
Submitted:
16 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Gamma-aminobutyric acid (GABA) is thought to have beneficial effects on human health and well-being. GABA enrichment by lactic acid bacteria (LAB) could be used in the development of functional fermented foods. In this work, four GABA-producing strains each of Lactococcus lactis and Streptococcus thermophilus were isolated from cowʼs milk, and their phenotypic, technological, and safety profiles were determined. Genome analysis provided genetic support for the majority of the analyzed traits. The operon harboring the glutamate decarboxylase gene (gadB) was chromosomally encoded in all strains and showed the same gene content and gene order as that reported, respectively, for L. lactis and S. thermophilus. In the latter species, the operon was flanked (as in most strains of this species) by complete or truncated copies of insertion sequences (IS), suggesting recent acquisition by horizontal gene transfer. The genomes of three L. lactis and two S. thermophilus strains showed a gene encoding a caseinolytic proteinase (PrtP in L. lactis and PrtS in S. thermophilus). These strains grew in milk forming a coagulum of good appearance and an appealing acidic flavor and taste. They also produced GABA from monosodium glutamate in this medium. Two L. lactis strains were identified as belonging to the biovar. diacetylactis, utilized citrate from milk and produced significant amounts of acetoin. None of the strains showed any noticeable antibiotic resistance, nor did their genomes harbor transferable antibiotic resistance genes, genes involved in toxicity, virulence, or pathogenicity. Altogether these results suggest all eight strains may be considered candidates for use as starter or components of mixed LAB cultures for the manufacture of GABA-enriched fermented dairy products.
Keywords:
Lactococcus lactis
; Streptococcus thermophilus starters
; diary cultures
; dairy products
; cheese
; yogurt
; milk fermentation
1. Introduction
In the human central nervous system, gamma-aminobutyric acid (GABA), a non-protein amino acid synthesized from glutamic acid, acts as an inhibitory neurotransmitter affecting nerve impulses. GABA reduces stress and improves the onset and duration of sleep; it thus acts as a tranquilizer [1]. In addition, GABA modulates kidney function and lowers blood pressure [2]. Some of these effects have been demonstrated in randomized clinical trials [1,3,4,5], strongly supporting the idea that GABA can have a beneficial influence on human health and well-being. These properties of GABA have attracted much interest about developing GABA-enriched functional foods [6].
The transformation of glutamic acid into GABA is carried out by the enzyme glutamate decarboxylase (GAD), using pyridoxal phosphate (the active form of vitamin B6) as a cofactor [7]. Caseins have a high concentration of glutamic acid, and the GABA precursor amino acid is therefore abundant in milk. GABA production in fermented dairy products could be achieved if free glutamic acid plus microorganisms (from starter cultures, adjuncts, or contaminating microbiota) with GAD activity are present [8,9,10]. Indeed, the use of GABA-producing lactic acid bacteria (LAB) as a starter and/or adjunct culture, might allow the production of GABA-enriched functional fermented dairy products [11,12,13]. GAD activity has been detected in strains of different LAB species, including Levilactobacillus brevis (formerly Lactobacillus brevis), Lacticaseibacillus paracasei (formerly Lactobacillus paracasei), Lactococcus lactis, Streptococcus thermophilus, and many others [11,13,14]. GAD activity seems to provide microorganisms with greater resistance to low pH, such as that found in most fermented foods [15,16]. In certain LAB, the decarboxylation of amino acids appears to be coupled to ATP biosynthesis. Therefore, the conversion of glutamic acid into GABA provides cells with extra energy that can be used for enhanced growth [17].
L. lactis and S. thermophilus are the most economically-important LAB species and are used worldwide as starters in cheese making and yogurt manufacturing, respectively [18]. However, only certain strains of these two species show GAD activity and are therefore capable of GABA production [10,12,13,14,19]. The enzyme in L. lactis is encoded by a single gene (gadB), that forms part of an operon (gadRCB), which includes two upstream genes coding for a GABA-glutamate antiporter (gadC) and a transcription regulator (gadR) [20,21]. Inactivation of either gadB or gadC results in reduced cell survival at low pH `20, 21]. The gadRCB operon in L. lactis is recognized as part of an acid stress regulon [21]. Genes equivalent to gadB and gadC in L. lactis are also found in S. thermophilus [15]. However, compared to L. lactis, the order of the genes in the operon is reversed and the gene encoding the regulator is absent [22]. Further, in S. thermophilus the gad operon is flanked by transposon- and insertion sequence-derived sequences, suggesting a horizontal acquisition [23]. Naturally, the confident use of GABA-producing strains of L. lactis and S. thermophilus as starters requires strains to be subjected to exhaustive biochemical and genetic characterization to ensure that they comply with technological and safety requirements [24].
This work reports a survey of GABA production by L. lactis and S. thermophilus strains isolated from raw milk. A set of eight GABA-producing strains (four L. lactis and four S. thermophilus) was subjected to a battery of procedures to determine their technological and safety properties, followed by genome sequencing that revealed the basis of many of their key traits, including GABA production.
2. Results
2.1. Molecular Identification and GABA Production
Molecular identification of 119 isolates from the PCA plates incubated at 32ºC proved lactococcal species to be the majority populations in raw milk, accounting for circa 35% of the isolates, including L. lactis (26 isolates), Lactococcus raffinolactis (8 isolates), and Lactococcus garvieae (7 isolates). Similarly, among 117 colonies selected from the GLM17 plates incubated at 42ºC, a majority of isolates were identified as S. thermophilus, accompanied by a few isolates of Staphylococcus epidermidis, Lactobacillus delbrueckii, Streptococcus gallolyticus subsp. macedonicus, and Streptococcus equinus. These isolates accounted for the species withstanding pasteurization and high incubation temperature. Thus, under the specific isolation conditions of this study, L. lactis (21.8%) and S. thermophilus (41.8%) proved to be, respectively, the majority mesophilic and thermophilic LAB species in milk.
Since L. lactis and S. thermophilus isolates were recovered from every milk sample, an isolate of each of the species per sample was tested for GABA production in GM17 (for L. lactis) or GLM17 (for S. thermophilus) broth supplemented with 5 mM MSG. GABA was detected in the supernatant of a majority of cultures, although tiny amounts were measured in some isolates. Isolates producing <0.64 mM of GABA in broth were considered non-producers. With this threshold, six L. lactis and eight S. thermophilus isolates converted most of the MSG precursor into GABA (>4.0 mM; >80% conversion). To avoid the presence of replicates, these 14 isolates were subjected to RAPD and rep-PCR typing. Taking into account the repeatability of the method (93%), the combined fingerprinting results suggested the presence of six different strains for each species (Figure S1). Among those, four unrelated strains each of L. lactis (Lc 5.5, Lc 14.4, Lc 19.3, and Lc 21.1) and S. thermophilus (St 8.1, St 9.1, St 18.1, and St 21.1) were selected for the characterization of relevant technological and safety properties, as well as for genome sequencing and analysis.
2.2. Biochemical and Technological Characterization
The carbohydrate fermentation profile of the strains, as determined by the API-50 CHL system, matched those of their respective species (Table S1). The L. lactis profile ranged from 13 to 21 carbohydrates. Lc 21.1 utilized the smallest number of compounds, which is typical of well-adapted-to-milk (“domesticated”) lactococcal strains. The profile of the S. thermophilus strains included the fermentation of 3-5 carbohydrates, a short but typical sugar-utilization profile for this species.
Species-specific profiles for enzyme activities, assayed using the API-ZYM system, were also scored for the L. lactis and S. thermophilus strains (Table S2). Strains of the two species showed strong leucine aminopeptidase and phosphohydrolase (against naphthol-AS-BI) activities. L. lactis strains showed strong acid phosphatase activity and moderate cysteine aminopeptidase activity. In general, the S. thermophilus strains showed strong β-galactosidase activity, moderate valine aminopeptidase and esterase activities, and weak lipase activity. Although with variable enzymatic levels, all S. thermophilus strains returned a positive urease test.
Two strains of each of the species (Lc 5.5 and Lc 14.4, and St 8.1 and St 18.1) grew well and coagulated the milk by 12 h of incubation (pH 4.43-4.70), forming a firm coagulum of good appearance and pleasant acidic flavor and taste. In contrast, the pH of the milk at this time was ≥6.0 for all other strains, and none of these clotted the milk even after 24 h of incubation. Minor differences in acidification were observed when the milk was supplemented with 10 mM MSG (Figure 1).
GABA production in supplemented milk was associated with growth (Figure 1). However, GABA synthesis in S. thermophilus was retarded compared to that recorded for L. lactis. At 12 h of incubation, only L. lactis Lc 5.5. and Lc 14.4 produced significant amounts of GABA, while at 24 h three L. lactis (Lc 5.5, Lc 14.4, and Lc 19.3) and two S. thermophilus (St 8.1 and St 18.1) strains did so. In laboratory media, the presence of MSG caused a small increased in the final pH of the cultures (0.23±0.06). However, this effect was not observed in milk. Finally, under the conditions of this study, none of the strains produced GABA in MSG-supplemented fecal homogenates.
The strains produced and consumed different amounts of organic acids and sugars. However, no clear-cut species-specific profiles were observed (Table 1). In L. lactis, major differences between the strains were associated with the utilization of citrate. Strains using this compound (Lc 14.4 and Lc 21.1) produced large amounts of acetic acid. In addition, strains Lc 5.5 and Lc 14.4 produced moderate amounts of formic and succinic acids. In S. thermophilus, profiles for sugar fermentation and the production of organic compounds seemed to be mostly influenced by growth in milk. Strains St 8.1 and St 18.1 grew well and generated large amounts of lactic acid, releasing large quantities of galactose. Strains St 9.1 and St 21.1 did not grow well in milk and, in general, their metabolic activity was rather poor. At the same time, they both released notable quantities of glucose, suggesting premature lysis of the cells.
After growing the strains in milk at either 32ºC (L. lactis strains) or 42ºC (S. thermophilus strains), 29 volatile compounds were detected (Table 2), 15 of which were not present in the starting milk. For the L. lactis strains, no species-specific profile was observed due to the different behaviour shown by the Lc 21.1 strain; unlike its counterparts it did not produce isovaleric acid, 4-octanone, or phenylacetaldehyde. Further, Lc 5.5 and Lc 14.4 produced 2-phenyl ethanol, isoamyl alcohol, and 5-methyl-2-phenyl-2-hexenal. Lc 14.4 produced acetoin (diacetyl cannot be detected with the solid-phase microextraction (SPME)-gas chromatography method utilized in this study), and large amounts of acetic acid. Lc 5.5 and Lc 19.3 both produced notable amounts of benzoic acid, while Lc 14.4 and Lc 19.3 produced 2,5-dimethyl-3-hexanone and 2,3-heptanodione. In contrast, with only minor differences between the strains, species-specific profiles were seen for the four S. thermophilus strains. Two species-specific volatile compounds were associated with S. thermophilus, namely tetradecanoic acid and 2-heptanone.
Under the conditions of the assay, none of the strains produced bacteriocin-like inhibitory substances active against L. sakei CECT906T.
2.3. Safety Evaluation
The MIC values of the tested antibiotics showed the strains to be susceptible to all relevant antibiotics (Table S3). All strains were strongly resistant to trimethoprim, a resistance intrinsic to cocci LAB. Deviating from the profile, L. lactis Lc 21.1 and S. thermophilus St 9.1 showed a high MIC value to rifampicin (>64 µg mL-1) and kanamycin (128 µg mL-1), respectively.
Under the culture conditions assayed, none of the isolates produced biogenic amines from any of the four precursors tested (histidine, tyrosine, agmatine, and ornithine).
2.4. Genome Sequence and Analysis
To understand the genetic basis of GABA production -and to help ensure their safe use- all eight strains were subjected to genome sequencing and analysis. The acquired genomic data served to confirm the taxonomic identification of the strains via in silico dDDH and orhtoANI analysis. The examined strains were compared with type strains of related species, including L. lactis subsp. lactis, L. lactis subsp. lactis biovar. diacetylactis, and L. cremoris subsp. cremoris, or the S. thermophilus type strain and the type strains of related species (Table S4).
These analyses confirmed the previous identification of the strains as either L. lactis or S. thermophilus. Further, L. lactis Lc 21.1. was identified as belonging to biovar. diacetylactis. Although lower than those of Lc 21.1, the dDDH and orhtoANI values indicated Lc 14.4 to be slightly closer to the L. lactis subsp. lactis biovar. diacetylactis type strain (GL2T) than to the L. lactis subsp. lactis type strain (ATCC 19435T).
Table 3 shows the general features of the genome of all the sequenced strains. Genomic analysis of the strains showed they all carried the gene coding for the GAD enzyme (gadB) and its corresponding associated antiporter (gadC) organized as an operon-like structure (Figure 2). The assembly of the genome of the L. lactis strain Lc 14.4 split the operon into four contigs, impeding the determination of its structure. One contig harboured genes encoding proteins involved in plasmid replication, which strongly suggest that there are two copies of the GABA operon in the genome of this strain, of which one is plasmid-associated. This structure may account for the assembly failure in Lc 14.4 (Figure 2A). As expected, the order of the gad genes in L. lactis and S. thermophilus was reversed, and the open reading frames (ORFs) flanking them were different in the two species (Figure 2B). The GABA operon in L. lactis was flanked by chromosomal genes encoding ribonuclease HII (rnhB) and a potassium efflux system (kefA). Located at the first position, the operon in L. lactis also comprised the regulatory gene (gadR) (Figure 2A).
The S. thermophilus operon also appeared to be encoded on the chromosome, and in the same position in all strains. However, the operon in this species lacked the gene encoding the regulator and was flanked by sequences coding for insertion elements and mobilization proteins (Figure 2B), suggesting a (recent) acquisition by horizontal transfer.
The genetic makeup for lactose and galactose utilization included clusters for lactose utilization via the Leloir pathway in both the L. lactis and S. thermophilus strains, and an operon for lactose utilization via the tagatose-6-phosphate pathway in the genome of all lactococcal strains (Figure S2). These pathways are well characterized in strains of many LAB species, including L. lactis and S. thermophilus [25].
Genome analysis of the citrate-associated genes in the two L. lactis strains utilizing citrate (Lc 14.4 and Lc 21.1) revealed striking differences (Figure S3). Three operons, each located in a different contig, were identified in Lc 21.1. Two operons were considered to be located on the bacterial chromosome, encoding the citrate lyase complex and the malolactic enzyme. Each of the operons harbored a gene encoding a citrate/H+ symporter-like permease, which contained mutations that might render the enzymes inactive. The third operon was found in a contig together with genes encoding plasmid-replication proteins, strongly suggesting a plasmid location. This operon also contained an ORF encoding the well-described citrate-specific transporter CitP. In contrast, only the chromosomal operons encoding citrate lyase and malolactic enzyme were identified in the genome of the Lc 14.4 strain. Surprisingly, the genes coding for the corresponding associated permeases were complete and contained no mutations. Genes encoding citrate synthase and isocitrate dehydrogenase were found in the genome of all L. lactis strains; these enzymes might intervene in the synthesis of citrate.
The analyzed genomes showed a complex repertoire of metabolic genes involved in the growth of the strains in milk and the formation of taste and aroma compounds (Tables S5 and S6). With only minor differences between strains, gene content species-specific profiles were established. Nonetheless, these minor differences might be critical for rapid development and/or for the synthesis of key flavor compounds in milk. In addition to the genetic machinery involved in diacetyl production from citrate, genes coding for the L. lactis and S. thermophilus caseinolytic proteinases (PrtP and PrtS, respectively) were identified in most strains, but not in Lc 21.1, St 9.1, and St 18.1. High cell densities and milk coagulation after overnight culture were only attained by the strains harboring either prtP or prtS genes. As expected from the biochemical characterization results, a complete urease operon consisting of three catalytic enzymes and seven accessory genes was found in all four S. thermophilus strains (Figure S4).
As expected, no CRISPR-Cas-associated loci were identified in the genome of the L. lactis strains; these systems are uncommon in this species. In agreement, a large proportion of phage-associated genes was identified in the genome of all L. lactis strains (Table 3). The Phaster software associated these genes with either complete (from one to three) or incomplete (from 4 to 13) phages. In contrast, all four S. thermophilus strains possessed three loci showing both CRISPR repeats and genes coding for Cas proteins. A short series of isolated CRISPR repeats was also found in all strains except in St 8.1. Only two loci showing an incomplete set of genes of phage origin were detected in all S. thermophilus strains. In addition, a complete phage of 43.9 kbp was identified in strain St 21.1. PlasmidFinder identified two plasmid-derived sequences in St 9.1, while one plasmid-replication protein was detected by BV-BRC in St 8.1, St 9.1, and St 18.1 (Table 3). PlasmidFinder has been reported not to be reliable for the detection of plasmids in LAB species [26]. Despite this, the software identified plasmid sequences in all L. lactis strains, which agrees well with the large number of genes encoding putative plasmid-associated proteins detected by BV-BRC in all lactococcal genomes (Table 3).
Despite the absence of antimicrobial activity in the agar well-diffusion assay, several loci were identified by BAGEL 4 as putative regions encoding bacteriocin-like peptides (Blp) or ribosomally synthesized and post-translationally modified peptides (RiPPs) (Table 3). Besides several lactococcins, Lc 14.4 encoded a protein with homology to enterolysin A (53% amino acid identity). Three lactococcal strains showed a locus for the synthesis of sactipeptides (sulfur-to-α-carbon-containing peptides; also classified as Class Ic bacteriocins). Finally, a locus for the synthesis of linaridin (a class of linear, dehydrated peptides) was detected in the genome of Lc 19.3. For the S. thermophilus strains, loci involved in the synthesis of several Blp antimicrobials (Class II bacteriocins with a double-glycine leader peptide) were identified. In addition, they all contained a locus for the synthesis of streptides (a class of 20-membered cyclic peptides). Finally, St 8.1 showed a locus with the capability to encode a bacteriocin homologous to the BhtR bacteriocin (a Class I two-component lantibiotic) identified in Streptococcus ratti [27].
Interrogation of the genomes with the CARD database for antibiotic resistance genes detected two strict hits in the genome of all four S. thermophilus strains with low homology (<33% identity) to the deduced proteins for the vanT and vanY genes in Enterococcus spp. These genes were annotated by BV-BRC as alanine racemase and D-alanyl-D-alanine carboxypeptidase, respectively. Neither genes nor mutations involved in antibiotic resistance were detected in the S. thermophilus genomes by ResFinder. The D-alanyl-D-alanine carboxypeptidase gene was also detected as a strict hit by CARD in all L. lactis strains. In addition, this database identified the lmrD gene in all strains as a perfect hit (100% identity, 100% length coverage). lmrD encodes a subunit of the well-characterized LmrCD heterodimeric efflux ABC transporter, which is involved in multidrug resistance in L. lactis [28].
Glutamate decarboxylase was the only amino acid carboxylase encoded in the genome of any strain. A few genes encoding housekeeping enzymes, multidrug efflux pumps, and proteins involved in heavy metal homeostasis and resistance, were found in the genomes of the strains on the Rast Server, except for a gene encoding a serine protease with partial similarity to the exfoliative toxin A from Staphylococcus aureus present in all L. lactis and S. thermophilus strains, no genes encoding toxins were identified.
3. Discussion
In the present work, four strains each of L. lactis and S. thermophilus, selected from among a set of GABA-producing LAB isolated from raw milk based on the typing results, were phenotypically and genomically characterized. In fermented foods, LAB starters increase the availability of milk nutrients and can contribute to the production of bioactive compounds with a beneficial effect on consumer health, such as GABA [29]. Consequently, GABA-producing LAB starter and adjunct cultures could be used in the development of functional fermented foods. However, the functional, technological, and safety traits of such cultures is paramount if safe products with appealing sensory properties are to be obtained [30]. The combined phenotype testing and genome sequencing and analysis performed revealed the underlying genetic basis of a majority of their key traits, including GABA production.
Genome analysis confirmed the identification of the strains at the species level (as L. lactis and S. thermophilus), and allocated two of the L. lactis strains (Lc 14.4 and Lc 21.1) to the biovar. diacetylactis. These two strains were able to use citrate during growth in milk and produced noticeable amounts of acetoin. Both strains harbored a complete operon encoding all the components of the citrate lyase complex. However, the absence of citP in Lc 14.4, which utilized citrate from milk more efficiently than Lc 21.1., was surprising. It may be that one of the two citrate/H+ symporters in Lc 14.4, or both, which are inactivated by mutations in Lc 21.1 (a strain that harbors a citP gene), takes over the internalization of citrate. Although not yet studied in detail, citrate-utilizing (and diacetyl-producing) L. lactis strains without a recognizable CitP permease, have been reported elsewhere [31,32]. None of the S. thermophilus strains utilized citrate from milk, but they all produced moderate amounts of acetoin. S. thermophilus is thought to produce diacetyl and acetoin by diverting the α-acetolactate that may result from an excess of branched-chain amino acids [33].
All eight strains converted MSG to GABA, both in laboratory media and in milk. The amount of GABA produced by the strains falls within the mid-high range compared to other L. lactis and S. thermophilus strains [12,14,34], but changes in manufacturing and/or ripening conditions may lead to improved GABA production in fermented dairy products [14,35]. The synthesis of GABA was sustained by the presence of operons harboring the genes encoding both the GAD enzyme (gadB) and the associated glutamate-GABA antiporter (gadC). The operon of L. lactis strains contained a gene (gadR) coding for a putative regulatory protein, while that of S. thermophilus strains did not. This is the usual gene content and gene order in these two LAB species, respectively [23,36]. Despite the assembly failure of the GABA operon in Lc 14.3, GABA production indicates that Lc 14.4 carries a fully functional biosynthetic pathway. Indeed, the strain was thought to carry GABA genes in both the chromosome and on a plasmid. Shuffling of genes between the bacterial chromosome and plasmids in L. lactis has been reported elsewhere [37]. The first role attributed to gene duplication is to provide enhanced enzymatic activity. Not surprisingly, Lc 14.4 showed the highest GABA production at 24 h.
In contrast to the results in laboratory media and milk, none of the strains produced GABA in faecal homogenates. LAB strains with this property could have a role as probiotics, allowing GABA production in the gastrointestinal tract after ingestion [29]. This trait has been found in some lactobacilli and Bifidobacterium spp. strains of gut origin [38,39]. The presence of large numbers of bacteria well adapted to the gut environment may have prevented the dairy strains from growing (and thus producing GABA) in the faecal cultures.
For rapid lactic acid production, and to attain a high cell density in milk, LAB require efficient systems for the utilization of lactose and caseins as a source of carbon and nitrogen [30]. The lactose operons found in the genome of the sequenced strains corresponded to those described for L. lactis and S. thermophilus, respectively [25]. However, genes encoding extracellular proteinases with caseinolytic activity (PrtP and PrtS for L. lactis and S. thermophilus, respectively) were not identified in Lc 21.1, St 9.1, and St 21.1. These three strains grew slowly and did not coagulate milk after 24 h of incubation at their respective optimal temperature. It is noteworthy that the PrtP sequence from L. lactis Lc 5.5 lacked a C-terminal region that included the final 163 amino acids. Deletions of the C-terminal part of the lactococcal caseinolytic protease retaining proteolytic activity had already been experimentally determined [40]. prtP and prtS genes are either located in plasmids or on the chromosome [41,42]. Chromosomal and plasmidic genes are thought to be acquired by horizontal transmission and can be mobilized by conjugation [41,42]. Surprisingly, the Lc 19.3 strain grew slowly in milk, even though it had a complete prtP gene. The reason for the slow growth in milk of this strain remains unknown. Changes in regulatory regions outside the coding regions may account for a poor expression of the protease and/or its essential maturation partner protein in L. lactis (PrtM) [43]. Nevertheless, the inclusion of protease-positive and protease-negative variants in the same starter mixture has been shown to reduce bitterness by certain L. lactis cultures [44].
Although BAGEL 4 identified loci for bacteriocins and RiPPs in all strains, none inhibited the highly susceptible strain used as an indicator. Some of these molecules (sactipeptides, streptides) have been identified in LAB only recently by mining their genomes with bioinformatic tools and may have if at all, a narrow-spectrum activity [45], which could explain the negative testing results. Further, classical bacteriocins may require specific target cells, functional quorum sensing systems, or the presence of autoinducers that have to be provided by a helper strain [46].
Apart from the gene encoding glutamate decarboxylase, none coding for amino acid carboxylases was detected in any strain, which is consistent with the negative results obtained for the production of biogenic amines. Neither were transmissible resistance genes identified, which agrees with the susceptibility of the strains to all relevant antibiotics tested. Further, despite the results of the BV-BRC annotation program, no genes coding for virulence factors nor toxins were identified in dedicated databases, except for those encoding exfoliative toxins. These proteins in LAB share significant structural homology with chymotrypsin, have a very limited substrate specificity, and their physiological role remains to be determined [47]. All these data suggest that the examined strains are free of genes of concern, which indicates they all are safe and can be confidently used in food systems. The presence of prophages in a genome that, under uncertain environmental conditions might be activated and thus induce the following of a lytic cycle, might impair a bacterial strain’s use in industrial settings [48]. However, the total absence of integrated phages, at least in L. lactis strains, is highly unlikely [49].
4. Materials and Methods
4.1. Milk Sampling and Bacterial Culture Conditions
Twenty-four raw cow’s milk samples from different areas of the Asturias region (Northern Spain) were used as a source of Lactococcus lactis and Streptococcus thermophilus isolates. For the recovery of mesophilic microorganisms, milk dilutions in Ringer (Merck, Darmstadt, Germany) were plated on PCA agar (Merck) and incubated for 48-72 h at 32ºC. For the recovery of thermophiles, 40 mL milk samples were pasteurized at 63ºC for 30 min and incubated overnight at 42ºC. Dilutions of coagulated milk samples with good appearance and odor were then plated on M17 (Formedium, Norfolk, UK) agar supplemented with 0.5% glucose and 0.5% lactose (GLM17). The plates were incubated in an anaerobic jar with Anaerocult A (Merck) at 42ºC for 24-48 h. Representative colonies produced at the two incubation temperatures were chosen at random and purified by subculturing on M17 agar supplemented with 1% glucose (GM17), or on GLM17, and incubated under the above conditions. Purified colonies were inoculated into broth of their respective isolation medium, and incubated overnight. Cells were then centrifuged, suspended in fresh medium supplemented with 20% glycerol (v/v), and stored at -80ºC.
4.2. Identification and Typing
The overnight cultures of the isolates used to prepare the frozen stocks were also used for total genomic DNA extraction and purification using the QIAmp DNA Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol for Gram-positive bacteria. Isolates were identified by amplification of their 16S rRNA genes with the universal primers 27F (5’-AGAGTTTGATCMTGGCTCAG-3’) and 1492R (5’-GGTTACCTTGTTACGACTT-3’), and partial sequencing with the 27F primer. The sequences obtained were compared with those in databases.
GABA-producing L. lactis and S. thermophilus isolates were typed by combining the fingerprinting profiles obtained with primers BoxA2R (5’-ACGTGGTTTGAAGAGATTTTCG-3’), OPA18 (5’-AGGTGACCGT-3’) and M13 (5’-GAGGGTGGCGGTTCT-3’), following the amplification conditions described by Rodríguez et al. [50]. Amplicons supplemented with EZ-Vision Two DNA Dye (VWR International) were separated in 2.5% agarose gels, visualized under UV light in a G-Box transilluminator (Syngene, Cambridge, UK), and the profiles obtained clustered using the Unweighted Pair Group Method with Arithmetic mean algorithm (UPGMA) and Jaccard similarity coefficient.
4.3. GABA Production and Quantification
The ability of the L. lactis and S. thermophilus isolates to synthesize GABA from its precursor was tested, respectively, in GM17 or GLM17 broth media supplemented with 5 mM monosodium glutamate (MSG) after overnight incubation at either 32ºC (for L. lactis) or 42ºC (for S. thermophilus). GABA was detected and quantified by ultra-high-performance liquid chromatography (UHPLC) as previously reported [19].
2.4. Phenotypic, Technological and Safety Profiles of GABA-Producing Strains
2.4.1. Phenotype Characterization
The carbohydrate fermentation profile of the L. lactis and S. thermophilus strains was examined using the API50 CHL system (bioMérieux, Montalieu-Vercieu, France). Enzyme activities were identified using the semi-quantitative API-ZYM system (bioMérieux). Urease activity in S. thermophilus was measured by assessing the release of ammonia from urea in the phenol red assay [51]. Briefly, 0.5 mL of an overnight culture in GLM17 was added to one volume of solution A (2 g of urea in 2 mL of ethanol and 4 mL of sterile water) and 19 volumes of solution B (KH2PO4 1 g L-1; K2HPO4 1 g L-1; NaCl 5 g L-1; phenol red 20 mg mL-1). This suspension was then incubated at 37°C for up to 2 h; the development of a violet-red color indicated positive activity.
2.4.2. Growth in Milk and Production of Organic Acids and Volatile Compounds
The L. lactis and S. thermophilus strains were grown in 10 mL of commercial UHT semi-skimmed milk (CAPSA, Siero, Spain) at 32ºC or 42ºC, respectively, for 12-24 h. Milk cultures were evaluated in triplicate for the production of organic acids, sugars and volatile compounds (VOCs).
Organic acids and sugars from milk cultures were extracted and determined by UHPLC following the method of Alegría et al. [52]. Briefly, organic acids and sugars were separated using an ICSep ICE-ION-300 ion-exchange column (ThermoFisher Scientific, Waltham, MA, USA), employing an 8.5 mM H2SO4 aqueous mobile phase, an operating temperature of 65ºC, and a flow rate of 0.4 mL/min. Organic acids were identified using a 996 Photodiode Array Detector (Waters, Milford, MA, USA) at 210 nm, and sugars using a 410 Differential Refractometer (Waters) at 280 nm, with quantification against calibration curves prepared with commercial standards.
The production of VOCs in milk was determined following the solid-phase microextraction (SPME)-gas chromatography method of Ziadi et al. [53]. Briefly, 5 mL of fermented milk were added to 20 mL screw-capped SPME vials (Agilent Technologies, Santa Clara, CA, USA). The vials were then sealed with a PTFE/silicone liner septum and equilibrated at 60ºC for 10 min with pulsed agitation for 5 s at 500 rpm using a PAL RSI 120 device (CTC Analytics, Zwingen, Switzerland). Volatile compounds were absorbed onto an ARR11-DVB-120/20 DVB/PDMS fibre (CTC Technologies) exposed to the headspace above the samples for 20 min at a depth of 40 mm (performed at 60ºC). All other GC-MS conditions were as in Ziadi et al. [53] and the eluted compounds were identified by their retention times and by comparison of their mass spectra with the Wiley Mass Spectral database (Wiley and Sons, NY, USA). Quantification was done by gas chromatography with a flame ionization detector (GC-FID) (HP5890 series II plus).
2.4.3. GABA Production in Milk and Feces
GABA production by the strains in milk was assessed in commercial UHT semi-skimmed milk supplemented with 10 mM MSG after 12-24 h incubation at the optimal temperature for each strain. GABA was detected and quantified as above. In addition, GABA production by the strains was assessed in fecal homogenates supplemented with 30 mg mL⁻¹ MSG following the procedure of Barrett et al. [38].
2.4.4. Production of Biogenic Amines
The production of biogenic amines was examined in the supernatants of cultures following incubation at the optimal temperature for 48 h in GM17 or GLM17 supplemented with 5 mM of the precursor amino acids and amino acid-derived compounds (tyrosine, histidine, agmatine, and ornithine) (all from Sigma-Aldrich, Merck). As positive controls, the tyramine and histamine producers Enterococcus faecalis BA6 and Lentilactobacillus parabuchneri 2ES-1 were used. Processing and sample analysis were performed following the method described by Redruello et al. [54].
2.4.5. Antibiotic Resistance
The antimicrobial resistance-susceptibility profile of the L. lactis and S. thermophilus strains to 16 antibiotics was assessed by broth microdilution using EULACBI1 and EULACBI2 plates (Trek Diagnostic System; ThermoFisher, Waltham, MASS, USA), as previously reported [50]. Minimum inhibitory concentration (MIC) values were compared to those reported by the European Food Safety Authority [55].
4.4.6. Production of Bacteriocins
The production of bacteriocin-like substances was examined by an agar-well diffusion test using Latilactobacillus sakei CECT906T as an indicator, following previously reported inoculation procedures and incubation conditions (Rodríguez et al., 2022).
2.5. Whole Genome Sequencing and Analysis
Next-generation sequencing libraries were prepared with total genomic DNA and using the TruSeq DNA PCR-free Sample Preparation Kit (Illumina, San Diego, CA, USA). Libraries were paired-end sequenced in a HiSeq 1500 System (Eurofins, Ebersberg, Germany). Genomes were annotated using the services of the Bacterial and Viral Bioinformatics Resource Center (BV-BRC; https://bv-brc.org/). For this, reads were first checked for quality with FastQC software, and contigs were assembled using the Unicycler program after comparing different variables using the Quality Assessment Tool for Genome Assemblies (QUAST). Errors were polished using Pilon and Racon software. Annotation with BV-BRC was performed using the RAST tool kit (RASTtk). Using BLAST tools (https://blast.ncbi.nlm.nih.gov/Blast.cgi), DNA and deduced protein sequences of interest were individually compared against those in the NCBI database. Whole-genome sequence data were used to confirm identifications through digital DNA-DNA hybridization (dDDH) and orthologous average nucleotide identity (orthoANI) analyses, as reported by Meier-Kolthoff and Göker [56] and Yoon et al. [57], respectively.
Genome sequences were searched for plasmids using PlasmidFinder (https://www.genomicepidemiology.org/services/). Ribosomally synthesized and post-translationally modified peptides and bacteriocins were sought using BAGEL 4 (http://bagel4.molgenrug.nl/). CRISPR-Cas systems were searched for using the CRISPRCasTyper v.1.6.4 online tool (https://crisprcastyper.crispr.dk). Phaster software was used to search the genome sequence for prophages (https://phaster.ca).
Virulence, Disease, and Defense proteins were searched among the subsystems of the Rast Server (https://rast.nmpdr.org/), as well as in the Virulence Factors (VFDB; http://www.mgc.ac.cn/VFs/main.htm) and Victors (http://www.phidias.us/victors) databases.
5. Conclusions
Genomic analysis of four GABA-producing strains each of L. lactis and S. thermophilus, isolated from milk, revealed the position and structure of the operon for the biosynthesis of this bioactive compound. In addition to laboratory media, all strains produced GABA in milk supplemented with MSG, but not in fecal homogenates. Genomic analysis determined the strains’ full biotechnological potential and contributed information guaranteeing their safety for use in food systems. Species-specific genes involved in the growth in milk and the formation of flavor compounds (taste and aroma) were recorded, which agrees with the differential profile of VOCs found in milk fermented by strains of the two species. The combined use of both L. lactis and S. thermophilus strains as a starter could result in fermented foods with improved sensory attributes. The individual strains might also be used to complement currently-in-use starter strains. Phenotype and genome analyses indicate the strains to be suitable and safe for the manufacture of GABA-enriched functional fermented milks supplemented with MSG. Testing of their suitability in other dairy systems (e.g., cheese) is currently underway.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Carbohydrate fermentation profile of the Lactococcus lactis and Streptococcus thermophilus GABA producer strains of this study; Table S2: Enzymatic activities of GABA-producing L. lactis and S. thermophilus strains by the API-ZYM system; Table S3: Minimum inhibitory concentration (MIC) of 16 antibiotics to GABA-producing L. lactis and S. thermophilus from mil; Table S4: Genomic distance analysis of the L. lactis and S. thermophilus (B) strains of this study with type strains of related species; Table S5 Presence of metabolic genes involved in growth in milk and the formation of flavor compounds (taste and aroma) in the genome of four GABA producer L. lactis strains; Table S6: Presence of metabolic genes involved in the growth in milk and formation of flavor compounds (taste and aroma) in the genome of four GABA producer S. thermophilus strains; Figure S1 RAPD and rep-PCR typing profiles of L. lactis and S. thermophilus strains; Figure S2: Genetic organization of the lactose/galactose operons found in the genome of L. lactis and S. thermophilus (B) strains; Figure S3: Genetic organization of citrate gene clusters found in the genome of L. lactis strains; Figure S4. Genetic organization of the urease operon in the genome of S. thermophilus strains.
Author Contributions
Conceptualization, B.M. and O.M.V.; methodology, B.M. and A.B.F.; investigation, J.A.V., J.R., and L.V.; genomic data curation and submission, A.B.F., J.A.V., and L.V.; writing—original draft preparation, B.M.; writing—review and editing, B.M., J.A.V., L.V., J.R., O.M.V. and A.B.F.; funding acquisition, B.M. and O.M.V. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by a Grant (Ref. PID2022-141271NB-I00) funded by MCIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe” project. J.A.V’s stay at IPLA-CSIC was sponsored by a scholarship from the “Programa de Movilidad Académica de la Asociación Universitaria Iberoamericana de Posgrado (AUIP)”. A PhD grant from the Severo Ochoa Program of Asturias Principality was awarded to J.R. (BP19-098).
Data Availability Statement
The draft genome sequences of the strains examined in this study were deposited in GenBank under the BioSample accession numbers SAMN39094998, SAMN39094999, SAMN39095000, and SAMN39095001 (L. lactis), and, SAMN39095002, SAMN39095003, SAMN39095004, and SAMN39095005 (S. thermophilus).
Acknowledgments
The biogenic amine producers Enterococcus faecalis BA6 and Lentilactobacillus parabuchneri 2ES-1 were kindly provided by V. Ladero (IPLA-CSIC).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of oral gamma-aminobutyric acid (GABA) administration on stress and sleep in humans: A systematic review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.H.; Vo, T.S. An updated review on pharmaceutical properties of gamma-aminobutyric acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-pressure-lowering effect of a novel fermented milk containing gamma-aminobutyric acid (GABA) in mild hypertensives. Eur. J. Clin. Nutr. 2003, 57, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Pouliot-Mathieu, K.; Gardner-Fortier, C.; Lemieux, S.; St-Gelais, D.; Champagne, C.P.; Vuillemard, J.C. Effect of cheese containing gamma-aminobutyric acid-producing lactic acid bacteria on blood pressure in men. Pharma Nutr. 2013, 1, 141–148. [Google Scholar] [CrossRef]
- Shimada, M.; Hasegawa, T.; Nishimura, C.; Kan, H.; Kanno, T.; Nakamura, T.; Matsubayashi, T. Anti-hypertensive effect of gamma-aminobutyric acid (GABA)-rich Chlorella on high-normal blood pressure and borderline hypertension in placebo-controlled double blind study. Clin. Exp. Hypertens. 2009, 31, 342–354. [Google Scholar] [CrossRef]
- Essa, M.M.; Bishir, M.; Bhat, A.; Chidambaram, S.B.; Al-Balushi, B.; Hamdan, H.; Govindarajan, N.; Freidland, R.P.; Qoronfleh, M.W. Functional foods and their impact on health. J. Food Sci. Technol. 2023, 60, 820–834. [Google Scholar] [CrossRef] [PubMed]
- Yogeswara, I.B.A.; Maneerat, S.; Haltrich, D. Glutamate decarboxylase from lactic acid bacteria-A key enzyme in GABA synthesis. Microorganisms 2020, 8, 1923. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R. Tuohy, K.M. Biodiversity and γ-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. BioMed Res. Inter. 2015, 2015, 25740. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Kimoto, H.; Someya, Y.; Fukurawa, S.; Suzuki, I. Production of γ-aminobutyric acid by cheese starters during cheese ripening. Dairy Foods 1998, 81, 1486–1491. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Tres, A.; Llombart, M.; Rafecas, M. Spanish cheese screening and selection of lactic acid bacteria with high gamma-aminobutyric acid production. LWT-Food Sci. Technol. 2014, 56, 351–355. [Google Scholar] [CrossRef]
- Linares, D.M.; O’Callaghan, T.F.; O’Connor, P.M.; Ross, R.P.; Stanton, C. Streptococcus thermophilus APC151 strain is suitable for the manufacture of naturally GABA-enriched bioactive yogurt. Front. Microbiol. 2016, 7, 1876. [Google Scholar] [CrossRef]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [PubMed]
- Renes, E.; Linares, D.M.; González, L.; Fresno, J.M.; Tornadijo, M.E.; Stanton, C. Production of conjugated linoleic acid and gamma-aminobutyric acid by autochthonous lactic acid bacteria and detection of the genes involved. J. Funct. Foods 2017, 34, 340–346. [Google Scholar] [CrossRef]
- Sezgin, E.; Tekin, B. Molecular evolution and population genetics of glutamate decarboxylase acid resistance pathway in lactic acid bacteria. Front. Genet. 2023, 14, 1027156. [Google Scholar] [CrossRef] [PubMed]
- Su, M.S.; Schlicht, S.; Gänzle, M.G. Contribution of glutamate decarboxylase in Lactobacillus reuteri to acid resistance and persistence in sourdough fermentation. Microb. Cell Fact. 2011, 10, S8. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Hayashi, H.; Abe, K. Exchange of glutamate and gamma-aminobutyrate in a Lactobacillus strain. J. Bacteriol. 1997, 179, 3362–3364. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.; Guinee, T.P.; Cogan, T.M.; and McSweeney, P.L.H. Starter cultures. In Fundamentals of Cheese Science; Fox, P., Guinee, T.P., Cogan, T.M., and McSweeney, P.L.H., Eds.; Springer Nature: New York, USA, 2017; pp. 121–183. [Google Scholar]
- Valenzuela, J.A.; Flórez, A.B.; Vázquez, L.; Vasek, O.M.; Mayo, B. Production of γ-aminobutyric acid (GABA) by lactic acid bacteria strains isolated from traditional, starter-free dairy products made of raw milk. Benef. Microbes 2019, 10, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Nakajima, I.; Fujita, Y.; Kobayashi, M.; Kimoto, H.; Suzuki, I.; Aso, H. Lactococcus lactis contains only one glutamate decarboxylase gene. Microbiol. 2000, 145, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.W.; Leenhouts, K.; Burghoorn, J.; Brands, J.R.; Venema, G.; Kok, J. A chloride-inducible acid resistance mechanism in Lactococcus lactis and its regulation. Mol Microbiol. 1998, 27, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.X.; Xie, C.; Gu, Z.X. Optimisation of fermentative parameters for GABA enrichment by Lactococcus lactis. Czech J. Food Sci. 2009, 27, 433–442. [Google Scholar] [CrossRef]
- Somkuti, G.A.; Renye, J.A., Jr.; Steinberg, D.H. Molecular analysis of the glutamate decarboxylase locus in Streptococcus thermophilus ST110. J. Ind. Microbiol. Biotechnol. 2012, 39, 957–963. [Google Scholar] [CrossRef]
- Pereira, G.V.M.; De Carvalho Neto, D.P.; Junqueira, A.C.D.O.; Karp, S.G.; Letti, L.A.; Magalhães Júnior, A.I.; Soccol, C.R. A review of selection criteria for starter culture development in the food fermentation industry. Food Rev. Int. 2020, 36, 135–167. [Google Scholar] [CrossRef]
- Iskandar, C.F.; Cailliez-Grimal, C.; Borges, F.; Revoll-Junelles, A.M. Review of lactose and galactose metabolism in lactic acid bacteria dedicated to expert genomic annotation. Trends Food Sci. Technol. 2019, 88, 121–132. [Google Scholar] [CrossRef]
- Flórez, A.B.; Vázquez, L.; Rodríguez, J.; Mayo, B. Phenotypic and safety assessment of the cheese strain Lactiplantibacillus plantarum LL441, and sequence analysis of its complete genome and plasmidome. Int. J. Mol. Sci. 2023, 24, 605. [Google Scholar] [CrossRef]
- Hyink, O.; Balakrishnan, M.; Tagg, J.R. Streptococcus rattus strain BHT produces both a class I two-component lantibiotic and a class II bacteriocin. FEMS Microbiol. Lett. 2005, 252, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.H.; Bakkes, P.J.; Lubelski, J.; Agustiandari, H.; Kuipers, O.P.; Driessen, A.J. The ABC-type multidrug resistance transporter LmrCD is responsible for an extrusion-based mechanism of bile acid resistance in Lactococcus lactis. J. Bacteriol. 2008, 190, 7357–7366. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.C.; Malcata, F.X.; Silva, C.C.G. Lactic acid bacteria in raw-milk cheeses: From starter cultures to probiotic functions. Foods 2022, 11, 2276. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Couderc, C.; Laroute, V.; Coddeville, M.; Caillaud, M.A.; Jard, G.; Raynaud, C.; Cocaign-Bousquet, M.; Tormo, H.; Daveran-Mingot, M.-L. Harnessing diversity of Lactococcus lactis from raw goat milk: Design of an indigenous starter for the production of Rocamadour, a French PDO cheese. Int. J. Food Microbiol. 2022, 379, 109837. [Google Scholar] [CrossRef] [PubMed]
- Fusieger, A.; Martins, M.C.F.; de Freitas, R.; Nero, L.A.; de Carvalho, A.F. Technological properties of Lactococcus lactis subsp. lactis bv. diacetylactis obtained from dairy and non-dairy niches. Braz. J. Microbiol. 2020, 51, 313–321. [Google Scholar] [CrossRef]
- Monnet, C.; Nardi, M.; Hols, P.; Gulea, M.; Corrieu, G.; Monnet, V. Regulation of branched-chain amino acid biosynthesis by alpha-acetolactate decarboxylase in Streptococcus thermophilus. Lett. Appl. Microbiol. 2003, 36, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Redruello, B.; Saidi, Y.; Sampedro, L.; Ladero, V.; Del Rio, B.; Álvarez, M.A. GABA-Producing Lactococcus lactis strains isolated from camel’s milk as starters for the production of GABA-enriched cheese. Foods 2021, 10, 633. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.X.; Chen, Z.; Gu, Z.X.; Han, Y. Isolation of γ-aminobutyric acid-producing bacteria and optimization of fermentative medium. Biochem. Eng. J. 2008, 41, 48–52. [Google Scholar] [CrossRef]
- Laroute, V.; Aubry, N.; Audonnet, M.; Mercier-Bonin, M.; Daveran-Mingot, M.L.; Cocaign-Bousquet, M. Natural diversity of lactococci in γ-aminobutyric acid (GABA) production and genetic and phenotypic determinants. Microb. Cell Fact. 2023, 22, 178. [Google Scholar] [CrossRef] [PubMed]
- Nardi, M.; Renault, P.; Monnet, V. Duplication of the pepF gene and shuffling of DNA fragments on the lactose plasmid of Lactococcus lactis. J. Bacteriol. 1997, 179, 4164–4171. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Duranti, S.; Ruiz, L.; Lugli, G.A.; Tames, H.; Milani, C.; Mancabelli, L.; Mancino, W.; Longhi, G.; Carnevali, L.; Sgoifo, A.; Margolles, A.; Ventura, M.; Ruas-Madiedo, P.; Turroni, F. Bifidobacterium adolescentis as a key member of the human gut microbiota in the production of GABA. Sci. Rep. 2020, 10, 14112. [Google Scholar] [CrossRef] [PubMed]
- Bruinenberg, P.G.; De Vos, W.M.; Siezen, R.J. Deletion of various carboxy-terminal domains of Lactococcus lactis SK11 proteinase: Effects on activity, specificity, and stability of the truncated enzyme. Appl. Environ. Microbiol. 2000, 66, 2859–2865. [Google Scholar] [CrossRef] [PubMed]
- Dandoy, D.; Fremaux, C.; de Frahan, M.H.; Horvath, P.; Boyaval, P.; Hols, P.; Fontaine, L. The fast milk acidifying phenotype of Streptococcus thermophilus can be acquired by natural transformation of the genomic island encoding the cell-envelope proteinase PrtS. Microb. Cell Fact. 2011, 10, S21. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Caplice, E.; McSweeney, A.; Fitzgerald, G.F.; Daly, C. pAMbeta1-associated mobilization of proteinase plasmids from Lactococcus lactis subsp. lactis UC317 and L. lactis subsp. cremoris UC205. Appl. Environ. Microbiol. 1990, 56, 195–201. [Google Scholar] [CrossRef]
- Vos, P.; van Asseldonk, M.; van Jeveren, F.; Siezen, R.; Simons, G.; de Vos, W.M. A maturation protein is essential for production of active forms of Lactococcus lactis SK11 serine proteinase located in or secreted from the cell envelope. J. Bacteriol. 1989, 171, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Juillard, V.; Furlan, S.; Foucaud, C.; Richard, J. Mixed cultures of proteinase-positive and proteinase-negative strains of Lactococcus lactis in milk. J. Dairy Sci. 1996, 79, 964–970. [Google Scholar] [CrossRef]
- Bushin, L.B.; Covington, B.C.; Rued, B.E.; Federle, M.J.; Seyedsayamdost, M.R. Discovery and biosynthesis of streptosactin, a sactipeptide with an alternative topology encoded by commensal bacteria in the human microbiome. J. Am. Chem. Soc. 2020, 142, 16265–16275. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Barba, J.L.; Caballero-Guerrero, B.; Maldonado-Barragán, A.; Jiménez-Díaz, R. Coculture with specific bacteria enhances survival of Lactobacillus plantarum NC8, an autoinducer-regulated bacteriocin producer, in olive fermentations. Food Microbiol. 2010, 27, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, M.; Wladyka, B.; Dubin, G. Exfoliative toxins of Staphylococcus aureus. Toxins (Basel) 2010, 2, 1148–1165. [Google Scholar] [CrossRef]
- Lunde, M.; Aastveit, A.H.; Blatny, J.M.; Nes, I.F. Effects of diverse environmental conditions on ΦLC3 prophage stability in Lactococcus lactis. Appl. Environ, Microbiol. 2005, 71, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Desiere, F.; Lucchini, S.; Canchaya, C.; Ventura, M.; and Brüssow, H. Comparative genomics of phages and prophages in lactic acid bacteria. Antonie van Leeuwenhoek 2002, 82, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; González-Guerra, A.; Vázquez, L.; Fernández-López, R.; Flórez, A.B.; de la Cruz, F.; Mayo, B. Isolation and phenotypic and genomic characterization of Tetragenococcus spp. from two Spanish traditional blue-veined cheeses made of raw milk. Int. J. Food Microbiol. 2022, 371, 109670. [Google Scholar] [CrossRef]
- Lanyi, B. Classical and rapid identification: Methods for medically important bacteria. In Methods in Microbiology, Vol. 19; Colwell, R.R., Grigorova, R., Eds.; Academic Press: New York, USA, 1987; pp. 1–67. [Google Scholar]
- Alegría, A.; González, P.; Delgado, S.; Flórez, A.B. Hernández-Barranco, A.; Rodríguez, A. Mayo, B. Characterisation of the technological behaviour of mixtures of mesophilic lactic acid bacteria isolated from traditional cheeses made of raw milk without added starters. Int. J. Dairy Technol. 2016, 69, 1–13. [Google Scholar] [CrossRef]
- Ziadi, M.; Wathelet, J.P.; Marlier, M.; Hamdi, M.; Thonart, P. Analysis of volatile compounds produced by 2 strains of Lactococcus lactis isolated from leben (Tunisian fermented milk) using solid-phase microextraction-gas chromatography. J. Food Sci. 2008, 73, S247–S252. [Google Scholar] [CrossRef]
- Redruello, B.; Ladero, V.; Cuesta, I.; Álvarez-Buylla, J.R.; Martín, M.C.; Fernández, M.; Álvarez, M.A. A fast, reliable, ultra-high performance liquid chromatography method for the simultaneous determination of amino acids, biogenic amines and ammonium ions in cheese, using diethyl ethoxymethylenemalonate as a derivatising agent. Food Chem. 2013, 139, 1029–1035. [Google Scholar] [CrossRef]
- EFSA FEEDAP Panel. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
Figure 1.
pH (A) and GABA production (B) of L. lactis (32°C) and S. thermophilus (42°C) strains in semi-skim milk supplemented with 10 mM monosodium glutamate (MSG).
Figure 1.
pH (A) and GABA production (B) of L. lactis (32°C) and S. thermophilus (42°C) strains in semi-skim milk supplemented with 10 mM monosodium glutamate (MSG).
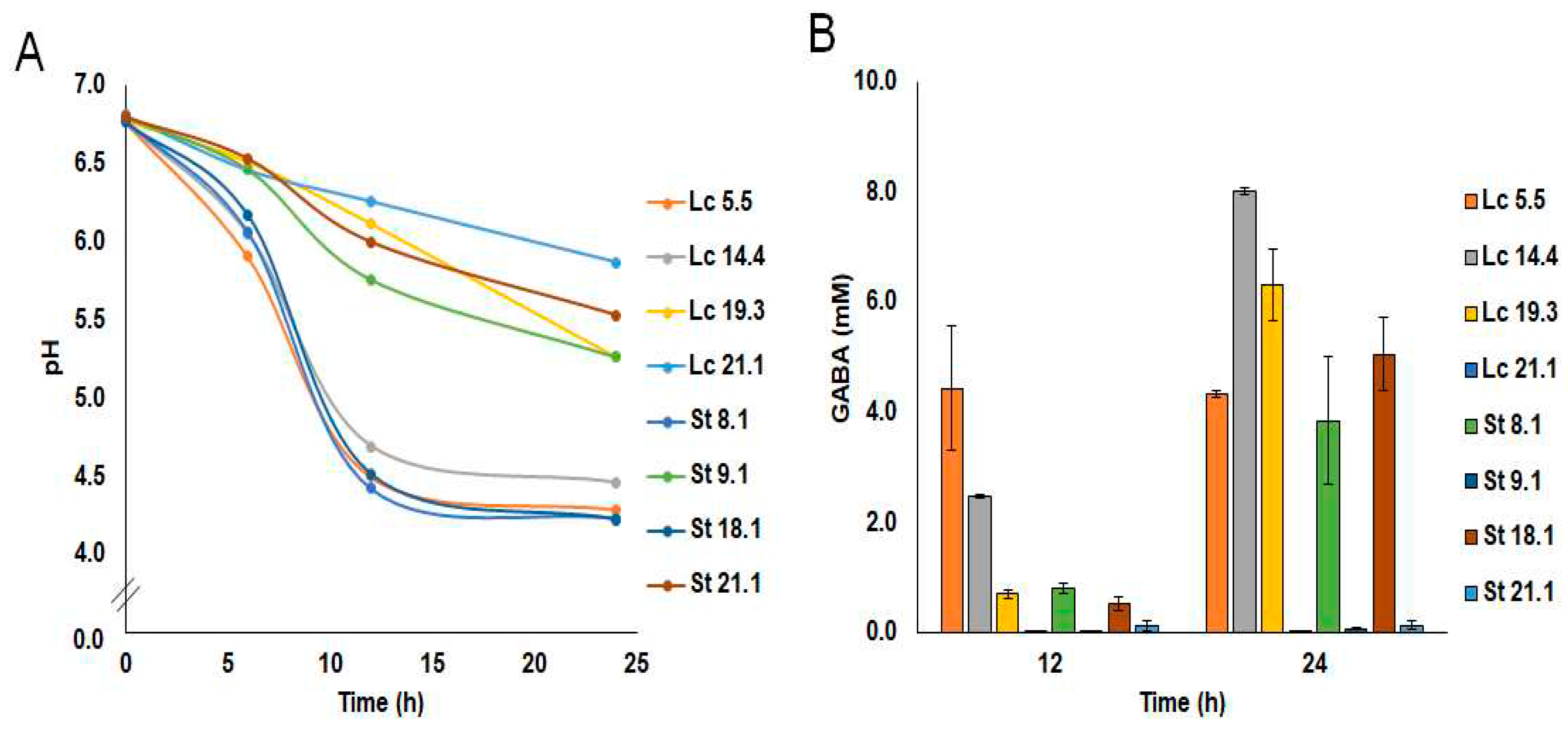
Figure 2.
Genetic organization of the GABA operons found in the genome of Lactococcus lactis (A) and Streptococcus thermophilus (B) strains. Color code: in red, genes coding for the glutamate decarboxylase (gadB); in green, genes encoding the GABA/H+ antiporters (gadC), in purple, genes coding for the regulatory protein (gadR); in yellow, genes coding for transposases; in dark blue, genes encoding plasmid replication functions; in white, genes encoding proteins of other systems or ORFs encoding hypothetical proteins. The GABA operon of L. lactis Lc 14.4 was found to be spread over four different contigs, of which one harbors genes encoding plasmid replication proteins. Dotted lines indicate that contigs extend beyond the depicted area.
Figure 2.
Genetic organization of the GABA operons found in the genome of Lactococcus lactis (A) and Streptococcus thermophilus (B) strains. Color code: in red, genes coding for the glutamate decarboxylase (gadB); in green, genes encoding the GABA/H+ antiporters (gadC), in purple, genes coding for the regulatory protein (gadR); in yellow, genes coding for transposases; in dark blue, genes encoding plasmid replication functions; in white, genes encoding proteins of other systems or ORFs encoding hypothetical proteins. The GABA operon of L. lactis Lc 14.4 was found to be spread over four different contigs, of which one harbors genes encoding plasmid replication proteins. Dotted lines indicate that contigs extend beyond the depicted area.
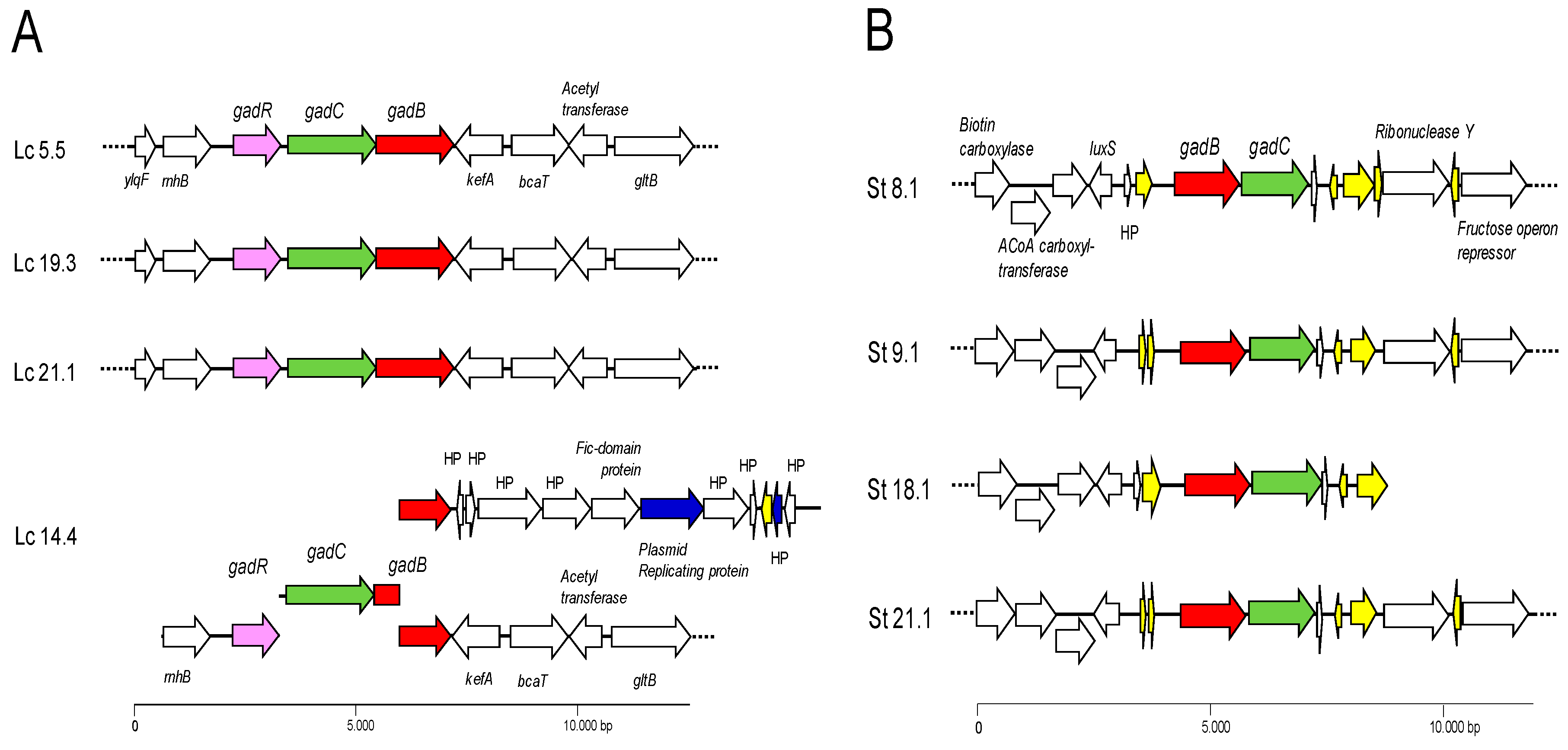

Table 1.
Sugar and organic acid profile of the GABA-producing Lactococcus lactis and Streptococcus thermophilus strains after growth in semi-skim milk.
Table 1.
Sugar and organic acid profile of the GABA-producing Lactococcus lactis and Streptococcus thermophilus strains after growth in semi-skim milk.
| Species/strain | Organic acid sugar a | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lactose | Glucose | Galactose | Orotic acid | Citric acid | Pyruvic acid | Succinic acid | Lactic acid | Formic acid | Acetic acid | Uric acid | Hippuric acid | |
| L. lactis | ||||||||||||
| Lc 5.5 | 6.68±2.3 | 2.3±0.1 | 44.4±12.2 | 9.2±2.5 | 272.5±86.4 | 7.5±1.7 | 2.1±3.0 | 938.2±224.4 | 7.3±1.4 | - | 3.1±1.0 | - |
| Lc 14.4 | 4.49±0.2 | 1.8±0.1 | 51.4±1.5 | 6.0±0.4 | - | 7.4±2.9 | 2.0±2.8 | 826.3±68.1 | 9.8±3.3 | 73.7±19.1 | 2.3±0.1 | 2.3±0.0 |
| Lc 19.3 | 5.36±1.2 | 1.9±0.1 | 59.7±22.0 | 9.4±2.5 | 218.4±47.7 | 2.8±0.8 | - | 405.7±132.6 | - | 6.6±9.4 | 2.5±0.7 | 0.6±0.3 |
| Lc 21.1 | 7.12±2.5 | 2.5±1.3 | 18.4±5.8 | 10.7±2.9 | 21.5±8.3 | 15.1±2.9 | - | 281.5±62.9 | - | 74.2±13.9 | 2.4±0.6 | 3.2±0.9 |
| S. thermophilus | ||||||||||||
| St 8.1 | 5.05±1.1 | 9.1±2.1 | 982.4±129.1 | 10.5±1.1 | 263.2±43.7 | 8.0±0.3 | - | 1173.3±55.5 | - | - | 2.9±0.4 | 2.6±0.5 |
| St 9.1 | 5.84±1.5 | 32.7±7.4 | 391.9±105.2 | 10.2±2.6 | 251.4±65.4 | 1.0±0.1 | - | 464.0±116.2 | - | - | 2.6±0.7 | 2.1±0.4 |
| St 18.1 | 5.07±3.0 | 9.0±5.4 | 972.2±547.8 | 10.8±5.8 | 265.5±147.3 | 8.3±3.4 | - | 1203.6±617.5 | - | - | 3.0±1.7 | 2.6±1.3 |
| St 21.1 | 6.84±2.3 | 77.0±24 | 365.5±105.8 | 11.2±3.1 | 287.7±92.2 | 1.6±0.1 | - | 375.6±90.4 | - | - | 2.6±0.6 | 2.0±0.5 |
| Milk (control) | 6.53±0.3 | 42.2±8.2 | 282.7±152.4 | 8.5±0.1 | 191.8±1.4 | 1.4±1.0 | - | 313.1±184.2 | - | - | 1.8±0.2 | 1.9±0.3 |
a Results expressed as mg per 100 mL of milk, except for lactose (g per 100 mL). -, not detected.
Table 2.
Relative amounts of volatile compounds produced in semi-skim milk by L. lactis and S. thermophilus strains.
Table 2.
Relative amounts of volatile compounds produced in semi-skim milk by L. lactis and S. thermophilus strains.
| Compounda | Control milk | L. lactis | S. thermophilus | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Lc 5.5 | Lc 14.4 | Lc 19.3 | Lc 21.1 | St 8.1 | St 9.1 | St 18.1 | St 21.1 | ||
| Alcohols | |||||||||
| 2-Phenyl ethanol | - | 19.13 | 33.26 | - | - | - | - | - | - |
| Isoamyl alcohol | - | 100.86 | 164.49 | - | - | - | - | - | - |
| Carboxylic acids | |||||||||
| Acetic acid | - | 3.63 | 14.36 | 3.28 | 6.2 | 1.39 | 0.84 | 1.63 | 0.64 |
| Benzoic acid | - | 13.44 | 0.86 | 12.26 | 0.57 | 2.55 | 5.19 | 0.95 | 4.24 |
| Butanoic acid | 0.26 | 4.43 | 3.85 | 4.67 | 2.39 | 6.09 | 5.09 | 6.38 | 4.89 |
| Hexanoic acid | 1.07 | 33.09 | 31.86 | 33.35 | 20.27 | 38.75 | 35.79 | 39.55 | 34.24 |
| Heptanoic acid | - | 0.98 | 1.01 | 1.04 | - | 0.85 | 0.81 | 0.94 | 0.81 |
| Octanoic acid | 1.87 | 35.99 | 37.83 | 36.51 | 30.9 | 41.25 | 40.64 | 42.48 | 39.72 |
| n-Decanoic acid | 2.62 | 20.69 | 21.14 | 19.67 | 18.07 | 21.6 | 20.34 | 22.58 | 20.23 |
| 9-Decenoic acid | - | 2.27 | 2.37 | 2.32 | 1.82 | 2.8 | 2.54 | 2.8 | 2.47 |
| Nonanoic acid | - | 0.48 | 0.71 | 0.7 | 0.56 | 0.63 | 0.76 | 0.84 | 0.73 |
| Dodecanoic acid | 0.52 | 3.62 | 3.48 | 3.11 | 2.73 | 4.38 | 3.64 | 4.09 | 4.05 |
| Tetradecanoic acid | - | - | - | - | - | 1.04 | 0.82 | 0.95 | 0.88 |
| Isovaleric acid | - | 14.99 | 31.97 | 7.96 | - | - | - | - | - |
| Ketones | |||||||||
| Acetoinb | - | - | 17.02 | - | 25.54 | 13.58 | 8.31 | 12.81 | 9.82 |
| 2-Heptanone | 2.67 | - | - | - | - | 3.86 | 3.26 | 4.42 | 3.38 |
| 2-Nonanone | 1.85 | 2.12 | 1.9 | 2.05 | 3.06 | 2.56 | 2.6 | 2.96 | 2.52 |
| 2-Pentadecanone | - | - | 0.55 | - | - | - | - | - | - |
| 2-Tridecanone | 0.37 | 0.52 | 0.58 | 0.51 | 0.47 | 0.56 | 0.65 | 0.66 | 0.65 |
| 2-Undecanone | 0.86 | 1.09 | 1.17 | 1.28 | 1.2 | 1.45 | 1.41 | 1.54 | 1.44 |
| 4-Octanone | - | 40.7 | 79.5 | 20.59 | - | - | - | - | - |
| 2,5-Dimethyl-3-hexanone | - | - | 3.99 | 0.67 | - | - | - | - | - |
| Lactones | |||||||||
| δ-Decalactone | 0.35 | 0.66 | 0.7 | 0.66 | 0.62 | 0.73 | 0.64 | 0.71 | 0.61 |
| γ-Dodecalactone | 0.35 | 0.45 | 0.43 | 0.4 | 0.4 | 0.47 | 0.66 | 0.48 | 0.5 |
| δ-Dodecalactone | 0.44 | 0.69 | 0.71 | 0.63 | 0.61 | 0.83 | 0.74 | 0.78 | 0.76 |
| Other | |||||||||
| Benzaldehyde | 0.60 | 1.49 | 1.63 | 1.35 | 0.72 | 0.77 | - | 0.81 | - |
| Phenyl acetaldehyde | 0.34 | 11.12 | 11.73 | 4.56 | - | - | - | - | |
| 2,3-Heptanedione | - | - | 1.72 | 0.89 | - | - | - | - | - |
| 5-Methyl-2-phenyl-2-hexenal | - | 1.45 | 1.62 | - | - | - | - | - | - |
a Values of the relative area of the chromatograms as compared to that of the styrene internal standard.
Table 3.
General features of the whole genome sequence of GABA-producing L lactis and S. thermophilus strains of this study.
Table 3.
General features of the whole genome sequence of GABA-producing L lactis and S. thermophilus strains of this study.
| Property/protein-encoding genes | Species and strains | |||||||
|---|---|---|---|---|---|---|---|---|
| L. lactis | S. thermophilus | |||||||
| Lc 5.5 | Lc 14.4 | Lc 19.3 | Lc 21.1 | St 8.1 | St 9.1 | St 18.1 | St 21.1 | |
| Genome size (bp) | 2,718,046 | 2,652,084 | 2,575,310 | 2,433,628 | 1,806,293 | 1,761,434 | 1,807,628 | 1,794,253 |
| G+C content | 35.17 | 35.16 | 35.17 | 34.79 | 39.77 | 39.40 | 39.89 | 39.47 |
| No. of contigs | 95 | 73 | 84 | 104 | 46 | 49 | 54 | 45 |
| No. of coding sequences | 2,882 | 2,762 | 2,721 | 2,558 | 2,043 | 1,989 | 2,043 | 2,041 |
| No. of BV-BRC subsystems | 207 | 207 | 208 | 204 | 202 | 202 | 202 | 202 |
| Penicillin binding proteins | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Efflux-related proteins | 28 | 24 | 29 | 27 | 27 | 34 | 28 | 34 |
| Virulence Factors (VFDB/Victors) | 2/8 | 2/9 | 1/8 | 1/7 | 1/39 | 1/41 | 1/39 | 1/41 |
| Antibiotic Resistance (BV-BRC/CARD) | 26/2 | 26/2 | 27/2 | 26/2 | 24/0 | 24/0 | 24/0 | 24/0 |
| Resistance to heavy metals | 1 | 2 | 2 | 1 | 2 | 2 | 2 | 2 |
| rRNA sequences (23S+16S+5S) | 4 (1+1+2) | 4 (1+1+2) | 4 (1+1+2) | 4 (1+1+2) | 3 (1+1+1) | 2 (1+1+0) | 3 (1+1+1) | 2 (1+1+0) |
| tRNA molecules | 52 | 50 | 50 | 51 | 37 | 30 | 35 | 30 |
| Proteases | 12 | 15 | 12 | 15 | 11 | 11 | 11 | 11 |
| Peptidases | 26 | 27 | 27 | 26 | 25 | 23 | 26 | 23 |
| Transposases/integrases/excisionases | 27 | 18 | 24 | 24 | 25 | 24 | 24 | 26 |
| Phage-derived proteins | 333 | 245 | 252 | 225 | 7 | 5 | 6 | 61 |
| Plasmid replication proteins | 13 | 11 | 14 | 12 | 1 | 1 (pUB110-type) | 1 | - |
| CRISPR-Cas loci | 0 | 0 | 0 | 0 | 3 | 3 | 3 | 3 |
| Bacteriocins | Lactococcin B and Q, sactipeptides | Enterolysin A, lactococcin A, B, and Q, sactipetides | 2 x lactococcin B, linaridin | Lactococcin B, sactipeptides | BhtR, BlpD, BlpK, BlpU, streptide | BlpD, BlpK, BlpU, streptide | BlpD, BlpK, BlpU, streptide | BlpD, BlpK, BlpU, streptide |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



