
Submitted:
22 January 2024
Posted:
23 January 2024
You are already at the latest version
Abstract
Skin wounds and their infections by antibiotic resistant bacteria (ARB)are very common in small animals, posing the risk of acquiring ARB by pet owners or antibiotic resistance gene (ARG) transfer to the owners’ microbiota. The aim of this study was to identify the most common pathogens infecting wounds of companion animals, assess their antibiotic resistance and determine the ARGs using culture-based, molecular and proteomic methods. A total of 136 bacterial strains were isolated from wound swabs. Their species was identified using chromogenic media, followed by MALDI-TOF spectrometry. Disk diffusion was used to test the antibiotic resistance and twelve ARGs were detected using PCRs. The dominant species included Staphylococcus pseudintermedius (9.56%), E. coli and E. faecalis (both n=11, 8.09%). Enterobacterales were mostly resistant to amoxicillin/clavulanic acid (68.3% strains), all Pseudomonas were resistant to ceftazidime, piperacillin/tazobactam, imipenem and tylosin, Acinetobacter was mostly resistant to tylosin (55.5%), all Enterococcus were resistant to imipenem, 39.2% of Staphylococci were resistant to clindamycin. Among ARGs, strA (streptomycin resistance), sul3 (sulfonamide resistance) and blaTEM, an ESBL determinant, were the most frequent. The risk of ARB and ARD transfer between animals and humans causes the need to search for new antimicrobial therapies in the future veterinary medicine.
Keywords:
antibiotic resistant bacteria
; antibiotic resistance genes
; companion animals
; veterinary medicine
; wound infections
1. Introduction
In recent years, the number of pets in the European countries has gradually increased with dogs and cats being the two most prevalent types of companion animals in households [1]. The social role of companion animals has changed and attention to their health and welfare has significantly increased, too [2]. Skin wounds are very common in small animals. Such wounds are frequently colonized by bacteria or show signs of bacterial infection. Both, colonization and infection often result in healing delay and difficulties, thus increasing cost of treatment [3]. Currently, the treatment of wounds in veterinary medicine is based on the administration of broad –spectrum antibiotics, the continuous administration of which contributes to the increasing antimicrobial resistance among both pathogenic bacteria and even commensal skin microbiota [4].
In this context, the threat of wound infection, especially by antimicrobial resistant bacteria (ARB), becomes even more serious problem. This is due to the risk of acquiring the ARB or the relative antibiotic resistance genes (ARGs) by the pet owners via contact with their companion animals [2]. The risk of interspecies transmission of bacteria, including antibiotic resistant or multidrug resistant bacteria is very high, because the majority of bacterial pathogens of companion animals are the bacterial species that can commonly occur in humans, too. These pathogens and opportunistic pathogens are: Staphylococcus (including S. aureus), Pseudomonas (including P. aeruginosa), beta-haemolytic Streptococcus, E. coli or Enterococcus (including E. faecalis or E. faecium) [1,2,3,4,5,6].
With respect to the high risk of interspecies transmission of pathogenic ARB, the risk of infection treatment failure is high not only among companion animals, but also among their owners. There is surprisingly small number of studies concerning the scope of AMR in companion animals, while research of animal wound infections is limited to a only a few texts [3,4]. For this reason, understanding the prevalence of antimicrobial resistant bacterial infections in wounds of companion animals, that can easily be transmitted to their owners is among the most important gaps to be filled. With this in mind, the aim of this study was to identify the most common pathogens infecting wounds of companion animals treated in veterinary clinics, to assess their resistance to antibacterial agents used in their treatment in both veterinary and human medicine and to determine the genes determining the resistance mechanisms. The combination of culture-based, molecular biology and proteomic methods was used in order to obtain the most reliable results.
2. Results
A total of 136 bacterial isolates were obtained from the collected swabs, including: 52 (38.24%) from cats, 80 (58.82%) from dogs and 4 (2.94%) from rabbits. There were 71 (52.21%) Gram-positive and 65 (47.79%) Gram-negative isolates. The dominant genera were Staphylococcus spp. (n=37, 27.21%), followed by Enterococcus spp. (n=17, 12.50%), Escherichia (n=11, 8.09%) and Acinetobacter (n=10, 7.35%), with the following dominant species: S. pseudintermedius (n=13, 9.56%), E. coli and E. faecalis (both n=11, 8.09%). The dominant bacterial species differed between the examined groups of animals (Table 1).
Antimicrobial resistance of 120 bacterial isolates was tested using 16 antimicrobial agents in five combinations, based on the target organism (Table 1). Among the group of Enterobacterales, the resistance to amoxicillin/clavulanic acid was the most prevalent (detected in 68.3% of strains), followed by ampicillin (52.6%) and tylosin (44.7%). The ESBL mechanism of resistance was detected in 13.16% of strains (Figure 4A, Figure 5A; Table 4). None of the examined antimicrobials were effective against all strains. All strains of Pseudomonas spp. (n=7) were resistant to ceftazidime, piperacillin/tazobactam, imipenem and tylosin. Out of the tested antibiotics, only amikacin proved effective against all tested strains (Figure 4B). Finally, Acinetobacter (n=9) showed the highest percentage of resistance to tylosin (55.5% of strains), followed by enrofloxacin and gentamicin (both 44.4%, Figure 5C). In the case of Gram-positive bacteria, all Enterococcus spp. strains (n=15) were resistant to imipenem, 66.7% were resistant to tigecycline and 46.7% to enrofloxacin (Figure 6A). In Staphylococcus spp., the resistance to clindamycin was the most frequently observed (i.e. in 39.2% of strains). Methicillin resistance, shown by the resistance to cefoxitin was the second most prevalent, and was observed in 33.3% of strains (Table 4, Figure 4B), along with the resistance to erythromycin (also detected in 33.3% of strains). Both MSb and MLSb constitutive type of resistance mechanism was observed in 13.7%, while inducible MLSb mechanism was observed in 3.9% of strains (Figure 4C and D; Figure 6B, Table 4).
Figure 1.
The resistance mechanisms observed in the examined groups of bacteria. A – Extended Spectrum Beta Lactamases (ESBL) in Enterobacterales; B – methicillin resistance in Staphylococcus pasteuri; C – inducible resistance to macrolides, lincosamids and streptogramins b (MLSb) in Staphylococcus pseudintermedius; D – constitutive MLSb in S. pseudintermedius.
Figure 1.
The resistance mechanisms observed in the examined groups of bacteria. A – Extended Spectrum Beta Lactamases (ESBL) in Enterobacterales; B – methicillin resistance in Staphylococcus pasteuri; C – inducible resistance to macrolides, lincosamids and streptogramins b (MLSb) in Staphylococcus pseudintermedius; D – constitutive MLSb in S. pseudintermedius.
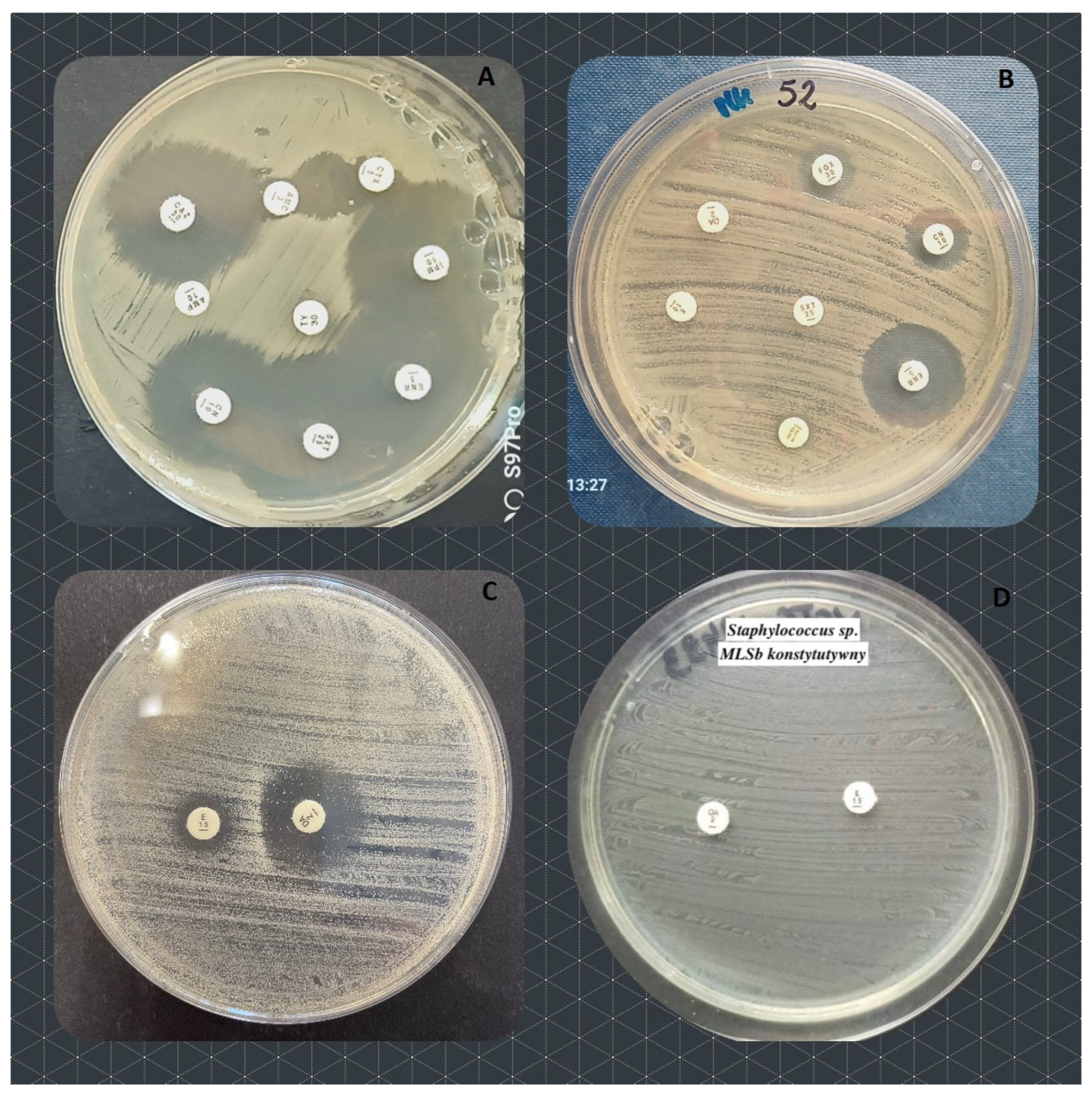
One Enterobacterales strain (canine Proteus mirabilis), out of all examined, was resistant (or insusceptible) to eight out of nine antimicrobial agents tested. Two other Gram-negative strains (K. pneumoniae and Aeromonas media) were resistant to 7 antimicrobials (Table 4). Among Gram-positives, three Staphylococci were resistant to seven out of eight antimicrobials tested (Table 4; feline S. pseudintermedius and S. aureus, and canine S. pseudintermedius).
Figure 2.
Share (%) of resistant (R) and susceptible (S) Gram-negative bacterial strains (A – Enterobacterales; B – Pseudomonas; C – Acinetobacter) isolated from wounds of companion animals. CN – gentamycin; SXT – trimethoprim/sulfamethoxazole; CTX – cefotaxime; AK – amikacin; AMC – amoxicillin/clavulanic acid; CAZ – ceftazidime; ENR – enrofloxacin; AMP – ampicillin; IMP – imipenem; TY – tylosin; TZP – piperacillin/tazobactam; ESBL – extended-spectrum-beta-lactamase-producing strains of Enterobacterales.
Figure 2.
Share (%) of resistant (R) and susceptible (S) Gram-negative bacterial strains (A – Enterobacterales; B – Pseudomonas; C – Acinetobacter) isolated from wounds of companion animals. CN – gentamycin; SXT – trimethoprim/sulfamethoxazole; CTX – cefotaxime; AK – amikacin; AMC – amoxicillin/clavulanic acid; CAZ – ceftazidime; ENR – enrofloxacin; AMP – ampicillin; IMP – imipenem; TY – tylosin; TZP – piperacillin/tazobactam; ESBL – extended-spectrum-beta-lactamase-producing strains of Enterobacterales.
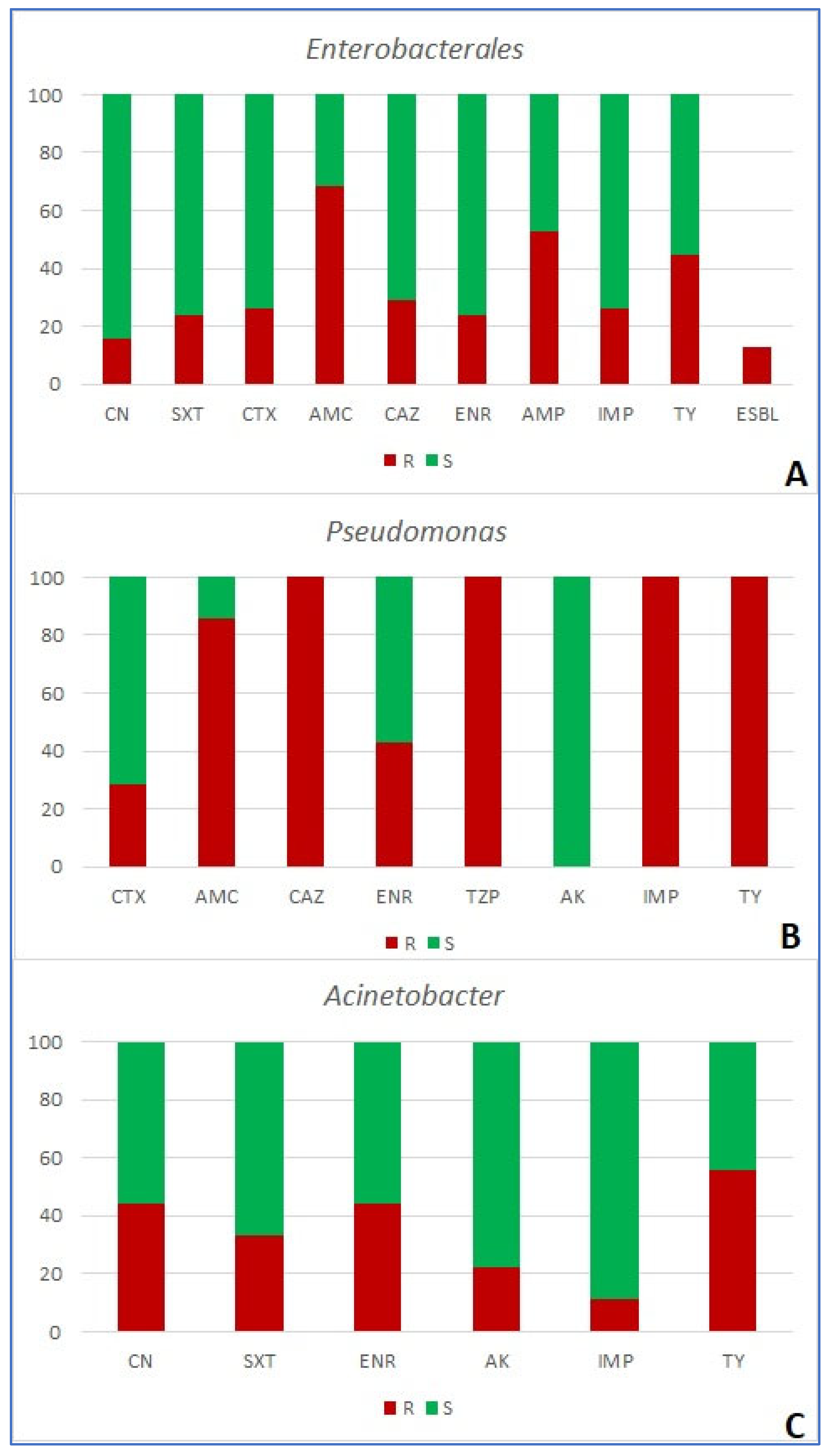
Figure 3.
Share (%) of resistant (R) and susceptible (S) Gram-positive bacterial strains (A –Enterococcus; B – Staphylococcus) isolated from wounds of companion animals. AMP – ampicillin; CN – gentamycin; DA – clindamycin; E – erythromycin; IMP – imipenem; ENR – enrofloxacin; FOX – cefoxitin; SXT – trimethoprim/sulfamethoxazole; TGC – tigecycline; TE – tetracycline; TY – tylosin; MSb – resistance mechanisms to macrolides and streptogramins b; MLSb ind – inducible mechanisms of resistance to macrolides, lincosamids and streptogramins b; MLSb const – constitutive mechanisms of resistance to macrolides, lincosamids and streptogramins b.
Figure 3.
Share (%) of resistant (R) and susceptible (S) Gram-positive bacterial strains (A –Enterococcus; B – Staphylococcus) isolated from wounds of companion animals. AMP – ampicillin; CN – gentamycin; DA – clindamycin; E – erythromycin; IMP – imipenem; ENR – enrofloxacin; FOX – cefoxitin; SXT – trimethoprim/sulfamethoxazole; TGC – tigecycline; TE – tetracycline; TY – tylosin; MSb – resistance mechanisms to macrolides and streptogramins b; MLSb ind – inducible mechanisms of resistance to macrolides, lincosamids and streptogramins b; MLSb const – constitutive mechanisms of resistance to macrolides, lincosamids and streptogramins b.
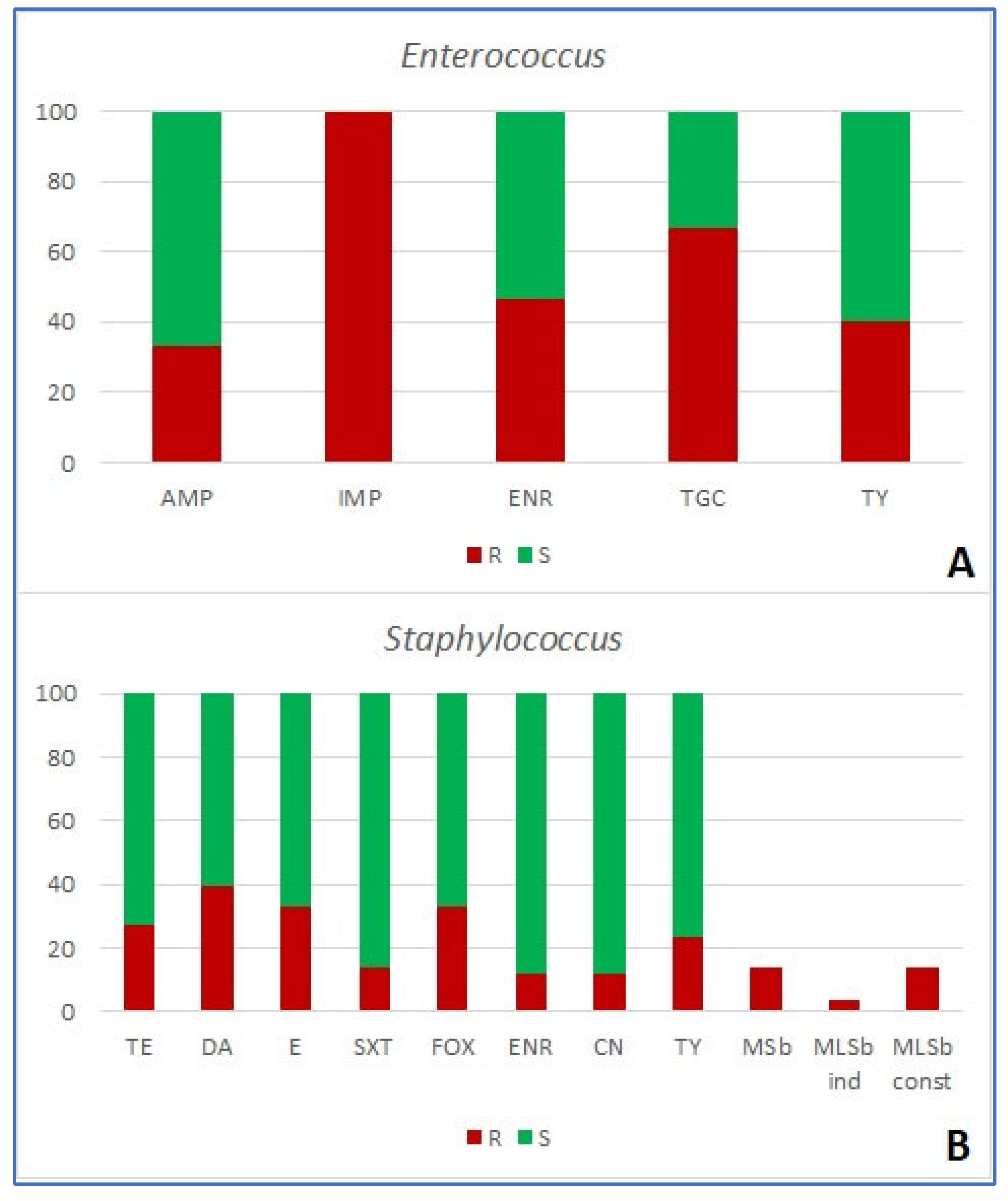

Table 2.
Prevalence of resistance profile in groups of bacteria isolated from wounds of companion animals.
Table 2.
Prevalence of resistance profile in groups of bacteria isolated from wounds of companion animals.
| number of antibiotics bacteria are resistant to (%) | resistance mechanism | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group of bacteria | R0 | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | MDR | ESBL | MRS | MSb | MLSb-i | MLSb-c |
| Enterobacterales (n=38) | 3 (7.9) | 2 (5.3) | 12 (31.6) | 7 (18.4) | 6 (15.8) | 3 (7.9) | 2 (5.3) | 2 (5.3) | 1 (2.6) | 16 (42.1) | 5 (13.2) | - | - | - | - |
| Pseudomonas (n=7) | 0 | 0 | 0 | 1 | 0 | 3 (42.9) | 3 (42.9) | 0 | 0 | 7 (100) | 0 | - | - | - | - |
| Acinetobacter (n=9) | 0 | 5 (55.6) | 1 (11.1) | 1 (11.1) | 1 (11.1) | 1 (11.1) | 0 | - | - | 3 (33.3) | - | - | - | - | - |
| Enterococcus (n=15) | 0 | 3 (20.0) | 3 (20.0) | 2 (13.3) | 7 (46.7) | 0 | - | - | - | 9 (60) | - | - | - | - | - |
| Staphylococcus (n=51) | 14 (27.5) | 15 (29.4) | 8 (15.7) | 3 (5.9) | 5 (9.8) | 0 | 3 (5.9) | 3 (5.9) | 0 | 14 (27.5) | - | 17 (33.3) | 7 (13.7) | 2 (3.9) | 7 (13.7) |
Finally, PCR tests were carried out to search for 12 genetic determinants of antibiotic resistance to all antimicrobial classes used in the treatment of Gram-positive and Gram-negative bacteria. Due to the fact that various resistance mechanisms may be characteristic of Gram-positive and Gram-negative bacteria, some genes examined in these two groups of bacteria varied (such as methicillin and MLSb resistance determinants in Gram-positives and ESBL determinants in Gram-negatives), while others (such as strA determining the aminoglycoside resistance and sul3 that determines the resistance to sulfonamides) were common for both these groups.
Figure 4.
Positive results of PCR test for the few exemplary bacterial isolates. Lanes marked as M – DNA Marker 3 (A&A Biotechnology, Poland); lanes 1-18 (A) and 1-24 (B) – bacterial DNA isolates; lanes 5-7 and 13-18 (A) and 10,17, 19, 20 (B) – DNA bands of searched length, confirming the presence of (A) blaTEM and (b) lnuA genes.
Figure 4.
Positive results of PCR test for the few exemplary bacterial isolates. Lanes marked as M – DNA Marker 3 (A&A Biotechnology, Poland); lanes 1-18 (A) and 1-24 (B) – bacterial DNA isolates; lanes 5-7 and 13-18 (A) and 10,17, 19, 20 (B) – DNA bands of searched length, confirming the presence of (A) blaTEM and (b) lnuA genes.
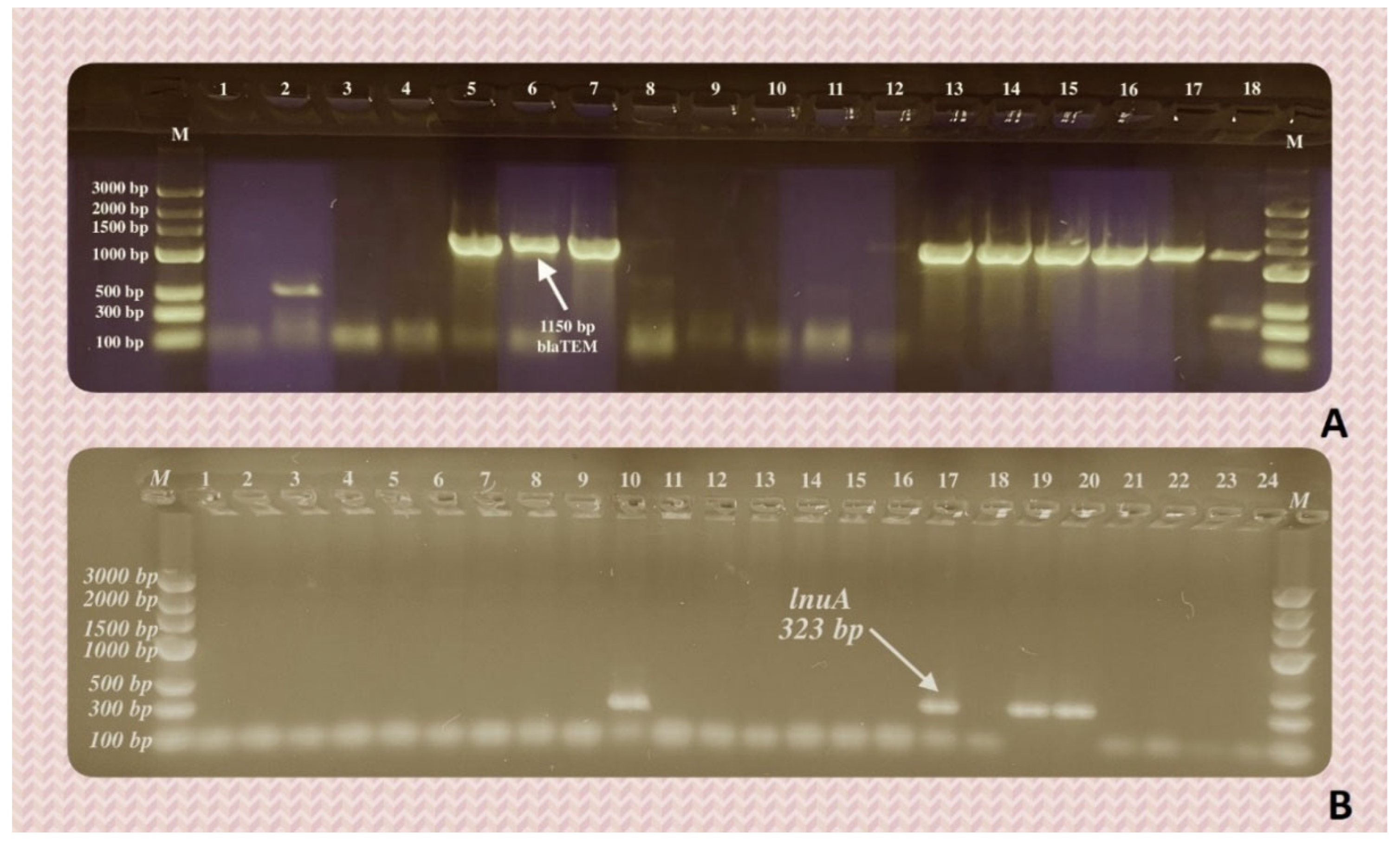
Ten out of the tested 12 genetic determinants of antimicrobial resistance were detected in the DNA extracted from bacterial isolates from cats, dogs and rabbit (Table 5, Table 6, Figure 7 A and B). Genes ereA (erythromycin esterase), qnrA and qnrD (plasmid-mediated quinolone resistance genes) were not detected in any of the examined samples. On the other hand, the streptomycin resistance gene, strA, was detected in 29 samples, in all groups thereof, in both Gram-positive and Gram-negative bacteria. In terms of the detection frequency, it was followed by sul3, a sulfonamide resistance gene (14 positive samples, in Gram-positive and Gram-negative isolates of both cats and dogs) and blaTEM, an ESBL determinant, characteristic of Gram-negative bacteria (13 positive samples in cats and dogs, Table 6, Figure 7A).
Table 3.
Positive results in Gram-positive (n =59) bacteria isolated from cats (n=25) and dogs (n=34).
Table 3.
Positive results in Gram-positive (n =59) bacteria isolated from cats (n=25) and dogs (n=34).
| Animal | n (%) | ||||||
| mecA | msrA | lnuA | strA | tetK | sul3 | ereA | |
| Cat | 0 | 3 (12) | 0 | 7 (28) | 6 (21.4) | 6 (21.4) | 0 |
| Dog | 4 (11.8) | 1 (2.9) | 4 (11.8) | 10 (29.4) | 3 (8.8) | 3 (8.8) | 0 |
Table 4.
Positive results in Gram-negative (n= 49) bacteria isolated from cats (n=12), dogs (n=33) and rabbit (n=4).
Table 4.
Positive results in Gram-negative (n= 49) bacteria isolated from cats (n=12), dogs (n=33) and rabbit (n=4).
| Animal | n (%) | ||||||
| blaTEM | blaSHV | blaCTX-M | blaOXA-1 | sul3 | qnrD | strA | |
| Cat | 3 (25) | 0 | 1 (8.3) | 0 | 3 (25) | 0 | 1 (8.3) |
| Dog | 10 (30.3) | 3 (9.1) | 1 (3) | 1 (3) | 2 (6.1) | 0 | 10 (30.3) |
| Rabbit | 0 | 1 (25) | 0 | 0 | 0 | 0 | 1 (25) |
Interestingly, there have been six canine and one feline bacterial isolates with phenotypic and molecular resistance profiles (Table 5) that could make them alarming, as they were resistant to all or nearly all examined antimicrobial agents and their DNA contained three or four different ARGs.
Table 5.
Selected phenotypic and molecular resistance profiles of canine and feline bacteria isolated from wounds.
Table 5.
Selected phenotypic and molecular resistance profiles of canine and feline bacteria isolated from wounds.
| Origin | Species | phenotype of resistance (antibiotic class) | resistance genes (type of resistance) |
| feline | Enterococcus faecalis | IMP (β-lactam -carbapeneme) ENR (fluoroquinolone) TGC (tetracycline) TY (macrolide) |
mecA (methicillin) msrA (macrolides) lnuA (lincosamides) tetK (tetracyclines) |
| canine | Enterococcus faecalis | IMP (β-lactam -carbapeneme) ENR(fluoroquinolone) TGC (tetracycline) TY (macrolide) |
msrA (macrolides) strA (aminoglycosides) tetK (tetracyclines) sul3 (sulfonamides) |
| canine | Staphylococcus sciuri | - |
lnuA (lincosamides) strA (aminoglycosides) tetK (tetracyclines) |
| canine | Staphylococus pseudintermedius | TE (tetracycline) DA (lincosamide) E (macrolide) SXT (diaminopyrimidines/sulfonamide) ENR (fluoroquinolone) CN (aminoglycoside) TY (macrolide) |
lnuA (lincosamides) strA (aminoglycosides) tetK (tetracyclines) |
| canine | Escherichia coli | CN (aminoglycoside) AMC (β-lactam/ β-lactamase inhibitor) AMP (β-lactam - aminopenicillin) IMP (β-lactam -carbapeneme) |
blaTEM (ESBL) blaSHV (ESBL) strA (aminoglycosides) |
| canine | Escherichia coli | CTX (β-lactam – 3rd gen. cephalosporin) AMC (β-lactam/ β-lactamase inhibitor) CAZ (β-lactam – 3rd gen. cephalosporin) AMP (β-lactam - aminopenicillin) TY (macrolide) |
blaTEM (ESBL) blaSHV (ESBL) strA (aminoglycosides) |
| canine | Proteus mirabilis | CN (aminoglycoside) SXT (diaminopyrimidines/sulfonamide) CTX (β-lactam – 3rd gen. cephalosporin) AMC (β-lactam/ β-lactamase inhibitor) CAZ (β-lactam – 3rd gen. cephalosporin) ENR (fluoroquinolone) AMP (β-lactam - aminopenicillin) IMP (β-lactam/carbapeneme) |
blaTEM (ESBL) blaOXA-1 (ESBL-carbapenemes) strA (aminoglycosides) |
3. Discussion
There is growing evidence that resistant bacteria (including multidrug resistant individuals) occur in companion animals and that many species among them are shared between animals and humans ([2]). With this in mind, the inappropriate use of antimicrobials in animals may result in the selection and spread of antimicrobial resistance, thus constituting a potential risk to public health [2]. This study provides data on bacteria colonizing wounds of companion animals, their antimicrobial resistance profiles and genetic determinants of resistance to all classes of antimicrobial agents among Gram-negative and Gram-positive bacteria. The distribution of bacterial taxa shows that Staphylococcus spp. (with S. pseudintermedius and S. aureus), Enterococcus spp. (with E. faecalis), E. coli, Acinetobacter spp. (with A. ursingii) and Pseudomonas (with P. aeruginosa) were the most frequently isolated from all samples. High prevalence of Staphylococcus spp. with higher numbers of S. pseudintermedius than of S. aureus was reported by [6,7]. [3] also observed that Staphylococcus (including S. intermedius, later reclassified to S. pseudintermedius) was the most frequent among wound-infecting Gram-positive bacteria, while E. coli was most frequently identified Gram-negative. Staphylococcus pseudintermedius is the most common canine bacterial pathogen, but it is, indeed, accompanied by a variety of other – both Gram-positive and Gram-negative bacteria.
In recent years, increased attention is being paid to the welfare of small, companion animals, which results in e.g. increased expenses on veterinary care. This involves frequent use of antimicrobial agents in pets, and many of these antimicrobials are commonly used in both human and veterinary medicine. However, identification of causal agents and their antimicrobial susceptibility is often neglected, leading to inappropriate empirical treatment [8]. In both cats and dogs, the most frequent causes of antimicrobial use are wound infections [8]. The most frequently used classes of antibiotics in animals are fluoroquinolones, β-lactams, cephalosporins, sulfonamides, macrolides and glycopeptides [9]. Among the consequences of antimicrobial use in companion animals is that the amounts and patterns of antibiotic administration reflect in the rate at which the resistance develops and spreads in the exposed bacterial population [8].
In our study, high percentage of Enterobacterales were resistant to β-lactam antibiotics (amoxicillin/clavulanic acid and ampicillin – 68.3% and 52.6%, respectively), while 100% of Pseudomonas isolates were insusceptible to ceftazidime (cephalosporin), piperacillin/tazobactam (β-lactam), imipeneme (carbapaneme) and tylosin (macrolide). However, the resistance to enrofloxacin (fluoroquinolone) ranged from 11.8% in Staphylococci to 46.7% in Enterococci and it was never the highest - only in Acinetobacter spp. it was the third highest (44.4%) among the observed resistance rates. Generally, enrofloxacin is commonly used systemically in infection treatment of small animals and although it is still efficient, there have been cases of treatment failure [8]. With respect to the fact that inappropriate use (e.g. pulse-dose, low dose) is very common in treatment of small animals, this might favor the development of resistant strains, particularly when long-term treatment is required [10]. As far as the antibiotic administration effect is concerned, it has been reported that enrofloxacin treatment promotes MDR E. coli colonization and that the proportion of dogs carrying resistant E. coli increased with the duration of hospital stay and with the antimicrobial treatment [2].
Even though the antimicrobial resistance is spread among environmental microorganisms, human and animal pathogens, of special concern seems the situation when the resistance against last-resort antimicrobial agents for human medicine is detected, particularly in microorganisms that can be easily transmitted between animals and humans [2,8]. Insusceptibility to imipeneme (carbapeneme antimicrobial) has been detected quite frequently in our study. All Enterococcus spp. and all Pseudomonas spp. isolates were resistant to this antimicrobial. Importantly, both canine E. faecalis MDR strains, positive for four genetic determinants of resistance and resistant to four antibiotics, mentioned in Table 7, were also resistant to imipeneme. In the case of Gram-negative isolates mentioned in the special concern group (Table 7), one canine E. coli and P. mirabilis were also imipeneme-resistant. All these species are listed as human pathogens, too. They cause, among many others, urinary tract infections [8,11]. Close interaction between pets and humans favors the transmission of bacteria by both direct contact and through the domestic environment and children are at greater risk than adults. This is due to their more common and closer physical contact with pets and with household environment [8]. The most important hazard for human health is therefore related to the transmission of antimicrobial resistant bacteria from pets to humans and the related zoonotic infections [2].
Notably, antimicrobial resistance can be transmitted by low bacterial numbers or even in the absence thereof, if only the genetic determinants are present in the environment. The resistance gene transfer frequently occurs horizontally. Most classes of antibiotics have long been used in both human and veterinary medicine, thus the same resistance genes are being identified in bacteria isolated from humans and animals [8]. In our study, ten genetic determinants of antimicrobial resistance have been identified in bacterial isolates of cats, dogs and rabbit. Streptomycin resistance (strA) was most frequently identified, followed by sulfonamide resistance (sul3) and one of the ESBL determinants (blaTEM). Studies on the genetic determinants in bacteria isolated from companion animals are scarce, however the listed genes have also been mentioned by other authors to occur in bacteria isolated from companion animals [8,12,13]. All four examined ESBL determinants were detected in this study in the following frequency: blaTEM > blaSHV > blaCTX-M > blaOXA. [12] on the other hand, detected blaTEM, blaCTX-M and blaSHV in similar proportions in E. coli isolates obtained from dogs and their owners. [13] examined the presence of nine genes determining the resistance to tetracyclines, quinolones, aminoglycosides, sulfonamides and trimethoprim in E. coli isolates from pet cats. Similarly as in our study, qnr genes (determining the quinolone resistance) were very rare or absent. In this study, the resistance to enrofloxacin, which is most frequently applied fluoroquinolone veterinary antibiotic varied and ranged from 11.7% in staphylococci to 46.7% in enterococci. In an earlier study focused on antimicrobial resistance and the molecular resistance mechanisms in chicken feces-derived E. coli, [14] observed that the fluoroquinolone resistance would reach even 93.3% of isolates. At the same time, the resistance to β-lactams was also very frequent, in some experimental groups reaching 100% of bacterial isolates. Consequently, qnrB and qnrS genes were very frequent (detected in more than 81% and 86% of isolates of some experimental groups, respectively). However, the selection pressure put on bacteria present in farm poultry is much higher than bacteria derived from pet animals, despite the restrictions resulting from the European Union regulations.
4. Materials and Methods
4.1. Collection of Samples
A total of 145 wound swabs were collected from companion animals (84 dogs, 60 cats and one rabbit) that underwent treatment at the University of Agriculture Veterinary Clinic. The samples were collected with a sterile swab by a qualified veterinarian (Figure 1), immediately transferred to the laboratory of Microbiology and Biomonitoring and inoculated on selective media for the isolation and preliminary identification of bacterial pathogens and opportunistic pathogens.
Figure 5.
Collection of swab sample from a skin wound of a dog.
4.2. Isolation and Identification of Bacteria
The culture media included: Chromogenic UTI Medium (ThermoFisher Scientific, UK) for presumptive identification and differentiation of Enterococcus, Escherichia coli, Proteus, Pseudomonas, Staphylococcus and Klebsiella; MacConkey agar (Biomaxima, Lublin, Poland) for Gram-negative Enterobacteriaceae; Tryptone-bile-X-glucuronide agar (Biomaxima, Lublin, Poland) for E. coli; Slanetz-Bartley agar (Biomaxima, Lublin, Poland) for Enterococcus; Baird Parker agar (Biomaxima, Lublin, Poland) for Staphylococcus aureus; Columbia CNA agar with 5% sheep blood (ThermoFisher Scientific, UK) for Gram-positive cocci and determination of their haemolysis type; Cetrimide agar for Pseudomonas. All cultures were conducted for 24–48 h at 37 ± 1 °C). After incubation, different morphotypes, characteristic of the examined groups of bacteria (Figure 2), were subcultured by plate streaking, followed by observations of Gram-stained preparations (Figure 3). The species identification was performed via MALDI-TOF (Matrix-assisted laser desorption/ionisation-time of-flight) technology on a Bruker microflex ® mass spectrometry instrument. This technique uses a laser to ionize and separate molecules by their mass-to-charge ratio. The resulting peptide mass fingerprint can be compared to a database of known spectra to find the best match. The high throughput, accuracy, and small required sample size of the MALDI-TOF MS have made it a widely used technique in the clinical microbiological laboratory. All identified bacteria had a log (score) value higher or equal to 2.0, which indicated ‘highly probable species identification’.
Figure 6.
Typical colonies of (A) E. coli, P. aeruginosa and S. aureus, (B) E. faecalis, K. pneumoniae, (C) Staphylococcus sp. (C) and S. aureus (D), grown on UTI medium (A and B) and on Baird-Parker medium (C and D).
Figure 6.
Typical colonies of (A) E. coli, P. aeruginosa and S. aureus, (B) E. faecalis, K. pneumoniae, (C) Staphylococcus sp. (C) and S. aureus (D), grown on UTI medium (A and B) and on Baird-Parker medium (C and D).

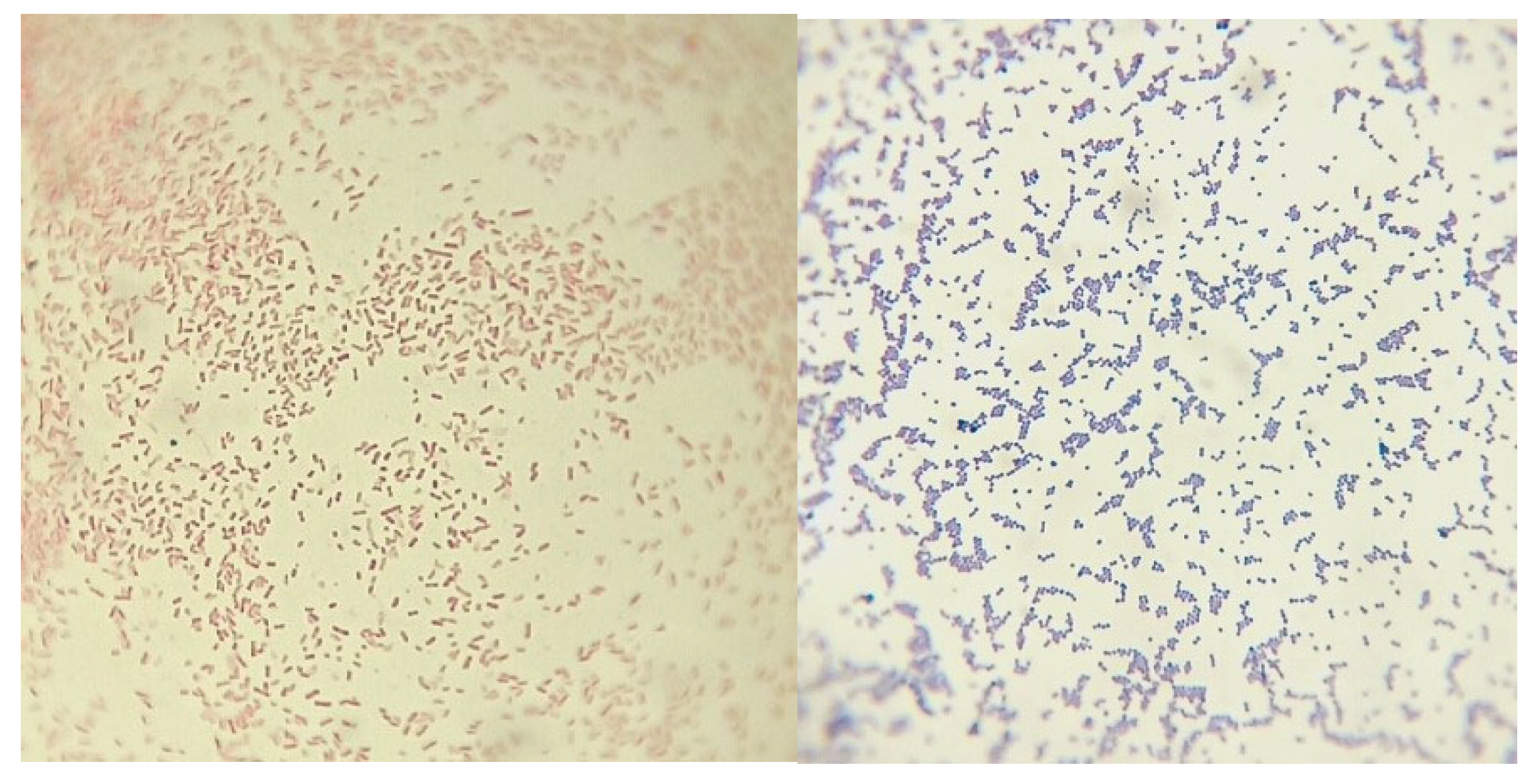
Figure 7.
Light microscope image (1000 ×) of Pseudomonas aeruginosa (on the left) and Staphylococcus aureus (on the right).
Figure 7.
Light microscope image (1000 ×) of Pseudomonas aeruginosa (on the left) and Staphylococcus aureus (on the right).
4.3. Antibacterial Susceptibility Tests
A total of 136 bacterial strains were isolated from the examined swabs. The antimicrobial susceptibility patterns were assessed using the disk-diffusion method, based on the recommendations of the Polish National Reference Centre for Antimicrobial Susceptibility [15]. The examinations were carried out on 120 isolates of Gram-negative (n=54) and Gram-positive (n=66) bacteria. As the examined bacteria were classified to 28 different genera, the recommendations for both Gram-positive and Gram-negative bacteria were applied and divided into five groups (Table 1). Antimicrobial disk cartridges were obtained from Oxoid (Great Britain). The ESBL (extended-spectrum beta-lactamase)-positive Enterobacteriaceae and Pseudomonas strains were confirmed with the double disk synergy test [16]. Macrolide, lincosamid and streptogramin b type of resistance was assessed according to [17].
Table 6.
Antimicrobial susceptibility tests according to groups of bacteria.
| Enterobacterales (E. coli, Klebsiella, Proteus, Enterobacter) | Pseudomonas | Acinetobacter | Enterococcus | Staphylococcus | |
| No of strains in total | 38 | 7 | 9 | 15 | 51 |
| Cats | 7 | 1 | 3 | 5 | 21 |
| Dogs | 28 | 6 | 5 | 10 | 30 |
| Rabbit | 3 | 0 | 1 | 0 | 0 |
| antimicrobial disks abbreviations * | ENR | ENR | ENR | ENR | ENR |
| AMC (ESBL)** | AMC (ESBL)** | AK | AMP | E (MLSb)** | |
| CAZ (ESBL)** | CAZ (ESBL)** | CN | MEM/IMP | DA (MLSb)** | |
| CTX (ESBL)** | CTX (ESBL)** | MEM/IMP | TGC | FOX (MRS)** | |
| AMP | AK | SXT | TY | CN | |
| CN | MEM/IMP | TY | SXT | ||
| SXT | TZP | TE | |||
| MEM/IMP | TY | TY | |||
| TY |
*ENR (enrofloxacin 5 µg), E (erythromycin 15 µg), DA (clindamycin 2 µg), FOX (cefoxitin 30 µg), CN (gentamicin 10 µg), SXT (trimethoprim/sulfamethoxazole 1.25/23.75 µg), TE (tetracycline 30 µg), TY (tylosin 30 µg), AMC (amoxicillin/clavulanic acid 20/10 µg), CAZ (ceftazidime 30 µg), CTX (cefotaxime 30 µg), AMP (ampicillin 10 µg), MEM (meropeneme 10 µg), IMP (imipeneme 10 µg), TZP (piperacillin/tazobactam 100/10 µg), TGC (tigecycline 15 µg). ** in brackets – mechanisms of resistance determined using the provided antimicrobial disks: ESBL – extended spectrum beta lactamases; MLSb – constitutive or inducible resistance to macrolides, lincosamids and streptogramins b; MRS – methicillin resistance.
After incubation for 18 – 24 h at 36 ± 1°C the growth inhibition zone diameters around disks were measured (in mm) and compared to the most recent breakpoint values provided by the European Committee on Antimicrobial Susceptibility Testing [18].
4.4. Assessment of Genes Conferring the Bacterial Resistance to Different Groups of Antimicrobials
Bacterial DNA was extracted from all 120 strains subjected to antimicrobial resistance testing and from the control strains: susceptible E. coli ATCC 25922, Methicillin-susceptible S. aureus ATCC 25923; environmental strains of methicillin resistant S. aureus and ESBL-positive E. coli were used as positive control. DNA extraction was conducted from overnight cultures, using the Genomic Mini DNA extraction set of reagents (A&A Biotechnology, Gdańsk, Poland), following the manufacturer’s recommendations.
PCR amplifications were conducted using specific primers (Table 7) to search for genes conferring the methicillin resistance in staphylococci, antibiotics from groups of macrolides, lincosamids and streptogramins in Gram-positive cocci, ESBL determinants in Enterobacteriaceae and Pseudomonas, aminoglycosides, carbapenems, fluoroquinolones, sulfonamides and tetracyclines in both Gram-negative and Gram-positive isolates. The reactions were performed in a volume of 25 μl containing 50 ng of DNA template, 12.5pM of each primer, 2× (12.5 μl) of PCR Mix Plus Green (A&A Biotechnology, Poland) filled up with ultrapure water up to 25 μl. The following temperature profile was used for the reactions: initial denaturation at 95 °C for 5 min, followed by 35 cycles of 94 °C for 45 s, annealing for 45 s at temperatures corresponding to individual primers, then extension at 72 °C for 1 min with the final extension at 72 °C for 10 min. The reactions were performed in a T100 Thermal Cycler (Bio-Rad, USA). The PCR products were electrophoresed for 60min in 1 × TBE SimplySafe (EurX, Poland)-stained 1% agarose gel and visualized in UV light. DNA size Marker 3 (A&A Biotechnology, Gdańsk, Poland) was used to assess the size of product bands.
Table 7.
PCR primers used in the study.
| No. | Gene | Primer | Sequence (5’-3’) | Annealing temp. (°C) | Product length (bp) | Reference |
|---|---|---|---|---|---|---|
| 1. | msrA | msrA-F | GGCACAATAAGAGTGTTTAAAGG AAGTTATATCATGAATAGATTGTCCTGTT |
50 | 940 | [19] |
| msrA-R | ||||||
| 2. | ereA | ereA-F | AACACCCTGAACCCAAGGGACG CTTCACATCCGGATTCGCTCGA |
57 | 420 | [20] |
| ereA-R | ||||||
| 3. | lnuA | lnuA-F | GGTGGCTGGGGGGTAGATGTATTAACTGG GCTTCTTTTGAAATACATGGTATTTTTCGATC |
57 | 323 | [19] |
| lnuA-R | ||||||
| 4. | mecA | mecA-F | GTAGAAAATGACTGAACGTCCGATAA CAATTCCACATTGTTTCGGTCTAA |
55 | 310 | [21] |
| mecA-R | ||||||
| 5. | tetK | tetK-F | TCGATAGGAACAGCAGTA CAGCAGATCCTACTCCTT |
55 | 169 | [22] |
| tetK-R | ||||||
| 6. | blaTEM | blaTEM-F | ATTCTTGAAGACGAAAGGGC ACGCTCAGTGGAACGAAAAC |
60 | 1150 | [23] |
| blaTEM-R | ||||||
| 7. | blaSHV | blaSHV-F | CACTCAAGGATGTATTGTG TTAGCGTTGCCAGTGCTCG |
52 | 885 | [23] |
| blaSHV-R | ||||||
| 8. | blaCTX-M | blaCTX-M-F | CGATGTGCAGTACCAGTAA TTAGTGACCAGAATCAGCGG |
55 | 585 | [24] |
| blaCTX-M-R | ||||||
| 9. | blaOXA-1 | blaOXA-1-F | ACACAATACATATCAACTTCGC AGTGTGTTTAGAATGGTGATC |
61 | 813 | [23] |
| blaOXA-1-R | ||||||
| 10. | sul3 | sul3-F | ACCACCGATAGTTTTTCCGA TGCCTTTTTCTTTTAAAGCC |
62 | 199 | [25] |
| sul3-R | ||||||
| 11. | qnrA | qnrA-F | GGGTATGGATATTATTGATAAAG CTAATCCGGCAGCACTATTA |
55 | 580 | [26] |
| qnrA-R | ||||||
| 12. | qnrD | qnrD-F | AGTGAGTGTTTAGCTCAAGGAG CAGTGCCATTCCAGCGATT |
53 | 175 | [27] |
| qnrD-R | ||||||
| 13. | strA | strA-F | TCAATCCCGACTTCTTACCG CACCATGGCAAACAACCATA |
52 | 126 | [28] |
| strA-R |
4.5. Statistical Analysis
The significance of differences in bacterial resistance to various antimicrobial agents and the presence of genetic determinants of antimicrobial resistance was assessed using the chi-square test (https://www.socscistatistics.com/tests/chisquare2/default2.aspx). The significance level was set at a p value of <0.05 for all tests.
5. Conclusions
The issues of infection control, antimicrobial resistance and spread of resistance genes are the same in companion animal and human hospitals, as these facilities are characterized by intensive use of antibiotics and high density of patients. For these reasons, they are high-risk environments for the occurrence and spread of nosocomial infections, resistant bacteria and genetic determinants thereof [2,29]. As shown in the current study, the composition of bacterial species that infect companion animals’ wounds can vary greatly and even though some species could be considered as more prevalent than others, none of them were prevalent enough to conclude that their presence in pets’ wounds could be assumed with high probability. Moreover, these bacteria can carry a number of resistance genes that determine the insusceptibility to a variety of antimicrobial agents. This study also resulted in two disturbing observations: the first one refers to the prevalence of multidrug resistant bacteria (in some groups reaching even 100% of isolates), while the second is the detection of bacteria that are also human pathogens (e.g. Proteus mirabilis), which were resistant to nearly all or all antimicrobial agents tested and carried three or four antibiotic resistance genes. The widely reported risk of transfer of resistant bacteria between animals and humans, increasing demand for advanced therapies in companion animals coupled with the spread of MDR bacteria, may cause the need for searching new antimicrobial therapies in the future in veterinary medicine.
Author Contributions
Conceptualization, A.L-B. and K.S.; methodology, A.L-B., K.S. and M.T; software, A.L-B and K.D.; validation, K.S. and K.D.; formal analysis, A.L-B and K.S.; investigation, N.C., A.R., K.B., M.H., D.S., I.S. and D.C.; resources, A.L-B., K.S., K.D., I.S. and M.T; data curation, K.S.; writing—original draft preparation, A.L.-B.; writing—review and editing, K.S. and K.D.; visualization, A.L.-B. and K.S.; supervision, A.L.-B.; project administration, A.L-B.; funding acquisition, A.L-B. and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the measures of the state budget Polish Ministry of Education and Science under the program “Student scientific circles create innovations”—Research on the effectiveness of innovative hyaluron bionanocomposites with nanoencapsulated ozonated olive oil, graphene, and nanosilver against bacteria that infect wounds in dogs and cats (Agreement No. SKN/SP/569551/2023) and by the statutory measures of the University of Agriculture in Kraków.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
“Author Kinga Dworak was employed by the company ‘Diagnostyka S.A. Medical Microbiological Laboratory, Na Skarpie 66, 31-913 Krakow, Poland’. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest”.
References
- Marco-Fuertes, A.; Marin, C.; Lorenzo-Rebenaque, L.; Vega, S.; Montoro-Dasi, L. Antimicrobial Resistance in Companion Animals: A New Challenge for the One Health Approach in the European Union. Veterinary Sciences 2022, 9. [Google Scholar] [CrossRef]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M.; et al. Public Health Risk of Antimicrobial Resistance Transfer from Companion Animals. Journal of Antimicrobial Chemotherapy 2017, 72, 957–968. [Google Scholar] [CrossRef]
- Kožár, M.; Hamilton, H.; Koščová, J. Types of Wounds and the Prevalence of Bacterial Contamination of Wounds in the Clinical Practice of Small Animals. Folia Veterinaria 2018, 62, 39–47. [Google Scholar] [CrossRef]
- Windahl, U.; Bengtsson, B.; Nyman, A.K.; Holst, B.S. The Distribution of Pathogens and Their Antimicrobial Susceptibility Patterns among Canine Surgical Wound Infections in Sweden in Relation to Different Risk Factors. Acta Veterinaria Scandinavica 2015, 57. [Google Scholar] [CrossRef] [PubMed]
- Scott Weese, J. Antimicrobial Resistance in Companion Animals. Animal health research reviews / Conference of Research Workers in Animal Diseases 2008, 9, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fernández, R.; Durán, I.; Molina-López, R.A.; Darwich, L. Antimicrobial Resistance in Bacteria Isolated From Cats and Dogs From the Iberian Peninsula. Frontiers in Microbiology 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Pires Dos Santos, T.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic Review on Global Epidemiology of Methicillin-Resistant Staphylococcus Pseudintermedius: Inference of Population Structure from Multilocus Sequence Typing Data. Frontiers in microbiology 2016, 7, 1599. [Google Scholar] [CrossRef]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet Animals as Reservoirs of Antimicrobial-Resistant Bacteria: Review. Journal of Antimicrobial Chemotherapy 2004, 54, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Ganière, J.P.; Médaille, C.; Limet, A.; Ruvoen, N.; André-Fontaine, G. Antimicrobial Activity of Enrofloxacin against Staphylococcus Intermedius Strains Isolated from Canine Pyodermas. Veterinary dermatology 2001, 12, 171–175. [Google Scholar] [CrossRef]
- Actor, J.K. 12 - Clinical Bacteriology. In; Actor, J.K.B.T.-E.I.R.I. and M. (Second E., Ed.; W.B. Saunders: Philadelphia, 2012; pp. 105–120 ISBN 978-0-323-07447-6.
- Carvalho, A.C.; Barbosa, A. V; Arais, L.R.; Ribeiro, P.F.; Carneiro, V.C.; Cerqueira, A.M.F. Resistance Patterns, ESBL Genes, and Genetic Relatedness of Escherichia Coli from Dogs and Owners. Brazilian Journal of Microbiology 2016, 47, 150–158. [Google Scholar] [CrossRef]
- Akhtardanesh, B.; Ghanbarpour, R.; Ganjalikhani, S.; Gazanfari, P. Determination of Antibiotic Resistance Genes in Relation to Phylogenetic Background in Escherichia Coli Isolates from Fecal Samples of Healthy Pet Cats in Kerman City. Veterinary research forum : an international quarterly journal 2016, 7, 301–308. [Google Scholar] [PubMed]
- Lenart-Boron, A.; Augustyniak, K.; Boron, P. Screening of Antimicrobial Resistance and Molecular Detection of Fluoroquinolone Resistance Mechanisms in Chicken Faeces-Derived Escherichia Coli. Veterinarni Medicina 2016, 61, 80–89. [Google Scholar] [CrossRef]
- Gniadkowski, M.; Żabicka, D.; Hryniewicz, W. Rekomendacje Doboru Testów Do Oznaczania Wrażliwości Bakterii Na Antybiotyki i Chemioterapeutyki 2009 Oznaczanie Wrażliwo ś Ci Pałeczek Gram-Ujemnych. 2009, 1–29.
- Drieux, L.; Brossier, F.; Sougakoff, W.; Jarlier, V. Phenotypic Detection of Extended-Spectrum β-Lactamase Production in Enterobacteriaceae: Review and Bench Guide. Clinical Microbiology and Infection 2008, 14, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Fiebelkorn, K.R.; Crawford, S.A.; McElmeel, M.L.; Jorgensen, J.H. Practical Disk Diffusion Method for Detection of Inducible Clindamycin Resistance in Staphylococcus Aureus and Coagulase-Negative Staphylococci. Journal of clinical microbiology 2003, 41, 4740–4744. [Google Scholar] [CrossRef]
- EUCAST European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters. http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_5.0_Breakpoint_Table_01.pdf 2023, 0–77.
- Lina, G.; Quaglia, A.; Reverdy, M.E.; Leclercq, R.; Vandenesch, F.; Etienne, J. Distribution of Genes Encoding Resistance to Macrolides, Lincosamides, and Streptogramins among Staphylococci. Antimicrobial agents and chemotherapy 1999, 43, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of Erythromycin-Resistant Determinants by PCR. Antimicrobial agents and chemotherapy 1996, 40, 2562–2566. [Google Scholar] [CrossRef]
- Geha, D.J.; Uhl, J.R.; Gustaferro, C.A.; Persing, D.H. Multiplex PCR for Identification of Methicillin-Resistant Staphylococci in the Clinical Laboratory. Journal of clinical microbiology 1994, 32, 1768–1772. [Google Scholar] [CrossRef]
- Pazda, M.; Rybicka, M.; Stolte, S.; Piotr Bielawski, K.; Stepnowski, P.; Kumirska, J.; Wolecki, D.; Mulkiewicz, E. Identification of Selected Antibiotic Resistance Genes in Two Different Wastewater Treatment Plant Systems in Poland: A Preliminary Study. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Sáenz, Y.; Briñas, L.; Domínguez, E.; Ruiz, J.; Zarazaga, M.; Vila, J.; Torres, C. Mechanisms of Resistance in Multiple-Antibiotic-Resistant Escherichia Coli Strains of Human, Animal, and Food Origins. Antimicrobial Agents and Chemotherapy 2004, 48, 3996–4001. [Google Scholar] [CrossRef]
- Batchelor, M.; Hopkins, K.; Threlfall, E.J.; Clifton-Hadley, F.A.; Stallwood, A.D.; Davies, R.H.; Liebana, E. BlaCTX-M Genes in Clinical Salmonella Isolates Recovered from Humans in England and Wales from 1992 to 2003. Antimicrobial Agents and Chemotherapy 2005, 49, 1319–1322. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowski, R.; Linke, B.; Krahn, I.; Gartemann, K.-H.; Gützkow, T.; Eichler, W.; Pühler, A.; Schlüter, A. Detection of 140 Clinically Relevant Antibiotic-Resistance Genes in the Plasmid Metagenome of Wastewater Treatment Plant Bacteria Showing Reduced Susceptibility to Selected Antibiotics. Microbiology 2009, 155, 2306–2319. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.-J.; Nordmann, P. Multiplex PCR for Detection of Plasmid-Mediated Quinolone Resistance Qnr Genes in ESBL-Producing Enterobacterial Isolates. The Journal of antimicrobial chemotherapy 2007, 60, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, L.; Ashbolt, N.; Wang, X.; Cui, Y.; Zhu, X.; Xu, Y.; Yang, Y.; Mao, D.; Luo, Y. Arctic Antibiotic Resistance Gene Contamination, a Result of Anthropogenic Activities and Natural Origin. Science of The Total Environment 2018, 621, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Walsh, F.; Ingenfeld, A.; Zampicolli, M.; Hilber-Bodmer, M.; Frey, J.E.; Duffy, B. Real-Time PCR Methods for Quantitative Monitoring of Streptomycin and Tetracycline Resistance Genes in Agricultural Ecosystems. Journal of microbiological methods 2011, 86, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Morley, P.S. Surveillance for Nosocomial Infections in Veterinary Hospitals. The Veterinary clinics of North America. Equine practice 2004, 20, 561–576. [Google Scholar] [CrossRef]
Table 1.
Species of bacteria isolated from wounds of three groups of animals.
| Species | total (n=136) | cats (n=52) | dogs (n=80) | rabbits (n=4) |
|---|---|---|---|---|
| Gram-negative | 65 (47.79) | 23 (44.23) | 38 (47.50) | 4 (100) |
| Acinetobacter baumanni | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Acinetobacter calcoaceticus | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Acinetobacter johnsonii | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Acinetobacter pittii | 2 (1.47) | 0 | 1 (1.25) | 1 (25) |
| Acinetobacter ursingii | 5 (3.68) | 5 (9.62) | 0 | 0 |
| Aeromonas media | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Brevundimonas diminuta | 2 (1.47) | 1 (1.92) | 1 (1.25) | 0 |
| Citrobacter freundi | 3 (2.21) | 2 (3.85) | 1 (1.25) | 0 |
| Enterobacter cloacae | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Enterobacter hormaechei | 2 (1.47) | 0 | 2 (2.50) | 0 |
| Escherichia coli | 11 (8.09) | 1 (1.92) | 8 (10.0) | 2 (50) |
| Hafnia alvei | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Klebsiella pneumoniae | 2 (1.47) | 0 | 1 (1.25) | 1 (25) |
| Klebsiella spp. | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Leclercia adecarboxylata | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Pantoea agglomerans | 2 (1.47) | 0 | 2 (2.50) | 0 |
| Pantoea sp. | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Moraxella osloensis | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Proteus mirabilis | 3 (2.21) | 0 | 3 (3.75) | 0 |
| Proteus vulgaris | 5 (3.68) | 4 (7.69) | 1 (1.25) | 0 |
| Pseudomonas aeruginosa | 3 (2.21) | 0 | 3 (3.75) | 0 |
| Pseudomonas putida | 3 (2.21) | 1 (1.92) | 2 (2.50) | 0 |
| Pseudomonas graminis | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Pseudomonas fulva | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Pseudomonas koreensis | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Psychrobacter sanguinis | 2 (1.47) | 0 | 2 (2.50) | 0 |
| Psychrobacter pulmonis | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Serratia marcescens | 3 (2.21) | 3 (5.77) | 0 | 0 |
| Serratia liquefaciens | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Stenotrophomonas maltophilia | 2 (1.47) | 0 | 2 (2.50) | 0 |
| Gram-positive | 71 (52.21) | 29 (55.77) | 42 (52.50) | 0 |
| Bacillus pumilus | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Curtobacterium flaccumfaciens | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Enterococcus avium | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Enterococcus faecalis | 11 (8.09) | 3 (5.77) | 8 (10) | 0 |
| Enterococcus faecium | 3 (2.21) | 2 (3.85) | 1 (1.25) | 0 |
| Enterococcus hirae | 2 (1.47) | 1 (1.92) | 1 (1.25) | 0 |
| Kocuria rhizophila | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Lactococcus garvieae | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Lysinibacillus fusiformis | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Macrococcus canis | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Micrococcus luteus | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Microbacterium maritypicum | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Microbacterium oxydans | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Microbacterium paraoxydans | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Microbacterium phyllosphaerae | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Peribacillus simplex | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Staphylococcus aureus | 5 (3.68) | 4 (7.69) | 1 (1.25) | 0 |
| Staphylococcus capitis | 2 (1.47) | 2 (3.85) | 0 | 0 |
| Staphylococcus cohnii | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Staphylococcus devriesei | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Staphylococcus condimenti | 2 (1.47) | 2 (3.85) | 0 | 0 |
| Staphylococcus epidermidis | 2 (1.47) | 1 (1.92) | 1 (1.25) | 0 |
| Staphylococcus felis | 2 (1.47) | 2 (3.85) | 0 | 0 |
| Staphylococcus haemolyticus | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Staphylococcus pasteuri | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Staphylococcus lentus | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Staphylococcus pseudintermedius | 13 (9.56) | 3 (5.77) | 10 (12.5) | 0 |
| Staphylococcus schleiferi | 2 (1.47) | 0 | 2 (2.50) | 0 |
| Staphylococcus saprophyticus | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Staphylococcus warneri | 1 (0.74) | 1 (1.92) | 0 | 0 |
| Staphylococcus sciuri | 1 (0.74) | 0 | 1 (1.25) | 0 |
| Streptococcus canis | 5 (3.68) | 2 (3.85) | 3 (3.75) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



