
Submitted:
24 January 2024
Posted:
25 January 2024
You are already at the latest version
Abstract
Overweight and Obesity are global health and economic concerns. According to the World Health Organization, the number of individuals with obesity has nearly tripled since 1975. This health issue is currently considered an epidemic, affecting over 1.9 billion adults and 340 million children and adolescents. It contributes to more than 5 million deaths each year and has a global economic burden that will reach $4.32 trillion annually by 2035, according to the World Obesity Federation. This disease can affect every system of the human body and can lead to complications such as metabolic syndrome, diabetes, cancer, dyslipidemia, cardiovascular diseases, and hypertension, among others. Treatment may sometimes include diet, exercise, drugs, and bariatric surgery. Nonetheless, not all people have access to these treatments, and public health strategies consider prevention the most important factor. In this regard, recent investigations are aiming to find alternatives and adjuvants for the treatment of obesity, its prevention, and the reversion of some of its complications, using natural sources of anti-obesogenic compounds like polyphenols, terpenes, alkaloids, saponins, among others. In this review, we gather the most current information using PubMed, Google Scholar, Scopus, Cochrane, and Web of Science. We present and discuss the current information about natural products that have shown anti-obesogenic effects at a molecular level. We also consider the impact of dietary habits and lifestyle in preventing overweight and obesity due to the evidence of the benefits of certain foods and compounds consumed regularly. We discuss mechanisms, pathways, and receptors involved in the modulation of obesity, especially those related to inflammation and oxidative stress linked to this disease, due to the relevance of these two aspects in developing complications.
Keywords:
natural products
; polyphenols
; alkaloids
; saponins
; obesity
; overweight
; anti-obesogenic compounds
1. Introduction
Over the past decades, the number of adults aged 18 and older who are overweight and obese has been increasing worldwide. Overweight and obesity are defined as having a body max index (BMI) greater than 25 kg/m2 and 30 kg/m2, respectively [1]. Overweight and obesity are health issues that greatly affect the public health and are linked to socioeconomic factors. Some groups are more affected than others. For example, in the United States, non-Hispanic black adults have the highest prevalence of obesity, followed by Hispanic adults, non-Hispanic white adults, and non-Hispanic Asian adults. Furthermore, the total prevalence of overweight and obesity among adults aged 20-39 is 39.8%. additionally, the burden of overweight and obesity is also influenced by the degree of education; for instance, it has been reported that men and women with college degrees have a lower prevalence of obesity compared with those with less education [2,3]. Other factors that are important in the incidence of weight disorders are genetic, behavioral, environmental, and lifestyle factors. Particularly, sedentarism and low metabolic rate, diet, genes, and metabolism are the main traits of this disease in current times [4].
Being overweight and obese, as public health problems represent a high burden on the public expenditure in health care systems. In this sense, people living with obesity have 30% higher medical costs than normal-weight individuals [5]. Currently, weight disorders have a strong association with non-communicable diseases such as diabetes mellitus, hypertension, the metabolic syndrome, stroke, sleep apnea, gastrointestinal cancers, and inflammatory diseases [6].
Global efforts to reduce the burden of weight disorders are focused on dietary and lifestyle modifications, such as restricted calories intake and increased physical activity are the most recommended. Within the food factors, there are some food ingredients such as phytochemicals that have been associated with weight loss agents or coadjuvants. Polyphenols, alkaloids, and saponins are among the most widely studied phytochemicals in interdisciplinary studies evaluating their antiobesity mechanisms of action. Here, we summarize current studies regarding the potential antiobesity potential of these three phytochemicals.
1.2. Polyphenols, alkaloids and saponins
1.2.1. Polyphenols
Phenolic compounds are a heterogeneous group of secondary metabolites derived from the phenylpropanoid and phenylpropanoid acetate pathways. They have a structure with an aromatic ring that is composed of one or more -OH radicals. Furthermore, they are ubiquitously present in the plant kingdom and their classification is based on the number of carbons in the molecules and can be classified into several categories, including flavonoids, phenolic acids, lignans and stilbenes. These compounds can be commonly found in the human diet including foods such as fruits and vegetables, cereals, herbs and spices and beverages such as wine, tea, and coffee. The concentration and type of polyphenols in different food products depend significantly on the origin, cultivation, environmental conditions, maturity, pre- and post-harvest, storage, and transportation. Furthermore, these compounds have been widely studied for their beneficial health effects, highlighting their antidiabetic, anticancer, and anti-inflammatory effects, among others [7,8,9].
1.2.2. Alkaloids
Alkaloids are natural organic compounds found in a wide variety of plants that contain nitrogen. Their common structure is a heterocyclic ring with one or more nitrogen atoms, and they are classified into several categories, including indole, isoquinoline, pyridine, piperidine, tropane and quinolizidine. Alkaloids are found in a wide variety of foods such as tea, coffee, fruits and vegetables, spices, legumes and nuts, and their distribution in the human diet varies according to geographic region and dietary habits. Furthermore, its bioavailability can be affected by several factors such as the food matrix, the way of processing and the intestinal microbiota, among others [10,11,12].
1.2.3. Saponins
Saponins are a group of chemical compounds found in a wide variety of plants that have a common steroid or triterpenoid ring structure with one or more hydroxyl groups. They can be classified into several categories including steroids and triterpenes. These compounds are found in a wide variety of foods, including legumes, cereals, fruits, vegetables, tea, coffee, cocoa, and spices. These secondary metabolites have been studied for their anti-inflammatory, antioxidant, antitumor, antidiabetic and antiviral properties, among others. Additionally, it has been shown that saponins can improve cardiovascular health, reduce the risk of chronic diseases, and improve cognitive function [13].
1.2.4. Terpenes
Terpenes are a group of secondary metabolites naturally present in plants, derived the mevalonic acid pathway. Terpenes have been also reported in insects and marine organisms. More than 55,000 compounds have been classified as terpenes, with a variety of characteristics, functions, and application. They have been part of folk medicine since ancient times, and for perfume making, cosmetics, flavoring and even for religious reasons. Based on the number of isoprene units present in their structure, they are classified as hemiterpenes, monoterpenes, sesquiterpenes, diterpenes, sesterpenes, triterpenes, tetraterpenes, polyterpenes. They are involved in the plant development, growth and defense against biotic and abiotic stress. Terpenoids also have synergic effects with other compounds and are commonly used as administration vehicles or additives. Most common human health-related effects attributable to terpenes are anticancer, immunomodulatory, antibacterial, antiviral, neuroprotective, antioxidant, anti-inflammatory, among others [14,15,16].
1.2. Relationship between lifestyle, oxidative stress, and inflammation in overweight and obesity
Obesity is a multifactorial chronic disease in which social, behavioral, genetic, psychological, metabolic, cellular, and molecular factors are involved. One of the main pillars for the appearance of obesity is the excessive consumption of macronutrients and low energy consumption by the body. Eating habits along with a low level of physical activity have led to a drastic increase in obesity [1]. In general, a diet high in processed foods, saturated fats, added sugars, and refined carbohydrates has been linked to an increased risk of obesity. On the other hand, a diet rich in fruits, vegetables, lean proteins, and whole grains have been linked to a lower risk of suffering from it. Likewise, research has shown that a diet high in fat and carbohydrates induces a significant increase in oxidative stress and inflammation in people with obesity [17]. The consumption of high-fat diets can generate alterations in oxygen metabolism and increase the production of reactive oxygen species (ROS). If the production of these ROS exceeds the antioxidant capacity of the cells, the oxidative stress resulting from lipid peroxidation could contribute to the development of atherosclerosis, diabetes, cardiovascular diseases, and other complications [18,19].
ROS and reactive nitrogen species (RNS) participate in various biological actions of the body, including defense against pathogenic microorganisms, which is mediated by the immune system and intracellular signaling. However, at elevated levels, they can damage DNA, lipids, and proteins, leading to tissue injury and cell death [20]. To maintain adequate levels of these reactive species, the body has antioxidant compounds and enzymes to reduce their toxicity. It has naturally occurred antioxidant compounds such as glutathione and ubiquinone and some proteins and enzymes that have antioxidant functions. However, in people with obesity these antioxidant compounds are decreased, and the production of free radicals is increased, so this balance is lost [20,21]. The increased production of free radicals in obesity may be due to several factors, such as excess adipose tissue, oxidation of fatty acids, excessive oxygen consumption caused by mechanical and metabolic load, accumulations of damaged cells, the type of diet that is eaten (high-fat diets can alter oxygen metabolism) and one of the most important, the damage to the mitochondria, due to the leak of electrons through the respiration chain and the excess mitochondrial triglycerides, which inhibit the translocation of adenine nucleotides and promote the generation of superoxide [17].
The increase in oxidative stress in obesity causes inflammatory processes in the body, since adipocytes and preadipocytes have been identified as a source of proinflammatory cytokines, such as TNF-α, IL-1 and IL-6; Therefore, obesity is considered a state of chronic low-grade inflammation. These cytokines are potent stimulators to produce reactive oxygen and nitrogen by macrophages and monocytes, so the increase in the concentration of cytokines could increase oxidative stress [17,19,22].
Hyperplasia and hypertrophy of adipocytes in the adipose tissue is an important characteristic in obese patients, this results in the release of adipokines into the bloodstream, such as leptin, which acts directly on macrophages to increase phagocytic activity and the production of proinflammatory cytokines which exert an effect on T cells, monocytes, neutrophils, and endothelial cells. Another important adipokine is TNF-α, which activates nuclear factor κB (NF-κ B), resulting in increased expression of adhesion molecules on the surface of endothelial cells and vascular smooth muscle cells, resulting in an inflammatory state in adipose tissue. Unlike the other adipokines that can be released by adipocytes, adiponectin does not increase its expression, it is reduced, and, since it has regulatory actions on energy homeostasis, glucose and lipid metabolism and anti-inflammatory action, this in a key factor in the development of complications such as insulin resistance, metabolic syndrome, hyperglycemia, and dyslipidemia [23].
2. Action Mechanisms of Phytochemicals against Overweight and Obesity
Nowadays literature provides us with several effects that are considered antiobesogenic, nonetheless, the main targets of anti-obesity compounds are lipase inhibition, regulation of adipogenesis, thermogenesis, and appetite suppression. Some of these effects have been evaluated in a molecular level, especially those related to adipogenesis and/or adipose tissue cells’ functions and homeostasis.
2.1. Lipase Inhibitors
There are various studies focused on the inhibition of lipase enzymes since they are the enzymes with a key function during the process of digestion and absorption of lipids. In this sense, pancreatic lipase is the enzyme secreted by the pancreas, which fulfills the function of hydrolyzing the ester bonds in triglycerides (TGs) to monoglycerides and fatty acids, which when mixed with cholesterol, lysophosphatidic acid, and bile salts in the intestine, form micelles that are absorbed in the enterocytes, where TGs are resynthesized and are stored in mature adipocytes. The increase of TGs in adipocytes is associate related to overweight and obesity [24,25]. Therefore, the inhibition of the activity of the pancreatic lipase enzyme is a key element in reducing the digestion and absorption of lipids.
2.2. Regulation of Adipogenesis
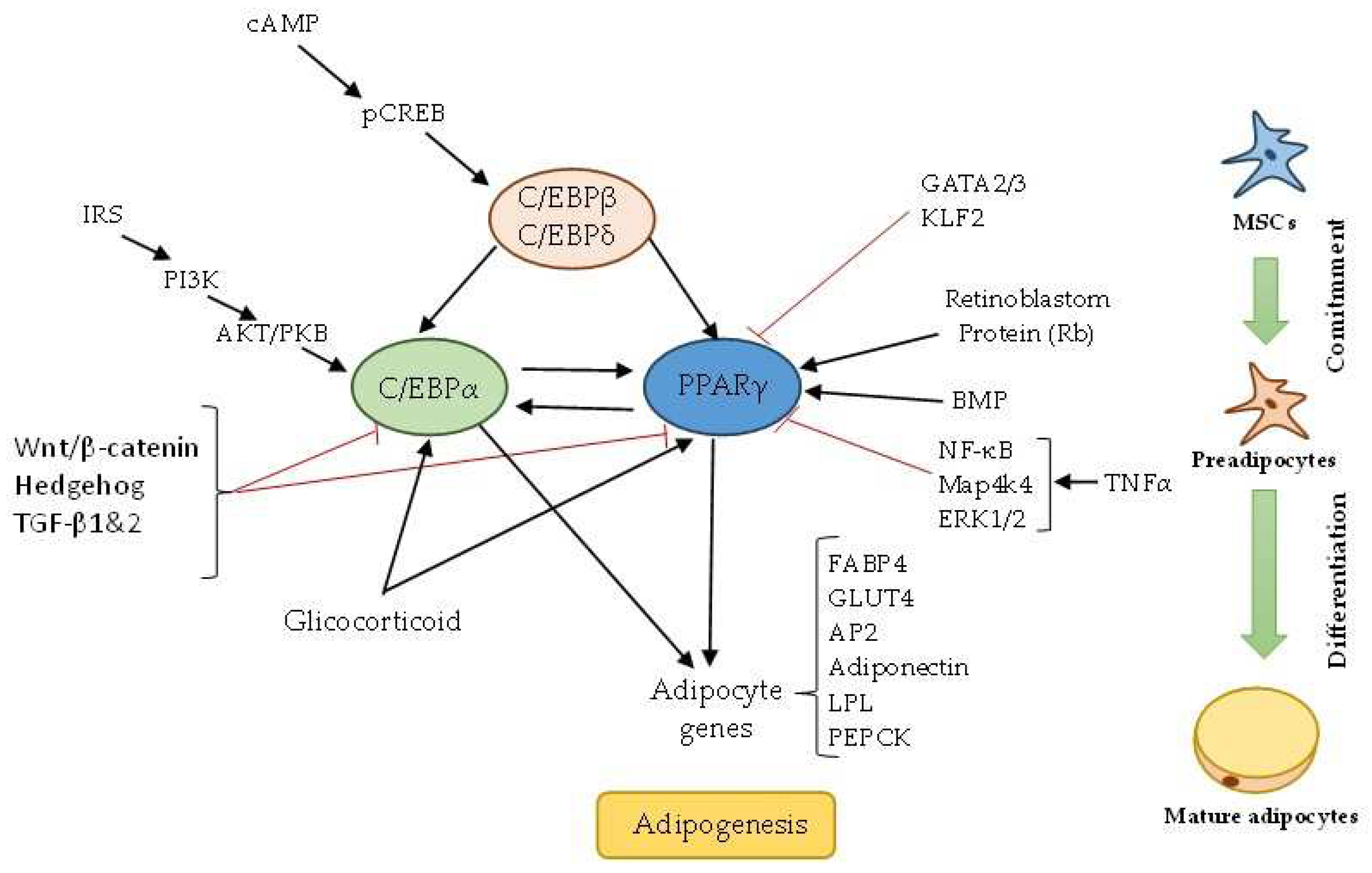
The energy imbalance between excessive caloric intake without a rise in energy expenditure leads the adipocyte to present a response of hyperplasia (formation new adipocytes) and hypertrophy (increase cell size of existing adipocytes) with an increase in adipose tissue [26,27]. Adipocytes hyperplasia, is known as adipogenesis which is a complex process by which preadipocytes (cells with fibroblastic morphology that derive mesenchymal cells) phase called commitment, differentiate into mature adipocytes (phase terminal) loaded with lipids [28,29,30]. In the process of adipogenesis, adipocytes undergo sequential phenotypic, functional and morphological changes, regulated by transcriptional cascade and signaling pathway [28,31]. The nuclear receptor PPAR-γ (peroxisome proliferator-activated receptor γ) plays an important role in differentiation, during adipogenesis CCAAT/enhancer-binding proteins (C/EBPβ/δ) stimulate C/EBPα and PPAR-γ and stimulate differentiation and induction of adipocytes-specific genes, furthermore adipocyte protein 2 (aP2), lipoprotein lipase, fatty acid synthase (FAS), and perilipin protein [28,32]. The expression of the transcription factor SREBP-1 (sterol regulatory element binding protein) also increase PPAR-γ activity in adipocyte differentiation [33].
In adipogenesis regulation (Figure 1) are involved multiple signals such as Insulin-Like Growth Factor 1 (IGF-1), Cyclic Adenosine Monophosphate (cAMP), Wnt, AMPK (adenosine 5‘-monophosphate-activated protein) glucocorticoid, BMP2, BMP4, BMP7 (Bone Morphogenic Proteins), Ras, Wnt (Wingless and INT-1 proteins), Hedgehog (Hh), Transforming Growth Factor beta 1 and 2 (TGF-β-1 and 2), Retinoblastoma Protein and Myostatin, p38/MAPK (Mitogen-activated protein kinase) and ERK/MAPK (Extracellular signal-regulated kinase/Mitogen-activated protein kinase), GATA 2/3 (Globin transcription protein-2 and -3), and TNFα (Tumor necrosis factor-α) [33,34,35,36]. Many other transcription factors are involved positively or negatively regulation of adipogenesis like Sterol regulatory element-binding protein 1 (SREBP1), and several proteins from Kruppel-like factor family (KLFs) such as KLF4, KLF5, KLF9, KLF15 and signal transducer and activator of transcription 5 (STAT5) [37,38,39].
2.3. Thermogenesis
Thermogenesis is the generation of heat by the body due to an external stimulus, is divided into shivering and non-shivering thermogenesis, both involved in energy homeostasis. In thermogenesis process are involved brown (BAT) and white (WAT) adipose tissue, mainly BAT whose function is transform energy into heat, through mitochondrial oxidative phosphorylation and an unusual mechanism to uncouple respiration to catabolize fatty acids and glucose to increase energy expenditure [40,41,42].
During lipolysis, fatty acids are release and active protein signaling in brown adipose tissue (BAT) called as thermogenin or uncoupling protein (UCP-1), this is a protein prevent the production of ATP, therefore the elevation of UCP-1 expression could prevent obesity [43]. In the other hand WAT is categorized as a visceral adipose, it function is store lipids and has a low thermogenesis capacity; however WAT can be converted to BAT-like cells (beige cells), process known as browing. Therefore, browing WAT, can be a target in the treatment of obesity [42,44,45].
2.5. Appetite Suppressants
Hypothalamus and brainstem are the key in control of food intake, perceiving metabolic signals to moderate eating behaviors [46]. However, appetite control is multifactorial event derived from the neurological and hormonal relationship [47]. Different signals converge on hypothalamus to create a network with gut, liver, pancreas, brainstem, and adipose tissue, to modulates appetite through higher cortical centers [48]. The arcuate nucleus (ARC) of the hypothalamus is considered the main appetite regulator and have neuronal subtypes. One type of neurons coexpresses orexigenic neuropeptides such as neuropeptide Y, AgRP (agouti-Related Peptide), and melanin-concentrating hormone, the other type of neurons express anorexigenic like pro-opiomelanocortin, cocaine, amphetamine-regulated transcript, nefastin-1, 5-HT (5-hydroxydopamine), dopamine and norepinephrine. The appetite regulation in controlled by the activation of anorexigenic peptides and suppression on orexigenic neuropeptides [46,48,49].
In this review we focused on the most recent publications of secondary metabolites with reported antiobesogenic effects, whose mechanisms have been elucidated in a molecular level in vivo and in vitro and have the potential use for treatment and prevention of obesity.
3. Alkaloids modulating overweight and obesity.
Alkaloids have been reported with a variety of pharmacological activities, including antibacterial, antidepressant, antihistaminic, anticancer, fungicidal, among others. Some alkaloids discovered in natural sources are nowadays used as modern drugs [50,51]. Recently, alkaloids have been related to obesity, overweight and their complications. In the past decade, along with polyphenols, alkaloids have been highly reported for their antiobesogenic effects in a molecular level, due to their structure which makes it easy to interact with molecules and receptors, especially those of the nervous system. Their effects have been evaluated in vitro and in vivo, and in several cases, mechanisms are determined in a molecular level: modulating adipogenesis, adipocyte differentiation, lipogenesis, lipolysis, fatty acid oxidation, oxidative stress and inflammation caused by obesity, and others [52]. Due to the vast information available about molecular mechanisms of alkaloids, we aimed to cite the most recent and novel articles.
The molecule bouchardatine, is an alkaloid isolated from the plant Bouchardatia neurococca, has showed interesting results regarding obesity. In 2017 a group of researchers [53] evaluated bouchardatine in 3T3-L1 cells (adipocytes) and in mice fed with a high fat diet. Results indicated that in adipocytes, this alkaloid facilitates activation of AMPK by increasing sirtuin 1 (SIRT1) activity to contribute liver kinase B1 (LKB1) activating AMPK and reduced lipid accumulation inside the cells. Additionally, in obese mice with chronic administration of bouchardatine (50 mg/kg), activation of the SIRT1-LKB1-AMPK pathway, was determined leading to attenuation of weight gain, dyslipidemia, and fatty liver. All these results in vivo were reported without any side effects or changes in food intake.
Morus alba L. is a source of alkaloids (1-deoxynojirimycin, 1,4-dideoxy-1,4-imino-D-arabinitol, fagomin) and it is already being used in tab China as treatment for diabetes mellitus type 2. To elucidate the molecular mechanisms of these alkaloids, researchers administrated the Morus alba L. powder to mice with fatty liver. The alkaloids significantly increased expression and secretion of adiponectin, both in mice and in 3T3L-1 cells. Several genes (436) were evaluated in the liver tissue of mice treated with alkaloids and the top 20 upregulated and downregulated include: reducing expression of lipid uptake genes (CD36), proinflammatory genes (C-X-C motif chemokine ligand 9 (Cxcl9) and interferon alpha-inducible protein 27-like protein 2A (Ifi27l2a). Another finding indicates that the mRNA levels of adiponectin receptor 1 (AdipoR1) and adiponectin receptor 2 (AdipoR2) were significantly higher compared to the control group of mice. In addition, the expression of the lipogenic gene PPARγ was significantly downregulated in a dose of 400 mg/kg. Morus alba L. alkaloids were an effective treatment for the activation of AMPK and upregulation of PPARα and PGC1α expression to help improving β-oxidation of fatty acids. These findings imply that genes and pathways of lipid metabolism and metabolic stress-induced liver injury in obese mice are regulated by alkaloids [54].
Berberine is one of the most reported alkaloids regarding its antiobesogenic effects. It is an isoquinoline alkaloid, extracted from plant of the Coptis and Phellodendron genera [55]. This compound is capable of increasing fatty acid oxidation by activiting AMPK in hepatic cell lines (HepG2), and, in the liver and adipose tissue of mice with obesity-associated non-alcoholic fatty liver disease, showed reduction of the phosphorylation state of JNK1 and the mRNA levels of proinflammatory cytokines [56]. It has also been reported as an alleviator of adipogenesis in 3T3-L1 cell lines, via regulation of via the AMPKα-SREBP pathway [57]. It also increases the ATP-binding cassette transporter, which mediates hepatic cholesterol modulatesprotein kinase C phosphorylation on Tyr311, alleviating hepatic lipid accumulation [58]. In mice white adipose tissue, berberine inhibited adipocyte differentiation, proliferation and adiposity, all through down-regulating galectin-3 [59]. In male mice fed with a high fat diet, berberine (25 mg/kg and 100 mg/kg) activates the energy metabolic sensing pathway AMPK/SIRT1 axis, increasing PPARγ deacetylation, finall leading to the remodeling of the adipose tissue and a significant increase of the thermogenic protein UCP-1 [60] Recently, berberine has shown a novel antiobesogenic effect: administrated orally to obese mice, it binds to the bitter-taste receptors present in the intestine (TAS2Rs). Via signaling the TAS2Rs pathway, berberine upregulates release of GLP-1 and helps ameliorating obesity [61].
In a different approach, eight alkaloids present in lotus (Nelumbo nucifera Gaertn.) leaves, were evaluated as dopamine receptors (D1 and D2) antagonists in kidney cells (human embryonic kidney 239 cell lines). Antagonizing these receptors is an strategy that has recently proven to be effective in long term maintenance of weight loss. Among the alkaloids tested, o-nornuciferine was the most potent as a dopamine inhibitor. This is an aporphine alkaoid, which means it is has an extra ring, connecting a tetrahydroisoquinoline and a benzyl. Analyzing the o-nornuciferine, it was determined that the R3 was the critical pharmacophore, and that methyl substation was is preferred than hydrogen at this position. In addition, this alkaloid is capable of fully desensity serotonin 2A, by antagonizing the receptor 5-HT2A [62]. Also extracted from lotus leaves, the aporphine alkaloid nuciferine has been widely studied due to its antiobesogenic effects, such as lowering blood lipid and glucose values and reducing inflammation [63]. One of the most recent publications, determined that this molecule ameliorates lipid accumulation, apoptosis, and impaired migration through the activation of LKB1/AMPK signaling pathway, thus, reducing the lipotoxicity related to overweight and obesity[64]. And in mice fed with high fat diet, it improves the microbiome dysbiosis, and downregulating expression of pro-inflammatory genes IL-6, IL-1β, and TNF-α [65].
Other alkaloids have been reported with a variety of molecular mechanisms related to obesity, adipogenesis, lipogenesis, oxidative stress and inflammation related to obesity, such as evodiamine, capable of lowering oxidative stress and inflammation cause by free fatty acids, by inhibiting enhanced expression of P2X7, and its dependent TNF-α expression and ERK 1/2 phosphorylation [66]. Betaine administered to mice with hepatic steatosis, a significant activation of AMPK and down-regulation of sterol regulatory element-binding protein 1C (SREBP-1c) was determined, enhancing lipid metabolism [67]. Curcumin is a potent alkaloid that has show an antiobesity mechanism through modulation of SIRT1/AMPKα/FOXO1 in bovine adipocytes and modulating UPC1 (uncoupling protein 1) in male mice with induced obesity [68].
The structure-relationship between most of these alkaloids and their molecular mechanisms against obesity has been discussed, and mainly attributed to the alkoxyl of the C1, hydroxyl at the C2 position and the alkyl substituent on the N-atom. Also, the lack of a methyl group in R1 and R3 position, has been mentioned as an important factor for their effectiveness. Although it is important to mention low bioavailability, absorption, and water solubility of alkaloids are some important factors to consider for their application, implying that micro or nano encapsulation are a convenient future solution for their administration [69,70,71].
4. Saponins modulating overweight and obesity.
Even though saponins are known for their antiobesogenic effects as inhibitors of lipid digestion and absorption on an intestinal level, there are other effects and mechanisms of these molecules in different cells or tissues, such as adipose and hepatic tissue which are key organs involved in obesity development.
Bupleurum chinensis, a plant whose root has been used to treat inflammation, fever, liver diseases, but also against lipid accumulation. The mechanism linked to its anti-obesogenic activity was determined isolating two saponins: saikosaponin A and saikosaponin D and evaluating their effect on adipocytes (3T3-L1 cells). Results indicated that, in doses that do not compromise the cell viability, these two saponins suppresing expression of adipogenic genes: peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT/enhancer binding protein alpha (C/EBPα), sterol regulatory element binding protein-1c (SREBP-1c), and adiponectin. They also downregulated expression of lipogenic genes: fatty acid binding protein (FABP4), fatty acid synthase (FAS), and lipoprotein lipase (LPL) [72].
A rich in saponins fraction obtained from green tea seeds was used to design a formulation with glucosides extracted from Stevia rebaudiana (stevia) and evaluated in 3T3-L1 cells. The combination showed a synergic behavior and significantly inhibited the expression of adipogenesis and lipogenesis related genes and signaling molecules: PPARγ, C/EBPα, aP2, and SREBP-1c (sterol regulatory element-binding protein)[73].
Ginsenoside Rg2 was extracted from red ginger to use as treatment in obese mice and in 3T3-L1 cells to ellucidate the antobesogenic mechanism of this saponin. Both in vivo and in vitro, Gingenoside Rg2 induced the activation of AMPK, subsecuenlty decreasing the expresion of adipogenic transcription factors (PPARγ, C/EBPα, and SREBP1-c) [74]. Similar research was conducted using only saponins extracted from the roots of ginsen and tested on obese mice with induced non-alcoholic fatty liver disease. Anti-inflammatory activity was observed in the mice liver tissue, modulating the expression inflammatory cytokines ARG1 (arginase), CCL2 (, and IL-1β.
Soy saponins were administred to mice and used as treatment in 3T3-L1 cells. Results indicated that these compounds are involved in the reduction of triacylglycerol accumulation and lipogenesis in adipose cells. While some saponins have demonstrated their antiobesogenic mechanism is linked to downregulating adipogenesis, soysaponins have an effect on lipogensis, significantly downregulating SREBP-1c and fatty acid synthase (FAS) [75].
Other molecular mechanisms were reported in 2019. Gynostemma pentaphyllum is a plant used in traditional medicine to treat diabetes mellitus, dyslipidemia, and inflammation. It was demonstrated that, among other mechanisms, the saponins present in its extract were able to significantly decrease the expression of adipocyte protein 2 (AP2), and sirtuin 1 (SIRT1). It also increased the expression of carnitine palmitoyltransferase (CPT1) and hormone- sensitive lipase (HSL), thus contributing to ameliorating obesity in male mice[76] .
It has been determined that the primary structure-activity relationship for saponins effects, is the ether bond between carbon 13 (C-13) and carbon 28 (C-28). Other structural factors for their bioactive effects are the number of sugars in the carbon 3 position and the aglycone structure, this may confer saponins the ability to interact with nuclar factors related to ameliorating obesity [77,78].
5. Terpenes modulating overweight and obesity.
Terpenes are a very extensive group of secondary metabolites with proven anti-obesogenic effects [79]. Previously it was reported that terpenes from plants exert action as modulators of receptors involved in the metabolism of lipids such as PPARs and LXRs (liver X receptors) [80]. Therefore, several plant species have been studied in order to identify their natural components, such as terpenes or their derivatives, and to understand their anti-obesogenic mechanisms to identify possible targets for obesity treatment [81].
Terpenes from Ilex aquifolium have been studied in order to understand their anti-obesity mechanism. Pachura, et al. [82] evaluated the effect of the terpenoid fraction of extracts from I. aquifolium leaves on the expression of genes related to the lipid metabolism and on the hepatic architecture using obese Zucker male rats as in vivo model. Firstly, the composition of I. aquifolium leaves was analyzed, the monoterpenes p-cymene, α-phellandrene and α-pinene, as well as the triterpenes oleanolic and ursolic acid were found. Secondly, rats were fed 10 mg of terpenoid fraction per kg of body weight during eight weeks and compared with the obese control group. Rats fed with the terpenoid fraction showed a significantly increase in the expression of the gene LXR1 (liver X receptor) factor, which is required for the elimination of cholesterol in mice [83]. Furthermore, intake of terpenoids from I. aquifolium reduced the accumulation of lipids in the liver, as compared with the obese rats in the control group. Additionally, the terpenes of this plant species exerted antioxidant and anti-inflammatory effects. This evidence indicates that the terpenes found in the leaves of I. aquifolium have potential as lipid metabolism modulators and could be used as alternative in obesity treatment.
Another plant species recently studied is Brucea javanica. Lahrita, et al. [84] evaluated the pro-lipolytic effect of terpenes, known as quassinoids, found in extracts from B. javanica fruit using 3T3-L1 cells differentiated to adipocytes as in vitro model. The extracts from the fruit contained the terpenes brucein A-C, brusatol, bruceantinol, hydroxybrucein A, and yadanzioside B. 3T3-L1 adipocytes were treated with the extracts at concentrations of 3.13-100 µg/mL for 24 h, and glycerol released was measured as a lipolysis marker. All concentrations evaluated significantly increased the glycerol levels, compared with the control. Furthermore, individual terpenes from the extract were evaluated, it was found that yadanzioside B (1-100 µM) did not significantly affect the glycerol levels. On the other hand, the rest of the terpenes brucein A-C, brusatol, bruceantinol, and hydroxybrucein A exerted pro-lipolytic activity in the cells with a dose-dependent effect (0.16-10 µM), and bruceantinol showed the highest lipolytic activity. According to the authors the presence of a hydroxyl group in the position 3, as well as shorts acyl chains esterified to the hydroxyl in the position 15 may be of relevance for the pro-lipolytic activity of these terpenes. These demonstrates the potential of quassinoids from B. javanica in the treatment of obesity.
In addition to plants, other natural products containing terpenes, such as marine resources, have been explored for their anti-obesogenic properties; such is the case of soft corals of the order Alcyonacea [85]. In this study, the effect of methanolic extract of Sarcophyton glaucum supplementation to obese rats on several obesity markers was studied. Obese male albino rats fed crude methanolic extract of S. glaucum (20 mg/kg) during eight weeks showed lower levels of fasting blood sugar, HOMA-IR (homeostatic model assessment for insulin resistance), cholesterol, triglycerides, and LDL-cholesterol, and higher levels of insulin and HDL-cholesterol, when compared with the obese control group (without S. glaucum extract supplementation).
Furthermore, levels of fetuin A, fetuin B, and PTP1Β (protein tyrosine phosphatase 1B) were significantly reduced; while adropin and omentin levels increased in the serum of rats fed the extract compared with the control group. Fetuin A is known for participating as inhibitor of the lysis of lipids which causes accumulation of triglycerides in the liver and might alter the adipogenesis. On the other hand, PTP1B contributes to insulin resistance. Also, treatment of obese rats with the extract increased the expression of PPARγ coactivator 1 alpha. The regulation of these markers is promising as target for the treatment of obesity. A total of 27 terpenoids were identified in the methanolic extract of S. glaucum, where two of them showed the highest anti-obesogenic potential, one tricyclic and one bicyclic diterpenes with cores II and III, respectively. These diterpenes shared an α, β-unsaturated ε-caprolactone fused to a cyclic system in their structures, which might be important for their activity.
Polyphenols have been the focus of many studies regarding the promotion of human health. Many reports associate polyphenols with antioxidant, anti-inflammatory, anticancer, with protective effects against neurodegenerative diseases, and as potential agents against overweight and obesity. Boix-Castejón, et al. [86], reported that a combination of Lippia citriodora and Hibiscus sabdariffa polyphenol-rich extracts administered at 500 mg/day to 54 overweight subjects could improve anthropometric measurements like body fat, decreases blood pressure and heart rate. Also, the extracts showed the levels of adipohormones hormones regulating hunger and satiety, the anorexigenic hormones like glucagon-like peptide-1 and decreased the levels of ghrelin, an orexigenic hormone. The authors suggested that AMP-activated protein kinases may play a role in the regulation of energy homeostasis, daily energy expenditure, and lipid metabolism. A further study in obese individuals in a randomized controlled trial, showed that administration of L. citriodora and H. sabdariffa extracts for 2 months decreased with, improved the anthropometric parameters of body weight from, body mass index, abdominal circumference, and percentage of body fat. Also, the treatment with the extracts in overweight and obese individuals reduced systolic and diastolic blood pressure by the first 30 days of treatment. Interestingly, no adverse effects were observed during the study. This effect was attributed to the most abundant compounds identified in the mixture: verbascoside, delphinidin-3-O-sambudioside, cyanidin-3-O-samudioside, and isoverbascoside. It was mentioned that synergy of these molecules could mediate leptin levels, regulating satiety and by modulation of the activation of AMPK [87].
López-Tenorio, et al. [88] also showed the potential synergistic action of natural products as they showed that the combination of morin (50 mg/kg weight) and polyunsaturated fatty acids (1 mL/kg weight) (EPA and DHA, 1:1 ratio) downregulated the expression of the inflammasome biomarker Nlrp3 mRNA when administered to high-fat fed Wistar rats, as the effects of the administered bioactive components was better than each individually. Furthermore, the mixture also improved biochemical parameters like decreased levels in triacylglycerides, increased HDL cholesterol and decreased LDL cholesterol levels.
Cocoa polyphenols have been associated with decreased levels of inflammation biomarkers in obesity. The study of Gu, et al. [89] administered cocoa powder to high-fat diet fed male C57BL/6J mice. The cocoa supplementation decreased weight gain, final body weight, insulin resistance by improvement of the HOMA-IR and decreased the severity of fatty liver disease by modulating the levels of plasma alanine aminotransferase and liver triglyceride. Furthermore, cocoa supplementation decreased the plasmatic levels of the pro-inflammatory biomarkers IL-6, MCP-1, and increased the levels of adiponectin by 33.6%, which was associated with the modulation of the gene expression of these proteins.
Terpenes and other natural products show potential as therapeutic agents against obesity due to their capacity to modulate diverse markers related to the lipid and glucose metabolism.
6. Conclusions and perspectives
Understanding the mechanisms of action and safe dosage of plants, fruits, extracts, compounds, or any type of nutraceutical is an essential step for designing effective nutraceuticals and biopharmaceuticals. This information can also help to determine when it is safe to use them, whether is a matter of age, or health condition. It can also be useful when deciding if using them as coadjuvants or for prevention, could have antagonism with other treatments that the individual is already having. On the other hand, analyzing all this recent information has led as to understand the importance of evaluating bioaccessibility and bioavailability of the antiobesogenic compounds. Evaluating these two important factors can help determine the best way to administrate them, whether being part of the diet or using the idoneous encapsulation model. Considering obesity is now considered an epidemic, and that not all people have access to the drugs available in the market, having other options or coadjuvants is crucial, especially considering that the anti-obesogenic effects of natural compounds have not yet show side effects in in vivo evaluations.
Author Contributions
“Conceptualization, C.A.E.R., E.P.G.G.; investigation, C.A.E.R., N.L.L., L.A.C.A., R.C.P.L., L.Z.R.A., J.L.F., S.A.B.O., E.P.G.G.; writing—review and editing, C.A.E.R., J.B.H., E.P.G.G. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization. Obesity and overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 January 2024).
- Centers for Disease Control and Prevention. Adult obesity facts. Available online: https://www.cdc.gov/obesity/data/adult.html (accessed on 10 January 2024).
- National Institute of Diabetes and Digestive and Kidney Diseases. Overweight and obesity statistics. Available online: https://www.niddk.nih.gov/health-information/health-statistics/overweight-obesity (accessed on 10 January 2024).
- Pledger, S.L.; Ahmadizar, F. Gene-environment interactions and the effect on obesity risk in low and middle-income countries: a scoping review. Frontiers in Endocrinology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Withrow, D.; Alter, D.A. The economic burden of obesity worldwide: a systematic review of the direct costs of obesity. Obesity Reviews 2011, 12, 131–141. [Google Scholar] [CrossRef] [PubMed]
- González-Castejón, M.; Rodriguez-Casado, A. Dietary phytochemicals and their potential effects on obesity: A review. Pharmacological Research 2011, 64, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Grijalva, E.P.; Leyva-López, N.; Vazquez-Olivo, G.; Heredia, J.B. Oregano as a potential source of antidiabetic agents. J Food Biochem 2022, 46, e14388. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Molath, A.; Choksi, H.; Kumar, S.; Mehra, R. Classifications of polyphenols and their potential application in human health and diseases. Int. J. Physiol. Nutr. Phys. Educ 2021, 6, 293–301. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Kukula-Koch, W.; Widelski, J. Alkaloids. 2017; pp. 163-198.
- Abookleesh, F.L.; Al-Anzi, B.S.; Ullah, A. Potential Antiviral Action of Alkaloids. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F. Alkaloids: Biochemistry, Ecology, and Medicinal Applications; Springer US: 2013.
- Aguilar Salguero, S.A. Relación de la estructura de las saponinas con sus aplicaciones, una revisión actualizada. 2022.
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes, Ramawat, K.G., Mérillon, J.-M., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2013; pp. 2665–2691. [Google Scholar]
- Roba, K. The role of terpene (secondary metabolite). Nat. Prod. Chem. Res. 2020, 9. [Google Scholar]
- Mabou, F.D.; Yossa, I.B.N. TERPENES: Structural classification and biological activities. IOSR J. Pharm. Biol. Sci. e-ISSN 2021, 16, 2319–7676. [Google Scholar]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int J Mol Sci 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Biobaku, F.; Ghanim, H.; Batra, M.; Dandona, P. Macronutrient-Mediated Inflammation and Oxidative Stress: Relevance to Insulin Resistance, Obesity, and Atherogenesis. J Clin Endocrinol Metab 2019, 104, 6118–6128. [Google Scholar] [CrossRef] [PubMed]
- Karam, B.S.; Chavez-Moreno, A.; Koh, W.; Akar, J.G.; Akar, F.G. Oxidative stress and inflammation as central mediators of atrial fibrillation in obesity and diabetes. Cardiovasc Diabetol 2017, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Guarner-Lans, V.; Rubio-Ruiz, M.E. Reductive Stress in Inflammation-Associated Diseases and the Pro-Oxidant Effect of Antioxidant Agents. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediators Inflamm 2010, 2010. [Google Scholar] [CrossRef]
- Sikaris, K.A. The clinical biochemistry of obesity. The Clinical Biochemist Reviews 2004, 25, 165. [Google Scholar]
- Ahmad, B.; Friar, E.P.; Vohra, M.S.; Garrett, M.D.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Mechanisms of action for the anti-obesogenic activities of phytochemicals. Phytochemistry 2020, 180, 112513. [Google Scholar] [CrossRef]
- Rahim, A.T.M.A.; Takahashi, Y.; Yamaki, K. Mode of pancreatic lipase inhibition activity in vitro by some flavonoids and non-flavonoid polyphenols. Food Research International 2015, 75, 289–294. [Google Scholar] [CrossRef]
- Ràfols, E.M. Tejido adiposo: heterogeneidad celular y diversidad funcional. Endocrinología y Nutrición 2014, 6, 100–112. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From Stem Cell to Adipocyte. Annual Review of Biochemistry 2012, 81, 715–736. [Google Scholar] [CrossRef]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes, Metabolic Syndrome and Obesity 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: adipocytes or osteoblasts? Cell Death & Differentiation 2016, 23, 1128–1139. [Google Scholar]
- Nic-Can, G.I.; Rodas-Junco, B.A.; Carrillo-Cocom, L.M.; Zepeda-Pedreguera, A.; Peñaloza-Cuevas, R.; Aguilar-Ayala, F.J.; Rojas-Herrera, R.A. Epigenetic Regulation of Adipogenic Differentiation by Histone Lysine Demethylation. International Journal of Molecular Sciences 2019, 20, 3918. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Liu, D.; Zeng, R.; Xian, T.; Lu, Y.; Zeng, G.; Sun, Z.; Huang, B.; Huang, Q. Epigallocatechin-3-gallate inhibits adipogenesis through down-regulation of PPARγ and FAS expression mediated by PI3K-AKT signaling in 3T3-L1 cells. European Journal of Pharmacology 2017, 795, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Balaji, M.; Ganjayi, M.S.; Hanuma Kumar, G.E.N.; Parim, B.N.; Mopuri, R.; Dasari, S. A review on possible therapeutic targets to contain obesity: The role of phytochemicals. Obesity Research & Clinical Practice 2016, 10, 363–380. [Google Scholar]
- Khalilpourfarshbafi, M.; Gholami, K.; Murugan, D.D.; Abdul Sattar, M.Z.; Abdullah, N.A. Differential effects of dietary flavonoids on adipogenesis. European Journal of Nutrition 2019, 58, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. International Journal of Molecular Sciences 2020, 21. [Google Scholar] [CrossRef]
- Jiang, N.; Li, Y.; Shu, T.; Wang, J. Cytokines and inflammation in adipogenesis: an updated review. Frontiers of Medicine 2019, 13, 314–329. [Google Scholar] [CrossRef]
- Son, Y.H.; Ka, S.; Kim, A.Y.; Kim, J.B. Regulation of Adipocyte Differentiation via MicroRNAs. enm 2014, 29, 122–135. [Google Scholar] [CrossRef]
- Lowe, C.E.; O'Rahilly, S.; Rochford, J.J. Adipogenesis at a glance. Journal of Cell Science 2011, 124, 2681–2686. [Google Scholar] [CrossRef]
- Tang, P.; Virtue, S.; Goie, J.Y.G.; Png, C.W.; Guo, J.; Li, Y.; Jiao, H.; Chua, Y.L.; Campbell, M.; Moreno-Navarrete, J.M.; et al. Regulation of adipogenic differentiation and adipose tissue inflammation by interferon regulatory factor 3. Cell Death & Differentiation 2021, 28, 3022–3035. [Google Scholar]
- Linda, C.; Urarat, N.; Rawiwun, K.; Sudarat, H.; Wanwisa, S. Inhibitory Effect of Carallia Brachiata Extract Through Regulation of Adipogenesis Pathways in 3T3-L1 Cells. Pharmacognosy Journal 2022, 14, 655–660. [Google Scholar]
- Audano, M.; Pedretti, S.; Caruso, D.; Crestani, M.; De Fabiani, E.; Mitro, N. Regulatory mechanisms of the early phase of white adipocyte differentiation: an overview. Cellular and Molecular Life Sciences 2022, 79, 139. [Google Scholar] [CrossRef] [PubMed]
- Pant, R.; Firmal, P.; Shah, V.K.; Alam, A.; Chattopadhyay, S. Epigenetic Regulation of Adipogenesis in Development of Metabolic Syndrome. Front Cell Dev Biol 2020, 8, 619888. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Matsushita, M.; Yoneshiro, T.; Okamatsu-Ogura, Y. Brown Adipose Tissue, Diet-Induced Thermogenesis, and Thermogenic Food Ingredients: From Mice to Men. Frontiers in Endocrinology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Kazak, L.; Spiegelman, B.M. New advances in adaptive thermogenesis: UCP1 and beyond. Cell metabolism 2019, 29, 27–37. [Google Scholar] [CrossRef]
- Pan, R.; Zhu, X.; Maretich, P.; Chen, Y. Combating Obesity With Thermogenic Fat: Current Challenges and Advancements. Front Endocrinol (Lausanne) 2020, 11, 185. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, X.; Chen, Y. Recruitment of Thermogenic Fat: Trigger of Fat Burning. Frontiers in Endocrinology 2021, 12. [Google Scholar] [CrossRef]
- Yu, J.H.; Kim, M.-S. Molecular Mechanisms of Appetite Regulation. dmj 2012, 36, 391–398. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bulletin of Faculty of Pharmacy, Cairo University 2014, 52, 269–284. [Google Scholar] [CrossRef]
- Hussain, S.S.; Bloom, S.R. The regulation of food intake by the gut-brain axis: implications for obesity. International Journal of Obesity 2013, 37, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Jiang, Y.; Guo, J.; Su, Z. Natural Products with Anti-obesity Effects and Different Mechanisms of Action. Journal of Agricultural and Food Chemistry 2016, 64, 9571–9585. [Google Scholar] [CrossRef] [PubMed]
- Thawabteh, A.; Juma, S.; Bader, M.; Karaman, D.; Scrano, L.; Bufo, S.A.; Karaman, R. The Biological Activity of Natural Alkaloids against Herbivores, Cancerous Cells and Pathogens. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Grijalva, E.P.; López-Martínez, L.X.; Contreras-Angulo, L.A.; Elizalde-Romero, C.A.; Heredia, J.B. Plant Alkaloids: Structures and Bioactive Properties. In Plant-derived Bioactives: Chemistry and Mode of Action, Swamy, M.K., Ed.; Springer Singapore: Singapore, 2020; pp. 85–117. [Google Scholar]
- Balaji, M.; Ganjayi, M.S.; Kumar, G.E.H.; Parim, B.N.; Mopuri, R.; Dasari, S. A review on possible therapeutic targets to contain obesity: The role of phytochemicals. Obesity research & clinical practice 2016, 10, 363–380. [Google Scholar]
- Rao, Y.; Yu, H.; Gao, L.; Lu, Y.-T.; Xu, Z.; Liu, H.; Gu, L.-Q.; Ye, J.-M.; Huang, Z.-S. Natural alkaloid bouchardatine ameliorates metabolic disorders in high-fat diet-fed mice by stimulating the sirtuin 1/liver kinase B-1/AMPK axis. British Journal of Pharmacology 2017, 174, 2457–2470. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Lian, C.-F.; Sun, Q.-W.; Wang, T.-T.; Liu, Y.-Y.; Ye, J.; Gao, L.-L.; Yang, Y.-F.; Liu, S.-N.; Shen, Z.-F.; et al. Ramulus Mori (Sangzhi) Alkaloids Alleviate High-Fat Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice. Antioxidants 2022, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- Pirillo, A.; Catapano, A.L. Berberine, a plant alkaloid with lipid- and glucose-lowering properties: From in vitro evidence to clinical studies. Atherosclerosis 2015, 243, 449–461. [Google Scholar] [CrossRef]
- Guo, T.; Woo, S.L.; Guo, X.; Li, H.; Zheng, J.; Botchlett, R.; Liu, M.; Pei, Y.; Xu, H.; Cai, Y.; et al. Berberine Ameliorates Hepatic Steatosis and Suppresses Liver and Adipose Tissue Inflammation in Mice with Diet-induced Obesity. Sci Rep 2016, 6, 22612. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, X.; Feng, X.; Liu, X.; Deng, C.; Hu, C.H. Berberine Alleviates Olanzapine-Induced Adipogenesis via the AMPKα-SREBP Pathway in 3T3-L1 Cells. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef]
- Liang, H.; Wang, Y. Berberine alleviates hepatic lipid accumulation by increasing ABCA1 through the protein kinase C δ pathway. Biochem Biophys Res Commun 2018, 498, 473–480. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Ma, S.R.; Zuo, Z.Y.; Wu, Y.B.; Kong, W.J.; Wang, A.P.; Jiang, J.D. Berberine inhibits adipocyte differentiation, proliferation and adiposity through down-regulating galectin-3. Sci Rep 2019, 9, 13415. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, T.; Ma, G.; Zheng, L.; Jiang, X.; Yang, F.; Wang, Z.; Li, N.; He, Z.; Song, X.; et al. Berberine modulates deacetylation of PPARγ to promote adipose tissue remodeling and thermogenesis via AMPK/SIRT1 pathway. Int J Biol Sci 2021, 17, 3173–3187. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Yang, Y.; Xiong, R.; Ni, Y.; Ma, X.; Hou, M.; Chen, L.; Xu, Z.; Chen, L.; Ji, M. Oral berberine ameliorates high-fat diet-induced obesity by activating TAS2Rs in tuft and endocrine cells in the gut. Life Sci. 2022, 311, 121141. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hou, T.; Gao, Z.; Guo, X.; Wang, C.; Wang, J.; Liu, Y.; Liang, X. Discovery of eight alkaloids with D1 and D2 antagonist activity in leaves of Nelumbo nucifera Gaertn. Using FLIPR assays. J. Ethnopharmacol. 2021, 278, 114335. [Google Scholar] [CrossRef]
- Wan, Y.; Xia, J.; Xu, J.-f.; Chen, L.; Yang, Y.; Wu, J.-J.; Tang, F.; Ao, H.; Peng, C. Nuciferine, an active ingredient derived from lotus leaf, lights up the way for the potential treatment of obesity and obesity-related diseases. Pharmacol. Res 2022, 175, 106002. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, C.; Liu, M.; Chen, L.; Zhu, Y.; Gao, W.; Du, X.; Song, Y.; Li, X.; Liu, G. Nuciferine ameliorates nonesterified fatty acid-induced bovine mammary epithelial cell lipid accumulation, apoptosis, and impaired migration via activating LKB1/AMPK signaling pathway. J. Agric. Food Chem 2022, 71, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.-T.; Liao, J.-B.; Yang, Z.-X.; Cui, H.-T.; Zhang, Z.-Y.; Wen, W.-B.; Wang, H.-W. Effect of nuciferine on gut microbiota and inflammatory response in obese model mice. Zhongyaocai 2021, 46, 2104–2111. [Google Scholar] [CrossRef]
- Xue, Y.; Guo, T.; Zou, L.; Gong, Y.; Wu, B.; Yi, Z.; Jia, T.; Zhao, S.; Shi, L.; Li, L.; et al. Evodiamine Attenuates P2X(7)-Mediated Inflammatory Injury of Human Umbilical Vein Endothelial Cells Exposed to High Free Fatty Acids. Oxid Med Cell Longev 2018, 2018, 5082817. [Google Scholar] [CrossRef]
- Xu, G.; Huang, K.; Zhou, J. Hepatic AMP Kinase as a Potential Target for Treating Nonalcoholic Fatty Liver Disease: Evidence from Studies of Natural Products. Curr Med Chem 2018, 25, 889–907. [Google Scholar] [CrossRef]
- Wang, H.N.; Xiang, J.Z.; Qi, Z.; Du, M. Plant extracts in prevention of obesity. Crit Rev Food Sci Nutr 2022, 62, 2221–2234. [Google Scholar] [CrossRef]
- Yang, Z.; Song, Z.; Xue, W.; Sheng, J.; Shu, Z.; Shi, Y.; Liang, J.; Yao, X. Synthesis and structure–activity relationship of nuciferine derivatives as potential acetylcholinesterase inhibitors. Med Chem Res 2014, 23, 3178–3186. [Google Scholar] [CrossRef]
- Behl, T.; Singh, S.; Sharma, N.; Zahoor, I.; Albarrati, A.; Albratty, M.; Meraya, A.M.; Najmi, A.; Bungau, S. Expatiating the Pharmacological and Nanotechnological Aspects of the Alkaloidal Drug Berberine: Current and Future Trends. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Cheng, C.; Li, Z.; Zhao, X.; Liao, C.; Quan, J.; Bode, A.M.; Cao, Y.; Luo, X. Natural alkaloid and polyphenol compounds targeting lipid metabolism: Treatment implications in metabolic diseases. Eur J Pharmacol 2020, 870, 172922. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Lee, H.S.; Han, H.-K.; Choi, C.-I. Saikosaponin A and D Inhibit Adipogenesis via the AMPK and MAPK Signaling Pathways in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2021, 22, 11409. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Khan, M.Z.; Shin, J.H.; Shin, T.S.; Lee, Y.B.; Kim, M.Y.; Kim, J.D. Pharmacological Approaches to Attenuate Inflammation and Obesity with Natural Products Formulations by Regulating the Associated Promoting Molecular Signaling Pathways. BioMed Research International 2021, 2021, 2521273. [Google Scholar] [CrossRef]
- Liu, H.; Liu, M.; Jin, Z.; Yaqoob, S.; Zheng, M.; Cai, D.; Liu, J.; Guo, S. Ginsenoside Rg2 inhibits adipogenesis in 3T3-L1 preadipocytes and suppresses obesity in high-fat-diet-induced obese mice through the AMPK pathway. Food & Function 2019, 10, 3603–3614. [Google Scholar] [CrossRef]
- Tsai, W.-T.; Nakamura, Y.; Akasaka, T.; Katakura, Y.; Tanaka, Y.; Shirouchi, B.; Jiang, Z.; Yuan, X.; Sato, M. Soyasaponin ameliorates obesity and reduces hepatic triacylglycerol accumulation by suppressing lipogenesis in high-fat diet-fed mice. J Food Sci. 2021, 86, 2103–2117. [Google Scholar] [CrossRef]
- Lee, H.S.; Lim, S.M.; Jung, J.I.; Kim, S.M.; Lee, J.K.; Kim, Y.H.; Cha, K.M.; Oh, T.K.; Moon, J.M.; Kim, T.Y.; et al. Gynostemma Pentaphyllum Extract Ameliorates High-Fat Diet-Induced Obesity in C57BL/6N Mice by Upregulating SIRT1. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Sun, C.L.; Geng, C.A.; Huang, X.Y.; Ma, Y.B.; Zheng, X.H.; Yang, T.H.; Chen, X.L.; Yin, X.J.; Zhang, X.M.; Chen, J.J. Bioassay-guided isolation of saikosaponins with agonistic activity on 5-hydroxytryptamine 2C receptor from Bupleurum chinense and their potential use for the treatment of obesity. Chin J Nat Med 2017, 15, 467–473. [Google Scholar] [CrossRef]
- Luo, Z.; Xu, W.; Zhang, Y.; Di, L.; Shan, J. A review of saponin intervention in metabolic syndrome suggests further study on intestinal microbiota. Pharmacol. Res 2020, 160, 105088. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Mubarak, M.S. Anti-obesity Effect of Plant Diterpenes and their Derivatives: A Review. Phytother. Res. 2020, 34, 1216–1225. [Google Scholar] [CrossRef]
- Goto, T.; Takahashi, N.; Hirai, S.; Kawada, T. Various Terpenoids Derived from Herbal and Dietary Plants Function as PPAR Modulators and Regulate Carbohydrate and Lipid Metabolism. PPAR Res. 2010, 2010, 483958. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, M.; Yadav, P.; Vashishth, D.; Sharma, K.; Kumar, A.; Chahal, J.; Dalal, S.; Kataria, S.K. A Review on Obesity Management through Natural Compounds and a Green Nanomedicine-Based Approach. Molecules. 2021, 26, 3278. [Google Scholar] [CrossRef] [PubMed]
- Pachura, N.; Kupczyński, R.; Lewandowska, K.; Włodarczyk, M.; Klemens, M.; Kuropka, P.; Nowaczyk, R.; Krzystek-Korpacka, M.; Bednarz-Misa, I.; Sozański, T.; et al. Biochemical and Molecular Investigation of the Effect of Saponins and Terpenoids Derived from Leaves of Ilex aquifolium on Lipid Metabolism of Obese Zucker Rats. Molecules. 2022, 27, 3376. [Google Scholar] [CrossRef] [PubMed]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.-M.A.; Hammer, R.E.; Mangelsdorf, D.J.J.C. Cholesterol and Bile Acid Metabolism are Impaired in Mice Lacking the Nuclear Oxysterol Receptor Lxrα. Cell. 1998, 93, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Lahrita, L.; Moriai, K.; Iwata, R.; Itoh, K.; Kato, E. Quassinoids in Brucea javanica are Potent Stimulators of Lipolysis in Adipocytes. Fitoterapia. 2019, 137, 104250. [Google Scholar] [CrossRef] [PubMed]
- Tammam, M.A.; Aly, O.; Pereira, F.; Mahdy, A.; El-Demerdash, A. Unveiling the Potential of Marine-Derived Diterpenes from the order Alcyonacea as Promising Anti-obesity Agents. Curr. Res. Biotechnol. 2024, 100175. [Google Scholar] [CrossRef]
- Boix-Castejón, M.; Herranz-López, M.; Pérez Gago, A.; Olivares-Vicente, M.; Caturla, N.; Roche, E.; Micol, V. Hibiscus and lemon verbena polyphenols modulate appetite-related biomarkers in overweight subjects: A randomized controlled trial. Food. Funct. 2018, 9, 3173–3184. [Google Scholar] [CrossRef]
- Herranz-López, M.; Olivares-Vicente, M.; Boix-Castejón, M.; Caturla, N.; Roche, E.; Micol, V. Differential effects of a combination of Hibiscus sabdariffa and Lippia citriodora polyphenols in overweight/obese subjects: A randomized controlled trial. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- López-Tenorio, I.I.; Domínguez-López, A.; Miliar-García, Á.; Escalona-Cardoso, G.N.; Real-Sandoval, S.A.; Gómez-Alcalá, A.; Jaramillo-Flores, M.E. Modulation of the mRNA of the Nlrp3 inflammasome by Morin and PUFAs in an obesity model induced by a high-fat diet. Food Res. Int. 2020, 137. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, S.; Lambert, J.D. Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. Eur. J. Nutr. 2014, 53, 149–158. [Google Scholar] [CrossRef]
Figure 1.
Schematic adipogenesis process. A cascade of transcription factors is activated to induce or inhibit the PPAR-γ and C/EBPα expression, which are the key proteins in adipogenesis. Black narrow indicates induce and red inhibit.
Figure 1.
Schematic adipogenesis process. A cascade of transcription factors is activated to induce or inhibit the PPAR-γ and C/EBPα expression, which are the key proteins in adipogenesis. Black narrow indicates induce and red inhibit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



