
Submitted:
29 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Hydrocephalus is caused by an overproduction of cerebrospinal fluid (CSF), a blockage of fluid circulation, or improper reabsorption. CSF accumulation in the brain’s ventricles causes ventriculomegaly and brain cell damage. Hydrocephalus can be caused by brain trauma, hemorrhage, infection, tumors, or genetic mutations. Currently, there is no cure for hydrocephalus. Treatments like shunting and endoscopic third ventriculostomies are used, but unfortunately, these techniques require brain surgery and have high failure rates. To advance the development of hydrocephalus treatments, physiologically relevant pre-clinical models are crucial. This review covers some of the current animal and cell culture methods used to study hydrocephalus. The choroid plexus epithelium (CPe) is thought to be the major producer of CSF in the brain. It is a polarized epithelium that regulates ion and water movement from a fenestrated capillary exudate to the ventricles. Despite decades of research, control of electrolyte movement in the CPe is still not fully understood. This review discusses important transporters on the CPe and how some of these could be potential targets for hydrocephalus treatment.
Keywords:
choroid plexus
; cerebrospinal fluid
; electrolyte transport
; ventriculomegaly
; preclinical hydrocephalic models
1. Hydrocephalus
Hydrocephalus was first described in the 5th century by Hippocrates, and he was thought to have attempted treatment with ventricular punctures [1,2]. In the Middle Ages, the Arabic surgeon Abulcasis wrote a discourse on medicine that included the treatment of hydrocephalus. Advancements in the understanding of hydrocephalus occurred after Vesalius refined the knowledge of the anatomy and physiology of the brain in the mid-1500s. In the late 1600s, Thomas Willis first hypothesized that the choroid plexus was the source of cerebrospinal fluid (CSF). In the early 1800s, Magendie advanced understanding of the brain by describing the flow of CSF throughout the organ. Even with these advancements, hydrocephalus therapies remained crude. Some of the treatments included head wrapping, bloodletting, and purgatives. In 1881, Wernicke introduced ventricular puncture with external CSF drainage. Soon after, the first permanent shunt was invented in 1893, which ushered in a revolution for hydrocephalus therapy [2]. The invention of artificial values and silicone in the 1960s opened the door for modern shunt technology that is still used today [2].
Hydrocephalus is a disease caused by excessive production of CSF, a blockage of flow, or decreased reabsorption. Hydrocephalus can occur with or without a change in intracranial pressure. In cases where the pressure is low or normal, it is thought that there is an expansion of the skull or contraction of the cortical tissue to compensate [3]. Hydrocephalus can be categorized into two groups: communicating and non-communicating (Figure 1). When the CSF flow is unobstructed yet fails to absorb, or there is an overproduction of CSF, it is categorized as communicating hydrocephalus. Non-communicating hydrocephalus is characterized by a blockage of CSF flow due to various causes [4]. The increased CSF in the brain can cause headaches, vision loss, vomiting, nausea, and, if it is left untreated, it can be fatal [4]. Hydrocephalus is treatable, however, current treatments are suboptimal and a burden to the healthcare system. A cross-sectional study looking at healthcare data from 2019 in the United States determined that across all patient demographics, the cost of shunting was around $2.06 billion [5]. This does not include any other costs associated with hydrocephalus care [5].
Hydrocephalus is typically an acquired disease. Worldwide, the most common types of hydrocephalus are post-hemorrhagic and post-infectious hydrocephalus [6,7]. Pediatric post-hemorrhagic hydrocephalus occurs most commonly in preterm neonates; especially in those less than 1500 grams [8]. In adults, this condition is commonly associated with traumatic brain injury, hypertension, stroke, or aneurysm rupture [4,5,9]. Post-infectious hydrocephalus is more common in developing countries, specifically Africa, Latin America, and Southeast Asia [7]. Infants in developing countries typically acquire hydrocephalus after being infected by African meningitis, tuberculosis, or zika virus [6]. In developed countries, infections in neonates are typically associated with toxoplasma gondii or cytomegalovirus [10,11]. In adults, hydrocephalus can be caused by Neisseria meningitidis and Streptococcus pneumoniae [12].
There are also some genetic mutations that can lead to either communicating or non-communicating hydrocephalus. Most genetic mutations cause pediatric hydrocephalus. X-linked hydrocephalus makes up 5% of hydrocephalus cases at birth [13]. One form of X-linked hydrocephalus is caused by a mutation in L1 which affects the cell adhesion protein L1 cell adhesion molecule (L1CAM) [13]. Males with L1 syndrome have a phenotypic spectrum from mild to severe, but in all cases they are born with hydrocephalus, malformations in the corpus callosum, and intellectual disabilities [14]. Another genetic disorder, called syndromic craniosynostosis, is associated with hydrocephalus due to mutations in fibroblast growth factor receptor 2 (FGFR2) or fibroblast growth factor receptor 3 (FGFR3) that leads to primary cerebral maldevelopment, brain atrophy, and structural obstruction of CSF outflow [13]. Neurofibromatosis 1 and 2 are autosomal dominant diseases that cause tumors in the central and peripheral nervous system [13]. Tumors can lead to blockages in the ventricles and cause non-communicating hydrocephalus. Tuberous sclerosis is also an autosomal dominant disorder that causes subependymal giant cell astrocytomas that can lead to non-communicating hydrocephalus [13]. Walker-Warburg syndrome is an autosomal recessive disorder that can be associated with hydrocephalus. The disease is most commonly caused by mutations in protein O-mannosyl-transferase 1 (POMT1), but it can also be caused by defects in protein O-mannosyl-transferase 2 (POMT2), protein O-linked mannose N-acetylglucosaminyltransferase 1 (POMGNT1), acetylglucosaminyltransferase-like protein (LARGE), isoprenoid synthase domain containing (ISPD), fukutin (FKTN), or fukutin-related (FKRP) [13]. Mutations in coiled-coil domain containing 88C (CCDC88C) cause non-syndromic autosomal recessive hydrocephalus due to defective neural tube development [15]. These are the major syndromes that are known to cause hydrocephalus, but there are many more mutations that can cause the disease. Using a genome-wide association study (GWAS) on CSF and blood samples, the authors showed that there are almost 1,500 genes that could be associated with hydrocephalus [16]. This study, and studies like it, are laying the groundwork to discover more genetic causes of hydrocephalus.
A meta-analysis of hydrocephalus cases worldwide determined that per 100,000 individuals, there are 88 cases of hydrocephalus patients under the age of 18 [17]. A child may be diagnosed with the disease for various reasons like premature birth resulting in germinal matrix hemorrhage, infection, some medications taken during pregnancy, genetic syndromes, aqueductal stenosis, or tumors [18,19]. Globally, the most common cause of pediatric hydrocephalus is infection, while in the United States the most common cause is hemorrhaging of prematurity [19]. It is estimated that 383,000 children will be born with hydrocephalus per year, worldwide [7].
Hydrocephalus is the least common in middle-aged adults. In the above mentioned meta-analysis of worldwide cases of hydrocephalus, the authors found that there are 11 cases of hydrocephalus in adults from the age of 19 to 64 years of age for every 100,000 individuals [17]. The causes of hydrocephalus in adults have some overlap with the causes in children, like infection, trauma, and hemorrhage. Obese young women have a tendency to develop idiopathic intracranial hypertension [20]. Rapid weight gain is a factor in this disease, but the exact mechanism behind the development of intracranial hypertension is poorly understood.
The elderly are the most severely affected group. The worldwide meta-analysis found that in people over the age of 64, there are 175 cases per 100,000 people [17]. The most common type of hydrocephalus diagnosed in the elderly population is idiopathic normal pressure hydrocephalus (iNPH). In fact, one study estimated that 5.9% of individuals 80 or older have iNPH [21]. This disease can be easily overlooked because the symptoms of iNPH are similar to symptoms of aging in general, including urinary incontinence, gait disturbances, and cognitive dysfunction that can progress to dementia [21].
2. Hydrocephalus Treatments
Currently, the most common treatment for hydrocephalus is shunting (Table 1). In the simplest form, shunts consist of a ventricular catheter that is surgically implanted into the lateral ventricle of the brain, a valve that regulates the CSF flow out of the brain, and a catheter that empties the CSF into a body cavity. The most common body cavities where the CSF is shunted into are the peritoneal cavity, the pleura, and the atrium of the heart. The most common are put into the peritoneal cavity and are, therefore, called ventriculoperitoneal shunts (VPS) [22].
VPS are commonly used to treat many types of hydrocephalus, but the shunts have varying results. When used to treat iNPH, they can successfully relieve symptoms in 60-80% of patients [23]. However, in children shunts commonly fail with a frequency of 50% within the first two years [24]. Even with advances in shunt technology, the failure rate remains the same. A metanalysis of patients with shunts found that pediatric shunt failure rates did not improve in the thirty years from 1970 to 2000 [25]. Shunts can fail for multiple reasons like mechanical problems, catheter migration, or blockages caused by cell overgrowth [24,25]. Every shunt failure necessitates an operation which increases the chance of infection or death for the patient [24].
There have been some recent advances in shunt technology. A new FDA approved device, called the Reflow® System, flushes the VPS and prevents shunt obstructions [26]. Pediatric clinical trials with the Reflow® System have promising results. Two years before the Reflow® System was put into a group of 9 patients, they had previously experienced 14 shunt failures [26]. After the addition of the device, there was only one shunt failure during a follow up period ranging from 2.8-4 years [26]. Even though these results are promising, the Reflow® System is still going through clinical trials. Time will tell if it will be able to improve shunt failure rates in a wider group of patients.
Another type of shunt is the lumboperitoneal shunt (LPS). The shunt is placed into the lumbar region of the spinal cord to remove excess CSF to treat communicating hydrocephalus. LPS is beneficial because it is not necessary to have brain surgery and greatly decreases the chance of brain hemorrhage [27]. A meta-analysis performed in 2023 showed that LPS have a lower complication rate compared to VPS. Out of 3,654 patients there was a 12.98% complication rate with the LPS and 23.8% complication rate in the VPS patients [27]. Since their inception, LPS techniques are improving. A new two-stage LPS procedure was safer for the patient and still outperformed VPS. In this study, only one out of 92 LPS needed revision, while 26 out of 192 VPS surgeries needed revision and 11 surgeries had infections [28]. On the downside, LPS cannot be used as a treatment for non-communicating hydrocephalus.
Another option for treating hydrocephalus is an endoscopic third ventriculostomy (ETV) which does not require a shunt. This treatment involves brain surgery where a hole is created between the third ventricle and the subarachnoid space [29]. This treatment is used for both communicating and non-communicating hydrocephalus. In some respects, ETVs are considered to be safer and more effective than VPS. A meta-analysis of patients treated with either a shunt or ETV, showed that there was a higher complication rate for the VPS. There was only 1 cases of infection out of 128 ETV patients verses 26 cases of infection for the VPS group of 122 patients [30]. In the same study, it was also noted that 6 VPS patients died after surgery while no patients were lost after ETV treatment [30]. Unfortunately, pediatric patients do not respond well to ETVs. Kulkarni et al. created a predictive model of ETV success rates in children, and if the child is under one year old there is only a 30% ETV success rate [31]. Patients 10 years or older have higher success rates [31].
Hydrocephalus can also be treated by ablating the choroid plexus (CP) in the ventricles of the brain. This technique is not used as frequently in developed countries, but it is used often in less developed countries because it avoids the cost of shunt hardware and patient shunt dependency [32,33]. Additionally, there is a lack of pediatric neurosurgery training programs in developing countries [32]. Choroid plexus cauterization (CPC) is commonly paired with ETV. An international meta-analysis of ETV versus ETV/CPC was compared in pediatric patients from Africa and North America. Overall, the data showed no difference between the two procedures, however, when examined in the subpopulations in Sub-Saharan Africa, ETV/CPC was more effective [34].
ETV/CPC seems to be more effective than VPS in pediatric patients. In a study that compared ETV/CPC with VPS outcomes in children under 12 months of age, the Kaplan-Meier survival analysis showed that the estimated need for reoperation was lower in the ETV/CPC group [33]. The study estimated that 59% of the patients would avoid shunt dependency for a length of 11 years [33]. However, the long-term effects of ablating the CP, which does not regenerate, have not been studied.
There have been past attempts to use nonsurgical hydrocephalus treatments. Osmotic diuretics were one such treatment. Theobromine sodium salicylate, also known as Diuretin, looked like a promising treatment for pediatric hydrocephalus. A clinical study from 1924 showed that Diuretin had good efficacy in 4 pediatric patients [35]. Unfortunately, no further clinical studies were performed and Diuretin was abandoned. Oral isosorbide, which is another osmotic diuretic, was also attempted as a treatment for hydrocephalus. It was found to control CSF levels and intracranial pressure temporarily, but it caused intolerable dehydration and hypernatremia [36,37], and was discontinued. Diamox and Lasix, two other diuretics, were also abandoned after a systematic review proved both drugs to be ineffective for the treatment of post-hemorrhagic hydrocephalus [38]. Oral cardiac glycosides, which inhibit Na+-K+-ATPase, were investigated for a time. They were tested in clinical trials but had unfavorable results and were not investigated further [39].
There has been research into anti-inflammatory treatments that may have potential for treatment of hydrocephalus. Minocycline is more commonly known for its anti-microbial properties, but this FDA-approved antibiotic has been used to treat symptoms of brain trauma. When it is given in high doses, minocycline has anti-inflammatory, anti-apoptotic, and anti-oxidant properties [40]. It is also able to reduce ferritin [41]. In pre-clinical trials with rats, minocycline inhibited immunoreactive astrocytes and microglia while also thickening the cerebral cortex [42]. The drug had no effect on healthy animals [42]. Clinical trials with minocycline as a treatment for traumatic brain injury have had mixed results. Many of them have failed to replicate the results from the pre-clinical trials, and that could be due to the inability to treat patients within a critical window after injury [40]. Although there are no clinical trials using minocycline to treat hydrocephalus directly, traumatic brain injury can lead to hydrocephalus.
Another anti-inflammatory treatment that is currently being investigated is erythropoietin and melatonin (EPO + MLT). EPO alone has shown to help improve cognitive function in cases of brain injury in rats [43]. In pre-clinical trials, EPO + MLT were able to reduce ventriculomegaly in induced hemorrhage neonatal rats [44]. In another study, EPO + MLT was used to restore gait in adult rats that previously had induced prenatal injury [45]. The results from these studies are very promising and are moving into clinical trials for preterm infants with severe intraventricular hemorrhage [46].
There are currently some drugs available to help patients with hydrocephalus. However, these treatments are not long-term solutions. For example, mannitol, urea, oral glycerol, or acetazolamide can be used as a short-term treatment to decrease cranial pressure before or after shunt surgeries [47,48]. Additionally, mannitol is used in adults to decrease intracranial pressure caused by intracerebral hemorrhage [48,49]. Steroids, notably glucocorticoids, have also been used to relieve disorders with increased intercranial pressure, but it only provides temporary relief for hydrocephalus [48].
3. Animal Models
There is a clear need for better nonsurgical treatments for hydrocephalus. Testing potential pharmaceutical interventions requires well characterized preclinical models. Animal models have been used to study multiple forms of hydrocephalus (Table 2).
Post-traumatic hydrocephalus caused by traumatic brain injury (TBI) can be modeled in animals. Zhao et al. used a fluid-percussion device to induce TBI in adult rats [50]. After the initial injury, they injected the animals with FeCl3. This modeled intracranial hemorrhage which is common in patients with TBI. The rats developed acute hydrocephalus within a 24 hour period [50].
Post-hemorrhagic models of hydrocephalus have also been developed. Some commonly used animal models include rodents, pigs, and dogs [8]. To induce hydrocephalus, blood is injected into the ventricles [8]. In other experiments, blood components, like red blood cells, iron, or hemoglobin have also been effective [8]. This technique is not guaranteed to produce hydrocephalus every time. For example, Pang et al. attempted to make 10 post-hemorrhagic models of hydrocephalus in dogs, but only 8 of those caused ventriculomegaly after 3 months [51]. These models can be done on young animals to model germinal matrix hemorrhages that occur with premature birth. Robinson et al. injected rat pups with lysed red blood cells which caused progressive macrocephaly after the animals were primed with the inflammatory mediator lipopolysaccharide [44]. This model is an efficient way to model post-hemorrhagic hydrocephalus, however, it does have some draw backs. The methods, including the nature of the “priming” material such as pre-treatment with inflammatory mediators and the level of consistency in the degree of hydrocephalus make it difficult to standardize techniques and expected results across laboratories.
Hydrocephalus can be induced chemically, and one popular chemical used to do so is kaolin. It has been used in experiments since the 1950s [52,53]. One benefit of using kaolin is that it is inexpensive and relatively reliable in inducing hydrocephalus. In a recent experiment with juvenile pigs, the animals were injected with kaolin into the intracisternal region which produced bilateral ventriculomegaly [54]. While the severity of the hydrocephalus varied among the animals, it proved to be a reliable model that was used to test shunting techniques [54]. Kaolin has been successfully used in rodents, dogs, cats, lambs, and primates [55,56,57,58,59]. One drawback of using kaolin to induce hydrocephalus is that it may kill some animals immediately after injection [55]. Depending on the animal model being used, it can take some time to determine what is the optimal, non-lethal concentration of the drug. Another downside of chemically induced hydrocephalus is that it does not model any naturally occurring form of hydrocephalus. Non-specific, caustic chemicals will not just ablate the choroid plexus epithelium (CPe), but also damage the ventricle lining.
6-aminonicotinamide (6-AN) is another chemically induced model that has been used in rats. 6-aminonicotinamide can be injected into the brain to induce symptoms similar to those described in Dandy-Walker syndrome [60]. 6-AN can also be injected into a pregnant animal to induce hydrocephalus in the fetuses [61]. Dandy-Walker syndrome causes malformations in the brain, particularly the cerebellum [62]. Almost 80% of patients with Dandy-Walker syndrome develop hydrocephalus [62]. 6-aminonicotinamide treatment mimics this syndrome by inducing cerebellum hypoplasia and other brain malformations [61]. When the rats are treated with 6-aminonicotinamide, ventriculomegaly is visible after 72 hours [63]. This is a useful model to replicate human disease, however this is another chemically induced method that will cause non-specific damage to the brain.
4. Genetic Animal Models
The Wpk rat (Tmem67-/-) rat, models pediatric communicating hydrocephalus [64,65]. The rats have a single nucleotide mutation in the Tmem67 protein which is essential for the development of primary cilia [64]. This models the human disease Meckel-Gruber syndrome type 3 [64]. The homozygous phenotype is characterized by severe hydrocephalus and polycystic kidney disease [64,65]. Using this genetic model has some limitations because the animals do not live very long. The homozygous phenotype is so severe that the animals typically die 18-21 days after birth [65]. These animals have been used successfully to test the efficacy of transient receptor potential vanilloid-type 4 (TRPV4) antagonists [66] and serum glucocorticoid-induced kinase 1 (SGK1) inhibitors [67].
The hydrocephalus Texas rat (H-Tx) is a good model for studying non-communicating hydrocephalus. The rats have defective cortical development and have aqueductal stenosis with detectable hydrocephalus around day 18 [68]. The lateral ventricles of the brain are enlarged, and in some animals the third ventricle is also enlarged [69]. It was noted that the aqueducts of the brain were abnormal due to a thickening of the midbrain [69].
LEW/Jms rats have a mutation that leads to the development of hydrocephalus immediately after birth [70]. It was noticed that more male than female rats developed symptoms, so the genetic cause may be sex-linked [70]. This model could be used to study human diseases like X-linked hydrocephalus. The L1CAM mouse is very similar to the LEW/Jms model. These mice lack the L1 gene which encodes L1CAM. This protein is a cell adhesion molecule that is important for axon growth, neuronal migration, and synaptic plasticity [71]. Mice without L1 have improper brain development and the disease disproportionately effects males [71]. This model can be used to study X-linked hydrocephalus and L1 syndrome.
The E2F5 mouse line is a homozygous knockout of the E2F5 transcription factor which is important for the choroid plexus maturation and cilia function [72]. The knockout line has a cilia malfunction that leads to the hydrocephalus phenotype which develops gradually [73]. The affected animals have normal size and appearance compared to their wildtype counterparts until they are 3-4 weeks of age [74]. This mutation is lethal, the mutant mice live to an average of 6 weeks [74].
Growth arrest specific 8 (Gas8) is another genetic mouse model used for hydrocephalus. Gas8 is important for hedgehog signaling and regulating cilia movement [75]. This gene also has some clinical significance. There are at least two human patients with missense mutations in Gas8 that lead to hydrocephalus: A391V and E199K [75]. Preliminary results indicate that, dependent on background, Gas8 mice develop hydrocephalus between postnatal day 12-16, making it a good model for pediatric hydrocephalus [76].
The coiled-coil domain containing 39 (CCDC39) mouse model causes neonatal hydrocephalus with abnormal motile cilia development [77]. The mice have short, thin cilia that cannot beat properly because of the loss of CCDC39. Interestingly, they also have situs inversus. They have normal brain development at birth but then progressively develop hydrocephalus until it becomes fatal at 3 weeks of age. This model mimics the timing of human congenital hydrocephalus [77].
Autosomal recessive mutation hydrocephalus-3 (Hy-3) mouse is a genetic mouse model that produces communicating hydrocephalus [78]. The mice have a mutation in the hydin protein which is a homolog to the human protein caldesmon. This protein is important for the cellular cytoskeleton. The mice develop hydrocephalus shortly after they are born. Is it thought that they develop communicating hydrocephalus due to loss of cilia [78].
The hydrocephalus with hop gait mouse (Hyh) model has a lethal recessive mutation on chromosome 7 that leads to hydrocephalus [79]. Homozygous mice have the characteristic domed heads of hydrocephalus and develop a unique hopping gait approximately two weeks after birth. The mice die between 4-10 weeks of age [79]. This model can be used for genetic hydrocephalus, as well as non-communicating hydrocephalus because there is a blockage of CSF movement in the fourth ventricle [79]. A similar model known as the hydrocephalus associated with polydactyly mice (Hpy) has recessive mutation that causes hydrocephalus. The mice have hopping gait, sterility in male mice, and scoliosis [80]. The mice develop communicating hydrocephalus around day 6 after birth and most die around day 14 [80]. However, the specific cause of hydrocephalus is unknown.
Mf1 mice model congenital hydrocephalus. Mesoderm/mesenchyme forkhead 1(Mf1) is a gene that encodes winged helix proteins that are DNA-binding proteins [81]. Mice with Mf1 knockout mice die perinatally from hemorrhagic hydrocephalus. They also have issues in other parts of the body where Mf1 is expressed. There are eye abnormalities and skeletal defects [81]. Similar to the Mf1 mouse, the hemorrhagic hydrocephalus mice (Hhy) develop hydrocephalus due to a recessive genetic mutation [60]. The symptoms of hydrocephalus occur between 3.5 – 19 weeks of age, with the majority of mice developing hydrocephalus by 15 weeks of age [82]. These models are useful for hemorrhagic hydrocephalus because there is no need to induce an injury or inject blood components into the animals to get the desired results.
Rho family GTPase 3 (Rnd3) is important for regulating cell migration and proliferation. When this protein is absent, in mice, the cerebral aqueduct is blocked resulting in non-communicating hydrocephalus [83]. Mice with this mutation are described as having extreme dilation on the third ventricles but not the fourth, thick ependymal cell layers, and upregulated Notch signaling [83].
The Msx1 mouse models have malformations of the brain that result in the collapse of the cerebral aqueduct [60]. The symptoms of hydrocephalus start to develop before birth due to the deletion of msh homeobox 1 (Msx1). The function of the Msx1 protein is not fully understood, but it is thought to be involved with the development of the fasciculus retroflexus [84]. The mice are characterized with brain abnormalities such as reduced corpus callosum, cerebral cortex, caudate putamen, and septum. Some of the homozygous mutant mice have ventriculomegaly in the third ventricles. The main cause of hydrocephalus in this animal model is thought to be ependymal detachment in the developing embryos [84].
The SUMS/NP mouse is a genetic model of congenital hydrocephalus that starts to develop 14 days after gestation [85]. The hydrocephalic phenotype is not noticeable until 4-5 days after birth, when there is a significant increase in the lateral ventricles. These mice also have absent or reduced cerebral aqueducts. This mouse model does not live very long, they typically die soon after they are weened [85].
With the ease of use of CRISPR, genetic manipulation of the CPe is possible. There are also many genetic knockout lines that can be created to study hydrocephalus in vivo and a recent review by Jang and Lehtinen does an excellent job of covering many of them [86]. Viral vectors are another method to genetically manipulate the CPe. Viral vectors can change gene expression within 24-48 hours without the need for breeding [86]. They can be used in many stages of life in animal, including in utero [86].
Animal models are useful for understanding the physiology of hydrocephalus and testing potential drug treatments, but they do have some drawbacks. Animals models do not perfectly model the human brain. The size of the ventricles vary across species, and some animal CPe cells may have different ion channels compared to humans. For example, it has been reported that mice CPe have the sodium transporter known as the epithelial Na+ channel (ENaC) while humans cells do not [87]. The animal models may not represent the symptoms of hydrocephalus perfectly and they may not respond to drug treatments in the same way as humans. Some large animal models are very costly as well. Additionally, when trying to observe molecular mechanisms, in vivo models can have other confounding physiological factors that can muddy observations. Using cell culture models in conjunction with animal models can help reduce confounding factors and provide depth to the experimental reliability.
5. The Choroid Plexus Epithelium
One major target in the search to find treatment options for hydrocephalus has been the functionality of the choroid plexus. The CPe is primarily responsible for the production of CSF. The CPe is a monolayer of epithelial cells that resides in all four ventricles of the brain [88] and receives nutrients and substrates from a fenestrated capillary network it surrounds. The epithelial cells contain tight junctions, desmosomes, and adherent junctions, thus forming the CSF-blood barrier [88,89]. The cells are cuboidal in shape and are ciliated [88,89]. Interestingly, the CPe is polarized differently than most epithelial cells. Typically, ion transporters like Na+-K+-ATPase, sodium potassium chloride co-transporter 1 (NKCC1) and Na/H exchanger 1 (NHE1) are located in the basolateral membrane [88,90]. On the CPe, these are located apically facing the CSF [88,90]. There are also some transporters found on the basolateral side of the CPe that are normally found on the apical side. These include anion exchanger 2 (AE2), K+-Cl- cotransporter (KCC3), and Na+-HCO3- cotransporter 1 (NBCn1) [88].
It is estimated that the adult human brain contains about 150 mL of CSF, with 125 mL located in the cranial and spinal subarachnoid spaces and the remaining 25 mL in the ventricles [91]. The CPe is thought to secrete around 80% of the total CSF while the remaining 20% is produced by leakage across the blood-brain barrier [88]. A healthy human adult produces between 400-600 mL of CSF per day [91]. The CPe produces CSF by controlling electrolyte and water flow which has been reported to be independent of the trans-epithelial osmotic gradient [92]. Control of electrolyte and water movement in the CPe is still not fully understood.
Along with producing CSF, the CPe has many other functions. One function is removing waste products from the CSF. Some of the waste products include β-amyloid, phosphatidylinositol, and uric acid [90]. The CPe is also important for immune function. It is a bridge where immune cells can cross into the CSF from the blood [90]. Additionally, it acts as a physical barrier so that some pathogens cannot enter the CSF [90]. The CPe is also an important entry point for drug delivery to the brain [90].
6. Cell Culture Models
Primary cultures derived from the CPe have been obtained from many species of animals including amphibians [93], mice [94], rats [95], rabbits [96], pigs [97], sheep [98], and cows [99]. The downside to primary cell culture is that it can difficult to obtain pure cultures and the cultures have short life spans [100]. The CPe needs to be removed soon after the animal is sacrificed and the cells need to be purified. Additionally, there is usually low cell abundance so many animals are required for cell culture [100].
CPe cell cultures can also be obtained from biotech companies. Human choroid plexus epithelial cells (HCPEpiC) can be bought from different companies and have been referenced in publications. For example, this cell line has been used to study the inflammatory response [101] and the effects of chemotherapy on CPe cells [102].
As an alternative to primary cultures, there are some continuous CPe cell lines (Figure 2). The porcine choroid plexus-Riems (PCP-R) cell line has been used to model the CPe [103]. It has been used to identify ion channels on the CPe using Ussing-style electrophysiology and to study changes in barrier function. Recent studies have, however, indicated that the PCP-R is mispolarized meaning that the ion channels are on the opposite side of the cell when compared to the native epithelium [104]. Therefore, this cell line is not ideal for studying CSF production. However, this cell line is still useful for other experiments. For example, the PCP-R has been used to test what kinds of xenobiotic drugs can cross the blood-CSF barrier [97]. Similarly, it has been used to study what immune cells can cross the blood-CSF barrier [105]. The cells also contain tight junctions like Claudin-1 and Claudin-2 [103]. This cell line also has high transepithelial electrical resistance (TEER) which is typical of mammalian choroid plexus cells [103,106]. On average the TEER was approximately 1000 Ω cm2 when the resistance was measured with Ussing-style electrophysiology [104].
The Z130 is an immortalized rat CPe cell line that was created through gene transfection [100]. When compared to primary rat cell cultures, the morphology was the same and it was able to mimic the blood-cerebrospinal fluid barrier [100]. The polarity of the cells are the same as primary rat cultures [100,107]. Z130 cells have tight junctions but can only maintain a maximum TEER value around 100 Ω cm2 [108]. Z130 cells have been used to study how heavy metal cross the CPe along with the toxic effects of these metals [109].
The human choroid plexus papilloma (HIBCPP) is another cell line that can be used to model the CPe. This immortalized cell line was derived from a malignant papilloma in the choroid plexus of a 29 year old Japanese woman [110]. It has been characterized to have structures similar to the CPe-like villus structures located apically [106]. Unlike the PCP-R cells, the important transport proteins are correctly polarized with reference to the native epithelium. Notably, TRPV4 and NKCC1 are located apically [106]. While the basolateral side of the cell contains sodium-driven chloride bicarbonate exchanger (NCBE) and AE2 [106]. Both sides of the cell have aquaporin channels [106]. This cell line has a moderately high resistance when it is grown in a monolayer with TEER measurements over 400 Ω cm2 [106]. The downside to this cell line is that it is relatively difficult to maintain in culture.
Another interesting model is the co-culture of immortalized human CP endothelial cell (iHCPEnC) line with HIBCPP cells. The cell types are cultivated on opposite sides of the same permeable support to produce a combined CP model [111]. The addition of the endothelial cells increases the tightness of the cells and more closely mimics the conditions found in the brain [111]. Using a tracer voltohmmeter technology the authors measured the TEER of the combined cell culture which reached a max of 700 Ω cm2 [111].
CP organoids can be derived from stem cells. Interestingly, the organoids can produce CSF-like fluid. When the CSF-like fluid from the organoid was compared with CSF proteins from human patients, the same proteins were present [112]. Another group made a CP organoid that had a similar morphology and expressed the genes LIM homeobox transcription factor 1 alpha (LMX1A), orthodenticle homeobox 2 (OTX2), and aquaporin 1 (AQP1) that are also expressed in the CP [102]. There are some limitations for organoids. They are expensive and time-consuming. There are also different protocols that produce organoids of different quality and composition [102]. The inconsistency makes it difficult to compare studies from different research groups.
7. Electrolyte Transporters and Potential Roles in Fluid/Electrolyte Homeostasis
The main driving force for transepithelial ion flux is Na+ [93,113]. In one potential scenario, Na+ enters the CPe from the blood stream by the NBCE and Na+-HCO3- cotransporter 2 (NBCn2) channels. Subsequently Na+-K+-ATPase pumps Na+ into the CSF [88,90,93]. The Na+-K+-ATPase is also important for K+ homeostasis. It removes excess K+ from the CSF by pumping the ions into the cell [114]. It has been suggested that the potassium voltage-gated channel subfamily Q member 1 (KCNQ1) and potassium voltage-gated channel, shaker-related subfamily, member 3 (KCNA3) can join with potassium voltage-gated channel subfamily E regulatory subunit 2 (KCNE2) to form K+ channels on the basolateral side of cell to excrete K+ into the blood [115]. The K+, Cl- co-transporter KCC3 also moves K+ from the CSF into the bloodstream [116]. While these multiple K+ channels have been implicated, which are critical, regulated transporters remain unknown.
NKCC1 is a co-transporter that regulates the movement of Na+, K+, and two Cl- ions across the CPe apical membrane [88] (Figure 3). The directionality of the electrolyte transport as well as the ability of the protein to transport water is controversial. For a synopsis of the controversy surrounding the directionality of flux the reader is referred to a point-counter-point exchange in the Journal of Physiology [117,118,119,120]. More recently, data suggest that the direction of electrolyte movement may be heavily influenced by local ion concentrations [121] or developmental stage [122]. It was previously reported that around 600 molecules could be moved by NKCC1 [123], but recent studies suggest that the transporter only moves around 86 water molecules [124]. The NKCC1 is not only expressed on the CPe, but in many tissues where it is important for maintaining cell volume [125,126,127]. When NKCC1 is mutated or knocked out it causes severe disease symptoms [128]. However, NKCC1 knockout models do not have altered ventricle size, but they do have brain malformations and smaller CPe cells compared to their wildtype counterparts [128,129]. There are some rare cases where human patients are deficient in NKCC1 due to a mutation in solute carrier family 12 member 2 (SLc12a2). Since NKCC1 is found throughout the body, patients suffer from multiple serious problems including severe intellectual disability, hearing loss, respiratory weakness, and gastrointestinal issues [128,130].
There are many groups that have investigated NKCC1’s role in CSF production, but their conclusions are conflicting. For example, when NKCC1 is pharmacologically blocked during inflammatory conditions in rats, less CSF is produced by the CPe [131]. However, when NKCC1 was overexpressed in a neonatal post-hemorrhagic hydrocephalic mouse model, CSF production diminished and the brain ventricle size returned to normal [132]. Another study investigated the role of NKCC1 under normal conditions in a rat model and found that when NKCC1 was blocked, CSF production was not reduced [133]. One explanation for the apparently contradictory behavior of this channel is that it can adapt to maintain homeostasis. The ion channel seems to be able to move ions to either side of the membrane depending on the external/internal environment [121,127]. Additional studies will be required to determine the factors involved in the control and mechanism of NKCC1 in the CPe. One way to investigate the role of NKCC1 would be to test cultured CPe cells for fluid secretion in different conditions. A recently characterized human cell line, HIBCPP, provides a unique epithelium in which fluid secretion can be easily determined [67]. Culturing the CPe in vitro removes any confounding factors and can help determine the role of NKCC1 in CSF production.
Pharmacologically blocking NKCC1 has been investigated in pre-clinical models. However, before this can be translated into the clinical sphere, better NKCC1 antagonists need to be developed. Bumetanide is a common clinical inhibitor but this drug has very low brain penetration [134]. Therefore, it needs to be infused directly into the ventricles for efficacy in decreasing CSF production [131]. Additionally, this drug effects both NKCC1 and NKCC2 and therefore has substantial renal effects, causing excessive diuresis [134]. Some other symptoms include low potassium and chloride in the blood, metabolic alkalosis, and elevated levels of uric acid in the blood. There has been some work to create better NKCC1 inhibitors, and a recent review written by Savardi et al. covers a few of them [134].
TRPV4 is an osmo-, shear-, temperature, and pressure sensitive channel [135,136,137,138,139,140] that is found on the apical membrane of the CPe (Figure 3). Activation results in Ca2+ and Na+ influx into the CPe and, secondarily, causes a substantial change in transepithelial permeability and electrolyte flux that appears to involve multiple transporters [106]. This channel may play a role in CSF production in pathological states. In an experiment with Tmem67-/- rats, a genetic form of hydrocephalus was reversed with treatment with the TRPV4 antagonists HC067047 and RN 1734 [66]. Blocking TRPV4 with an antagonist seems to be a promising form of treatment for communicating hydrocephalus. TRPV4 knockout mice have no obvious physiological or behavioral phenotypes under normal conditions [140,141]. Therefore, using a TRPV4 antagonist would likely be safe for patients. In fact, oral TRPV4 antagonists were tested in clinical trials as a treatment for heart failure [142]. The drug, GSK2798745, was well tolerated by healthy and ill patients alike [142,143].
There may be a relationship between TRPV4 and NKCC1. When TRPV4 transports Ca2+ into the cell, with no lysine kinase (WNK) is stimulated. A study in human salivary glands found that WNK can be activated when there is an increase in cellular levels of Ca2+ [144]. WNK then phosphorylates the SPS1-related proline/alanine-rich kinase (SPAK) which secondarily activates NKCC1 [114,145].
KCC3 on the basolateral membrane of the CPe (Figure 3) is associated with inherited forms of hydrocephalus. When the channel is mutated, it leads to decreased K+ transport in the brain [88,146]. KCC3 dysfunction is also associated with Mendelian disease agenesis of the corpus callosum with peripheral neuropathy which causes malformation of the corpus callosum [147]. NKCC1 is related to KCC3; both ion channels are regulated by the WNK/SPAK pathway [148]. KCC3 is inhibited by WNK/SPAK activation. When it is phosphorylated, it moves Na+ and Cl- ions out of the cell [148,149,150].
The cystic fibrosis transmembrane conductance regulator (CFTR) is another channel whose role in CPe is controversial. CFTR secretes Cl- ions from epithelial cells [151,152]. It has been reported that there is no CFTR mRNA in the CPe of rats [87,88]. However, unpublished data from our laboratory indicates the presence of CFTR in both human and porcine epithelial cells at the mRNA level and an effect of CFTR inhibitors on transepithelial ion flux. In other tissues of the body, like the lungs, TRPV4 has been shown to be important for activating CFTR [153].
Additionally, the CPe is important for pH regulation. There are many acid-base transporter proteins like the NCBE (Na+-driven chloride bicarbonate exchanger), AE2, and NCBE2 (Na+-driven bicarbonate cotransporter) [88] Other important transporters for pH homeostasis include acid-base transporters Cl−−/H+, sodium/hydrogen exchanger 6 (NHE6), and chloride-proton exchanger 7 (CLC-7) [89].
NCBE on the basolateral membrane of the CPe (Figure 3) may be an important transporter for understanding the pathophysiology of hydrocephalus. The transporter is likely regulated by inflammation in the brain. When mouse CPe cells were treated with inflammatory cytokines TNFα and IL-1β, NCBE expression decreased [154]. Inhibiting carbonic anhydrase with acetazolamide prevents CSF secretion, pointing to the importance of HCO3- in CSF production [155]. There may be interplay between NCBE and multiple transporters that impact CSF secretion. When the gene responsible for NCBE, solute carrier family 4 member 10 (Slc4a10), is knocked out in mouse models, aquaporin 1 and Na+-K+-ATPase expression is drastically decreased [156]. Additionally, the WNK/SPAK pathway regulates the functional activity of NCBE1 [157]. It was shown in pancreatic epithelial cells that SPAK inhibits the expression of NCBE1-B [158]. As previously discussed, the WNK/SPAK pathway regulates ion channels like TRPV4 and NKCC1 that may be important for the regulation of CSF. It has also been noted that mice with Slc4a10 mutations have small ventricles [156,159]. Humans with de novo Slc4a10 mutations have small ventricles as well as cognitive impairments and epilepsy [160,161].
Even though the CP is a small tissue, it is essential for the production of CSF, but it is also important for removing waste, pH regulation, drug delivery to the brain, and immune function. CPe cells are uniquely polarized so they can produce large amounts of CSF to maintain homeostasis. The CPe is able to complete this task by precisely controlling the flow of electrolytes and water with the help of a complex system of ion channels and intracellular regulators. Some of these ion channels, specifically TRPV4, NKCC1, NCBE, and KCC3 seem to be involved in CSF production and ventricle size. These channels as well as the biochemical pathways that regulate their activity are prime targets to explore in the search for potential pharmaceutical interventions to treat hydrocephalus.
8. Conclusion
Hydrocephalus is a complex disease, with many origins. There have been myriad attempts to find better treatments for hydrocephalus, but currently invasive shunt surgeries or ETVs remain the standard of care. To facilitate the development of pharmacological interventions that can be used on an as-needed basis, it is key to increase our understanding of how CSF production and composition are regulated. An important component of this research is to develop and characterize animal and cell culture models that are widely used within the research community. If there is agreement as to standardized models, replication of results may facilitate the resolution of existing controversies. Both animal and cell culture models are necessary so that these can be used in an iterative fashion particularly in the realm of drug testing.
Author Contributions
Original draft preparation including figures, V.N.; review and editing, B.BY. and V.N.; funding acquisition, B.BY. Both authors have read and agreed to the published version of the manuscript.
Funding
The research in the authors’ laboratory is funded by grants from the United States Department of Defense office of the Congressionally Directed Medical Research Programs (CDMRP): Expansion Award, PR221811 (BBY) and Focused Program Award, PR221805 (BBY).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lifshutz, J.I.; Johnson, W.D. History of hydrocephalus and its treatments. Neurosurgical Focus 2001, 11, 1–5. [Google Scholar] [CrossRef]
- Aschoff, A.; Kremer, P.; Hashemi, B.; Kunze, S. The scientific history of hydrocephalus and its treatment. Neurosurgical Review 1999, 22, 67–93. [Google Scholar] [CrossRef]
- Mokri, B. The Monro–Kellie hypothesis: applications in CSF volume depletion. Neurology 2001, 56, 1746–1748. [Google Scholar] [CrossRef] [PubMed]
- Hochstetler, A.; Raskin, J.; Blazer-Yost, B.L. Hydrocephalus: historical analysis and considerations for treatment. European Journal of Medical Research 2022, 27, 1–17. [Google Scholar] [CrossRef]
- Koschnitzky, J.E.; Yap, E.; Zhang, Y.; Chau, M.J.; Yerneni, K.; Somera, A.L.; Luciano, M.; Moghekar, A. Inpatient healthcare burden and variables influencing hydrocephalus-related admissions across the lifespan. Journal of Neurosurgery 2022, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karimy, J.K.; Reeves, B.C.; Damisah, E.; Duy, P.Q.; Antwi, P.; David, W.; Wang, K.; Schiff, S.J.; Limbrick Jr, D.D.; Alper, S.L. Inflammation in acquired hydrocephalus: pathogenic mechanisms and therapeutic targets. Nature Reviews Neurology 2020, 16, 285–296. [Google Scholar] [CrossRef]
- Dewan, M.C.; Rattani, A.; Mekary, R.; Glancz, L.J.; Yunusa, I.; Baticulon, R.E.; Fieggen, G.; Wellons, J.C.; Park, K.B.; Warf, B.C. Global hydrocephalus epidemiology and incidence: systematic review and meta-analysis. Journal of Neurosurgery 2018, 130, 1065–1079. [Google Scholar] [CrossRef]
- Chen, Q.; Feng, Z.; Tan, Q.; Guo, J.; Tang, J.; Tan, L.; Feng, H.; Chen, Z. Post-hemorrhagic hydrocephalus: recent advances and new therapeutic insights. Journal of the Neurological Sciences 2017, 375, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Cioca, A.; Gheban, D.; Perju-Dumbrava, D.; Chiroban, O.; Mera, M. Sudden death from ruptured choroid plexus arteriovenous malformation. The American Journal of Forensic Medicine and Pathology 2014, 35, 100–102. [Google Scholar] [CrossRef]
- Karimy, J.K.; Duran, D.; Hu, J.K.; Gavankar, C.; Gaillard, J.R.; Bayri, Y.; Rice, H.; DiLuna, M.L.; Gerzanich, V.; Simard, J.M. Cerebrospinal fluid hypersecretion in pediatric hydrocephalus. Neurosurgical Focus 2016, 41, E10. [Google Scholar] [CrossRef]
- Bale, J.F. Fetal infections and brain development. Clinics in Perinatology 2009, 36, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Thigpen, M.C.; Whitney, C.G.; Messonnier, N.E.; Zell, E.R.; Lynfield, R.; Hadler, J.L.; Harrison, L.H.; Farley, M.M.; Reingold, A.; Bennett, N.M. Bacterial meningitis in the United States, 1998–2007. New England Journal of Medicine 2011, 364, 2016–2025. [Google Scholar] [CrossRef] [PubMed]
- Varagur, K.; Sanka, S.A.; Strahle, J.M. Syndromic hydrocephalus. Neurosurgery Clinics 2022, 33, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Stumpel, C.; Vos, Y.J. L1 Syndrome; University of Washington: Seattle, Seattle (WA), 1993. [Google Scholar]
- Marguet, F.; Vezain, M.; Marcorelles, P.; Audebert-Bellanger, S.; Cassinari, K.; Drouot, N.; Chambon, P.; Gonzalez, B.J.; Horowitz, A.; Laquerriere, A. Neuropathological hallmarks of fetal hydrocephalus linked to CCDC88C pathogenic variants. Acta Neuropathologica Communications 2021, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hale, A.T.; Bastarache, L.; Morales, D.M.; Wellons, J.C.; Limbrick, D.D.; Gamazon, E.R. Multi-omic analysis elucidates the genetic basis of hydrocephalus. Cell Reports 2021, 35, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, A.M.; Riva-Cambrin, J.; Yavin, D.; Hockley, A.; Pringsheim, T.M.; Jette, N.; Lethebe, B.C.; Lowerison, M.; Dronyk, J.; Hamilton, M.G. Age-specific global epidemiology of hydrocephalus: Systematic review, metanalysis and global birth surveillance. PloS One 2018, 13, e0204926. [Google Scholar] [CrossRef]
- Tully, H.M.; Dobyns, W.B. Infantile hydrocephalus: a review of epidemiology, classification and causes. European Journal of Medical Genetics 2014, 57, 359–368. [Google Scholar] [CrossRef]
- Wright, Z.; Larrew, T.W.; Eskandari, R. Pediatric hydrocephalus: current state of diagnosis and treatment. Pediatrics in Review 2016, 37, 478–490. [Google Scholar] [CrossRef]
- Ardissino, M.; Moussa, O.; Tang, A.; Muttoni, E.; Ziprin, P.; Purkayastha, S. Idiopathic intracranial hypertension in the British population with obesity. Acta Neurochirurgica 2019, 161, 239–246. [Google Scholar] [CrossRef]
- Andersson, J.; Rosell, M.; Kockum, K.; Lilja-Lund, O.; Söderström, L.; Laurell, K. Prevalence of idiopathic normal pressure hydrocephalus: a prospective, population-based study. PloS One 2019, 14, e0217705. [Google Scholar] [CrossRef]
- Fowler, J.B.; De Jesus, O.; Mesfin, F.B. Ventriculoperitoneal Shunt; StatPearls Publishing: StatPearls [Internet]. Treasure Island (FL), 2023. [Google Scholar]
- Akyol, M.E.; Cetin, E. Effects of shunt types used in idiopathic normal pressure hydrocephalus on patients’ clinical outcomes. Annals of Medical Research 2023, 30, 146–281. [Google Scholar] [CrossRef]
- Shannon, C.N.; Carr, K.R.; Tomycz, L.; Wellons, J.C.; Tulipan, N. Time to first shunt failure in pediatric patients over 1 year old: a 10-year retrospective study. Pediatric Neurosurgery 2015, 49, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Stein, S.C.; Guo, W. Have we made progress in preventing shunt failure? A critical analysis. Journal of Neurosurgery: Pediatrics 2008, 1, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Vinzani, M.; Alshareef, M.; Eskandari, R. Use of a Prophylactic Retrograde-Flushing Device in High-Risk Pediatric Patients with Ventriculoperitoneal Shunts: A Technical Note. Pediatric Neurosurgery 2023, 58, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.J.; Chiang, W.C.; Huang, H.Y.; Lin, S.Z.; Tsai, S.T. Effectiveness and safety of ventriculoperitoneal shunt versus lumboperitoneal shunt for communicating hydrocephalus: A systematic review and meta-analysis with trial sequential analysis. CNS Neuroscience & Therapeutics 2023, 29, 804–815. [Google Scholar]
- Yang, T.-H.; Chang, C.-S.; Sung, W.-W.; Liu, J.-T. Lumboperitoneal shunt: a new modified surgical technique and a comparison of the complications with ventriculoperitoneal shunt in a single center. Medicina 2019, 55, 1–10. [Google Scholar] [CrossRef]
- Yadav, Y.R.; Parihar, V.; Pande, S.; Namdev, H.; Agarwal, M. Endoscopic third ventriculostomy. Journal of Neurosciences in Rural Practice 2012, 3, 163–173. [Google Scholar] [CrossRef]
- Lu, L.; Chen, H.; Weng, S.; Xu, Y. Endoscopic third ventriculostomy versus ventriculoperitoneal shunt in patients with obstructive hydrocephalus: meta-analysis of randomized controlled trials. World Neurosurgery 2019, 129, 334–340. [Google Scholar] [CrossRef]
- Kulkarni, A.V.; Drake, J.M.; Mallucci, C.L.; Sgouros, S.; Roth, J.; Constantini, S.; Group, C.P.N.S. Endoscopic third ventriculostomy in the treatment of childhood hydrocephalus. The Journal of Pediatrics 2009, 155, 254–259.e251. [Google Scholar] [CrossRef]
- Hernandez, N.; Wang, S.; Ragheb, J. The Role of the Pediatric Neurosurgeon in the Management of Hydrocephalus Internationally. Journal of Global Neurosurgery 2022, 2. [Google Scholar] [CrossRef]
- Warf, B.C.; Weber, D.S.; Day, E.L.; Riordan, C.P.; Staffa, S.J.; Baird, L.C.; Fehnel, K.P.; Stone, S.S. Endoscopic third ventriculostomy with choroid plexus cauterization: predictors of long-term success and comparison with shunt placement for primary treatment of infant hydrocephalus. Journal of Neurosurgery: Pediatrics 2023, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ellenbogen, Y.; Brar, K.; Yang, K.; Lee, Y.; Ajani, O. Comparison of endoscopic third ventriculostomy with or without choroid plexus cauterization in pediatric hydrocephalus: a systematic review and meta-analysis. Journal of Neurosurgery: Pediatrics 2020, 26, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Marriott, W.M. The use of theobromin sodio salicylate (diuretin) in the treatment of hydrocephalus. American Journal of Diseases of Children 1924, 28, 479–483. [Google Scholar] [CrossRef]
- Hayden, P.W.; Foltz, E.L.; Shurtleff, D.B. Effect of an oral osmotic agent on ventricular fluid pressure of hydrocephalic children. Pediatrics 1968, 41, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Shurtleff, D.B.; Hayden, P.W. The treatment of hydrocephalus with isosorbide, an oral hyperosmotic agent. Journal of Clinical Pharmacology & New Drugs 1972, 108–114. [Google Scholar]
- Group, I.P.D.T. International randomised controlled trial of acetazolamide and furosemide in posthaemorrhagic ventricular dilatation in infancy. The Lancet 1998, 352, 433–440. [Google Scholar]
- Bass, N.H.; Fällström, S.; Lundborg, P. Digoxin-induced arrest of the cerebrospinal fluid circulation in the infant rat: implications for medical treatment of hydrocephalus during early postnatal life. Pediatric Research 1979, 13, 26–30. [Google Scholar] [CrossRef]
- Bergold, P.J.; Furhang, R.; Lawless, S. Treating traumatic brain injury with minocycline. Neurotherapeutics 2023, 20, 1546–1564. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Tan, C.; Wang, Y.; Tang, Z.; Zhang, Z.; Liu, J.; Xiao, G. Novel therapeutics for hydrocephalus: Insights from animal models. CNS Neuroscience & Therapeutics 2021, 27, 1012–1022. [Google Scholar]
- McAllister, J.P.; Miller, J.M. Minocycline inhibits glial proliferation in the H-Tx rat model of congenital hydrocephalus. Cerebrospinal Fluid Research 2010, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Winer, J.L.; Kitase, Y.; Brigman, J.L.; Jantzie, L.L. Neonatal administration of erythropoietin attenuates cognitive deficits in adult rats following placental insufficiency. Journal of Neuroscience Research 2022, 100, 2112–2126. [Google Scholar] [CrossRef]
- Robinson, S.; Conteh, F.S.; Oppong, A.Y.; Yellowhair, T.R.; Newville, J.C.; Demerdash, N.E.; Shrock, C.L.; Maxwell, J.R.; Jett, S.; Northington, F.J. Extended combined neonatal treatment with erythropoietin plus melatonin prevents posthemorrhagic hydrocephalus of prematurity in rats. Frontiers in Cellular Neuroscience 2018, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Jantzie, L.L.; Muthukumar, S.; Kitase, Y.; Vasan, V.; Fouda, M.A.; Hamimi, S.; Burkhardt, C.; Burton, V.J.; Gerner, G.; Scafidi, J. Infantile Cocktail of Erythropoietin and Melatonin Restores Gait in Adult Rats with Preterm Brain Injury. Developmental Neuroscience 2022, 44, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Safety of Erythropoietin and Melatonin for Very Preterm Infants With Intraventricular Hemorrhage (SCEMPI). Available online: https://clinicaltrials.gov/study/NCT05617833?term=robinson&intr=erythropoietin&rank=1 (accessed on 1/14/24).
- Indrawijaya, Y.Y.A.; Sumarno, S.; Suryaningtyas, W.; Husna, N.A. Drug utilization study of diuretics in children with hydrocephalus. In Proceedings of the Proceedings of International Pharmacy Ulul Albab Conference and Seminar (PLANAR), 2021; pp. 77–87.
- Del Bigio, M.R.; Di Curzio, D.L. Nonsurgical therapy for hydrocephalus: a comprehensive and critical review. Fluids and Barriers of the CNS 2015, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Diringer, M.N.; Edwards, D.F.; Zazulia, A.R. Hydrocephalus: a previously unrecognized predictor of poor outcome from supratentorial intracerebral hemorrhage. Stroke 1998, 29, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, Z.; Xi, G.; Keep, R.; Hua, Y. Deferoxamine attenuates acute hydrocephalus after traumatic brain injury in rats. Transl Stroke Res 2014, 5, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Zhao, G. Adult posthaemorrhagic hydrocephalus animal models. Journal of the Neurological Sciences 2017, 379, 39–43. [Google Scholar] [CrossRef] [PubMed]
- NS, V. Experimental hydrocephalus in young rats. Arkhiv Patologii 1957, 19, 44–52. [Google Scholar]
- Pudenz, R.H. Experimental and clinical observations on the shunting of cerebrospinal fluid into the circulatory system. Neurosurgery 1958, 5, 98–115. [Google Scholar] [CrossRef]
- McAllister, J.P.; Talcott, M.R.; Isaacs, A.M.; Zwick, S.H.; Garcia-Bonilla, M.; Castaneyra-Ruiz, L.; Hartman, A.L.; Dilger, R.N.; Fleming, S.A.; Golden, R.K. A novel model of acquired hydrocephalus for evaluation of neurosurgical treatments. Fluids and Barriers of the CNS 2021, 18, 1–17. [Google Scholar] [CrossRef]
- da Silva Lopes, L.; Slobodian, I.; Del Bigio, M.R. Characterization of juvenile and young adult mice following induction of hydrocephalus with kaolin. Experimental Neurology 2009, 219, 187–196. [Google Scholar] [CrossRef]
- Gonzalez-Darder, J.; Barbera, J.; Cerda-Nicolas, M.; Segura, D.; Broseta, J.; Barcia-Salorio, J.L. Sequential morphological and functional changes in kaolin-induced hydrocephalus. Journal of Neurosurgery 1984, 61, 918–924. [Google Scholar] [CrossRef]
- Nakayama, D.K.; Harrison, M.R.; Berger, M.S.; Chinn, D.H.; Halks-Miller, M.; Edwards, M.S. Correction of congenital hydrocephalus in utero I. The model: intracisternal kaolin produces hydrocephalus in fetal lambs and rhesus monkeys. Journal of Pediatric Surgery 1983, 18, 331–338. [Google Scholar] [CrossRef]
- Marlin, A.; Wald, A.; Hochwald, G.; Malhan, C. Kaolin-induced hydrocephalus impairs CSF secretion by the choroid plexus. Neurology 1978, 28, 945–945. [Google Scholar] [CrossRef]
- Liu, C.; Chen, Y.; Cui, W.; Cao, Y.; Zhao, L.; Wang, H.; Liu, X.; Fan, S.; Huang, K.; Tong, A. Inhibition of neuronal necroptosis mediated by RIP1/RIP3/MLKL provides neuroprotective effects on kaolin-induced hydrocephalus in mice. Cell proliferation 2021, 54, e13108. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Guo, J.; Yu, C.; Yang, J. Molecular mechanisms and risk factors for the pathogenesis of hydrocephalus. Frontiers in Genetics 2022, 12, 777926. [Google Scholar] [CrossRef]
- Yamada, H.; Oi, S.; Tamaki, N.; Matsumoto, S.; Taomoto, K. Embryopathoetiology of Congenital Hydrocephalus in Experimental Models: A Comparative Morphological Study in Two Different Models. In Proceedings of the Hydrocephalus; 1991; pp. 27–35. [Google Scholar]
- Stambolliu, E.; Ioakeim-Ioannidou, M.; Kontokostas, K.; Dakoutrou, M.; Kousoulis, A.A. The most common comorbidities in Dandy-Walker syndrome patients: a systematic review of case reports. Journal of Child Neurology 2017, 32, 886–902. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, J. Early neurovascular abnormalities underlying 6-aminonicotinamide (6-AN)-induced congenital hydrocephalus in rats. Teratology 1970, 3, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Gattone, V.H.; Tourkow, B.A.; Trambaugh, C.M.; Yu, A.C.; Whelan, S.; Phillips, C.L.; Harris, P.C.; Peterson, R.G. Development of multiorgan pathology in the wpk rat model of polycystic kidney disease. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology: An Official Publication of the American Association of Anatomists 2004, 277, 384–395. [Google Scholar] [CrossRef]
- Shim, J.W.; Territo, P.R.; Simpson, S.; Watson, J.C.; Jiang, L.; Riley, A.A.; McCarthy, B.; Persohn, S.; Fulkerson, D.; Blazer-Yost, B.L. Hydrocephalus in a rat model of Meckel Gruber syndrome with a TMEM67 mutation. Scientific Reports 2019, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hochstetler, A.E.; Smith, H.M.; Preston, D.C.; Reed, M.M.; Territo, P.R.; Shim, J.W.; Fulkerson, D.; Blazer-Yost, B.L. TRPV4 antagonists ameliorate ventriculomegaly in a rat model of hydrocephalus. Jci Insight 2020, 5, 1–14. [Google Scholar] [CrossRef]
- Hochstetler, A.; Smith, H.; Reed, M.; Hulme, L.; Territo, P.; Bedwell, A.; Persohn, S.; Perrotti, N.; D’Antona, L.; Musumeci, F. Inhibition of serum-and glucocorticoid-induced kinase 1 ameliorates hydrocephalus in preclinical models. Fluids and Barriers of the CNS 2023, 20, 61. [Google Scholar] [CrossRef]
- Mashayekhi, F.; Draper, C.E.; Bannister, C.M.; Pourghasem, M.; Owen-Lynch, P.J.; Miyan, J.A. Deficient cortical development in the hydrocephalic Texas (H-Tx) rat: a role for CSF. Brain 2002, 125, 1859–1874. [Google Scholar] [CrossRef]
- Jones, H.; Bucknall, R. INHERITED PRENATAL HYDROCEPHALUS IN THE H–Tx RAT: A MORPHOLOGICAL STUDY. Neuropathology and Applied Neurobiology 1988, 14, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.C.; Carter, B.J.; Morel, L. Characteristics of hydrocephalus expression in the LEW/Jms rat strain with inherited disease. Child's Nervous System 2003, 19, 11–18. [Google Scholar] [CrossRef]
- Itoh, K.; Fushiki, S. The role of L1 cam in murine corticogenesis, and the pathogenesis of hydrocephalus. Pathology International 2015, 65, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Swetloff, A.; Ferretti, P. Changes in E2F5 intracellular localization in mouse and human choroid plexus epithelium with development. International Journal of Developmental Biology 2004, 49, 859–865. [Google Scholar] [CrossRef]
- Banizs, B.; Pike, M.M.; Millican, C.L.; Ferguson, W.B.; Komlosi, P.; Sheetz, J.; Bell, P.D.; Schwiebert, E.M.; Yoder, B.K. Dysfunctional cilia lead to altered ependyma and choroid plexus function, and result in the formation of hydrocephalus. Development 2005, 132, 5329–5339. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, G.J.; Dagnino, L.; Gaubatz, S.; Xu, Y.; Bronson, R.T.; Warren, H.B.; Livingston, D.M. A specific, nonproliferative role for E2F-5 in choroid plexus function revealed by gene targeting. Genes & Development 1998, 12, 1092–1098. [Google Scholar]
- Lewis, W.R.; Malarkey, E.B.; Tritschler, D.; Bower, R.; Pasek, R.C.; Porath, J.D.; Birket, S.E.; Saunier, S.; Antignac, C.; Knowles, M.R. Mutation of growth arrest specific 8 reveals a role in motile cilia function and human disease. PLoS Genetics 2016, 12, e1006220. [Google Scholar] [CrossRef]
- Hochstetler, A.E.; Whitehouse, L.; Antonellis, P.; Berbari, N.F.; Blazer-Yost, B.L. Characterizing the Expression of TRPV4 in the Choroid Plexus Epithelia as a Prospective Component in the Development of Hydrocephalus in the Gas8GT Juvenile Mouse Model. The FASEB Journal 2018, 32, 750.712–750.712. [Google Scholar] [CrossRef]
- Abdelhamed, Z.; Vuong, S.M.; Hill, L.; Shula, C.; Timms, A.; Beier, D.; Campbell, K.; Mangano, F.T.; Stottmann, R.W.; Goto, J. A mutation in Ccdc39 causes neonatal hydrocephalus with abnormal motile cilia development in mice. Development 2018, 145, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Davy, B.E.; Robinson, M.L. Congenital hydrocephalus in hy3 mice is caused by a frameshift mutation in Hydin, a large novel gene. Human Molecular Genetics 2003, 12, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Bronson, R.T.; Lane, P.W. Hydrocephalus with hop gait (hyh): a new mutation on chromosome 7 in the mouse. Developmental Brain Research 1990, 54, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Di Curzio, D.L. Animal models of hydrocephalus. Open Journal of Modern Neurosurgery 2017, 8, 57–71. [Google Scholar] [CrossRef]
- Kume, T.; Deng, K.-Y.; Winfrey, V.; Gould, D.B.; Walter, M.A.; Hogan, B.L. The forkhead/winged helix gene Mf1 is disrupted in the pleiotropic mouse mutation congenital hydrocephalus. Cell 1998, 93, 985–996. [Google Scholar] [CrossRef]
- Kuwamura, M.; Kinoshita, A.; Okumoto, M.; Yamate, J.; Mori, N. Hemorrhagic hydrocephalus (hhy): a novel mutation on mouse chromosome 12. Developmental Brain Research 2004, 152, 69–72. [Google Scholar] [CrossRef]
- Lin, X.; Liu, B.; Yang, X.; Yue, X.; Diao, L.; Wang, J.; Chang, J. Genetic deletion of Rnd3 results in aqueductal stenosis leading to hydrocephalus through up-regulation of Notch signaling. Proceedings of the National Academy of Sciences 2013, 110, 8236–8241. [Google Scholar] [CrossRef]
- Ramos, C.; Fernández-Llebrez, P.; Bach, A.; Robert, B.; Soriano, E. Msx1 disruption leads to diencephalon defects and hydrocephalus. Developmental Dynamics: An Official Publication of the American Association of Anatomists 2004, 230, 446–460. [Google Scholar] [CrossRef]
- Jones, H.; Dack, S.; Ellis, C. Morphological aspects of the development of hydrocephalus in a mouse mutant (SUMS/NP). Acta Neuropathologica 1987, 72, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Jang, A.; Lehtinen, M.K. Experimental approaches for manipulating choroid plexus epithelial cells. Fluids and Barriers of the CNS 2022, 19, 1–15. [Google Scholar] [CrossRef]
- Millar, I.D.; Bruce, J.I.; Brown, P.D. Ion channel diversity, channel expression and function in the choroid plexuses. Cerebrospinal Fluid Research 2007, 4, 1–16. [Google Scholar] [CrossRef]
- Damkier, H.H.; Brown, P.D.; Praetorius, J. Cerebrospinal fluid secretion by the choroid plexus. Physiological Reviews 2013, 93, 1847–1892. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Z.; Chen, Y.; Liao, J.; Wang, Y.; Liu, J.; Lin, Z.; Xiao, G. Choroid plexus epithelium and its role in neurological diseases. Frontiers in Molecular Neuroscience 2022, 15, 949231. [Google Scholar] [CrossRef]
- Yang, Y.; He, J.; Wang, Y.; Wang, C.; Tan, C.; Liao, J.; Tong, L.; Xiao, G. Targeting choroid plexus epithelium as a novel therapeutic strategy for hydrocephalus. Journal of Neuroinflammation 2022, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sakka, L.; Coll, G.; Chazal, J. Anatomy and physiology of cerebrospinal fluid. European Annals of Otorhinolaryngology, Head and Neck Diseases 2011, 128, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Oernbo, E.K.; Steffensen, A.B.; Razzaghi Khamesi, P.; Toft-Bertelsen, T.L.; Barbuskaite, D.; Vilhardt, F.; Gerkau, N.J.; Tritsaris, K.; Simonsen, A.H.; Lolansen, S.D. Membrane transporters control cerebrospinal fluid formation independently of conventional osmosis to modulate intracranial pressure. Fluids and Barriers of the CNS 2022, 19, 65. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M. Mechanisms of ion transport across the choroid plexus. The Journal of Physiology 1972, 226, 545–571. [Google Scholar] [CrossRef] [PubMed]
- Bouillé, C.; Mesnil, M.; Barriere, H.; Gabrion, J. Gap junctional intercellular communication between cultured ependymal cells, revealed by lucifer yellow CH transfer and freeze-fracture. Glia 1991, 4, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhao, Q.; Graziano, J.H. Primary culture of choroidal epithelial cells: characterization of an in vitro model of blood-CSF barrier. In Vitro Cellular & Developmental Biology-Animal 1998, 34, 40–45. [Google Scholar]
- Mayer, S.E.; Sanders-Bush, E. Sodium-dependent antiporters in choroid plexus epithelial cultures from rabbit. Journal of Neurochemistry 1993, 60, 1308–1316. [Google Scholar] [CrossRef]
- Baehr, C.; Reichel, V.; Fricker, G. Choroid plexus epithelial monolayers–a cell culture model from porcine brain. Cerebrospinal Fluid Research 2006, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Harter, D.; Hsu, K.; Rose, H. Immunofluorescence and cytochemical studies of visna virus in cell culture. Journal of Virology 1967, 1, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Crook, R.B.; Kasagami, H. , Prusiner, S.B. Culture and characterization of epithelial cells from bovine choroid plexus. Journal of Neurochemistry 1981, 25, 215–227. [Google Scholar]
- Monnot, A.D.; Zheng, W. Culture of choroid plexus epithelial cells and in vitro model of blood–CSF barrier. Epithelial Cell Culture Protocols: Second Edition 2013, 13–29. [Google Scholar]
- Fejes, Z.; Pócsi, M.; Takai, J.; Erdei, J.; Tóth, A.; Balogh, E.; Rusznyák, Á.; Fenyvesi, F.; Nagy, A.; Kappelmayer, J. Preterm intraventricular hemorrhage-induced inflammatory response in human choroid plexus epithelial cells. International Journal of Molecular Sciences 2021, 22, 8648. [Google Scholar] [CrossRef] [PubMed]
- Guy, B.; Zhang, J.S.; Duncan, L.H.; Johnston Jr, R.J. Human neural organoids: Models for developmental neurobiology and disease. Developmental Biology 2021, 478, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Schroten, M.; Hanisch, F.-G.; Quednau, N.; Stump, C.; Riebe, R.; Lenk, M.; Wolburg, H.; Tenenbaum, T.; Schwerk, C. A novel porcine in vitro model of the blood-cerebrospinal fluid barrier with strong barrier function. PLoS One 2012, 7, e39835. [Google Scholar] [CrossRef] [PubMed]
- Hochstetler, A.; Hulme, L.; Delpire, E.; Schwerk, C.; Schroten, H.; Preston, D.; Simpson, S.; Blazer-Yost, B.L. Porcine choroid plexus-riems cell line demonstrates altered polarization of transport proteins compared with the native epithelium. American Journal of Physiology-Cell Physiology 2022, 323, C1–C13. [Google Scholar] [CrossRef]
- Lauer, A.N.; März, M.; Meyer, S.; Meurer, M.; de Buhr, N.; Borkowski, J.; Weiß, C.; Schroten, H.; Schwerk, C. Optimized cultivation of porcine choroid plexus epithelial cells, a blood–cerebrospinal fluid barrier model, for studying granulocyte transmigration. Laboratory Investigation 2019, 99, 1245–1255. [Google Scholar] [CrossRef]
- Hulme, L.; Hochstetler, A.; Schwerk, C.; Schroten, H.; Ishikawa, H.; Tung, C.-Y.; Perrin, B.; Blazer-Yost, B. Characterization of TRPV4-mediated signaling pathways in an optimized human choroid plexus epithelial cell line. American Journal of Physiology-Cell Physiology 2022, 323, C1823–C1842. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Dziegielewska, K.M.; Ek, C.J.; Habgood, M.D.; Bauer, H.; Bauer, H.-C.; Lindsay, H.; Wakefield, M.J.; Strazielle, N.; Kratzer, I. Correction: mechanisms that determine the internal environment of the developing brain: a transcriptomic, functional and ultrastructural approach. PLoS One 2016, 11, e0147680. [Google Scholar] [CrossRef]
- Shi, L.Z.; Zheng, W. Establishment of an in vitro brain barrier epithelial transport system for pharmacological and toxicological study. Brain Research 2005, 1057, 37–48. [Google Scholar] [CrossRef]
- Zheng, W. Toxicology of choroid plexus: Special reference to metal-induced neurotoxicities. Microscopy Research and Technique 2001, 52, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, I.; Ishiwat, C.; Ishiwata, E.; Sato, Y.; Kiguchi, K.; Tachibana, T.; Hashimoto, H.; Ishikawa, H. Establishment and characterization of a human malignant choroids plexus papilloma cell line (HIBCPP). Human Cell 2005, 18, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Muranyi, W.; Schwerk, C.; Herold, R.; Stump-Guthier, C.; Lampe, M.; Fallier-Becker, P.; Weiß, C.; Sticht, C.; Ishikawa, H.; Schroten, H. Immortalized human choroid plexus endothelial cells enable an advanced endothelial-epithelial two-cell type in vitro model of the choroid plexus. Iscience 2022, 25, 1–23. [Google Scholar] [CrossRef]
- Giovannucci, T.A.; Leckey, C.A.; Moncur, E.; Tariq, K.; Thorne, L.; Watkins, L.; Toma, A.; Fox, N.C.; Bateman, R.J.; Mills, K. Choroid plexus protein turnover in human choroid plexus organoids recapitulates turnover in humans measured using stable isotope labeling kinetics (SILK). Alzheimer's & Dementia 2023, 19, e074240. [Google Scholar]
- Praetorius, J.; Nielsen, S. Distribution of sodium transporters and aquaporin-1 in the human choroid plexus. American Journal of Physiology-Cell Physiology 2006, 291, C59–C67. [Google Scholar] [CrossRef]
- MacAulay, N.; Toft-Bertelsen, T.L. Dual function of the choroid plexus: cerebrospinal fluid production and control of brain ion homeostasis. Cell Calcium 2023, 102797. [Google Scholar] [CrossRef] [PubMed]
- Roepke, T.K.; Kanda, V.A.; Purtell, K.; King, E.C.; Lerner, D.J.; Abbott, G.W. KCNE2 forms potassium channels with KCNA3 and KCNQ1 in the choroid plexus epithelium. The FASEB Journal 2011, 25, 4264. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.; Lu, J.; Mount, D.; Delpire, E. Localization of the K+–Cl− cotransporter, KCC3, in the central and peripheral nervous systems: expression in the choroid plexus, large neurons and white matter tracts. Neuroscience 2001, 103, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Leefmans, F.J. CrossTalk proposal: Apical NKCC1 of choroid plexus epithelial cells works in the net inward flux mode under basal conditions, maintaining intracellular Cl− and cell volume. The Journal of Physiology 2020, 598, 4733–4736. [Google Scholar] [CrossRef] [PubMed]
- MacAulay, N.; Rose, C. CrossTalk opposing view: NKCC1 in the luminal membrane of choroid plexus is outwardly directed under basal conditions and contributes directly to cerebrospinal fluid secretion. The Journal of Physiology 2020, 598, 4737–4739. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Leefmans, F.J. Rebuttal from Francisco J. Alvarez-Leefmans. The Journal of Physiology 2020, 598, 4741–4742. [Google Scholar] [CrossRef]
- MacAulay, N.; Rose, C.R. Rebuttal from Nanna MacAulay and Christine R. Rose. The Journal of Physiology 2020, 598, 4743–4743. [Google Scholar] [CrossRef]
- Delpire, E.; Gagnon, K.B. Elusive role of the Na-K-2Cl cotransporter in the choroid plexus. American Journal of Physiology-Cell Physiology 2019, 316, C522–C524. [Google Scholar] [CrossRef]
- Fame, R.M.; Xu, H.; Pragana, A.; Lehtinen, M. Age-appropriate potassium clearance from perinatal cerebrospinal fluid depends on choroid plexus NKCC1. Fluids and Barriers of the CNS 2023, 20, 1–10. [Google Scholar] [CrossRef]
- Zeuthen, T.; MacAulay, N. Cotransport of water by Na+–K+–2Cl− cotransporters expressed in Xenopus oocytes: NKCC1 versus NKCC2. The Journal of Physiology 2012, 590, 1139–1154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhou, J.; Zhang, Y.; Liu, T.; Friedel, P.; Zhuo, W.; Somasekharan, S.; Roy, K.; Zhang, L.; Liu, Y. The structural basis of function and regulation of neuronal cotransporters NKCC1 and KCC2. Communications Biology 2021, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Macvicar, B.A.; Feighan, D.; Brown, A.; Ransom, B. Intrinsic optical signals in the rat optic nerve: role for K+ uptake via NKCC1 and swelling of astrocytes. Glia 2002, 37, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.-L.; Yu, M.-J.; Kassai, E.M.; Morris, R.G.; Hoffert, J.D.; Wall, S.M.; Knepper, M.A. Roles of basolateral solute uptake via NKCC1 and of myosin II in vasopressin-induced cell swelling in inner medullary collecting duct. American Journal of Physiology-Renal Physiology 2008, 295, F192–F201. [Google Scholar] [CrossRef] [PubMed]
- Blazer-Yost, B.L. Consideration of Kinase Inhibitors for the Treatment of Hydrocephalus. International Journal of Molecular Sciences 2023, 24, 1–13. [Google Scholar] [CrossRef]
- Koumangoye, R.; Bastarache, L.; Delpire, E. NKCC1: newly found as a human disease-causing ion transporter. Function 2021, 2, 1–16. [Google Scholar] [CrossRef]
- Gregoriades, J.M.; Madaris, A.; Alvarez, F.J.; Alvarez-Leefmans, F.J. Genetic and pharmacological inactivation of apical Na+-K+-2Cl− cotransporter 1 in choroid plexus epithelial cells reveals the physiological function of the cotransporter. American Journal of Physiology-Cell Physiology 2019, 316, C525–C544. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, J.; Minichiello, L. NKCC1 deficiency in forming hippocampal circuits triggers neurodevelopmental disorder: role of BDNF-TrkB signalling. Brain Sciences 2022, 12, 502. [Google Scholar] [CrossRef]
- Karimy, J.K.; Zhang, J.; Kurland, D.B.; Theriault, B.C.; Duran, D.; Stokum, J.A.; Furey, C.G.; Zhou, X.; Mansuri, M.S.; Montejo, J. Inflammation-dependent cerebrospinal fluid hypersecretion by the choroid plexus epithelium in posthemorrhagic hydrocephalus. Nature Medicine 2017, 23, 997–1003. [Google Scholar] [CrossRef]
- Sadegh, C.; Xu, H.; Sutin, J.; Fatou, B.; Gupta, S.; Pragana, A.; Taylor, M.; Kalugin, P.N.; Zawadzki, M.E.; Alturkistani, O. Choroid plexus-targeted NKCC1 overexpression to treat post-hemorrhagic hydrocephalus. Neuron 2023, 111, 1591–1608.e1594. [Google Scholar] [CrossRef]
- Bothwell, S.W.; Omileke, D.; Patabendige, A.; Spratt, N.J. CSF secretion is not altered by NKCC1 nor TRPV4 antagonism in healthy rats. Brain Sciences 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Savardi, A.; Borgogno, M.; De Vivo, M.; Cancedda, L. Pharmacological tools to target NKCC1 in brain disorders. Trends in Pharmacological Sciences 2021, 42, 1009–1034. [Google Scholar] [CrossRef]
- Shibasaki, K. TRPV4 ion channel as important cell sensors. Journal of Anesthesia 2016, 30, 1014–1019. [Google Scholar] [CrossRef]
- Liedtke, W.; Choe, Y.; Martí-Renom, M.A.; Bell, A.M.; Denis, C.S.; Hudspeth, A.; Friedman, J.M.; Heller, S. Vanilloid receptor–related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T. TRPV4 calcium entry channel: a paradigm for gating diversity. American Journal of Physiology-Cell Physiology 2004, 286, C195–C205. [Google Scholar] [CrossRef] [PubMed]
- Strotmann, R.; Harteneck, C.; Nunnenmacher, K.; Schultz, G.; Plant, T.D. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nature Cell Biology 2000, 2, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Darby, W.; Grace, M.; Baratchi, S.; McIntyre, P. Modulation of TRPV4 by diverse mechanisms. The International Journal of Biochemistry & Cell Biology 2016, 78, 217–228. [Google Scholar]
- Liedtke, W.; Friedman, J.M. Abnormal osmotic regulation in trpv4-/-mice. Proceedings of the National Academy of Sciences 2003, 100, 13698–13703. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, K.; Suzuki, M.; Mizuno, A.; Hara, A. Hearing impairment in TRPV4 knockout mice. Neuroscience Letters 2005, 382, 304–308. [Google Scholar] [CrossRef]
- Goyal, N.; Skrdla, P.; Schroyer, R.; Kumar, S.; Fernando, D.; Oughton, A.; Norton, N.; Sprecher, D.L.; Cheriyan, J. Clinical pharmacokinetics, safety, and tolerability of a novel, first-in-class TRPV4 ion channel inhibitor, GSK2798745, in healthy and heart failure subjects. American Journal of Cardiovascular Drugs 2019, 19, 335–342. [Google Scholar] [CrossRef]
- Lawhorn, B.G.; Brnardic, E.J.; Behm, D.J. TRPV4 antagonists: A patent review (2015–2020). Expert Opinion on Therapeutic Patents 2021, 31, 773–784. [Google Scholar] [CrossRef]
- Park, S.; Ku, S.K.; Ji, H.W.; Choi, J.-H.; Shin, D.M. Ca(2+) is a Regulator of the WNK/OSR1/NKCC Pathway in a Human Salivary Gland Cell Line. The Korean Journal of Physiology and Pharmacology 2015, 19, 249–255. [Google Scholar] [CrossRef]
- Toft-Bertelsen, T.L.; Barbuskaite, D.; Heerfordt, E.K.; Lolansen, S.D.; Andreassen, S.N.; Rostgaard, N.; Olsen, M.H.; Norager, N.H.; Capion, T.; Rath, M.F. Lysophosphatidic acid as a CSF lipid in posthemorrhagic hydrocephalus that drives CSF accumulation via TRPV4-induced hyperactivation of NKCC1. Fluids and Barriers of the CNS 2022, 19, 1–17. [Google Scholar] [CrossRef]
- Jin, S.C.; Furey, C.G.; Zeng, X.; Allocco, A.; Nelson-Williams, C.; Dong, W.; Karimy, J.K.; Wang, K.; Ma, S.; Delpire, E. SLC12A ion transporter mutations in sporadic and familial human congenital hydrocephalus. Molecular Genetics & Genomic Medicine 2019, 7, e892. [Google Scholar]
- Kahle, K.T.; Flores, B.; Bharucha-Goebel, D.; Zhang, J.; Donkervoort, S.; Hegde, M.; Begum, G.; Duran, D.; Liang, B.; Sun, D. Peripheral motor neuropathy is associated with defective kinase regulation of the KCC3 cotransporter. Science Signaling 2016, 9, ra77–ra77. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Alessi, D.R. The regulation of salt transport and blood pressure by the WNK-SPAK/OSR1 signaling pathway. Journal of Cell Science 2008, 121, 3293–3304. [Google Scholar] [CrossRef] [PubMed]
- Lauf, P.K.; Bauer, J. Direct evidence for chloride-dependent volume reduction in macrocytic sheep reticulocytes. Biochemical and Biophysical Research Communications 1987, 144, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Mercado, A.; Broumand, V.; Zandi-Nejad, K.; Enck, A.H.; Mount, D.B. A C-terminal domain in KCC2 confers constitutive K+-Cl-cotransport. Journal of Biological Chemistry 2006, 281, 1016–1026. [Google Scholar] [CrossRef]
- Quinton, P.M. Chloride impermeability in cystic fibrosis. Nature 1983, 301, 421–422. [Google Scholar] [CrossRef]
- Bear, C.E.; Li, C.; Kartner, N.; Bridges, R.J.; Jensen, T.J.; Ramjeesingh, M.; Riordan, J.R. Purification and functional reconstitution of the cystic fibrosis transmembrane conductance regulator (CFTR). Cell 1992, 68, 809–818. [Google Scholar] [CrossRef]
- Genovese, M.; Borrelli, A.; Venturini, A.; Guidone, D.; Caci, E.; Viscido, G.; Gambardella, G.; di Bernardo, D.; Scudieri, P.; Galietta, L.J. TRPV4 and purinergic receptor signalling pathways are separately linked in airway epithelia to CFTR and TMEM16A chloride channels. The Journal of Physiology 2019, 597, 5859–5878. [Google Scholar] [CrossRef]
- Johnsen, L.Ø.; Friis, K.A.; Damkier, H.H. In vitro investigation of the effect of proinflammatory cytokines on mouse choroid plexus membrane transporters Ncbe and NKCC1. Fluids and Barriers of the CNS 2023, 20, 1–14. [Google Scholar] [CrossRef]
- Praetorius, J.; Damkier, H.H. Transport across the choroid plexus epithelium. American Journal of Physiology-Cell Physiology 2017, 312, C673–C686. [Google Scholar] [CrossRef]
- Damkier, H.H.; Praetorius, J. Genetic ablation of Slc4a10 alters the expression pattern of transporters involved in solute movement in the mouse choroid plexus. American Journal of Physiology-Cell Physiology 2012, 302, C1452–C1459. [Google Scholar] [CrossRef]
- Christensen, I.B.; Wu, Q.; Bohlbro, A.S.; Skals, M.G.; Damkier, H.H.; Hübner, C.A.; Fenton, R.A.; Praetorius, J. Genetic disruption of slc4a10 alters the capacity for cellular metabolism and vectorial ion transport in the choroid plexus epithelium. Fluids and Barriers of the CNS 2020, 17, 1–18. [Google Scholar] [CrossRef]
- Yang, D.; Li, Q.; So, I.; Huang, C.-L.; Ando, H.; Mizutani, A.; Seki, G.; Mikoshiba, K.; Thomas, P.J.; Muallem, S. IRBIT governs epithelial secretion in mice by antagonizing the WNK/SPAK kinase pathway. The Journal of Clinical Investigation 2011, 121, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.; Ruusuvuori, E.; Sipilä, S.T.; Haapanen, A.; Damkier, H.H.; Kurth, I.; Hentschke, M.; Schweizer, M.; Rudhard, Y.; Laatikainen, L.M. Mice with targeted Slc4a10 gene disruption have small brain ventricles and show reduced neuronal excitability. Proceedings of the National Academy of Sciences 2008, 105, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Gurnett, C.A.; Veile, R.; Zempel, J.; Blackburn, L.; Lovett, M.; Bowcock, A. Disruption of sodium bicarbonate transporter SLC4A10 in a patient with complex partial epilepsy and mental retardation. Archives of Neurology 2008, 65, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Fasham, J.; Huebner, A.K.; Liebmann, L.; Khalaf-Nazzal, R.; Maroofian, R.; Kryeziu, N.; Wortmann, S.B.; Leslie, J.S.; Ubeyratna, N.; Mancini, G.M. SLC4A10 mutation causes a neurological disorder associated with impaired GABAergic transmission. Brain 2023, 146, 4547–4561. [Google Scholar] [CrossRef]
Figure 1.
Schematic of hydrocephalus categorization. Created with BioRender.com.
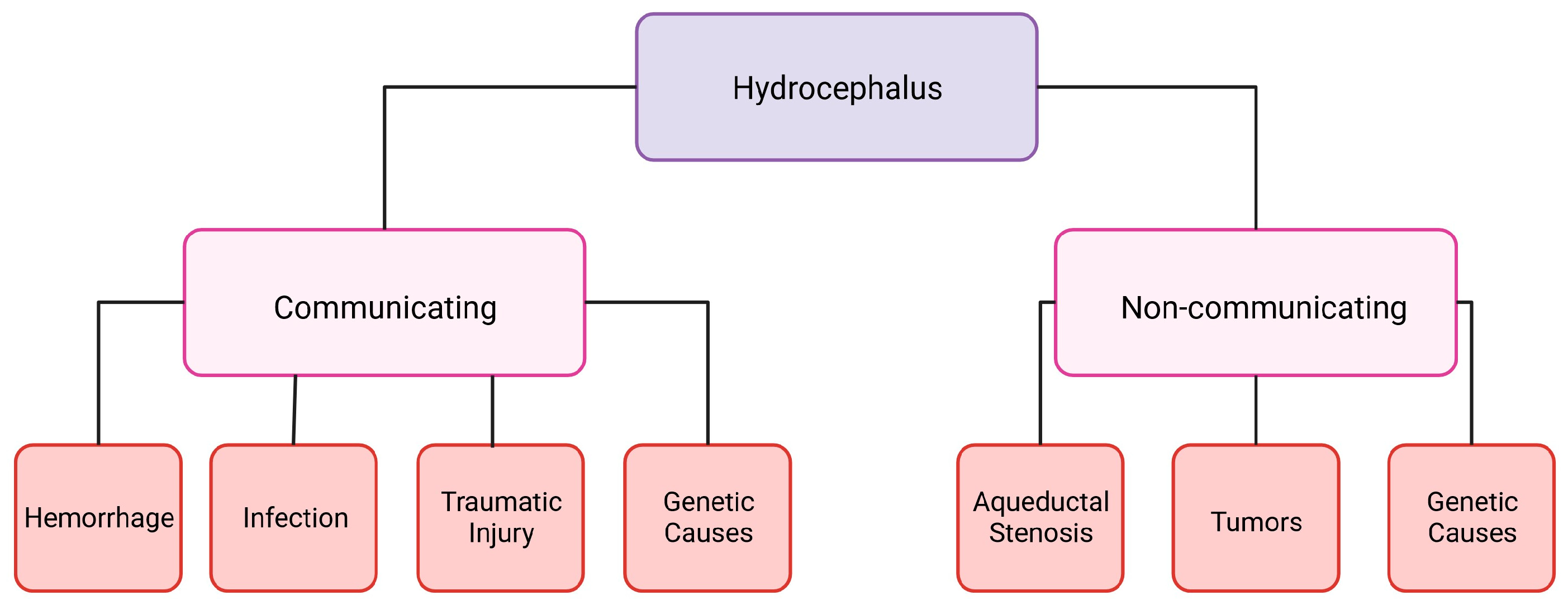
Figure 2.
Schematic of different choroid plexus models. The transporters and channels indicated in each cell are not meant to be comprehensive but to indicate those transport proteins that have been conclusively identified. PCP-R porcine choroid plexus-Riems, NKCC1 sodium potassium chloride co-transporter 1, TRPV4 transient receptor potential vanilloid-type 4, NCBE2 sodium-driven chloride bicarbonate exchanger 2, HIBCPP human choroid plexus papilloma, AE2 anion exchanger 2, NCBE sodium-driven chloride bicarbonate exchanger, iHCPEnC immortalized human choroid plexus endothelial cell. Created with BioRender.com.
Figure 2.
Schematic of different choroid plexus models. The transporters and channels indicated in each cell are not meant to be comprehensive but to indicate those transport proteins that have been conclusively identified. PCP-R porcine choroid plexus-Riems, NKCC1 sodium potassium chloride co-transporter 1, TRPV4 transient receptor potential vanilloid-type 4, NCBE2 sodium-driven chloride bicarbonate exchanger 2, HIBCPP human choroid plexus papilloma, AE2 anion exchanger 2, NCBE sodium-driven chloride bicarbonate exchanger, iHCPEnC immortalized human choroid plexus endothelial cell. Created with BioRender.com.
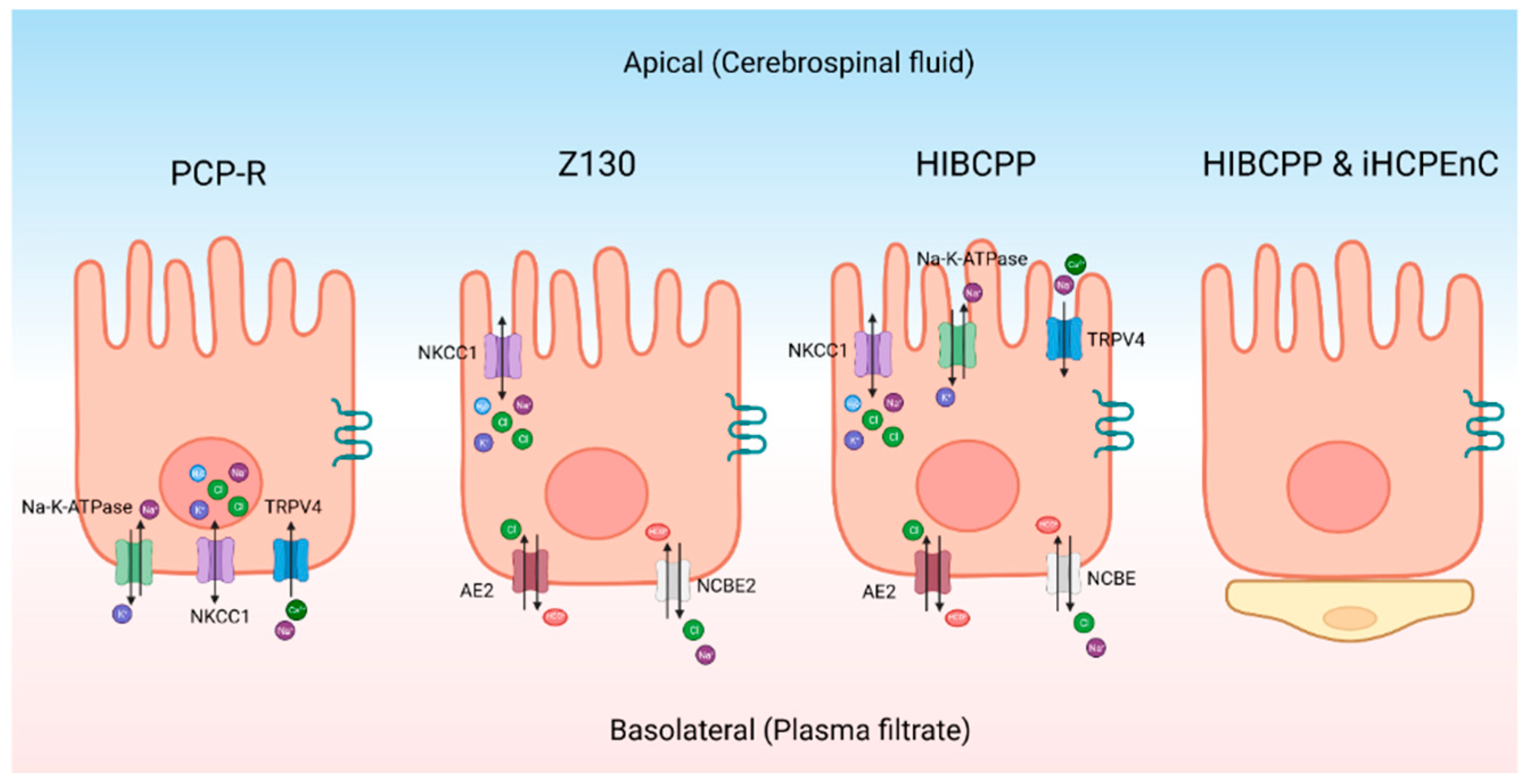
Figure 3.
Schematic of a choroid plexus cell with the ion transporters that may be involved with hydrocephalus. NKCC1 Sodium potassium chloride co-transporter 1, CSF cerebral spinal fluid, TRPV4 transient receptor potential vanilloid-type 4, KCC3 potassium chloride cotransporter 3. Created with BioRender.com.
Figure 3.
Schematic of a choroid plexus cell with the ion transporters that may be involved with hydrocephalus. NKCC1 Sodium potassium chloride co-transporter 1, CSF cerebral spinal fluid, TRPV4 transient receptor potential vanilloid-type 4, KCC3 potassium chloride cotransporter 3. Created with BioRender.com.
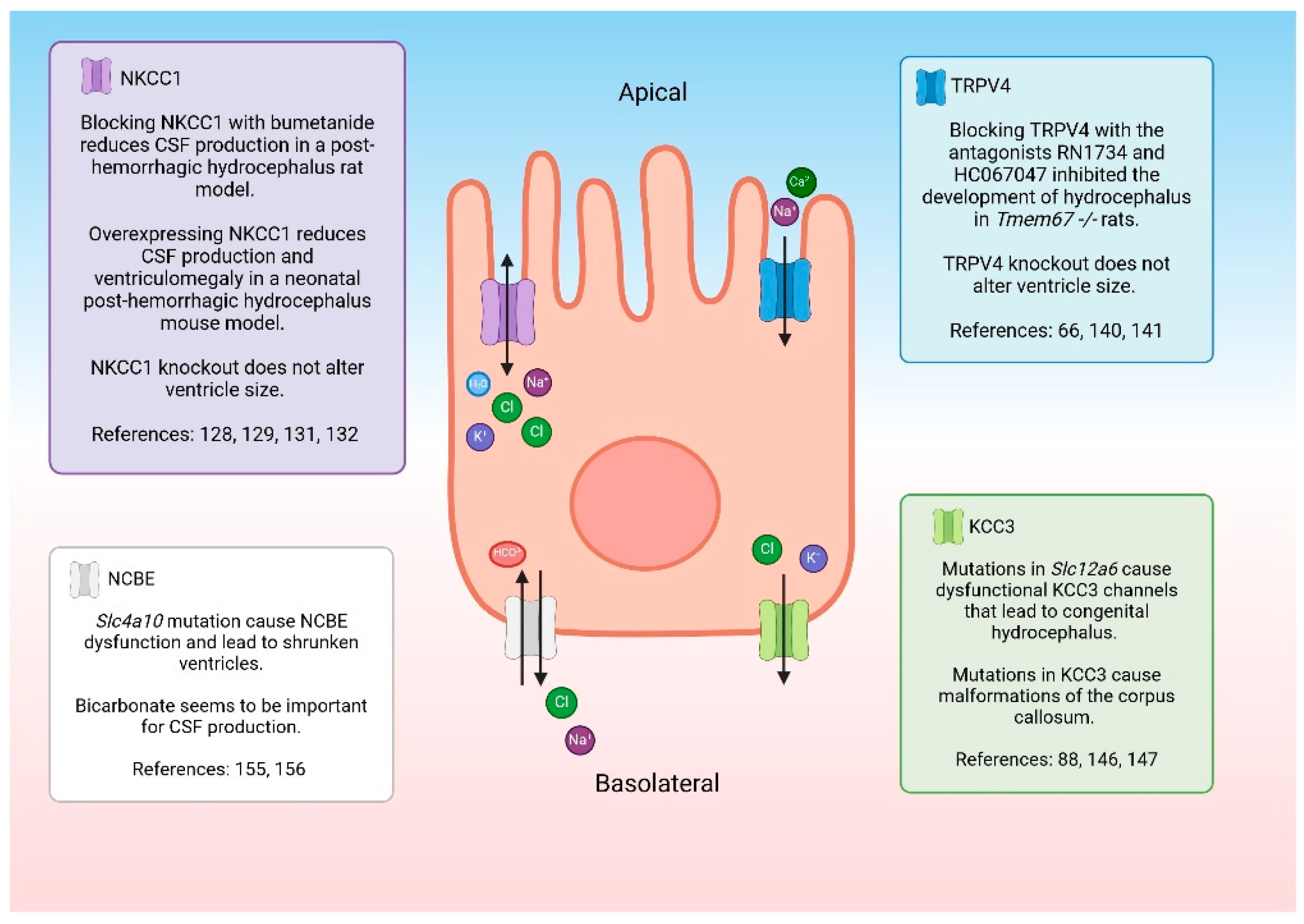

Table 1.
Benefits and complications associated with current hydrocephalus treatments.
| Treatment Type | Benefits | Complications | References |
|---|---|---|---|
| Ventriculoperitoneal shunts (VPS) | Treats both communicating and non-communicating hydrocephalus Effective for patients suffering from iNPH |
Failure due to mechanical problems, catheter migration, cell overgrowth Fails frequently in pediatric patients; surgery required for shunt revisions Possibility of infection |
[23,24,25] |
| Ventriculoperitoneal shunts and the Reflow® System | Fewer shunt obstructions | Still in clinical trials Can fail from other complications typical of VPS |
[26] |
| Lumboperitoneal shunts (LPS) | Does not require brain surgery Decreases chances of brain hemorrhage |
Cannot be used for non-communicating hydrocephalus Failure due to mechanical problems, catheter migration, cell overgrowth Possibility of infection |
[27,28] |
| Endoscopic third ventriculostomy (ETV) | Does not require a shunt | Pediatric patients do not respond to treatment Requires brain surgery Possibility of infection |
[29,30,31] |
| Endoscopic third ventriculostomy/choroid plexus cauterization (ETV/CPC) | Lower need for re-operation when compared to ETV alone May be more beneficial in developing countries |
Long term effects of CPC have not been studied Possibility of infection |
[32,33,34] |
Table 2.
Current animal models used for studying hydrocephalus.
| Disease Model | Animal Models | Type of Hydrocephalus | References |
|---|---|---|---|
| Hydrocephalus caused by traumatic brain injury | Fluid percussion injury model with injection of FeCl3 | Communicating | [50] |
| Post-hemorrhagic hydrocephalus | Induced models using injections of blood, red blood cells, iron, hemoglobin | Communicating | [8,44,51] |
| Chemically induced hydrocephalus | Kaolin injected models, 6-AN rats | Communicating | [52,53,54,55,56,57,58,59,60,61,62,63] |
| Genetic hydrocephalus models | Rat: Wpk, LEW/Jms Mouse: L1CAM, E2F5, Gas8, CCDC39, Hy-3, Hpy, Hyh, Msx1, SUMS/NP |
Communicating | [64,65,70,71,72,73,74,75,76,77,78,79,80,84,85,] |
| Genetic hydrocephalus models | Rat: H-Tx Mouse: Rnd3, Hhy |
Non-communicating | [60,68,69,82,83] |
| Genetic model of post-hemorrhagic hydrocephalus | Mouse: Mf1 Mouse: Hhy |
Communicating Non-communicating |
[60,81,82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



