
Submitted:
16 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Burkholderia are a group of Gram-negative bacteria that can cause a variety of diseases in at-risk populations. B. pseudomallei and B. mallei, the etiological agents of melioidosis and glanders, respectively, are the two clinically relevant members of the B. pseudomallei complex (Bpc). The development of vaccines against Bpc species has been accelerated in recent years, resulting in numerous promising subunits and glycoconjugate vaccines incorporating a variety of antigens. However, a second group of pathogenic Burkholderia species exists known as the Burkholderia cepacia complex (Bcc), a group of opportunistic bacteria which tend to affect individuals with weakened immunity or cystic fibrosis. To date there have been few attempts to develop vaccines to Bcc species. Therefore, the primary goal of this review is to provide a broad overview of the various subunit antigens that have been tested in Bpc species, their protective efficacy, study limitations, and known or suspected mechanisms of protection. Then, we assess the reviewed Bpc antigens for their amino acid sequence conservation to homologous proteins found in Bcc species. We propose that protective Bpc antigens with a high degree of Bpc-to-Bcc sequence conservation could serve as components of a pan-Burkholderia vaccine capable of protecting against both disease-causing groups.
Keywords:
Burkholderia pseudomallei
; Burkholderia mallei
; Burkholderia cepacia complex
; melioidosis
; glanders
; cystic fibrosis
; vaccines
; subunit
; glycoconjugate
1. Introduction
Burkholderia are a ubiquitous genus of Gram-negative bacteria primarily consisting of saprophytes and rhizobacteria. Given their specialization for this environmental niche, most of the 120 or so species of Burkholderia are relatively innocuous to humans and are not known to cause overt disease [1,2]. However, there are two prominent disease-causing groups: the B. pseudomallei complex (Bpc) and the B. cepacia complex (Bcc). The former notably contains B. pseudomallei and B. mallei, the causative agents of melioidosis and glanders, respectively, which are designated as tier 1 select agents due to their status as potential biothreat agents [3]. Additionally, B. pseudomallei is associated with significant global disease incidence and mortality rates [4], making it the Burkholderia species of highest concern from a public health and safety standpoint. While Bcc species are neither biothreat agents nor are they associated with particularly high incidence rates, they can still significantly affect the quality of life of at-risk individuals and can even lead to death [5,6,7].
B. pseudomallei is found in tropical and subtropical climates where soil conditions are conducive for growth [8], with Southeast Asia and Northern Australia being considered the major hotspots of the disease [4]. However, a comprehensive modeling study suggests that disease incidence is significantly underreported in regions of South and Central America, the Middle East, and Africa [4]. Others have predicted that global climate change could lead to an increase in the incidence rate of melioidosis and could allow the bacteria to spread to areas where it was previously not considered endemic [9,10,11]. This latter prediction has materialized into reality in recent years, with B. pseudomallei being isolated for the first time from environmental samples in Mississippi, USA [12] and Southeast Queensland, Australia [13], suggesting a spread of the bacteria to new areas of endemicity. In terms of disease presentation, melioidosis can have a particularly wide array of symptoms affecting various bodily systems [14], leading some to refer to the pathogen as “the great mimicker” [15,16]. In general, roughly 9 out of 10 melioidosis patients experience an acute form of disease, the most common manifestation of which is a pneumonia-like illness that often proceeds to bacteremia [14,17]. The chronic form of disease generally presents as a more tuberculosis-like illness and is often associated with development of ulcers and abscesses at various sites of the body [14,17]. While chronic infection has a relatively low mortality rate, the acute form of disease is lethal in roughly 10% of cases under ideal treatment conditions [14,17]. In regions where reliable, accurate diagnosis cannot be achieved or proper antibiotics cannot be prescribed, mortality to acute infection increases to over 40% [14,17]. Taking into consideration the underreporting of cases, melioidosis is estimated to result in between 36,000 and 227,000 deaths annually across the globe [4].
B. mallei represents a clonal derivative of B. pseudomallei that has undergone successive rounds of reductive genome evolution, allowing it to become an obligate mammalian pathogen whose primary reservoir is solipeds [18]. Thanks to stringent global culling and monitoring practices, glanders has been eradicated in most developed countries, though sporadic outbreaks occur in other parts of the world [19,20]. Thus, human cases of glanders are rare and usually only observed in lab workers and in people who work closely with horses in regions of the world where the disease is still endemic [21]. It is generally held that the disease course of human glanders is like that of melioidosis, though due to the rarity of human glanders cases there could be subtle differences yet to be elucidated [21]. Despite the low disease incidence, quite a few B. mallei vaccines have been tested in animal models because B. mallei, like B. pseudomallei, is considered a tier 1 select agent, and there is therefore a level of vaccine development support for national defense purposes. Additionally, it should be noted that nearly all functional B. mallei genes are identical or near identical in translated amino acid sequence to their direct homologs in B. pseudomallei [18,22]. Thus, eliciting cross-reactivity between B. pseudomallei and B. mallei using a single vaccine is likely feasible. For this reason, many of the subunit and glycoconjugate vaccines that have been described for B. mallei have also been tested against B. pseudomallei.
The Bcc group consists of at least 26 species [23], only a fraction of which are associated with disease. Unlike members of the Bpc, Bcc species are ubiquitously distributed across the globe and thus pose a risk to susceptible patients everywhere. Members of the Bcc are consistently identified as one of the most common contaminants of pharmaceutical products, resulting in numerous product recalls and sporadic nosocomial outbreaks [24,25]. As such, individuals with weakened immune systems who are exposed to contaminated material can become sick with a range of disease manifestations largely depending on the route of entry into the body. Cystic fibrosis (CF) patients represent a particularly relevant risk group, as the unique environment of their lungs is conducive to chronic Bcc colonization [6,7]. Affected CF patients can be asymptomatic, though many experience an accelerated decline in lung function. A small subset of patients will develop a rapid-onset necrotizing pneumonia known as Cepacia syndrome, which almost always results in death [5,6,7,26]. Bcc lung colonization is noteworthy for being the only Burkholderia-associated disease known to be transmissible from person-to-person, albeit only amongst CF patients [27]. As a result, Bcc-positive CF patients are requested to isolate themselves from other CF patients [25]. Bcc colonization is remarkably difficult to clear with antibiotics, which can cause affected individuals to be given lower priority for potentially life-saving lung transplants [28,29]. Previously, B. cenocepacia was considered the most common cause of CF lung colonization though B. multivorans has become the dominant species for the past couple of decades [30,31]. Despite this, B. cenocepacia is still quite common and is associated with the highest degree of disease mortality [26]. Other clinically relevant species in the Bcc group include B. ambifaria, B. cepacia, B. contaminans, B. dolosa, and B. vietnamensis [25]. Only a very small number of vaccination studies have been conducted against Bcc species, which can be at least partially explained by the lack of animal models that effectively recapitulate the human disease [32]. Additionally, the correlates of protection for Bcc colonization of the CF lung are not well-defined, and it is particularly unclear to what extent Bcc-specific adaptive immune responses (i.e., antibodies and T cells) are capable of effectively preventing infection in the unique environment of the CF lung. Finally, because it is primarily CF patients who stand to benefit from a prophylactic Bcc vaccine, there is very little interest at the industry level in pushing a Bcc-specific vaccine through human clinical trials due to the relatively small patient population.
Therefore, some researchers have proposed repurposing Bpc-specific vaccines to treat Bcc infections as a feasible and economically viable solution [33,34,35]. One study recently broke down the cost effectiveness of developing and testing a melioidosis-specific vaccine, ultimately concluding it to be a cost-effective strategy of reducing disease incidence [36]. By extension, a melioidosis vaccine that could additionally protect CF patients from Bcc infection could only provide more incentive for industry-supported vaccine development. Taking an interest in this concept of a pan-Burkholderia vaccine, we decided to take a closer look at which, if any, of the proteinaceous B. pseudomallei and B. mallei vaccine antigens in the literature may potentially be cross-protective to Bcc species based on amino acid sequence conservation. To assess this, we first downloaded the annotated RefSeq proteome files from the NIH National Center for Biotechnology Information assembly database for all B. pseudomallei (Taxonomy ID 28450), B. mallei (TaxID 13373), and B. cepacia complex (TaxID 87882) strains/isolates that were not flagged as anomalous and that had an assembly level of “complete genome” or “chromosome.” We manually removed some of the resulting files if they appeared to be duplicates of the same strain/isolate. This resulted in a dataset consisting of 147 B. pseudomallei, 30 B. mallei, and 183 Bcc strains/isolates. A breakdown of the Bcc isolates in terms of their geographic origin and isolation source is provided in Figure 1. It is worth noting that there is some geographic and species bias in the Bcc dataset, as nearly half the proteomes represent B. multivorans isolates and nearly half of all isolates were sampled in Europe. By comparison, there is very little representation in Africa and South America due to a lack of high-quality sequences from these regions (Figure 1). The amino acid sequence FASTA files for each previously described Burkholderia vaccine antigen were downloaded from UniProt to serve as reference sequences. Such sequences were always taken from B. pseudomallei strain K96243 to ensure consistency in our comparisons and because it is one of the most used strains in B. pseudomallei vaccine studies. These FASTA files were individually compared to the downloaded RefSeq proteome files using the blastp function of the BLAST+ executables suite v2.14.0 downloaded from the NCBI website [37,38,39]. When running this command, the -evalue parameter was set to 0.00005 and the -max_target_seqs parameter was set to 1. For each individual antigen, an output for each strain/isolate comparison was created, and these results were compiled into one dataset using R v4.3.1 [40]. These comparisons' results are reported in Table 1, Table 2 and Table 3, which are referenced throughout the review as we discuss individual antigens. To keep these tables to a manageable size, we only included the average sequence conservation of the Bcc protein homologs compared to the reference B. pseudomallei strain K96243 sequences. Unless stated otherwise in the review, all antigens described had >90% sequence coverage, >99% sequence similarity, and were present in >90% of the annotated proteomes analyzed when comparing sequence conservation across B. pseudomallei and B. mallei strains/isolates.
2. Flagellar Proteins
B. pseudomallei and members of the Bcc group are motile due to the presence of one or more flagella. B. mallei is notably non-motile, having lost the ability to express a functional flagellum likely due to the presence of an insertion sequence in the fliP gene [41]. Flagella have been implicated as potential virulence factors in mouse models of melioidosis [42,43] and in Bcc colonization of the lung [44,45]. They are also potent mediators of inflammation due to their TLR5 and NLRC4 inflammasome agonistic activity [45,46,47,48,49]. The flagellum consists of three major domains: a basal body, a hook, and a long filament that spans into the extracellular milieu. The transmembrane basal body component serves as a motor, utilizing the electrochemical gradient of ions across the bacterial membrane to generate thrust. The hook sits on the surface of the bacterial outer membrane and acts as a flexible linker that transmits torque from the basal body to the filament. Lastly, the filament is the largest and most visible component of the flagellum, consisting of thousands of repeating flagellin (FliC) subunits that serve as a sort of molecular propeller [50]. FliC represents the first proteinaceous antigen characterized for Burkholderia, and a handful of studies have explored it as a vaccine antigen either alone or as a carrier protein in lipopolysaccharide (LPS) glycoconjugate vaccines [51,52,53,54,55,56]. Our lab has also evaluated the hook-associated protein FlgL as a candidate vaccine antigen [57,58,59].
2.1. FliC (BPSL3319)
FliC has numerous properties that make it an ideal target for humoral immunity. The relatively high level of expression and repetitive pattern of polymerized FliC subunits likely allows them to efficiently cross-link B cell receptors, leading to more efficient B cell activation [60]. Additionally, the flagellar filament is localized deep in the extracellular environment, far away from the bacterial glycocalyx which could otherwise interfere with B cell receptor and antibody binding [60]. FliC is also the component of flagella that activates TLR5 and the NLRC4 inflammasome, meaning that FliC has self-adjuvanting properties when used in a subunit vaccine [46,47,48,49]. Unsurprisingly given these favorable B cell stimulatory qualities, FliC-specific antibody titers are an established serodiagnostic marker of melioidosis, though such titers do not appear to correlate with improved disease outcome [61,62,63,64,65]. The FliC of both B. pseudomallei and B. cenocepacia has additionally been found to contain post-translational modifications (PTMs) in the form of species- and strain-specific patterns of glycosylation [66,67]. In B. cenocepacia these glycans appear to partially block the induction of TLR5 and serve to reduce FliC’s immunogenicity [67]. It is currently unclear how these glycans affect antigenicity, either by blocking underlying B cell epitopes or by serving as B cell epitopes themselves.
Brett et al. first isolated FliC from B. pseudomallei strain 319a and demonstrated its antigenic potential [51]. This seminal study determined that diabetic rats passively immunized with polyclonal antibodies specific to FliC were partially or fully protected from i.p.-delivered B. pseudomallei strain 316c, with survival rates ranging from 20 to 100% depending on the inoculum size and route of antibody delivery. It was further demonstrated in vitro that these antibodies could inhibit the bacteria’s motility, suggesting a possible mechanism of action [51]. Subsequent studies on this mechanism are quite contentious, with an early study by DeShazer et al. indicating that flagella are dispensable in Syrian hamster and diabetic rat i.p. models of infection, implying that flagella-mediated motility is not necessary [68]. However, Chua et al. found that a flagellin knockout B. pseudomallei strain KWH was significantly attenuated in both i.p. and i.n. infection routes using BALB/c mice, suggesting different virulence mechanisms in these rodent models [42]. Further adding to the controversy, another study was then unable to reproduce these results, finding that flagella was essentially dispensable in BALB/c mouse models of infection [69]. Regardless of the exact mechanism, whether FliC-specific antibodies are mediating protection by inhibiting bacterial motility or through Fc-mediated mechanisms of immunity, this initial passive transfer study by Brett et al. clearly demonstrated that FliC-specific antibodies provide some level of protection against B. pseudomallei, at least in the diabetic rat infection model [51].
Another group later created a DNA vaccine encoding the B. pseudomallei fliC gene [53,54]. The authors of these studies immunized female BALB/c mice intramuscularly with the fliC-encoding plasmid or subcutaneously with recombinant FliC in Freund’s adjuvant. An intravenous challenge with 105 CFU B. pseudomallei was administered, resulting in 83% protection in the DNA-vaccinated mice, 50% in mice given the recombinant protein, and 0% in the control mice. While statistical significance was not reported for this study, the DNA vaccine appeared to be the superior vaccine formulation despite eliciting lower overall IgG titers than the recombinant protein. Interestingly, the animals receiving the DNA vaccine had relatively balanced IgG1/IgG2a titers, whereas the animals receiving the recombinant protein were markedly more IgG1-skewed, suggesting Th2, humoral-biased immunity. Furthermore, splenocytes from the DNA vaccinated group exhibited a significantly higher propensity to proliferate and secrete IFN-γ upon ex vivo stimulation with FliC compared to splenocytes recovered from mice immunized with the protein, providing additional support that Th1 responses were stronger in animals receiving the DNA vaccine [53]. These results implicate T cell responses as playing a significant role in protecting these mice from lethal challenge. The group then added immunostimulatory CpG motifs to their DNA vaccine to skew the immune response further towards a Th1 phenotype, though they did not report mouse survival data for these experiments. Instead, they demonstrated that the inclusion of these CpG motifs led to a statistically significant decrease in liver but not spleen colonization using the same intravenous challenge model [54]. Providing additional support that FliC acts as a potent T cell antigen, this protein has also been repeatedly used as the carrier protein in glycoconjugate vaccines to Burkholderia [52,55,56]. Such vaccines have exhibited a strong ability to generate high antigen-specific IgG titers to Burkholderia surface polysaccharides, suggesting that T cell dependent activation of these B cell subsets has been achieved [52,55,56]. However, when compared side-by-side to other potential glycoconjugate carrier proteins, FliC performed worse than Hcp1 and similarly to TetHc in its ability to induce LPS-specific antibody titers, suggesting that it may not be particularly special in its T cell antigenic properties [56]. Further exploration of this topic is needed before claims can be made about FliC-specific T cells and their role in protection against melioidosis.
All published studies that specifically set out to explore a protective role of FliC in vaccination against B. pseudomallei have used outdated challenge models unlikely to recapitulate disease in humans, namely intraperitoneal challenge of diabetic rats [51] and intravenous challenge of mice [53,54]. The more recently explored FliC-incorporating glycoconjugate vaccines have utilized the current “gold standard” model of respiratory-acquired glanders, though similar reports are surprisingly lacking for respiratory-acquired melioidosis. In these glanders studies, there was little evidence to suggest that FliC-specific immunity played any meaningful role in protection, as seen by the similar levels of organ colonization and survival in the animals immunized with FliC-LPS versus those immunized with TetHc-LPS, as TetHc is a non-Burkholderia protein. This finding is somewhat unsurprising given that B. mallei lacks an assembled flagellum [55,56]. As there are no peer-reviewed examples of FliC being used to protect against respiratory-acquired B. pseudomallei, definitive conclusions cannot be drawn as to FliC’s potential as a melioidosis vaccine antigen. That said, another study discussed unpublished data suggesting that FliC is not protective in such respiratory models, casting some doubt on the antigen’s usefulness as a melioidosis vaccine antigen [70]. It is worth highlighting that Brett et al.’s initial characterization of FliC is the only published study that used B. pseudomallei-derived FliC [51], which would have native glycosylation patterns. All subsequent studies have used host-cell translated FliC [53,54] or E. coli-expressed FliC [55,56], neither of which would necessarily be expected to retain native-like glycosylation patterns. The extent to which this plays a role in humoral immunity is entirely unexplored and worthy of future consideration.
Regardless, even if FliC does have potential as a melioidosis vaccine antigen, it is significantly hampered compared to most other antigens discussed in this review by the fact that it is unlikely to elicit cross-protection to glanders. Similarly, FliC is expected to have only moderate applicability as a Bcc cross-reactive antigen. This is because Bcc species are known to express at least two evolutionarily distinct FliC variants that vary significantly in size and sequence [71]. However, we did find in our bioinformatic screen that B. cepacia, B. cenocepacia, and most strains of B. multivorans, which represent three of the most clinically relevant species of Bcc, express a FliC variant that is similar to that of B. pseudomallei. The amino acid sequences of these Bcc FliC variants are 74.0% identical and 84.6% similar to B. pseudomallei strain K96243 FliC, suggesting a moderate degree of conservation compared to the other antigens discussed herein (Table 1). Supporting this finding, Musson et al. generated T cell hybridomas specific for a B. pseudomallei FliC epitope which cross-reacted strongly with peptides derived from B. cepacia-, B. cenocepacia-, and B. multivorans-derived FliC, suggesting that T cell-mediated cross-protection could be elicited by this protein [72]. We found in our BLAST analysis that the variants of FliC found in other Bcc species appear to exhibit a generally lower degree of sequence conservation to the N- and C-terminal ends (residues 1-160 and 286-388) of B. pseudomallei FliC, though the middle region (residues 161-285) lacks any reported sequence conservation. It is therefore questionable as to whether FliC could elicit cross-reactive immunity to other Bcc species. In summary, FliC’s role as a protective antigen in respiratory-acquired melioidosis is contentious, it is unlikely to protect against glanders to any meaningful capacity, and the extent of cross-reactivity to Bcc FliC is likely to be highly species-dependent but theoretically possible. Further investigation is needed to confirm these assertions.
2.2. FlgL (BPSL0281)
Our laboratory identified the hook protein FlgL in a reverse vaccinology approach designed to screen for potentially antigenic surface proteins [57]. This protein has been included in three separate studies in which different carrier proteins were tested in gold nanoparticle-vectored LPS glycoconjugate vaccines to melioidosis [57,59] and glanders [58]. In general, the results when using FlgL have been somewhat mixed. In the first study, it was found that no difference was observed between the carrier proteins in their ability to induce LPS-specific IgG when administered intranasally [57]. Despite this, the construct containing FlgL as the carrier protein outperformed every other construct in its ability to protect mice from a lethal intranasal challenge of B. pseudomallei strain K96243, with 90% of mice surviving the challenge compared to 10-20% in the groups receiving LPS conjugated to Hcp1, BpaE, or bovine serum albumin (BSA) [57]. This suggested that FlgL-specific immunity was playing an important role in protecting these mice. However, in a follow-up experiment we tested a wider array of carrier proteins and the FlgL construct was not similarly protective [59]. One potential explanation of this discrepancy could be the different adjuvant systems used in these separate studies. Somewhat unsurprisingly, the FlgL-LPS construct was also not protective against a lethal intranasal challenge of glanders, which can likely be attributed to the lack of flagellum assembly in this species [58]. Further studies are needed to confirm whether FlgL is protective against melioidosis. If it is, future researchers may explore whether protection is mediated primarily by FlgL-specific antibodies or T cells, as FlgL is likely much less accessible for antibody binding than FliC. Further exploration of FlgL may be worthwhile given FlgL’s higher degree of Bpm-to-Bcc conservation compared to FliC (Table 1).
3. Type 3 Secretion Systems
Type 3 secretion systems (T3SS), also known as injectosomes, are syringe-like structures capable of delivering substrates across the plasma membrane of nearby cells or across the endosomal membrane of infected eukaryotic cells [73,74]. This largely implicates them as virulence factors in a wide variety of Gram-negative bacteria including Yersinia pestis, pathogenic E. coli, Salmonella enterica, Shigella, and Pseudomonas aeruginosa. While most of the T3SS components are localized within the cell envelope where they cannot be readily recognized by antibodies, there are at least two components that are surface-exposed. These include a needle-like complex that extends from the bacterial surface towards the target cell and a hetero-oligometric translocation pore that sits at its tip [73,74]. Some groups have suggested that the outer membrane-embedded ring may be targetable by antibodies as well, though to date targeting this domain in other bacterial species has had little success [75,76,77,78]. Antibodies towards the translocation pore would be predicted to have the highest likelihood of blocking the activity of the T3SS, with antibodies towards the needle or outer membrane ring being perhaps more likely to elicit Fc-mediated mechanisms of protection.
B. pseudomallei expresses two T3SSs belonging to the Hrp-2 group (T3SS-1 and T3SS-2) and one from the Inv/Mxi-Spa group (T3SS-3) [79]. T3SS-2 and -3 are conserved in B. mallei but T3SS-1 is not [79,80]. Bcc T3SSs have minimal overlap with those of the B. pseudomallei complex, and there is only one T3SS that is broadly conserved across Bcc species, termed the Bcc injectisome (BCI) [79]. Evolutionary cousins of the T3SS-1, -2, and -3 are found in some Bcc isolates, but only the T3SS-2 is found with any significant degree of sequence conservation in B. ubonensis and B. stagnalis, two species of little clinical significance [79]. B. multivorans and B. cenocepacia, the two most clinically relevant Bcc species in CF lung colonization, are reported to only express the BCI [79]. Thus, T3SSs are predicted to have no applicability as Bcc cross-protective antigens.
Despite this, the T3SS-3 is one of the most critical and well-studied virulence determinants in B. pseudomallei and B. mallei, making it an attractive target for vaccines or therapeutics against melioidosis and glanders. This topic has been extensively reviewed elsewhere [81]. T3SS-3 secreted effectors are required for endosomal escape during the intracellular replication cycle [82] and dissemination of the bacteria from the lungs in intranasally challenged animals [83]. There are seven confirmed T3SS-3 effector proteins including BopA, BopC, BopE, BapA, BapC, BprD, and CHBP [81]. While such effectors are unlikely to play a significant role in humoral immunity given their intracellular localization, they may represent particularly promising T cell antigens. This is because during the intracellular phase of infection, T3SS substrates are often the first bacterial proteins that make their way into the cytoplasm where they can be processed by the proteasome into MHCI fragments to subsequently stimulate CD8 T cell immunity. This strategy of using T3SS substrates as T cell antigens has been employed with success in Salmonella enterica vaccines [84,85] and a similar approach has been utilized with type IV secretion system substrates in Coxiella burnetii [86], which are similarly injected into the cytoplasm from within the endosome. However, a key difference in these organisms is that S. enterica and C. burnetii tend to survive and replicate within the endosome for hours or days, whereas B. pseudomallei escapes the endosome within minutes of being internalized [87,88,89,90]. Given this extremely short period of time, it is possible, perhaps even likely, that Burkholderia T3SS-3 substrates do not have any advantage as T cell antigens over other proteins. Regardless, based on this information two general strategies for vaccinating against the T3SS-3 have been employed: inducing humoral immunity to the translocation pore [91,92] and inducing T cell immunity towards the T3SS-3 effector proteins [70].
3.1. T3SS-3 Translocation Pore Proteins
The translocation pore of the T3SS-3 consists of BipB (BPSS1532), BipC (BPSS1531), and BipD (BPSS1529). Stevens et al. first attempted to use BipD as a vaccine antigen, pairing it with Sigma Adjuvant System (SAS) and administering it intraperitoneally in BALB/c mice [91]. The authors ultimately did not observe any protection from a lethal i.p. challenge of B. pseudomallei strain K96243, despite robust BipD-specific IgG titers [91]. Druar et al. followed up with a more complete investigation of the T3SS-3 needle tip proteins as antigens, testing BipB, BipC (N-terminal half or C-terminal half), and BipD in Freund’s adjuvant [92]. A single dose of any of these four antigens delivered intraperitoneally did not confer any detectable protection against 30x LD50 of i.p.-delivered B. pseudomallei strain Ashdown, again despite robust IgG titers [92]. Curiously, serum from experimentally infected mice did not react to these proteins while serum from recovered human melioidosis patients did [92]. This could indicate that the lack of protective efficacy in mice may be attributed to the intraperitoneal mouse challenge model employed by the authors. Supportive evidence suggest that the T3SS-3 is expressed at low levels when grown in the lab, but then becomes transcriptionally upregulated when human serum is present, within the environment of the murine lung, or when salt concentrations are high [92,93,94,95]. This may suggest that the amount of T3SS-3 available for antibody binding during the extracellular phase of infection may be dependent on the initial growth conditions of the bacteria as well as its route of entry into the body. Future studies should investigate whether intranasally-infected mice develop antibody titers to the needle tip proteins, or whether needle tip protein vaccination is protective in intranasal models of infection. Evaluating the needle component, BsaL, as a vaccine antigen may also be worthy of consideration. While BsaL antibodies are less likely to antagonize T3SS-3 activity, they could still prove efficacious through Fc-mediated mechanisms of protection. Additionally, BsaL, like FliC, may have self-adjuvanting properties on account of its NLRC4 stimulating activity [46,96].
3.2. T3SS-3 Effector Proteins
BopA (BPSS1524) plays a significant role in intracellular survival, possibly by inhibiting autophagy [81], and is the only T3SS-3 effector that has been tested as a vaccine antigen [70]. In this study, B. mallei-derived BopA was adjuvanted with CpG ODN 2395 and immune-stimulating complex AbISCO 100 and intranasally administered into BALB/c mice. The animals were then challenged intranasally with 2x LD50 of B. mallei strain ATCC 23344 and were completely protected from mortality over the course of 21 days. By comparison, only one out of eight animals given BSA in the same adjuvant survived past day 5 [70]. The authors also attempted a heterologous vaccination/challenge model in which the animals were immunized intranasally with the B. mallei-derived BopA in cationic liposome-DNA complex adjuvant before being challenged intranasally with 2x LD50 of B. pseudomallei strain 1026b. In this model, 60% of the immunized animals survived to the endpoint of the study at day 60 whereas control animals all succumbed within the first week after challenge. Furthermore, the other two antigens tested in this study, BimA and LolC, failed to elicit long-term protection to the same degree [70]. Altogether, this indicates that BopA-specific immunity is protective against respiratory-acquired B. mallei and B. pseudomallei, likely through T cell-mediated mechanisms though this was not directly confirmed by the authors. Surprisingly, BopA has not appeared in any subsequent vaccine studies despite exhibiting promising results in “gold standard” infection models of melioidosis and glanders. Future studies may try pairing promising B cell antigens, such as LPS or capsular polysaccharide (CPS), with BopA to determine if the two complement each other. T cell responses should also be measured to confirm their role in mediating protection. Furthermore, since BopA showed such promise in this initial study, it may warrant exploring other T3SS-3 effectors as protective antigens.
4. Type 5 Secretion Systems
Type 5 secretion systems (T5SS), also known as autotransporters (ATs), are remarkably efficient proteins capable of inserting themselves into the outer membrane to subsequently carry out some effector function in the extracellular space [97,98]. These proteins are involved in many functions, ranging from adhesion, invasion, biofilm formation, cytotoxicity, and immunomodulation. They consist of a translocator domain, which inserts itself into the outer membrane to serve as a pore, and a passenger domain, which is extruded across the membrane to carry out the AT’s function [97,98]. Notably, the translocator domain consists of a β-barrel motif. Such motifs make the expression of recombinant proteins in their native conformations particularly challenging, a topic discussed in greater detail later in the section on general β-barrel proteins. As a result, most studies of ATs as subunit vaccine antigens have instead opted to express only the passenger domain of the protein.
There are 11 predicted ATs in B. pseudomallei [99]. These can be classified into two distinct groups: classical ATs and trimeric ATs. Nine of the predicted ATs fall into the latter category including BpaA, BpaB, BpaC, BpaD, BpaE, BpaF, BoaA, BoaB, and BimA [99]. In Gram-negative bacteria, trimeric ATs are almost always involved in adhesion and invasion [99]. BimA is one of the very few exceptions to this rule, whose function is to induce actin polymerization in infected host cells as a form of intracellular motility leading to dissemination of the bacteria into adjacent cells [100]. BimA is thus one of the most well-studied ATs in B. mallei and B. pseudomallei and represents a critical virulence factor. The remaining two ATs, BcaA and BatA, are classical ATs which tend to have a more diverse array of possible functions. Our current understanding of these two proteins suggests that the former is involved in host cell invasion [101], while the latter appears to have lipase activity and may be required for efficient intracellular replication and/or survival [102].
ATs in Bcc species are not as well characterized as those in the Bpc, though one study screened eight different species of the Bcc and identified eight evolutionarily distinct lineages of trimeric ATs [103]. We looked further into the B. cenocepacia representatives of six of these lineages and found that none had any significant sequence conservation to trimeric ATs of the Bpc, exhibiting poor sequence coverage and low sequence identity (34-57%) to BpaA, BpaC, or BpaE. We were unable to find any information regarding non-trimeric ATs in the Bcc. The extreme variability of the trimeric ATs in Bcc and their apparently poor conservation between the Bpc and Bcc leads us to the conclusion that they would make poor cross-reactive vaccine antigens. That said, AT sequences were largely incompatible with our BLAST analysis pipeline due to the presence of multiple amino acid tandem repeat sequences of highly varying length. As such, we were not able to compare the Bcc AT sequences to those of B. pseudomallei strain K96243 using the same pipeline as the other proteins discussed in this review. For consistency, we have opted not to include these comparisons in Table 1. To date, three of the above B. pseudomallei ATs have been tested as vaccine antigens to Bpc pathogens including BimA [70], BpaE [57,58,59], and BatA [104].
4.1. BimA (BPSS1492)
BimA is a trimeric AT harboring proline-rich motifs and two regions with significant homology to WASP domain-2. These domains allow the bacteria to recruit host cell actin to its pole [100]. BimA also harbors a variable number of PDASX repeats that are directly involved in inducing actin polymerization, and variants of BimA harboring a higher number of repeats appear to polymerize actin more efficiently [105]. B. pseudomallei and B. mallei utilize these polymerizing actin filaments to propel themselves across the cytoplasm of the infected cell and to penetrate and infect adjacent cells, thus playing a vital role in dissemination and persistence of the bacteria [106]. While the majority of B. pseudomallei isolates appear to express a variant of BimA that is distinct from that of B. mallei, a subset of B. pseudomallei isolates from Northern Australia, India, and Sri Lanka harbor a B. mallei¬-like variant [107,108,109]. Interestingly, the B. mallei-like variant of BimA in melioidosis appears to be associated with increased intracellular persistence [110], increased incidence of neurological symptoms [111], and worse disease outcomes in patients experiencing neurological symptoms [112]. In addition to the two evolutionarily distinct variants of BimA, there is a relatively high amount of variation in the number of PDASX repeats among B. pseudomallei isolates as well as in the number of NIPVPPPMPGGGA repeats found adjacent to the proline-rich domain. Taking this into account, BimA likely represents the protein antigen with the highest degree of B. pseudomallei intra-species variability discussed in this review and is the only antigen that is present in B. mallei but lacks significant sequence identity to the B. pseudomallei strain K96243 homolog (~71% sequence identity). This represents a potentially significant limitation to vaccinating against BimA, as cross-reactivity between species or even strains are likely to be low compared to the other antigens.
Due to these limitations, only one study has attempted to use BimA as a vaccine antigen. Whitlock et al. expressed BimA from B. mallei strain ATCC 23344 without its signal sequence or the C-terminal translocator domain and formulated it with the adjuvants CpG ODN2395 and immune-stimulating complex AbISCO100. The formulation was administered to BALB/c mice via the i.n. route before challenging via the same route with 2x LD50 B. mallei strain ATCC 23344. All vaccinated animals survived to 21 days post-challenge compared to only 12.5% in control animals. The authors performed a follow-up experiment in which they adjuvanted this B. mallei variant of BimA with cationic liposome-DNA complex and subsequently challenged animals with 2x LD50 of B. pseudomallei strain 1026b. This heterologous challenge model resulted in ~80% protection to the acute phase of infection and ~20% long-term protection. While this was notably better than the control mice which all succumbed during the acute phase of infection, BimA provided inferior long-term protection compared to vaccination with BopA (60%) [70]. It has not yet been determined if antibodies to BimA can inhibit intracellular motility and/or intercellular dissemination, or if some other mechanism is responsible for the reported protection.
4.2. BpaE (BPSS0908)
BpaE (hemagglutinin) is a somewhat typical trimeric AT which plays a role in adherence and invasion [113]. A B. pseudomallei BpaE mutant exhibited significantly reduced adherence, invasion, and plaque formation when incubated with A549 cells [113] and exhibited a 40-fold higher median lethal dose in a murine melioidosis infection model [114]. Together this implicates BpaE as a moderate virulence factor in B. pseudomallei. BpaE has also been confirmed as an immunogenic protein during natural infection, as human melioidosis serum and peripheral blood mononuclear cells (PBMCs) are reactive towards it [57,63,114]. BpaE is another one of the proteins identified by our group’s reverse vaccinology screen as a potentially immunogenic surface protein, which it was referred to in past publications as “hemagglutinin” due to its annotation as a hemagglutinin-like protein. The full-length recombinant protein from B. mallei strain ATCC 23344 was expressed and chemically conjugated to LPS as a glycoconjugate vaccine [57]. Given the predicted antigenicity of BpaE, the BpaE-LPS construct consistently elicited high LPS-specific antibody titers [57,58,59]. Despite this, in two separate studies in which respiratory challenges with B. pseudomallei were performed, the BpaE-LPS construct failed to elicit meaningful protection [57,59]. However, in a glanders infection model, 90% of mice were protected to a low dose challenge of 2x LD50 and 50% of mice were protected from a 25x higher challenge dose. The control mice in these two experiments exhibited 50% and 0% survival, respectively. In the high dose challenge experiment, we also noted that the animals receiving the BpaE construct appeared to have the highest rate of sterilizing immunity in the lungs, liver, and spleen, though statistical significance could not be established [58]. This may suggest that BpaE-specific immunity is playing a role in limiting dissemination, though given the scope of this experiment, we could not definitively determine if such protection was due to BpaE-specific immunity or LPS-specific immunity.
4.3. BatA (BPSL2237)
BatA, one of the only two known classical ATs in the Bpc, was the final B. pseudomallei AT to be functionally characterized from the initial list of 11. In murine models of melioidosis infection, BatA appears to play little to no role in virulence [114]. Furthermore, PBMCs from melioidosis seropositive human donors do not significantly respond to antigen recall with BatA, nor does seropositive donor serum react to recombinant BatA, suggesting that the protein is poorly immunogenic or minimally expressed during B. pseudomallei infection [104,114]. In sharp contrast, a BatA knockout strain of B. mallei exhibited a reduction in virulence by more than two orders of magnitude in aerosol challenge models in mice, and both horses and mice experimentally challenged with wild type B. mallei elicited significant BatA-specific antibody titers [102]. These results may highlight a potential difference in virulence mechanisms between B. pseudomallei and B. mallei. The authors of the B. mallei studies attempted to understand this discrepancy by functionally characterizing BatA in B. mallei, finding that it has lipolytic activity and plays a role in intracellular survival and/or replication in J774 murine macrophages [102].
Because of these findings, the B. mallei-derived BatA passenger domain was subsequently tested by Lafontaine et al. as a recombinant vaccine antigen [104]. The group opted to use a somewhat atypical adjuvant for their initial experiment, that being RECOMBITEK Lyme, a veterinary-approved Lyme disease vaccine primarily consisting of lipidated OspA protein from Borreliella burgdorferi. Simultaneous administration of this vaccine via both the subcutaneous and intranasal routes on three separate days resulted in an extremely IgG1-biased immune response indicative of primarily humoral-driven immunity [104]. After three immunizations, the animals were challenged i.n. with 10x LD50 B. mallei strain ATCC 23344, resulting in 26% protection to acute infection and 21% protection past two weeks [104]. By comparison, the control animals receiving only the adjuvant were 10% protected from acute infection and only 5% survived more than 2 weeks [104]. Three of the four surviving vaccinated animals had no detectable bacteria in their lungs, though sterility was not observed in the spleens [104]. Altogether this experiment suggested only lackluster protection afforded by recombinant BatA passenger domain, though it is possible that the lipolytic activity of BatA may have interfered with OspA’s adjuvanting capacity by cleaving its lipid tail, a possibility that was not addressed by the study.
The BatA passenger domain was also cloned into parainfluenza virus 5 (PIV5) to serve as a vector for the antigen [104]. A single dose of this construct resulted in 85% and 74% protection against acute and chronic phases of glanders infection, respectively. Sterility was observed in the lungs of nearly half the surviving mice, but again not in the spleens. By comparison, a PIV5 vector expressing a non-specific tuberculosis protein only conferred 35% and 25% protection against acute and chronic phases, respectively. The authors also explored whether BatA-specific immunity could protect against a heterologous intranasal B. pseudomallei strain K96243 challenge. 80% and 60% protection to acute and chronic stages of infection was achieved, respectively, with PIV5-vectored BatA, whereas the PIV5 control vector afforded only 33% and 20% protection. The authors reported almost complete sterility in the lungs of surviving mice, and nearly half the surviving mice lacked bacteria in the spleen. Despite these promising results, the authors reported that PIV5-BatA-vaccinated animals did not elicit significant antibody titers towards recombinant BatA passenger domain, possibly suggesting that humoral immunity is not responsible for this protection [104]. If antibody titers to BatA were indeed low, BatA-specific T cell responses would likely be responsible for the protection reported by the authors. While an IFN-γ ELISpot of BatA-recalled splenocytes resulted in a significant number of spots compared to those from naïve mice, splenocytes from mice vaccinated with the non-specific PIV5 control construct also exhibited a significant number of spots that was not clearly discernible from the PIV5-BatA group. Thus, it is still unclear how the PIV5-BatA vaccine is mediating protection and evidence for BatA’s use in a melioidosis or glanders vaccine is promising but inconclusive.
5. Type 6 Secretion Systems
The type 6 secretion system (T6SS) is sometimes described as a molecular “harpoon” that is most often used by bacteria to deliver harmful effectors to other prokaryotes or eukaryotic cells [115,116]. Less commonly, they can also play a role in scavenging nutrients and metal ions [115,116,117]. The T6SS consists primarily of the harpoon-like structure, a sheath surrounding it, and a membrane-spanning complex and base plate. The shaft of the harpoon consists of polymerized Hcp (TssD) proteins that end in a tip made of VgrG (TssI) and PAAR. Cargo molecules can bind to the shaft or tip for delivery into the recipient cell, though Hcp, VgrG, and PAAR can have effector functions as well [118,119,120]. The sheath consists of polymerized TssB and TssC, the length of which is controlled by TssA at the base. This sheath contracts to thrust the harpoon into an adjacent cell via a contact-dependent manner (virulence) or into the extracellular milieu in a contact-independent manner (nutrient acquisition). Both the sheath and shaft reside primarily in the cytoplasm until the shaft is fired across the cell envelope. Finally, the membrane-spanning complex and base plate consist of TssE-G and TssJ-M, providing a channel for the harpoon-like structure to pass the bacterial envelope. Only TssJ is found in the outer membrane, and it appears to be located exclusively within the lipid bilayer and periplasm [115,116]. In summary, none of the components of contact dependent T6SSs are predicted to be exposed to the extracellular space before or after secretion, indicating that they are likely serving as T cell antigens and not antibody targets when used in vaccination.
The B. pseudomallei genome harbors six T6SS loci (T6SS-1-6). There are two distinct numberings of these systems used in the literature which can be a source of confusion. In this review, we have opted to use the nomenclature introduced by Schell et al [121]. The T6SS-2 was characterized as a contact-independent system involved in Zn2+ and Mn2+ acquisition to combat oxidative stress [117]. The T6SS-4 was recently implicated as being involved in biofilm formation, flagella biosynthesis, and intracellular survival in A549 cells, though the exact mechanistic role of the T6SS-4 in these wide-ranging functions has yet to be elucidated [122]. The T6SS-6 is highly conserved across B. pseudomallei and all Bcc species, leading Spiewak et al. to conclude that it is the prototypical T6SS of Burkholderia [123]. It is currently held that this system is used to impede the growth of other Gram-negative bacteria [123]. Very little is known about the T6SS-3 and -5; however, they along with the T6SS-2, -4, and -6 do not appear to play any significant role in virulence in mammalian models of infection [123,124,125]. For the sake of developing a cross-protective vaccine, it is worth noting that B. mallei has lost the entire T6SS-5 locus as well as seven T6SS-6 genes, likely rendering the T6SS-6 non-functional [121]. Additionally, only the T6SS-6 is broadly conserved across Bcc species and is thus the only T6SS that could theoretically be targeted for B. pseudomallei-to-Bcc cross-protection (Table 2) [123].
The T6SS-1 is the most well-studied T6SS in B. pseudomallei and B. mallei on account of its clearly defined role in virulence [124,125]. It was first identified in a screen for B. pseudomallei proteins that are upregulated during macrophage invasion and intracellular survival [126]. Around the same time, studies in B. mallei indicated that overexpression of the VirAG virulence-associated transcriptional regulatory system resulted in upregulation of the T6SS-1 [121]. In the absence of this system, B. mallei and B. pseudomallei’s ability to replicate in macrophages and induce the formation of multinucleated giant cells (MNGCs), a characteristic disease manifestation, is decreased [125,127]. To date, no cargo molecules have been identified for the T6SS-1. Instead, our current understanding of the T6SS-1’s mechanism of action indicates that the tip protein VgrG [118,119], and potentially PAAR as well [120], are directly responsible for inducing host cell membrane fusion with neighboring cells, leading to the formation of MNGCs.
To date, vaccination towards T6SSs in B. mallei and B. pseudomallei have primarily focused on the shaft protein of the T6SS-1, Hcp1 (BPSS1498), though the shaft proteins of the T6SS-2-6 as well as the TssM protein from the T66S-1 locus have also been tested [125,128]. These antigens are not generally thought to be exposed to the extracellular space for recognition by humoral immunity and would primarily be thought to serve as T cell antigens. Likely due to the generally held belief that both B and T cell immunity are needed for a vaccine to confer durable protection to melioidosis and glanders, most of the studies utilizing Hcp1 have also included a B cell antigen such as CPS or LPS.
5.1. T6SS Shaft Proteins
Multiple studies have demonstrated that Hcp1 is highly immunogenic, reacting particularly strongly to human melioidosis sera [125,129] and glanders sera from mice, horses, and one human [121,130]. PBMCs from melioidosis seropositive donors also respond to stimulation with Hcp1, correlating with patient survival [131]. By comparison, Hcp2 (BPSS0518), Hcp3 (BPSS2098), Hcp4 (BPSS0171), Hcp5 (BPSS0099), and Hcp6 (BPSL3105) are minimally reactive to human melioidosis serum [125], suggesting they may not be similarly immunogenic during an infection. Burtnick et al. screened these Hcp proteins as potential melioidosis vaccine antigens [125]. Female BALB/c mice were intraperitoneally immunized with one of each of the Hcp proteins formulated in SAS. The animals were subsequently challenged i.p. with ~50,000 CFU of B. pseudomallei strain K96243, ~50 times the median lethal dose. In this first trial, Hcp2 exhibited the greatest protective efficacy with 5 of 6 mice surviving to day 42 post-challenge. Hcp1, 3, 4, and 6 all had 2 or 3 animals survive to day 42, while Hcp5 did not confer any protection. However, in a subsequent replicate of this study using only Hcp1, 2, 3, and 6, Hcp6 conferred the highest level of protection with 4 of 12 animals surviving to day 42. In the Hcp1, 2, and 3 groups, only 2, 1, and 3 animals survived, respectively, which was approximately the same degree of protection as afforded by the adjuvant alone group in which 2 animals survived. This study appears to be the only published use of Hcp2 through Hcp6 in vaccination. Given that Hcp6 provided the highest level of protection in the follow-up experiment, even if only marginally, it may be worth revisiting as a potentially cross-protective antigen to Bcc species due to its extremely high degree of sequence conservation between B. pseudomallei and Bcc species (Table 2) [123].
Likely due to the T6SS-1’s clear role in virulence and Hcp1’s potent immunogenicity during natural infection, only Hcp1 has been explored further as a subunit vaccine antigen, despite the somewhat lackluster protection afforded by the protein in the initial screen [125]. In another study, Whitlock et al. tested whether B. mallei-derived Hcp1 was protective against a respiratory glanders challenge [70]. When female BALB/c mice were immunized i.n. with the antigen paired with CpG ODN 2395 and immune-stimulating complex AbISCO 100, mice were 80% protected from a 2x LD50 challenge of B. mallei strain ATCC 23344, in-line with the other antigens tested in this study (LolC, BimA, BopA) [70]. In this study, Hcp1 conferred markedly higher protection than reported in the Hcp screen [125], though it is unclear if this is due to the different pathogen, immunization/challenge routes, adjuvant systems, or challenge doses.
In recent years, Hcp1 has become a particularly popular component of glycoconjugate vaccines, either directly conjugated to B. thailandensis-derived LPS [56,57,58,59] or admixed with CPS that has been conjugated to a non-Burkholderia carrier protein [128,132]. Gregory et al. were the first to chemically link LPS to Hcp1, which was in turn linked to a gold nanoparticle vector. When this gold nanoparticle construct was administered in an intranasal immunization/challenge model in mice, it was noted that it elicited markedly higher LPS-specific IgG titers than when FliC or a tetanus toxoid carrier were used as carrier proteins [56]. When mice were subsequently challenged with a low dose of 1.9-2.6x LD50 B. mallei strain China7, 89% of the Hcp1-LPS-vaccinated animals survived, which was superior to but not significantly different from animals receiving the tetanus toxoid- or FliC-containing constructs [56]. In a subsequent experiment using a higher dose of 7x LD50 of the same challenge strain, the animals given the Hcp1-LPS formulation were 22% protected, largely in-line with the other two groups [56]. In both studies, the three groups receiving protein linked LPS exhibited significantly higher protection and lower spleen colonization than the negative control animals. However, the lack of any significant difference in protection between the Hcp1-LPS vaccinated group and those receiving the FliC or tetanus toxoid carriers may indicate that Hcp1-specific immune responses played little to no direct role in the observed protection, which was instead likely mediated primarily by LPS-specific immunity. Regardless, this study at least confirmed that Hcp1 could elicit robust T cell responses, as indirectly indicated by higher LPS-specific IgG titers.
Our group later used the same gold nanoparticle platform to test a wider array of B. mallei-derived carrier proteins for their ability to protect against melioidosis and glanders challenges when conjugated to LPS [57,58,59]. In all three of these studies, we did not observe any significant improvement in LPS-specific antibody titers afforded by the inclusion of Hcp1, at least compared to the other carrier proteins that were screened [57,58,59]. On the contrary, it was observed that the Hcp1-containing glycoconjugate elicited the lowest LPS-specific titers in one such study [59]. When a variety of these different constructs were tested for their ability to protect against a respiratory glanders challenge, the Hcp1 glycoconjugate elicited 89% protection, which was more-or-less in line with all the other carrier protein formulations that were assessed [58]. The lack of differences in protection against glanders afforded by the different carrier proteins may again suggest that protection is primarily being mediated by LPS-specific antibodies rather than carrier protein-specific responses. One of the primary arguments for including a strong T cell antigen such as Hcp1 in a B. mallei or B. pseudomallei vaccine formulation is that it may provide superior sterilizing immunity by providing additional protection from the intracellular phase of infection. However, in the glanders studies significant differences were not observed in spleen and lung colonization in surviving animals between the different carrier protein groups, and sterilizing immunity was not achieved [58]. When these constructs were used to immunize against a heterologous melioidosis challenge, the Hcp1-containing glycoconjugates failed to elicit any detectable protection [57,59].
Burtnick et al. paired B. pseudomallei-derived Hcp1 with a CPS glycoconjugate consisting of CPS chemically linked to the potent T cell antigen CRM197, a non-toxic mutant of diptheria toxin [128]. The authors demonstrated via IFN-γ ELISpot that PBMCs from mice immunized with Hcp1 or Hcp1 paired with CPS-CRM197 were strongly reactive to stimulation with the protein, further supporting Hcp1’s T cell antigenic properties [128]. When the authors assessed Hcp1’s protective efficacy in vaccination, they found that the combination of Hcp1 and CPS-CRM197 conferred 100% protection against an intranasal challenge of 10x LD50 B. pseudomallei strain K96243 compared to 67% in the CPS-CRM197 group, 30% in the Hcp1 group, and 0% in the adjuvant group [128]. This study thus provides some of the strongest support for the inclusion of Hcp1 in a subunit vaccine, demonstrating its protective efficacy with and without the B cell antigen CPS. Most surviving animals in both the combination and CPS-CRM197 groups had sterilizing immunity in the lungs, livers, and spleens, which may indicate that CPS-specific responses were primarily responsible [128]. Based on these results, Biryukov et al., recently performed a similar immunization study using CPS-CRM197+Hcp1 with or without AhpC, another robust T cell antigen discussed later [132]. Somewhat counterintuitively, the inclusion of AhpC lowered the degree of protection to an aerosol challenge from 80% to 50%, though both groups performed far better than the adjuvant alone group at 0% survival. Furthermore, bacterial loads in the spleens and lungs of these animals were significantly reduced 3 days post-challenge compared to controls [132]. When splenocytes from these two immunization groups were recalled with either AhpC or Hcp1, it was determined that AhpC was the more potent T cell antigen, as PBMCs recalled with AhpC elicited 1-2 orders of magnitude higher levels of various Th1- and Th2-associated cytokines compared to stimulation with Hcp1 [132]. Finally, these authors also noted that the Hcp1+CPS-CRM197 formulation conferred protection to B. pseudomallei strain MSHR5855 and a heterologous challenge with B. mallei strain FMH, though it is unclear to what extent Hcp1 was responsible for this strain and species cross-protection given the lack of Hcp1 and CPS-CRM197 alone controls [132].
Another novel approach to Hcp1 vaccination was recently conducted by Zhu et al., who were able to produce Staphylococcus aureus-derived membrane vesicles loaded with Hcp1 [133]. This was achieved by fusing the hcp1 sequence to the S. aureus pyruvate dehydrogenase E1 component subunit beta (pdhB), a protein known to be efficiently loaded into membrane vesicles. BALB/c mice were immunized with recombinant Hcp1 or the Hcp1-loaded vesicles, either alone or adjuvanted with Freund’s adjuvant. Animals were simultaneously immunized subcutaneously, intramuscularly, and intraperitoneally. Only the animals receiving the adjuvanted vesicles elicited significant Hcp1-specific antibody titers, though this did not translate to any significant difference in protective efficacy between the adjuvanted (70% survival) and unadjuvanted (60%) groups receiving the vesicles after a challenge with 5x LD50 of B. pseudomallei strain BPC006. Given that Hcp1 is predicted to mediate protection primarily through T cell responses, it may have been more relevant to perform ELISpots or similar assays using Hcp1-recalled splenocytes from the immunized animals. By comparison, the recombinant Hcp1 conferred no protection, though this can likely be attributed at least in part to the lack of adjuvant in this group. Finally, five days post-challenge, the adjuvanted and unadjuvanted groups receiving the vesicles had no colonization of the lungs, liver, or spleens [133].
Overall, there appears to be just as much evidence in support of including Hcp1 in a vaccine to B. mallei or B. pseudomallei as there is weighing against it. As previously stated, the wide range of efficacies reported for this protein could be due to a variety of reasons such as the route of immunization/infection, the adjuvant used, inclusion of other antigens, etc. That said, there is another potential drawback to using Hcp proteins as antigens that may provide additional context for why Hcp1-mediated protection appears so variable. Specifically, it has been demonstrated previously that the Hcp protein of the sci-1 T6SS in enteroaggegative E. coli is capable of spontaneously associating into hexameric rings even in the absence of other T6SS components [134]. Furthermore, these hexamers were capable of self-assembling into a shaft-like structure in an unregulated manner such that they could arrange in head-to-head, tail-to-tail, or head-to-tail orientations [134]. This finding has significant implications for using the protein as a subunit vaccine antigen, assuming Hcp1 is similarly capable of this spontaneous assembly. Even if Hcp1 is only being used as a soluble, unmodified antigen, the degree of spontaneous polymerization could result in significant batch-to-batch variability, and in turn, could significantly impact the pharmacokinetics and pharmacodynamics of the protein. For example, larger particulates are much more readily detected by innate immune cells than non-aggregated antigen due to size-dependent differences in immune recognition [135,136,137]. Furthermore, large particulates have a decreased ability to passively diffuse to secondary lymphoid tissues [135]. In the case of nanoparticle-vectored vaccines such as the gold nanoparticle platform, conjugation efficiency may be directly affected by the degree of Hcp1 polymerization. While these observations are speculative at this point, exploration of this matter appears justified if Hcp1 is to ever make it to regulated pre-clinical or clinical studies, particularly if post-expression modifications are to be utilized.
5.2. TssM (BPSS1512)
Outside of Burkholderia, TssM proteins are generally characterized as important structural components of the membrane-spanning complex of T6SSs. However, this function has not technically been confirmed for the TssM of B. mallei and B. pseudomallei’s T6SS-1 genomic locus. In fact, knocking out this gene had no effect on intracellular survival or MNGC formation in RAW 264.7 cells [138], suggesting this structural role may not be conserved in the T6SS-1. Instead, most studies of this protein have characterized it as a deubiquitinase that is secreted during the intracellular phase of infection [139]. This protein appears to be directly involved in interfering with NF-kβ and type I IFN responses of infected cells [139] and its secretion is reportedly mediated by the type 2 secretion system [140]. Human melioidosis serum and PBMCs react strongly to TssM, and the latter appears to correlate with survival [63,131]. Burtnick et al. have performed the only vaccination study using TssM, pairing it with CPS-CRM197 [128]. Upon intranasal challenge, the TssM+CPS-CRM197-immunized animals were 80% protected, which was in-line with the group receiving only CPS-CRM197, which were 70% protected. By comparison, the Hcp1+CPS-CRM197 group was 100% protected, though a significant difference between these groups was not evident. Animals receiving TssM alone were only 20% protected, which was comparable to the animals receiving only Hcp1 [128]. Together this supports the notion that CPS-specific immunity is primarily responsible for the reported protection, and that the inclusion of TssM appears to be only slightly beneficial.
6. Type IV Pili
Type IV pili are long, thin projections that are involved in several functions such as mediating twitching and gliding motility, uptake of DNA, and adherence to eukaryotic cells. These structures primarily consist of repeating subunits known as major pilins, though minor pilins are also present at lower abundance [141]. Like flagellin, polymerized pilin subunits may be particularly antigenic to B cells on account of their repetitive pattern and distance from the bacterial glycocalyx. The B. pseudomallei genome harbors eight distinct type IV pilus-associated loci (TFP1-8) that remain only partially characterized [142]. To date, only the minor pilin subunit of TFP7 (BPSS1593; PilV) has been tested in a subunit vaccination approach. While the authors reported that PilV was particularly immunogenic, subcutaneous vaccination with the B. pseudomallei strain K96243-derived protein did not protect BALB/c mice from an intraperitoneal challenge of B. pseudomallei strain G207 [143]. This failure to confer protection could be due to a multitude of reasons, such as structural differences in the recombinantly expressed protein or low expression of the TFP7 locus in mouse infection models. As it stands, PilV does not appear to be an appropriate subunit vaccine candidate, though the other type IV pilus proteins remain to be characterized.
7. General Outer Membrane Proteins: β-Barrels
Not including multi-subunit complexes such as flagella and the various secretion systems, the outer membrane proteome of Gram-negative bacteria largely consists of two major classes of proteins: lipoproteins (discussed in the following section) and proteins containing a β-barrel motif [144,145]. As the name suggests, β-barrels consist of 8-36 antiparallel β-strands that loop back and forth through the outer membrane, arranging into a cylindrical “barrel” shape held together by hydrogen bonds [144]. The core of these proteins is generally hollow, serving as a channel for various cargos (nutrients, waste products, antibiotics, toxins, etc.) in both passive and active transport. The periplasmic and extracellular regions that link the β-strands are often referred to as “loops.” Such loops at the extracellular face are the most likely regions to serve as functional B cell epitopes, given their relative accessibility to the extracellular environment. Conversely, the transmembrane barrel and periplasmic loops would primarily be thought to serve as T cell epitopes.
Because these proteins are embedded in the outer membrane, they consist of a relatively high number of hydrophobic, externally oriented residues, making it difficult to express these recombinant proteins due to solubility issues. Further complicating matters, natively expressed outer membrane β-barrels generally require specialized bacterial machinery, known as the β-barrel-assembly machinery (BAM) complex, as well as a class of chaperones known as holdases to be stably inserted into the outer membrane in their native conformation [144,146]. Structural biologists interested in β-barrels have occasionally been able to circumvent these issues using detergents [147,148], liposomes [149], or lipid nanodiscs [148,150], which mimic the environment created by the bacterial outer membrane and associated holdases. As one might expect from these structural limitations, the study of β-barrel proteins as subunit vaccine antigens is particularly challenging. In the absence of stabilizing agents, protein yields tend to be exceedingly low, the resultant protein is prone to aggregation, and any protein recovered may not be expected to maintain a native-like three-dimensional structure. From an immunological perspective, these last two issues are particularly problematic for B cell responses, as many of the epitopes recognized by B cells are non-contiguous and would not be expected to be retained in aggregated or misfolded protein. Some animal vaccination studies have included detergents to stabilize β-barrel proteins, though many detergents are of questionable biocompatibility and only polysorbate 20 and 80 have been included in FDA-approved vaccines. Furthermore, it is unclear if detergents can retain the protein’s folding conformation in vivo long enough for the antigen to engage with B cell receptors. Despite these limitations, quite a few β-barrel proteins have been explored as subunit vaccine antigens for melioidosis and glanders vaccines.
7.1. BamA (BPSL2151)
The first such protein studied as a Burkholderia subunit vaccine antigen was an Omp85 family protein (BPSL2151) [151]. Su et al. initially performed a high-throughput screen for human melioidosis serum-reactive peptides derived from B. pseudomallei strain D286. They ultimately identified 109 reactive peptides, one of which was derived from BPSL2151 [152]. Protein BLAST of the BPSL2151 sequence largely implicates this Omp85 family protein as being the Burkholderia homolog of BamA [153], one of the most highly conserved proteins found in the outer membrane proteome of virtually all known Gram-negative bacteria [154]. This high conservation is due to the integral role of the BAM complex, which serves to mediate insertion of other β-barrel proteins into the outer membrane [154]. Studies in E. coli have demonstrated that bacteria are not viable without this system [155], likely due to the vital functions of the myriad β-barrel proteins expressed on the outer membrane. The BamA protein has thus been explored recently as a potential vaccine candidate and target of monoclonal antibodies in Acinetobacter baumannii [156,157] and pathogenic E. coli [158,159160].
Su et al. expressed the B. pseudomallei strain D286-derived full-length protein in E. coli, resulting in inclusion bodies that had to be denatured and refolded [151]. While the protein was not verified to be refolded into a native-like structure, the study demonstrated that human melioidosis serum reacted to the recombinant protein [151]. However, this does not confirm proper folding of the protein, as the reactive antibodies could be limited to linear B cell epitopes on BamA. Given that this protein was first identified by serum reactivity to such linear B cell epitope peptides, melioidosis serum may be expected to react to recombinant BPSL2151 whether it is properly folded or not. Most structural studies of E. coli BamA report that detergents or lipid-based carriers are required to maintain the protein in a native-like form. Furthermore, these studies often do not utilize full-length BamA, instead truncating it such that the bulky periplasmic domains are removed [147,149,150,160]. This vaccination study did not report using any such stabilizing agent nor did they truncate the protein, suggesting the protein may not have been conformationally intact [151].
Regardless, they immunized BALB/c mice intraperitoneally with the recombinant protein and Freund’s adjuvant before challenging intraperitoneally with 10x LD50 of B. pseudomallei strain D286. Immunized animals were afforded 70% protection whereas only 10% of the control animals survived. Spleen, lung, liver, and blood colonization was significantly reduced in the vaccinated group, though sterilizing immunity was not achieved. Finally, the authors provided a potential mechanism of immunity by demonstrating that polyclonal antibodies from the vaccinated animals were capable of opsonizing bacteria and were bactericidal in the presence of complement, suggesting involvement of the classical complement activation pathway [151]. Interestingly, a BamA-specific monoclonal antibody has been described in E. coli which was bactericidal even in the absence of complement. This antibody was capable of directly antagonizing BamA’s ability to mediate the insertion of other β-barrel proteins into the outer membrane, resulting in cell death [159]. We speculate that such direct bactericidal activity in the absence of complement may be achievable using Burkholderia-derived BamA if conformational B cell epitopes can be maintained.
Despite these promising initial results and an interest in BamA in other Gram-negative bacteria, Burkholderia BamA has received little further interest as a subunit vaccine component. Of all the surface-expressed proteins discussed in this review, BamA exhibits the greatest degree of sequence conservation between the Bpc and Bcc species (Table 1), and thus represents one of the most promising targets for humoral-mediated cross-protection to these pathogens. Specifically, the Bcc homologs of BamA had an average sequence identity and similarity of 94.0% and 97.0%, respectively, compared to B. pseudomallei-derived BamA (Table 1). In fact, when limit this analysis of sequence conservation to only the extracellular loops of BamA, Bcc-to-B. pseudomallei sequence identity increases to 98.2%, indicating that such potential antibody targets are almost entirely conserved across pathogenic Burkholderia species (Figure 2a,b; Table 3). A significant caveat to this is that BamA’s conservation is so high that its use as a vaccine antigen may impact the healthy gut microbiome, as has been reported when mice were immunized with A. baumannii BamA [157]. Su et al. did address this concern to some extent, showing that the B. pseudomallei BamA immunization serum did not cross-react to E. coli [151], though future studies should expand on this by directly looking at potential changes in the gut microbiome of vaccinated mice. Despite this potential drawback, revisiting BamA as a Bcc cross-reactive antigen appears justified.
7.2. OmpW1 (BPSL1552)
Another outer membrane β-barrel protein that has been explored as a potential Burkholderia subunit vaccine antigen is a member of the OmpW family of proteins. Unlike most of the proteins discussed in this review, this protein was first identified in Bcc species and subsequently tested as a Bcc-specific vaccine antigen. McClean et al. found this antigen upon performing a screen for B. cenocepacia and B. multivorans proteins involved in adhesion to cystic fibrosis-like lung epithelia cells (CFBE41o-) [164]. They demonstrated that expression of the B. multivorans strain LMG13010 OmpW-like protein in E. coli resulted in significantly increased adherence to CFBE41o- cells, and that pooled serum from Bcc-colonized cystic fibrosis patients was reactive to the protein. Intraperitoneal immunization of BALB/c mice with this protein and alum followed by intranasal challenges with either B. cenocepacia strain BC7 or B. multivorans strain LMG13010 resulted in a significant reduction in lung colonization but not spleen colonization. The authors also demonstrated that splenocytes from immunized animals could be recalled with recombinant OmpW1, resulting in secretion of IFN-γ, IL-4, IL-17, and IL-6 [164]. This would indicate that Th1, Th2, and Th17 responses may have been elicited by the protein, a somewhat unexpected finding given the highly Th2-skewing properties of alum.
Given these promising results as well as the relatively high level of sequence similarity to the B. pseudomallei homolog, it was explored if the B. pseudomallei homolog was protective against a B. pseudomallei challenge. BALB/c mice were immunized intraperitoneally with the protein homolog and either SAS or alum and subsequently challenged intraperitoneally with B. pseudomallei strain 576. OmpW1 paired with either alum or SAS did not confer any long-term protection, though the group that received OmpW1 with SAS did experience a delay in median time of death. However, the SAS alone control suggests this was primarily due to effects of the adjuvant rather than the antigen, as they also had a marked increase in median time of death. A subsequent experiment using an i.p./i.p. model in C57BL/6 mice demonstrated more promising results, with OmpW1+SAS affording 75% protection on day 80 compared to 12.5% in the adjuvant alone group [165]. Splenocytes from vaccinated animals secreted significant levels of IFN-γ upon ex vivo stimulation with OmpW1, suggesting a strong Th1 response was mounted [166]. A notable downside of these experiments is that the study reported significant issues with protein solubility that could only be overcome by solubilizing the protein in urea. Running their protein on a gel without urea resulted in significant smearing, which may be indicative of protein aggregation [165]. Given this information, it is possible that the somewhat lackluster results from protection studies in BALB/c mice was a direct result of the protein not being in its native-like, unaggregated state. Such issues could potentially be addressed by the inclusion of a stabilizing detergent or other membrane-mimicking platform. Overcoming these potential technical problems appears warranted, as OmpW1 is another one of the highly conserved proteins between Bpc and Bcc species. This protein was identified in all the Bcc strains in our dataset and exhibited an average Bcc-to-B. pseudomallei sequence identity of 78.3% and similarity of 88.0% (Table 1). However, when this analysis was restricted to only the extracellular loops, sequence identity drops markedly to 67.7%, potentially limiting this protein’s usefulness as a cross-reactive antibody target (Figure 2c,d; Table 3).
7.3. OpcP (BPSS0879), OpcP1 (BPSS0708), and OmpW2 (BPSL2704)
As mentioned already, our group has bioinformatically identified a variety of B. mallei-derived proteins that are predicted to be highly antigenic and surface-exposed [57]. We have tested a handful of these proteins as gold nanoparticle-coupled LPS glycoconjugate carrier proteins, including three general β-barrel proteins: OpcP, OpcP1, and OmpW2 [58,59]. Very little is known about these proteins, though OpcP has been functionally characterized as a general diffusion porin [167]. All three of the B. mallei strain ATCC 23344-derived β-barrel glycoconjugate constructs elicited robust LPS-specific IgG titers, confirming the proteins’ T cell antigenic properties [58,59]. When these various glycoconjugate vaccine formulations were tested for their ability to protect against a low dose of intranasally-delivered B. mallei strain ATCC 23344, we found that the OpcP and OmpW2 constructs were the only ones that elicited complete protection, though statistically significant differences were not observed between them and the other constructs. In both groups, more than half the surviving animals did not exhibit lung colonization at the endpoint of the study. The OpcP1 construct only performed marginally worse at 77% protection [58]. When the challenge dose was increased, OpcP and OmpW2 both elicited significant levels of protection with a 77% survival rate. Most of the surviving animals exhibited colonization of the lungs, liver, and spleen of these animals, suggesting dissemination was not prevented [58]. It is worth noting that while B. mallei strain ATCC 23344 harbors the opcP1 gene, 21 of the 30 B. mallei isolates queried in our bioinformatic screen did not, calling into question OpcP1’s potential for use in a glanders vaccine (Table 1). Whether this is related to OpcP1’s relatively reduced performance compared to OpcP and OmpW2 is currently unclear.
We have also used these constructs in a similar immunization scheme followed by a heterologous challenge with B. pseudomallei strain K96243. In contrast to the findings in the B. mallei challenge model, the OmpW2 construct failed to elicit any meaningful protection against melioidosis [59]. The OpcP and OpcP1 constructs on the other hand were capable of conferring protection to a lethal B. pseudomallei challenge at 90% and 30% survival, respectively. Furthermore, protection elicited to these constructs was mostly sterilizing, with most animals having no detectable bacteria in the lungs, liver, and spleen at the experimental endpoint. Upon repeating this heterologous challenge study, vaccination with the OpcP or OpcP1 constructs resulted in 90% and 80% protection, respectively. Organ colonization data was like the first trial, though sterilizing immunity was not achieved in the lungs [59]. Because the other constructs tested in this study failed to confer protection to B. pseudomallei despite robust LPS-specific antibody titers, it is believed that OpcP- and OpcP1-specific immune responses are playing a significant role in protection to respiratory melioidosis. Our lab is still investigating whether such protection is mediated by protein-specific antibody or T cell responses; however, we noted that splenocytes recalled with OpcP or OpcP1 elicited significant levels of IFN-γ, IL-17, and IL-2, indicating robust Th1 and Th17 immunity had been elicited by the vaccines [59].
OpcP and OpcP1 are also notable for being moderately well-conserved in Bcc species, both having ~76% Bcc-to-B. pseudomallei sequence identity and ~85% similarity (Table 1). OpcP is also reportedly one of the most highly expressed porins in B. cepacia and B. cenocepacia [168,169], suggesting it is highly expressed for immune recognition. However, the conservation of the extracellular loops of these proteins is considerably lower at 69.8% identity for OpcP1 and 59.8% for OpcP (Table 3). While OmpW2 and its extracellular loops are remarkably well conserved across most Bcc species, B. multivorans notably lacks the gene (Table 1 and Table 3). Given that B. multivorans is one of the most clinically relevant species in the Bcc, OmpW2’s potential as a universal Burkholderia vaccine candidate is likely limited.
7.4. Bucl8 (BPSL1972)
Resistance nodulation cell division (RND) efflux pumps are tripartite complexes that play a significant role in extruding clinically relevant antibiotics in Gram-negative bacteria. These pumps assemble as homotrimers and span the outer membrane with a trimeric β-barrel motif. Given their role in antibiotic resistance, they are particularly popular targets for therapeutic development. Three such pumps have been described in B. pseudomallei and B. mallei [170], though none of these have been tested as vaccine targets. However, one group has recently identified a putative new RND efflux pump, Bucl8, in a screen for B. pseudomallei and B. mallei proteins containing collagen-like motifs of repeating GXY residues [171]. While it was subsequently determined that these collagen-like motifs allow Bucl8 to bind fibrinogen, it was not determined whether this plays any role during infection [172]. Fusaric acid is currently the only known substrate of Bucl8 [172].
Likely recognizing the inherent difficulty of recombinantly expressing a multimeric integral membrane protein, the study instead opted to try a peptide-based vaccination strategy [173]. They first used ElliPro to bioinformatically predict that the Bucl8 extracellular β-barrel loops could serve as linear B cell epitopes for antibody binding. Then, immunization studies were performed with each of Bucl8’s two extracellular loop peptides chemically conjugated to an inactivated diphtheria toxoid carrier protein and formulated in AddaVax. These vaccines delivered subcutaneously were capable of eliciting robust, IgG1-skewed antibody titers that reacted to the respective peptide or an attenuated B. pseudomallei strain [173]. However, this subcutaneous immunization was not protective against a heterologous intranasal challenge of 8 x 106 CFU of B. thailandensis strain E264 [174]. This result can likely be attributed at least in part to the lack of mucosal immunity that would be expected from such an immunization scheme. A follow-up experiment was performed in which CD-1 mice were intranasally immunized with diphtheria toxin linked to one of the two Bucl8 peptides and fluorinated cyclic diguanosine monophosphate, a STING agonist that has not been previously used as an adjuvant in any Burkholderia vaccine studies. These animals were challenged intranasally with 8 x 105 CFU of B. thailandensis strain E264 expressing a luciferase gene. The vaccinated animals had significantly reduced weight changes, significantly lower luminescent signals at the site of infection at day 7, and significantly lower bacterial burdens in the lungs and nasal wash at days 3 and 7, respectively [174]. While significant differences in protection were not reported, this is not entirely surprising given that the studies were done in an outbred mouse model using a heterologous challenge model and only immunizing with a single peptide epitope. The fact that any protection at all was observed in such a model is quite promising, and it would be interesting to see how this antigen performs when administered as a full protein or against a B. pseudomallei challenge. Future studies should confirm that Bucl8 vaccination is protective in more clinically relevant models of glanders, melioidosis, or even Bcc colonization of the lung. Bcc cross-protection studies may be particularly warranted given that Bucl8 is broadly conserved in Bcc species and exhibits a relatively high level of sequence conservation in the extracellular loops at 88.0% sequence identity (Table 1 and Table 3).
8. General Outer Membrane Proteins: Lipoproteins
Besides the integral β-barrel proteins, lipoproteins make up the other major class of proteins expressed on the Gram-negative outer membrane. Because most lipoproteins are expressed on the cytoplasmic membrane or on the inner leaflet of the outer membrane, lipoproteins are a relatively underappreciated class of potential antibody targets in Gram-negative bacteria. While early studies identified a handful of surface-exposed lipoproteins in E. coli [175,176], Treponema pallidum [177], and Borrelia burgdorferi [178], such examples were generally regarded as anomalous. However, recent evidence suggests that such surface-exposed lipoproteins are much more widespread than previously believed, and thus represent an emerging field of study that is particularly relevant for vaccine researchers working with Gram-negative bacteria [179]. While outer membrane lipoproteins are relatively easy to identify bioinformatically due to their differences in signal sequence, it is not so easy to determine which outer membrane lipoproteins are surface exposed, and instead they must be identified empirically. Schell et al. performed studies to characterize the outer membrane proteome of B. pseudomallei and B. mallei, and identified several lipoproteins in the process, but the approach employed would not necessarily distinguish between surface- and periplasm-facing lipoproteins [180]. However, Sousa et al., recently performed surface trypsin shaving studies of B. cenocepacia strain J2315 and identified 55 surface-exposed proteins [181]. When we analyzed these proteins further using DTU Health Tech’s SignalP v6.0 program [182], we found that 11 were predicted lipoproteins. These proteins corresponded to the following locus tags: BCAL0340, BCAL1493, BCAL2166, BCAL1893, BCAL0151, BCAL2687, BCAL3204, BCAL2645, BCAL1288, BCAL1881, and BCAL0304. The BCAL3204 predicted lipoprotein is particularly noteworthy, as the B. pseudomallei homolog is extremely well-conserved and has shown great promise as a melioidosis vaccine antigen [183,184,185]. Another of these proteins, BCAL2645, has yet to be used in a Burkholderia vaccine to date, though it seems a particularly strong candidate given some of its properties which will be discussed here.
8.1. Omp7 (BPSL765)
The first study of a Burkholderia lipoprotein antigen was performed by Hara et al. [183]. The authors bioinformatically identified 12 OmpA family proteins (designated Omp1-12) and attempted to purify them as recombinant antigens to test for melioidosis serum reactivity and as subunit vaccine antigens. Of these 12 proteins, five are predicted by SignalP v6.0 to be lipoproteins including Omp1 (BPSL0999), Omp4 (BPSL2062), Omp5 (BPSL1659), Omp7 (BPSL2765), and Omp9 (BPSS0909) [182]. Only 6 of the 12 proteins were successfully cloned from B. pseudomallei strain D286 and expressed in E. coli, including the putative lipoproteins Omp4, Omp5 and Omp7. Omp7, as well as the non-lipoprotein Omp3 (discussed later), reacted particularly strongly to human melioidosis serum and as a result were down selected for testing as vaccine antigens. BALB/c mice were immunized intraperitoneally with Omp3 or Omp7 in Fruend’s adjuvant before challenging i.p. with 10x LD50 of B. pseudomallei strain D286, resulting in 50% protection in both vaccination groups compared to 0% in the control animals [183].
Given these promising results, Omp7 has been utilized in subsequent vaccination studies. One such study used a similar model as Hara et al., albeit with a much higher challenge dose of 100x LD50 and using SAS as the adjuvant [184]. In this study, Omp7 was pooled with a trio of T cell antigens associated with chronic infection: IscA, TadE, and AcoD (discussed later). This antigen pool was further mixed with either the well-studied T cell antigen LolC (discussed later), the potent B cell antigen CPS, or neither. Between 30 and 50% protection level was observed, depending on the exact combination of antigens, compared to 0% in the control animals [184]. However, given the complex nature of these formulations, it is impossible to ascertain to what extent Omp7 was responsible for this protection.
More recently, Dyke et al. extensively characterized Omp7, identifying it as a peptidoglycan-associated lipoprotein (Pal) [185]. The authors went on to show that B. mallei Omp7 is required for virulence, with the isogenic mutant being largely attenuated in BALB/c mice. The mutant also appeared to be more susceptible to osmotic stress, polymyxin B, and killing by macrophages, potentially due to disruptions in the cell envelope given Pal’s known role in cell envelope stability in other species [185]. The study also demonstrated that intranasal vaccination with B. mallei derived Omp7 expressed by a PIV5 vector elicited 80% long-term protection to an intranasal challenge with 10x LD50 B. mallei strain ATCC 23344, though sterilizing immunity was not achieved in most animals, with both lungs and spleens harboring bacteria [185]. Studies outlining the mechanism of protection elicited by Omp7 are somewhat sparse, though a study by Gourlay et al. provides some insight [186]. Generating antibodies against a specific epitope of Omp7, they demonstrated that such antibodies promoted the killing of B. pseudomallei by neutrophils and caused the bacteria to agglutinate [186]. While Pal-like proteins are generally thought to be found on the periplasmic side of the outer membrane, these functional antibody studies may indicate that some proportion of Omp7 is located on the extracellular face. This would align with studies of Pal from E. coli [187] and Haemophilus influenzae [188], which have been shown to be expressed on both the periplasmic and extracellular side of the outer membrane.
Running in parallel with the Bpc studies, the B. cenocepacia homolog BCAL3204 has also been intensely studied. This protein is one of the most highly immunodominant B cell antigens in Bcc species, being readily recognized by sera from human CF patients with Bcc colonization of the lung [189]. This was confirmed by Makidon et al. by performing immunization studies with a nanoemulsion containing outer membrane proteins purified directly from B. cenocepacia. One of the most immunodominant proteins in this formulation was determined to be BCAL3204 [190]. Subsequent studies characterizing the protein found that an isogenic B. cenocepacia mutant had increased susceptibility to polymyxin B, caused reduced IL-8 secretion in CFBE41o- cells, and had a reduced ability to adhere to CFBE41o- cells [191]. Additionally, it has been demonstrated that sera from human CF patients with Bcc colonization of the lung is cross-reactive to peptides derived from B. pseudomallei Omp7, providing strong empirical evidence for the use of this antigen as a pan-Burkholderia vaccine antigen [192]. Our own screen for potential cross-reactive antigens adds additional support, as Omp7 is the third most conserved Burkholderia surface antigen discussed in this review, with the Bcc homologs exhibiting an average of 85.5% identity and 93.1% similarity to the B. pseudomallei strain K96243 ortholog (Table 1; Figure 3a,b).
8.2. Omp1 (BPSL0999)
While Hara et al. was ultimately unable to express Omp1 nor has it ever been used in a published Burkholderia vaccine study to date, recent evidence in support of it warrants discussion. This protein was first identified as a seroreactive marker in high-throughput screens of sera from human melioidosis patients [63,64] and a human glanders patient [193]. Additionally, Omp1-derived peptides were found to be some of the most robust stimulators of IFN-γ production by PBMCs derived from melioidosis seropositive donors, with nearly all donors responding to the stimulus [194]. The B. cenocepacia homolog, BCAL2645, is another one of the lipoproteins identified via Sousa et al’s surface trypsin shaving study, providing evidence that it is a potential surface-exposed target for antibody binding [181]. Further, Seixas et al. recently demonstrated that polyclonal antibodies towards BCAL2645 impair adhesion and invasion of CFBE41o- cells by B. cenocepacia [195]. The ability of these antibodies to impede adherence suggests a possible mechanism of protection if this antigen were to be used in vaccination. Additionally, Omp1 is the second most well-conserved surface antigen between Bpc and Bcc species discussed in this review, following only BamA (Table 1). The Bcc homologs of this protein appear to have an average sequence identity of 93.4% and similarity of 95.9% when compared to the B. pseudomallei strain K96243 sequence (Table 1; Figure 3c,d). Unlike BamA, Omp1 does not appear to be widely conserved across Gram-negative species, making it far less likely to have an impact on the healthy human microbiome. Together, this implicates Omp1 as a top candidate for a pan-Burkholderia vaccine, assuming protective efficacy can be demonstrated in an active immunization model.
9. ABC-Binding Cassette (ABC) Transporters
ABC transporters are a superfamily of multicomponent protein complexes found in all kingdoms of life whose purpose is to transfer various cargo molecules across cell membranes. ABC transporters share a similar architecture consisting of two hydrophobic integral membrane domains and two highly conserved, hydrophilic nucleotide-binding domains within the cytoplasm that hydrolyze ATP to provide energy to the system [197,198,199]. In Gram-negative bacteria, ABC importers also associate with a periplasmic receptor protein that binds the cargo prior to transport while ABC exporters associate with a membrane fusion protein and an outer membrane factor that serve as a pore for cargo to reach the extracellular environment [197,198,199]. In bacteria, ABC systems are known to play vital roles in survival and virulence with functions relating to nutrient uptake, toxin/antibiotic efflux, lipoprotein localization to the outer membrane, and siderophore secretion [199]. From an immunological perspective, only the outer membrane factor component of ABC exporters is exposed to the extracellular space for antibody binding. All other components of ABC transporters in Gram-negative bacteria are localized to the periplasm, inner membrane, or cytoplasm and would therefore be expected to act primarily as T cell antigens when used as vaccine antigens. To our knowledge, there are currently no examples of vaccination with the outer membrane factor of a Burkholderia ABC exporter.
Harland et al. created a comprehensive annotation of B. pseudomallei and B. mallei ABC proteins [200]. Based on this annotation, they selected three ABC transporter proteins to test as potential subunit vaccine antigens: LolC (BPSL2277), PotF (BPSS0467), and OppA (BPSS2141) [201]. LolC is one of the integral membrane components of the LolCDE ABC exporter which has been extensively studied in E. coli [202]. This ABC exporter is essential to most Gram-negative species, as it is required for transporting lipoproteins from the inner membrane to the outer membrane [202]. PotF is the periplasmic receptor protein of PotFGHI, an ABC importer of putrescine [203]. Notably, B. mallei has lost the direct homolog of this potF gene through genome reduction. Lastly, OppA is the periplasmic receptor protein of the OppABCDF importer of oligopeptides. Such oligopeptides serve as an amino acid and carbon source for multiple species of bacteria [204,205]. To demonstrate the proteins’ potential as T cell antigens, the group challenged naïve BALB/c mice with B. pseudomallei strain 2D2 and stimulated splenocytes recovered from the animals’ ex vivo with recombinant LolC, PotF, or OppA. Antigen recalled splenocytes secreted detectable levels of IFN-γ, suggesting robust Th1 responses to these proteins over the course of natural infection, a finding that has been replicated by others [201,206,207,208].
Harland et al. also demonstrated the antigens’ protective potential by intraperitoneally vaccinating BALB/c mice with each individual antigen paired with monophosphoryl lipid A-trehalose dicorynomycolate as an adjuvant before challenge via the i.p. route with 40x LD50 of B. pseudomallei strain K96243. Mice immunized with PotF or LolC displayed 50% and ~85% survival, respectively, while OppA vaccination afforded only ~20% protection [201]. In a follow-up experiment in which LolC was paired with different adjuvants, the authors reported that the same LolC/adjuvant combo resulted in only 33% protection, though this could be attributed to the higher challenge dose of 70x LD50. The other adjuvants tested with LolC included various combinations of Emulsigen, CpG ODN 10103, and AbISCO-100, all of which were administered subcutaneously instead of intraperitoneally. The protection afforded by these different formulations ranged from 17 to 66% with the higher challenge dose [201].
A subsequent study performed by Scott et al. found that LolC paired with Alhydrogel and CpG ODN 2006 did not protect BALBc mice from mortality when administered subcutaneously followed by an intraperitoneal challenge [209]. They did, however, report a significant increase in median time of death from 3.5 to 10 days, suggesting at least some level of protection afforded by the antigen. This group also found that LolC paired with B. pseudomallei CPS provided slightly improved protection and reduced disease severity compared to mice vaccinated with CPS alone, though statistically significant differences were not reported. That said, vaccination with CPS alone was significantly more protective than vaccination with LolC alone, again supporting the dominant role of humoral immunity in protection in murine models of infection [209].
Finally, Whitlock et al. compared LolC’s protective efficacy to two other Burkholderia subunit antigens, BimA and BopA, in a clinically relevant intranasal immunization and challenge model. Using a cationic liposome-DNA complex as an adjuvant, the results mirrored that of Scott et al., finding that none of the LolC-immunized mice survived a 2x LD50 challenge with B. pseudomallei strain 1026b. They likewise found that LolC-vaccinated mice had an increase in median time of death with all mice surviving for 1-7 weeks compared to the naïve control group which all succumbed within a week of infection. By comparison, mice immunized with BimA and BopA were approximately 20% and 60% protected with similarly increased median times of death [70]. Interestingly, when the authors attempted a heterologous challenge with B. mallei strain ATCC 23344, LolC-vaccinated mice were 83% protected [70]. These data would suggest that LolC is only mildly protective in melioidosis models, though perhaps shows promise as a protective glanders antigen. Because of its essential function in shuttling lipoproteins to the outer membrane, LolC is unsurprisingly one of the most highly conserved antigens between Bpc and Bcc species discussed in this review at 93.3% sequence identity and 97.1% sequence similarity (Table 2). As such, LolC may be another candidate for the development of a pan-Burkholderia vaccine.
10. Miscellaneous Antigens
10.1. Omp3 (BPSL2522)
In the Hara et al. study mentioned previously, they identified two OmpA domain-containing proteins, Omp3 and Omp7, that reacted strongly to human melioidosis serum and conferred similar levels of protection in i.p./i.p. vaccination/challenge murine models [183]. Despite this, only Omp7 received further attention as a potential candidate vaccine antigen while Omp3 has only seen use as a serodiagnostic marker, with numerous studies identifying it as highly reactive to human melioidosis serum [63,64,65,183,196,210] and horse glanders serum [130]. In addition to these B cell antigenic properties, PBMCs derived from healthy melioidosis seropositive donors and recovered melioidosis donors react strongly to Omp3-derived peptides, suggesting that robust T cell-immunity is also mounted to this protein during natural exposure to B. pseudomallei [194].
Despite lacking typical motifs observed in outer membrane proteins of Gram-negative bacteria (i.e., β-barrel domain or lipobox sequence), there is evidence to suggest that this protein is expressed on the outer membrane surface and is therefore a potential target for humoral immunity. Schell et al. identified Omp3 in trypsin shaving studies of B. pseudomallei and B. mallei outer membrane preparations, confirming it as an outer membrane-associated protein [180]. Additionally, the highly conserved B. cenocepacia homolog, BCAL2958, is one of the 55 proteins identified in Sousa et al’s surface trypsin shaving studies, strongly suggesting it is extracellularly oriented, at least in B. cenocepacia [181]. Looking closer at the primary sequence of this protein, we noted the presence of an N-terminal patch of hydrophobic tryptophan residues that was distant from the highly hydrophilic C-terminal OmpA domain. Such tryptophan-rich patches are known to occasionally serve as membrane anchoring motifs [211], providing a potential explanation for the proteins apparent surface localization.
The B. cenocepacia Omp3 homolog has gained attention in recent years as a potential therapeutic target for Bcc colonization of the CF lung. Two separate studies investigating this protein demonstrated that it is highly reactive to serum from CF patients that are positive for Bcc lung colonization [181,212]. They also noted that antibodies to B. cenocepacia-derived Omp3 are highly cross-reactive to 12 strains of Bcc from seven distinct species, suggesting it is highly conserved [212]. In fact, our own analysis confirms that Omp3 is extremely well-conserved not just among all the Bcc species in our dataset, but also between the Bpc and Bcc groups (Table 1). Of all the putative targets of humoral immunity discussed here, Omp3 ranks as having the fourth highest degree of sequence conservation, with the Bcc homologs of Omp3 exhibiting an average of 91.6% sequence identity and 92.9% sequence similarity to Bcc homologs (Table 1; Figure 3e,f). It has also been demonstrated that recombinant B. cenocepacia-derived Omp3 is highly stimulatory to healthy human primary neutrophils, causing them to release significantly higher levels of TNFα, elastase, nitric oxide, catalase, and myeloperoxidase [212]. The mechanism and purpose of this function is still unknown. Considering how highly conserved this protein is across Burkholderia species, it seems likely that it plays some other role related to survival in the environment or cell envelope integrity/barrier function. The immunodominance of this protein in Bpc- and Bcc-associated diseases, its surface localization, and its high degree of Bpc-to-Bcc sequence conservation make it a key target for a pan-Burkholderia vaccine.
10.2. MprA (BPSS1993)
MprA is a secreted serine protease generally regarded as the most active secreted protease produced by B. pseudomallei [213,214,215]. However, mprA expression appears to be primarily restricted to the stationary growth phase and the encoded enzyme does not appear to play any significant role in virulence in mouse models of infection [214]. Furthermore, melioidosis patient sera is only mildly reactive to the protein with less than half of samples reacting to it in one study [196]. As such, MprA may not be an ideal target for neutralizing antibodies. Chin et al. immunized BALB/c mice with MprA in Freund’s adjuvant, resulting in high antigen-specific antibody titers [215]. Animals challenged i.p. with 10x LD50 B. pseudomallei strain D286 had significantly increased survival times compared to the control animals but succumbed to infection past day 25. These animals exhibited bacteremia, splenomegaly, and abscess formation after the challenge, indicating that the bacteria were not prevented from disseminating [215]. As such, MprA appears to confer some low level of protection, at least in i.p. models of infection in mice, but is overall less protective than many of the other antigens discussed in this review. Furthermore, an MprA homolog was not identified in any of the B. mallei proteomes in our dataset, nor was it identified in any Bcc species except B. stagnalis and B. ubonensis, neither of which are particularly relevant to human disease. As such, MprA has little potential as a glanders or Bcc cross-reactive antigen.
10.3. AhpC (BPSL2096)
Alkyl hydroperoxide reductases such as AhpC in B. pseudomallei are frequently upregulated in bacteria during the intracellular phase of infection to survive host-derived hydrogen peroxide [216]. As a result, natural exposure to such intracellular bacteria is often associated with robust immune responses to these proteins, making them popular candidate antigens for a variety of pathogenic bacteria [217,218,219]. This trend is also observed with B. pseudomallei AhpC, which acts as a highly serodiagnostic antigen in human melioidosis [63,131,220] and horse glanders [130]. PBMCs from human melioidosis seropositive donors also respond strongly to AhpC-derived peptides or full-length protein and the strength of this response appears to correlate with improved disease outcomes [131,194,220,221]. Together, these studies suggest that AhpC is strongly immunogenic during natural melioidosis infection, eliciting robust B and T cell responses. Furthermore, AhpC is extremely well-conserved and has the highest level of sequence identity between the Bpc and Bcc of any antigen discussed here (Table 2), making it a very promising T cell antigen for a Bcc cross-reactive vaccine.
Given these promising findings, Schmidt et al. performed a vaccination study using AhpC paired with the CPS-CRM197 glycoconjugate antigen [222]. To account for AhpC’s enzymatic activity and any detrimental effect it might have on immunity or animal health, the authors introduced a cysteine-to-glycine substitution at residue 57 which renders the enzyme nonfunctional. C57BL/6 mice were immunized subcutaneously with the AhpC mutant and CPS-CRM197, along with CpG ODN 2006 and Alhydrogel as adjuvants. This resulted in robust IgG1 and IgG2a/b titers, suggesting a Th1/Th2-balanced response [222]. These antibodies were functionally characterized via opsonophagocytosis assay and were capable of significantly increasing the uptake of B. thaliandensis by macrophages, though this was largely attributed to CPS-specific antibodies [222]. Splenocytes from immunized animals were recalled with full-length AhpCC57G or AhpC-derived peptides, resulting in significant induction of IFN-γ, IL-5, and IL-17 [222]. Finally, in mouse challenge studies, The AhpCC57G and CPS-CRM197-vaccinated animals exhibited 70% protection against an intranasal challenge of B. pseudomallei strain K96243 compared to 0% in the adjuvant group. A follow-up study further confirmed that AhpC elicits particularly robust T cell responses after vaccination, though they were unable to establish if this had any role in protection [132]. On the contrary, mice immunized with Hcp1+AhpC+CPS-CRM197 were less protected than those immunized with Hcp1+CPS-CRM197, albeit not to a statistically significant degree [132]. Finally, it is also worth noting that AhpC vaccination has reportedly been associated with reactogenicity after exposure to B. pseudomallei [223], which could significantly limit the usefulness of the antigen due to safety concerns.
10.4. IscA (BPSL2287), TadE (BPSL1897), and AcoD (BPSL3369)
Because many B. pseudomallei vaccines fail to confer sterilizing immunity in mice resulting in chronic infection, Champion et al., used qRT-PCR to identify upregulated genes in chronically infected mouse tissue [184]. Six such genes were identified, but only IscA, TadE, and AcoD could be recombinantly expressed. These genes are notably predicted by pSORTb to be cytoplasmic or in the inner membrane [184], meaning they are likely functioning primarily as T cell antigens in a subunit vaccine. These antigens were mixed with Omp7 and administered intraperitoneally to C57BL/6 mice as a cocktail paired with LolC, CPS, or alone. These antigens were adjuvanted with SAS and the vaccine was administered i.p. before challenging i.p. with 100x LD50 of B. pseudomallei strain K96243. All formulations containing the chronic antigen cocktail, regardless of CPS or LolC presence, exhibited 30-50% long-term protection compared to 0% in naïve animals [184]. However, because Omp7 was included in all reported formulations, it cannot be said whether the reported protection was due to the inclusion of the chronic disease-associated T cell antigens, the putative B cell antigen Omp7, or a combination of both. Another significant limitation of this study is that organ colonization was not reported for the surviving animals, so it is unclear if these antigens were efficacious in preventing chronic infection in the surviving animals, which was ultimately the goal of this study. If such concerns could be addressed, these proteins could serve as potentially promising T cell antigens in a Bcc cross-protective vaccine given their high degree of sequence conservation in Bcc species, particularly IscA (Table 2).
11. Conclusions and Future Directions
Numerous protective subunit vaccine antigens have been explored for Bpc species, some of which appear to be broadly conserved across members of the Bcc. Further exploration of these antigens as possible pan-Burkholderia vaccine components appear warranted given that such an approach may be the only economically viable way to produce a Bcc vaccine. We have identified some of the most promising candidates as BamA, Omp1, Omp3, Omp7, OmpW1, and OpcP, which are all putative targets of humoral immunity, as well as AhpC and LolC, which are likely T cell-restricted antigens. Of note, many of the antigens described here have only been tested in what are now generally considered outdated vaccination/challenge models, chiefly those that utilized the intraperitoneal or intravenous administration routes. BamA, Omp3, and OmpW1 have all been tested only in such i.p./i.p. models. Future studies should explore whether their protective capabilities extend to respiratory infection models, which more closely recapitulate the human disease. Another common issue is that most research groups do not report confirming the secondary structures of their proteins. Aggregated or misfolded proteins are significantly impaired in their ability to elicit antibodies to conformational epitopes, thus lowering the likelihood of seeing protection when immunizing with the purified protein. This issue was particularly evident in studies that utilized β-barrel protein antigens such as BamA, OpcP, and OmpW1, which is altogether unsurprising given that these proteins have evolved to be stably inserted into the hydrophobic environment of the lipid bilayer. Such studies generally reported solubility issues with their protein, and it is plausible to believe that many or all these studies used protein that is not conformationally intact, limiting their ability to elicit protective antibody responses. Unfortunately, there do not appear to be widely adopted solutions to this problem, as membrane protein stabilizing agents (i.e., detergents, lipid nanodiscs) are not commonly used in vivo and their ability to maintain a protein’s structure in vivo is unclear. Finally, while the general dogma in B. pseudomallei and B. mallei vaccine research holds that T cell immunity is critical to controlling the intracellular phase of infection and preventing the bacteria from establishing itself as a chronic disease, we found that none of the three most well-studied T cell-restricted antigens (Hcp1, LolC, AhpC) have been able to demonstrate this definitively [56,57,58,59,128,209]. On the contrary, the B cell-restricted polysaccharide antigens LPS and CPS consistently appeared to be more important for mediating protection in murine models of respiratory-acquired melioidosis and glanders. Furthermore, adding a T cell-restricted antigen to a formulation containing CPS or LPS has never been shown to confer a statistically significant improvement to survival or organ colonization [56,57,128,209], though one study came close to demonstrating significantly improved survival [128]. This would seem to indicate that a vaccine’s ability to prevent the bacteria from colonizing during the extracellular phase of infection is much more important for conferring protection than attempting to clear the bacteria after it has proceeded to the intracellular phase of infection. Regardless, we hope that this review has provided some useful insight into the rational design of subunit and glycoconjugate Burkholderia vaccines. Designing a vaccine capable of conferring protection to both Bpc and Bcc species has been alluded to in the past but the results of our bioinformatic screen appear to lend additional support to this strategy and hopefully sparks the interest for other researchers to develop a pan-Burkholderia vaccine.
Author Contributions
“Conceptualization, A.J.B. and A.G.T.; methodology, A.J.B.; validation, A.J.B..; formal analysis, A.J.B.; investigation, A.J.B. and A.G.T..; data curation, A.J.B.; writing—original draft preparation, A.J.B.; writing—review and editing, A.J.B. and A.G.T.; visualization, A.J.B.; supervision, A.G.T.; project administration, A.G.T..; funding acquisition, A.G.T. All authors have read and agreed to the published version of the manuscript.”
Funding
A.J.B. was funded by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health, award number T32AI060549. A.G.T. received institutional funds from UTMB Health.
Data Availability Statement
All data is available from published manuscripts. Additional analysis of the data is presented in this review and available upon request.
Acknowledgments
We thank Andy Li for providing scripts to assist in automating the BLAST analysis for use with large datasets.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Eberl, L.; Vandamme, P. Members of the genus Burkholderia: Good and bad guys. F1000Res 2016, 5, F1000 Faculty Rev–1007. [Google Scholar] [CrossRef]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C., Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int J Syst Evol Microbiol 2020, 70, 5607-5612. (accessed 2023 November 11). [CrossRef]
- Center for Disease Control and Prevention. Available from: https://www.selectagents.gov/sat/list.htm (accessed 2023 November 11). 11 November.
- Limmathurotsakul, D.; Golding, N.; Dance, D.A.B.; Messina, J.P.; Pigott, D.M.; Moyes, C.L.; Rolim, D.B.; Bertherat, E.; Day, N.P.J.; Peacock, S.J.; et al. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat Microbiol 2016, 1, 15008. [Google Scholar] [CrossRef] [PubMed]
- Zlosnik, J.E.A.; Costa, P.S.; Brant, R.; Mori, P.Y.B.; Hird, T.J.; Fraenkel, M.C.; Wilcox, P.G.; Davidson, AGF, Speert DP. Mucoid and Nonmucoid Burkholderia cepacia Complex Bacteria in Cystic Fibrosis Infections. Am J Resp Crit Care Med 2011, 183(1). [CrossRef]
- Folescu, T.W.; da Costa, C.H.; Cohen, R.W.F.; Cohen, R.; da Conceição Neto, O.C.; Albano, R.M.; Marques, E.A. Burkholderia cepacia complex: Clinical course in cystic fibrosis patients. BMC Pulm Med 2015, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Häfliger, E.; Atkinson, A.; Marschall, J. Systematic review of healthcare-associated Burkholderia cepacia complex outbreaks: Presentation, causes and outbreak control. Infect Prevent Practice 2020, 2(3), 100082. [Google Scholar] [CrossRef] [PubMed]
- Pongmala, K.; Pierret, A.; Oliva, P.; Pando, A.; Davong, V.; Rattanavong, S.; Silvera, N.; Luangraj, M.; Boithias, L.; Xayyathip, K; et al. Distribution of Burkholderia pseudomallei within a 300-cm deep soil profile: Implications for environmental sampling. Sci Rep 2022, 12, 8674. [Google Scholar] [CrossRef] [PubMed]
- Merritt, A.J.; Inglis, T.J.J. The Role of Climate in the Epidemiology of Melioidosis. Curr Trop Med Resp 2017, 4(4), 185–191. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.Y.I.; Fisher, D. Earth, wind, rain, and melioidosis. Lancet Planet Health 2018, 2(8), E329–E330. [Google Scholar] [CrossRef]
- Birnie, E.; Biemond, J.J.; Wiersinga, W.J. Drivers of melioidosis endemicity: Epidemiological transition, zoonosis, and climate change. Curr Opin Infect Dis 2022, 35(3), 196–204. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention. Available from: https://emergency.cdc.gov/han/2022/han00470.asp (accessed 2023 November 11). 11 November.
- Gassiep, I.; Grey, V.; Thean, L.J.; Farquhar, D.; Clark, J.E.; Ariotti, L.; Graham, R.; Jennison, A.V.; Bergh, H.; Anuradha, S.; et al. Expanding the Geographic Boundaries of Melioidosis in Queensland, Australia. Am J Trop Med Hyg 2023, 108, 1215–1219. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Virk, H.S.; Torres, A.G.; Currie, B.J.; Peacock, S.J.; Dance, D.A.B.; Limmathurotsakul, D. Melioidosis. Nat Rev Dis Primers 2018, 4, 17107. [Google Scholar] [CrossRef]
- Singh, M.; Mahmood, M. Melioidosis: The great mimicker. J Community Hosp Intern Med Perspect 2017, 7(4), 245–247. [Google Scholar] [CrossRef]
- Khakhum, N.; Chapartegui-González, I.; Torres, A.G. Combating the great mimicker: Latest progress in the development of Burkholderia pseudomallei vaccines. Expert Rev Vaccines, 2020, 19(7), 653-660. [CrossRef]
- UpToDate. Available from: https://www.uptodate.com/contents/melioidosis-epidemiology-clinical-manifestations-and-diagnosis (accessed 2023 November 11). 11 November.
- Losada, L.; Ronning, C.M.; DeShazer, D.; Woods, D.; Fedorova, N.; Kim, H.S.; Shabalina, S.A.; Pearson, T.R.; Brinkac, L; Tan, P. ; et al. Continuing Evolution of Burkholderia mallei Through Genome Reduction and Large-Scale Rearrangements. Genome Biol Evol 2010, 2, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Rojas, J.L.G.; Schmidt, C.G.; Herskin, M.; et al. Assessment of the control measures of the category A diseases of Animal Health Law: Burkholderia mallei (Glanders). EFSA J 2022, 20(1), e07069. [Google Scholar] [CrossRef] [PubMed]
- Merck Veterinary Manual. Available from: https://www.merckvetmanual.com/generalized-conditions/glanders/glanders-in-horses-and-other-animals (accessed 2023 November 11). 11 November.
- Van Zandt, K.E.; Greer, M.T.; Gelhaus, H.C. Glanders: An overview of infection in humans. Orphanet J Rare Dis 2013, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Godoy, D.; Randle, G.; Simpson, A.J.; Aanensen, D.M.; Pitt, T.L.; Kinoshita, R.; Spratt, B.G. Multilocus Sequence Typing and Evolutionary Relationships among the Causative Agents of Melioidosis and Glanders, Burkholderia pseudomallei and Burkholderia mallei. J Clin Microbiol 2003, 41(5), 2068–2079. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ruíz L.M.; Rodríguez-Cisneros, M.; Kerber-Díaz, J.C.; Rojas-Rojas, F.U.; Ibarra, J.A.; Los Santos, P.E. Burkholderia orbicola sp. nov., a novel species within the Burkholderia cepacia complex. Arch Microbiol 2022, 204(3), 178. [CrossRef]
- Center for Disease Control and Prevention. Available from: https://www.cdc.gov/hai/organisms/bcepacia.html (accessed 2023 November 11). 11 November.
- Tavares, M.; Kozak, M.; Balola, A.; Sá-Correia, I. Burkholderia cepacia Complex Bacteria: A Feared Contamination Risk in Water-Based Pharmaceutical Products. Clin Microbiol Rev 2020, 33(3), e00139–19. [Google Scholar] [CrossRef] [PubMed]
- Daccò, V.; Alicandro, G.; Consales, A.; Rosazza, C.; Sciarrabba, C.S.; Cariani, L.; Colombo, C. Cepacia syndrome in cystic fibrosis: A systematic review of the literature and possible new perspectives in treatment. Pediatr Pulmonol 2023, 58(5), 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat Rev Microbiol 2005, 3, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Horsley, A.; Webb, K.; Bright-Thomas, R.; Govan, J.; Jones, A. Can early Burkholderia cepacia complex infection in cystic fibrosis be eradicated with antibiotic therapy? Front Cell Infect Microbiol 2011, 1, 18. [Google Scholar] [CrossRef]
- Lord, R.; Jones, A.M.; Horsley, A. Antibiotic treatment for Burkholderia cepacia complex in people with cystic fibrosis experiencing a pulmonary exacerbation. Cochrane Database Sys Rev 2020, 2020(4), CD009529. [Google Scholar] [CrossRef]
- LiPuma, J.J. The Changing Microbial Epidemiology in Cystic Fibrosis. Clin Microbiol Rev 2010, 23(2), 299–323. [Google Scholar] [CrossRef] [PubMed]
- Zlosnik, J.E.A.; Henry, D.A.; Hird, T.J.; Hickman, R.; Campbell, M.; Cabrera, A.; Chiavegatti, G.L.; Chilvers, M.A.; Sadarangani, M. Epidemiology of Burkholderia Infections in People with Cystic Fibrosis in Canada between 2000 and 2017. Ann Am Thorac Soc 2020, 17(12). [CrossRef]
- Pradenas, G.A.; Ross, B.N.; Torres, A.G. Burkholderia cepacia Complex Vaccines: Where Do We Go from here? Vaccines (Basel). 2016, 4(2), 10. [Google Scholar] [CrossRef]
- Grund, M.E.; Soo, J.C.; Cote, C.K.; Berisio, R.; Lukomski, S. Thinking Outside the Bug: Targeting Outer Membrane Proteins for Burkholderia Vaccines. Cells 2021, 10(3), 495. [Google Scholar] [CrossRef]
- Irudal, S.; Scoffone, V.C.; Trespidi, G.; Barbieri, G.; D’Amato, M.; Viglio, S.; Pizza, M.; Scarselli, M.; Riccardi, G.; Buroni, S. Identification by Reverse Vaccinology of Three Virulence Factors in Burkholderia cenocepacia That May Represent Ideal Vaccine Antigens. Vaccines (Basel) 2023, 11(6), 1039. [Google Scholar] [CrossRef]
- Cocorullo, M.; Chiarelli, L.R.; Stelitano, G. Improving Protection to Prevent Bacterial Infections: Preliminary Applications of Reverse Vaccinology against the Main Cystic Fibrosis Pathogens. Vaccines (Basel) 2023, 11(7), 1221. [Google Scholar] [CrossRef] [PubMed]
- Luangasanatip, N.; Flasche, S.; Dance, D.A.B.; Limmathurotsakul, D.; Currie, B.J.; Mukhopadhyay, C.; Atkins, T.; Titball, R.; Jit, M. The global impact and cost-effectiveness of a melioidosis vaccine. BMC Med 2019, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J Mol Biol 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform 2008, 10, 421. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2021 available from: https://www.R-project.org.
- Scholz, H.C.; Joseph, M.; Tomaso, H.; Dahouk, S.A.; Witte, A.; Kinne, J.; Hagen, R.M.; Wernery, R.; Wernery, U.; Neubauer, H. Detection of the reemerging agent Burkholderia mallei in a recent outbreak of glanders in the United Arab Emirates by a newly developed fliP-based polymerase chain reaction assay. Diag Microbiol Infect Dis 2006, 54(4), 241–247. [Google Scholar] [CrossRef]
- Chua, K.L.; Chan, Y.Y.; Gan, Y.H. Flagella Are Virulence Determinants of Burkholderia pseudomallei. Infect Immun 2003, 71(4), 1622–1629. [Google Scholar] [CrossRef]
- Chuaygud, T.; Tungpradabkul, S.; Sirisinha, S.; Chua, K.L.; Utaisincharoen, P. A role of Burkholderia pseudomallei flagella as a virulent factor. Trans R Soc Trop Med Hyg 2008, 102, Suppl 1:S140–S144. [Google Scholar] [CrossRef]
- Tomich, M.; Herfst, C.A.; Golden, J.W.; et al. Role of Flagella in Host Cell Invasion by Burkholderia cepacia. Infect Immun 2002, 70(4), 1799–1806. [Google Scholar] [CrossRef]
- Urban, T.A.; Griffith, A.; Torok, A.M.; Smolkin, M.E.; Burns, J.L.; Goldberg, J.B. Contribution of Burkholderia cenocepacia Flagella to Infectivity and Inflammation. Infect Immun 2004, 72(9). [CrossRef]
- Ceballos-Olvera, I.; Sahoo, M.; Miller, M.A.; del Barria, L.; Re, F. Inflammasome-dependent Pyroptosis and IL-18 Protect against Burkholderia pseudomallei Lung Infection while IL-1β Is Deleterious. PLoS Pathog 2011, 7(12), e1002452. [Google Scholar] [CrossRef]
- West, T.E.; Myers, N.D.; Chantratita, N.; Chierakul, W.; Limmathurotsakul, D.; Wuthiekanun, V.; Miao, E.A.; Hajjar, A.M.; Peacock, S.J.; Liggitt, H.D.; et al. NLRC4 and TLR5 Each Contribute to Host Defense in Respiratory Melioidosis. PLoS Negl Trop Dis 2014, 8(9), e3178. [Google Scholar] [CrossRef] [PubMed]
- Koosakulnirand, S.; Phokrai, P.; Jenjaroen, K.; Roberts, R.A.; Utaisincharoen, P.; Dunachie, S.J.; Brett, P.J.; Burtnick, M.N.; Chantratita, N. Immune response to recombinant Burkholderia pseudomallei FliC. PloS One 2018, 13(6), e0198906. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, K.; Dankmeyer, J.L.; Bernhards, R.C.; Fetterer, D.P.; Waag, D.M.; Worsham, P.L.; DeShazer, D. Activation of Toll-Like Receptors by Live Gram-Negative Bacterial Pathogens Reveals Mitigation of TLR4 Responses and Activation of TLR5 by Flagella. Front Cell Infect Microbiol 2021, 11, 745325. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9(7), 279. [Google Scholar] [CrossRef]
- Brett, P.J.; Mah, D.C.; Woods, D.E. Isolation and characterization of Pseudomonas pseudomallei flagellin proteins. Infect Immun 1994, 62(5), 1914–1919. [Google Scholar] [CrossRef]
- Brett, P.J.; Woods, D.E. Structural and Immunological Characterization of Burkholderia pseudomallei O-Polysaccharide–Flagellin Protein Conjugates. Infect Immun 1996, 64(7), 2824–2828. [Google Scholar] [CrossRef]
- Chen, Y.S.; Hsiao, Y.S.; Lin, H.H.; Yen, C.M.; Chen, S.C.; Chen, Y.L. Immunogenicity and anti-Burkholderia pseudomallei activity in Balb/c mice immunized with plasmid DNA encoding flagellin. Vaccine 2006, 24(6), 750–758. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Hsiao, Y.S.; Lin, H.H.; Liu, Y.; Chen, Y.L. CpG-Modified Plasmid DNA Encoding Flagellin Improves Immunogenicity and Provides Protection against Burkholderia pseudomallei Infection in BALB/c Mice. Infect Immun 2006, 74(3), 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Gregory, A.E.; Hatcher, C.L.; Vinet-Oliphant, H.; Morici, L.A.; Titball, R.W.; Roy, C.J. Protection of non-human primates against glanders with a gold nanoparticle glycoconjugate vaccine. Vaccine 2015, 33(5), 686–692. [Google Scholar] [CrossRef]
- Gregory, A.E.; Judy, B.M.; Qazi, O.; Blumentritt, C.A.; Brown, K.A.; Shaw, A.M.; Torres, A.G.; Titball, R.W. A gold nanoparticle-linked glycoconjugate vaccine against Burkholderia mallei. Nanomedicine: NBM 2015, 11(2), 447-456. [CrossRef]
- Muruato, L.A.; Tapia, D.; Hatcher, C.L.; Kalita, M.; Brett, P.J.; Gregory, A.E.; Samuel, J.E.; Titball, R.W.; Torres, A.G. Use of Reverse Vaccinology in the Design and Construction of Nanoglycoconjugate Vaccines against Burkholderia pseudomallei. Clin Vaccine Immunol 2017, 24(11), e00206–17. [Google Scholar] [CrossRef]
- Tapia, D.; Sanchez-Villamil, J.I.; Torres, A.G. Multicomponent gold nano-glycoconjugate as a highly immunogenic and protective platform against Burkholderia mallei. npj Vaccines 2020, 5, 82. [Google Scholar] [CrossRef]
- Tapia, D.; Sanchez-Villamil, J.I.; Stevenson, H.L.; Torres, A.G. Multicomponent Gold-Linked Glycoconjugate Vaccine Elicits Antigen-Specific Humoral and Mixed TH1-TH17 Immunity, Correlated with Increased Protection against Burkholderia pseudomallei. mBio 2021, 12(3), e01227–21. [Google Scholar] [CrossRef]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin—a potent immunomodulatory agent. Exp Mol Med 2017, 49(9), e373. [Google Scholar] [CrossRef]
- Charuchaimontri, C.; Suputtamongol, Y.; Nilakul, C.; Chaowagul, W.; Chetchotisakd, P.; Lertatanasuwun, N.; Intaranongpai, S.; Brett, P.J.; Woods, D.E. Antilipopolysaccharide II: An antibody protective against fatal melioidosis. Clin Infect Dis 1999, 29(4), 813–818. [Google Scholar] [CrossRef]
- Chen, Y.S.; Shiuan, D.; Chen, S.C.; Chye, S.M.; Chen, Y.L. Recombinant Truncated Flagellin of Burkholderia pseudomallei as a Molecular Probe for Diagnosis of Melioidosis. Clin Diagn Lab Immunol 2003, 10(3), 423–425. [Google Scholar] [CrossRef] [PubMed]
- Felgner, P.L.; Kayala, M.A.; Vigil, A.; Burk, C.; Nakajima-Sasaki, R.; Pablo, J.; Molina, D.M.; Hirst, S.; Chew, J.S.W.; Wang, D.; et al. A Burkholderia pseudomallei protein microarray reveals serodiagnostic and cross-reactive antigens. PNAS 2009, 106(32), 13499–13504. [Google Scholar] [CrossRef]
- Suwannasaen, D.; Mahawantung, J.; Chaowagul, W.; Limmathurotsakul, D.; Felgner, P.L.; Davies, H.; Bancroft, G.J.; Titball, R.W.; Lertmemongkolchai, G. Human Immune Responses to Burkholderia pseudomallei Characterized by Protein Microarray Analysis. J Infect Dis. 2011, 203(7), 1002–1011. [Google Scholar] [CrossRef]
- Kohler, C.; Dunachie, S.J.; Müller, E.; Kohler, A.; Jenjaroen, K.; Teparrukkul, P.; Baier, V.; Ehricht, R.; Steinmetz, I. Rapid and Sensitive Multiplex Detection of Burkholderia pseudomallei-Specific Antibodies in Melioidosis Patients Based on a Protein Microarray Approach. PLoS Negl Trop Dis 2016, 10(7), e0004847. [Google Scholar] [CrossRef]
- Scott, A.E.; Twine, S.M.; Fulton, K.M.; Titball, R.W.; Essex-Lopresti, A.E.; Atkins, T.P.; Prior, J.L. Flagellar Glycosylation in Burkholderia pseudomallei and Burkholderia thailandensis. J Bacteriol 2011, 193(14), 3577–3587. [Google Scholar] [CrossRef]
- Hanuszkiewicz, A.; Pittock, P.; Humphries, F.; Moll, H.; Rosales, A.R.; Molinaro, A.; Moynagh, P.N.; Lajoie, G.A.; Valvano, M.A. Identification of the Flagellin Glycosylation System in Burkholderia cenocepacia and the Contribution of Glycosylated Flagellin to Evasion of Human Innate Immune Responses. J Biol Chem 2014, 289(27), 19231–19244. [Google Scholar] [CrossRef]
- DeShazer, D.; Brett, P.J.; Carlyon, R.; Woods, D.E. Mutagenesis of Burkholderia pseudomallei with Tn5-OT182: Isolation of Motility Mutants and Molecular Characterization of the Flagellin Structural Gene. J Bacteriol 1997, 179(7), 2116–2125. [Google Scholar] [CrossRef] [PubMed]
- Wikraiphat, C.; Charoensap, J.; Utaisincharoen, P.; Wongratanacheewin, S.; Taweechaisupapong, S.; Woods, D.E.; Bolscher, J.G.M.; Sirisinha, S. Comparative in vivo and in vitro analyses of putative virulence factors of Burkholderia pseudomallei using lipopolysaccharide, capsule and flagellin mutants. FEMS Immunol Med Microbiol 2009, 56(3), 253–259. [Google Scholar] [CrossRef]
- Whitlock, G.C.; Deeraksa, A.; Qazi, O.; Judy, B.M.; Taylor, K.; Propst, K.L; Duffy, A.J.; Johnson, K.; Kitto, G.B.; Brown, K.A. Protective response to subunit vaccination against intranasal Burkholderia mallei and B. pseudomallei challenge. Procedia Vaccinol 2010, 2(1). [CrossRef]
- Hales, B.A.; Morgan, J.A.W.; Hart, C.A.; Winstanley, C. Variation in Flagellin Genes and Proteins of Burkholderia cepacia. J Bacteriol 1998, 180(5), 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Musson, J.A.; Reynolds, C.J.; Rinchai, D.; Nithichanon, A.; Khaenam, P.; Favry, E.; Spink, N.; Chu, K.K.Y.; de Soyza, A.; Bancroft, G.J.; et al. CD4+ T Cell Epitopes of FliC Conserved between Strains of Burkholderia: Implications for Vaccines against Melioidosis and Cepacia Complex in Cystic Fibrosis. J Immunol 2014, 193(12), 6041–6049. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat Rev Microbiol 2017, 15, 323–337. [Google Scholar] [CrossRef]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial type III secretion systems: A complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol Lett 2018, 365(19), fny20. [Google Scholar] [CrossRef]
- Goodin, J.L.; Raab, R.W.; McKown, R.L.; Coffman, G.L.; Powell, B.S.; Enama, J.T.; Ligon, J.A.; Andrews, G.P. Yersinia pestis outer membrane type III secretion protein YscC: Expression, purification, characterization, and induction of specific antiserum. Protein Expr Purif 2005, 40(1), 152–156. [Google Scholar] [CrossRef] [PubMed]
- Fasciano, A.C.; Shaban, L.; Mecsas, J. Promises and Challenges of the Type Three Secretion System-Injectisome as an Anti-Virulence Target. EcoSal Plus 2019, 8(2). [CrossRef]
- Hotinger, J.A.; May, A.E. Antibodies Inhibiting the Type III Secretion System of Gram-Negative Pathogenic Bacteria. Antibodies (Basel) 2020, 9(3). [CrossRef]
- Hotinger, J.A.; Pendergrass, H.A.; May, A.E. Molecular Targets and Strategies for Inhibition of the Bacterial Type III Secretion System (T3SS); Inhibitors Directly Binding to T3SS Components. Biomolecules 2021, 11(2), 316. [Google Scholar] [CrossRef]
- Wallner, A.; Moulin, L.; Busset, N.; Rimbault, I.; Béna, G. Genetic Diversity of Type 3 Secretion System in Burkholderia s.l. and Links With Plant Host Adaptation. Front Microbiol 2021, 12, 761215. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, L.; Hart, C.A.; Winstanley, C. Distribution of type III secretion gene clusters in Burkholderia pseudomallei, B. thailandensis and B. mallei. J Med Microbiol 2002, 51(5), 374–384. [Google Scholar] [CrossRef]
- Vander Broek, C.W.; Stevens, J.M. Type III Secretion in the Melioidosis Pathogen Burkholderia pseudomallei. Front Cell Infect Microbiol 2017, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.P.; Wood, M.W.; Taylor, L.A.; Monaghan, P.; Hawes, P.; Jones, P.W.; Wallis, T.S.; Galyov, E.E. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Molec Microbiol 2002, 46(3), 649–659. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Pfeffer, T.L.; Warawa, J.M. Type 3 Secretion System Cluster 3 Is a Critical Virulence Determinant for Lung-Specific Melioidosis. PLoS Negl Trop Dis 2015, 9(1), e3441. [Google Scholar] [CrossRef]
- Kurtz, J.R.; Petersen, H.E.; Frederick, D.R.; Morici, L.A.; McLachlan, J.B. Vaccination with a Single CD4 T Cell Peptide Epitope from a Salmonella Type III-Secreted Effector Protein Provides Protection against Lethal Infection. Infect Immun 2014, 82(6), 2424–2433. [Google Scholar] [CrossRef]
- Lee, S.J.; Benoun, J.; Sheridan, B.S.; Fogassy, Z.; Pham, O.; Pham, Q.M.; Puddington, L.; McSorley, S.J. Dual immunization with SseB/flagellin provides enhanced protection against Salmonella infection mediated by circulating memory cells. J Immunol 2017, 199(4), 1353–1361. [Google Scholar] [CrossRef]
- Xiong, X.; Jiao, J.; Gregory, A.E.; Wang, P.; Bi, Y.; Wang, X.; Jiang, Y.; Wen, B.; Portnoy, D.A.; Samuel, J.E.; et al. Identification of Coxiella burnetii CD8+ T-Cell Epitopes and Delivery by Attenuated Listeria monocytogenes as a Vaccine Vector in a C57BL/6 Mouse Model. J Infect Dis 2017, 215(10), 1580–1589. [Google Scholar] [CrossRef]
- Harley, V.S.; Dance, D.A.; Tovey, G.; McCrossan, M.V.; Drasar, B.S. An ultrastructural study of the phagocytosis of Burkholderia pseudomallei. Microbios 1998, 94(377), 35–45. [Google Scholar]
- Steele-Mortimer, O. The Salmonella-containing Vacuole – Moving with the Times. Curr Opin Microbiol 2008, 11(1), 38–45. [Google Scholar] [CrossRef]
- Allwood, E.M.; Devenish, R.J.; Prescott, M.; Adler, B.; Boyce, J.D. Strategies for Intracellular Survival of Burkholderia pseudomallei. Front Microbiol 2011, 2, 170. [Google Scholar] [CrossRef]
- Latomanski, E.A.; Newton, H.J. Interaction between autophagic vesicles and the Coxiella-containing vacuole requires CLTC (clathrin heavy chain). Autophagy 2018, 14(10), 1710–1725. [Google Scholar] [CrossRef]
- Stevens, M.P.; Haque, A.; Atkins, T.; Hill, J.; Wood, M.W.; Easton, A.; Nelson, M.; Underwood-Fowler, C.; Titball, R.W.; Bancroft, G.J.; et al. Attenuated virulence and protective efficacy of a Burkholderia pseudomallei bsa type III secretion mutant in murine models of melioidosis. Microbiol 2004, 150(8), 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- Druar, C.; Yu, F.; Barnes, J.L.; Okinaka, R.T.; Chantratita, N.; Beg, S.; Stratilo, C.W.; Olive, A.J.; Soltes, G.; Russell, M.L.; et al. Evaluating Burkholderia pseudomallei Bip proteins as vaccines and Bip antibodies as detection agents. FEMS Microbiol Immunol 2007, 52(1), 78–87. [Google Scholar] [CrossRef] [PubMed]
- Pumirat, P.; Cuccui, J.; Stabler, R.A.; Stevens, J.M.; Muangsombut, V.; Singsuksawat, E.; Stevens, M.P.; Wren, B.W.; Korbsrisate, S. Global transcriptional profiling of Burkholderia pseudomallei under salt stress reveals differential effects on the Bsa type III secretion system. BMC Microbiol 2010, 10, 171. [Google Scholar] [CrossRef] [PubMed]
- Ooi, W.F.; Ong, C.; Nandi, T.; Kreisberg, J.F.; Chua, H.H.; Sun, G.; Chen, Y.; Mueller, C.; Conejero, L.; Eshaghi, M.; et al. The Condition-Dependent Transcriptional Landscape of Burkholderia pseudomallei. PLoS Genet 2013, 9(9), e1003795. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Wong, R.R.; Ghazali, A.K.; Hara, Y.; Aziz, T.N.T.; Nathan, S. Transcriptional landscape of Burkholderia pseudomallei cultured under environmental and clinical conditions. Microb Genom 2023, 9(4). [CrossRef]
- Lichtenegger, S.; Stiehler, J.; Saiger, S.; Zauner, A.; Kleinhappl, B.; Bernecker, C.; Schlenke, P.; Wagner, G.E.; Krause, K.; Gastager, M.; et al. Burkholderia pseudomallei triggers canonical inflammasome activation in a human primary macrophage-based infection model. PLoS Negl Trop Dis 2020, 14(11), e0008840. [Google Scholar] [CrossRef]
- Leyton, D.L.; Rossiter, A.E.; Henderson, I.R. From self sufficiency to dependence: Mechanisms and factors important for autotransporter biogenesis. Nat Rev Microbiol 2012, 10, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Hor, L; Pilapitiya, A. ; Luirink, J.; Paxman, J.J.; Heras, B. Phylogenetic Classification and Functional Review of Autotransporters. Front Immunol 2022, 13, 921272. [Google Scholar] [CrossRef] [PubMed]
- Adler, N.R.L.; Stevens, J.M.; Stevens, M.P.; Galyov, E.E. Autotransporters and Their Role in the Virulence of Burkholderia pseudomallei and Burkholderia mallei. Front Microbiol 2011, 2, 151. [Google Scholar] [CrossRef]
- Stevens, M.P.; Stevens, J.M.; Jeng, R.L.; Taylor, L.A.; Wood, M.W.; Hawes, P.; Monaghan, P.; Welch, M.D.; Galyov, E.E. Identification of a bacterial factor required for actin-based motility of Burkholderia pseudomallei. Mol Microbiol 2005, 56(1), 40–53. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.G.; Borst, L.; Cotter, P.A. Characterization of BcaA, a putative classical autotransporter protein in Burkholderia pseudomallei. Infect Immun 2013, 81(4), 1121–1128. [Google Scholar] [CrossRef]
- Zimmerman, S.M.; Dyke, J.S.; Jelesijevic, T.P.; Michel, F.; Lafontaine, E.R.; Hogan, R.J. Antibodies against In Vivo-Expressed Antigens Are Sufficient To Protect against Lethal Aerosol Infection with Burkholderia mallei and Burkholderia pseudomallei. Infect Immun 2017, 85(8), e00102–17. [Google Scholar] [CrossRef]
- Mil-Homens, D.; Fialho, A.M. Trimeric autotransporter adhesins in members of the Burkholderia cepacia complex: A multifunctional family of proteins implicated in virulence. Front Cell Infect Microbiol 2011, 1. [Google Scholar] [CrossRef]
- Lafontaine, E.R.; Chen, Z.; Huertas-Diaz, M.C.; Dyke, J.S.; Jelesijevic, T.P.; Michel, F.; Hogan, R.J.; He, B. The autotransporter protein BatA is a protective antigen against lethal aerosol infection with Burkholderia mallei and Burkholderia pseudomallei. Vaccine X 2019, 1, 100002. [Google Scholar] [CrossRef]
- Sitthidet, C.; Korbsrisate, S.; Layton, A.N.; Field, T.R.; Stevens, M.P.; Stevens, J.M. Identification of Motifs of Burkholderia pseudomallei BimA Required for Intracellular Motility, Actin Binding, and Actin Polymerization. J Bacteriol 2011, 193(8), 1901–1910. [Google Scholar] [CrossRef]
- Kespichayawattana, W.; Rattanachetkul, S.; Wanun, T.; Utaisincharoen, P.; Sirisinha, S. Burkholderia pseudomallei Induces Cell Fusion and Actin-Associated Membrane Protrusion: A Possible Mechanism for Cell-to-Cell Spreading. Infect Immun 2000, 68(9), 5377–5384. [Google Scholar] [CrossRef]
- Sitthidet, C.; Stevens, J.M.; Chantratita, N.; Currie, B.J.; Peacock, S.J.; Korbsrisate, S.; Stevens, M.P. Prevalence and Sequence Diversity of a Factor Required for Actin-Based Motility in Natural Populations of Burkholderia Species. J Clin Microbiol 2008, 46(7), 2418–2422. [Google Scholar] [CrossRef]
- Mukhopadhyay, C.; Kaestli, M.; Vandana, K.E.; Sushma, K.S.; Mayo, M.; Richardson, L.; Tuanyok, A.; Keim, P.; Godoy, D.; Spratt, B.G.; et al. Molecular Characterization of Clinical Burkholderia pseudomallei Isolates from India. Am J Trop Med Hyg 2011, 85(1), 121–123. [Google Scholar] [CrossRef] [PubMed]
- Corea, E.M.; de Silva, A.D.; Thevanesam, V. Melioidosis in Sri Lanka. Trop Med Infect Dis. 2018, 3(1), 22. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.L.; Fane, A.; Sarovich, D.S.; Price, E.P.; Rush, C.M.; Govan, B.L.; Parker, E.; Mayo, M.; Currie, B.J.; Ketheesan, N. Increased Neurotropic Threat from Burkholderia pseudomallei Strains with a B. mallei–like Variation in the bimA Motility Gene, Australia. Emerg Infect Dis 2017, 23(5), 740-749. [CrossRef]
- Sarovich, D.S.; Price, E.P.; Webb, J.R.; Ward, L.M.; Voutsinos, M.Y.; Tuanyok, A.; Mayo, M.; Kaestli, M.; Currie, B.J. Variable Virulence Factors in Burkholderia pseudomallei (Melioidosis) Associated with Human Disease. PLoS One 2014, 9(3), e91682. [Google Scholar] [CrossRef] [PubMed]
- Gora, H.; Hasan, T.; Smith, S.; Wilson, I.; Mayo, M.; Woerle, C.; Webb, J.R.; Currie, B.J.; Hanson, J.; Meumann, E.M. Melioidosis of the Central Nervous System: Impact of the bimABm Allele on Patient Presentation and Outcome. Clin Infect Dis 2022, ciac111. [Google Scholar] [CrossRef]
- Campos, C.G.; Byrd, M.S.; Cotter, P.A. Functional Characterization of Burkholderia pseudomallei Trimeric Autotransporters. Infect Immun 2013, 81(8), 2788–2799. [Google Scholar] [CrossRef]
- Adler, N.R.L.; Stevens, M.P.; Dean, R.E.; Saint, R.J.; Pankhania, D.; Prior, J.L.; Atkins, T.P.; Kessler, B.; Nithichanon, A.; Lertmemongkolchai, G.; et al. Systematic Mutagenesis of Genes Encoding Predicted Autotransported Proteins of Burkholderia pseudomallei Identifies Factors Mediating Virulence in Mice, Net Intracellular Replication and a Novel Protein Conferring Serum Resistance. PLoS One 2015, 10(4), e0121271. [Google Scholar] [CrossRef]
- Coulthurst, S. The Type VI secretion system: A versatile bacterial weapon. Microbiol (Reading) 2019, 165(5). [CrossRef]
- Singh, R.P.; Kumari, K. Bacterial type VI secretion system (T6SS): An evolved molecular weapon with diverse functionality. Biotech Letters 2023, 45, 309–331. [Google Scholar] [CrossRef]
- DeShazer, D. A novel contact-independent T6SS that maintains redox homeostasis via Zn2+ and Mn2+ acquisition is conserved in the Burkholderia pseudomallei complex. Microbiol Res 2019, 226, 48–54. [Google Scholar] [CrossRef]
- Toesca, I.J.; French, C.T.; Miller, J.F. The Type VI Secretion System Spike Protein VgrG5 Mediates Membrane Fusion during Intercellular Spread by Pseudomallei Group Burkholderia Species. Infect Immun 2014, 82(4), 1436–1444. [Google Scholar] [CrossRef]
- Schwarz, S.; Singh, P.; Robertson, J.D.; LeRoux, M.; Skerrett, S.J.; Goodlett, D.R.; West, T.E.; Mougous, J.D. VgrG-5 Is a Burkholderia Type VI Secretion System-Exported Protein Required for Multinucleated Giant Cell Formation and Virulence. Infect Immun 2014, 82(4), 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Kostow, N.; Welch, M.D. Plasma membrane protrusions mediate host cell–cell fusion induced by Burkholderia thailandensis. Mol Biol Cell 2022, 33(8), ar70. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A.; Ulrich, R.L.; Ribot, W.J.; Brueggemann, E.E.; Hines, H.B.; Chen, D.; Lipscomb, L.; Kim, H.S.; Mrázek, J.; Nierman, W.C.; et al. Type VI secretion is a major virulence determinant in Burkholderia mallei. Mol Microbiol 2007, 64(6), 1466–1485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Ye, F.; Wang, Y.; Liu, R.; Huang, Z.; Chen, C.; Liu, L.; Kang, X.; Dong, S.; Rajaofera, M.J.N.; et al. Role of type VI secretion system protein TssJ-3 in virulence and intracellular survival of Burkholderia pseudomallei. Biochem Biophys Res Common 2023, 682(19), 397–406. [Google Scholar] [CrossRef] [PubMed]
- Spiewak, H.L.; Shastri, S.; Zhang, L.; Schwager, S.; Eberl, L.; Vergunst, A.C.; Thomas, M.S. Burkholderia cenocepacia utilizes a type VI secretion system for bacterial competition. Microbiologyopen 2019, 8(7), e00774. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; West, T.E.; Boyer, F.; Chiang, W.C.; Carl, M.A.; Hood, R.D.; Rohmer, L.; Tolker-Nielsen, T.; Skerrett, S.J.; Mougous, J.D. Burkholderia Type VI Secretion Systems Have Distinct Roles in Eukaryotic and Bacterial Cell Interactions. PLoS Pathog 2010, 6(8), e1001068. [Google Scholar] [CrossRef]
- Burtnick, M.N.; Brett, P.J.; Harding, S.V.; Ngugi, S.A.; Ribot, W.J.; Chantratita, N.; Scorpio, A.; Milne, T.S.; Dean, R.E.; Fritz, D.L.; et al. The Cluster 1 Type VI Secretion System Is a Major Virulence Determinant in Burkholderia pseudomallei. Infect Immun 2011, 79(4), 1512–1525. [Google Scholar] [CrossRef]
- Shalom, G.; Shaw, J.G.; Thomas, M.S. In vivo expression technology identifies a type VI secretion system locus in Burkholderia pseudomallei that is induced upon invasion of macrophages. Microbiol (Reading) 2007, 153, 2689–2699. [Google Scholar] [CrossRef]
- Burtnick, M.N.; DeShazer, D.; Nair, V.; Gherardini, F.C.; Brett, P.J. Burkholderia mallei Cluster 1 Type VI Secretion Mutants Exhibit Growth and Actin Polymerization Defects in RAW 264.7 Murine Macrophages. Infect Immun 2010, 78(1), 88–99. [Google Scholar] [CrossRef]
- Burtnick, M.N.; Shaffer, T.L.; Ross, B.N.; Muruato, L.A.; Sbrana, E.; DeShazer, D.; Torres, A.G.; Brett, P.J. Development of Subunit Vaccines That Provide High-Level Protection and Sterilizing Immunity against Acute Inhalational Melioidosis. Infect Immun 2018, 86(1), e00724–17. [Google Scholar] [CrossRef]
- Tran, Q.T.L.; Nguyen, H.V.; Pham, H.T.; Mai, T.V.; Nguyen, Q.H.M.; Le, D.V.; Bui, L.N.H.; Hoang, L.T.H.; Hoang, T.Q.; Trinh, T.T. Clinical Utility of Combined Whole-cell Antigen and Recombinant Hemolysis Co-regulated Protein 1-Enzyme-linked Immunosorbent Assays Reveals Underdiagnosed Cases of Melioidosis in Vietnam. Am J Trop Med Hyg 2022, 107(3), 585–591. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.E.; Berner, A.; Lipp, M.; Kohler, C.; Assig, K.; Lichtenegger, S.; Saqib, M.; Müller, E.; Trinh, T.T.; Gad, A.M. et al. Protein Microarray-Guided Development of a Highly Sensitive and Specific Dipstick Assay for Glanders Serodiagnostics. J Clin Microbiol 2023, 61(1), e01234-22. [CrossRef]
- Sengyee, S.; Yarasai, A.; Janon, R.; Morakot, C.; Ottiwet, O.; Schmidt, L.K.; West, T.E.; Burtnick, M.N.; Chantratita, C.; Brett, P.J. Melioidosis Patient Survival Correlates With Strong IFN-γ Secreting T Cell Responses Against Hcp1 and TssM. Front Immunol 2021, 12, 698303. [Google Scholar] [CrossRef]
- Biryukov, S.S.; Cote, C.K.; Klimko, C.P.; Dankmeyer, J.L.; Rill, N.O.; Shoe, J.L.; Hunter, M.; Shamsuddin, Z.; Velez, I.; Hedrick, Z.M.; et al. Evaluation of two different vaccine platforms for immunization against melioidosis and glanders. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Li, G.; Li, J.; Zheng, M.; Peng, X.; Rao, Y.; Li, M.; Zhou, R.; Rao, X. Hcp1-loaded staphylococcal membrane vesicle vaccine protects against acute melioidosis. Front Immunol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Brunet, Y.R.; Hénin, J.; Celia, H.; Cascales, E. Type VI secretion and bacteriophage tail tubes share a common assembly pathway. EMBO Rep 2014, 15(3), 315–321. [Google Scholar] [CrossRef]
- Backmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat Rev Immunol 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Kumar, S.; Anselmo, A.C.; Banerjee, A.; Zakrewsky, M.; Mitragotri. Shape and size-dependent immune response to antigen-carrying nanoparticles. J Control Release 2015, 220(Pt A), 141-148. [CrossRef]
- Baranov, M.V.; Kumar, M.; Sacanna, S.; Thutupalli, S.; van den Bogaart, G. Modulation of Immune Responses by Particle Size and Shape. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Shanks, J.; Burtnick, M.N.; Brett, P.J.; Waag, D.M.; Spurgers, K.N.; Ribot, W.J.; Schell, M.A.; Panchal, R.G.; Gherardini, F.C.; Wilkinson, K.D. , et al. Burkholderia mallei tssM Encodes a Putative Deubiquitinase That Is Secreted and Expressed inside Infected RAW 264.7 Murine Macrophages. Infect Immun 2009, 77(4), 1636–1648. [Google Scholar] [CrossRef]
- Tan, K.S.; Chen, Y.; Lim, T.C.; Tan, G.Y.G.; Liu, Y.; Lim, Y.T.; Macary, P.; Gan, Y.H. Suppression of Host Innate Immune Response by Burkholderia pseudomallei through the Virulence Factor TssM. J Immunol 2010, 184(9), 5160–5171. [Google Scholar] [CrossRef]
- Burtnick, M.N.; Brett, P.J.; DeShazer, D. Proteomic Analysis of the Burkholderia pseudomallei Type II Secretome Reveals Hydrolytic Enzymes, Novel Proteins, and the Deubiquitinase TssM. Infect Immun 2014, 82(8). [CrossRef]
- Jacobsen, T.; Bardiaux, B.; Francetic, O.; Izadi-Pruneyre, N.; Nilges, M. Structure and function of minor pilins of type IV pili. Med Microbiol Immunol 2020, 209(3), 301–308. [Google Scholar] [CrossRef]
- Essex-Lopresti, A.E.; Boddey, J.A.; Thomas, R.; Smith, M.P.; Hartley, M.G.; Atkins, T.; Brown, N.F.; Tsang, C.H.; Peak, I.R.A.; Hill, J; et al. A Type IV Pilin, PilA, Contributes to Adherence of Burkholderia pseudomallei and Virulence In Vivo. Infect Immun 2005, 73(2), 1260-1264. [CrossRef]
- Sangdee, K.; Waropastrakul, S.; Wongratanachewin, S.; Homchampa, P. Heterologously type IV pilus expressed proteins of Burkholderia pseudomallei is immunogenic but fails to elicit protective immunity in mice. Southeast Asian J Trop Med Public Health 2011, 42(5), 1190–1196. [Google Scholar]
- Wu, R.; Stephenson, R.; Gichaba, A.; Noinaj, N. The Big BAM Theory: An Open and Closed Case? Biochim Biophys Acta Biomembr 2020, 1862(1), 183062. [Google Scholar] [CrossRef]
- Rayes, J.E.; Rodríguez-Alonso, R.; Collet, J.F. Lipoproteins in Gram-negative bacteria: New insights into their biogenesis, subcellular targeting and functional roles. Curr Opin Microbiol 2021, 61, 25–34. [Google Scholar] [CrossRef]
- Chatorvedi, D.; Mahalakshmi, R. Transmembrane β-barrels: Evolution, folding and energetics. Biochim Biophys Acta Biomembr 2017, 1859(12), 2467–2482. [Google Scholar] [CrossRef]
- Ni, D.; Huang, Y. The Expression, Purification, and Structure Determination of BamA from E. coli. In: Buchanan, S.; Noinaj, N. (eds) The BAM Complex. Methods Mol Biol 2015, 1329, 169–178. [Google Scholar] [CrossRef]
- Mouhib, M.; Benediktsdottir, A.; Nilsson, C.S.; Chi, C.N. Influence of Detergent and Lipid Composition on Reconstituted Membrane Proteins for Structural Studies. ACS Omega 2021, 6(38), 24377–24381. [Google Scholar] [CrossRef]
- Doerner, P.A.; Sousa, M.C. Extreme Dynamics in the BamA β-Barrel Seam. Biochem 2017, 56(24), 3142–3149. [Google Scholar] [CrossRef]
- Iadanza, M.G.; Schiffrin, B.; White, P.; Watson, M.A.; Horne, J.E.; Higgins, A.J.; Calabrese, A.N.; Brockwell, D.J.; Tuma, R.; Kalli, A.C.; et al. Distortion of the bilayer and dynamics of the BAM complex in lipid nanodiscs. Commun Biol 2020, 3, 766. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Wan, K.L.; Mohamed, R.; Nathan, S. Immunization with the recombinant Burkholderia pseudomallei outer membrane protein Omp85 induces protective immunity in mice. Vaccine 2010, 28(31), 5005–5011. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Wan, K.L.; Mohamed, R.; Nathan, S. A genome level survey of Burkholderia pseudomallei immunome expressed during human infection. Microbes Infect 2008, 10(12-13), 1335-1345. [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res 2008, 36(Suppl_2), W5-W9. [CrossRef]
- Xu, Q.; Guo, M.; Yu, F. β-Barrel Assembly Machinery (BAM) Complex as Novel Antibacterial Drug Target. Molecules 2023, 28(9), 3758. [Google Scholar] [CrossRef] [PubMed]
- Malinverni, J.C.; Werner, J.; Kim, S.; Sklar, J.G.; Kahne, D.; Misra, R.; Silhavy, T.J. YfiO stabilizes the YaeT complex and is essential for outer membrane protein assembly in Escherichia coli. Mol Microbiol 2006, 61, 151–164. [Google Scholar] [CrossRef]
- Singh, R.; Capalash, N.; Sharma, P. Immunoprotective potential of BamA, the outer membrane protein assembly factor, against MDR Acinetobacter baumannii. Sci Rep 2017, 7, 12411. [Google Scholar] [CrossRef]
- De Araujo, A.E.V.; Conde, L.V.; da Silva, H.C. Jr; Machado, L.A.; Lara, F.A.; Chapeaurouge, A.; Pauer, H.; Hardoim, C.C.P.; Antunes, L.C.M.; Carvalho-Assef, A.P.D.; et al. Cross-reactivity and immunotherapeutic potential of BamA recombinant protein from Acinetobacter baumannii. Microbes Infect 2021, 23(4-5), 104801. [CrossRef]
- Guan, Q.; Wang, X.; Wang, X.; Teng, D.; Wang, J. In silico analysis and recombinant expression of BamA protein as a universal vaccine against Escherichia coli in mice. Apply Microbiol Biotechnol 2016, 100(11), 5089–5098. [Google Scholar] [CrossRef]
- Storek, K.M.; Auerbach, M.R.; Shi, H.; Garcia, N.K.; Sun, D.; Nickerson, N.N.; Vij, R.; Lin, Z.; Chiang, N.; Schneider, K; et al. Monoclonal antibody targeting the β-barrel assembly machine of Escherichia coli is bactericidal. PNAS 2018, 115(14), 3692-3697. [CrossRef]
- Vij, R.; Lin, Z.; Chiang, N.; Vernes, J.M.; Storek, K.M.; Park, S.; Chan, J; Meng, Y. G.; Comps-Agrar, L.; Luan, P.; et al. A targeted boost-and-sort immunization strategy using Escherichia coli BamA identifies rare growth inhibitory antibodies. Sci Rep 2018, 8, 7136. [Google Scholar] [CrossRef]
- Meng, E.C.; Goddard, T.D.; Pettersen, E.F.; Couch, G.S.; Pearson, Z.J.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci 2023, 32(11), e4792. [Google Scholar] [CrossRef]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Armenteros, J.J.A.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. bioRxiv 2023. [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- McClean, S.; Healy, M.E.; Collins, C.; Carberry, S.; O’Shaughnessy, L.; Dennehy, R.; Adams, Á; Kennelly, H.; Corbett, J.M.; Carty, F.; et al. Linocin and OmpW Are Involved in Attachment of the Cystic Fibrosis-Associated Pathogen Burkholderia cepacia Complex to Lung Epithelial Cells and Protect Mice against Infection. Infect Immun 2016, 84(5), 1424-1437. [CrossRef]
- Casey, W.T.; Spink, N.; Cia, F.; Collins, C.; Romano, M.; Berisio, R.; Bancroft, G.J.; McClean, S. Identification of an OmpW homologue in Burkholderia pseudomallei, a protective vaccine antigen against melioidosis. Vaccine 2016, 34(23), 2616–2621. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Cortázar, J.; Bossi, L.; Quinn, C.; Reynolds, C.J.; Butler, D.K.; Corcoran, N.; Murchú, M.Ó.; McMahon, E.; Singh, M.; Rongkard, P; et al. BpOmpW Antigen Stimulates the Necessary Protective T-Cell Responses Against Melioidosis. Front Immunol 2021, 12, 767359. [Google Scholar] [CrossRef] [PubMed]
- Siritapetawee, J.; Prinz, H.; Krittanai, C.; Sugninta, W. Expression and refolding of Omp38 from Burkholderia pseudomallei and Burkholderia thailandensis, and its function as a diffusion porin. Biochem J 2004, 384(Pt 3), 609–617. [Google Scholar] [CrossRef]
- Kiekens, S.; Sass, A.; van Nieuwerburgh, F.; Deforce, D.; Coenye, T. The Small RNA ncS35 Regulates Growth in Burkholderia cenocepacia J2315. mSphere 2018, 3(1). [CrossRef]
- Sass, A.M.; Coenye, T. The Small RNA NcS25 Regulates Biological Amine-Transporting Outer Membrane Porin BCAL3473 in Burkholderia cenocepacia. mSphere 2023, 8(2), e00083–23. [Google Scholar] [CrossRef] [PubMed]
- Podnecky, N.L.; Rhodes, K.A.; Schweizer, H.P. Efflux pump-mediated drug resistance in Burkholderia. Front Microbiol 2015, 6, 305. [Google Scholar] [CrossRef] [PubMed]
- Bachert, B.A.; Choi, S.J.; Snyder, A.K.; Rio, R.V.M.; Durney, B.C.; Holland, L.A.; Amemiya, K.; Welkos, S.L.; Bozue, J.A.; Cote, C.K.; et al. A Unique Set of the Burkholderia Collagen-Like Proteins Provides Insight into Pathogenesis, Genome Evolution and Niche Adaptation, and Infection Detection. PLoS One 2015, 10(9), e0137578. [Google Scholar] [CrossRef] [PubMed]
- Grund, M.E.; Choi, S.J.; McNitt, D.H.; Barbier, M.; Hu, G.; LaSala, P.R.; Cote, C.K.; Berisio, R.; Lukomski, S. Burkholderia collagen-like protein 8, Bucl8, is a unique outer membrane component of a putative tetrapartite efflux pump in Burkholderia pseudomallei and Burkholderia mallei. PLoS One 2020, 15(11), e0242593. [Google Scholar] [CrossRef]
- Grund, M.E.; Kramarska, E.; Choi, S.J.; McNitt, D.H.; Klimko, C.P.; Rill, N.O.; Dankmeyer, J.L.; Shoe, J.L.; Hunter, M.; Fetterer, D.P.; et al. Predictive and Experimental Immunogenicity of Burkholderia Collagen-like Protein 8-Derived Antigens. Vaccines (Basel) 2021, 9(11), 1219. [Google Scholar] [CrossRef] [PubMed]
- Grund, M.; Choi, S.J.; Powell, L.; Lukomski, S. Intranasal immunization with a Bucl8-based vaccine ameliorates bacterial burden and pathological inflammation, and promotes an IgG2a/b dominant response in an outbred mouse model of Burkholderia infection. Front Immunol 2023, 14. [Google Scholar] [CrossRef]
- Manning, P.A.; Beutin, L.; Achtman, M. Outer membrane of Escherichia coli: Properties of the F sex factor traT protein which is involved in surface exclusion. J Bacteriol 1980, 142(1), 285–294. [Google Scholar] [CrossRef]
- Perumal, N.B.; Minkley, E.G. Jr. The product of the F sex factor traT surface exclusion gene is a lipoprotein. J Biol Chem 1984, 259(9), 5357–5360. [Google Scholar] [CrossRef]
- Chamberlain, N.R.; Brandt, M.E.; Erwin, A.L.; Radolf, J.D.; Norgard, M.V. Major integral membrane protein immunogens of Treponema pallidum are proteolipids. Infect Immun 1989, 57(9), 2872–2877. [Google Scholar] [CrossRef]
- Brandt, M.E.; Riley, B.S.; Radolf, J.D.; Norgard, M.V. Immunogenic integral membrane proteins of Borrelia burgdorferi are lipoproteins. Infect Immun 1990, 58(4), 983–991. [Google Scholar] [CrossRef]
- Wilson, M.M.; Bernstein, H.D. Surface Exposed Lipoproteins: An Emerging Secretion Phenomenon in Gram-negative Bacteria. Trends Microbiol 2016, 24(3), 198–208. [Google Scholar] [CrossRef]
- Schell, M.A.; Zhao, P.; Wells, L. Outer membrane proteome of Burkholderia pseudomallei and Burkholderia mallei from diverse growth conditions. J Proteome Res 2011, 10(5), 2417–2424. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.A.; Seixas, A.M.M.; Mandal, M.; Rodríguez-Ortega, M.J.; Leitão, J.H. Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome. Vaccines (Basel) 2020, 8(3), 509. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat Biotechnol 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Hara, Y.; Mohamed, R.; Nathan, S. Immunogenic Burkholderia pseudomallei Outer Membrane Proteins as Potential Candidate Vaccine Targets. PLoS One 2009, 4(8), e6496. [Google Scholar] [CrossRef]
- Champion, O.L.; Gourlay, L.J.; Scott, A.E.; Lassaux, P.; Conejero, L.; Perletti, L.; Hemsley, C.; Prior, J.; Bancroft, G.; Bolognesi, M.; et al. Immunisation with proteins expressed during chronic murine melioidosis provides enhanced protection against disease. Vaccine 2016, 34(14), 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Dyke, J.S.; Huertas-Diaz, M.C.; Michel, F.; Holladay, N.E.; Hogan, R.J.; He, B.; Lafontaine, E.R. The Peptidoglycan-associated lipoprotein Pal contributes to the virulence of Burkholderia mallei and provides protection against lethal aerosol challenge. Virulence 2020, 11(1), 1024–1040. [Google Scholar] [CrossRef]
- Gourlay, L.J.; Peri, C.; Ferrer-Navarro, M.; Conchillo-Solé, O.; Gori, A.; Rinchai, D.; Thomas, R.J.; Champion, O.L.; Michell, S.L.; Kewcharoenwong, C.; et al. Exploiting the Burkholderia pseudomallei Acute Phase Antigen BPSL2765 for Structure-Based Epitope Discovery/Design in Structural Vaccinology. Chem Biol 2013, 20(9), 1147–1156. [Google Scholar] [CrossRef]
- Michel, L.V.; Shaw, J.; MacPherson, V.; Barnard, D.; Bettinger, J.; D’Arcy, B.; Surendran, N.; Hellman, J.; Pichichero, M.E. Dual orientation of the outer membrane lipoprotein Pal in Escherichia coli. Microbiol (Reading) 2015, 161(Pt 6), 1251–1259. [Google Scholar] [CrossRef]
- Michel, L.V.; Snyder, J.; Schmidt, R.; Milillo, J.; Grimaldi, K.; Kalmeta, B.; Khan, M.N.; Sharma, S.; Wright, L.K.; Pichichero, M.E. Dual Orientation of the Outer Membrane Lipoprotein P6 of Nontypeable Haemophilus influenzae. J Bacteriol 2013, 195(14), 3252–3259. [Google Scholar] [CrossRef]
- Shinoy, M.; Dennehy, R.; Coleman, L.; Carberry, S.; Schaffer, K.; Callaghan, M.; Doyle, S.; McClean, S. Immunoproteomic Analysis of Proteins Expressed by Two Related Pathogens, Burkholderia multivorans and Burkholderia cenocepacia, during Human Infection. PLoS One 2013, 8(11), e80796. [Google Scholar] [CrossRef] [PubMed]
- Makidon, P.E.; Knowlton, J.; Groom, J.V. II; Blanco, L.P.; LiPuma, J.J.; Bielinska, A.U.; Baker, J.R. Jr. Induction of immune response to the 17 kDa OMPA Burkholderia cenocepacia polypeptide and protection against pulmonary infection in mice after nasal vaccination with an OMP nanoemulsion-based vaccine. Med Microbiol Immunol 2010, 199, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, R.; Romano, M.; Ruggiero, A.; Mohamed, Y.F.; Dignam, S.L.; Troncoso, C.M.; Callaghan, M.; Valvano, M.A.; Berisio, R.; McClean, S. The Burkholderia cenocepacia peptidoglycan-associated lipoprotein is involved in epithelial cell attachment and elicitation of inflammation. Cell Microbiol 2016, 19(5), e12691. [Google Scholar] [CrossRef]
- Peri, C.; Gori, A.; Gagni, P.; Sola, L.; Girelli, D.; Sottotetti, S.; Cariani, L.; Chiari, M.; Cretich, M.; Colombo, G. Evolving serodiagnostics by rationally designed peptide arrays: The Burkholderia paradigm in Cystic Fibrosis. Sci Rep 2016, 6, 32873. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.J.; Vigil, A.; DeShazer, D.; Waag, D.M.; Felgner, P.; Goldberg, J.B. Distinct human antibody response to the biological warfare agent Burkholderia mallei. Virulence 2012, 3(6), 510–514. [Google Scholar] [CrossRef] [PubMed]
- Nithichanon, A.; Rinchai, D.; Buddhisa, S.; Saenmuang, P.; Kewcharoenwong, C.; Kessler, B.; Khaenam, P.; Chetchotisakd, P.; Maillere, B.; Robinson, J.; et al. Immune Control of Burkholderia pseudomallei––Common, High-Frequency T-Cell Responses to a Broad Repertoire of Immunoprevalent Epitopes. Front Immunol 2018, 9, 484. [Google Scholar] [CrossRef]
- Seixas, A.M.M.; Sousa, S.A.; Feliciano, J.R.; Gomes, S.C.; Ferreira, M.R.; Moreira, L.M.; Leitão, J.H. A Polyclonal Antibody Raised against the Burkholderia cenocepacia OmpA-like Protein BCAL2645 Impairs the Bacterium Adhesion and Invasion of Human Epithelial Cells In Vitro. Biomedicines 2021, 9(12), 1788. [Google Scholar] [CrossRef]
- Hara, Y.; Chin, C.Y.; Mohamed, R.; Puthucheary, S.D.; Nathan, S. Multiple-antigen ELISA for melioidosis - a novel approach to the improved serodiagnosis of melioidosis. BMC Infect Dis 2013, 13, 165. [Google Scholar] [CrossRef]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, Function, and Evolution of Bacterial ATP-Binding Cassette Systems. Microbiol Mol Biol Rev 2008, 72(2), 317–364. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat Rev Mol Cell Biol 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Garmory, H.S.; Titball, R.W. ATP-Binding Cassette Transporters Are Targets for the Development of Antibacterial Vaccines and Therapies. Infect Immun 2004, 72(12), 6757–6763. [Google Scholar] [CrossRef]
- Harland, D.N.; Dassa, E.; Titball, R.W.; Brown, K.A.; Atkins, H.S. ATP-binding cassette systems in Burkholderia pseudomallei and Burkholderia mallei. BMC Genomics 2007, 8, 83. [Google Scholar] [CrossRef]
- Harland, D.N.; Chu, K.; Haque, A.; Nelson, M.; Walker, N.J.; Sarkar-Tyson, M.; Atkins, T.P.; Moore, B.; Brown, K.A.; Bancroft, G.; et al. Identification of a LolC Homologue in Burkholderia pseudomallei, a Novel Protective Antigen for Melioidosis. Infect Immunol 2007, 75(8), 4173–4180. [Google Scholar] [CrossRef]
- Sharma, S.; Zhou, R.; Wan, L.; Feng, S.; Song, K.; Xu, C.; Li, Y.; Liao, M. Mechanism of LolCDE as a molecular extruder of bacterial triacylated lipoproteins. Nat Commun 2021, 12, 4687. [Google Scholar] [CrossRef]
- Terui, Y.; Saroj, S.D.; Sakamoto, A.; Yoshida, T.; Higashi, K.; Kurihara, S.; Suzuki, H.; Toida, T.; Kashiwagi, K.; Igarashi, K. Properties of putrescine uptake by PotFGHI and PuuP and their physiological significance in Escherichia coli. Amino Acids 2013, 46, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.C.; Blevins, T.C.; Short, S.A. Regulation of peptide transport in Escherichia coli: Induction of the trp-linked operon encoding the oligopeptide permease. J Bacteriol 1986, 165(2), 428–433. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, M.V.; Maier, R.J. Peptide Transport in Helicobacter pylori: Roles of Dpp and Opp Systems and Evidence for Additional Peptide Transporters. J Bacteriol 2007, 189(9), 3392–402. [Google Scholar] [CrossRef] [PubMed]
- Tippayawat, P.; Saenwongsa, W.; Mahawantung, J.; Suwannasaen, D.; Chetchotisakd, P.; Limmathurotsakul, D.; Peacock, S.J.; Felgner, P.L.; Atkins, H.S.; Titball, R.W.; et al. Phenotypic and Functional Characterization of Human Memory T Cell Responses to Burkholderia pseudomallei. PLoS Negl Trop Dis 2009, 3(4), e407. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.K.; Tippayawat, P.; Walker, N.J.; Harding, S.V.; Atkins, H.S.; Maillere, B.; Bancroft, G.J.; Lertmemongkolchai, G.; Altmann, D.M. CD4+ T cell immunity to the Burkholderia pseudomallei ABC transporter LolC in melioidosis. Eur J Immunol 2011, 41(1), 107–115. [Google Scholar] [CrossRef] [PubMed]
- Tippayawat, P.; Pinsiri, M.; Rinchai, D.; Riyapa, D.; Romphruk, A.; Gan, T.H.; Houghton, R.L.; Felgner, P.L.; Titball, R.W.; Stevens, M.P.; et al. Burkholderia pseudomallei Proteins Presented by Monocyte-Derived Dendritic Cells Stimulate Human Memory T Cells In Vitro. Infect Immun 2011, 79(1), 305–313. [Google Scholar] [CrossRef]
- Scott, A.E.; Burtnick, M.N.; Stokes, M.G.M.; Whelan, A.O.; Williamson, E.D.; Atkins, T.P.; Prior, J.L.; Brett, P.J. Burkholderia pseudomallei Capsular Polysaccharide Conjugates Provide Protection against Acute Melioidosis. Infect Immun 2014, 82(8), 3206–3213. [Google Scholar] [CrossRef]
- Settles, E.W.; Sonderegger, D.; Shannon, A.B.; Celona, K.R.; Lederer, R.; Yi, J.; Seavey, C.; Headley, K.; Mbegbu, M.; Harvey, M.; et al. Development and evaluation of a multiplex serodiagnostic bead assay (BurkPx) for accurate melioidosis diagnosis. PLoS Negl Trop Dis 2023, 17(2), e0011072. [Google Scholar] [CrossRef]
- Khemaissa, S.; Sagan, S.; Walrant, A. Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins. Crystals 2021, 11(9), 1032. [Google Scholar] [CrossRef]
- Sousa, S.; Morad, M.; Feliciano, J.R.; Pita, T.; Nady, S.; El-Hennamy, R.E.; Abdel-Rahman, M.; Cavaco, J.; Pereira, L.; Barreto, C.; et al. The Burkholderia cenocepacia OmpA-like protein BCAL2958: Identification, characterization, and detection of anti-BCAL2958 antibodies in serum from B. cepacia complex-infected Cystic Fibrosis patients. AMB Express 2016, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.A.; Liu, Y. Sequencing and characterization of a novel serine metalloprotease from Burkholderia pseudomallei. FEMS Microbiol Lett 2000, 192(1), 67–72. [Google Scholar] [CrossRef] [PubMed]
- Valade, E.; Thibault, F.M.; Gauthier, Y.P.; Palencia, M.; Popoff, M.Y.; Vidal, D.R. The PmlI-PmlR Quorum Sensing System in Burkholderia pseudomallei Plays a Key Role in Virulence and Modulates Production of the MprA Protease. J Bacteriol 2004, 186(8), 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.Y.; Tan, S.C.; Nathan, S. Immunogenic recombinant Burkholderia pseudomallei MprA serine protease elicits protective immunity in mice. Front Cell Infect Microbiol 2012, 2, 85. [Google Scholar] [CrossRef] [PubMed]
- Welkos, S.; Blanco, I.; Okaro, U.; Chua, J.; DeShazer, D. A DUF4148 family protein produced inside RAW264.7 cells is a critical Burkholderia pseudomallei virulence factor. Virulence 2020, 11(1), 1041–1058. [Google Scholar] [CrossRef] [PubMed]
- O'Riordan, A.A.; Morales, V.A.; Mulligan, L.; Faheem, N.; Windle, H.J.; Kelleher, D.P. Alkyl hydroperoxide reductase: A candidate Helicobacter pylori vaccine. Vaccine 2012, 30, 3876–3884. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.H.; Wang, H.F.; Nian, Z.G.; Wang, Y.D.; Zeng, Q.Y.; Zhang, G. Immunization with alkyl hydroperoxide reductase subunit C reduces Fusobacterium nucleatum load in the intestinal tract. Sci Rep 2017, 7, 10566. [Google Scholar] [CrossRef]
- Lou, H.; Li, X.; Sheng, X.; Fang, S.; Wan, S.; Sun, A.; Chen, H. Development of a Trivalent Construct Omp18/AhpC/FlgH Multi Epitope Peptide Vaccine Against Campylobacter jejuni. Front Microbiol 2022, 12. [Google Scholar] [CrossRef]
- Dunachie, S.J.; Jenjaroen, K.; Reynold, C.J.; Quigley, K.J.; Sergeant, R.; Sumonwiriya, M; Chaichana, P. ; Chumseng, S.; Ariyaprasert, P.; Lassaux, P.; et al. Infection with Burkholderia pseudomallei – immune correlates of survival in acute melioidosis. Sci Rep 2017, 7, 12143. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.; Goudet, A.; Jenjaroen, K.; Sumonwiriya, M.; Rinchai, D.; Musson, J.; Overbeek, S.; Makinde, J.; Quigley, K.; Manji, J.; et al. T Cell Immunity to the Alkyl Hydroperoxide Reductase of Burkholderia pseudomallei: A Correlate of Disease Outcome in Acute Melioidosis. J Immunol 2015, 194(10), 4814–4824. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.K.; Orne, C.E.; Shaffer, T.L.; Wilson, S.M.; Khakhum, N.; Torres, A.G.; Brett, P.J.; Burtnick, M.N. Development of Melioidosis Subunit Vaccines Using an Enzymatically Inactive Burkholderia pseudomallei AhpC. Infect Immun 2022, 90(8), e00222–22. [Google Scholar] [CrossRef] [PubMed]
- Klimko, C.P.; Shoe, J.L.; Rill, N.O.; Hunter, M.; Dankmeyer, J.L.; Talyansky, Y.; Schmidt, L.K.; Orne, C.E.; Fetterer, D.P.; Biryukov, S.S.; et al. Layered and integrated medical countermeasures against Burkholderia pseudomallei infections in C57BL/6 mice. Front Microbiol 2022, 13, 965572. [Google Scholar] [CrossRef]
Figure 1.
Breakdown of the Bcc isolates used in the BLAST screen for Bcc-to-B. pseudomallei antigen sequence conservation based on (a) species, (b) continent of origin, and (c) isolation source. Graphs were created using Graphpad Prism 10.
Figure 1.
Breakdown of the Bcc isolates used in the BLAST screen for Bcc-to-B. pseudomallei antigen sequence conservation based on (a) species, (b) continent of origin, and (c) isolation source. Graphs were created using Graphpad Prism 10.
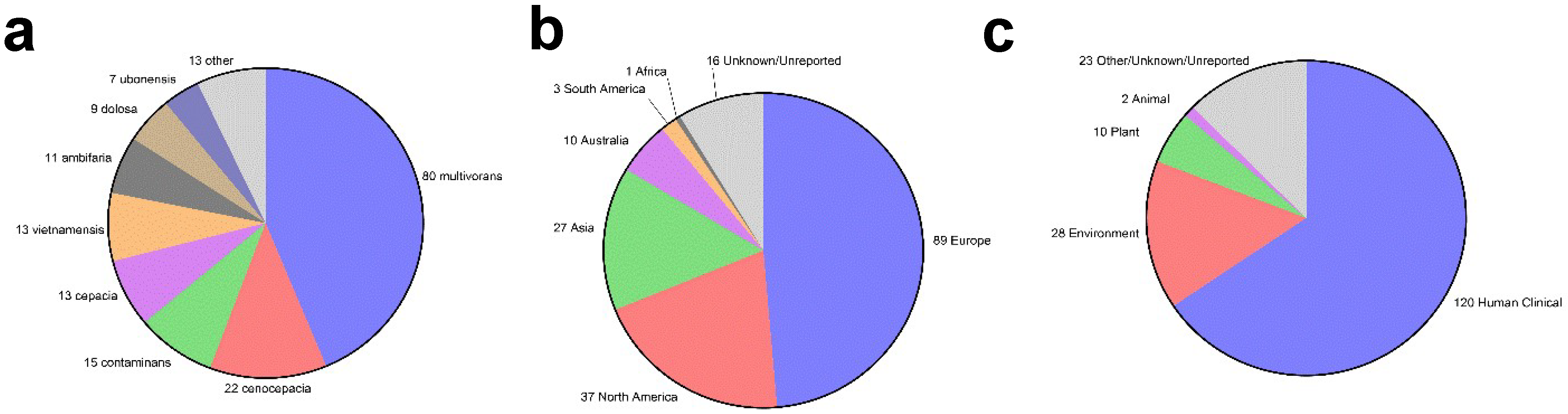
Figure 2.
ChimeraX-generated ribbon structures of β-barrel antigens with a high degree of Bcc-to-B. pseudomallei amino acid sequence identity [161]. Residues not predicted by DeepTMHMM v1.0.24 to be extracellularly oriented are colored in translucent gray [162]. Extracellularly oriented residues that are 100% conserved between the Bcc homologs and the B. pseudomallei K96243 reference are colored light gray. Residues which are substituted or missing in Bcc homologs are colored according to the scale bar to represent the degree of sequence conservation (identity). (a) and (b) are front and back views of the BamA extracellular loops (AFDB accession AF-Q63T20-F1) [163]. (c) and (d) are front and back views of the OmpW1 extracellular loops (AFDB accession AF-Q63UP4-F1) [163].
Figure 2.
ChimeraX-generated ribbon structures of β-barrel antigens with a high degree of Bcc-to-B. pseudomallei amino acid sequence identity [161]. Residues not predicted by DeepTMHMM v1.0.24 to be extracellularly oriented are colored in translucent gray [162]. Extracellularly oriented residues that are 100% conserved between the Bcc homologs and the B. pseudomallei K96243 reference are colored light gray. Residues which are substituted or missing in Bcc homologs are colored according to the scale bar to represent the degree of sequence conservation (identity). (a) and (b) are front and back views of the BamA extracellular loops (AFDB accession AF-Q63T20-F1) [163]. (c) and (d) are front and back views of the OmpW1 extracellular loops (AFDB accession AF-Q63UP4-F1) [163].
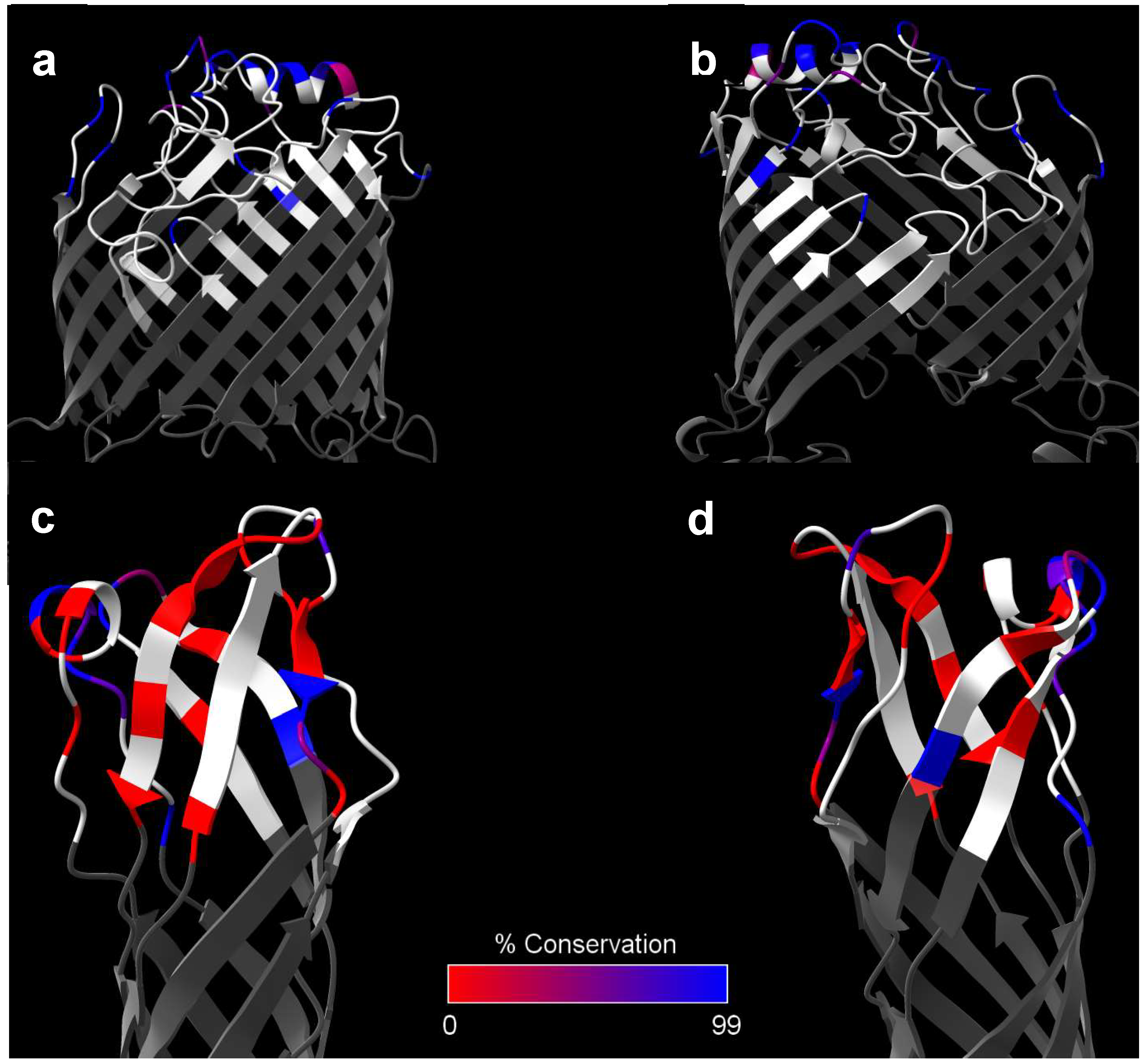
Figure 3.
ChimeraX-generated surface structures of OmpA domain-containing antigens [161]. Residues that are 100% conserved between the Bcc homologs and the B. pseudomallei K96243 reference are colored light gray. Residues which are substituted or missing in Bcc homologs are colored according to the scale bar to represent the degree of sequence conservation (identity). (a) and (b) are front and back views of Omp7 (AFDB accession AF-Q63RA7-F1) [163]. (c) and (d) are front and back views of Omp1 (AFDB accession AF-Q63W89-F1) [163]. (e) and (f) are front and back views of Omp3 (AFDB accession AF-Q63RZ9-F1) [163]. The OmpA domain, which is the most likely region to be targetable by antibodies, is the globular domain in the upper half of each panel.
Figure 3.
ChimeraX-generated surface structures of OmpA domain-containing antigens [161]. Residues that are 100% conserved between the Bcc homologs and the B. pseudomallei K96243 reference are colored light gray. Residues which are substituted or missing in Bcc homologs are colored according to the scale bar to represent the degree of sequence conservation (identity). (a) and (b) are front and back views of Omp7 (AFDB accession AF-Q63RA7-F1) [163]. (c) and (d) are front and back views of Omp1 (AFDB accession AF-Q63W89-F1) [163]. (e) and (f) are front and back views of Omp3 (AFDB accession AF-Q63RZ9-F1) [163]. The OmpA domain, which is the most likely region to be targetable by antibodies, is the globular domain in the upper half of each panel.

Table 1.
Bcc-to-B. pseudomallei amino acid sequence conservation of antigens predicted or known to be accessible for antibody binding.
Table 1.
Bcc-to-B. pseudomallei amino acid sequence conservation of antigens predicted or known to be accessible for antibody binding.
| B. pseudomallei K96243 Locus | Protein Name |
Antigen Class |
Absent in B. mallei? |
Bcc % Identity to K96243 |
Bcc % Similarity to K96243 |
Bcc % Coverage to K96243 |
% of Bcc Strains with Gene |
|---|---|---|---|---|---|---|---|
| BPSL2151 | BamA | β-barrel | 94.0 | 96.9 | 99.8 | 100 | |
| BPSL0999 | Omp1 | Lipoprotein | 93.5 | 95.9 | 99.2 | 100 | |
| BPSL2765 | Omp7 | Lipoprotein | 85.5 | 93.1 | 100 | 100 | |
| BPSL2522 | Omp3 | Other | 91.6 | 92.9 | 99.7 | 100 | |
| BPSL1552 | OmpW1 | β-barrel | 78.3 | 88.0 | 98.8 | 100 | |
| BPSS0879 | OpcP | β-barrel | 75.4 | 82.4 | 100 | 100 | |
| BPSL0281 | FlgL | Flagella | * | 67.9 | 80.8 | 100 | 100 |
| BPSL1972 | Bucl8 | β-barrel | 78.0‡ | 88.2‡ | 98.1‡ | 99 | |
| BPSS0708 | OpcP1 | β-barrel | Partially | 75.7 | 85.5 | 99.9 | 98 |
| BPSL3319 | FliC | Flagella | * | 74.0† | 84.6† | 98.2† | 59† |
| BPSL2704 | OmpW2 | β-barrel | 87.8 | 94.1 | 91.0 | 56 | |
| BPSS1593 | PilV | Pilin | Partially | 42.8 | 56.0 | 78.3 | 25 |
| BPSS1532 | BipB | T3SS | 33.2 | 53.2 | 55.6 | 5 | |
| BPSS1993 | MprA | Other | Absent | 83.3 | 89.6 | 99.3 | 4 |
| BPSS1529 | BipD | T3SS | 32.2 | 55.1 | 58.0 | 1 | |
| BPSS1531 | BipC | T3SS | 0 | 0 | 0 | 0 |
* B. mallei does not express any flagella but retains these genes. ‡ Only compared residues 1-495. C-terminal collagen-like domain was excluded. † Only Bcc FliC homologs with full gene coverage (>95%) were included in these calculations.
Table 2.
Bcc-to-B. pseudomallei amino acid sequence conservation of antigens predicted to serve primarily as T cell antigens.
Table 2.
Bcc-to-B. pseudomallei amino acid sequence conservation of antigens predicted to serve primarily as T cell antigens.
| B. pseudomallei K96243 Locus | Protein Name |
Antigen Class |
Absent in B. mallei? |
Bcc % Identity to K96243 |
Bcc % Similarity to K96243 |
Bcc % Coverage to K96243 |
% of Bcc Strains with Gene |
|---|---|---|---|---|---|---|---|
| BPSL2096 | AhpC | Other | 98.9 | 99.7 | 100 | 100 | |
| BPSL2287 | IscA | Other | 95.7 | 97.2 | 100 | 100 | |
| BPSL2277 | LolC | ABC Transporter | 93.3 | 97.1 | 100 | 100 | |
| BPSL3105 | Hcp6 | T6SS | Absent | 92.2 | 97.1 | 100 | 100 |
| BPSS0467 | PotF | ABC Transporter | Absent | 86.1 | 92.1 | 99.4 | 100 |
| BPSS2141 | OppA | ABC Transporter | 82.8 | 89.4 | 96.4 | 100 | |
| BPSL3369 | AcoD | Other | 74.3 | 84.7 | 100 | 100 | |
| BPSL1897 | TadE | Other | 48.3 | 60.7 | 92.6 | 100 | |
| BPSS0171 | Hcp4 | T6SS | 89.2 | 95.7 | 100 | 48 | |
| BPSS2098 | Hcp3 | T6SS | 93.1 | 97.7 | 100 | 16 | |
| BPSS1498 | Hcp1 | T6SS | 25.6 | 42.0 | 97.9 | 12 | |
| BPSS0099 | Hcp5 | T6SS | Absent | 88.6 | 95.1 | 100 | 8 |
| BPSS0518 | Hcp2 | T6SS | Partially | 95.6 | 98.3 | 100 | 2 |
| BPSS1512 | TssM | T6SS* | 0 | 0 | 0 | 0 | |
| BPSS1524 | BopA | T3SS | 0 | 0 | 0 | 0 |
* TssM is considered part of the T6SS-1 genomic locus but does not appear to play a functional role in the T6SS-1.
Table 3.
Outer Membrane β-barrel antigen sequence identity in extracellular loop domains.
| B. pseudomallei K96243 Locus | Protein Name |
Bcc % Identity to K96243 (Full Protein) | Bcc % Identity to K96243 (Extracellular Loops) |
|---|---|---|---|
| BPSL2151 | BamA | 94.0 | 98.2 |
| BPSL2704 | OmpW2 | 87.8 | 88.9 |
| BPSL1972 | Bucl8 | 78.0 | 88.0 |
| BPSS0708 | OpcP1 | 75.7 | 69.8 |
| BPSL1552 | OmpW1 | 78.3 | 67.7 |
| BPSS0879 | OpcP | 75.4 | 59.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



