
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Due to the ongoing climate change plants face multiple stress factors (mainly the abiotic ones) simultaneously, which means that the reduced yields of critical crops (including wheat, rice, or barley) can be expected. Therefore, our understanding of the light and temperature sensing mechanisms and related signaling is of primary importance. Herein we present a review focused on the direct sensing (ensured via photoreceptors) of light, which is the main abiotic factor driving the development, phenology, and especially resistance against oxidative stress via the increased accumulation of protective metabolites of crops. Thus, in the following text, special attention is paid to the description of the plant photoreceptors, their mode of action, ranges of perception, and related signaling pathways leading to the activation of target transcription factors. These finally lead to alterations in the expression of target genes related to photoprotection. Due to the focus of our research, genes related to the production of phenolic compounds (PheCs) were selected as a representative group of target genes having a UV-screening, and antioxidative function meaning that their activation and activity of resulting proteins can significantly contribute to the increased tolerance of plants against stress cues.
Keywords:
light
; phenolic compounds
; photoreceptors
; temperature
; transcription factors
1. Introduction
Under natural conditions, plants have to cope with the coaction of multiple abiotic environmental factors (for example light, temperature, or drought) [1]. Plants developed overlapping and cross-talking sensing mechanisms and related signaling pathways for light and temperature. Due to the primary importance of light for plant growth, development, and reproduction timing [2,3], we decided to focus more in-depth on the light perception and related signaling pathways leading to the activation of the transcription factors directly controlling the target gene expression.
Although the vast majority of light-sensing studies in plants are focused on the mechanisms of direct light perception, we can distinguish also the indirect one. Indirect light sensing does not include any specialized proteins and includes less studied, yet very interesting (and important) mechanisms - sensing of the incident radiation via the reactive oxygen species (ROS), related redox changes, and also sugar sensing - TOR kinase, or SnRK1-based signaling [4]. Several examples of the indirect light sensing mechanisms in plants can be found in Chapter 2., but (concerning the main scope of this work) this part is very brief, and therefore it is strongly recommended to read original resources.
In comparison to indirect sensing, direct light sensing requires specialized proteins called photoreceptors, which have (at least in most cases) bound nonprotein chromophores, which are responsible for the light absorption itself. After the light absorption, the photoreceptor turns into its active state, generally caused by conformational changes promoting the photoreceptor interactive potential - usually, this activation is followed by the downstream protein-protein interactions finally leading to the activation of the transcriptional factors, which directly regulate the expression of target genes. In plants, we can distinguish several groups of the photoreceptors, based on their absorption range and other (structural) features. Therefore, the detailed mechanism of the direct photo perception per each group is described below in Chapter 3.
We decided to summarize current knowledge about the direct light sensing ensured via the photoreceptors and describe the following interactions of the active (excited) receptors, via the protein-protein interactions reaching the transcription factors, which directly regulate the expression of the target genes. As a model group of such gene expression which is regulated by photoreceptors, these, related to the PheCs production were selected.
2. Indirect Light Sensing
Although the vast majority of incident radiation plants perceive directly (by the photoreceptors), there also exist alternative sensory mechanisms and pathways, which are related to light sensing yet are not so at the center of research interest. Therefore, we selected those, that are very important and contribute to light perception, we briefly describe their mode of action, and we summarized literature documenting their importance in light sensing.
The first example of a nonspecific light sensory system is via reactive oxygen species (ROS). We can distinguish radical and nonradical ROS based on, whether they have an unpaired electron in the valence orbital. ROS can be formed physiologically in the cells - specifically in the mitochondria, chloroplasts, or peroxisomes during the metabolic processes [5]. On the other hand, the high irradiance conditions can lead to damage of the photosynthetic apparatus, and their amount in the cells steeply increases, which means, that other interactions with the biomolecules (lipid, proteins, DNA, or RNA) can occur leading to the cell damage. Another link between ROS and direct light signalings was discovered recently - it was documented, that plant cryptochromes during their photocycle release ROS in vitro, which opens up a question, of whether the same occurs in vivo and how the crosstalk between the photoreceptors and ROS are coordinated [6].
Due to the impressive reactive potential to interact with other biomolecules, the levels of ROS have to be precisely regulated. For such purposes, plants developed a complex antioxidative network including both low molecular weight (tocopherol, carotenoids, phenolic compounds, or proline) as well as high molecular weight antioxidants - enzymes (such as SOD, CAT, APX) [5]. In addition, it was documented, that the activity of antioxidative enzymes can be significantly affected by the interaction with plant phytohormones. Such an example can be the interaction of salicylic acid with catalase which probably reduces the catalase activity [7].
Incident light is directly related to photosynthesis and therefore to synthesis of carbohydrates. Resulting carbohydrates (sugars) directly affect the activity of the kinases, which transduce the signal via protein-protein interactions and directly contribute to the control of the target gene expression [8]. The first example of the kinase related to the gene expression control in response to the sugar formation can be TOR (Target of rapamycin), which is part of the TORC complex [9]. In general , there were documented TORC1 and TORC2 complexes, which differ in the core protein composition [9]. While in plants TORC1 complex is documented, whereas TORC2 is absent, but the existence of other TORCs cannot be excluded [9]. TORC1 complex in plants consists of TOR kinase, RAPTOR (Regulatory-associated protein of TOR), and LST8 (Lethal with SEC13 protein 8) core components [9]. Based on the available data, the TORC1 complex in Arabidopsis can interact with more than 180 proteins [10] depending on the conditions plants are exposed to, and can lead to the increased expression of genes related to various biosynthetic processes (including photosynthesis, biosynthesis of amino acids, or nucleotides). Although our understanding of the complex role of plant TORC1 in transcription control increases, our knowledge is still very limited. For example, it has been documented, that in vitro TORC1 can phosphorylate E2Fa and E2Fb [9,10]. These transcription factors play a role in orchestrating the expression of genes related to thermotolerance and thermal memory [10].
Another such sensory system is hexokinase1 (HXK1), this enzyme phosphorylates either glucose or fructose (preferentially glucose) and connects nutrient, light, and hormone signaling [11,12]. To ensure further signaling steps, HXK1 requires interaction with other proteins. It was documented, that HXK1 can form the complex with the VHA-B (vacuolar proton pump - subunit B), or components of Polycomb Repressive Complex 2 - SWN (SWINGER), and - CLF (CURLY LEAF) [13]. Interaction of HXK1 with the CLF and SWN leads to the trimethylation of H3 lysine in the promoter region of the target genes and therefore stimulates the expression of target genes [13]. In the nucleus of Arabidopsis thaliana, the presence of glucose leads to increased degradation of EIN3 (Ethylene insensitive 3) by AtHXK1, resulting in a diminished response to ethylene-induced signaling [13]. In addition, MdHXK1 phosphorylates Ser361 of the MdbHLH3 protein and thus stabilizes the interaction between MdbHLH3 and MYB1-9-11, which leads to the stimulation of the anthocyanin's biosynthesis under the low-temperature conditions in Malus domestica [13].
The last indirect light sensing mechanism is via the SnRK1 kinase (Sucrose nonfermenting related kinase 1). In contrast to the TORC1 and HXK1, SnRK1 is activated under the low availability of carbohydrates [14] (so-called "carbon starvation"). This particular kinase phosphorylates enzymes from several biosynthetic pathways such as HMG-Coenzyme A (mevalonate pathway), sucrose phosphate synthase (enzyme from sucrose biosynthetic pathway), nitrate reductase (the enzyme responsible for the reduction of nitrate anions to nitrite), 6-phosphofructo-2-kinase/2,6-fructose-bis-phosphatase (enzyme driving the glycolytic pathway), or trehalose-6-phosphate synthase (which catalyzes the formation of trehalose-6-phosphate) [14]. Besides the phosphorylation of the enzymes directly related to the metabolism, SnRK1 can interact, phosphorylate, and therefore control the activity of the transcription factors. Such examples can be bZIP63, WRI, WRKY3, or KFB proteins [14], which induce the ubiquitination (and therefore decomposition) of the PAL (Phenylalanine ammonia-lyase; [15]). The activity of SnRK1 is inhibited by phosphorylated carbohydrates such as glucose-6-phosphate, glucose-1-phosphate, or trehalose-6-phosphate [16]. These phosphorylated carbohydrates strongly affect the SnRK1 activity and are direct products of the TORC1 pathway activity, which indicates the tight coordination of these sensory systems in the monitoring of plant energy status and related responses.
In terms of indirect light perception, all these above-mentioned sensory systems have irreplaceable roles. For example, ROS (induced by the methyl viologen) can lead to the increased expression of genes related to antioxidative protection in Arabidopsis thaliana - such examples can be ascorbate peroxidase 1 (APX1), glutathione-s-transferase 6 (GST6), or heat-shock protein 70-3 (HSP70-3) [17]. In addition, these genes responded to the increased light irradiance, therefore a link between ROS and light sensing has been for a long time presumed [17]. This assumption was further strengthened by the transcriptomic study from 2018 by Zandalinas et al [18], which identified several hundreds of genes responding directly to the applied solution of hydrogen peroxide, and some of these genes even overlapped with genes responding to the excess light (also tested in this study). Furthermore, Zandalinas et al. described ROS-induced TFs, which might be involved in the transcription control of ROS-responsive genes, and some of them are probably involved in the light stress responses as well [18]. Similarly, sugar kinases have a crucial role in light/dark perception and photoperiod sensing - for example, HXK1 defective mutants displayed reduced growth in response to light in comparison with the WT Arabidopsis thaliana [19] plants. In a similar manner as the HXK1, also the SnRK1 activity during the light/dark phases fluctuates, which was monitored in vivo using the artificial reporter sequence in Arabidopsis thaliana plants. SnRK1 activity was inhibited as the concentration/amount of the Tre-6-P increased [20]. In addition, TORC1 is tightly related to the light/dark perception as well. Hajibezhad et al in 2023 documented TORC feedback loop integrating signals about the energy status and light with an impact on the Arabidopsis thaliana habit [21]. In the presence of light, the TORC perceives signals about the energy status from the available sugars, as described above, and therefore positively affects the expression of target genes (including ATH1), while in the dark COP1 complex leads to the decreased activity of TORC and therefore reduced expression of target genes [21].
3. Direct Light Sensing
3.1. Basic Description of Photoreceptors and Mechanism of its Activation
Direct light sensing is ensured via the photoreceptors operating in specific spectral intervals of solar radiation (Figure 1.). Blue light is sensed by a diverse group of photoreceptors - specifically cryptochromes (CRYs), phototropines (PHOTs), and zeitlupes (ZTLs). CRYs are relatively short proteins (over 480 amino acids in Hordeum vulgare; [11]) and bind flavin adenine dinucleotide which ensures the ability to absorb light (it is assumed, that flavin radical plays major role in the photocycle [22]) in the range of 320 - 500nm [23]. Within the CRYs, CRY1 and CRY2 can be distinguished. Interestingly, CRYs-related signaling can modulate the expression of 10 - 20% of the Arabidopsis thaliana coding genes [24]. In monocot plants, CRY1 is encoded by 2 different genes (sharing 81 % nucleotide identity in Hordeum vulgare [25]) providing 2 different transcripts designated as CRY1a and CRY1b. CRY2 is encoded by an independent gene – the main difference compared to CRY1 is in the length of gene sequence, exon-intron structure, presence or absence of 5´UTR (five prime untranslated region of RNA), and amino acid composition of the resulting protein [25]. From the structural point of view, they contain PHR (Photolyase Homologous Region) and CCE (CRY C-terminal Extension, also referred to as CCT domain) domains.
The general model of CRYs sensing is based on the blue light absorption via the incorporated FAD (which has its own photocycle and can be oxidized if needed; electron donors are proximal tryptophan residues), which leads to the conformational changes of protein causing the formation of CRY homodimers (in the darkness, the excited CRY dimers can be recovered spontaneously in a temperature-dependent manner). The active dimer is phosphorylated (which seems not to be related to the signaling itself) and later can interact with COP1/SPA (CONSTITUTIVE PHOTOMORPHOGENIC 1 and SUPPRESSOR OF PHYTOCHROME A-105; Figure 2.) E3 ubiquitin ligase complex (cryptochrome is ubiquitinated and later on degraded). COP1/SPA complex is therefore occupied, HY5 transcription factor is no longer repressed and the target gene expression can be initiated [26]. In addition, the transition of CRYs from monomers to dimers can be repressed by the interaction with BIC1 (BLUE-LIGHT INHIBITOR OF CRYPTOCHROMES 1) and BIC2 (BLUE-LIGHT INHIBITOR OF CRYPTOCHROMES 2) proteins which provide the additional level of the signaling control. On the other hand, this model does not reflect the most recent knowledge about the CRYs potential to form tetramers [27,28,29].
The second group of blue light photoreceptors are PHOTs. Those proteins are about 900 amino acids long (PHOT1 – 996 amino acids, PHOT2 – 915 amino acids in Arabidopsis thaliana) [11] and they dispose of two LOV (Light-oxygen-voltage-sensing domain) domains (LOV1 and LOV2; PAS family domain members) with a photosensory center at N terminus, serine/threonine kinase domain located at C terminus (responsible for AGC /automatic gain control/ type signaling - cAMP-dependent, cGMP-dependent, and phospholipid-dependent proteinkinase C type; [11]) and Jα helix (links LOV2 domain with kinase). In the dark, the serine/threonine kinase domain is in close proximity to LOV domains, but after the light absorption by its chromophores FMNs (two flavine mononucleotides bound to the LOV domains [30]), the PHOTs conformational changes occur (kinase domain is dissociated from the LOVs proximity and the Jα helix is disorganized), the kinase domain is activated and phosphorylates specific amino acid residues in the PHOTs sequence (but also other substrates can be phosphorylated; [30,31,32,33]. Although the PHOT signaling pathway has not been completely described yet, several proteins able to interact with PHOTs have been identified. The first example is the NPH3 (NON-PHOTOTROPIC HYPOCOTYL 3) protein which directly interacts with PHOT1 and is crucial for the photoreceptor complex constitution (Figure 2.; [34]). The second protein capable of interacting with with PHOT1 is RPT2 (ROOT PHOTOTROPISM 2) which is also (similarly to NPH3) involved in the phototropism and stomata opening [35,36].
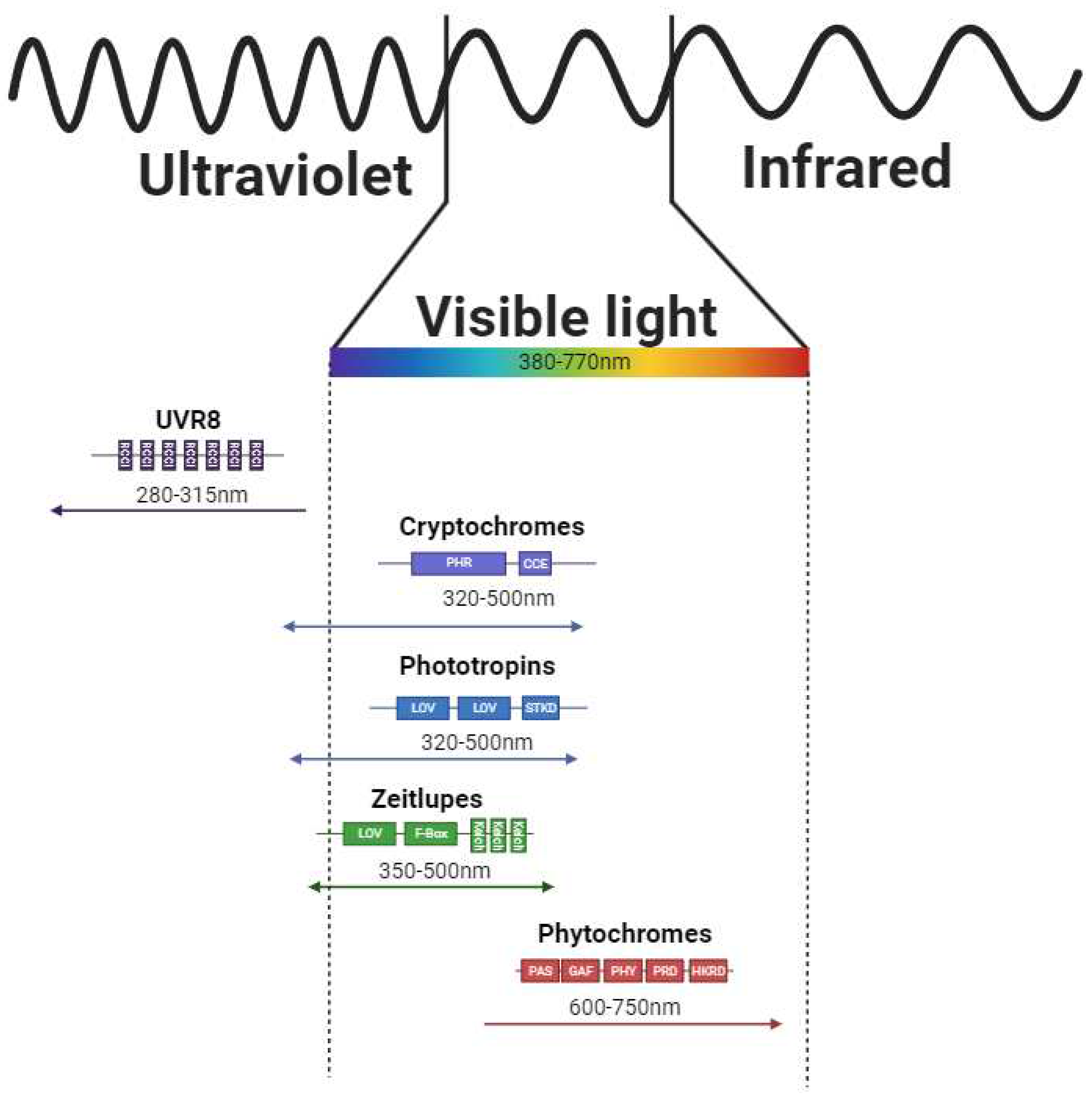
Another group of photoreceptors capable of light sensing are ZTLs. Those proteins are responsible for sensing in blue/green light regions (some publications call them even “green light photoreceptors”, while others include them in blue light receptors). The absorption range of the ZTLs is from 350 - 500 nm [41], differs in light and darkness, and spans UVA, blue, cyan, and green spectral components. ZTLs dispose of LOV (described above), F-box, and kelch domains. The LOV domain contains (similarly to the PHOTs) flavine (FAD, FMN, or riboflavin; [42]) bound to LOV protein via cysteine residues. The main function of ZTLs (together with the FKF1 - Flavin-binding kelch repeat F-box protein 1 and LKP - LOV kelch protein) is the sensing of the photoperiod duration and flowering time [41]. The ZTLs and FKF1 (Flavin-Kelch-Fbox-1) proteins can form heterodimers with the GI (GIGANTEA) protein. The formation of such (ZTL/GI and FKF1/GI) complexes is induced by the light (specifically its blue component). The reduction of light dose ([41]; especially its blue component) during the light/dark transition causes the ZTL release from the complex with GI and enables interaction of ZTL with the TOC1 (Timing Of Cab expression 1; transcription repressor) leading to its degradation during the dark period resulting in the dynamic changes in the transcription of the target genes [41,43]. Similarly, the decreasing light dose (especially its blue component) leads to the release of the FKF1 protein from the heterodimer with the GI, therefore, the FKF1 can interact with the CDF (Cycling Dof Factor) which is a transcriptional repressor and therefore the transcription of the related genes can be triggered [41,43].
The specific group of photoreceptors are UVR8s. The absorption range is from approximately 280 - 315 nm ([44]; and differs for the monomeric and dimeric state [45]), therefore these photoreceptors are able to absorb UV-A and also UV-B radiation.
The specificity of UVR8 lie in the fact that it does not contain a nonprotein chromophore, but instead it has several RCC1 domains resulting in a beta-propeller structure with aro-matic amino acids (tryptohans) situated at the propeller's core. Tryptophans, being aromatic amino acids, possess the capability to directly absorb light and transmit excitation energy. Within the UVR8 protein, these amino acids aggregate into three tryptophan clusters, with six tryptophans forming the distal cluster, three in the proximal cluster, and four in the central cluster, effectively fulfilling the role typically played by nonprotein ligands (such as FAD or FMN) in other photoreceptors. In Arabidopsis, the transport of excitation energy within UVR8 is facilitated through resonance energy transfer [46], starting from the distal tryptophan cluster (W39, W92, W144, W196, W300, W352), then passing through the proximal cluster (W198, W250, W302), and finally reaching the central cluster (W233, W285, W337, W94). Although the ultimate destiny of the excitation energy remains unknown, it is established that a conformational change occurs within the entire UVR8 structure, leading to UVR8 monomerization and consequent alterations in its potential interactions with other proteins.
Inactive UVR8 forms dimers, but once the UV radiation photon is absorbed, the dissociation occurs and UVR8 forms an active monomer. Active UVR8 monomer binds the COP1/SPA/E3 ubiquitin ligase complex and induces COP1/SPA dissociation from the enzymatic complex (CUL4/DDB1/E3 ubiquitin ligase; Figure 2.). Active complex UVR8/COP1/SPA then migrates to the nucleus. Here UVR8/COP1/SPA complex directly interacts with the HY5 transcription factor and therefore increases its stability. In addition, HY5s binding to its own promoter [47] can be induced.
The only known photoreceptors responsible for red and far-red perception are phytochromes (PHYs). Phytochromes are approximately 124kDa proteins that form homodimers and contain covalently bound pigments (tetrapyrroles - specifically phytochromobilin [48]). Their tertiary structure contains N-terminal extension, PAS (per-ARNT-sim), GAF (cGMP phosphodiesterase/adenylyl cyclase/FhlA domain), PHY (phytochrome specific domain), PRD (PAS-related domain) and HKRD (histidine kinase-related domain) domains [49]. Based on the phylogenetic analysis we can distinguish phytochrome subfamily members, which seem to be genus-specific and therefore the number of members varies amongst plant species. For example, in Arabidopsis thaliana we can recognize PhyA-PhyE, while in spring barley PhyA-PhyC are described [11]. On the other hand, several studies showed that the most important for environmental sensing are PhyA [50] and PhyB [51,52], but the role of remaining PHY family members remains to be understood. The precise mechanism of the Phy-related signaling will be described below.
4. Photoreceptor Signaling and Thermosensing
In the moderate range of temperatures, the majority of the thermal sensing is usually attributed to the phytochromes (specifically PhyB; Figure 3.; [51,53], which seems to be the main perceptive molecule for temperature and thermomorphogenesis. Phytochromes can be found in two different states - inactive (Pr) and active (Pfr), whose transitions are caused by red and far-red radiation, as depicted in Figure 2 and their ratio respectively. While the red light activates PhyB, the far-red light inactivates it (and the same process is induced by the increased temperature). Such transitions are accompanied by the conformational changes responsible for shifts of absorption spectra, and it changes the potential for interaction with other proteins. Specifically, when the phytochrome is in its active state, it is able to interact with phytochrome interacting factors (PIFs - PIF4 and 7, responsible for positive control of the gene expression - transcriptional activators), ubiquitinate and degrade the 26S proteasome follows and therefore the gene expression is reduced. In contrast, once the phytochromes are inactivated (by the increased temperature or far-red light), they can´t interact with PIFs and therefore the transcription of the target genes can be initiated.
PIFs target genes containing in the promoters G-boxes (for example genes related to the PheCs biosynthesis contain such sequence motif) or G/PBE boxes and can form homodimers or heterodimers both with different PIFs or completely different proteins [54]. Such an example can be interaction with HY5 ([54]; Protein LONG HYPOCOTYL 5), involved in light-related signaling.
Besides the direct interaction with activated phytochromes, PIFs (PIF4 and PIF7) transcription (PIF4) and translation (PIF7) are precisely finetuned by the alternative regulatory mechanisms. For example, the PIF4 expression under lower temperatures is blocked by the ELF3 protein, but once the temperature increases it leads to the phase separation of the ELF3 and therefore to the initiation of target genes transcription. Similarly, in the 5´-UTR (five prime untranslated region of the mRNA) of the PIF7 mRNA, there is localised a hairpin, which changes the structure with increased temperature and is directly linked with the increased synthesis of PIF7 proteins [56].
Moreover, all of the before mentiond photoreceptors sensing in the UV and blue /green region could be involved in the thermosensing process (PHOTs, CRYs, ZTLs, and UVR8 [57]). For example, PHOTs in Marchantia polymorpha displayed a prolonged duration of the active state [58] under decreased temperature. Similarly, in Arabidopsis thaliana the greater population of active CRYs was documented under the lower temperatures compared to the moderate ones [59]. The active state of UVR8 photoreceptors seems to be also affected by the temperature. It was documented that the proportion of inactive UVR8 dimers from the total UVR8 population is reduced under lower temperatures [60], which suggests increased monomerization. While above 18 °C the observed changes in the monomer/dimer equilibrium were rather negligible. In addition, the number of active UVR8 monomers can be mediated by RUP (REPRESSOR OF UV-B PHOTOMORPHOGENESIS1 and REPRESSOR OF UV-B PHOTOMORPHOGENESIS2) proteins [60]. ZTL inactivation seems to be progressing with increased temperatures, but it is assumed that ZTL proteins are involved in the heat stress tolerance, which has been documented directly with the ZTL-deficient and ZTL-overexpressing mutant lines of Arabidopsis thaliana [61]. Therefore, it seems that besides their primary photosensory role, plant photoreceptors also have a shared thermosensory role, which is based on the regulation of the active state by the temperature. Lower temperature increases the active state lifetime, whereas the higher temperature decreases it. Although the increasing number of publications supports such an assumption, direct experiments to confirm or refute such a hypothesis are needed. In addition to mentioned interplay of photoreceptors in thermosensing plants possess many other thermosensing mechanisms [62], however these are out of the scope of this review.
5. COP1/SPA Complex
The vast majority of the activated photoreceptors at some point interact with the COP1/SPA (Constitutive photomorphogenesis protein 1/ Protein SPA1-RELATED) protein complex (Figure 2.), which consists of the COP1 protein (which gene has a single copy in Arabidopsis thaliana) and one out of four SPA proteins. It was demonstrated that COP1/SPA is a component of the larger complex together with the CUL4 (Cullin-4), DDB1 (DNA damage-binding protein 1), and RBX (RING-box protein) [63].
In Arabidopsis thaliana this enzymatic complex can induce the degradation of several dozens of different proteins including TFs (BIT1, HY5, HYH, PIF1, PIF5, PIF8, PIL1) and photoreceptors (CRYs, PHYs, [63] involved in diverse processes (photomorphogenesis, light perception, anthocyanin biosynthesis, GA signaling, etc.)). In general, in the darkness, different substrates (mostly TFs) are ubiquitinated, resulting in the suppression of the target genes, but once the light activates the photoreceptors, COP1/SPA complex activity is suppressed (alternative substrates can be cleaved) resulting in the increased population of TFs and therefore activation of the downstream genes. Although, in general, the COP1/SPA complex shows rather an inhibitory role (induces the degradation of its substrates), once interacting with the activated UVR8 it leads to the stabilization of the HY5 transcription factor, or when associated with the PHYs, it can promote the PIFs (transcription repressors) degradation as well.
6. Transcription Factors Involved in PheCs Biosynthesis Control
PheCs biosynthetic pathway forms an important branch of higher plant secondary metabolism. PheCs content is effectively regulated by several environmental cues and their co-action (such as PAR, UV, LT, low N, high CO2, etc.) and they are often studied for their possible contribution to stress-related responses and enhancement of plant tolerance. Below, we intend to describe in detail how the transcriptional control of genes is directly involved in the production of such universal/multipurpose substances. TFs responsible for the coordination of the gene expression contain one of the following motifs: bHLH, bZIP, HTH, or WRKY, and are listed in Table 1.
6.1. Proteins Containing Basic Helix-Loop-Helix Motifs (So-Called bHLH)
This diverse group of proteins contains (as suggested by the acronym bHLH) a basic Helix Loop Helix motif and comprises 26 protein subfamilies [64], and all of them are responsible for direct interaction with the DNA and transcription control. These proteins are present in almost all plant species and in some of them, more than 100 different bHLH proteins were documented. The estimated origin of this group of proteins dates back 440 million years ago to the first land plants [64]. They are involved in the various processes under both optimum [65] and stress conditions [66].
An important protein containing a bHLH structural motif is TT8 (TRANSPARENT TESTA 8; also known as AtbHLH42, bHLH42, or Transcription factor EN32) [11]. According to the literature, this protein is 518 amino acids long transcription activator, when associated with MYB75 (PAP1) or MYB90 (PAP2) [11]. In addition, this protein controls the expression of the DFR (DIHYDROFLAVONOL 4-REDUCTASE; Table 1) and BAN (BANYULS) genes [67] in Arabidopsis thaliana siliques. DFR gene codes for the DFR protein which catalyzes the formation of leucopelargonidin from dihydrokaempferol or the formation of leucocyanidin from dihydroquercetin [67]. BANYULS gene codes for a DFR-like protein [68] which is a negative regulator of flavonoid biosynthesis. Plants with knocked-out BAN displayed a higher accumulation of anthocyanins in the seed coat. In addition, it was even proposed that this gene codes for leucoanthocyanin reductase (LAR), but no direct evidence was reported. On the contrary, it was documented that BAN genes of Medicago truncatula and Arabidopsis thaliana encode for anthocyanidin reductase catalyzing the formation of 2,3-cis-flavan-3-ols from anthocyanins and therefore the reduction of coloration [69].
6.2. Basic ZIPper Containing Proteins (bZIP)
The second group of the TFs related to the PheCs production are basic (leucine) zipper (bZIP) containing proteins (Table 1). Common ancestor (the divergence point varies depending on the plant species, for monocots and eudicots are estimated 140-150 million years ago) probably had four bZIP genes, and those proteins were probably involved in the oxidative stress responses and also in the responses to light [70]. This assumption support also current literature - where bZIP family members are referred to as key players in the transcription control of the genes triggered in the response to light and photomorphogenesis. Moreover, this diverse group of TFs is involved also in responses to other abiotic stresses [71], as well as in the responses to pathogens [72].
The most important member of this protein family is AtZIP56 (Arabidopsis thaliana basic leucine zipper 56) also known as HY5 (ELONGATED HYPOCOTYL 5). This TF interacts directly with the G-box motif (-GCCACGTGC/GC-; but also interactions with E-box, GATA-box, Z-box, C-box, and other hybrid boxes are documented [73]) of the DNA including promoters related to the PheCs production. Due to the interaction with the COP/SPA protein complex which is involved in the signaling of the environmental cues (both light intensity and spectral quality, as well as temperature), HY5 is designated as an "integrator of light and temperature signals" [74]. In Arabidopsis thaliana, HY5 is 168 amino acids long protein containing disordered, bZIP, leucine zipper, basic motif, and interaction site for COP1 (-ESDEEIRRVPEF-; [11]). In the dark, HY5 is ubiquitinated after the interaction with the COP1 and degraded at the 26S proteasome complex [75]. On the contrary, in the light COP1 is inhibited, and therefore the proteins HY5/HYH/CAM7 (HY5 - ELONGATED HYPOCOTYL 5, HYH - HY5 HOMOLOG, CAM7 - Calmodulin-7) can interact with the HY5 promoter, and induce HY5s transcription. This results in temporary accumulation of HY5 proteins which can trigger the expression of the genes with the G box [75] containing promoters, including those related to the PheCs production.
The second and equally important member of the bZIP family is HYH (HY5 HOMOLOG). In Arabidopsis thaliana it is 149 amino acids long protein with the disordered, bZIP, basic motif, leucine zipper regions, and interaction site for COP1 [11]. HY5 and HYH share an identity of 49% (73 out of 149 amino acid residues), and in the DNA binding region the homology in the amino acid sequence steeply increases to 87,5 % (21 out of 24 amino acid residues) [76]. Similarly to HY5, also HYH is involved in the regulation of the expression of genes related to the biosynthesis of PheCs. It was documented that HYH directly interacts with the COP1´s WD-40 domain and the main consequence is the degradation of HYH protein. In addition, both proteins (HY5 and HYH) are involved in the regulation of the transcription in the same pathway - biosynthesis of anthocyanins under the low temperature in the Arabidopsis thaliana [77].
6.3. Transcription Factors Containing Helix-Turn-Helix Motifs (HTH TFs)
The third group of the TFs is helix-turn-helix motif-containing proteins (HTHs). To this diverse group of proteins belong for example MYB proteins (MYBs; the acronym is derived from Avian myeloblastosis virus, where the first protein containing such structure was reported). MYBs have a conserved DNA-binding domain that consists of amino acid repeats (R meaning one repeat in the structure descriptions below) which can be imperfect and are approximately 50 amino acids long. Each of the repeat forms α-helices, but the most important are the second and third helices of each repeat - those directly recognize (the third one), interact (the second one) with the DNA, and form HLH structure [78]. In Arabidopsis thaliana 197 genes coding for MYB TFs were identified in all five chromosomes which vary in the number of the repeats and therefore form 4 subfamilies [79]: MYB-R1R2R3, MYB-R2R3, MYB-related genes, and atypical MYB genes.
TT2 (Protein TRANSPARENT TESTA2, also known as AtMYB123) is a 258 amino acids long transcription factor. When associated with bHLH2/EGL3/MYC146 (basic Helix Loop Helix protein 2, also known as Protein ENHANCER OF GLABRA 3, or Transcription factor EN 30), bHLH12/EN52/MYC1 (basic Helix Loop Helix protein 12, also known as Transcription factor EN 58, or Myelocytomatosis protein 1), or bHLH42/tt8 (basic Helix Loop Helix protein 42, also known as TRANSPARENT TESTA8) work as transcription activator involved in the control of late anthocyanin genes (Table 1; [80]).
In Arabidopsis thaliana, PAP1 (Probable plastid-lipid-associated protein 1, also known as FBN1a, FIB1a, PGL35) is 318 amino acids long protein involved in the transcription control (genes controlled by the PAP1 are mentioned below). Expression of this protein stimulates the jasmonic acid biosynthesis and it was documented that increased accumulation of PAP1 is linked with the increased tolerance of the PSII against photoinhibition in Arabidopsis [81]. On top of that, the PAP1 protein is a key player in the PheCs regulation for example in Arabidopsis thaliana [82], Salvia miltiorrhiza [83], Brassica napus [84], and Nicotiana tabacum [85].
PAP2 (also known as FBN1b, FIB1b) is 310 amino-acid residues long transcription factor in Arabidopsis thaliana. Similarly to PAP1, PAP2 is involved in stress responses and its expression also stimulates jasmonic acid biosynthesis [86]. In addition, PAP2 in Arabidopsis thaliana is involved in the protection of PS II against photooxidative stress (induced by high light in combination with cold). According to the Plant Transcription Factor Database (Table 1), PAP2 stimulates the expression of TTG2 in Arabidopsis thaliana. Therefore, both proteins (PAP1 and PAP2) are connected with growth regulation, PSII protection, but also with anthocyanin accumulation under stress conditions [86].
According to the current knowledge, also other members of the MYB family are equally important. Such example is the trio MYB11, MYB12, and MYB111, which are involved in the control of genes involved in PheCs biosynthesis. All three proteins have the same type of the MYB domain - R2R3, have a similar length (MYB11 – 343 amino acids, MYB12 – 371 amino acids, MYB111 – 342 amino acids), and are designated as flavonol-specific TFs, because all of them positively regulate the transcription of the CHS, CHI, F3H, and FLS1 genes in Arabidopsis thaliana

Table 1.
Summary of selected plant transcription factors of Arabidopsis thaliana related to the control of the genes involved in PheCs biosynthesis (data were extracted from the PlantTFDB database [87]; question marks represent no available data in the source database dealing with the activated or repressed target genes). Dash represents none detected repressed genes, while question marks no available data in the database.
Table 1.
Summary of selected plant transcription factors of Arabidopsis thaliana related to the control of the genes involved in PheCs biosynthesis (data were extracted from the PlantTFDB database [87]; question marks represent no available data in the source database dealing with the activated or repressed target genes). Dash represents none detected repressed genes, while question marks no available data in the database.
| TF | TT8 | HY5 | HYH_1 | HYH_2 | HYH_3 | HYH_4 | TT2 | PAP1 | MYB11 | MYB12 | MYB111 | WRKY23 | WRKY36 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Motif | bHLH | bZIP | bZIP | bZIP | bZIP | bZIP | HTH | HTH | HTH | HTH | HTH | WRKY | WRKY |
| ID | AT4G09820 | AT5G11260 | AT3G17609.1 | AT3G17609.2 | AT3G17609.3 | AT3G17609.4 | AT5G35550.1 | AT1G56650.1 | AT3G62610.1 | AT5G49330.1 | AT5G49330.1 | AT2G47260.1 | AT1G69810.1 |
| Target genes activated | BAN, TT8, MYBL2, DFR, GL2, MES6, MES4, TTG2 | RBCS1A, ELIP1, PHR1, LZF1, IAA7, CHS, CAB2, ABI5, IAA14, DFR, CAB1, UGT84A1, PSBD, NIA2, ELF4, LDOX, HEMA1, MYB12, HB-8 | PHR1, NIA2, ELIP1, PEX11B, CHS | PHR1, NIA2, ELIP1, PEX11B, CHS | PHR1, NIA2, ELIP1, PEX11B, CHS | PHR1, NIA2, ELIP1, PEX11B, CHS | ANS, TT8, TT2, TTG2, GL2, DFR, BAN | CHS, CHI, DFR, MYB3, TT8, UF3GT, PAP2, A5GT, UGT78D2, 5MAT, MBD2.2, GST | CHS, CHI, F3H, FLS1 | CHS, CHI, F3H, FLS1 | CHS, CHI, F3H, FLS1 | ??? | ??? |
| Target genes repressed | - | NPF6.3, FHY1, FHL | NPF6.3 | NPF6.3 | NPF6.3 | NPF6.3 | - | scpl10 | - | - | - | ??? | ??? |
6.4. WRKY Proteins
This group of TFs is also involved in very diverse processes related to various types of abiotic stresses [88]. The characteristic of those TFs is that all members of the WRKY family have -WRKYGQ- repetitions near the N-terminus and can directly interact with W-box ((C/T) TGAC (T/C)) containing promoters [88]. In Arabidopsis thaliana, there were identified 74 genes coding for the WRKY TFs [88]. W-box motif in the promoter region is present mainly in genes involved in the lignin [89], flavonols, or tannins [90] biosynthesis. WRKY TFs therefore contribute to the control of the secondary metabolism. For example, in the Arabidopsis thaliana, WRKY23 probably controls the expression of TT4, TT5, TT6, and TT7 genes [91] (TRANSPARENT TESTA proteins 4-7; genes coding them are also known as CHS, Chalcone-flavanone isomerase, Naringenin,2-oxoglutarate 3-dioxygenase, and Flavonoid 3'-monooxygenase) [11] in the root tip. WRKYGQ repeat tract is responsible for the interaction with the DNA, which directly interacts with the W-box sequence in the major groove as was documented in Solanum lycopersicum (WRKY3 and WRKY4) [92].
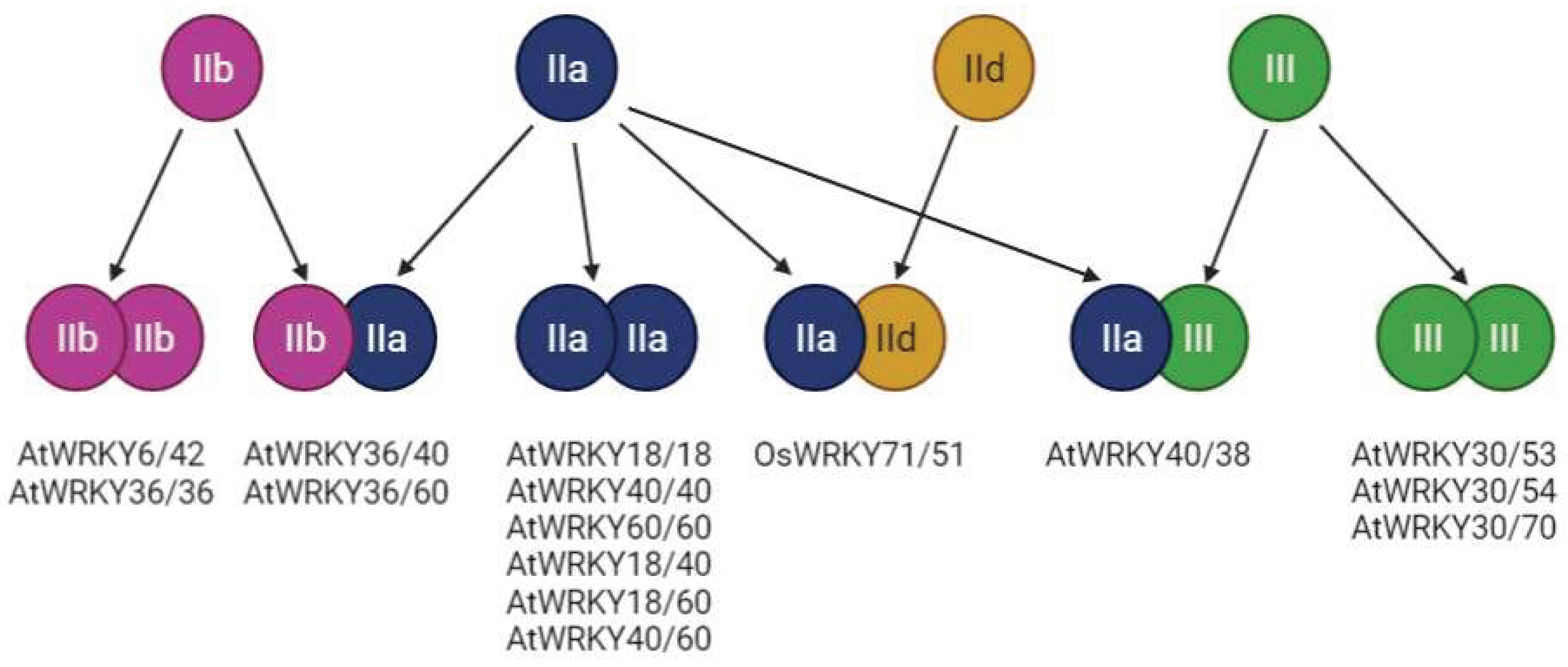
Several subclasses of the WRKY proteins were described based on their structure - I, II, and III. Group I WRKY proteins have two WRKY domains, group II proteins have one WRKY domain and one zinc finger (Cys2-His2) motif, and group three have one WRKY domain and one zinc finger (but different from the group II - Cys2-His/Cys Cys2-His2 zinc-finger motif) [93]. Besides the numeric description of the domain architecture, also other higher-order structure features can be indicated by the small letters a-e. The wide structural diversity of the WRKY proteins therefore enables them to interact with each other and further increase their regulatory potential (Figure 4). For example, in the Arabidopsis thaliana, members of the IIb group can interact with each other (AtWRKY6/ AtWRKY42), but also with the IIa group (AtWRKY36/ AtWRKY40). Similarly, the WRKY proteins belonging to the IIa group can both interact with each other or with the IIb group members (AtWRKY36/ AtWRKY60). Lastly, the members of group III can interact with each other (AtWRKY30/ AtWRKY53), but also with the IIa group members [93] (AtWRKY40/ AtWRKY38). Quite interesting is the limited regulatory potential of the IId members, whose exclusive interaction with the IIa group (OsWRKY71/OsWRKY51) was documented in Oryza sativa (Figure 4). In addition to this (WRKY-WRKY protein interactions), the interactions with other proteins were documented. For example with the VQ motif-containing proteins (HAIKU1, MAPK4, MKS1, or even with the chromatin remodelling proteins (HDA19) [94].
An interesting member of the WRKY TFs family involved in light-related signaling control is WRKY36, which is the transcriptional inhibitor of the HY5 gene coding for the HY5 protein. It was documented that UVR8 can directly interact with the WRKY36 in Arabidopsis thaliana and also that UVB exposure increases the amount of UVR8/WRKY36 dimers [95]. In combination with the fact that UVB leads to increased UVR8 accumulation in the nucleus, authors proposed [95] that active UVR8 monomers (formed after the UVB exposure) migrate to the nucleus and pull the WRKY36 transcription inhibitor from the promoter of the HY5 gene, and thus lead to the increased transcription of the central light and temperature integrator (HY5) which controls both other TFs (e. g. MYB12) as well as PheCs-related genes.
6.5. WD-40
WD-40 proteins are characterized by a WD-40 domain containing multiple repeats [96], where each repeat contains 44-60 amino acids [97]. Each unit contains dipeptides (either glycine-histidine (GH; in the proximity of the N-termini) or tryptophan-asparagine (WD; in the proximity of the C-termini)) and can be folded into the four-stranded antiparallel beta-sheet, which is one segment of the beta-propeller structure (similar structure to the UVR8 photoreceptor in the monomeric state; [97]). Each WD-40 protein has at least four such segments [96].
In Arabidopsis thaliana, 269 WD-40 proteins were identified and can be distinguished in approximately 113 subfamilies which have significant homology with their human or fruit fly counterparts suggesting their importance [97].
For example, the COP1 (CONSTITUTIVE PHOTOMORPHOGENIC 1) protein is a member of the WD-40 family having seven WD repeats in Arabidopsis [96]. This particular protein is involved in the light signaling and control of photomorphogenesis via the direct repression of the transcription activators [96]. Recently, it was also documented that it stabilizes the transcription repressors (PIFs, EIN3, or EIL1; [98]).
6.6. MYB-bHLH-WD-40 complex
For a late group of PheCs-related genes, transcription is controlled at multiple levels. Besides the direct interaction of the promoters with the various TFs (for example TT2), the MBW complex (consisting of the MYB-bHLH-WD-40 proteins) also contributes to the coordination of the gene expression. In addition, Arabidopsis thaliana has two different MBW complexes - the first one for the control of the anthocyanin (PAPs-EGL3/GL3/TT8-TTG1) (Lloyd et al., 2017) and the second one for the control of proanthocyanidin (TT2/TT8/TTG1) biosynthetic genes [100] in the plant body (anthocyanins) and seeds (proanthocyanidins) - which further highlight the importance of MBW complex. In this complex (MBW) the PAP1 and TT8 are responsible for the DNA binding, and consequent regulation of transcriptional activity, while the TTG1 is necessary for the MBW complex activity; and it is assumed that also prevents the promoter interaction with the transcriptional inhibitors [74].
MBW complex itself positively regulates transcription of the PheCs-related genes, but interactions with other proteins are documented and result in the alteration (usually decrease) of the late genes expression. For example, MYBL2 (MYB-like 2 protein) probably competes with the R2R3 MYBs at the MBW complex and interacts with the bHLH proteins which leads to decreased anthocyanin production [101]. Similarly, the SPL9 (SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 9) protein directly competes with the TT8 protein for interaction in the anthocyanin-related MBW complex. If the SPL9 is incorporated, the reduced transcription of the target genes related to the anthocyanin biosynthesis is observed (DFR, BAN, etc.) in Arabidopsis thaliana [101], and therefore (FLS and DFR compete for substrates) the flavonol biosynthesis [101] is increased. In addition, expression of the SPL9 is controlled at the posttranscriptional level by the miR156 and miR157 via PTGS [101].
7. Future Perspectives
Current studies dealing with light perception are mostly focused on the direct photosensory mechanism of light sensing and only a few studies dealt with the following protein-protein interactions. Our knowledge about the interactions of the active phototropins and zeitlupes is very limited and even less is known about the following protein-protein interactions. Therefore, it would be very useful to shed light on these interactomes and also to inspect the crosstalks between them.
In addition, very helpful would be an overview (let´s say a map) of the important genes (including their promoters with highlighted specific sequence motifs) related to the stress with their master regulators (light-activated TFs) - such overview to the best of our knowledge is missing and searching of the regulatory motifs in the literature hardens the effort to understand how are these target genes regulated.
Similar issues one has to face when dealing with the aliases, synonyms, or structural motifs of the plant transcription factors. A uniform, widely used nomenclature is missing and further complicates orientation in the light-induced coordination of the target genes.
In terms of the experimental work, systematic studies focused on the integration of the cues from different photoreceptors are missing. Specifically - how is the crosstalk between different blue light receptors ensured? Are these signals unified? If so, at which point? And how are signals related to the different spectral qualities of incident light integrated? Can they amplify each other? Can "opposite" signals (for example from cryptochromes and phytochromes) mute the response? These questions are crucial to address because although a big progress has been made, our understanding remains limited.
8. Conclusions
In this work, we summarized the current knowledge related to direct light sensing, which is ensured by the photoreceptors, and related signaling via the protein-protein interactions leading to the activation of transcription factors ensuring the control of the target genes expression. Special attention was paid to the transcription control of the genes related to PheCs production, which seem to be accumulated more in responses to environmental changes - especially those related to light.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, A.V.; resources, A.V.; writing—original draft preparation, A.V.; writing—review and editing, J.Č., R. P., J.N., V.Š.; visualization, A.V.; supervision, J.Č., V.Š..; funding acquisition, V.Š., R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Czech Science Foundation (GACR 21-18532S to V.Š., A.V., R.P., and J.N.), the European Union under the LERCO project (number CZ.10.03.01./00/22_003/0000003) via the Operational Programme Just Transition, the University of Ostrava (SGS/PřF/2024 to A.V., R.P., J.N.), and the Moravian-Silesian Region (RRC/10/2021 to R.P.). In addition, V.Š. was partially financially supported by the project of the Ministry of Education, Youth and Sports of the Czech Republic (AdAgriF; CZ.02.01.01/00/22_008/0004635).
Acknowledgments
We would like to thank ChatGPT (https://openai.com/) for rephrasing problematic sections in this article. All schemes accompanying this publication were created using BioRender software (https://biorender.com/). Our gratitude also deserves Martin Bartas, Ph.D. who performed a thorough review of this text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- J.J. Casal, Environmental cues affecting development, Current Opinion in Plant Biology 5 (2002) 37–42. [CrossRef]
- M. Landi, M. Zivcak, O. Sytar, M. Brestic, S.I. Allakhverdiev, Plasticity of photosynthetic processes and the accumulation of secondary metabolites in plants in response to monochromatic light environments: A review, Biochimica et Biophysica Acta (BBA) - Bioenergetics 1861 (2020) 148131. [CrossRef]
- G. Kudo, T.Y. Ida, T. Tani, Linkages Between Phenology, Pollination, Photosynthesis, and Reproduction in Deciduous Forest Understory Plants, Ecology 89 (2008) 321–331. [CrossRef]
- J. Saile, T. Wießner-Kroh, K. Erbstein, D.M. Obermüller, A. Pfeiffer, D. Janocha, J. Lohmann, A. Wachter, SNF1-RELATED KINASE 1 and TARGET OF RAPAMYCIN control light-responsive splicing events and developmental characteristics in etiolated Arabidopsis seedlings, The Plant Cell 35 (2023) 3413–3428. [CrossRef]
- K. Apel, H. Hirt, Reactive oxygen species: Metabolism, oxidative stress, and signal transduction, Annu. Rev. Plant Biol. 55 (2004) 373–399. [CrossRef]
- M. El-Esawi, L.-D. Arthaut, N. Jourdan, A. d’Harlingue, J. Link, C.F. Martino, M. Ahmad, Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome, Scientific Reports 7 (2017) 13875. [CrossRef]
- E. Horváth, T. Janda, G. Szalai, E. Páldi, In vitro salicylic acid inhibition of catalase activity in maize: differences between the isozymes and a possible role in the induction of chilling tolerance, Plant Science 163 (2002) 1129–1135. [CrossRef]
- S. Riegler, L. Servi, M.R. Scarpin, M.A. Godoy Herz, M.G. Kubaczka, P. Venhuizen, C. Meyer, J.O. Brunkard, M. Kalyna, A. Barta, E. Petrillo, Light regulates alternative splicing outcomes via the TOR kinase pathway, Cell Reports 36 (2021) 109676. [CrossRef]
- K. McCready, V. Spencer, M. Kim, The Importance of TOR Kinase in Plant Development, Frontiers in Plant Science 11 (2020). https://www.frontiersin.org/articles/10.3389/fpls.2020.00016 (accessed January 12, 2024).
- M. Jamsheer K, P. Awasthi, A. Laxmi, The social network of target of rapamycin complex 1 in plants, Journal of Experimental Botany 73 (2022) 7026–7040. [CrossRef]
- The UniProt Consortium, UniProt: a worldwide hub of protein knowledge, Nucleic Acids Research 47 (2019) D506–D515. [CrossRef]
- AT4G29130(HXK1), (n.d.). https://www.arabidopsis.org/servlets/TairObject?name=AT4G29130&type=locus (accessed January 12, 2024).
- R. Wu, X. Lin, J. He, A. Min, L. Pang, Y. Wang, Y. Lin, Y. Zhang, W. He, M. Li, Y. Zhang, Y. Luo, X. Wang, H. Tang, Q. Chen, Hexokinase1: A glucose sensor involved in drought stress response and sugar metabolism depending on its kinase activity in strawberry, Frontiers in Plant Science 14 (2023). https://www.frontiersin.org/articles/10.3389/fpls.2023.1069830 (accessed January 12, 2024).
- B. Peixoto, E. Baena-González, Management of plant central metabolism by SnRK1 protein kinases, Journal of Experimental Botany 73 (2022) 7068–7082. [CrossRef]
- B. Wang, X. Zhao, Y. Zhao, J. Shanklin, Q. Zhao, C.-J. Liu, Arabidopsis SnRK1 negatively regulates phenylpropanoid metabolism via Kelch domain-containing F-box proteins, New Phytologist 229 (2021) 3345–3359. [CrossRef]
- S. Hulsmans, M. Rodriguez, B.D. Coninck, F. Rolland, The SnRK1 Energy Sensor in Plant Biotic Interactions, Trends in Plant Science 21 (2016) 648–661. [CrossRef]
- J.B. Rossel, I.W. Wilson, B.J. Pogson, Global Changes in Gene Expression in Response to High Light in Arabidopsis, Plant Physiology 130 (2002) 1109–1120. [CrossRef]
- S.I. Zandalinas, S. Sengupta, D. Burks, R.K. Azad, R. Mittler, Identification and characterization of a core set of ROS wave-associated transcripts involved in the systemic acquired acclimation response of Arabidopsis to excess light, The Plant Journal 98 (2019) 126–141. [CrossRef]
- B. Moore, L. Zhou, F. Rolland, Q. Hall, W.-H. Cheng, Y.-X. Liu, I. Hwang, T. Jones, J. Sheen, Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal Signaling, Science 300 (2003) 332–336. [CrossRef]
- O. Avidan, T.A. Moraes, V. Mengin, R. Feil, F. Rolland, M. Stitt, J.E. Lunn, In vivo protein kinase activity of SnRK1 fluctuates in Arabidopsis rosettes during light-dark cycles, Plant Physiology 192 (2023) 387–408. [CrossRef]
- S. Shokrian Hajibehzad, S.S. Silva, N. Peeters, E. Stouten, G. Buijs, S. Smeekens, M. Proveniers, Arabidopsis thaliana rosette habit is controlled by combined light and energy signaling converging on transcriptional control of the TALE homeobox gene ATH1, New Phytologist 239 (2023) 1051–1067. [CrossRef]
- C. Orth, N. Niemann, L. Hennig, L.-O. Essen, A. Batschauer, Hyperactivity of the Arabidopsis cryptochrome (cry1) L407F mutant is caused by a structural alteration close to the cry1 ATP-binding site, Journal of Biological Chemistry 292 (2017) 12906–12920. [CrossRef]
- L. Lopez, C. Fasano, G. Perrella, P. Facella, Cryptochromes and the Circadian Clock: The Story of a Very Complex Relationship in a Spinning World, Genes 12 (2021) 672. [CrossRef]
- C. Lin, T. Todo, The cryptochromes, Genome Biology 6 (2005) 220. [CrossRef]
- J.M. Barrero, A.B. Downie, Q. Xu, F. Gubler, A Role for Barley CRYPTOCHROME1 in Light Regulation of Grain Dormancy and Germination, The Plant Cell 26 (2014) 1094–1104. [CrossRef]
- N. Tissot, R. Ulm, Cryptochrome-mediated blue-light signalling modulates UVR8 photoreceptor activity and contributes to UV-B tolerance in Arabidopsis, Nature Communications 11 (2020) 1323. [CrossRef]
- L. Ma, Z. Guan, Q. Wang, X. Yan, J. Wang, Z. Wang, J. Cao, D. Zhang, X. Gong, P. Yin, Structural insights into the photoactivation of Arabidopsis CRY2, Nature Plants 6 (2020) 1432–1438. [CrossRef]
- M. Palayam, J. Ganapathy, A.M. Guercio, L. Tal, S.L. Deck, N. Shabek, Structural insights into photoactivation of plant Cryptochrome-2, Communications Biology 4 (2021) 1–11. [CrossRef]
- K. Shao, X. Zhang, X. Li, Y. Hao, X. Huang, M. Ma, M. Zhang, F. Yu, H. Liu, P. Zhang, The oligomeric structures of plant cryptochromes, Nature Structural & Molecular Biology 27 (2020) 480–488. [CrossRef]
- J.M. Christie, Phototropin Blue-Light Receptors, Annual Review of Plant Biology 58 (2007) 21–45. [CrossRef]
- J.M. Christie, T.E. Swartz, R.A. Bogomolni, W.R. Briggs, Phototropin LOV domains exhibit distinct roles in regulating photoreceptor function, The Plant Journal 32 (2002) 205–219. [CrossRef]
- S. Inoue, A. Takemiya, K. Shimazaki, Phototropin signaling and stomatal opening as a model case, Current Opinion in Plant Biology 13 (2010) 587–593. [CrossRef]
- Y. Nakasone, M. Ohshima, K. Okajima, S. Tokutomi, M. Terazima, Photoreaction Dynamics of Full-Length Phototropin from Chlamydomonas reinhardtii, The Journal of Physical Chemistry B 123 (2019) 10939–10950. [CrossRef]
- A. Motchoulski, E. Liscum, Arabidopsis NPH3: A NPH1 Photoreceptor-Interacting Protein Essential for Phototropism, Science 286 (1999) 961–964. [CrossRef]
- S. Inada, M. Ohgishi, T. Mayama, K. Okada, T. Sakai, RPT2 Is a Signal Transducer Involved in Phototropic Response and Stomatal Opening by Association with Phototropin 1 in Arabidopsis thaliana, Plant Cell 16 (2004) 887–896. [CrossRef]
- T. Sakai, T. Wada, S. Ishiguro, K. Okada, RPT2: A Signal Transducer of the Phototropic Response in Arabidopsis, The Plant Cell 12 (2000) 225–236. [CrossRef]
- S.K. Gupta, M. Sharma, F. Deeba, V. Pandey, Plant Response, in: UV-B Radiation, John Wiley & Sons, Ltd, 2017: pp. 217–258. [CrossRef]
- S.-G. Kong, K. Okajima, Diverse photoreceptors and light responses in plants, Journal of Plant Research 129 (2016) 111–114. [CrossRef]
- M. Legris, Y.Ç. Ince, C. Fankhauser, Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants, Nature Communications 10 (2019) 5219. [CrossRef]
- Paik, E. Huq, Plant photoreceptors: Multi-functional sensory proteins and their signaling networks, Seminars in Cell & Developmental Biology 92 (2019) 114–121. [CrossRef]
- A. Pudasaini, B.D. Zoltowski, Zeitlupe Senses Blue-Light Fluence To Mediate Circadian Timing in Arabidopsis thaliana, Biochemistry 52 (2013) 7150–7158. [CrossRef]
- A. Pudasaini, J.S. Shim, Y.H. Song, H. Shi, T. Kiba, D.E. Somers, T. Imaizumi, B.D. Zoltowski, Kinetics of the LOV domain of ZEITLUPE determine its circadian function in Arabidopsis, ELife 6 (2017) e21646. [CrossRef]
- S. Ito, Y.H. Song, T. Imaizumi, LOV Domain-Containing F-Box Proteins: Light-Dependent Protein Degradation Modules in Arabidopsis, Molecular Plant 5 (2012) 573–582. [CrossRef]
- J.M. Christie, A.S. Arvai, K.J. Baxter, M. Heilmann, A.J. Pratt, A. O’Hara, S.M. Kelly, M. Hothorn, B.O. Smith, K. Hitomi, G.I. Jenkins, E.D. Getzoff, Plant UVR8 Photoreceptor Senses UV-B by Tryptophan-Mediated Disruption of Cross-Dimer Salt Bridges, Science 335 (2012) 1492–1496. [CrossRef]
- T. Mathes, M. Heilmann, A. Pandit, J. Zhu, J. Ravensbergen, M. Kloz, Y. Fu, B.O. Smith, J.M. Christie, G.I. Jenkins, J.T.M. Kennis, Proton-Coupled Electron Transfer Constitutes the Photoactivation Mechanism of the Plant Photoreceptor UVR8, Journal of the American Chemical Society 137 (2015) 8113–8120. [CrossRef]
- X. Li, H. Ren, M. Kundu, Z. Liu, F.W. Zhong, L. Wang, J. Gao, D. Zhong, A leap in quantum efficiency through light harvesting in photoreceptor UVR8, Nature Communications 11 (2020) 4316. [CrossRef]
- V.E. Tossi, J.J. Regalado, J. Iannicelli, L.E. Laino, H.P. Burrieza, A.S. Escandón, S.I. Pitta-Álvarez, Beyond Arabidopsis: Differential UV-B Response Mediated by UVR8 in Diverse Species, Frontiers in Plant Science 10 (2019). [CrossRef]
- N.C. Rockwell, Y.-S. Su, J.C. Lagarias, PHYTOCHOME STRUCTURE AND SIGNALING MECHANISMS, Annual Review of Plant Biology 57 (2006) 837–858. [CrossRef]
- J. Li, G. Li, H. Wang, X.W. Deng, Phytochrome Signaling Mechanisms, The Arabidopsis Book 2011 (2011). [CrossRef]
- K.A. Franklin, T. Allen, G.C. Whitelam, Phytochrome A is an irradiance-dependent red light sensor, The Plant Journal 50 (2007) 108–117. [CrossRef]
- M. Legris, C. Klose, E.S. Burgie, C.C.R. Rojas, M. Neme, A. Hiltbrunner, P.A. Wigge, E. Schäfer, R.D. Vierstra, J.J. Casal, Phytochrome B integrates light and temperature signals in Arabidopsis, Science 354 (2016) 897–900. [CrossRef]
- Y. Qiu, M. Li, R.J.-A. Kim, C.M. Moore, M. Chen, Daytime temperature is sensed by phytochrome B in Arabidopsis through a transcriptional activator HEMERA, Nature Communications 10 (2019) 140. [CrossRef]
- R. Bianchetti, B. De Luca, L.A. de Haro, D. Rosado, D. Demarco, M. Conte, L. Bermudez, L. Freschi, A.R. Fernie, L.V. Michaelson, R.P. Haslam, M. Rossi, F. Carrari, Phytochrome-Dependent Temperature Perception Modulates Isoprenoid Metabolism, Plant Physiology 183 (2020) 869–882. [CrossRef]
- V.N. Pham, P.K. Kathare, E. Huq, Phytochromes and Phytochrome Interacting Factors, Plant Physiology 176 (2018) 1025–1038. [CrossRef]
- T. Sakamoto, S. Kimura, Plant Temperature Sensors, Sensors 18 (2018) 4365. [CrossRef]
- B.Y.W. Chung, M. Balcerowicz, M. Di Antonio, K.E. Jaeger, F. Geng, K. Franaszek, P. Marriott, I. Brierley, A.E. Firth, P.A. Wigge, An RNA thermoswitch regulates daytime growth in Arabidopsis, Nature Plants 6 (2020) 522–532. [CrossRef]
- S. Hayes, J. Schachtschabel, M. Mishkind, T. Munnik, S.A. Arisz, Hot topic: Thermosensing in plants, Plant, Cell & Environment 44 (2021) 2018–2033. [CrossRef]
- Y. Fujii, H. Tanaka, N. Konno, Y. Ogasawara, N. Hamashima, S. Tamura, S. Hasegawa, Y. Hayasaki, K. Okajima, Y. Kodama, Phototropin perceives temperature based on the lifetime of its photoactivated state, Proceedings of the National Academy of Sciences 114 (2017) 9206–9211. [CrossRef]
- M. Pooam, N. Dixon, M. Hilvert, P. Misko, K. Waters, N. Jourdan, S. Drahy, S. Mills, D. Engle, J. Link, M. Ahmad, Effect of temperature on the Arabidopsis cryptochrome photocycle, Physiologia Plantarum 172 (2021) 1653–1661. [CrossRef]
- K.M.W. Findlay, G.I. Jenkins, Regulation of UVR8 photoreceptor dimer/monomer photo-equilibrium in Arabidopsis plants grown under photoperiodic conditions, Plant, Cell & Environment 39 (2016) 1706–1714. [CrossRef]
- P.A. Salomé, In the Heat of the Moment: ZTL-Mediated Protein Quality Control at High Temperatures, The Plant Cell 29 (2017) 2685–2686. [CrossRef]
- M. Noguchi, Y. Kodama, Temperature Sensing in Plants: On the Dawn of Molecular Thermosensor Research, Plant and Cell Physiology 63 (2022) 737–743. [CrossRef]
- Ponnu, U. Hoecker, Illuminating the COP1/SPA Ubiquitin Ligase: Fresh Insights Into Its Structure and Functions During Plant Photomorphogenesis, Frontiers in Plant Science 12 (2021). [CrossRef]
- N. Pires, L. Dolan, Origin and Diversification of Basic-Helix-Loop-Helix Proteins in Plants, Molecular Biology and Evolution 27 (2010) 862–874. [CrossRef]
- Y. Hao, X. Zong, P. Ren, Y. Qian, A. Fu, Basic Helix-Loop-Helix (bHLH) Transcription Factors Regulate a Wide Range of Functions in Arabidopsis, International Journal of Molecular Sciences 22 (2021) 7152. [CrossRef]
- Y. Qian, T. Zhang, Y. Yu, L. Gou, J. Yang, J. Xu, E. Pi, Regulatory Mechanisms of bHLH Transcription Factors in Plant Adaptive Responses to Various Abiotic Stresses, Frontiers in Plant Science 12 (2021). https://www.frontiersin.org/articles/10.3389/fpls.2021.677611 (accessed October 11, 2022).
- N. Nesi, I. Debeaujon, C. Jond, G. Pelletier, M. Caboche, L. Lepiniec, The TT8 Gene Encodes a Basic Helix-Loop-Helix Domain Protein Required for Expression of DFR and BAN Genes in Arabidopsis Siliques, Plant Cell 12 (2000) 1863–1878.
- M. Devic, J. Guilleminot, I. Debeaujon, N. Bechtold, E. Bensaude, M. Koornneef, G. Pelletier, M. Delseny, The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development, The Plant Journal 19 (1999) 387–398. [CrossRef]
- D.-Y. Xie, S.B. Sharma, N.L. Paiva, D. Ferreira, R.A. Dixon, Role of Anthocyanidin Reductase, Encoded by BANYULS in Plant Flavonoid Biosynthesis, Science 299 (2003) 396–399. [CrossRef]
- L.G.G. Corrêa, D.M. Riaño-Pachón, C.G. Schrago, R.V. dos Santos, B. Mueller-Roeber, M. Vincentz, The Role of bZIP Transcription Factors in Green Plant Evolution: Adaptive Features Emerging from Four Founder Genes, PLOS One 3 (2008) e2944. [CrossRef]
- Y. Yu, Y. Qian, M. Jiang, J. Xu, J. Yang, T. Zhang, L. Gou, E. Pi, Regulation Mechanisms of Plant Basic Leucine Zippers to Various Abiotic Stresses, Frontiers in Plant Science 11 (2020). https://www.frontiersin.org/articles/10.3389/fpls.2020.01258 (accessed October 11, 2022).
- M.S. Alves, S.P. Dadalto, A.B. Gonçalves, G.B. De Souza, V.A. Barros, L.G. Fietto, Plant bZIP Transcription Factors Responsive to Pathogens: A Review, International Journal of Molecular Sciences 14 (2013) 7815–7828. [CrossRef]
- S.N. Gangappa, J.F. Botto, The Multifaceted Roles of HY5 in Plant Growth and Development, Molecular Plant 9 (2016) 1353–1365. [CrossRef]
- N.H. Nguyen, HY5, an integrator of light and temperature signals in the regulation of anthocyanins biosynthesis in Arabidopsis, AIMS Molecular Science 7 (2020) 70–81. [CrossRef]
- D. Xu, Y. Jiang, J. Li, F. Lin, M. Holm, X.W. Deng, BBX21, an Arabidopsis B-box protein, directly activates HY5 and is targeted by COP1 for 26S proteasome-mediated degradation, Proceedings of the National Academy of Sciences 113 (2016) 7655–7660. [CrossRef]
- M. Holm, L.-G. Ma, L.-J. Qu, X.-W. Deng, Two interacting bZIP proteins are direct targets of COP1-mediated control of light-dependent gene expression in Arabidopsis, Genes & Development 16 (2002) 1247–1259. [CrossRef]
- Y. Zhang, S. Zheng, Z. Liu, L. Wang, Y. Bi, Both HY5 and HYH are necessary regulators for low temperature-induced anthocyanin accumulation in Arabidopsis seedlings, Journal of Plant Physiology 168 (2011) 367–374. [CrossRef]
- C. Dubos, R. Stracke, E. Grotewold, B. Weisshaar, C. Martin, L. Lepiniec, MYB transcription factors in Arabidopsis, Trends in Plant Science 15 (2010) 573–581. [CrossRef]
- A. Katiyar, S. Smita, S.K. Lenka, R. Rajwanshi, V. Chinnusamy, K.C. Bansal, Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis, BMC Genomics 13 (2012) 544. [CrossRef]
- N. Nesi, C. Jond, I. Debeaujon, M. Caboche, L. Lepiniec, The Arabidopsis TT2 Gene Encodes an R2R3 MYB Domain Protein That Acts as a Key Determinant for Proanthocyanidin Accumulation in Developing Seed, Plant Cell 13 (2001) 2099–2114. [CrossRef]
- Y. Yang, R. Sulpice, A. Himmelbach, M. Meinhard, A. Christmann, E. Grill, Fibrillin expression is regulated by abscisic acid response regulators and is involved in abscisic acid-mediated photoprotection, Proceedings of the National Academy of Sciences of the United States of America 103 (2006) 6061–6066. [CrossRef]
- M.-Z. Shi, D.-Y. Xie, Features of anthocyanin biosynthesis in pap1-D and wild-type Arabidopsis thaliana plants grown in different light intensity and culture media conditions, Planta 231 (2010) 1385–1400. [CrossRef]
- Y. Zhang, Y.-P. Yan, Z.-Z. Wang, The Arabidopsis PAP1 Transcription Factor Plays an Important Role in the Enrichment of Phenolic Acids in Salvia miltiorrhiza, Journal of Agricultural and Food Chemistry 58 (2010) 12168–12175. [CrossRef]
- X. Li, M.-J. Gao, H.-Y. Pan, D.-J. Cui, M.Y. Gruber, Purple Canola: Arabidopsis PAP1 Increases Antioxidants and Phenolics in Brassica napus Leaves, Journal of Agricultural and Food Chemistry 58 (2010) 1639–1645. [CrossRef]
- T. Mitsunami, M. Nishihara, I. Galis, K.M. Alamgir, Y. Hojo, K. Fujita, N. Sasaki, K. Nemoto, T. Sawasaki, G. Arimura, Overexpression of the PAP1 Transcription Factor Reveals a Complex Regulation of Flavonoid and Phenylpropanoid Metabolism in Nicotiana tabacum Plants Attacked by Spodoptera litura, PLOS ONE 9 (2014) e108849. [CrossRef]
- A. Youssef, Y. Laizet, M.A. Block, E. Maréchal, J.-P. Alcaraz, T.R. Larson, D. Pontier, J. Gaffé, M. Kuntz, Plant lipid-associated fibrillin proteins condition jasmonate production under photosynthetic stress, The Plant Journal 61 (2010) 436–445. [CrossRef]
- J. Jin, F. Tian, D.-C. Yang, Y.-Q. Meng, L. Kong, J. Luo, G. Gao, PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants, Nucleic Acids Research 45 (2017) D1040–D1045. [CrossRef]
- J. Jiang, S. Ma, N. Ye, M. Jiang, J. Cao, J. Zhang, WRKY transcription factors in plant responses to stresses, Journal of Integrative Plant Biology 59 (2017) 86–101. [CrossRef]
- S. Guillaumie, R. Mzid, V. Méchin, C. Léon, I. Hichri, A. Destrac-Irvine, C. Trossat-Magnin, S. Delrot, V. Lauvergeat, The grapevine transcription factor WRKY2 influences the lignin pathway and xylem development in tobacco, Plant Molecular Biology 72 (2010) 215–234. [CrossRef]
- U.J. Phukan, G.S. Jeena, R.K. Shukla, WRKY Transcription Factors: Molecular Regulation and Stress Responses in Plants, Frontiers in Plant Science 7 (2016) 760. [CrossRef]
- W. Grunewald, I. De Smet, D.R. Lewis, C. Löfke, L. Jansen, G. Goeminne, R. Vanden Bossche, M. Karimi, B. De Rybel, B. Vanholme, T. Teichmann, W. Boerjan, M.C.E. Van Montagu, G. Gheysen, G.K. Muday, J. Friml, T. Beeckman, Transcription factor WRKY23 assists auxin distribution patterns during Arabidopsis root development through local control on flavonol biosynthesis, Proceedings of the National Academy of Sciences of the United States of America 109 (2012) 1554–1559. [CrossRef]
- M. Aamir, V.K. Singh, M. Meena, R.S. Upadhyay, V.K. Gupta, S. Singh, Structural and Functional Insights into WRKY3 and WRKY4 Transcription Factors to Unravel the WRKY–DNA (W-Box) Complex Interaction in Tomato (Solanum lycopersicum L.). A Computational Approach, Frontiers in Plant Science 8 (2017) 819. [CrossRef]
- L. Chen, Y. Song, S. Li, L. Zhang, C. Zou, D. Yu, The role of WRKY transcription factors in plant abiotic stresses, Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 1819 (2012) 120–128. [CrossRef]
- Y. Chi, Y. Yang, Y. Zhou, J. Zhou, B. Fan, J.-Q. Yu, Z. Chen, Protein–Protein Interactions in the Regulation of WRKY Transcription Factors, Molecular Plant 6 (2013) 287–300. [CrossRef]
- Y. Yang, T. Liang, L. Zhang, K. Shao, X. Gu, R. Shang, N. Shi, X. Li, P. Zhang, H. Liu, UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis, Nature Plants 4 (2018) 98–107. [CrossRef]
- S. van Nocker, P. Ludwig, The WD-repeat protein superfamily in Arabidopsis: conservation and divergence in structure and function, BMC Genomics 4 (2003) 50. [CrossRef]
- A.K. Mishra, S. Puranik, M. Prasad, Structure and regulatory networks of WD40 protein in plants, Journal of Plant Biochemistry and Biotechnology 21 (2012) 32–39. [CrossRef]
- Y. Pan, H. Shi, Stabilizing the Transcription Factors by E3 Ligase COP1, Trends in Plant Science 22 (2017) 999–1001. [CrossRef]
- S. Laubinger, K. Fittinghoff, U. Hoecker, The SPA Quartet: A Family of WD-Repeat Proteins with a Central Role in Suppression of Photomorphogenesis in Arabidopsis, Plant Cell 16 (2004) 2293–2306. [CrossRef]
- A. Lloyd, A. Brockman, L. Aguirre, A. Campbell, A. Bean, A. Cantero, A. Gonzalez, Advances in the MYB–bHLH–WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-option of an Anthocyanin MYB for Betalain Regulation, Plant and Cell Physiology 58 (2017) 1431–1441. [CrossRef]
- S. Li, Transcriptional control of flavonoid biosynthesis, Plant Signaling & Behavior 9 (2014) e27522. [CrossRef]
Figure 2.
The mechanism of the photoreceptors sensing, signaling, and regeneration - for each plant photoreceptor the basic mechanism of the photo sensing is indicated (it can be either monomerization, dimerization, or conformational changes), together with the light region which induced the active state of photoreceptors, and the signaling pathway where the light signal is further propagated (related literature is cited in the text accompanying this scheme).
Figure 2.
The mechanism of the photoreceptors sensing, signaling, and regeneration - for each plant photoreceptor the basic mechanism of the photo sensing is indicated (it can be either monomerization, dimerization, or conformational changes), together with the light region which induced the active state of photoreceptors, and the signaling pathway where the light signal is further propagated (related literature is cited in the text accompanying this scheme).
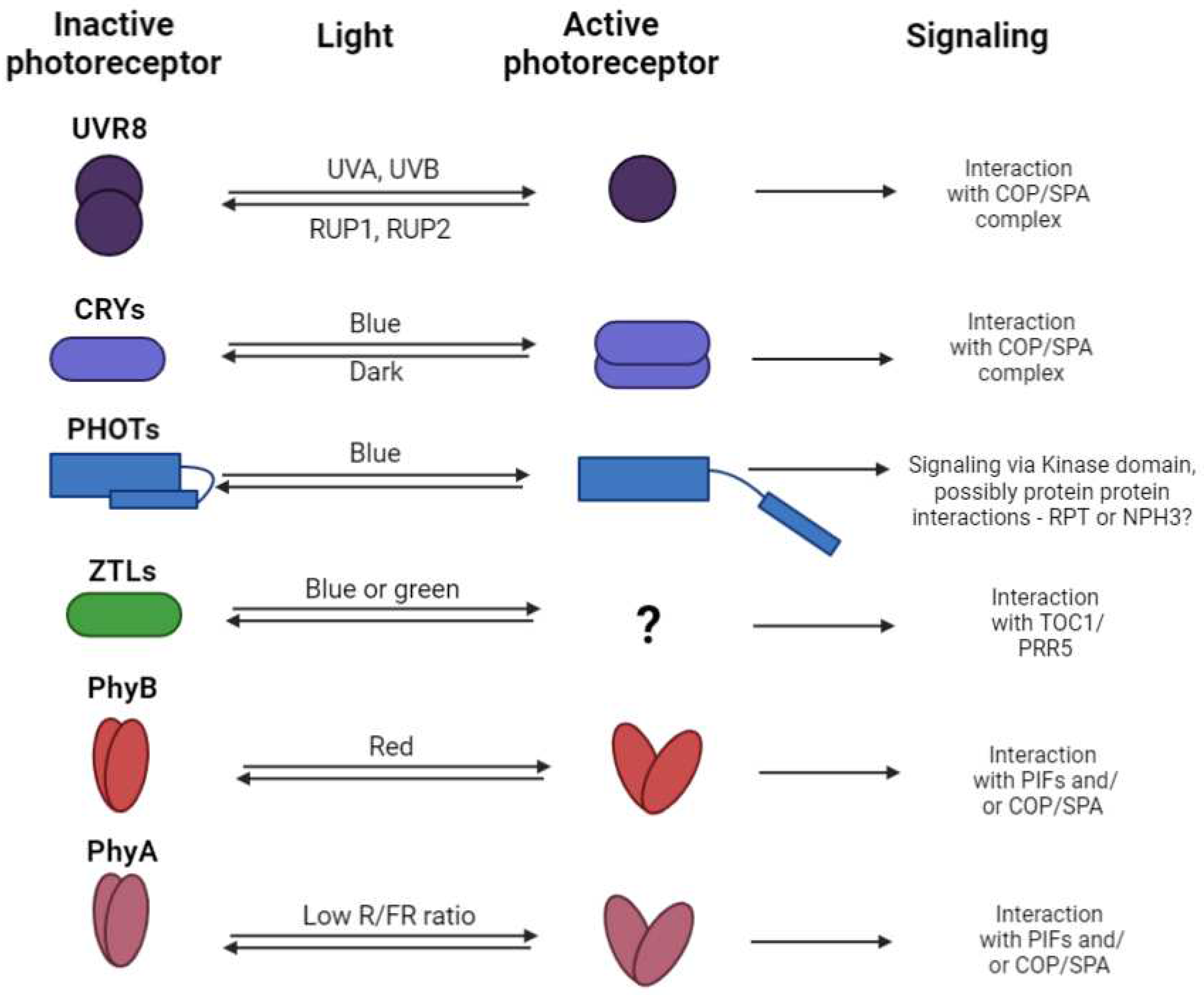
Figure 3.
Mechanism of the transcription control of target genes by PhyB interaction with PIFs (scheme was adopted from [55]).
Figure 3.
Mechanism of the transcription control of target genes by PhyB interaction with PIFs (scheme was adopted from [55]).
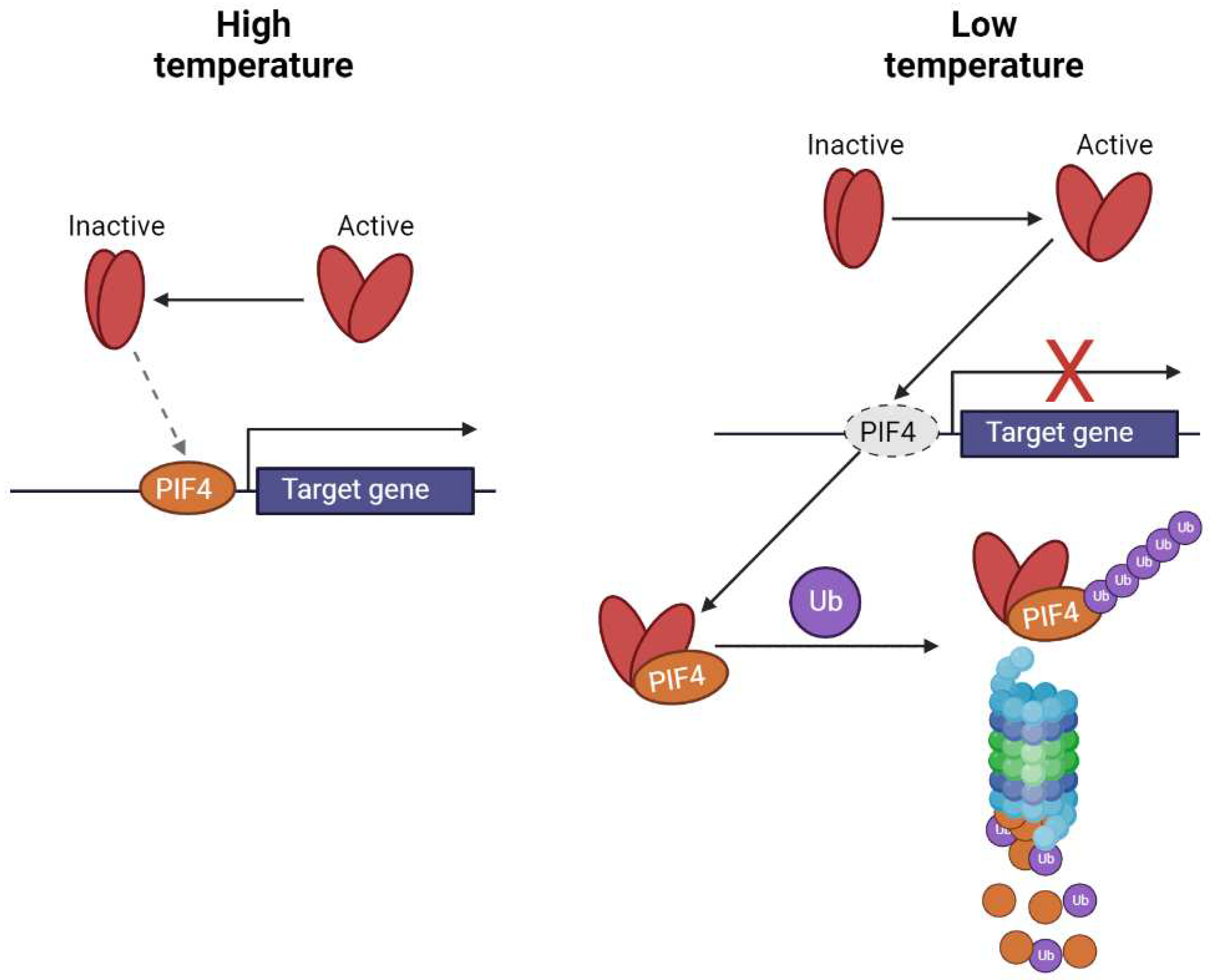
Figure 4.
Scheme representing the WRKY-WRKY interactions within and between different subfamilies (Adopted from [94]).
Figure 4.
Scheme representing the WRKY-WRKY interactions within and between different subfamilies (Adopted from [94]).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



