
Submitted:
22 January 2024
Posted:
25 January 2024
You are already at the latest version
Abstract
Injuries from exposure to blasts are of significant concern in military operational settings, including tactical training, and is associated with self-reported concussion-like symptomology and physiological changes as increased intestinal permeability (IP) investigated herein. Timeseries gene expression and IP biomarker data were generated from “breachers” exposed to controlled, low-level explosive blast during training. Samples from 30 male participants at pre, post, follow-up blast exposure the next day were assayed via RNA-seq and ELISA. A battery of symptom data was also collected at each of these time points that acutely showed elevated symptom reporting related to headache, concentration, dizziness, and slowed thinking, dissipating ~16hrs following blast exposure. Evidence for bacterial translocation into circulation following blast exposure was detected by significant stepwise increase in microbial diversity (measured via alpha-diversity p=0.049). Alterations in levels of IP protein biomarkers (i.e., Zonulin, LBP, Claudin-3, I-FABP) assessed in a subset of these participants (n=23) further evidenced blast exposure associates with IP. The observed symptom profile was consistent with mTBI and was further associated with changes in bacterial translocation and intestinal permeability, suggesting that IP may be linked to a decrease in cognitive performance. These preliminary findings show for the first
Keywords:
Intestinal permeability
; leaky gut
; blast
; military
; mTBI
; brain-gut axis
; microbiome
INTRODUCTION
Across all 9/11 active duty and Veteran service members, ~20% have reported at least one mild traumatic brain injury (mTBI), with more than one-third of these having been attributed to blast related injuries from improvised explosive devices (IEDs) or rocket propelled grenades [1]. While data on the effect of blast on the gut microbiota is limited, intestinal dysfunction and systemic inflammation have been noted following TBI. Patients with TBI present with abdominal pain, gastric distension, altered intestinal motility, constipation, and/or ulcers, with these individuals sustaining more complications and experiencing worse outcomes with chronic dysfunction of the gastrointestinal system and disability [2].
TBI can also lead to increased gut permeability. Intestinal permeability (IP) following TBI is associated with a concomitant reduction in intestinal tight junction protein expression. Intestinal epithelial cells function as a physical barrier between the intestinal lumen and the underlying vasculature and lymphatics that is supported by the structural integrity of cellular tight junctions, which are composed of a series of interacting transmembrane proteins, including claudins, Occludin, and junctional adhesion molecules. Emerging findings from preclinical animal models of TBI have shown that decreased expression of the intestinal tight junction proteins ZO-1 and Occludin are linked to loss of tight junction barrier integrity and IP [3]. IP has been reported as early as 2hr following TBI and, as such, disruption of the anatomic and functional integrity of the gut can result in bacterial translocation into the circulatory system.
Several established IP biomarkers used in clinical settings include Zonulin (haptoglobin 2 precursor), Lipopolysaccharide-Binding Protein (LBP), Claudin-3 and intestinal Fatty Acid Biding Protein (I-FABP). Zonulin, an intestine synthesized protein [4], serves as an IP marker by directly enhancing the permeability of intestinal tight junctions between cells of the digestive tract [5,6]. LBP is a soluble acute-phase protein synthesized by intestinal cells that can be used to monitor gut leakage by detecting the presence of lipopolysaccharide (LPS). LBP can recognize and bind to LPS [7,8] the main component of the outer membrane of gram-native bacteria [9]. After binding, LBP then presents LPS to cell surface pattern recognition receptors (PRR) such as CD14 and TLR4, to elicit subsequent immune response to infection [10,11]. Claudin-3, also known as CLDN3, is a protein which is encoded by an intron-less gene CLDN3 in humans. It is an integral membrane protein and a component of tight junction strands and therefore serves as a marker of paracellular barrier integrity loss [12]. I-FABP is a cytosolic protein found within mature enterocytes and a marker of enterocyte turnover and intestinal cell wall damage. Moreover, investigations of the human blood microbiome in neurodegenerative and psychiatric disorders have also provided evidence of bacterial translocation and potential for intestinal permeability. However, since blood has been generally considered a sterile environment that lacks microbes, studies of the human blood microbiome have received little recognition until recently with the availability of large compilation of reference microbial genomes, where increased microbial community variability in blood samples of schizophrenia and Alzheimer’s disease patients have been reported [13,14].
The present study investigates microbial variability and alterations in intestinal permeability biomarkers using these approaches in blood samples collected from military service members participating in breaching exercises during training. “Breachers” and “breaching” refers to an occupational discipline in the military and law enforcement in which explosives can be used to make entry to secured structures, and breachers, during training for this discipline, are repeatedly exposed to blast shockwaves. Breachers, and others who fire heavy weapons, report mTBI-like symptomology during these training exposures. Within this cohort of breachers and corresponding blood sample specimens collected during the course of the breacher training, we aim to identify transcriptional and protein alterations associated with blast induced IP and associated symptom reporting. We hypothesize that exposure to blast will: (1) induce increased bacterial diversity in blood (2) induce alterations in levels of gut permeability protein biomarkers in circulation including Zonulin, LBP, Claudin-3, and I-FABP, and (3) that these molecular alterations related to leaky gut will be associated with acute blast-related exposure symptomology.
METHODS
All subjects consented to participate in the study and the human use protocol for interaction with the subjects was approved by the Institutional Review Board (IRB) of the Walter Reed Army Institute of Research (WRAIR, Silver Spring, MD; FYSA: WRAIR #2304) and chains of command prior to data collection. The procedures were followed in accordance with the ethical standards of the IRB, Army Regulation 70-25, and the Helsinki Declaration.
Sample demographics and symptom reporting. Data reflects three timepoints collected at one military training site. Thirty-three male subjects participated in a routine training protocol using explosives in the demolition of a concrete wall where cumulative exposure was measured as cumulative impulse (measured in pounds per square inch [psi] x milliseconds [ms]) for each participant in the field with wearable sensors [15]; however, blood sample specimens were available only for 30 participants investigated in the present study. Participant average age was 30.2 +/- 7.4 years. Note that all participants met military physical requirement standards and were fit for duty. Although the training protocol was open to participation of both male and female service members, no females participated during the data collection for this study. Participants’ self-reported history of mTBI and number of career breaching events were recorded prior to engagement in training and are shown in Figure 1; ranging from 0 to 400+ self-reported career breaching events. Blood samples were collected serially at pre-blast (morning 7:30AM to 9:00AM), post-blast (afternoon 4:30PM to 5:30PM) on the training day, and follow-up the next day (morning 7:30AM to 9:00AM) and stored in -80 °C for downstream experiments. On average the post blast time point corresponds to 1hr, and follow-up to 16hr following exposures to blast during training. Additionally, during the course of training, participants completed a 32-item, paper-and-pencil health symptom inventory at pre-, post-, and follow-up blast exposure timepoints in conjunction with each blood draw. Items on the symptom survey were the same as that of the Rivermead Post Concussion Symptoms Questionnaire [16,17], with additional items included relevant to capture additional effects previously observed as reported in blast exposure environments [18]. These items are consistent with concussion symptomology present in current clinical and research findings [19,20]. Items on this survey are on a 5-point Likert scale (0 “not experienced at all,” 1 “no more of a problem than before training,” 2 “mild problem–present but don’t really notice and doesn’t concern me,” 3 “moderate problem–I can continue what I am doing but I notice the problem,” 4 “severe problem–constantly present, feels like it could affect my performance”).
Total RNA sample/ library preparation sequencing. Blood samples were collected in Paxgene blood RNA tubes (PreAnalytix) according to manufacturer’s instructions were stored at -80˚C. RNA was extracted with Paxgene Blood RNA Kit (PreAnalytix) and globin mRNA removed with Globin Clear Human Globin mRNA Removal kit (Ambion). Total RNA sequencing libraries were prepared from RNA samples with RNA integrity numbers, RINs>6.0 (measured via BioAnalyzer) using the Illumina Stranded Total RNA Library Prep Kit with Ribo-Zero Gold in accordance with the manufacturer’s instructions. Briefly, 290ng-500ng of total RNA was used for ribosomal depletion and fragmented by divalent cations under elevated temperatures. The fragmented RNA underwent first strand synthesis using reverse transcriptase and random primers followed by second strand synthesis to generate cDNA. The cDNA fragments undergwent end repair, adenylation, and ligation of Illumina sequencing adapters. The cDNA library was enriched using 11 cycles of PCR and purified. Final libraries were then evaluated using PicoGreen (Life Technologies) and Fragment Analyzer (Advanced Analytics) and sequenced on an Illumina HiSeq2500 sequencer (v4 chemistry) using 2 x 125bp read lengths.
RNA-seq data preprocessing and microbial bioinformatics analysis. RNA sequencing reads were aligned to the human reference hg19 using STAR aligner (v2.7.5b) [21]. Following alignment of RNA sequencing reads, read pairs that mapped to the human genome were eliminated. We then performed normalization by sub-sampling the remaining reads for each sample to 100,000 reads, and then used FASTX and Prinseq to filter out low-quality and low-complexity reads; that is, reads with at least 75% of their base pairs having quality lower than 30 (FASTX) [22] and reads with sequences of consecutive repetitive nucleotides (Prinseq) [23]. Next, the remaining reads were realigned to the reference human genome and transcriptome (Ensembl GRCh38 transcriptome and Ensembl hg38 build) using Megablast aligner (Blast v.2.9.0+) to filter out any remaining potentially human reads. After QC there were an average of 41,770 reads per sample. These remaining reads were used as candidate microbial reads. Phylogenetic classification was then performed using Kraken2 [24] to assign the filtered candidate microbial reads to the microbial genes from 33 distinct taxa on the phylum level.
For analysis of microbial diversity, alpha diversity on phylum level within each sample was determined using the inverse Simpson index as follows,
where R is richness (the total number of types of phyla in the sample) and pi is proportional abundance of ith phylum. This index simultaneously assesses both richness (corresponding to the number of distinct taxa) and relative abundance of the microbial communities within each sample. In particular, it enables effective differentiation between the microbial communities shaped by the dominant taxa and the communities with many taxa with even abundances.
Intestinal permeability biomarkers assayed via ELISA. Several IP biomarkers including Zonulin, LBP (LPS binding protein), Claudin-3 and Intestinal-Fatty Acid Binding Protein, were assayed by a number of specific ELISA kits with high sensitivity and specificity to measure concentrations as low as ng/ml in human blood samples, analogously to previous reports that included studies of military service members and Veterans [25,26,27]. Plasma levels of Zonulin, LBP and Claudin-3 were quantified using ELISA kits from MyBiosorce (catalog # MBS706368, MBS2024051 and MBS2023694 respectively). It should be noted that, since commercial ELISA kits such as Immundiagnostik Kit (Bensheim, Germany) are found to recognize off-target molecules like properdin instead of Zonulin [28], in the present study to avoid such inaccurate measurements, we chose Zonulin ELISA MBS706368 kit (MyBioSource, San Diego, USA) as well as applied internal positive controls with a series of standards for measurement of Zonulin levels. The plasma level of I-FABP was measured using the ELISA kit from R&D Systems (catalog # DFBP20). All procedures followed the manufacturer’s protocols. In brief, 100ul of each sample (undiluted plasma samples for Zonulin and Claudin-3 Elisa plates, 5-fold diluted and 500-fold diluted plasma samples for I-FABP and LBP ELISA plates respectively) and a series of standards were added to each well with 2 hours incubation at room temperature (RT), followed by three washes to remove unbound substances, detection antibody was then added to each well and incubated for another 2 hours at RT. Three-time wash was then repeated to remove extra detection antibody. 100ul of HRP conjugate was added to each well, and incubated for 20 minutes at RT. To remove the unbounded HRP conjugate, the wells were washed another three times, followed by the addition of 100ul of substrate solution to each well and incubation for 20 min at RT to react with HRP conjugate. Finally, 50ul or 100ul stop solution was added to each well to stop the HRP reaction.
The Concentrations of target molecules was then calculated based on the optical density of each, well determined by a microplate reader set to 450 nm and 540 nm, and calculated with CurveExpert professional [29,30] to find the best fit curve formula and corresponding concentrations based on the optical density of samples, standards and blank controls. All IP biomarkers were measured in duplicate, and the average concentrations were calculated for subsequent analyses.
Data and statistical analyses. Statistical and data analyses were performed using R 4.3.0 [31]. For each of top 3 most prevalent phyla, t-test was used to investigate the pre- to post-exposure change in its prevalence and whether the change in prevalence is associated with prior history of mTBI or cumulative history of blast exposures. Ordered logistic regression model was used, with alpha diversity of each sample as outcome and timepoint as an ordinal response to test whether alpha diversity changed stepwise over time following exposure to blast. Average concentration of IP biomarkers was used in mixed effect models as repeated measures, with time as predictor, adjusting for age as covariate, and subject-specific random intercepts. To compare amongst the timepoints pre vs. post and pre vs. follow-up, the pairwise comparisons were run using the “emmeans” function in R. Significance levels were adjusted for multiple testing by applying a Benjamini-Hochberg adjustment [32]. In exploratory analyses, we calculated and tested the significance of correlation of IP biomarker change from pre to post and follow-up with the cumulative exposure measured in cumulative impulse (psi*ms). To Investigate association between elevated-reporting symptoms with measures of alpha-diversity and IP biomarkers, for each symptom we assigned participants to either of two groups based on changes in their self-reported questionnaire (i.e., increasing vs. non-increasing by comparing the symptom scales reported at pre vs. post per subject). Then the increasing vs. the non-increasing groups by symptom were compared for differences in alpha-diversity and levels of IP biomarkers (separately) across timepoints (specifically post-pre). For these comparisons, given small sample sizes, we computed the effect sizes measured by Cohen’s d, rather than performing significance testing.
RESULTS
Microbial variability in blood samples was collected from 30 military service members participating in breaching tactical and training exercises (Figure 1). Following removal of blood RNA sequencing reads that mapped to the human transcriptome, candidate microbial sequences were identified, and relative prevalence of the top 3 microbial phyla found in blood throughout breacher training were characterized (Figure 2A), showing no significant differences in prevalence of these microbial phyla following exposures to blast (p=0.5933, 0.8177, 0.7190 for Proteobacteria, Actinobacteria and Firmicutes, respectively). Microbial diversity (measured by alpha diversity) on phylum level were assessed by performing ordered logistic regression analysis showing significant stepwise increases in alpha diversity following exposure to blast (p=0.049, Figure 2B). For a subset of participants (n=24), changes in intestinal permeability biomarkers (Zonulin, LBP, Claudin-3, I-FABP) were assessed during training. Using linear mixed effect models, accounting for age as a covariate with random intercepts for each subject, significant differences in Zonulin, LBP, Claudin-3 and I-FABP levels were observed across the three timepoints (Figure 3) and pairwise post-hoc analysis, i.e., post vs. pre and follow-up vs. pre analyses showed significant elevation in levels of these biomarkers following exposure to blast. (Table 1).
We found both pre-post and pre-follow-up change in LBP levels showed significant correlations with cumulative blast exposure (r= .4923 and .7564, p=.0398 and .0004 respectively, Figure 4 and Table S1). Exploratory investigations involving blast-related symptoms in association with measures of alpha-diversity and IP biomarkers were also performed. Examination of symptom data collected serially concomitant with blood collection sampling during the training, determined elevated reporting of symptoms (more than 10 subjects reporting at post timepoint) related to headache, concentration, dizziness, and slowed thinking acutely following blast (Figure 5) that dissipated 16hr following. For each symptom, participants were assigned to either of two groups based on changes in their self-reported symptoms (i.e., increasing vs. non-increasing) by comparing the symptom scales reported at pre vs. post per subject. Then the increasing vs. the non-increasing groups by symptom were compared for differences in alpha-diversity and levels of IP biomarkers (separately) across timepoints (specifically post-pre), with effect size measured by Cohen’s d (Figure 6, and supplemental Table S2).
DISCUSSION
This study presents data acquired from military Breachers that show evidence of bacterial translocation into circulation, corroborated by an additional line of preliminary data that shows alterations in IP protein biomarkers, 1-to-16 hrs following military occupational training where individuals were exposed to blast. These data also show elevated reporting of symptoms of headache, slowed thinking, dizziness and concentration by participants acutely following blast that dissipated 16 hrs following. These constellations of symptoms are in line with symptoms typically reported by individuals experiencing mTBI [33,34], and degradations in cognitive performance amongst individuals exposed to blast in military operational breaching during training [35]. Symptoms were further shown to be associated with changes in bacterial translocation and IP as measured by alpha diversity and levels of IP biomarkers, suggesting that intestinal permeability may be linked to decreased cognitive performance. To our knowledge this is the first study that shows exposures to blast in a military operational setting contributes to intestinal permeability along with associated cognitive symptoms, establishing the role of the gut-brain axis in blast related sequalae.
This study has several limitations. As an observational human study conducted within military operational settings, clinician-administered assessments were not performed since it was not feasible and would have interfered with the operational duty of the participants. Also, as a human study, inclusion of a control group is challenging. Since the protocol requires multiple days of military training exercises, typically there are no available service members with comparable military occupational specialties in handling explosives and heavy weapons that could serve as controls. Hence this study is based on a longitudinal design with repeated measures, where each participant’s pre-exposure is their own control when comparing symptom and biomarker measures pre vs. post-and-follow-up exposures to blast. Further, there was a lack of representation of both sexes in the study participants. Although the training protocol was open to both sexes, there were no females in this training cohort. As an observational study, data is collected where blast exposure occurs in existing operational training environments, and presently represents the bias in predominantly male service members that participate in these training protocols. There were also no longer-term longitudinal data on both bacterial species and IP measures following blast exposures, which were not feasible given the scope of the parent protocol under which the symptom and biospecimen data were collected, as well as the participants’ duties while in military service. In terms of biomarker detection, it is possible that the experimental approaches were not sensitive enough to detect subtle changes in IP biomarkers that could be investigated in future studies using other approaches as digital ELISA (or single molecule enzyme-linked immunosorbent assay).
Military service members frequently operate in extreme environments that challenge their health, cognition, and physical function. There is a growing recognition of the role of microbiota dysbiosis and deleterious health outcomes related to military operational exposures (e.g., psychological stress, sleep deprivation, environmental extremes (high altitude, heat, and cold), noise, diet (reviewed here [36])), and, notably, exposures to blast supported by the present study. These exposures can induce central stress responses that lead to altered gastrointestinal and immune function that potentially elicit unfavorable changes in gut microbiota composition, function, and metabolic activity, resulting in dysbiosis that further compromises gastrointestinal function and mucosal barrier, and facilitates translocation of gut microbes into circulation. The gastrointestinal tract tightly controls antigen trafficking by providing a dynamic barrier through both the transcellular and paracellular pathways [37]. Intercellular tight junctions are the key structures regulating paracellular trafficking of macromolecules. Zonulin is a known physiological modulator of intercellular tight junctions synthesized within intestinal and liver cells [38] that modulates the permeability of tight junctions between cells of the wall of the digestive tract. Zonulin has been used as an IP biomarker for autoimmune diseases such as inflammatory bowel disease (Crohn’s disease [39]), as well as multiple sclerosis [40] and Schizophrenia [41]. In the present study, plasma level of Zonulin was significantly increased at 1hr and 16hr post blast exposure, indicating blast may contribute to impairment of the gut barrier in the paracellular pathway (Figure 3 and Table 1), which was further supported by elevated bacteria and alpha diversity in the blood transcriptome (Figure 2B and Table 1).
To additionally capture the potentially complex effect of blast on gut permeability, we assayed levels of other molecules produced by the intestinal epithelial cells – such as fatty acid-binding proteins [42] (FABPs), lipopolysaccharide binding protein (LBP), and Claudin-3. Fatty acid binding protein (FABP) is one of the intracellular proteins, with a low molecular weight of approximately 15 kDa [43], that plays important roles in the transport and metabolism of long-chain fatty acids [44,45]. FABP family of proteins and specifically intestinal-FABP (iFABP) is found primarily in the enterocytes of the jejunum and in the colon and is rapidly released into the circulation following intestinal mucosal tissue injury. Significant elevation of circulating levels of plasma I-FABP were detected 16hrs following exposures to blast (Figure 3 and Table 1). It is possible that the cumulating I-FABP levels in circulation are not acutely detectable with the experimental assays used, with no observed differences pre vs. 1hr post blast exposure (Table 1). Circulating Claudin-3 IP biomarker levels have also been used for detection of intestinal tight junction damage, paracellular localization and expression throughout the jejunum, ileum and colon [46]. Knockout studies of Claudin genes result in loss of tight junction barrier function, with dysregulation of Claudin-3 gene reported in IBD (both Crohn’s disease [47,48] and ulcerative colitis [49,50]) and celiac disease [51]. Although increased circulating Claudin-3 was reported to be indicative of impaired gut barrier, data from the present study showed decreased levels of circulating Claudin-3 levels acutely 1hr post blast that returned to baseline prior to blast exposure within 16hrs (Figure 3, and Table 1). It is possible that as an IP biomarker, circulating Claudin-3 expression might be reflective of chronic inflammatory conditions, rather than acute injury associated with exposures to blast. LBP is synthesized in hepatocytes and released into the bloodstream after glycosylation [52], and is commonly used as a biomarker for the lipopolysaccharide (LPS) response following gut leakage and infection [53,54]. LBP thereby has been shown to serve as a gut permeability biomarker for several inflammatory disorders including celiac disease [55], systemic inflammatory response syndrome [56], heart failure [57] and also obesity [58]. As the primary binding protein for LPS in circulation, LBP enhances the proinflammatory response and clearance of LPS [8]. Findings from the present study showed that plasma LBP expression correlated with cumulative magnitude of blast exposure (Figure 4). LBP levels decreased within 2 hours following blast exposure (Figure 3 and Table 1), suggesting possible elevations of inflammation post blast in response to bacterial translocation due to leaky gut and blast exposure. Indeed, inflammatory response to blast exposure is well-documented [59,60,61].
Immunological as well as endocrine, metabolic, and neural pathways are critically involved in the bidirectional communication pathways between gut microbiota and CNS, referred to as the “microbiota-gut-brain” axis. Broadly, the microbiota-gut-brain axis has been implicated in the pathogenesis of neurological disorders (e.g., Alzheimer’s and Parkinson’s), neuropsychiatric disorders (e.g., PTSD and depression), autoimmune disease (e.g., multiple sclerosis), CNS injuries (e.g., stroke, TBI and spinal cord injury) [62,63], and headaches [64,65,66,67], with some of these disorders having greater prevalence in our warfighters and Veterans that may be attributable to their military experience [36]. Notably, combat-deployed service members with concussive blast injury and followed longitudinally show reductions in fractional anisotropy through imaging studies, which is indicative of chronic brain injury [68]. This suggests that injuries from subconcussive blast exposures may contribute to evolving brain injury pathology that manifests in a range of clinical symptoms [68,69] including chronic headaches [70,71,72] and cognitive impairment [68,69,73]. These are also reflective of symptoms of headache and cognitive problems reported acutely by military service members exposed to blast in operational settings, and specifically by participants in the present study with similar exposures to blast, which are consistent with concussion symptomology [20,74,75]. The elevated reporting of symptoms by these participants related to headache, slowed thinking, dizziness and concentration acutely following blast that dissipated 12 hr following and are in line with constellations of symptoms typically reported by individuals experiencing mTBI [33,34]. Further, data from present study show that these symptoms are associated with possible bacterial translocation from the gut and intestinal permeability measured via changes in alpha-diversity and IP biomarkers, respectively (Figure 6), with translational impact linking the effect of blast on the gut-brain axis with associated clinical symptoms.
In conclusion, findings from this novel course of investigation suggest a possible role of blast exposure in gut permeability and the importance of gut-brain axis in blast injury. These findings have major long-term potential for therapeutic impact in the manner by which we target clinical symptoms associated with exposure to blast and long term sequalae, through adoption of a multi-targeted neuroprotective approach. Instead of prioritizing treatments directed toward single molecules or symptoms, findings from this study support investigations of interventions that modify multiple targets, explicitly by stabilizing the gut microbiota, mucosal barrier, and systemic inflammation. This can be achieved through the deployment of the body’s own immunomodulatory machinery that may help to inform a personalized medicine approach by optimizing the type of probiotic strain, dosage, and timing of delivery of psychobiotic treatments [63]. Specifically, findings from this study may have a widespread clinical impact, leading to a paradigm shift in the manner by which the military can mitigate injuries related to blast exposures through delivery of probiotics for the treatment of microbiota dysbiosis and IP prophylactically [76].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgements
Fatemeh Haghighi, PhD., a recipient of the VA CSR&D Research Career Scientist Award (CX002074) laboratory and work, is supported by CX001728, CX001395, BX003794, and RX003818 at the James J. Peters VA Medical Center. This work is also supported by the VA Rocky Mountain Mental Illness Research Education and Clinical Center. Data collection was conducted under U.S. Army Medical Research and Development Command (USAMRDC) Military Operational Medicine Research Program and Office of the Assistant Secretary of Defense for Health Affairs, BAA Award No. W81XWH-16-2-0001. Walter Carr, PhD and a research team at the Walter Reed Army Institute of Research have been supported through intramural awards from the Military Operational Medicine Research Program at the US Army Medical Research and Development Command and the Defense Health Agency. The authors acknowledge the military service members who accommodated this research activity and made this work possible as well as to their service to this nation every day. The authors also acknowledge CDR Josh L. Duckworth and the CONQUER program (Combat and Training Queryable Exposure/Event Repository) for sharing this mission and for material support in the measurement of blast overpressure exposure.
Conflicts of Interest
The authors declare no conflicts of interest.
Disclaimer
Material has been reviewed by the Walter Reed Army Institute of Research. There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the authors, and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. The investigators have adhered to the policies for protection of human subjects as prescribed in AR 70–25.
References
- Regasa, L.E.; Agimi, Y.; Stout, K.C. Traumatic brain injury following military deployment: evaluation of diagnosis and cause of injury. The Journal of Head Trauma Rehabilitation 2019, 34, 21-29. [CrossRef]
- Zhu, C.S.; Grandhi, R.; Patterson, T.T.; Nicholson, S.E. A review of traumatic brain injury and the gut microbiome: insights into novel mechanisms of secondary brain injury and promising targets for neuroprotection. Brain sciences 2018, 8, 113. [CrossRef]
- Hanscom, M.; Loane, D.J.; Shea-Donohue, T. Brain-gut axis dysfunction in the pathogenesis of traumatic brain injury. Journal of Clinical Investigation 2021, 131, e143777. [CrossRef]
- Tripathi, A.; Lammers, K.M.; Goldblum, S.; Shea-Donohue, T.; Netzel-Arnett, S.; Buzza, M.S.; Antalis, T.M.; Vogel, S.N.; Zhao, A.; Yang, S.; et al. Identification of human zonulin, a physiological modulator of tight junctions, as prehaptoglobin-2. Proceedings of the National Academy of Sciences 2009, 106, 16799-16804, doi:doi:10.1073/pnas.0906773106.
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E. Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease. Lancet 2000, 355, 1518-1519. [CrossRef]
- Wang, W.; Uzzau, S.; Goldblum, S.E.; Fasano, A. Human zonulin, a potential modulator of intestinal tight junctions. Journal of cell science 2000, 113, 4435-4440. [CrossRef]
- Tobias, P.S.; Soldau, K.; Ulevitch, R.J. Isolation of a lipopolysaccharide-binding acute phase reactant from rabbit serum. J Exp Med 1986, 164, 777-793. [CrossRef]
- Schumann, R.R.; Leong, S.R.; Flaggs, G.W.; Gray, P.W.; Wright, S.D.; Mathison, J.C.; Tobias, P.S.; Ulevitch, R.J. Structure and function of lipopolysaccharide binding protein. Science 1990, 249, 1429-1431. [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.d.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis. Frontiers in Microbiology 2021, 12. [CrossRef]
- Ryu, J.-K.; Kim, S.J.; Rah, S.-H.; Kang, J.I.; Jung, H.E.; Lee, D.; Lee, H.K.; Lee, J.-O.; Park, B.S.; Yoon, T.-Y.; Kim, H.M. Reconstruction of LPS Transfer Cascade Reveals Structural Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity 2017, 46, 38-50. [CrossRef]
- Park, B.S.; Lee, J.-O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Experimental & molecular medicine 2013, 45, e66-e66. [CrossRef]
- Findley, M.K.; Koval, M. Regulation and roles for claudin-family tight junction proteins. IUBMB Life 2009, 61, 431-437. [CrossRef]
- Olde Loohuis, L.M.; Mangul, S.; Ori, A.P.; Jospin, G.; Koslicki, D.; Yang, H.T.; Wu, T.; Boks, M.P.; Lomen-Hoerth, C.; Wiedau-Pazos, M. Transcriptome analysis in whole blood reveals increased microbial diversity in schizophrenia. Translational psychiatry 2018, 8, 1-9. [CrossRef]
- Maheshwari, P.; Eslick, G.D. Bacterial infection and Alzheimer’s disease: a meta-analysis. Journal of Alzheimer’s Disease 2015, 43, 957-966. [CrossRef]
- Biometrics, B. Blast Gauge®.
- Cicerone, K.D.; Kalmar, K. Persistent postconcussion syndrome: The structure of subjective complaints after mild traumatic brain injury. The Journal of head trauma rehabilitation 1995, 10, 1-17. [CrossRef]
- King, N.S.; Crawford, S.; Wenden, F.J.; Moss, N.E.G.; Wade, D.T. The Rivermead Post Concussion Symptoms Questionnaire: a measure of symptoms commonly experienced after head injury and its reliability. Journal of Neurology 1995, 242, 587-592. [CrossRef]
- Aguet, F.; Brown, A.A.; Castel, S.E.; Davis, J.R.; He, Y.; Jo, B.; Mohammadi, P.; Park, Y.; Parsana, P.; Segrè, A.V.; et al. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204-213. [CrossRef]
- Carr, W.; Polejaeva, E.; Grome, A.; Crandall, B.; LaValle, C.; Eonta, S.E.; Young, L.A. Relation of Repeated Low-Level Blast Exposure With Symptomology Similar to Concussion. The Journal of Head Trauma Rehabilitation 2015, 30. [CrossRef]
- Vartanian, O.; Rhind, S.G.; Nakashima, A.; Tenn, C.; Lam, T.K.; Shiu, M.; Caddy, N.; King, K.; Natale, A.; Jetly, R. Blast effects on post-concussive and mental health outcomes: Data from Canadian Armed Forces breachers and snipers. Journal of Military, Veteran and Family Health 2022, 8, 82-96. [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15-21. [CrossRef]
- Lab, H. FASTX-Toolkit. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 20 March 2024).
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863-864. [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome biology 2019, 20, 1-13. [CrossRef]
- Weber, N.S.; Gressitt, K.L.; Cowan, D.N.; Niebuhr, D.W.; Yolken, R.H.; Severance, E.G. Monocyte activation detected prior to a diagnosis of schizophrenia in the US Military New Onset Psychosis Project (MNOPP). Schizophr Res 2018, 197, 465-469. [CrossRef]
- Giron, L.B.; Dweep, H.; Yin, X.; Wang, H.; Damra, M.; Goldman, A.R.; Gorman, N.; Palmer, C.S.; Tang, H.-Y.; Shaikh, M.W.; et al. Plasma Markers of Disrupted Gut Permeability in Severe COVID-19 Patients. Front Immunol 2021, 12. [CrossRef]
- McKenna, Z.; Houck, J.; Ducharme, J.; Li, Z.; Berkemeier, Q.; Fennel, Z.; Wells, A.; Mermier, C.; Deyhle, M.; Laitano, O. The effect of prolonged interval and continuous exercise in the heat on circulatory markers of intestinal barrier integrity. European Journal of Applied Physiology 2022, 122, 2651-2659. [CrossRef]
- Scheffler, L.; Crane, A.; Heyne, H.; Tönjes, A.; Schleinitz, D.; Ihling, C.H.; Stumvoll, M.; Freire, R.; Fiorentino, M.; Fasano, A. Widely used commercial ELISA does not detect precursor of haptoglobin2, but recognizes properdin as a potential second member of the zonulin family. Frontiers in endocrinology 2018, 9, 22. [CrossRef]
- Xu, J.; Tanaka, H.; Shoyama, Y. One-step immunochromatographic separation and ELISA quantification of glycyrrhizin from traditional Chinese medicines. Journal of Chromatography B 2007, 850, 53-58. [CrossRef]
- Altamura, S.; Kopf, S.; Schmidt, J.; Müdder, K.; da Silva, A.R.; Nawroth, P.; Muckenthaler, M.U. Uncoupled iron homeostasis in type 2 diabetes mellitus. Journal of Molecular Medicine 2017, 95, 1387-1398. [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www. R-project.org/ 2013.
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal statistical society: series B (Methodological) 1995, 57, 289-300. [CrossRef]
- McMahon, P.J.; Hricik, A.; Yue, J.K.; Puccio, A.M.; Inoue, T.; Lingsma, H.F.; Beers, S.R.; Gordon, W.A.; Valadka, A.B.; Manley, G.T. Symptomatology and functional outcome in mild traumatic brain injury: results from the prospective TRACK-TBI study. Journal of neurotrauma 2014, 31, 26-33. [CrossRef]
- McInnes, K.; Friesen, C.L.; MacKenzie, D.E.; Westwood, D.A.; Boe, S.G. Mild Traumatic Brain Injury (mTBI) and chronic cognitive impairment: A scoping review. PloS one 2017, 12, e0174847. [CrossRef]
- LaValle, C.R.; Carr, W.S.; Egnoto, M.J.; Misistia, A.C.; Salib, J.E.; Ramos, A.N.; Kamimori, G.H. Neurocognitive performance deficits related to immediate and acute blast overpressure exposure. Frontiers in neurology 2019, 10, 949. [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of psychological, environmental and physical stressors on the gut microbiota. Frontiers in microbiology 2018, 2013. [CrossRef]
- Fasano, A. Intestinal permeability and its regulation by zonulin: diagnostic and therapeutic implications. Clinical Gastroenterology and Hepatology 2012, 10, 1096-1100. [CrossRef]
- Tripathi, A.; Lammers, K.M.; Goldblum, S.; Shea-Donohue, T.; Netzel-Arnett, S.; Buzza, M.S.; Antalis, T.M.; Vogel, S.N.; Zhao, A.; Yang, S. Identification of human zonulin, a physiological modulator of tight junctions, as prehaptoglobin-2. Proceedings of the National Academy of Sciences 2009, 106, 16799-16804. [CrossRef]
- Caviglia, G.P.; Dughera, F.; Ribaldone, D.G.; Rosso, C.; Abate, M.L.; Pellicano, R.; Bresso, F.; Smedile, A.; Saracco, G.M.; Astegiano, M. Serum zonulin in patients with inflammatory bowel disease: a pilot study. Minerva Medica 2019, 110, 95-100. [CrossRef]
- Camara-Lemarroy, C.R.; Silva, C.; Greenfield, J.; Liu, W.-Q.; Metz, L.M.; Yong, V.W. Biomarkers of intestinal barrier function in multiple sclerosis are associated with disease activity. Multiple Sclerosis Journal 2020, 26, 1340-1350. [CrossRef]
- Gokulakrishnan, K.; Nikhil, J.; Vs, S.; Holla, B.; Thirumoorthy, C.; Sandhya, N.; Nichenametla, S.; Pathak, H.; Shivakumar, V.; Debnath, M. Altered Intestinal Permeability Biomarkers in Schizophrenia: A Possible Link with Subclinical Inflammation. Annals of Neurosciences 2022, 29, 151-158. [CrossRef]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; De Vos, W.M. Homeostasis of the gut barrier and potential biomarkers. American Journal of Physiology-Gastrointestinal and Liver Physiology 2017, 312, G171-G193. [CrossRef]
- Coe, N.R.; Bernlohr, D.A. Physiological properties and functions of intracellular fatty acid-binding proteins. Biochimica et biophysica acta 1998, 1391, 287-306. [CrossRef]
- Zimmerman, A.; Veerkamp, J. New insights into the structure and function of fatty acid-binding proteins. Cellular and Molecular Life Sciences CMLS 2002, 59, 1096-1116. [CrossRef]
- Chmurzyńska, A. The multigene family of fatty acid-binding proteins (FABPs): function, structure and polymorphism. Journal of applied genetics 2006, 47, 39-48. [CrossRef]
- Grootjans, J.; Thuijls, G.; Verdam, F.; Derikx, J.P.; Lenaerts, K.; Buurman, W.A. Non-invasive assessment of barrier integrity and function of the human gut. World Journal of Gastrointestinal Surgery 2010, 2, 61. [CrossRef]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61-72. [CrossRef]
- Goswami, P.; Das, P.; Verma, A.K.; Prakash, S.; Das, T.; Nag, T.; Ahuja, V.; Gupta, S.D.; Makharia, G.K. Are alterations of tight junctions at molecular and ultrastructural level different in duodenal biopsies of patients with celiac disease and Crohn’s disease? Virchows Archiv 2014, 465, 521-530. [CrossRef]
- Prasad, S.; Mingrino, R.; Kaukinen, K.; Hayes, K.L.; Powell, R.M.; MacDonald, T.T.; Collins, J.E. Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells. Laboratory investigation 2005, 85, 1139-1162. [CrossRef]
- Thuijls, G.; Derikx, J.P.; de Haan, J.-J.; Grootjans, J.; de Bruïne, A.; Masclee, A.A.; Heineman, E.; Buurman, W.A. Urine-based detection of intestinal tight junction loss. Journal of clinical gastroenterology 2010, 44, e14-e19. [CrossRef]
- Lu, Z.; Ding, L.; Lu, Q.; Chen, Y.-H. Claudins in intestines: Distribution and functional significance in health and diseases. Tissue barriers 2013, 1, e24978. [CrossRef]
- Zweigner, J.; Schumann, R.R.; Weber, J.R. The role of lipopolysaccharide-binding protein in modulating the innate immune response. Microbes and Infection 2006, 8, 946-952. [CrossRef]
- Jensen, S.B.; Latysheva, N.; Hindberg, K.; Ueland, T. Plasma lipopolysaccharide-binding protein is a biomarker for future venous thromboembolism: Results from discovery and validation studies. Journal of Internal Medicine 2022, 292, 523-535. [CrossRef]
- Pugin, J.M.; Schürer-Maly, C.; Leturcq, D.; Moriarty, A.; Ulevitch, R.J.; Tobias, P.S. Lipopolysaccharide activation of human endothelial and epithelial cells is mediated by lipopolysaccharide-binding protein and soluble CD14. Proceedings of the National Academy of Sciences 1993, 90, 2744-2748. [CrossRef]
- Ferreira, S.; Masi, J.; Giménez, V.; Carpinelli, M.-M.; Laterza, O.; Hermoso, M.; Ortiz-Villalba, J.; Chamorro, M.-E.; Langjahr, P. Effect of gluten-free diet on levels of soluble CD14 and lipopolysaccharide-binding protein in adult patients with celiac disease. Central European Journal of Immunology 2021, 46, 225-230. [CrossRef]
- Myc, A.; Buck, J.; Gonin, J.; Reynolds, B.; Hammerling, U.; Emanuel, D. The level of lipopolysaccharide-binding protein is significantly increased in plasma in patients with the systemic inflammatory response syndrome. Clinical Diagnostic Laboratory Immunology 1997, 4, 113-116. [CrossRef]
- Trøseid, M.; Andersen, G.Ø.; Broch, K.; Hov, J.R. The gut microbiome in coronary artery disease and heart failure: Current knowledge and future directions. EBioMedicine 2020, 52. [CrossRef]
- Gonzalez-Quintela, A.; Alonso, M.; Campos, J.; Vizcaino, L.; Loidi, L.; Gude, F. Determinants of serum concentrations of lipopolysaccharide-binding protein (LBP) in the adult population: the role of obesity. PloS one 2013, 8, e54600. [CrossRef]
- Wang, Z.; Wilson, C.M.; Mendelev, N.; Ge, Y.; Galfalvy, H.; Elder, G.; Ahlers, S.; Yarnell, A.M.; LoPresti, M.L.; Kamimori, G.H.J.J.o.N. Acute and chronic molecular signatures and associated symptoms of blast exposure in military breachers. 2019. [CrossRef]
- Gill, J.; Motamedi, V.; Osier, N.; Dell, K.; Arcurio, L.; Carr, W.; Walker, P.; Ahlers, S.; LoPresti, M.; Yarnell, A. Moderate blast exposure results in increased IL-6 and TNFα in peripheral blood. Brain, behavior, and immunity 2017, 65, 90-94. [CrossRef]
- Edwards, K.A.; Leete, J.J.; Smith, E.G.; Quick, A.; Modica, C.M.; Wassermann, E.M.; Polejaeva, E.; Dell, K.C.; LoPresti, M.; Walker, P. Elevations in tumor necrosis factor alpha and interleukin 6 from neuronal-derived extracellular vesicles in repeated low-level blast exposed personnel. Frontiers in Neurology 2022, 13. [CrossRef]
- Malan-Muller, S.; Valles-Colomer, M.; Foxx, C.L.; Vieira-Silva, S.; van den Heuvel, L.L.; Raes, J.; Seedat, S.; Lowry, C.A.; Hemmings, S.M. Exploring the relationship between the gut microbiome and mental health outcomes in a posttraumatic stress disorder cohort relative to trauma-exposed controls. European Neuropsychopharmacology 2022, 56, 24-38. [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V. The microbiota-gut-brain axis. Physiological reviews 2019. [CrossRef]
- Arzani, M.; Jahromi, S.R.; Ghorbani, Z.; Vahabizad, F.; Martelletti, P.; Ghaemi, A.; Sacco, S.; Togha, M.; Federation, S.o.A.S.o.t.E.H. Gut-brain axis and migraine headache: a comprehensive review. The journal of headache and pain 2020, 21, 1-12. [CrossRef]
- Aamodt, A.; Stovner, L.; Hagen, K.; Zwart, J. Comorbidity of headache and gastrointestinal complaints. The Head-HUNT Study. Cephalalgia 2008, 28, 144-151.
- Crawford, J.; Liu, S.; Tao, F. Gut microbiota and migraine. Neurobiology of Pain 2022, 11, 100090. [CrossRef]
- Sgro, M.; Ray, J.; Foster, E.; Mychasiuk, R. Making Migraine Easier to Stomach: The Role of the Gut Brain Immune Axis in Headache Disorders. European Journal of Neurology 2023. [CrossRef]
- Mac Donald, C.L.; Barber, J.; Andre, J.; Evans, N.; Panks, C.; Sun, S.; Zalewski, K.; Sanders, R.E.; Temkin, N. 5-Year imaging sequelae of concussive blast injury and relation to early clinical outcome. Neuroimage: clinical 2017, 14, 371-378. [CrossRef]
- Mac Donald, C.L.; Barber, J.; Patterson, J.; Johnson, A.M.; Dikmen, S.; Fann, J.R.; Temkin, N. Association between 5-year clinical outcome in patients with nonmedically evacuated mild blast traumatic brain injury and clinical measures collected within 7 days postinjury in combat. JAMA network open 2019, 2, e186676-e186676. [CrossRef]
- Hoffman, J.M.; Lucas, S.; Dikmen, S.; Braden, C.A.; Brown, A.W.; Brunner, R.; Diaz-Arrastia, R.; Walker, W.C.; Watanabe, T.K.; Bell, K.R. Natural history of headache after traumatic brain injury. Journal of neurotrauma 2011, 28, 1719-1725. [CrossRef]
- Walker, W.C.; Marwitz, J.H.; Wilk, A.R.; Ketchum, J.M.; Hoffman, J.M.; Brown, A.W.; Lucas, S. Prediction of headache severity (density and functional impact) after traumatic brain injury: A longitudinal multicenter study. Cephalalgia 2013, 33, 998-1008. [CrossRef]
- Stacey, A.; Lucas, S.; Dikmen, S.; Temkin, N.; Bell, K.R.; Brown, A.; Brunner, R.; Diaz-Arrastia, R.; Watanabe, T.K.; Weintraub, A. Natural history of headache five years after traumatic brain injury. Journal of neurotrauma 2017, 34, 1558-1564. [CrossRef]
- Mac Donald, C.L.; Barber, J.; Jordan, M.; Johnson, A.M.; Dikmen, S.; Fann, J.R.; Temkin, N. Early clinical predictors of 5-year outcome after concussive blast traumatic brain injury. JAMA neurology 2017, 74, 821-829. [CrossRef]
- Carr, W.; Dell, K.; Yanagi, M.; Hassan, D.; LoPresti, M. Perspectives on repeated low-level blast and the measurement of neurotrauma in humans as an occupational exposure risk. Shock waves 2017, 27, 829-836. [CrossRef]
- Caplan, B.; Bogner, J.; Brenner, L.; Carr, W.; Polejaeva, E.; Grome, A.; Crandall, B.; LaValle, C.; Eonta, S.E.; Young, L.A. Relation of repeated low-level blast exposure with symptomology similar to concussion. Journal of Head Trauma Rehabilitation 2015, 30, 47-55. [CrossRef]
- Brenner, L.A.; Forster, J.E.; Stearns-Yoder, K.A.; Stamper, C.E.; Hoisington, A.J.; Brostow, D.P.; Mealer, M.; Wortzel, H.S.; Postolache, T.T.; Lowry, C.A. Evaluation of an immunomodulatory probiotic intervention for veterans with co-occurring mild traumatic brain injury and posttraumatic stress disorder: a pilot study. Frontiers in neurology 2020, 11, 1015. [CrossRef]
Figure 1.
Number of self-reported career breaching experience at baseline prior to start of training exercises including 30 participants with/out history of mTBI (denoted in red and blue, respectively).
Figure 1.
Number of self-reported career breaching experience at baseline prior to start of training exercises including 30 participants with/out history of mTBI (denoted in red and blue, respectively).
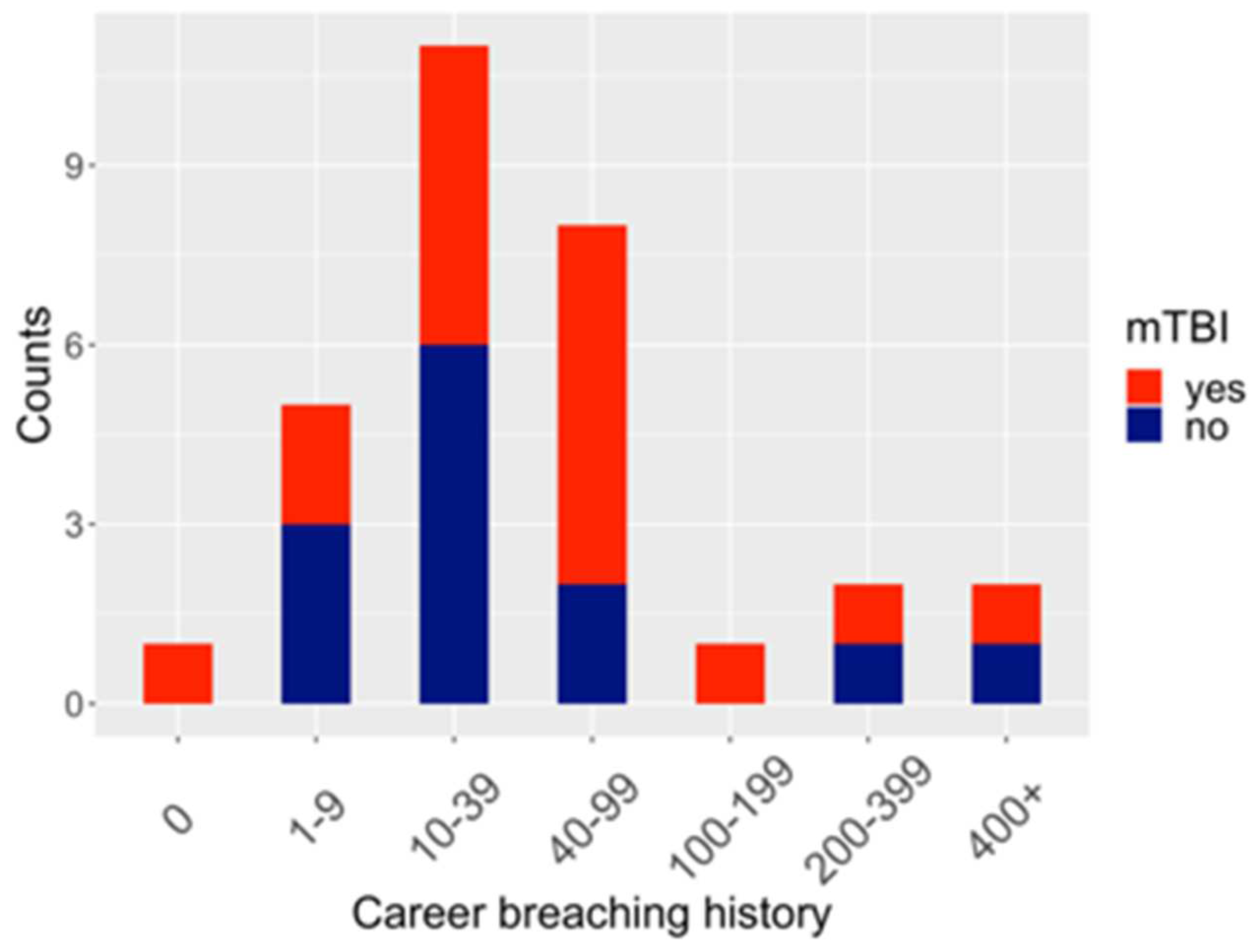
Figure 2.
(a) Distribution of relative abundances of microbiota at phylum level at three time points i.e., pre-blast, post-blast and follow-up. Phylogenetic classification was performed to assign filtered candidate microbial reads to the microbial genes from 33 distinct taxa on the phylum level. Top three phyla, which include Proteobacteria, Firmicutes and Actinobacteria, are shown in different colors depicting their proportions of RNA-seq reads mapped. From the vertex arranged clockwise are samples from 30 participants corresponding to pre-, post-training and follow-up (1hr and 16hr post blast respectively). Each participant is assigned a sequential ID number starting with ID number from 01 to 37 and ending with either of the letters A, B, or Z, corresponding to the pre-, post- and follow up time points respectively. All “other” low abundant bacteria are grouped together. (b) Boxplot shows distribution of alpha diversity at phylum-level at each timepoint using time series blood RNA-seq data i.e., pre-post and follow-up blast timepoints (i.e., 1hr, and 16hr post blast respectively). Significant (p=0.049) stepwise increase in alpha diversity is observed following exposure to blast. Alpha diversity is derived using the inverse Simpson index, that simultaneously assesses both richness (corresponding to the number of distinct taxa) and relative abundance of the microbial communities within each sample.
Figure 2.
(a) Distribution of relative abundances of microbiota at phylum level at three time points i.e., pre-blast, post-blast and follow-up. Phylogenetic classification was performed to assign filtered candidate microbial reads to the microbial genes from 33 distinct taxa on the phylum level. Top three phyla, which include Proteobacteria, Firmicutes and Actinobacteria, are shown in different colors depicting their proportions of RNA-seq reads mapped. From the vertex arranged clockwise are samples from 30 participants corresponding to pre-, post-training and follow-up (1hr and 16hr post blast respectively). Each participant is assigned a sequential ID number starting with ID number from 01 to 37 and ending with either of the letters A, B, or Z, corresponding to the pre-, post- and follow up time points respectively. All “other” low abundant bacteria are grouped together. (b) Boxplot shows distribution of alpha diversity at phylum-level at each timepoint using time series blood RNA-seq data i.e., pre-post and follow-up blast timepoints (i.e., 1hr, and 16hr post blast respectively). Significant (p=0.049) stepwise increase in alpha diversity is observed following exposure to blast. Alpha diversity is derived using the inverse Simpson index, that simultaneously assesses both richness (corresponding to the number of distinct taxa) and relative abundance of the microbial communities within each sample.
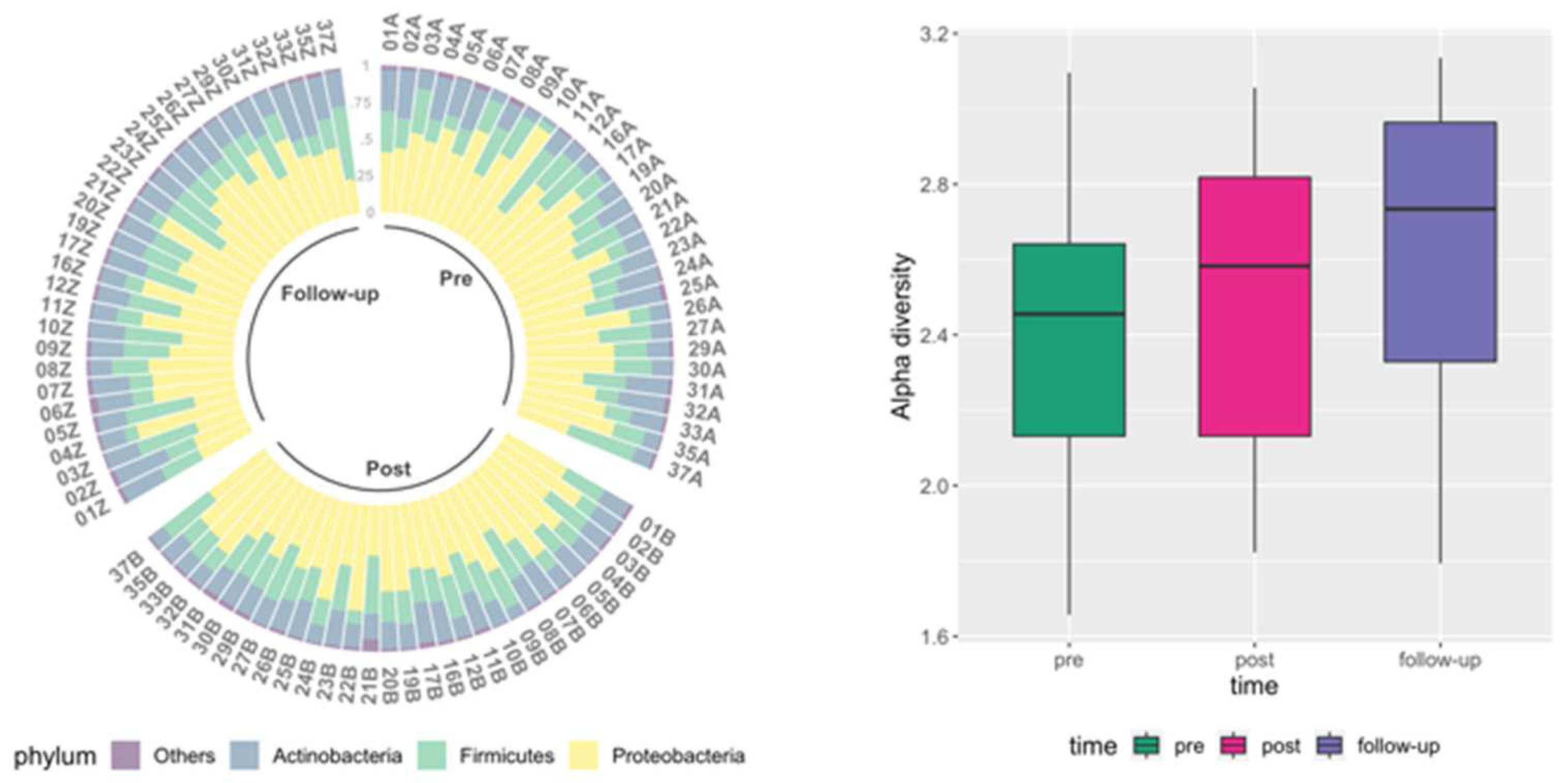
Figure 3.
Boxplots show distribution of IP biomarkers measured at each timepoint. Data are shown for (a) Zonulin, (b) LBP, (c) Claudin-3 and (d) I-FABP. A spaghetti plot connecting each subject’s measures at different timepoints was added to the boxplot as another layer to visualize the observed trends in individual changes in levels of these IP biomarkers.
Figure 3.
Boxplots show distribution of IP biomarkers measured at each timepoint. Data are shown for (a) Zonulin, (b) LBP, (c) Claudin-3 and (d) I-FABP. A spaghetti plot connecting each subject’s measures at different timepoints was added to the boxplot as another layer to visualize the observed trends in individual changes in levels of these IP biomarkers.
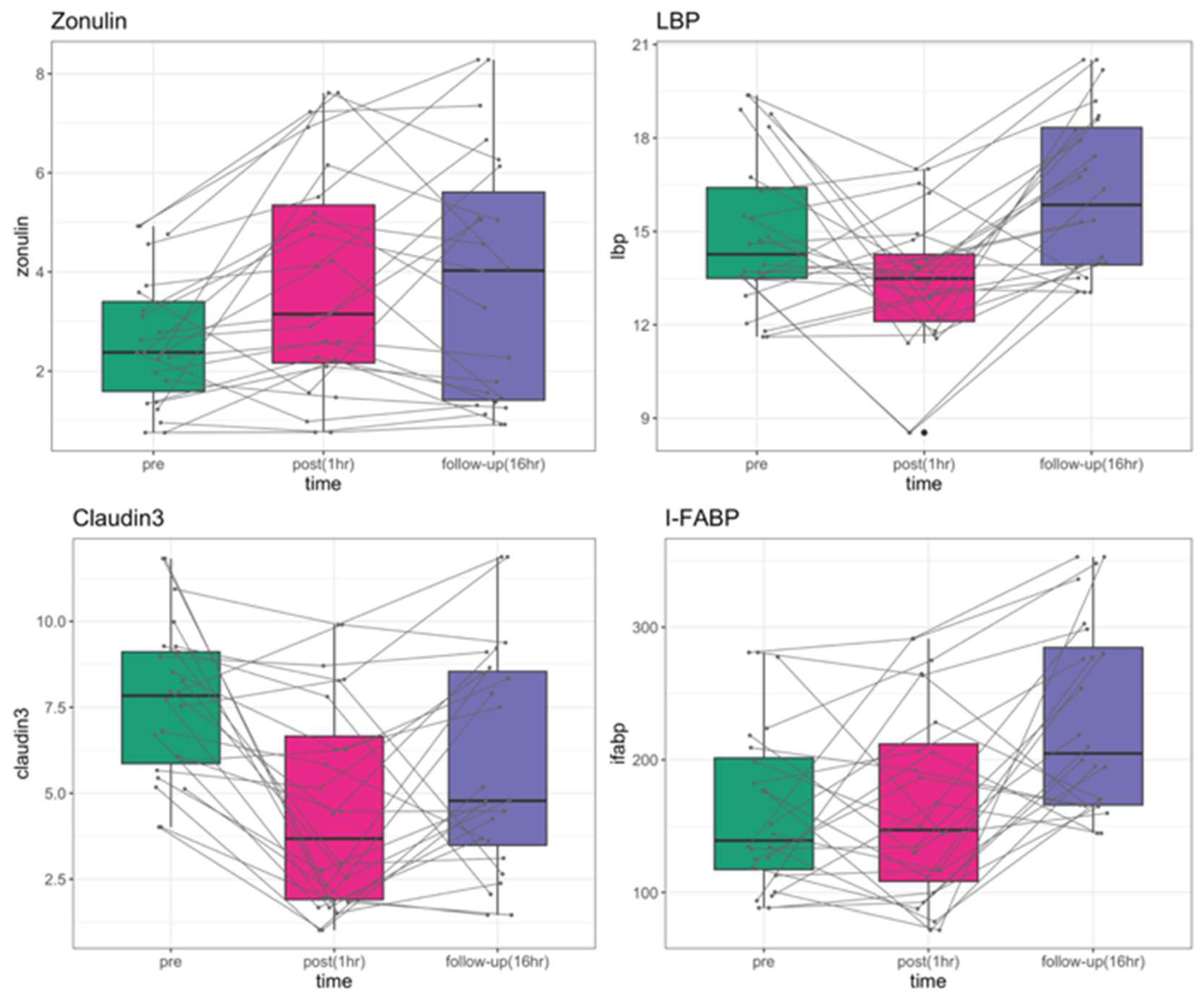
Figure 4.
Correlation of cumulative exposure (measured in psi*ms) showing LBP IP biomarker levels changing from pre- to post- (black) and follow-up (orange). Both pre-post and pre-follow-up change in LBP levels showed significant correlation with cumulative exposure (r= .4923 and .7564, p=.0398 and .0004, respectively).
Figure 4.
Correlation of cumulative exposure (measured in psi*ms) showing LBP IP biomarker levels changing from pre- to post- (black) and follow-up (orange). Both pre-post and pre-follow-up change in LBP levels showed significant correlation with cumulative exposure (r= .4923 and .7564, p=.0398 and .0004, respectively).
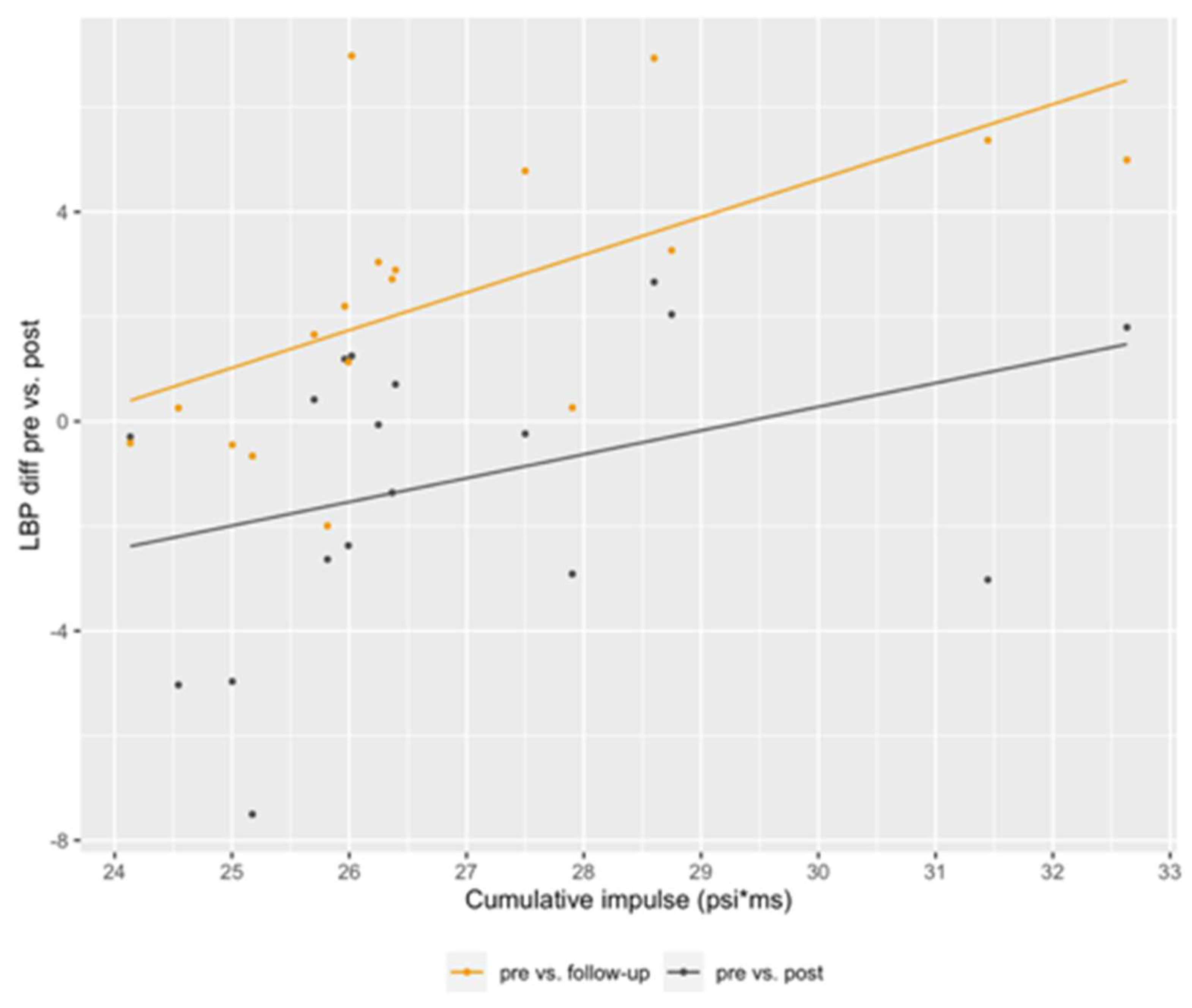
Figure 5.
Symptom response frequencies from self-report surveys at each time point throughout the training (pre-blast, post-blast, and follow-up), showing elevated symptoms related to headache, slowed thinking, concentration, and dizziness, directly following exposure to blast (shown in pink bars).
Figure 5.
Symptom response frequencies from self-report surveys at each time point throughout the training (pre-blast, post-blast, and follow-up), showing elevated symptoms related to headache, slowed thinking, concentration, and dizziness, directly following exposure to blast (shown in pink bars).
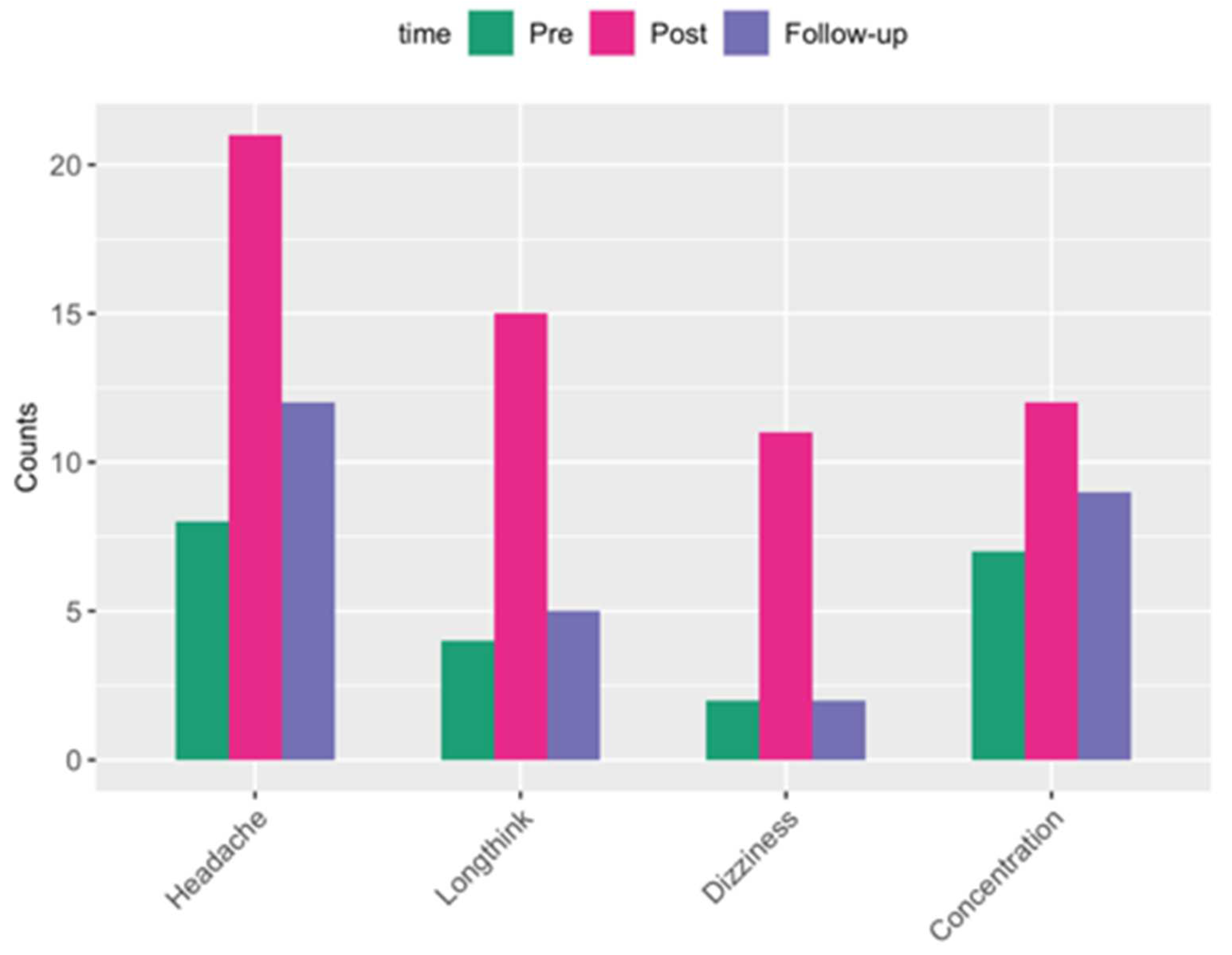
Figure 6.
Association of changes in self-reported symptoms vs. changes in alpha-diversity and IP biomarkers, shown as effect sizes measured by Cohen’s d. For each elevated symptom self-reported, participants were assigned to either of two groups i.e., increasing vs. non-increasing by comparing the symptom scales reported at pre vs. post and pre vs. follow-up per subject. Then the increasing vs. the non-increasing groups by symptom were compared for differences in alpha-diversity and levels of IP biomarkers (separately) across timepoints (specifically post - pre and follow-up - pre). Note, negative value corresponds to lower levels of an IP biomarker or alpha diversity post-blast. Dashed lines are cutoffs between small and medium magnitude effect sizes.
Figure 6.
Association of changes in self-reported symptoms vs. changes in alpha-diversity and IP biomarkers, shown as effect sizes measured by Cohen’s d. For each elevated symptom self-reported, participants were assigned to either of two groups i.e., increasing vs. non-increasing by comparing the symptom scales reported at pre vs. post and pre vs. follow-up per subject. Then the increasing vs. the non-increasing groups by symptom were compared for differences in alpha-diversity and levels of IP biomarkers (separately) across timepoints (specifically post - pre and follow-up - pre). Note, negative value corresponds to lower levels of an IP biomarker or alpha diversity post-blast. Dashed lines are cutoffs between small and medium magnitude effect sizes.
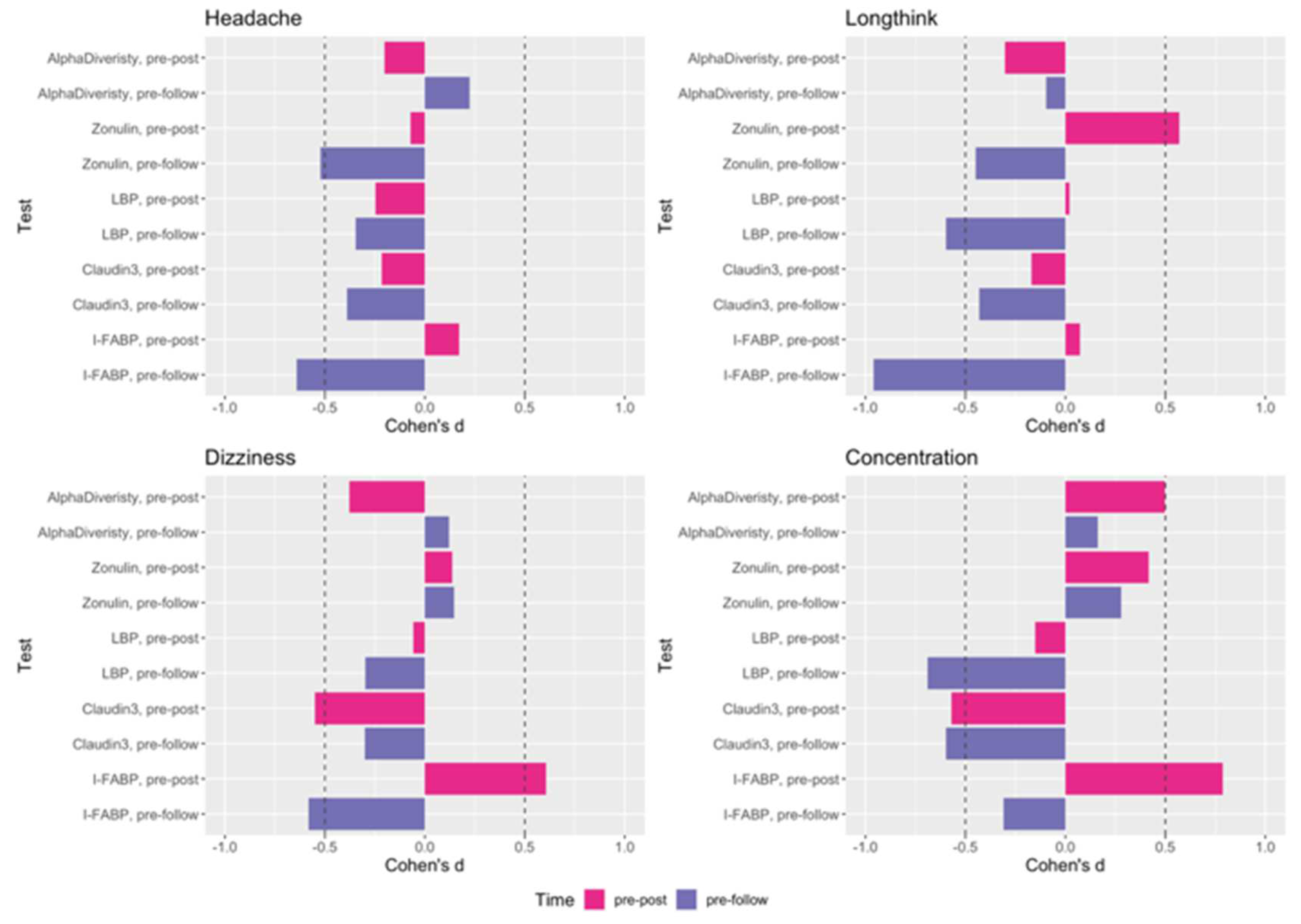

Table 1.
Linear mixed effect models, accounting for age as a covariate with random intercept for each subject, were used to test differences between timepoints in Zonulin, LBP, Claudin-3 and I-FABP levels. P-values of ANOVA and pairwise post-hoc comparisons, i.e., post vs. pre and follow-up vs. pre analyses are reported.
Table 1.
Linear mixed effect models, accounting for age as a covariate with random intercept for each subject, were used to test differences between timepoints in Zonulin, LBP, Claudin-3 and I-FABP levels. P-values of ANOVA and pairwise post-hoc comparisons, i.e., post vs. pre and follow-up vs. pre analyses are reported.
| P-value | Zonulin | LBP | Claudin-3 | I-FABP |
| ANOVA | 0.0018 | 0.0002 | 6.24E-05 | 8.34E-05 |
| pre vs. post | 0.0029 | 0.0222 | 3.8E-05 | 0.7870 |
| pre vs. follow-up | 0.0029 | 0.0513 | 0.0115 | 0.0003 |
| post vs. follow-up | 0.9680 | 0.0001 | 0.0412 | 0.0003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



