
Submitted:
19 January 2024
Posted:
23 January 2024
You are already at the latest version
Abstract
Human language and social cognition are two key disciplines that have traditionally been studied as separate domains. However, emerging research from detailed studies of comparative animal behavior, human social behavior, language acquisition in children, social cognitive neuroscience, and the cognitive neuroscience of language suggest that language and social cognition are two deeply interrelated capacities of the human species. For instance, recent developmental studies on the acquisition of language in infants and young children, draw attention to the important crucial benefits associated with several different aspects of social stimulation for youngsters, including the quantity and quality of linguistic input, dyadic infant/child-to-parent verbal and non-verbal social interactions, and other important social cues integral for facilitating language learning and social bonding. Studies of the adult human brain further suggest a high degree of specialization for sociolinguistic information processing, memory retrieval, and comprehension, suggesting the function of these regions may link language with social cognition and social bonding. In effect, the social brain and social complexity hypotheses may jointly help to explain how neurotypical children and adults learn language, why autistic adults and children exhibit simultaneous deficits in language and social cognition, and why nonhuman primates and other animals with significant computational abilities do not acquire the capacity for language. But perhaps most critically, the following article argues that this and related research will allow scientists to generate a holistic profile and deeper understanding of the healthy adult social brain, while developing more innovative and effective diagnoses, prognoses, and treatments for maladies and deficits also associated with the social brain.
Keywords:
social brain
; language brain
; social brain hypothesis
; social complexity hypothesis for animal communication
; social bonding
; social cognitive neuroscience
; language acquisition
1. Introduction
Human beings are an incredibly sophisticated species: technologically, scientifically, and cerebrally - why is this? The social brain hypothesis posits that the cognitive demands of living in complex social animal groups, selected for overall increases in brain volume, explaining in large part the unusually large brains of many primate species [1,2,3]. The initial evidence for this proposal came primarily from the finding that neocortex size correlates with social group size for a variety of primate species, including humans see Figure 1; for a review, see [4]. Since this initial finding, a considerable body of empirical work now indicates that, among primates, neocortex size correlates with many measures of social complexity: the frequency of coalitions, tactical deception, and social learning, the prevalence of social play, number of females in the group, grooming clique size (often used to facilitate social bonding), and of course, social group size [1]. In primates, this behavioral flexibility is partially reflected in the use of intense social bonds (often facilitated by social grooming) to prevent social groups from dissolving under these pressures.
A natural extension of the social brain hypothesis, the social complexity hypothesis for animal communication (also known, more colloquially, as the social bonding hypothesis), posits that animal groups with complex social systems require more complex communicative systems to regulate interactions among group members, including humans, and facilitate social bonding [5,6]. The initial evidence for this hypothesis originated from a comparative analysis of over forty species of non-human primates, in which it was found that both social group size and social grooming time (as a measure of social bonding) were positively associated with vocal repertoire size across these primates see Figure 2; [7]. Subsequent studies have further found larger vocal repertoire sizes to be associated with the long-term courtship bonds found among large, structurally complex gelada baboon social groupings, when compared to the more transient courtships of much smaller, less complex chacma baboon groupings [8]. In humans, more recent studies have confirmed two key predictions of the social complexity hypothesis, in that group size and social density processes appear to be related to communicative complexity and social bonding in human social networks [9,10].
Intriguingly, additional evidence for the social bonding function of human language, arguably the most complex of all primate communication systems, comes from detailed studies of human social behavior. In particular, studies of human conversational content reveal, both in traditional cultures as well as in Western industrial societies, that social gossip topics predominate in daily dialogue, typically encompassing over two-thirds of daily conversation time [11,12]. Moreover, further analyses have shown that the spread of information, relevant to personal reputation via gossip, promotes prosocial behavior by encouraging mindful partner selection when situations require cooperation [13]. In addition, feelings of relationship closeness have also been found to be promoted via social gossip, whereby the influence is generally strongest when individuals share a weakly held, negative view of an absent third party [14]. Lastly, agent-based simulation models have revealed similar findings, namely, that the evolution of communicative complexity depends upon an ecological demand for large social groups, as biological fitness improves with increasing group size as the social network gains more independent sources of information [15].
2. The social brain and social cognitive neuroscience
Despite the clear anthropological and evolutionary connection between the social brain, social cognition, and social behavior, described above, this framework has not yet been fully integrated into our current understanding of social cognitive neuroscience. In truth, the complexity of the neurological processes that underlie social living in primates is enormous, including activities such as coalition formation, tactical deception, organizing grooming cliques, social play, and social learning [1]. In humans, a complex network of brain regions underlie important activities such as the detection and processing of social stimuli, recognising faces, evaluating mental states (i.e. mentalizing or theory of mind), perceiving emotions, sharing attention, determining friends from foes, evaluating others’ perceptions and beliefs, social learning, relationship formation, and social bonding [1,16,17].
In an early and influential model of the social brain, neuroscientist Leslie Brothers [18] emphasized contributions of the amygdala, superior temporal sulcus (STS), orbitofrontal cortex (OFC), and fusiform gyrus (FFG) to social information processing and social perception. More recently, functional magnetic resonance imaging (fMRI) technologies of the brain have provided further identification of a complex interconnected network of neural regions linking the parietal and temporal lobes to the prefrontal lobes of the brain [4]. In particular, these include the parietal association cortex, OFC, dorsolateral prefrontal cortex, amygdala, anterior cingulate cortex (ACC), and superior temporal gyrus (STG) see Figure 3; [16,19].
Speaking broadly, the OFC has been strongly implicated in social reinforcement and social reward processing [16,19]. More specifically, the STS region, particularly the right-hemisphere posterior STS (pSTS) area, has been found to analyze biological motion cues, including the hand, eye, and other salient body motions in order to predict and interpret the intentions and behaviors of other agents [16,19]. In addition to this area, the right inferior temporal gyrus, occipital part of the fusiform gyrus, right parietal lobule, and middle temporal gyrus in both hemispheres are differentially activated by processing the direction of gaze [16,19]. Several areas have been further implicated in empathy and emotional perception. For example, a complex structure that is highly interconnected with cortical and subcortical structures of the brain, the amygdala, has been implicated in recognizing others’ emotional states through facial expression processing and analysis, as well as in the regulation and experience of internal emotional states [16,19]. Furthermore, the FFG, located in the ventral occipitotemporal cortex, contains an area referred to as the fusiform face area (FFA), which has been implicated specifically in face detection and recognition see Figure 3; [16,19].
Lastly, the amygdala, precuneus, medial prefrontal cortex (mPFC), pSTS, right temporoparietal junction (rTPJ), and temporopolar cortex play an important role in evaluating mental states (i.e. mentalizing or theory of mind) [16,19]. Further, several studies now show that the social brain hypothesis not only explains variation in brain volume between various primate species, but also individual differences within the human species, with respect to many aspects of social cognition and social networking. In particular, gray matter volume in the OFC, ACC, ventromedial prefrontal cortex (vmPFC), amygdala, and temporal lobe, including the STS, is correlated with individual differences in higher-order intentionality capacity (i.e. advanced mentalizing or theory of mind) and social network size [20,21,22,23]. Lastly, recent findings from human brain lesioning studies, have revealed that psychometric general intelligence, emotional intelligence, and social problem solving, are supported by a significantly shared neural network of temporal, frontal, and parietal areas of the brain, including white-matter association tracts that connect these regions into a coordinated system [24].
Finally, the recent discovery of so-called ‘mirror neurons’—neurons that fire both when an animal acts and when an animal observes the same action performed by another—has been argued to be important for mentalizing or theory of mind, language, empathy, understanding the actions and intentions of other agents, and learning new skills by imitation [25,26]. More specifically, observing an action and producing the same action oneself, appear to be neurally equivalent in adult monkeys, and at least in monkeys, this ability plays a role in imitation and social understanding [25,26]. Though mirror neurons have been directly observed in non-human primate species (most notably in the inferior parietal lobule and inferior frontal gyrus in macaques), in humans, brain activity consistent with that of mirror neurons has been found in the primary somatosensory cortex, superior and inferior parietal lobe, inferior frontal cortex, premotor cortex, and supplementary motor area [27]. In summary, the human brain shows a high degree of specialization for social stimuli and social information processing, encompassing regulation from the neural network level to the neurotransmitter level, including distinctly social neurotransmitters such as oxytocin and endorphins [4,19,28,29,30,31,32].
3. The social brain and cognitive neuroscience of language
In similar fashion, despite the clear anthropological and evolutionary connection between the social brain and social communication, described above, this framework has not yet been fully integrated into our current understanding of the cognitive neuroscience of human language [33,34]. Perhaps most critically, a complex neurological system of communication - for regulating interactions and social bonding with important members in the group - appears to be crucial for many non-human primates, including human social relationships [6]. In humans, a complex network of brain regions underlies the processing of language, including speech comprehension and production, and substantive integration with the social brain, including social-semantic working memory, and encompassing regulation from the neural network level to the neurotransmitter level, including social neurotransmitters such as oxytocin and dopamine [35,36,37,38,39].
In an early and influential model of the cognitive neuroscience of language, Pierre Paul Broca determined in 1861 that language functions are located primarily in the left cerebral hemisphere of the brain [40]. In later years, much research, including anatomical studies by Geschwind and Galaburda, further suggested left hemisphere dominance in brain regions dedicated to language [41,42], including larger pyramidal cells and myelinated axons in the left hemisphere, allowing for faster and more efficient processing of linguistic information [43,44]. Eventually, it became well-established that, at least in healthy adults, expressive language processing, also known as Broca’s area, and receptive language processing, also known as Wernicke’s area, is primarily associated with the left cerebral hemisphere, as well as white matter association tracts, including the left arcuate fasciculus (AF) that connect Broca’s and Wernicke’s areas into an integrated system [45,46]. Moreover, Broca’s area has been found to be related to both speech and the mirror neuron system, suggesting that there may in fact be substantial overlap between the neural networks for language, social cognition, and other related social brain networks see Figure 4; [34,47,48].
Nonetheless, more recent work has further shown that additional areas, including the putamen, caudate nucleus, and internal capsule appear to play additional roles in language processing [49], while very young children also show significant activation in inferior frontal and superior temporal areas of the right-hemisphere - homologues of traditional left-hemisphere language areas - with a right-hemisphere activation profile that appears to diminish with age [50]. Intriguingly, homologous brain regions of both Broca’s and Wernicke’s areas have also been identified in social, group-adapted, nonhuman primate brains, strongly suggesting a shared evolutionary or phylogenetic history [51,52]. Though their function in nonhuman primates is poorly understood, an evolutionary perspective would suggest that they are probably central to nonhuman primate vocalization processing, in ways similar to human language processing [53,54,55,56,57,58,59].
Recent studies on the social cognitive neuroscience of language have further demonstrated that neural activity during sentence processing in two canonical language areas, the left ventral temporoparietal junction (vTPJ) and lateral anterior temporal lobe (lATL), are associated with social-semantic working memory, in opposition to previous studies primarily implicating their role in general semantic or syntactic processing. In other words, these regions were sensitive to sentences only if the sentences conveyed social meaning. Moreover, these same regions appeared to maintain activity even after the linguistic stimuli were taken away [39]. In addition, several studies have shown that both chanting [60] and conversational speech [37] activate neural markers associated with social bonding: specifically, the temporoparietal junction (TPJ), associated with mentalizing, as well as the hypothalamus and amygdala, associated with social reward and motivation. Additional studies have revealed that self-disclosure - an important aspect of social bonding in humans - is intrinsically self-rewarding, due to activation of the mesolimbic dopamine system in the brain [38]. In summary, language and social cognition are two deeply interrelated capacities of the human species that have traditionally been studied as two separate domains. In effect, the human brain shows a high degree of specialization for sociolinguistic memory retrieval, information processing, and comprehension, suggesting that the function of these regions may link language with social cognition and social bonding [6].
4. The social brain and first language acquisition
Historically speaking, traditional theories of social learning have often emphasized the importance of social interaction for children’s acquisition of language [61,62]. Recent empirical studies further indicate that learning language often relies on children’s attunement to others’ communicative intentions, sensitivity to joint visual attention, and desire to imitate [61,63,64,65,66,67]. Developmental psychologists have even more recently extended these same theories and models to even earlier speech learning [68].
In particular, it has been argued that the earliest phases of language acquisition require social interaction; in other words, the social brain ‘gates’ the computational mechanisms involved in the acquisition of language [68,69,70,71]. Interestingly, randomized clinical trials appear to support this view, in finding consistent improvements in children’s expressive language skills with increases in parent–child interactions [72,73,74,75,76].
4.1. Social signals that facilitate early language acquisition
Social interaction skills, including play, reading, reference or joint attention of infant and parent to an outward object, and the face-to-face interactions involved in infant-directed speech in natural language environments, crucially aid the early acquisition of language see Figure 5; [36,68,72,77,78,79,80]. Infant-directed speech aids language acquisition by providing relevant social cues (e.g., gestures, facial and emotional expressions, and directed eye-gaze) that provoke infant attention and emphasize important pragmatic signals. Crucially, social interaction appears to impact the development of both speech perception or comprehension [81], as well as speech production learning [82,83,84]. For instance, in one study on speech perception, a foreign-language intervention was used to compare the language learning efficacy of a live social interaction, as opposed to televised or audio-only presentations. Accordingly, findings indicated an effect of social context, in that infants exposed to Mandarin via interaction with a live human being showed robust learning effects, while linguistic stimuli delivered via television or audio showed no evidence of language learning [81]. In an alternative study on speech production, infants exposed to mothers which responded immediately to their infants’ vocalizations by smiling, moving closer to, and touching their infants, produced more vocalizations than infants in a similarly-matched control group [84].
Several important developments accompany the capacity to understand reference, or joint attention of infant and parent to an outward thing or object [68]. By 9 months of age, infants start to engage in triadic person–object–person activities in which they devote attention to objects with gazes that provoke interest from another individual, also known as joint attention [65,66]. This shared perception of communicative intentions has been argued to be crucial for the infant’s acquisition of language [65,66,85], as well as understanding others as intentional agents [65,86]. Crucially, the developmental timing of these important social abilities generally coincides with the beginnings of attention to linguistic units, such as phonemes, as well as later word perception, comprehension and production (see Figure 5).
In addition, the quantity and quality of the speech input (e.g., vocabulary diversity, number of word units, and mean length of utterance (MLU), are further related to the growth of infant vocabulary [87,88,89]. Unfortunately, while most language acquisition research has been conducted on families of high socioeconomic status (SES), infants raised in poorer communities with multiple challenges can affect caregiver interactions, leading to greater variability in language abilities [90,91,92].
4.2. Infant-/child-directed speech and face-to-face communication
Infant-directed speech (IDS) or child-directed speech (CDS), also known as face-to-face communication between an infant or child and parent or guardian, is intrinsically multimodal and many nonverbal social signals are present during this sort of communication (e.g., gestures, facial and emotional expressions, and directed eye-gaze; see Figure 5). Several studies have demonstrated that eye gaze is a key form of nonverbal communication as it facilities language acquisition in a number of respects, including language processing, the development of vocabulary, and perceptual mapping of form-to-object [64,93,94]. For instance, gaze following and directed eye contact provoke arousal and attention by emphasizing important social stimuli and facilitating the infant or child’s social engagement [95,96]. Furthermore, the capacity and willingness to engage in this sort of sustained attention, has been shown to predict future cognitive development and general language skills [97,98,99]. For instance, infants who visually attend longer than those who engage in briefer states of attention demonstrate improved memory recall during object-naming tasks [100,101,102,103].
Gaze following as a tool for learning language is typically characterized by an early developmental trajectory where infants show a preference for open eyes on upright faces, involving the specialization of areas of the cortex associated with gaze processing [93,104,105,106,107]. Infants develop the capacity to follow gaze beginning from 3-4 months of age and gaze-following becomes a consistent social communication signal from 6-8 months of age [104,106]. However, it is not until 9–12 months of age, that gaze begins to become an important tool used for indicating reference, and facilitating language acquisition by providing directed eye gaze signaling [64,108,109,110,111]. Moreover, gaze is an important facilitator of social bonding between a mother and infant, with studies showing a positive association of maternal oxytocin to the duration of mother-to-infant gaze [112,113,114]. Further, directed attention to the mouth also plays an important role in language acquisition as it delivers important cues about mouth shape and associated interpretations of speech sounds [115]. Mouth attention becomes especially pronounced in infants 12-months-old when exposed to novel words [116], non-native sounds [117], or a bilingual environment [118]. Attention to the mouth further occurs in infants from 14-18 months of age, coinciding with the first burst of vocabulary [119].
The quality and quantity of speech during social interactions are also important factors in the acquisition of language, especially the growth of vocabulary [120,121,122,123]. For instance, studies have shown the amount of child-directed speech at 19 months predicts vocabulary growth at 24 months (Weisleder and Fernald, 2013). Parental engagement, namely, vocal responses to babbling with either vowels or words, rapidly influences the infant’s vocal productions, as infants begin to assimilate the phonological sound patterns produced by the parent, facilitating the acquisition of new vocal forms [124]. Additional instances of infant-/child-directed speech and ‘parentese’ (i.e., infant-/child-directed speech with higher or exaggerated prosody, an enlarged vowel space, and shorter utterances [125,126,127,128] enhances infant babbling from 6-14 months of age and facilities larger vocabularies at 14 months of age [72]. Lastly, dyadic infant/child to parent verbal and non-verbal social engagement, underpinned by various social neurotransmitters, including oxytocin, serotonin, and endorphins, appears to be critical for maternal attachment, and facilitating further dyadic social interactions, and social bonding between a mother and infant [28,129,130].
Child-parent social interactions are further affected by a number of environmental factors, such as socioeconomic status (SES). More specifically, SES affects both the quantity and quality of parental speech stimuli [120,131]; for instance, children of lower-SES families often display more sluggish real-time effectiveness of language processing and subsequent growth of vocabulary [91]. Low SES communities and families often display significant variation in the number of child-parent interactions, impacting processing efficiency of familiar words and predicting later expressive vocabulary scores [89]. Several factors account for the impact of low SES on social and cognitive development, including differences in healthcare, sanitation, psychological and physical stress, nutrition, and environmental pollution [132]. Differences in language acquisition, as a consequence of SES, can emerge as early as 9 months of age and predict later performance in school [133,134]. In summary, a number of developmental studies indicate that many different types of verbal and non-verbal social stimulation, including the quantity and quality of linguistic input, appear to benefit infants and children at different points in the developmental timetable and during various critical periods, including the primacy of gaze, dyadic infant/child-to-parent verbal and non-verbal social interactions, and other social cues for facilitating language acquisition and social bonding [68,123,135].
5. The social brain, cognitive neuroscience of language, and first language acquisition
Given the central importance of child–parent interactions for the acquisition of language, neuroimaging research has recently begun to investigate how just this sort of communication may impact the developing brain. As previously discussed, studies have found that, early in development, the quantity of language input, measured by the number of adult words, appears to be strongly predictive of myelin in white matter association tracts most associated with language - including the left arcuate fasciculus (AF) and superior longitudinal fasciculus (SLF) in younger children at 30 months of age - as well as children’s emerging language skills [136]. In contrast, the quality of language input - richness of words, utterance length, and conversational experience - appears to be more crucial for older children 4-6 years of age, who show greater white matter connectivity involving left AF and SLF [137,138] and increased cortical thickness in the left inferior frontal gyrus (IFG) and supramarginal gyri [139], as well as older children 5-9 years of age, who show increased cortical areas in the left perisylvian areas [140]. Additional social cognitive neuroscience studies have revealed that the neural circuits underlying the perception of mother’s voice - also known as ‘motherese’, as an important component of social bonding - include auditory and voice-selective structures in the temporal cortex, reward circuit areas in the nucleus accumbens (NAc), orbitofrontal cortex (OFC), and ventromedial prefrontal cortex (vmPFC), affective processing areas, including the amygdala, and regions associated with visual face processing, including the fusiform cortex - predict social communication and language function abilities in older children at 7-12 years of age [35]. Undoubtedly, these physiological mechanisms, also including social neurotransmitters such as oxytocin, dopamine, serotonin and endorphins, facilitate further dyadic interactions, maternal attachment, and social bonding between a mother and infant [28,129,130].
Until recently, most neuroimaging research on early social interactions has focused on neural activation in the infant or child brain in response to a unidirectional social stimulus. However, a newly utilized technique, known as ‘hyperscanning’, allows for the simultaneous data collection of neural activation from more than one individual at once, concurrently taking part in a social interaction [141]. More specifically, real-time temporal dynamics of social interactions can be associated with the temporal alignment of neural oscillations between a parent and child. For instance, several recent studies have shown that during a social interaction, neural synchronization occurs as a consequence of both verbal and non-verbal signals [142,143]. On the other hand, in a recent neuroimaging study involving a live two-way social interaction between adults and infants 9-15 months of age, which involved singing, playing, and reading, adult and infant brains differentially engaged with a range of social signals (e.g., speech prosody, eye gaze, and joint attention) as a function of their social importance. For example, activation of the prefrontal cortex in both the infant and adult brains, was significantly coupled to the time course of mutual gaze, suggesting that both individuals anticipated joint eye contact, while an increase in the infant’s prefrontal activation was consistently followed by pitch variability in the adult’s speech, likely the result of the adult producing extreme pitch contours in response to a range of infant behaviors, like highlighting a particular word [144]. Altogether, dual-brain imaging studies present new avenues for studying infant development in reciprocal social interactions and how infant’s learning strategies evolve over the course of natural communication. In summary, a number of social cognitive neuroscience studies indicate that many different types of verbal and non-verbal social stimulation, appear to benefit infants and children at different points in the developmental timetable and during various critical periods, including the quantity and quality of linguistic input, dyadic infant/child-to-parent verbal and non-verbal social interactions, and other social cues for facilitating language acquisition and social bonding [68,123,135].
6. The social brain and second language acquisition
A relatively recent number of studies have further explored the effects of bilingualism on mentalizing or perspective-taking, as well as empathy, on language processing in young children. For instance, at least one recent study found bilingual children were more accurate than monolingual children, in a task which required analyzing an observer’s perspective from different positions [145]. Moreover, a recent meta-analysis appears to indicate these general findings are robust [146]. Further studies have suggested that empathy appears to be associated with second language learning, including pronunciation accuracy [147,148,149]. Although difficult to predict the sociological impact of this work at such an early stage in scientific development, further studies on the effects of bilingual education in young children could have important implications for academic achievement in linguistic as well as non-linguistic social cognitive areas.
7. The social brain, developmental disorders, and psychopathologies
In recent decades, an increasing number of neuroscientists and psychologists are coming to understand that many psychopathologies and developmental disorders can be largely attributed to dysfunctions of the evolved social brain [150]. In the majority of cases, such dysfunctions typically involve significant deficits in social communication, social cognition, and linguistic abilities. In particular, autism spectrum disorder (ASD) is a heterogeneous disorder of social cognition, generally involving various social deficits, including dysfunctions in social communication (e.g., tone of voice and facial expressions), social interactions (e.g., eye gaze, joint attention, and gesture), imitation and social norms, mentalizing or theory of mind, empathy, metaphor (e.g., sarcasm and jokes), unfamiliar or new situations, planning for or predicting the future, and social imagination (e.g., make believe or play in children) [151]. Intriguingly, due to the strong genetic and neurodevelopmental etiology, and serious dysfunctions in social cognition by which ASD is defined, ASD offers the opportunity for neuroscientists, anthropologists, and psychologists to study the neurobiological origins of social cognition and social behaviors inherent to human nature. Additionally, numerous other psychopathologies and developmental disorders have been equally implicated as distinct disorders of social cognition and social behavior, including borderline personality disorder, social isolation and depression, narcissistic personality disorder, bipolar disorder, schizophrenia, psychosis, and dementia. In summary, such findings have led several scientists to argue that nearly all forms of psychopathology and cognitive developmental dysfunction should be specifically considered disorders of the social brain [150].
8. The social brain and autism spectrum disorder
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by chronic impairments in social interaction, verbal and non-verbal communication, and social cognition, including deficits in mentalizing or theory of mind [151,152]. In many ways, the complex interrelated social, genetic, and neurodevelopmental pathways and deficits found in ASD present one of the clearest and most compelling connections, perhaps, between the social brain, language function, social cognition, and social bonding [19]. As the name suggests, autism is situated on a spectrum, with some individuals whose verbal capacities exist within the normal range of functioning, while others never learn to speak [153]. Interestingly, in those with adequate language and cognitive capacities, such as those with high-functioning autism (HFA) and Asperger syndrome, specifically social communicative abilities ostensibly remain impaired. In other words, language is typically used instrumentally, one-sidedly, non-reciprocally, and non-socially, rather than for socially related functions [154]. Neurological studies on cortical development of the frontal and temporal language-related areas have been further associated with language impairments in ASD, including asymmetrical reversal of the frontal cortex [155,156,157], anterior and superior shifting of the left inferior frontal sulcus and superior temporal sulcus [158], bilateral decreases of gray matter volume in the superior temporal sulcus [159], and apparently overall reduced left hemispheric dominance. Intriguingly, though difficult to disentangle the relative contributions of deficits in social cognition in autism to linguistic deficits in autism, several recent studies in both autistic and neurotypical adults and children appear to suggest that mentalizing may be important for the syntactic ability to build subordinate and recursive imbedded clauses (e.g. ‘‘Mary thinks that Sandra believes the broom is in the closet’’) see Figure 6; [160,161,162], suggesting another direct link between social cognition and language ability.
As has been previously discussed so far, neurotypical infants and children must be attracted to and interested in infant/child-to-parent directed speech (IDS/CDS) in order to reliably acquire language. In general, neurotypical infants and children tend to prefer infant/child-to-parent directed speech (IDS/CDS) over adult-directed speech (ADS) when given the option [163,164], while neurotypical infants further tend to prefer speech over nonspeech signals [165]. Curiously, however, youngsters with ASD generally do not prefer IDS or CDS. For instance, in one recent study, neurotypical and ASD toddlers were permitted to choose between brief ‘motherese’ speech samples or nonspeech analogs of these same signals; yet, it was only toddlers with ASD that appeared to show a preference for the nonspeech signals [166]. Moreover, the severity of autism symptoms and degree of delay in verbal scores, predicted the degree of preference for nonspeech signals in children with ASD [166]. As has already been noted, a lack of interest or engagement in social interaction typically has a profound impact on the acquisition of language. Among the few rare documented cases in which children have been raised in complete social isolation, such circumstances have had a severe negative impact on the development of language, to where normal language abilities are not fully acquired [167]. In summary, the upshot of these studies, in neurotypical and neuroatypical adults and children, suggests that language acquisition crucially depends upon a social interest in people and the signals they produce.
As has been previously discussed so far, neurotypical infants and children must be attracted to and interested in infant/child-to-parent directed speech (IDS/CDS) in order to reliably acquire language. In general, neurotypical infants and children tend to prefer infant/child-to-parent directed speech (IDS/CDS) over adult-directed speech (ADS) when given the option [163,164], while neurotypical infants further tend to prefer speech over nonspeech signals [165]. Curiously, however, youngsters with ASD generally do not prefer IDS or CDS. For instance, in one recent study, neurotypical and ASD toddlers were permitted to choose between brief ‘motherese’ speech samples or nonspeech analogs of these same signals; yet, it was only toddlers with ASD that appeared to show a preference for the nonspeech signals [166]. Moreover, the severity of autism symptoms and degree of delay in verbal scores, predicted the degree of preference for nonspeech signals in children with ASD [166]. As has already been noted, a lack of interest or engagement in social interaction typically has a profound impact on the acquisition of language. Among the few rare documented cases in which children have been raised in complete social isolation, such circumstances have had a severe negative impact on the development of language, to where normal language abilities are not fully acquired [167]. In summary, the upshot of these studies, in neurotypical and neuroatypical adults and children, suggests that language acquisition crucially depends upon a social interest in people and the signals they produce.
9. Early biomarkers of language-related abilities and relevant clinical applications
Describing the early development of neurotypical and neuroatypical language neurobiology is critical for the early identification and potential treatment of clinical language disorders. Crucially, delays in language and speech in infants and children can negatively affect important social and academic skills such as attention, social interactions, reading, writing, and, of course, later educational outcomes [168]. For instance, delays in language acquisition from 2-5 years of age have been implicated in poor reading comprehension in the classroom [169,170]. If such language delays persist after 5 years of age, there are often related challenges in the consequent development of attention, directed eye gaze, and socialization [168,171]. The majority of language delays are often noticed during parental observations or clinical check-ups when an important developmental landmark does not appear to be present, like syntactic challenges or speech onset delays. As a consequence of this rather crude ‘sit-and-wait’ approach, most youngsters are unfortunately not characterized as having had a disorder or delay of language until 2-3 years of age, that is often noted by the absence of combinatorial speech, or the ability to formulate words into phrases and sentences [168,172]. Therefore, by the time a diagnosis has been made, language delays or disorders may be magnified due to the combined effects of accumulating negative experiences, resulting in atypical development within a substandard physical and social environment. Crucially, by 3 years of age, critical neurodevelopmental milestones that support language acquisition have essentially occurred, therefore missing any opportunities for early identification and clinical intervention. Although most children eventually do catch up with their peers before school age, approximately 7-10% of children enter the classroom with chronic impairments in language development [173].
An alternative approach emphasizes the development of early indications, or biomarkers, of probable language outcomes early enough to ensure that clinical interventions into speech and language delays and disorders might provide the greatest benefits. Perhaps surprisingly, there are currently no standardized or universally agreed upon recommendations regarding screening for speech and language impairments. Nonetheless, the most promising clinical interventions will most likely depend upon the very earliest identification of particular cognitive or behavioral traits, presumably underpinned by the hopeful discovery of critical neural or genetic biomarkers, that may allow for the early characterization and potential treatment of likely language outcomes before the emergence of language neurodevelopmental disorders. Biomarkers provide objective indicators of a clinical state that can be measured accurately and reliably [174], and contribute to the early recognition of atypical behavioral or neural patterns related to a later clinical disorder. For instance, a particular pattern of neural activation at 6 months of age could perhaps serve as an early predictive biomarker of socio-pragmatic difficulties at a later stage of development. More broadly, biomarkers could potentially exploit the broad heterogeneity observed in various language outcomes and serve as reliable measures predictive of later atypical patterns of development.
As might be expected, the diagnosis of language delays and language disorders are usually based on comparisons with developmental landmarks expected in neurotypical language development [175]. Children with language delays typically follow a normal developmental trajectory, but at a slower rate than would be expected [176], whereas children with language disorders tend to display regressions in language development (e.g., loss of words from 15-21 months of age in ASD), persistent and severe delays in language learning (e.g., syntactic challenges in children with specific language impairment (SLI), or impairments in at least two domains of development (e.g., motor function and language impairments in global developmental delay (GDD) [175,177,178]. As a general rule-of-thumb, language delays typically require clinical intervention when the development rate falls below 3/4 of the expected rate; for instance, when a developmental landmark expected at 24 months of age is not present in a child at 30 months of age [179]. In fortunate cases, comprehensive social and language evaluations are then performed to assess whether the delayed developmental pattern(s) may be associated with a primary language disorder, such as SLI, or a secondary developmental disorder, such as ASD or GDD.
Nonetheless, speech and language interventions should arguably begin even earlier in development. In fact, speech processing already begins in utero, in spite of the fact that the more observable first two years of life are characterized by the more obvious mappings of form-to-meaning at 6 months of age and native sound discrimination proficiency from 6-12 months of age [180]. In summary, even pediatricians and speech and language therapists with the best of intentions often miss critical early developmental opportunities in which the infant brain is most plastic and malleable in response to the environment and experience, and during which the implementation of targeted clinical interventions could potentially provide the greatest therapeutic benefits.
10. Discussion
10.1. Early biomarkers of probable language outcomes and clinical interventions
Though biomarker approaches to language delays and disorders research is still in its early stages of development, recent findings in language acquisition research have pinpointed particular behavioral, cognitive, and neural measures in early language learning that can be used to predict subsequent language outcomes. For instance, measures of real-time language processing efficiency, especially when used in conjunction with vocabulary growth measures, are a potentially promising tool for recognizing ‘late talkers’ more likely to catch up, versus those with continual delays. For example, at least one recent study in infants 18 months of age, found that both accuracy and speed measures in a familiar word recognition task, predicted later variation in vocabulary development (i.e., acceleration and rate of acquisition) in both ‘late talkers’ and typically developing children from 18-30 months of age [87]. Interestingly, this same technique further permitted the recognition of ‘late talkers’ more likely to experience accelerated vocabulary growth over the next 12 months. Moreover, the speed of linguistic processing efficiency, measured in children 18 months of age, further appeared to predict later development in children 54 months of age on particular language measures, namely, non-verbal intelligence, receptive vocabulary, and global language abilities [181]. In similar fashion, in infants at high-risk for SLI, discrimination of particular tone sequences at 7 months of age appears to be a significant predictor of language learning for language outcome measures from 12-36 months of age. At 36 months of age, this particular auditory task, in conjunction with sex, explains an even greater degree of performance on later language outcome measures [182].
Additional biomarkers further demonstrate the potential for the early diagnosis, prognosis, and implementation of crucial clinical interventions. In particular, neural biomarkers for early language impairments can be important for permitting clinical prognosis before relevant cognitive and behavioral symptoms emerge. For example, at least one recent study was able to use event-related potentials (ERPs: tiny voltages generated in the brain in response to specific events or stimuli) related to word processing in ASD children 2 years of age, to isolate early neural markers capable of predicting language outcomes at 4-6 years of age [183]. More specifically, the ERP signals related to familiar words was a significant predictor of adaptive behavior, cognitive ability, and receptive language measures. Perhaps surprisingly, the predictive accuracy of this measure even increased over time, further predicting scores at 6 years of age. Additionally, neural biomarkers related to discrimination of native sounds are further predictive of individual differences in later language abilities; in particular, ERP signals have been found to index neural speech discrimination in infants 7 months of age, accurately predicting later language growth rates [184,185]. Moreover, these same neural patterns at 11 months of age further predict individual differences in spoken syntactic abilities at 6 years of age, as well as the likelihood of acquiring a more serious language or speech disorder [186]. Ultimately, the accurate recognition and isolation of early neural biomarkers that predict language abilities will require a deeper understanding of the mechanisms underpinning the association between a given biomarker and various language outcomes. Future studies on the cognitive neuroscience of language and speech impairments should structure large sample longitudinal studies to ensure that previously identified biomarkers are consistent and reliable indicators of relevant language outcomes. In particular, studies conducted within the crucial first two years of age have the potential to isolate important neural biomarkers that may predict language delays, disorders, and abnormal neurodevelopmental trajectories. At best, this approach can potentially facilitate the implementation of relevant interventions at the earliest stages of development, when social and language skills have been less affected by experience, and the brain may be more malleable to crucial social and clinical interventions.
10.2. The social brain and cognitive neuroscience of language
Though many behavioral studies have demonstrated the importance of synchronous speech (i.e. chanting) [187], as well as conversational dialogue for facilitating social bonding [14,188], the methods used in behavioral studies are unable to provide detailed information about the brain areas and functional circuits underlying synchronous speech or conversational discourse. Moreover, though chanting has been observed in every human culture, less than 5% of speech involves joint speech, and there are good reasons to suspect that chanting may engage additional, or even different, cortical systems than conversational speech [6,189]. Indeed, this issue is crucially important, as it is not fully understood the mechanisms by which such vocalizations, in general, shape the developing brain [190]. In particular, this includes the manifestation of both pathological and healthy social development, such as autism and other disorders of social cognition, where the perception of and social bonding to socially salient voices may be impaired. In summary, the neural mechanisms associated with social bonding during conversational speech are not well understood.
Intriguingly, Rauchbauer et al. [37] has recently shown that conversational speech activates neural markers associated with social bonding: specifically, the temporo- parietal junction (TPJ), associated with mentalizing, as well as the hypothalamus and amygdala, associated with social reward and motivation. However, these results remain incomplete, as they did not control for the well-documented social bonding effects of:
1) conversational content, 2) eye-gaze, facial expressions, and body language, 3) joint attention, and 4) voice inflection, prosody, and other emotion-related aspects of speech [6,113,191]. Although a recent study by Jasmin et al. [60] avoided many of these pitfalls, these results also remain incomplete, as they primarily focused on chanting, not conversational speech. Moreover, neither Rauchbauer et al. [37] nor Jasmin et al. [60] included crucial measures of social bonding, thereby creating problems of reverse inference [192]. It thus remains unclear whether conversational speech, in general, actually facilitates social bonding, or whether the social bonding effects of speech can be explained by these aforementioned potentially confounding variables. In summary, it is not well understood whether the social neuroscience of conversational discourse involves the activation of specific neural regions of interest (ROIs) related to social reward, empathy and social bonding. Future studies on the social cognitive neuroscience of speech and social bonding should address these aforementioned concerns, as well as specifically compare neurotypical and neuroatypical participants, infants and children with adults, and male participants with female participants to determine the specific neurological profile in healthy neurotypical adults, and how that varies with specific to various neuroatypical social and linguistic deficits.
11. Conclusions
In conclusion, the human brain shows a high degree of specialization for social stimuli, social communication, language, and social information processing, encompassing regulation from the neural network level to the neurotransmitter level, including social neurotransmitters such as oxytocin, dopamine, serotonin and endorphins [4,28,29,30,31,32]. In particular, studies investigating the acquisition of language suggest that infants and children appear to benefit from many different aspects of social stimulation, depending on the particular point in the developmental timetable and during various critical periods, including the quantity and quality of linguistic input, dyadic infant/child-to-parent verbal and non-verbal social interactions, and other social cues for facilitating language acquisition and social bonding [68,123,135].
Detailed studies of comparative animal behavior, human social behavior, social cognitive neuroscience, and the cognitive neuroscience of human language in adults suggest a similar profile, in demonstrating that language and social cognition are two deeply interrelated capacities of the human species that have traditionally been studied as two separate domains [6]. In particular, the human brain shows a high degree of specialization, as well as functional overlap of neural areas dedicated to social and linguistic memory retrieval, information processing, and comprehension, suggesting that the evolutionary function of these regions may link language with social cognition and social bonding (see Figure 7). In effect, both the social brain hypothesis and social complexity hypothesis may help to explain how neurotypical children and adults learn language, why autistic children and adults exhibit simultaneous deficits in social cognition and language, and why nonhuman primates and other animals with significant computational abilities do not acquire the capacity for language [4,68].
This and related research, in conjunction with studies of early development, will allow scientists to generate a holistic profile, understanding, and potential treatment of maladies and deficits associated with the social brain.
Author Contributions
Conceptualization, N.O.; investigation, N.O.; writing—original draft preparation, N.O.; writing—review and editing, N.O.; supervision, D.S.; project administration, D.S.; funding acquisition, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thanks all anonymous reviewers for their support.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the manuscript; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Dunbar, R.I.M.; Shultz, S. Evolution in the social brain. Science 2007, 317, 1344–47. [Google Scholar] [CrossRef]
- Semendeferi, K.; Armstrong, E.; Schleicher, A.; Zilles, K.; Van Hoesen, G.W. Prefrontal cortex in humans and apes: A comparative study of area 10. Am. J. Phys. Anthropol. 2001, 114, 224–41. [Google Scholar] [CrossRef]
- Semendeferi, K.; Lu, A.; Schenker, N.; Damasio, H. Humans and great apes share a large frontal cortex. Nat. Neurosci. 2002, 5, 272–6. [Google Scholar] [CrossRef]
- Oesch, N. Social brain hypothesis. In H. Callan (Ed.), International encyclopedia of anthropology 2018, (pp. 1-11). New York: John Wiley and Sons. [CrossRef]
- Freeberg, T.M.; Dunbar RI, M.; Ord, T.J. Social complexity as a proximate and ultimate factor in communicative complexity. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1785–1801. [Google Scholar] [CrossRef] [PubMed]
- Oesch, N. Music and language in social interaction: Synchrony, antiphony and functional origins. Front. Psychol. 2019, 10. [Google Scholar] [CrossRef]
- McComb, K.; Semple, S. Coevolution of vocal communication and sociality in primates. Biol. Lett. 2005, 1, 381–85. [Google Scholar] [CrossRef] [PubMed]
- Gustison, M.L.; le Roux, A.; Bergman, T.J. Derived vocalizations of geladas (Theropithecus gelada) and the evolution of vocal complexity in primates. Philos. Trans. R. Soc. B 2012, 367, 1847–59. [Google Scholar] [CrossRef] [PubMed]
- King, A.J.; Myatt, J.P.; Fürtbauer, I.; Oesch, N.; Dunbar RI, M.; Sumner, S.; Usherwood, J.R.; Hailes, S.; Brown, M.R. Social density processes regulate the functioning and performance of foraging human teams. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Oesch, N.; Dunbar RI, M. Group size, communication, and familiarity effects in foraging human teams. Ethology 2018, 124, 483–495. [Google Scholar] [CrossRef]
- Dahmardeh, M.; Dunbar RI, M. What shall we talk about in Farsi?: Content of everyday conversations in Iran. Hum. Nat. 2017, 28, 423–33. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Marriott, A.; Duncan, N.D. Human conversational behavior. Hum. Nat. 1997, 8, 231–246. [Google Scholar] [CrossRef]
- Feinberg, M.; Willer, R.; Shultz, M. Gossip and ostracism promote cooperation in groups. Psychol. Sci. 2014, 25, 656–64. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.R.; Bosson, J.K. I feel like I know you: sharing negative attitudes of others promotes feelings of familiarity. Pers. Soc. Psychol. Bull. 2011, 37, 481–491. [Google Scholar] [CrossRef]
- Laidre, M.E.; Lamb, A.; Shultz, S.; Olsen, M. Making sense of information in noisy networks: Human communication, gossip, and distortion. J. Theor. Biol. 2013, 317, 152–160. [Google Scholar] [CrossRef]
- Adolphs, R. The social brain: Neural basis of social knowledge. Annu. Rev. Psychol. 2009, 60, 693–716. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.M. The social brain hypothesis and its implications for social evolution. Ann. Hum. Biol. 2009, 36, 562–72. [Google Scholar] [CrossRef] [PubMed]
- Brothers, L. The social brain: A project for integrating primate behavior and neurophysiology in a new domain. Concepts Neurosci. 1990, 1, 27–51. [Google Scholar]
- Johnson, M.H.; Griffin, R.; Csibra, G.; Halit, H.; Farroni, T.; de Haan, M.; Tucker, L.A.; Baron-Cohen, S.; Richards, J. The emergence of the social brain network: Evidence from typical and atypical development. Dev. Psychopathol. 2005, 17, 599–619. [Google Scholar] [CrossRef]
- Lewis, P.A.; Birch, A.; Hall, A.; Dunbar RI, M. Higher order intentionality tasks are cognitively more demanding. Soc. Cogn. Affect. Neurosci. 2017, 12, 1063–1071. [Google Scholar] [CrossRef]
- Kanai, R.; Bahrami, B.; Roylance, R.; Rees, G. Online social network size is reflected in human brain structure. Proc. R. Soc. B 2012, 279, 1327–1334. [Google Scholar] [CrossRef]
- Kwak, S.; Joo, W.; Youm, Y.; Chey, J. Social brain volume is associated with in-degree social network size among older adults. Proc. R. Soc. B 2018, 285, 20172708. [Google Scholar] [CrossRef]
- Von Der Heide, R.; Vyas, G.; Olson, I.R. The social network-network: Size is predicted by brain structure and function in the amygdala and paralimbic regions. Soc. Cogn. Affect. Neurosci. 2014, 9, 1962–1972. [Google Scholar] [CrossRef]
- Barbey, A.K.; Colom, R.; Paul, E.J.; Chau, A.; Solomon, J.; Grafman, J.H. Lesion mapping of social problem solving. Brain 2014, 137, 2823–2833. [Google Scholar] [CrossRef]
- Meltzoff, A.N.; Decety, J. What imitation tells us about social cognition: A rapprochement between developmental psychology and cognitive neuroscience. Philos. Trans. R. Soc. Lond. B, Biol. Sci. 2003, 358, 491–500. [CrossRef]
- Rizzolatti, G. The mirror neuron system and imitation. In S. Hurley and N. Chater (Eds.), Perspectives on imitation: From neuroscience to social science – I: Mechanisms of imitation and imitation in animals 2005, (pp. 55–76). Cambridge: MIT Press.
- Kilner, J.M.; Lemon, R.N. What we know currently about mirror neurons. Curr. Biol. 2013, 23, R1057–R1062. [Google Scholar] [CrossRef] [PubMed]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social reward requires coordinated activity of nucleus accumbens oxytocin and serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.M. The social brain hypothesis and human evolution. In Oxford Research Encyclopedias: Psychology 2016, (pp. 1-31). Oxford: Oxford University Press. [CrossRef]
- Krach, S.; Paulus, F.M.; Bodden, M.; Kircher, T. The rewarding nature of social interactions. Front. Behav. Neurosci. 2010, 4. [Google Scholar] [CrossRef]
- Machin, A.J.; Dunbar RI, M. The brain opioid theory of social attachment: A review of the evidence. Behaviour 2011, 148, 985–1025. [Google Scholar] [CrossRef]
- Watanabe, N.; Yamamoto, M. Neural mechanisms of social dominance. Front. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R. Cognitive neurosciences of human social behavior. Nat. Rev. Neurosci. 2003, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Pulvermuller, F. Brain mechanisms linking language to action. Nat. Rev. Neurosci. 2005, 6, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Abrams, D.A.; Chen, T.; Odriozola, P.; Cheng, K.M.; Baker, A.E.; Padmanabhan, A.; Ryali, S.; Kochalkaa, J.; Feinstein, C.; Menon, V. Neural circuits underlying mother’s voice perception predict social communication abilities in children. PNAS 2016, 113, 6295–6300. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Jeong, H. The social brain of language: Grounding second language learning in social interaction. Npj Science of Learning 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Rauchbauer, B.; Nazarian, B.; Bourhis, M.; Ochs, M.; Prévot, L.; Chaminade, T. Brain activity during reciprocal social interaction investigated using conversational robots as control condition. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374:20180033. [CrossRef]
- Tamir, D.I.; Mitchell, J.P. Disclosing information about the self is intrinsically rewarding. PNAS 2012, 109, 8038–8043. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, Y.; Wang, X.; Li, J.; Shi, W.; Bi, Y.; Lin, N. A social-semantic working-memory account for two canonical language areas. Nat. Hum. Behav. 2023, 7, 1980–1997. [Google Scholar] [CrossRef] [PubMed]
- Fancher, R.E. Pioneers of Psychology, 2nd ed. 1990, (pp. 72-93). New York: W. W. Norton and Co.
- Galaburda, A.M.; LeMay, M.; Kemper, T.L.; Geschwind, N. Right-left asymmetrics in the brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.J.; Belin, P. Spectral and temporal processing in human auditory cortex. Cereb. Cortex 2001, 11, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Galuske, R.A.; Schlote, W.; Bratzke, H.; Singer, W. Interhemispheric asymmetries of the modular structure in human in human temporal cortex. Science 2000, 289, 1946–1949. [Google Scholar] [CrossRef]
- Hutsler, J.J. The specialized structure of human language cortex: Pyramidal cell size asymmetries within auditory and language-associated regions of the temporal lobes. Brain Lang. 2003, 86, 226–242. [Google Scholar] [CrossRef]
- Pujol, J.; Deus, J.; Losilla, J.M.; Capdevila, A. Cerebral lateralization of language in normal left-handed people studied by functional MRI. Neurology 1999, 52, 1038–1043. [Google Scholar] [CrossRef]
- Springer, J.A.; Binder, J.R.; Hammeke, T.A.; Swanson, S.J.; Frost, J.A.; Bellgowan, P.S.; Mueller, W.M. Language dominance in neurologically normal and epilepsy subjects: A functional MRI study. Brain 1999, 122, 2033–2046. [Google Scholar] [CrossRef]
- Imada, T.; Zhang, Y.; Cheour, M.; Taulu, S.; Ahonen, A.; Kuhl, P.K. Infant speech perception activates Broca’s area: A developmental magnetoencephalography study. Neuroreport 2006, 17, 957–962. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef]
- Lieberman, P. Human language and our reptilian brain: The subcortical bases of speech, syntax, and thought. 2000. Cambridge: Harvard University Press.
- Oluladea, O.A.; Seydell-Greenwalda, A.; Chambersa, C.E.; Turkeltauba, P.E.; Dromericka, A.W.; Berlb, M.M.; Gaillard, W.D.; Newport, E.L. The neural basis of language development: Changes in lateralization over age. PNAS 2020, 117, 23477–23483. [Google Scholar] [CrossRef]
- Schenker, N.M.; Hopkins, W.D.; Spocter, M.A.; Garrison, A.R.; Stimpson, C.D.; Erwin, J.M.; Hof, P.R.; Sherwood, C.C. Broca’s area homologue in chimpanzees (Pan troglodytes): Probabilistic mapping, asymmetry, and comparison to humans. Cereb. Cortex 2010, 20, 730–742. [Google Scholar] [CrossRef]
- Striedter, G.F. Principles of brain evolution. 2005. Sunderland, MA: Sinauer Associates.
- Aboitiz, F. A brain for speech. Evolutionary continuity in primate and human auditory-vocal processing. Front. Neurosci. 2018, 12. [CrossRef]
- Ardesch, D.J.; Scholtens, L.H.; Longchuan, L.; Preuss, T.M.; Rilling, J.K.; van den Heuvel, M.P. Evolutionary expansion of connectivity between multimodal association areas in the human brain compared with chimpanzees. PNAS 2019, 116, 7101–7106. [Google Scholar] [CrossRef]
- Hage, S.R.; Nieder, A. Dual neural network model for the evolution of speech and language. Trends Neurosci. 2016, 39, 813–829. [Google Scholar] [CrossRef]
- Holstege, G.; Subramanian, H.H. Two different motor systems are needed to generate human speech. J. Comp. Neurol. 2016, 524, 1558–1577. [Google Scholar] [CrossRef]
- Kumar, V.; Croxson, P.L.; Simonyan, K. Structural organization of the laryngeal motor cortical network and its implication for evolution of speech production. J. Neurosci. 2016, 36, 4170–4181. [Google Scholar] [CrossRef] [PubMed]
- Petkov, C.I.; Jarvis, E.D. Birds, primates, and spoken language origins: Behavioral phenotypes and neurobiological substrates. Front. Evol. Neurosci. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Schoenemann, P.T. Conceptual complexity and the brain: Understanding language origins. In W. S.-Y. Wang and J. W. Minett (Eds.), Language acquisition, change and emergence: Essays in evolutionary linguistics 2005, (pp. 47–94). Hong Kong: City University of Hong Kong Press.
- Jasmin, K.M.; McGettigan, C.; Agnew, Z.K.; Lavan, N.; Josephs, O.; Cummins, F.; Scott, S.K. Cohesion and joint speech: Right hemisphere contributions to synchronized vocal production. J. Neurosci. 2016, 36, 4669–80. [Google Scholar] [CrossRef] [PubMed]
- Bruner, J. Child’s talk: Learning to use language. 1983. New York: W.W. Norton.
- Vygotsky, L.S. Thought and language. 1962. Cambridge, MA: MIT Press.
- Baldwin, D.A. Understanding the link between joint attention and language. In C. Moore and P.J. Dunham (Eds.), Joint attention: Its origins and role in development 1995, (pp. 131–158). Hillsdale, NJ: Lawrence Erlbaum Associates.
- Brooks, R.; Meltzoff, A.N. The development of gaze following and its relation to language. Dev. Sci. 2005, 8, 535–543. [Google Scholar] [CrossRef]
- Tomasello, M. Constructing a language. (2003a). Cambridge, MA: Harvard University Press.
- Tomasello, M. The key is social cognition. In D. Gentner and S. Kuczaj (Eds.), Language and thought (2003b), (pp. 47–58). Cambridge, MA: MIT Press.
- Tomasello, M.; Farrar, M.J. Joint attention and early language. Child Dev. 1986, 57, 1454–1463. [Google Scholar] [CrossRef]
- Kuhl, P.K. Is speech learning ‘gated’ by the social brain? Dev. Sci. 2007, 10, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Doupe, A.J.; Kuhl, P.K. Birdsong and human speech: Common themes and mechanisms. Annu. Rev. Neurosci. 1999, 22, 567–631. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.S.; Marler, P. Language and animal communication: Parallels and contrasts. In H.L. Roitblat and J.-A. Meyer (Eds.), Comparative approaches to cognitive science: Complex adaptive systems 1995, (pp. 341–382). Cambridge: MIT Press.
- Marler, P. The instinct to learn. In S. Carey and R. Gelman (Eds.), The epigenesis of mind: Essays on biology and cognition 1991, (pp. 37–66). Hillsdale, NJ: Lawrence Erlbaum Associates.
- Ferjan Ramírez, N.; Lytle, S.R.; Fish, M.; Kuhl, P.K. Parent coaching at 6 and 10 months improves language outcomes at 14 months: A randomized controlled trial. Dev. Sci. 2019, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Heidlage, J.K.; Cunningham, J.E.; Kaiser, A.P.; Trivette, C.M.; Barton, E.E.; Frey, J.R.; Roberts, M.Y. The effects of parent-implemented language interventions on child linguistic outcomes: A meta-analysis. Early Child. Res. Q. 2020, 50, 6–23. [Google Scholar] [CrossRef]
- Leung, C.Y.Y.; Hernandez, M.W.; Suskind, D.L. Enriching home language environment among families from low-SES backgrounds: A randomized controlled trial of a home visiting curriculum. Early Child. Res. Q. 2020, 50, 24–35. [Google Scholar] [CrossRef]
- McGillion, M.; Pine, J.M.; Herbert, J.S.; Matthews, D. A randomised controlled trial to test the effect of promoting caregiver contingent talk on language development in infants from diverse socioeconomic status backgrounds. J. Child Psychol. Psychiatry Allied Discip. 2017, 58, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; Thomas, C.; Boben, M. Providence talks: A citywide partnership to address early childhood language development. Stud. Educ. Eval. 2020, 64, 1–13. [Google Scholar] [CrossRef]
- Kuhl, P.K. Brain mechanisms in early language acquisition. Neuron 2010, 67, 713–727. [Google Scholar] [CrossRef]
- Kuhl, P.K. Early language learning and the social brain. Cold Spring Harb. Symp. Quant. Biol. 2014, 79, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Lytle, S.R.; Garcia-Sierra, A.; Kuhl, P. Two are better than one: Infant language learning from video improves in the presence of peers. PNAS 2018, 115, 9859–9866. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Esparza, N.; García-Sierra, A.; Kuhl, P. Look who’s talking NOW! Parentese speech, social context, and language development across time. Front. Psychol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K.; Tsao, F.-M.; Liu, H.-M. Foreign-language experience in infancy: Effects of short-term exposure and social interaction on phonetic learning. PNAS 2003, 100, 9096–9101. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K. Social elicitation of infant vocal behavior. J. Exp. Child Psychol. 1975, 20, 51–58. [Google Scholar] [CrossRef]
- Bloom, K.; Esposito, A. Social conditioning and its proper control procedures. J. Exp. Child Psychol. 1975, 19, 209–222. [Google Scholar] [CrossRef]
- Goldstein, M.; King, A.; West, M. Social interaction shapes babbling: Testing parallels between birdsong and speech. PNAS 2003, 100, 8030–8035. [Google Scholar] [CrossRef]
- Akhtar, N.; Tomasello, M. Intersubjectivity in early language learning and use. In S. Bråten (Ed.), Intersubjective communication and emotion in early ontogeny 1998, (pp. 316–335). Cambridge: Cambridge University Press.
- Meltzoff, A.N. Understanding the intentions of others: Re-enactment of intended acts by 18-month-old children. Dev. Psychol. 1995, 31, 838–850. [Google Scholar] [CrossRef]
- Fernald, A.E.; Marchman, V.A. Causes and consequences of variability in early language learning. In I. Arnon and E. Clark (Eds.), Experience, variation and generalization: Learning a first language 2011, (pp. 181-202). New York: John Benjamins. [CrossRef]
- Fernald, A.; Perfors, A.; Marchman, V.A. Picking up speed in understanding: Speech processing efficiency and vocabulary growth across the 2nd year. Dev. Psychol. 2006, 42, 98–116. [Google Scholar] [CrossRef]
- Weisleder, A.; Fernald, A. Talking to children matters: Early language experience strengthens processing and builds vocabulary. Psychol. Sci. 2013, 24, 2143–2152. [Google Scholar] [CrossRef]
- Fernald, L.C.H.; Weber, A.; Galasso, E.; Ratsifandrihamanana, L. Socioeconomic gradients and child development in a very low income population: Evidence from Madagascar. Dev. Sci. 2011, 14, 832–847. [Google Scholar] [CrossRef]
- Fernald, A.; Marchman, V.A.; Weisleder, A. SES differences in language processing skill and vocabulary are evident at 18 months. Dev. Sci. 2013, 16, 234–248. [Google Scholar] [CrossRef]
- Henrich, J.; Heine, S.J.; Norenzayan, A. The weirdest people in the world? Behav. Brain Sci. [CrossRef]
- Carpenter, M.; Nagell, K.; Tomasello, M.; Butterworth, G.; Moore, C. Social cognition, joint attention, and communicative competence from 9 to 15 months of age. Monogr. Soc. Res. Child Dev. 1998, 63, i–vi, 1. [Google Scholar] [CrossRef]
- Morales, M.; Mundy, P.; Delgado CE, F.; Yale, M.; Messinger, D.; Neal, R.; Schwartz, H.K. Responding to joint attention across the 6-through 24-month age period and early language acquisition. J. Appl. Dev. Psychol. 2000, 21, 283–298. [Google Scholar] [CrossRef]
- Çetinçelik, M.; Rowland, C.F.; Snijders, T.M. Do the eyes have it? A systematic review on the role of eye gaze in infant language development. Front. Psychol. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Smith, L.B. Hand-eye coordination predicts joint attention. Child Dev. 2017, 88, 2060–2078. [Google Scholar] [CrossRef] [PubMed]
- Kannass, K.N.; Oakes, L.M. The development of attention and its relations to language in infancy and toddlerhood. J. Cogn. Dev. 2008, 9, 222–246. [Google Scholar] [CrossRef]
- Lawson, K.R.; Ruff, H.A. Early focused attention predicts outcome for children born prematurely. J. Dev. Behav. Pediatr. 2004, 25, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Ruff, H.A.; Lawson, K.R. Development of sustained, focused attention in young children during free play. Dev. Psychol. 1990, 26, 85–93. [Google Scholar] [CrossRef]
- Macroy-Higgins, M.; Montemanaro, E. Attention and word learning in toddlers who are late talkers. J. Child Lang. 2016, 43, 1020–1037. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.F.; Smith, L.B.; Yu, C. A bottom-up view of toddler word learning. Psychon. Bull. Rev. 2014, 21, 178–185. [Google Scholar] [CrossRef]
- Salley, B.; Panneton, R.K.; Colombo, J. Separable attentional predictors of language outcome. Infancy 2013, 18, 462–489. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Suanda, S.H.; Smith, L.B. Infant sustained attention but not joint attention to objects at 9 months predicts vocabulary at 12 and 15 months. Dev. Sci. 2019; 22, e12735. [Google Scholar] [CrossRef]
- D’Entremont, B.; Hains SM, J.; Muir, D.W. A demonstration of gaze following in 3- to 6-month-olds. Infant Behav. Dev. 1997, 20, 569–572. [Google Scholar] [CrossRef]
- Farroni, T.; Csibra, G.; Simion, F.; Johnson, M.H. Eye contact detection in humans from birth. PNAS 2002, 99, 9602–9605. [Google Scholar] [CrossRef] [PubMed]
- Gredebäck, G.; Fikke, L.; Melinder, A. The development of joint visual attention: A longitudinal study of gaze following during interactions with mothers and strangers. Dev. Sci. 2010, 13, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Perra, O.; Gattis, M. Attention engagement in early infancy. Infant Behav. Dev. 2012, 35, 635–644. [Google Scholar] [CrossRef]
- Butler, S.C.; Caron, A.J.; Brooks, R. Infant understanding of the referential nature of looking. J. Cogn. Dev. 2000, 1, 359–377. [Google Scholar] [CrossRef]
- Caron, A.J.; Butler, S.; Brooks, R. Gaze following at 12 and 14 months: Do the eyes matter? Br. J. Dev. Psychol. 20. [CrossRef]
- Johnson, S.C.; Ok, S.-J.; Luo, Y. The attribution of attention: 9-month-olds? Interpretation of gaze as goal-directed action. Dev. Sci. 2007, 10, 530–537. [Google Scholar] [CrossRef]
- Woodward, A.L. Infants’ developing understanding of the link between looker and object. Dev. Sci. 2003, 6, 297–311. [Google Scholar] [CrossRef]
- Dickstein, S.; Thompson, R.A.; Estes, D.; Malkin, C.; Lamb, M.E. Social referencing and the security of attachment. Infant Behav. Dev. 1984, 7, 507–516. [Google Scholar] [CrossRef]
- Kim, S.; Fonagy, P.; Koos, O.; Dorsett, K.; Strathearn, L. Maternal oxytocin response predicts mother-to-infant gaze. Brain Res. 2014, 580, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ross, H.E.; Young, L.J. Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior. Front. Neuroendocrinol. 2009, 30, 534–547. [Google Scholar] [CrossRef]
- Tsang, T.; Atagi, N.; Johnson, S.P. Selective attention to the mouth is associated with expressive language skills in monolingual and bilingual infants. J. Exp. Child Psychol. 2018, 169, 93–109. [Google Scholar] [CrossRef]
- Tenenbaum, E.J.; Sobel, D.M.; Sheinkopf, S.J.; Malle, B.F.; Morgan, J.L. Attention to the mouth and gaze following in infancy predict language development. J. Child Lang. 2015, 42, 1173–1190. [Google Scholar] [CrossRef] [PubMed]
- Lewkowicz, D.J.; Hansen-Tift, A.M. Infants deploy selective attention to the mouth of a talking face when learning speech. PNAS 2012, 109, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Pons, F.; Bosch, L.; Lewkowicz, D.J. Bilingualism modulates infants’ selective attention to the mouth of a talking face. Psychol. Sci. 2015, 26, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Hillairet de Boisferon, A.; Tift, A.H.; Minar, N.J.; Lewkowicz, D.J. The redeployment of attention to the mouth of a talking face during the second year of life. J. Exp. Child Psychol. 2018, 172, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Hoff, E. How social contexts support and shape language development. Dev. Rev. 2006, 26, 55–88. [Google Scholar] [CrossRef]
- Hurtado, N.; Marchman, V.A.; Fernald, A. Does input influence uptake? Links between maternal talk, processing speed and vocabulary size in Spanish-learning children. Dev. Sci. 2008, 11, F31–F39. [Google Scholar] [CrossRef]
- Huttenlocher, J.; Waterfall, H.; Vasilyeva, M.; Vevea, J.; Hedges, L.V. Sources of variability in children’s language growth. Cogn. Psychol. 2010, 61, 343–365. [Google Scholar] [CrossRef]
- Rowe, M.L. A longitudinal investigation of the role of quantity and quality of child-directed speech in vocabulary development. Child Dev. 2012, 83, 1762–1774. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.H.; Schwade, J.A. Social feedback to infants’ babbling facilitates rapid phonological learning. Psychol. Sci. 2008, 19, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.P.; Aslin, R.N. Preference for infant-directed speech in the first month after birth. Child Dev. 1990, 61, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Fernald, A. Four-month-old infants prefer to listen to motherese. Infant Behav. Dev. 1985, 8, 181–195. [Google Scholar] [CrossRef]
- Fisher, C.; Tokura, H. Acoustic cues to grammatical structure in infant-directed speech: Cross-linguistic evidence. Child Dev. 1996, 67, 3192–3218. [Google Scholar] [CrossRef]
- Pegg, J.E.; Werker, J.F.; McLeod, P.J. Preference for infant-directed over adult-directed speech: Evidence from 7-week-old infants. Infant Behav. Dev. 1992; 15, 325–345. [Google Scholar] [CrossRef]
- Nagasawa, M.; Okabe, S.; Mogi, K.; Kikusui, T. Oxytocin and mutual communication in mother-infant bonding. Front. Hum. Neurosci. 2012, 6. [Google Scholar] [CrossRef]
- Rilling, J.K.; Young, L.J. The biology of mammalian parenting and its effect on offspring social development. Science 2014, 345, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Hoff, E. The specificity of environmental influence: Socioeconomic status affects early vocabulary development via maternal speech. Child Dev. 2003, 74, 1368–1378. [Google Scholar] [CrossRef]
- Bradley, R.H.; Corwyn, R.F. Socioeconomic status and child development. Annu. Rev. Psychol. 2002, 53, 371–399. [Google Scholar] [CrossRef]
- Halle, T.; Forry, N.; Hair, E.; Perper, K.; Wandner, L.; Wessel, J.; Vick, J. Disparities in early learning and development: Lessons from the early childhood longitudinal study - birth cohort (ECLS-B). 2009. Washington, DC: Child Trends, 1-39. [CrossRef]
- Walker, D.; Greenwood, C.; Hart, B.; Carta, J. Prediction of school outcomes based on early language production and socioeconomic factors. Child Dev. 1994, 65, 606–21. [Google Scholar] [CrossRef]
- Sanchez-Alonso, S.; Aslin, R.N. Towards a model of language neurobiology in early development. Brain Lang. 2022, 224, 105047. [Google Scholar] [CrossRef]
- Fibla, L.; Forbes, S.H.; McCarthy, J.; Mee, K.; Magnotta, V.; Deoni, S.; Cameron, D.; Spencer, J.P. Language exposure and brain myelination in early development. J. Neurosci. 2023, 43, 4279–4290. [Google Scholar] [CrossRef]
- Romeo, R.R.; Leonard, J.A.; Robinson, S.T.; West, M.R.; Mackey, A.P.; Rowe, M.L.; Gabrieli JD, E. Beyond the 30-Million-word gap: Children’s conversational exposure is associated with language-related brain function. Psychol. Sci. 2018, 29, 700–710. [Google Scholar] [CrossRef]
- Romeo, R.R.; Segaran, J.; Leonard, J.A.; Robinson, S.T.; West, M.R.; Mackey, A.P.; Yendiki, A.; Rowe, M.L.; Gabrieli, J.D. Language exposure relates to structural neural connectivity in childhood. J. Neurosci. 2018, 38, 7870–7877. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.R.; Leonard, J.; Grotzinger, H.; Robinson, S.T.; Takada, M.E.; Mackey, A.; Scherer, E.; Rowe, M.; West, M.R.; Gabrieli, J. Neuroplasticity associated with changes in conversational turn-taking following a family-based intervention. Dev. Cogn. Neurosci. 2021, 49, 100967. [Google Scholar] [CrossRef]
- Merz, E.C.; Maskus, E.A.; Melvin, S.A.; He, X.; Noble, K.G. Socioeconomic disparities in language input are associated with children’s language-related brain structure and reading skills. Child Dev. 2020, 91, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Montague, P.R.; Berns, G.S.; Cohen, J.D.; McClure, S.M.; Pagnoni, G.; Dhamala, M.; Fisher, R.E. Hyperscanning: Simultaneous fMRI during linked social interactions. NeuroImage 2002, 16, 1159–1164. [Google Scholar] [CrossRef]
- Dumas, G.; Lachat, F.; Martinerie, J.; Nadel, J.; George, N. From social behaviour to brain synchronization: Review and perspectives in hyperscanning. IRBM 2011, 32, 48–53. [Google Scholar] [CrossRef]
- Hasson, U.; Ghazanfar, A.A.; Galantucci, B.; Garrod, S.; Keysers, C. Brain-to-brain coupling: A mechanism for creating and sharing a social world. Trends Cogn. Sci. 2012, 16, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Piazza, E.A.; Hasenfratz, L.; Hasson, U.; Lew-Williams, C. Infant and adult brains are coupled to the dynamics of natural communication. Psychol. Sci. 2020, 31, 6–17. [Google Scholar] [CrossRef]
- Greenberg, A.; Bellana, B.; Bialystok, E. Perspective-taking ability in bilingual children: Extending advantages in executive control to spatial reasoning. Cogn. Dev. 2013, 28, 41–50. [Google Scholar] [CrossRef]
- Schroeder, S.R. Do bilinguals have an advantage in theory of mind? A meta-analysis. Front. Commun. 2018, 3. [Google Scholar] [CrossRef]
- Guiora, A.Z.; Brannon RC, L.; Dull, C.Y. Empathy and second language learning. Lang. Learn. 1972, 22, 111–130. [Google Scholar] [CrossRef]
- Kleinmann, H.H. Avoidance behaviour in adult second language learning. Lang. Learn. 1977, 27, 93–101. [Google Scholar] [CrossRef]
- Mishan, F. Designing authenticity into language learning materials. 2005. New York: Intellect Books.
- Brüne, M.; Ribbert, H.; Schiefenhövel, W. The social brain: Evolution and pathology. 2003. West Sussex, UK: John Wiley and Sons.
- Groen, W.B.; Zwiers, M.P.; van der Gaag, R.-J.; Buitelaar, J.K. The phenotype and neural correlates of language in autism: An integrative review. Neurosci. Biobehav. Rev. 2008, 32, 1416–1425. [Google Scholar] [CrossRef]
- Klin, A.; Pauls, D.; Schultz, R.; Volkmar, F. Three diagnostic approaches to Asperger syndrome: Implications for research. J. Autism Dev. Disord. 2005, 35, 221–234. [Google Scholar] [CrossRef]
- Lord, C.; Paul, R. Language and communication in autism. In D. Cohen and F. R. Volkmar (Eds.), Handbook of autism and pervasive developmental disorders. 1997. New York: John Wiley.
- Fine, J.; Bartolucci, G.; Szatmari, P.; Ginsberg, G. Cohesive discourse in pervasive developmental disorders. J. Autism Dev. Disord. 1994, 24, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Abell, F.; Krams, M.; Ashburner, J.; Passingham, R.; Friston, K.; Frackowiak, R.; Happe, F.; Frith, C.; Frith, U. The neuroanatomy of autism: A voxel-based whole brain analysis of structural scans. Neuroreport 1999, 10, 1647–1651. [Google Scholar] [CrossRef]
- DeFosse, L.; Hodge, S.M.; Makris, N.; Kennedy, D.N.; Caviness Jr, V.S.; McGrath, L.; Steele, S.; Ziegler, D.A.; Herbert, M.R.; Frazier, J.A.; Tager-Flusberg, H.; Harris, G.J. Language-association cortex asymmetry in autism and specific language impairment. Ann. Neurol. 2004, 56, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.R.; Harris, G.J.; Adrien, K.T.; Ziegler, D.A.; Makris, N.; Kennedy, D.N.; Lange, N.T.; Chabris, C.F.; Bakardjiev, A.; Hodgson, J.; Takeoka, M.; Tager-Flusberg, H.; Caviness Jr, V.S. Abnormal asymmetry in language association cortex in autism. Ann. Neurol. 2002, 52, 588–596. [Google Scholar] [CrossRef]
- Levitt, J.G.; Blanton, R.E.; Smalley, S.; Thompson, P.M.; Guthrie, D.; McCracken, J.T.; Sadoun, T.; Heinichen, L.; Toga, A.W. Cortical sulcal maps in autism. Cereb. Cortex 2003, 13, 728–735. [Google Scholar] [CrossRef]
- Boddaert, N.; Chabane, N.; Gervais, H.; Good, C.D.; Bourgeois, M.; Plumet, M.H.; Barthelemy, C.; Mouren, M.C.; Artiges, E.; Samson, Y.; Brunelle, F.; Frackowiak, R.S.; Zilbovicius, M. Superior temporal sulcus anatomical abnormalities in childhood autism: A voxel-based morphometry MRI study. Neuroimage 2004, 23, 364–369. [Google Scholar] [CrossRef]
- Colle, L.; Baron-Cohen, S.; Hill, J. Do children with autism have a theory of mind? A non-verbal test of autism vs. specific language impairment. J. Autism Dev. Disord. 2007, 37, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Oesch, N.; Dunbar RI, M. The emergence of recursion in human language: Mentalising predicts recursive syntax task performance. J. Neurolinguistics 2017, 43, 95–106. [Google Scholar] [CrossRef]
- Varley, R.; Siegal, M.; Want, S. Severe impairment in grammar does not preclude theory of mind. Neurocase 2001, 7, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Fernald, A. Four-month-old infants prefer to listen to motherese. Infant Behav. Dev. 1985, 8, 181–195. [Google Scholar] [CrossRef]
- Fernald, C.A.; Kuhl, P.K. Acoustic determinants of infant preference for motherese speech. Infant Behav. Dev. 1987, 10, 279–293. [Google Scholar] [CrossRef]
- Vouloumanos, A.; Werker, J.F. Listening to language at birth: Evidence for a bias for speech in neonates. Dev. Sci. 2007, 10, 159–164. [Google Scholar] [CrossRef]
- Kuhl, P.K.; Coffey-Corina, S.; Padden, D.; Dawson, G. Links between social and linguistic processing of speech in preschool children with autism: Behavioral and electrophysiological measures. Dev. Sci. 2005, 8, F1–F12. [Google Scholar] [CrossRef]
- Fromkin, V.; Krashen, S.; Curtiss, S.; Rigler, D.; Rigler, M. The development of language in Genie: A case of language acquisition beyond the "critical period. " Brain Lang. 1974, 1, 81–107. [Google Scholar] [CrossRef]
- McLaughlin, M.R. Speech and language delay in children. Am. Fam. Physician 2011, 83, 1183–1188. [Google Scholar]
- Catts, H.W.; Fey, M.E.; Tomblin, J.B.; Zhang, X. A longitudinal investigation of reading outcomes in children with language impairments. J. Speech Lang. Hear. Res. 2002, 45, 1142–1157. [Google Scholar] [CrossRef]
- Durand, V.N.; Loe, I.M.; Yeatman, J.D.; Feldman, H.M. Effects of early language, speech, and cognition on later reading: A mediation analysis. Front. Psychol. 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Snowling, M.J.; Bishop DV, M.; Stothard, S.E.; Chipchase, B.; Kaplan, C. Psychosocial outcomes at 15 years of children with a preschool history of speech-language impairment. J. Child Psychol. Psychiatry 2006, 47, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, S.; Simon, P. Speech and language disorders in children. In S. Rosenbaum and P. Simon (Eds.), Speech and language disorders in children: Implications for the social security administration’s supplemental security income program. 2016. New York: National Academies Press.
- Tomblin, J.B.; Records, N.L.; Buckwalter, P.; Zhang, X.; Smith, E.; O’Brien, M. Prevalence of specific language impairment in kindergarten children. J. Speech Lang. Hear. Res. 1997, 40, 1245–1260. [Google Scholar] [CrossRef] [PubMed]
- Strimbu, K.; Tavel, J.A. What are biomarkers? Curr. Opin. HIV AIDS 2010, 5, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Schum, R.L. Language screening in the pediatric office setting. Pediatr. Clin. N. Am. 2007, 54, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Rescorla, L. Age 17 language and reading outcomes in late-talking toddlers: Support for a dimensional perspective on language delay. J. Speech Lang. Hear. Res. 2009, 52, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Tager-Flusberg, H.; Paul, R.; Lord, C. Language and communication in autism spectrum disorders. In F. R. Volkmar, R. Paul, A. Klin, and J. D. Cohen (Eds.), Handbook of autism and pervasive developmental disorders: Diagnosis, development, neurobiology, and behavior 2005 (pp. 335–364). New York: John Wiley and Sons Inc.
- Zengin-Akkuş, P., Çelen-Yoldaş, T., Kurtipek, G., and Özmert, E. N. Speech delay in toddlers: Are they only ‘late talkers’?. Turk. J. Pediatr. (2018), 60(2), 165-172. 2018; 60, 165–172. [CrossRef]
- Feldman, H.M. How young children learn language and speech. Pediatr. Rev. 2019, 40, 398–411. [Google Scholar] [CrossRef]
- Moon, C.; Lagercrantz, H.; Kuhl, P.K. Language experienced in utero affects vowel perception after birth: A two-country study. Acta Paediatr. 2013, 102, 156–60. [Google Scholar] [CrossRef]
- Marchman, V.A.; Loi, E.C.; Adams, K.A.; Ashland, M.; Fernald, A.; Feldman, H.M. Speed of language comprehension at 18 Months old predicts school-relevant outcomes at 54 months old in children born preterm. J. Dev. Behav. Pediatr. 2018, 39, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Benasich, A.A.; Tallal, P. Infant discrimination of rapid auditory cues predicts later language impairment. Behav. Brain Res. 2002, 136, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K.; Coffey-Corina, S.; Padden, D.; Munson, J.; Estes, A.; Dawson, G. Brain responses to words in 2-year-olds with autism predict developmental outcomes at age 6. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K.; Rivera-Gaxiola, M. Neural substrates of language acquisition. Annu. Rev. Neurosci. 2008, 31, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K. Early language acquisition: Cracking the speech code. Nat. Rev. Neurosci. 2004, 5, 831–843. [Google Scholar] [CrossRef]
- Zhao, T.C.; Boorom, O.; Kuhl, P.K.; Gordon, R. Infants’ neural speech discrimination predicts individual differences in grammar ability at 6 years of age and their risk of developing speech-language disorders. Dev. Cogn. Neurosci. 2021, 48, 100949. [Google Scholar] [CrossRef]
- Zimmermann, J.; Richardson, D.C. Verbal synchrony and action dynamics in large groups. Front. Psychol. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Collins, N.L.; Miller, L.C. Self-disclosure and liking: A meta-analytic review. Psychol. Bull. 1994, 116, 457–475. [Google Scholar] [CrossRef]
- Cummins, F. Towards an enactive account of action: Speaking and joint speaking as exemplary domains. Adapt. Behav. 2013, 13, 178–186. [Google Scholar] [CrossRef]
- Scott, S. From speech and talkers to the social world: The neural processing of human spoken language. Science 2019, 366, 58–62. [Google Scholar] [CrossRef]
- Kreysa, H.; Kessler, L.; Schweinberger, S.R. Direct speaker gaze promotes trust in truth-ambiguous statements. PLoS ONE 2016, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Poldrack, R.A. Inferring mental states from neuroimaging data: From reverse inference to large-scale decoding. Neuron 2011, 72, 692–97. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mean social group size correlates with relative neocortex volume in anthropoid primates, indexed as the ratio of neocortex volume to the rest of the brain. Several distinct taxonomies exist in primates, including prosimians (triangles), monkeys (open circles), and apes (solid circles).
Figure 1.
Mean social group size correlates with relative neocortex volume in anthropoid primates, indexed as the ratio of neocortex volume to the rest of the brain. Several distinct taxonomies exist in primates, including prosimians (triangles), monkeys (open circles), and apes (solid circles).
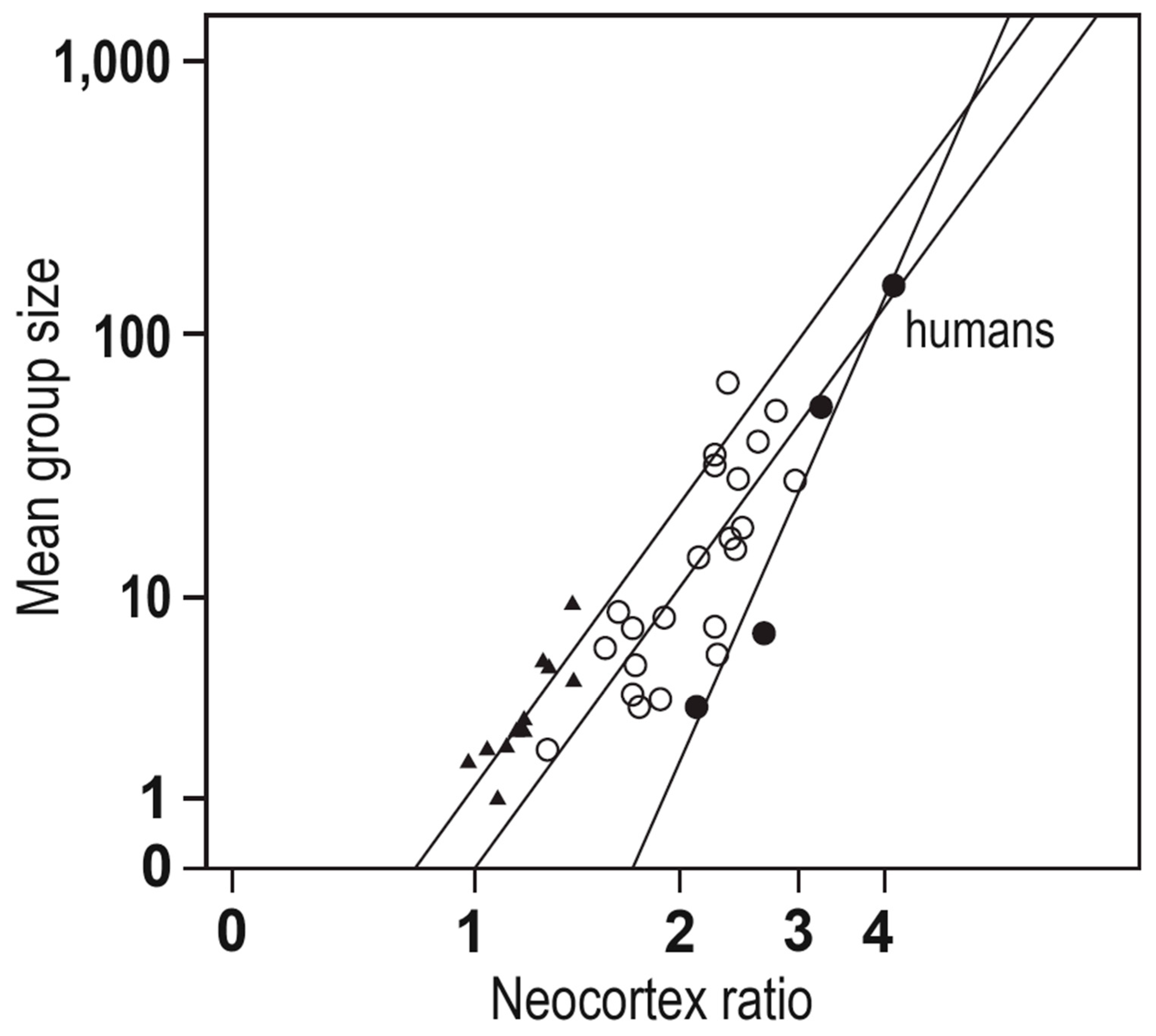
Figure 2.
Differences in character states, or contrasts, between sister taxa at each point in the primate phylogeny; namely, (a) contrasts in repertoire size and group size, and (b) contrasts in repertoire size and social grooming time, as a measure of social bonding.
Figure 2.
Differences in character states, or contrasts, between sister taxa at each point in the primate phylogeny; namely, (a) contrasts in repertoire size and group size, and (b) contrasts in repertoire size and social grooming time, as a measure of social bonding.
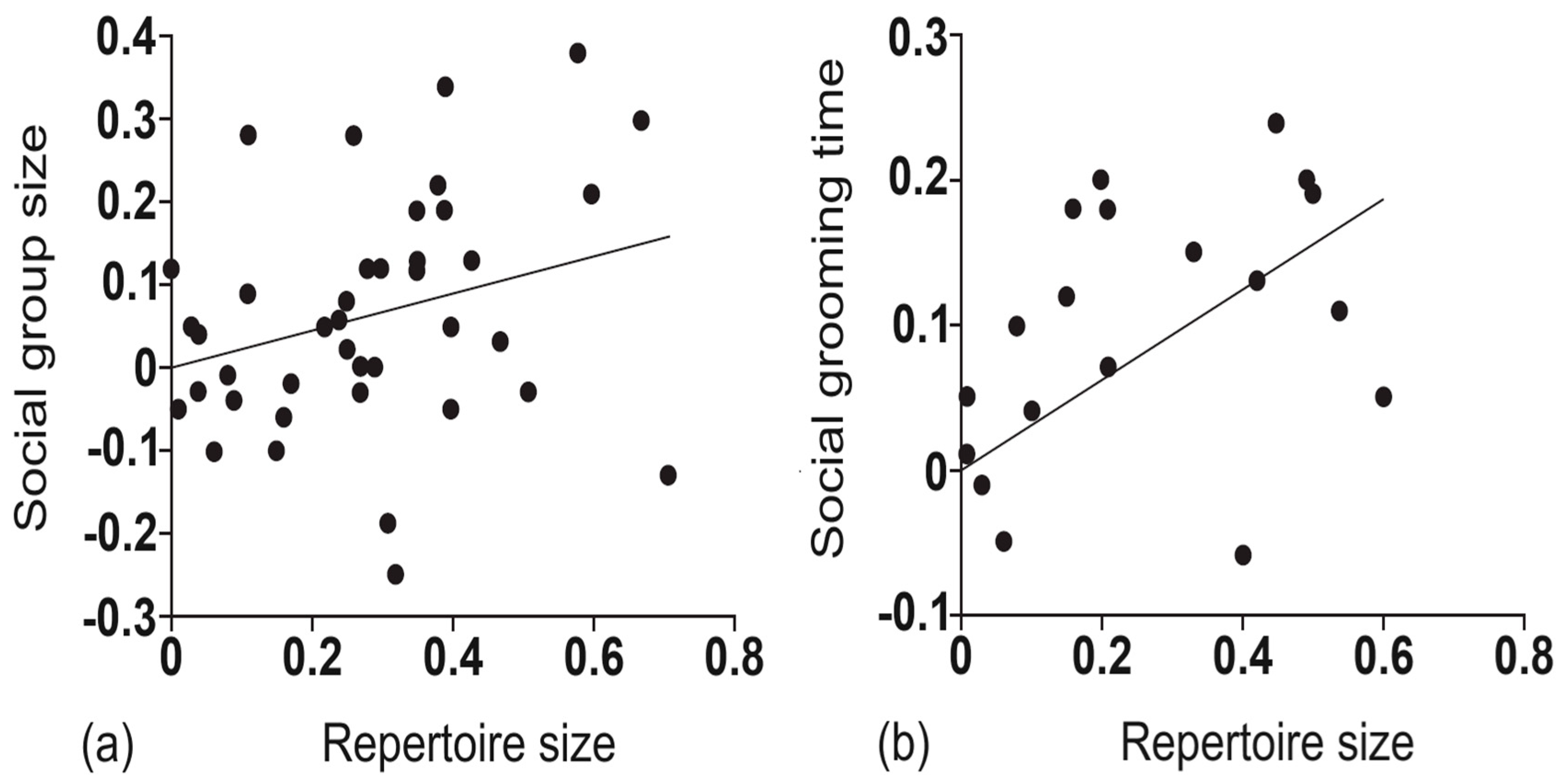
Figure 3.
Areas of the social brain include the amygdala (AMG), anterior cingulate cortex (ACC), prefrontal cortex (PFC), fusiform gyrus (FFG), and temporal and parietal lobes of the brain. In particular, this includes the medial prefrontal cortex (mPFC), ventromedial prefrontal cortex (vmPFC), and orbitofrontal cortex (OFC) of the PFC, motor cortex (MC) and premotor cortex (PreMC) of the parietal lobe, fusiform face area (FFA) and superior temporal gyrus (STG) of the temporal lobe, and action-perception network (APN) and temporal parietal junction (TPJ) of the temporal and parietal lobes. Note that, for simplicity and clarity, not all regions implicated in the network are shown; several subcortical and brainstem structures are not illustrated here.
Figure 3.
Areas of the social brain include the amygdala (AMG), anterior cingulate cortex (ACC), prefrontal cortex (PFC), fusiform gyrus (FFG), and temporal and parietal lobes of the brain. In particular, this includes the medial prefrontal cortex (mPFC), ventromedial prefrontal cortex (vmPFC), and orbitofrontal cortex (OFC) of the PFC, motor cortex (MC) and premotor cortex (PreMC) of the parietal lobe, fusiform face area (FFA) and superior temporal gyrus (STG) of the temporal lobe, and action-perception network (APN) and temporal parietal junction (TPJ) of the temporal and parietal lobes. Note that, for simplicity and clarity, not all regions implicated in the network are shown; several subcortical and brainstem structures are not illustrated here.
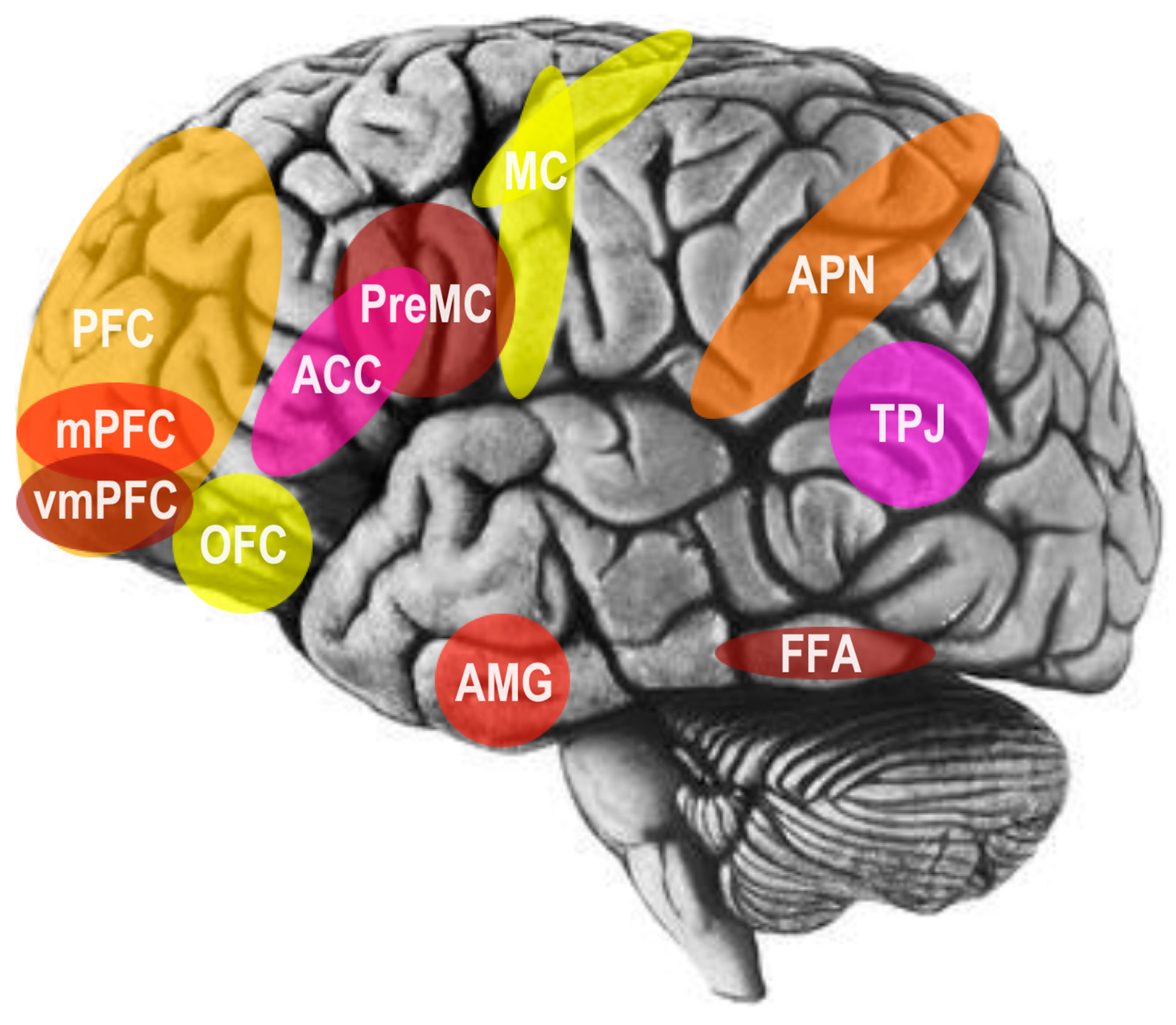
Figure 4.
Areas of the language brain, unlike the social brain, are primarily associated with the left cerebral hemisphere, and traditionally include Broca’s area (BA), related to expressive language processing, Wernicke’s area (WA), related to receptive language processing, and white matter association tracts, including the left arcuate fasciculus (AF) that connect Broca’s and Wernicke’s areas into an integrated system. Broca’s area has been found to be related to both speech and the mirror neuron system, suggesting substantive overlap between the neural networks for language, social cognition, and related social brain networks, including the inferior frontal gyrus (IFG), angular gyrus (AG), primary auditory cortex (PAC), and motor cortex (MC). Recent studies have also shown neural activity during sentence processing in two canonical social brain areas, namely, the temporoparietal junction (TPJ) and left temporal lobe, associated with social-semantic working memory, further suggesting substantive overlap and integration with the language brain and social brain. Note that, for simplicity and clarity, not all regions implicated in the network are shown; several subcortical and brainstem structures are not illustrated here.
Figure 4.
Areas of the language brain, unlike the social brain, are primarily associated with the left cerebral hemisphere, and traditionally include Broca’s area (BA), related to expressive language processing, Wernicke’s area (WA), related to receptive language processing, and white matter association tracts, including the left arcuate fasciculus (AF) that connect Broca’s and Wernicke’s areas into an integrated system. Broca’s area has been found to be related to both speech and the mirror neuron system, suggesting substantive overlap between the neural networks for language, social cognition, and related social brain networks, including the inferior frontal gyrus (IFG), angular gyrus (AG), primary auditory cortex (PAC), and motor cortex (MC). Recent studies have also shown neural activity during sentence processing in two canonical social brain areas, namely, the temporoparietal junction (TPJ) and left temporal lobe, associated with social-semantic working memory, further suggesting substantive overlap and integration with the language brain and social brain. Note that, for simplicity and clarity, not all regions implicated in the network are shown; several subcortical and brainstem structures are not illustrated here.
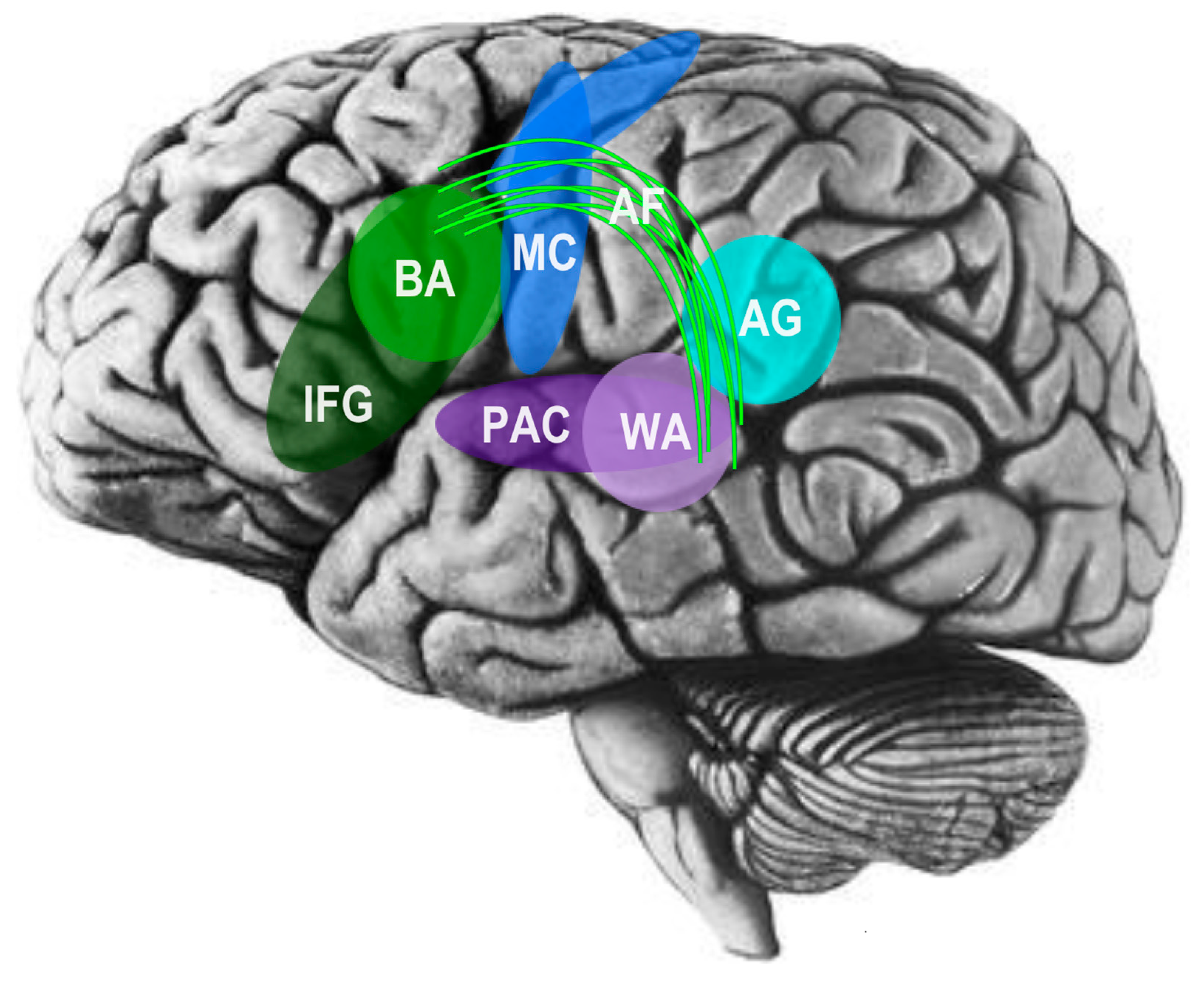
Figure 5.
Social interaction skills, including play, reading, or reference, and the face-to-face interactions involved in infant-directed speech (IDS) in natural language environments, crucially aid the early acquisition of language. IDS aids language acquisition by providing relevant social signals (e.g. gestures, facial and emotional expressions, and directed eye-gaze) that provoke infant attention and emphasize important pragmatic signals. Social interaction appears to impact the development of speech perception or comprehension as well as speech production. The capacity to understand reference, also known as joint attention - of infant and parent to a thing or object - further appears to be integral for the infant’s acquisition of language, as well as understanding others as intentional agents.
Figure 5.
Social interaction skills, including play, reading, or reference, and the face-to-face interactions involved in infant-directed speech (IDS) in natural language environments, crucially aid the early acquisition of language. IDS aids language acquisition by providing relevant social signals (e.g. gestures, facial and emotional expressions, and directed eye-gaze) that provoke infant attention and emphasize important pragmatic signals. Social interaction appears to impact the development of speech perception or comprehension as well as speech production. The capacity to understand reference, also known as joint attention - of infant and parent to a thing or object - further appears to be integral for the infant’s acquisition of language, as well as understanding others as intentional agents.

Figure 6.
Several recent studies in both autistic and neurotypical adults and children appear to suggest that mentalizing may be important for the syntactic ability to build subordinate and recursive embedded clauses (e.g. “Mary thinks that Sandra believes the broom is in the closet”), suggesting a direct link between social cognition and language ability.
Figure 6.
Several recent studies in both autistic and neurotypical adults and children appear to suggest that mentalizing may be important for the syntactic ability to build subordinate and recursive embedded clauses (e.g. “Mary thinks that Sandra believes the broom is in the closet”), suggesting a direct link between social cognition and language ability.

Figure 7.
The human brain shows a high degree of specialization, as well as functional overlap .of neural areas dedicated to social and linguistic memory retrieval, information processing, and comprehension, suggesting the evolutionary function of these regions may link language with social cognition and social bonding. In particular, Broca’s area (BA) has been found to be related to both speech and the mirror neuron system, suggesting substantial overlap between the neural networks for language, social cognition, and other related social brain networks, including the inferior frontal gyrus (IFG), angular gyrus (AG), motor cortex (MC). Moreover, the left ventral temporoparietal junction (vTPJ) and lateral anterior temporal lobe (lATL), are associated with social-semantic working memory, in opposition to previous studies primarily implicating their role in general semantic or syntactic processing. Here, traditional areas of the social brain are highlighted in warm colors, while traditional areas of the language brain are highlighted in cool colors, further illustrating substantive loverlap and integration of relevant language and social brain areas. Note that, for simplicity and clarity, not all regions implicated in the networks are shown; several subcortical and brainstem structures are not illustrated here.
Figure 7.
The human brain shows a high degree of specialization, as well as functional overlap .of neural areas dedicated to social and linguistic memory retrieval, information processing, and comprehension, suggesting the evolutionary function of these regions may link language with social cognition and social bonding. In particular, Broca’s area (BA) has been found to be related to both speech and the mirror neuron system, suggesting substantial overlap between the neural networks for language, social cognition, and other related social brain networks, including the inferior frontal gyrus (IFG), angular gyrus (AG), motor cortex (MC). Moreover, the left ventral temporoparietal junction (vTPJ) and lateral anterior temporal lobe (lATL), are associated with social-semantic working memory, in opposition to previous studies primarily implicating their role in general semantic or syntactic processing. Here, traditional areas of the social brain are highlighted in warm colors, while traditional areas of the language brain are highlighted in cool colors, further illustrating substantive loverlap and integration of relevant language and social brain areas. Note that, for simplicity and clarity, not all regions implicated in the networks are shown; several subcortical and brainstem structures are not illustrated here.
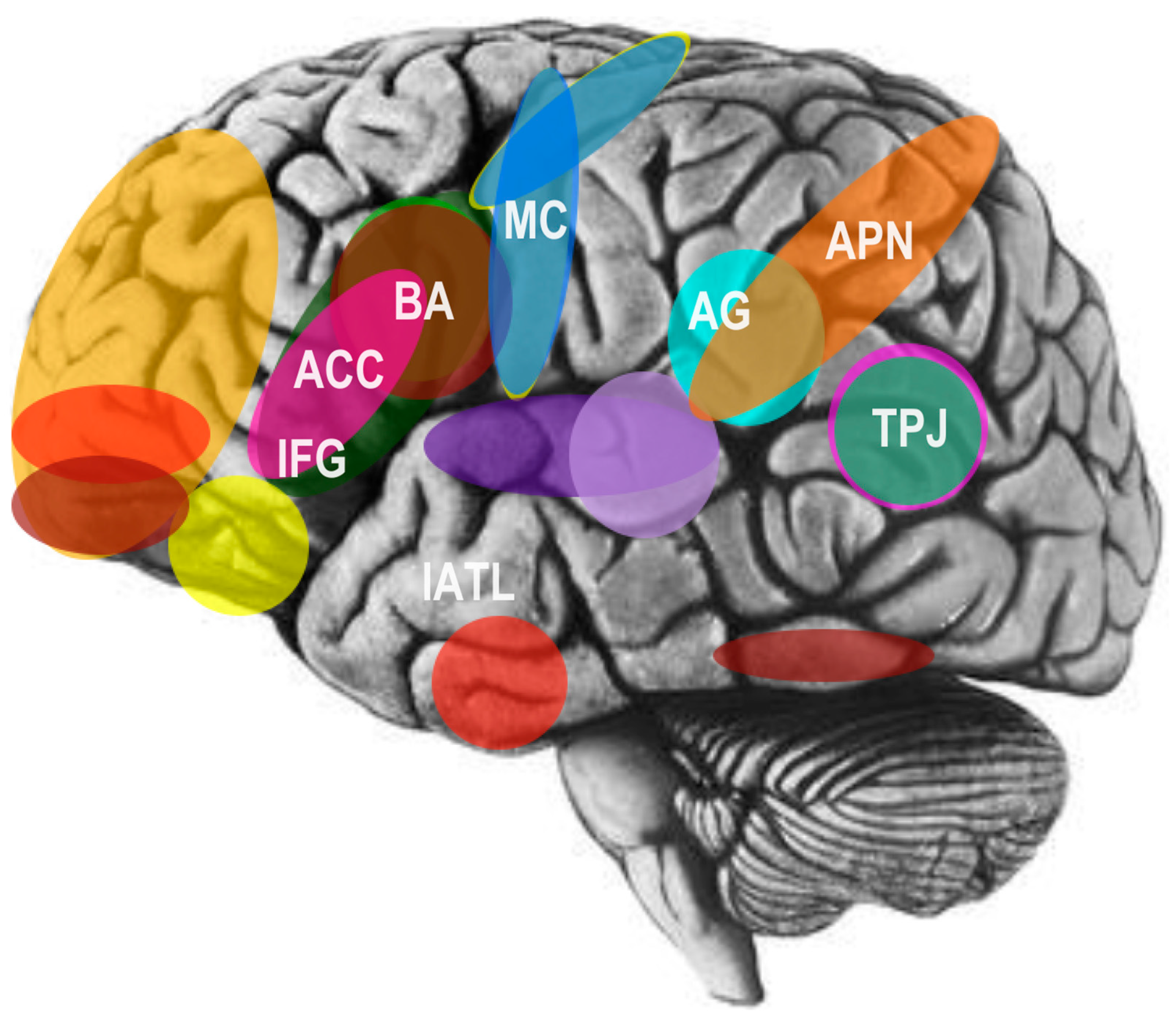
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



