
Submitted:
26 January 2024
Posted:
29 January 2024
You are already at the latest version
Abstract
This study describes the seasonal course of denitrifying and nitrate reducing bacteria in a dimictic mesotrophic lake (Lake Scharmützelsee, Brandenburg, Germany) within a three-year period from 2011 to 2013. The bacterial cell numbers were quantified by fluorescence microscopy, most probable number (MPN) and PCR dependent quantification of the chromosomal 16S rDNA and of the nirS and nirK gene copy number. The highest seasonal differences (up to three orders of magnitudes) has been measured by MPN in the epilimnion. This variation was not reflected by PCR dependent approaches or direct microscopical enumeration. At adverse conditions (low temperature and/or low nitrate concentrations), the differences between MPN and gene copy numbers increased up to five orders of magnitudes and decreased to one magnitude at favorable environmental conditions. These results can be explained best by an increasing ratio of viable but not cultivable (VBNC) cells or dead cells at impairing conditions. In the hypolimnion, the courses of MPN and nir gene copy numbers were similar. This can be explained by higher feeding pressure and therefore smaller amounts of dormant cells. In the pelagial in general, the total cell numbers enumerated by either microscopical or molecular approaches were similar. In the sediment, more than 99 % of the DNA was obviously not related to viable bacteria but was rather “dead DNA” in dead cells or adsorbed to particle surfaces.
Keywords:
denitrification
; nitrite reductase
; MPN
; vbnc state
; PCR
; microscopical cell number
; dimictic lake
; epilimnion
; hypolimnion
; sediment
1. Introduction
Urbanization and human land uses such as agriculture have resulted in increased levels of bioavailable nitrogen in lakes, streams and rivers. Denitrification is the most important way of nitrogen elimination in limnic ecosystems. The determination of the abundance, function and activity of the responsible group of bacteria is an invaluable aspect of microbial ecology ([1] and can be a useful approach to estimate in situ turnover rates [2,3,4] parallel to direct in situ measurements, which however are often difficult [5].
Many studies on denitrification rates and the responsible bacterial populations have been carried out mainly in marine and coastal ecosystems and especially estuaries (e.g. [6,7,8]), but samples were often taken only few times per year [9] and eventually analyzed either by cultivation based techniques ([10]) or by molecular methods [9,11,12,13,14]. Although fresh water ecosystems can serve as an important nitrogen sink in temperate regions, only little is known about the seasonal variability of the denitrifying bacterial populations in these systems [10]. The same is true for the relative impacts of the pelagial and sediment to the overall denitrification capacity. One reason could be the lack of adequate enumeration techniques for the different groups of bacteria in the nitrogen cycle and the many drawbacks of individual quantification methods.
Although several methods are available, the exact enumeration of denitrifying bacteria is still difficult [14]. Cultivation based techniques have been applied for decades for selective counting of functional bacterial groups. Their advantages and disadvantages have been often discussed [15]. They might only catch a small fraction of the bacterial group of interest [16,17], are time and material consuming [18,19] and quite imprecise [20]. It is argued, that cultivation depending enumeration will most likely underestimate the number of bacteria by magnitudes compared to molecular techniques [15,16,21,22]. With advances in molecular techniques, the selective detection and quantification of denitrifying bacteria based on the PCR of genomic DNA of enzymes of this metabolic pathway became possible [11,13,23,24]. PCR based methods are faster with less material effort and more precise, especially when using q-PCR or even more advanced molecular techniques. However, molecular techniques encounter several difficulties as well [25,26,27,28,29]. A major disadvantage in ecological means is that PCR techniques based on ribosomal or genomic DNA cannot distinguish between inactive or non viable cells or dead DNA and metabolically active cells [24,30,31,32,33], but deliver a proportion of total cell counts, copy number, or relative signal intensities rather than an absolute number of cells or biomass [1]. Only in systems with fresh biomass the MPN counts and relative numbers of gene copies are closely correlated [34]. Thus, several authors concluded that molecular approaches are not sufficiently discussed in comparison with traditional methods [35]. A comparison between cultivation based and molecular cell counting techniques and a correlation of these results with in situ activities is still difficult. Whereas [36] and [5] found a correlation between gene copy number and in stream denitrification activity, [14] suggest the use of gene abundance mainly as indication of potential activity.
It has been shown previously, that bacteria can enter a so called viable but non cultivable state (VBNC state) [37] as a response to harsh environmental conditions such as nutrient starvation, extreme temperatures, sharp changes in pH or salinity or oxygen availability [38]. In a lab study with a pure Pseudomonas aerogina strain, less than 0.1 % of all bacteria were cultivable under nitrate starvation whereas optimal growth conditions resulted in nearly identical cell counts by MPN and molecular techniques [24]. The discrepancies between MPN and molecular techniques may indicate the portion of bacteria that are not involved in substrate turnover processes. Especially in environmental systems with changing growth conditions, molecular based techniques might not reflect the variability of active cell abundance [24].
Thus, the understanding of the nitrogen turnover and the study of interrelationships among and between bacterial populations and their environment still needs adequate enumeration techniques for metabolically active bacteria [29].
In this study the abundances and seasonal variations of nitrate reducing bacteria in the pelagial and upper sediment of a mesotrophic dimictic fresh water lake (Lake Scharmützelsee, Germany) has been followed over a period of more than two years. Because of the difficulties in bacterial enumeration we used and compared the cultivation based MPN technique with the molecular quantification of the nitrite reductase encoding nirS and nirK gene copy number and 16S rDNA as well as with total concentrations of bacterial cells counted by filtration and fluorescence microscopy. Our results can help to understand the seasonal dynamic of denitrification processes in comparable systems and might improve the estimation of the denitrification capacities of the entire system and its different compartments.
2. Materials and Methods
2.1. Sampling Site and Physical and Chemical Parameters
The lake Scharmützelsee is a dimictic mesotrophic lake of the type LCB 1 of the European intercalibration lake type in the Central Baltic region [39]. It is located in NE Germany (52°15´ N, 14°03´ E), has a size of aprox. 12 km², a mean depth of 8.8 m, a maximal depth of 29.5 m and a mean residence time of 16 years [40]. The depth of the thermocline varies between 6 and 10 meters.
In the dimictic Lake Scharmützelsee the environmental parameters temperature, oxygen and nitrate concentrations show typical annual courses with high similarities between the three studied years (Figure 1 and 4). Temperature was slowly increasing during summer and declined after mixing in fall. Oxygen dropped from high concentrations in winter to zero in the hypolimnion after stratification during spring. The nitrate concentrations showed distinct annual courses as well with peak concentrations in winter, a small decline in spring, another peak in early summer and depletion in fall.
In more detail the water temperature was lowest in winter down to 1°C (Figure 1a). After stratification in spring, temperature in the epilimnion increased up to 22 °C. In the hypolimnion, the temperature increased much slower and reached maximum temperatures of 10°C in fall (Figure1a).
The oxygen in the epilimnion and the mixed water column showed a seasonal course with maximum values in late winter and early spring (13. 7 mg L-1 in March 2011, 14.5 mg L-1 in February and March 2012) and minimum values in fall (5.8 mg L-1 in 2011 and 6.3 in 2012, Figure 1b). In the hypolimnion, oxygen dropped very fast to zero in late June after stratification and remained zero till circulation in fall (Figure 1b).
The nitrate concentration showed a strong seasonal course as well (Figure 1c) with maximum concentrations (about 400 µg∙L-1) always in winter. Concentrations are displayed in logarhytmic scale to better visualize low concentrations. In the epilimnion, nitrate dropped down in spring 2011 to 11 µg L-1 and reached a minimum value in July of 1.9 µg L-1 (Figure 1c). In 2012 it dropped to 1.2 µg L-1 in May and remained below 14 µg L-1 during summer. In 2013, nitrate concentration decreased to 2.3 µg L-1 in June and fluctuated between lower and higher values up to 40 µg L-1 in summer (Figure 1c). In fall and especially after mixing, nitrate concentration increased rapidly to the winter maximum (Figure 1c). In the hypolimnion, nitrate decreased in spring as well but remained at higher values than compared to the epilimnion (64, 48 and 92 µg∙L-1 in 2011, 2013 and 2013 respectively). In summer, an additional short maximum in June or July of up to 300 µg∙L-1 could be observed (Figure 1c). Till October it dropped down to minimum concentrations of around 3 µg∙L-1 in 2011 and 2012 and 1.7 µg∙L-1 in 2013, before it rapidly increased after mixing to maximum concentrations in winter (Figure 1c). Dissolved organic Carbon (DOC) as another major parameter for denitrifying bacteria varied between 6 and 8 mg∙L-1 in the epilimnion with two exceptions in May 2012 (14 mg∙L-1) and October 2012 (4.4 mg∙L-1). In the hypolimnion, DOC ranged in 2011 between 7 and 10 mg∙L-1 without a noticeable seasonality. In 2012 the values decreased from June to October from 9.8 to 3.7 mg∙L-1.
2.2. Sampling
Pelagic samples were taken in the years 2011, 2012 and 2013 at the long time monitoring site Rietz (RIE, 29 m depth) as pooled water samples of the entire mixed water column or separately from epi- and hypolimnion during stratification. Limnological parameters were measured and kindly provided by the Chair of Freshwater Conservation (Brandenburg University of Technology). Dissolved oxygen and temperature profiles were measured with a multiparameter probe (HYDROLAB H20, Austin, TX, USA) connected to a field computer (HUSKY Hunter, Itronix Corp., USA) in 0.5 m intervals. Mean values for the seasonal courses were calculated either for the epi- and hypolimnion separately (during stratification) or for the entire mixed water column (thereafter discussed together with epilimnion data). Nitrate and Nitrite were analysed as a combined parameter according to standard methods (DEV 1976) as described by Grüneberg et al. (2011).Nitrate and nitrite concentrations were summarized to NO3-N, since NO2 concentrations were always negligible. Dissolved organic carbon (DOC) concentrations were either provided by the Leibniz Institute of Freshwater Ecology and Inland Fisheries ore were measured by a DIMATOC® 100 (DIMATEC Analysentechnik GmbH) according to DIN 38 409 Teil 3, EN 1484.
Intact sediment cores were taken at the same site according to [41] with an Uwitec®-corer (flutter valve, acrylic glass tubes, i.d.=57 mm) parallel to the water samples and sliced into the first and second cm layer. The dry weight of the sediment at this site was between 5 and 6.5 %. The portion of organic materials (oTS, determined as loss of ignition at 550 °C) was between 24 % and 30 % of the dry weight. Water and sediment samples were cooled at 4-6°C and analysed within 12 hours.
2.3. Total Cell Counts
The total amount of viable bacterial cells was counted by fluorescence microscopy (Nikon Eclipse LV 100 with NIS Elements BR) at 20 and 40 fold magnification. 1 mL of a water sample or 0.5 to 1 ml of an appropriate diluted sediment sample (usually from the 10-3 dilution step of the MPN dilution series) was incubated with 100 µL of a Syto 9- Propidium iodide mixture (LIVE/DEAD® BacLight™, 0,33mM and 2 mM, Invitrogen Deutschland) and filtered through a counting filter (Isopore Membrane Filters, 0,2 µm GTBP, Merck, Irland) using a filtration unit (Sartorius, Göttingen, Deutschland) according to [42] and [43]. The filter was directly used for cell counting.
2.4. Most Probable Number
The number of denitrifying and nitrate reducing bacteria in water and sediment samples was measured as Most Probable Number (MPN) according to [44] from one tenfold dilution series per sample with three tubes per dilution step in Nutrient Broth II (Sifin, Berlin, Germany) with 1.5 mg L-1 KNO3 in Hungate tubes. Durham tubes were added for gas capture. Gas production and decrease or loss of nitrate, analysed according to [45], confirmed denitrification. Nitrate reduction was confirmed by Nitrite accumulation, measured by the Griess-Ilosvay methode ([46] without gas production. MPN and 95 % confidence intervals were calculated according to FDA recommendations [47].
2.5. Molecular approach for microbial enumeration
For the estimation of the total bacterial cell number and the number of denitrifying bacteria, a semi quantitative PCR method described by Rathsack et al. (2014) was used, which correlates the amount of PCR products with a calibration curve generated with known cell numbers.
For DNA extraction, 50 ml of pelagic samples were filtered as mentioned above to get enough PCR product for quantification. Filters and native sediment samples were stored at -20°C until extraction. Cells from filters were resuspended with purified water. 1.5 ml of a 1:100 dilution of the defrosted sediment were centrifuged at 12000 g and the pellet was used for DNA extraction. DNA was extracted by several application steps of extraction buffer/ chloroform and isopropanol according to [43]. The DNA samples were resuspended with 100 µL of purified water and stored at -80 °C. As a control for a constant extraction and PCR efficiency, DNA of a known cell number of Pseudomonas aeruginosa ATCC 27853 was extracted and amplified parallel.
The chromosomal 16S rDNA gene was amplified using the universal primer set 27f (5`-AgAgTTTgATC(A/C)TggCTCA-3`) and 1525r (5´-AggAggTgATCCAgCC-3`). For enumeration of denitrifying bacteria, the chromosomal DNA of the nitrite reductase nirS and nirK genes were amplified with the primer set cd3aF (5´-gT(C/G)AACgT(C/G)AAggA(A/G)AC(C/G)gg-3`) and R3cd (5´-gA(C/G)TTCgg(A/G)Tg(C/G)gTCTTgA-3`, [48]. A PCR-protocol described by [24] was used. Agarose gels were stained with enthidium bromide and the amount of the PCR-products were quantified with the software GelixOne G230 (biostep®, Jahnsdorf, Germany) in comparison with a quantification marker (MassRuler DNA Ladder Mix, Thermo Scientific, Schwerte, Germany). The initial number of gene copies was calculated with a calibration curve, which was established with known cell numbers (microscopical cell counting after cell staining with Syto 9) of P. aeruginosa ATCC 27853, which represents a widespread group of denitrifying bacteria. NirS and nirK gene copy numbers were summarised to nir gene copy number.
2.6. Statistical variance of the different methods
The MPN method is known to have a high inherent statistical error. According to Corry et al. (2012) MPN values were considered significantly different, when the 95% confidence intervals did not overlap which is in most cases a difference of roughly two log units (least significant difference =1.01). In our study, the variance of the different methods was tested with five parallel samples of a P. aeruginosa culture at 4 different cell densities each. The standard deviation of the log transformed microscopical counting was 0.05 and for the MPN results 0.41. The enumeration by PCR had a standard deviation of 0.15 log units generated from the same samples, which were extracted and amplified separately. Using the students-t statistics a difference of 1.8 log units for MPN values, 0.22 log units for microscopical counting and 0.6 log units between two gene copy numbers can be considered as significant (p=0.05).
For correlation between the log transformed cell numbers of the different enumeration approaches and between cell numbers and environmental parameters the correlation coefficient (R) of Spearman was calculated by the software Origin Pro 8.5 (Origin Lab Corporation).
3. Results
Total cell counts, numbers of cultivable nitrate reducing and denitrifying bacteria as well as the abundances of the chromosomal 16S rDNA gene copies and the nirS and nirK gene copies have been documented over the annual courses of three consecutive years from 2011 to 2013.
3.1. Epilimnion
In the epilimnion, the total microscopic cell counts ranged from 5∙105 to 7∙106 mL-1 (Figure 2a). Besides a small decline in summer 2012 and an increasing tendency by one magnitude from fall 2013 to spring 2014 there was no obvious seasonality.
The total cell numbers in the epilimnion calculated from the 16S rDNA gene copy number varied between 1.5∙105 and 2∙107 mL-1 (Figure 2a) and were comparable to the microscopic cell counts. In 2012, a slight increase from spring to summer by one magnitude was calculated followed by lower values in fall and winter. The data suggest another decline in winter 2013-2014 from 2∙107 to 2.6∙106 mL-1. However, as shown for the microscopical cell counts before, the combined results from 2011 to 2014 did not reveal an obvious seasonality in the annual courses for the 16S gene copy numbers as well (Figure 3a). Both methods detected in average the same amount of bacterial cells (difference = 0.1 log units), although the 16S cell number showed a bit more fluctuation most likely caused by the higher methodical error.
Compared to the total cell counts, a strong seasonal variation was observed in the MPN of nitrate reducing and denitrifying bacteria (Figure 2b). From April to August 2011 the MPN of nitrate reducing bacteria raised from 1.1∙ 102 to 3.8∙103 mL-1 and dropped down to 9.3∙101 mL-1 in late September, a difference of more than one magnitude (Figure 2b). In 2012 the number of nitrate reducers raised continuously from 2.1∙101 mL-1 in March to 2.4∙103 mL-1 in October, a difference of two orders of magnitudes. In winter, the MPN dropped down again by more than two orders of magnitudes from 9.3∙103 mL-1 in December 2013 to 2.3∙101 mL-1 in April 2014 (Figure 2b).
The MPN of denitrifying bacteria increased from minimal 3.6∙10-1 mL-1 in April 2011 to 2.0∙102 mL-1 in August (Figure 2b), a difference of almost three orders of magnitudes. In 2012 MPN increased again from low values (4.3 and 3.0 mL-1 in March and May respectively) to 2.3∙102 mL-1 in August with a decline afterwards to 2.8∙101 mL-1 in January, a difference of two and one orders of magnitudes respectively. Similar to the nitrate reducing bacteria, denitrifyer concentration decreased in winter from 9.3∙103 mL-1 in December 2013 to 2.3∙101 mL-1 in February 2014 (Figure 2b).
The combined results from the three years study showed a significant seasonality in the MPN of nitrate reducing and denitrifying bacteria with a maximum in winter and a decrease in spring (Figure 3b). In summer, the MPN increased to maximum values in August and October.
The difference between nitrate reducing and denitrifying bacteria calculated by the MPN approach varied between zero and more than two orders of magnitudes without an obvious seasonal course. In average (median), the difference of the log transformed values was 0.3 log units.
The number of denitrifying bacteria calculated from the sum of nirS and nirK gene copy numbers was in median 0.79 log units smaller than the 16S cell number. It ranged between 3.7∙104 (spring 2012) and 2∙106 mL-1 (fall 2012). Similar to the total cell numbers calculated from 16S rDNA, an increasing trend from spring to fall 2012 with a difference of almost two orders of magnitudes was detected (Figure 2b), although the values showed large fluctuations. During winter 2013-2014, the number of nir gene copies remained in average almost constant (Figure 2b). There were roughly four times more nirS gene copies detected than nirK.
Combining the results from 2012 to 2014, the nir gene copy numbers didn’t show any significant seasonality. There was only a slight minimum in early summer, which however might be caused by the few number of data at this time (Figure 3b). This was contrary to the significant seasonality in the MPN approach.
The average differences between microscopical and molecular enumeration on the one hand and the MPN values of denitrifyers on the other were roughly 4 log units (Figure 3c). However, the differences showed a seasonal course similar to the seasonal course of the MPN itself with an increase from 3 log units to 5 log units during winter till late spring and a decrease in summer to 3 log units (Figure 3c).
3.2. Hypolimnion
In the hypolimnion, the development of the environmental parameters and cell numbers were in parts different to the epilimnion (Figure 1, Figure 4). The displayed data of temperature, oxygen and nitrate concentrations (Figure 1a, b, c) reveal high similarities of these parameters between the three studied years. Temperature was slowly increasing during summer and declined after mixing in fall. Oxygen dropped from high concentrations in winter to zero during spring. The nitrate concentrations showed distinct annual courses as well with peak concentrations in winter, a small decline in spring, another peak in early summer and depletion in fall.
As a result of the annual variation in the physical and chemical parameters, the MPN of nitrate reducing and denitrifyining bacteria showed a significant annual course as well. The MPN numbers were highest in winter, declined in spring, had another maximum in late summer followed again by a decline in fall till the end of stratification (Figure 4b and Figure 5a).
In detail, the annual course can be described as follows. After a smaller drop of the nitrate concentration in spring, temperature and nitrate concentration increased in June. The MPN numbers of nitrate reducing bacteria raised correspondingly from 1.1∙102 to 9.3 ∙102 mL-1 in 2011 and from 2.1∙101 to 9.3∙102 mL-1 in 2012 (Figure 4b), a difference of almost two magnitudes. The MPN of denitrifying bacteria increased as well, from less than 1 ml-1 in 2011 and 2 ml-1 in 2012 to 1.5∙102 and 2.3∙102 ml-1 in late summer 2011 and 2012 respectively. This was a significant difference of two magnitudes. In late summer and fall, the nitrate concentration dropped and the MPN of nitrate reducing bacteria also declined to minimum values of 1.5∙102 mL-1 in 2011 and 9.3∙101 mL-1 in 2012 before autumn circulation (Figure 4b). The number of denitrifyers decreased as well to 4.3 ml-1 in October 2012. Low values, 4.3∙101 ml-1 for nitrate reducing bacteria and 2.3∙101 ml-1 for denitrifying bacteria, were also detected in October 2013 (Figure 4b). In winter, the number of nitrate reducing bacteria reached another maximum of 1.5∙103 (January 2013) and 9.3∙103 mL-1 (December 2013). This maximum was also detectable for denitrifying bacteria, at least in 2013. In spring 2014 both groups dropped to low values of 4.3 and 2.3 mL-1, a difference of more than two magnitudes.
Contrary to the epilimnion, the 16S and nir cell numbers showed a similar seasonal trend as the MPN (Figure 4a,b), but with a larger amplitude. The lowest cell numbers were measured in spring 2012 with 1∙105 mL-1 (16S) and 4∙104 mL-1 (nir). During the following stratification, the 16S and nir cell numbers increased up to 1.7∙106 mL-1 and 2.4∙106 mL-1 in August and decreased thereafter till mixing in fall down to 2∙105 and 4∙104 respectively. In 2013, the 16S and nir cell numbers reached maximal values of 1∙107 and 1∙108 in August, maybe due to higher nitrate concentrations compared to 2012. Combining all three years, a maximum in summer was always followed by a strong decline to a minimum in fall (Figure 5a). There was eventually another slight 16S cell number peak in winter (Figure 5a). Over all, the 16S cell number showed a variability of two orders of magnitudes, the nir cell number of 3.5 magnitudes. The total microscopic cell counts ranged from 7∙105 to 5∙106 mL-1 without a detectable seasonal course.
The differences between denitrifying and nitrate reducing bacteria ranged from zero to 3 log units (average 0.9 log units) without a seasonal dependency. The microscopical and molecular enumerations on one hand and the MPN values on the other differed by roughly 4 log units. This was similar to the epilimnion. Between the nir gene copy number and the number of cultivable denitrifying bacteria, a seasonality in the difference is obvious (Figure 5b). In winter and fall, when the MPN was highest, the difference was lowest (1.2 log units in December, 2.6 in September Figure 5b). At low MPN in spring 2011 and 2013, the difference was highest (4.8 log units).
Similar to the epilimnion, the microscopical and 16S cell numbers in the hypolimnion were almost equal (difference of the log transformed values = 0.1 log units). The nir gene copy numbers were in average 0.45 log units smaller than the related 16S gene copy numbers.
3.3. Sediment
As expected, the bacterial cell numbers in the sediment were much higher than in the pelagial. In 2011, MPN of nitrate reducing bacteria were higher in summer (4.8∙105 mL-1) than in November (6.8∙104 mL-1). In 2011 a trend of decline from 2∙106 mL-1 in spring to 4∙104 mL-1 in fall could be observed, a difference of almost two orders of magnitudes (Figure 6). The number of denitrifying bacteria ranged between 2.6∙105 mL-1 in June 2011, 1.5∙104 mL-1 in November 2011 and 2.1∙104 mL-1 in May 2012. A seasonal course could not be deduced. The total microscopic cell numbers were in a range between 8∙107 to 8∙109 mL-1 with no detectable seasonal course as well (Figure 6). In contrary to the pelagial, the cell numbers calculated from the 16S rDNA and nir gene copy numbers in the sediments were two to four orders of magnitudes higher than the corresponding microscopic cell counts (Figure 6) and varied between 5.6∙1011 mL-1 and 2.5∙1014 mL-1. More than 99 % of the extracted DNA was obviously not related to intact bacteria. Although the nir results suggest higher values between fall and spring and lower values in late summer with a difference of two orders of magnitudes, a seasonal dependency is difficult to detect. In median the nir values are similar to the 16S values (median difference less than 0.1 log units), although in some cases nir genes were higher than 16S, which eventually is caused by methodical uncertainties.
A statistical analysis revealed some correlations in the data set. The gene copy numbers of 16S and nir were correlated (epilimnion: 0.65, p=0.004, n=17, hypolimnion: 0.57, p=0.006, n=20) as well as the MPN of nitrate reducing and denitrifying bacteria (epilimnion: 0.69, p=0.02, n=16, hypolimnion: 0.63, p=0.04, n=18). Surprisingly, microscopical cell numbers and MPN of denitrifying and nitrate reducing bacteria were negatively correlated (epilimnion: -0.5, p=0.02 and 0.03, n=17, hypolimnion: -0.4 and -0.5, p=0.07 and 0.02, n=18). In the hypolimnion from March 2012 to January 2013, MPN of nitrate reducing bacteria and denitrifying bacteria were correlated with nir gene copy numbers (0.8, p=0.027 and 0.7, p=0.08, n=7, Figure 5b). Beside this, no further correlation between log transformed microscopical, PCR based and MPN cell numbers could be found (p>0.15, n=14 to 18). Significant correlations between log transformed cell numbers and the environmental factors temperature, DOC and nitrate could only be found between MPN of denitrifying bacteria and temperature in the epilimnion (0.65, p= 0.01, n=14), when excluding one outlier in December 2013 (Figure 2, otherwise 0.47, p=0.07, n=15), and between MPN of nitrate reducing bacteria at the sediment surface and nitrate concentration of the hypolimnion (0.6. p=0.04, n=10, Figure 6). In addition, from May to October 2012 MPN of nitrate reducing bacteria and nitrate concentration in the epilimnion were correlated as well (0.9, p=0.03, n=5, Figure 2).
4. Discussion
4.1. Seasonal course of bacteria and dependency on environmental parameters
The seasonal courses of denitrifying and nitrate reducing bacteria in different compartments of a dimictic lake have been measured by different methods. As shown, the seasonality was best displayed by the cultivation based MPN technique. Although this method has a high inherent statistical error, significant differences of more than two orders of magnitude could be detected within the annual courses in the epi- and hypolimnion. The seasonal courses of the cell numbers differed between epilimnion, hypolimnion and sediment, due to different driving environmental parameters.
In the epilimnion, the MPN of nitrate reducing and denitrifying bacteria increased from minimal values in spring at low temperatures till fall up to two orders of magnitudes at parallel increasing temperatures and increasing nitrate concentrations. Although both parameters are correlated with the MPN at this time, the growth during summer seems to be driven by temperature and oxygen rather than by the availability of nitrate, since many denitrifying bacteria prefer oxygen, which was present throughout the year. In winter at low temperatures the MPN of nitrate reducing bacteria dropped back to minimal values.
DOC as another major factor for heterotrophic bacteria was fairly constant at limiting values between 6 and 8 mg∙L-1, much lower than the Km-value (about 70 mg L-1, [49]. Thus, it could not be proven to be responsible for the seasonal cell number variation. However, the increase of the MPN in summer and fall might at least partially be influenced by higher carbon supply from growing and degrading phytoplankton. This can be concluded from the reported variation in the Chlorophyll concentration [50]. Here the combination of increased supply, immediate consumption and increasing bacterial concentrations would keep the DOC values at constant low concentrations.
The described seasonal course in the hyplimnion was later confirmed by a more detailed sampling campaign in 2015 [51]. In contrast to the epilimnion, in the hypolimnion nitrate was the main electron acceptor for denitrifying bacteria, since oxygen was depleted very fast in spring in part by nitrification [51]. At increasing temperature and nitrate concentrations, the MPN cell numbers of nitrate reducing bacteria in the hypolimnion also increased by almost two orders of magnitudes from spring till summer and fell back to low values parallel to nitrate depletion in late summer. However, no direct correlation could be found between denitrifying bacteria and environmental parameters. This is most likely caused by changing limiting parameters and offsets between environmental conditions and the slow responds of the cell numbers in the colder hypolimnion. This can be seen e.g. in the delay of the MPN compared to the course of nitrate in the combined annual course (Figure 5).
For the upper layer of the sediment, similar conclusions can be drawn. The number of nitrate reducing bacteria was correlated to the nitrate concentration and declined by almost two orders of magnitudes in summer at depleting nitrate concentrations.
4.2. Explanations on the seasonal courses
The seasonal course of the nir cell number in the epilimnion was less pronounced than the MPN. In summer 2012 it increased by 1.8 orders of magnitudes compared to two and three in the MPN. At impairing environmental conditions and low temperatures, the MPN significantly decreased while the nir cell number remained constant (best to see in winter 2014). As a result, the differences between nir gene copy numbers and the MPN of and nitrate reducing and denitrifying bacteria respectively varied between one and 1.9 log units at the end of favourable conditions and up to three and even five orders of magnitudes at impairing conditions. This observation can be explained at least in two different ways.
A first explanation could be that only a fraction of all nitrate reducing and denitrifying species are cultivable thus causing the difference between MPN and nir gene copy numbers. If moreover is assumed that bacteria multiplied at favourable condition and died at non favourable conditions, the multiplication of cultivable cells would only slightly affect the nir cell number, due to the large difference between MPN and nir cell numbers at the starting point. However, in Lake Scharmützelsee the ratios between the nir gene abundance and the number of cultivable bacteria were changing by more than two orders of magnitudes during the seasonal course. To interpret these findings in the context of the explanation above, a faster growth of cultivable species compared to non cultivable cells would be necessary. On the other hand, mainly active cultivable cells need to die at non favourable conditions in winter to cause the increasing difference between nir cell numbers and MPN. Such assumptions seem to be rather improbable.
The multiplication and degradation of cells in front of a larger pool of cell free intact DNA or dead DNA seems also less likely. The DNA has been degraded in early spring parallel to constant or even increasing microscopical cell counts as can be concluded from the decreasing 16S gene copy numbers.
A second explanation seems more likely. We assume that a large portion of denitrifying bacteria changes from a viable into a dormant or VBNC state [52] during non-favourable conditions (e.g. at cold temperatures as seen in spring 2014). As a result, the MPN counts varied strongly between adequate and reverse conditions within the annual course, whereas the cell numbers detected by molecular or microscopic methods showed less seasonal variability.
According to our assumption, dormant denitrifying bacteria become active again at favourable conditions as discussed in several other studies before [43,52]. This explanation is supported by laboratory tests reported by [43] with pure cultures of denitrifying Pseusdomonas at growing and starving conditions. Alternatively, several investigators also discussed the possibility that a relative small number of cells remained vital under unfavourable conditions or become active independent of environmental factors and start to multiply at adequate environmental conditions [53]. This would also result in a relatively smaller increase in the nir gene copy number due to the large background.
Another explanation, the formation of micro colonies at non favourable conditions as assumed by [54] for soil nitrifying bacteria can be excluded. The microscopical analysis did not reveal those clusters at least in pelagic samples.
Combining the results from the cited laboratory tests and this study it can be argued, that the often reported large difference between cultivation based and other cell counting methods (e.g. [29]) are caused at least in parts by a proportion of cells in the VBNC or dormant state and not exclusively by the occurrence of non-cultivable species. At the end of growing periods, cell counts obtained either from MPN or gene copy numbers matched quite well.
In contrast to the epilimnion, the seasonal increase and decrease of the MPN, nir and 16S gen copy numbers in the hypolimnion were comparable. This could be explained by a higher feeding pressure by bacterivourus placton like ciliates and flagellates in the hypolimnion compared to the epilimnion, as reported by [55]. High feeding pressure would remove non active and therefore non multiplying bacteria including their DNA resulting in only low amounts of dormant or VBNC cells and a parallel course of MPN and nir cell numbers.
For the sediment it is difficult to draw conclusions. There was a decline in the nir gene copy number and the MPN from December 2011 to summer 2012 at decreasing nitrate concentrations while the 16S barely changes. Death and eventually lysis of denitrifying cells and a switch between active and dormant cells could not be distinguished, since the total viable cell number was several orders of magnitudes higher than the number of denitrifying ones. However, similar to the pelagial the MPN values reflected the changing environmental conditions best.
In the pelagial there was a good match between microscopic and molecular total cell numbers with generally less than one magnitude difference. This can be explained by a low amount of cell free intact genetic material. Therefore, both methods seem to be appropriate to estimate the total cell number in pelagic environments and other systems with fast degradation of free DNA. The nir gene copy number reflects the total amount of denitrifying cells independent of their cultivable status.
In contrast to the pelagial, microscopic cell numbers and gene copy numbers in the sediment differed by at least two orders of magnitudes. One reason might be a possible underestimation of cells in sediment particles, although this effect proved to be very small [56]. Another more likely explanation is a high abundance of intact DNA in dead cells or extracellular DNA bound to particles. Previous studies reported that the amount of extracellular DNA adsorbed to sediment particles can be 10 to 70 times higher than the cellular DNA concentrations [57]. In those environments with slow degradation of free DNA, the estimation of cell abundance by measuring genomic gene copy number with calibration curves made by log phase cultures [58]) will lead to an overestimation by eventually several orders of magnitudes. These limits of molecular techniques for measuring active metabolizing cells have been already described by other investigators before. [59] demonstrated that the success of disinfection could be monitored by the depletion of cultivable cells while the number of gene copies only slightly decreased. [11] described a correlation between gene copies and potential enzymatic activity for only some reactions in glacier soil and argued with inactive populations due to substrate shortage on one side and with uncertainties in the potential activity measurement due to shifting population on the other. The discrepancies in correlation of nir gene copies and denitrification activities reported by [5] might in part also be explained by an inactive portion of genetic material. Here other molecular techniques like the detection of m-RNA [60] seems a better choice, since its half-time is short [61]. However, the m-RNA amount per cell can change depending on starvation and resuscitation [62] and cannot directly be related to metabolic activity [63]. Therefore a combination of both cultivation and molecular techniques seems necessary as recommended earlier [22,30]. As an alternative, several investigators tried to separate extracellular DNA during the extraction procedure [57].
As shown in this paper, the determination of total cell numbers either by direct cell counting or by the 16S rDNA approach might be helpful for the interpretation of the results especially in sediments. In controlled systems like laboratory experiments, the microscopic cell number seems to be an efficient parameter too.
The study demonstrates that the population of denitrifying bacteria can change by several orders of magnitudes within a seasonal course. There are differences between the epi- and hypolimnion due to different driving factors. To estimate the seasonal variability of the active metabolizing cell number especially in highly dynamic systems with changing environmental conditions like nutrient or oxygen concentrations, cultivation based techniques might give a better understanding than molecular approaches. These results can be of particular importance, when using the cell number for the calculation of in situ metabolic activities and turnover rates [51]. On the other hand, to estimate the growth dynamic of a bacterial population, the quantification of the gene copy number might be an important tool as well, if a major portion of the cells simply switch between dormant and active condition, because the MPN number will increase without multiplication of cells. However, this will only be suitable for environments with a small background of cell free or dead DNA.
thors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
5. Conclusions
The study demonstrates that the population of denitrifying bacteria in a dimictic lake can change by several orders of magnitudes within a seasonal course. There are differences between the epi- and hypolimnion due to different driving factors. To estimate the seasonal variability of the active metabolizing cell number especially in highly dynamic systems with changing environmental conditions like nutrient or oxygen concentrations, cultivation based techniques might give a better understanding than classic molecular approaches. These results can be of particular importance, when using the cell number for the calculation of in situ metabolic activities and turnover rates [51]. On the other hand, to estimate the growth dynamic of a bacterial population, the quantification of the gene copy number might be an important tool as well, if a major portion of the cells simply switch between dormant and active condition, because the MPN number will increase without multiplication of cells. However, this will only be suitable for environments with a small background of cell free or dead DNA.
Author Contributions
Conceptualization: Marion Martienssen; methodology: Jörg Böllmann Marion Martienssen; validation: Jörg Böllmann, Marion Martienssen; formal analysis, Jörg Böllmann; investigation: Jörg Böllmann; resources: Marion Martienssen; data curation: Jörg Böllmann; writing—original draft preparation: Jörg Böllmann; writing—review and editing: Jörg Böllmann, Marion Martienssen; visualization: Jörg Böllmann; supervision: Marion Martienssen; project administration: Marion Martienssen; funding acquisition: Marion Martienssen. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the German Federal Ministry of Education and Research (Fkz. 033L041D). The APC was funded in parts by the Brandenburgische Technische Universität Cottbus-Senftenberg, Germany.
Data Availability Statement
Raw Data on Cell Numbers and Physico-Chemical Parameters are available at DOI: 10.13140/RG.2.2.28332.85122 .
Acknowledgments
We thank the colleges of the Department of Freshwater Conservation in Bad Saarow (Björn Grünerberg, Jacqueline Rücker, Ingo Henschke) for sampling, data collection and good collaboration.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Coskuner, G.; Ballinger, S.J.; Davenport, R.J.; Pickering, R.L.; Solera, R.; Head, I.M.; Curtis, T.P. Agreement between Theory and Measurement in Quantification of Ammonia-Oxidizing Bacteria. Appl. Environ. Microbiol. 2005, 71, 6325–6334. [Google Scholar] [CrossRef] [PubMed]
- Muela, A.; Gorostiza, I.; Iriberri, J.; Egea, L. Indicative Parameters of Denitrification in River Sediments. Acta Hydrochim. Et Hydrobiol. 1988, 16, 157–163. [Google Scholar] [CrossRef]
- Baldwin, B.R.; Nakatsu, C.H.; Nies, L. Detection and Enumeration of Aromatic Oxygenase Genes by Multiplex and Real-Time PCR. Appl. Environ. Microbiol. 2003, 69, 3350–3358. [Google Scholar] [CrossRef]
- Röling, W.F.M. Do Microbial Numbers Count? Quantifying the Regulation of Biogeochemical Fluxes by Population Size and Cellular Activity. FEMS Microbiol. Ecol. 2007, 62, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Trippett, C.; Dodds, W.K.; O’Brien, J.M.; Banner, E.B.K.; Head, I.M.; Smith, M.S.; Yang, R.K.; Knapp, C.W. Correlations between in Situ Denitrification Activity and Nir-Gene Abundances in Pristine and Impacted Prairie Streams. Environ. Pollut. 2010, 158, 3225–3229. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.B.; Rysgaard, S.; Sloth, N.P.; Dalsgaard, T.; Schwaerter, S. Sediment Mineralization, Nutrient Fluxes, Denitrification and Dissimilatory Nitrate Reduction to Ammonium in an Estuarine Fjord with Sea Cage Trout Farms. Aquat. Microb. Ecol. 2000, 21, 73–84. [Google Scholar] [CrossRef]
- Magalhães, C.; Bano, N.; Wiebe, W.J.; Bordalo, A.A.; Hollibaugh, J.T. Dynamics of Nitrous Oxide Reductase Genes (nosZ) in Intertidal Rocky Biofilms and Sediments of the Douro River Estuary (Portugal), and Their Relation to N-Biogeochemistry. Microb Ecol 2008, 55, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Gihring, T.M.; Canion, A.; Riggs, A.; Huettel, M.; Kostka, J.E. Denitrification in Shallow, Sublittoral Gulf of Mexico Permeable Sediments. Limnol. Oceanogr. 2010, 55, 43–54. [Google Scholar] [CrossRef]
- Chon, K.; Chang, J.-S.; Lee, E.; Lee, J.; Ryu, J.; Cho, J. Abundance of Denitrifying Genes Coding for Nitrate (narG), Nitrite (nirS), and Nitrous Oxide (nosZ) Reductases in Estuarine versus Wastewater Effluent-Fed Constructed Wetlands. Ecol. Eng. 2011, 37, 64–69. [Google Scholar] [CrossRef]
- McCrackin, M.L.; Elser, J.J. Denitrification Kinetics and Denitrifier Abundances in Sediments of Lakes Receiving Atmospheric Nitrogen Deposition (Colorado, USA). Biogeochemistry 2012, 108, 39–54. [Google Scholar] [CrossRef]
- Brankatschk, R.; Töwe, S.; Kleineidam, K.; Schloter, M.; Zeyer, J. Abundances and Potential Activities of Nitrogen Cycling Microbial Communities along a Chronosequence of a Glacier Forefield. ISME J. 2010, 5, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Abell, G.; Ross, D.; Keane, J.; Oakes, J.; Eyre, B.; Robert, S.; Volkman, J. Nitrifying and Denitrifying Microbial Communities and Their Relationship to Nutrient Fluxes and Sediment Geochemistry in the Derwent Estuary, Tasmania. Aquat. Microb. Ecol. 2013, 70, 63–75. [Google Scholar] [CrossRef]
- Bellini, M.I.; Gutiérrez, L.; Tarlera, S.; Scavino, A.F. Isolation and Functional Analysis of Denitrifiers in an Aquifer with High Potential for Denitrification. Syst. Appl. Microbiol. 2013, 36, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Veraart, A.J.; Audet, J.; Dimitrov, M.R.; Hoffmann, C.C.; Gillissen, F.; de Klein, J.J.M. Denitrification in Restored and Unrestored Danish Streams. Ecol. Eng. 2013, 66, 129–140. [Google Scholar] [CrossRef]
- Michotey, V.; Mejean, V.; Bonin, P. Comparison of Methods for Quantification of Cytochrome Cd1-Denitrifying Bacteria in Environmental Marine Samples. Appl. Environ. Microbiol. 2000, 66, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.J.; Harris, D.; Dollhopf, S.L.; Gross, K.L.; Prosser, J.I.; Paul, E.A. Effects of Agronomic Treatments on Structure and Function of Ammonia-Oxidizing Communities. Appl. Environ. Microbiol. 2000, 66, 5410–5418. [Google Scholar] [CrossRef] [PubMed]
- Torsvik, V.; Øvreås, L. Microbial Diversity and Function in Soil: From Genes to Ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Both, G.J.; Gerards, S.; Laanbroek, H.J. Enumeration of Nitrite-Oxidizing Bacteria in Grassland Soils Using a Most Probable Number Technique: Effect of Nitrite Concentration and Sampling Procedure. FEMS Microbiol. Lett. 1990, 74, 277–285. [Google Scholar] [CrossRef]
- Philippot, L. Use of Functional Genes to Quantify Denitrifiers in the Environment. Biochem. Soc. Trans. 2006, 34, 101–103. [Google Scholar] [CrossRef]
- Noble, R.T.; Weisberg, S.B.; Leecaster, M.K.; McGee, C.D.; Ritter, K.; Walker, K.O.; Vainik, P.M. Comparison of Beach Bacterial Water Quality Indicator Measurement Methods. Environ. Monit. Assess. 2003, 81, 301–312. [Google Scholar] [CrossRef]
- Belser, L.; Mays, E. Use of Nitrifier Activity Measurements to Estimate the Efficiency of Viable Nitrifier Counts in Soils and Sediments. Appl. Environ. Microbiol. 1982, 43, 945–948. [Google Scholar] [CrossRef]
- Degrange, V.; Bardin, R. Detection and Counting of Nitrobacter Populations in Soil by PCR. Appl. Environ. Microbiol. 1995, 61, 2093–2098. [Google Scholar] [CrossRef]
- Scala, D.J.; Kerkhof, L.J. Nitrous Oxide Reductase (nosZ) Gene-Specific PCR Primers for Detection of Denitrifiers and Three nosZ Genes from Marine Sediments. FEMS Microbiol. Lett. 1998, 162, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Rathsack, K.; Böllmann, J.; Martienssen, M. Comparative Study of Different Methods for Analyzing Denitrifying Bacteria in Fresh Water Ecosystems. J. Water Resour. Prot. 2014, 06, 609–617. [Google Scholar] [CrossRef]
- von Wintzingerode, F.; Göbel, U.B.; Stackebrandt, E. Determination of Microbial Diversity in Environmental Samples: Pitfalls of PCR-Based rRNA Analysis. FEMS Microbiol. Rev. 1997, 21, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Freeman, W.M.; Walker, S.J.; Vrana, K.E. Quantitative RT-PCR: Pitfalls and Potential. BioTechniques 1999, 26, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Ginzinger, D.G. Gene Quantification Using Real-Time Quantitative PCR: An Emerging Technology Hits the Mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef]
- Smith, C.J.; Nedwell, D.B.; Dong, L.F.; Osborn, A.M. Evaluation of Quantitative Polymerase Chain Reaction-Based Approaches for Determining Gene Copy and Gene Transcript Numbers in Environmental Samples. Environ. Microbiol. 2006, 8, 804–815. [Google Scholar] [CrossRef]
- Schulz, S.; Peréz-de-Mora, A.; Engel, M.; Munch, J.C.; Schloter, M. A Comparative Study of Most Probable Number (MPN)-PCR vs. Real-Time-PCR for the Measurement of Abundance and Assessment of Diversity of alkB Homologous Genes in Soil. J. Microbiol. Methods 2010, 80, 295–298. [Google Scholar] [CrossRef]
- Klein, D. Quantification Using Real-Time PCR Technology: Applications and Limitations. Trends Mol. Med. 2002, 8, 257–260. [Google Scholar] [CrossRef]
- Rompré, A.; Servais, P.; Baudart, J.; de-Roubin, M.-R.; Laurent, P. Detection and Enumeration of Coliforms in Drinking Water: Current Methods and Emerging Approaches. J. Microbiol. Methods 2002, 49, 31–54. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Hallin, S. Finding the Missing Link between Diversity and Activity Using Denitrifying Bacteria as a Model Functional Community. Curr. Opin. Microbiol. 2005, 8, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Bjornsdottir-Butler, K.; Jones, J.L.; Benner Jr., R. A.; Burkhardt III, W. Quantification of Total and Specific Gram-Negative Histamine-Producing Bacteria Species in Fish Using an MPN Real-Time PCR Method. Food Microbiol. 2011, 28, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Kayashima, T.; Suzuki, H.; Maeda, T.; Ogawa, H.I. Real-Time PCR for Rapidly Detecting Aniline-Degrading Bacteria in Activated Sludge. Chemosphere 2013, 91, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Picozzi, C.; Foschino, R.; Di Pilato, P. Exploiting a MPN-PCR Technique to Quantify Escherichia Coli in Minced Meat. Ann. Microbiol. 2004, 54, 343–349. [Google Scholar]
- O’Connor, B.L.; Hondzo, M.; Dobraca, D.; LaPara, T.M.; Finlay, J.C.; Brezonik, P.L. Quantity-Activity Relationship of Denitrifying Bacteria and Environmental Scaling in Streams of a Forested Watershed. J. Geophys. Res. 2006, G04014, 2005–2012. [Google Scholar] [CrossRef]
- Barer, M.R.; Harwood, C.R. Bacterial Viability and Culturability. Adv. Microb. Physiol. 1999, 41, 93–137. [Google Scholar] [PubMed]
- Oliver, J.D. Recent Findings on the Viable but Nonculturable State in Pathogenic Bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Nixdorf, B.; Rektins, A.; Mischke, U. Standards and Thresholds of the EU Water Framework Directive (WFD) — Phytoplankton and Lakes. In Standards and Thresholds for Impact Assessment; Schmidt, M., Glasson, J., Emmelin, L., Helbron, H., Eds.; Environmental Protection in the European Union; Springer Berlin Heidelberg: Berlin, Heidelberg, 2008; ISBN 978-3-540-31140-9. [Google Scholar]
- Grüneberg, B.; Rücker, J.; Nixdorf, B.; Behrendt, H. Dilemma of Non-Steady State in Lakes - Development and Predictability of in-Lake P Concentration in Dimictic Lake Scharmützelsee (Germany) after Abrupt Load Reduction. Int. Rev. Hydrobiol. 2011, 96, 599–621. [Google Scholar] [CrossRef]
- Kleeberg, A.; Grüneberg, B. Phosphorus Mobility in Sediments of Acid Mining Lakes, Lusatia, Germany. Ecol. Eng. 2005, 24, 89–100. [Google Scholar] [CrossRef]
- Hobbie, J.E.; Daley, R.J.; Jasper, S. Use of Nuclepore Filters for Counting Bacteria by Fluorescence Microscopy. Appl Env. Microbiol 1977, 33, 1225–1228. [Google Scholar] [CrossRef]
- Rathsack, K.; Böllmann, J.; Martienssen, M. Comparative Study of Different Methods for Analyzing Denitrifying Bacteria in Fresh Water Ecosystems. JWARP 2014, 06, 609–617. [Google Scholar] [CrossRef]
- Michels, J.; Stuhrmann, M.; Frey, C.; Koschitzky, H.-P. Handlungsempfehlungen Mit Methodensammlung, Natürliche Schadstoffminderung Bei Der Sanierung von Altlasten; Dechema e.V., VEGAS, Institut für Wasserbau, Universität Stuttgart: Frankfurt a.M, 2008; ISBN -13. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Bremner, J.M. Inorganic Forms of Nitrogen. In Agronomy Monographs; Norman, A.G., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1965; pp. 1179–1237. ISBN 978-0-89118-204-7. [Google Scholar]
- Garthright, W.E.; Blodgett, R.J. FDA’s Preferred MPN Methods for Standard, Large or Unusual Tests, with a Spreadsheet. Food Microbiol. 2003, 20, 439–445. [Google Scholar] [CrossRef]
- Throbäck, I.N.; Enwall, K.; Jarvis, Ãs. ; Hallin, S. Reassessing PCR Primers Targeting nirS, nirK and nosZ Genes for Community Surveys of Denitrifying Bacteria with DGGE. FEMS Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Martienssen, M.; Schöps, R. Population Dynamics of Denitrifying Bacteria in a Model Biocommunity. Water Res. 1999, 33, 639–646. [Google Scholar] [CrossRef]
- Grüneberg, B.; Rücker, J.; Nixdorf, B.; Behrendt, H. Dilemma of Non-Steady State in Lakes - Development and Predictability of in-Lake P Concentration in Dimictic Lake Scharmützelsee (Germany) after Abrupt Load Reduction. Int. Rev. Hydrobiol. 2011, 96, 599–621. [Google Scholar] [CrossRef]
- Martienssen, M.; Böllmann, J.; Nixdorf, B.; Rathsack, K. Calculation of Hypolimnic Denitrification in a Dimictic Freshwater Lake during Summer Stratification. Aquat. Microb. Ecol. 2019, 83, 189–201. [Google Scholar] [CrossRef]
- Fakruddin, Md.; Mannan, K.S.B.; Andrews, S. Viable but Nonculturable Bacteria: Food Safety and Public Health Perspective. ISRN Microbiol. 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Epstein, S. The Phenomenon of Microbial Uncultivability. Curr. Opin. Microbiol. 2013, 16, 636–642. [Google Scholar] [CrossRef]
- Berg, P. Nitrifier Populations and Nitrification Rates in Agricultural Soil; Rapport - Sveriges lantbruksuniversitet, Institutionen för mikrobiologi, Ed.; Sveriges Lantbruksuniversitet: Uppsala, 1986; ISBN 91-576-2647-2. [Google Scholar]
- Bręk-Laitinen, G.; López Bellido, J.; Ojala, A. Response of a Microbial Food Web to Prolonged Seasonal Hypoxia in a Boreal Lake. Aquat. Biol. 2012, 14, 105–120. [Google Scholar] [CrossRef]
- Böllmann, J.; Rathsack, K.; Martienssen, M. The Precision of Bacterial Quantification Techniques on Different Kinds of Environmental Samples and the Effect of Ultrasonic Treatment. J. Microbiol. Methods 2016, 126, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Danovaro, R.; Dell’Anno, A. Simultaneous Recovery of Extracellular and Intracellular DNA Suitable for Molecular Studies from Marine Sediments. Appl. Environ. Microbiol. 2005, 71, 46–50. [Google Scholar] [CrossRef]
- Fredslund, L.; Ekelund, F.; Jacobsen, C.S.; Johnsen, K. Development and Application of a Most-Probable-Number-PCR Assay To Quantify Flagellate Populations in Soil Samples. Appl. Environ. Microbiol. 2001, 67, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Buttner, M.P.; Cruz, P.; Stetzenbach, L.D.; Klima-Comba, A.K.; Stevens, V.L.; Cronin, T.D. Determination of the Efficacy of Two Building Decontamination Strategies by Surface Sampling with Culture and Quantitative PCR Analysis. Appl. Environ. Microbiol. 2004, 70, 4740–4747. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, F.H.; Banat, I.M.; Marchant, R. Detection and Quantification of Gene Expression in Environmental Bacteriology. Appl. Environ. Microbiol. 2004, 70, 3795–3806. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Kjelleberg, S. The Role of RNA Stability during Bacterial Stress Responses and Starvation. Minireview. Environ. Microbiol. 2000, 2, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, A.; Schmidt, I.; Saunders, A.M.; Nicolaisen, M.H. Influence of Starvation on Potential Ammonia-Oxidizing Activity and amoA mRNA Levels of Nitrosospira Briensis. Appl. Environ. Microbiol. 2005, 71, 1276–1282. [Google Scholar] [CrossRef]
- Bustin, S.A. Quantification of mRNA Using Real-Time Reverse Transcription PCR (RT-PCR): Trends and Problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef]
Figure 1.
Courses of temperature (a), oxygen (b) and nitrate concentration (c) in the epilimnion (grey line) and hypolimnion (black line) of lake Scharmützelsee from 2012 to 2013.
Figure 1.
Courses of temperature (a), oxygen (b) and nitrate concentration (c) in the epilimnion (grey line) and hypolimnion (black line) of lake Scharmützelsee from 2012 to 2013.
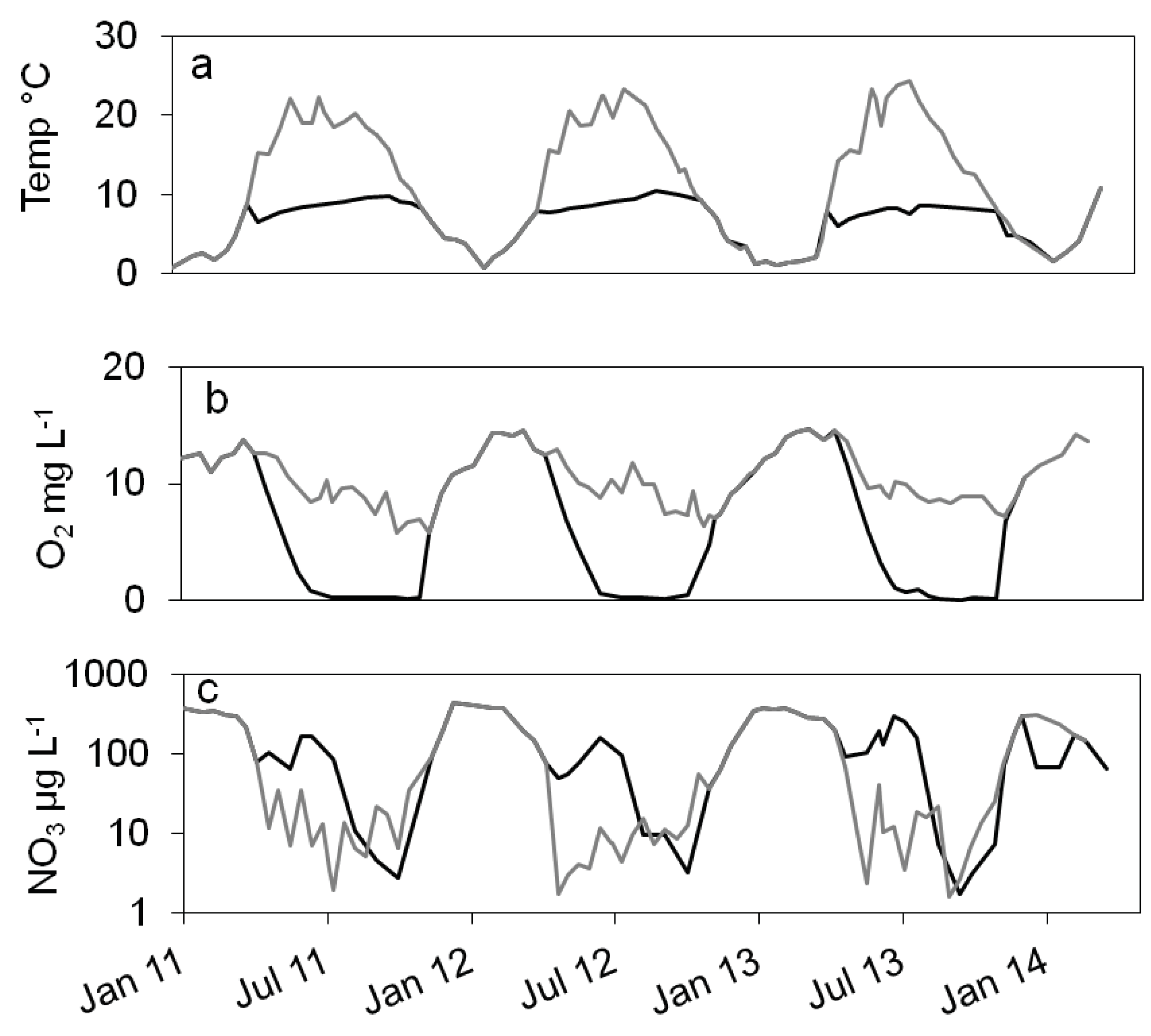
Figure 2.
Courses of nitrate concentration and cell numbers in the epilimnion of lake Scharmützelsee from 2012 to 2014: a: microscopical cell counting ( ), 16S rDNA gene copy number (
), 16S rDNA gene copy number ( ), b: MPN of nitrate reducing bacteria (
), b: MPN of nitrate reducing bacteria ( ) and denitrifying bacteria (
) and denitrifying bacteria ( ), sum of nirS and nirK (
), sum of nirS and nirK ( ), nitrite concentration (grey line).
), nitrite concentration (grey line).
), 16S rDNA gene copy number (), b: MPN of nitrate reducing bacteria () and denitrifying bacteria (), sum of nirS and nirK (), nitrite concentration (grey line).
Figure 2.
Courses of nitrate concentration and cell numbers in the epilimnion of lake Scharmützelsee from 2012 to 2014: a: microscopical cell counting (), 16S rDNA gene copy number (), b: MPN of nitrate reducing bacteria () and denitrifying bacteria (), sum of nirS and nirK (), nitrite concentration (grey line).
), 16S rDNA gene copy number (), b: MPN of nitrate reducing bacteria () and denitrifying bacteria (), sum of nirS and nirK (), nitrite concentration (grey line).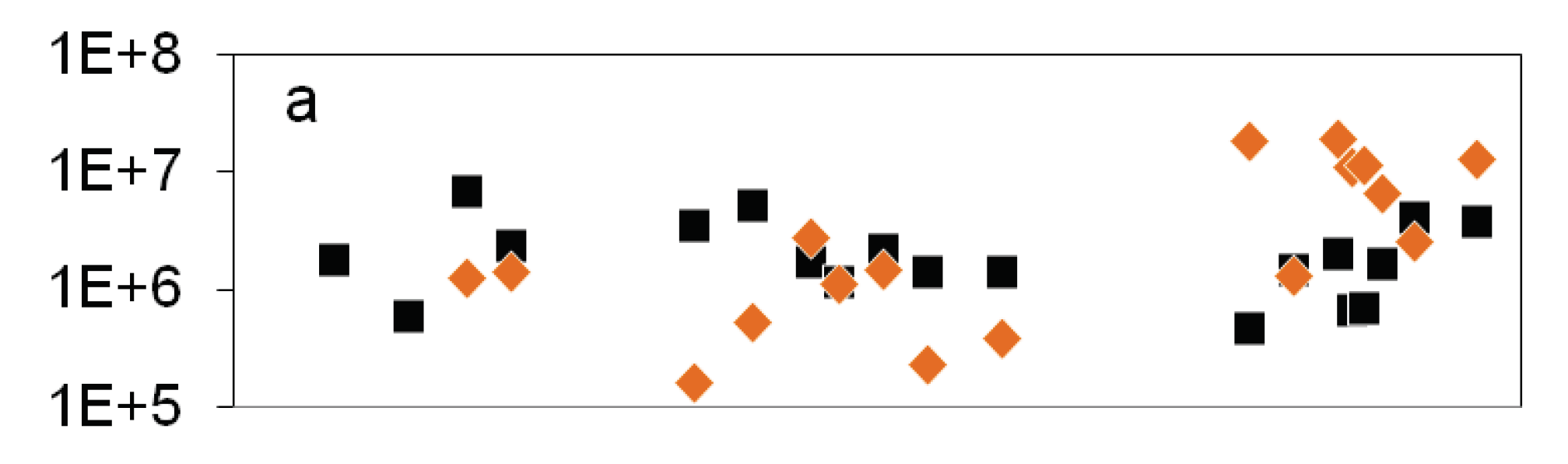
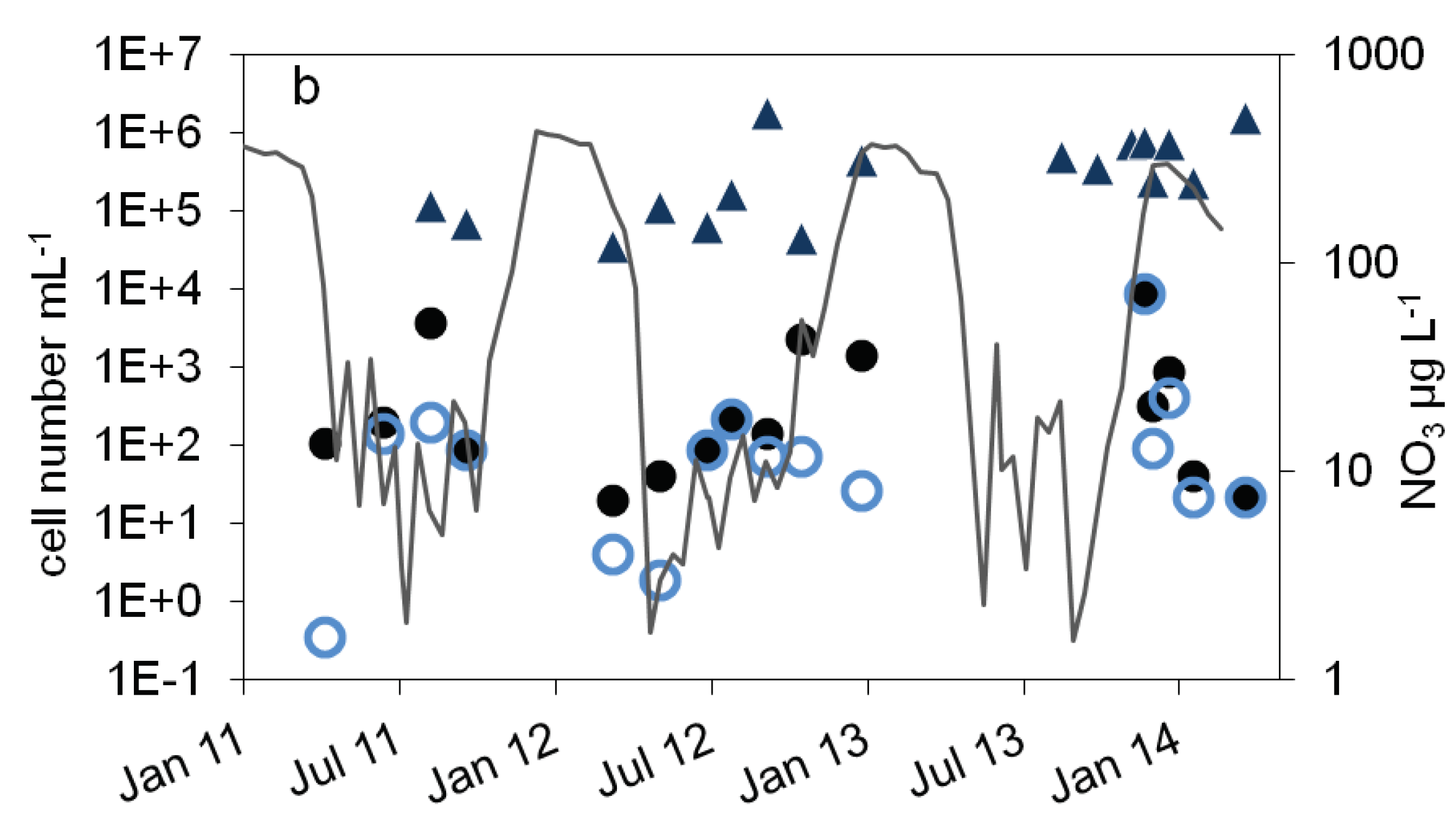
Figure 3.
Combined annual courses of cell numbers and nitrate concentrations in the epilimnion of lake Scharmützelsee: a: microscopical cell counts (),16S rDNA cell number estimation (), temperature 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line) b: numbers of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (), c: difference between log10 transformed cell numbers (nir)and MPN of denitrifying bacteria (+), nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line).
),16S rDNA cell number estimation (), temperature 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line) b: numbers of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (), c: difference between log10 transformed cell numbers (nir)and MPN of denitrifying bacteria (+), nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line).
Figure 3.
Combined annual courses of cell numbers and nitrate concentrations in the epilimnion of lake Scharmützelsee: a: microscopical cell counts (),16S rDNA cell number estimation (), temperature 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line) b: numbers of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (), c: difference between log10 transformed cell numbers (nir)and MPN of denitrifying bacteria (+), nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line).
),16S rDNA cell number estimation (), temperature 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line) b: numbers of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (), c: difference between log10 transformed cell numbers (nir)and MPN of denitrifying bacteria (+), nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line).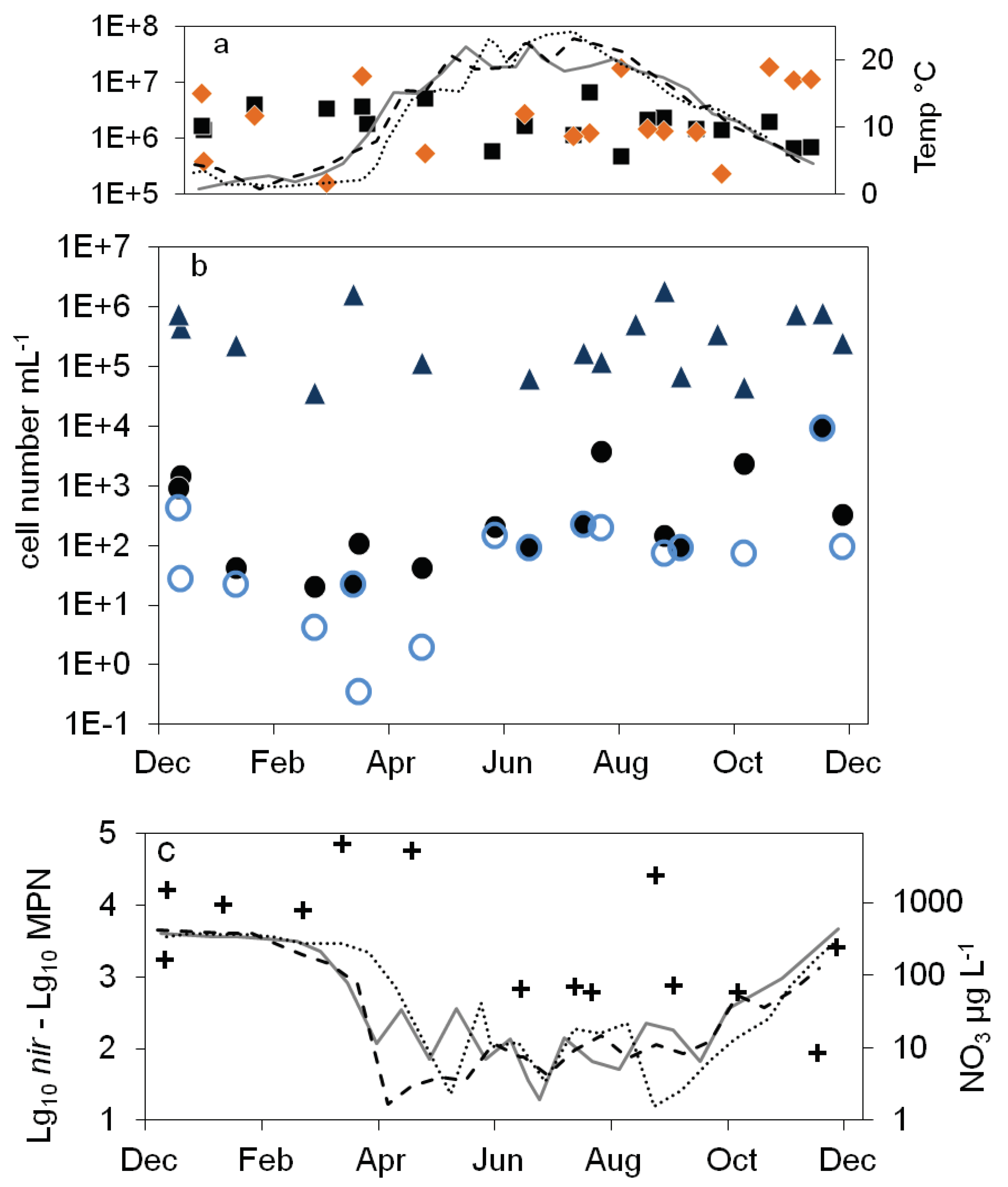
Figure 4.
Courses of nitrate concentration and cell numbers in the hypolimnion of lake Scharmützelsee from 2012 to 2014: a: microscopical cell counting (),16S rDNA cell number estimation () b: number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
),16S rDNA cell number estimation () b: number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
Figure 4.
Courses of nitrate concentration and cell numbers in the hypolimnion of lake Scharmützelsee from 2012 to 2014: a: microscopical cell counting (),16S rDNA cell number estimation () b: number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
),16S rDNA cell number estimation () b: number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).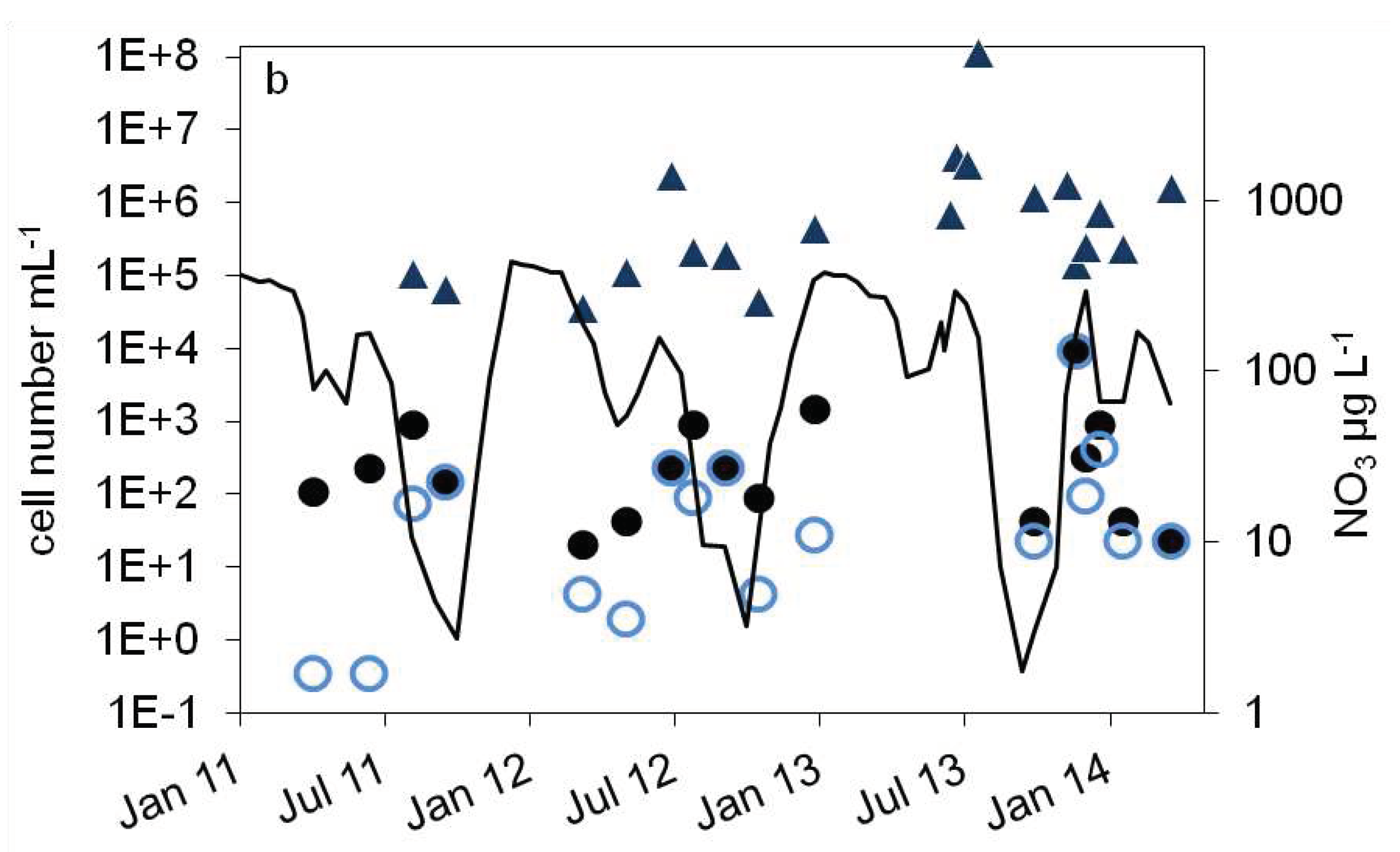
Figure 5.
Combined annual courses of cell numbers and nitrate concentrations in the hypolimnion of lake Scharmützelsee: a: 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (),nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line), b: difference between log10 transformed cell numbers (nir) and MPN of denitrifying bacteria (+).
), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (),nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line), b: difference between log10 transformed cell numbers (nir) and MPN of denitrifying bacteria (+).
Figure 5.
Combined annual courses of cell numbers and nitrate concentrations in the hypolimnion of lake Scharmützelsee: a: 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (),nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line), b: difference between log10 transformed cell numbers (nir) and MPN of denitrifying bacteria (+).
), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrate reducing bacteria () and denitrifying bacteria (),nitrate concentration 2011 (solid grey line), 2012 (dashed black line), 2013 (dotted black line), b: difference between log10 transformed cell numbers (nir) and MPN of denitrifying bacteria (+).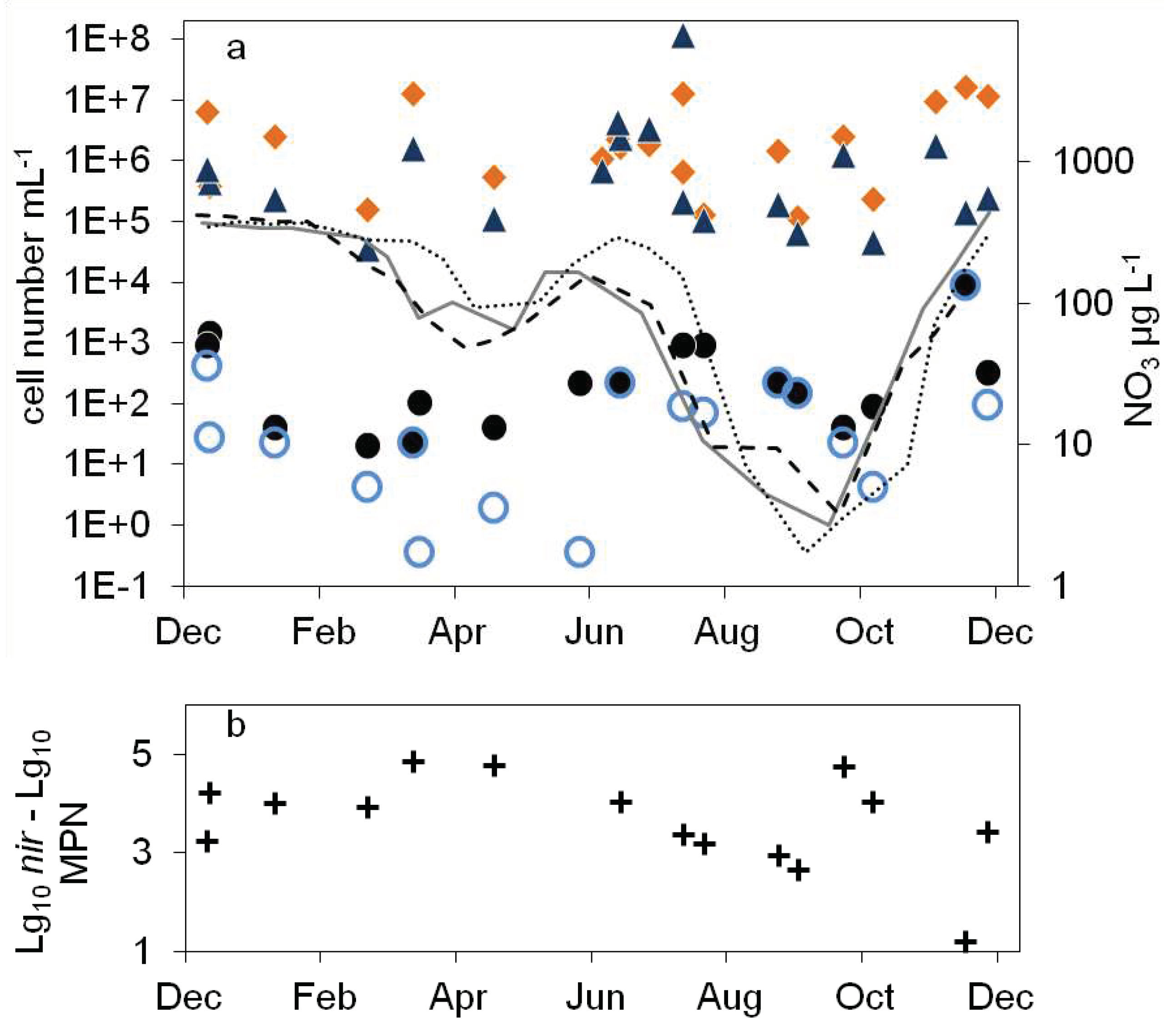
Figure 6.
Courses of nitrate concentration in the hypolimnion and cell numbers in the upper sediment layer of lake Scharmützelsee from 2012 to 2014: microscopical cell counting (), 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
), 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
Figure 6.
Courses of nitrate concentration in the hypolimnion and cell numbers in the upper sediment layer of lake Scharmützelsee from 2012 to 2014: microscopical cell counting (), 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).
), 16S rDNA cell number estimation (), number of denitrifying bacteria estimated by the sum of nirS and nirK gene copies (), MPN of nitrite reducing bacteria () and denitrifying bacteria (), nitrate concentration (black line).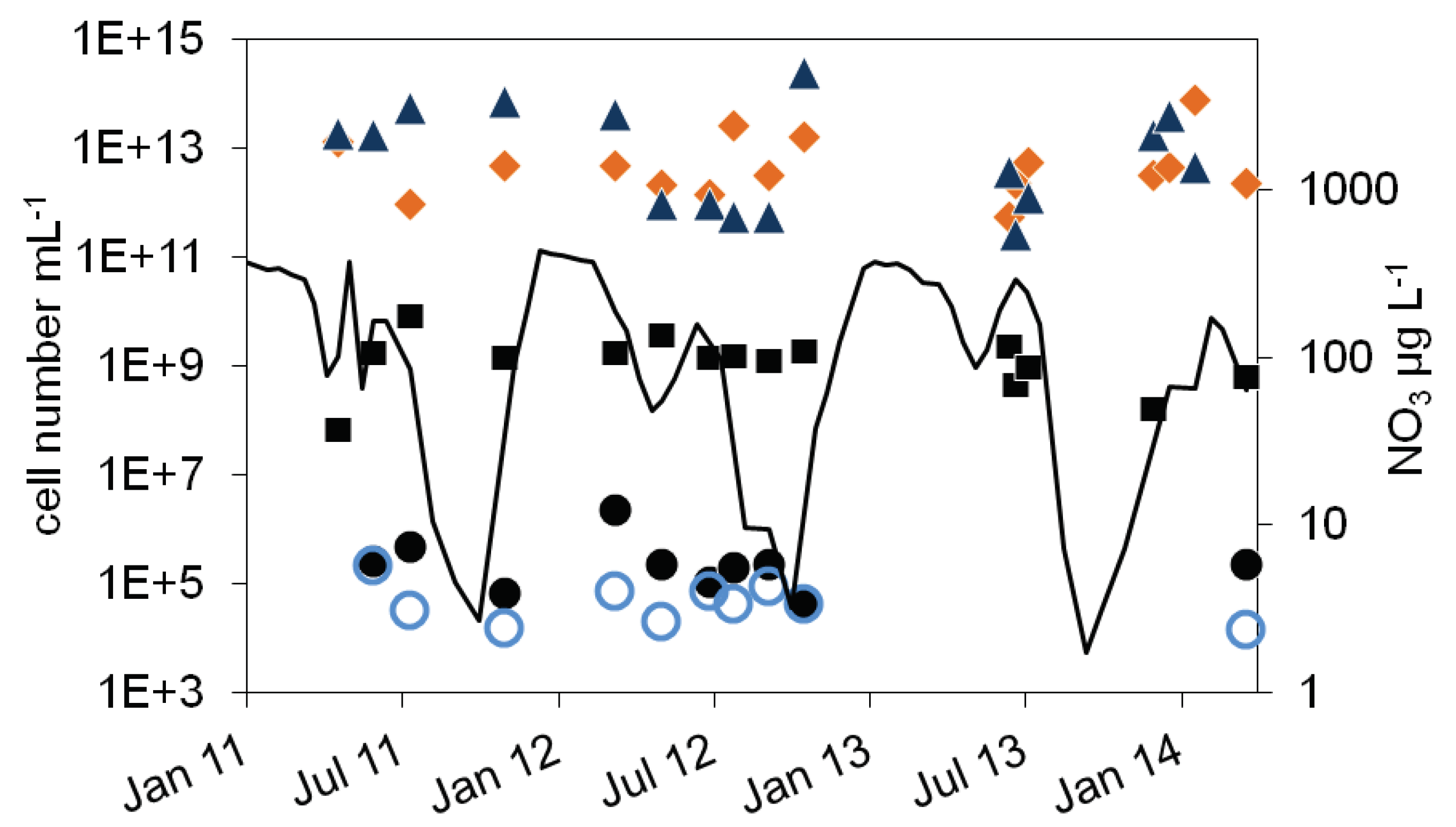
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



