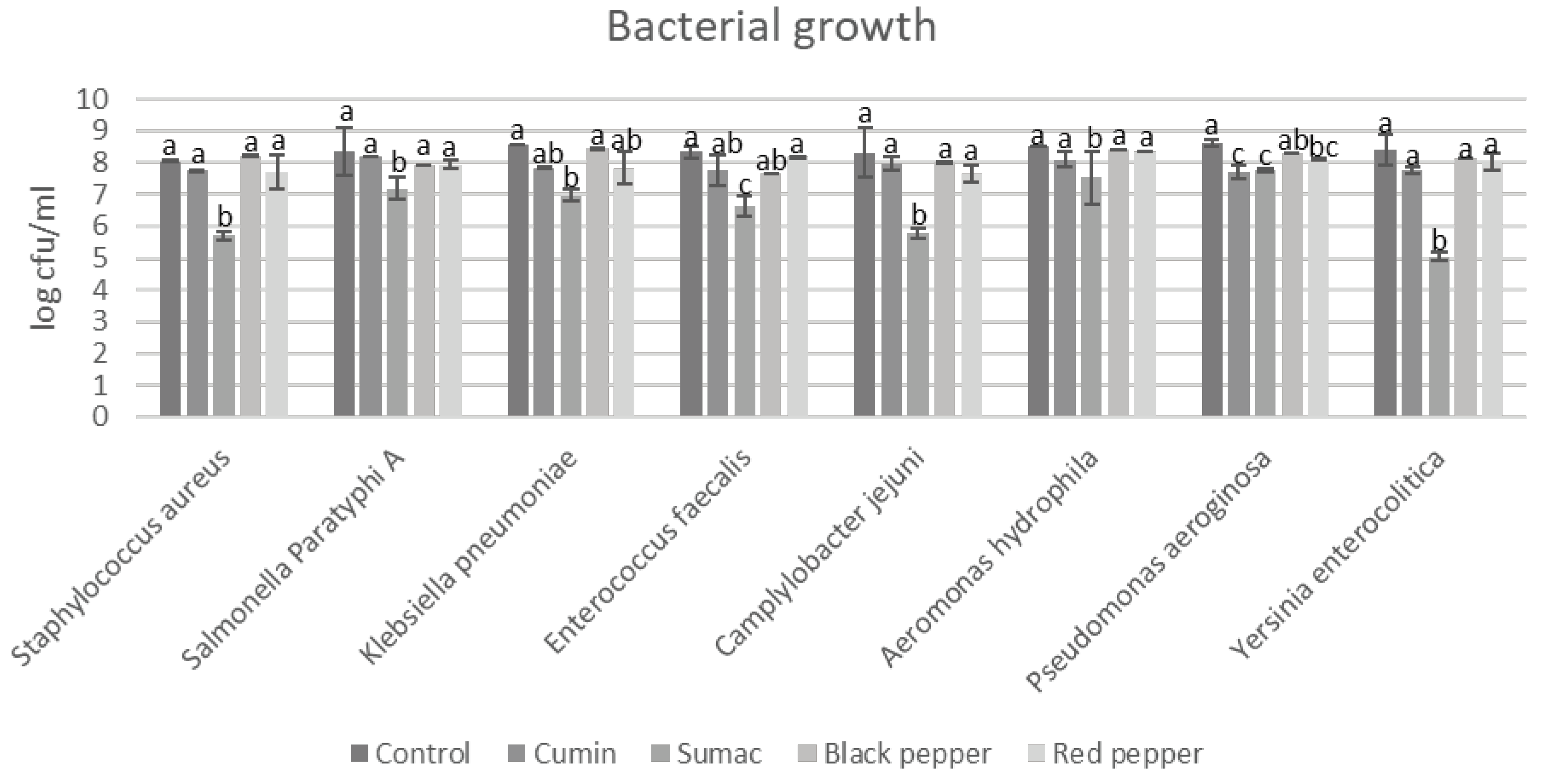
Figure 2.
Histamine production by food-borne pathogen bacteria in the presence of spice extracts in tyrosine decarboxylase broth. a–e indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
Figure 2.
Histamine production by food-borne pathogen bacteria in the presence of spice extracts in tyrosine decarboxylase broth. a–e indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
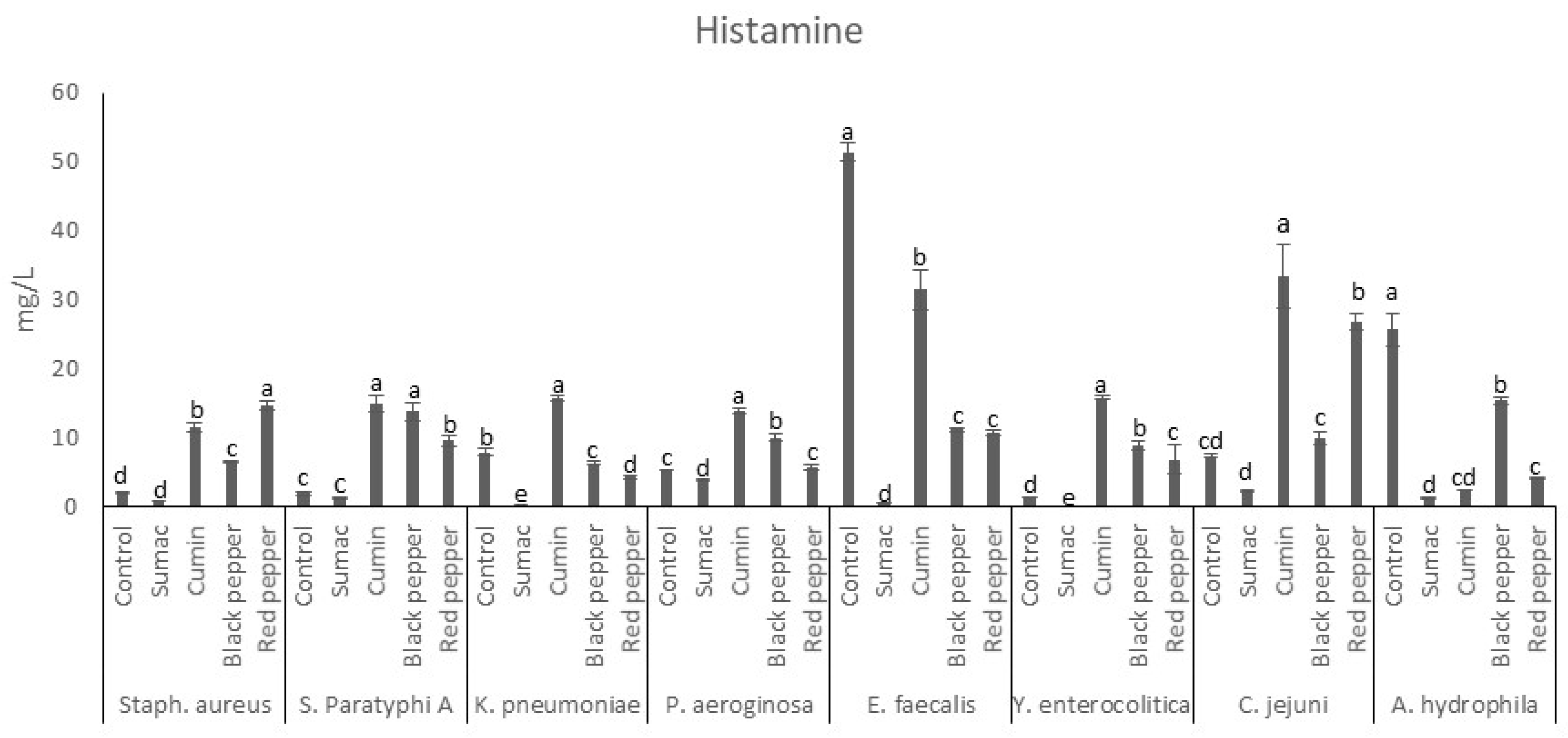
Figure 3.
Tyramine production by food-borne pathogen bacteria in the presence of spice extracts in tyrosine decarboxylase broth. a–e indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
Figure 3.
Tyramine production by food-borne pathogen bacteria in the presence of spice extracts in tyrosine decarboxylase broth. a–e indicate significant differences (P < 0.05) among groups. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).

Table 1.
Ammonia and biogenic amine production by food-borne pathogens spice extracts in tyrosine decarboxylase broth (mg/L).
Table 1.
Ammonia and biogenic amine production by food-borne pathogens spice extracts in tyrosine decarboxylase broth (mg/L).
| AMN | PUT | CAD | SPD | TRP | PHEN | SPM | SER | TMA | DOP | AGM | Group | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Staphylococcus aureus (ATCC29213) |
808.05±68.56a 84.60±5.66c 323.35±17.46b 344.10±34.97b 419.86±30.89b |
15.85±0.54b 5.64±0.22c 81.09±4.21a 15.77±1.13b 8.34±0.48c |
3.42±0.21cd 2.01±0.32d 27.99±2.27a 16.87±0.97b 6.22±0.40c |
16.70±1.36b 2.10±0.20c 90.42±8.11a 1.01±0.01c 15.25±1.27b |
0.28±0.01c 0.00±0.00c 17.80±0.27a 0.28±0.03c 5.80±0.18b |
0.25±0.01b 0.00±0.00b 12.77±0.51a 0.00±0.00b 0.41±0.01b |
16.16±0.37c 5.50±0.13d 42.13±1.91b 9.32±0.07d 69.93±3.29a |
7.69±0.25d 95.31±1.66a 34.03±2.62b 14.27±1.37c 8.99±0.60d |
3.21±0.03d 3.96±0.21d 22.34±0.09b 12.79±0.34c 30.01±2.15a |
95.24±1.63c 190.39±13.94b 503.00±0.34a 494.54±16.05a 24.37±1.38d |
39.18±0.27c 42.49±3.77c 71.83±1.28b 111.65±8.84a 47.11±1.62c |
C SUM CUM BP RP |
|
Salmonella Paratyphi A (NCTC13) |
836.47±62.76a 71.51±5.94c 286.12±5.54b 78.34±0.79c 287.30±18.56b |
21.49±1.04b 1.44±0.02c 57.39±4.55a 5.89±0.39c 4.23±0.07c |
4.39±0.40c 1.73±0.04d 27.41±1.19a 3.07±0.13cd 7.86±0.42b |
22.24±2.08b 5.34±0.48c 60.55±2.34a 0.00±0.00d 21.25±2.47b |
0.00±0.00b 0.00±0.00b 16.54±0.67a 0.16±0.02b 0.77±0.01b |
0.00±0.00b 0.00±0.00b 12.81±0.85a 0.00±0.00b 0.97±0.10b |
17.81±1.53c 2.96±0.16e 58.75±0.63b 7.28±0.74d 112.37±2.42a |
31.40±2.52b 82.42±2.93a 23.96±0.51c 8.56±0.35d 19.71±1.09c |
18.65±0.92a 4.24±0.09c 17.17±1.29a 3.29±0.16c 14.36±1.09b |
468.63±41.47a 355.11±30.40b 482.23±37.95a 45.24±4.30c 322.84±21.54b |
70.90±5.82a 26.40±1.56c 80.87±6.91a 32.90±1.59bc 43.26±1.82b |
C SUM CUM BP RP |
|
Klebsiella pneumoniae (ATCC700603) |
699.88±45.21a 133.15±10.39c 259.56±24.37b 181.69±7.46c 267.97±18.00b |
26.36±1.53a 16.88±1.31c 21.25±0.75b 13.18±1.03d 24.88±0.96a |
3.03±0.04c 3.10±0.13c 32.05±2.53a 8.08±0.14b 9.14±0.01b |
16.66±0.93b 12.04±0.83b 55.52±4.94a 4.67±0.23c 16.63±0.41b |
0.00±0.00b 0.00±0.00b 16.32±0.29a 0.30±0.01b 0.39±0.55b |
0.00±0.00b 0.00±0.00b 14.56±0.92a 0.00±0.00b 0.66±0.08b |
31.99±2.78a 2.39±0.02c 30.87±0.79a 11.13±0.46b 31.02±0.18a |
57.20±1.80b 65.51±3.73a 44.31±2.82c 6.27±0.60d 6.48±0.10d |
5.40±0.26b 1.70±0.04c 35.04±1.48a 5.06±0.21b 0.91±0.02c |
815.25±53.59a 434.63±38.38b 457.67±40.57b 97.64±4.07c 160.19±5.44c |
74.48±3.94b 18.67±1.33c 93.58±6.11a 25.20±1.11c 22.55±1.58c |
C SUM CUM BP RP |
|
Pseudomonas aeruginosa (ATCC27853) |
844.24±46.07a 240.06±8.52c 328.78±9.13b 166.48±6.82d 193.23±18.26d |
2.20±0.18c 1.07±0.02c 60.45±7.54a 16.40±1.12b 3.30±0.33c |
3.42±0.33c 3.26±0.26c 59.49±0.92a 3.44±0.05c 11.09±0.52b |
46.77±2.93b 2.88±0.17d 104.70±8.03a 0.77±0.10d 35.86±1.68c |
0.00±0.00c 0.00±0.00c 21.56±1.46a 0.00±0.00c 4.05±0.01b |
0.28±0.03b 0.00±0.00b 25.06±1.62a 0.00±0.00b 0.36±0.05b |
21.51±0.47c 11.87±0.39cd 67.97±3.48b 7.69±0.15d 109.52±10.36a |
16.91±0.69c 51.04±1.59a 46.76±2.20b 4.52±0.19d 14.83±0.39c |
20.83±1.54b 4.34±0.02c 48.89±0.29a 1.34±0.08d 2.01±0.07d |
668.71±55.08a 453.12±41.40b 357.57±12.44c 118.81±6.11d 175.72±4.57d |
64.32±4.17b 24.99±0.05d 92.37±4.40a 33.98±2.08c 36.93±0.56c |
C SUM CUM BP RP |
|
Enterococcus faecalis (ATCC29212) |
689.23±56.42a 87.75±1.91d 290.61±7.15bc 329.76±4.75b 234.01±14.94c |
4.88±0.45c 1.46±0.05c 23.78±1.44b 26.72±2.27b 33.09±1.69a |
3.50±0.16c 2.24±0.09c 50.75±3.56a 9.12±0.11b 3.74±0.42c |
90.22±4.82a 4.43±0.04cd 0.00±0.00d 8.14±0.61c 27.99±0.94b |
0.95±0.07b 0.88±0.04b 20.35±2.08a 0.24±0.00b 0.37±0.05b |
0.35±0.01b 0.00±0.00b 15.75±0.52a 0.51±0.01b 0.00±0.00b |
7.50±0.09c 2.97±0.04d 57.36±4.62a 7.90±0.50c 41.42±0.93b |
20.98±0.94c 75.87±2.93a 25.57±0.80b 13.65±1.38d 9.92±0.71d |
36.16±2.58b 5.56±0.28d 49.73±0.54a 3.86±0.28d 10.79±0.09c |
998.43±15.67a 517.62±38.93b 532.55±25.63b 94.04±4.44c 516.60±29.40b |
55.08±4.19b 22.97±1.63d 112.61±4.92a 22.39±1.63d 43.36±1.28c |
C SUM CUM BP RP |
|
Yersinia enterocolitica (NCTC 11175) |
570.71±10.84a 55.10±1.52d 278.62±5.22b 135.34±6.13c 143.63±14.69c |
27.70±1.05a 2.81±0.27c 26.75±0.58ab 2.67±0.12c 25.85±0.65b |
4.95±0.08bc 2.27±0.00d 26.27±1.62a 1.63±0.04d 3.67±0.26c |
48.16±0.45a 0.00±0.00e 38.72±1.66b 4.41±0.15d 21.71±2.23c |
0.00±0.00c 0.00±0.00c 17.19±0.27a 0.38±0.03b 0.63±0.01b |
0.00±0.00b 0.00±0.00b 14.43±0.68a 0.00±0.00b 0.00±0.00b |
24.81±0.65c 2.59±0.11e 66.23±1.43a 8.70±0.47d 51.04±0.82b |
135.38±10.36a 14.06±0.29c 97.55±1.46b 6.28±0.44c 15.28±3.22c |
8.23±0.53b 1.28±0.08d 16.08±1.42a 1.79±0.10d 5.67±3.73c |
1159.63±114.29a 440.95±28.32c 599.80±23.20b 102.60±3.14d 214.46±120.13d |
65.27±4.36b 25.79±0.08d 97.11±1.07a 10.70±0.10e 31.45±7.25c |
C SUM CUM BP RP |
|
Campylobacter jejuni (ATCC 33560) |
691.20±66.03a 309.57±8.03b 317.87±9.43b 168.40±9.99c 211.38±17.40c |
35.49±3.54b 7.84±0.24c 71.68±2.93a 13.43±1.16c 9.30±0.37c |
3.23±0.00c 3.27±0.01c 24.72±0.56a 13.98±0.24b 2.58±0.08c |
37.71±2.94a 4.22±0.39c 0.00±0.00d 3.96±0.12c 20.91±0.99b |
1.07±0.09b 0.00±0.00c 20.85±0.29a 0.89±0.01b 0.21±0.01c |
0.00±0.00b 0.00±0.00b 26.80±1.32a 0.00±0.00b 0.00±0.00b |
46.78±2.95b 6.34±0.21c 70.20±2.55a 7.65±0.61c 4.89±0.27c |
15.33±1.02b 11.06±1.31bc 125.78±4.12a 10.91±0.53bc 7.84±0.29c |
14.35±0.12b 6.39±0.28c 22.05±0.29a 6.23±0.32c 2.15±0.06d |
636.14±24.06a 279.47±7.78c 628.98±61.35a 16.78±1.17d 494.09±3.03b |
81.69±7.40b 19.04±1.00cd 95.69±0.04a 11.24±0.01d 26.79±0.87c |
C SUM CUM BP RP |
|
Aeromonas hydrophila (NCIMB1135) |
543.77±52.09a 99.15±2.96c 58.50±3.53c 83.68±5.21c 256.36±11.04b |
3.29±0.10b 5.71±6.63b 25.64±1.16a 1.74±0.06b 3.76±0.21b |
4.61±0.77c 5.33±0.28c 9.06±1.11b 4.57±0.14c 11.18±0.23a |
78.29±3.54a 6.36±0.44d 32.21±1.76b 1.29±0.13e 20.45±0.68c |
0.66±0.08b 0.00±0.00c 5.98±0.28a 0.28±0.02c 0.70±0.07b |
0.97±0.11b 0.00±0.00c 4.04±0.04a 0.00±0.00c 0.00±0.00c |
37.21±1.51b 6.73±0.22d 31.71±1.03c 8.91±0.19d 93.57±2.23a |
36.74±3.60b 7.36±0.17d 66.56±2.16a 5.07±0.07d 24.02±0.25c |
53.66±0.98a 2.76±0.23d 8.27±0.64b 4.89±0.42c 1.42±0.11d |
746.78±5.75a 578.68±34.14b 707.41±69.65a 57.53±4.14c 144.53±6.51c |
254.91±17.94a 51.44±1.79c 164.72±4.99b 13.96±0.95d 29.24±1.79d |
C SUM CUM BP RP |
Different lowercase letters in superscript (a–d) in a column indicate significant differences (P < 0.05) between the Control (C) and a Group for each food borne pathogen (FBP). Abbreviations of extracts, ammonia and biogenic amines (BA’s): C, Control; SUM, Sumac; CUM, Cumin; BP, Black pepper; RP, Red pepper; 0.00, Not Detected. AMN, Ammonia; PUT, Putrescine, CAD, Cadaverine; SPD, Spermidine; TRP, Tryptamine; PHEN, Phenylethylamine; SPM, Spermine; SER, Serotonin; TMA, Trimethylamine; DOP, Dopamine; AGM – Agmatine. Spice extracts were added to the in tyrosine decarboxylase broth at a rate of 1% (w/v).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



