
Submitted:
16 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Heat-stressed lactating dairy cattle exhibit unique metabolic symptoms; many of which are undoubtedly involved in heat-induced subfertility. Because of its known systemic effects, we hypothesized that γ-aminobutyric acid (GABA) participates in the regulation of insulin and progesterone during heat stress. Multiparous lactating Holstein cows (n=6) were studied during four experimental periods: 1) thermoneutral (TN; d 1-5), 2) TN + hyperinsulinemic-hypoglycemic clamp (d 6-10), 3) heat stress (HS; d 16-20), and 4) HS + euglycemic clamp (d 21-25). Blood samples were collected once daily via coccygeal venipuncture into heparinized evacuated tubes. Analysis of GABA concentrations from all four treatment periods yielded no differences. In direct comparison to TN concentrations, plasma GABA tended to be decreased during HS (16.57±2.64 vs 13.87±2.28 ng/ml, respectively, P = 0.06). Both milk production and plasma insulin were moderately correlated with plasma GABA (r=0.35, P<0.01; r = -0.32, P<0.01). Plasma progesterone was correlated with plasma GABA concentrations during TN but not HS. These results are the first to indicate that peripheral GABA could be involved in regulating aspects of production and reproduction during heat stress. More research is needed to determine its precise role(s).
Keywords:
γ-aminobutyric acid
; dairy cow
; heat stress
; insulin
; glucose
; progesterone
; milk production
1. Introduction
γ-Aminobutyric acid (GABA) is the primary inhibitory neurotransmitter in the central nervous system. It is not thought to cross the blood brain barrier [1], but it is also produced by several peripheral systems in quantities comparable to central nervous system (CNS) concentrations and is therefore found in systemic circulation [2]. The two tissues that are best documented as sources of peripheral GABA are ovarian and pancreatic tissues [3-4].
In the pancreas, GABA is secreted by beta cells separately and independently from insulin and stimulates insulin production in a paracrine manner via the GABAA receptor [5-6]. This effect contrasts with the known inhibitory action of GABA in the CNS and is due to the differential expression of chloride cation cotransporters in the two tissues [7]. γ-Aminobutyric acid also has an inhibitory effect on the secretion of glucagon from pancreatic alpha cells via the GABAA receptor and insulin acts in a paracrine manner to upregulate the expression of these receptors on the alpha cell surface [8-9]. Glucagon works counter to insulin when conditions necessitate an increase in blood glucose by stimulating gluconeogenic pathways.
Binding sites for GABA have been documented in the mammalian ovary, in particular on mammalian granulosa cells [10]. Additionally, in preliminary work, inhibition of GABAA type receptors with muscimol was shown to decrease the expression of bone morphogenetic proteins 2 and 6 in granulosa cells [11]. Effects on bone morphogenetic proteins are noteworthy because they are important for the early phases of follicular growth [12]. Inhibition of GABAA type receptors with muscimol also decreased expression of the luteinizing hormone receptor in granulosa cells [11]. Peluso and Pappalardo [13] found that using bicuculline to inhibit GABAA type receptors inhibited progesterone’s ability to prevent insulin-dependent mitosis, as well as apoptosis, of granulosa cells. Taken together, these findings suggest multiple roles for GABA and its receptors in mediating granulosa cell growth and follicular development.
The secretion and actions of GABA may be of particular interest for lactating dairy cows experiencing heat stress. Reproductive performance of dairy cows is known to be hindered during the summer months as a result of heat stress [14-16]. This is most directly observable on farms in the form of depressed conception rates. Successful establishment of pregnancy is a multi-faceted process involving numerous tissues, hormones and signaling pathways. Exposure to heat stress alters many of the processes affected by GABA, including follicular development and dynamics, estrous cycle length, and progesterone production [17-19].
Further evidence for the involvement of GABA in the heat stress response is the fact that lactating dairy cows typically exhibit a state of hyperinsulinemia during heat stress. This increase in circulating insulin is peculiar because it exists concurrently with a reduction in dry matter intake and hypoglycemia [20-21]. It is currently unknown how this altered glycemic state is established or regulated, and what physiological purpose it may have. Perturbations in circulating insulin and glucose are likely involved in the disruption of reproductive processes during heat stress, as both are important for fertility. Furthermore, given its involvement in the regulation of ovarian and pancreatic processes, we hypothesized that GABA is involved in the upregulation of circulating insulin concentrations commonly observed during periods of heat stress, and that GABA may be related to circulating progesterone concentrations.
The objectives of this study were to determine the effect of heat stress on plasma GABA concentrations in multiparous lactating Holstein cows and to investigate the relationships between plasma GABA, progesterone, and glucose and insulin, alongside other physiological parameters.
2. Materials and Methods
2.1. IACUC Statement
All animal procedures performed during this experiment were approved by the Virginia Tech Institutional Animal Care and Use Committee.
2.2. Animals and Experimental Design
This experiment was conducted in two replicates using cows from the Virginia Tech Dairy Science Complex. These were a subset of an on-going experiment described in [22]. Six multiparous lactating Holstein cows (63.33±2.35 DIM, 3.17±0.40 lactations) were paired and randomly placed in one of two environmentally controlled rooms for four experimental periods: 1) thermoneutral (TN; d 1-5), 2) TN + hyperinsulinemic-hypoglycemic clamp (HHC; d 6-10), 3) HS (d 16-20), and 4) HS + euglycemic clamp (EC; d 21-25) as previously described [22]. Two transition periods were instated to allow the cows time to acclimate to the experimental conditions and husbandry procedures. The first was applied for three days before the start of the experiment (d -3 to -1). The second acclimation period allowed acclimation to heat stress and was applied for five days after the termination of the HHC period. A diagram of the treatment protocol can be found in Figure 1. Cows were milked twice daily (1230 and 0030h), and milk yields were recorded at each milking. All cows were individually fed a TMR (Total Mixed Rations) twice daily (1230 and 0030h) and refusals were weighed before the 1230 feeding each day. Prior to placing cows into the environmentally controlled rooms, two indwelling jugular catheters were placed in each cow on either side of the neck (four total per cow) for infusion of insulin and dextrose, as well as for blood sampling. These catheters were flushed with heparinized saline at least once daily to maintain patency.
Rectal temperatures (°C) and respiration rates (breaths/min) were monitored throughout the experiment. Each were measured once daily (1230 h) during the TN and HHC periods and 3 times daily (0030, 0900, and 1230 h) during the HS and EC periods.
2.3. Thermoneutral and Hyperinsulinemic-Hypoglycemic Clamp Periods
The TN and HHC periods were both performed under thermoneutral conditions for lactating dairy cattle (THI = 67.1±0.26 for TN and 64.1±0.36 for HHC). No other treatments were applied during the TN period. The HHC period began on day 6 of the experiment and lasted for 96 hours. Hand-held blood glucose monitors were used to monitor blood glucose concentrations throughout this period (Contour Next EZ, Ascensia Diabetes Care US, Inc., Parsippany, NJ). Before beginning glucose and insulin infusions, 6 baseline blood glucose measurements were recorded for each cow over a 12-hour period to calculate an appropriate blood glucose baseline for each cow.
The insulin infusate was prepared by adding purified bovine insulin (I5500, Sigma-Aldrich, Inc., St. Louis, MO) dissolved in 0.01 M HCl to a saline solution containing a volume of each cow’s plasma equivalent to 1.25% of the solution. This insulin solution was infused at a rate of 0.3 ug/kg of BW/h using syringe pumps (Genie Plus, Kent Scientific, Torrington, CT). A 50% dextrose solution (Nova-Tech, Inc., Grand Island, NE) was simultaneously infused (Plum A+, Hospira, Lake Forest, IL). The rate of glucose infusion varied according to each cow’s baseline blood glucose measurements in order to maintain a hypoglycemic state (90±10% of the average of the baseline blood glucose readings for each cow). Blood glucose was measured every 5 minutes at the onset of the infusions until the glucose infusion rate and blood glucose measurements stabilized, at which point blood glucose was measured at intervals of every 30 minutes or less. For cows that did not respond to the low-dose insulin infusion, the infusion rate of insulin was slowly increased to a maximum of 0.7 ug/kg/ of BW/h until blood glucose fell within the desired range.
2.4. Heat Stress and Euglycemic Clamp
As previously mentioned, heat stress conditions were applied for a five-day acclimation period before formally beginning the HS period. Environmental conditions sufficient to induce heat stress in lactating dairy cows continued to be applied throughout the HS and EC periods (THI = 72.2±0.36 for HS and 71.1±0.36 for EC). Heat stress conditions were set to vary diurnally to mimic a summer day. The room temperature reached a maximum of 32.2°C during the afternoon, after which it cooled slowly until it reached a minimum temperature of 21.1°C overnight. In cases where a cow failed to acclimate acceptably to the HS treatment (multiple and repeated intervention when rectal temperature was 40.5°C or higher), the maximum room temperature was lowered to 29.4°C.
No other treatments were applied during the HS period. The EC period commenced on day 21 of the experiment, and the related infusion lasted 96 hours. Six baseline blood glucose measurements for the HS period were again recorded over 12 hours prior to the start of the EC period. Glucose was infused (Plum A+, Hospira, Lake Forest, IL) at variable rates to return blood glucose concentrations to 100±10% of the average TN baseline measurement for each cow. Blood glucose was initially measured every five minutes until infusion rates stabilized, at which point it was measured at intervals no greater than every 60 minutes.
2.5. Blood Sampling
Blood samples were collected once daily (1300h) via coccygeal venipuncture into heparinized evacuated tubes. Blood samples were kept cold (refrigerated) until centrifugation at 2500 RPM for 20 minutes. Once centrifuged, plasma for GABA concentration analysis was collected and stored at -80°C. Plasma for insulin concentration analysis was collected and stored at -20°C.
2.6. Hormone and Metabolite Analysis
Plasma GABA concentrations were determined using a competitive inhibition ELISA (LSBio Gamma-Aminobutyric Acid (Competitive EIA) ELISA Kit - LS-F10676) validated for use in bovine samples. The inter- and intra-assay coefficients of variation were 13.9% and 6.3% respectively. Plasma insulin concentrations were determined using a sandwich ELISA (Mercodia Bovine Insulin ELISA (10-1201-01)). The inter- and intra-assay coefficients of variation were 11.1% and 5.9% respectively. All samples and standards were run in triplicate. GraphPad Prism 9.0.0 was used to interpolate the standard curves to determine the concentrations in the individual samples. Blood glucose concentrations were determined at the time of blood collection using handheld glucometers as previously stated (Contour Next EZ). Circulating concentrations of progesterone were analyzed by radioimmunoassay (RIA) using methodology described by [23]. Intra- and interassay coefficients of variation for progesterone assays were 3.5% and 8.4%, respectively,
2.7. Statistical Analyses
The experiment was conducted so that each cow served as its own control. The order of treatment administration could not be randomized due to the nature of the treatments. All models were fit as generalized linear mixed models using PROC GLIMMIX (SAS 9.4). In all models, the period and day of period were included as fixed effects and day of period was removed from the model when it was not significant. The interaction of the period and time was not able to be modeled due to sample size limitations. The random effects were modeled hierarchically to account for the multiple subject effects. A random intercept for each cow was included to account for dependence among all observations within a cow. The autocorrelation of observations from each cow within a period was modeled as an R-side effect with a first-order autoregressive covariance structure. This was necessary because cows served as their own controls. For analyses of plasma GABA concentration, a GLMM with a normal distribution and logarithmic link function was used. For analyses of plasma progesterone, a GLMM with a gamma distribution and logarithmic link function was used. The gamma distribution was selected due to the highly skewed nature of the progesterone data. For analyses of plasma insulin, rectal temperature, and respiration rate, a normal distribution and identity link function were used. This is identical to using a linear mixed model since the model assumptions were met for plasma insulin. All post hoc pairwise comparisons were adjusted using the Tukey-Kramer adjustment for multiple comparisons.
Additionally, RCORR (SAS 9.4) was used to evaluate correlational relationships between plasma GABA concentrations and the other production parameters, and to evaluate the relationship between the change in plasma GABA concentrations from thermoneutral to heat stress conditions alongside the corresponding change in circulating glucose and insulin concentrations.
Because plasma GABA concentrations in heat stressed dairy cattle have never been compared to a thermoneutral control, two versions of the data were used for analysis. The first version included data from all four treatment periods. The second analysis excluded data from HHC and EC periods so that GABA concentrations during TN were directly compared to concentrations during HS. The results of each analysis are reported explicitly. The SAS code for all analyses can be found in the supplementary material.
3. Results
Respiration rates differed between the TN and HS periods as expected (TN = 58.1 ± 3.7 bpm, HS = 71.8 ± 3.8 bpm, p = 0.0005). Rectal temperatures also differed between the TN and HS periods as expected (TN = 38.9 ± 0.2 C, HS = 39.7 ± 0.2 C, p = 0.01).
Plasma insulin differed by period both in the analyses with and without the HHC and EC periods included. In the analysis with all four periods included, plasma insulin concentrations during the HS and EC periods were greater than that during the TN period (Table 1). Recall that insulin and glucose were experimentally manipulated during the HHC, and thus, insulin tended to be elevated during the HHC compared to TN (Table 2, p = 0.067). In the analysis including only data from the TN and HS periods, plasma insulin was increased during the HS period (Table 1).
In the analysis including all four experimental periods, no significant period differences were found among mean plasma GABA concentrations (Figure 2). In the analysis that directly compared the TN period to HS without including either clamp period, plasma GABA concentrations during HS tended to be decreased during the TN period (Figure 3).
The models of daily plasma progesterone measurements by period with and without the HHC and EC periods yielded no significant differences (Table 2).
A correlational analysis was also conducted on this data set (Figure 4) and plasma GABA concentrations were moderately correlated with daily milk production (r = 0.35, p = 0.0002; Figure 5) and daily plasma insulin measurements (r = -0.32, p = 0.0009; Figure 6). None of the other variables tested (plasma progesterone, daily feed intake, average daily rectal temperature, or average daily respiration rate) were significantly correlated to daily plasma GABA concentrations (Figure 4). As expected for heat-stressed dairy cattle, average daily rectal temperatures and respiration rates were negatively correlated with feed intake (r = -0.48, p < 0.0001 and r = -0.36, p = 0.0002, respectively; Figure 4). Plasma progesterone was correlated with daily feed intake (r = -0.41, p < 0.0001), average daily respiration rate (r = 0.24, p = 0.0141), and average daily rectal temperature (r = 0.40, p < 0.0001) (Figure 4). A separate correlational analysis of the available blood glucose measurements with the corresponding plasma GABA measurements at the end of the TN and HS periods yielded a strong negative correlation (r = -0.63, p = 0.0273; Figure 7). It is worth noting that the sample size available for this analysis was small, however.
Correlational analyses were also conducted separately for the TN and HS periods to identify changes in relationships between the physiological variables. In this analysis, it was found that the correlation between plasma GABA and daily milk production was greater during HS than TN (TN: r = 0.333, p = 0.08; HS: r = 0.583, p = 0.003). Interestingly, plasma progesterone was moderately correlated with plasma GABA during TN, but not during HS (TN: r = 0.453, p = 0.01367; HS: r = -0.280, p = 0.1858). The full results of this analysis can be found in the supplementary material (Figure S1)
4. Discussion
Undeniably, reproductive- and lactation-based responses to heat stress in cattle are quite complex, and there is no one solution capable of improving productivity and fertility in heat-stressed cattle. For these reasons, it is necessary to develop a complete understanding of the physiological adaptations that occur when cattle are exposed to heat stress. The hyperinsulinemic-hypoglycemic state that is typically observed in heat-stressed cattle is one such adaptation that requires a deeper understanding. In particular, the increase in circulating insulin concentrations is counterintuitive, as cattle exposed to heat stress reduce their dry matter intake. Unfortunately, the mechanism(s) driving the increase in circulating insulin concentrations have not been elucidated.
As expected, circulating insulin concentrations in the current experiment were greater during the HS period than during the TN period. Insulin is known to have positive effects on postpartum follicular development [24-25], but damages oocyte quality. Diets designed to increase circulating insulin concentrations in cows and heifers decrease in vitro blastocyst development [26-28]. Furthermore, states of hyperinsulinemia are known to decrease oocyte quality in cattle, mice and humans [26, 29-31]. These data suggest that insulin has opposing effects on two different aspects of fertility: positive effects on postpartum follicular development, and negative effects on the developmental competence of oocytes. Garnsworthy and colleagues [25] exploited these opposing effects by target-feeding dairy cattle for high or low circulating insulin concentrations during the postpartum period. As expected, pregnancy rates were immensely improved by feeding an insulin-stimulating diet until the first postpartum ovulation and then switching cows to a lower insulin diet (high-low treatment). At 120 d postpartum, 60% of these cows were pregnant compared to only 27% of the cows on other dietary treatments (high-high, low-high or low-low diets). These results provide striking evidence for opposing effects of insulin on follicular development and oocyte quality.
We hypothesized GABA is one of the factors involved in the regulation of insulin during heat stress. Concentrations of GABA in the plasma of dairy cattle and the physiological consequences of changes therein are not well characterized. The majority of published studies pertaining to GABA in lactating dairy cows are based on supplementing a rumen-protected form of GABA to heat stressed cows. Dietary supplementation of GABA in lactating dairy cows has resulted in increased feed intake, decreased rectal temperatures, modest increases in milk production, and various changes in immune molecule concentrations [32-33]. A similar study performed under thermoneutral conditions also documented increased feed intake and modest increases in milk production [34]. In the current study, we did not observe a significant correlation between plasma GABA and feed intake. This may imply that increased feed intake following dietary supplementation of GABA is a pharmacological effect of exogenously increasing plasma GABA. Plasma GABA was positively correlated with daily milk production in the current study, however, which is consistent with the results of the GABA supplementation studies demonstrating modestly increased milk production.
Overall, plasma GABA concentrations tended to be decreased during the HS period compared to during TN. Knowing that insulin increased during heat stress, and GABA is generally stimulatory for insulin secretion, this result was unexpected. We expected plasma GABA to increase during heat stress, which would suggest a role for GABA in the stimulation of insulin secretion during heat stress. Upon examining the GABA concentrations of individual cows, we found that four out of six cows enrolled in this experiment experienced a decrease in plasma GABA during the HS period (in comparison to TN). Interestingly, the two cows that responded with increased plasma GABA also experienced a more extreme hypoglycemic response during heat stress, based on the percent change in blood glucose. This response would be consistent with the expected increase in circulating insulin in response to greater GABA concentrations and highlights the variability in individual cow responses to heat stress. Clearly, more work is needed to further explore the relationship between endogenous circulating GABA and insulin in lactating dairy cattle.
Although it was not the expected result, the overall reduction in plasma GABA during heat stress could be indicative of an increase in receptor populations and, thus, greater uptake by the pancreas and other GABA-responsive tissues. Previous work demonstrated that GABAA receptor subunits are not found in substantial quantities on the surface of beta cells, which could suggest that GABA does not have much impact on the release of insulin from the pancreas at physiological concentrations (Wendt et al., 2004). If heat stress causes an increase in receptor numbers within the pancreas, the increased sensitivity to available GABA could be a mechanism driving the increase in insulin during heat stress. In the current study, plasma insulin was weakly correlated with plasma GABA. This finding provides further evidence that plasma GABA may not be representative of GABA activity and/or concentrations within the pancreas. This makes sense if most of the GABA produced in the pancreas is used locally in an autocrine or paracrine fashion as is widely considered to be the case [35]. Unfortunately, there was no opportunity to collect pancreatic tissue for analysis of receptor populations because the cows enrolled in this study were not sacrificed. Future studies are needed for evaluation of receptor populations in the pancreas and other GABA-responsive tissues during heat stress.
Several studies have demonstrated that perfusing the pancreatic cells with GABA or GABAA receptor agonists does impact the release of insulin from the pancreas [36-38] and Feng and co-workers [39] showed that administering GABA to mice treated with streptozotocin to induce type I diabetes largely restored insulin levels. Taking these into account, there is reason to think that alterations in plasma GABA levels, no matter their origin, should impact insulin secretion from the pancreas. However, the correlational analysis of plasma GABA and plasma insulin presented in this work suggests that the impact may be much smaller than that of other endocrine or nervous changes yet to be investigated.
In the case that plasma GABA is directly representative of pancreatic GABA, the lack of substantial correlation between plasma insulin and plasma GABA supports findings from Braun and co-workers [6] that the release of GABA from the pancreas is independent of the release of insulin both temporally and spatially. Some studies have implicated GABA in the insulin production process via its ability to increase beta cell mass rather than beta cell action [40-41]. Investigating changes in beta cell mass and GABAA receptor expression on the beta cell surface during heat stress in lactating cows would provide more clarification regarding whether changes in plasma GABA are indicative of an effect on insulin production.
The relationship between blood glucose and plasma GABA observed in this study is of particular interest going forward. While the precise relationship between GABA and pancreatic beta cells is somewhat unclear, there is substantial unified evidence that GABAA receptors exist on the surface of alpha cells and allow GABA to mediate an inhibitory effect on glucagon release [8-9, 42]. Additionally, Xu and co-workers [9] provided evidence that insulin induces GABAA receptor translocation to the alpha cell membrane and that this is the mechanism by which insulin can suppress glucagon release. Therefore, the state of heat-stress-induced hyperinsulinemia may increase the GABAA receptor density on the alpha cell surface. This mechanism would also explain why the two cows with the greatest decrease in plasma GABA during heat stress were also those cows who experienced increased plasma glucose during heat stress, since the reduced GABA concentrations could be related to increased utilization rather than decreases production.
Interactions between GABA and progesterone are bidirectional and highly dependent upon the tissues and systems of interest. These interactions have been implicated in normal reproductive processes such as regulation of LH secretion [43-44] and sperm hyperactivation [45], as well as a number of maladies, including premenstrual dysphoric disorder, peripartum depression, catamenial epilepsy and polycystic ovary syndrome [46-49]. While the effects of GABA on reproductive processes have been best defined within the central nervous system, some direct actions on female reproductive tissues have been described as well. As previously mentioned, GABA and its binding sites are found in substantial quantities within the ovary [10]. In vitro treatment of luteal cells with GABA has yielded mixed results, with some finding no effect [50] and others finding that GABA reduces luteal-cell progesterone production [51]. Perhaps more importantly, progesterone and GABA receptor agonists are thought to work cooperatively to enhance oviductal cilia activity that is necessary for gamete transport [52]. In the current study, plasma progesterone was positively correlated with plasma GABA in thermoneutral conditions, which is consistent with cooperative effects to promote fertility. This relationship was not present during heat stress, however, indicating a disruption in any concerted actions.
Even though daily plasma GABA concentrations were not related to feed intake, they were correlated with daily milk production. This correlation was especially pronounced during heat stress. Limited research is available regarding the action of GABA within mammary tissue in mammals and none exists that is specific to bovines of any kind. The available research predominantly pertains to the role of GABA in breast cancer metastasis [53-54] or to regulation of prolactin secretion by GABA in the hypothalamus and pituitary gland [55-56]. Because GABA does not cross the blood brain barrier, more research is necessary to determine if GABA is produced within mammary tissue and to understand if it is capable of directly affecting the bovine mammary gland or if it acts elsewhere to influence milk production.
5. Conclusions
The results of this experiment provide initial information regarding relationships between plasma GABA and pertinent physiological parameters in heat stressed dairy cows. Additionally, this study was the first to determine how plasma GABA changes during heat stress compared to a thermoneutral control and demonstrates a tendency for plasma GABA to be reduced during heat stress. Taken together, the results provide compelling support for the further investigation of the role of GABA in the heat stress response in lactating dairy cows regarding the altered glycemic state, fertility and overall productivity during heat stress.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. Correlograms showing the correlations between plasma ɣ-aminobutyric acid (GABA) concentration, plasma progesterone (P4) concentration, daily feed intake, average daily respiration rates (RR), average daily rectal temperatures (RT), and daily milk production in kilograms, and plasma insulin concentration for the TN and HS periods separately. The bottom half of the graph shows scatterplots of each variable against the others, and the top half shows the Pearson correlation coefficient for each pairing. A correlation value marked with * shows that the p-value is <0.05; *** shows that the p-value is <0.01. The diagonal shows a density plot representative of the distribution of each variable.
Author Contributions
Conceptualization, A.G.A. and M.L.R.; Methodology, A.G.A. and M.L.R.; Validation, A.G.A., G.A.P. and M.L.R.; Formal Analysis, A.G.A., J.W.S., G.A.P. and M.L.R.; Investigation, A.G.A., J.W.S., M.K.H.B., G.A.P. and M.L.R.; Resources, A.G.A., J.W.S., M.K.H.B., G.A.P. and M.L.R.; Data Curation, A.G.A., J.W.S., M.K.H.B., G.A.P. and M.L.R.; Writing – Original Draft Preparation, A.G.A. and M.L.R.; Writing – Review & Editing, A.G.A., J.W.S., M.K.H.B., G.A.P. and M.L.R.; Visualization, A.G.A. and M.L.R.; Supervision, G.A.P. and M.L.R.; Project Administration, A.G.A., J.W.S., M.K.H.B., G.A.P. and M.L.R.; Funding Acquisition, M.L.R.
Funding
The authors gratefully acknowledge funding of this work by the Agriculture and Food Research Initiative: Competitive Grant no. 2016-67015-24897 from the USDA National Institute of Food and Agriculture (Washington, DC). Additional support was provided by the Virginia Agricultural Experiment Station (Blacksburg) and the Hatch Program of the National Institute of Food and Agriculture, USDA.
Institutional Review Board Statement
All animal procedures performed during this experiment were approved by the Virginia Tech Institutional Animal Care and Use Committee (protocol number 19-004, approved 09-03-19).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Many thanks to the staff and students from the Virginia Tech Dairy Science Complex and the School of Animal Sciences (formerly the Department of Animal and Poultry Sciences and the Department of Dairy Science).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kuriyama, K.; Sze, P. Y. Blood-brain barrier to H3-gamma-aminobutyric acid in normal and amino oxyacetic acid-treated animals. Neuropharmacology 1971, 10, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Erdö, S.L.; Wolff, J.R. γ-Aminobutyric Acid Outside the Mammalian Brain. J Neurochem 1990, 54, 363–72. [Google Scholar] [CrossRef]
- del Rio, R.M; Caballero, A.L. Presence of γ-Aminobutyric Acid in Rat Ovary. Journal of Neurochemistry 1980, 34, 1584–1586. [Google Scholar] [CrossRef]
- Erdö, S.L.; Rosdy, B.; Szporny, L. Higher GABA Concentrations in Fallopian Tube Than in Brain of the Rat. Journal of Neurochemistry 1982, 38, 1174–1176. [Google Scholar] [CrossRef]
- Kawai, K.; Unger, R.H. Effects of γ -Aminobutyric Acid on Insulin, Glucagon, and Somatostatin Release from Isolated Perfused Dog Pancreas. Endocrinology 1983, 113, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Wendt, A.; Birnir, B.; Broman, J.; Eliasson, L.; Galvanovskis, J.; Gromada, J.; Mulder, H.; Rorsman, P. Regulated Exocytosis of GABA-containing Synaptic-like Microvesicles in Pancreatic β-cells. Journal of General Physiology 2004, 123, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Kursan, S.; McMillen, T.S.; Beesetty, P.; Dias-Junior, E.; Almutairi, M.M.; Sajib, A.A.; Kozak, J.A.; Aguilar-Bryan, L.; di Fulvio, M. The neuronal K+Cl- co-transporter 2 (Slc12a5) modulates insulin secretion. Scientific Reports 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Wendt, A.; Birnir, B.; Buschard, K.; Gromada, J.; Salehi, A.; Sewing, S.; Rorsman, P.; Braun, M. Glucose Inhibition of Glucagon Secretion from Rat α-Cells Is Mediated by GABA Released from Neighboring β-Cells. Diabetes 2004, 53, 1038–1045. [Google Scholar] [CrossRef]
- Xu, E.; Kumar, M.; Zhang, Y.; Ju,W. ; Obata,T.; Zhang, N.; Liu, S.; Wendt, A.; Deng, S.; Ebina, Y.; Wheeler, M.B.; Braun, M.; Wang, Q. Intra-islet insulin suppresses glucagon release via GABA-GABAA receptor system. Cell Metabolism 2006, 3, 47–58. [Google Scholar] [CrossRef]
- Schaeffer, J. M.; Hsueh, A. J. W. Identification of gamma-aminobutyric acid and its binding sites in the rat ovary. Life Sciences 1982, 30, 1599–1604. [Google Scholar] [CrossRef]
- Danzy, L.; Hou, L.; Yalcinkaya, T.M. Bone morphogenetic proteins (BMP)-2 and -6 are potential targets of gaba-A receptor activation in human luteinized granulosa cells (GC). Fertility and Sterility 2011, 96. [Google Scholar] [CrossRef]
- Yoshino, O.; Shi, J.; Osuga, Y.; Harada, M.; Nishii, O.; Yano, T.; Taketani, Y. The function of bone morphogenetic proteins in the human ovary. Reprod Med Biol 2011, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Pappalardo, A. Progesterone Mediates its Anti-Mitogenic and Anti-Apoptotic Actions in Rat Granulosa Cells Through a Progesterone-Binding Protein With Gamma Aminobutyric AcidA Receptor-Like Features. Biology of Reproduction 1998, 58, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.E.; Halbach, T.J.; Armstrong, D.V. Season and Lactation Number Effects on Milk Production and Reproduction of Dairy Cattle in Arizona. Journal of Dairy Science 1992, 75, 2976–2983. [Google Scholar] [CrossRef] [PubMed]
- De Rensis, F.; Scaramuzzi, R.J. Heat stress and seasonal effects on reproduction in the dairy cow—a review. Theriogenology 2003, 60, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M. L. Review: Reproductive consequences of whole-body adaptations of dairy cattle to heat stress. Animal 2023, 17, 100847. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Thatcher, W.W.; Badinga, L.; Savio, J.D.; Meidan, R.; Lew, B.J.; Braw-Tal, R.; Berman, A. Effect of Heat Stress on Follicular Development during the Estrous Cycle in Lactating Dairy Cattle’. Biol Reprod 1995, 52, 1106–13. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.J. , Marion, R.S.; Spain, J.N.; Spiers, D.E.; Keisler, D.H.; Lucy, M.C. Effects of Controlled Heat Stress on Ovarian Function of Dairy Cattle. 1. Lactating Cows. Journal of Dairy Science 1998, 81, 2124–2131. [Google Scholar] [CrossRef]
- Roth, Z.; Meidan, R.; Braw-Tal, R.; Wolfenson, D. Immediate and delayed effects of heat stress on follicular development and its association with plasma FSH and inhibin concentration in cows. J Reprod Fertil 2000. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L. H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. Journal of Dairy Science 2009, 92, 1986–1997. [Google Scholar] [CrossRef]
- Wheelock, J.B. , Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. Journal of Dairy Science 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Stewart, J. W.; Arneson, A.G.; Byrd, M. K. H.; Negron-Perez, V. M.; Newberne, H. M.; White, R. R.; El-Kadi, S.W.; Ealy, A.D.; Rhoads, R.P.; Rhoads, M. L. Comparison of production-related responses to hyperinsulinemia and hypoglycemia induced by clamp procedures or heat stress of lactating dairy cattle. J Dairy Sci 2022, 105, 8439–8453. [Google Scholar] [CrossRef]
- Engel, C. L.; Patterson, H.H.; Perry, G.A. Effect of dried corn distillers grains plus solubles compared with soybean hulls, in late gestation heifer diets, on animal and reproductive performance. Journal of Animal Science 2008, 86, 1697–1708. [Google Scholar] [CrossRef]
- Gong, J. G. , Lee, W.J.; Garnsworthy, P.C.; Webb, R. Effect of dietary-induced increases in circulating insulin concentrations during the early postpartum period on reproductive function in dairy cows. Reproduction 2002, 123, 419–427. [Google Scholar] [CrossRef]
- Garnsworthy, P. C.; Fouladi-Nashta, A. A.; Mann, G. E.; Sinclair, K.D.; Webb, R. Effect of dietary-induced changes in plasma insulin concentrations during the early post partum period on pregnancy rate in dairy cows. Reproduction 2009, 137, 759–768. [Google Scholar] [CrossRef]
- Adamiak, S. J.; Mackie, K.; Watt, R.G.; Webb, R.; Sinclair, K.D. Impact of nutrition on oocyte quality: cumulative effects of body composition and diet leading to hyperinsulinemia in cattle. Biol Reprod 2005, 73, 918–926. [Google Scholar] [CrossRef]
- Fouladi-Nashta, A. A.; Gutierrez, C.G.; Garnsworthy, P.C.; Webb, R. Effects of dietary carbohydrate source on oocyte/embryo quality and development in high-yielding, lactating dairy cattle. Biol Reprod 2005, 135–136. [Google Scholar]
- Adamiak, S. J.; Powell, K.; Rooke, J.A.; Webb, R.; Sinclair, K.D. Body composition, dietary carbohydrates and fatty acids determine post-fertilisation development of bovine oocytes in vitro. Reproduction 2006, 131, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, E. , Pavone, M. E.; Hirshfeld-Cytron, J.E. Metabolic syndrome and oocyte quality. Trends Endocrinol Metab 2011, 22, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Feng, H.L. Extra- and intra-ovarian factors in polycystic ovary syndrome: impact on oocyte maturation and embryo developmental competence. Human reproduction update 2011, 17, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Ou, X. H.; Li, S.; Wang, Z.B.; Li, M.; Quan, S.; Xing, F.; Guo, L.; Chao, S.B.; Chen, Z.; Liang, X.W.; Hou, Y.; Schatten, H.; Sun, Q.Y. Maternal insulin resistance causes oxidative stress and mitochondrial dysfunction in mouse oocytes. Hum Reprod 2012, 27, 2130–2145. [Google Scholar] [CrossRef]
- Cheng, J.B.; Bu, D.P.; Wang, J.Q.; Sun, X.Z.; Pan, L.; Zhou, L.Y.; Liu, W. Effects of rumen-protected γ-aminobutyric acid on performance and nutrient digestibility in heat-stressed dairy cows. Journal of Dairy Science 2014, 97, 5599–5607. [Google Scholar] [CrossRef]
- Cheng, J.; Zheng, N.; Sun, .; Li, S.; Wang, J.; Zhang,Y. Feeding rumen-protected gamma-aminobutyric acid enhances the immune response and antioxidant status of heat-stressed lactating dairy cows. Journal of Thermal Biology 2016, 60, 103–108. [CrossRef] [PubMed]
- Wang, D.M.; Wang, C.; Liu, H.Y.; Liu, J.X.; Ferguson, J.D. Effects of rumen-protected γ-aminobutyric acid on feed intake, lactation performance, and antioxidative status in early lactating dairy cows. Journal of Dairy Science 2013, 96, 96:3222–3227. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, A. Paracrine and autocrine interactions in the human islet: More than meets the eye. Seminars in Cell and Developmental Biology 2013, 24, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Kumar, M.; Zhang, Y.; Gyulkhandanyan, A.; Xiang, Y.Y.; Ye, B.; Perrella, J.; Hyder, A.; Zhang, N.; Wheeler, M.; Lu, W.Y.; Wang, Q. Gamma-aminobutyric acid up- and downregulates insulin secretion from beta cells in concert with changes in glucose concentration. Diabetologia 2006, 49, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Delgado, J.; Braun, M.; Hernández-Fisac, I.; Martín-Del-Río, R.; Tamarit-Rodriguez, J. ; Glucose promotion of GABA metabolism contributes to the stimulation of insulin secretion in β-cells. Biochemical Journal 2010, 431, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P. , Wang, S.; Liu, S.; Xiang, Y.Y.; Lu, W.Y.; Wang, Q. GABA Coordinates with Insulin in Regulating Secretory Function in Pancreatic INS-1 β-Cells. PLoS ONE 2011, 6, e26225. [Google Scholar] [CrossRef] [PubMed]
- Feng, A.L.; Xiang, Y.Y.; Gui, L.; Kaltsidis, G.; Feng, Q.; Lu, W.Y. Paracrine GABA and insulin regulate pancreatic alpha cell proliferation in a mouse model of type 1 diabetes. Diabetologia 2017, 60, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Purwana, I.; Zheng, J.; Li, X.; Deurloo, M.; Son, D.O.; Zhang, Z.; Liang, C.; Shen, E.; Tadkase, A.; Feng, Z.P.; Li, Y.; Hasilo, C.; Paraskevas, S.; Bortell, R.; Greiner, D.L.; Atkinson, M.; Prud’homme, G.J.; Wang, Q. GABA promotes human β-cell proliferation and modulates glucose homeostasis. Diabetes 2014, 63, 4197–4205. [Google Scholar] [CrossRef]
- Untereiner, A.; Abdo, S.; Bhattacharjee, A.; Gohil, H.; Pourasgari, F.; Ibeh, N.; Lai, M.; Batchuluun, B.; Wong, A.; Khuu, N.; Liu, Y.; al Rijjal, D.; Winegarden, N.; Virtanen, C.; Orser, B. A.; Cabrera, O.; Varga, G.; Rocheleau, J.; Dai, F.F.; Wheeler, M.B. GABA promotes β-cell proliferation, but does not overcome impaired glucose homeostasis associated with diet-induced obesity. The FASEB Journal 2019, 33, 3968–3984. [Google Scholar] [CrossRef] [PubMed]
- Rorsman, P. , Berggren, P.O.; Bokvist, K.; Ericson, H.; Möhler, H.; Östenson, C.G.; Smith, P.A. Glucose-inhibition of glucagon secretion involves activation of GABAA-receptor chloride channels. Nature 1989, 341, 233–236. [Google Scholar] [CrossRef]
- Robinson, J.E.; Kendrick, K.M. Inhibition of Luteinizing Hormone Secretion in the Ewe by Progesterone: Associated Changes in the Release of Gamma-Aminobutyric Acid and Noradrenaline in the Preoptic Area as Measured by Intracranial Microdialysis. Journal of Neuroendocrinology 1992, 4, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; Shen, X.; Ivanova, D.; Lass, G.; He, W.; Chen, Q.; Yu, S.; Wang, Y.; Long, H.; Wang, L.; Lyu, Q.; Kuang, Y.; O’Byrne, K.T. Dynorphin and GABAA Receptor Signaling Contribute to Progesterone’s Inhibition of the LH Surge in Female Mice. Endocrinology 2020, 161, bqaa036. [Google Scholar] [CrossRef]
- Kon, H.; Takei, G. L.; Fujinoki, M.; Shinoda, M. Suppression of Progesterone-enhanced Hyperactivation in Hamster Spermatozoa by γ-aminobutyric Acid. Journal of Reproduction and Development 2014, 60, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Stiernman, L.; Dubol, M.; Comasco, E.; Sundström-Poromaa, I.; Boraxbekk, C-J. ; Johansson, M.; Bixo, M. Emotion-induced brain activation across the menstrual cycle in individuals with premenstrual dysphoric disorder and associations to serum levels of progesterone-derived neurosteroids. Transl Psychiatry 2023, 13. [Google Scholar] [CrossRef]
- Deligiannidis, K.M.; Kroll-Desrosiers, A.R. ; Mo,S. ; Nguyen, H.P.; Svenson, A.; Jaitly, N.; Hall, J.E.; Barton, B.A.; Rothschild, A.J.; Shaffer, S.A. Peripartum neuroactive steroid and γ-aminobutyric acid profiles in women at-risk for postpartum depression, Psychoneuroendocrinology 2016, 70, 98–107. [Google Scholar] [CrossRef]
- Joshi, S.; Kapur, J. Neurosteroid regulation of GABAA receptors: A role in catamenial epilepsy. Brain Research 2019, 1703, 31–40. [Google Scholar] [CrossRef]
- Hedström, H.; Bäckström, T.; Bixo, M.; Nyberg, S.; Wang, M.; Gideonsson, I.; Turkmen, S. Women with polycystic ovary syndrome have elevated serum concentrations of and altered GABAA receptor sensitivity to allopregnanolone. Clin Endocrinol 2015, 83, 643–650. [Google Scholar] [CrossRef]
- Battista, P.J.; Condon, W.A. Serotonin-induced stimulation of progesterone production by cow luteal cells in vitro. Reproduction 1986, 76, 231–238. [Google Scholar] [CrossRef]
- Zhang, H.; Ni, J.; Zhang, W.; Tian, S. J. GABA inhibits progesterone production of luteal cells of rat ovary. Acta physiologica Sinica 2000, 52, 185–187. [Google Scholar]
- Jung, C.; Fernandez-Dueñas, V.; Plata, C.; Garcia-Elias, A.; Ciruela, F.; Fernandez-Fernandez, J.M.; Valverder, M.A. Functional coupling of GABAA/B receptors and the channel TRPV4 mediates rapid progesterone signaling in the oviduct. Science Signaling 2018, 11. [Google Scholar] [CrossRef]
- Opolski, A.; Mazurkiewicz, M.; Wietrzyk, J.; Kleinrok, Z.; Radzikowski, C. The role of GABA-ergic system in human mammary gland pathology and in growth of transplantable murine mammary cancer. Journal of Experimental and Clinical Cancer Research 2000, 19, 383–390. [Google Scholar] [PubMed]
- Sizemore, G.M.; Sizemore, S.T.; Seachrist, D.D.; Keri, R.A. GABA(A) Receptor Pi (GABRP) Stimulates Basal-like Breast Cancer Cell Migration through Activation of Extracellular-regulated Kinase 1/2 (ERK1/2). Molecular Bases of Disease 2014, 289, 24102–24113. [Google Scholar] [CrossRef] [PubMed]
- Lamberts, S.W.J.; MacLeod, R.M. Studies on the Mechanism of the GABA-Mediated Inhibition of Prolactin Secretion. Experimental Biology and Medicine 1978, 158, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Mansky, T.; Mestres-Ventura, P.; Wuttke, W. Involvement of GABA in the feedback action of estradiol on gonadotropin and prolactin release: Hypothalamic GABA and catecholamine turnover rates. Brain Research 1982, 231, 353–364. [Google Scholar] [CrossRef]
Figure 1.
Visual Representation of the Experimental Design. Cows (n=6) were subjected to four experimental periods: 1) thermoneutral (TN; d 1-5), 2) TN + hyperinsulinemic-hypoglycemic clamp (HHC; d 6-10), 3) HS (d 16-20), and 4) HS + euglycemic clamp (EC; d 21-25). Cows acclimated to tie stall housing and experimental procedures from d −3 to −1. Days 11 to 15 of the experiment served as a period for acclimation to HS.
Figure 1.
Visual Representation of the Experimental Design. Cows (n=6) were subjected to four experimental periods: 1) thermoneutral (TN; d 1-5), 2) TN + hyperinsulinemic-hypoglycemic clamp (HHC; d 6-10), 3) HS (d 16-20), and 4) HS + euglycemic clamp (EC; d 21-25). Cows acclimated to tie stall housing and experimental procedures from d −3 to −1. Days 11 to 15 of the experiment served as a period for acclimation to HS.
Figure 2.
Plasma ɣ-aminobutyric acid (GABA) concentrations for all four experimental periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp). Different point styles and colors represent different individual cows, as represented in the figure legend. The gray box represents the model estimated means (center line) for each group and their associated 95% confidence intervals. There were no significant differences detected when all four periods were included in the analysis (p = 0.11).
Figure 2.
Plasma ɣ-aminobutyric acid (GABA) concentrations for all four experimental periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp). Different point styles and colors represent different individual cows, as represented in the figure legend. The gray box represents the model estimated means (center line) for each group and their associated 95% confidence intervals. There were no significant differences detected when all four periods were included in the analysis (p = 0.11).
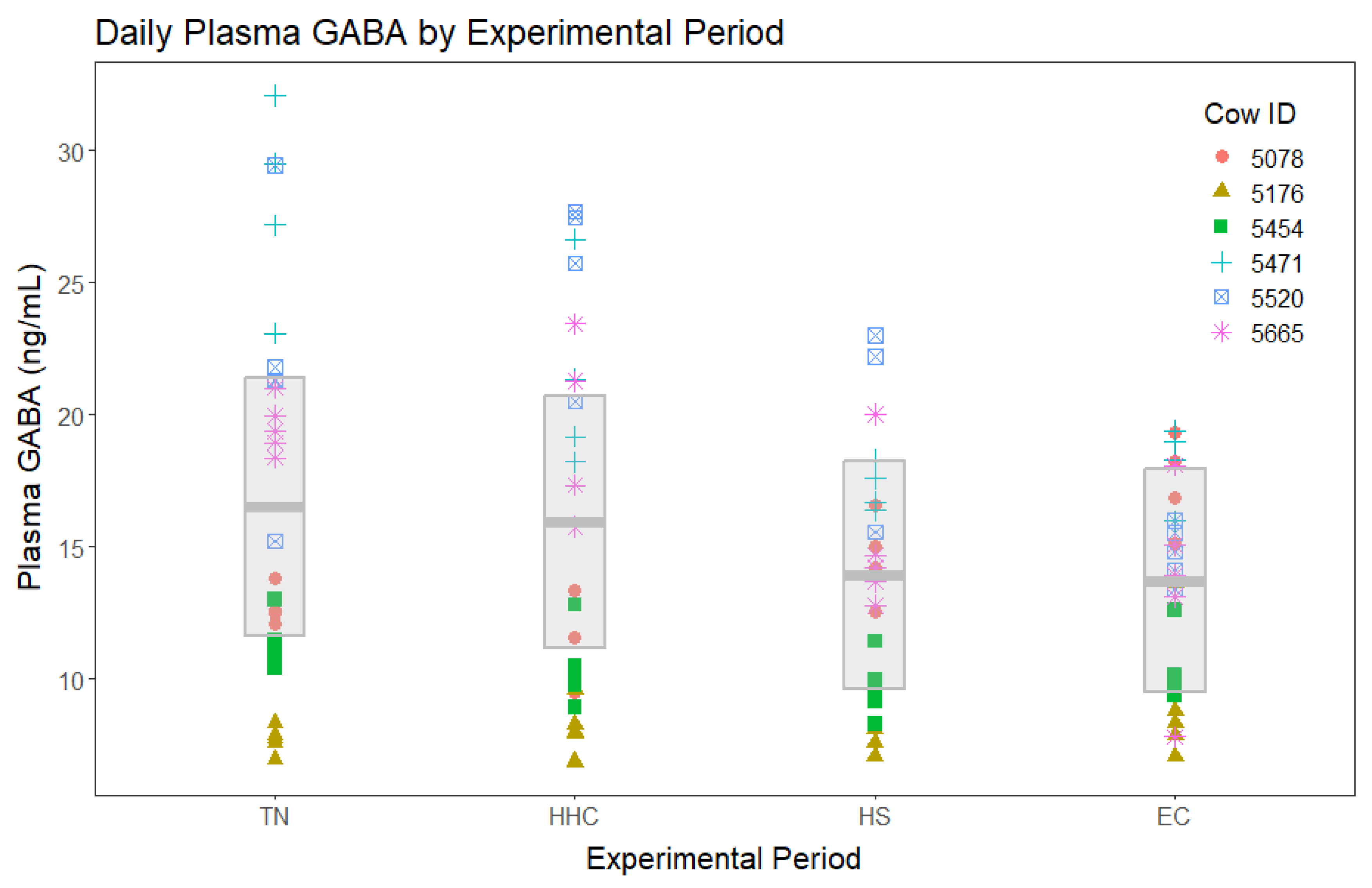
Figure 3.
Plasma ɣ-aminobutyric acid (GABA) concentrations by period for the thermoneutral (TN) and heat stress (HS) periods only. Points of different shapes and colors represent different cows. Small points show the raw data. Large points show the raw mean values for each cow. The gray box represents the model estimated means (center line) for each group and their associated 95% confidence intervals. Plasma GABA during HS tended to be reduced compared to the TN period (p = 0.06).
Figure 3.
Plasma ɣ-aminobutyric acid (GABA) concentrations by period for the thermoneutral (TN) and heat stress (HS) periods only. Points of different shapes and colors represent different cows. Small points show the raw data. Large points show the raw mean values for each cow. The gray box represents the model estimated means (center line) for each group and their associated 95% confidence intervals. Plasma GABA during HS tended to be reduced compared to the TN period (p = 0.06).
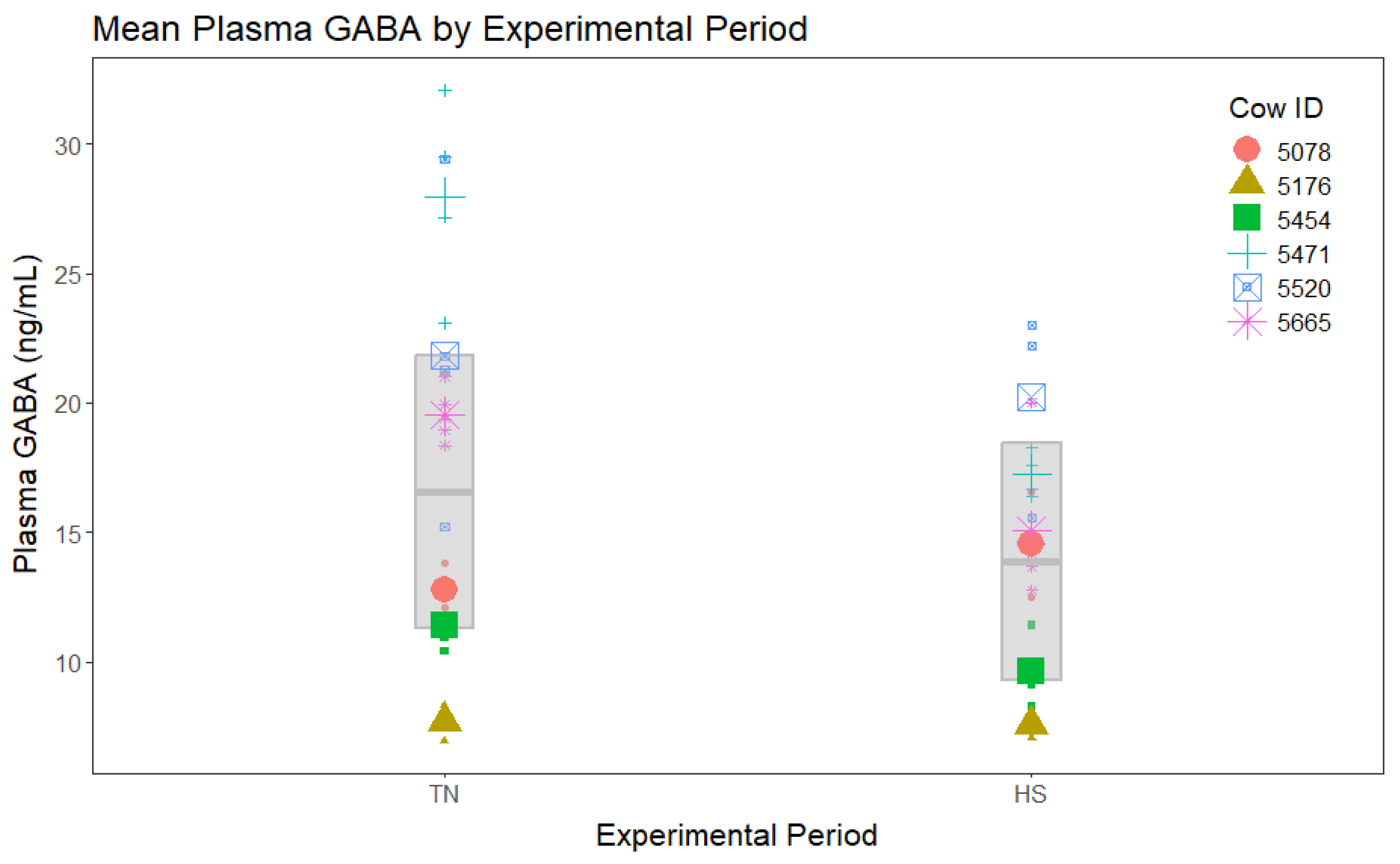
Figure 4.
Correlogram showing the correlations between plasma ɣ-aminobutyric acid (GABA) concentration, plasma progesterone (P4) concentration, daily feed intake, average daily respiration rates (RR), average daily rectal temperatures (RT), and daily milk production in kilograms, and plasma insulin concentration. The bottom half of the graph shows scatterplots of each variable against the others, and the top half shows the Pearson correlation coefficient for each pairing. A correlation value marked with * shows that the p-value is <0.05; *** shows that the p-value is <0.01. The diagonal shows a density plot representative of the distribution of each variable.
Figure 4.
Correlogram showing the correlations between plasma ɣ-aminobutyric acid (GABA) concentration, plasma progesterone (P4) concentration, daily feed intake, average daily respiration rates (RR), average daily rectal temperatures (RT), and daily milk production in kilograms, and plasma insulin concentration. The bottom half of the graph shows scatterplots of each variable against the others, and the top half shows the Pearson correlation coefficient for each pairing. A correlation value marked with * shows that the p-value is <0.05; *** shows that the p-value is <0.01. The diagonal shows a density plot representative of the distribution of each variable.
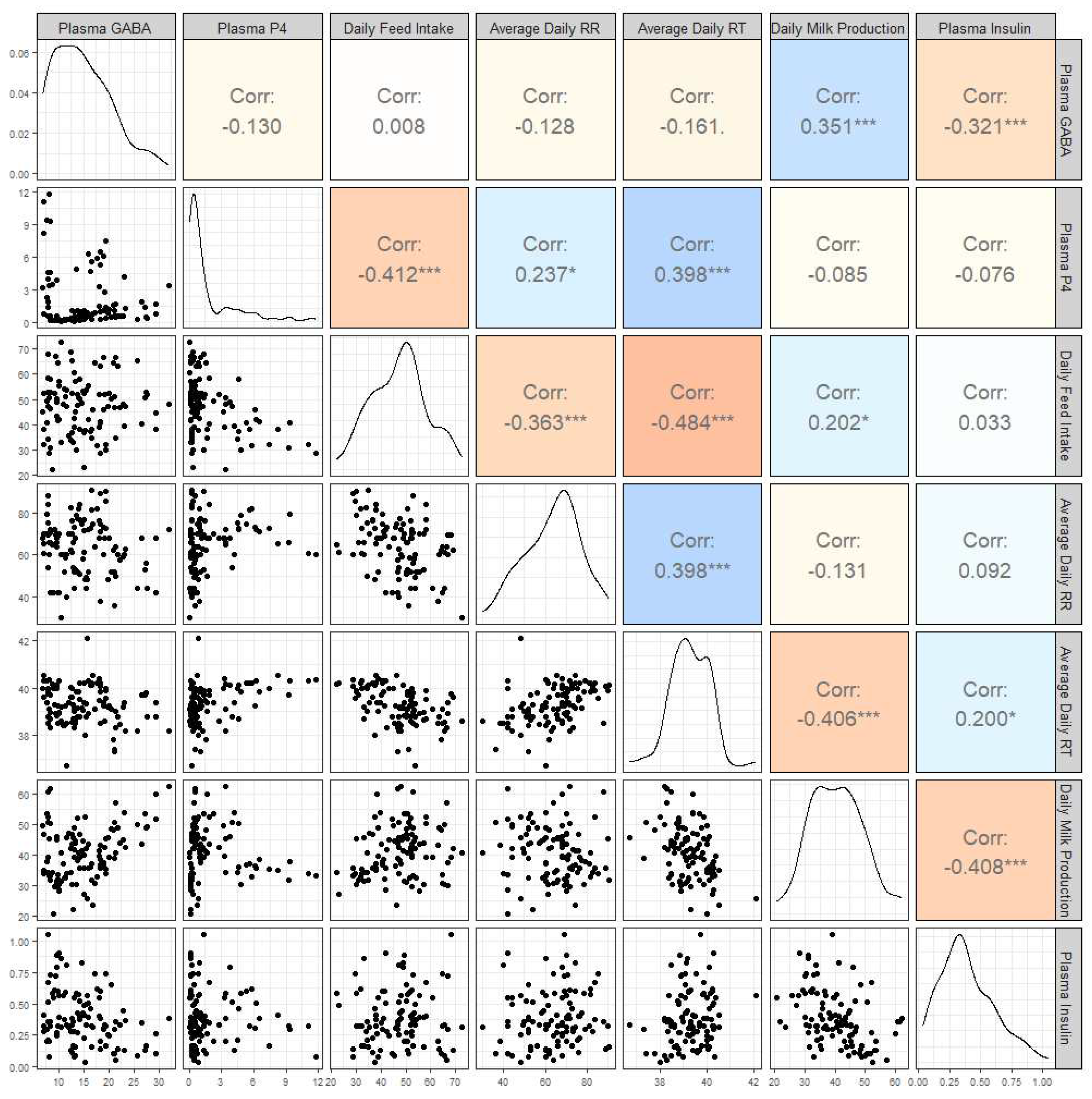
Figure 5.
The linear fit of total daily milk production in kilograms versus plasma ɣ-aminobutyric acid (GABA) concentration from all four experimental periods. Plasma GABA was a significant predictor of milk production, but not sufficient by itself to explain a substantial portion of the variability in milk production.
Figure 5.
The linear fit of total daily milk production in kilograms versus plasma ɣ-aminobutyric acid (GABA) concentration from all four experimental periods. Plasma GABA was a significant predictor of milk production, but not sufficient by itself to explain a substantial portion of the variability in milk production.
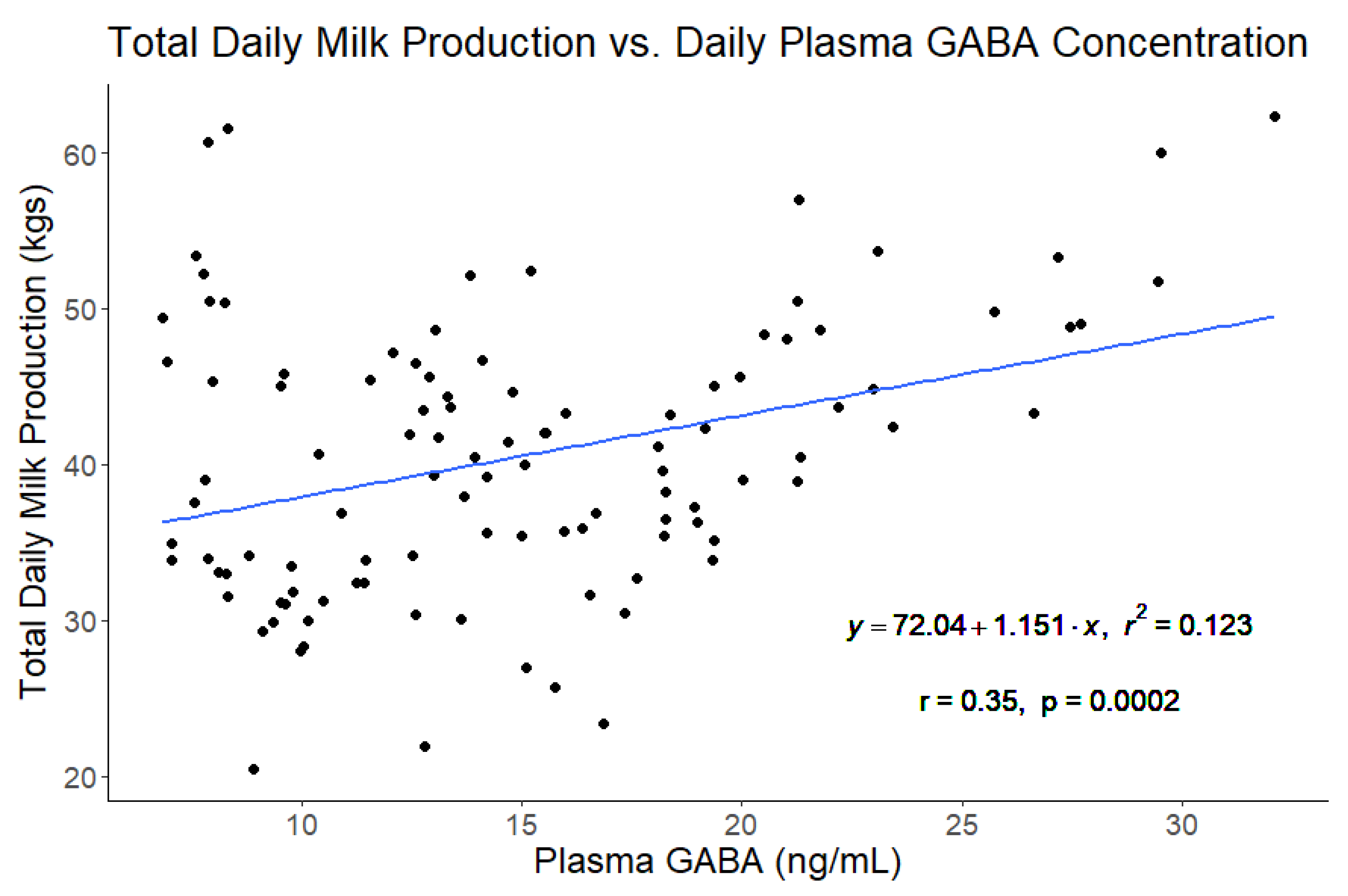
Figure 6.
The linear fit of daily plasma insulin concentrations versus daily plasma ɣ-aminobutyric acid (GABA) concentrations including data points from all four experimental periods. The two are lowly correlated and plasma GABA is not explanatory of a substantial portion of variability in plasma insulin concentrations.
Figure 6.
The linear fit of daily plasma insulin concentrations versus daily plasma ɣ-aminobutyric acid (GABA) concentrations including data points from all four experimental periods. The two are lowly correlated and plasma GABA is not explanatory of a substantial portion of variability in plasma insulin concentrations.

Figure 7.
The linear fit of blood glucose versus plasma ɣ-aminobutyric acid (GABA) from day 5 of the thermoneutral and heat stress periods. No other samples were included because glucose measurements were only collected concurrently with GABA measurements on these days. This comparison yielded a moderate correlation and the model including baseline GABA was significantly better than the intercept only model.
Figure 7.
The linear fit of blood glucose versus plasma ɣ-aminobutyric acid (GABA) from day 5 of the thermoneutral and heat stress periods. No other samples were included because glucose measurements were only collected concurrently with GABA measurements on these days. This comparison yielded a moderate correlation and the model including baseline GABA was significantly better than the intercept only model.
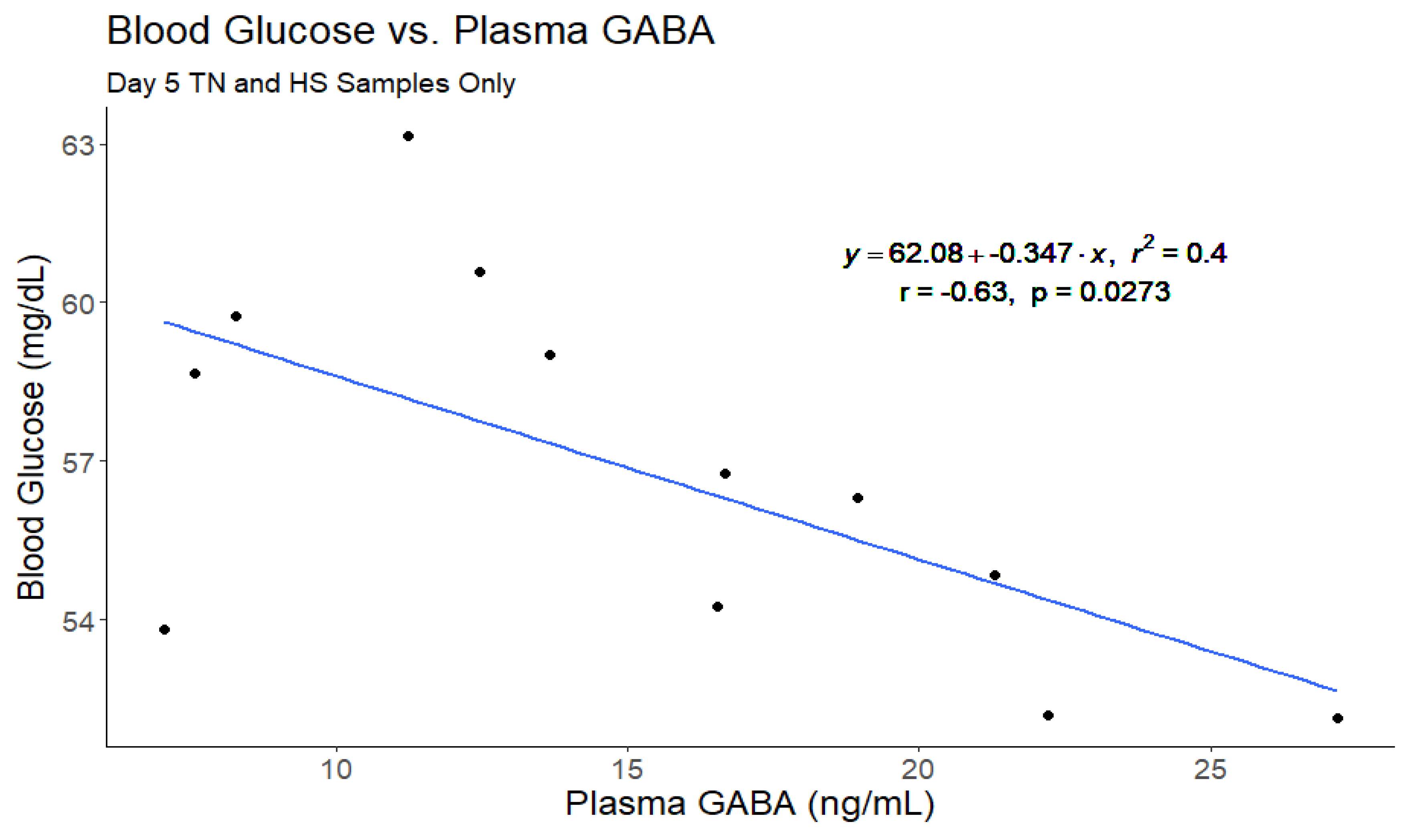

Table 1.
LS Means for Plasma Insulin by Period. The results from both the analyses including all periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp) and including only TN and HS periods are shown. Different numbers next to the names of the periods designate significant adjusted pairwise differences between the treatments.
Table 1.
LS Means for Plasma Insulin by Period. The results from both the analyses including all periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp) and including only TN and HS periods are shown. Different numbers next to the names of the periods designate significant adjusted pairwise differences between the treatments.
| Means from analysis with all periods | |||
| Plasma Insulin (ug/L) | Standard Error | P-value | |
| TN1 | 0.27 | 0.054 | 0.0131 |
| HHC1 2 | 0.42 | 0.057 | |
| HS2 | 0.43 | 0.057 | |
| EC2 | 0.43 | 0.055 | |
| Means from analysis with all TN and HS periods only | |||
| Plasma Insulin (ug/L) | Standard Error | P-value | |
| TN1 | 0.27 | 0.060 | 0.0026 |
| HS2 | 0.43 | 0.062 | |
Table 2.
LS Means for Plasma Progesterone by Period. The results from both the analyses including all periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp) and including only TN and HS periods are shown.
Table 2.
LS Means for Plasma Progesterone by Period. The results from both the analyses including all periods (TN=thermoneutral, HHC=hyperinsulinemic-hypoglycemic clamp, HS=heat stress, EC=euglycemic clamp) and including only TN and HS periods are shown.
| Means from analysis with all periods | |||
| Plasma Progesterone (ng/mL) | Standard Error | P-value | |
| TN | 0.78 | 0.44 | 0.1151 |
| HHC | 0.76 | 0.43 | |
| HS | 1.05 | 0.60 | |
| EC | 1.17 | 0.66 | |
| Means from analysis with all TN and HS periods only | |||
| Plasma Progesterone (ng/mL) | Standard Error | P-value | |
| TN | 0.73 | 0.40 | 0.2336 |
| HS | 1.11 | 0.62 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



