
Submitted:
12 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Animal trypanosomiasis (AT) is a complex of veterinary diseases known under various names such as nagana, surra, dourine and mal de caderas, depending on the country, the infecting trypanosome species and the host. AT is caused by parasites of the genus Trypanosoma, and the main species infecting domesticated animals are T. brucei brucei, T. b. rhodesiense, T. congolense, T. vivax, T. evansi and T. equiperdum. AT transmission, again depending on species, is through tsetse flies or common Stomoxys and tabanid flies or through copulation. Therefore, the geographical spread of all forms of AT together is not restricted to the habitat of a single vector like the tsetse fly and currently includes almost all of Africa, and most of South America and Asia. The disease is a threat to millions of companion and farm animals in these regions, creating a financial burden in the billions of dollars to developing economies as well as serious impacts on livestock rearing and food production. Despite the scale of these impacts, control of AT is neglected and under-resourced, with diagnosis and treatments being woefully inadequate and not improving for decades. As a result, neither the incidence of the disease, nor the effectiveness of treatment is documented in most endemic countries, although it is clear that there are serious issues of resistance to the few old drugs that are available. In this review we particularly look at the drugs, their application to the various forms of AT, and their mechanisms of action and resistance. We also discuss the spread of veterinary trypanocide resistance and its drivers, and highlight current and future strategies to combat it.
Keywords:
Nagana
; Surra
; animal trypanosomiasis
; drug resistance
; history
; trypanocide
; drug mechanism of action
1. Introduction
Protozoan parasites of the genus Trypanosoma, most prominently including T. brucei spp, T. congolense, T. vivax, T. evansi and T. equiperdum, cause a wide range of diseases affecting humans and animals. With the exception of T. equiperdum, which is transmitted by copulation, pathogenic trypanosomes are mostly transmitted by insect vectors (Magez et al., 2021) and in parts of South America also by vampire bats (Herrera et al., 2004). In Africa south of the Sahara, trypanosomiasis is mostly transmitted by the tsetse fly and known as African animal trypanosomiasis (AAT) or nagana, whereas beyond the tsetse belt, T. vivax and T. evansi are transmitted by other haematophagous flies such as tabanids and Stomoxys spp. The disease is commonly known as surra from North Africa to Asia, while in South America it is known by a variety of names including mal de caderas and derrengadera. The wasting disease caused by T. equiperdum in horses and other equids is known as dourine. The mode of transmission and the availability of susceptible hosts concentrates the disease in rural agricultural regions or communities with poor housing conditions around the world. While the human trypanosomiases – sleeping sickness and Chagas disease – lead to major economic losses through Disability-Adjusted Life Years (DALYs) (Lee et al., 2013; Sutherland et al., 2017), the veterinary trypanosomiases constitute a big challenge to the rural economies and food security of developing countries (Giordani et al., 2016).
The control of African animal trypanosomiasis (AAT) and surra depends largely on chemotherapy (Peregrine, 1994), making dourine the only veterinary trypanosomiasis for which test-and-slaughter is the only option (Gizaw, Megersa and Fayera, 2017). However, all the six veterinary trypanocides in current use – diminazene, isometamidium, homidium, quinapyramine, suramin and cymelarsan – are all between half a century and over a century old, and their usage is associated with toxicity and treatment failure (Anene et al., 2001; Giordani et al., 2016). Moreover, each of these drugs has its own spectrum of which trypanosome species it is effective against, and susceptibility varies widely (Ungogo et al., 2022) and as such the number of drugs used in a given geographical region is actually a fraction of the total.
As the overdependence on the few antitrypanosomal drugs persists, a continuous exposure of Trypanosoma populations to suboptimal dosages of the drugs resulted in a widespread resistance of the parasites to the drugs. While the recent discovery of a vaccine candidate for T. vivax (Autheman et al., 2021), and the prospects for development of the benzoxaborole AN11736 into a new veterinary trypanosomiasis drug (Giordani et al., 2020) are among reasons for cautious optimism, the introduction and implementation of new drugs or vaccines remains many years away. Thus, there remains a need to study drug resistance in animal trypanosomes, and use the knowledge to optimise current veterinary practice (Richards et al., 2021), and shape efforts to develop new drugs. In this light, investigations into the current drugs, how they act in each of the divergent animal trypanosomes, and how each species becomes resistant to the drugs, as well as the mechanisms of cross-resistance will provide useful insights.
The sustained efforts into T. brucei brucei over decades have furthered the understanding of its biology and pharmacology. Importantly, how this parasite internalises and interacts with the drugs used against it have been fully elucidated (Alsford et al., 2012; Munday et al., 2015b; Thomas et al., 2018; De Koning, 2020). The research into T. b. brucei as a model for trypanosomatid species was in part sustained because most findings in the veterinary pathogen T. b. brucei equally apply to the subspecies affecting humans, T. b. rhodesiense and T. b. gambiense, as well as the closely related animal trypanosomes T. evansi and T. equiperdum, all in the subgenus Trypanozoon (Alsford et al., 2012; Stewart et al., 2010; Witola et al., 2004). This might explain the efficacy of suramin and melaminophenyl arsenicals in the treatment of both the human sleeping sickness and the veterinary disease surra (Desquesnes et al., 2013; Wiedemar et al., 2020), while being much less effective against nagana, which is predominantly caused by T. congolense (subgenus Nannomonas) and T. vivax (subgenus Duttonella) (Ungogo et al., 2022).
With the successes reported in the fight against HAT in Africa and the WHO goal of eradication of the disease being realised, at least for the overwhelmingly anthroponotic T. b. gambiense infections (Franco et al., 2018b), it is important to now focus on the animal trypanosomiases (Morrison et al., 2016). It is also timely to evaluate all the knowledge gathered about T. brucei to see what does or does not apply to veterinary trypanosomes. This includes the effectiveness, mode of action and mode of resistance of drugs, which are potentially different for trypanosomes of the different sub-genera (Giordani et al., 2016). For example, T. brucei internalises diminazene through P2/AT1 aminopurine transporter which determines sensitivity and resistance to several drugs (Matovu et al., 2003; Ward et al., 2011; Graf et al., 2013). In contrast, T. congolense and T. vivax lack the orthologue of P2/AT1 (Jackson et al., 2013; Munday et al., 2013; Ungogo et al., 2023), and diminazene resistance in T. congolense is not determined by its capacity to transport the drug (Carruthers et al., 2021).
Here we review the problem of drug resistance in animal trypanosomiases, highlight its molecular mechanisms, to the extent known, and discuss current and emerging strategies to combat this challenge.
2. Animal trypanosomiases: a global problem
2.1. The globalisation of animal trypanosomiasis
Trypanosomiasis is a broad term used for a collection of human and veterinary diseases caused by the kinetoplastid protozoa of the genus Trypanosoma. In humans, there are two divergent diseases caused by different Trypanosoma species and transmitted by unrelated insect vectors in different continents; Human African Trypanosomiasis (HAT or sleeping sickness) and American Trypanosomiasis or Chagas Disease (Barrett et al., 2003; Namangala and Odongo, 2014). HAT is caused by salivarian parasites Trypanosoma brucei gambiense and T. b. rhodesiense and is named ‘Human African Trypanosomiasis’ due to its restriction to sub-Saharan Africa, courtesy of relying exclusively on the Glossina (tsetse fly) vector that hardly survives outside a defined tropical habitat, and ’sleeping sickness’ due to a late-stage nervous condition partly characterised by daytime somnolence and an instant inordinate sleep impulse (Barrett et al., 2003). Chagas Disease is caused by the stercorarian species T. cruzi, which is transmitted by haematophagous reduviid bugs (Lidani et al., 2019).
In animals, however, trypanosomiasis affects an expanding area on the world map and takes different forms, with varied means of transmission. Importantly, a classical wasting disease of livestock, called African Animal Trypanosomiasis (AAT, or nagana), which is very much related to HAT and transmitted by similar tsetse vectors, is caused predominantly by T. congolense, T. vivax and T. b. brucei. The non-tsetse transmitted animal trypanosomiases (NTTAT), endemic in multiple continents include surra, caused by T. evansi and transmitted mechanically by biting flies, and the sexually transmitted dourine caused by T. equiperdum in equids (Giordani et al., 2016; Desquesnes et al., 2013). Known principally as a disease of man, Chagas disease may also result in clinical illness in young animals which can turn fatal if left untreated (Barr, 2009). The ability of both T. b. gambiense and T. b. rhodesiense to establish infection in animals has been demonstrated (Yesufu, 1971; Njiokou et al., 2010; Hamill et al., 2013), but whereas T. b. rhodesiense is a zoonosis with large reservoirs in many wild animals, transmission of T. b. gambiense is believed to be almost exclusively between humans (Franco et al., 2018b), although Group1 T. b. gambiense has also been detected I a variety of domesticated animals, including, recently, in Nigerian dogs (Umeakuana et al., 2019). On the other hand, the number of reports on atypical human infections with Trypanosoma species that were understood to only infect animals including T. b. brucei, T. vivax, T. congolense, T. evansi, T. lewisi, and T. lewisi-like, though rare, is rising (Truc et al., 1998, 2013; Pumhom et al., 2015), although it is impossible to say whether this is because of improved (molecular) detection and case finding, or a genuine increase in cases. Although a human infection with T. evansi was linked to a specific immune defect in the patient, the lack of Apolipoprotein L-1 (Vanhollebeke et al., 2006), infection with T. lewisi and related species, for which the dominant hosts are rodents, are not believed to be linked to an immune deficiency (Truc et al., 2013), nor was there any indication of an immune system deficiency or HIV in the case of human T. congolense infection (Truc et al., 1998). There are thus legitimate questions as to how absolute the delineation of the diseases and species into ‘animal’ and ‘human’ really is, and this comes with the attendant public health significance.
Despite significant differences, animal trypanosomiases share several clinical presentations including anaemia and emaciation as well as their negative impact on the rural and agricultural economies (Namangala and Odongo, 2014). Blood is the principal clinical sample for the diagnosis of animal trypanosomiases. In addition, dourine can be diagnosed using a vaginal swab, conjunctival swab, joint fluid, skin biopsy and cerebrospinal fluid (Pascucci et al., 2013). Diagnostic methods include direct microscopic examination to identify the parasite and molecular methods such as PCR, complement fixation and indirect immunofluorescence tests (Desquesnes et al., 2022).
Nagana or AAT is a debilitating disease of domestic animals endemic in 37 out of 54 African countries, covering about 10 million km2 of arable land, more than one-third of the continent, where it wreaks serious economic havoc (Wilkowsky, 2018; Yaro et al., 2016). An estimated 55 million cattle and approximately 100 million other farm animals are at risk of AAT, and the disease kills approximately 3 million livestock animals annually (Kabayo, 2002). Restricted only to pigs, a fourth trypanosome species, T. simiae, causes a highly fatal disease transmitted by tsetse in Africa. T. vivax can also be mechanically transmitted by other vectors, particularly horseflies (Tabanus spp.) and stable flies (Stomoxys spp.), because its short life cycle does not require development in the tsetse midgut (Namagala and Odongo, 2014; Giordani et al., 2016); this explains their prevalence outside the tsetse habitat, particularly in South America (Gonzatti et al., 2014). A recent report documented an alarming prevalence of T. vivax in camels in Iran, often as co-infection with T. evansi (Asghari and Rassouli, 2022) – the first report of T. vivax in Asia according to these authors.
While AAT is characterised by a cocktail of clinical signs including lethargy, lymphadenopathy, splenomegaly, oedema, emaciation, immunosuppression, abortion and reduced milk production, anaemia is the most prominent finding and causes the highest impact (Taylor and Authié, 2004). In the absence of treatment, mortality can ensue within weeks of onset, else the acute disease leads to a chronic illness characterised by lethargy and infertility that may subsist for several years before death (Taylor and Authié, 2004). While subtle differences exist in the presentation of diseases caused by the different AAT-causing trypanosomes, symptoms are largely indistinguishable, and mixed infection is very common (Giordani et al., 2016).
Surra is a wasting disease of animals caused by T. evansi, transmitted mechanically by multiple genera of biting flies, with almost the same clinical presentation as AAT (Brun et al., 1998; Giordani et al., 2016). In addition to mechanical transmission by biting flies, T. evansi, which do not require biological development in a vector, may be spread by vampire bats, as well as through sexual, horizontal, vertical and iatrogenic transmission (Desquesnes et al., 2013). T. evansi infects most domestic animals and numerous wildlife species and is the animal trypanosome with the widest geographical reach, covering North Africa, the Middle East, much of Asia, Latin America as well as some parts of Europe, making surra a significant challenge to agriculture around the world (Brun et al., 1998; Desquesnes et al., 2013). The nonpathognomonic nature of trypanosome infection clearly complicates diagnosis, requiring molecular analysis for certainty in species identification and it is possible that infections in the Middle East or Asia have been ascribed to T. evansi by default, letting T. vivax spread go unnoticed until recently (Asghari and Rassouli, 2022). As T. vivax and T. evansi have quite different sensitivity to some drugs, T. vivax for instance being much less sensitive to suramin (Leach and Roberts, 1981), the issue of correct species identification is of prime importance for successful treatment and thus for the avoidance of resistance by underdosing as well.
Dourine, caused by T. equiperdum, is a sexually transmitted disease of equids that is found to be very fatal in horses but usually sub-clinical in donkeys and mules (Brun et al., 1998). The disease is characterised by anaemia, cachexia, oedema, swelling of the genitalia and mammary glands, cutaneous plaques, and neurological signs including incoordination and paralysis of face and lips (Gizaw et al., 2017). Dourine is an endemic disease in Africa and Asia but has also been reported in the Middle-East, South-East Europe and South America (Brun et al., 1998; Giordani et al., 2016; Gizaw et al., 2017). As it requires no vector, and there is no vaccine or prophylactic treatment, the spread of the disease must be avoided through rigorous screening of imported equids and by careful, controlled breeding programmes.
T. cruzi infection can result in clinical disease in young dogs, cats, and pigs. The parasite is transmitted to animals by triatomine bugs, which deposit infected faeces on wounds during a bloodmeal (Namangala and Odongo, 2014). In addition, there is increased focus on oral transmission (Shikanai Yasuda, 2022) through contaminated food and drink and not just in humans; de Noya and González describe this as the ‘ancestral form of transmission to wild and domestic mammals’ (de Noya and González, 2015). The disease is historically endemic in poor rural areas of Central and South America, but has spread northward and infected vectors are now endemic in Mexico and the southern United States (Garcia et al., 2015; Mills, 2020). The acute disease can be fatal (Namangala and Odongo, 2014), but recovered animals proceed to the mainly asymptomatic chronic (but still infectious) form (Barr, 2009; Stevens et al., 2011).
2.2. Economic Importance of Animal Trypanosomiasis
The local name for AAT, ‘nagana’ (which means ‘useless’, or ‘powerless’ in Zulu), accurately illustrates the impact of animal trypanosomiases on both the animal and its owner. Losses due to nagana and surra are incurred directly through morbidity, impaired fertility, mortality, treatment costs and the disease and vector control programmes as well as indirectly from animal culling and impaired animal traction (Shaw et al., 2014; Desquesnes et al., 2013; Kumar et al., 2017). AAT is considered one of the most important livestock diseases in sub-Saharan Africa, causing agricultural economic losses estimated to run to $4.75 billion annually, with devastating social implications on rural farming communities (Kristjanson et al., 1999; Shaw et al., 2014). While precise data on the overall global economic impact of surra is lacking, many studies have explored national and regional economic impacts of the disease (Desquesnes et al., 2013), and in India alone, direct and indirect losses due to surra are estimated at $671.1 million per annum (Kumar et al., 2017). Dourine poses a serious challenge to equine production (Mizushima et al., 2020), and is a great barrier to the international trade and movement of horses (Gizaw et al., 2017). T. vivax, which can also be transmitted mechanically, has spread beyond the African tsetse belt (Fetene et al., 2021) and is already causing important economic losses in South America. Although there is a dearth of accurate data on the extent of the economic losses caused by T. vivax infection in South America, a study has shown that a single epidemic in a 250,000 km2 area of the Brazilian Pantanal and the Bolivian lowlands in 1995 caused losses amounting to $160 million (Seidl et al., 1999).
An estimated 35-50 million doses of trypanocidal drugs are administered to animals annually in tropical Africa for curative and preventive purposes (Kristjanson et al., 1999; Richards et al., 2021). Each treatment costs approximately $0.5 – 3.55 and the total loss due to the purchase of treatments alone runs to $90 million annually (Sutcliffe et al., 2014; Richards et al., 2021). However, many studies reported that trypanocides are sold informally on African markets and when these are additionally considered, the number of trypanocide doses sold may be as high as 70 million doses per annum. In cases of treatment failure, repeat treatment using a higher dose of the same drug or a different one is usually carried out (Anene et al., 2001). As such, the total doses of trypanocides used and the cost involved would likely be considerably lower in the absence of drug resistance and treatment failure (Richards et al., 2021).
2.3. Control of Animal Trypanosomiasis
Unlike other protozoan diseases, like malaria and leishmaniasis, the chances of vaccine development for trypanosomiasis in the near future are slim. This should be attributed in large extent to antigenic variation, which allows the parasite to continuously change its surface glycoproteins, coupled with the parasite’s ability to remove surface-bound immune effectors, thereby undermining production of effective antibodies by the host (Horn, 2014; Quintana et al., 2018). A T. vivax invariant surface glycoprotein (ISG) antigen named IFX was recently reported to induce protective immunity in a mouse model (Autheman et al., 2021). However, the IFX antigen failed to protect goats from T. vivax infection (Ramirez, 2020), and the lack of homologues of the glycoprotein in other African animal trypanosomes (Autheman et al., 2021) would raise concern for applicability since mixed infection is common in endemic areas: T. vivax is co-endemic with T. evansi in South America and with T. b. brucei and T. congolense in Africa. Thus, the control of vector-transmitted trypanosomiases depends mainly on targeting the disease vector(s), and on chemotherapy against the parasite in its mammalian hosts (Giordani et al., 2016).
Vector control measures include an array of traditional and modern techniques aimed at preventing exposure of animals to the tsetse and other insect vectors, reducing vector population in a specific area or even eradicating it completely from a defined region (Kabayo, 2002; Yaro et al., 2016). Outdated and environmentally detrimental vector control strategies included bush clearing (Ford et al., 1970), killing of wild animal reservoir (Ford, 1970) as well as aerial and ground spraying with insecticides (Potts and Jackson, 1952; WHO, 2013). More selective and environmentally friendly methods used currently include bait methods, using different screens to attract tsetse; use of insecticides on the animal host, either ‘pour on’ or impregnated on curtains and dog-collars; as well as the sterile insect technique (Dame and Schmidt, 1970; Gooding and Krafsur, 2005; Vreysen et al., 2013). With the advances in genomics, new frontiers of targeting genes, proteins and molecules in vectors, their endosymbionts and the trypanosomes are being explored (Farikou et al., 2010). However, the tsetse-specific control programmes in Africa, however important, are insufficient considering the adaptation of T. vivax to mechanical transmission (Desquesnes and Dia, 2003; Ooi et al., 2016). This underscores the need for a more holistic approach to vector control in the control of nagana, although this is not to argue that tsetse control in specific foci is not of significant value and would reduce the incidence of nagana.
Chemotherapy for trypanosomiases is more than a century old and the whole field of medicinal chemistry finds its roots in the efforts to provide a cure for the disease, which devastated the then European colonies in sub-Saharan Africa (Steverding, 2010). Resistance has now been reported to most of the available drugs for the treatment of human and animal trypanosomiases, and this, coupled with marked toxicity, has fuelled calls for the development of newer, safer and more effective drugs (Delespaux and De Koning, 2007). However, there is little investment in the development of new veterinary trypanocides mainly due to the high costs of development and ultimate registration in dozens of countries, limited market and low prospects for profits (Geerts et al., 2001; Maudlin, 2006). Moreover, the current trypanocides like diminazene and isometamidium are exceedingly well known by livestock owners in Africa for at least two generations and are cheap and widely available. In addition, both the choice and the administration of treatment are overwhelmingly in the hands of the animal’s owner rather than a licenced veterinarian. Thus, the road from discovery of a new veterinary trypanocide to the successful implementation across the huge regions affected is a daunting one indeed.
Some African livestock breeds that are indigenous to tsetse-endemic areas, such as N’Dama cattle, West African Dwarf goat and Djallonke sheep, possess a remarkable inherent capacity to thrive despite the challenges of nagana – a phenomenon known as trypanotolerance (Dwinger et al., 1994; Yaro et al., 2016). This can provide part of a mitigation strategy for living with nagana, considering the success recorded by programmes utilising similar genetic-based disease tolerance in other parts of the world.
The control of Chagas disease on the other hand relies heavily on reducing the bug population around domestic settings, through improved housing conditions and hygienic practices, such as the use of metal roofing instead of thatched roofs or the replacement of piled brushwood with wire fencing in animal enclosures (Stevens et al., 2011). In addition to chemotherapy and vector control, the control of surra incorporates movement restriction of affected animals from infected to non-infected areas through screening of carriers and quarantine. Transmission to sensitive equines can be minimised from bovine and canine reservoir by separating cattle herds and horse stables kilometres apart, and by not feeding dogs with carcasses of infected animals, respectively (Desquesnes et al., 2013). Vector control for surra mainly targets the horsefly and stable fly as well as vampire bats in Latin America where they significantly transmit the disease (Foil and Hogsette, 1994; Arellano-Sota and Arellano-Sota, 1988; Desquesnes et al., 2013).
The control of the sexually transmitted dourine, however, is hinged on a different strategy, with chemotherapy highly discouraged since recovered animals are potential carriers – and transmitters – of the parasite (Giordani et al., 2016). The World Organization for Animal Health (OiE) puts forward a stamping-out policy for dourine recommending compulsory notification and slaughter of infected animals, movement control and hygienic practices during assisted mating (OIE, 2013). However, this stern ‘test and slaughter’ eradication policy is unrealistic especially in endemic developing countries considering the huge cost implications and it has not been successful in stopping the spread of the disease. Moreover, recent attempts to treat dourine have shown promising results (Gizaw et al., 2017; Davkharbayar et al., 2020).
3. Veterinary trypanocides: mechanisms of action and resistance
The molecular mechanisms of drug resistance in animal trypanosomes are still being elucidated, although advances have been made in the highly studied T. brucei. The entry of the drug into the trypanosome appears to be the first determinant of drug activity (Figure 1). Of great importance in drug uptake and resistance of existing trypanocides are protein transporters on the surface membrane (Munday et al., 2015a). Specifically, the P2/AT1 transports diamidines (Stewart et al., 2005; Ward et al., 2011), melaminophenyl arsenicals (Carter and Fairlamb, 1993; De Koning et al., 2000) and pentamidine (Carter et al., 1995; De Koning and Jarvis, 2001) in the Trypanozoon subgenus parasites: T. brucei, T. evansi and T. equiperdum. Moreover, the aquaglyceroporin AQP2 effectively functions as a high affinity pentamidine transporter and is responsible for the uptake of melaminophenyl arsenicals as well (Alghamdi et al., 2020; Baker et al., 2012; Munday et al., 2014). Suramin is mainly transported via endocytosis after specific binding to an invariant surface glycoprotein, ISG75 (Zoltner et al., 2016, 2020) and there is substantial evidence pointing to the role of specific transporters in the uptake isometamidium (Sutherland, Mounsey and Holmes, 1992; Wilkes et al., 1995, 1997; Eze et al., 2016). While there is strong cellular uptake of isometamidium and from there rapid uptake into the mitochondrion, there is also a substantial component of energy-dependent efflux from the cell (Eze et al., 2016; Sutherland and Holmes, 1993). This makes the dynamics of the local isometamidium concentration very complicated as it is governed by multiple, different interdependent processes (Sutherland et al., 1992a): influx end efflux across the cell membrane and the same across the mitochondrial and other organellar membranes and the binding to target molecules/structures and off-target structures including nuclear and mitochondrial DNA (Boibessot et al., 2002; Wilkes et al., 1995). Moreover, (most of) the isometamidium transport processes are energy dependent (Sutherland et al., 1992a; Sutherland and Holmes, 1993), making the plasma membrane potential and mitochondrial membrane potential (MMP) part of the driving force as well as one of the affected parameters of the drug action (Wilkes et al., 1997; Eze et al., 2016). This example highlights the complexities in drug uptake/efflux/distribution in a target cell. Following entry into the trypanosome, mediated by one or more transport mechanisms, trypanocides target specific and general biomolecules and organelles to disrupt essential metabolic processes and/or facilitate cell killing. A number of trypanocides target the mitochondrion and the kinetoplast, a distinct structure connected to the flagellar basal body and containing the mitochondrial DNA. Mutations that disrupt drug transporters and/or targets (Figure 1) have been shown to result in drug resistance, defined as decreased or loss of sensitivity of the parasite to a drug at a dose that would otherwise be toxic to it.
3.1. Diamidines
The discovery of diamidines resulted from a systematic search for agents that would lower the level of glucose in host blood, based on findings that this nutrient was essential in the parasite (Schoenbach and Greenspan, 1948; Yorke et al., 1929). The hypoglycaemic drug synthalin (Figure 2A) was soon found to exert potent antitrypanosomal activity in mice and rats (Lourie and Yorke, 1937), and although it was shown not to act through any effects on host glucose levels (King et al., 1937), the discovery stimulated the synthesis and screening of related compounds. Further modifications of the structure of synthalin led to more effective and safer derivatives containing two terminal benzamidines (Lourie and Yorke, 1939). Out of several diamidines with different substitutions, stilbamidine, pentamidine, propamidine and phenamidine (Figure 2A) were promoted to clinical investigation in the 1940’s. From these, pentamidine and stilbamidine were taken into clinical use for early stage sleeping sickness but stilbamidine was discontinued due to neurotoxicity in some patients (Napier and Gupta, 1942). The development of the veterinary trypanocides in the 1950’s, the phenanthridines and quinapyramine, benefitted from the experimental methods for in vivo and in vitro screening established in the development of the diamidines.
Diamidines are not active against late-stage trypanosomiasis, where the parasite is present in the central nervous system (CNS). Nevertheless, there are some reports of pentamidine being effective against ‘early late-stage’ sleeping sickness (Lejon, 2003; Singaro et al., 1996) and [3H]-pentamidine was shown to cross the blood-brain barrier (BBB); however, P-glycoproteins and multi-drug resistance efflux pumps on the BBB kept the pentamidine concentration in the CNS at sub-curative concentrations (Sanderson et al., 2009). Interestingly, although furamidine (DB75) and its prodrug pafuramidine, were also not effective against cerebral trypanosomiasis (Paine et al., 2010), its aza analogue DB829 (later renamed CPD0801) was highly effective against cerebral trypanosomiasis in a vervet monkey model, curing 5/5 animals with intramuscular administration of just 2.5 mg/kg/day for 5 consecutive days (Thuita et al., 2015). As the physical parameters of DB75 and DB829 are virtually identical (Figure 2B), the improved penetration into the CNS of the latter cannot be ascribed to superior diffusion across the BBB. In contrast, it is highly likely that DB829 employs a transporter of the BBB endothelial cells, just like pentamidine does, while being a poorer substrate for the efflux transporters. DB829 should be considered for further clinical or veterinary development but the discontinuation of the pafuramidine clinical trials after the observation of severe renal complications in five patients (Paine et al., 2010; Pohlig et al., 2016) gave the furamidine series a severe disadvantage and may have doomed the series. This is perhaps undeserved as it is likely that it was the N-methoxy prodrug strategy that caused the furamidine toxicity, by trapping the prodrug in renal cells after its intracellular activation.
Diminazene is an aromatic diamidine (4,4′-(diazoamino) benzamidine) that is active against African trypanosomes, T. evansi and Babesia spp (Peregrine and Mamman, 1993). Diminazene aceturate (DA) is the most widely used trypanocide in SSA for the treatment of nagana due to its higher therapeutic index and its activity against trypanosomes resistant to other trypanocides (Peregrine, 1994). Despite poor efficacy or tolerance in some animals, especially in horses and dogs, DA is still used to treat surra in many parts of the world (Desquesnes et al., 2013). There were reports of off-label use of DA to treat early-stage T. b rhodesiense and T. b. gambiense sleeping sickness in humans with some appreciable success and low toxicity (Delespaux and De Koning, 2007), but the indiscriminate use of veterinary and human trypanocides was strongly discouraged, especially since there is cross-resistance between diminazene and pentamidine in T. brucei (Bridges et al., 2007).
The entry of diminazene into trypanosomes via passive diffusion is unlikely given the highly charged nature of the compound (it is a di-cation at neutral pH), and as such specific transporters are required (Delespaux and De Koning, 2007). The role of the TbAT1/P2 adenosine/adenine transporter in the uptake of diamidines in T. brucei has been demonstrated (Mäser et al., 1999; Matovu et al., 2003; De Koning et al., 2004). The presence of a P2/AT1 transport system similar to T. brucei, and its role in diminazene uptake, has also been demonstrated in T. evansi and T. equiperdum (Berger et al., 1995; Witola et al., 2004; Stewart et al., 2010). For the most important species causing AAT, T. congolense and T. vivax, however, it appears that the mechanism of diminazene uptake is different, owing to the lack of authentic orthologues of TbAT1 in these species (Jackson et al., 2013; Munday et al., 2013; Ungogo et al., 2023). Previously, T. congolense gene TcIL3000_9_2500 was thought to be an orthologue to the T. brucei AT1 gene in T. congolense and was named TcoAT1 (Delespaux et al., 2006). The gene was proposed to be responsible for diminazene uptake in T. congolense after a strong correlation between a Single Strand Conformation Polymorphism (SSCP) pattern and drug sensitivity was established using a single dose mouse test. It was therefore also proposed that an SNP in TcoAT1 confers resistance to diminazene in T. congolense. Subsequently, based on this understanding, a PCR-restriction fragment length polymorphism (PCR-RFLP) test to determine the SSCP in TcoAT1 was developed for the diagnosis of diminazene resistance in T. congolense (Delespaux et al., 2006) and enhanced using the DpnII restriction enzyme (Vitouley et al., 2011). However, Munday et al., (2013) have shown that the expression of the TcoAT1 gene did not restore the sensitivity of drug resistant T. brucei B48 to diminazene. Further analysis and characterisation indicated that TcoAT1 is neither an orthologue of TbAT1 nor even a P2-type transporter, but a P1-type adenosine/inosine transporter that is not involved in diminazene uptake and hence highly unlikely to be related to resistance in T. congolense (Munday et al., 2013). Giordani et al. (2016) suggested that the nomenclature of the gene, TcoAT1, is in itself a misnomer, and that the gene should be renamed TcoNT10, as it is in fact orthologous and syntenic to the P1-type T. brucei NT10 transporter (Munday et al., 2013). Indeed, an exhaustive investigation of [3H]-diminazene uptake in T. congolense shows it to be not inhibited by substrates of P2-type transporters; instead it is accumulated through several transporters with only low-affinity for the drug, including the folate transporters (Carruthers et al., 2021). Most recently, TcoAT1/NT10 was expressed in a Leishmania mexicana cell line from which all nucleoside transporters had been deleted (Aldfer et al., 2022) and its interactions with substrates was characterised in detail (Ungogo et al., 2023), showing exactly the same binding patten previously determined for P1-type transporters in T. b. brucei (Al-Salabi et al., 2007; De Koning and Jarvis, 1999).
Following entry into trypanosomes, diamidines bind to the minor groove of double-stranded DNA via specific interaction with sites rich in adenine-thymine (A-T) base pairs in the kinetoplast and nuclear DNA (Brack et al., 1972; Brack and Delain, 1975). A fluorescent diamidine, furamidine (DB75) (Figure 2B), was shown to localise to the kinetoplast and nucleus (Mathis et al., 2006; Stewart et al., 2005; Carruthers et al., 2021). Binding of diamidines to the DNA minor groove occurs by attachment to specific sites via electrostatic and hydrogen-bond forces (Gresh and Pullman, 1984; Lane et al., 1991). This interaction leads to the inhibition of RNA primer synthesis and kDNA replication (Brack et al., 1972) and results in the complete and irreversible loss of trypanosomal kDNA (Riou and Benard, 1980). Similarly, pentamidine accumulates to very high levels in the mitochondrion of T. brucei (Damper and Patton, 1976a; Damper and Patton, 1976b) and induces the loss of the kinetoplast in T. b. brucei (Thomas et al., 2018). Moreover, diminazene also interferes with DNA replication by inhibiting the mitochondrial type II topoisomerase (Shapiro and Englund, 1990). Beyond trypanocidal activity, diminazene has also been shown to ameliorate immunosuppression and inflammation, two essential features in trypanosomiasis (Rurangirwa et al., 1979; Otesile and Tabel, 1987; Uzonna et al., 1999). Diminazene treatment resulted in an increase in plasma levels of parasite-specific IgG2a and IgG3 in T. congolense-infected mice (Uzonna et al., 1999). It has been shown that treatment of mice infected with T. congolense with diminazene significantly lowered the serum levels of several pro-inflammatory cytokines (IL-6, TNF, IL-12, and IFN-γ), reduced the secretion of IL-6 and TNF by Kupffer cells and reduced the expression of CD25 on lymphocytes and Foxp3+ in CD4+ T cells from the spleen (Kuriakose et al., 2012).
In T. brucei, T. evansi and T. equiperdum, the P2/AT1 aminopurine transporter is almost exclusively responsible for diminazene uptake. Sensitivity to diamidines in general and diminazene specifically is associated with its uptake into the trypanosome and subsequent accumulation in cellular targets (Barrett et al., 1995; De Koning et al., 2004; De Koning, 2020). The T. brucei TbAT1−/− knockout line, from which the P2/TbAT1 gene was deleted, showed a high level of resistance to diminazene and a lower level of resistance to pentamidine (Matovu et al., 2003). Likewise, the re-expression of the P2 transporter into multidrug resistant line B48 restored sensitivity to diminazene (Munday et al., 2013). In addition, B48 exhibited cross resistance to other diamidines, furamidine and CPD0801 (Ward et al., 2011). The combination of two or more specific amino acid mutations in this transporter, found in field isolates (Mäser et al., 1999; Matovu et al., 2001), can distort the binding site for diamidines, and results in significant levels of resistance (Munday et al., 2015b). The role of P2/TbAT1 transporter in diminazene resistance has been validated with several clinical isolates and laboratory-generated drug resistant lines (Nerima et al., 2007; Munday et al., 2015; Graf et al., 2016). In T. evansi, a knock-down of the TevAT1 gene by RNA-interference (RNAi) was shown to result in significant resistance to diminazene (Witola et al., 2004). In another study, a diminazene-resistant T. equiperdum strain demonstrated impaired diminazene uptake (Barrett et al., 1995) and loss of RNA expression for the TeqAT1 gene (Stewart et al., 2010). In the brucei group of trypanosomes (Trypanozoon subgenus) a shared mechanism of uptake was shown to be responsible for the cross-resistance between diminazene and other diamidines with melaminophenyl arsenicals (Matovu et al., 2003; Bridges et al., 2007; Baker et al., 2013), a fact corroborated by a lack of cross-resistance with unrelated compounds.
The effect of the deletion of the TbAT1/P2 gene in T. brucei on the sensitivity to pentamidine and melaminophenyl arsenicals was significant but resulted in a relatively low level of resistance, unlike the very substantial loss of sensitivity to diminazene and some other diamidines (Matovu et al., 2003; Ward et al., 2011). These low levels of resistance were insufficient to explain the extent of melarsoprol-pentamidine cross-resistance (MPXR) observed in some strains, confirming the existence of an additional, adenosine-insensitive high affinity pentamidine transporter (HAPT), which was demonstrated through a detailed analysis of the transport kinetics of [3H]-pentamidine (De Koning, 2001a; Bridges et al., 2007). Through a genome-wide RNAi screen, the aquaglyceroporin AQP2/AQP3 locus was identified in T. brucei to be the genetic determinant for MPXR (Baker et al., 2012). Further analyses identified the aquaglyceroporin 2 (AQP2) as the HAPT (Alghamdi et al., 2020; Munday et al., 2014). Expression of the TbAQP2 allele was found to fully restore sensitivity to pentamidine and melaminophenyl arsenicals in even the most resistant strains of T. brucei, and greatly increased the sensitivity of Leishmania mexicana promastigotes to both pentamidine and melarsoprol (Munday et al., 2014). Recent studies have described the structure activity relationship (SAR) of pentamidine for TbAQP2 and highlighted mutations that result in drug resistance (Alghamdi et al., 2020). It was also shown that rearrangements in the TbAQP locus lead to a chimeric TbAQP2/3 with impaired localisation and oligimerisation (Quintana et al., 2020), which is unable to mediate pentamidine or melarsen oxide uptake (Graf et al., 2015; Munday et al., 2014). The structure of TbAQP2, with pentamidine and with melarsoprol in the channel has now been elucidated by cryo-electron microscopy (Matusevicius et al., 2024).
In T. congolense, diminazene resistance is not related to its uptake into the cell as there was no significant difference in DA uptake or efflux between the DA-resistant T. congolense strain and the wild type (Carruthers et al., 2021). The mechanism of resistance to diminazene in T. congolense is instead associated with mitochondrial function as diminazene-resistant T. congolense strains all showed reduced mitochondrial membrane potential compared to the control. Genome analysis has shown missense mutations in a vacuolar-type Ca2+-ATPase in all the resistant lines sequenced. In addition, some of the resistant clones showed mutations in some of the T. congolense folate transporters. Functional expression of the T. congolense folate transporters (FT1–3) allowed for the conclusion that, although they partially contribute to low affinity diminazene uptake, they do not mediate resistance to the drug, at least in the in vitro-generated clones investigated in this study (Carruthers et al., 2021). The mechanism of resistance to diminazene in T. vivax is largely unclear, mainly because laboratory adapted strains and tools for genetic manipulation are still largely unavailable. Nevertheless, diminazene-resistant T. vivax isolates have been widely reported (Sonibare et al., 2016; Sow et al., 2012).
3.2. Phenanthridines
The phenanthridine compounds, isometamidium and homidium, are among the oldest trypanocides used in the treatment of animal trypanosomiasis. Yet, as related by James Williamson (1970) they derive from a series of compounds starting with a diaminoacridine developed by Paul Ehrlich, that he named trypaflavine (Figure 3) because of promising trypanocidal activity (Williamson, 1970), although its antibacterial properties were actually more impressive (Browning and Gilmour, 1913). Although it was soon noticed that the isomeric phenanthridine had superior trypanocidal properties than the acridine structure, it was not until 1938 that phenidium was shown to have highly promising activity against T. congolense (Browning et al., 1938) and field trials confirmed cure with a single injection of a small dose. The low therapeutic index however, led to the further development of dimidium, which was used extensively until approximately 1952, when mass-treatment led to resistance. Browning (1949) reports that the administration of over 2 million doses to African cattle had led to widespread resistance while increasing doses led to unacceptable toxicity. It was therefore replaced by the currently-still-in-use ethidium bromide (‘homidium’) which had a ten-fold improved therapeutic index, obtained by the simple change of methyl for ethyl as the quaternary ammonium substitution (Figure 3) (Watkins and Woolfe, 1952), which was further derived to isometamidium at the laboratories of the chemical company May and Baker (Wragg et al., 1958).
Isometamidium is an amphiphilic cationic phenanthridine compound formed by coupling of homidium chloride with m-amidinobenzenediazonium chloride, a part of the diminazene molecule (Berg, 1960). Commercially, isometamidium (Samorin ®) exists as an impure mixture of four compounds that appears as either di-cationic chloride salts or a dihydro form with a hydrogen addition to C-6, depending on solvent and pH (Figure 4) (Igoli et al., 2015). Earlier studies dismissed the other three components of isometamidium as impurities with inferior biological activity and advanced the compound 8-(3-m-amidinophenyl-2-triazeno)-3-amino-5-ethyl-6-phenylphenanthridinium chloride hydrochloride (M&B4180A; ‘isometamidium’) as the active component (Schad et al., 2008). However, a recent study has shown impure Samorin® to be significantly more efficacious than pure M&B4180A as well as all the other individual components in their pure form suggesting that the hitherto considered impurities are important as well, and may act synergistically in some way (Eze et al., 2019). In addition, all the individual components were found to be cross-resistant to Samorin® when their efficacy was determined against an isometamidium resistant T. brucei line (ISMR1) and compared to the wild type (Eze et al., 2019). Homidium (ethidium) was marketed as bromide and chloride salts (Ethidium® and Novidium®) (Watkins and Woolfe, 1952; Leach and Roberts, 1981). Isometamidium and homidium are both prophylactic and curative against African animal trypanosomes (Peregrine, 1994). However, isometamidium is preferred due to its better prophylactic activity and remarkable economic value while the use of homidium has dwindled because of widespread resistance in addition to food safety concerns linked to its carcinogenic features (Kinabo and Bogan, 1988).
The transport of isometamidium across trypanosome membrane through facilitated diffusion across its cytoplasmic membrane was hypothesised (Wilkes et al., 1997) but other studies concluded that the uptake was energy-dependent as it was sensitive to metabolic inhibitors including of Trypanosome Alternative Oxidase (TAO) (Sutherland et al., 1992a; Sutherland and Holmes, 1993). As discussed above, however, the kinetics of isometamidium accumulation are particularly complex and the inhibition of TAO and the resulting reduction in ATP levels could equally impact on the drug’s uptake into the mitochondrion, as this is driven by the mitochondrial membrane potential. Should that process be impeded, the drug will accumulate to a higher extent in the cytoplasm and could be exported from there through an active efflux system (Eze et al., 2016) or indeed through a facilitated diffusion carrier.
Within a kinetoplastid cell, phenanthridine compounds accumulate in the kinetoplast, and appear bound to kDNA (Ardelli and Woo, 2001; Boibessot et al., 2002), resulting in inhibition of the topoisomerase and consequently the linearization of the kDNA minicircle (Shapiro and Englund, 1990). Nevertheless, isometamidium was shown to be active against dyskinetoplastic trypanosomes as well, suggesting at least the presence of other targets outside the kinetoplast (Kaminsky et al., 1997). In contrast to the mitochondrial accumulation of isometamidium, homidium was observed to be distributed throughout the trypanosome cell (Boibessot et al., 2002). Yet, it kills trypanosomes by specific inhibition of replication initiation of kDNA minicircles through helix distortion in addition to the inhibition of nuclear DNA replication (Roy Chowdhury et al., 2010). Not unexpectedly, Eze et al. (2016) reported strong cross-resistance between isometamidium and ethidium bromide in T. brucei.
The role of the TbAT1/P2 transporter in the uptake of isometamidium was concluded to be likely inconsequential, despite its high affinity for isometamidium (De Koning, 2001b). One argument is that resistance to diminazene in T. b. brucei does not necessarily result in cross-resistance to isometamidium and that the diamidine resistant cell T. brucei B48 is not resistant to isometamidium (Eze et al., 2019). Similarly, RNAi knockdown of AT1 in T. evansi resulted in only minor cross-resistance with isometamidium in that species (Witola et al., 2004). Moreover, T. congolense, lacking an AT1 transporter (Munday et al., 2013), was more sensitive than T. brucei to isometamidium in vitro (Ungogo et al., 2022). However, isometamidium-resistant isolates of T. brucei exhibited mutations in the TbAT1 gene suggesting a possible link between the transporter and isometamidium resistance (Afework et al., 2006). Drug accumulation was found to be lower in isometamidium-resistant T. brucei than in the wild type (Eze et al., 2016), an observation that is consistent with the comparison between isometamidium-resistant T. congolense and the respective sensitive parental lines (Wilkes et al., 1997; Tihon et al., 2017). However, specific transporters involved in isometamidium cellular uptake and accumulation in mitochondrion are yet to be identified and characterised genetically. It is possible that mutations in a putative isometamidium transporter gene will cause decreased cellular drug uptake, reduced mitochondrial accumulation or both (Wilkes et al., 1997; Delespaux et al., 2005; Delespaux et al., 2008). Delespaux et al. have proposed four possible mechanisms of isometamidium resistance in trypanosomes; (a) reduction in entry of the drug into mitochondrion due to low mitochondrial membrane potential; (b) changes in a putative mitochondrial membrane transporter (c) increased drug extrusion through a cytoplasmic membrane transporter or (d) a combination of all these processes (Delespaux et al., 2008).
Using genome-scale RNA-interference (RNAi) screening in T. brucei, thirty potential candidate genes related to isometamidium resistance were identified, most of which form part of either a V-type H+-ATPase (V-ATPase); endoplasmic-reticulum membrane complex (EMC) and Adaptin-3 complexes (Baker et al., 2015). It appears that mutations involving the mitochondrial F1-ATPase y-subunit of the vacuolar ATPase or absence of transport proteins that facilitate interaction between both ATPases allow T. brucei to dispose of its kDNA, thereby conferring resistance to phenanthridines as well as cross-resistance to diamidines (Gould and Schnaufer, 2014; Eze et al., 2016). However, a different adaptation mechanism possibly allows naturally dyskinetoplastic T. evansi and T. equiperdum to maintain sensitivity to isometamidium (Kaminsky, Schmid and Lun, 1997; Birhanu et al., 2016).
Earlier studies have associated isometamidium resistance to mutations in the gene coding for the topoisomerase II enzyme, which is highly conserved between T. brucei and T. congolense. However, there was no correlation found between the gene’s sequence polymorphisms and isometamidium resistance phenotypes (Delespaux and De Koning, 2007; Delespaux, Geysen and Geerts, 2007). Using Amplified fragment-length polymorphism (AFLP) technology to compare isometamidium sensitive and resistant T. congolense isogenic strains, a conserved GAA codon insertion for an extra lysine was identified in a gene coding for an ATP-binding cassette (ABC) transporter in the resistant clone (Delespaux et al., 2005). However, some strains of T. congolense that were resistant to isometamidium in mouse tests did not contain the GAA insertion (Delespaux et al., 2005). Thus, it may be that resistance to this compound is multi-factorial, and that multiple resistance mechanism could be involved, each contributing solely or jointly to a partial loss of sensitivity (Delespaux et al., 2008).
More recent genomic analyses have identified several changes in read depth frequency of alleles at heterozygous loci in genes that code for ABC transporters and, in addition, a Drug/Metabolite Transporter (DMT) when isometamidium-sensitive and drug induced-resistant T. congolense lines were compared. Although no novel SNPs were detected, these mutations were found to be consistent in the naturally resistant isolates evaluated in the study (Tihon et al., 2017). However, no significant changes within the genes coding for the F1-subunit γ or other proteins associated with the mitochondrial F1Fo-ATP synthase were observed in that study (Tihon et al., 2017), although an earlier study associated isometamidium resistance with reduced mitochondrial membrane potential (Wilkes et al., 1997). This suggests that some peculiar and complex isometamidium resistance mechanisms are in play in T. congolense, different from T. b. brucei. It has proven to be impossible to induce isometamidium resistance in T. congolense in vitro (Ungogo and De Koning, unpublished) or even in an immune-competent mouse model (Tihon et al., 2017), despite the successful in vitro development of isometamidium resistance in T. brucei (Eze et al., 2016). Partly because the use of homidium salts is on the decline, the mechanism of action of trypanosomes to homidium is not actively studied, but the consistent cross-resistance observed suggests that it is likely to be similar to the more studied mechanism of isometamidium.
3.3. Suramin
The whole field of medicinal chemistry, which is concerned with the ‘identification, synthesis and development’ of chemical compounds for use as drugs, owes its development to efforts to find suitable agents against HAT in late 19th and early 20th centuries (Steverding, 2010). As the synthetic (textile) dye industry developed in Europe, chemists and scientists in the 19th century found inspiration in the specificity of dyes to particular materials and the interestingly selective toxicity of dyes to biological entities (Travis, 1991) – a principle widely used in histology. Motivated by the great interest of colonial administrations in trypanosomiasis epidemics affecting both humans and animals in the then African colonies, Paul Ehrlich began testing dyes for possible chemotherapeutic potential against trypanosomes. Ehrlich investigated more than 100 synthetic dyes for activity against T. b. brucei and T. equinum (Sneader, 2005). Nagana Red (Figure 5), a benzopurpurine compound was found to affect parasitaemia and increase the survivability of mice infected with T. b. brucei. The low solubility of Nagana Red was suggested to be responsible for the poor efficacy, and Ludwig Benda synthesised a derivative of Nagana Red, with extra sulphonic acid function named Trypan Red, which was found to possess both prophylactic and curative activity in mice infected with T. equinum. However, Trypan red was found to be ineffective against other trypanosome species and several of its derivatives tested did not show any promise (Steverding, 2010; Wainwright, 2010). Out of several benzopurpurine derivatives supplied by the German pharmaceutical company, Bayer, a blue compound named Trypan Blue was found to successfully eliminate parasitaemia in trypanosome-infected animals (Steverding, 2010; De Koning, 2020), but left unacceptable blue stains on their skin and was therefore discarded (Sneader, 2005). Efforts were thereafter focused on colourless compounds and a series of naphthalene urea derivatives amounting to more than 1000 molecules was synthesised and screened for over a decade by Bayer (Dressel, 1961). It was not until 1917, that a colourless naphthalene urea compound, Bayer 205 (renamed Germanin) was found to cure trypanosomiasis impressively in both animals and in humans (Dressel, 1961). The formula of Germanin was kept secret for potential German colonial advantage (Jacobi, 2010; Steverding and Troeberg, 2024) but was elucidated and published in 1924 by the French chemist Ernest Fourneau (Fourneau et al., 1924), who renamed the drug Fourneau 270. The compound was later renamed suramin (Ewins, 1944) and despite toxic side effects, has been at the forefront in the treatment of early-stage rhodesiense-HAT in humans and surra in camels and horses as well as T. b. brucei and T. equiperdum infections in equids for a century (Wiedemar et al., 2020).
Suramin, a negatively charged polysulfonated naphthyl urea (Figure 5) synthesised in 1916, is the oldest trypanocide still in use and has in fact been in use against sleeping sickness since 1922 (Dressel, 1961; Steverding, 2010; Giordani et al., 2016; Wiedemar et al., 2020). It is active against T. b. rhodesiense, T. b. gambiense, T. b. brucei, T. evansi and T. equiperdum (i.e. the Trypanozoon subgenus) and is still in use in the treatment of early-stage T. b. rhodesiense sleeping sickness in humans, surra in camels and horses as well as in dourine and T. b. brucei infections in equids (Giordani et al., 2016; Wiedemar et al., 2020). However, suramin is ineffective against T. congolense and T. vivax, which is why it is rarely used against animal trypanosomiasis in sub-Saharan Africa and in South America. Against this background, the recent report of T. vivax and T. evansi co-infections in camels in Iran (Asghari and Rassouli, 2022), the first such in Asia, is particularly problematic and need urgent confirmation.
The transport of suramin across the trypanosomal plasma membrane through passive diffusion or a membrane transporter seem unlikely due to its high molecular weight and six negative charges (Delespaux and De Koning, 2007; De Koning, 2020). An earlier study indicated that the suramin enters T. brucei through a non-saturable process (Fairlamb and Bowman, 1980). Subsequently, the possibility of specific expression of a ‘suramin-receptor’ in bloodstream forms of trypanosomes was suggested. Of specific importance, low-density lipoproteins (LDL) were proposed to be the carriers of suramin based on the relative abundance of the LDL receptors at the flagellum and flagellar pocket in trypanosomal membrane (Vansterkenburg et al., 1993; Coppens et al., 1987). However, Pal et al. (2002) have shown that suramin uptake into trypanosomes is not correlated to LDL (Pal et al., 2002). More recent evidence strongly suggests that suramin is transported via endocytosis following binding to the invariant surface glycoprotein ISG75 (Zoltner et al., 2016). A genome-wide loss-of-function study identified multiple genes that sensitize trypanosomes to suramin many of which have roles and/or locations at the endocytic pathway (Alsford et al., 2012). The most significant of the genes identified included Invariant Surface Glycoprotein ISG75, a putative transporter of the major facilitator superfamily (MFST) and the lysosomal proteins cathepsin L, CBP1 and GLP-1 (Alsford et al., 2012). The overexpression of ISG75 yielded an increased cellular accumulation of suramin, while, conversely, RNAi silencing of the glycoprotein resulted in reduced internalisation of the drug (Zoltner et al., 2020).
Endocytosis in trypanosomes is not only highly specialised and adapted to transport of particular substances into the cell, but greatly contributes in clearing immune effectors that bind to the cell surface as part of how African trypanosomes evade the host immune response (Zoltner et al., 2016). Trypanosomal endomembrane systems typically include a Golgi complex, early and recycling endosomes, late endosomes incorporating the ESCRT/multi-vesicular body system, a terminal endosome or lysosome, as well as other typical organelles. In addition, the endosomal surface is specialised with abundant GPI-anchored homodimers known as variant surface glycoproteins (VSG), instead of the conserved heterotetrameric (AP)-2 adaptin complex in mammals, as well as other surface proteins bearing diverse and trypanosome-specific transmembrane domains (Gadelha et al., 2015). Unlike in higher eukaryotes where multiple endocytic mechanisms are available, endocytosis in trypanosomes depends exclusively on clathrin, with several clathrin adaptor proteins present, containing epsin N-terminal homology (ENTH) and AP180 N-terminal homology (ANTH) domain phosphoinositide-binding proteins (Allen, 2003; Doherty and McMahon, 2009; Manna et al., 2015). The endocytic transport process varies significantly among life stages and species of trypanosome and this difference may be an important basis of suramin sensitivity and resistance (and possibly other drugs). In T. brucei, for example, the endocytic transport rate is many times higher in the mammalian bloodstream forms than in the insect forms, and this correlates with sensitivity to suramin, with the procyclic insect forms being much less sensitive to the drug (Zoltner et al., 2016).
A 6-step mode of action of suramin was proposed, involving; i) Binding to ISG75; ii) internalisation; iii) delivery to lysosome; iv) digestion of ISG75 by cathepsin L; v) suramin escape from the lysosome via a major facilitator super-family (MFS) transporter into the cytosol and vi) trypanosome killing by the now cytosolic suramin (Zoltner et al., 2016). It appears that there are multiple targets of suramin toxicity in the trypanosomal cytosol (De Koning, 2020; Wiedemar et al., 2020). Trypanocidal activity may be related to the polyanion’s inhibitory effect on a number of essential trypanosomal enzymes including dihydrofolate reductase, glycolytic enzymes, RNA-editing ligase-1 and thymidine kinase (Chello and Jaffe, 1972; Willson et al., 1993; Delespaux and De Koning, 2007; Zimmermann et al., 2016). Suramin was shown to block receptor-mediated uptake of low density lipoprotein (LDL), thereby possibly depriving trypanosomes of cholesterol and phospholipids (Vansterkenburg et al., 1993; Wiedemar et al., 2019). Treatment with suramin significantly reduced cytosolic ATP in T. brucei with a concomitant increase in mitochondrial ATP and provokes metabolic reprogramming with pyruvate accumulation and proline catabolism (Zoltner et al., 2020). Although the FoF1-ATPase activity is not different between untreated and suramin-treated cells, the mitochondrial membrane potential is significantly reduced in suramin-treated cells (Zoltner et al., 2020), although the drug most likely does not enter the mitochondrion, given its six negative charges. It is, however, possible that suramin inhibits an essential carrier, perhaps the ADP/ATP exchanger, which also recognises highly negative substrates. In a different study, exposure to suramin in T. brucei resulted in an increase in the number of trypanosomes with double nuclei, suggesting that the drug inhibits cytokinesis (Thomas et al., 2018). On balance of all the evidence, it appears that the trypanocidal activity of suramin, once free in the cell, is multifaceted and multifactorial but that it may be the rate of internalisation that, much like the diamidines and melaminophenyl arsenicals, determines the sensitivity to the drug, as shown with the knockdown/overexpression of ISG75, discussed above. The one observation arguing against this, is that T. congolense, although hardly sensitive to suramin, internalises the drug robustly over 10 minutes (Ungogo et al., 2022). It could still be argued that it is not the internalisation, as such, that matters, but the escape from the endocytic/lysosomal system, but expressing the T. b. brucei MFS transporter potentially implicated in that process (Alsford et al., 2012) in T. congolense did not alter that species’ sensitivity to suramin, or to pentamidine, diminazene or isometamidium (Ungogo et al., 2022). Yet more research remains necessary to untie this Gordian knot.
The mechanism of resistance to suramin is not yet fully elucidated. Suramin has high cure rates in the early stage of rhodesiense-HAT, even after 100 years of uninterrupted use, and resistance to the drug has not been reported in T. b. rhodesiense (Franco et al., 2018a). However, there are many reports of suramin resistance in the veterinary pathogen T. evansi, and development of resistant clones of both T. brucei and T. evansi in the laboratory through drug pressure appeared to be easy (Brun and Lun, 1994; Ungogo and De Koning, unpublished). Franco et al. (2018a) suggested that the discrepancy in suramin resistance might result from the different dose and dosage regimen employed in humans (five doses of 20 mg/kg) compared to animals (single dose of 10 mg/kg).
Drug sensitivity may be dependent upon entry into the trypanosome and blocking uptake of suramin, or its passage via the endocytic pathway, is sufficient to render parasites resistant to the drug (Giordani et al., 2016). Following the endocytosis model, it is possible for the drug to accumulate inside the cell since once internalized, the highly charged suramin molecule is highly unlikely to escape from the cell (Zoltner et al., 2016) unless it is a substrate of an ABC-type or other multi-drug resistance transporter. Overexpression of TbMRP-E did indeed resulted in two- to three-fold resistance to suramin in T. brucei, suggesting a possible role of MRPs in suramin resistance (Shahi et al., 2002). However, it has not been demonstrated that MRP-E is in fact overexpressed in any suramin-resistant strain. Nor has it been shown that ABC/MRP transporters are associated with resistance to any drug in any trypanosome strain, unless experimentally overexpressed (Bridges et al., 2007; Lüscher et al., 2006).
Surprisingly, quantitative transcriptomics by mRNA-Seq did not reveal any differential expression of genes encoding for the ISG75, the putative MFST or the lysosomal proteins in rapid laboratory-induced suramin resistant T. b rhodesiense (Wiedemar et al., 2018). However, all the selected resistant lines in the study had undergone an expression switch to a different variant surface glycoprotein (VSG) named VSGsur (Wiedemar et al., 2018). The transgenic expression of VSGSUR in sensitive cells resulted in 58-fold decrease in suramin sensitivity. In addition, VSGSUR-expressing cells exhibited reduced uptake of LDL and transferrin, shown to result from an impaired endocytosis. Interestingly, the knockdown of ISG75 in VSGSUR expressing cells led to reduced suramin sensitivity suggesting that ISG75 and VSGSUR affect suramin uptake through distinct pathways (Wiedemar et al., 2019). Through structural studies, suramin was shown to bind tightly to VSGSUR supporting a model that VSGSUR diverts trafficked suramin from the endocytic pathway thus effectively decreasing its intracellular concentration (Zeelen et al., 2021).
3.4. Melaminophenyl arsenicals
The use of arsenical compounds as drugs and poisons dates nearly 2500 years (Fairlamb and Horn, 2018). The association of arsenicals with African trypanosomiasis began in 1858 when David Livingstone improved the condition of a ‘fly-struck’ mare with nagana by treatment with arsenic; this was long before the causative agent of nagana was discovered but clearly the explorer believed the horse’s condition to be due to an infection transmitted by flies (Livingstone, 1858). Likewise, David Bruce reported treating an affected horse with arsenic in his seminal treatise in 1895 (Bruce, 1895). In 1902, inorganic arsenite As(III)O33- was demonstrated to be active against ‘nagana and surra’ in laboratory-infected rodents, although relapse ensued as soon as treatment was discontinued, and the treatment was highly toxic (Laveran and Mesnil, 1902). In 1904 Laveran followed this up with T. gambiense in rats and expressed confidence that arsenite could cure gambiense sleeping sickness although ‘in human Trypanosomiasis arsenical compounds have been often tried and have yielded only a passing amelioration, but in general the doses prescribed have been too feeble’ (Laveran, 1904). Over the following years, several promising organo-arsenical compounds were synthesised and tested against trypanosomiasis, most of which had to be disqualified due to serious neurotoxicity, such as the popular drug Atoxyl (Figure 6) (OIE, 2013). The story of Atoxyl, overstated to be 40 – 50-fold less toxic than any inorganic form of arsenic – a claim made by the very name of the drug – is told in detail by Walter Sneader (2005).
The next arsenical of importance was synthesised in 1915 by Jacobs and Heidelberger (Jacobs and Heidelberger, 1919) and initially known as compound A63 but soon thereafter known as tryparsamide (Pearce, 1921). As shown in Figure 6, it is simply atoxyl with an ethyl amide coupled to the amino group. Atoxyl and tryparsamide allowed for the first time the mass case finding and treatment of human trypanosomiasis, particularly in French colonial Africa, achieving a dramatic reduction in sleeping sickness where it was intensively applied. According to a retrospective study by Dukes (1984), ‘in Cameroon alone, some 1000 kg of atoxyl and 600 kg of tryparsamide were employed: a total of 460 kg of arsenic in a six-year period’. However, tryparsamide resistance became a serious problem by the early 1940s (Friedheim, 1949; Van Hoof et al., 1944).
It was therefore fortunate that around the same time a new arsenical trypanocide, melarsoprol (Arsobal, MelB), was produced by combining the heavy metal chelator and arsenic antidote, British anti-Lewisite (BAL, dimercaprol) with melarsen oxide (Friedheim, 1948; 1949), and that this compound was not cross-resistant with the earlier arsenicals (Friedheim, 1951; Williamson and Rollo, 1959). Since tryparsamidine was less active against the late stage of sleeping sickness and not at all against rhodesiense HAT, melarsoprol was also the first cure for late-stage East African sleeping sickness (Apted, 1970) and it has remained the only registered drug for this condition up to this date. However, today, 15 December 2023, it was announced that the European Medicines Agency’s (EMA) Committee for Medicinal Products for Human Use (CHMP) has ‘formed a positive scientific opinion’ of as the first oral drug against rhodesiense HAT was announced by the Drugs for Neglected Diseases initiative (DNDi) who conducted the trials (DNDi, 2023) and it is hoped that full registration will soon follow.
Since then, melarsoprol has been actively used in the treatment of late-stage HAT. Despite remarkable efficacy against T. evansi, T. equiperdum and T. b. brucei, melarsoprol is inconvenient and too expensive for veterinary use because of its multiple dose regimen and I.V. route of administration, which is due to its almost complete insolubility in water (it is administered as a 3.6% solution in propylene glycol). This necessitated the synthesis of a more soluble melaminophenyl arsenite, melarsomine (Cymelarsan, MelCy), by the conjugation of melarsen oxide and cysteamine, for use in animals (Touratier, 1992).
Melarsomine (cymelarsan), a melaminophenyl thioarsenite introduced in 1992 is the latest veterinary trypanocide in clinical use (Peregrine, 1994). It is indicated for the treatment of surra in camels, cattle and buffalo as well as for the treatment of heartworm (Dirofilaria immitis) in dogs (Desquesnes et al., 2013). Melarsomine is currently used in a wide range of domestic animals including cats (Tarello, 2005). There are reports indicating extra-label use of melarsomine to treat dourine, caused by T. equiperdum, despite WHO’s test-and-slaughter policy for the disease. However, in a recent study, melarsomine failed to clear T. equiperdum from the cerebrospinal fluid (CSF) of infected horses, which implies that the drug is unable to cross blood-brain barrier (BBB) in horses, or at least does not reach a therapeutically efficacious concentration in the brain at tolerated doses (Hébert et al., 2018). It may be that, by making melarsoprol more water-soluble, its valuable ability to cross the blood-brain barrier has been compromised. While it is also possible that melarsomine does not cross the BBB for lack of uptake transporters expressed by the BBB endothelial cells, an alternative explanation is that it is (also) rapidly extruded from the cerebro-spinal fluid by efflux transporters, which these cells express in large numbers. These transporters also exclude pentamidine from the brain (Sanderson et al., 2009; Sekhar et al., 2014).
With melarsomine rapidly disintegrating in aqueous medium (Berger and Fairlamb, 1994) and melarsoprol exhibiting a plasma half-life of less than an hour (Keiser, 2000), the mechanism of action of these melaminophenyl arsenicals is tied at least in part to a common active trypanocidal metabolite, melarsen oxide. This classifies both melarsomine and melarsoprol as prodrugs although both are also active themselves. Indeed, cross-resistance between melarsomine and melarsoprol has been reported in vivo and in vitro (Scott et al., 1997, 1996).
The uptake of melaminophenyl arsenicals into trypanosomes has been shown to be facilitated by the AT1/P2 transporter, which also transports diamidines (Carter and Fairlamb, 1993; De Koning et al., 2000), and aquaglyceroporin-2, a membrane transporter for water and glycerol (Baker et al., 2012; Munday et al., 2014). A similar AT1/P2 transport system is present in T. b. brucei, T. evansi and T. equiperdum (Berger et al., 1995; Witola et al., 2004; Stewart et al., 2010), but lacking in T. congolense and T. vivax (Munday et al., 2013). It has been suggested that the lipid-soluble melarsoprol might diffuse through the cell membrane. However, rapid metabolism of the drug into melarsen oxide in vitro would make this form of transport inconsequential (Scott et al., 1996). Once taken up by trypanosomes, melarsoprol/melarsen oxide forms an adduct with trypanothione that is referred to as Mel-T (Fairlamb et al., 1989) and that is implied to be toxic (Alsford et al., 2012). Cell cycle analysis using DNA configuration has shown that melarsoprol exposure inhibits mitosis, resulting in cells bearing replicated but unsegregated nuclei. In addition, the effect of the drug can be partially suppressed by putative mitogen-activated protein kinases (MAPKs) (Thomas et al., 2018). In another study, inhibition of DNA synthesis was observed in melarsoprol-treated T. brucei cells in addition to mitotic inhibition, and this can be alleviated by the overexpression of γ-glutamylcysteine synthetase (GSH1) (Larson et al., 2021), a key enzyme in the synthesis of the cellular anti-oxidant trypanothione.
The molecular mechanism of resistance to melaminophenyl arsenicals lies in their uptake transporters, AT1/P2 and AQP2. The inhibition of trypanocidal activity of melarsen oxide by other substrates of the TbAT1 transporter was demonstrated, and melarsen-resistant T. brucei appeared to have lost P2 adenosine transport activity (Carter and Fairlamb, 1993; Carter et al., 1999). This is corroborated by the detection of a mutated TbAT1 allele in a melarsoprol-resistant T. b. brucei laboratory strain (Mäser et al., 1999) and field isolates of T. b. gambiense and T. b. rhodesiense resistant to melarsoprol (Matovu et al., 2001; Nerima et al., 2007). After the withdrawal of melarsoprol for treatment from the Omugo site reported on by Matovu et al. (2001), the resistance allele was no longer detected in as many as 105 isolates from Omugo, whereas it was still present in isolates from the distant Moyo treatment site (both in Uganda) that still used melarsoprol (Kazibwe et al., 2009). Several mutations of these resistance alleles were systematically introduced in a wild-type TbAT1 allele and although the effect of any single point mutation was limited, the introduction of several of them or the deletion of codon F316, strongly impaired the transport of both adenosine and diminazene, and also resulted in high levels of resistance to cymelarsan (Munday et al., 2015b).
TbAQP2, a high affinity pentamidine transporter, also transports melaminophenyl arsenicals. Deletion of TbAQP2 confers significant resistance to pentamidine and melaminophenyl arsenicals, whereas re-expression in resistant lines restored sensitivity (Baker et al., 2012, 2013; Munday et al., 2014). Similarly, expression of TbAQP2 in Leishmania promastigotes highly sensitised these parasites to pentamidine and particularly to melaminophenyl arsenicals. In addition, specific mutagenesis or chimerisation of the TbAQP2 gene with neighbouring TbAQP3 results in significant resistance to pentamidine and melarsoprol (Quintana et al., 2020; Alghamdi et al., 2020). Furthermore, laboratory strains and clinical isolates resistant to melaminophenyl arsenicals showed mutations, deletions or chimerisation of TbAQP2 (Graf et al., 2013; Munday et al., 2015a; Pyana Pati et al., 2014). Beyond transporters, other potential mechanisms of resistance to melaminophenyl arsenicals in trypanosomes have been described. In T. brucei, engineered overexpression of ornithine decarboxylase and gamma-glutamyl-cysteine synthetase, two key enzymes in trypanothione biosynthesis, resulted in four-fold resistance to melarsoprol (Shahi et al., 2002). In addition, the overexpression of T. brucei multidrug-resistance associated protein A (TbMRPA), a putative thiol conjugate transporter, yielded an up to 10-fold increase in the EC50 of melarsoprol (Shahi et al., 2002; Alibu et al., 2006). However, it has never been shown that these mechanisms are involved in arsenical resistance in a clinical or veterinary context in the field.
3.5. Quinapyramine
Introduced in the early 1950’s, quinapyramine or Antrycide (Figure 7) is an aminoquinaldine derivative that is used as di-methosulphate salt for curative treatment, whereas its chloride salt (antrycide prosalt) is used for prophylaxis and therapy of animal trypanosomiases (Davey, 1950; Leach and Roberts, 1981). The use of quinapyramine in cattle in Africa became discouraged and was finally discontinued altogether in 1974 (Schillinger and Röttcher, 1986) due to the possible induction of cross-resistance to isometamidium, homidium and diminazene (Hawking, 1963; Peregrine et al., 1997). For instance, MacLennan (1968) reported that Antrycide was discontinued for use in Northern Nigeria as early as 1963 because of widespread resistance, after trypanosomiasis treatments in that region having averaged ‘about 400,000 per annum for the last 10 years’, and replaced with Berenil (diminazene aceturate) even though there was concern about resistance to that drug as well (Maclennan and Jones-Davies, 1967). Moreover, Whiteside (1962) showed that the higher the level of quinapyramine resistance was allowed to go, the more the cross-resistance, and to more drugs, became apparent.
The discontinuation of quinapyramine use for African cattle was very likely justified and saved to a large extent the diamidine and phenanthridine drugs for use up to this day, albeit still with significant resistance problems of their own (see above). Nevertheless, the complete cessation of quinapyramine production by May & Baker (Dagenham, UK) had the unintended consequence of further limiting the treatment options for surra to essentially only suramin, particularly in camels, this being before the introduction of melarsomine, and with suramin resistance in T. evansi being an increasing problem, this left many animals untreatable. Thus, quinapyramine was reintroduced in 1984 under the trade names Trypacide and Quintricyde, for both therapeutic and prophylactic use (Schillinger and Röttcher, 1986). However, the drug remains restricted to the treatment of surra in camel and horses, off-label treatment of dourine and the treatment of T. vivax infection in South America (Desquesnes et al., 2013; Dávila and Silva, 2006). Interestingly, however, two reports from the 1950s are clear that quinapyramine-resistant T. equiperdum tended to display increased sensitivity to suramin, rather than cross-resistance (Hawking, 1958; Ormerod, 1952). As importantly, suramin-resistant T. evansi remains fully sensitive to quinapyramine (Leach and Roberts, 1981; Luckins et al., 1979; Schillinger and Röttcher, 1986).
There is scant information about the molecular mechanisms of quinapyramine uptake and action. The investigation of the ‘mode of action of antrycide’ by Ormerod (1951a) revealed that the entry of the drug into the trypanosomal cell is slow and the drug’s target is neither on the surface membrane nor the nucleus (Ormerod, 1951b). In a study that compared the effect of treatment with different trypanocides in T. evansi, uptake of melarsoprol and stilbamidine was observed to be rapid and resulted in the trypanosome population to decline immediately and become fully cleared within 27 hours. In contrast, trypanosomes treated with suramin and quinapyramine continued to replicate for up to ~7 cell division cycles (72 hours), remaining constant in number, and only then started to die, suggesting a very slow uptake and/or action (Hawking and Sen, 1960). Considering that quinapyramine fluorescence was found to be associated with kinetoplasts of T. equiperdum 1 h after a high dosage of the drug was administered to a mouse followed by staining of cytoplasmic granules after 4 h (Ormerod, 1952), the latter would seem to be the more likely alternative, even if uptake may be slower than for diamidines and melaminophenyl arsenicals.
Thus, Hawking and Sen (1960) divided trypanocides into two classes; group 1 (trivalent arsenicals and diamidines) with rapid absorption and trypanosomal killing and group 2 (quinapyramine, suramin, homidium and other phenanthridines) characterised by slow uptake and trypanosomal killing. Interestingly, decades later, the uptake of melaminophenyl arsenicals and diamidines was found to mediated by specific membrane transporters, at least in the Trypanozoon subgenus (Munday et al., 2015a), while the transport of suramin is now known to be through endocytosis (Zoltner et al., 2016). Therefore, the slow uptake of quinapyramine may at least be partly mediated by means other than a regular membrane (nutrient) transporter or if by a nutrient transporter, it is transported with low efficiency and (probably) affinity, as appears to be the case for diminazene uptake by T. congolense (Carruthers et al., 2021). In a more recent study, the percentage of live cells in a culture treated with quinapyramine for 72 h remained above 50% (Rani et al., 2021). Quinapyramine has been reported to bind to ‘basophilic inclusion bodies’ or ‘granules of ribonucleoprotein’ that appear in the trypanosome after 24 h incubation with quinapyramine or dimidium (Ormerod, 1951a, 1951b; Newton, 1964). These granular bodies have been speculated to be ribosomes but in the micrographs presented the structures are much too big to be ribosomes. However, the inhibition of protein synthesis as the mechanism of action would be a potential explanation for the slow and mostly trypanostatic effects of quinapyramine. Treatment of Amoeba discoides with quinapyramine resulted in 50% loss of ribosomal RNA at 4 h and a complete loss after 17 h while an increase in the level of low molecular weight RNAs was observed, which suggests possible inhibition of ribosomal RNA synthesis or degradation of already formed rRNA (Summers and Hawkins, 1978).
Although resistance to quinapyramine is widely reported in laboratory-induced trypanosomes and clinical isolates, there is a paucity of information on the mechanism of quinapyramine resistance (Gill, 1971; Ndoutamia et al., 1993; Liao and Shen, 2010). However, it can be speculated, on the basis of the high level of cross-resistance between quinapyramine, diminazene, isometamidium and homidium (Ormerod, 1952; Ndoutamia et al., 1993), that quinapyramine resistance may be related at least in part to the TbAT1 transporter and/or the mitochondrial membrane potential (Munday et al., 2015a; Eze et al., 2016; Carruthers et al., 2021). In addition, the observed cross-resistance among quinapyramine, diamidines and phenanthridines in Leptomonas (Bacchi et al., 1975) suggests involvement of a target conserved in trypanosomatids, such as the kinetoplast located in the mitochondrion, where all these cationic trypanocides tend to accumulate. Based on investigations on the quinapyramine-resistant T. evansi strains in China, Liao and Shen (2010) reported the loss of the P2/TevAT1 gene (note: they call the gene TbTA1) in a quinapyramine-resistant isolate. While this correlation does not prove a functional link for P2 in quinapyramine accumulation, it adds to various other such hints and our own investigations show that quinapyramine is indeed an inhibitor of P2/TbAT1 with a moderate affinity (Ungogo and De Koning, manuscript in preparation).
4. Multidrug resistance and cross-resistance between trypanocides
Trypanosomes resistant to more than one drug have been isolated from many countries (Na’Isa, 1967; Mulugeta et al., 1997; Sinyangwe et al., 2004; Jamal et al., 2005). Multiple drug resistance can arise from cross-resistance between the drugs as a result of adaptations in a shared mechanism of action or uptake, or the co-existence of resistance to two or more drugs induced separately (Giordani et al., 2016). Considering the indiscriminate usage of multiple drugs, sometimes at sub-standard quality and rarely supervised by qualified veterinarians, data from the field on genuine cross-resistance (as opposed to treatment failure) is mostly unreliable. The development of drug resistance in trypanosomes through increased drug pressure has been demonstrated in vivo and in vitro (Carruthers et al., 2021; Eze et al., 2016; Osman et al., 1992; Tihon et al., 2017), and the characterisation of the resistant lines gives more reliable information on cross-resistance and the mechanism of resistance of each drug (Table 1).
Studies on drug resistant field isolates and laboratory induced resistant clones of T. b. brucei and T. evansi, have revealed cross resistance between diamidines and melaminophenyl arsenicals (Baker et al., 2013; Bridges et al., 2007; Osman et al., 1992; Ross and Barns, 1996). Likewise, laboratory induced melarsomine-resistant T. evansi have shown cross resistance to Berenil® (DA) and Arsobal® (melarsoprol) while remaining sensitive to suramin and antrycide (Ross and Barns, 1996). A shared mechanism of uptake through the TbAT1/P2 adenosine transporter was shown to be responsible for this cross-resistance. Through exhaustive studies of field isolates, laboratory induced resistant lines and generated mutants, the role of TbAT1/P2 gene in T. brucei in the resistance to diamidines, particularly diminazene, and melaminophenyl arsenicals has been elucidated (Matovu et al., 2003; Munday et al., 2015a, 2015b). These studies also led to the observations that suggest the existence of another, adenosine-insensitive transporter that helps to determine high-level melarsoprol-pentamidine cross-resistance (MPXR), initially dubbed the High Affinity Pentamidine Transporter (De Koning, 2001a), which was subsequently identified as the aquaglyceroporin TbAQP2 (Baker et al., 2012; Munday et al., 2014).
Many field studies have reported isolates of T. congolense, T. b. brucei and T. vivax that are resistant to both isometamidium and diminazene, the two major drugs currently used in the treatment of nagana (Anene et al., 2001; Geerts et al., 2001). The use of quinapyramine in cattle is discouraged due to a high incidence of cases of resistance and of cross-resistance to isometamidium, homidium and diminazene (Peregrine, 1994). Quinapyramine cross-resistance was demonstrated in experimentally induced quinapyramine-resistant T. congolense, which showed 5.5-fold resistance to diminazene, 6-fold resistance to isometamidium and 28-fold in resistance to homidium in infected mice (Ndoutamia et al., 1993). Similarly, a T. congolense clone that was adapted to 94-fold isometamidium resistance in mice exhibited multiple drug resistance to homidium chloride (33-fold), diminazene aceturate (3.4-fold) as well as quinapyramine sulphate (4.2-fold) (Peregrine et al., 1997). In vitro-induced isometamidium resistant T. brucei showed considerable cross-resistance to the chemically related phenanthridine ethidium bromide and to diminazene, as well as minor cross-resistance to pentamidine, likely mediated by the loss of kinetoplast and reduction of mitochondrial membrane potential (MMP) (Eze et al., 2016). However, similarly-generated T. congolense clones that were adapted to 6-7-fold resistance to diminazene showed negligible cross-resistance to isometamidium despite reduced MMP (Carruthers et al., 2021), showing that cross-resistance between diminazene and isometamidium in T. congolense is not inevitable and may, as with quinapyramine, depend on which drug the parasite was forced to adapt to, as well as on the species of trypanosome and the level of drug resistance. Giordani et al., (2016) suggested that since all these trypanocides share a mitochondrial target, and any single alteration affecting the MMP or the loss of organic cation carriers in the inner mitochondrial membrane, may give rise to cross-resistance to all the four trypanocides. Diamidine-phenanthridine cross-resistance has also been linked to mechanisms that support dyskinetoplasty, including mutations in F1-ATPase γ-subunit (Eze et al., 2016; Gould and Schnaufer, 2014), and the depletion of V-type H+-ATPases and endoplasmic-reticulum membrane complex (EMC) subunits (Baker et al., 2015). In addition, cross-resistance between diamidines, phenanthridines and possibly quinapyramine may be related to the P2/AT1 transporter in brucei group trypanosomes (Afework et al., 2006; De Koning, 2001b; Liao and Shen, 2010). However, given the rarity of dyskinetoplasty in T. congolense, and the lack of P2/AT1 orthologues in T. congolense and T. vivax, some multidrug resistance mechanisms in these species may be different from those in T. brucei.
Interestingly, cross resistance of suramin to other trypanocides used commonly to treat surra or HAT has not been reported (Wiedemar et al., 2018). This is possibly due the variety of intracellular targets the drug interacts with and its ability to bind to a VSG (Wiedemar et al., 2018, 2019, 2020; Zeelen et al., 2021) and to ISG75, leading to its internalisation through endocytosis (Zoltner et al., 2020, 2016). Clearly, the interactions and cellular distribution of suramin, with six negative charges, must be very different from the other trypanocides, which are all neutral (arsenicals) or cations (quinapyramine, diamidines, phenanthridines) at physiological pH. A recent study has shown cross resistance with Trypan-blue only, which is not very closely related chemically, but formed the backbone through which suramin was developed (Wiedemar et al., 2018). Interestingly, trypan blue is virtually the definition of a substance that is not taken up by cells and has been used for decades in the ‘trypan blue exclusion test’ to distinguish live from dead cells (Strober, 2015), but is readily taken up by trypanosomes (De Koning, unpublished observation) – presumably in the same way as suramin is.
5. Epidemiology of veterinary trypanocide resistance: incidence and spread in the field
Trypanocidal drug resistance is said to occur when the sensitivity of trypanosomes to a standard quality trypanocide is reduced or absent at the recommended dosage and route of administration (Assefa and Shibeshi, 2018). In highly endemic areas, however, treatment failure and relapse parasitaemia do not only arise from drug resistance as parasitaemia in the treated animal host may arise from a new infection, and not necessarily the initial one (Rowlands et al., 2001) or because of substandard drugs (see below) or faulty administration by unqualified practitioners, usually the farmers/herders themselves.
Using DNA markers to genotype T. congolense in Ethiopia following treatment with diminazene in cattle has shown a near-equal proportion of the parasite from new infection (40%) and actual relapse (37·5%) (Moti et al., 2015). Natural variation in drug sensitivity among Trypanosoma species and strains irrespective of previous drug exposure has been reported. For example, T. congolense has shown a higher rate of relapse infections compared to T. vivax following treatment with homidium. In contrast, T. vivax strains from West Africa have exhibited a higher level of resistance to diminazene than T. congolense (Leach and Roberts, 1981). Acquired resistance to trypanocides is usually developed by trypanosomes following drug underdosing arising from a variety of circumstances (Leach and Roberts, 1981). Once resistance is developed to a trypanocide, there is a chance that the resistant parasite strain would show decreased sensitivity to other drugs with a shared mechanism of uptake or action (Assefa and Shibeshi, 2018; De Koning, 2020).
Resistance of animal trypanosomes to diminazene and isometamidium was reported as early as the 1960s in Nigeria (Jones-Davies and Folkers, 1966; Na’Isa, 1967), barely a decade after their introduction. Trypanocide resistance is currently reported in 21 out of the 37 countries in the tsetse belt of sub-Saharan Africa, 10 countries of which have also documented cases of multiple drug resistance (Assefa and Shibeshi, 2018). It is highly probable that there is trypanocidal resistance in many other African countries but that is yet to be documented, largely because no studies on the local situation have been carried out or published (Melaku and Birasa, 2013). Nevertheless, some caution must be applied in interpreting the literature. The available data is patchy both geographically and in time and, although many reports on trypanosomiasis treatment relate treatment failure or resistance, it must be remembered that the study area may well have been selected because of local treatment failure, thereby self-selecting to find just that. Moreover, findings of drug resistance are more likely to be written, and accepted, for publication than a similar survey finding no problems. Therefore, the nature of the incidental reporting from small areas might paint an overly pessimistic picture with regards to drug resistance.
Overreliance on the same old treatments for decades, often inexpertly, results in drug resistance and treatment failure with all the attendant consequences (Delespaux and De Koning, 2007), just as happened with the early trypanocides, as documented above. Prevalence of trypanocide resistance is highly variable, and can be up to 100% as reported in T. congolense isolates from dogs in Nigeria (Obi et al., 2022). However, a recent meta-analysis of 30 studies reported an overall prevalence of 29% for drug resistance in African animal trypanosomes (Okello et al., 2022b). A widespread incidence of trypanocide resistance was reported in the cotton belt of west Africa between eastern Guinea across southern Mali to the southwest of Burkina Faso, possibly linked to a high use of trypanocides (Clausen et al., 2010; Grace et al., 2008). In the south-central African state of Zambia, drug resistance may have resulted from extensive use of isometamidium treatments as part of a trypanosomiasis eradication program (Chitanga et al., 2011; Sinyangwe et al., 2004). There are also several reports of drug resistance in T. evansi from several parts of the world (Brun et al., 1998).
The bulk of the available information on drug resistance emanates from local case reports and the baseline information on the exact prevalence of drug resistance is unavailable (Mamoudou et al., 2008). Delespaux et al. (2008) attributed this dearth of baseline information to the fact that the two widely used in vivo methods for the detection of drug resistance – the laboratory and field methods – are not only expensive but laborious (Eisler et al., 2001). The alternative in vitro tests are also challenging, as the Glossina infectivity test (DIGIT) is greatly dependent on the availability of tsetse flies while the drug incubation infectivity test (DIIT) still requires infecting mice after drug incubation (Kaminsky et al., 1990; Knoppe et al., 2006). With advances in molecular biology, biomarker genes of drug resistance in T. brucei species, such as TbAT1 and TbAQP2, were discovered and validated, and have been applied as biomarkers in the more time-efficient molecular methods. However, in T. congolense, these markers are not present and other proposed drug resistance genes were found to be unreliable. The T. congolense topoisomerase II enzyme and an ATP-binding cassette (ABC) transporter were found not to be definitive biomarkers of isometamidium resistance (Delespaux and De Koning, 2007; Delespaux et al., 2007; Delespaux et al., 2005, 2008). In addition, the claim that T. congolense TcoAT1/NT10 was a diminazene resistance marker was proven to be erroneous (Munday et al., 2013; Ungogo et al., 2023). There are currently no markers at all (proven or proposed) for molecular detection of drug resistance for T. vivax. It should be noted that the discovery of a few validated trypanocide resistance markers has not, to date, translated into the development of rapid diagnostic tests. Unfortunately, in the absence of any much-needed on-the-field trypanocidal sensitivity tests, rational chemotherapy relies upon the characterisation and prevalence of drug resistance in trypanosome isolates from a particular area, which requires laborious studies and long periods to establish (Peregrine and Mamman, 1993). It goes without saying that for decades there has been a need for a rapid and reliable test based on molecular or phenotypic biomarkers capable of detecting drug sensitivity and/or resistance in trypanosomes to inform the choice of drugs by veterinarians and clinicians alike.

Table 2.
Incidence of trypanosomal drug resistance reported by country.
| Country | Animal host | Trypanosoma spp | Drug | Diagnostic | Reference |
|---|---|---|---|---|---|
| Nigeria | Cattle | T. congolense | Homidium chloride | Test in cattle | Jones-Davies and Folkers, 1966 |
| Cattle | T. congolense | DA, ISM | Test in cattle | Na’Isa, 1967 | |
| Cattle | T. congolense | Homidium chloride | Test in mice | Joshua, 1988 | |
| Cattle | T. b brucei | Homidium chloride, DA | Test in mice | Joshua, 1988 | |
| Dog | T. b brucei | DA, pentamidine | Test in rats | Anene et al., 2006 | |
| Cattle | T. vivax | Homidium chloride | Test in sheep | Ogbaje et al., 2015 | |
| Cattle | T. vivax | ISM, DA, Homidium chloride | Test in sheep | Ogbaje et al., 2015 | |
| Dog | T. congolense, T. b brucei | DA, ISM | Test in mice | Obi et al., 2022 | |
| Ivory Coast | Cattle | T. congolense, T. vivax | Trypamidium, Homidium | (Küpper and Wolters, 1983) | |
| Burkina Faso/ Upper Volta | Cattle | T. congolense | DA, ISM | Test in mice | Authie et al., 1984 |
| T. congolense | ISM | Test in mice | Pinder and Authie, 1984 | ||
| Cattle | T. congolense | DA, ISM, Homidium | Test in goats and cattle | Clausen et al., 1992 | |
| Cattle | T. vivax | DA, ISM | Test in cattle | Sow et al., 2012 | |
| Cattle | T. congolense | ISM | DIIT, Standard mouse test | Knoppe et al., 2006 | |
| Senegal | Cattle | T. vivax | DA, Ethidium | Test in goat | Diaité et al., 1997 |
| Somalia | cattle | T. congolense | DA, ISM, | Test in mice and cattle | Ainanshe et al., 1992 |
| T. vivax | ISM, homidium and quinapyramine | Schönefeld et al., 1987 | |||
| Kenya | T. vivax | ISM, homidium and quinapyramine | Schönefeld et al., 1987 | ||
| Camel | T. evansi | ISM, homidium and quinapyramine | Single dose RCT in mice | Mdachi et al., 2023 | |
| Camel | T. evansi | Quinapyramine prosalt, melarsomine, ISM | Test in mice | Waitumbi et al., 1994 | |
| Cattle | T. congolense | DA, Homidium | Test in mice | Okello et al., 2022a | |
| Tanzania | Cattle | T. congolense | DA | Test in mice and cattle | Mbwambo, Mella and Lekaki, 1988 |
| Uganda | Cattle | T. brucei, T. vivax | Homidium | Test in mice, goats cattle | Olila et al., 2002 |
| Zambia | Cattle | T. brucei, T. congolense, T. vivax | ISM, DA | Test in mice and cattle | Sinyangwe et al., 2004 |
| Sudan | Cattle | T. brucei, T. congolense, T. vivax | Homidium bromide, ISM, DA | Test in goats | Mohamed-Ahmed et al., 1992 |
| Cattle | T. brucei, T. congolense, T. vivax | Homidium bromide | Test in sheep, goats and cattle | Abdel Gadir et al., 1981 | |
| Camel | T. evansi | Suramin, QPR | hypoxanthine incorporation test, in vivo drug sensitivity test, mouse | El Rayah et al., 1999 | |
| Ethiopia | Cattle | T. congolense | DA, ISM, Homidium | Block treatment, mouse test, BCT | Mulugeta et al., 1997 |
| cattle | T. congolense | DA, ISM | Block treatment, single dose mouse test | Afewerk et al., 2000 | |
| Donkeys | T. congolense | ISM | BCT, mice | Assefa and Abebe, 2001 | |
| Cattle | T. congolense | DA, ISM | In vivo test in mice and cattle | Chaka and Abebe, 2003 | |
| Cattle, goats | T. vivax | DA, ISM | Block treatment (goats) | W/yohannes et al., 2010 | |
| Cattle | T. vivax | DA, ISM | Block treatment (cattle) | Dagnachew et al., 2015 | |
| Cattle | T.congolense, T. vivax | DA, ISM | Block treatment (cattle) | Dagnachew et al., 2017 | |
| Cattle | T.congolense, T. brucei, T. vivax | DA, ISM | Block treatment (cattle) | Degneh et al., 2019 | |
| Mozambique | cattle | T. congolense | DA, ISM, Homidium | Multi-dose mouse test, BCT | Jamal et al., 2005 |
| Cattle | T. congolense | DA, ISM | Block treatment in cattle | Mulandane et al., 2018 | |
| Cameroun | cattle | T. congolense, T. b. brucei | DA, ISM | PCR-RFLP, AS-PCR, single-dose mouse test | Mamoudou et al., 2008 |
| Zambia, Zimbabwe, South Africa | Buffalo, tsetse fly | DA | PCR, MC, Mouse test | Chitanga et al., 2011 | |
| Mali | Cattle | T. congolense, T, vivax | DA, ISM | MIC | Mungube et al., 2012 |
| Togo | Togo | T. vivax | DA, ISM | Test in goat | Boma et al., 2022 |
| French Guyana | Cattle | T. vivax | DIM | Test in cattle and sheep | Desquesnes et al., 1995 |
| China | Buffalo, horse, mule, camel | T. evansi | Suramin and Quinapyramine prosalt | Growth inhibition test, single-dose mouse test | Zhou et al., 2004 |
| Buffaloes, horses, camels, mules | T. evansi | Quinapyramine prosalt | Liao and Shen, 2010 | ||
| Philippines | Water buffaloes | T. evansi | Likely P2/AT1-related drugs | PCR showed P2/AT1 deletion | Mingala et al., 2019 |
RCT, randomised controlled trial; DA, diminazene aceturate; ISM, isometamidium chloride; two salts of Homidium are reported, Homidium chloride (Novidium); and Homidium bromide (Ethidium); Quinapyramine prosalt is reported as Antrycide in some studies.
6. Strategies to combat the challenge of trypanocide resistance
6.1. Tackling substandard trypanocides
There is a persisting problem of poor-quality drugs in African markets and a variety of trypanocides in circulation with reduced or even no trypanocidal activity were reported. An investigation financed by the Ministry of Animal Resources of Burkina Faso in collaboration with FAO revealed that about half of the trypanocides on the local markets in the country did not meet the standard quality requirements (Sow et al., 2012). Similarly, in another study, 40% of sampled trypanocides from Togolese markets were found to contain lower amounts of active ingredients than specified, among other deviations from quality reference standards based on HPLC (Tchamdja et al., 2016). Furthermore, a recent larger study comprising of 6 African counties (Togo, Burkina Faso, Benin, Mali, Côte d'Ivoire, and Niger) showed that 51.90% of the 308 trypanocides sampled did not comply to standards and were containing a lower quantity of the active ingredients than stated in the packaging (Bengaly et al, 2018). The poor quality of trypanocides (and other marketed medications) can be linked to the large size of the unofficial market. For example, most of the substandard trypanocides (53.57%) detected in one study were from unofficial markets, and only 25% were from the official markets (Tchamdja et al., 2016). According to reports from the Burkinabè Ministry for Animal Resources (MRA), out of the over 2.8 million doses of trypanocides used annually in Burkina Faso, only 23% are officially imported (Sow et al., 2012). The International Federation for Animal Health (IFAH) has estimated that the annual trade in unregistered and substandard veterinary drugs in Africa is as large as the official drug market and is worth $400 million (FAO, 2012). This kind of market value creates a lot of stakeholders that are heavily invested in the continued local production and distribution of these drugs, without much regard for quality and safety protocols; and the generic product is generally cheaper than the imported drugs.
The use of sub-standard drugs often results in an unintended under-dosing, which in turn leads to treatment failure and the development of drug resistance; unintentional overdosing, where too much active ingredient was included, has also been anecdotally reported. Hence, measures need to be taken to ensure that only trypanocides that meet quality standards find their way to the markets. Taking Nigeria as a case, Kingsley (2015) reported on the intricate processes through which the veterinary drug market operates and how these make it possible for substandard drugs and their attendant consequences to thrive despite existing regulations.
It is very clear that the impact of issuing regulations by a central government, though essential, is nonetheless limited. The structural neglect of the livestock sector and pastoralist communities has bred a disconnect between the regulators and cattle owners. Thus, there is a need for a more holistic understanding of, and engagement with, the key stake holders beyond the regulate-and-enforce approach (Kingsley, 2015). Furthermore, lack of an effective regional strategy and collaboration between national governments may contribute to the reasons that substandard drugs are routinely traded across African borders and misused by unqualified people. Therefore, there is need for a regional approach to the quality control, importation, supply and use of veterinary medicines (Bengaly et al., 2018). Without improvements in this area, any new trypanocide will be at risk once again of rapid resistance because of misuse. Unfortunately, it will need to be done at both the governmental level and at a local level, which means at thousands of local markets and farm shops throughout Africa.
6.2. Rational drug use
In the face of limited coverage and affordability of licensed veterinary services in Sub-Saharan Africa, farmers, herders and other unqualified people administer treatments to sick animals, often illegally (Grace et al., 2008; Kasozi et al., 2022). A survey on trypanosomosis management reported that up to 50% of farmers treated their animals themselves, without veterinary support (Liebenehm et al., 2016). Likewise, another survey, engaging salespersons/owners of veterinary drug outlets in two rural areas in Nigeria, revealed that 66.7% of respondents had no form of professional training in animal health. In most of the shops studied, suitable storage facilities such as refrigerators and standard procedures such as record keeping for transactions and expiration date of drugs were lacking (Elelu, 2017) and at unofficial rural markets this is not likely to be any better. The use of trypanocides by unqualified people has been reported to come with inappropriate usage and incorrect dosage (Delespaux et al., 2002; Machila et al., 2003; Van den Bossche et al., 2000), thus aiding the development of drug resistance.
Rational drug use (RDU) is identified as the best-bet strategy to control drug resistance against trypanosomiasis, especially in areas where resistance is not yet well established (Clausen et al., 2010). RDU involves using the appropriate drug at the appropriate dose and administering it correctly, and only when clinically needed (Clausen et al., 2010). Reduction of the need and use of drugs can be achieved through integrating chemotherapy with other disease prevention strategies such as vector control (Holmes, 1997). A recent study has shown that in an integrated trypanosomiasis control strategy, intervention herds with rational drug usage had reduced risk of trypanosomiasis in villages of Northern Togo where drug resistance had been reported. Furthermore, trypanocidal treatments per animal per year were found to be much lower in the intervention herds compared to control herds (0.3 vs 5 for DA and 0.8 vs 2 for ISM). However, it is difficult to conclude that rational drug usage alone was responsible for the success in this study, due to other measures also taken that might have confounding effects (Tchamdja et al., 2019).
In west Africa, for example, a study has shown that the understanding of the problem of resistance was low among both policymakers and stakeholders while an enabling environment for RDU is lacking (Clausen et al., 2010). Achieving RDU entails engagement, training and collaboration between policy makers and the stakeholders handling trypanocides including animal health workers and farmers. In order to solve the challenge of an inadequate animal health workforce, there were many instances where farmers were successfully trained to become community animal health workers so they can make correct diagnoses and administer drugs (Catley and Leyland, 2001). In a cluster-randomised controlled trial that involved training farmers on rational drug-use in Mali, 84% farmer treatment success was reported in the intervention group, compared to 73% in the control group. In addition, farmers who received RDU training were reported to give higher dosages of isometamidium chloride 5 months post-training, an encouraging finding considering the role of under-dosing in the development of drug resistance (Grace et al., 2008). However, even after training, local farmers may struggle to work to RDU standards when lacking basic equipment such as scales that are needed so the correct dose in g/kg body weight can be calculated.
7. Optimising use of the existing trypanocides
With slim chances of new veterinary trypanocides in the near future, optimising the performance of the current drugs must be relied upon to increase treatment successes.
7.1. Adjustment of dosage and dosage regimen
With the growing reports of treatment failure and drug resistant trypanosomes, adjustment of dosage and dosage regimen of the existing trypanosomes becomes paramount. Ijomanta et al. (2016) compared the efficacy of 3 graded doses of DA in combination with fixed dose of levamisole in mice experimentally infected with T. b. brucei. While relapse parasitaemia was observed in the group treated with 7 mg/Kg DA at day 30 post-treatment, there was no relapse in groups treated with 14 mg/Kg or 21 mg/Kg, although higher mortality was recorded in the 21 mg/Kg DA group – evidence of a narrow therapeutic index. Therefore, since the same dose of levamisole was used and treatment with levamisole alone had no effect on parasitaemia, it can be concluded that increased doses of DA provided prolonged protection (Ijomanta et al., 2016). In the treatment of Surra, increasing the dosage of melarsomine (from 0.25 mg/kg to 0.5 mg/kg bw) and DA (from 3.5 mg/kg to 7 mg/kg) has been recommended in order to achieve a fully curative (sterilising) effect (Desquesnes et al., 2013). However, administration of increased doses of diminazene (14 mg/Kg and 21 mg/Kg bw) in pregnant rats resulted in adverse reproductive effects; a significant increase in number of resorbed foetuses, significant reduction in the post-implantation survival index of embryos, increased gestation length and a significant decrease in the litter weights (Oguejiofor et al., 2010). Thus, although increasing the doses of trypanocides might overcome low-level drug resistance, it should be done with caution due to possibility of increased toxicity, as the therapeutic index of these old trypanocides is regrettably very low.
Another approach for optimising effectiveness of trypanocides in the face of resistance challenges is a multiple dosing regimen. There is a striking difference in cure rates of rhodesiense-HAT and surra treated with suramin. This might arise from the difference in dosage regimens used; five doses of 20 mg/kg in humans compared to a single dose of 10 mg/kg for animals (Franco et al., 2018a). Treatment of rats infected with T. b. brucei at a single dose of 7 mg/Kg DA were compared with multiple dosing at weekly intervals (repeated once, twice or thrice). While relapse parasitaemia was observed in rats treated once, twice and thrice, the four-times treatment appeared to protect against relapse and the serum levels of liver enzymes suggest that the multiple therapy was safe (Ezeh et al., 2016), the spacing of the dosages being informed by the clearance time of diminazene, which has a fairly long elimination half life time, of ~100 h, although not long enough for prolonged prophylactictic activity (Peregrine and Mamman, 1993). Successful treatment of three cats diagnosed with T. evansi infection with a 4-day course of melarsomine has been reported, with one of the cats testing negative of the parasite during a 1-month post-treatment follow-up (Tarello, 2005). However, multiple-dose administration regimens may prove difficult in livestock veterinary practice in developing countries (Giordani et al., 2016).
7.2. Enhancement of drug delivery
The challenges surrounding trypanocidal chemotherapies have highlighted the use of drug delivery strategies (DDS) with the aim of improving drug absorption, distribution, metabolism and elimination (ADME) (Kroubi et al., 2011). DDS could potentially increase the efficacy and safety of trypanocides, and possibly reduce the risk of development of resistance, or even bypass it. DDS strategies for optimising the current trypanocides are under development and include formulations for slow release, for improved solubility, and for improved targeting and uptake into trypanosomes.
Many slow-release preparations of trypanocides have been developed to reduce toxicity and extend their prophylactic activity (Peregrine, 1994). For example, prophylactic and therapeutic use of 1 mg of diminazene encapsulated in stearylamine-bearing liposomes resulted in a significant increase in the survival of mice infected with T. evansi compared with 1 mg of conventional diminazene (Yongsheng et al., 1996). Another liposome-encapsulated diminazene formulation showed increased in vitro activity against T. evansi compared to conventional DA, although in vivo studies in rats did not show superiority of the liposome-based preparation, likely due to the low dosage used (Oliveira et al., 2014). However, trypanocide formulations with prolonged prophylactic activity are likely to facilitate the development of drug resistance (De Deken et al., 1989; Peregrine, 1994).
With advancements in nanoscience, there is great potential for developing nanoparticulate formulations of existing trypanocidal drugs to optimise their efficacy and safety in a cost-effective manner (Kroubi et al., 2011). Loading diminazene in 70DGNP+, which is made of porous cationic nanoparticles (NP+) with a lipid core, resulted in faster killing of T. b. brucei in vitro (Kroubi et al., 2010). Similarly, coupling pentamidine to a drug nanocarrier that was covered by a single-domain antibody fragment which specifically targets a cryptic epitope within the densely packed variant surface glycoprotein (VSG) surface coat resulted in an approximately sevenfold and tenfold increase in sensitivity in vitro and in vivo respectively, compared to the free drug. The nanoparticle was designed to move to the flagellar pocket and become internalized with continuous VSG turnover, thus by-passing the transporter-mediated uptake (Arias et al., 2015). An improved nanoparticle formulation of pentamidine, pentamidine-loaded functionalized PEGylated-chitosan nanoparticles coated with NbAn33 (NbAn33-pentamidine-chNPs) bypassed drug resistance in a pentamidine-resistant AQP2-mutated T. brucei strain, decreasing the IC5O by 11.6-fold in vitro compared to the free pentamidine (Unciti-Broceta et al., 2015). In addition, treatment with four daily doses of 2.5 mg/kg of NbAn33-pentamidine-chNPs in immunosuppressed mice infected with T. brucei resulted in 60% survival, while all the mice treated with a similar dosage of free pentamidine died after infection (Unciti-Broceta et al., 2015). Thus, nanoparticle-based formulations constitute a promising approach for avoiding drug resistance associated with mutations of surface transporters by completely bypassing the transporters (Garcia-Salcedo et al., 2016; Stijlemans et al., 2017). Another promising delivery approach that bypasses the membrane transporters is involves increasing the solubility of trypanocides by formation of water-soluble inclusion complexes. Coupling melarsoprol in two different cyclodextrin inclusion complexes with β-cyclodextrin and randomly methylated-β-cyclodextrin (Rodgers et al., 2011) allowed for the oral administration of the mainly IV-administered drug to achieve cure in murine model of late stage rhodesiense-HAT (Rodgers et al., 2011). Garcia-Salcedo et al. (2016) suggested that the uptake of melarsoprol cyclodextrin inclusion complexes into trypanosomes is likely via endocytosis, and not the AQP2 channel for free melarsoprol, thus the complexes could possibly evade melarsoprol resistance (Garcia-Salcedo et al., 2016), exactly as demonstrated for the pentamidine nanoformulation. Considering what we now know of TbAQP2 as a drug carrier and its substrate limitations (Alghamdi et al., 2020; Matusevicius et al., 2024), this seems highly likely.
7.3. Sanative pair
Alternate use of a pair of unrelated trypanocides with low risk of cross-resistance, known as a sanative pair, has been proposed to control drug resistance in AT. In this context, Whiteside et al. (1962) used the concept "sanative pair" to refer to 2 drugs each capable of curing infection resistant to the other and popularised the use of a sanative pair of diminazene and homidium in the treatment of nagana. An infected animal is first treated with one of the pair, and when relapse parasitaemia is observed, the animal is then treated again with the other drug in the pair (Leach and Roberts, 1981). The use of 'sanative' drug combinations has been employed to prevent the development of drug resistance (Peregrine, 1994) or, equally, to prevent resistant strains to gain a foothold and spread. One modification of this strategy involved rotating the use of homidium for 6 months and diminazene for another 6 months, and this formed the trypanosomiasis drug policy of the now-defunct Northern Region of Nigeria (Eghianruwa and Oridupa, 2018). As the use of Homidium becomes rarer, a sanative pair of diminazene and isometamidium is currently the one most frequently used in the treatment of nagana.
7.4. Combination therapy
The use of combination therapy has been shown to improve therapeutic success and/or reduce toxicity in some treatment regimens. Combination therapy can be used to optimise the value of an existing drug and to potentially control the problem of resistance, in a cheaper and time-saving manner compared to the development of a new drug (Cheesman et al., 2017). The potential of the synergistic use of two trypanocides to provide better cure rates has been investigated. The nifurtimox-eflornithine combination therapy (NECT) has proved to be a successful front-line treatment for second stage-gambiense-HAT that is easy to administer and potentially protective against the development of resistance (Priotto et al., 2009). This combination was chosen over another combination that was being trialled at the same time: melarsoprol and nifurtimox. Although that combination was also more effective than the standard monotherapies, fatal encephalopathic syndromes did still occur in the group receiving the combination treatment (Bisser et al., 2007).
The act of suramin being highly anionic was utilised to form suraminate complexes with the cationic trypanocides diminazene, isometamidium chloride, homidium and quinapyramine, with some of these complexes found to prolong prophylactic activity of the trypanocides, probably because they form poorly-soluble deposits that are only slowly released (Williamson and Desowitz, 1956; Williamson, 1957). While the use of homidium suraminate in cattle produced localised toxicity (Stephen, 1958), quinapyramine suraminate successfully cleared parasitaemia in pigs experimentally infected with T. simiae with no apparent toxicity at the injection site (Stephen, 1962).
In order to investigate synergy between trypanocides and the possible superiority of combination therapy, Williamson et al. (1982) tested 27 different drug pairs from 12 trypanocides in vitro and in vivo. Out of these combinations, suramin and tryparsamide, suramin and puromycin, and suramin and diminazene demonstrated statistically significant synergy. In addition, 6 other combinations including suramin and ISM, ISM and homidium, and ISM and quinapyramine showed increased cure rates in mice infected with T. b. rhodesiense (Williamson et al., 1982). Another strong advocate of combination therapy, especially for cerebral trypanosomiasis, was F. W. Jennings, who tested a substantial number of combinations in mouse models, and achieved highly promising results, but in the end lamented – with some justification – how difficult it was to progress from the mouse to human trials (Jennings, 1993, 1990).
In the treatment of T. evansi infection in horses and dogs, a combination of diminazene or melarsomine and quinapyramine has been recommended since the goal would be to completely clear the potentially fatal infection (Desquesnes et al., 2013). Recently, the combination of diminazene and quinapyramine sulphate was used to successfully treat dourine in a horse, with no relapse for 2.5 years (Davkharbayar et al., 2020).
Using mathematical modelling, Wangwe et al. (2019) have demonstrated that using a combination of 2 trypanocides achieved better results and lower levels of trypanocide resistance compared to using a single drug in the treatment of African animal trypanosomiasis. In addition, combination treatment in up to 80 – 100% of the cattle in a herd is recommended as this progressively reduces the number of cattle with drug resistant infections (Wangwe et al., 2019). With no cross resistance between suramin and any other trypanocide documented, combinations of suramin with melarsomine or quinapyramine should be reconsidered, especially in the treatment of surra. It would also be worth to start searching for candidates for combination therapy with existing trypanocides from the vast array of potentially active and safe natural and synthesised compounds screened against Trypanosoma parasites.
Apart from a combination of existing trypanocides, many potential combinations with experimental drugs have shown promise, including existing trypanocides combined with antibiotics, antiparasitic agents, plant extract/fractions and so on. For instance, Delespaux et al. (2010) have investigated the effect of combined ISM-antibiotic treatment in ISM-resistant T. congolense infections. Mice infected with ISM-resistant T. congolense and treated with a combination of ISM and tetracycline survived significantly longer than the group treated with ISM alone. The experiment in cattle infected with same T. congolense strain resulted in a similar observation with 50% cure in the groups treated with a combination of ISM/tetracycline or ISM/enrofloxacine compared to relapse parasitaemia in all the cattle treated with ISM alone (Delespaux et al., 2010). A combination of DA (3.5 mg/Kg) and secnidazole (100 or 200 mg) cleared parasitaemia with no relapse for up to 40 days post treatment in dogs experimentally infected with T. b. brucei while DA alone failed to clear the parasites. In addition, compared to the DA single therapy, dogs treated with the combination showed significantly higher PCV (days 5 – 18 post-treatment), total leukocyte count (at day 14 post-treatment), and haemoglobin concentration (at day 7 post-treatment) (Eke et al., 2020). Synergistic activities of combinations of natural compounds from many plants with conventional antibiotics against resistant bacterial strains have also been reported (Chusri et al., 2009).The combination of DA and the methanolic extract of Azadarachta indica has been shown to provide better results compared to a single diminazene aceturate therapy in rats infected with T. b. brucei (Omoja et al., 2011).
There is thus overwhelming evidence that combination chemotherapy can be beneficial in many ways: reducing administration times and/or dosage of the individual agents, thereby reducing toxicity; improving efficacy for agents that each individually are insufficiently active; and potentially delaying or preventing the onset of resistance. Although combinations could also introduce complications such as unanticipated drug interactions, successful examples such as nifurtimox/eflornithine for late stage gambiense HAT and artemisinin combination therapy in malaria should inspire similar efforts for AAT. However, it has been argued that drugs combinations should only be introduced before drug resistance has emerged to either of the elements (De Koning, 2017), as resistance to one of the agents will often make the combination fail and thus amount to an underdosing of the second agent as well – and that could lead to resistance of both drugs simultaneously.
7.5. Use of vitamins and immune modulators
Among the classical pathologies of trypanosomiases is immunosuppression. Host immunosuppression has also been shown to be essential in in vivo development of resistance to some trypanocides, specifically isometamidium (Tihon et al., 2017). Many researchers have indeed argued that some known trypanocides, particularly eflornithine, are actually trypanostatic and they only inhibit the multiplication of the parasite for an immunocompetent host to be able to fight it. This suggests a possible role for vitamins and other immunomodulators for use in combination with existing trypanocides. In addition, some studies have shown antitrypanosomal activity of vitamins and their antioxidant property may help ameliorate toxicity, which is a characteristic of many trypanocides. The antitrypanosomal activities of vitamin C, vitamin D3 and vitamin E been demonstrated in vitro with IC5O of 58.6 μM, 4.58 μM and 28.6 μM respectively in T. b. brucei (Jamal et al., 2016). Likewise, in mice infected with T. b. brucei, treatment with 250 and 500 ng/Kg vitamin D3 resulted in significant decrease in parasitaemia, significant suppression of splenomegaly and recovery from declined RBC compared with non-treated control mice, although there was no detectable difference in body weight (Jamal et al., 2016). In a different study, both a combination of 100 mg/Kg each of vitamins A and E, and that of 10,000 i.u/Kg vitamin C and 100 mg/Kg vitamin E resulted in a significantly lower parasitaemia in rats infected with T. b. brucei compared to the infected untreated controlled (Umar et al., 2008). The 2 vitamin combinations also prevented induced elevation of serum urea and creatinine, and in addition, the vitamin A and E combination significantly prevented the increase in alanine aminotransferase (ALT) and aspartate aminotransferase (AST), and the decrease in PCV associated with trypanosomiasis, suggesting protective roles of the vitamins against the disease (Umar et al., 2008). However, treatment with up to 1 mg/Kg vitamin E in rats infected with T. b. brucei did not result in a significant difference in the reduction of serum glutathione and in glutathione reductase concentrations compared to the DA treated group and the untreated control (Adegoke et al., 2018), possibly due to the low dosages of the vitamin used. In T. cruzi, vitamin C has shown trypanocidal activity similar to that of benznidazole in 3 forms of the parasite as well as reduced the toxicity of benznidazole in mammalian cells in vitro. Although there was a significant decrease in parasitaemia in a murine model of acute Chagas’ disease treated with vitamin C compared to the untreated infected control, the combination of benznidazole and vitamin C did not result in significant reduction in parasitaemia compared to benznidazole alone. However, the combined therapy resulted in 100% survival and reduced weight loss (Puente et al., 2018), with the vitamin apparently ameliorating the pathology of the infection rather than clearing it, with nonetheless substantial clinical benefit.
7.6. Use of drug Resistance Modulators
Intracellular accumulation is essential for many antiparasitic drugs to be effective. Thus, efflux transporters have been associated with drug resistance. Calcium antagonists such as verapamil and cyproheptadine have been shown to interfere with drug efflux by inhibiting P-glycoproteins to reverse multiple drug resistance in cancer cells (Tsuruo et al., 1981 1983). Likewise, verapamil reversed chloroquine resistance in Plasmodium falciparum (Martin et al., 1987), nifurtimox resistance in T. cruzi (Neal et al., 1989), regulated arsenite sensitivity in Leishmania donovani (Kaur and Dey, 2000) and potentiated pentamidine activity against L. major (Coelho et al., 2003). However, verapamil did not reverse multidrug resistance with diminazene aceturate or isometamidium chloride in T. b. brucei, either in vitro or in vivo (Kaminsky and Zweygarth, 1991). Similarly, in another study, calcium antagonists verapamil, desipramine, cyproheptidine and chlorpromazine failed to reverse resistance to melarsomine, diminazene aceturate and suramin in T. evansi (Anene et al., 1996). This apparent failure to reverse resistance in some trypanosome species is possibly due to the fact that their mechanisms of resistance are different from cancer cells, Plasmodium, Leishmania or T. cruzi (Anene et al., 1996; Kaminsky and Zweygarth, 1991) and specifically, do not involve the upregulation of ABC-type drug efflux transporters. Nevertheless, the active extrusion of isometamidium by T. b. brucei has certainly been demonstrated, although this process was not upregulated in in vitro-induced resistant strains (Eze et al., 2016). On balance, although efflux inhibitors have not yet been shown to be effective in reversing trypanocide resistance or potentiating drug action, this should be further investigated.
8. Drug Repurposing
Drug repurposing, the use of existing drugs in new medical indications is increasingly becoming popular because it potentially incurs lower costs and minimises safety concerns and development timelines compared to new drugs (Pushpakom et al., 2019). Repurposing strategies are especially relevant to drug discovery for NTDs, where time, resources, and information are limited (Klug et al., 2016). There is a great potential in repurposing antimicrobial and anticancer agents in the treatment for trypanosomiasis either as monotherapy or in combination with the current trypanocides. One success story of drug repurposing is eflornithine (difluoromethylornithine or DFMO), an inhibitor of polyamine biosynthesis enzyme ornithine decarboxylase, originally developed for cancer therapy and now utilized in the treatment of HAT (Burri and Brun, 2003). Although effective as a 2-week monotherapy, its combination with another repurposed drug, nifurtimox (long used against Chagas disease) has big advantages. Secnidazole, a 5-nitroimidazole antiprotozoal used in the treatment of giardiasis, trichomoniasis and amoebiasis has shown antitrypanosomal effects in vitro with an MIC of 1.4 mg/ml in vitro, and displayed in vivo activity in rats, which was curative in combination with DA (Eke et al., 2017). A study that tested the antitrypanosomal activity of ionophores and their derivatives reported remarkable in vitro activity against BSF T. b. brucei with a 50% growth inhibition (GI50) of 0.029 ± 0.002 μM; 0.040 ± 0.007 μM and 0.057 ± 0.029 μM for monensin, salinomycin n-butyl amide and salinomycin 2,2,2-trifluoroethyl ester respectively; very similar to the positive control, suramin (0.035 ± 0.002 μM) (Steverding et al., 2016). Salinomycin and monensin have been used in the treatment of another protozoan diseases, especially coccidiosis and salinomycin is currently proposed as a potential cancer therapy.
Several antibiotics are currently used in the treatment of protozoan diseases. Although no antibiotic is currently used clinically in the chemotherapy of trypanosomiasis, recent studies have shown great promise for utilising selected antibiotics in that respect. The trypanocidal activity of azithromycin was demonstrated in vitro against T. congolense, T. b. brucei and T. evansi, and in vivo against T. congolense with no relapse for up to 90 days in infected mice with 100% survival rate (Molefe et al., 2017). Another antibiotic, nitrofurantoin achieved complete cure at doses higher than 30 mg/Kg when orally administered for 7 days in mice infected with T. congolense (Suganuma et al., 2022). Terconazole, a synthetic triazole derivative used as antifungal agent, was found to exert promising activity against T. cruzi with IC50 of 4.56 ± 0.32 µM in trypomastigotes and 5.96 ± 0.35 μM in amastigotes, which is more effective than the control drug benznidazole (IC50 values of 12.71 ± 1.53 μM for trypomastigotes and 11.4 ± 0.5 μM for amastigotes) (Reigada et al., 2019).
9. Avoiding cross-resistance in new trypanocides
With advances in molecular parasitology and in the availability of tools for genetic manipulation, the mechanism of resistance is becoming clearer for more drugs and in more Trypanosoma species. Where drug resistance genes are identified, the knock-out and overexpression clones of the parasite can easily be used to screen for cross-resistance to new compounds in early development, and this should be standard practice. In addition, more drug resistant clones of the parasites obtained from gradual drug pressure are becoming available, and can be utilised to further understand the mechanism of action and possible resistance to other drugs and drug candidates.
While two new drugs, fexinidazole and acoziborole, have gone through clinical trials within the last decade for the treatment of HAT (Dickie et al., 2020), and fexinidazole has now been approved for clinical use as the first completely oral drug for sleeping sickness (DNDi, 2023), the last drug introduced for the animal trypanosomiases, melarsomine, is thirty years old and is not effective against T. congolense or T. vivax, so limited to use in T. evansi surra, in a few host species. As drug resistance and toxicity continue to undermine the role chemotherapy plays in the control of the animal trypanosomiases, the need for better and safer drugs cannot be overemphasized. Despite all the efforts to control human sleeping sickness, there is no evidence that the incidence of nagana is falling, and surra continues to spread for lack of almost any control measures.
Coincidentally, multiple trypanocides target the mitochondria, resulting in a reduced mitochondrial membrane potential by multiple mechanisms including through the modification of F1Fo ATP synthase complex (Fidalgo and Gille, 2011). Based on the success of pentamidine and diminazene, there were several efforts to redesign diamidines with improved targeting of membrane transporters and antitrypanosomal activity. Out of several diamidine derivatives, pafuramidine, a prodrug that is metabolised to the toxic 2,5-bis(4-amidinophenyl)-furan (furamidine) by cytochrome P450 enzymes, emerged as an efficacious drug candidate for HAT (Das and Boykin, 1977). Unfortunately, pafuramidine was withdrawn after Phase III clinical trials, following delayed renal toxicity in a small number of patients (Dickie et al., 2020). While the activity of the diamidine series was evaluated in veterinary trypanocides, the high potential for cross-resistance to diminazene has discouraged further development, although it is now clear that, in T. congolense at least, many diamidines including pentamidine are not cross-resistant with diminazene (Carruthers et al., 2021). Moreover, a recent study has shown that the mono-N-arylation of pentamidine yielded compounds that by-pass the cross-resistance mechanisms even in T. brucei (Robertson et al., 2021), since binding of pentamidine to its high affinity transporter AQP2 is in part facilitated by its two amidine groups (Alghamdi et al., 2020).
A separate effort focuses on the design and synthesis of a new class of compounds that, like diamidines, bind to the minor groove of DNA in trypanosomes, but are structurally distinct so as to avoid cross-resistance. These minor groove binders (MGBs) derived from distamycin displayed a remarkable antitrypanosomal activity (Scott et al., 2016; Giordani et al., 2019). In addition, the compounds showed activity against both T. congolense and T. vivax, exhibited no cross resistance to diamidines and isometamidium and low toxicity to mammalian cells. Furthermore, two selected MGBs were curative in a mouse model of AAT using a non-toxic dosage regimen (Giordani et al., 2019).
A group of compounds containing the oxaborole heterocycle coupled to a benzene ring, and thus known as benzoxaboroles, emerged in the last decade and showed potent antitrypanosomal activity (Ding et al., 2010). Following SAR studies and in vivo tests, SCYX-7158 (Figure 8 (1)) was shown to cure stage 2 HAT in a mouse model (Jacobs et al., 2011), and was advanced to clinical trial after its safety was established in mice and dogs (Wring et al., 2014). Another benzoxaborole, AN11736 (Figure 8 (2)) showed potent activity against African animal trypanosomes and was advanced as a lead candidate for development as a veterinary drug (Akama et al., 2018). Through development of resistance in vitro, investigations into the mechanism of action of benzoxaboroles identified RNA cleavage and polyadenylation specificity factor subunit 3 (CPSF3) as the target of the drug in multiple parasites (Steketee et al., 2018; Wall et al., 2018).
The role of the adenosine/adenine P2/AT1 in the uptake of multiple trypanocides has stimulated investigations into kinetoplastid nucleoside transporters and how they can be utilised as conduits for nucleoside drugs (Geiser et al., 2005; Berg et al., 2010; Ranjbarian et al., 2017; Hulpia et al., 2019). Series of nucleoside analogues with in vitro and in vivo activity against trypanosomes have been reported including cordycepin (3’-deoxyadenosine) and its analogues (Rottenberg et al., 2005; Vodnala et al., 2013), tubercidin (7-deazaadenosine) derivatives (Hulpia et al., 2020a; Lin et al., 2019), and inosine analogues (Hulpia et al., 2020b). We recently reported modified nucleoside analogues that target the highly conserved trypanosomatid P1 nucleoside transporters with high activity against the five most clinically relevant African veterinary trypanosomes (Mabille et al., 2022; Ungogo et al., 2023).
10. Conclusion
Having laid out in the pages above the current state of animal trypanosomiasis, its control and treatment options, we revisit the question posed by Professor Anene of the University of Nigeria, Nsukka, and his colleagues: “Drug resistance in pathogenic African trypanosomes: what hopes for the future?” (Anene et al., 2001). But we should start with the question “what progress in over 20 years?.”
The assessment of Anene et al. in 2001 seemed realistic at the time and has certainly proven to be so: “for now the management and control of trypanosomosis will continue to depend on proper usage of the few available trypanocides, especially strategic deployment of the sanative drugs in order to reduce the development of drug resistance, in addition to the continued use of environmentally friendly vector control programmes such tsetse trapping.” This has been the reality for decades now and on the ground little has changed. The science of AAT has progressed a lot, e.g. genomics, metabolomics, transcriptomics, RNAi and other such investigative tools have elucidated more of the biology and biochemistry of the various Trypanosoma species and the mechanisms of drug action and resistance. Following the discovery and robust validation of drug resistance genes in T. brucei, these genes have been applied as reliable biomarkers in prevalence studies and in preclinical drug screening in order to discard new trypanocides with same mode of resistance to the current drugs from the development pipeline. Importantly, it is now understood that mechanisms and biomarkers of drug resistance are different between the highly studied brucei group trypanosomes and T. congolense, determined by key biological differences (Carruthers et al., 2021; Munday et al., 2013; Steketee et al., 2021; Ungogo et al., 2022), stimulating interest into far more in-depth studies of T. congolense and T. vivax (Morrison et al., 2016). In addition, improved diagnostic options have been developed, even if not widely implemented yet. New active trypanocidal agents have been identified and treatment options have been proposed. But as of this moment, none of this will have changed the veterinary practice and AAT control very much in most endemic countries.
The biggest bottleneck is therefore not so much the science or the discovery of new diagnostic techniques and chemotherapeutic agents (which is nevertheless very important), but their implementation on the ground, where it is needed. The distance between the academic laboratory in the global north and the rural pastures of the global south remains mostly unbridged. Genuine north-south partnerships with the ability and funding to field test potential new trypanocides are needed and must have serious commitment from pharmaceutical companies and governments (north and south) towards development and implementation. The scale of training programmes in local communities about the rational and safe use of the available drugs, and the new diagnostic tools such as loop-mediated isothermal amplification (LAMP) and recombinase polymerase amplification (RPA)-based methods (Kim et al., 2023) should be increased, and the production/distribution of counterfeit and substandard drugs curbed. Efforts should be made to increase awareness and monitoring of animal trypanosomiasis in endemic countries of Northern Africa, Asia and South America. We must be clear that control of animal trypanosomiasis is part of sustainable agriculture and food security aims.
Most of the above list, unfortunately, can still be classified as ‘hopes for the future’. AAT needs to become a greater priority for governments, funders and NGOs. However, the Global Alliance for Livestock and Veterinary Medicine (GALVmed) has an active programme on animal trypanosomiasis and is implementing a point-of-care diagnostic that recognises T. congolense and T. vivax (VerY Diag; GALVmed, 2023) and is part of the consortium developing oxaborole AN11736 for AAT. It is also hoped that the increasing interest in T. congolense and T. vivax will generate the much-needed, reliable molecular biomarkers for the diagnosis and surveillance of drug resistance in these species. As a scientific and veterinary community working on animal trypanosomes and trypanosomiasis, it will to a large extent be up to us to ensure that over the next 20 years, things will have finally changed for the better for those dealing with AAT locally in the endemic areas.
Acknowledgments
This work was made possible through an Impact Accelerator Award from the Biotechnology and Biological Sciences Research Council of the UK (Grant ref: BB/S506734/1).
References
- Abdel Gadir, F. , Osman, O.M., Abdalla, H.S., Abdel Razig, M.T., 1981. Ethidium bromide-resistant trypanosomes in southern Darfur. Sudan J. Vet. Res. 3, 63–65.
- Adegoke, O. Adebayo, Chukwu I. Johnpaul, Anichukwu Faith Nnenna, 2018. Effect of Vitamin E on glutathione reductase and reduced glutathione levels of male wistar albino rats infected with Trypanosoma brucei brucei. Int. J. Biomed. Adv. Res. 9, 371–375.
- Afewerk, Y. , Clausen, P.-H., Abebe, G., Tilahun, G., Mehlitz, D., 2000. Multiple-drug resistant Trypanosoma congolense populations in village cattle of Metekel district, north-west Ethiopia. Acta Trop. 76, 231–238.
- Afework, Y. , Mäser, P., Etschmann, B., von Samson-Himmelstjerna, G., Zessin, K.-H., Clausen, P.-H., 2006. Rapid identification of isometamidium-resistant stocks of Trypanosoma b. brucei by PCR–RFLP. Parasitol. Res. 99, 253–261.
- Ainanshe, O.A. , Jennings, F.W., Holmes, P.H., 1992. Isolation of drug-resistant strains of Trypanosoma congolense from the Lower Shabelle Region of southern Somalia. Trop. Anim. Health Prod. 24, 65–73.
- Akama, T. , Zhang, Y.-K., Freund, Y.R., Berry, P., Lee, J., Easom, E.E., Jacobs, R.T., Plattner, J.J., Witty, M.J., Peter, R., Rowan, T.G., Gillingwater, K., Brun, R., Nare, B., Mercer, L., Xu, M., Wang, J., Liang, H., 2018. Identification of a 4-fluorobenzyl l-valinate amide benzoxaborole (AN11736) as a potential development candidate for the treatment of Animal African Trypanosomiasis (AAT). Bioorg. Med. Chem. Lett. 28, 6–10.
- Al-Salabi, M.I. , Wallace, L.J., Luscher, A., Maser, P., Candlish, D., Rodenko, B., Gould, M.K., Jabeen, I., Ajith, S.N., De Koning, H.P., 2007. Molecular interactions underlying the unusually high adenosine affinity of a novel Trypanosoma brucei nucleoside transporter. Mol. Pharmacol. 71, 921–929.
- Aldfer, M.M. , AlSiari, T.A., Elati, H.A.A., Natto, M.J., Alfayez, I.A., Campagnaro, G.D., Sani, B., Burchmore, R.J.S., Diallinas, G., De Koning, H.P., 2022. Nucleoside Transport and Nucleobase Uptake Null Mutants in Leishmania mexicana for the Routine Expression and Characterization of Purine and Pyrimidine Transporters. Int. J. Mol. Sci. 23, 8139.
- Alghamdi, A.H. , Munday, J.C., Campagnaro, G.D., Gurvic, D., Svensson, F., Okpara, C.E., Kumar, A., Quintana, J., Abril, M.E.M., Milić, P., Watson, L., Paape, D., Settimo, L., Dimitriou, A., Wielinska, J., Smart, G., Anderson, L.F., Woodley, C.M., Kelly, S.P.Y., Ibrahim, H.M.S., Hulpia, F., Al-Salabi, M.I., Eze, A.A., Sprenger, T., Teka, I.A., Gudin, S., Weyand, S., Field, M., Dardonville, C., Tidwell, R.R., Carrington, M., O’neill, P., Boykin, D.W., Zachariae, U., De Koning, H.P., 2020. Positively selected modifications in the pore of TBAQP2 allow pentamidine to enter Trypanosoma brucei. eLife 9, e56416.
- Alibu, V.P. , Richter, C., Voncken, F., Marti, G., Shahi, S., Renggli, C.K., Seebeck, T., Brun, R., Clayton, C., 2006. The role of Trypanosoma brucei MRPA in melarsoprol susceptibility. Mol. Biochem. Parasitol. 146, 38–44.
- Allen, C.L. , 2003. Clathrin-mediated endocytosis is essential in Trypanosoma brucei. EMBO J. 22, 4991–5002.
- Alsford, S. , Eckert, S., Baker, N., Glover, L., Sanchez-Flores, A., Leung, K.F., Turner, D.J., Field, M.C., Berriman, M., Horn, D., 2012. High-throughput decoding of antitrypanosomal drug efficacy and resistance. Nature 482, 232–236.
- Anene, B.M. , Onah, D.N., Nawa, Y., 2001. Drug resistance in pathogenic African trypanosomes: what hopes for the future? Vet. Parasitol. 96, 83–100.
- Anene, B.M. , Ezeokonkwo, R.C., Mmesirionye, T.I., Tettey, J.N.A., Brock, J.M., Barrett, M.P., De Koning, H.P., 2006. A diminazene-resistant strain of Trypanosoma brucei brucei isolated from a dog is cross-resistant to pentamidine in experimentally infected albino rats. Parasitology 132, 127–133.
- Anene, B.M. , Ross, C.A., Anika, S.M., Chukwu, C.C., 1996. Trypanocidal resistance in Trypanosoma evansi in vitro: effects of verapamil, cyproheptidine, desipramine and chlorpromazine alone and in combination with trypanocides. Vet. Parasitol. 62, 43–50.
- Apted, F.I.C. , 1970. Treatment of human trypanosomiasis. In: Mulligan, H.W. (Ed.), The African Trypanosomiases. George Allen and Unwin Ltd, pp. 684–710.
- Ardelli, B.F. , Woo, P.T.K., 2001. The in vitro effects of isometamidium chloride (Samorin) on the piscine hemoflagellate Cryptobia salmositica (Kinetoplastida, Bodonina). J. Parasitol. 87, 194–202.
- Arellano-Sota, C. , Arellano-Sota, C., 1988. Biology, ecology, and control of the vampire bat. Rev. Infect. Dis. 10, S615–S619.
- Arias, J.L. , Unciti-Broceta, J.D., Maceira, J., del Castillo, T., Hernández-Quero, J., Magez, S., Soriano, M., García-Salcedo, J.A., 2015. Nanobody conjugated PLGA nanoparticles for active targeting of African Trypanosomiasis. J. Control. Release 197, 190–198.
- Asghari, M.M. , Rassouli, M., 2022. First identification of Trypanosoma vivax among camels (Camelus dromedarius) in Yazd, central Iran, jointly with Trypanosoma evansi. Parasitol. Int. 86, 102450.
- Assefa, E. , Abebe, G., 2001. Drug-resistant Trypanosoma congolense in naturally infected donkeys in north Omo Zone, southern Ethiopia. Vet. Parasitol. 99, 261–271.
- Assefa, S. , Shibeshi, W., 2018. Drug resistance in African animal trypanosomes: a review. African J. Microbiol. Res. 12, 380–386.
- Autheman, D. , Crosnier, C., Clare, S., Goulding, D.A., Brandt, C., Harcourt, K., Tolley, C., Galaway, F., Khushu, M., Ong, H., Romero-Ramirez, A., Duffy, C.W., Jackson, A.P., Wright, G.J., 2021. An invariant Trypanosoma vivax vaccine antigen induces protective immunity. Nature 595, 96–100.
- Authie, E. , Somda, B., Dossama, M., Toutou, P., 1984. [Demonstration of resistance to trypanocides among the species of Trypanosoma congolense recently isolated in Burkina]. Rev. Elev. Med. Vet. Pays Trop. 37 Spec No, 219–235.
- Bacchi, C.J. , Lambros, C., Ellenbogen, B.B., Penkovsky, L.N., Sullivan, W., Eyinna, E.E., Hutner, S.H., 1975. Drug-Resistant Leptomonas : Cross-Resistance in Trypanocide-Resistant Clones. Antimicrob. Agents Chemother. 8, 688–692.
- Baker, N. , De Koning, H.P., Mäser, P., Horn, D., 2013. Drug resistance in African trypanosomiasis: the melarsoprol and pentamidine story. Trends Parasitol. 29, 110–118.
- Baker, N. , Glover, L., Munday, J.C., Aguinaga Andrés, D., Barrett, M.P., De Koning, H.P., Horn, D., 2012. Aquaglyceroporin 2 controls susceptibility to melarsoprol and pentamidine in African trypanosomes. Proc. Natl. Acad. Sci. 109, 10996–11001.
- Baker, N. , Hamilton, G., Wilkes, J.M., Hutchinson, S., Barrett, M.P., Horn, D., 2015. Vacuolar ATPase depletion affects mitochondrial ATPase function, kinetoplast dependency, and drug sensitivity in trypanosomes. Proc. Natl. Acad. Sci. 112, 9112–9117.
- Barr, S.C. , 2009. Canine Chagas’ Disease (American Trypanosomiasis) in North America. Vet. Clin. North Am. Small Anim. Pract. 39, 1055–1064.
- Barrett, M.P. , Burchmore, R.J.S., Stich, A., Lazzari, J.O., Frasch, A.C., Cazzulo, J.J., Krishna, S., 2003. The trypanosomiases. In: Lancet. pp. 1469–1480.
- Barrett, M.P. , Zhang, Z.Q., Denise, H., Giroud, C. and Baltz, T., 1995. A diamidine-resistant Trypanosoma equiperdum clone contains a P2 purine transporter with reduced substrate affinity. Mol. Biochem. Parasitol. 73, 223–229.
- Bengaly, Z. , Vitouley, S.H., Somda, M.B., Zongo, A., Têko-Agbo, A., Cecchi, G., Adam, Y., Sidibé, I., Bayala, B., Belem, A.M.G., Van Den Abbeele, J., Delespaux, V., 2018. Drug quality analysis of isometamidium chloride hydrochloride and diminazene diaceturate used for the treatment of African animal trypanosomosis in West Africa. BMC Vet. Res. 14, 361.
- Berg, M. , Kohl, L., Van der Veken, P., Joossens, J., Al-Salabi, M.I., Castagna, V., Giannese, F., Cos, P., Versées, W., Steyaert, J., Grellier, P., Haemers, A., Degano, M., Maes, L., De Koning, H.P., Augustyns, K., 2010. Evaluation of Nucleoside Hydrolase Inhibitors for Treatment of African Trypanosomiasis. Antimicrob. Agents Chemother. 54, 1900–1908.
- Berg, S.S. , 1960. Structure of Isometamidium (M. and B. 4180A), 7-m-Amidinophenyldiazoamino-2-amino-10-ethyl-9-phenylphenanthridinium Chloride Hydrochloride, the Red Isomer present in Metamidium. Nature 188, 1106–1107.
- Carter, N.S. , Barrett, M.P., De Koning, H.P. 1999. A drug resistance determinant from Trypanosoma brucei. Trends Microbiol. 7, 469–471.
- Berger, B. J, Carter, N.S., Fairlamb, A.H. 1995. Characterisation of pentamidine-resistant Trypanosoma brucei brucei. Mol. Biochem. Parasitol. 69, 289–298.
- Berger, B.J. , Fairlamb, A.H., 1994. Properties of melarsamine hydrochloride (Cymelarsan) in aqueous solution. Antimicrob. Agents Chemother. 38, 1298–1302.
- Birhanu, H. , Gebrehiwot, T., Goddeeris, B.M., Büscher, P., Van Reet, N., 2016. New Trypanosoma evansi Type B Isolates from Ethiopian Dromedary Camels. PLoS Negl. Trop. Dis. 10, e0004556.
- Bisser, S. , N’Siesi, F., Lejon, V., Preux, P., Van Nieuwenhove, S., Miaka Mia Bilenge, C., Büscher, P., 2007. Equivalence Trial of Melarsoprol and Nifurtimox Monotherapy and Combination Therapy for the Treatment of Second-Stage Trypanosoma brucei gambiense Sleeping Sickness. J. Infect. Dis. 195, 322–329.
- Boibessot, I. , Turner, C.M.R., Watson, D.G., Goldie, E., Connel, G., McIntosh, A., Grant, M.H., Skellern, G.G., 2002. Metabolism and distribution of phenanthridine trypanocides in Trypanosoma brucei. Acta Trop. 84, 219–228.
- Boma, S. , Vitouley, S.H., Somda, M.B., Bengaly, Z., Houaga, I., Lombo, Y., Tchamdja, E., Dayo, G.-K., 2022. In vivo analysis of trypanocidal drug resistance in sahelian goats infected by Trypanosoma vivax strains collected in northern Togo. Vet. Parasitol. 306, 109723.
- Brack, C. , Delain, E., 1975. Electron-microscopic mapping of AT-rich regions and of E. coli RNA polymerase-binding sites on the circular kinetoplast DNA of Trypanosoma cruzi. J. Cell Sci. 17, 287–306.
- Brack, C. , Delain, E., Riou, G., Festy, B., 1972. Molecular organization of the kinetoplast DNA of Trypanosoma cruzi treated with berenil, a DNA interacting drug. J. Ultrastruct. Res. 39, 568–579.
- Bridges, D.J. , Gould, M.K., Nerima, B., Mäser, P., Burchmore, R.J.S., De Koning, H.P., 2007. Loss of the High-Affinity Pentamidine Transporter Is Responsible for High Levels of Cross-Resistance between Arsenical and Diamidine Drugs in African Trypanosomes. Mol. Pharmacol. 71, 1098–1108.
- Browning, C. , Gilmour, W., 1913. Bactericidal action and chemical constitution with special reference to basic benzol derivatives. J. Pathol. Bacteriol. 18, 144–145.
- Browning, C.H. , 1949. Chemotherapy of T. congolense Infections with Phenanthridine Compounds : Biological Aspects. Nature 163, 590–591.
- Browning, C.H. , Morgan, G.T., Robb, J.V.M., Walls, L.P., 1938. The trypanocidal action of certain phenanthridinium compounds. J. Pathol. Bacteriol. 46, 203–204.
- Brun, R. , Hecker, H., Lun, Z.R., 1998. Trypanosoma evansi, 79.
- Brun, R. , Lun, Z.-R., 1994. Drug sensitivity of Chinese Trypanosoma evansi and Trypanosoma equiperdum isolates. Vet. Parasitol. 52, 37–46.
- Burri, C. , Brun, R., 2003. Eflornithine for the treatment of human African trypanosomiasis. Parasitol. Res. 90, S49–S52.
- Carruthers, L. V. , Munday, J.C., Ebiloma, G.U., Steketee, P., Jayaraman, S., Campagnaro, G.D., Ungogo, M.A., Lemgruber, L., Donachie, A., Rowan, T.G., Peter, R., Morrison, L.J., Barrett, M.P., De Koning, H.P., 2021. Diminazene resistance in Trypanosoma congolense is not caused by reduced transport capacity but associated with reduced mitochondrial membrane potential. Mol. Microbiol. 116, 564–588.
- Carter, N.S. , Berger, B.J., Fairlamb, A.H., 1995. Uptake of diamidine drugs by the P2 nucleoside transporter in melarsen- sensitive and -resistant Trypanosoma brucei brucei. J. Biol. Chem. 270, 28153–28157.
- Carter, N.S. , Fairlamb, A.H., 1993. Arsenical-resistant trypanosomes lack an unusual adenosine transporter. Nature 361, 173–176.
- Catley, A. , Leyland, T., 2001. Community participation and the delivery of veterinary services in Africa. Prev. Vet. Med. 49, 95–113.
- Chaka, H. , Abebe, G., 2003. Drug resistant trypanosomes: a threat to cattle production in the Southwest of Ethiopia. Rev. Elev. Med. Vet. Pays Trop. 56, 33–3.
- Cheesman, M. , Ilanko, A., Blonk, B., Cock, I., 2017. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with conventional antibiotics the solution? Pharmacogn. Rev. 11, 57–72.
- Chello, P.L. , Jaffe, J.J., 1972. Isolation, Partial Purification, and Properties of Thymidine Kinase from Trypanosoma (Trypanozoon) brucei rhodesiense. J. Parasitol. 58, 298–305.
- Chitanga, S. , Marcotty, T., Namangala, B., Van den Bossche, P., Van Den Abbeele, J., Delespaux, V., 2011. High Prevalence of Drug Resistance in Animal Trypanosomes without a History of Drug Exposure. PLoS Negl. Trop. Dis. 5, e1454.
- Chusri, S. , Villanueva, I., Voravuthikunchai, S.P., Davies, J., 2009. Enhancing antibiotic activity: a strategy to control Acinetobacter infections. J. Antimicrob. Chemother. 64, 1203–1211.
- Clausen, P.-H. , Bauer, B., Zessin, K.-H., Diall, O., Bocoum, Z., Sidibe, I., Affognon, H., Waibel, H., Grace, D., Randolph, T., 2010. Preventing and Containing Trypanocide Resistance in the Cotton Zone of West Africa. Transbound. Emerg. Dis. 57, 28–32.
- Clausen, P.-H. , Sidibe, I., Kabore´, I., Bauer, B., 1992. Development of multiple drug resistance of Trypanosoma congolense in Zebu cattle under high natural tsetse fly challenge in the pastoral zone of Samorogouan, Burkina Faso. Acta Trop. 51, 229–236.
- Coelho, A.C. , Beverley, S.M., Cotrim, P.C., 2003. Functional genetic identification of PRP1, an ABC transporter superfamily member conferring pentamidine resistance in Leishmania major. Mol. Biochem. Parasitol. 130, 83–90.
- Coppens, I. , Opperdoes, F.R., Courtoy, P.J., Baudhuin, P., 1987. Receptor-Mediated Endocytosis in the Bloodstream Form of Trypanosoma brucei 1. J. Protozool. 34, 465–473.
- Dagnachew, S. , Terefe, G., Abebe, G., Barry, D., McCulloch, R., Goddeeris, B., 2015. In vivo experimental drug resistance study in Trypanosoma vivax isolates from tsetse infested and non-tsetse infested areas of Northwest Ethiopia. Acta Trop. 146, 95–100.
- Dagnachew, S. , Tsegaye, B., Awukew, A., Tilahun, M., Ashenafi, H., Rowan, T., Abebe, G., Barry, D.J., Terefe, G., Goddeeris, B.M., 2017. Prevalence of bovine trypanosomosis and assessment of trypanocidal drug resistance in tsetse infested and non-tsetse infested areas of Northwest Ethiopia. Parasite Epidemiol. Control 2, 40–49.
- Dame, D.A. , Schmidt, C.H., 1970. The Sterile-Male Technique Against Tsetse Flies, Glossina Spp. Bull. Entomol. Soc. Am. 16, 24–30.
- Damper, D, Patton, C.L., 1976. Pentamidine transport and sensitivity in brucei-group trypanosomes. J. Protozool. 23, 349–56.
- Damper, Dianne, Patton, C.L., 1976. Pentamidine transport in Trypanosoma brucei—Kinetics and specificity. Biochem. Pharmacol. 25, 271–276.
- Das, B.P. , Boykin, D.W., 1977. Synthesis and antiprotozoal activity of 2,5-bis(4-guanylphenyl)furans. J. Med. Chem. 20, 531–536.
- Davey, D.G. , 1950. Experiments with “antrycide” in the Sudan and East Africa. Trans. R. Soc. Trop. Med. Hyg. 43, 583–616.
- Dávila, A.M.R. , Silva, R.A.M.S., 2006. Animal Trypanosomiasis in South America: Current Status, Partnership, and Information Technology. Ann. N. Y. Acad. Sci. 916, 199–212.
- Davkharbayar, B. , Davaasuren, B., Narantsatsral, S., Battur, B., Punsantsogvoo, M., Battsetseg, B., Mizushima, D., Inoue, N., Suganuma, K., 2020. Treatment Efficiency of Combination Therapy With Diminazene Aceturate and Quinapyramine Sulfate in a Horse With Dourine. J. Equine Vet. Sci. 87, 102905.
- De Deken, R. , Geerts, S., Kageruka, P., Ceulemans, F., Brandt, J., Schacht, E., Pascucci, C., Lootens, C., 1989. Chemoprophylaxis of trypanosomiasis, due to Trypanosoma (Nannomonas) congolense, in rabbits using a slow release device containing homidium bromide. Ann. Soc. Belg. Med. Trop. (1920). 69, 291–296.
- De Koning, H.P. , 2001b. Transporters in African trypanosomes: role in drug action and resistance. Int. J. Parasitol. 31, 512–522.
- De Koning, H.P. , 2017. Drug resistance in protozoan parasites. Emerg. Top. Life Sci. 1, 627–632.
- De Koning, H. , 2020. The Drugs of Sleeping Sickness: Their Mechanisms of Action and Resistance, and a Brief History. Trop. Med. Infect. Dis. 5, 14.
- De Koning, H.P. , 2001a. Uptake of Pentamidine in Trypanosoma brucei brucei is Mediated by Three Distinct Transporters: Implications for Cross-Resistance with Arsenicals. Mol. Pharmacol. 59, 586–592.
- De Koning, H.P. , Anderson, L.F., Stewart, M., Burchmore, R.J.S., Wallace, L.J.M., Barrett, M.P., 2004. The Trypanocide Diminazene Aceturate Is Accumulated Predominantly through the TbAT1 Purine Transporter: Additional Insights on Diamidine Resistance in African Trypanosomes. Antimicrob. Agents Chemother. 48, 1515–1519.
- De Koning, H.P. , Jarvis, S.M., 1999. Adenosine Transporters in Bloodstream Forms of Trypanosoma brucei brucei : Substrate Recognition Motifs and Affinity for Trypanocidal Drugs. Mol. Pharmacol. 56, 1162–1170.
- De Koning, H.P. , Jarvis, S.M., 2001. Uptake of pentamidine in Trypanosoma brucei brucei is mediated by the P2 adenosine transporter and at least one novel, unrelated transporter. Acta Trop. 80, 245–250.
- De Koning, H.P. , MacLeod, A., Barrett, M.P., Cover, B., Jarvis, S.M., 2000. Further evidence for a link between melarsoprol resistance and P2 transporter function in African trypanosomes. Mol. Biochem. Parasitol. 106, 181–185.
- de Noya, B.A. , González, O.N., 2015. An ecological overview on the factors that drives to Trypanosoma cruzi oral transmission. Acta Trop. 151, 94–102.
- Degneh, E. , Ashenafi, H., Kassa, T., Kebede, N., Shibeshi, W., Asres, K., Terefe, G., 2019. Trypanocidal drug resistance: A threat to animal health and production in Gidami district of Kellem Wollega Zone, Oromia Regional State, Western Ethiopia. Prev. Vet. Med. 168, 103–107.
- Delespaux, V. , Chitanga, S., Geysen, D., Goethals, A., Van den Bossche, P., Geerts, S., 2006. SSCP analysis of the P2 purine transporter TcoAT1 gene of Trypanosoma congolense leads to a simple PCR-RFLP test allowing the rapid identification of diminazene resistant stocks. Acta Trop. 100, 96–102.
- Delespaux, V. , De Koning, H., 2007. Drugs and drug resistance in African trypanosomiasis. Drug Resist. Updat. 10, 30–50.
- Delespaux, V. , Geerts, S., Brandt, J., Elyn, R., Eisler, M.C., 2002. Monitoring the correct use of isometamidium by farmers and veterinary assistants in Eastern Province of Zambia using the isometamidium-ELISA. Vet. Parasitol. 110, 117–122.
- Delespaux, V. , Geysen, D., Geerts, A.S., 2007. Point mutations in mitochondrial topoisomerase enzymes of Trypanosoma congolense are not involved in isometamidium resistance. Mol. Biochem. Parasitol. 151, 137–140.
- Delespaux, V. , Geysen, D., Majiwa, P.A.O., Geerts, S., 2005. Identification of a genetic marker for isometamidium chloride resistance in Trypanosoma congolense. Int. J. Parasitol. 35, 235–243.
- Delespaux, V. , Geysen, D., Van den Bossche, P., Geerts, S., 2008. Molecular tools for the rapid detection of drug resistance in animal trypanosomes. Trends Parasitol. 24, 236–42.
- Delespaux, V. , Vitouley, H.S., Marcotty, T., Speybroeck, N., Berkvens, D., Roy, K., Geerts, S., Van den Bossche, P., 2010. Chemosensitization of Trypanosoma congolense Strains Resistant to Isometamidium Chloride by Tetracyclines and Enrofloxacin. PLoS Negl. Trop. Dis. 4, e828.
- Desquesnes, M. , (de) La Rocque, S., Peregrine, A.S., 1995. French guyanan stock of Trypanosoma vivax resistant to diminazene aceturate but sensitive to isometamidium chloride. Acta Trop. 60, 133–136.
- Desquesnes, M. , Dargantes, A., Lai, D.-H., Lun, Z.-R., Holzmuller, P., Jittapalapong, S., 2013. Trypanosoma evansi and Surra: A Review and Perspectives on Transmission, Epidemiology and Control, Impact, and Zoonotic Aspects. Biomed Res. Int. 2013, 1–20.
- Desquesnes, M. , Dia, M.L., 2003. Trypanosoma vivax: mechanical transmission in cattle by one of the most common African tabanids, Atylotus agrestis. Exp. Parasitol. 103, 35–43.
- Desquesnes, M. , Gonzatti, M., Sazmand, A., Thévenon, S., Bossard, G., Boulangé, A., Gimonneau, G., Truc, P., Herder, S., Ravel, S., Sereno, D., Jamonneau, V., Jittapalapong, S., Jacquiet, P., Solano, P., Berthier, D., 2022. A review on the diagnosis of animal trypanosomoses. Parasit. Vectors 15, 64.
- Diaité, A. , Seye, M., Mane, A., Ndiaye, T., Seye, M.M., 1997. Detection and characterisation of trypanosome strains supposedly resistant to trypanocidal drugs In Senegal. In: Workshop on Epidemiological Tools for Monitoring Trypanosomosis and Tsetse Control Programmes. p. IAEA-TECDOC--925.
- Dickie, E.A. , Giordani, F., Gould, M.K., Mäser, P., Burri, C., Mottram, J.C., Rao, S.P.S., Barrett, M.P., 2020. New Drugs for Human African Trypanosomiasis: A Twenty First Century Success Story. Trop. Med. Infect. Dis. 5, 29.
- Ding, D. , Zhao, Y., Meng, Q., Xie, D., Nare, B., Chen, D., Bacchi, C.J., Yarlett, N., Zhang, Y.-K., Hernandez, V., Xia, Y., Freund, Y., Abdulla, M., Ang, K.-H., Ratnam, J., McKerrow, J.H., Jacobs, R.T., Zhou, H., Plattner, J.J., 2010. Discovery of Novel Benzoxaborole-Based Potent Antitrypanosomal Agents. ACS Med. Chem. Lett. 1, 165–169.
- DNDi (2023) EMA gives positive opinion to Fexinidazole Winthrop as first oral treatment of acute form of sleeping sickness (rhodesiense) found in East and Southern Africa. Press release, available at https://dndi.org/press-releases/2023/ema-gives-positive-opinion-fexinidazole-winthrop-first-oral-treatment-sleeping-sickness-rhodesiense/ (accessed 16/12/2023).
- Doherty, G.J. , McMahon, H.T., 2009. Mechanisms of Endocytosis. Annu. Rev. Biochem. 78, 857–902.
- Dressel, J. 1961. The discovery of Germanin by Oskar Dressel and Richard Kothe. J. Chem. Educ. 38, 620.
- Dukes, P. , 1984. Arsenic and old taxa: subspeciation and drug sensitivity in Trypanosoma brucei. Trans. R. Soc. Trop. Med. Hyg. 78, 711–725.
- Dwinger, R.H. , Agyemang, K., Snow, W.F., Rawlings, P., Leperre, P., Bah, M.L., 1994. Productivity of trypanotolerant cattle kept under traditional management conditions in the Gambia. Vet. Q. 16, 81–86.
- Eghianruwa, K.I. , Oridupa, O.A., 2018. Chemotherapeutic control of trypanosomosis - A review of past measures, current status and future trends. Vet. Arh. 88, 245–270.
- Eisler, M.C. , Brandt, J., Bauer, B., Clausen, P.-H., Delespaux, V., Holmes, P.H., Ilemobade, A., Machila, N., Mbwambo, H., McDermott, J., Mehlitz, D., Murilla, G., Ndung’u, J.M., Peregrine, A.S., Sidibé, I., Sinyangwe, L., Geerts, S., 2001. Standardised tests in mice and cattle for the detection of drug resistance in tsetse-transmitted trypanosomes of African domestic cattle. Vet. Parasitol. 97, 171–183.
- Eke, I.G. , Ezeh, I.O., Ezeudu, T.A., Eze, U.U., Anaga, A.O., Onyeyili, P.A., 2017. Anti-trypanosomal activity of secnidazole in vitro and in vivo. Trop. J. Pharm. Res. 16, 535–541.
- Eke, I.G. , Ezeh, I.O., Ezeudu, T.A., Eze, U.U., Anaga, A.O., Onyeyili, P.A., 2020. Efficacy of secnidazole-diminazene aceturate combination therapy in the late treatment of Trypanosoma brucei brucei infection in dogs. Brazilian J. Pharm. Sci. 56.
- El Rayah, I.E. , Kaminsky, R., Schmid, C., El Malik, K.H., 1999. Drug resistance in Sudanese Trypanosoma evansi. Vet. Parasitol. 80, 281–287.
- Elelu, N. , 2017. Assessment of veterinary drug retail outlets in two rural areas of Kwara state, north-central Nigeria. Sokoto J. Vet. Sci. 15, 54.
- Ewins, A.J. 1944. Chemotherapy in Tropical Medicine: A lecture delivered before the Chemical Society on th, 1944. J. Chem. Soc., 1944, 351–355. 17 February.
- Eze, A.A. , Gould, M.K., Munday, J.C., Tagoe, D.N.A., Stelmanis, V., Schnaufer, A., De Koning, H.P., 2016. Reduced Mitochondrial Membrane Potential Is a Late Adaptation of Trypanosoma brucei brucei to Isometamidium Preceded by Mutations in the γ Subunit of the F1Fo-ATPase. PLoS Negl. Trop. Dis. 10, e0004791.
- Eze, A.A. , Igoli, J., Gray, A.I., Skellern, G.G., De Koning, H.P., 2019. The individual components of commercial isometamidium do not possess stronger trypanocidal activity than the mixture, nor bypass isometamidium resistance. Int. J. Parasitol. Drugs Drug Resist. 9, 54–58.
- Ezeh, I.O. , Ugwu, E.N., Enemuo, O. V., Obi, C.F., Iheagwam, C.N., Ezeokonkwo, R.C., Onah, D.N., 2016. Efficacy of repeated doses of diminazene aceturate (Dinazene®) in the treatment of experimental Trypanosoma brucei infection of Albino rats. Iran. J. Vet. Res. 17, 124–129.
- Fairlamb, A.H. , Bowman, I.B.R., 1980. Uptake of the trypanocidal drug suramin by bloodstream forms of Trypanosoma brucei and its effect on respiration and growth rate in vivo. Mol. Biochem. Parasitol. 1, 315–333.
- Fairlamb, A.H. , Henderson, G.B., Cerami, A., 1989. Trypanothione is the primary target for arsenical drugs against African trypanosomes. Proc. Natl. Acad. Sci. 86, 2607–2611.
- Fairlamb, A.H. , Horn, D., 2018. Melarsoprol Resistance in African Trypanosomiasis. Trends Parasitol. 34, 481–492.
- Farikou, O. , Njiokou, F., Mbida Mbida, J.A., Njitchouang, G.R., Djeunga, H.N., Asonganyi, T., Simarro, P.P., Cuny, G., Geiger, A., 2010. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes—An epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect. Genet. Evol. 10, 115–121.
- Fetene, E. , Leta, S., Regassa, F., Büscher, P., 2021. Global distribution, host range and prevalence of Trypanosoma vivax: a systematic review and meta-analysis. Parasit. Vectors 14, 80.
- Fidalgo, L.M. , Gille, L., 2011. Mitochondria and Trypanosomatids: Targets and Drugs. Pharm. Res. 28, 2758–2770.
- Foil, L.D. , Hogsette, J.A., 1994. Biology and control of tabanids, stable flies and horn flies. Rev. Sci. Tech. l’OIE 13, 1125–1158.
- Food and Agricultural Organisation (FAO), 2012. Alliance to combat black market in counterfeit veterinary drugs.
- Ford, J. , 1970. Control by destruction of the larger fauna. In: Mulligan, H.W. (Ed.), The African Trypanosomiases. George Allen and Unwin Ltd, London, pp. 557–563.
- Ford, J. , Nash, T.A.M., Welch, J. R., 1970. Control by clearing of vegetation. In: Mulligan, H.W. (Ed.), The African Trypanosomiases. George Allen and Unwin Ltd, London, pp. 543–556.
- Fourneau, E. Tréfouel, J., Vallée, J., 1924. Recherches de chimiothérapie dans la série du Bayer 205. Urées des acides aminobenzoylaminonaphthaléniques. Ann Inst Pasteur 38, 81–114.
- Franco, J. , Scarone, L., Comini, M.A., 2018a. Drugs and Drug Resistance in African and American Trypanosomiasis. In: Annual Reports in Medicinal Chemistry. Pp. 97–133.
- Franco, J.R. , Cecchi, G., Priotto, G., Paone, M., Diarra, A., Grout, L., Simarro, P.P., Zhao, W., Argaw, D., 2018b. Monitoring the elimination of human African trypanosomiasis: Update to 2016. PloS Negl. Trop. Dis. 12, e0006890.
- Friedheim, E.A. , 1949. Mel B in the treatment of human trypanosomiasis. Am. J. Trop. Med. Hyg. 29(2), 173–180.
- Friedheim, E.A.H. , 1948. Melarsen Oxide in the Treatment of Human Trypanosomiasis. Ann. Trop. Med. Parasitol. 42, 357–363.
- Friedheim, E.A.H. , 1949. Mel B in the Treatment of Human Trypanosomiasis 1,2. Am. J. Trop. Med. Hyg. S1-29, 173–180.
- Friedheim, E.A.H. , 1951. Mel B in the Treatment of Tryparsamide Resistant T. Gambiense Sleeping Sickness: Observations on Drug Resistance in the Trypanosomes of the French Cameroun 1,2. Am. J. Trop. Med. Hyg. S1-31, 218–235.
- Gadelha, C. , Zhang, W., Chamberlain, J.W., Chait, B.T., Wickstead, B., Field, M.C., 2015. Architecture of a Host–Parasite Interface: Complex Targeting Mechanisms Revealed Through Proteomics. Mol. Cell. Proteomics 14, 1911–1926.
- Garcia, M.N. , Woc-Colburn, L., Aguilar, D., Hotez, P.J. and Murray, K.O., 2015. Historical perspectives on the epidemiology of human Chagas disease in Texas and recommendations for enhanced understanding of clinical Chagas disease in the Southern United States. PLoS Negl. Trop. Dis. 9, e0003981.
- Garcia-Salcedo, J.A. , Unciti-Broceta, J.D., Valverde-Pozo, J., Soriano, M., 2016. New Approaches to Overcome Transport Related Drug Resistance in Trypanosomatid Parasites. Front. Pharmacol. 7, 351.
- Geerts, S. , Holmes, P.H., Eisler, M.C., Diall, O., 2001. African bovine trypanosomiasis: the problem of drug resistance. Trends Parasitol. 17, 25–28.
- Geiser, F. , Lüscher, A., De Koning, H.P., Seebeck, T., Mäser, P., 2005. Molecular Pharmacology of Adenosine Transport in Trypanosoma brucei : P1/P2 Revisited. Mol. Pharmacol. 68, 589–595.
- Gill, B.S. , 1971. Resistance of Trypanosoma evansi to quinapyramine, suramin, stilbamidine and tryparsamide and analysis of cross-resistance. Trans. R. Soc. Trop. Med. Hyg. 65, 352–357.
- Giordani, F. , Khalaf, A.I., Gillingwater, K., Munday, J.C., De Koning, H.P., Suckling, C.J., Barrett, M.P., Scott, F.J., 2019. Novel Minor Groove Binders Cure Animal African Trypanosomiasis in an in Vivo Mouse Model. J. Med. Chem. 62, 3021–3035.
- Giordani, F. , Paape, D., Vincent, I.M., Pountain, A.W., Fernández-Cortés, F., Rico, E., Zhang, N., Morrison, L.J., Freund, Y., Witty, M.J., Peter, R., Edwards, D.Y., Wilkes, J.M., van der Hooft, J.J.J., Regnault, C., Read, K.D., Horn, D., Field, M.C., Barrett, M.P., 2020. Veterinary trypanocidal benzoxaboroles are peptidase-activated prodrugs. PLOS Pathog. 16, e1008932.
- Giordani, Morrison, L.J., Rowan, T.G., De Koning, H.P., Barrett, M.P., 2016. The animal trypanosomiases and their chemotherapy: a review. Parasitology 143, 1862–1889.
- Gizaw, Y. , Megersa, M., Fayera, T., 2017. Dourine: a neglected disease of equids. Trop. Anim. Health Prod. 49, 887–897.
- Gonzatti, M.I. , González-Baradat, B., Aso, P.M., Reyna-Bello, A., 2014. Trypanosoma (Duttonella) vivax and Typanosomosis in Latin America: Secadera/Huequera/Cacho Hueco. In: Trypanosomes and Trypanosomiasis. Springer Vienna, Vienna, pp. 261–285.
- Gooding, R.H. , Krafsur, E.S., 2005. Tsetse genetics: Contributions to Biology, Systematics, and Control of Tsetse Flies. Annu. Rev. Entomol. 50, 101–123.
- Gould, M.K. , Schnaufer, A., 2014. Independence from Kinetoplast DNA Maintenance and Expression Is Associated with Multidrug Resistance in Trypanosoma brucei In Vitro. Antimicrob. Agents Chemother. 58, 2925–2928.
- Grace, D. , Randolph, T., Diall, O., Clausen, P.-H., 2008. Training farmers in rational drug-use improves their management of cattle trypanosomosis: A cluster-randomised trial in south Mali. Prev. Vet. Med. 83, 83–97.
- Graf, F.E. , Baker, N., Munday, J.C., De Koning, H.P., Horn, D., Mäser, P., 2015. Chimerization at the AQP2-AQP3 locus is the genetic basis of melarsoprol-pentamidine cross-resistance in clinical Trypanosoma brucei gambiense isolates. Int. J. Parasitol. Drugs Drug Resist. 5, 65–68.
- Graf, F.E. , Ludin, P., Arquint, C., Schmidt, R.S., Schaub, N., Kunz Renggli, C., Munday, J.C., Krezdorn, J., Baker, N., Horn, D., Balmer, O., Caccone, A., De Koning, H.P., Mäser, P., 2016. Comparative genomics of drug resistance in Trypanosoma brucei rhodesiense. Cell. Mol. Life Sci. 73, 3387–3400.
- Graf, F.E. , Ludin, P., Wenzler, T., Kaiser, M., Brun, R., Pyana, P.P., Büscher, P., De Koning, H.P., Horn, D., Mäser, P., 2013. Aquaporin 2 Mutations in Trypanosoma brucei gambiense Field Isolates Correlate with Decreased Susceptibility to Pentamidine and Melarsoprol. PloS Negl. Trop. Dis. 7, e2475.
- Gresh, N. , Pullman, B., 1984. A theoretical study of the nonintercalative binding of berenil and stilbamidine to double-stranded (dA-dT)n oligomers. Mol. Pharmacol. 25, 452–8.
- Hamill, L.C. , Kaare, M.T., Welburn, S.C., Picozzi, K., 2013. Domestic pigs as potential reservoirs of human and animal trypanosomiasis in Northern Tanzania. Parasit. Vectors 6, 322.
- Hawking, F. , 1958. The Action of Berenil on Trypanosomes, Including Strains Resistant to Antrycide and to Stilbamidine. J. Comp. Pathol. Ther. 68, 295–299.
- Hawking, F. , 1963. Drug-resistance of Trypanosoma congolense and other trypanosomes to quinapyramine, phenanthridines, Berenil and other compounds in mice. Ann. Trop. Med. Parasitol. 57, 262–282.
- Hawking, F. , Sen, A.B., 1960. The trypanocidal action of homidium, quinapyramine and suramin. Br. J. Pharmacol. Chemother. 15, 567–570.
- Hébert, L. , Guitton, E., Madeline, A., Géraud, T., Zientara, S., Laugier, C., Hans, A., Büscher, P., Cauchard, J., Petry, S., 2018. Melarsomine hydrochloride (Cymelarsan®) fails to cure horses with Trypanosoma equiperdum OVI parasites in their cerebrospinal fluid. Vet. Parasitol. 264, 47–51.
- Herrera, H.M. , Dávila, A.M.R., Norek, A., Abreu, U.G., Souza, S.S., D’andrea, P.S., Jansen, A.M., 2004. Enzootiology of Trypanosoma evansi in pantanal, Brazil. Vet. Parasitol. 125, 263–275.
- Holmes, P.H. , 1997. New approaches to the integrated control of trypanosomosis. Vet. Parasitol. 71, 121–135.
- Horn, D. , 2014. Antigenic variation in African trypanosomes. Mol. Biochem. Parasitol. 195, 123–129.
- Hulpia, F. , Bouton, J., Campagnaro, G.D., Alfayez, I.A., Mabille, D., Maes, L., De Koning, H.P., Caljon, G., Van Calenbergh, S., 2020a. C6–O-alkylated 7-deazainosine nucleoside analogues: Discovery of potent and selective anti-sleeping sickness agents. Eur. J. Med. Chem. 188, 112018.
- Hulpia, F. , Campagnaro, G.D., Alzahrani, K.J., Alfayez, I.A., Ungogo, M.A., Mabille, D., Maes, L., De Koning, H.P., Caljon, G., Van Calenbergh, S., 2020b. Structure–Activity Relationship Exploration of 3′-Deoxy-7-deazapurine Nucleoside Analogues as Anti- Trypanosoma brucei Agents. ACS Infect. Dis. 6, 2045–2056.
- Hulpia, F. , Mabille, D., Campagnaro, G.D., Schumann, G., Maes, L., Roditi, I., Hofer, A., De Koning, H.P., Caljon, G., Van Calenbergh, S., 2019. Combining tubercidin and cordycepin scaffolds results in highly active candidates to treat late-stage sleeping sickness. Nat. Commun. 10, 5564.
- Igoli, J.O. , Blackburn, G., Gray, A.I., Sutcliffe, O.B., Watson, D.G., Euerby, M.R., Skellern, G.G., 2015. Chromatographic and spectroscopic analysis of the components present in the phenanthridinium trypanocidal agent isometamidium. Anal. Bioanal. Chem. 407, 1171–1180.
- Ijomanta, M.K. , Ugochukwu, C.I., Ugochukwu, E.I., 2016. Comparative effects of graded doses of diaminanzine ace-turate and fixed doses of levamisole in the treatment of albino mice experimentally infected with Trypanosoma brucei brucei. J. Exp. Appl. Anim. Sci. 2, 59–70.
- Jackson, A.P. , Allison, H.C., Barry, J.D., Field, M.C., Hertz-Fowler, C., Berriman, M., 2013. A Cell-surface Phylome for African Trypanosomes. PloS Negl. Trop. Dis. 7, e2121.
- Jacobi, E.A. , 2010. Das Schlafkrankheitsmedikament Germanin als Propagandainstrument: Rezeption in Literatur und Film zur Zeit des Nationalsozialismus. Wurzbg Medizinhist Mitt 29, 43–72.
- Jacobs, R.T. , Nare, B., Wring, S.A., Orr, M.D., Chen, D., Sligar, J.M., Jenks, M.X., Noe, R.A., Bowling, T.S., Mercer, L.T., Rewerts, C., Gaukel, E., Owens, J., Parham, R., Randolph, R., Beaudet, B., Bacchi, C.J., Yarlett, N., Plattner, J.J., Freund, Y., Ding, C., Akama, T., Zhang, Y.-K., Brun, R., Kaiser, M., Scandale, I., Don, R., 2011. SCYX-7158, an Orally-Active Benzoxaborole for the Treatment of Stage 2 Human African Trypanosomiasis. PloS Negl. Trop. Dis. 5, e1151.
- Jacobs, W.A. , Heidelberger, M., 1919. Aromatic arsenic compounds. V. N-substituted glycylarsanilic acids. J. Am. Chem. Soc. 41, 1809–1821.
- Jamal, R. , Shimogawara, R., Yamamoto, K., Ohta, N., 2016. Anti-trypanosome effects of nutritional supplements and vitamin D3: in vitro and in vivo efficacy against Trypanosoma brucei brucei. Trop. Med. Health 44, 26.
- Jamal, S. , Sigauque, I., Macuamule, C., Neves, L., Marcotty, T., Penzhorn, B.L., Van den Bossche, P., 2005. The susceptibility of Trypanosoma congolense isolated in Zambézia Province, Mozambique, to isometamidium chloride, diminazene aceturate and homidium chloride. Onderstepoort J Vet Res 72.
- Jennings, F.W. , 1990. Future prospects for the chemotherapy of human trypanosomiasis. Trans. R. Soc. Trop. Med. Hyg. 84, 618–621.
- Jennings, F.W. , 1993. Combination chemotherapy of CNS trypanosomiasis. Acta Trop. 54, 205–213.
- Jones-Davies, W.J. , Folkers, C., 1966. The prevalence of homidium-resistant strains of trypanosomes in cattle in northern Nigeria. Bull. Epizoot. Dis. Afr. 14, 65–72.
- Joshua, R.A. , 1988. Drug resistance in recent isolates of Trypanosoma brucei and Trypanosoma congolense. Rev. Elev. Med. Vet. Pays Trop. 41, 359–64.
- Kabayo, J.P. , 2002. Aiming to eliminate tsetse from Africa. Trends Parasitol. 18, 473–475.
- Kaminsky, R. , Gumm, I.D., Zweygarth, E., Chuma, F., 1990. A drug incubation infectivity test (DIIT) for assessing resistance in trypanosomes. Vet. Parasitol. 34, 335–343.
- Kaminsky, R. , Schmid, C., Lun, Z.R., 1997. Susceptibility of dyskinetoplastic Trypanosoma evansi and T. equiperdum to isometamidium chloride. Parasitol. Res. 83, 816–818.
- Kaminsky, R. , Zweygarth, E., 1991. The effect of verapamil alone and in combination with trypanocides on multidrug-resistant Trypanosoma brucei brucei. Acta Trop. 49, 215–225.
- Kasozi, K.I. , MacLeod, E.T., Waiswa, C., Mahero, M., Ntulume, I., Welburn, S.C., 2022. Systematic Review and Meta-Analysis on Knowledge Attitude and Practices on African Animal Trypanocide Resistance. Trop. Med. Infect. Dis. 7, 205.
- Kaur, J. , Dey, C.S., 2000. Putative P-Glycoprotein Expression in Arsenite-Resistant Leishmania donovani Down-Regulated by Verapamil. Biochem. Biophys. Res. Commun. 271, 615–619.
- Kazibwe, A.J.N. , Nerima, B., De Koning, H.P., Mäser, P., Barrett, M.P., Matovu, E., 2009. Genotypic Status of the TbAT1/P2 Adenosine Transporter of Trypanosoma brucei gambiense Isolates from Northwestern Uganda following Melarsoprol Withdrawal. PloS Negl. Trop. Dis. 3, e523.
- Keiser, J. , 2000. Investigations of the metabolites of the trypanocidal drug melarsoprol. Clin. Pharmacol. Ther. 67, 478–488.
- Kim, J. ,Álvarez-Rodríguez, A.,Li, Z., Radwanska, M., Magez, S., 2023. Recent progress in the detection of Surra, a neglected disease caused by Trypanosoma evansi with a One Health impact in large parts of the tropic and sub-tropic world. Microorganisms (in press).
- Kinabo, L.D.B. , Bogan, J. A., 1988. The pharmacology of isometamidium. J. Vet. Pharmacol. Ther. 11, 233–245.
- King, H. , Lourie, E.M., Yorke W. 1937. New trypanocidal substances. 1360. [Google Scholar]
- Kingsley, P. , 2015. Inscrutable medicines and marginal markets: tackling substandard veterinary drugs in Nigeria. Pastoralism 5, 2.
- Klug, D.M. , Gelb, M.H., Pollastri, M.P., 2016. Repurposing strategies for tropical disease drug discovery. Bioorg. Med. Chem. Lett. 26, 2569–2576.
- Knoppe, T.N. , Bauer, B., McDermott, J.J., Peregrine, A.S., Mehlitz, D., Clausen, P.-H., 2006. Isometamidium sensitivity of Trypanosoma congolense stocks from cattle in West Africa tested in mice and the drug incubation infectivity test. Acta Trop. 97, 108–116.
- Kristjanson, P.M. , Swallow, B.M., Rowlands, G.J., Kruska, R.L., de Leeuw, P.N., 1999. Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research. Agric. Syst. 59, 79–98.
- Kroubi, M. , Daulouede, S., Karembe, H., Jallouli, Y., Howsam, M., Mossalayi, D., Vincendeau, P., Betbeder, D., 2010. Development of a nanoparticulate formulation of diminazene to treat African trypanosomiasis. Nanotechnology 21, 505102.
- Kroubi, M. , Karembe, H., Betbeder, D., 2011. Drug delivery systems in the treatment of African trypanosomiasis infections. Expert Opin. Drug Deliv. 8, 735–747.
- Kumar, R. , Jain, S., Kumar, Saroj, Sethi, K., Kumar, Sanjay, Tripathi, B.N., 2017. Impact estimation of animal trypanosomosis (surra) on livestock productivity in India using simulation model: Current and future perspective. Vet. Parasitol. Reg. Stud. Reports 10, 1–12.
- Küpper, W. , Wolters, M., 1983. Observation on drug resistance of Trypanosoma (Nannomonas) congolense and Trypanosoma (Duttonella) vivax in cattle at a feedlot in the Northern Ivory Coast. Tropenmed. Parasitol. 34, 203–5.
- Kuriakose, S. , Muleme, H.M., Onyilagha, C., Singh, R., Jia, P., Uzonna, J.E., 2012. Diminazene Aceturate (Berenil) Modulates the Host Cellular and Inflammatory Responses to Trypanosoma congolense Infection. PloS One 7, e48696.
- Lane, A.N. , Jenkins, T.C., Brown, T., Neidle, S., 1991. Interaction of berenil with the EcoRI dodecamer d(CGCGAATTCGCG)2 in solution studied by NMR. Biochemistry 30, 1372–1385.
- Larson, S. , Carter, M., Hovel-Miner, G., 2021. Effects of trypanocidal drugs on DNA synthesis: new insights into melarsoprol growth inhibition. Parasitology 148, 1143–1150.
- Laveran, A. , 1904. The action of human serum on certain pathogenic Trypanosomes; action of arsenious acid upon Trypanosoma gambiense. Ann. Mag. Nat. Hist. Zool. Bot. Geol. 13, 401–403.
- Laveran, A. , Mesnil, F., 1902. Recherches sur le traitement et la prévention du nagana. Ann. Inst. Pasteur (Paris). 16, 786–817.
- Leach, T.M. , Roberts, C.J., 1981. Present status of chemotherapy and chemoprophylaxis of animal trypanosomiasis in the eastern hemisphere. Pharmacol. Ther. 13, 91–147.
- Lee, B.Y. , Bacon, K.M., Bottazzi, M.E., Hotez, P.J., 2013. Global economic burden of Chagas disease: a computational simulation model. Lancet Infect. Dis. 13, 342–348.
- Lejon, V. , 2003. Neuro-inflammatory risk factors for treatment failure in “early second stage” sleeping sickness patients treated with Pentamidine. J. Neuroimmunol. 144, 132–138.
- Liao, D. , Shen, J., 2010. Studies of quinapyramine-resistance of Trypanosoma brucei evansi in China. Acta Trop. 116, 173–177.
- Lidani, K.C.F. , Andrade, F.A., Bavia, L., Damasceno, F.S., Beltrame, M.H., Messias-Reason, I.J., Sandri, T.L., 2019. Chagas Disease: From Discovery to a Worldwide Health Problem. Front. Public Heal. 7, 166.
- Liebenehm, S. , Bett, B., Verdugo, C., Said, M., 2016. Optimal Drug Control under Risk of Drug Resistance – The Case of African Animal Trypanosomosis. J. Agric. Econ. 67, 510–533.
- Lin, C. , Hulpia, F., da Silva, C.F., Batista, D.D.G.J., Van Hecke, K., Maes, L., Caljon, G., Soeiro, M.D.N.C., Van Calenbergh, S., 2019. Discovery of Pyrrolo[2,3- b ]pyridine (1,7-Dideazapurine) Nucleoside Analogues as Anti- Trypanosoma cruzi Agents. J. Med. Chem. 62, 8847–8865.
- Livingstone, D. , 1858. Arsenic as a Remedy for the Tsetse Bite. BMJ s4-1, 360–361.
- Lourie, E.M. , Yorke, W., 1937. Studies in Chemotherapy. XVI. The Trypanocidal Action of Synthalin. Ann. Trop. Med. Parasitol. 31, 435–445.
- Lourie, E.M. , Yorke, W., 1939. Studies in Chemotherapy XXI. The Trypanocidal Action of Certain Aromatic Diamidines.. Ann. Trop. Med. Parasitol. 33, 289–304.
- Luckins, A.G. , Boid, R., Rae, P., Mahmoud, M.M., el Malik, K.H., Gray, A.R., 1979. Serodiagnosis of infection withTrypanosoma evansi in camels in the Sudan. Trop. Anim. Health Prod. 11, 1–12.
- Lüscher, A. , Nerima, B., Mäser, P., 2006. Combined contribution of TbAT1 and TbMRPA to drug resistance in Trypanosoma brucei. Mol. Biochem. Parasitol. 150, 364–366.
- Mabille, D. , Ilbeigi, K., Hendrickx, S., Ungogo, M.A., Hulpia, F., Lin, C., Maes, L., De Koning, H.P., Van Calenbergh, S., Caljon, G., 2022. Nucleoside analogues for the treatment of animal trypanosomiasis. Int. J. Parasitol. Drugs Drug Resist. 19, 21–30.
- Machila, N. , Wanyangu, S.W., McDermott, J., Welburn, S.C., Maudlin, I., Eisler, M.C., 2003. Cattle owners’ perceptions of African bovine trypanosomiasis and its control in Busia and Kwale Districts of Kenya. Acta Trop. 86, 25–34.
- Maclennan, K. , Jones-Davies, W., 1967. The occurrence of a berenil-resistant Trypanosoma congolense strain in Northern Nigeria. Vet. Rec. 80, 389–390.
- MacLennan, K.J.R. , 1968. Some recent findings concerning the use of Berenil for the treatment of bovine trypanosomiasis in Northern Nigeria. Trans. R. Soc. Trop. Med. Hyg. 62, 139–140.
- Magez, S. , Pinto Torres, J.E., Oh, S., Radwanska, M., 2021. Salivarian Trypanosomes Have Adopted Intricate Host-Pathogen Interaction Mechanisms That Ensure Survival in Plain Sight of the Adaptive Immune System. Pathogens 10, 679.
- Mamoudou, A. , Delespaux, V., Chepnda, V., Hachimou, Z., Andrikaye, J.P., Zoli, A., Geerts, S., 2008. Assessment of the occurrence of trypanocidal drug resistance in trypanosomes of naturally infected cattle in the Adamaoua region of Cameroon using the standard mouse test and molecular tools. Acta Trop. 106, 115–118.
- Manna, P.T. , Gadelha, C., Puttick, A.E., Field, M.C., 2015. ENTH and ANTH domain proteins participate in AP2-independent clathrin-mediated endocytosis. J. Cell Sci. 128, 2130–2142.
- Martin, S.K. , Oduola, A.M.J., Milhous, W.K., 1987. Reversal of chloroquine resistance in Plasmodium falciparum by verapamil. Science. 235, 899–901.
- Mäser, P. , Sütterlin, C., Kralli, A., Kaminsky, R., 1999. A Nucleoside Transporter from Trypanosoma brucei Involved in Drug Resistance. Science. 285, 242–244.
- Mathis, A.M. , Holman, J.L., Sturk, L.M., Ismail, M.A., Boykin, D.W., Tidwell, R.R., Hall, J.E., 2006. Accumulation and Intracellular Distribution of Antitrypanosomal Diamidine Compounds DB75 and DB820 in African Trypanosomes. Antimicrob. Agents Chemother. 50, 2185–2191.
- Matovu, E. , Geiser, F., Schneider, V., Mäser, P., Enyaru, J.C.K., Kaminsky, R., Gallati, S., Seebeck, T., 2001. Genetic variants of the TbAT1 adenosine transporter from African trypanosomes in relapse infections following melarsoprol therapy. Mol. Biochem. Parasitol. 117, 73–81.
- Matovu, E. , Stewart, M.L., Geiser, F., Brun, R., Mäser, P., Wallace, L.J.M., Burchmore, R.J., Enyaru, J.C.K., Barrett, M.P., Kaminsky, R., Seebeck, T., De Koning, H.P., 2003. Mechanisms of arsenical and diamidine uptake and resistance in Trypanosoma brucei. Eukaryot. Cell 2, 1003–1008.
- Matusevicius, M. , Corey, R., Gragera, M., Yamashita, K., Sprenger, T., Ungogo, M.A., Blaza, J., Castro-Hartmann, P., Chirgadze, D., Chaitania, V.S., Afanasyev, P., Melero, R., Warshamanage, Carazo, J.M., Carrington, M., Blundell, T., Murshudov, G., Stansfeld, P., Sansom, M., De Koning, H.P., Tate, C.G., Weyand, S.N., 2023. Insights from aquaporin structures into drug-resistant sleeping sickness. Nat Comm., submitted.
- Maudlin, I. , 2006. African trypanosomiasis. Ann. Trop. Med. Parasitol. 100, 679–701.
- Mbwambo, H.A. , Mella, P.N., Lekaki, K.A., 1988. Berenil (diminazene aceturate)-resistant Trypanosoma congolense in cattle under natural tsetse challenge at Kibaha, Tanzania. Acta Trop. 45, 239–44.
- Mdachi, R.E. , Ogolla, K.O., Auma, J.E., Wamwiri, F.N., Kurgat, R.K., Wanjala, K.B., Mugunieri, L.G., Alusi, P.M., Chemuliti, J.K., Mukiria, P.W., Okoth, S.O., 2023. Variation of sensitivity of Trypanosoma evansi isolates from Isiolo and Marsabit counties of Kenya to locally available trypanocidal drugs. PloS One 18, e0281180.
- Melaku, A. , Birasa, B., 2013. Drugs and Drug Resistance in African Animal Trypanosomosis : A Review. Eur. J. Appl. Sci. 5, 82–89.
- Mills, R.M. , 2020. Chagas disease: Epidemiology and barriers to treatment. Am. J. Med. 133, 1262–1265.
- Mingala, C.N. , Pasag, A.C.P., Salinas, M.B.S., Balbin, M.M., Villanueva, M.A., 2019. Characterization of drug resistance-associated TevAT1 gene of Trypanosoma evansi from Philippine water buffaloes (Bubalus bubalis). Ann. Parasitol. 65, 381–386.
- Mizushima, D. , Amgalanbaatar, T., Davaasuren, B., Kayano, M., Naransatsral, S., Myagmarsuren, P., Otgonsuren, D., Enkhtaivan, B., Davkharbayar, B., Mungun-Ochir, B., Baatarjargal, P., Nyamdolgor, U., Soyolmaa, G., Altanchimeg, A., Zoljargal, M., Nguyen, T.-T., Battsetseg, B., Battur, B., Inoue, N., Yokoyama, N., Suganuma, K., 2020. Nationwide serological surveillance of non-tsetse-transmitted horse trypanosomoses in Mongolia. Parasite Epidemiol. Control 10, e00158.
- Mohamed-Ahmed, M.M. , Rahman, A.H., Abdel Karim, E.I., 1992. Multiple drug-resistant bovine trypanosomes in South Darfur Province, Sudan. Trop. Anim. Health Prod. 24, 179–181.
- Molefe, N.I. , Yamasaki, S., Macalanda, A.M.C., Suganuma, K., Watanabe, K., Xuan, X., Inoue, N., 2017. Oral administration of azithromycin ameliorates trypanosomosis in Trypanosoma congolense-infected mice. Parasitol. Res. 116, 2407–2415.
- Morrison, L.J. , Vezza, L., Rowan, T., Hope, J.C., 2016. Animal African Trypanosomiasis: Time to Increase Focus on Clinically Relevant Parasite and Host Species. Trends Parasitol. 32, 599–607.
- Moti, Y. , De Deken, R., Thys, E., Van Den Abbeele, J., Duchateau, L., Delespaux, V., 2015. PCR and microsatellite analysis of diminazene aceturate resistance of bovine trypanosomes correlated to knowledge, attitude and practice of livestock keepers in South-Western Ethiopia. Acta Trop. 146, 45–52.
- Mulandane, F.C. , Fafetine, J., Van Den Abbeele, J., Clausen, P.-H., Hoppenheit, A., Cecchi, G., Oosthuizen, M., Delespaux, V., Neves, L., 2018. Resistance to trypanocidal drugs in cattle populations of Zambezia Province, Mozambique. Parasitol. Res. 117, 429–436.
- Mulugeta, W. , Wilkes, J., Mulatu, W., Majiwa, P.A.O., Masake, R., Peregrine, A.S., 1997. Long-term occurrence of Trypanosoma congolense resistant to diminazene, isometamidium and homidium in cattle at Ghibe, Ethiopia. Acta Trop. 64, 205–217.
- Munday, J.C. , Eze, A.A., Baker, N., Glover, L., Clucas, C., Aguinaga Andres, D., Natto, M.J., Teka, I.A., McDonald, J., Lee, R.S., Graf, F.E., Ludin, P., Burchmore, R.J.S., Turner, C.M.R., Tait, A., MacLeod, A., Maser, P., Barrett, M.P., Horn, D., De Koning, H.P., 2014. Trypanosoma brucei, 69.
- Munday, J.C. , Rojas López, K.E., Eze, A.A., Delespaux, V., Van Den Abbeele, J., Rowan, T., Barrett, M.P., Morrison, L.J., De Koning, H.P., 2013. Functional expression of TcoAT1 reveals it to be a P1-type nucleoside transporter with no capacity for diminazene uptake. Int. J. Parasitol. Drugs Drug Resist. 3, 69–76.
- Munday, J.C. , Settimo, L., De Koning, H.P., 2015a. Transport proteins determine drug sensitivity and resistance in a protozoan parasite, Trypanosoma brucei. Front. Pharmacol. 6, 32.
- Munday, J.C. , Tagoe, D.N.A., Eze, A.A., Krezdorn, J.A.M., Rojas López, K.E., Alkhaldi, A.A.M., McDonald, F., Still, J., AlzahraFni, K.J., Settimo, L., De Koning, H.P., 2015b. Functional analysis of drug resistance-associated mutations in the Trypanosoma brucei adenosine transporter 1 (TbAT1) and the proposal of a structural model for the protein. Mol. Microbiol. 96, 887–900.
- Mungube, E.O. , Vitouley, H.S., Allegye-Cudjoe, E., Diall, O., Boucoum, Z., Diarra, B., Sanogo, Y., Randolph, T., Bauer, B., Zessin, K.-H., Clausen, P.-H., 2012. Detection of multiple drug-resistant Trypanosoma congolense populations in village cattle of south-east Mali. Parasit. Vectors 5, 155.
- Mutugi, M.W. , Boid, R., Luckins, A.G., 1994. Experimental induction of suramin-resistance in cloned and uncloned stocks of Trypanosoma evansi using immunosuppressed and immunocompetent mice. Trop. Med. Parasitol. 45, 232–6.
- Na’Isa, B.K. , 1967. Follow-up of a survey on the prevalence of homidium-resistant strains of trypanosomes in cattle in Northern Nigeria and drug cross-resistance tests on the strains with Samorin and Berenil. Bull. Epizoot. Dis. Afr. 15, 231–41.
- Namangala, B. , Odongo, S., 2014. Animal African Trypanosomosis in Sub-Saharan Africa and Beyond African Borders. In: Trypanosomes and Trypanosomiasis. Springer Vienna, Vienna, pp. 239–260.
- Napier, L.E. , Gupta, P.C. Sen, 1942. A Peculiar Neurological Sequel to Administration of 4: 4’-Diamidino-Diphenyl-Ethylene (M.&B. 744). Ind. Med. Gaz.
- Ndoutamia, G. , Moloo, S.K., Murphy, N.B., Peregrine, A.S., 1993. Derivation and characterization of a quinapyramine-resistant clone of Trypanosoma congolense. Antimicrob. Agents Chemother. 37, 1163–1166.
- Neal, R.A. , van Bueren, J., McCoy, N.G., Iwobi, M., 1989. Reversal of drug resistance in Trypanosoma cruzi and Leishmania donovani by verapamil. Trans. R. Soc. Trop. Med. Hyg. 83, 197–198.
- Nerima, B. , Matovu, E., Lubega, G.W., Enyaru, J.C.K., 2007a. Detection of mutant P2 adenosine transporter (TbAT1) gene in Trypanosoma brucei gambiense isolates from northwest Uganda using allele-specific polymerase chain reaction. Trop. Med. Int. Heal. 12, 1361–1368.
- Newton, B.A. , 1964. Mechanisms of Action of Phenanthridine and Aminoquinaldine Trypanocides. In: Advances in Chemotheraphy. Elsevier, pp. 35–83.
- Njiokou, F. , Nimpaye, H., Simo, G., Njitchouang, G.R., Asonganyi, T., Cuny, G., Herder, S., 2010. Domestic animals as potential reservoir hosts of Trypanosoma brucei gambiense in sleeping sickness foci in Cameroon. Parasite 17, 61–66.
- Obi, C.F. , Okpala, M.I., Ezeh, I.O., Onyeabor, A., Ezeokonkwo, R.C., 2022. Drug-resistant trypanosome isolates populations in dogs in Enugu North Senatorial Zone, Southeastern Nigeria. Parasitol. Res. 121, 423–431.
- Ogbaje, C.I. , Lawal, I.A., Ajanusi, O.J., 2015. Sensitivity of Nigerian field isolates of Trypanosoma vivax and Trypanosoma congolense to commonly available trypanocides. Asian Pacific J. Trop. Dis. 5, 214–218.
- Oguejiofor, C. , Ochiogu, I., Umeoduagu, C., 2010. Increasing doses of diminazene aceturate: adverse reproductive effects in female Wistar rats. Asian Pac. J. Trop. Med. 3, 887–889.
- OIE, 2013. Dourine (Infection with Trypanosoma equiperdum) Aetiology Epidemiology Diagnosis Prevention and Control References. (https://www.woah.org/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/DOURINE.pdf). Accesed last on Dec. 10th, 2023.
- Okello, I. , Mafie, E., Eastwood, G., Nzalawahe, J., Mboera, L.E.G., 2022a. African Animal Trypanosomiasis: A Systematic Review on Prevalence, Risk Factors and Drug Resistance in Sub-Saharan Africa. J. Med. Entomol. 59, 1099–1143.
- Okello, I. , Mafie, E., Nzalawahe, J., Eastwood, G., Mboera, L.E.G., Hakizimana, J.N., Ogola, K., 2022b. Trypanosoma Congolense Resistant to Trypanocidal Drugs Homidium and Diminazene and their Molecular Characterization in Lambwe, Kenya. Acta Parasitol.
- Olila, D. , McDermott, J.J., Eisler, M.C., Mitema, E.S., Patzelt, R.J., Clausen, P.-H., Poetzsch, C.J., Zessin, K.-H., Mehlitz, D., Peregrine, A.S., 2002. Drug sensitivity of trypanosome populations from cattle in a peri-urban dairy production system in Uganda. Acta Trop. 84, 19–30.
- Oliveira, C.B. , Rigo, L.A., Rosa, L.D., Gressler, L.T., Zimmermann, C.E.P., Ourique, A.F., Da Silva, A.S., Miletti, L.C., Beck, R.C.R., Monteiro, S.G., 2014. Liposomes produced by reverse phase evaporation: in vitro and in vivo efficacy of diminazene aceturate against Trypanosoma evansi. Parasitology 141, 761–769.
- Omoja, V. , Anaga, A., Obidike, I., Ihedioha, T., Umeakuana, P., Mhomga, L., Asuzu, I., Anika, S., 2011. The effects of combination of methanolic leaf extract of Azadirachta indica and diminazene diaceturate in the treatment of experimental Trypanosoma brucei brucei infection in rats. Asian Pac. J. Trop. Med. 4, 337–341.
- Ooi, C.-P. , Schuster, S., Cren-Travaillé, C., Bertiaux, E., Cosson, A., Goyard, S., Perrot, S., Rotureau, B., 2016. The Cyclical Development of Trypanosoma vivax in the Tsetse Fly Involves an Asymmetric Division. Front. Cell. Infect. Microbiol. 6, 115.
- Ormerod, W.E. , 1952. A study of resistance to antrycide in a strain of Trypanosoma equiperdum. Br. J. Pharmacol. Chemother. 7, 674–684.
- Ormerod, W.E. , 1951a. The mode of action of antrycide. Br. J. Pharmacol. Chemother. 6, 325–333.
- Ormerod, W.E. , 1951b. A study of basophilic inclusion bodies produced by chemotherapeutic agents in trypanosomes. Br. J. Pharmacol. Chemother. 6, 334–341.
- Osman, A.S. , Jennings, F.W., Holmes, P.H., 1992. The rapid development of drug-resistance by Trypanosoma evansi in immunosuppressed mice. Acta Trop. 50, 249–257.
- Otesile, E.B. , Tabel, H., 1987. Enhanced Resistance of Highly Susceptible Balb/c Mice to Infection with Trypanosoma congolense after Infection and Cure. J. Parasitol. 73, 947.
- Paine, M.F. , Wang, M.Z., Generaux, C.N., Boykin, D.W., Wilson, W.D., De Koning, H.P., Olson, C.A., Pohlig, G., Burri, C., Brun, R., Murilla, G.A., Thuita, J.K., Barrett, M.P., Tidwell, R.R., 2010. Diamidines for human African trypanosomiasis. Curr. Opin. Investig. Drugs 11, 876–83.
- Pal, A. , Hall, B.S., Field, M.C., 2002. Evidence for a non-LDL-mediated entry route for the trypanocidal drug suramin in Trypanosoma brucei. Mol. Biochem. Parasitol. 122, 217–221.
- Pascucci, I. , Di Provvido, A., Cammà, C., Di Francesco, G., Calistri, P., Tittarelli, M., Ferri, N., Scacchia, M., Caporale, V., 2013. Diagnosis of dourine in outbreaks in Italy. Vet. Parasitol. 193, 30–38.
- Pearce, L. , 1921. Studies on the treatment of human trypanosomiasis with tryparsamide (the sodium salt of n-phenylglycineamide-p-arsonic acid). J. Exp. Med. 34, 1–104.
- Peregrine, A.S. , 1994. Chemotherapy and delivery systems: haemoparasites. Vet. Parasitol. 54, 223–248.
- Peregrine, A.S. , Gray, M.A., Moloo, S.K., 1997. Cross-resistance associated with development of resistance to isometamidium in a clone of Trypanosoma congolense. Antimicrob. Agents Chemother. 41, 1604–1606.
- Peregrine, A.S. , Mamman, M., 1993. Pharmacology of diminazene: a review. Acta Trop. 54, 185–203.
- Pinder, M. , Authie, E., 1984. The appearance of isometamidium resistant Trypanosoma congolense in West Africa. Acta Trop. 41, 247–52.
- Pohlig, G. , Bernhard, S.C., Blum, J., Burri, C., Mpanya, A., Lubaki, J.-P.F., Mpoto, A.M., Munungu, B.F., N’tombe, P.M., Deo, G.K.M., Mutantu, P.N., Kuikumbi, F.M., Mintwo, A.F., Munungi, A.K., Dala, A., Macharia, S., Bilenge, C.M.M., Mesu, V.K.B.K., Franco, J.R., Dituvanga, N.D., Tidwell, R.R., Olson, C.A., 2016. Efficacy and Safety of Pafuramidine versus Pentamidine Maleate for Treatment of First Stage Sleeping Sickness in a Randomized, Comparator-Controlled, International Phase 3 Clinical Trial. PloS Negl. Trop. Dis. 10, e0004363.
- Potts, W.H. , Jackson, C.H.N., 1952. The Shinyanga game destruction experiment. Bull. Entomol. Res. 53, 365–374.
- Priotto, G. , Kasparian, S., Mutombo, W., Ngouama, D., Ghorashian, S., Arnold, U., Ghabri, S., Baudin, E., Buard, V., Kazadi-Kyanza, S., Ilunga, M., Mutangala, W., Pohlig, G., Schmid, C., Karunakara, U., Torreele, E., Kande, V., 2009. Nifurtimox-eflornithine combination therapy for second-stage African Trypanosoma brucei gambiense trypanosomiasis: a multicentre, randomised, phase III, non-inferiority trial. Lancet 374, 56–64.
- Puente, V. , Demaria, A., Frank, F.M., Batlle, A., Lombardo, M.E., 2018. Anti-parasitic effect of vitamin C alone and in combination with benznidazole against Trypanosoma cruzi. PloS Negl. Trop. Dis. 12, e0006764.
- Pumhom, P. , Morand, S., Tran, A., Jittapalapong, S., Desquesnes, M., 2015. Trypanosoma from rodents as potential source of infection in human-shaped landscapes of South-East Asia. Vet. Parasitol. 208, 174–180.
- Pushpakom, S. , Iorio, F., Eyers, P.A., Escott, K.J., Hopper, S., Wells, A., Doig, A., Guilliams, T., Latimer, J., McNamee, C., Norris, A., Sanseau, P., Cavalla, D., Pirmohamed, M., 2019. Drug repurposing: progress, challenges and recommendations. Nat. Rev. Drug Discov. 18, 41–58.
- Pyana Pati, P. , Van Reet, N., Mumba Ngoyi, D., Ngay Lukusa, I., Karhemere Bin Shamamba, S., Büscher, P., 2014. Melarsoprol Sensitivity Profile of Trypanosoma brucei gambiense Isolates from Cured and Relapsed Sleeping Sickness Patients from the Democratic Republic of the Congo. PloS Negl. Trop. Dis. 8, e3212.
- Quintana, J.F. , Bueren-Calabuig, J., Zuccotto, F., De Koning, H.P., Horn, D., Field, M.C., 2020. Instability of aquaglyceroporin (AQP) 2 contributes to drug resistance in Trypanosoma brucei. PloS Negl. Trop. Dis. 14, e0008458.
- Quintana, J.F. , Pino, R.C. Del, Yamada, K., Zhang, N., 2018. Adaptation and Therapeutic Exploitation of the Plasma Membrane of African Trypanosomes. Genes (Basel). 9, 368.
- Ramirez, A.I.R. , 2020. Antigen discovery in Trypanosoma vivax. University of Liverpool.
- Rani, R. , Narasimhan, B., Varma, R.S., Kumar, R., 2021. Naphthoquinone derivatives exhibit apoptosis-like effect and anti-trypanosomal activity against Trypanosoma evansi. Vet. Parasitol. 290, 109367.
- Ranjbarian, F. , Vodnala, M., Alzahrani, K.J.H., Ebiloma, G.U., De Koning, H.P., Hofer, A., 2017. 9-(2′-Deoxy-2′-Fluoro-β-D-Arabinofuranosyl) Adenine Is a Potent Antitrypanosomal Adenosine Analogue That Circumvents Transport-Related Drug Resistance. Antimicrob. Agents Chemother. 61.
- Reigada, C. , Sayé, M., Valera-Vera, E., Miranda, M.R., Pereira, C.A., 2019. Repurposing of terconazole as an anti Trypanosoma cruzi agent. Heliyon 5, e01947.
- Richards, S. , Morrison, L.J., Torr, S.J., Barrett, M.P., Manangwa, O., Mramba, F., Auty, H., 2021. Pharma to farmer: field challenges of optimizing trypanocide use in African animal trypanosomiasis. Trends Parasitol. 37, 831–843.
- Riou, G. , Benard, J., 1980. Berenil induces the complete loss of kinetoplast DNA sequences in Trypanosoma equiperdum. Biochem. Biophys. Res. Commun. 96, 350–354.
- Robertson, J. , Ungogo, M.A., Aldfer, M.M., Lemgruber, L., McWhinnie, F.S., Bode, B.E., Jones, K.L., Watson, A.J.B., De Koning, H.P., Burley, G.A., 2021. Direct, Late-Stage Mono- N -arylation of Pentamidine: Method Development, Mechanistic Insight, and Expedient Access to Novel Antiparastitics against Diamidine-Resistant Parasites. ChemMedChem 16, 3396–3401.
- Rodgers, J. , Jones, A., Gibaud, S., Bradley, B., McCabe, C., Barrett, M.P., Gettinby, G., Kennedy, P.G.E., 2011. Melarsoprol Cyclodextrin Inclusion Complexes as Promising Oral Candidates for the Treatment of Human African Trypanosomiasis. PloS Negl. Trop. Dis. 5, e1308.
- Ross, C.A. , Barns, A.M., 1996. Alteration to one of three adenosine transporters is associated with resistance to Cymelarsan in Trypanosoma evansi. Parasitol. Res. 82, 183–188.
- Rottenberg, M.E. , Masocha, W., Ferella, M., Petitto-Assis, F., Goto, H., Kristensson, K., McCaffrey, R., Wigzell, H., 2005. Treatment of African Trypanosomiasis with Cordycepin and Adenosine Deaminase Inhibitors in a Mouse Model. J. Infect. Dis. 192, 1658–1665.
- Rowlands, G.J. , Leak, S.G.A., Peregrine, A.S., Nagda, S.M., Mulatu, W., D’Ieteren, G.D.M., 2001. The incidence of new and the prevalence and persistence of recurrent trypanosome infections in cattle in southwest Ethiopia exposed to a high challenge with drug-resistant parasites. Acta Trop. 79, 149–163.
- Roy Chowdhury, A. , Bakshi, R., Wang, J., Yildirir, G., Liu, B., Pappas-Brown, V., Tolun, G., Griffith, J.D., Shapiro, T.A., Jensen, R.E., Englund, P.T., 2010. The Killing of African Trypanosomes by Ethidium Bromide. PloS Pathog. 6, e1001226.
- Rurangirwa, F.R. , Tabel, H., Losos, G.J., Tizard, I.R., 1979. Suppression of antibody response to Leptospira biflexa and Brucella abortus and recovery from immunosuppression after Berenil treatment. Infect. Immun. 26, 822–826.
- Sanderson, L. , Dogruel, M., Rodgers, J., De Koning, H.P., Thomas, S.A., 2009. Pentamidine Movement across the Murine Blood-Brain and Blood-Cerebrospinal Fluid Barriers: Effect of Trypanosome Infection, Combination Therapy, P-Glycoprotein, and Multidrug Resistance-Associated Protein. J. Pharmacol. Exp. Ther. 329, 967–977.
- Schad, G.J. , Allanson, A., Mackay, S.P., Cannavan, A., Tettey, J.N.A., 2008. Development and validation of an improved HPLC method for the control of potentially counterfeit isometamidium products. J. Pharm. Biomed. Anal. 46, 45–51.
- Schillinger, D. , Röttcher, D., 1986. Treatment of camels for Trypanosoma brucei evansi infection (surra). World Anim Rev 60, 32.
- Schoenbach, E.B. , Greenspan, E.M., 1948. The pharmacology, mode of action and therapeutic potentialities of stilbamidine, pentamidine, propamidine and other aromatic diamidines—a review. Medicine (Baltimore). 27, 327–377.
- Schönefeld, A. , Röttcher, D., Moloo, S.K., 1987. The sensitivity to trypanocidal drugs of Trypanosoma vivax isolated in Kenya and Somalia. Trop. Med. Parasitol. 38, 177–80.
- Scott, A.G. , Tait, A., Michael, C., Turner, R., 1996. Characterisation of clones lines of Trypanosoma brucei expressing stable resistance to MelCy and suramin. Acta Trop. 60, 251–262.
- Scott, A.G. , Tait, A., Turner, C.M.R., 1997. Trypanosoma brucei:Lack of Cross-resistance to Melarsoprolin Vitroby Cymelarsan-Resistant Parasites. Exp. Parasitol. 86, 181–190.
- Scott, F.J. , Khalaf, A.I., Giordani, F., Wong, P.E., Duffy, S., Barrett, M., Avery, V.M., Suckling, C.J., 2016. An evaluation of Minor Groove Binders as anti-Trypanosoma brucei brucei therapeutics. Eur. J. Med. Chem. 116, 116–125.
- Seidl, A. , Dávila, A.M.R., Silva, R.A.M.S., 1999. Estimated Financial Impact of Trypanosoma vivax on the Brazilian Pantanal and Bolivian Lowlands. Mem. Inst. Oswaldo Cruz 94, 269–272.
- Sekhar, G.N. , Watson, C.P., Fidanboylu, M., Sanderson, L., Thomas, S.A., 2014. Delivery of Antihuman African Trypanosomiasis Drugs Across the Blood-Brain and Blood-CSF Barriers. In: Advances in Pharmacology. Pp. 245–275.
- Shahi, S.K. , Krauth-Siegel, R.L., Clayton, C.E., 2002. Overexpression of the putative thiol conjugate transporter TbMRPA causes melarsoprol resistance in Trypanosoma brucei. Mol. Microbiol. 43, 1129–1138.
- Shapiro, T.A. , Englund, P.T., 1990. Selective cleavage of kinetoplast DNA minicircles promoted by antitrypanosomal drugs. Proc. Natl. Acad. Sci. 87, 950–954.
- Shaw, A.P.M. , Cecchi, G., Wint, G.R.W., Mattioli, R.C., Robinson, T.P., 2014. Mapping the economic benefits to livestock keepers from intervening against bovine trypanosomosis in Eastern Africa. Prev. Vet. Med. 113, 197–210.
- Shikanai Yasuda, M.A. , 2022. Emerging and reemerging forms of Trypanosoma cruzi transmission. Mem. Inst. Oswaldo Cruz. 117, e210033.
- Singaro, J.R.S. , Yapo, F.B., Doua, F., Miezan, T.W., Baltz, T., 1996. The Efficacy of Pentamidine in the Treatment of Early-Late Stage Trypanosoma brucei gambiense Trypanosomiasis. Am. J. Trop. Med. Hyg. 55, 586–588.
- Sinyangwe, L. , Delespaux, V., Brandt, J., Geerts, S., Mubanga, J., Machila, N., Holmes, P.H., Eisler, M.C., 2004. Trypanocidal drug resistance in eastern province of Zambia. Vet. Parasitol. 119, 125–135.
- Sneader, W. , 2005. Drug Discovery, Drug Discovery: A History. John Wiley and Sons Ltd, Chichester, England.
- Sonibare, A. , Famuyide, I., Takeet, M., Oyewusi, I., Egbetade, A., Abapka, S., Adediran, O., Solanke, A., 2016. Diminazene-resistant Trypanosoma vivax in West African Dwarf lamb, south-west, Nigeria: A case report. Niger. J. Parasitol. 37, 62.
- Sow, A. , Sidibé, I., Bengaly, Z., Marcotty, T., Séré, M., Diallo, A., Vitouley, H.S., Nebié, R.L., Ouédraogo, M., Akoda, G.K., Van den Bossche, P., Van Den Abbeele, J., De Deken, R., Delespaux, V., 2012. Field detection of resistance to isometamidium chloride and diminazene aceturate in Trypanosoma vivax from the region of the Boucle du Mouhoun in Burkina Faso. Vet. Parasitol. 187, 105–111.
- Steketee, P.C. , Dickie, E.A., Iremonger, J., Crouch, K., Paxton, E., Jayaraman, S., Alfituri, O.A., Awuah-Mensah, G., Ritchie, R., Schnaufer, A., Rowan, T., De Koning, H.P., Gadelha, C., Wickstead, B., Barrett, M.P., Morrison, L.J., 2021. Divergent metabolism between Trypanosoma congolense and Trypanosoma brucei results in differential sensitivity to metabolic inhibition. PLOS Pathog. 17, e1009734.
- Steketee, P.C. , Vincent, I.M., Achcar, F., Giordani, F., Kim, D.-H., Creek, D.J., Freund, Y., Jacobs, R., Rattigan, K., Horn, D., Field, M.C., MacLeod, A., Barrett, M.P., 2018. Benzoxaborole treatment perturbs S-adenosyl-L-methionine metabolism in Trypanosoma brucei. PloS Negl. Trop. Dis. 12, e0006450.
- Stephen, L.E. , 1958. Suramin Complexes. Ann. Trop. Med. Parasitol. 52, 417–426.
- Stephen, L.E. , 1962. Suramin Complexes. Ann. Trop. Med. Parasitol. 56, 406–414.
- Stevens, L. , Dorn, P.L., Schmidt, J.O., Klotz, J.H., Lucero, D., Klotz, S.A., 2011. Kissing Bugs. The Vectors of Chagas. In: Advances in Parasitology. Pp. 169–192.
- Steverding, D. , 2010. The development of drugs for treatment of sleeping sickness: a historical review. Parasit. Vectors 3, 15.
- Steverding, D. , Antoszczak, M., Huczyński, A., 2016. In vitro activity of salinomycin and monensin derivatives against Trypanosoma brucei. Parasit. Vectors 9, 409.
- Steverding, D. , Troeberg L., 2024. 100 years since the publication of the suramin formulation. Parasitol Res. 123:11.
- Stewart, M.L. , Burchmore, R.J.S., Clucas, C., Hertz-Fowler, C., Brooks, K., Tait, A., MacLeod, A., Turner, C.M.R., De Koning, H.P., Wong, P.E., Barrett, M.P., 2010. Multiple Genetic Mechanisms Lead to Loss of Functional TbAT1 Expression in Drug-Resistant Trypanosomes. Eukaryot. Cell 9, 336–343.
- Stewart, M.L. , Krishna, S., Burchmore, R.J.S., Brun, R., De Koning, H.P., Boykin, D.W., Tidwell, R.R., Ed Hall, J., Barrett, M.P., 2005. Detection of arsenical drug resistance in Trypanosoma brucei with a simple fluorescence test. Lancet 366, 486–487.
- Stijlemans, B. , De Baetselier, P., Caljon, G., Van Den Abbeele, J., Van Ginderachter, J.A., Magez, S., 2017. Nanobodies As Tools to Understand, Diagnose, and Treat African Trypanosomiasis. Front. Immunol. 8, 724.
- Strober, W. , 2015. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 111, A3.B.1–A3.B.3.
- Suganuma, K. , N’Da, D.D., Watanabe, K., Tanaka, Y., Mossaad, E., Elata, A., Inoue, N., Kawazu, S., 2022. Therapeutic Efficacy of Orally Administered Nitrofurantoin against Animal African Trypanosomosis Caused by Trypanosoma congolense Infection. Pathogens 11, 331.
- Summers, J.M. , Hawkins, S.E., 1978. The effect of antrycide of patterns of RNA synthesis in Amoeba discoides. J. Cell Sci. 30, 237–250.
- Suswam, E.A. , Taylor, D.W., Ross, C.A., Martin, R.J., 2001. Changes in properties of adenosine transporters in Trypanosoma evansi and modes of selection of resistance to the melaminophenyl arsenical drug, Mel Cy. Vet. Parasitol. 102, 193–208.
- Sutcliffe, O.B. , Skellern, G.G., Araya, F., Cannavan, A., Sasanya, J.J., Dungu, B., Van Gool, F., Munstermann, S., Mattioli, R.C., 2014. Animal trypanosomosis: making quality control of trypanocidal drugs possible. Rev. Sci. Tech. L’OIE 33, 813–830.
- Sutherland, C.S. , Stone, C.M., Steinmann, P., Tanner, M., Tediosi, F., 2017. Seeing beyond 2020: an economic evaluation of contemporary and emerging strategies for elimination of Trypanosoma brucei gambiense. Lancet Glob. Heal. 5, e69–e79.
- Sutherland, I.A. , Holmes, P.H., 1993. Alterations in drug transport in resistant Trypanosoma congolense. Acta Trop. 54, 271–278.
- Sutherland, I.A. , Mounsey, A., Eisler, M., Holmes, P.H., 1992a. Kinetic modelling of isometamidium chloride (Samorin) uptake by Trypanosoma congolense. Parasitology 105, 91–95.
- Sutherland, I.A. , Mounsey, A., Holmes, P.H., 1992b. Transport of isometamidium (Samorin) by drug-resistant and drug-sensitive Trypanosoma congolense. Parasitology 104, 461–467.
- Tarello, W. , 2005. Trypanosoma evansi infection in three cats. Rev. Med. Vet. (Toulouse). 156, 133–134.
- Taylor, K. , Authié, E.M.-L., 2004. Pathogenesis of animal trypanosomiasis. CAB International, Wallingford, UK.
- Tchamdja, E. , Clausen, P.-H., Kulo, A.E., Batawui, K., Bauer, B., Den Abbeele, J. Van, Delespaux, V., Hoppenheit, A., 2019. How rational drug use reduces trypanosome infections in cattle in chemo-resistance hot-spot villages of northern Togo. Acta Trop. 190, 159–165.
- Tchamdja, E. , Kulo, A.E., Akoda, K., Teko-Agbo, A., Assoumy, A.M., Niang, E.M.M., Batawui, K., Adomefa, K., Bankolé, A.A., Kombiagou, K., Hoppenheit, A., Clausen, P.-H., Mattioli, R.C., Peter, R., Napier, G.B., De Deken, R., Marcotty, T., Van Den Abbeele, J., Delespaux, V., 2016. Drug quality analysis through high performance liquid chromatography of isometamidium chloride hydrochloride and diminazene diaceturate purchased from official and unofficial sources in Northern Togo. Prev. Vet. Med. 126, 151–158.
- Teka, I.A. , Kazibwe, A.J.N., El-Sabbagh, N., Al-Salabi, M.I., Ward, C.P., Eze, A.A., Munday, J.C., Mäser, P., Matovu, E., Barrett, M.P., De Koning, H.P., 2011. The Diamidine Diminazene Aceturate Is a Substrate for the High-Affinity Pentamidine Transporter: Implications for the Development of High Resistance Levels in Trypanosomes. Mol. Pharmacol. 80, 110–116.
- Thomas, J.A. , Baker, N., Hutchinson, S., Dominicus, C., Trenaman, A., Glover, L., Alsford, S., Horn, D., 2018. Insights into antitrypanosomal drug mode-of-action from cytology-based profiling. PloS Negl. Trop. Dis. 12, e0006980.
- Thuita, J.K. , Wolf, K.K., Murilla, G.A., Bridges, A.S., Boykin, D.W., Mutuku, J.N., Liu, Q., Jones, S.K., Gem, C.O., Ching, S. and Tidwell, R.R., 2015. Chemotherapy of second stage human African trypanosomiasis: comparison between the parenteral diamidine DB829 and its oral prodrug DB868 in vervet monkeys. PLoS Negl. Trop. Dis. 9, e0003409.
- Tihon, E. , Imamura, H., Van den Broeck, F., Vermeiren, L., Dujardin, J.-C., Van Den Abbeele, J., 2017. Genomic analysis of Isometamidium Chloride resistance in Trypanosoma congolense. Int. J. Parasitol. Drugs Drug Resist. 7, 350–361.
- Touratier, L. , 1992. Eleventh International meeting on Trypanosoma evansi : report of the working group. Rev. Sci. Tech. l’OIE 11, 275–304.
- Travis, A.S. , 1991. A hundred years of chemotherapy 1891-1991. Biochem. (Lond). 13, 9–12.
- Truc, P. , Büscher, P., Cuny, G., Gonzatti, M.I., Jannin, J., Joshi, P., Juyal, P., Lun, Z.-R., Mattioli, R., Pays, E., Simarro, P.P., Teixeira, M.M.G., Touratier, L., Vincendeau, P., Desquesnes, M., 2013. Atypical human infections by animal trypanosomes. PloS Negl. Trop. Dis. 7, e2256.
- Truc, P. , Jamonneau, V., N’Guessan, P., N’Dri, L., Diallo, P.B., Cuny, G., 1998. Trypanosoma brucei, m: T. congolense; 92.
- Tsuruo, T. , Iida, H., Nojiri, M., Tsukagoshi, S., Sakurai, Y., 1983. Circumvention of vincristine and Adriamycin resistance in vitro and in vivo by calcium influx blockers. Cancer Res. 43, 2905–10.
- Tsuruo, T. , lida, H., Tsukagoshi, S., Sakurai, Y., 1981. Overcoming of Vincristine Resistance in p388 Leukemia in Vivo and in Vitro Through Enhanced Cytotoxicity of Vincristine and Vinblastine by Verapamil. Cancer Res. 41, 1967–1972.
- Umar, I.A. , Rumah, B.L., Kamla, S.L., Jobin, Ibrahim, Isah, M.A., 2008. Effects of intraperitoneal administration of vitamins C and E or A and E combinations on the severity of Trypanosoma brucei brucei infection in rats. African J. Biochem. Res. 2, 88–91.
- Umeakuana, P.U. , Gibson, W., Ezeokonkwo, R.C., Anene, B.M, 2019. Identification of Trypanosoma brucei gambiense in naturally infected dogs in Nigeria. Parasit. Vectors 12, 420.
- Unciti-Broceta, J.D. , Arias, J.L., Maceira, J., Soriano, M., Ortiz-González, M., Hernández-Quero, J., Muñóz-Torres, M., De Koning, H.P., Magez, S., Garcia-Salcedo, J.A., 2015. Specific Cell Targeting Therapy Bypasses Drug Resistance Mechanisms in African Trypanosomiasis. PLOS Pathog. 11, e1004942.
- Ungogo, M.A. , Aldfer, M.M., Natto, M.J., Zhuang, H., Chisholm, R., Walsh, K., Mcgee, M., Ilbeigi, K., Asseri, J.I., Burchmore, R.J.S., Caljon, G., Van, S., De Koning, H.P., 2023. Cloning and characterization of Trypanosoma congolense and T. vivax nucleoside transporters reveal the potential of P1-type carriers for the discovery of broad-spectrum nucleoside-based therapeutics against Animal African trypanosomiasis. Int. J. Mol. Sci. 24, 3144.
- Ungogo, M.A. , Campagnaro, G.D., Alghamdi, A.H., Natto, M.J., De Koning, H.P., 2022. Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals. Int. J. Mol. Sci. 23, 2844.
- Uzonna, J.E. , Kaushik, R.S., Gordon, J.R., Tabel, H., 1999. Cytokines and antibody responses during Trypanosoma congolense infections in two inbred mouse strains that differ in resistance. Parasite Immunol. 21, 57–71.
- Van den Bossche, P. , Doran, M., Connor, R.J., 2000. An analysis of trypanocidal drug use in the Eastern Province of Zambia. Acta Trop. 75, 247–258.
- Van Hoof, L. , Henrard, C., Peel, E., 1944. Pentamidine in the prevention and treatment of trypanosomiasis. Trans. R. Soc. Trop. Med. Hyg. 37, 271–280.
- Vanhollebeke, B. , Truc, P., Poelvoorde, P., Pays, A., Joshi, P.P., Katti, R., Jannin, J.G., Pays, E., 2006. Human Trypanosoma evansi Infection Linked to a Lack of Apolipoprotein L-I. N. Engl. J. Med. 355, 2752–2756.
- Vansterkenburg, E.L.M. , Coppens, I., Wilting, J., Bos, O.J.M., Fischer, M.J.E., Janssen, L.H.M., Opperdoes, F.R., 1993. The uptake of the trypanocidal drug suramin in combination with low-density lipoproteins by Trypanosoma brucei and its possible mode of action. Acta Trop. 54, 237–250.
- Vitouley, H.S. , Mungube, E.O., Allegye-Cudjoe, E., Diall, O., Bocoum, Z., Diarra, B., Randolph, T.F., Bauer, B., Clausen, P.-H., Geysen, D., Sidibe, I., Bengaly, Z., Van den Bossche, P., Delespaux, V., 2011. Improved PCR-RFLP for the Detection of Diminazene Resistance in Trypanosoma congolense under Field Conditions Using Filter Papers for Sample Storage. PloS Negl. Trop. Dis. 5, e1223.
- Vodnala, S.K. , Lundbäck, T., Yeheskieli, E., Sjöberg, B., Gustavsson, A.-L., Svensson, R., Olivera, G.C., Eze, A.A., De Koning, H.P., Hammarström, L.G.J., Rottenberg, M.E., 2013. Structure–Activity Relationships of Synthetic Cordycepin Analogues as Experimental Therapeutics for African Trypanosomiasis. J. Med. Chem. 56, 9861–9873.
- Vreysen, M.J.B. , Seck, M.T., Sall, B., Bouyer, J., 2013. Tsetse flies: Their biology and control using area-wide integrated pest management approaches. J. Invertebr. Pathol. 112, S15–S25.
- W/yohannes, D. , Kebede, E., Abebe, G., 2010. Study on the assessment of drug resistance on Trypanosoma vivax in Tselemti woreda, Tigray, Ethiopia. Ethiop. Vet. J. 14, 15–30.
- Wainwright, M. , 2010. Dyes, trypanosomiasis and DNA: a historical and critical review. Biotech. Histochem. 85, 341–354.
- Waitumbi, J.N. , Murphy, N.B., Peregrine, A.S., 1994. Genotype and drug-resistance phenotype of Trypanosoma evansi isolated from camels in northern Kenya. Ann. Trop. Med. Parasitol. 88, 677–683.
- Wall, R.J. , Rico, E., Lukac, I., Zuccotto, F., Elg, S., Gilbert, I.H., Freund, Y., Alley, M.R.K., Field, M.C., Wyllie, S., Horn, D., 2018. Clinical and veterinary trypanocidal benzoxaboroles target CPSF3. Proc. Natl. Acad. Sci. 115, 9616–9621.
- Wangwe, I.I. , Wamwenje, S.A., Mirieri, C., Masila, N.M., Wambua, L., Kulohoma, B.W., 2019. Modelling appropriate use of trypanocides to restrict wide-spread multi-drug resistance during chemotherapy of animal African trypanosomiasis. Parasitology 146, 774–780.
- Ward, C.P. , Wong, P.E., Burchmore, R.J., De Koning, H.P., Barrett, M.P., 2011. Trypanocidal Furamidine Analogues: Influence of Pyridine Nitrogens on Trypanocidal Activity, Transport Kinetics, and Resistance Patterns. Antimicrob. Agents Chemother. 55, 2352–2361.
- Watkins, T.I. , Woolfe, G., 1952. Effect of Changing the Quaternizing Group on the Trypanocidal Activity of Dimidium Bromide. Nature 169, 506–506.
- Whiteside, E.F. , 1962. Interactions between drugs, trypanosoomes and cattle in the field. In: Drugs, Parasites and Hosts. In: Goodwin, L.G., Nimmo-Smith.J., R.H., Churchill, A. (Eds.), Drugs, Parasites and Hosts. London, pp. 116–141.
- Wiedemar, N. , Graf, F.E., Zwyer, M., Ndomba, E., Kunz Renggli, C., Cal, M., Schmidt, R.S., Wenzler, T., Mäser, P., 2018. Beyond immune escape: a variant surface glycoprotein causes suramin resistance in Trypanosoma brucei. Mol. Microbiol. 107, 57–67.
- Wiedemar, N. , Hauser, D.A., Mäser, P., 2020. 100 Years of Suramin. Antimicrob. Agents Chemother. 64(3), e01168-19.
- Wiedemar, N. , Zwyer, M., Zoltner, M., Cal, M., Field, M.C., Mäser, P., 2019. Expression of a specific variant surface glycoprotein has a major impact on suramin sensitivity and endocytosis in Trypanosoma brucei. FASEB BioAdvances 1, 595–608.
- Wilkes, J.M. , Mulugeta, W., Wells, C., Peregrine, A.S., 1997. Modulation of mitochondrial electrical potential: a candidate mechanism for drug resistance in African trypanosomes. Biochem. J. 326, 755–761.
- Wilkes, J.M. , Peregrine, A.S., Zilberstein, D., 1995. The accumulation and compartmentalization of isometamidium chloride in Trypanosoma congolense, monitored by its intrinsic fluorescence. Biochem. J. 312, 319–327.
- Wilkowsky, S.E. , 2018. Trypanosoma. In: Parasitic Protozoa of Farm Animals and Pets. Springer International Publishing, Cham, pp. 271–287.
- Williamson, J. , 1957. Suramin complexes. I. Prophylactic activity against Trypanosoma congolense in small animals. Ann Trop Med Parasitol 51, 440–456.
- Williamson, J. , 1970. Review of chemotherapeutic and chemotherapeutic agents. In: Mulligan, H.W. (Ed.), The African Trypanosomiases. George Allen and Unwin Ltd, London, pp. 684–710.
- Williamson, J. , Desowitz, R.S., 1956. Prophylactic Activity of Suramin Complexes in Animal Trypanosomiasis. Nature 177, 1074–1075.
- Williamson, J. , March, J.C., Scott-Finnigan, T.J., 1982. Drug synergy in experimental African trypanosomiasis. Tropenmed. Parasitol. 33, 76–82.
- Williamson, J. , Rollo, I.M., 1959. Drug resistance in trypanosomes; cross-resistance analyses. Br. J. Pharmacol. Chemother. 14, 423–430.
- Willson, M. , Callens, M., Kuntz, D.A., Perié, J., Opperdoes, F.R., 1993. Synthesis and activity of inhibitors highly specific for the glycolytic enzymes from Trypanosoma brucei. Mol. Biochem. Parasitol. 59, 201–210.
- Witola, W.H. , Inoue, N., Ohashi, K., Onuma, M., 2004. RNA-interference silencing of the adenosine transporter-1 gene in Trypanosoma evansi confers resistance to diminazene aceturate. Exp. Parasitol. 107, 47–57.
- World Health Organization (WHO), 2013. Control and surveillance of human African trypanosomiasis. World Health Organ. Tech. Rep. Ser. 1–237.
- Wragg, W.R. , Washbourn, K., Brown, K.N., Hill, J., 1958. Metamidium: a New Trypanocidal Drug. Nature 182, 1005–1006.
- Wring, S. , Gaukel, E., Nare, B., Jacobs, R., Beaudet, B., Bowling, T., Mercer, L., Bacchi, C., Yarlett, N., Randolph, R., Parham, R., Rewerts, C., Platner, J., Don, R., 2014. Pharmacokinetics and pharmacodynamics utilizing unbound target tissue exposure as part of a disposition-based rationale for lead optimization of benzoxaboroles in the treatment of Stage 2 Human African Trypanosomiasis. Parasitology 141, 104–118.
- Yongsheng, Y. , Yongchun, O., Chengmai, R., Yuanguo, C. and Fenqin, Z., 1996. Trypanocidal value of liposomal diminazene in experimental Trypanosoma brucei evansi infection in mice. Vet. Parasitol. 61, 349–352.
- Yaro, M. , Munyard, K.A., Stear, M.J., Groth, D.M., 2016. Combatting African Animal Trypanosomiasis (AAT) in livestock: The potential role of trypanotolerance. Vet. Parasitol. 225, 43–52.
- Yesufu, H.M. , 1971. Experimental transmission of Trypanosoma gambiense to domestic animals. Ann. Trop. Med. Parasitol. 65, 341–347.
- Yorke, W. , Adams, A.R.D., Murgatroyd, F., 1929. Studies in Chemotherapy I. A Method for Maintaining Pathogenic Trypanosomes Alive in Vitro at 37° C. for 24 Hours. Ann. Trop. Med. Parasitol. 23, 501–518.
- Zeelen, J. , van Straaten, M., Verdi, J., Hempelmann, A., Hashemi, H., Perez, K., Jeffrey, P.D., Hälg, S., Wiedemar, N., Mäser, P., Papavasiliou, F.N., Stebbins, C.E., 2021. Structure of trypanosome coat protein VSGsur and function in suramin resistance. Nat. Microbiol. 6, 392–400.
- Zhang, Z.Q. , Giroud, C., Baltz, T., 1993. Trypanosoma evansi: In Vivo and in Vitro Determination of Trypanocide Resistance Profiles. Exp. Parasitol. 77, 387–394.
- Zhou, J. , Shen, J., Liao, D., Zhou, Y., Lin, J., 2004. Resistance to drug by different isolates Trypanosoma evansi in China. Acta Trop. 90, 271–275.
- Zimmermann, S. , Hall, L., Riley, S., Sørensen, J., Amaro, R.E., Schnaufer, A., 2016. A novel high-throughput activity assay for the Trypanosoma brucei editosome enzyme REL1 and other RNA ligases. Nucleic Acids Res. 44, e24–e24.
- Zoltner, M. , Campagnaro, G.D., Taleva, G., Burrell, A., Cerone, M., Leung, K.-F., Achcar, F., Horn, D., Vaughan, S., Gadelha, C., Zíková, A., Barrett, M.P., De Koning, H.P., Field, M.C., 2020. Suramin exposure alters cellular metabolism and mitochondrial energy production in African trypanosomes. J. Biol. Chem. 295, 8331–8347.
- Zoltner, M. , Horn, D., De Koning, H.P., Field, M.C., 2016. Exploiting the Achilles’ heel of membrane trafficking in trypanosomes. Curr. Opin. Microbiol. 34, 97–103.
Figure 1.
Illustration of drug action and resistance mechanisms in trypanosomes. A. Drug importer such as P2/AT1 transporter for diamidines. B. Drug efflux pump such as multidrug resistance-associated protein (MRAP) in resistance to melaminophenyl arsenicals. C. Mechanisms that relate to delivery and accumulation in or efflux of drug from target organelle such as mitochondrion which houses the kinetoplast, which in itself is a target of multiple trypanocides. E. Surface binding protein, endocytic mechanism or a transporter that delivers endocytosed drug to the cell membrane such as a lysosomal transporter in suramin action. F. Target organelle or molecule that drug binds to. Disruption of these drug transport or action mechanisms may result in drug resistance.
Figure 1.
Illustration of drug action and resistance mechanisms in trypanosomes. A. Drug importer such as P2/AT1 transporter for diamidines. B. Drug efflux pump such as multidrug resistance-associated protein (MRAP) in resistance to melaminophenyl arsenicals. C. Mechanisms that relate to delivery and accumulation in or efflux of drug from target organelle such as mitochondrion which houses the kinetoplast, which in itself is a target of multiple trypanocides. E. Surface binding protein, endocytic mechanism or a transporter that delivers endocytosed drug to the cell membrane such as a lysosomal transporter in suramin action. F. Target organelle or molecule that drug binds to. Disruption of these drug transport or action mechanisms may result in drug resistance.
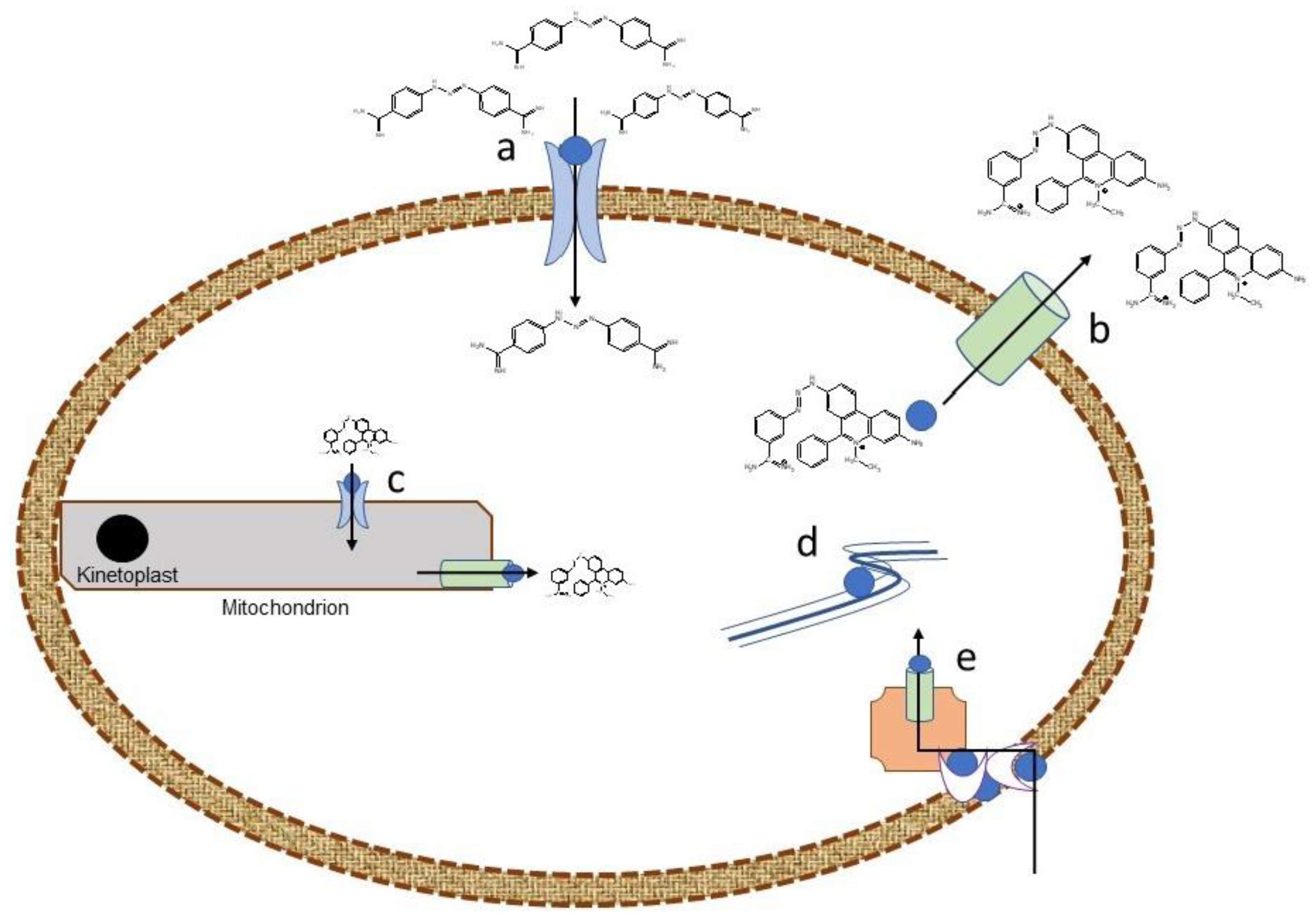
Figure 2A.
Structures of synthalin and the main diamidine drugs against trypanosomiasis.
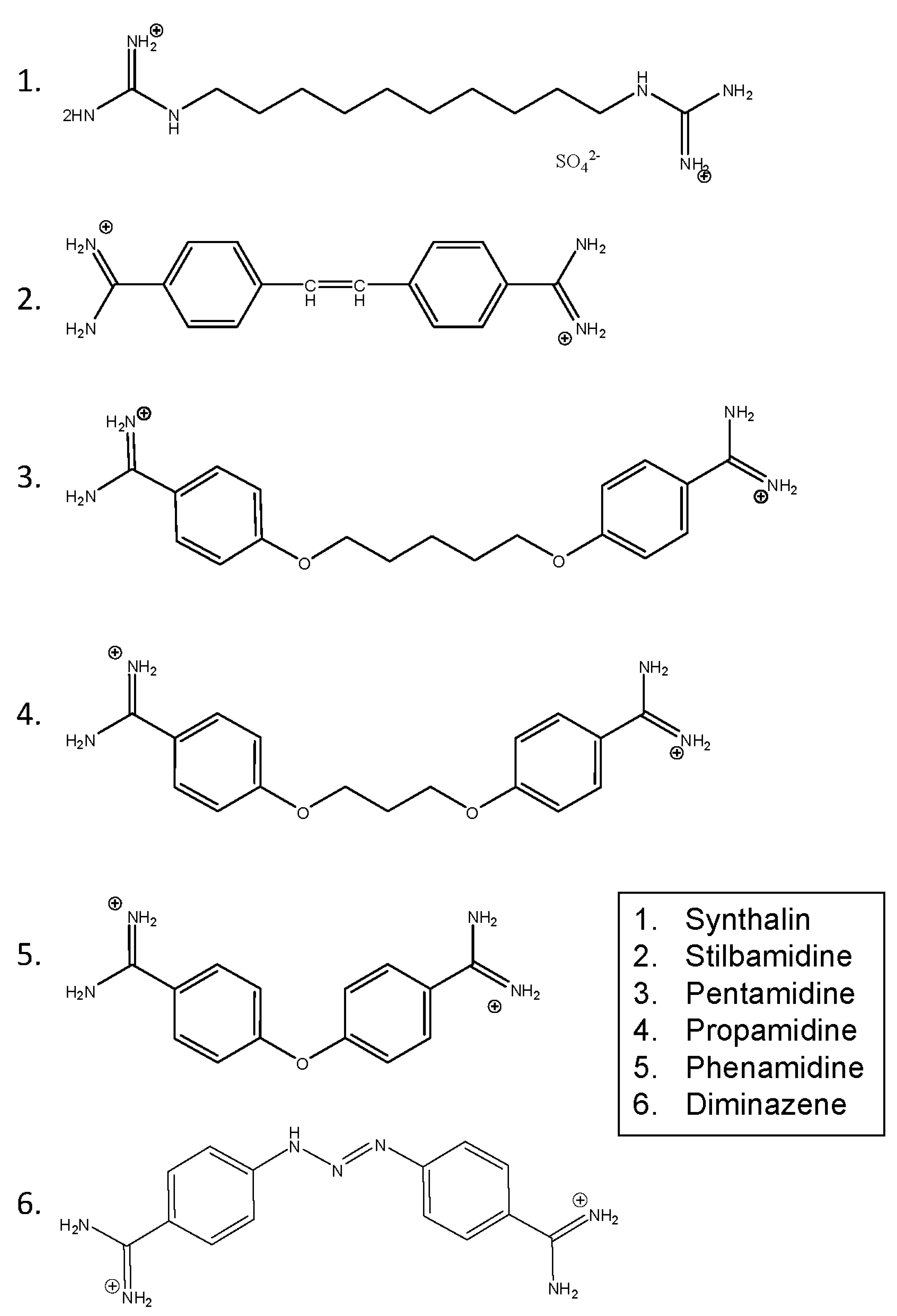
Figure 2B.
Structures of the main furamidines.
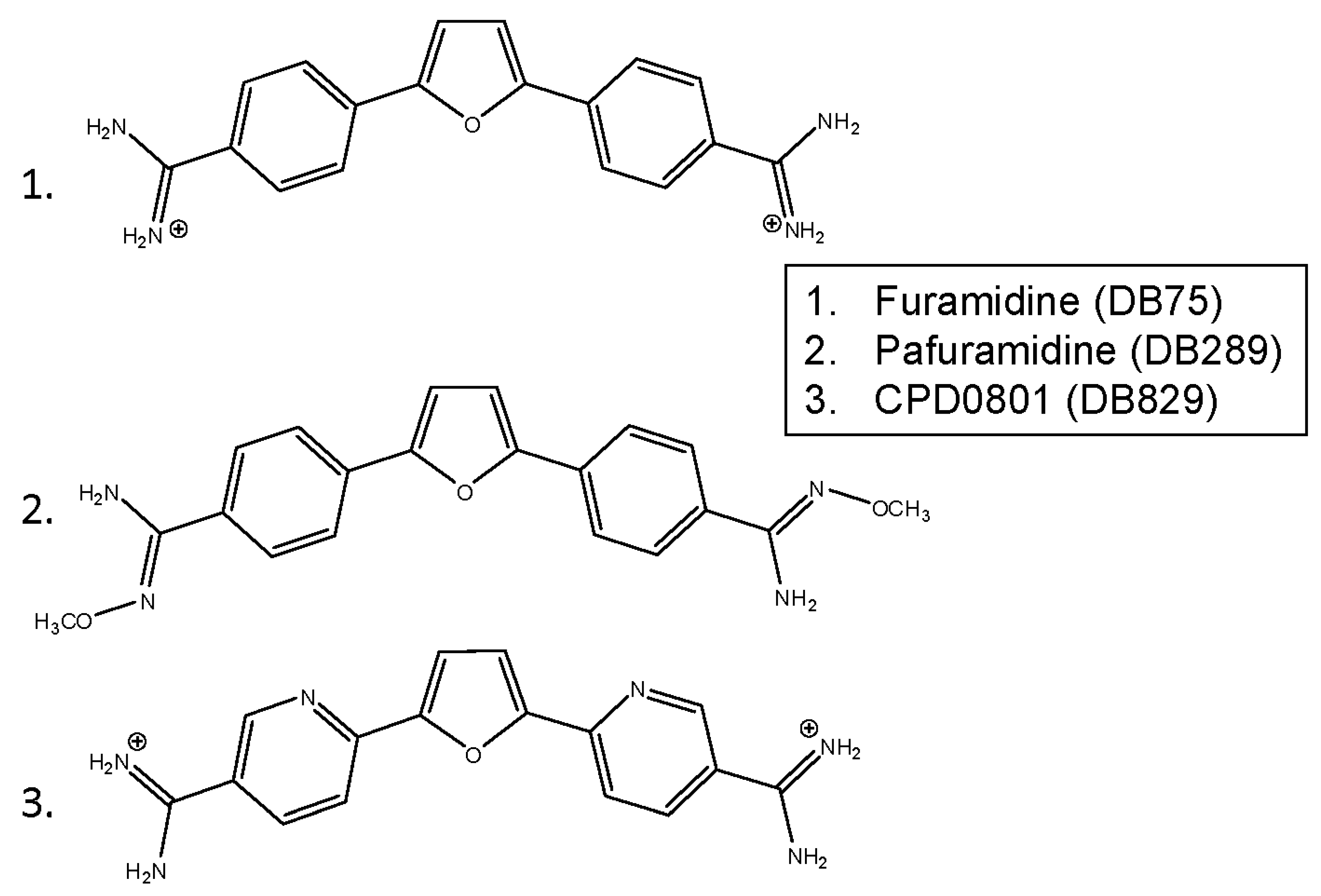
Figure 3.
Structures of an early flavin trypanocide, trypaflavine, and the phenanthridine drugs that were developed therefrom.
Figure 3.
Structures of an early flavin trypanocide, trypaflavine, and the phenanthridine drugs that were developed therefrom.
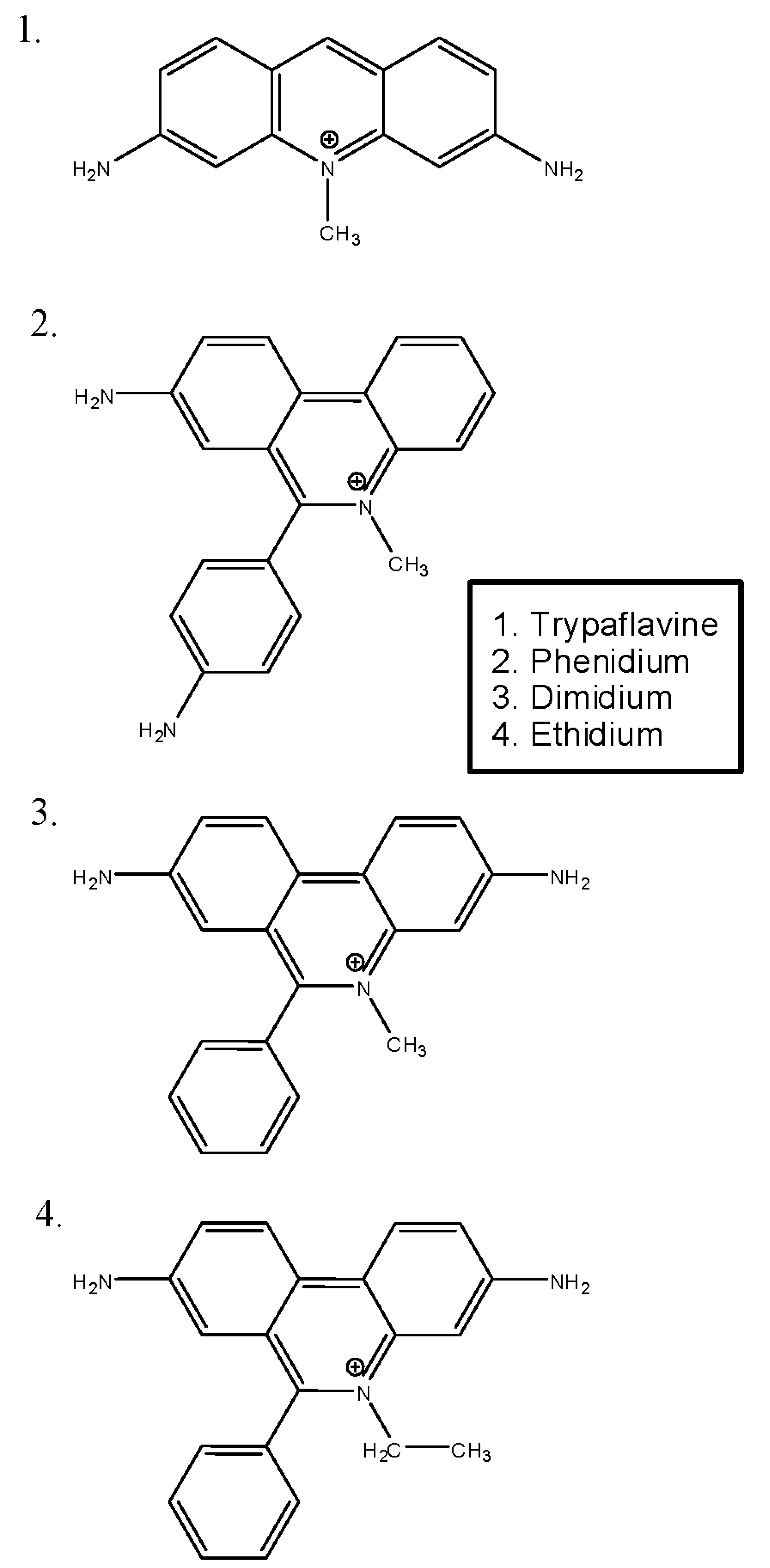
Figure 4.
Structures of the main constituents of commercial isometamidium (Samorin). ISM, isometamidium, generally believed to be the (most) active agent in the mixture. The original May & Baker numbers are given.
Figure 4.
Structures of the main constituents of commercial isometamidium (Samorin). ISM, isometamidium, generally believed to be the (most) active agent in the mixture. The original May & Baker numbers are given.
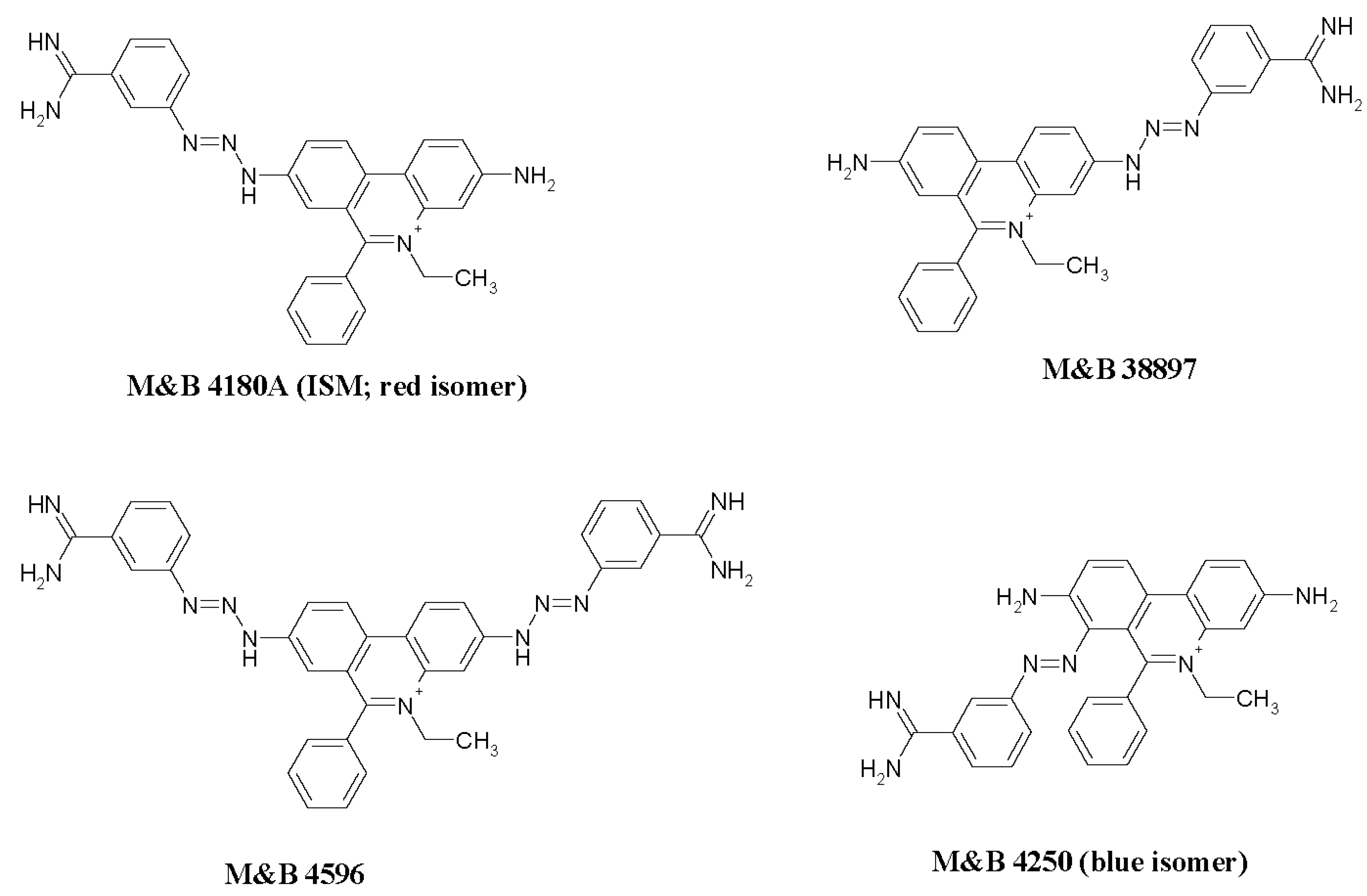
Figure 5.
Structure of suramin and the early trypanocidal dyes from which suramin was ultimately developed.
Figure 5.
Structure of suramin and the early trypanocidal dyes from which suramin was ultimately developed.
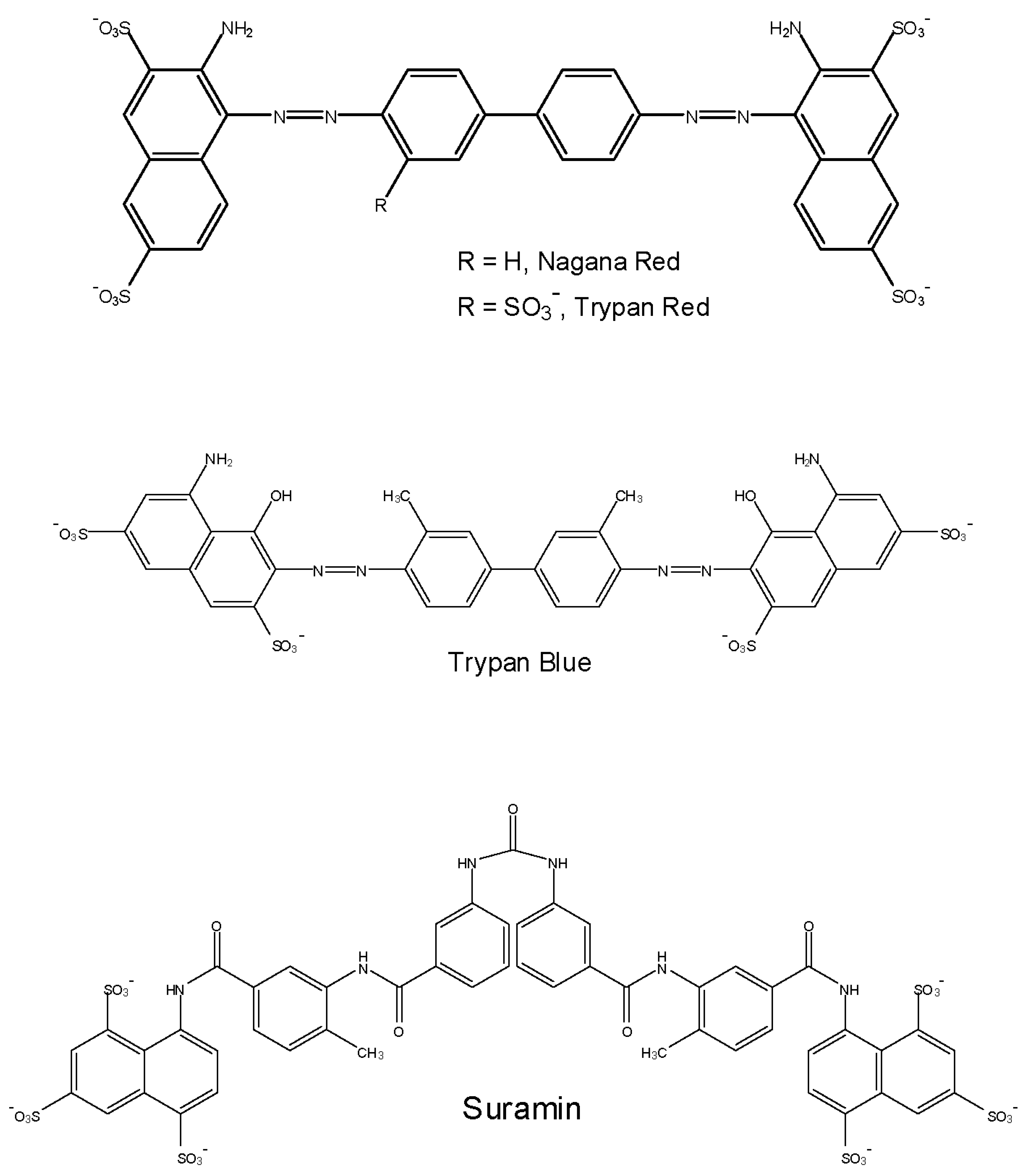
Figure 6.
Structures of some of the arsenical drugs that have been (1, 2) or are still used against trypanosomiasis (3, 4). M, melaminophenyl group. BAL, British anti-Lewisite (dimercaprol).
Figure 6.
Structures of some of the arsenical drugs that have been (1, 2) or are still used against trypanosomiasis (3, 4). M, melaminophenyl group. BAL, British anti-Lewisite (dimercaprol).
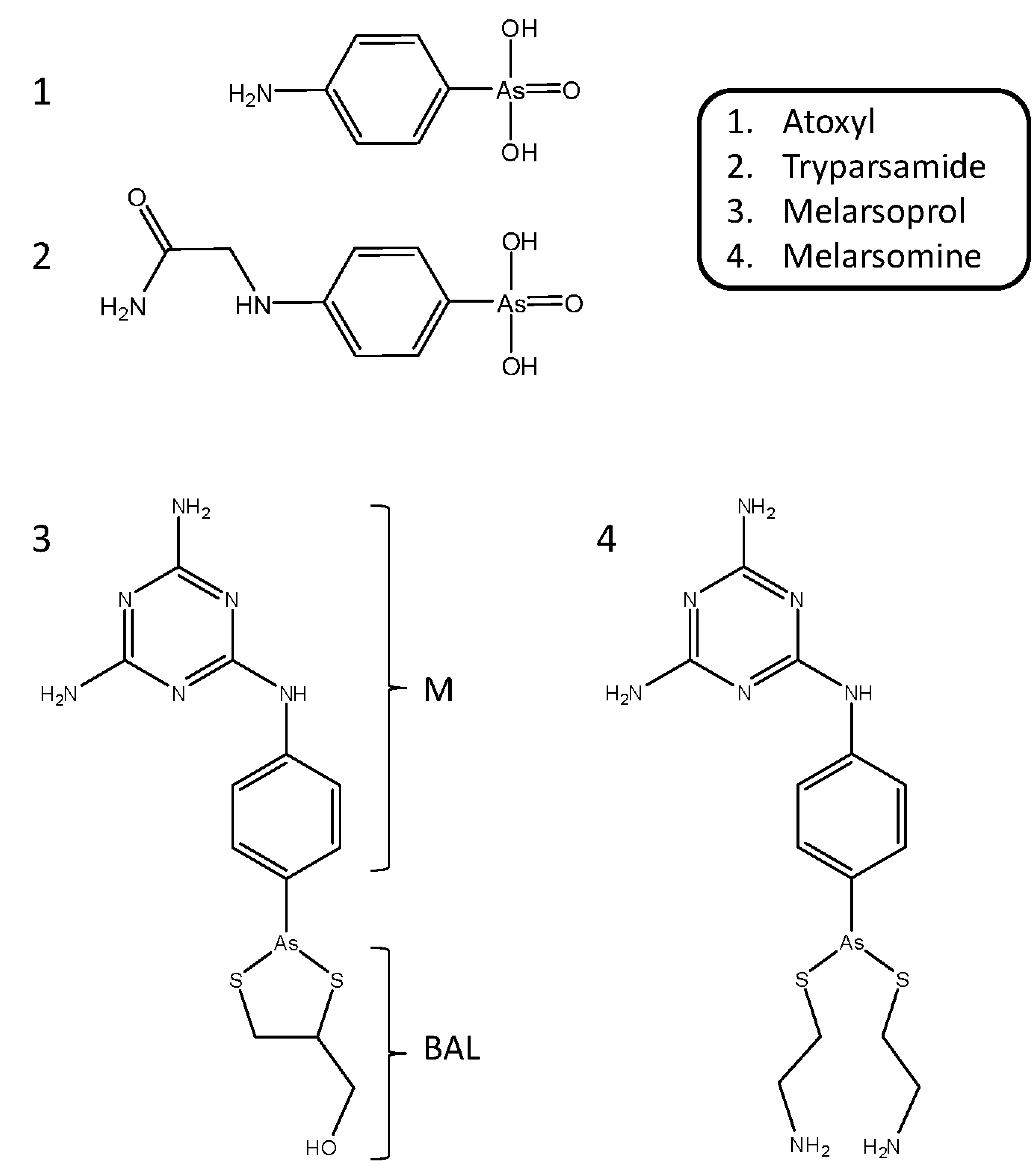
Figure 7.
Structure of quinapyramine.
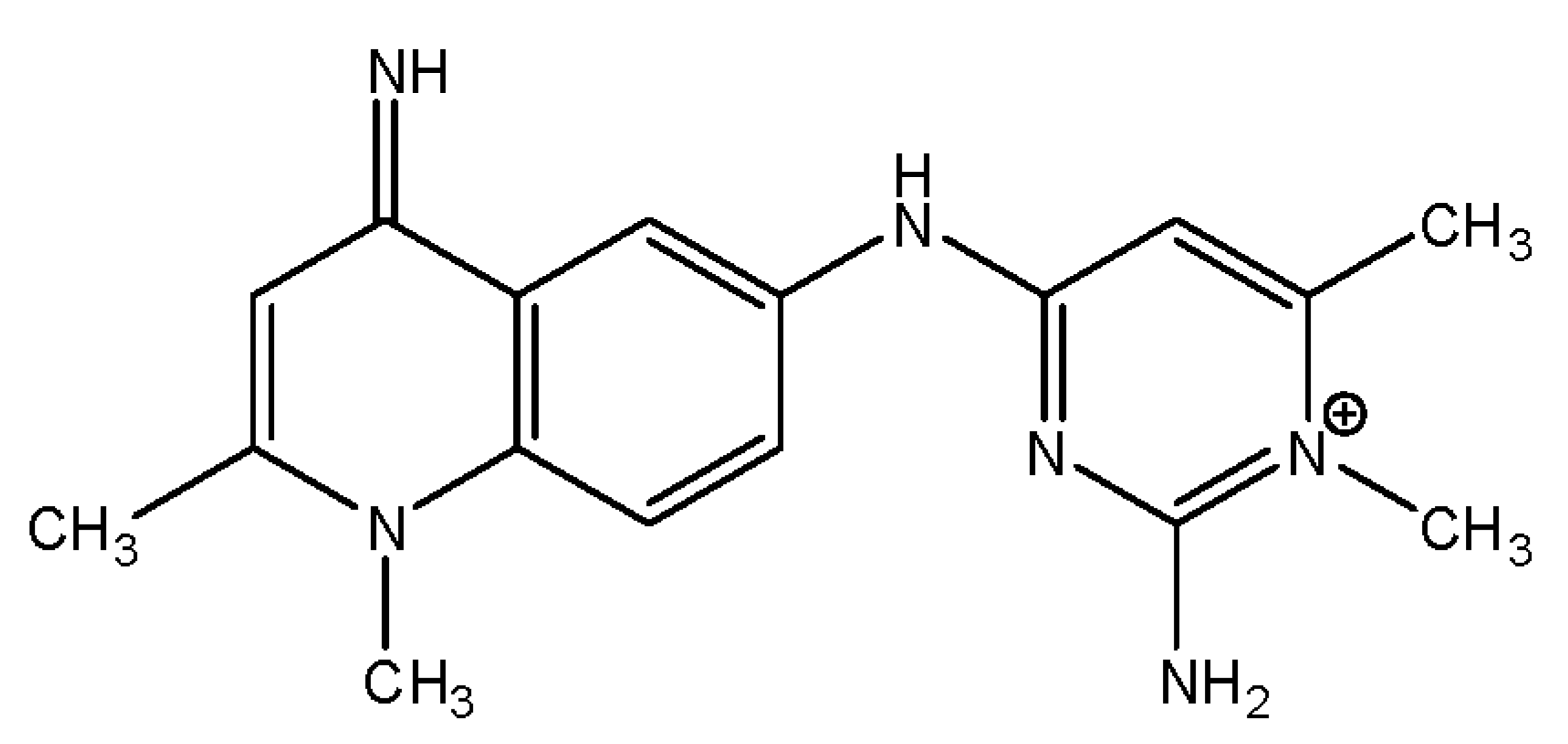
Figure 8.
Structures of two oxaboroles proposed as veterinary trypanocides.
Table 1.
Cross-resistance pattern in veterinary trypanosomes with laboratory generated drug resistance.
Table 1.
Cross-resistance pattern in veterinary trypanosomes with laboratory generated drug resistance.
| Drug resistance developed | Species | Type of study | Drugs Cross-resistant to | Drugs not cross-resistant to | Reference |
|---|---|---|---|---|---|
| Melarsomine | T. evansi | In vivo (mice) | Diminazene and pentamidine | Suramin, isometamidium | Osman et al., 1992 |
| T. evansi | In vitro | Diminazene and Arsobal | Suramin and antrycide | Ross and Barn, 1996 | |
| T. brucei | In vivo (mice) | Melarsoprol | Suramin | Scott et al., 1996 | |
| T. brucei | In vivo (mice) | Melarsoprol (in vivo only), trimelarsen, melarsen oxide diminazene, pentamidine | Melarsoprol in vitro | Scott et al., 1997 | |
| T. evansi | In vitro and in vivo | Downregulation of diminazene transporter P2/AT1 | Suswam et al., 2001 | ||
| Tryparsamide | T. evansi | In vivo | Oxophenarsine | All others | Gill, 1971 |
| Diminazene | T. evansi | In vivo | Melarsomine | Zhang et al., 1993 | |
| T. brucei | In vitro | Pentamidine, furamidine, and melarsomine | Isometamidium, AN7973 and SCYX7158. | Teka et al., 2011 | |
| T. congolense | In vitro | -- | Isometamidium, AN7973 and SCYX7158. | Carruthers et al., 2021 | |
| Stilbamidine (diamidine) | T. evansi | In vivo | Oxophenarsine (aromatic arsenical), Melarsoprol, MsB, acriflavine, tryparsamide, diminazene, and metamidium | Quinapyramine, suramin | Gill, 1971 |
| Isometamidium | T. congolense | Diminazene, homidium chloride, quinapyramine sulphate | Diminazene | Peregrine et al., 1997 | |
| T. brucei | In vitro | Ethidium bromide, diminazene, and pentamidine | - | Eze et al., 2016 | |
| T. congolense | In vivo | - | - | Tihon et al., 2017 | |
| Suramin | T. evansi | In vivo | ---- | Oxophenarsine (aromatic arsenical), Melarsoprol, MsB, acriflavine, tryparsamide, diminazene, metamidium, quinapyramine | Gill, 1971 |
| T. evansi | In vivo | ---- | --- | Mutugi et al., 1994) | |
| T. brucei | In vivo | ---- | Melarsomine | Scott et al., 1996 | |
| T. brucei | In vivo | Trypan blue | Melarsoprol, Pentamidine | Wiedemar et al., 2018 | |
| Quinapyramine | T. equiperdum | In vivo | Dimidium and stilbamidine | Suramin | Ormerod et al., 1952 |
| T. evansi | In vivo | None | Oxophenarsine (aromatic arsenical), Melarsoprol, MsB, acriflavine, tryparsamide, diminazene, metamidium, suramin | Gill, 1971 | |
| T. congolense | In vivo (mice) | Diminazene, isometamidium, homidium | - | Ndoutamia et al., 1993 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



