
Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
Glucosinolates, the main secondary metabolites accumulated in cruciferous flora, have a major impact on fortifying plant immunity against diverse pathogens. Although Botrytis cinerea exhibits varying sensitivity to these compounds, current research has yet to fully understand the intricate mechanisms governing its response to glucosinolates. Different species of the genus Botrytis were exposed to glucosinolate-breakdown products revealing that B. fabae, B. deweyae and B. convoluta, species lacking the mfsG transporter gene (Bcin06g00026), were more sensitive to glucosin-olate-breakdown products than Botrytis species containing that gene such as B. cinerea, B. pseudo-cinerea and B. byssoidea. This finding was further corroborated by the inability of species lacking the mfsG gene to infect plants with a high concentration of glucosinolate breakdown products. These results challenge established correlations, revealing varying aggressiveness on different plant substrates. Expression analysis highlighted the gene's induction in the presence of isothiocyanate, and bioinformatic investigation identified homologous genes in other Botrytis species. Our study underscored the importance of advanced biotechnology to help understand these proteins and thus offer innovative solutions for agriculture
Keywords:
Fungi
; Botrytis
; Pathogenicity
; Glucosinolates
1. Introduction
One of the fundamental strategies in plant defence against pathogenic microorganisms involves the synthesis of phytoalexins [1,2,3] and phytoanticipins [4,5]. Compounds like camalexin and glucosinolates (GLSs), derived from amino acids [6], are notable examples. Pathogens employ diverse pathogenetic and virulence strategies to neutralize or eradicate these phytoanticipins and phytoalexins [5,7,8]. Fungal species, in particular, utilize enzymatic pathways for the effective detoxification of these compounds. Additionally, active efflux mechanisms contribute to the elimination process, an example being the way Botrytis cinerea handles phytoalexins such as resveratrol and eugenol [7,8,9,10,11].
Glucosinolates (GLSs) are a structurally well-defined group of anionic natural products which serve as vital defence compounds in plants belonging to the Brassicales order that include various brassicaceous vegetables such as cabbage, radish, and broccoli [12]. These GLSs undergo hydrolysis facilitated by specific myrosinase enzymes known as thio-β-glucosidases. This hydrolysis process leads to the formation of unstable aglycones, yielding primarily isothiocyanates (ITCs), with the common structure R-N=C=S (Figure 1), nitriles R-CN, or thiocyanates R-S=C=N. In this process the specific type of GLSs, the local environment, and the presence of specifier proteins have a bearing on the outcome [13,14]. The standard product of the reaction is isothiocyanate; the other two compounds mainly occur in the presence of specialized plant proteins that alter the final product of the reaction [13]. These resulting breakdown products operate as deterrents or toxins against potential aggressors. Notably, GLSs and myrosinases are segregated within distinct cellular compartments and only interact upon tissue disruption, such as that caused by chewing insects, triggering the hydrolysis process [15].
In various fungal species, the active efflux mediated by ATP-binding cassette (ABC) or major facilitator superfamily (MFS) transporters serves as a defence mechanism against an array of toxic compounds encompassing secondary metabolites, antibiotics, and fungicides [10,16,17]. Although essential to plant pathogenicity, exploration of ABC transporters in pathogenesis remains sparce. Studies include Magnaporthe grisea ABC1, ABC3, and ABC4 in rice [18,19,20], B. cinerea atrB in grapevine and Arabidopsis thaliana [10,21], Gibberella pulicaris ABC1 in potato [22], and Mycosphaerella graminicola atr4 in wheat [23].
B. cinerea, a necrotrophic plant pathogen with a broad range of hosts (more than 500 genera of plants), has a devastating effect on many plants, including economically significant cruciferous crops. It is a grey mould causing substantial economic losses during crop growth and postharvest stages [24,25]. Effective management of B. cinerea relies heavily on repeated applications of chemical fungicides which accelerates the development of fungicide resistance [26,27,28]. Therefore, deciphering the mechanisms employed by this pathogen to neutralize its host’s defence systems not only sheds light on fundamental cellular processes but also paves the way for innovative strategies to efficiently manage this pathogen.
Recent studies have shown important variability in B. cinerea‘s susceptibility to GLSs and their breakdown products, while Alternaria brassicicola, a specialized Brassica pathogen, demonstrated heightened tolerance to GLS and their breakdown components [29]. Recent studies have shown that an mfs transporter was vital to the detoxification process of isothiocyanates (ITCs) [30]. This transporter exhibited different expression patterns in response to various ITCs under controlled in vitro conditions with heightened expression levels during interaction with wild-type A. thaliana [30]. When the mfsG gene was deleted in B. cinerea B05.10 (ΔmfsG), its ability to withstand ITCs in vitro decreased, resulting in reduced virulence toward A. thaliana [30].
2. Results
2.1. Evaluation of the Tolerance of Botrytis Species to Glucosinolate Breakdown Products
Six different Botrytis species were evaluated for their tolerance to isothiocyanates. Three of these six species (B. cinerea, B. pseudocinerea and B. byssoidea) contain the mfsG gene, while the other three (B. deweyae, B. fabae and B. convoluta) do not. The experiment was performed as indicated in the Materials and Methods Section and tested against propyl isothiocyanate (PITC), benzyl isothiocyanate (BITC) and 2-phenethyl isothiocyanate (PhITC) (Figure 1 and Figure 2).
The concentrations evaluated for each compound were 0, 20, 50, 100 and 200 µM. As shown in Figure 2, Botrytis species containing this gene exhibited higher tolerance (lower percentage of inhibition) than the species lacking it. Of the species containing the mfsG gene, B. pseudocinerea proved to be the most resistant to these compounds, followed by B. cinerea, with B. byssoidea being the most sensitive of the three species with the transporter. Regarding the species without the gene, B. deweyae was the most tolerant. Of the species most sensitive to these compounds, B. convoluta was slightly more resistant when compared to B. fabae.
However, Figure 2 shows that the tolerance of all Botrytis species to these compounds depends on the concentration of the isothiocyanate derivatives. BITC is the most toxic of the compounds used followed by PhITC. PITC was the least toxic compound, even failing to completely inhibit growth at the concentrations used on the species lacking the mfsG gene such as B. deweyae. For a more detailed analysis, see the variability in the minimum and maximum of the descriptive statistics of the inhibition data obtained in this experiment (Supplementary Table S2).
These inhibition data for Botrytis species without the gene coincide with those reported by Vela-Corcía in 2019, with inhibition to these compounds similar to that presented by the mfsG knock-out gene in B. cinerea B05.10 [30].
Infectivity assays were performed to assess the significance of the resistance conferred by this gene to these plant defence compounds as shown in Figure 3. Brassica oleracea and Raphanus sativus leaves, Raphanus sativus tuber and Brassica oleracea var. italica vegetables were infected. All of these have previously been reported to contain high glucosinolate levels [31].
In all the samples analysed, species containing the mfsG gene were able to infect these substrates in contrast to Botrytis species lacking the mfsG gene. Surprisingly, this was not the case with broccoli where just the opposite occurred, i.e. the species lacking the mfsG gene were the most successful in infecting the substrate.
These data contradict those reported by Vela-Corcía in 2019 who tested infectivity on different species of the genus Arabidopsis [30]. In that sense, the presence of the mfsG gene confirms tolerance to glucosinolate degradation products, being important during infection. However, this fact cannot be extrapolated to the whole Brassicaceae family (characterized by producing high levels of these products) as in the case of broccoli infections. In this case, Botrytis species lacking the mfsG gene are the most pathogenic. Infection of broccoli plants by B. convoluta, B. fabae, B. deweyae, B. byssoidea and B. pseudocinerea is reported here for first time.
Regarding infection of cabbage and radish leaves and radish tubers, B. pseudocinerea was the most aggressive species, followed by B. cinerea and B. byssoidea, except in the case of radish in tuber form where B. cinerea was the most aggressive, followed by B. pseudocinerea and B. byssoidea. Species lacking the mfsG gene were unable to infect these substrates in vitro. In radish leaves, a small lesion was caused by B. deweyae and B. fabae infections (with no significant differences between them) which were unable to develop further. This is also the case for B. deweyae infection in radish tubers where the lesion remained small. As in the previous case, the infection of these species by B. pseudocinerea and B. byssoidea has been described in vitro for the first time.
2.2. Glucosinolate Breakdown Products and Infection Processes Induce the mfsG Gene
The results obtained shed light on the significant impact of pure compounds—specifically, propyl isothiocyanate (PITC), benzyl isothiocyanate (BITC), and phenethyl isothiocyanate (PhITC)— and the genetic response due to infection in different plants. Notably, the presence of these compounds induces mfsG gene expression at a higher level compared to the expression triggered during infection process.
Despite the diversity of the plant species studied, a consistent pattern emerged from the axenic cultures supplemented with isothiocyanates and from infection experiments. In all instances and across all the species analysed, 60 times more of these genes were induced compared to the control axenic culture (cultured in yeast glucose glycerol medium without the addition of isothiocyanate).
This remarkable and uniform increase in gene expression suggests the robust and widespread influence of PITC, BITC, and PhITC on the genetic machinery of plants. This induction could be indicative of a sophisticated interaction between these isothiocyanate compounds and the genetic regulatory mechanisms of plants. The findings suggest the potential role of these compounds in enhancing plant defences or triggering other vital physiological responses.
The results not only underscore the potency of these pure compounds in modulating gene expression but also highlight their consistency across diverse plant species. This response uniformity implies a fundamental and perhaps evolutionarily aspect of the plant’s molecular reaction to these isothiocyanates. This study opens intriguing avenues for further research, inviting exploration into the specific molecular pathways affected and the broader implications for plant health, stress responses, and defence mechanisms in the context of these isothiocyanate compounds.
2.3. mfsG Gene Present in Only a Few Botrytis Species
A bioinformatic analysis was performed to elucidate the presence of homologous genes in other Botrytis species and showed the occurrence of these genes exclusively in B. cinerea, B. pseudocinerea, B. byssoidea, B. fragariae, and B. medusae, as detailed in Table 1.
Taking this a step further, the conservation of amino acids within the catalytic centre proposed by Vela-Corcía in 2019 [30] (Trp54, Phe254, Gln344, and Phe369) was assessed using the amino acid sequences from B. cinerea, B. fragariae, and B. byssoidea. This analysis revealed a notable conservation of these amino acids across the species, suggesting their potential significance in the protein’s activity (Figure 5).
The study also looked into conservation of the amino acids involved in the hydrogen binding of BITC and PITC (Trp54) and PhITC (Arg290). The results appear in Figure 5 and show that most of these amino acids in various Botrytis species are conserved. This suggests the potential importance of these conserved amino acids in the overall activity of the protein.
Bioinformatic exploration provides valuable insights into the evolutionary conservation of key amino acids within the Botrytis species studied. The consistent presence and conservation of these amino acids across different species implies functional relevance, highlighting their potential role in the activity of the protein under investigation.
This comprehensive approach, integrating experimental findings with bioinformatic analyses, contributes to a more nuanced understanding of the molecular landscape across Botrytis species. The conservation patterns identified pave the way for future research, providing a basis with which to explore the functional implications of these conserved amino acids and their role in the broader context of plant-pathogen interactions.
3. Discussion
In this study, six Botrytis species were systematically assessed for their responsiveness to isothiocyanates, with a particular focus on the involvement of the mfsG gene. Of the examined species, i.e. B. cinerea, B. pseudocinerea, B. byssoidea, B. deweyae, B. fabae, and B. convoluta, three were identified as having the mfsG gene, while the remaining three lacked this genetic element. The experimental design involved exposure to propyl isothiocyanate (PITC), benzyl isothiocyanate (BITC), and 2-phenethyl isothiocyanate (PhITC) at concentrations ranging from 0 to 200 µM.
The results shown in Figure 2, indicate a clear link between the presence of the mfsG gene and increased tolerance to these isothiocyanate compounds. Growth of species containing the gene was less inhibited compared to those lacking it. Notably, B. pseudocinerea was the most resistant of the mfsG-containing species, followed by B. cinerea, whereas B. byssoidea exhibited heightened sensitivity. In contrast, among the mfsG-absent species, B. deweyae exhibited elevated tolerance, while B. convoluta was marginally more resistant than B. fabae. These findings support recent evidence that suggests that our understanding of the glucosinolate breakdown mechanism in plants is incomplete, as is the list of antifungal agents. Previous research has concluded that unknown decomposition products may contribute to in vivo antifungal effects [37,38,39]. However, our data unequivocally reinforce that isothiocyanates (ITCs) are the most potent antifungal decomposition products of glucosinolates. These results underscore the importance of fully understanding glucosinolate breakdown processes and their relation to plant antifungal resistance. Further research is clearly needed to explore potential yet unidentified compounds that may contribute to the in vivo antifungal response and enhance the overall understanding of plant-fungus interactions [40].
Additionally, concentration-dependent responses were evident across all Botrytis species, with BITC exhibiting the highest toxicity, followed by PhITC. PITC demonstrated the least toxicity, even at concentrations where growth inhibition was negligible in mfsG-absent species, such as B. deweyae.
Subsequent infectivity assays showed that mfsG-containing species could successfully infect B. oleracea, R. sativus leaves, R. sativus tuber, and B. oleracea var. italica vegetable. A notable exception was observed in the case of broccoli, where mfsG-absent species exhibited heightened infectivity. Sclerotinia sclerotiorum, infecting Brassicaceae plants, cleverly manipulates the glucosinolate-myrosinase system by metabolizing isothiocyanates through two pathways. While one involves conjugation to glutathione, the fungus excels in the second, more efficient route, utilizing an isothiocyanate hydrolase analogous to a bacterial enzyme [41]. This enzyme transforms isothiocyanates into non-toxic compounds, promoting fungus growth despite the host’s defences. This adaptation contributes significantly to the virulence of S. sclerotiorum on glucosinolate-producing plants, revealing a sophisticated fungal strategy in navigating and thriving amidst the biochemical complexities of plant-fungus interactions [41].
Contrary to previous findings [30], this study challenges the established correlation between the mfsG gene and tolerance, especially evident in the case of broccoli. This nuanced observation underscores the intricate nature of plant-pathogen interactions, necessitating a refined interpretation of genetic associations. Furthermore, this study reports the previously undocumented infection of broccoli plants by B. convoluta, B. fabae, B. deweyae, B. byssoidea, and B. pseudocinerea. In cabbage and radish leaves, as well as radish in tuber form, B. pseudocinerea consistently emerged as the most aggressive species among mfsG-containing species. However, in radish tubers, B. cinerea exhibited higher aggression. Interestingly, mfsG-absent species failed to infect these substrates in vitro.
Expression analysis of the mfsG gene elucidated significant induction in the presence of isothiocyanates, both in axenic culture and after infection on Brassicaceae family plants. This induction was more than 60 times greater than control levels, emphasizing the pivotal role of this gene in responding to these compounds. Alternate research has suggested dual roles for ABC and MFS transporters, potentially serving as effectors or contributors to the secretion of pathogenicity factors, influencing fungal virulence. Dos Santos, et al.’s research into MFS transporters in yeast proposed an additional function—providing drug resistance through indirect regulation of stress response and membrane potential control. This prompts consideration of analogous impacts in plant-pathogen transporters, where similar multifaceted effects on virulence and resistance mechanisms may be at play. These insights broaden our understanding of transporter functions, indicating their potential significance in both pathogenicity and drug resistance across diverse biological systems [42,43,44,45].
A bioinformatic investigation identified homologous mfsG genes exclusively in B. cinerea, B. pseudocinerea, B. byssoidea, B. fragariae, and B. medusae. Conservation analysis of amino acids within the catalytic centre proposed by Vela-Corcía in 2019 [30] demonstrated notable conservation across diverse Botrytis species, suggesting the potential significance of these amino acids in protein activity.
This comprehensive inquiry provides valuable insights into the intricate dynamics between Botrytis species and isothiocyanates, underscoring the vital role of the mfsG gene in conferring tolerance. These findings call for continued research in this domain, advocating the use of advanced biotechnological tools to unravel the intricate biology of these proteins and deepen our understanding of plant-pathogen interactions. The findings also generate heightened interest in the use of diverse biotechnological tools to deepen our understanding of the biology surrounding these proteins. Biotechnological approaches such as advanced genomic techniques, proteomics, and bioinformatics, offer avenues for more detailed investigation. Genomic tools can aid in unravelling the genetic architecture, identifying regulatory elements, and understanding the broader genetic networks associated with these proteins [46]. Proteomic techniques can provide insights into the protein’s structure, interactions, and potential post-translational modifications [47,48]. Moreover, bioinformatics can facilitate the integration of large datasets, allowing for a comprehensive analysis of the molecular landscape. Also, CRISPR-Cas9 gene editing technology can enable targeted modifications, allowing researchers to validate the functional significance of specific genes and amino acids identified in the study [49,50]. This biotechnological toolkit empowers scientists to dissect the intricate mechanisms at play, shedding light on the functional roles of these proteins in plant-pathogen interactions.
The need for continued research, coupled with the integration of advanced biotechnological tools, not only contributes to our understanding of these specific proteins but also has broader implications for agricultural practices. Unravelling the biology of these proteins can inform the development of targeted strategies to enhance plant resistance, optimize crop yield, and mitigate the impact of plant diseases.
In conclusion, the multifaceted data presented in this report advocate for a sustained and diversified research approach, leveraging biotechnological tools to decipher the biology of these proteins. This scientific journey promises not only a deeper understanding of plant-pathogen interactions but also the development of innovative solutions with tangible benefits for agriculture and crop management.
4. Materials and Methods
4.1. Organisms, Media and Culture Conditions
The Botrytis species used in this study were: B. cinerea B05.10 [32,33], B. pseudocinerea VD165 [51], B. deweyae B1 [52], B. byssoidea MUCL94 [53], B. fabae 2220 and B. convoluta MUCLII595 [54]. All fungal cultures underwent standard cultivation in a YGG medium (composed of 2% glucose, 0.5% yeast extract, 0.3% Gamborg’s B5 medium from Duchefa Biochemie, and, when necessary, 1.5% agar) at a temperature of 20°C for three days. Conidia were produced on tomato agar plates (comprised of 25% homogenized tomato fruit by weight, 1.5% agar, pH 5.5). These conidial stocks were preserved in a 10% glycerol solution at -80°C. For non-sporulate species, mycelium plugs were preserved in a 20% and 60% glycerol stock solution at -80˚C.
4.2. Vegetative Growth
To examine vegetative growth inhibition of the fungi in the presence of different glucosinolate breakdown products, Petri dishes were prepared with YGG-agar medium and subsequently inoculated with three mycelial agar plugs measuring 3 mm in diameter. Incubation was at 20°C and any deviation in fungal growth patterns was monitored. Daily measurements of colony radius were recorded over a period of three days. The radial growth rate assessment involved plotting the colony radius against time and fitting the data into a linear model which displayed a robust correlation (Pearson’s correlation coefficient value (r2 ≥ 0.98). To evaluate the strains’ responsiveness to stress stimuli, the YGG-agar medium was supplemented with different concentrations (20µM, 50µM, 100µM and 200µM) of isothiocyanate products (PITC, BITC and PhITC) to induce stress caused by the presence of plant defence products which are glucosinolate breakdown products. The radial mycelial growth radius (R, in mm) was measured at 72 h for treatments with isothiocyanate products (Rt) and controls without any supplementation (Rc) in YGG medium. The fungitoxic effects of PITC, BITC and PhITC were quantified as the percentage of growth inhibition (GI) using the formula: GI(%) = (Rc—Rt)/Rc × 100. This comprehensive experiment comprised a total of 50 measurements, distributed across 5 independent replicates, each consisting of 10 measurements.
Results were represented as the mean of growth inhibition percentages in cm, presented alongside their respective standard deviations, derived from a minimum of forty independent experiments.
4.3. Virulence Assay
Virulence assessments were conducted using detached B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable. These substrates were inoculated with agar plugs measuring 3-mm in diameter in a 10µL TGGK solution (comprised of 60mM KH2PO4, 10mM glycine, 0.01% Tween 20, and 100mM glucose). Incubation ensued in darkness at 20°C, maintaining high humidity levels, with photographic documentation every 24 hours. ImageJ software [55] facilitated the measurement of lesion radii on B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable. Results are presented as the mean lesion diameters from fifty biological replicates from the infection of the different Botrytis species after 3 days of infection.
4.4. Quantitative Real-Time PCR (qRT-PCR)
Total RNA extraction from fungal mycelia entailed a filtration process from 30 mL of YGG medium (control) or YGG supplemented with 80µM of PITC, BITC and PhITC inoculated with 5 agar plugs measuring 0.3 cm in diameter. This culture was left to incubate at 20°C in darkness for three days. To explore gene expression within plant hosts, detached B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable were inoculated with 5 µL droplets containing a mycelial plug measuring 0.3 cm in diameter in a 10µL TGGK solution composed of 60 mM KH2PO4, 10 mM glycine, 0.01% Tween 20, and 100 mM glucose. All inoculated species were incubated for 72 hours at 22°C in a dark environment, maintaining a humidity level of 70%. After infection, segments of the affected leaf areas were excised and promptly frozen at -80°C for subsequent analysis. Both mycelia and infected plant tissues were homogenized, and total RNA was isolated using Trizol Reagent (Sigma-Aldrich, T 9424) following the manufacturer’s protocol.
The subsequent step involved cDNA synthesis from 1 µg of total RNA, using the iScript cDNA Synthesis Kit (Bio-Rad, USA), following the instructions provided. Quantitative real-time PCR (qRT-PCR) was conducted on an iCycler iQ system (Bio-Rad, USA) using the iQ SYBR Green Supermix (Bio-Rad, USA) along with primers detailed in Supplementary Table S1. To ensure normalization of the cDNA samples, the actA (Bcin16g02020) and B-tub (Bcin01g08040) genes, responsible for encoding actin and tubulin, respectively, were used as internal controls. Relative mRNA quantities were determined using the ΔΔCt method [56], averaging three independent determinations of the threshold cycle (Ct). The resulting data were presented as mean values of 2ΔΔCt±SD normalized concerning the expression of each gene in YGG without the addition of the glucosinolate breakdown products PITC, BITC and PhITC.
Furthermore, the relative expression of the mfsG gene was measured in the species containing the indicated transporter gene (B. cinerea, B. pseudocinerea and B. byssoidea). The study was carried out in axenic culture after 72 hours of incubation, with the isothiocyanates PITC, BITC and PhITC, at a single concentration of 70 µM, and after 72 hours following infection on Brassicaceae family plants. The axenic culture of these Botrytis species after 72 hours of incubation in YGG medium without the addition of any other compound was used as a control. This control was used to normalize data and ascertain the extent to which this gene was induced under each condition compared to the control.
4.5. Bioinformatic Analysis
The nucleotide sequence of the mfsG gene was used to search for homologous genes in other Botrytis species using the BLASTn tool [57,58] and the nucleotide collection database (nr/nt). For species that are poorly sequenced and for which there is no gene databank, the Whole-genome shotgun contigs (wgs) database was used. To analyse amino acid conservation of the MfsG protein sequence, a blastp was used to find homologous proteins in other Botrytis species using the non-redundant protein sequences (nr) database [58]. The protein sequences from B. cinerea B05.10, B. fragariae BVB16 and B. byssoidea MUCL94 were aligned with the structural alignment (Expresso) from the T-coffe tool [59]. Results were visualized with the Espript 3.0 tool [60] (https://espript.ibcp.fr). The conserved amino acid proposed by Vela-Corcía et. al. in 2019 [30] to be the binding region (Trp54, Phe254, Gln344, and Phe369), and the amino acid involved in the hydrogen binding of BITC and PITC (Trp54) and PhITC (Arg290) from B. cinerea B05.10 in B. byssoidea MUCL94 and B. fragariae BVB16, were displayed in the alignment.
4.6. Statistical Analysis
Graphpad Prism 8 was used for statistical analyses. The Kolmogorov-Smirnov test (for samples >50) and the Shapiro-Wilk test (for samples <50) were used to assess normality. Depending on normality outcomes, the T-test or Mann-Whitney test was applied to compare normally distributed or nonparametric data, respectively. Statistical significance was established at a p-value < 0.05.
5. Conclusions
In this study, six Botrytis species were systematically studied for their response to isothiocyanates, particularly focusing on the mfsG gene. The presence of this gene was associated with increased tolerance to isothiocyanate compounds as demonstrated by reduced growth inhibition percentages in mfsG-containing species. However, the correlation between the mfsG gene and tolerance was challenged, particularly in the case of broccoli, suggesting the complex nature of plant-pathogen interactions.
Concentration-dependent responses were observed across all Botrytis species, with benzyl isothiocyanate (BITC) exhibiting the highest toxicity. Infectivity assays revealed varying aggressiveness among species on different plant substrates, challenging previous findings and highlighting the need for a nuanced interpretation of genetic associations.
Expression analysis of the mfsG gene showed significant induction in the presence of isothiocyanates, emphasizing its pivotal role in responding to these compounds. Bioinformatic investigations identified homologous mfsG genes in specific Botrytis species, with conserved amino acids in the catalytic centre suggesting their functional significance.
The study underscores the importance of advanced biotechnological tools, including genomics, proteomics, bioinformatics, and CRISPR-Cas9 gene editing, in deciphering the intricate biology of these proteins. This comprehensive approach not only enhances our understanding of plant-pathogen interactions but also holds promise for developing targeted strategies to improve plant resistance and optimize crop yield. In conclusion, continued and diversified research using advanced biotechnological tools is essential for unlocking the full potential of these proteins, offering innovative solutions for agriculture and crop management.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
“Conceptualization, V. C-R and I.G.C.; methodology, J.A. and V. C-R; investigation, V. C-R.; resources, J.A. and I.G.C.; writing—original draft preparation, V.C-R.; writing—review and editing, J.A. and I.G.C.; supervision, J.A. and I.G.C.; project administration, I.G.C.; funding acquisition, J.A. and I.G.C. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was supported by a grant from MCIN (PID2021-1228990-B-C21 MCIN/AEI/FEDER, EU).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Smith, D.A.; Banks, S.W. Formation and biological properties of isoflavonoid phytoalexins. Prog. Clin. Biol. Res. 1986, 213, 113–24. [Google Scholar]
- Arbona, V.; Gómez-Cadenas, A. Omics in Plant Disease Resistance; Caister Academic Press, 2016; ISBN 9781910190357. [Google Scholar]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: phytoalexins versus “Phytoanticipins” . Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef]
- JPMorrissey, A.O. Fungal resistance to plant antibiotics as a mechanism of pathogenesis. Microbiol. Mol. Biol. Rev. 1999, 63, 708–724. [Google Scholar] [CrossRef]
- Kliebenstein, D.J. Secondery metabolites and plant/environment interactions: a view through Arabidopsis thaliana tinged glasses. Plant Cell Environ. 2004, 27, 675–684. [Google Scholar] [CrossRef]
- VanEtten, H.D.; Matthews, D.E.; Matthews, P.S. Phytoalexin detoxification: importance for pathogenicity and practical implications. Annu. Rev. Phytopathol. 1989, 27, 143–164. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Ahiahonu, P.W.K. Metabolism and detoxification of phytoalexins and analogs by phytopathogenic fungi. Phytochemistry 2005, 66, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Ahiahonu, P.W.K. Probing the phytopathogenic stem rot fungus with phytoalexins and analogues: unprecedented glucosylation of camalexin and 6-methoxycamalexin. Bioorg. Med. Chem. 2002, 10, 3307–3312. [Google Scholar] [CrossRef] [PubMed]
- Schoonbeek, H.; Del Sorbo, G.; De Waard, M.A. The ABC transporter BcatrB affects the sensitivity of Botrytis cinerea to the phytoalexin resveratrol and the fungicide fenpiclonil. Mol. Plant Microbe Interact. 2001, 14, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Schoonbeek, H.J.; Raaijmakers, J.M.; De Waard, M.A. Fungal ABC transporters and microbial interactions in natural environments. Mol. Plant Microbe Interact. 2002, 15, 1165–1172. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Burow, M.; Wittstock, U. Regulation and function of specifier proteins in plants. Phytochem. Rev. 2009, 8, 87–99. [Google Scholar] [CrossRef]
- Borek, V.; Morra, M.J.; Brown, P.D.; McCaffrey, J.P. Allelochemicals produced during sinigrin decomposition in Soil. J. Agric. Food Chem. 1994, 42, 1030–1034. [Google Scholar] [CrossRef]
- Andréasson, E.; J#xD8;rgensen, L.B.; Höglund, A.-S.; Rask, L.; Meijer, J. Different myrosinase and idioblast distribution in Arabidopsis and Brassica napus. Plant Physiol. 2001, 127, 1750–1763. [Google Scholar] [CrossRef] [PubMed]
- Del Sorbo, G.; Schoonbeek, H.J.; De Waard, M.A. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Genet. Biol. 2000, 30, 1–15. [Google Scholar] [CrossRef]
- Rogers, B. The pleitropic drug ABC transporters from Saccharomyces cerevisiae. J. Mol. Microbiol. Biotechnol. 2001, 3, 207–214. [Google Scholar] [PubMed]
- Urban, M.; Bhargava, T.; Hamer, J.E. An ATP-driven efflux pump is a novel pathogenicity factor in rice blast disease. EMBO J. 1999, 18, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.B.; Suresh, A.; Deng, Y.Z.; Naqvi, N.I. A multidrug resistance transporter in Magnaporthe is required for host penetration and for survival during oxidative stress. Plant Cell 2006, 18, 3686–3705. [Google Scholar] [CrossRef]
- Gupta, A.; Chattoo, B.B. Functional analysis of a novel ABC transporter ABC4 from Magnaporthe grisea. FEMS Microbiol. Lett. 2008, 278, 22–28. [Google Scholar] [CrossRef]
- Stefanato, F.L.; Abou-Mansour, E.; Buchala, A.; Kretschmer, M.; Mosbach, A.; Hahn, M.; Bochet, C.G.; Métraux, J.P.; Schoonbeek, H.J. The ABC transporter BcatrB from Botrytis cinerea exports camalexin and is a virulence factor on Arabidopsis thaliana. Plant J. 2009, 58, 499–510. [Google Scholar] [CrossRef]
- Fleißner, A.; Sopalla, C.; Weltring, K.M. An ATP-binding cassette multidrug-resistance transporter is necessary for tolerance of Gibberella pulicaris to phytoalexins and virulence on potato tubers. Mol. Plant Microbe Interact. 2002, 15, 102–108. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Van Nistelrooy, J.G.M.; Kema, G.H.J.; De Waard, M.A. Multiple mechanisms account for variation in base-line sensitivity to azole fungicides in field isolates of Mycosphaerella graminicola. Pest Manag Sci. 2003, 59, 1333–1343. [Google Scholar] [CrossRef]
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis: Biology, pathology and control. Botrytis Biol. Pathol. Control 2007, 1–403. [Google Scholar] [CrossRef]
- Fillinger, S.; Elad, Y. Botrytis – the Fungus, the Pathogen and its management in agricultural systems. Botrytis - Fungus, Pathog. its Manag. Agric. Syst. 2016, 1–486. [Google Scholar] [CrossRef]
- Leroux, P.; Fritz, R.; Debieu, D.; Albertini, C.; Lanen, C.; Bach, J.; Gredt, M.; Chapeland, F. Mechanisms of resistance to fungicides in field strains of Botrytis cinerea. Pest Manag. Sci. 2002, 58, 876–888. [Google Scholar] [CrossRef]
- Sofianos, G.; Samaras, A.; Karaoglanidis, G. Multiple and multidrug resistance in Botrytis cinerea: molecular mechanisms of MLR/MDR strains in Greece and effects of co-existence of different resistance mechanisms on fungicide sensitivity. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Fillinger, S.; Walker, A.-S. Chemical control and resistance management of Botrytis diseases. In Botrytis – the Fungus, the Pathogen and its Management in Agricultural Systems; Springer International Publishing: Cham, 2016; pp. 189–216. [Google Scholar]
- Buxdorf, K.; Yaffe, H.; Barda, O.; Levy, M. The effects of glucosinolates and their breakdown products on necrotrophic fungi. PLoS One 2013, 8, e70771. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Aditya Srivastava, D.; Dafa-Berger, A.; Rotem, N.; Barda, O.; Levy, M. MFS transporter from Botrytis cinerea provides tolerance to glucosinolate-breakdown products and is required for pathogenicity. Nat. Commun. 2019, 10, 2886. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liang, Z.; Zang, Y.; Zhu, Z.; Yang, J. Diversity of glucosinolates among common Brassicaceae vegetables in China. Hortic. Plant J. 2023, 9, 365–380. [Google Scholar] [CrossRef]
- Amselem, J.; Cuomo, C.A.; van Kan, J.A.L.L.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; de Vries, R.P.; Dyer, P.S.; Fillinger, S.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef] [PubMed]
- Van Kan, J.A.L.L.; Stassen, J.H.M.M.; Mosbach, A.; Van Der Lee, T.A.J.J.; Faino, L.; Farmer, A.D.; Papasotiriou, D.G.; Zhou, S.; Seidl, M.F.; Cottam, E.; et al. A gapless genome sequence of the fungus Botrytis cinerea. Mol. Plant Pathol. 2017, 18, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Saski, C.; Schnabel, G.; Xiao, S.; Hu, M. A high-quality genome resource of Botrytis fragariae, a new and rapidly spreading fungal pathogen causing strawberry gray mold in the United States. Phytopathology 2021, 111, 496–499. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Steentjes, M.B.F.; Slot, J.C.; Shi-Kunne, X.; Scholten, O.E.; van Kan, J.A.L. Dynamics in secondary metabolite gene clusters in otherwise highly syntenic and stable genomes in the fungal genus Botrytis. Genome Biol. Evol. 2020, 12, 2491–2507. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, M.C.; Harper, L.A.; Lopez-Ruiz, F.J. Positive Selection of Transcription Factors Is a Prominent Feature of the Evolution of a Plant Pathogenic Genus Originating in the Miocene. Genome Biol. Evol. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Frerigmann, H.; Piślewska-Bednarek, M.; Sánchez-Vallet, A.; Molina, A.; Glawischnig, E.; Gigolashvili, T.; Bednarek, P. Regulation of pathogen-triggered tryptophan metabolism in Arabidopsis thaliana by MYB transcription factors and indole glucosinolate conversion products. Mol. Plant 2016, 9, 682–695. [Google Scholar] [CrossRef]
- Yun, H.S.; Kang, B.G.; Kwon, C. Arabidopsis immune secretory pathways to powdery mildew fungi. Plant Signal. Behav. 2016, 11, e1226456. [Google Scholar] [CrossRef]
- He, Y.; Xu, J.; Wang, X.; He, X.; Wang, Y.; Zhou, J.; Zhang, S.; Meng, X. The Arabidopsis Pleiotropic drug resistance transporters PEN3 and PDR12 mediate camalexin secretion for resistance to Botrytis cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef] [PubMed]
- Plaszkó, T.; Szűcs, Z.; Vasas, G.; Gonda, S. Effects of glucosinolate-derived isothiocyanates on fungi: A comprehensive review on direct effects, mechanisms, structure-activity relationship data and possible agricultural applications. J. Fungi 2021, 7, 539. [Google Scholar] [CrossRef]
- Chen, J.; Ullah, C.; Reichelt, M.; Beran, F.; Yang, Z.L.; Gershenzon, J.; Hammerbacher, A.; Vassão, D.G. The phytopathogenic fungus Sclerotinia sclerotiorum detoxifies plant glucosinolate hydrolysis products via an isothiocyanate hydrolase. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- de Waard, M.A.; Andrade, A.C.; Hayashi, K.; Schoonbeek, H.; Stergiopoulos, I.; Zwiers, L. Impact of fungal drug transporters on fungicide sensitivity, multidrug resistance and virulence. Pest Manag. Sci. 2006, 62, 195–207. [Google Scholar] [CrossRef]
- Hayashi, K.; Schoonbeek, H. jan; De Waard, M.A. Bcmfs1, a novel major facilitator superfamily transporter from Botrytis cinerea, provides tolerance towards the natural toxic compounds camptothecin and cercosporin and towards fungicides. Appl Environ. Microbiol. 2002, 68, 4996–5004. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Schoonbeek, H. jan; De Waard, M.A. Modulators of membrane drug transporters potentiate the activity of the DMI fungicide oxpoconazole against Botrytis cinerea. Pest Manag. Sci. 2003, 59, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Gielkens, M.M.C.; Goodall, S.D.; Venema, K.; De Waard, M.A. Molecular cloning and characterisation of three new ATP-binding cassette transporter genes from the wheat pathogen Mycosphaerella graminicola. Gene 2002, 289, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Shafran, H.; Miyara, I.; Eshed, R.; Prusky, D.; Sherman, A. Development of new tools for studying gene function in fungi based on the Gateway system. Fungal Genet. Biol. 2008, 45, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Acero, F.J.; Cantoral, J.M.; Javier Fernández-Acero, F.; Carbú, M.; Garrido, C.; Vallejo, I.; Cantoral, J.M. Proteomic advances in phytopathogenic fungi. Current Proteomics 2007, 4(2), 79–88. [Google Scholar] [CrossRef]
- Fernández Acero, F.J.; Carbú, M.; El-Akhal, M.R.; Garrido, C.; González-Rodríguez, V.E.; Cantoral, J.M. Development of proteomics-based fungicides: New strategies for environmentally friendly control of fungal plant diseases. Int. J. Mol. Sci. 2011, 12, 795–816. [Google Scholar] [CrossRef]
- Hahn, M.; Scalliet, G. One cut to change them all: CRISPR/Cas, a groundbreaking tool for genome editing in Botrytis cinerea and other fungal plant pathogens. Phytopathology® 2021, 111, 474–477. [Google Scholar] [CrossRef]
- Chen, J.; Lai, Y.; Wang, L.; Zhai, S.; Zou, G.; Zhou, Z.; Cui, C.; Wang, S. CRISPR/Cas9-mediated efficient genome editing via blastospore-based transformation in entomopathogenic fungus Beauveria bassiana. Sci. Rep. 2017, 7, 45763. [Google Scholar] [CrossRef]
- Walker, A.S.; Gautier, A.; Confais, J.; Martinho, D.; Viaud, M.; Pêcheur, P. Le; Dupont, J.; Fournier, E. Botrytis pseudocinerea, a new cryptic species causing gray mold in french vineyards in sympatry with Botrytis cinerea. Phytopathology 2011, 101, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- van Kan, J.A.L.; Shaw, M.W.; Grant-Downton, R.T. Botrytis species: Relentless necrotrophic thugs or endophytes gone rogue? Mol. Plant Pathol. 2014, 15, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.C. Two undescribed species of Botrytis associated with the neck rot disease of onion bulbs. Phytopathology 1925, 15, 708–713. [Google Scholar]
- Whetzel & Drayton Botrytis convoluta. Mycologia 1932, 24(6), 475.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. 2012. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: architecture and applications. BMC Bioinformatics 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–17. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
Figure 1.
Glucosinolates (GLSs) and their enzymatic and spontaneous conversion to BITC, PITC and PhITC.
Figure 1.
Glucosinolates (GLSs) and their enzymatic and spontaneous conversion to BITC, PITC and PhITC.
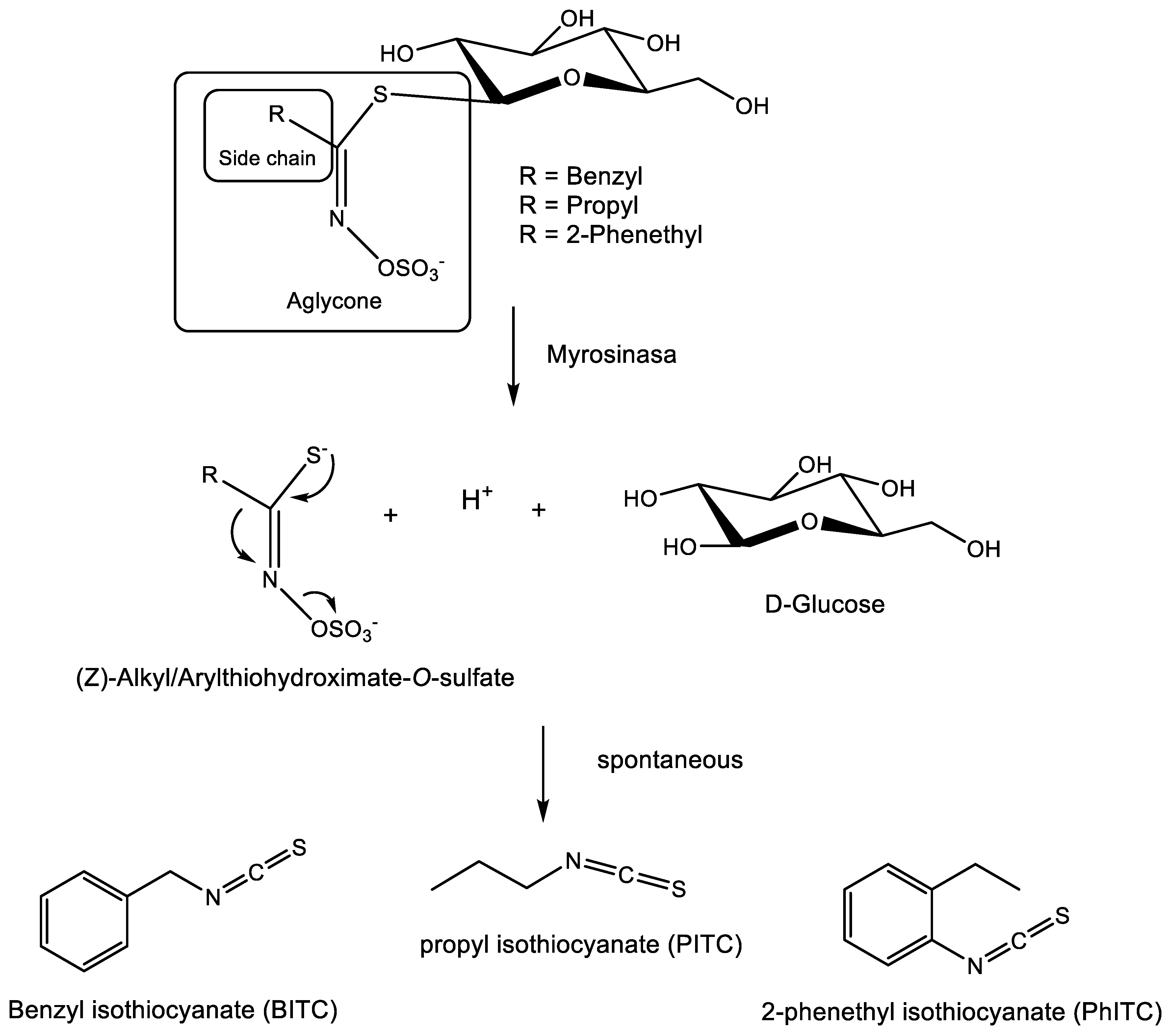
Figure 2.
Percentage inhibition to different isothiocyanates (PITC, BITC and PhITC) of different Botrytis species (B. cinerea, B. pseudocinerea, B. byssoidea, B. deweyae, B. fabae and B. convoluta). Data shown are the mean of five independent biological replicates, each consisting of 10 measurements.
Figure 2.
Percentage inhibition to different isothiocyanates (PITC, BITC and PhITC) of different Botrytis species (B. cinerea, B. pseudocinerea, B. byssoidea, B. deweyae, B. fabae and B. convoluta). Data shown are the mean of five independent biological replicates, each consisting of 10 measurements.
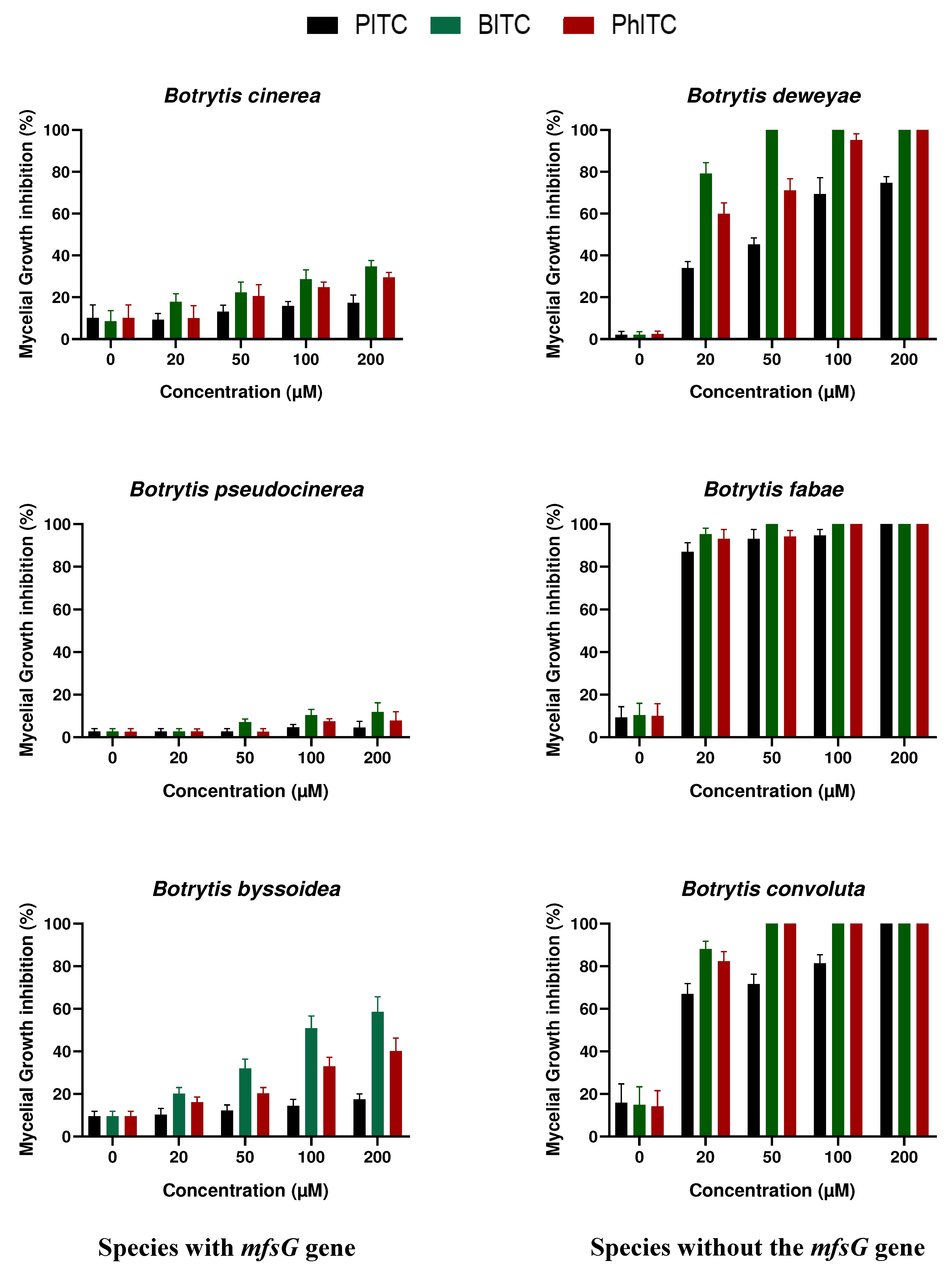
Figure 3.
Lesion diameter after 3 days of inoculation with B. cinerea, B. pseudocinerea, B. byssoidea, B. deweyae, B. fabae and B. convoluta in different members of the Brassicaceae family: B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable. The data shown in B, D, F and H correspond to the mean of 50 biological replicates. Different letters above each bar indicate significant differences (p<0.05) between the different Botrytis species analysed.
Figure 3.
Lesion diameter after 3 days of inoculation with B. cinerea, B. pseudocinerea, B. byssoidea, B. deweyae, B. fabae and B. convoluta in different members of the Brassicaceae family: B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable. The data shown in B, D, F and H correspond to the mean of 50 biological replicates. Different letters above each bar indicate significant differences (p<0.05) between the different Botrytis species analysed.
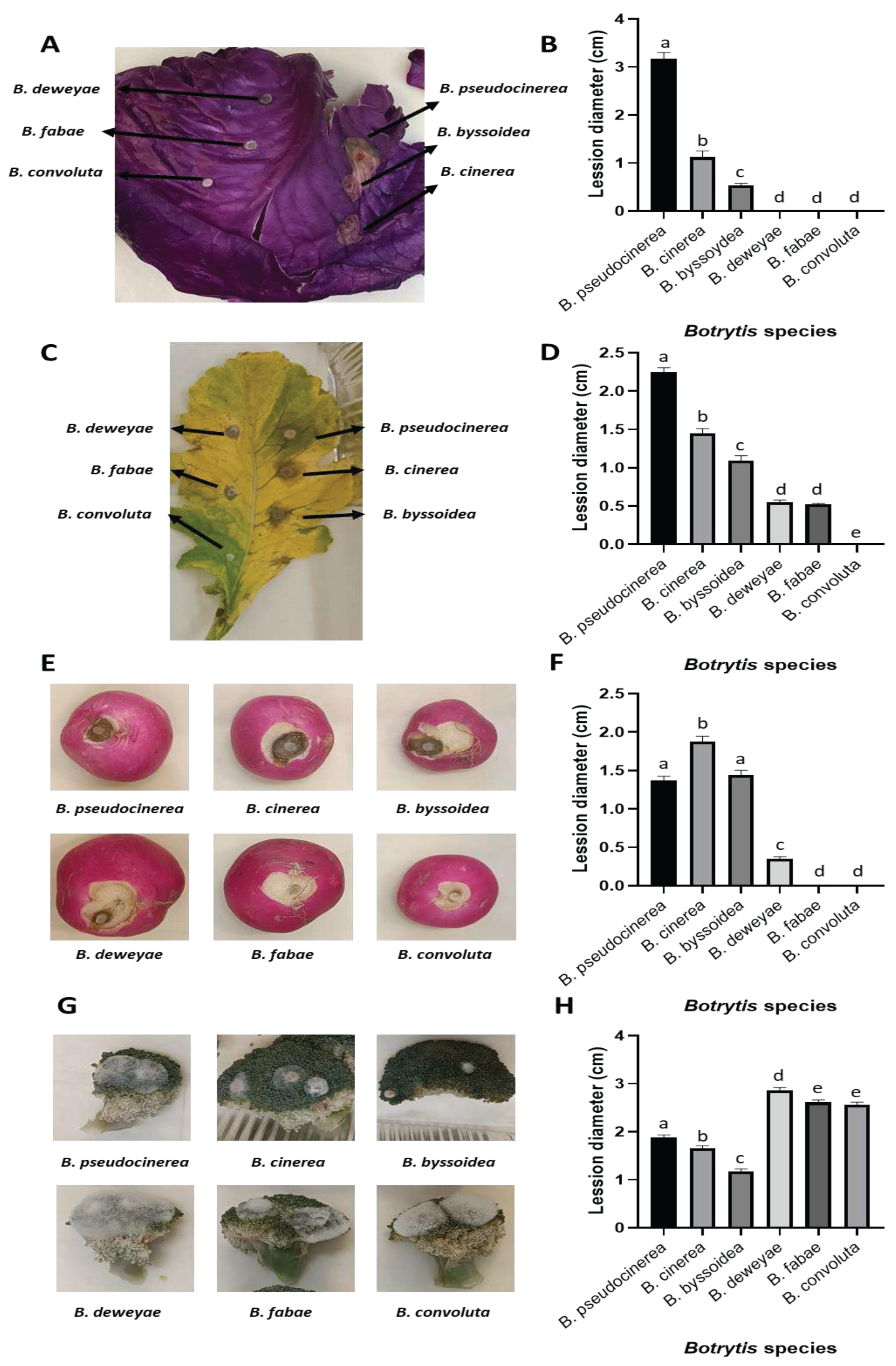
Figure 4.
Relative expression of the mfsG gene of B. cinerea, B. pseudocinerea, and B. byssoidea in reaction to the presence of thiocyanates in axenic culture (PITC, BITC, and PhITC), and during infection in high glucosinolate (Brassicaceae family) seedlings such as: B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable.
Figure 4.
Relative expression of the mfsG gene of B. cinerea, B. pseudocinerea, and B. byssoidea in reaction to the presence of thiocyanates in axenic culture (PITC, BITC, and PhITC), and during infection in high glucosinolate (Brassicaceae family) seedlings such as: B. oleracea and R. sativus leaves, R. sativus tuber and B. oleracea var. italica vegetable.
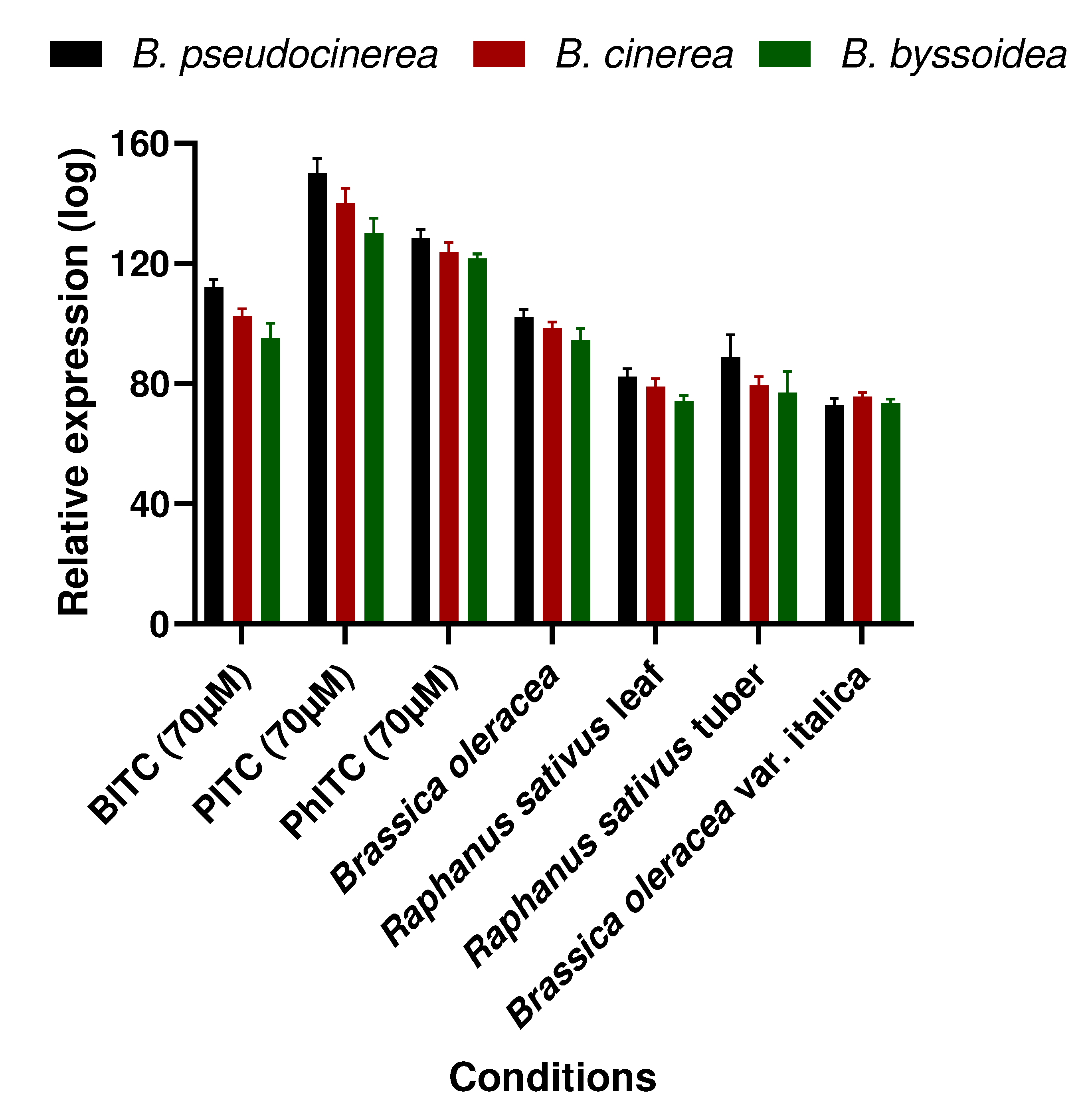
Figure 5.
Alignment of the MfsG homologous protein in other species of the Botrytis genus. Alignment in red represents highly conserved amino acids among the different MfsG sequences. XP_024549048.1 represent the MfsG protein sequence from B. cinerea, XP_037186706.1 represents the protein MfsG sequence from B. pseudocinerea and XP_038726455.1 represents the MfsG protein sequence from B. byssoidea. Green dots represent the conservation of the proposed amino acid involved in the binding region (Trp 54, Phe254, Gln344 and Phe369), yellow dots represent the predicted amino acids involved in the hydrogen binding to PITC and BITC, while blue dots are for the predicted amino acids involved in the PhITC hydrogen binding previously described in [30].
Figure 5.
Alignment of the MfsG homologous protein in other species of the Botrytis genus. Alignment in red represents highly conserved amino acids among the different MfsG sequences. XP_024549048.1 represent the MfsG protein sequence from B. cinerea, XP_037186706.1 represents the protein MfsG sequence from B. pseudocinerea and XP_038726455.1 represents the MfsG protein sequence from B. byssoidea. Green dots represent the conservation of the proposed amino acid involved in the binding region (Trp 54, Phe254, Gln344 and Phe369), yellow dots represent the predicted amino acids involved in the hydrogen binding to PITC and BITC, while blue dots are for the predicted amino acids involved in the PhITC hydrogen binding previously described in [30].


Table 1.
Identification of the mfsG gene among different Botrytis species.
| Species | Isolate | NCBI Gene Accession Number |
NCBI Protein Accession Number | Gene Identity with B. cinerea B05.10 (%) | Length (pb) | Reference |
|---|---|---|---|---|---|---|
| B. cinerea | B05.10 | XM_024693262.1 | XP_024549048.1 | 100% | 1379 | [32,33] |
| B. fragariae | BVB16 | XM_037342786.1 | XP_037186706.1 | 86.72% | 1296 | [34] |
| B. byssoidea | MUCL 94 | XM_038882714.1 | XP_038726455.1 | 84.47% | 1296 | [35] |
|
B. pseudocinerea |
BP362 | JAHXJK010000103.1: 18998-20539 | - | 99.33% | 1541 | [36] |
| B. medusae | B555 | JAHXJK010000103.1: 15498-16733 | - | 90.47% | 1235 | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



