
Submitted:
18 January 2024
Posted:
19 January 2024
You are already at the latest version
Abstract
The orexin/hypocretin neuropeptide family has emerged as a focal point of neuroscientific research following the discovery that this family plays a crucial role in a variety of physiological and behavioral processes. These neuropeptides serve as powerful neuromodulators, intricately shaping autonomic, endocrine, and behavioral responses across species. Notably, they serve as master regulators of vigilance and stress responses; however, their role in complex processes such as food intake, metabolism, and thermoregulation warrant further investigation. This narrative review provides a journey through the evolution of our understanding of the orexin system, from its initial discovery to the promising progress made in developing orexin derivatives. It goes beyond conventional boundaries, striving to synthesize the multifaceted activities of orexins. Special emphasis is placed on domains such as stress response, fear, anxiety, and learning, in which the authors contributed to the literature with original publications. This article demonstrates the orchestrating role of the orexins in several intricate and integrative physiological processes. It also overviews the advancement of orexin pharmacology, which has already yielded some promising successes, particularly in the treatment of sleep disorders. This interdisciplinary approach to studying neuropeptides unveils their significance as vital connectors between transient functional changes and lasting structural adaptations. Ultimately, it advances our understanding of the intricate physiological and behavioral contributions of the orexin/hypocretin neuropeptide family.
Keywords:
orexins
; neuropeptides
; neurotransmitters
; stress
; appetite: body temperature regulation
; fear
; anxiety
; learning
; sleep wake disorders
1. Introduction: the neuropeptides as the modulators of the connectome
Neuropeptide research started more than a hundred years ago with the discovery of the effects of vasopressin [1], oxytocin [2], and the tissue extraction of the first classical neuropeptide: substance-P [3]. Since then, more and more distinct features of neuropeptides have been identified that clearly separate them from the classical neurotransmitters [4,5,6]. First, some obvious biochemical features justify the differentiation. They are much larger molecules than the classical neurotransmitters, therefore the energy requirement of their synthesis and transport well exceed that of the neurotransmitters. Functionally, their release is not confined to synapses although some portions of their pool are also frequently co-secreted together with classic neurotransmitters. In contrast, they can be released from the dense core vesicles of practically any portion of the neurons [4,5,6]. Unlike classic neurotransmitters, neuropeptides are not taken back economically by a reuptake system but are metabolized by peptidases. Nonetheless, this process frequently yields biologically active compounds [6]. Further, due to their prolonged half-life, the neuropeptides can diffuse to long distances, therefore they act not only post- and presynaptically (i.e., paracrine, and autocrine fashion) but also in an endocrine manner[6], through the operation of G-protein coupled receptors (GPCRs) [6]. Due to these characteristics, their effect develops slowly, but it is usually longer lasting. Their impact, compared to that of the classic small molecular and even gaseous neurotransmitters, is almost always less robust, which strongly resembles the activity of hormones [7]. That is why in the literature some members are still mentioned as neurohormones and their activity has been formulated as “neuromodulation” [4,5,6], which represents a “mild” or “functionally buffered” way of signal transduction [4,5,6]. A unique feature of neuromodulation is that it is realized through a much broader arsenal of receptors than that of the neurotransmitters [5]. Also, the ligands themselves show immense structural versatility because in some families almost infinite splice variants can be produced from several copies of an ancestral gene [5]. Therefore, it is not surprising, that neuroscience has been struggling to formulate a rigid functional definition of neuromodulation, in contrast with that of neurotransmission and neurotransmitters [8].
Accordingly, neuropeptides appear to represent an individual and separate way of the transfer of biological information, somewhere in between the classical neurotransmitters and peripheral hormones. The secretion of these modulators gives rise to less acute but, in the long run, more profound changes in the neural connectome, the operative framework of neurophysiology [9]. The multi-faceted activity and redundancy of these signaling molecules provide an indispensable component of the plasticity and resilience of the CNS [10,11]. This is further augmented by the bewildering diversity and extremely broad distribution of several neuropeptide families. Some groups and their receptors are expressed at every level of neuroendocrine control [4,5,6].
It is apparent that the challenges to which the CNS is exposed ultimately will give rise to changes in translational processes. This will manifest itself in the modification of receptor and enzyme expression, synaptic plasticity, dendritic pattern, and structure and finally the complete wiring of the connectome [9,12]. The neuropeptides represent an essential but so far overlooked element of the translational machinery, bridging the gap between volatile functional alterations and permanent structural changes [13]. As being peptides, they represent one of the earliest steps (along with the neurotransmission-evoked peptide and protein synthesis) in the translational processes of information signaling in the CNS. Their modulatory action by the extremely versatile ligands, the corresponding receptors, and the multitude of stimulated signaling cascades make them the most flexible line of neuroendocrine plasticity and adaptation [4,5,6]. For instance, experiments on the orexin/hypocretin system especially support this view: its cooperation with other neuropeptides appears to harmonize the stress response to arousal-related stimuli [14,15,16,17,18,19,20,21,22,23].
The objective of this review paper is to provide an overview of the orexin/hypocretin peptide family and its role in the regulation of sleep and arousal states. Neuropeptides are a distinct class of signaling molecules that play a critical role in the modulation of neural activity and the regulation of behavior [24,25,26,27,28,29,30,31]. The orexin/hypocretin system is a recently described hypothalamic neuropeptide system that has been shown to be involved in the regulation of sleep and arousal states. The paper will explore the biochemical features of neuropeptides that differentiate them from classical neurotransmitters, the role of neuropeptides in the translational machinery of the CNS, and the flexibility of neuropeptides in neuroendocrine plasticity and adaptation. The review will also examine the cooperation of the orexin/hypocretin system with other neuropeptides in harmonizing the stress response to arousal-related stimuli. Overall, the review aims to provide a comprehensive understanding of the orexin/hypocretin peptide family and its role in the regulation of sleep and arousal states.
2. The hypocretin/orexin peptide family: an indispensable neuroendocrine switch of circadian activities.
The hypocretins/orexin system represents an extremely complex neuropeptide network in the CNS [32,33]. The seminal papers [34,35,36], which dealt with the discovery of the ligands (orexin-A, orexin-B) and their receptors (OX1 and OX2) also demonstrated the hyperphagic [34] and neuroexcitatory activity [35] of the orexins, and the unique distribution pattern of the system. The orexin/hypocretin system, similarly to the melanin-concentrating hormone (MCH) positive neurons [37], has a well-circumscribed expression in the hypothalamus [35,38]. Its cell bodies are restricted to the lateral, dorsal, dorsomedial, and perifornical areas. However, its axon terminals reach distant regions, and its receptors are scattered throughout the whole CNS [36].
Figure 1.
The amino acid sequences of the orexin/hypocretin family peptides.
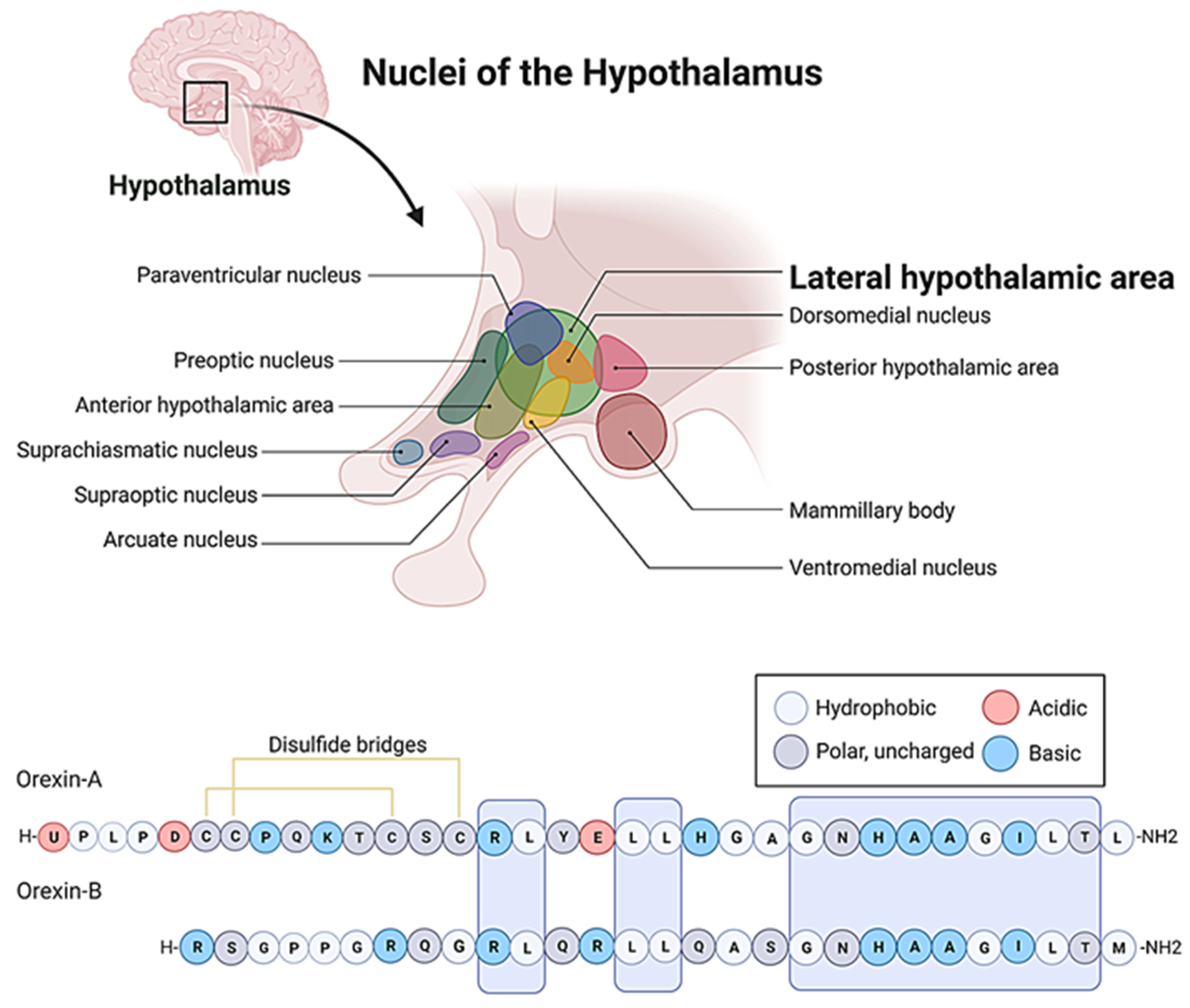
The function of the target areas of the orexin neurons suggested numerous clues to the feasible actions of the orexin neurons. Since then, many important physiological functions of the system have been verified [14,39]. Although at first, the orexins proved to play an important role in hedonic feeding [34,40], later publications unveiled that the most important aspect of their functional spectrum could be the regulation of arousal [41]. The orexin network is connected to several important centers of the ascending reticular activation system (ARAS) such as the nucleus raphe (NR), the locus coeruleus (LC), and the periaqueductal grey (PG) [36]. These nuclei operate with classical neurotransmitters (serotonin, norepinephrine, dopamine, respectively) and foster the most important inputs to the dualistic centers of sleep-wakefulness regulation: the tuberomammillary nucleus (TMN) and the ventrolateral preoptic nucleus (VLPO) of the hypothalamus. The histaminergic TMN and the galanin and (γ-amino-butyric-acid) GABAergic VLPO control sleep onset by a flip-flop mechanism [42,43,44]. It seems that the orexin-positive cells facilitate arousal by indirect disinhibition: they block the GABAergic output of the VLPO through the stimulation of the above-mentioned monoaminergic nuclei of the ARAS [36,45,46,47]. This way the orexin system could be easily categorized not only as the master regulator of arousal [39] but also as the modulator of the sleep-wake cycle and the circadian rhythms [41,42,48].
The most important physiological processes, which are under the circadian control of the orexin system are food and fluid intake [49,50,51], metabolism and thermoregulation [52], the activity of the hypothalamic-pituitary-adrenal cortex (HPA) axis [14,16,39,53,54,55,56], and reproduction [57,58]. The anatomical connections, which provide the foundation of these physiological actions have also been verified: the orexinergic axons target the arcuate, infundibular, preoptic, supraoptic, and paraventricular (PVN) nuclei of the hypothalamus [12,42]. Due to the hyperphagic activity and the specific localization of the orexin neurons, at least a group of their population can certainly be identified with some portion of the classic glucose-sensitive feeding center of the ventrolateral hypothalamus [59,60,61,62,63]. However, their role is more complex than suggested by the initial publications in the regulation of body weight [64]. While acutely they stimulate feeding [34], in the long run, it is their deficiency that is associated with weight gain [65], which is attributed to their two-pronged action on thermoregulation. The orexins concurrently activate heat loss and thermogenesis. They stimulate heat dissipation [15,66] through sympathetic vasodilation [67], together with the acceleration of metabolism in the brown adipose tissue [68]. Obviously, these thermoregulatory processes prevent excessive accumulation of fat. The orexins also modulate the activity of the reproductive axis [57,58,69,70], providing the link between self-preservation and species-preservation, since a well-fed, but not overweight subject can optimally guarantee the survival of its offspring. As far as self-preservation is concerned, the orexins also proved to be one of the most important orchestrators of the stress response [54,55]. Further, they modulate all threat-related adaptive behavioral processes [71,72,73], even in fear-related learning (Table 1) [74,75,76,77].
However, the orexin system has proven to act as an important regulator even in those physiological and pathophysiological processes which possess less apparent temporal characteristics. To mention a few of them: pain sensation [78], anxiety, mood, reward processes, and addiction (Table 1) [32,53,71,79,80]. The orexin system represents the poster child of neuropeptide regulation: its perikarya are confined to a small region but it signals diverse evolutionarily conserved functions to distant targets. In summary, we can say, that its principal role must be a temporal gating of brainstem functions [12,42].
During the investigation of the pathophysiological alterations of the hypocretin/orexin network, some unique features of the system were unveiled. It is a well-known phenomenon, in neuropeptide pathophysiology, that the deficiency of a given neuropeptide, or its secretory neurons usually does not bring about significant functional disturbances in the affected organism. It is due to the functional overlapping and redundancy of different neuropeptides or neuropeptides families. Typically, in congenital cases, during embryonic and fetal development, other neuropeptides can functionally compensate for the deficiency of the affected transcript even in knock-out animals. Obviously, acquired abnormalities are less prone to correction, due to the much more limited adaptation of the adult brain. Therefore, neuropeptide deficiencies do not cause such dramatic pathophysiological and clinical changes as in the case of congenital or acquired disorders of neurotransmission such as dopamine (phenylketonuria, Parkinson’s disease, and Sydenham’s chorea minor) GABA (Huntington’s disease, some forms of congenital epilepsies) or acetylcholine (myasthenia gravis, Lambert-Eton myasthenic syndrome, and Alzheimer’s disease) metabolism disorders [81]. However, the orexin/hypocretin system is different in this respect. Not only the acquired but also the congenital deficiency inevitably leads to severe pathophysiological changes exemplified by narcolepsy (Table 1) [82,83]. This might be attributed to the fact that the orexin-positive neuron population does not exceed 50,000-80,000 cells in the hypothalamus [84], which makes them peculiarly sensitive to injuries. Furthermore, the orexins bear weak structural resemblance only to few members of the incretin family [34,35]. Even orexin-A and orexin-B are different in 50% of their primary structure, and they exhibit significantly different receptor affinities [34,35]. Therefore, it is not surprising that hardly any other neuronal network can take over the function of the orexinergic system. Some functional overlapping might be provided by other GPCRs, since certain neuropeptide receptors, such as the type-2 NPY receptor, the thyrotropin-releasing hormone (TRH) receptor, the cholecystokinin type-A receptor, and the NK2 neurokinin receptor show some similarities (26%, 25%, 23%, and 20% identity, respectively) to the orexin receptors (OX1 and OX2) [34]. However, their binding affinity to the orexins is negligible[85]. The highest structural similarity is exhibited by the neuropeptide-FF (NPFF) receptor of the RF-amide peptide family, which is identical to OX1 in 37% and OX2 in 35%, respectively, and shows significant affinity to the orexins [86,87].
The indispensable role of the orexin system in the maintenance of vigilance is strongly supported by the finding that narcolepsy and cataplexy observed in the dog [88], and the mouse [89] can be solely attributed to the deficiency of the orexin/hypocretin system. Moreover, by now it has been supported by several observations, that human cases of narcolepsy with cataplexy [82,83,84] can also be attributed to either the abnormal development [90], or the acquired immunological destruction [91] of the orexin-hypocretin system. These observations offer great therapeutic opportunities not only for sleep disorders but also for the abnormalities of those physiological functions, which are under the circadian control of the orexin/hypocretin network.
3. The role of orexins in the regulation of the stress response.
The reaction of our neuroendocrine regulation to adverse challenges is provided by the interaction between the sympathoadrenal (SA) system and the HPA axis [92]. Although, they represent two distinct pathways, the line between them is frequently blurred even in scientific literature. Perhaps, due to their interwoven functions as they complement each other’s activity while trying to maintain the homeostatic balance of the challenged individual. However, the SA response described by Cannon [93] is carried out by the cooperation of the autonomic nervous system and the adrenal gland, while the stress response, discovered by Selye [94], relies on the reaction of the HPA system, one of the central neuroendocrine axes later described by Schally, Guillemin [95], and Vale [96]. Unfortunately, by now, the terminology has been oversimplified, and the stress response (though with several stages) is frequently used as an umbrella term for both responses. Only in meticulous descriptions are these two neuroendocrine reactions clearly separated. To avoid confusion, for the HPA response, the most suitable term is the synonym (general adaptation syndrome: GAS) coined later by Selye [97]. Nonetheless, the distinction between the two pathways is of crucial importance, because it helps clarify many contradictions in the literature. Some conflicting responses to certain stress paradigms could be easily resolved by clear discrimination between the two potential targets of adverse stimuli, that is the SA system and the HPA axis (Table 2).
It is well-known that many neuropeptides modulate the activity of the HPA axis. For instance, NPY, neurotensin (NT), ghrelin, apelin, and endomorphins activate [17,20,23,106,107,108,109], while oxytocin and the natriuretic peptides inhibit the system [110,111,112,113]. The output of the HPA axis is quite uniform: it begins with the pituitary translation and cleavage of pro-opiomelanocortin (POMC) yielding adrenocorticotropic hormone (ACTH), which upon secretion stimulates glucocorticoids release from the adrenal cortex [94,97]. However, in sharp contrast with the output, the input of the HPA axis is extremely diverse and involves a multitude of neuropeptides in signal transduction [107]. Therefore, it is not surprising that the modality (systemic and neurogenic ones) and the schedule (acute, repeated, and chronic) of the stressors strongly influence the extent of the HPA response. The systemic challenges (e. g. osmotic, immune, etc.) disturb the homeostatic balance of the organism, which is directly projected to the brainstem, while the neurogenic paradigms (fear, pain) are processed by cerebral centers [92]. The responses to these two types of challenges are signaled in a dichotomized manner in the brain. The corticotrope-releasing hormone (CRH) positive neurons of the PVN are responsible for the acute and processed stimuli, while parvocellular arginine-vasopressin (AVP) cells in the PVN and SON maintain responsiveness to chronic, repeated, and homeostatic challenges [114]. It is also worth noting, that neuropeptide modulation perfectly complements the built-in brakes of the GAS: the stepwise ultrashort, short and long loop feedback mechanisms provided by CRH, ACTH, and the glucocorticoids, as well as the potent anti-inflammatory action of glucocorticoids [92]. These mechanisms are called stress coping or stress resilience, and they harness the severe inflammatory response (SIRS), which, otherwise, could consume the organism [92,115,116].
As far as the effect of the orexins on the HPA axis and the SA system is concerned, the two responses work hand in hand. Namely, in both responses, the orexins play a predominantly stimulatory role [54,55]. However, according to literature data, they are activated separately. The orexins seem to elicit a uniform response of the SA system, initiated at the nucleus of the solitary tract (NST) and the LC [39,42,49,54,117]. Ultimately, it is not a far-fetched idea to state that the perifornical, dorsal, dorsomedial, and lateral hypothalamic foci of orexin-positive neurons can be identified with those in the caudal hypothalamic region, which was demonstrated essential for an intact “fight or flight” and the “sham rage” response by Philip Bard and Walter Hess [8,71].
However, as for the HPA axis, the picture is more complex. Soon after the discovery of the dense orexinergic innervation of the hypothalamic centers (PVN and SON) of the GAS, the scientific rivalry in this highly coveted topic begot several important papers, which established that the orexin neurons can activate the HPA axis predominantly at a hypothalamic level [14,39,118]. The main targets of the orexin neurons are the CRH-positive perikarya of the PVN [36,118]. Nonetheless, later publications showed, that the connection between the orexin- and CRH-positive neuron population is bidirectional since abundant CRH-positive fibers land on orexinergic perikarya of the hypothalamus [119,120,121]. Apparently, the orexin-evoked HPA activation also involves the release of noradrenaline and NPY [16,122,123,124], which can significantly diversify its processing [115,116].
As far as the input of the HPA axis is concerned, the activity of orexins appears to be stressor and schedule specific [54,55]. In an acute setting, the processed challenges with heightened arousal (aversive odor, novelty, and contextual fear) give rise to more conspicuous activation of the orexin neurons (verified by c-fos expression) than the systemic challenges (e.g., cold exposure) or long-lasting procedures (e.g., restraint and immobilization) [54,55,73,125]. Nevertheless, while acute stress mostly activates the orexin neurons, experiments with chronic or repeated stressors returned mixed results [54,55], which findings may reflect an adaptation to unavoidable and permanent challenges. Further studies revealed that the involvement of the orexins in the GAS depends on not only the modalities and the schedule of the applied stressor but also the species and the gender of investigated subjects. Females and strains with better stress resilience phenotype release less orexin to adverse stimuli [54,55].
Regarding the output of the HPA axis, the orexins have been proven to stimulate the HPA axis not only at a hypothalamic but also at a pituitary and adrenal level [54,55]. This finding is of crucial importance as peripheral activation stabilizes the HPA response to prolonged stimuli. It nurtures sufficient basal activity but also prevents an exaggerated hypothalamic response by maintaining negative feedback through the release of ACTH and glucocorticoids. Apparently, the orexin/hypocretin system also plays a crucial role in the cooperation and seamless integration of the GAS and the “fight or flight” response. Even the earliest publications, which dealt with the orexin system, demonstrated the dense innervation of the BNST, a limbic center, which harmonizes the activity of the SA system and the GAS [36]. Therefore, it is not unrealistic at all to conceive of the orexins as the coordinators of the stress response to challenges with heightened arousal[54,55].
4. The role of the orexins in the regulation of behavior.
There is a consensus in the scientific community, that the orexin system is the most important peptidergic mediator of the ARAS [39,41,42] and thus the arousal processes. Its indispensable role in the regulation of vigilance was ultimately confirmed by the observations that its deficit leads to irreversible functional consequences both in congenital and acquired disorders of arousal and sleep: narcolepsy and cataplexy (Table 1) [82,83,84,88,90]. However, over time it became obvious that not only the maintenance of wakefulness but also the fine-tuning of arousal-related behavior belongs to the functional repertoire of the orexin system [74]. This concept is supported by experimental data, which demonstrated that the orexins stimulate attention, rearing, and locomotion as well as such anxiety-related stereotyped behaviors as grooming and freezing [39,41,42,126,127,128,129]. Further experiments are needed to verify its role in such ancient behavioral patterns as the thanatosis [130,131].
It is well-known that threats belong to the strongest activators of vigilance. They evoke alertness and attention, then provoke an emotional response, i.e., fear. And fear has a huge impact on all aspects of behavior, which manifests itself in anxiety, alterations of mood, and cognition among others [132,133]. This strong association considered, it is not surprising, that the appropriate behavioral responses to both transient and permanent threats are accompanied by the stimulation of the orexin system [134]. It seems that in the central processing of threat, first, the robust stimulation of the ARAS will involve the activation of the orexinergic network [39,41,42]. In turn, its hypothalamic foci will fine-tune the neurotransmitter release [72,76,77,134,135,136] of those brainstem and limbic centers, which are responsible for the regulation of emotions, affections, mood, and learning processes [54,134,137]. However, this activation, similarly to the neuroendocrine parallels, is bidirectional, as inputs from the limbic structures (the septum, BNST, basal forebrain, amygdala, and the hippocampus) account for most of the telencephalic inputs to the orexin neurons [138]. In rodents, a multitude of experiments demonstrated that anxiogenic stimuli, such as exposure to cat odor or novelty [71,72,73,139] gave rise to the activation of the orexin network. This way it is not surprising that orexin treatment enhances the startle-response [140] and passive avoidance [77].
The function of the amygdala, which monitors emotional learning, and arousal-driven memory consolidation [141], is controlled by the orexin network indirectly and directly. It receives direct orexinergic projections [36], but the indirect pathway (through the LC) is more important: it carries rich noradrenergic projections to the lateral amygdala [134]. Therefore, soon after the discovery of the orexin network, several experiments were carried out to test the putative effect of the orexins on learning. These studies revealed that the orexins play an especially important role in cue-dependent fear memory formation, mainly through the release of noradrenaline [74,75,76]. For instance, orexin-A proved to promote emotional learning, memory consolidation, and retrieval processes in a passive avoidance paradigm and in social learning [77,142]. Hence, facilitation of learning ensures the avoidance of potentially harmful events, which ultimately is a preventive and therefore the most effective technique of stress coping.
Recently more and more attention has been paid to specific aspects of the orexin-mediated behavioral responses: reward and addiction. Functionally, the orexins are the prime examples of hedonistic neuropeptides [143]. In the past two decades, the orexins have been proven to take part in the control of such strong natural rewards as food, fluid intake [34,40,49,50,51], and reproduction [57,70]. They especially stimulate binge eating of palatable food [64,144]. This physiological data has already been substantiated by histological findings, as well. The cells of the two principal dopaminergic pathways (mesolimbic and mesocortical tracts) of the reward system [8] bear orexin-positive boutons [36]. The beginning (the ventral tegmental area (VTA)) of the pathways, the relay centers (the BNST, the amygdala, the hippocampus) and the nucleus accumbens (NAc), the target of the mesolimbic tract, and the prefrontal cortex, the ending of the mesocortical fibers receive rich orexinergic inputs [36].
In addition to these primary hedonistic features, the orexins appear to control stress resilience especially in chronic and repeated experimental conditions [54,55]. This concept was reinforced by histological data when the interaction between the orexin network and another hedonistic peptide the ghrelin was verified [62,145,146]. It was suggested that they should augment each other’s activity in stress coping, which could dampen the detrimental psychological consequences of adverse stimuli. Therefore, it was postulated early on that they could take part in the mediation and modulation of behavioral responses evoked by not only natural but also pharmacological rewards. Since then, numerous publications revealed that the orexin connectomics shows significant alterations not only in obesity [62,145,146] but also in drug dependence [147]. This way several pathophysiological responses in addiction, such as reinforcement, drug-seeking, and self-administration were attributed to its modified activity [148]. The orexin network is unambiguously upregulated in cocaine [149,150,151] and opiate abuse [152,153,154], while in the case of other substances, the reaction is more complex [148]. Acute alcohol consumption increased, while chronic alcohol, Δ19-tetrahydrocannabinol, and nicotine abuse decreased orexin expression in the hypothalamus [148,155,156,157]. The acute changes must be related to the direct effect of the addictive substance. However, chronic changes may reflect the response to a specific stress paradigm: drug withdrawal. Ultimately, these schedule-dependent bimodal changes also appear to reflect adaptation, a form of stress coping.
5. The cooperation between the orexinergic and other peptidergic neuronal networks.
As it has been discussed, the somas of the orexin/hypocretin neurons are restricted to the caudal portion of the hypothalamus [34], but their axon terminals reach distant regions, and their receptors are scattered throughout the whole CNS [36]. This spatial concentration of the cell bodies is not unique in the CNS since it can be observed among other neuropeptides such as MCH [158], ghrelin [159,160], and neuromedin-S [161]. Like in the case of other neuropeptides, the feasible interaction of the orexin system and other networks greatly broadens the regulatory repertoire of the orexin/hypocretin neurons. The potential partners are the MCH [162], NPY [163], apelin [17,164,165,166], ghrelin [18,19,20,167], and neuromedin [23,168] positive networks, which show marked functional overlapping with the orexins in the regulation of food intake, the sleep-wake cycle, stress response, and behavior. These networks may cooperate with each other, but the intact function of the orexin system is required for the normal processing of arousal-related processes [82,83,84,88,89].
So far, besides releasing hormones of the hypothalamic-pituitary-target organ axis, only the orexin system has proven to be the only essential neuropeptide in the regulation of the CNS. However, the interpolations of other neuropeptides and neurotransmitters in the signal transduction of the orexin neurons lend immense diversity and flexibility to orexin-regulated responses [57]. It is because the neuropeptide ligand and receptor families typically consist of several members, which may have numerous splice variants, can also be modified by peptidases after secretion, and can act on an arsenal of receptors [4,5,6,107,115,169]. With the different binding affinities and activities taken into consideration, the number of potential interactions between this abundance of ligands and receptors is infinite and may span from full agonism to complete antagonism [4,5,7,169].
Cooperation both in the afferent and efferent pathways of the orexin system has been verified. In the input, monoamines [36], NPY [170,171], and ghrelin [62,145,146] seem to play the most important role, while in the output NPY [15,16,124,170,172], POMC[173], and also monoamines [36,72,77,136,174] have been identified. Unambiguously, the pathways between the LC and the lateral hypothalamus are the most important connection of the orexin network in the regulation of arousal-related behavioral and endocrine responses [35,36]. Also, circulating peripheral or centrally released ghrelin significantly contributes to the hyperphagic activity of the orexins [62]. In the efferentation, the orexin connectomics cooperates with the corticotrope-releasing hormone (CRH) – urocortin system [14], the network of NPY positive [15,16] and monoaminergic [174] cells in the orchestration of the neuroendocrine responses to processed and homeostatic challenges [14,16]. Such interactions were established in other functions such as thermoregulation, mood, anxiety, learning [15,72,77,121,136,175], and reproduction [124]. Regarding arousal, one of the most important connections between the orexinergic system of the organism and the environment could be established through the suprachiasmatic nucleus (SCN) [176]. Neuromedin-S released from SCN of the hypothalamus might interpret those photic stimulations, which arrive at the SCN through the retinohypothalamic pathway[161]. Nevertheless, this aspect of hypocretin physiology must be further scrutinized and confirmed by experimental data.
6. Aspects of human pathophysiology: present and future therapeutic potential of orexin receptor ligands.
Even the first results of experiments done on the orexin/hypocretin system suggested that several human pathophysiological conditions could be explained by the alterations of the orexin neurons [177]. Dysfunctions of the ARAS and sleep disorders, such as obstructive sleep apnea-hypopnea syndrome, were the first and somewhat obvious culprits, which were thoroughly and successfully investigated. Since then, both in narcolepsy with cataplexy [82,83,84,90,178] and in obstructive sleep apnea-hypopnea syndrome [179,180,181] the dysfunction of the orexin system has been established. Moreover, the acquired form of narcolepsy proved to be a classic example of neuroinflammatory disorders. It seems to be evoked by H1N1 influenza virus infection or vaccination, which gives rise to an autoimmune reaction against hypocretin neurons [91,182,183]. As far as hyperactivity is concerned, the role of the orexin system has also been verified, since the exaggerated startle response in anxious patients could be effectively reduced with an orexin receptor antagonist [184].
Increased tone of the orexinergic system, especially in cooperation with the ghrelin network, has also been suspected in disorders of the reward system. It appears that their synergistic hyperactivity is accountable for a rare form of monogenic obesity, Prader-Willi syndrome (PWS) [185]. However, the picture is more complex, as the orexin system is a double-edged sword; it increases feeding and energy expenditure simultaneously depending on the environmental cues. Accordingly, it has been implicated in both weight gain and weight loss [64], as well as in such disorders of food consumption as binge eating [186], bulimia, and anorexia nervosa [187]. Furthermore, in Kleine-Levin syndrome [104,188] the alternation of hypersomnia with alert and hyperphagic stages has been connected to fluctuation of orexin levels in the CSF. It seems, that like the HPA response, the actual eating disorder is determined by the schedule and the modality of the psychological stressor [189], and it can manifest itself in seemingly opposite conditions (Table 1).
Regarding reproductive processes, the hypoactivity of the orexin system was observed in mothers suffering from gestational diabetes [190] and in patients diagnosed with polycystic ovary syndrome (PCOS) [191,192]. This might be related to the concomitant increase in body weight and leptin level of the patients, which downregulates orexin expression [193]. As for the pharmacological rewards, it is the substance withdrawal, that represents the common mechanism of orexin upregulation in different forms of drug addictions. Apparently, withdrawal symptoms are managed by the individual as stress stimuli and they increase arousal, attention, and drug-seeking behavior [143,194,195] (Table 1) .
In humans, the dysfunction of the orexin system in the regulation of the ARAS may also bring about the development of such diseases as attention deficit hyperactivity disorder (ADHD), anxiety, epilepsy, panic, and phobia (Table 1) [71,134,137,143,196]. These conditions go with the pathophysiological regulation of neuronal excitation and show clear circadian fluctuation, which reinforces the view that the alteration of the orexin/hypocretin system plays a causative role in their development. Long periods of overexcitation have detrimental impact on neurons. In the burnt-out phase, these diseases give way to such chronic conditions as major depression, post-traumatic stress disorder (PTSD), psychosomatic problems, like hypertension [197,198,199,200] and even neurodegenerative disorders [201]. Some further conditions such as cognitive disorders [202] and abnormal pain sensation [203,204,205] may also be related to the alterations of alertness and can be connected to the orexin/hypocretin system (Table 1). However, it is important to emphasize that orexin receptors show some structural similarities to those of the RF-amides [202]. Accordingly, their direct action may be mediated by not only their own receptors expressed on the crucial gating elements (LC, PAG) of pain signaling but also can be reinforced indirectly via the RF-amide receptors, which are expressed in both the ascending and descending pathways of pain sensation [206]. Abnormalities of orexin physiology have already been identified in chronic pain disorders, such as fibromyalgia [207], and especially in primary headaches such as migraine and cluster headaches (Table 1) [78,169,208]. The latter condition deserves special attention since their attacks show clear diurnal pattern, and in its pathogenesis, the role of the SCN has already been verified [208]. Additionally, in the context of neurological and psychiatric disorders linked to the orexin system, emerging research explores the potential of non-invasive brain stimulation techniques (NIBS), such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS), as therapeutic interventions. These NIBS approaches aim to modulate neural activity and may hold promise in mitigating symptoms associated with conditions like migraine, anxiety, depression, and cognitive disorders that are influenced by orexin dysfunction [209,210,211] .
The previously mentioned conformational overlapping between the RF-amide and orexin receptors may also account for the reproductive and antineoplastic activities of the orexins [212,213,214]. It is because RF-amides play a well-known role in the inhibition of metastasis formation [215] and in the past few years they have emerged as the metabolic regulators of the gonadal axis [216]. Since the activity of the RF-amide system shows clear periodic activity, it is quite reasonable to imply that the orexin system may modulate its function either directly or indirectly [212].
Finally, it must be mentioned that the dysfunction of the orexin system was demonstrated in such common neuropsychiatric, neuroinflammatory, and neurodegenerative disorders such as schizophrenia, Parkinson’s disease, Alzheimer’s disease, Huntington’s disease, multiple sclerosis (MS), and amyotrophic lateral sclerosis (ALS) (Table ) [201,217]. Nevertheless, in these pathologic conditions, the dysfunction of the orexin system is apparently not specific but can be attributed to the widespread devastation of the connectome. In these conditions, ultimately, all neural networks will be affected, but the orexin system is specifically frail and sensitive to focal injuries since it has a limited number of neurons, which are confined to a relatively small region. Therefore, it belongs to the first centers, which succumb to the detrimental effects of misfolded protein aggregation and neuroinflammation. That is why some shared, conspicuous symptoms of the above-mentioned fatal disorders were identified to result from the failure of the orexin network. Cataplexy and dysfunction of postural reflexes can be observed in Parkinson’s, Huntington’s, and prion diseases. Alteration of sleep patterns and vigilance are common findings in Alzheimer’s, MS, and prion diseases. Rapid fluctuation of mood, unwarranted anxiety, irrational fears, and extreme irritability are the common behavioral symptoms [218,219] in the above-mentioned diseases. Later in all these symptom categories either the hyper- or hypoactivity of the orexin system has been suspected or already verified [201,217].
7. Promising results in translational pharmacology
It is a well-known hindrance of neuropeptide pharmacology, that often those compounds, which possess the most promising biochemical features (affinity, activity, half-life, etc.) cannot bypass the blood-brain barrier (BBB) [7,220]. In several instances, only circumscribed areas (lamina cribrosa, the circumventricular organs) provide access to the cerebrospinal fluid (CSF) for native ligands [7], or sophisticated nanocarriers (liposomes, nanoparticles, etc.) are required to surmount this pharmacokinetic obstacle [221]. However, in the case of the previously discussed feed-promoting neuropeptides such as ghrelin and the orexin system both the natural ligands and their chemically designed analogs can freely bypass the BBB [7,222,223]. The derivatives of the orexins are especially successful in this regard because some of the antagonists have already been approved by the Food and Drug Administration (FDA) in the treatment of insomnia [100]. Other antagonists, which are suggested for the treatment of panic, major depressive disorder, anxiety, and binge eating are under investigation [100]. The studies with the antagonists, which are recommended for the treatment of narcolepsy are also in the clinical phase of pharmacological trials (Table 3). At present the most coveted aim in neuroendocrine research is to engineer orexin derivatives, which could relieve the abnormalities of sleep-wake cycle in neurodegenerative disorders. They would be game changers in palliative therapy as traditional hypnotics are strong depressants and further deteriorate the function of the otherwise failing CNS. Therefore, present, and future orexin derivatives belong to the pioneering and most successful compounds of neuropeptide pharmacology and have huge potential in pharmaceutical development [100].
8. Discussion
Orexin/hypocretin neuropeptides are pivotal players in regulating various physiological processes, such as food intake, metabolism, the HPA axis, reproduction, and behavior [12,32,34,40,56,57,64,74,112,224,225]. They were primarily described as orchestrating such parameters of homeostatic balance as feeding, thermogenesis, and heat dissipation [15,34,40,64,66]. Later research shed light on their intricate involvement in the mediation and modulation of such behavioral paradigms as arousal, anxiety, fear, and the stress response [14,39,41,56,71,76,134,135]. The orexin system is also implicated in the regulation of pain sensation and behavioral changes evoked by natural and pharmacological rewards such as addiction [53,71,78,101,152,169,194,226,227,228]. In this way, dysfunctions of the orexin system have been associated with various human pathophysiological conditions, such as obesity, addictive disorders, narcolepsy, obstructive sleep apnea-hypopnea syndrome, anxiety, cognitive disorders, and abnormal mood fluctuations [12,74,79,184,186,199,229]. This review tries to seamlessly integrate the diverse activities of the orexins and provides a more in-depth understanding of those fields, such as stress response, fear, anxiety, and learning, in which the authors have significantly contributed to the literature [14,15,16,72,77,136].
Regarding the limits of this article, it is important to note that the review is based on existing literature and does not present new experimental data. As a result, the caliber and scope of the studies included in the analysis limit the conclusions drawn from this review. Additionally, the review focuses on preclinical research on the orexin/hypocretin system, and the translation of these findings into clinical practice may be challenging. However, the review has several merits, including its comprehensive and interdisciplinary approach to understanding the orexin/hypocretin system synthesizing information from various fields, including neuroscience, endocrinology, and pharmacology [12,32,57,64,74,100,230,231,232,233,234]. It could provide a ‘food for thought’ for researchers and clinicians interested in the orexin/hypocretin system, and it could inspire future research by identifying knowledge gaps and areas that require further investigation.
The ultimate goal of the research on the orexin/hypocretin system is to develop effective therapeutic interventions for various disorders, such as sleep disorders, obesity, addiction, and anxiety [34,42,48,64,71,73,88,89,104,134,135,137,235,236,237,238,239]. However, this goal presents several challenges, including the need to understand the complex and multifaceted role of orexins in physiology and behavior, as well as the need to develop safe and effective drugs that target the orexin system. To achieve this goal, researchers need to have a deep understanding of the orexin/hypocretin system, including its molecular and cellular mechanisms as well as its interactions with other systems in the body. They also need to develop advanced technologies for studying the orexin system, such as optogenetics, chemogenetics, and advanced imaging techniques. In addition, researchers need to develop safe and effective drugs that target selectively the orexin system, which requires a thorough understanding of the pharmacokinetics and pharmacodynamics of orexin derivatives. Overall, this line of research has the potential to improve the lives of millions of people worldwide, making it a crucial area of investigation. as well as their potential therapeutic applications [240,241,242]. Nevertheless, since several derivatives of the orexins with high affinity and activity for their receptors are able to bypass the blood-brain barrier, some antagonists have already been approved by the FDA for the treatment of insomnia, and other antagonists and agonists are under investigation for the treatment of various disorders of food intake [100,231,243].
9. Conclusion
The interdisciplinary approach of this review has enhanced our understanding of the orexin/hypocretin neuropeptide family and its potential therapeutic applications. However, there are still several theoretical and methodological avenues that require refinement, such as the need for more precise and selective orexin receptor agonists and antagonists. Future research directions could focus on developing innovative drug delivery systems that can target the orexin system effectively while minimizing off-target effects. Additionally, further research is needed to understand the complex interactions between the orexin system and other physiological and behavioral processes, such as the immune system and circadian rhythms. Overall, the orexin/hypocretin system is a fascinating area of research with significant theoretical and translational implications. By understanding the complex and multifaceted role of the orexin system, researchers can identify new drug targets and develop innovative drug delivery systems that can effectively treat various disorders. We hope that this review serves as a valuable resource for researchers and clinicians interested in the orexin/hypocretin system and the potential pharmacons targeting the system.
Author Contributions
Conceptualization, M.J.; writing—original draft preparation, M.J. and M.T.; writing—review and editing, M.J.; B.T.; Z. B.; M.T. L. V.; visualization, M.T.; supervision, M.J. and M.T.; project administration, M.J. and M.T.; funding acquisition, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research, Development, and Innovation Office—NKFIH K138125, TUDFO/47138-1/2019-ITM, and the HUN-REN Hungarian Research Network.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The figure was created using BioRender.com
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACTH | adrenocorticotropic hormone |
| ALS | ascending reticular activation system |
| ARAS | ascending reticular activation system |
| ARC | arcuate nucleus |
| BBB | blood brain barrier |
| BNST | bed nucleus of stria terminalis |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| CRH | corticotrope-releasing hormone |
| DMH | dorsomedial hypothalamus |
| DR | dorsal raphe |
| FDA | Food and Drug Administration |
| GABA | -amino-butyric-acid |
| GAS | General adaptation syndrome |
| GPCRs | G-protein-coupled receptors |
| HPA | hypothalamic-pituitary-adrenal cortex |
| LA | lateral amygdala |
| LC | locus coeruleus |
| LHA | lateral hypothalamic area |
| MCH | melanin-concentrating hormone |
| MS | multiple sclerosis |
| NAc | nucleus accumbens |
| NIBS | non-invasive brain stimulation techniques |
| NM | neuromedin |
| NMS | neuromedin S |
| NPY | neuropeptide Y |
| NST | nucleus of the solitary tract |
| NT | neurotensin |
| OX | orexin receptors |
| PAG | periaqueductal grey |
| PFA | perifornical area |
| POA | preoptic area |
| POMC | pro-opiomelanocortin |
| PPT | pedunculopontine tegmental nucleus |
| PVN | paraventricular nuclei |
| RVLM | rostral ventrolateral medulla |
| RVMM | rostral ventromedial medulla |
| SA | sympathoadrenal |
| SCN | suprachiasmatic nucleus |
| SON | supraoptic nucleus |
| tDCS | transcranial direct current stimulation |
| TMs | transcranial magnetic stimulation |
| TMN | tuberomammillary nucleus |
| VTA | ventral tegmental area |
| VLPO | ventrolateral preoptic nucleus |
References
- Oliver, G.; Schafer, E.A. On the Physiological Action of Extracts of Pituitary Body and certain other Glandular Organs: Preliminary Communication. J Physiol 1895, 18, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Dale, H.H. On some physiological actions of ergot. J Physiol 1906, 34, 163–206. [Google Scholar] [CrossRef]
- US, V.E.; Gaddum, J.H. An unidentified depressor substance in certain tissue extracts. J Physiol 1931, 72, 74–87. [Google Scholar] [CrossRef]
- van den Pol, A.N. Neuropeptide transmission in brain circuits. Neuron 2012, 76, 98–115. [Google Scholar] [CrossRef]
- Elphick, M.R.; Mirabeau, O.; Larhammar, D. Evolution of neuropeptide signalling systems. J Exp Biol 2018, 221. [Google Scholar] [CrossRef]
- Marvar, P.J.; Andero, R.; Hurlemann, R.; Lago, T.R.; Zelikowsky, M.; Dabrowska, J. Limbic Neuropeptidergic Modulators of Emotion and Their Therapeutic Potential for Anxiety and Post-Traumatic Stress Disorder. J Neurosci 2021, 41, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Hokfelt, T.; Bartfai, T.; Bloom, F. Neuropeptides: opportunities for drug discovery. Lancet Neurol 2003, 2, 463–472. [Google Scholar] [CrossRef]
- Purves, D. Neuroscience, Sixth edition. ed.; Oxford University Press: New York, 2018; p. 1 volume (various pagings). [Google Scholar]
- Seguin, C.; Sporns, O.; Zalesky, A. Brain network communication: concepts, models and applications. Nat Rev Neurosci 2023. [Google Scholar] [CrossRef]
- Tanaka, M.; Spekker, E.; Szabó, Á.; Polyák, H.; Vécsei, L. Modelling the neurodevelopmental pathogenesis in neuropsychiatric disorders. Bioactive kynurenines and their analogues as neuroprotective agents—In celebration of 80th birthday of Professor Peter Riederer. Journal of Neural Transmission 2022, 129, 627–642. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Editorial of special issue ‘dissecting neurological and neuropsychiatric diseases: Neurodegeneration and neuroprotection’. 2022, 23, 6991. [CrossRef]
- Sakurai, T.; Mieda, M. Connectomics of orexin-producing neurons: interface of systems of emotion, energy homeostasis and arousal. Trends Pharmacol Sci 2011, 32, 451–462. [Google Scholar] [CrossRef]
- Tanaka, M.; Diano, M.; Battaglia, S. Insights into structural and functional organization of the brain: evidence from neuroimaging and non-invasive brain stimulation techniques. Frontiers in Psychiatry 2023, 14, 1225755. [Google Scholar] [CrossRef] [PubMed]
- Jaszberenyi, M.; Bujdoso, E.; Pataki, I.; Telegdy, G. Effects of orexins on the hypothalamic-pituitary-adrenal system. J Neuroendocrinol 2000, 12, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- Jaszberenyi, M.; Bujdoso, E.; Kiss, E.; Pataki, I.; Telegdy, G. The role of NPY in the mediation of orexin-induced hypothermia. Regul Pept 2002, 104, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Jaszberenyi, M.; Bujdoso, E.; Telegdy, G. The role of neuropeptide Y in orexin-induced hypothalamic-pituitary-adrenal activation. J Neuroendocrinol 2001, 13, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Jaszberenyi, M.; Bujdoso, E.; Telegdy, G. Behavioral, neuroendocrine and thermoregulatory actions of apelin-13. Neuroscience 2004, 129, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Palotai, M.; Bagosi, Z.; Jaszberenyi, M.; Csabafi, K.; Dochnal, R.; Manczinger, M.; Telegdy, G.; Szabo, G. Ghrelin and nicotine stimulate equally the dopamine release in the rat amygdala. Neurochem Res 2013, 38, 1989–1995. [Google Scholar] [CrossRef]
- Palotai, M.; Bagosi, Z.; Jaszberenyi, M.; Csabafi, K.; Dochnal, R.; Manczinger, M.; Telegdy, G.; Szabo, G. Ghrelin amplifies the nicotine-induced dopamine release in the rat striatum. Neurochem Int 2013, 63, 239–243. [Google Scholar] [CrossRef]
- Jaszberenyi, M.; Bujdoso, E.; Bagosi, Z.; Telegdy, G. Mediation of the behavioral, endocrine and thermoregulatory actions of ghrelin. Horm Behav 2006, 50, 266–273. [Google Scholar] [CrossRef]
- Tanaka, M.; Telegdy, G. Neurotransmissions of antidepressant-like effects of neuromedin U-23 in mice. Behav Brain Res 2014, 259, 196–199. [Google Scholar] [CrossRef]
- Telegdy, G.; Adamik, A. Anxiolytic action of neuromedin-U and neurotransmitters involved in mice. Regul Pept 2013, 186, 137–140. [Google Scholar] [CrossRef]
- Jaszberenyi, M.; Bagosi, Z.; Thurzo, B.; Foldesi, I.; Telegdy, G. Endocrine and behavioral effects of neuromedin S. Horm Behav 2007, 52, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Telegdy, G. Neurotransmissions of antidepressant-like effects of neuromedin U-23 in mice. Behavioural brain research 2014, 259, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Schally, A.; Telegdy, G. Neurotransmission of the antidepressant-like effects of the growth hormone-releasing hormone antagonist MZ-4-71. Behavioural brain research 2012, 228, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Csabafi, K.; Telegdy, G. Neurotransmissions of antidepressant-like effects of kisspeptin-13. Regulatory peptides 2013, 180, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Telegdy, G.; Tanaka, M.; Schally, A.V. Effects of the growth hormone-releasing hormone (GH-RH) antagonist on brain functions in mice. Behavioural brain research 2011, 224, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Rákosi, K.; Masaru, T.; Zarándi, M.; Telegdy, G.; Tóth, G.K. Short analogs and mimetics of human urocortin 3 display antidepressant effects in vivo. Peptides 2014, 62, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.N.; Nguyen, N.P.K.; Nguyen, L.T.H.; Shin, H.-M.; Yang, I.-J. Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils. Biomedicines 2023, 11, 1248. [Google Scholar] [CrossRef]
- Tanaka, M.; Kádár, K.; Tóth, G.; Telegdy, G. Antidepressant-like effects of urocortin 3 fragments. Brain Research Bulletin 2011, 84, 414–418. [Google Scholar] [CrossRef]
- Tanaka, M.; Telegdy, G. Antidepressant-like effects of the CRF family peptides, urocortin 1, urocortin 2 and urocortin 3 in a modified forced swimming test in mice. Brain research bulletin 2008, 75, 509–512. [Google Scholar] [CrossRef]
- Xia, L.; Liu, H.Y.; Wang, B.Y.; Lin, H.N.; Wang, M.C.; Ren, J.X. A review of physiological functions of orexin: From instinctive responses to subjective cognition. Medicine (Baltimore) 2023, 102, e34206. [Google Scholar] [CrossRef]
- Soya, S.; Sakurai, T. Evolution of Orexin Neuropeptide System: Structure and Function. Front Neurosci 2020, 14, 691. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [PubMed]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci U S A 1998, 95, 322–327. [Google Scholar] [CrossRef]
- Peyron, C.; Tighe, D.K.; van den Pol, A.N.; de Lecea, L.; Heller, H.C.; Sutcliffe, J.G.; Kilduff, T.S. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J Neurosci 1998, 18, 9996–10015. [Google Scholar] [CrossRef]
- Bittencourt, J.C.; Presse, F.; Arias, C.; Peto, C.; Vaughan, J.; Nahon, J.L.; Vale, W.; Sawchenko, P.E. The melanin-concentrating hormone system of the rat brain: an immuno- and hybridization histochemical characterization. J Comp Neurol 1992, 319, 218–245. [Google Scholar] [CrossRef]
- Lopez-Lopez, C.; Dietrich, M.O.; Metzger, F.; Loetscher, H.; Torres-Aleman, I. Disturbed cross talk between insulin-like growth factor I and AMP-activated protein kinase as a possible cause of vascular dysfunction in the amyloid precursor protein/presenilin 2 mouse model of Alzheimer’s disease. J Neurosci 2007, 27, 824–831. [Google Scholar] [CrossRef]
- Hagan, J.J.; Leslie, R.A.; Patel, S.; Evans, M.L.; Wattam, T.A.; Holmes, S.; Benham, C.D.; Taylor, S.G.; Routledge, C.; Hemmati, P.; et al. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc Natl Acad Sci U S A 1999, 96, 10911–10916. [Google Scholar] [CrossRef]
- Saper, C.B.; Chou, T.C.; Elmquist, J.K. The need to feed: homeostatic and hedonic control of eating. Neuron 2002, 36, 199–211. [Google Scholar] [CrossRef]
- Burlet, S.; Tyler, C.J.; Leonard, C.S. Direct and indirect excitation of laterodorsal tegmental neurons by Hypocretin/Orexin peptides: implications for wakefulness and narcolepsy. J Neurosci 2002, 22, 2862–2872. [Google Scholar] [CrossRef]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic regulation of sleep and circadian rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wu, F.; Zhang, M.; Ding, D.; Fan, S.; Chen, G.; Zhang, J.; Wang, L. The Interaction Between the Ventrolateral Preoptic Nucleus and the Tuberomammillary Nucleus in Regulating the Sleep-Wakefulness Cycle. Front Neurosci 2020, 14, 615854. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, E.; Fuller, P.M. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Barcomb, K.; Olah, S.S.; Kennedy, M.J.; Ford, C.P. Properties and modulation of excitatory inputs to the locus coeruleus. J Physiol 2022, 600, 4897–4916. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the ventrolateral preoptic nucleus. J Neurosci 2002, 22, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, E.; Yanagisawa, M.; Sakurai, T.; Mieda, M. Orexin neurons suppress narcolepsy via 2 distinct efferent pathways. J Clin Invest 2014, 124, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.; Yamanaka, A. The role of orexin neuron activity in sleep/wakefulness regulation. Peptides 2023, 165, 171007. [Google Scholar] [CrossRef]
- Zheng, H.; Patterson, L.M.; Berthoud, H.R. Orexin-A projections to the caudal medulla and orexin-induced c-Fos expression, food intake, and autonomic function. J Comp Neurol 2005, 485, 127–142. [Google Scholar] [CrossRef]
- Hurley, S.W.; Arseth, H.A.; Johnson, A.K. Orexin neurons couple neural systems mediating fluid balance with motivation-related circuits. Behav Neurosci 2018, 132, 284–292. [Google Scholar] [CrossRef]
- Backberg, M.; Hervieu, G.; Wilson, S.; Meister, B. Orexin receptor-1 (OX-R1) immunoreactivity in chemically identified neurons of the hypothalamus: focus on orexin targets involved in control of food and water intake. Eur J Neurosci 2002, 15, 315–328. [Google Scholar] [CrossRef]
- Plazzi, G.; Moghadam, K.K.; Maggi, L.S.; Donadio, V.; Vetrugno, R.; Liguori, R.; Zoccoli, G.; Poli, F.; Pizza, F.; Pagotto, U.; Ferri, R. Autonomic disturbances in narcolepsy. Sleep Med Rev 2011, 15, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Peleg-Raibstein, D.; Burdakov, D. Do orexin/hypocretin neurons signal stress or reward? Peptides 2021, 145, 170629. [Google Scholar] [CrossRef]
- Grafe, L.A.; Bhatnagar, S. Orexins and stress. Front Neuroendocrinol 2018, 51, 132–145. [Google Scholar] [CrossRef]
- Sargin, D. The role of the orexin system in stress response. Neuropharmacology 2019, 154, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Spinazzi, R.; Andreis, P.G.; Rossi, G.P.; Nussdorfer, G.G. Orexins in the regulation of the hypothalamic-pituitary-adrenal axis. Pharmacol Rev 2006, 58, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Tena-Sempere, M.; Dieguez, C. Cross-talk between orexins (hypocretins) and the neuroendocrine axes (hypothalamic-pituitary axes). Front Neuroendocrinol 2010, 31, 113–127. [Google Scholar] [CrossRef]
- Kohsaka, A.; Watanobe, H.; Kakizaki, Y.; Suda, T.; Schioth, H.B. A significant participation of orexin-A, a potent orexigenic peptide, in the preovulatory luteinizing hormone and prolactin surges in the rat. Brain Res 2001, 898, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.W.; Ranson, S.W. Hypothalamic lesions and adiposity in the rat. The Anatomical Record 1940, 78, 149–172. [Google Scholar] [CrossRef]
- Anand, B.K.; Brobeck, J.R. Hypothalamic control of food intake in rats and cats. Yale J Biol Med 1951, 24, 123–140. [Google Scholar]
- Oomura, Y.; Ono, T.; Ooyama, H.; Wayner, M.J. Glucose and osmosensitive neurones of the rat hypothalamus. Nature 1969, 222, 282–284. [Google Scholar] [CrossRef]
- Funahashi, H.; Takenoya, F.; Guan, J.L.; Kageyama, H.; Yada, T.; Shioda, S. Hypothalamic neuronal networks and feeding-related peptides involved in the regulation of feeding. Anat Sci Int 2003, 78, 123–138. [Google Scholar] [CrossRef]
- Elmquist, J.K.; Elias, C.F.; Saper, C.B. From lesions to leptin: hypothalamic control of food intake and body weight. Neuron 1999, 22, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T. Roles of orexins in the regulation of body weight homeostasis. Obes Res Clin Pract 2014, 8, e414–420. [Google Scholar] [CrossRef]
- Schuld, A.; Hebebrand, J.; Geller, F.; Pollmacher, T. Increased body-mass index in patients with narcolepsy. Lancet 2000, 355, 1274–1275. [Google Scholar] [CrossRef]
- Szekely, M.; Petervari, E.; Balasko, M. Thermoregulation, energy balance, regulatory peptides: recent developments. Front Biosci (Schol Ed) 2010, 2, 1009–1046. [Google Scholar] [CrossRef] [PubMed]
- Federici, L.M.; Caliman, I.F.; Molosh, A.I.; Fitz, S.D.; Truitt, W.A.; Bonaventure, P.; Carpenter, J.S.; Shekhar, A.; Johnson, P.L. Hypothalamic orexin’s role in exacerbated cutaneous vasodilation responses to an anxiogenic stimulus in a surgical menopause model. Psychoneuroendocrinology 2016, 65, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Folgueira, C.; Beiroa, D.; Porteiro, B.; Duquenne, M.; Puighermanal, E.; Fondevila, M.F.; Barja-Fernandez, S.; Gallego, R.; Hernandez-Bautista, R.; Castelao, C.; et al. Hypothalamic dopamine signaling regulates brown fat thermogenesis. Nat Metab 2019, 1, 811–829. [Google Scholar] [CrossRef]
- Sasson, R.; Dearth, R.K.; White, R.S.; Chappell, P.E.; Mellon, P.L. Orexin A induces GnRH gene expression and secretion from GT1-7 hypothalamic GnRH neurons. Neuroendocrinology 2006, 84, 353–363. [Google Scholar] [CrossRef]
- Di Sebastiano, A.R.; Wilson-Perez, H.E.; Lehman, M.N.; Coolen, L.M. Lesions of orexin neurons block conditioned place preference for sexual behavior in male rats. Horm Behav 2011, 59, 1–8. [Google Scholar] [CrossRef]
- Johnson, P.L.; Molosh, A.; Fitz, S.D.; Truitt, W.A.; Shekhar, A. Orexin, stress, and anxiety/panic states. Prog Brain Res 2012, 198, 133–161. [Google Scholar] [CrossRef]
- Palotai, M.; Telegdy, G.; Jaszberenyi, M. Orexin A-induced anxiety-like behavior is mediated through GABA-ergic, alpha- and beta-adrenergic neurotransmissions in mice. Peptides 2014, 57, 129–134. [Google Scholar] [CrossRef]
- Vanderhaven, M.W.; Cornish, J.L.; Staples, L.G. The orexin-1 receptor antagonist SB-334867 decreases anxiety-like behavior and c-Fos expression in the hypothalamus of rats exposed to cat odor. Behav Brain Res 2015, 278, 563–568. [Google Scholar] [CrossRef]
- Sakurai, T. The role of orexin in motivated behaviours. Nat Rev Neurosci 2014, 15, 719–731. [Google Scholar] [CrossRef]
- Sears, R.M.; Fink, A.E.; Wigestrand, M.B.; Farb, C.R.; de Lecea, L.; Ledoux, J.E. Orexin/hypocretin system modulates amygdala-dependent threat learning through the locus coeruleus. Proc Natl Acad Sci U S A 2013, 110, 20260–20265. [Google Scholar] [CrossRef]
- Soya, S.; Shoji, H.; Hasegawa, E.; Hondo, M.; Miyakawa, T.; Yanagisawa, M.; Mieda, M.; Sakurai, T. Orexin receptor-1 in the locus coeruleus plays an important role in cue-dependent fear memory consolidation. J Neurosci 2013, 33, 14549–14557. [Google Scholar] [CrossRef]
- Telegdy, G.; Adamik, A. The action of orexin A on passive avoidance learning. Involvement of transmitters. Regul Pept 2002, 104, 105–110. [Google Scholar] [CrossRef]
- Holland, P.; Goadsby, P.J. The hypothalamic orexinergic system: pain and primary headaches. Headache 2007, 47, 951–962. [Google Scholar] [CrossRef]
- Villano, I.; La Marra, M.; Di Maio, G.; Monda, V.; Chieffi, S.; Guatteo, E.; Messina, G.; Moscatelli, F.; Monda, M.; Messina, A. Physiological Role of Orexinergic System for Health. Int J Environ Res Public Health 2022, 19. [Google Scholar] [CrossRef]
- Vaseghi, S.; Zarrabian, S.; Haghparast, A. Reviewing the role of the orexinergic system and stressors in modulating mood and reward-related behaviors. Neurosci Biobehav Rev 2022, 133, 104516. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Szabó, Á.; Spekker, E.; Polyák, H.; Tóth, F.; Vécsei, L. Mitochondrial impairment: A common motif in neuropsychiatric presentation? The link to the tryptophan–kynurenine metabolic system. Cells 2022, 11, 2607. [Google Scholar] [CrossRef] [PubMed]
- Scammell, T.E. Narcolepsy. N Engl J Med 2015, 373, 2654–2662. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, C.E.; Cogswell, A.; Koralnik, I.J.; Scammell, T.E. The neurobiological basis of narcolepsy. Nat Rev Neurosci 2019, 20, 83–93. [Google Scholar] [CrossRef]
- Thannickal, T.C.; Moore, R.Y.; Nienhuis, R.; Ramanathan, L.; Gulyani, S.; Aldrich, M.; Cornford, M.; Siegel, J.M. Reduced number of hypocretin neurons in human narcolepsy. Neuron 2000, 27, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Scammell, T.E.; Winrow, C.J. Orexin receptors: pharmacology and therapeutic opportunities. Annu Rev Pharmacol Toxicol 2011, 51, 243–266. [Google Scholar] [CrossRef]
- Bonini, J.A.; Jones, K.A.; Adham, N.; Forray, C.; Artymyshyn, R.; Durkin, M.M.; Smith, K.E.; Tamm, J.A.; Boteju, L.W.; Lakhlani, P.P.; et al. Identification and characterization of two G protein-coupled receptors for neuropeptide FF. J Biol Chem 2000, 275, 39324–39331. [Google Scholar] [CrossRef] [PubMed]
- Laemmle, B.; Schindler, M.; Beilmann, M.; Hamilton, B.S.; Doods, H.N.; Wieland, H.A. Characterization of the NPGP receptor and identification of a novel short mRNA isoform in human hypothalamus. Regul Pept 2003, 111, 21–29. [Google Scholar] [CrossRef]
- Lin, L.; Faraco, J.; Li, R.; Kadotani, H.; Rogers, W.; Lin, X.; Qiu, X.; de Jong, P.J.; Nishino, S.; Mignot, E. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell 1999, 98, 365–376. [Google Scholar] [CrossRef]
- Chemelli, R.M.; Willie, J.T.; Sinton, C.M.; Elmquist, J.K.; Scammell, T.; Lee, C.; Richardson, J.A.; Williams, S.C.; Xiong, Y.; Kisanuki, Y.; et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell 1999, 98, 437–451. [Google Scholar] [CrossRef]
- Peyron, C.; Faraco, J.; Rogers, W.; Ripley, B.; Overeem, S.; Charnay, Y.; Nevsimalova, S.; Aldrich, M.; Reynolds, D.; Albin, R.; et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med 2000, 6, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Liblau, R.S.; Vassalli, A.; Seifinejad, A.; Tafti, M. Hypocretin (orexin) biology and the pathophysiology of narcolepsy with cataplexy. Lancet Neurol 2015, 14, 318–328. [Google Scholar] [CrossRef]
- Melmed, S.; Auchus, R.J.; Goldfine, A.B.; Koenig, R.J.; Rosen, C.J. Williams textbook of endocrinology, 14th edition. ed.; Elsevier: Philadelphia, PA, 2020; pp. xiv, 1777 pages. [Google Scholar]
- Cannon, W.B. THE EMERGENCY FUNCTION OF THE ADRENAL MEDULLA IN PAIN AND THE MAJOR EMOTIONS. American Journal of Physiology-Legacy Content 1914, 33, 356–372. [Google Scholar] [CrossRef]
- Selye, H. A Syndrome produced by Diverse Nocuous Agents. Nature 1936, 138, 32–32. [Google Scholar] [CrossRef]
- Wade, N. Guillemin and schally: a race spurred by rivalry. Science 1978, 200, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Vale, W.; Spiess, J.; Rivier, C.; Rivier, J. Characterization of a 41-residue ovine hypothalamic peptide that stimulates secretion of corticotropin and beta-endorphin. Science 1981, 213, 1394–1397. [Google Scholar] [CrossRef]
- Selye, H. The general adaptation syndrome and the diseases of adaptation. J Allergy 1946, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Badami, V.M.; Rice, C.D.; Lois, J.H.; Madrecha, J.; Yates, B.J. Distribution of hypothalamic neurons with orexin (hypocretin) or melanin concentrating hormone (MCH) immunoreactivity and multisynaptic connections with diaphragm motoneurons. Brain Res 2010, 1323, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Fadel, J.; Pasumarthi, R.; Reznikov, L.R. Stimulation of cortical acetylcholine release by orexin A. Neuroscience 2005, 130, 541–547. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Hoyer, D.; de Lecea, L. Hypocretins (orexins): The ultimate translational neuropeptides. J Intern Med 2022, 291, 533–556. [Google Scholar] [CrossRef]
- Kang, X.; Tang, H.; Liu, Y.; Yuan, Y.; Wang, M. Research progress on the mechanism of orexin in pain regulation in different brain regions. Open Life Sci 2021, 16, 46–52. [Google Scholar] [CrossRef]
- Li, S.B.; de Lecea, L. The hypocretin (orexin) system: from a neural circuitry perspective. Neuropharmacology 2020, 167, 107993. [Google Scholar] [CrossRef]
- Nambu, T.; Sakurai, T.; Mizukami, K.; Hosoya, Y.; Yanagisawa, M.; Goto, K. Distribution of orexin neurons in the adult rat brain. Brain Res 1999, 827, 243–260. [Google Scholar] [CrossRef]
- Pizza, F.; Barateau, L.; Dauvilliers, Y.; Plazzi, G. The orexin story, sleep and sleep disturbances. J Sleep Res 2022, 31, e13665. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, S.; Rodrigo-Angulo, M.L.; Núñez, A.; Garzón, M. Synaptic interactions between perifornical lateral hypothalamic area, locus coeruleus nucleus and the oral pontine reticular nucleus are implicated in the stage succession during sleep-wakefulness cycle. Front Neurosci 2013, 7, 216. [Google Scholar] [CrossRef]
- Bujdoso, E.; Jaszberenyi, M.; Tomboly, C.; Toth, G.; Telegdy, G. Behavioral and neuroendocrine actions of endomorphin-2. Peptides 2001, 22, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, G. Regulation of the hypothalamic-pituitary-adrenal axis by neuropeptides. Horm Mol Biol Clin Investig 2011, 7, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Bujdoso, E.; Jaszberenyi, M.; Tomboly, C.; Toth, G.; Telegdy, G. Effects of endomorphin-1 on open-field behavior and on the hypothalamic-pituitary-adrenal system. Endocrine 2001, 14, 221–224. [Google Scholar] [CrossRef]
- Rostene, W.H.; Alexander, M.J. Neurotensin and neuroendocrine regulation. Front Neuroendocrinol 1997, 18, 115–173. [Google Scholar] [CrossRef]
- Perras, B.; Schultes, B.; Behn, B.; Dodt, C.; Born, J.; Fehm, H.L. Intranasal atrial natriuretic peptide acts as central nervous inhibitor of the hypothalamo-pituitary-adrenal stress system in humans. J Clin Endocrinol Metab 2004, 89, 4642–4648. [Google Scholar] [CrossRef]
- Kuppusamy, T.; Ramaswamy, P.; Perumal, M.; Silambanan, S.; Prabu Kumar, A. A short note on oxytocin and stress attenuation. Bioinformation 2021, 17, 921–923. [Google Scholar] [CrossRef]
- Jaszberenyi, M.; Bujdoso, E.; Telegdy, G. Effects of C-type natriuretic peptide on pituitary-adrenal activation in rats. Neuroreport 1998, 9, 2601–2603. [Google Scholar] [CrossRef]
- Jaszberenyi, M.; Bujdoso, E.; Telegdy, G. Effects of brain natriuretic peptide on pituitary-adrenal activation in rats. Life Sci 2000, 66, 1655–1661. [Google Scholar] [CrossRef]
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur J Pharmacol 2003, 463, 235–272. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Kastin, A.J. Urocortin and the brain. Prog Neurobiol 2008, 84, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Henckens, M.J.; Deussing, J.M.; Chen, A. Region-specific roles of the corticotropin-releasing factor-urocortin system in stress. Nat Rev Neurosci 2016, 17, 636–651. [Google Scholar] [CrossRef]
- de Oliveira, C.V.; Rosas-Arellano, M.P.; Solano-Flores, L.P.; Ciriello, J. Cardiovascular effects of hypocretin-1 in nucleus of the solitary tract. Am J Physiol Heart Circ Physiol 2003, 284, H1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Samson, W.K.; Taylor, M.M.; Follwell, M.; Ferguson, A.V. Orexin actions in hypothalamic paraventricular nucleus: physiological consequences and cellular correlates. Regul Pept 2002, 104, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Winsky-Sommerer, R.; Yamanaka, A.; Diano, S.; Borok, E.; Roberts, A.J.; Sakurai, T.; Kilduff, T.S.; Horvath, T.L.; de Lecea, L. Interaction between the corticotropin-releasing factor system and hypocretins (orexins): a novel circuit mediating stress response. J Neurosci 2004, 24, 11439–11448. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, F.; Yamada, S.; Ueta, Y. Centrally administered orexin-A activates corticotropin-releasing factor-containing neurons in the hypothalamic paraventricular nucleus and central amygdaloid nucleus of rats: possible involvement of central orexins on stress-activated central CRF neurons. Regul Pept 2004, 118, 183–191. [Google Scholar] [CrossRef]
- Blasiak, A.; Gundlach, A.L.; Hess, G.; Lewandowski, M.H. Interactions of Circadian Rhythmicity, Stress and Orexigenic Neuropeptide Systems: Implications for Food Intake Control. Front Neurosci 2017, 11, 127. [Google Scholar] [CrossRef]
- Hirota, K.; Kushikata, T.; Kudo, M.; Kudo, T.; Lambert, D.G.; Matsuki, A. Orexin A and B evoke noradrenaline release from rat cerebrocortical slices. Br J Pharmacol 2001, 134, 1461–1466. [Google Scholar] [CrossRef]
- Brunton, P.J.; Bales, J.; Russell, J.A. Neuroendocrine stress but not feeding responses to centrally administered neuropeptide Y are suppressed in pregnant rats. Endocrinology 2006, 147, 3737–3745. [Google Scholar] [CrossRef]
- Russell, S.H.; Small, C.J.; Dakin, C.L.; Abbott, C.R.; Morgan, D.G.; Ghatei, M.A.; Bloom, S.R. The central effects of orexin-A in the hypothalamic-pituitary-adrenal axis in vivo and in vitro in male rats. J Neuroendocrinol 2001, 13, 561–566. [Google Scholar] [CrossRef]
- Furlong, T.M.; Vianna, D.M.; Liu, L.; Carrive, P. Hypocretin/orexin contributes to the expression of some but not all forms of stress and arousal. Eur J Neurosci 2009, 30, 1603–1614. [Google Scholar] [CrossRef]
- Kotz, C.M.; Wang, C.; Teske, J.A.; Thorpe, A.J.; Novak, C.M.; Kiwaki, K.; Levine, J.A. Orexin A mediation of time spent moving in rats: neural mechanisms. Neuroscience 2006, 142, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Kotz, C.M. Integration of feeding and spontaneous physical activity: role for orexin. Physiol Behav 2006, 88, 294–301. [Google Scholar] [CrossRef]
- Villano, I.; Messina, A.; Valenzano, A.; Moscatelli, F.; Esposito, T.; Monda, V.; Esposito, M.; Precenzano, F.; Carotenuto, M.; Viggiano, A.; et al. Basal Forebrain Cholinergic System and Orexin Neurons: Effects on Attention. Front Behav Neurosci 2017, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, S.; Wei, C.; Wang, H.; Sui, N.; Kirouac, G.J. Changes in emotional behavior produced by orexin microinjections in the paraventricular nucleus of the thalamus. Pharmacol Biochem Behav 2010, 95, 121–128. [Google Scholar] [CrossRef]
- Humphreys, R.K.; Ruxton, G.D. A review of thanatosis (death feigning) as an anti-predator behaviour. Behav Ecol Sociobiol 2018, 72, 22. [Google Scholar] [CrossRef]
- Peinkhofer, C.; Martial, C.; Cassol, H.; Laureys, S.; Kondziella, D. The evolutionary origin of near-death experiences: a systematic investigation. Brain Commun 2021, 3, fcab132. [Google Scholar] [CrossRef]
- Steimer, T. The biology of fear- and anxiety-related behaviors. Dialogues Clin Neurosci 2002, 4, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Preclinical modeling in depression and anxiety: Current challenges and future research directions. Adv. Clin. Exp. Med 2023, 32, 505–509. [Google Scholar] [CrossRef]
- Soya, S.; Sakurai, T. Orexin as a modulator of fear-related behavior: Hypothalamic control of noradrenaline circuit. Brain Res 2020, 1731, 146037. [Google Scholar] [CrossRef]
- Soya, S.; Takahashi, T.M.; McHugh, T.J.; Maejima, T.; Herlitze, S.; Abe, M.; Sakimura, K.; Sakurai, T. Orexin modulates behavioral fear expression through the locus coeruleus. Nat Commun 2017, 8, 1606. [Google Scholar] [CrossRef] [PubMed]
- Palotai, M.; Telegdy, G.; Ekwerike, A.; Jaszberenyi, M. The action of orexin B on passive avoidance learning. Involvement of neurotransmitters. Behav Brain Res 2014, 272, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.; Saravia, R.; Maldonado, R.; Berrendero, F. Orexins and fear: implications for the treatment of anxiety disorders. Trends Neurosci 2015, 38, 550–559. [Google Scholar] [CrossRef]
- Yoshida, K.; McCormack, S.; Espana, R.A.; Crocker, A.; Scammell, T.E. Afferents to the orexin neurons of the rat brain. J Comp Neurol 2006, 494, 845–861. [Google Scholar] [CrossRef]
- Avolio, E.; Alo, R.; Carelli, A.; Canonaco, M. Amygdalar orexinergic-GABAergic interactions regulate anxiety behaviors of the Syrian golden hamster. Behav Brain Res 2011, 218, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.A.; Lecourt, H.; Rakotoariniaina, A.; Jenck, F. Favoured genetic background for testing anxiolytics in the fear-potentiated and light-enhanced startle paradigms in the rat. Behav Brain Res 2011, 221, 34–42. [Google Scholar] [CrossRef]
- Camina, E.; Guell, F. The Neuroanatomical, Neurophysiological and Psychological Basis of Memory: Current Models and Their Origins. Front Pharmacol 2017, 8, 438. [Google Scholar] [CrossRef]
- Bahramzadeh Zoeram, S.; Elahdadi Salmani, M.; Lashkarbolouki, T.; Goudarzi, I. Hippocampal orexin receptor blocking prevented the stress induced social learning and memory deficits. Neurobiol Learn Mem 2019, 157, 12–23. [Google Scholar] [CrossRef]
- Katzman, M.A.; Katzman, M.P. Neurobiology of the Orexin System and Its Potential Role in the Regulation of Hedonic Tone. Brain Sci 2022, 12. [Google Scholar] [CrossRef]
- Piccoli, L.; Micioni Di Bonaventura, M.V.; Cifani, C.; Costantini, V.J.; Massagrande, M.; Montanari, D.; Martinelli, P.; Antolini, M.; Ciccocioppo, R.; Massi, M.; et al. Role of orexin-1 receptor mechanisms on compulsive food consumption in a model of binge eating in female rats. Neuropsychopharmacology 2012, 37, 1999–2011. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, F.; Juarez-Aguilar, E.; Santiago-Garcia, J.; Cardinali, D.P. Ghrelin and its interactions with growth hormone, leptin and orexins: implications for the sleep-wake cycle and metabolism. Sleep Med Rev 2014, 18, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Toshinai, K.; Date, Y.; Murakami, N.; Shimada, M.; Mondal, M.S.; Shimbara, T.; Guan, J.L.; Wang, Q.P.; Funahashi, H.; Sakurai, T.; et al. Ghrelin-induced food intake is mediated via the orexin pathway. Endocrinology 2003, 144, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Quarta, D.; Smolders, I. Rewarding, reinforcing and incentive salient events involve orexigenic hypothalamic neuropeptides regulating mesolimbic dopaminergic neurotransmission. Eur J Pharm Sci 2014, 57, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Matzeu, A.; Martin-Fardon, R. Understanding the Role of Orexin Neuropeptides in Drug Addiction: Preclinical Studies and Translational Value. Front Behav Neurosci 2021, 15, 787595. [Google Scholar] [CrossRef]
- James, M.H.; Stopper, C.M.; Zimmer, B.A.; Koll, N.E.; Bowrey, H.E.; Aston-Jones, G. Increased Number and Activity of a Lateral Subpopulation of Hypothalamic Orexin/Hypocretin Neurons Underlies the Expression of an Addicted State in Rats. Biol Psychiatry 2019, 85, 925–935. [Google Scholar] [CrossRef]
- Shaw, J.K.; Ferris, M.J.; Locke, J.L.; Brodnik, Z.D.; Jones, S.R.; Espana, R.A. Hypocretin/orexin knock-out mice display disrupted behavioral and dopamine responses to cocaine. Addict Biol 2017, 22, 1695–1705. [Google Scholar] [CrossRef]
- Steiner, N.; Rossetti, C.; Sakurai, T.; Yanagisawa, M.; de Lecea, L.; Magistretti, P.J.; Halfon, O.; Boutrel, B. Hypocretin/orexin deficiency decreases cocaine abuse liability. Neuropharmacology 2018, 133, 395–403. [Google Scholar] [CrossRef]
- McGregor, R.; Thannickal, T.C.; Siegel, J.M. Pleasure, addiction, and hypocretin (orexin). Handb Clin Neurol 2021, 180, 359–374. [Google Scholar] [CrossRef]
- Mohammadkhani, A.; Fragale, J.E.; Pantazis, C.B.; Bowrey, H.E.; James, M.H.; Aston-Jones, G. Orexin-1 Receptor Signaling in Ventral Pallidum Regulates Motivation for the Opioid Remifentanil. J Neurosci 2019, 39, 9831–9840. [Google Scholar] [CrossRef]
- Mohammadkhani, A.; James, M.H.; Pantazis, C.B.; Aston-Jones, G. Persistent effects of the orexin-1 receptor antagonist SB-334867 on motivation for the fast acting opioid remifentanil. Brain Res 2020, 1731, 146461. [Google Scholar] [CrossRef] [PubMed]
- Morganstern, I.; Chang, G.Q.; Barson, J.R.; Ye, Z.; Karatayev, O.; Leibowitz, S.F. Differential effects of acute and chronic ethanol exposure on orexin expression in the perifornical lateral hypothalamus. Alcohol Clin Exp Res 2010, 34, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Rotter, A.; Bayerlein, K.; Hansbauer, M.; Weiland, J.; Sperling, W.; Kornhuber, J.; Biermann, T. Orexin A expression and promoter methylation in patients with cannabis dependence in comparison to nicotine-dependent cigarette smokers and nonsmokers. Neuropsychobiology 2012, 66, 126–133. [Google Scholar] [CrossRef]
- Bayerlein, K.; Kraus, T.; Leinonen, I.; Pilniok, D.; Rotter, A.; Hofner, B.; Schwitulla, J.; Sperling, W.; Kornhuber, J.; Biermann, T. Orexin A expression and promoter methylation in patients with alcohol dependence comparing acute and protracted withdrawal. Alcohol 2011, 45, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Skofitsch, G.; Jacobowitz, D.M.; Zamir, N. Immunohistochemical localization of a melanin concentrating hormone-like peptide in the rat brain. Brain Res Bull 1985, 15, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschop, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003, 37, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, F.; Salio, C.; Lossi, L.; Merighi, A. Ghrelin in central neurons. Curr Neuropharmacol 2009, 7, 37–49. [Google Scholar] [CrossRef]
- Mori, K.; Miyazato, M.; Ida, T.; Murakami, N.; Serino, R.; Ueta, Y.; Kojima, M.; Kangawa, K. Identification of neuromedin S and its possible role in the mammalian circadian oscillator system. EMBO J 2005, 24, 325–335. [Google Scholar] [CrossRef]
- Saito, Y.; Nagasaki, H. The melanin-concentrating hormone system and its physiological functions. Results Probl Cell Differ 2008, 46, 159–179. [Google Scholar] [CrossRef]
- DiBona, G.F. Neuropeptide Y. Am J Physiol Regul Integr Comp Physiol 2002, 282, R635–636. [Google Scholar] [CrossRef]
- Antushevich, H.; Wojcik, M. Review: Apelin in disease. Clin Chim Acta 2018, 483, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Telegdy, G.; Adamik, A.; Jaszberenyi, M. Involvement of neurotransmitters in the action of apelin-13 on passive avoidance learning in mice. Peptides 2013, 39, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Telegdy, G.; Jaszberenyi, M. Transmitter mediation of the anxiolytic action of apelin-13 in male mice. Behav Brain Res 2014, 263, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Kangawa, K. Ghrelin: structure and function. Physiol Rev 2005, 85, 495–522. [Google Scholar] [CrossRef]
- Malendowicz, L.K.; Rucinski, M. Neuromedins NMU and NMS: An Updated Overview of Their Functions. Front Endocrinol (Lausanne) 2021, 12, 713961. [Google Scholar] [CrossRef]
- Holland, P.R. Biology of Neuropeptides: Orexinergic Involvement in Primary Headache Disorders. Headache 2017, 57 Suppl 2, 76–88. [Google Scholar] [CrossRef]
- Horvath, T.L.; Diano, S.; van den Pol, A.N. Synaptic interaction between hypocretin (orexin) and neuropeptide Y cells in the rodent and primate hypothalamus: a novel circuit implicated in metabolic and endocrine regulations. J Neurosci 1999, 19, 1072–1087. [Google Scholar] [CrossRef]
- Kageyama, H.; Takenoya, F.; Hirako, S.; Wada, N.; Kintaka, Y.; Inoue, S.; Ota, E.; Ogawa, T.; Shioda, S. Neuronal circuits involving neuropeptide Y in hypothalamic arcuate nucleus-mediated feeding regulation. Neuropeptides 2012, 46, 285–289. [Google Scholar] [CrossRef]
- Russell, S.H.; Small, C.J.; Kennedy, A.R.; Stanley, S.A.; Seth, A.; Murphy, K.G.; Taheri, S.; Ghatei, M.A.; Bloom, S.R. Orexin A interactions in the hypothalamo-pituitary gonadal axis. Endocrinology 2001, 142, 5294–5302. [Google Scholar] [CrossRef]
- Funahashi, H.; Yamada, S.; Kageyama, H.; Takenoya, F.; Guan, J.L.; Shioda, S. Co-existence of leptin- and orexin-receptors in feeding-regulating neurons in the hypothalamic arcuate nucleus-a triple labeling study. Peptides 2003, 24, 687–694. [Google Scholar] [CrossRef]
- Horvath, T.L.; Peyron, C.; Diano, S.; Ivanov, A.; Aston-Jones, G.; Kilduff, T.S.; van Den Pol, A.N. Hypocretin (orexin) activation and synaptic innervation of the locus coeruleus noradrenergic system. J Comp Neurol 1999, 415, 145–159. [Google Scholar] [CrossRef]
- Chen, B.; Xu, J.; Chen, S.; Mou, T.; Wang, Y.; Wang, H.; Zhang, Z.; Ren, F.; Wang, Z.; Jin, K.; Lu, J. Dysregulation of striatal dopamine D2/D3 receptor-mediated by hypocretin induces depressive behaviors in rats. J Affect Disord 2023, 325, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Huang, H.; Chen, X.; Deng, S.; Zhu, C.; Huang, H.; Li, G. Structural and functional characterization of neuromedin S in the teleost fish, zebrafish (Danio rerio). Comp Biochem Physiol B Biochem Mol Biol 2016, 191, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Fronczek, R.; Schinkelshoek, M.; Shan, L.; Lammers, G.J. The orexin/hypocretin system in neuropsychiatric disorders: Relation to signs and symptoms. Handb Clin Neurol 2021, 180, 343–358. [Google Scholar] [CrossRef]
- Dauvilliers, Y.; Arnulf, I.; Mignot, E. Narcolepsy with cataplexy. Lancet 2007, 369, 499–511. [Google Scholar] [CrossRef]
- Wang, W.; Pan, Y.; Li, Q.; Wang, L. Orexin: a potential role in the process of obstructive sleep apnea. Peptides 2013, 42, 48–54. [Google Scholar] [CrossRef]
- Sakurai, S.; Nishijima, T.; Arihara, Z.; Takahashi, K. Plasma orexin-A levels in obstructive sleep apnea-hypopnea syndrome. Chest 2004, 125, 1963. [Google Scholar] [CrossRef]
- Nishijima, T.; Sakurai, S.; Arihara, Z.; Takahashi, K. Plasma orexin-A-like immunoreactivity in patients with sleep apnea hypopnea syndrome. Peptides 2003, 24, 407–411. [Google Scholar] [CrossRef]
- Seifinejad, A.; Ramosaj, M.; Shan, L.; Li, S.; Possovre, M.L.; Pfister, C.; Fronczek, R.; Garrett-Sinha, L.A.; Frieser, D.; Honda, M.; et al. Epigenetic silencing of selected hypothalamic neuropeptides in narcolepsy with cataplexy. Proc Natl Acad Sci U S A 2023, 120, e2220911120. [Google Scholar] [CrossRef]
- Partinen, M.; Kornum, B.R.; Plazzi, G.; Jennum, P.; Julkunen, I.; Vaarala, O. Narcolepsy as an autoimmune disease: the role of H1N1 infection and vaccination. Lancet Neurol 2014, 13, 600–613. [Google Scholar] [CrossRef]
- Gorka, S.M.; Khorrami, K.J.; Manzler, C.A.; Phan, K.L. Acute orexin antagonism selectively modulates anticipatory anxiety in humans: implications for addiction and anxiety. Transl Psychiatry 2022, 12, 308. [Google Scholar] [CrossRef]
- Rahman, Q.F.A.; Jufri, N.F.; Hamid, A. Hyperphagia in Prader-Willi syndrome with obesity: From development to pharmacological treatment. Intractable Rare Dis Res 2023, 12, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mehr, J.B.; Mitchison, D.; Bowrey, H.E.; James, M.H. Sleep dysregulation in binge eating disorder and "food addiction": the orexin (hypocretin) system as a potential neurobiological link. Neuropsychopharmacology 2021, 46, 2051–2061. [Google Scholar] [CrossRef]
- Berner, L.A.; Brown, T.A.; Lavender, J.M.; Lopez, E.; Wierenga, C.E.; Kaye, W.H. Neuroendocrinology of reward in anorexia nervosa and bulimia nervosa: Beyond leptin and ghrelin. Mol Cell Endocrinol 2019, 497, 110320. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Han, F.; Dong, S.X.; Li, J.; An, P.; Zhang, X.Z.; Chang, Y.; Zhao, L.; Zhang, X.L.; Liu, Y.N.; et al. Cerebrospinal Fluid Orexin A Levels and Autonomic Function in Kleine-Levin Syndrome. Sleep 2016, 39, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Razzoli, M.; Bartolomucci, A. The Dichotomous Effect of Chronic Stress on Obesity. Trends Endocrinol Metab 2016, 27, 504–515. [Google Scholar] [CrossRef]
- Yilmaz, E.; Celik, O.; Celik, N.; Celik, E.; Turkcuoglu, I.; Simsek, Y.; Minareci, Y.; Boz, M.; Aydin, S. Maternal and fetal serum orexin-A levels in gestational diabetes mellitus. J Obstet Gynaecol Res 2013, 39, 139–145. [Google Scholar] [CrossRef]
- Yilmaz, E.; Celik, O.; Celik, N.; Simsek, Y.; Celik, E.; Yildirim, E. Serum orexin-A (OXA) level decreases in polycystic ovarian syndrome. Gynecol Endocrinol 2013, 29, 388–390. [Google Scholar] [CrossRef]
- Celik, O.; Aydin, S.; Celik, N.; Yilmaz, M. Peptides: Basic determinants of reproductive functions. Peptides 2015, 72, 34–43. [Google Scholar] [CrossRef]
- Jequier, E. Leptin signaling, adiposity, and energy balance. Ann N Y Acad Sci 2002, 967, 379–388. [Google Scholar] [CrossRef]
- Hopf, F.W. Recent perspectives on orexin/hypocretin promotion of addiction-related behaviors. Neuropharmacology 2020, 168, 108013. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Proudnikov, D.; Yuferov, V.; Kreek, M.J. Drug-induced and genetic alterations in stress-responsive systems: Implications for specific addictive diseases. Brain Res 2010, 1314, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Berteotti, C.; Calvello, C.; Liguori, C. Role of the orexin system in the bidirectional relation between sleep and epilepsy: New chances for patients with epilepsy by the antagonism to orexin receptors? Epilepsia 2023. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.R.; Molosh, A.I.; Johnson, P.L.; Shekhar, A. Role of medial hypothalamic orexin system in panic, phobia and hypertension. Brain Res 2020, 1731, 145942. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Lakis, G.A.; Zhoba, H. Sleep-wake and arousal dysfunctions in post-traumatic stress disorder: Role of orexin systems. Brain Res Bull 2022, 186, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Brundin, L.; Bjorkqvist, M.; Petersen, A.; Traskman-Bendz, L. Reduced orexin levels in the cerebrospinal fluid of suicidal patients with major depressive disorder. Eur Neuropsychopharmacol 2007, 17, 573–579. [Google Scholar] [CrossRef]
- Salomon, R.M.; Ripley, B.; Kennedy, J.S.; Johnson, B.; Schmidt, D.; Zeitzer, J.M.; Nishino, S.; Mignot, E. Diurnal variation of cerebrospinal fluid hypocretin-1 (Orexin-A) levels in control and depressed subjects. Biol Psychiatry 2003, 54, 96–104. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Abdulhadi, M.H.; Hussien, N.R.; Al-Niemi, M.S.; Rasheed, H.A.; Al-Gareeb, A.I. Involvement of orexinergic system in psychiatric and neurodegenerative disorders: A scoping review. Brain Circ 2020, 6, 70–80. [Google Scholar] [CrossRef]
- Toor, B.; Ray, L.B.; Pozzobon, A.; Fogel, S.M. Sleep, Orexin and Cognition. Front Neurol Neurosci 2021, 45, 38–51. [Google Scholar] [CrossRef]
- Mobarakeh, J.I.; Takahashi, K.; Sakurada, S.; Nishino, S.; Watanabe, H.; Kato, M.; Naghdi, N.; Yanai, K. Enhanced antinociception by intracerebroventricularly administered orexin A in histamine H1 or H2 receptor gene knockout mice. Pain 2005, 118, 254–262. [Google Scholar] [CrossRef]
- Watanabe, S.; Kuwaki, T.; Yanagisawa, M.; Fukuda, Y.; Shimoyama, M. Persistent pain and stress activate pain-inhibitory orexin pathways. Neuroreport 2005, 16, 5–8. [Google Scholar] [CrossRef]
- Li, S.B.; Jones, J.R.; de Lecea, L. Hypocretins, Neural Systems, Physiology, and Psychiatric Disorders. Curr Psychiatry Rep 2016, 18, 7. [Google Scholar] [CrossRef]
- Kalliomaki, M.L.; Panula, P. Neuropeptide FF, but not prolactin-releasing peptide, mRNA is differentially regulated in the hypothalamic and medullary neurons after salt loading. Neuroscience 2004, 124, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Roehrs, T.; Withrow, D.; Koshorek, G.; Verkler, J.; Bazan, L.; Roth, T. Sleep and pain in humans with fibromyalgia and comorbid insomnia: double-blind, crossover study of suvorexant 20 mg versus placebo. J Clin Sleep Med 2020, 16, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.; May, A. Diagnosis, pathophysiology, and management of cluster headache. Lancet Neurol 2018, 17, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Schmidt, A.; Hassel, S.; Tanaka, M. Editorial: Case reports in neuroimaging and stimulation. Front Psychiatry 2023, 14, 1264669. [Google Scholar] [CrossRef]
- Tanaka, M.; Diano, M.; Battaglia, S. Editorial: Insights into structural and functional organization of the brain: evidence from neuroimaging and non-invasive brain stimulation techniques. Front Psychiatry 2023, 14, 1225755. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Battaglia, S.; Sciamanna, G.; Tortora, F.; Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci Biobehav Rev 2021, 127, 334–352. [Google Scholar] [CrossRef] [PubMed]
- Navarro, V.M. Metabolic regulation of kisspeptin - the link between energy balance and reproduction. Nat Rev Endocrinol 2020, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Couvineau, A.; Nicole, P.; Gratio, V.; Voisin, T. The Orexin receptors: Structural and anti-tumoral properties. Front Endocrinol (Lausanne) 2022, 13, 931970. [Google Scholar] [CrossRef]
- Voisin, T.; Nicole, P.; Gratio, V.; Chassac, A.; Mansour, D.; Rebours, V.; Couvelard, A.; Couvineau, A. The Orexin-A/OX1R System Induces Cell Death in Pancreatic Cancer Cells Resistant to Gemcitabine and Nab-Paclitaxel Treatment. Front Oncol 2022, 12, 904327. [Google Scholar] [CrossRef]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; Le Poul, E.; Brezillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J Biol Chem 2001, 276, 34631–34636. [Google Scholar] [CrossRef]
- Wahab, F.; Atika, B.; Shahab, M.; Behr, R. Kisspeptin signalling in the physiology and pathophysiology of the urogenital system. Nat Rev Urol 2016, 13, 21–32. [Google Scholar] [CrossRef]
- Ten-Blanco, M.; Flores, A.; Cristino, L.; Pereda-Perez, I.; Berrendero, F. Targeting the orexin/hypocretin system for the treatment of neuropsychiatric and neurodegenerative diseases: From animal to clinical studies. Front Neuroendocrinol 2023, 69, 101066. [Google Scholar] [CrossRef]
- Ropper, A.H.; Samuels, M.A.; Klein, J.; Prasad, S. Adams and Victor’s principles of neurology. 2023, 1 online resource.
- McKnight, R.; Price, J.; Geddes, J. Psychiatry, Fifth edition. ed.; Oxford University Press: New York, NY, 2019; pp. x, 542 pages. [Google Scholar]
- Tanaka, M.; Szabó, Á.; Vécsei, L.; Giménez-Llort, L. Emerging translational research in neurological and psychiatric diseases: from in vitro to in vivo models. 2023, 24, 15739. [CrossRef]
- Tiwari, S.B.; Amiji, M.M. A review of nanocarrier-based CNS delivery systems. Curr Drug Deliv 2006, 3, 219–232. [Google Scholar] [CrossRef]
- Cummings, D.E. Ghrelin and the short- and long-term regulation of appetite and body weight. Physiol Behav 2006, 89, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A.J.; Pan, W.; Maness, L.M.; Banks, W.A. Peptides crossing the blood-brain barrier: some unusual observations. Brain Res 1999, 848, 96–100. [Google Scholar] [CrossRef]
- Milbank, E.; Lopez, M. Orexins/Hypocretins: Key Regulators of Energy Homeostasis. Front Endocrinol (Lausanne) 2019, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Biswas, D.A. Physiological Role of Orexin/Hypocretin in the Human Body in Motivated Behavior: A Comprehensive Review. Cureus 2023, 15, e34009. [Google Scholar] [CrossRef] [PubMed]
- Baimel, C.; Borgland, S.L. Orexin Signaling in the VTA Gates Morphine-Induced Synaptic Plasticity. J Neurosci 2015, 35, 7295–7303. [Google Scholar] [CrossRef] [PubMed]
- Baimel, C.; Bartlett, S.E.; Chiou, L.C.; Lawrence, A.J.; Muschamp, J.W.; Patkar, O.; Tung, L.W.; Borgland, S.L. Orexin/hypocretin role in reward: implications for opioid and other addictions. Br J Pharmacol 2015, 172, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Baimel, C.; Bartlett, S.E.; Chiou, L.C.; Lawrence, A.J.; Muschamp, J.W.; Patkar, O.; Tung, L.W.; Borgland, S.L. Orexin/hypocretin role in reward: implications for opioid and other addictions. British journal of pharmacology 2015, 172, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Brundin, L.; Petersen, A.; Bjorkqvist, M.; Traskman-Bendz, L. Orexin and psychiatric symptoms in suicide attempters. J Affect Disord 2007, 100, 259–263. [Google Scholar] [CrossRef]
- Tanaka, M.; Vecsei, L. Editorial of Special Issue "Crosstalk between Depression, Anxiety, and Dementia: Comorbidity in Behavioral Neurology and Neuropsychiatry". Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Muehlan, C.; Roch, C.; Vaillant, C.; Dingemanse, J. The orexin story and orexin receptor antagonists for the treatment of insomnia. J Sleep Res 2023, 32, e13902. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Editorial of special issue “crosstalk between depression, anxiety, and dementia: comorbidity in behavioral neurology and neuropsychiatry”. 2021, 9, 517. [CrossRef]
- Tanaka, M.; Vécsei, L. Monitoring the kynurenine system: Concentrations, ratios or what else? Advances in Clinical and Experimental Medicine 2021, 30, 775–778. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Monitoring the redox status in multiple sclerosis. Biomedicines 2020, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Mattar, P.; Uribe-Cerda, S.; Pezoa, C.; Guarnieri, T.; Kotz, C.M.; Teske, J.A.; Morselli, E.; Perez-Leighton, C. Brain site-specific regulation of hedonic intake by orexin and DYN peptides: role of the PVN and obesity. Nutr Neurosci 2022, 25, 1105–1114. [Google Scholar] [CrossRef]
- Butterick, T.A.; Billington, C.J.; Kotz, C.M.; Nixon, J.P. Orexin: pathways to obesity resistance? Rev Endocr Metab Disord 2013, 14, 357–364. [Google Scholar] [CrossRef]
- Perez-Leighton, C.E.; Butterick-Peterson, T.A.; Billington, C.J.; Kotz, C.M. Role of orexin receptors in obesity: from cellular to behavioral evidence. Int J Obes (Lond) 2013, 37, 167–174. [Google Scholar] [CrossRef]
- Seale, P. Orexin turns up the heat on obesity. Cell Metab 2011, 14, 441–442. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Chen, C. Towards a mechanistic understanding of depression, anxiety, and their comorbidity: perspectives from cognitive neuroscience. Frontiers in behavioral neuroscience 2023, 17. [Google Scholar] [CrossRef] [PubMed]
- Tajti, J.; Szok, D.; Csáti, A.; Szabó, Á.; Tanaka, M.; Vécsei, L. Exploring novel therapeutic targets in the common pathogenic factors in migraine and neuropathic pain. International Journal of Molecular Sciences 2023, 24, 4114. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Szabó, Á.; Körtési, T.; Szok, D.; Tajti, J.; Vécsei, L. From CGRP to PACAP, VIP, and Beyond: Unraveling the Next Chapters in Migraine Treatment. Cells 2023, 12, 2649. [Google Scholar] [CrossRef]
- Tanaka, M.; Bohár, Z.; Vécsei, L. Are kynurenines accomplices or principal villains in dementia? Maintenance of kynurenine metabolism. Molecules 2020, 25, 564. [Google Scholar]
- Krause, A.; Lott, D.; Brussee, J.M.; Muehlan, C.; Dingemanse, J. Population pharmacokinetic modeling of daridorexant, a novel dual orexin receptor antagonist. CPT Pharmacometrics Syst Pharmacol 2023, 12, 74–86. [Google Scholar] [CrossRef]

Table 1.
Diseases and disorders linked to the orexin/hypocretin system.
| Diseases and disorders |
|---|
| narcolepsy, cataplexy |
| obstructive sleep apnea-hypopnea syndrome |
| obesity |
| Prader-Willi syndrome |
| bulimia, anorexia nervosa |
| Kleine-Levin syndrome |
| gestational diabetes |
| polycystic ovary syndrome |
| attention deficit hyperactivity disorder |
| epilepsy |
| Panic disorder |
| phobia |
| major depressive disorder |
| post-traumatic stress disorder |
| psychosomatic disorders |
| hypertension |
| fibromyalgia |
| chronic pain disorders (migraine and cluster headaches) |
| schizophrenia |
| Parkinson’s disease |
| Alzheimer’s disease |
| Huntington’s disease |
| multiple sclerosis |
| amyotrophic lateral sclerosis |
Table 2.
The outline of the orexin/hypocretin connectome.
| Cell body location | Target region | Receptor | Function |
| LHA, DMH | VMH, ARC, PVN, NAc | OX1 | Stimulation of feeding |
| LHA, PFA | PAG, NTS, PVN, RVLM, RVMM, VTA | OX1, OX2 | Autonomic responses (RR↑, (tachycardia, thermoregulation) |
| PFA, DMH | LA, LC, PPT | OX1 | Emotions (anxiety, fear, mood) |
| LHA, DMH | VTA, NAc, DR | OX1, OX2 | Reward, addiction |
| LHA, DMH | PVN, POA | OX1, OX2 | Endocrine responses (stress response, gonadal functions) |
| PFA, LHA | LC, DR, VTA, TMN | OX2, OX1 | Arousal, Wakefulness |
ARC: arcuate nucleus, DMH: dorsomedial hypothalamus, DR: dorsal raphe, LA: lateral amygdala, LC: locus coeruleus, LHA: lateral hypothalamic area, NAc: nucleus accumbens, NTS: nucleus of the solitary tract, OX1: orexin 1 receptor, OX2: orexin 2 receptor, PFA: perifornical area, POA: preoptic area, PVN: paraventricular nucleus, PPT: pedunculopontine tegmental nucleus, RVLM: rostral ventrolateral medulla, RVMM: rostral ventromedial medulla, TMN: tuberomammillary nucleus [46,55,64,74,98,99,100,101,102,103,104,105].
Table 3.
Classes and indications of hypocretinergic drugs under clinical studies.
| Classes | Indications |
| Hypocretin receptors 1 & 2 antagonists | Insomnia, narcolepsy |
| Hypocretin receptors 1 antagonists | Panic disorder |
| Major depressive disorder | |
| anxiety | |
| Hypocretin receptors 2 antagonist | Binge eating disorder |
| Hypocretin receptors 2 agonist | narcolepsy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



