Submitted:
26 January 2024
Posted:
29 January 2024
You are already at the latest version
Abstract
This study investigated the impact of seven-strain probiotic supplementation on the frequency of T helper 17 (Th17) cells, regulatory T (T-reg) cells, Treg/Th17 ratio, and their gene expression in individuals with severe traumatic brain injury (STBI). Probiotics can influence the gut microbiota, modulate the immune response, and prevent or alleviate certain immune diseases. However, it is necessary to clarify whether probiotics can effectively impact Th cells in STBI patient. Despite the great attention to improving the immune system in most diseases, there is currently no accurate mechanism for the use of probiotics in diseases with damage to the head injury, which can be a basis for the use of probiotics. The primary objective of this study is to address this research gap. The present study was a randomized, double-blind, placebo-controlled trial that enrolled 48 STBI patients. They were treated with probiotics or placebo: 4 capsule/day for 14 days. The pre- and post-study evaluation included examining the frequency of Treg, Th17 cells, as well as the expression of associated immunological parameters (Foxp3 and RORγt) and Treg/Th17 ratio in both groups. No significant differences were found between the two groups in terms of subject characteristics (P>0.05). GCS scores at 14 days were significantly higher in the probiotic compared to the placebo group. The probiotic group showed a significant decrease in Th17 cell and RORγt gene expression (P = 0.022 and P< 0.001, respectively), a notable increase in T-reg and FoxP3 gene expression (P= 0.025 and P< 0.001, respectively), and a higher Treg/Th17 ratio compared to the placebo group. Our study provides promising evidence of the beneficial effects of seven-strain probiotic supplementation on Th cells in individuals with STBI.
Keywords:
severe traumatic brain injury
; regulatory T cell
; forkhead box protein 3
; T helper 17
; retinoic acid receptor-related orphan receptor gamma t
; gut microbiome
; probiotic
1. Introduction
Traumatic brain injury (TBI) is a complex disorder that influences over 2 million emergency room visits, 280,000 hospitalizations, and 50,000 deaths worldwide annually [1]. TBI causes the Blood-Brain Barrier (BBB), which regulates cellular and molecular exchange between the brain and the intravascular compartment, to become dysfunctional, leading to the leakage of proteins, fluid, and transmigration of immune cells and inflammatory molecules in the brain, triggering a systemic inflammatory response [2]. This inflammatory response contributes to secondary brain injury and worsens neurological outcomes [3].
T helper (Th) cells play a critical role in immune responses, but their impress in the CNS and their contributions to the pathogenesis of TBI are poorly understood [4]. A few studies have shown that Th cells could have a crucial role in TBI pathophysiology and beneficial effects on tissue repair following brain injury [5,6,7]. Different subsets of T cells, such as Th17 and Treg cells, can be implicated in the immune-inflammatory response after TBI [8,9]. These cells can inhibit the activation and proliferation of immune cells through the expression of the retinoic acid receptor-related orphan receptor gamma t (RORγt) and the transcription factor forkhead box protein P3 (Foxp3), respectively [10]. Th17 cells play a crucial role in promoting tissue inflammation, while Treg cells exhibit neuroprotective effects by modulating Th cell function. Tregs play a beneficial role in TBI pathogenesis by suppressing brain inflammation and reducing brain pathology [11]. Imbalances in Th cell subsets such as Th17 and Treg have been associated with the severity of TBI and can affect the outcome and complications of the injury [12]. Overall, there is an infiltration of Th cells into the brain, which can contribute to brain edema and acute damage following TBI. Th17/Treg imbalance has been reported to be associated with the severity of the injury, and the balance between Th17 and Treg is critical for maintaining health by controlling inflammatory and autoimmune disorders in trauma patients [12,13].
In STBI patients, reducing inflammation is essential, as it cannot only hinder the healing process but also increase hospitalization duration and mortality risk. Therefore, effective management of inflammation is crucial [14]. Intestinal microbes can be a major source of systemic infection and immune dysfunction in patients with STBI. In contrast, endogenous probiotic bacteria of the gut, such as Bifidobacterium and Lactobacillus, play a critical role in maintaining the intestinal mucosal barrier and enhancing immune responses [15]. Probiotics have recently been recognized as living microorganisms that play a role in immune balance through their effect on the brain-gut axis, which is a critical pathway for central nervous system (CNS) and gastrointestinal (GI) homeostasis [16] and also have an impact on the differentiation of Tregs and Th17 cells in the context of TBI (Figure 1) [17]. Despite decades of well-designed clinical trials and great scientific advances in the medical management of TBI patients, clear gaps remain in the effectiveness of probiotics on T cells in these patients. The results of in vitro studies showed that Lactobacillus (L) and Bifidobacterium (B) strains modulate immune responses by regulating Th cell responses and increasing the production of Treg and Th17 cells, leading to improvement of intestinal inflammation [18]. In animal models of TBI, studies have found that probiotics can promote the differentiation of Tregs and inhibit the differentiation of Th17 cells, which modulate the immune system and cytokine secretions, such as IL-2, IL-6, and TGF-β, that are involved in the regulation of Tregs and Th17 cells [19,20].
Due to the inflammatory response and the urgent need to reduce inflammation, mortality, and enhance immune function in STBI patients admitted to the ICU, there is a significant gap in the scientific literature regarding the effect of probiotics on the frequency and function of T17 and Treg cells. Since studies examining the effect of probiotics on the Th17 and Treg cells in STBI patients are evidently scarce, this research gap necessitates for comprehensive research to elucidate the potential mechanisms of probiotic supplementation in these patients. Therefore, the main hypothesis of this research is that the supplementation of probiotics leads to a favorable adjustment of the frequency and function of Th17 and Treg cells in patients with STBI.
2. Material and Methods
2.1. Study design
This double-blind, placebo-controlled, randomized clinical trial was conducted in northwest Iran from December 2020 to November 2021. Patients with STBI admitted into two ICU were enrolled in this study. The inclusion criteria were age between 18 and 60 years, suffering from STBI, consciousness based on GCS<8, feeding through a nasogastric tube [21], and being under mechanical ventilation. The excluded criteria were pregnancy, chronic diseases such as metabolic, cardiovascular, kidney, liver, autoimmune diseases, cancers and receiving total parenteral nutrition (TPN) for more than 2 days and being treated by immunosuppressive medications. After a full explanation of the study aims and protocol, a written informed consent was signed by one of the first-degree relatives of the patients.
2.2. Sample size
Based on the mean difference of Treg reported by Min Li [22], β= 0.20 and α= 0.05, the sample size was calculated 24 patients per group, and then, by considering a 15% dropout rate, the sample size was increased to 30 patients pin each group.
2.3. Randomization, Blinding, and Study Protocol
A person not involved in the research randomly assigned 48 patients with STBI into two groups (1:1) using the blocking method. The four-sized blocks were created by STATA 16.0 statistical software and identified by the letters A, B, C and D. The patients in the placebo group (N=24) received four placebo capsules (containing maltodextrin, silicon dioxide, microcrystalline cellulose, and sodium starch glycolate), whereas those in the probiotic group (N=24) received four seven-strain probiotic capsules (1010 CFU) (containing beneficial and safe bacterial strains, L. casei, L. rhamnosus, L. acidophilus, B. breve, L. bulgaricus, B. longum and Streptococcus thermophilus, for 14 days (Figure 2). The capsules had the same smell, taste, and appearance, as well as shape, color, size, and all other aspects that allow allocation concealment. The researcher, clinicians, and assessors were blinded to the study allocation.
2.4. Blood sampling and cell culture
After 12 hours overnight fasting, 10 ml blood samples were collected from each patient on days 1 and 14 of the study. Five ml of blood samples were applied for serum preparation and then stored at – 70∘C until the end of study. PBMCs were separated from remaining heparinized venous blood via density-gradient centrifugation using a 1.077 g/ml Ficol (Lymphosep; Biosera, East Sussex, UK) by 450g centrifugation for 25 min, and followed by twice washing for RPMI-1640 medium (Sigma-Aldrich, Schnelldorf, Germany). Fresh PBMCs parts were used for flow cytometry analysis. The remaining separated cells were cultured in 10% heat-inactivated fetal bovine serum, 200 mM L-glutamine and phorbolmyristate acetate, and 1% penicillin-streptomycin solution, and then, incubated for 48 hours at 37°C and 5% CO2. Cultured cells were then collected for the extraction of total RNA and gene expression assessment using quantitative real-time PCR.
2.5. Flow cytometry analysis
Th17and Treg cell number assessments were performed by Flow cytometry. For stimulation, freshly collected PBMCs were incubated for 5 hours with ionomycin (1 μg/ml) and PMA (25 ng/ml) in the presence of monensin, with the aim of enhancing the intracellular IL-17A stain (1.7 μg/ml; all from eBioscience, San Diego, CA). A 10-minute-long centrifugation at 300g provided a pellet, which was washed with FACS (Fluorescence-Activated Cell Sorting) buffer. Subsequently, a 15-minute-long incubation at 4°C was performed on the cells with anti-CD4-FITC (BD Biosciences, San Jose, CA), which were then washed and permeabilized with fixation/permeabilization buffer (eBioscience). For intracellular staining, a 20-minute-long exposure of the cells to phycoerythrin (PE)- labeled anti human IL-17A (eBioscience) was performed at room temperature. To evaluate the number of Treg cells, a 15-minute-long incubation at 4°C was performed with FITC-labeled anti-human CD4 (BD Biosciences, San Jose, CA), PE-labeled anti-human CD25, and APC conjugated anti-human CD127 (BD Biosciences) antibodies. Next, twice washing of the cells and re-suspension in FACS solution (phosphate-buffered saline) were done, following which the analyses were performed on the same day using a FACS Calibur (BD Biosciences) flow cytometer. FACSCalibur flow cytometry, and FlowJo software (Tree Star, Ashland, OR) analyzed the data. Lymphocyte gating was accomplished according to their forward and side scatters.
2.6. RNA extraction and the synthesis of complementary DNA (cDNA)
Following sample homogenization, total RNA extraction from PBMCs was accomplished using RNX-PLUS Solution (SinaClon, Tehran, Iran). Next, RNA quantity and quality evaluations were done in 260/280 and 260/230 wavelength ratios by spectrophotometer (Nano Drop; Agilent Technologies, Santa Clara, CA). cDNA synthesis from mRNAs was done using random hexamers, oligo(dT) primers, and the M-MLV Revert Aid Reverse Transcriptase Kit (Thermo Fisher, Waltham, MA) for the evaluation of cytokine transcription factors (RORγt and FoxP3).
2.7. Statistical analysis
The Kolmogorov–Smirnov test was implemented for assessing distribution of continuous variables using Graph Pad Prism (ver. 7.00 for Windows; Graph Pad Software, La Jolla, CA; www.graphpad.com). Descriptive continuous data was reported as the mean ± SD (standard deviation). Between-group comparison was performed using an independent samples t-test. Moreover, paired samples t-test was used for intra-group comparisons. The Between-group differences in qualitative data was assessed using the Chi-square test. At the end of the trial, no effect of the intervention was assessed using analysis of covariance (ANCOVA) to adjust for the effect of baseline values and confounding factors including gender, age, baseline BMI, and baseline GCS. P < 0.05 was considered as statistically significant.
3. Results
3.1. Clinical and anthropometric information
Forty- eight out of sixty subjects were randomly allocated into the study groups (probiotic group, N= 24; placebo group, N= 24) and completed the study. Three patients from each placebo and probiotic group were excluded due to unrelated to the intervention (food intolerance, gastrointestinal bleeding, and death) during the study.
The clinical and anthropometric information of the patients is presented in Table 1. Furthermore, there were no significant between- group differences in terms of sex, age, weight, and BMI. Weight and BMI did not also change after the study. Although GCS score increased in both groups; however, the increases in GCS were statistically significant in the probiotic group after intervention for 14 days (p= 0.008).
3.2. Th17 cell frequency
Figure 3 shows that there was no significant inter-group difference in Th17 cell frequency at baseline (p > 0.05). This shows that the randomization process was successfully achieved in two groups before the intervention. Although Th17 cells frequency decreased significantly in the probiotic group (p = 0.022), no statistical change was found in the placebo group (p = 0.061). After the trial and adjusted for baseline and GCS, an inter-group significant difference in Th17 cell frequency was observed (p = 0.045). This evidence shows that a seven-strain probiotic can reduce pro-inflammatory Th17 cells and has immunomodulatory effects during the 14-day supplementation period in STBI patients.
3.3. T-reg cell frequency
No significant difference in Treg cell frequency was found between the groups at baseline (p < 0.05) (Figure 4). Although Treg cells frequency increased significantly in the probiotic group (p = 0.025) but no change was observed in the placebo group (p = 0.057). At day 14 and adjusted for baseline and GCS,there was a significant difference inter-group in Treg cell frequency (p = 0.047). This provides evidence that probiotic supplementation can effectively increase levels of systemic anti-inflammatory Treg cells in patients with STBI.
3.4. The Treg/ Th17 ratio
As Figure 5 demonstrates, the Treg/Th17 ratio did not differ considerably between the groups at baseline. Over the trial, there were significant increases in the Treg/ Th17 ratio in both groups. A significant difference in Treg/ Th17 ratio was found between the groups (P< 0.001); and around 39.5% increase in the probiotic group compared placebo group. Analysis of the integrated Treg/Th17 ratio provides further evidence that probiotics modulate systemic inflammatory responses by favoring anti-inflammatory Treg cells over Th17 cells. This immunomodulation and suppression inflammation is a key mechanism underlying the neurological benefits of probiotics in STBI.
3.5. FoxP3 and RORγt expression
Figure 6 shows that RORγt expression decreased while FoxP3 expression increased in the probiotic group. After adjusting the baseline values and GCS, a significant between- group differences in RORγt and FoxP3 expression were found at the end of the study (p< 0.001 and p< 0.001, respectively). Analysis of T cell transcription factors shows that probiotic supplementation alters the balance of FoxP3 and RORγt to shift T cell differentiation away from inflammatory Th17 cells and toward anti-inflammatory Treg cells. This transcriptional reprogramming is likely an immune mechanism to improve cellular immunity in STBI.
4. Discussion
To the best of knowledge, the present trial appears to be the first study assessing the effects of probiotic supplementation on Th17 and Treg cells in patients with STBI. We found that the administration of seven-strain probiotic supplementation in STBI patients decreased Th17 cell frequency and mRNA expression of RORγt, as well as increased Treg cell frequency and mRNA expression of FoxP3, indicating the important role of Th17 and Treg cell balance in suppressing autoimmune responses.
In the present study, a significant effect of probiotics on the decrease in Th17 frequency and increase frequency of Treg cells was observed. Recent studies have reported a direct correlation between the frequency of Treg cells in circulation and neurological recovery in both animal models and TBI patients [22,23]. Kipnis et al. have shown that introducing Treg cells from an external source into an animal model with a weakened immune system after a CNS injury can lead to neuroprotection [24]. Reducing the presence of Th17 cells with pro-inflammatory properties and increasing the presence of Treg cells with anti-inflammatory properties could be a potential strategy for developing inflammation treatments in TBI patients [25]. Despite consuming immune-modulatory therapies, TBI subjects confront severe inflammatory attacks during the disease [26]. Furthermore, these treatments are associated with intolerable side effects, and thereby leading to searching for an alternative preventive medicine [27]. Probiotics may reduce brain inflammation by reducing Th17 cell responses. This controlled trial provides evidence that a short 2-week course of probiotic supplementation can significantly reduce the frequency of circulating pro-inflammatory Th17 cells in patients with STBI [28,29]. Several mechanisms have been proposed to inhibit inflammation by probiotics, which can reduce the inflammation process by suppressing Th17 activity, inducing Th17 cells, and enriching Tregs in inflamed areas through their experience in modulating DC functions and the signaling pathway of Toll-like receptors (TLR).The primary response to these bacteria is triggered by pattern recognition receptors (PRR), which bind pathogen-associated molecular patterns (PAMP). PRR comprises TLR, nucleotide-binding oligomerization domains, adhesion molecules and lectins [30]. The results of a study by K. Zhou et al. showed that the number of Tregs in the brain increases after intracerebral hemorrhage (ICH). Deletion of Tregs using a CD25 antibody or Foxp3 DTR-mice led to neurological deficient scores (NDS), higher levels of inflammatory factors, larger hematoma volumes, and increased neuronal degeneration. Conversely, boosting Tregs using a CD28 super-agonist antibody reduced the inflammatory injury. Additionally, Tregs depletion led to a shift in microglia/macrophage polarization towards the M1 phenotype, while promoting Tregs shifted this transition towards the M2 phenotype. Therefore, Tregs played a role in mitigating ICH-induced inflammatory injury by modulating microglia/macrophage polarization towards the M2 phenotype via the IL-10/GSK3b/PTEN axis [31]. So far, there have been no studies that specifically investigate the effects of probiotics on T cells in STBI, so more studies are needed.
In this study, a 39.5% increase in Treg/ Th17 ratio was found between the two groups. As the balance between Th17 and Treg cells is crucial for regulating immune responses [32], therefore, Th17/Treg cells’ imbalance seems to play a significant role in the progression of inflammatory responses [33]. Gupta et al. have suggested that a higher ratio of Th17 cells to Treg could result in more severe post-traumatic complications and worsen acute and chronic inflammatory reactions to TBI. Therefore, to manage the secondary damage caused by TBI, it may be beneficial to regulate the balance between Th17 and Treg cells [12]. The ratio of anti-inflammatory Treg cells to pro-inflammatory Th17 cells provides an integrated measure of the overall inflammatory balance [34]. Our findings should be interpreted in light of the following points. First, it is possible that probiotic supplementation in the current study had a small protective role, which was not detected. Second, the lack of significant effect of probiotics on Th cells possibly resulted from the STBI in which inflammatory response leads to irreversible damage. Third, it should be kept in mind that studies were performed on specialized populations with different health conditions and the effect of probiotics may be different on each of the diseases. Fourth, conflicting results of studies may be related to alternate dose, genus, species, or strain of probiotics.
In the present study, a significant effect of probiotics on decreased of RORγt expression and FoxP3 increased of expression in the probiotic group was observed. Cui et al. showed, in a rat model of post-surgical brain injury, that oral administration of brain proteins and probiotics significantly reduced CD4+T/CD8+T ratios and induced T-cell differentiation into CD4+CD25+Foxp3+Treg cells. This treatment could decrease secondary inflammatory responses following craniocerebral trauma [17]. The study of Kwon, Ho-Keun, et al. showed that the intervention effects of the probiotics mixture (IRT5) on mice at a concentration of 5 × 109 CFU of each of the following strains: BB, LA, ST, LC, and Lactobacillus leuteri were: significantly increased the level of CD4 + Foxp3 + Tregs in the mesenteric lymph node (MLN) by augmenting Foxp3 + levels in CD4 + CD25 -T cells, significantly increased Foxp3 mRNA expression, induced hypoproliferative CD4 + T cells and B cells without inducing apoptosis [35].
This study has several strengths. This is the first randomized controlled clinical trial that investigated the effects of probiotic supplementation on Th17 and Treg in STBI patients. This trial has several limitations. First, a fixed dosage of probiotic was used in this study and higher doses or longer administration times may have produced different results. Second, enrollment was limited to TBI patients and inclusion of patients from other ICU settings may have yielded different results. Third, the relatively small sample size limited our ability to perform subgroup analysis based on disease severity and other demographic characteristics. Fourth, clinicians and research staff were aware of supplement assignments. Fifth, some patients were lost to follow-up, but we used ITT analysis to address this problem.This study has some limitations such as not considering the effect of commonly used medications, the relatively short duration of the study, and the synergistic effect of the strains used, nevertheless, the administration of high dose and multi-strain of probiotics is considered as the strengths of the study.
5. Conclusion
It is concluded that seven- strain probiotics as an adjunctive therapy- in patients with STBI are encouraging and suggest that administration of probiotics in STBI patients is associated with improvement of cellular immunity function in relation to matched placebo-treated patients: They significantly reduce the levels of Th17 pro-inflammatory. Furthermore, a significant increase in Treg could be detected. Therefore, probiotics could reduce inflammation in STBI patients and might be considered as an adjunctive therapy in the treatment of STBI patients. However, further clinical trials for a longer duration, using various types of probiotics on a larger sample size, are required.
References
- Katzenberger, R.J., B. Ganetzky, and D.A. Wassarman, The gut reaction to traumatic brain injury. Fly (Austin), 2015. 9(2): p. 68-74. [CrossRef] [PubMed]
- Moretti, R., et al., Blood-brain barrier dysfunction in disorders of the developing brain. Frontiers in neuroscience, 2015. 9: p. 40. [CrossRef] [PubMed]
- Witcher, K.G., D.S. Eiferman, and J.P. Godbout, Priming the inflammatory pump of the CNS after traumatic brain injury. Trends in neurosciences, 2015. 38(10): p. 609-620. [CrossRef] [PubMed]
- Corps, K.N., T.L. Roth, and D.B. McGavern, Inflammation and neuroprotection in traumatic brain injury. JAMA neurology, 2015. 72(3): p. 355-362. [CrossRef] [PubMed]
- Hofstetter, H.H., et al., Autoreactive T cells promote post-traumatic healing in the central nervous system. Journal of neuroimmunology, 2003. 134(1-2): p. 25-34. [CrossRef] [PubMed]
- Jones, T., E. McDaniel, and P. Popovich, Inflammatory-mediated injury and repair in the traumatically injured spinal cord. Current pharmaceutical design, 2005. 11(10): p. 1223-1236. [CrossRef]
- Popovich, P.G. and T.B. Jones, Manipulating neuroinflammatory reactions in the injured spinal cord: back to basics. Trends in pharmacological sciences, 2003. 24(1): p. 13-17. [CrossRef] [PubMed]
- Morgan, M.E., et al., Effective treatment of collagen-induced arthritis by adoptive transfer of CD25+ regulatory T cells. Arthritis & Rheumatism, 2005. 52(7): p. 2212-2221. [CrossRef] [PubMed]
- Paulos, C.M., et al., Microbial translocation augments the function of adoptively transferred self/tumor-specific CD8+ T cells via TLR4 signaling. The Journal of clinical investigation, 2007. 117(8): p. 2197-2204. [CrossRef]
- Mickael, M.E., S. Bhaumik, and R. Basu, Retinoid-related orphan receptor RORγt in CD4+ T-Cell–mediated intestinal homeostasis and inflammation. The American Journal of Pathology, 2020. 190(10): p. 1984-1999. [CrossRef]
- Xu, X., et al., Anti-inflammatory and immunomodulatory mechanisms of atorvastatin in a murine model of traumatic brain injury. Journal of neuroinflammation, 2017. 14(1): p. 1-15. [CrossRef] [PubMed]
- Gupta, D.L., et al., Coexistence of Th1/Th2 and Th17/Treg imbalances in patients with post traumatic sepsis. Cytokine, 2016. 88: p. 214-221. [CrossRef] [PubMed]
- Luo, T., et al., Th17/Treg imbalance induced by dietary salt variation indicates inflammation of target organs in humans. Scientific reports, 2016. 6(1): p. 26767. [CrossRef] [PubMed]
- Lucas, S.M., N.J. Rothwell, and R.M. Gibson, The role of inflammation in CNS injury and disease. British journal of pharmacology, 2006. 147(S1): p. S232-S240. [CrossRef] [PubMed]
- Yang, W., et al., Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury. Microorganisms, 2022. 10(10): p. 2082. [CrossRef] [PubMed]
- Caputi, V. and M.C. Giron, Microbiome-gut-brain axis and toll-like receptors in Parkinson’s disease. International journal of molecular sciences, 2018. 19(6): p. 1689. [CrossRef] [PubMed]
- Cui, Y., et al., Orally administered brain protein combined with probiotics increases Treg differentiation to reduce secondary inflammatory damage following craniocerebral trauma. Frontiers in Immunology, 2022. 13: p. 928343. [CrossRef] [PubMed]
- Ng, S., et al., Mechanisms of action of probiotics: recent advances. Inflammatory bowel diseases, 2009. 15(2): p. 300-310. [CrossRef]
- Celorrio, M., et al., Innate and Peripheral Immune Alterations after Traumatic Brain Injury Are Regulated in a Gut Microbiota-Dependent Manner in Mice. Journal of neurotrauma, 2023. 40(7-8): p. 772-787. [CrossRef]
- Liu, M., J. Mao, and S. Zhang, Effect of intervention of probiotics in advance on Treg/Th17 in premature mice. BioMed Research International, 2022. 2022. [CrossRef]
- Vonder Haar, C., et al., Frontal traumatic brain injury increases impulsive decision making in rats: a potential role for the inflammatory cytokine interleukin-12. Journal of neurotrauma, 2017. 34(19): p. 2790-2800. [CrossRef] [PubMed]
- Li, M., et al., Role of regulatory T cell in clinical outcome of traumatic brain injury. Chinese medical journal, 2015. 128(08): p. 1072-1078. [CrossRef] [PubMed]
- Yu, Y., et al., Regulatory T cells exhibit neuroprotective effect in a mouse model of traumatic brain injury Retraction in/10.3892/mmr. 2017.6299. Molecular medicine reports, 2016. 14(6): p. 5556-5566. [CrossRef] [PubMed]
- Kipnis, J., et al., Dual effect of CD4+ CD25+ regulatory T cells in neurodegeneration: a dialogue with microglia. Proceedings of the National Academy of Sciences, 2004. 101(suppl_2): p. 14663-14669. [CrossRef]
- Bao, W., Y. Lin, and Z. Chen, The peripheral immune system and traumatic brain injury: Insight into the role of T-helper cells. International Journal of Medical Sciences, 2021. 18(16): p. 3644. [CrossRef]
- Bigler, E.D., Neuroimaging biomarkers in mild traumatic brain injury (mTBI). Neuropsychology review, 2013. 23: p. 169-209. [CrossRef] [PubMed]
- Muhammad, S., et al., Brain immune interactions—novel emerging options to treat acute ischemic brain injury. Cells, 2021. 10(9): p. 2429. [CrossRef] [PubMed]
- Suganya, K. and B.-S. Koo, Gut–brain axis: role of gut microbiota on neurological disorders and how probiotics/prebiotics beneficially modulate microbial and immune pathways to improve brain functions. International journal of molecular sciences, 2020. 21(20): p. 7551. [CrossRef]
- Zhu, C.S., et al., A review of traumatic brain injury and the gut microbiome: insights into novel mechanisms of secondary brain injury and promising targets for neuroprotection. Brain sciences, 2018. 8(6): p. 113. [CrossRef]
- Gómez-Llorente, C., S. Munoz, and A. Gil, Role of Toll-like receptors in the development of immunotolerance mediated by probiotics. Proceedings of the Nutrition Society, 2010. 69(3): p. 381-389. [CrossRef]
- Zhou, K., et al., RETRACTED: Regulatory T cells ameliorate intracerebral hemorrhage-induced inflammatory injury by modulating microglia/macrophage polarization through the IL-10/GSK3β/PTEN axis. Journal of Cerebral Blood Flow & Metabolism, 2017. 37(3): p. 967-979. [CrossRef]
- Zhang, W., et al., Transcriptional and posttranslational regulation of Th17/Treg balance in health and disease. European Journal of Immunology, 2021. 51(9): p. 2137-2150. [CrossRef] [PubMed]
- Weaver, C.T., et al., The Th17 pathway and inflammatory diseases of the intestines, lungs, and skin. Annual Review of Pathology: Mechanisms of Disease, 2013. 8: p. 477-512. [CrossRef] [PubMed]
- Liu, Y.-J., et al., Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner. Theranostics, 2020. 10(12): p. 5225. [CrossRef]
- Kwon, H.-K., et al., Generation of regulatory dendritic cells and CD4+ Foxp3+ T cells by probiotics administration suppresses immune disorders. Proceedings of the National Academy of Sciences, 2010. 107(5): p. 2159-2164. [CrossRef]
- Kreymann, K., et al., ESPEN guidelines on enteral nutrition: intensive care. Clinical nutrition, 2006. 25(2): p. 210-223. [CrossRef] [PubMed]
- McMahon, P.J., et al., Symptomatology and functional outcome in mild traumatic brain injury: results from the prospective TRACK-TBI study. Journal of neurotrauma, 2014. 31(1): p. 26-33. [CrossRef] [PubMed]
Figure 1.
Interplay between the Gut-Brain Axis, TBI, and Probiotics.
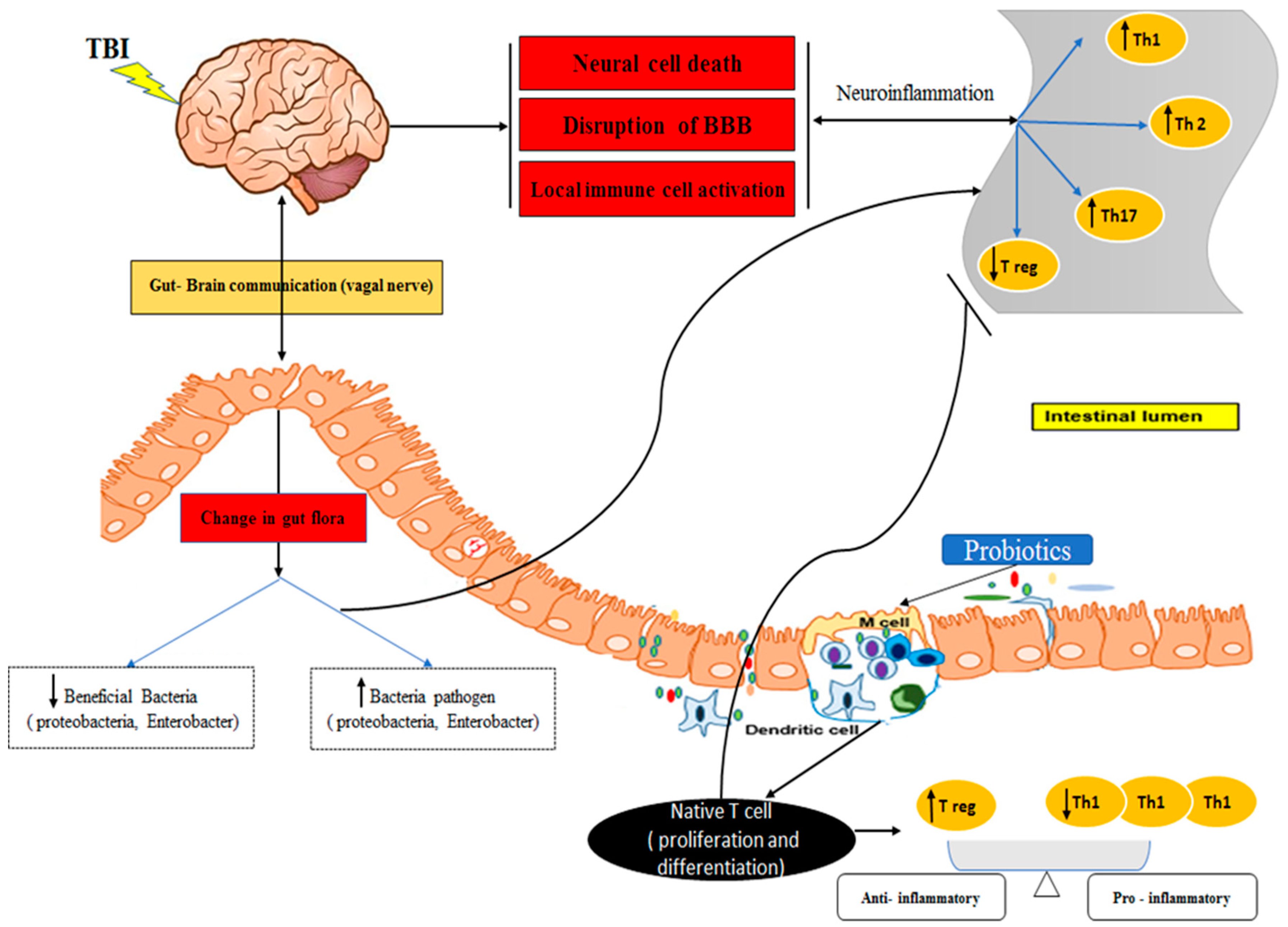
Figure 2.
Study flow chart (EN: Enteral Nutrition).
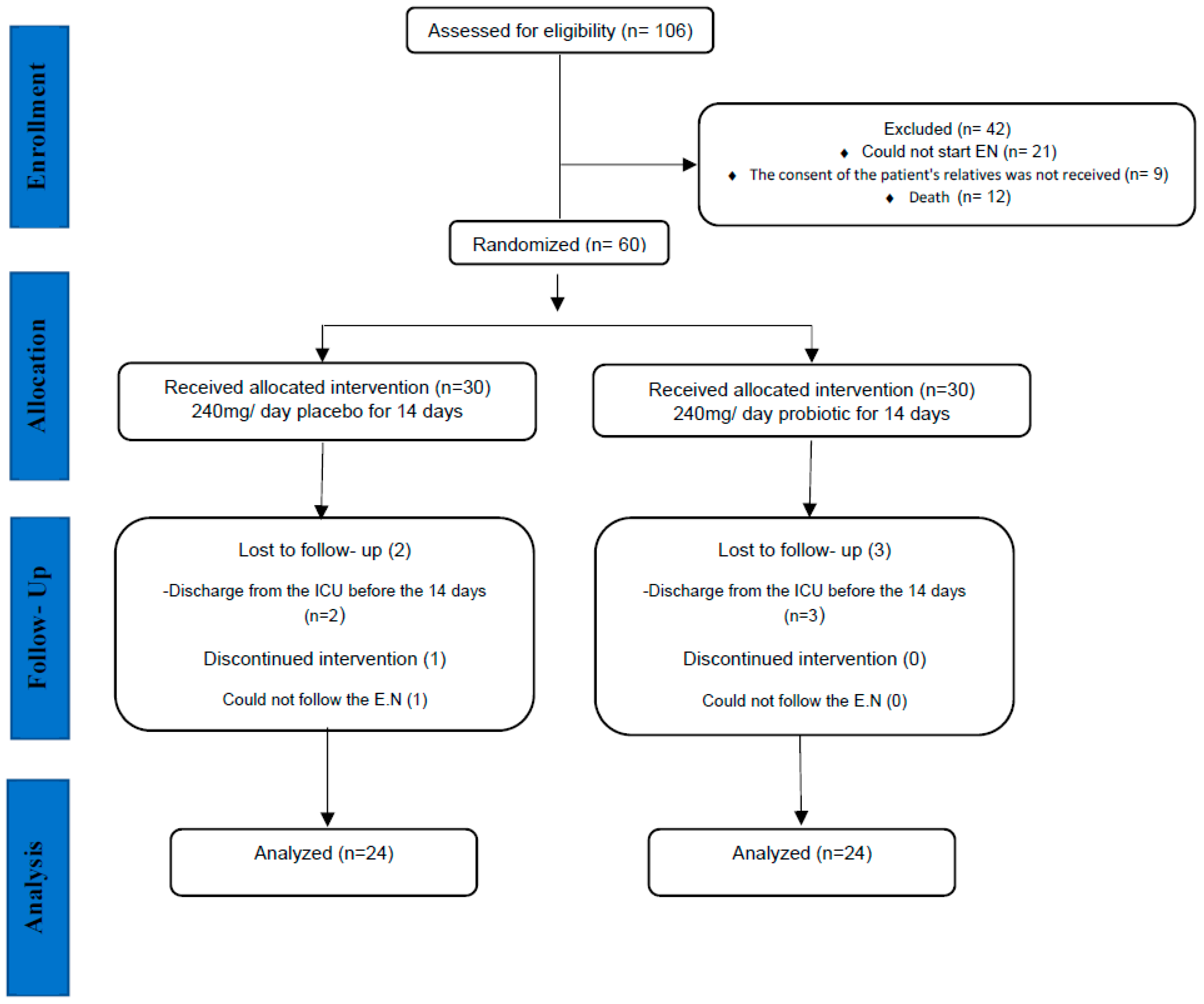
Figure 3.
Representative flow cytometry plots for identifying CD4+ Th 17 cells in whole blood and percentage of circulating Th 17 cells in study groups.
Figure 3.
Representative flow cytometry plots for identifying CD4+ Th 17 cells in whole blood and percentage of circulating Th 17 cells in study groups.
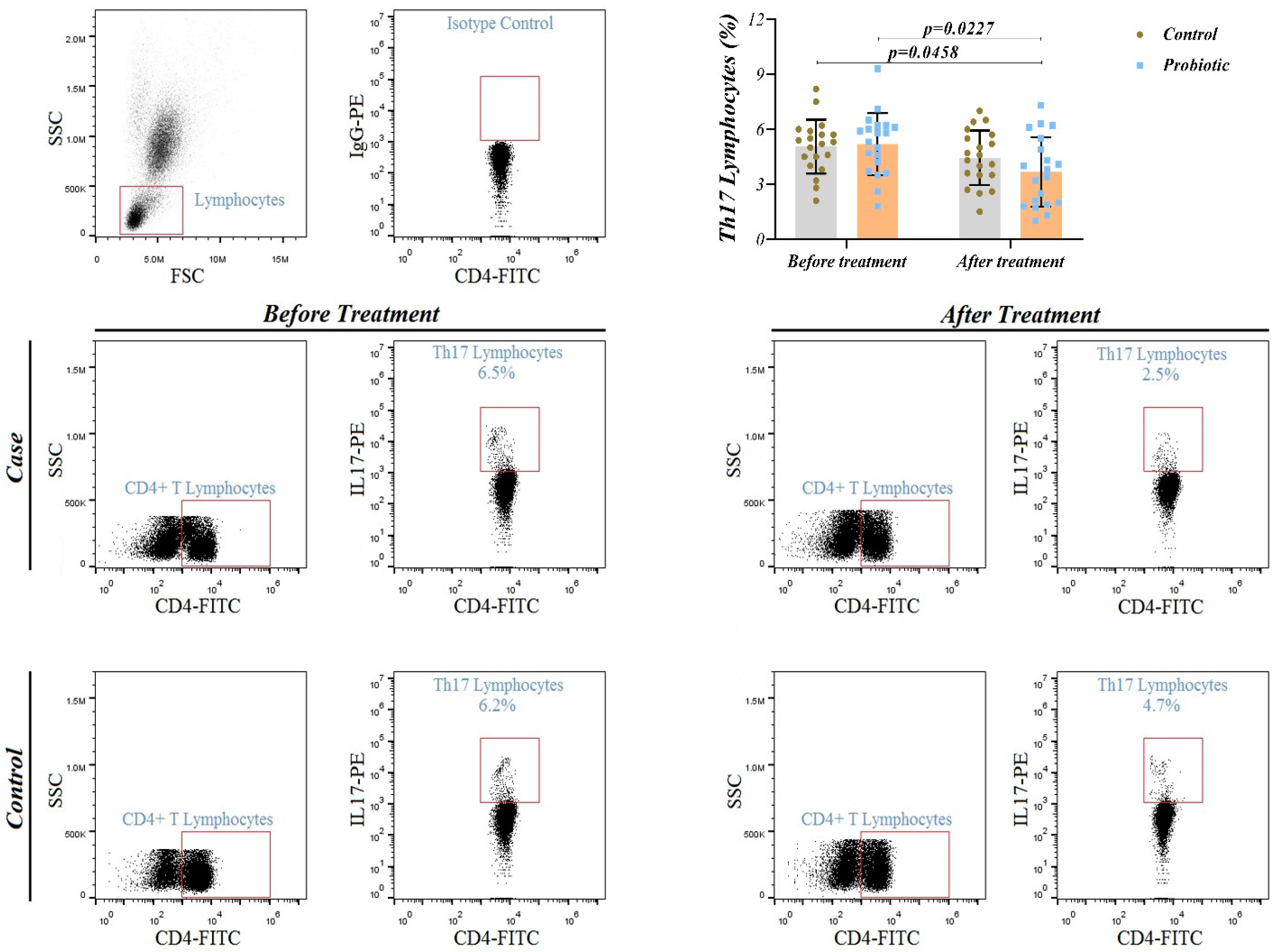
Figure 4.
Representative flow cytometry plots for identifying CD4+ CD25+ regulatory T (T-reg) cells in whole blood and percentage of circulating T-reg cells in study groups.
Figure 4.
Representative flow cytometry plots for identifying CD4+ CD25+ regulatory T (T-reg) cells in whole blood and percentage of circulating T-reg cells in study groups.
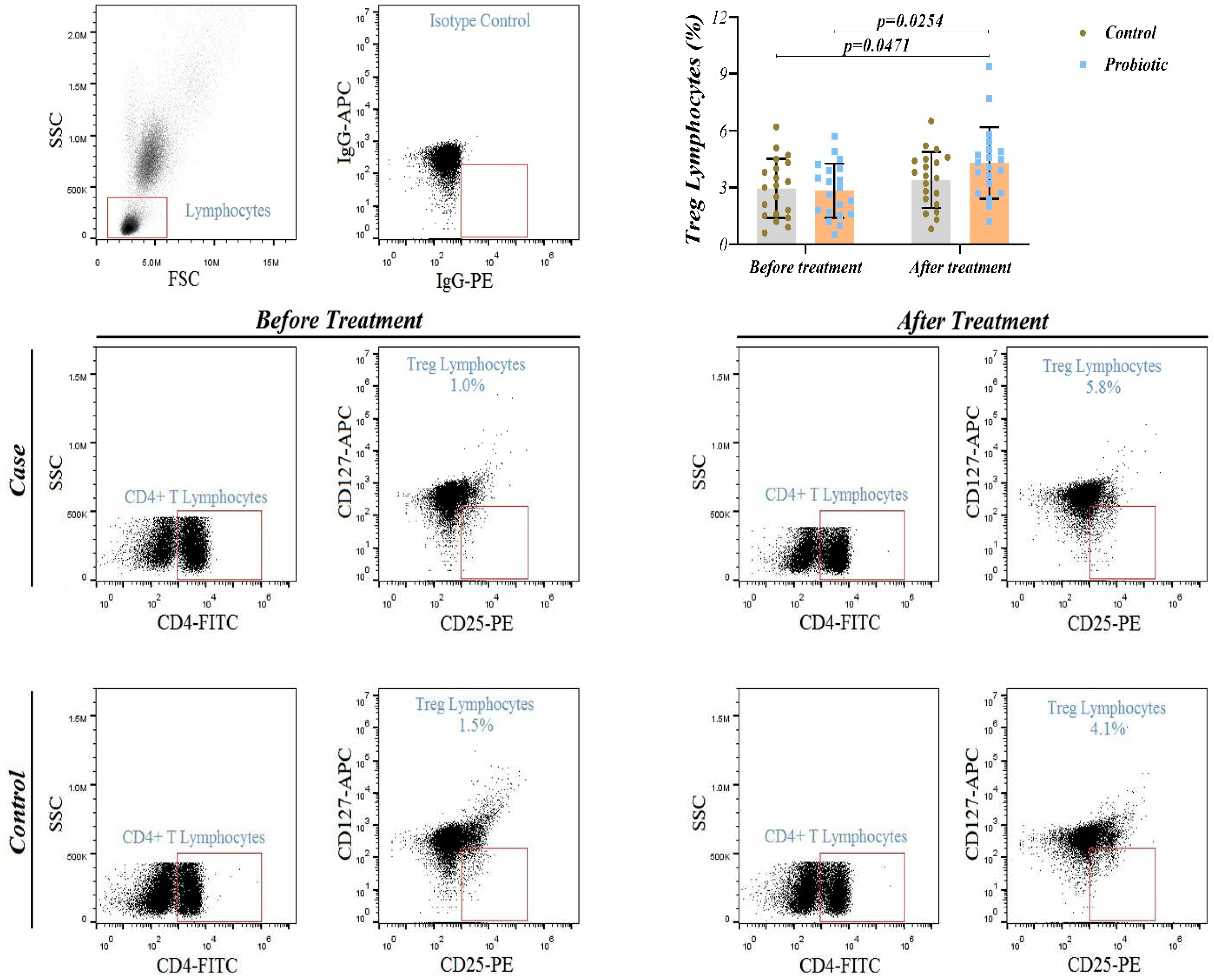
Figure 5.
Treg/Th17 ratio in control and probiotic group. Higher ratio of Treg /Th17 was observed in probiotic group enrolled in our study, compared to control group (P < 0.001).
Figure 5.
Treg/Th17 ratio in control and probiotic group. Higher ratio of Treg /Th17 was observed in probiotic group enrolled in our study, compared to control group (P < 0.001).
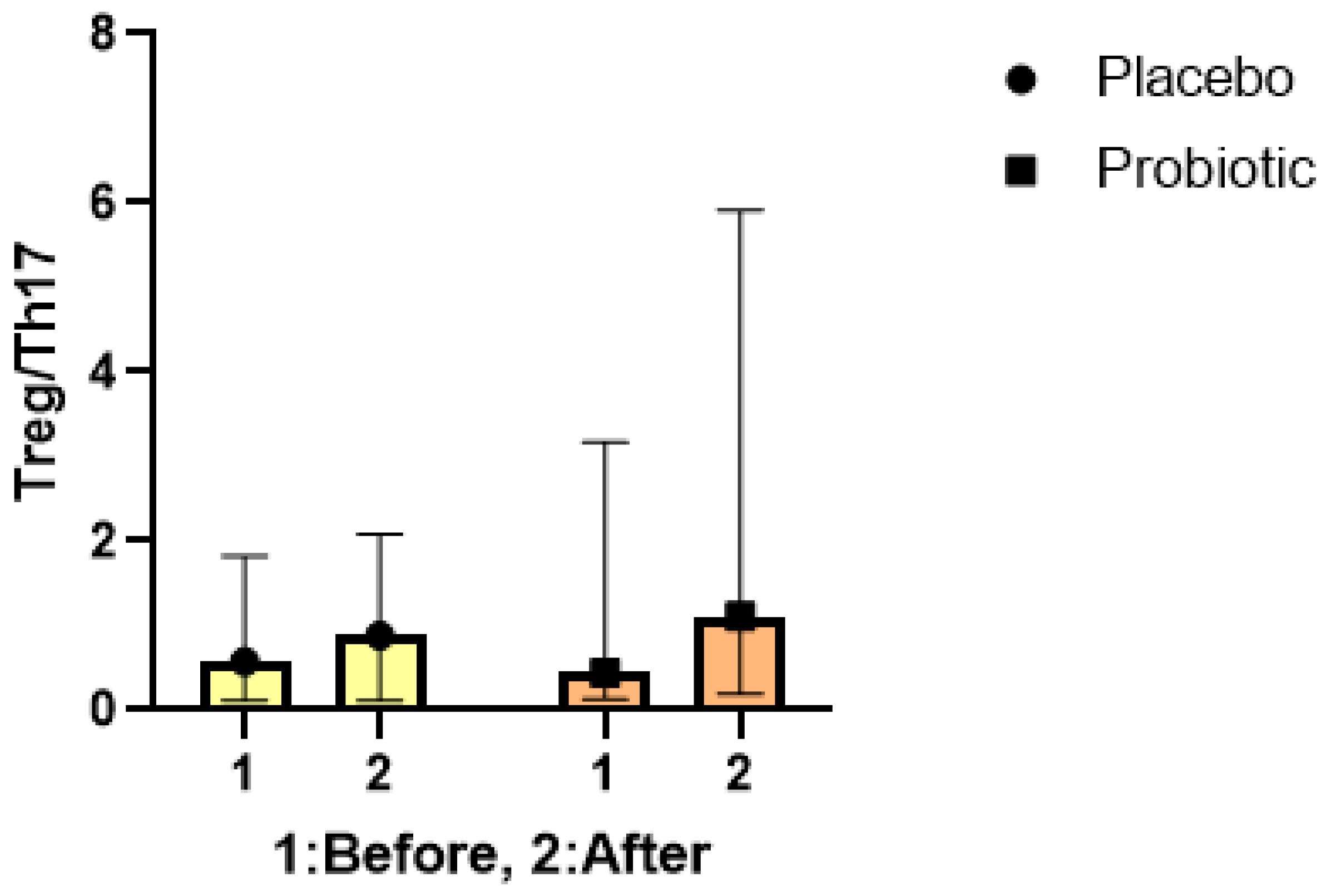
Figure 6.
The expression levels of Th 17 cells- related orphan receptor gamma t (RORγt) and regulatory T cells-related transcription factor FoxP3.
Figure 6.
The expression levels of Th 17 cells- related orphan receptor gamma t (RORγt) and regulatory T cells-related transcription factor FoxP3.
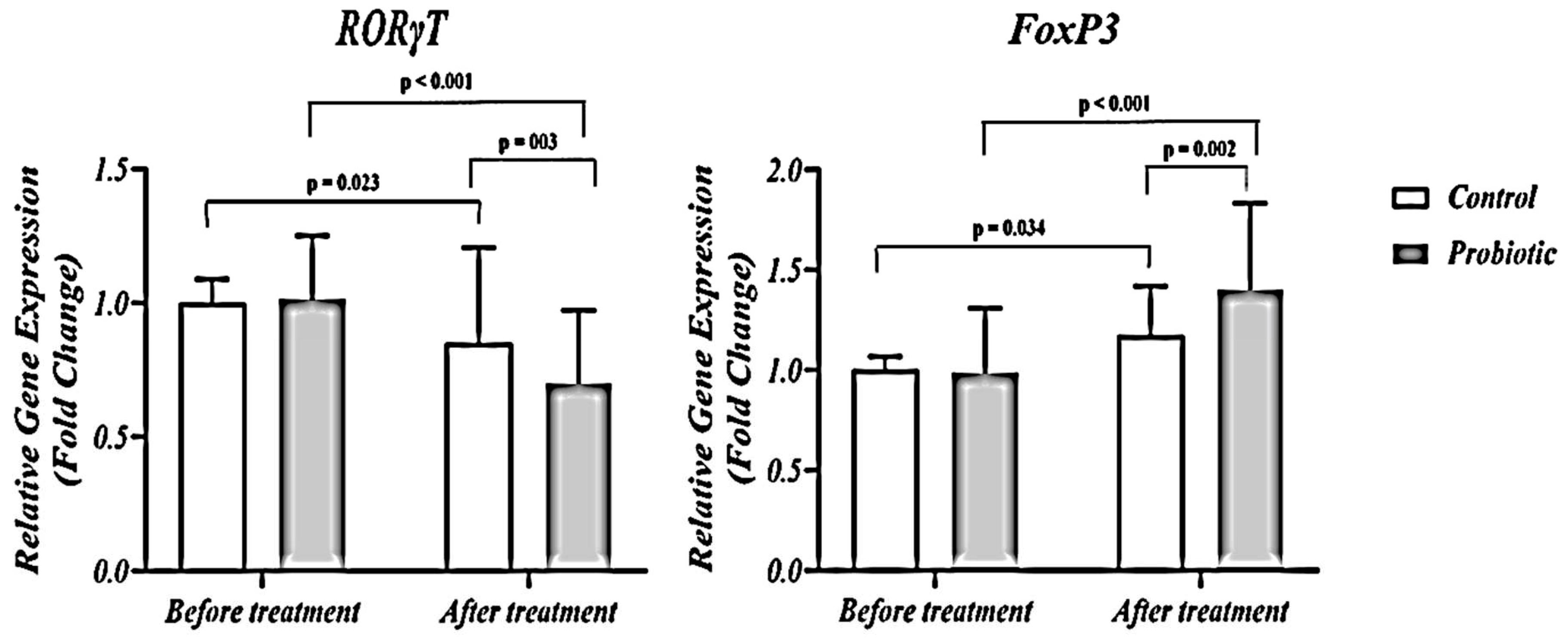

Table 1.
Clinical and anthropometric information for all patients.
| Probiotic (n=24) | Placebo (n=24) | Pc | |
|---|---|---|---|
| Age (years) a | 36.08 (13.65) | 39.33 (16.32) | 0.059 |
| Gender, n (%) b | |||
| Males | 20 (83.30) | 17 (70.80) | 0.073 |
| Females | 4 (16.70) | 7 (29.20) | 0.054 |
| Marital status, n (%) b | 11 (45.80) | 13 (54.20) | 0.062 |
| BMI(Kg/m2) a | 24.17 (4.21) | 24.94 (4.50) | 0.648 |
| MAC (cm) a | 27.00 (2.68) | 27.16 (3.05) | 0.929 |
| TSF (cm) a | 17.70 (4.55) | 16.25 (4.96) | 0.853 |
| GCS score a | 5.21 ± 1.37 | 7.01 ± 1.94 | 0.008 |
a Values are expressed as means (SD) based on independent sample t-test. b Values are expressed as numbers (percent) based on chi-square. c Independent t-test. BMI: Body mass index; MAC: Mid-arm circumference; TSF: Triceps skinfold thicknes; GCS: Glasgow Coma Scale.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



