
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Prophylactic vaccines are important in the control of visceral leishmaniasis (VL) and as economic measures for public health. Despite the efforts to develop a vaccine against human VL caused by Leishmania infantum, none is available, and the focus has shifted to developing vaccines against canine visceral leishmaniasis (CVL). Currently, commercially available vaccines are targeted at CVL but are not effective. Different strategies have been applied in developing and improving vaccines, such as using chimeric proteins to expand vaccine coverage. The search for patents can be a way of tracking vaccines that have the potential to be marketed. In this context, the present work presents a summary of immunological aspects relevant to VL vaccine development with a focus on the composition of chimeric protein vaccines for CVL deposited in patent banks as an important approach for biotechnological development. The resulting data could facilitate the screening and selection of antigens to compose vaccine candidates with high performance against VL.
Keywords:
Visceral Leishmaniasis
; vaccine
; chimeric proteins
; patents
1. Introduction
1.1. General aspects of visceral leishmaniasis
Visceral leishmaniasis (VL) is a neglected disease caused by a protozoan of Leishmania donovani complex (L. donovani, L. infantum or L. chagasi) transmitted by the bite of female sandflies [1]. Due to the level of molecular similarity, L. chagasi and L. infantum are considered synonymous by some researchers, although other researchers prefer to separate them at a subspecific level, with the names L. (L.) infantum infantum and L. (L.) infantum chagasi [2,3]. VL has a worldwide distribution and has represented a major impact on public health in developing countries for decades [4,5] with a predominance of cases in the regions of East Africa, India, and Brazil, and a global incidence of around 50,000 to 90,000 new human cases [6]. In 2020, more than 90% of the number of new cases reported were concentrated in 10 countries: Brazil, China, Ethiopia, Eritrea, India, Kenya, Somalia, South Sudan, Sudan, and Yemen [6].
VL caused by L. infantum is a zoonotic disease [2], whereupon the dog (Canis lupus familiaris) plays a prominent role in the parasite’s transmission cycle and maintenance of the parasite by presenting intensive cutaneous parasitism [7,8,9]. Due to the ease of transmission of the parasite dogs to the vector, and their proximity to humans reinforces the rise in the number of cases of the disease [10].
Euthanasia of seropositive dogs is one of the control methods recommended by Brazil’s Ministry of Health [9], but is considered a controversial measure. There is no consensus on reducing the number of cases in humans and dogs [11,12,13,14,15]. However, the impact of dog euthanasia and reduction of human VL has recently been demonstrated [16]. Early diagnosis and treatment of human cases and control of the insects by the use of insecticides are other recommended measures [9]. Although other countries, such as China, have previously adopted these control measures, Brazil is currently the only country to continue using euthanasia [17,18].
However, these control measures have limitations and present a challenge to reducing the number of canine and human cases [19,20,21]. In this context, the search for the development of effective vaccines against leishmaniasis becomes essential to improving disease control measures worldwide [22,23].
1.2. Development of Vaccines Against Canine Visceral Leishmaniasis
Despite the existence of candidate vaccines being tested for the development of a vaccine against human VL, they are still not available for use. Additionally, efforts have been made to find more effective vaccines against canine visceral leishmaniasis (CVL) due to the role dogs play in maintaining the parasite transmission [24,25].
Several studies have reported the potential of different vaccine candidates to trigger immunoprotective mechanisms against CVL. Some of these trials are in progress to characterize the protective immunity of candidates for use in a canine vaccine against L. infantum infection, including live or dead Leishmania parasites, purified Leishmania antigens, live recombinant bacteria expressing Leishmania antigens, and antigen-encoding DNA plasmid [26,27,28,29,30]. Recently, the sandfly antigens have been considered an important approach to blocking the VL transmission from infected dogs [23,31,32,33].
The host's immune response to the parasite may present a pattern of resistance or susceptibility, and the intrinsic characteristics of these profiles are the key to the successful search for vaccines. In the resistance profile, a Th1-type cellular immune response prevails with the production of pro-inflammatory cytokines, such as interferon-gamma (IFN-γ), interleukin-12 (IL-12), Tumor Necrosis Factor-alpha (TNF-α), interleukin-2 (IL-2), and Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) [31,34,35,36,37,38,39]. Nitric oxide (NO) produced by macrophages activated by IFN-γ or TNF-α is a harmful agent to Leishmania parasites [40]. In contrast, the predominance of Th2-type cellular immune responses, with the production of cytokines such as IL-4 (interleukin-4), IL-10 (interleukin-10), and Transforming Growth Factor-beta (TGF-β), negatively regulates the activation of phagocytic cells, favoring the proliferation of parasites and disease development [31,41]. The study of susceptibility and resistance biomarkers in CVL could guide the search for a vaccine capable of inducing a protective response. According to this logic, an ideal vaccine against CVL should have immunological features that include the elicitation of a long-lasting cell-mediated immune response that contributes to controlling the infection [21,23,31,42].
1.2.1. Commercially available vaccines for Canine Visceral Leishmaniasis
Dogs are the target of studies for the development of vaccines against VL since the control of infection by Leishmania infantum is essential to curtailing its spread.
Two vaccines were commercially available in Europe: CaniLeish® and LetiFend®. The CaniLeish® (Virbac, France) was composed of purified excreted-secreted proteins from L. infantum (LiESP) and a purified fraction of saponin Quilaja saponaria (QA-21) as an adjuvant. The vaccine protocol consisted of three doses at 21-day intervals and an annual booster [43]. This vaccine induced a Th1-type cellular response with IFN-γ production, a humoral response with increased IgG2, and inducible nitric oxide (iNOS) and nitrogen dioxide synthesis, essential for parasite elimination. A field study showed its effectiveness in preventing clinical signs was 68.4% and 92.7% vaccine protection [44]. However, CaniLeish® has been off the market since 2021 [45,46]. The LetiFend® (Laboratorios LETI, Spain) is a more recent vaccine, and unlike the others, it is the only commercial vaccine composed of a chimeric protein containing five antigenic fragments of four L. infantum proteins (histone H2A and ribosomal proteins LiP2a, LiP2b, and LiP0), with 72% efficiency [44].
Leish-Tec® (CEVA, Brazil), has been commercially available in Brazil since 2015, composed of recombinant protein A2 from L. donovani amastigote containing saponin as an adjuvant [44,47]. The initial vaccination protocol consisted of three doses administered subcutaneously with a 21-day interval between each dose and an annual booster [14]. The A2 recombinant protein found in the amastigote stages of L. donovani, L. amazonensis, and L. infantum induced partial protection against these species in immunized mice [48,49,50]. The vaccine also induced a highly specific humoral immune response in these animals and the cellular immune response was of the mixed Th1/Th2 type [51].
In a canine model study, beagle dogs immunized (n = 21) with Leish-Tec® showed high levels of anti-A2 IgG2 antibodies after vaccination, but the vaccine induced only partial protection against L. chagasi (syn. L. infantum). Clinical signs in immunized and infected dogs appeared late, about one year after infection, as compared to the control group [44,52]. The first field study was conducted in a heterogeneous native canine population (more than 500 dogs) distributed between vaccine and control groups, with a significant reduction in CVL cases observed in the vaccinated group. However, this study was unable to demonstrate a significant reduction in infectivity in immunized dogs due to the absence of statistically significant differences in the prevalence of positive sandfly pools after feeding in each experimental group [53]. In another more recent field study, different canine populations were used in the control (beagle or mongrel dogs recruited from VL-free areas) and vaccine (dogs from endemic areas) groups. The results demonstrated that the incidence of infection among vaccinated animals (27%, 40/151) and control animals (42%, 33/78) were statistically significant. However, immunized seropositive dogs (44%, 18/40) had twice as high the number of sick animals compared to the control group (21.2%, 7/33). Although studies have shown that this vaccine induces a significant increase in IFN-γ levels, in the humoral immune response with IgG2 antibodies, and a partial protective response against the parasite, it lacks of evidence of efficacy [14,44,54]. Leish-Tec® was suspended in 2023 after finding that the A2 protein content is lower than the minimum limit established in the Product License [55].
Despite several studies on vaccines licensed for CVL, they have demonstrated that there are methodological limitations, such as a lack of standardization in the experimental design [44]. Therefore, there remains a pressing need for research to test vaccines and develop new vaccine candidates against CVL
1.2.2. Chimeric proteins used as potential vaccine candidates for visceral leishmaniasis
Due to the variability of the parasite and its interaction with a mammalian immune system, characterized by its genetic polymorphism, some studies have suggested that vaccines composed of polypeptides would be more likely to induce protective immune responses against leishmaniasis in different individuals [56,57]. In this context, the use of vaccines consisting of recombinant proteins would have a low cost, standardized achievement, and stability [58].
“Chimaera “is a Greek word used to refer to a creature with the head of a lion, the body of a goat, and the tail of a snake. Inspired by this mythological figure, chimeric proteins are those whose construction is based on two or more individual proteins or peptides fused together, producing a single polypeptide chain [59,60].
The first candidate for a chimeric protein vaccine against VL was KSAC, a polyprotein comprising kinetoplastid membrane protein 11 (KMP11), sterol 24-c-methyltransferase (SMT), amastigote protein A2, and cysteine proteinase B (CPB). A study that evaluated the vaccine found that it was able to induce antigen-specific multifunctional Th1-type cells, protecting the mice used in the study against the L. infantum challenge. Induction of pro-inflammatory cytokines, such as TNF-α and IFN-γ, has also been observed [56]. Furthermore, this vaccine significantly reduced the parasitic burden on the animals’ spleen and liver. The choice of these proteins was based on their known protective efficacy against VL [61,62,63,64]. LetiFend® (Laboratorios LETI, Spain) is the only commercially available vaccine composed of a chimeric protein [44].
Although the protective action of these proteins is recognized, it is not guaranteed to trigger efficient protection, either from the human or canine population, due to the difference between those host immune responses [56]. Therefore, numerous strategies have been used to identify antigens to compose and improve vaccines, such as the use of bioinformatics tools (reverse vaccinology). These tools provide a diversification of vaccines, and those that include polyepitopes are being explored [65]. In this context, the present review aimed to identify patents for prophylactic vaccines consisting of chimeric proteins as potential candidates against VL.
2. Patents of Chimeric Proteins (2010-2023)
Patent documents protect and grant intellectual property rights that contain important commercial, technological, and scientific information in technological development [66,67]. Also, unlike peer-reviewed scientific articles, patents provide important information on possible short-term marketable products and their more direct applicability [67].
The patent search was conducted using the tools from the National Institute of Industrial Property (INPI) [68], Espacenet [69], and the World Intellectual Property Organization (WIPO) [70] databases. The keywords were Leishmani*, Vaccine, Chimer*, multi or polyepitope, truncated, combined with Boolean operators “AND” and “OR”, and/or the codes of the International Patent Classification (ICP). Truncation allows the localization of words with a common stem but different suffixes, represented by the character (*). The codes were selected according to the WIPO classification, which allowed the association of patents linked to vaccines against VL. The codes used were subgroup A61K 39/008 (Medical preparations containing antigens or antibodies – Leishmania antigens), A61P 33/02 (Antiparasitic agents - antiprotozoals), C07K 14/44 (Peptides having more than 20 amino acids – protozoa), G01N 33/569 (Investigating or analyzing materials for microorganisms, e.g., protozoa, bacteria, viruses). C12N 15/30 (Mutation or genetic engineering, DNA or RNA concerning genetic engineering, vectors, e.g., plasmids, or their isolation, preparation or purification - Genes encoding protozoal proteins, e.g., from Plasmodium, Trypanosoma, Eimeria).
Selected patents had results by prophylactic vaccines composed of chimeric proteins and had in vivo and/or in vitro assays.
Nine patents made up of chimeric proteins were found from 2010 to 2022, four from Brazil, three from the United States of America, and two from Spain. All the patents included here were developed by bioinformatics tools for identifying proteins that presented epitopes for human and/or mice T cells as a prerequisite to the construction of chimeric proteins. The results demonstrated that they have a predominantly Th1 type profile, with the induction of proinflammatory cytokines and reduced parasitic load (when evaluated).
The patents are summarized in Table 1, including the country of priority filing, the chimeric vaccine proteins, and the summary of results.
The arrows (↓ and ↑) indicate the decrease and increase in biomarker levels and/or parasite load. * Major Histocompatibility complex (MHC). ** Bone marrow-derived dendritic cells (BMDC). A2: specific amastigote protein; CPA, CPB, and CPC: cysteine peptidase proteins; HRF: IgE-dependent histamine-releasing factor; HSP70: heat shock protein; H2A, H2B, H3, H4: nucleosomal histones; K39: kinesin-related protein; KSA and KSAC: kinetoplastid membrane protein (KM11), sterol 24-c-methyltransferase (SMT), specific amastigote protein A2 and cysteine peptidase proteins B; LACK: activated kinase C receptor homologous Leishmania protein; LiHyp 1, LiHyp5, LiHyp6, and LiHyV: hypothetical protein; LiP2a and LiP0: acidic ribosomal protein; NS: nonspecific nucleoside hydrolase (NH) and sterol 24-c-methyltransferase (SMT); NSC: nonspecific nucleoside hydrolase (NH), sterol 24-c-methyltransferase (SMT) and cysteine polypeptidase B (CPB); Prohibitin: surface protein; PSA-50S: promastigote surface antigen; SGT: Small glutamine-rich TPR proteins.
2.1. Protein targets
The chimeras described in Table 1 primarily consisted of known proteins with recognition of major histocompatibility complex (MHC), in addition to previous studies demonstrating their potential to be used in vaccines against leishmaniasis, either individually or in association with other proteins. The proteins and some findings of their protective potential against leishmaniasis have been described below and their localization is shown in the schematic picture (Figure 1).
Cysteine proteases (CP) are enzymes with important functions in the pathogenesis of several parasites during their interaction with the host. They are associated with cell and tissue invasion, protein hydrolysis, autophagy, and modulation of the host's immune response [71]. The CPA, CPB, and CPC subtypes have been explored as vaccine candidates, capable of inducing protective immunity against cutaneous and visceral leishmaniasis, with the induction of NO and IFN-γ production in mice [72,73,74]. In addition, CPs were able to induce the production of IFN-γ, TNF-α, and IgG2 and low production of IgG1 antibodies and IL-10 in vaccinated dogs [75].
Heat shock protein HSP70 is a ubiquitous 70kDa molecular chaperone, highly conserved, and important in the folding and remodeling processes of cellular proteins [76]. It can then be present in mitochondria and the endoplasmic reticulum [77]. Its expression is high during the process of passing from the invertebrate vector to the mammalian host, which is important in differentiating between the forms of the parasite [78]. HSP-70 can induce high levels of IgG2a, IFN-γ, and IL-2 in mice immunized against L. donovani and suppress the Th2-type immune response [79], as well as the maturation capacity of splenic dendritic cells in mice [80].
K39 is an immunodominant epitope in a kinesin-related protein, consisting of 39 amino acids, highly conserved in a Leishmania donovani complex, present in the amastigote forms, and involved in the intracellular process [81,82]. This protein has been used to detect anti-Leishmania antibodies in several diagnostic platforms with a high degree of accuracy. The Kalazar Detect™ (InBios International, USA), the IT LEISH® (BIO-RAD Laboratories Inc., France), and the OnSite™ Leishmania IgG/IgM Combo test (CTK Biotech, USA) are immunochromatographic tests that have been used in the diagnosis of human VL and are, available in the Brazilian public health system [83]. Furthermore, the Dual-Path Platform (DPP – Bio-Manguinhos/Fiocruz, Brazil), presents a fused protein of rK26/rK39 used for the diagnosis of CVL, also available in Brazil [84]. Regarding immunoenzymatic assays, there are several studies evaluating the performance of rK39 in the diagnosis of both human and canine VL [85,86,87,88]. Despite its wide use in the diagnostic field, only one patent (BR102021000794) was found using rK39 as a possible vaccine candidate.
Kinetoplastid membrane protein 11 (KMP11) is mainly expressed on the cell surface of amastigotes and promastigotes, but it is also found in membrane structures, intracellular vesicles, and flagellar pockets [89]. This protein can produce strong antigenicity for T cells in humans and mice, being a candidate for the vaccine [90]. Its expression increased in metacyclogenesis and largely in the amastigote form [91]. Results of studies with this protein revealed its ability to produce IFN-γ, IL-10, and IgG1 and IgG2a antibodies. Despite stimulating the production of IL-10 and IgG1, their levels were lower than those of the IgG2a and IFN-γ, indicating a more prominent activation of the Th1-type response, resulting in a parasite load reduction in the spleen and lymph nodes in mice [92].
Leishmania homologue of activated C kinase (LACK) is a highly conserved protein, immunodominant present in all Leishmania species, and expressed in both amastigote and promastigote forms [93,94]. Studies have demonstrated its ability to induce CD8+ T cells and IFN-γ in L. major-infected mice [95]. In addition, the production of IFN-γ and IL-10 by peripheral blood mononuclear cells (PBMC) was reported in patients with cutaneous leishmaniasis caused by L. amazonensis, L. guyanensis, and L. braziliensis [96,97,98,99]. Similarly, asymptomatic individuals and VL cured patients presented the production of IFN-γ and TNF-α [29,100]. LACK induced the production of IL-4 [101], and different approaches are needed to redirect the initial IL-4 responses to Th1, such as the use of cytokines or DNA vectors [102,103]. However, some authors have reported failure to protect mice against L. infantum and L. donovani, even when using a DNA plasmid expressing LACK for immunization [104,105]. In contrast, Fernández et al. [106] reported promising results with the use of LACK together with the attenuated Vaccinia virus, in which the protein was recognized by the T cells of asymptomatic individuals infected with L. infantum and VL cured patients. These data on the LACK antigen reported contradictory results and did not support its use in vaccine formulations against VL.
LiP2a and LiP0 are acidic ribosomal proteins of L. infantum characterized by being immunodominant antigens recognized by the serum of humans and other animals infected by L. infantum [107,108]. These proteins were able to trigger the immune response and induce protection against infection by L. infantum and L. major in mice, with increased CD4+ and CD8+ T cells, and significant production of antigen-specific IL-12 [109]. These proteins also stimulate IFN-γ production by splenocytes in mice immunized with LiP2a [110].
Hypothetical proteins have been described in the genome of the Leishmania spp. but without a defined biological role [111]. Through an immunoproteomic study, LiHyp1 and LiHyp2 were recognized by antibodies in serum from VL patients but not by serum of healthy individuals [112]. The use of hypothetical proteins is still a field to be explored. Some authors have demonstrated that hypothetical proteins are capable of reducing the parasitic load on the liver, spleen, bone marrow, and lymph nodes, with an immunogenic profile related to high levels of IFN-γ, IL-12, GM-CSF, and specific IgG2 production [113,114].
Nucleoside hydrolases (NH) are vital enzymes in the metabolism of DNA, being essential for the replication of parasites, especially during the initial stages of infection. These enzymes are present in all species of Leishmania spp., justifying their use as phylogenetic markers. Additionally, they share high identification levels among many microorganisms but are absent in mammals [115]. These characteristics make NH targets for an anti-VL vaccine by inducing high immunogenicity [115]. For instance, the NH36 of L. donovani is a nonspecific nucleoside hydrolase that is the main antigen of Leishmune®, a vaccine previously sold in Brazil from 2004-2014, which was discontinued due to noncompliance with the requirements of phase III studies for efficacy (Brazil, Technical Note 038/2014). This vaccine had an efficacy and protection greater than 80%, being able to induce high levels of IgG2, a predominantly Th1 immune response with high production of IFN-γ, TNF-α, and IL-17 [44]. Other studies have affirmed the potential of NH [115,116].
Nucleosomal histones are important proteins in the DNA packaging process, transcription, and gene regulation. Evidence suggests that histones from Leishmania spp. are relevant immunogens during parasite/host interaction [117]. Therefore, histones H2A, H2B, H3, and H4 have been studied as potential vaccines against leishmaniasis. A profile similar to the Th1 response was detected in murine models immunized with histones against L. major and L. donovani infection with IFN-γ and TNF-α induction and low IL-4 production [117,118,119,120]. Moreover, the immunization was able to stimulate immune responses in PBMCs of cured individuals and patients infected with Leishmania, as well as reduce the parasitic burden by more than 80% in the spleen, liver, and bone marrow in hamsters [120].
Prohibitins (PHB) are conserved proteins found in all eukaryotic cells in the inner membrane of the mitochondria. These proteins are important in several functions linked to this organelle and the stabilization of its membrane. In Leishmania spp., prohibitins are involved in cell proliferation, greater infectivity, and maintenance of the parasite`s mitochondrial integrity [121,122]. The presence of anti-PHB antibodies in patients infected with L. donovani demonstrated that these proteins are relevant when the disease is active [121]. A study by Lage et al. [123] verified the induction of a Th1-type cellular response with high levels of IFN-γ, IL-12, and GM-CSF in immunized animals. Moreover, a significant reduction in the parasite load was reported in the spleen, liver, lymph nodes, and bone marrow in mice. High levels of IFN-γ in PBMC from healthy individuals and cured VL patients have also been observed [124].
Promastigote surface antigens (PSA) are members of a family of membrane-bound or secreted proteins from Leishmania spp., involved in resistance to lysis promoted by the complement system during interaction with host cells [125,126]. PSA is highly conserved and, despite being recognized by immune response cells, preferably Th1 in humans, it does not share homology with mammalian cells [127]. Studies with PSA subtypes have shown that they are capable of inducing Th1-type responses with IFN-γ production in mice against L. major infection [128] and proliferation of PBMCs in patients with cutaneous leishmaniasis in response to PSA-2 [129]. Chamakh-Ayari et al. [127] demonstrated that the PSA-38S of L. amazonensis induced a mixed Th1/Th2 response in individuals with immunity to L. major and L. infantum, in addition to inducing granzyme B production. Petitdidier et al. [130] demonstrated that PSA can induce IFN-γ, nitric oxide, and IgG2 antibodies in vaccinated dogs.
Small glutamine-rich TPR proteins (SGT) are co-chaperones that interact with HSP70 and HSP90 chaperones to ensure their proper functions, which are essential to the parasite's life cycle and survival [131]. Dias et al. [132] identified the potential of L. infantum's SGT for vaccine and diagnostic approaches against VL. Immunized mice with SGT developed a specific Th1 responses by producing IFN-γ, IL-12, and IgG2a, which induced a resistance profile against the infection.
Specific amastigote protein A2 corresponds to a family of specific amastigote genes necessary for the parasite’s survival and result in a virulence factore [133]. A2 was the main protein in the Leish-Tec® vaccine and has been explored as a possible vaccine candidate as it has been shown to induce a Th1 immune response represented by biomarkers IFN-γ, TNF-α, and IgG2 antibodies that conferred partial protection against CVL [14,47,52,134,135].
Sterol 24-c-methyltransferase (SMT) is an enzyme of the transferase family, important in steroid production, especially ergosterol biosynthesis present in the Leishmania spp. membrane [136]. Mice immunized with SMT showed IFN-γ induction and lower spleen and liver parasite loads [64]. SMT, together with the nucleoside hydrolase NH36 of L. donovani, composed a vaccine developed against human VL, reaching Phase I clinical trial [137].
3. Discussion and future perspectives
Vaccines are considered essential tools in preventing VL. Classic vaccines against leishmaniasis were based on live or attenuated parasites or their subunits. Additionally, the search for more specific vaccines could be more promising using Leishmania peptides that display high immunogenicity and protection profiles. As discussed here, the development of vaccines consisting of chimeric proteins could be more effective by linking several of the parasite’s antigens in a peptide chain. The genetic polymorphism of mammals and the variability of parasites would no longer be a hindrance. In addition, peptides offer such advantages as good stability, absence of potentially harmful materials, low antigen complexity, and low cost of amplification.
The construction of these proteins is only possible with the use of reverse vaccinology using bioinformatics tools. This approach can predict specific T-cells epitopes, an important action for targeting VL antigens and triggering an effective adaptive immune response. That is why chimeric vaccines show so much promise [138]. However, peptides are poorly immunogenic and the use of a potent adjuvant or nanotechnology by antigen encapsulation could overcome this drawback.
Chimeric vaccines must be constructed from Leishmania spp. associated with parasite invasion and survival in the vertebrate host (mammals) [139]. As has been demonstrated, the vaccines had different compositions and, with the exception of patent number. BR1020160061210, composed of hypothetical proteins (unknown), all contained one or more proteins related to the invasion, survival, and/or metabolic processes important for parasites in the vertebrate host.
Regarding the results here described, all vaccines composed of chimeric proteins induced high levels of proinflammatory cytokines, mainly IFN-γ, in cell cultures, while reducing the parasitic load of the spleen, liver, and/or lymph nodes. Furthermore, the compositions of patent BR1020180081977 were able to induce CD4+ and T CD8+ T-cells with a central memory phenotype.
4. Conclusions
The patents reporting the use of Leishmania antigens demonstrated that vaccines composed of chimeric proteins have the potential to control VL. Therefore, this review is offered as a way to facilitate the identification and selection of chimeric proteins for the improvement and development of new VL vaccines.
Author Contributions
Conceptualization, D.S.O. and R.C.G.; writing—original draft preparation, D.S.O; M.F.Z.; A.A.M.G.; L.A.R.O.; R.M.S.M.; D.F.S.P.; I.S.S.C.; M.A.F.C.; D.F.L.; D.F.V.B.; E.N.N.; I.S.A.; W.M.G.; D.S-L.; A.S.G.; R.A.P.N.; W.O.D. writing—review and editing, D.S.O.; A.A.M.G.; M.F.Z.; M.A.F.C. and L.A.R.O.; supervision, D.S.O. and R.C.G.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data collected were reported in the text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ready, P. D. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annu. Rev. Entomol., 2013, 58, 227–250. [Google Scholar] [CrossRef]
- Maurício, I. L.; Stothard, J. R.; Miles, M. A. The Strange Case of Leishmania Chagasi. Parasitol. Today Pers. Ed, 2000, 16, 188–189. [Google Scholar] [CrossRef]
- Lainson, R.; Shaw, J. J. Parasitology, 10th ed.; Hodder Arnold: London, 2005. [Google Scholar]
- Desjeux, P. Leishmaniasis: Current Situation and New Perspectives. Comp. Immunol. Microbiol. Infect. Dis., 2004, 27, 305–318. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I. D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PloS One, 2012, 7, e35671. [Google Scholar] [CrossRef]
- Leishmaniasis https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed Jan 8, 2024).
- Dantas-Torres, F. The Role of Dogs as Reservoirs of Leishmania Parasites, with Emphasis on Leishmania (Leishmania) Infantum and Leishmania (Viannia) Braziliensis. Vet. Parasitol., 2007, 149, 139–146. [Google Scholar] [CrossRef]
- Verçosa, B. L. A.; Melo, M. N.; Puerto, H. L. D.; Mendonça, I. L.; Vasconcelos, A. C. Apoptosis, Inflammatory Response and Parasite Load in Skin of Leishmania (Leishmania) Chagasi Naturally Infected Dogs: A Histomorphometric Analysis. Vet. Parasitol., 2012, 189, 162–170. [Google Scholar] [CrossRef]
- Prado, P. F. do; Rocha, M. F.; Sousa, J. F. de; Caldeira, D. I.; Paz, G. F.; Dias, E. S. Epidemiological Aspects of Human and Canine Visceral Leishmaniasis in Montes Claros, State of Minas Gerais, Brazil, between 2007 and 2009. Rev. Soc. Bras. Med. Trop., 2011, 44, 561–566. [Google Scholar] [CrossRef]
- Courtenay, O.; Quinnell, R. J.; Garcez, L. M.; Shaw, J. J.; Dye, C. Infectiousness in a Cohort of Brazilian Dogs: Why Culling Fails to Control Visceral Leishmaniasis in Areas of High Transmission. J. Infect. Dis., 2002, 186, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Costa, C. H. N.; Tapety, C. M. M.; Werneck, G. L. Controle da leishmaniose visceral em meio urbano: estudo de intervenção randomizado fatorial. Rev. Soc. Bras. Med. Trop., 2007, 40, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C. M.; Pires, M. M.; da Silva, K. M.; Assis, F. D.; Gonçalves Filho, J.; Perri, S. H. V. Relationship between Dog Culling and Incidence of Human Visceral Leishmaniasis in an Endemic Area. Vet. Parasitol., 2010, 170, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, G.; Teva, A.; dos-Santos, C. B.; Santos, F. N.; Pinto, I. de-Souza; Fux, B.; Leite, G. R.; Falqueto, A. Field Trial of Efficacy of the Leish-Tec® Vaccine against Canine Leishmaniasis Caused by Leishmania Infantum in an Endemic Area with High Transmission Rates. PLoS ONE, 2017, 12 (9), e0185438. [CrossRef]
- Coura-Vital, W.; Ker, H. G.; Roatt, B. M.; Aguiar-Soares, R. D. O.; Leal, G. G. de A.; Moreira, N. das D.; Oliveira, L. A. M.; de Menezes Machado, E. M.; Morais, M. H. F.; Corrêa-Oliveira, R.; et al. Evaluation of Change in Canine Diagnosis Protocol Adopted by the Visceral Leishmaniasis Control Program in Brazil and a New Proposal for Diagnosis. PloS One, 2014, 9 (3), e91009. [CrossRef]
- França-Silva, J. C.; Giunchetti, R. C.; Mariano, R. M. da S.; Machado-Coelho, G. L. L.; Teixeira, L. de A. S.; Barata, R. A.; Michalsky, É. M.; Rocha, M. F.; Fortes-Dias, C. L.; Dias, E. S. The Program for the Control of Visceral Leishmaniasis in Brazil: The Effect of the Systematic Euthanasia of Seropositive Dogs as a Single Control Action in Porteirinha, a Brazilian City with an Intense Transmission of Visceral Leishmaniasis. Pathog. Basel Switz., 2023, 12 (8), 1060. [CrossRef]
- Costa, C. H. N. How Effective Is Dog Culling in Controlling Zoonotic Visceral Leishmaniasis? A Critical Evaluation of the Science, Politics and Ethics behind This Public Health Policy. Rev. Soc. Bras. Med. Trop., 2011, 44, 232–242. [Google Scholar] [CrossRef]
- Otranto, D.; Dantas-Torres, F. The Prevention of Canine Leishmaniasis and Its Impact on Public Health. Trends Parasitol., 2013, 29, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Maia-Elkhoury, A. N. S.; Alves, W. A.; Sousa-Gomes, M. L. de; Sena, J. M. de; Luna, E. A. Visceral Leishmaniasis in Brazil: Trends and Challenges. Cad. Saúde Pública, 2008, 24, 2941–2947. [Google Scholar] [CrossRef] [PubMed]
- Gramiccia, M.; Gradoni, L. The Current Status of Zoonotic Leishmaniases and Approaches to Disease Control. Int. J. Parasitol., 2005, 35, 1169–1180. [Google Scholar] [CrossRef]
- Reis, A. B.; Giunchetti, R. C.; Carrillo, E.; Martins-Filho, O. A.; Moreno, J. Immunity to Leishmania and the Rational Search for Vaccines against Canine Leishmaniasis. Trends Parasitol., 2010, 26, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Mohebali, M.; Khamesipour, A.; Mobedi, I.; Zarei, Z.; Hashemi-Fesharki, R. Double-Blind Randomized Efficacy Field Trial of Alum Precipitated Autoclaved Leishmania Major Vaccine Mixed with BCG against Canine Visceral Leishmaniasis in Meshkin-Shahr District, I.R. Iran. Vaccine, 2004, 22, 4097–4100. [Google Scholar] [CrossRef]
- Leite, J. C.; Gonçalves, A. A. M.; de Oliveira, D. S.; Resende, L. A.; Boas, D. F. V.; Ribeiro, H. S.; Pereira, D. F. S.; da Silva, A. V.; Mariano, R. M. da S.; Reis, P. C. C.; et al. Transmission-Blocking Vaccines for Canine Visceral Leishmaniasis: New Progress and Yet New Challenges. Vaccines, 2023, 11 (10), 1565. [CrossRef]
- Le Rutte, E. A.; Coffeng, L. E.; Malvolti, S.; Kaye, P. M.; de Vlas, S. J. The Potential Impact of Human Visceral Leishmaniasis Vaccines on Population Incidence. PLoS Negl. Trop. Dis., 2020, 14, e0008468. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martínez, A. F.; Newman, S.; Ramanan, P.; Suarez, J. A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Curr. Trop. Med. Rep., 2021, 8, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Abdian, N.; Gholami, E.; Zahedifard, F.; Safaee, N.; Rafati, S. Evaluation of DNA/DNA and Prime-Boost Vaccination Using LPG3 against Leishmania Major Infection in Susceptible BALB/c Mice and Its Antigenic Properties in Human Leishmaniasis. Exp. Parasitol., 2011, 127, 627–636. [Google Scholar] [CrossRef]
- Araújo, M. S. S.; de Andrade, R. A.; Sathler-Avelar, R.; Magalhães, C. P.; Carvalho, A. T.; Andrade, M. C.; Campolina, S. S.; Mello, M. N.; Vianna, L. R.; Mayrink, W.; et al. Immunological Changes in Canine Peripheral Blood Leukocytes Triggered by Immunization with First or Second Generation Vaccines against Canine Visceral Leishmaniasis. Vet. Immunol. Immunopathol., 2011, 141, 64–75. [Google Scholar] [CrossRef]
- Nagill, R.; Kaur, S. Vaccine Candidates for Leishmaniasis: A Review. Int. Immunopharmacol., 2011, 11, 1464–1488. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sundar, S. Leishmaniasis: Vaccine Candidates and Perspectives. Vaccine, 2012, 30, 3834–3842. [Google Scholar] [CrossRef] [PubMed]
- Thomaz-Soccol, V.; Ferreira da Costa, E. S.; Karp, S. G.; Junior Letti, L. A.; Soccol, F. T.; Soccol, C. R. Recent Advances in Vaccines Against Leishmania Based on Patent Applications. Recent Pat. Biotechnol., 2018, 12, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Giunchetti, R. C.; Silveira, P. C.; Resende, L. A.; Leite, J. C.; Melo-Júnior, O.; Rodrigues-Alves, M.; Costa, L. M.; Lair, D.; Chaves, V.; Soares, I.; et al. Canine visceral leishmaniasis biomarkers and their employment in vaccines - ScienceDirect https://www.sciencedirect.com/science/article/pii/S0304401719301025?via%3Dihub (accessed Jan 8, 2024).
- Graciano, R. C. D.; Ribeiro, J. A. T.; Macêdo, A. K. S.; de S Lavareda, J. P.; de Oliveira, P. R.; Netto, J. B.; Nogueira, L. M.; Machado, J. M.; Camposda-Paz, M.; Giunchetti, R. C.; et al. Recent Patents Applications in Red Biotechnology: A Mini-Review. Recent Pat. Biotechnol., 2019, 13, 170–186. [Google Scholar] [CrossRef] [PubMed]
- de Lana, M.; Giunchetti, R. C. Dogs as a Model for Chemotherapy of Chagas Disease and Leishmaniasis. Curr. Pharm. Des., 2021, 27, 1741–1756. [Google Scholar] [CrossRef] [PubMed]
- Murray, H. W.; Cervia, J. S.; Hariprashad, J.; Taylor, A. P.; Stoeckle, M. Y.; Hockman, H. Effect of Granulocyte-Macrophage Colony-Stimulating Factor in Experimental Visceral Leishmaniasis. J. Clin. Invest., 1995, 95, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Agallou, M.; Smirlis, D.; Soteriadou, K. P.; Karagouni, E. Vaccination with Leishmania Histone H1-Pulsed Dendritic Cells Confers Protection in Murine Visceral Leishmaniasis. Vaccine, 2012, 30, 5086–5093. [Google Scholar] [CrossRef]
- Das, A.; Ali, N. Vaccine Prospects of Killed but Metabolically Active Leishmania against Visceral Leishmaniasis. Expert Rev. Vaccines, 2012, 11, 783–785. [Google Scholar] [CrossRef]
- Dumas, C.; Muyombwe, A.; Roy, G.; Matte, C.; Ouellette, M.; Olivier, M.; Papadopoulou, B. Recombinant Leishmania Major Secreting Biologically Active Granulocyte-Macrophage Colony-Stimulating Factor Survives Poorly in Macrophages in Vitro and Delays Disease Development in Mice. Infect. Immun., 2003, 71, 6499–6509. [Google Scholar] [CrossRef]
- Lage, D. P.; Martins, V. T.; Duarte, M. C.; Garde, E.; Chávez-Fumagalli, M. A.; Menezes-Souza, D.; Roatt, B. M.; Tavares, C. a. P.; Soto, M.; Coelho, E. a. F. Prophylactic Properties of a Leishmania-Specific Hypothetical Protein in a Murine Model of Visceral Leishmaniasis. Parasite Immunol., 2015, 37, 646–656. [Google Scholar] [CrossRef]
- Martins, V. T.; Lage, D. P.; Duarte, M. C.; Costa, L. E.; Garde, E.; Rodrigues, M. R.; Chávez-Fumagalli, M. A.; Menezes-Souza, D.; Roatt, B. M.; Tavares, C. A. P.; et al. A New Leishmania-Specific Hypothetical Protein, LiHyT, Used as a Vaccine Antigen against Visceral Leishmaniasis. Acta Trop., 2016, 154, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Green, S. J.; Mellouk, S.; Hoffman, S. L.; Meltzer, M. S.; Nacy, C. A. Cellular Mechanisms of Nonspecific Immunity to Intracellular Infection: Cytokine-Induced Synthesis of Toxic Nitrogen Oxides from L-Arginine by Macrophages and Hepatocytes. Immunol. Lett., 1990, 25, 15–19. [Google Scholar] [CrossRef]
- Blackwell, J. M.; Fakiola, M.; Ibrahim, M. E.; Jamieson, S. E.; Jeronimo, S. B.; Miller, E. N.; Mishra, A.; Mohamed, H. S.; Peacock, C. S.; Raju, M.; et al. Genetics and Visceral Leishmaniasis: Of Mice and Man. Parasite Immunol., 2009, 31, 254–266. [Google Scholar] [CrossRef]
- Kaye, P. M.; Aebischer, T. Visceral Leishmaniasis: Immunology and Prospects for a Vaccine. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis., 2011, 17, 1462–1470. [Google Scholar] [CrossRef]
- Moreno, J.; Vouldoukis, I.; Martin, V.; McGahie, D.; Cuisinier, A.-M.; Gueguen, S. Use of a LiESP/QA-21 Vaccine (CaniLeish) Stimulates an Appropriate Th1-Dominated Cell-Mediated Immune Response in Dogs. PLoS Negl. Trop. Dis., 2012, 6, e1683. [Google Scholar] [CrossRef] [PubMed]
- Velez, R.; Gállego, M. Commercially Approved Vaccines for Canine Leishmaniosis: A Review of Available Data on Their Safety and Efficacy. Trop. Med. Int. Health TM IH, 2020, 25, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Baxarias, M.; Homedes, J.; Mateu, C.; Attipa, C.; Solano-Gallego, L. Use of Preventive Measures and Serological Screening Tools for Leishmania Infantum Infection in Dogs from Europe. Parasit. Vectors, 2022, 15, 134. [Google Scholar] [CrossRef] [PubMed]
- CaniLeish | Agência Europeia de Medicamentos https://www.ema.europa.eu/en/medicines/veterinary/EPAR/canileish#ema-inpage-item-product-info (accessed Jan 14, 2024).
- Fernandes, C. B.; Junior, J. T. M.; de Jesus, C.; Souza, B. M. P. da S.; Larangeira, D. F.; Fraga, D. B. M.; Tavares Veras, P. S.; Barrouin-Melo, S. M. Comparison of Two Commercial Vaccines against Visceral Leishmaniasis in Dogs from Endemic Areas: IgG, and Subclasses, Parasitism, and Parasite Transmission by Xenodiagnosis. Vaccine, 2014, 32 (11), 1287–1295. [CrossRef]
- Coelho, E. A. F.; Tavares, C. A. P.; Carvalho, F. A. A.; Chaves, K. F.; Teixeira, K. N.; Rodrigues, R. C.; Charest, H.; Matlashewski, G.; Gazzinelli, R. T.; Fernandes, A. P. Immune Responses Induced by the Leishmania (Leishmania) Donovani A2 Antigen, but Not by the LACK Antigen, Are Protective against Experimental Leishmania (Leishmania) Amazonensis Infection. Infect. Immun., 2003, 71, 3988–3994. [Google Scholar] [CrossRef] [PubMed]
- Zanin, F. H. C.; Coelho, E. A. F.; Tavares, C. A. P.; Marques-da-Silva, E. A.; Silva Costa, M. M.; Rezende, S. A.; Gazzinelli, R. T.; Fernandes, A. P. Evaluation of Immune Responses and Protection Induced by A2 and Nucleoside Hydrolase (NH) DNA Vaccines against Leishmania Chagasi and Leishmania Amazonensis Experimental Infections. Microbes Infect., 2007, 9, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Resende, D. M.; Caetano, B. C.; Dutra, M. S.; Penido, M. L. O.; Abrantes, C. F.; Verly, R. M.; Resende, J. M.; Piló-Veloso, D.; Rezende, S. A.; Bruna-Romero, O.; et al. Epitope Mapping and Protective Immunity Elicited by Adenovirus Expressing the Leishmania Amastigote Specific A2 Antigen: Correlation with IFN-Gamma and Cytolytic Activity by CD8+ T Cells. Vaccine, 2008, 26, 4585–4593. [Google Scholar] [CrossRef] [PubMed]
- de Mendonça, L. Z.; Resende, L. A.; Lanna, M. F.; Aguiar-Soares, R. D. de O.; Roatt, B. M.; Castro, R. A. de O. E.; Batista, M. A.; Silveira-Lemos, D.; Gomes, J. de A. S.; Fujiwara, R. T.; et al. Multicomponent LBSap Vaccine Displays Immunological and Parasitological Profiles Similar to Those of Leish-Tec® and Leishmune® Vaccines against Visceral Leishmaniasis. Parasit. Vectors, 2016, 9 (1), 472. [CrossRef]
- Fernandes, A. P.; Costa, M. M. S.; Coelho, E. A. F.; Michalick, M. S. M.; de Freitas, E.; Melo, M. N.; Luiz Tafuri, W.; Resende, D. de M.; Hermont, V.; Abrantes, C. de F.; et al. Protective Immunity against Challenge with Leishmania (Leishmania) Chagasi in Beagle Dogs Vaccinated with Recombinant A2 Protein. Vaccine, 2008, 26 (46), 5888–5895. [CrossRef]
- Regina-Silva, S.; Feres, A. M. L. T.; França-Silva, J. C.; Dias, E. S.; Michalsky, É. M.; de Andrade, H. M.; Coelho, E. A. F.; Ribeiro, G. M.; Fernandes, A. P.; Machado-Coelho, G. L. L. Field Randomized Trial to Evaluate the Efficacy of the Leish-Tec® Vaccine against Canine Visceral Leishmaniasis in an Endemic Area of Brazil. Vaccine, 2016, 34, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Wylie, C. E.; Carbonell-Antoñanzas, M.; Aiassa, E.; Dhollander, S.; Zagmutt, F. J.; Brodbelt, D. C.; Solano-Gallego, L. A Systematic Review of the Efficacy of Prophylactic Control Measures for Naturally-Occurring Canine Leishmaniosis, Part I: Vaccinations. Prev. Vet. Med., 2014, 117, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Mapa suspende fabricação e venda e determina o recolhimento de lotes de vacina contra Leishmaniose https://www.gov.br/agricultura/pt-br/assuntos/noticias/mapa-suspende-fabricacao-e-venda-e-determina-o-recolhimento-de-lotes-de-vacina-contra-leishmaniose-apos-fiscalizacao (accessed Jan 14, 2024).
- Goto, Y.; Bhatia, A.; Raman, V. S.; Liang, H.; Mohamath, R.; Picone, A. F.; Vidal, S. E. Z.; Vedvick, T. S.; Howard, R. F.; Reed, S. G. KSAC, the First Defined Polyprotein Vaccine Candidate for Visceral Leishmaniasis▿. Clin. Vaccine Immunol. CVI, 2011, 18, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Agallou, M.; Margaroni, M.; Kotsakis, S. D.; Karagouni, E. A Canine-Directed Chimeric Multi-Epitope Vaccine Induced Protective Immune Responses in BALB/c Mice Infected with Leishmania Infantum. Vaccines, 2020, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Jain, N. K. Vaccines for Visceral Leishmaniasis: A Review. J. Immunol. Methods, 2015, 422, 1–12. [Google Scholar] [CrossRef]
- Bonde, J. S.; Bülow, L. Chimeric Genes, Proteins. In Brenner’s Encyclopedia of Genetics (Second Edition); Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, 2013; pp. 519–520. [Google Scholar] [CrossRef]
- Strohl, W. Chimeric Genes, Proteins☆. In Reference Module in Life Sciences; Elsevier, 2017. [CrossRef]
- Ghosh, A.; Zhang, W. W.; Matlashewski, G. Immunization with A2 Protein Results in a Mixed Th1/Th2 and a Humoral Response Which Protects Mice against Leishmania Donovani Infections. Vaccine, 2001, 20, 59–66. [Google Scholar] [CrossRef]
- Basu, R.; Bhaumik, S.; Basu, J. M.; Naskar, K.; De, T.; Roy, S. Kinetoplastid Membrane Protein-11 DNA Vaccination Induces Complete Protection against Both Pentavalent Antimonial-Sensitive and -Resistant Strains of Leishmania Donovani That Correlates with Inducible Nitric Oxide Synthase Activity and IL-4 Generation: Evidence for Mixed Th1- and Th2-Like Responses in Visceral Leishmaniasis1. J. Immunol., 2005, 174, 7160–7171. [Google Scholar] [CrossRef]
- Rafati, S.; Zahedifard, F.; Nazgouee, F. Prime-Boost Vaccination Using Cysteine Proteinases Type I and II of Leishmania Infantum Confers Protective Immunity in Murine Visceral Leishmaniasis. Vaccine, 2006, 24, 2169–2175. [Google Scholar] [CrossRef]
- Goto, Y.; Bogatzki, L. Y.; Bertholet, S.; Coler, R. N.; Reed, S. G. Protective Immunization against Visceral Leishmaniasis Using Leishmania Sterol 24-c-Methyltransferase Formulated in Adjuvant. Vaccine, 2007, 25, 7450–7458. [Google Scholar] [CrossRef]
- Brito, R. C. F. D.; Ruiz, J. C.; Cardoso, J. M. de O.; Ostolin, T. L. V. D. P.; Reis, L. E. S.; Mathias, F. A. S.; Aguiar-Soares, R. D. de O.; Roatt, B. M.; Corrêa-Oliveira, R.; Resende, D. de M.; et al. Chimeric Vaccines Designed by Immunoinformatics-Activated Polyfunctional and Memory T Cells That Trigger Protection against Experimental Visceral Leishmaniasis. Vaccines, 2020, 8 (2), 252. [CrossRef]
- Chang, P.-L.; Wu, C.-C.; Leu, H.-J. Using Patent Analyses to Monitor the Technological Trends in an Emerging Field of Technology: A Case of Carbon Nanotube Field Emission Display. Scientometrics, 2009, 82, 5–19. [Google Scholar] [CrossRef]
- Mucke, H. A. M. What Patents Tell Us about Drug Repurposing for Cancer: A Landscape Analysis. Semin. Cancer Biol., 2021, 68, 3–7. [Google Scholar] [CrossRef]
- INPI https://www.gov.br/inpi/pt-br/copy2_of_nova-home-page (accessed Jan 8, 2024).
- Espacenet – patent search https://worldwide.espacenet.com/ (accessed Jan 8, 2024).
- WIPO - Search International and National Patent Collections https://patentscope.wipo.int/search/en/search.jsf (accessed Jan 8, 2024).
- Siqueira-Neto, J. L.; Debnath, A.; McCall, L.-I.; Bernatchez, J. A.; Ndao, M.; Reed, S. L.; Rosenthal, P. J. Cysteine Proteases in Protozoan Parasites. PLoS Negl. Trop. Dis., 2018, 12, e0006512. [Google Scholar] [CrossRef] [PubMed]
- Agallou, M.; Athanasiou, E.; Koutsoni, O.; Dotsika, E.; Karagouni, E. Experimental Validation of Multi-Epitope Peptides Including Promising MHC Class I- and II-Restricted Epitopes of Four Known Leishmania Infantum Proteins. Front. Immunol., 2014, 5, 268. [Google Scholar] [CrossRef]
- Seyed, N.; Taheri, T.; Vauchy, C.; Dosset, M.; Godet, Y.; Eslamifar, A.; Sharifi, I.; Adotevi, O.; Borg, C.; Rohrlich, P. S.; et al. Immunogenicity Evaluation of a Rationally Designed Polytope Construct Encoding HLA-A*0201 Restricted Epitopes Derived from Leishmania Major Related Proteins in HLA-A2/DR1 Transgenic Mice: Steps toward Polytope Vaccine. PLoS ONE, 2014, 9, e108848. [Google Scholar] [CrossRef] [PubMed]
- Noormehr, H.; Zavaran Hosseini, A.; Soudi, S.; Beyzay, F. Enhancement of Th1 Immune Response against Leishmania Cysteine Peptidase A, B by PLGA Nanoparticle. Int. Immunopharmacol., 2018, 59, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.; Zahedifard, F.; Saljoughian, N.; Doroud, D.; Jamshidi, S.; Mahdavi, N.; Shirian, S.; Daneshbod, Y.; Hamid Zarkesh-Esfahani, S.; Papadopoulou, B.; et al. Immunological Comparison of DNA Vaccination Using Two Delivery Systems against Canine Leishmaniasis. Vet. Parasitol., 2015, 212, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Nillegoda, N. B.; Mayer, M. P.; Bukau, B. The Hsp70 Chaperone Network. Nat. Rev. Mol. Cell Biol., 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Clare, D. K.; Saibil, H. R. ATP-Driven Molecular Chaperone Machines. Biopolymers, 2013, 99, 846–859. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A.; Maier, A. G.; Przyborski, J. M.; Blatch, G. L. Intracellular Protozoan Parasites of Humans: The Role of Molecular Chaperones in Development and Pathogenesis. Protein Pept. Lett., 2011, 18, 143–157. [Google Scholar] [CrossRef]
- Kaur, T.; Sobti, R. C.; Kaur, S. Cocktail of Gp63 and Hsp70 Induces Protection against Leishmania Donovani in BALB/c Mice. Parasite Immunol., 2011, 33, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, M.; Karimi, M. H.; Kalani, M.; Ebrahimnezhad, S.; Namayandeh, M.; Moravej, A. Immunostimulatory Effects of Leishmania Infantum HSP70 Recombinant Protein on Dendritic Cells in Vitro and in Vivo. Immunotherapy, 2014, 6, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Burns, J. M.; Shreffler, W. G.; Benson, D. R.; Ghalib, H. W.; Badaro, R.; Reed, S. G. Molecular Characterization of a Kinesin-Related Antigen of Leishmania Chagasi That Detects Specific Antibody in African and American Visceral Leishmaniasis. Proc. Natl. Acad. Sci. U. S. A., 1993, 90, 775–779. [Google Scholar] [CrossRef] [PubMed]
- de Souza, L. M. B.; Carvalho, J.; Bates, M. D.; Petterle, R. R.; Thomaz-Soccol, V.; Bates, P. A. Production of a Kinesin-Related Recombinant Protein (Lbk39) from Leishmania Braziliensis by Leishmania Tarentolae Promastigotes and Its Application in the Serodiagnosis of Leishmaniasis. One Health, 2019, 8, 100111. [Google Scholar] [CrossRef] [PubMed]
- Freire, M. L.; Assis, T. S. M. de; Avelar, D. M. de; Rabello, A.; Cota, G. Evaluation of a New Brand of Immunochromatographic Test for Visceral Leishmaniasis in Brazil Made Available from 2018. Rev. Inst. Med. Trop. Sao Paulo, 2018, 60, e49. [Google Scholar] [CrossRef]
- Garcia, V. S.; Guerrero, S. A.; Gugliotta, L. M.; Gonzalez, V. D. G. A Lateral Flow Immunoassay Based on Colored Latex Particles for Detection of Canine Visceral Leishmaniasis. Acta Trop., 2020, 212, 105643. [Google Scholar] [CrossRef]
- Assis, T. S. M. de; Braga, A. S. da C.; Pedras, M. J.; Barral, A. M. P.; Siqueira, I. C. de; Costa, C. H. N.; Costa, D. L.; Holanda, T. A.; Soares, V. Y. R.; Biá, M.; et al. Validação Do Teste Imunocromatográfico Rápido IT-LEISH® Para o Diagnóstico Da Leishmaniose Visceral Humana. Epidemiol. E Serviços Saúde, 2008, 17 (2), 107–116. [CrossRef]
- Pedras, M. J.; de Gouvêa Viana, L.; de Oliveira, E. J.; Rabello, A. Comparative Evaluation of Direct Agglutination Test, rK39 and Soluble Antigen ELISA and IFAT for the Diagnosis of Visceral Leishmaniasis. Trans. R. Soc. Trop. Med. Hyg., 2008, 102, 172–178. [Google Scholar] [CrossRef]
- Romero, H. D.; Silva, L. de A.; Silva-Vergara, M. L.; Rodrigues, V.; Costa, R. T.; Guimarães, S. F.; Alecrim, W.; Moraes-Souza, H.; Prata, A. Comparative Study of Serologic Tests for the Diagnosis of Asymptomatic Visceral Leishmaniasis in an Endemic Area. Am. J. Trop. Med. Hyg., 2009, 81 (1), 27–33.
- Medeiros, F. A. C.; Souza Filho, J. A. de; Barbosa, J. R.; Donato, L. E.; Figueiredo, F. B.; Werneck, G. L.; Paz, G. F.; Thompson, M.; Marcelino, A. P. Phase II Validation Study of the rK39 ELISA Prototype for the Diagnosis of Canine Visceral Leishmaniasis in Brazil. Cad. Saúde Pública, 2021, 37, e00041320. [Google Scholar] [CrossRef]
- Salari, S.; Sharifi, I.; Keyhani, A. R.; Ghasemi Nejad Almani, P. Evaluation of a New Live Recombinant Vaccine against Cutaneous Leishmaniasis in BALB/c Mice. Parasit. Vectors, 2020, 13, 415. [Google Scholar] [CrossRef]
- Jardim, A.; Hanson, S.; Ullman, B.; McCubbin, W. D.; Kay, C. M.; Olafson, R. W. Cloning and Structure-Function Analysis of the Leishmania Donovani Kinetoplastid Membrane Protein-11. Biochem. J., 1995, 305 Pt 1, 315–320. [Google Scholar] [CrossRef]
- Lacerda, D. I.; Cysne-Finkelstein, L.; Nunes, M. P.; De-Luca, P. M.; Genestra, M. da S.; Leon, L. L. P.; Berrêdo-Pinho, M.; Mendonça-Lima, L.; Matos, D. C. de S.; Medeiros, M. A.; et al. Kinetoplastid Membrane Protein-11 Exacerbates Infection with Leishmania Amazonensis in Murine Macrophages. Mem. Inst. Oswaldo Cruz, 2012, 107, 238–245. [CrossRef]
- Mortazavidehkordi, N.; Fallah, A.; Abdollahi, A.; Kia, V.; Khanahmad, H.; Najafabadi, Z. G.; Hashemi, N.; Estiri, B.; Roudbari, Z.; Najafi, A.; et al. A Lentiviral Vaccine Expressing KMP11-HASPB Fusion Protein Increases Immune Response to Leishmania Major in BALB/C. Parasitol. Res., 2018, 117, 2265–2273. [Google Scholar] [CrossRef]
- Bourreau, E.; Collet, M.; Prévot, G.; Milon, G.; Ashimoff, D.; Hasagewa, H.; Parra-Lopez, C.; Launois, P. IFN-γ-Producing CD45RA+ CD8+ and IL-10-Producing CD45RA– CD4+ T Cells Generated in Response to LACK in Naive Subjects Never Exposed to Leishmania. Eur. J. Immunol., 2002, 32, 510–520. [Google Scholar] [CrossRef]
- Pérez-Jiménez, E.; Kochan, G.; Gherardi, M. M.; Esteban, M. MVA-LACK as a Safe and Efficient Vector for Vaccination against Leishmaniasis. Microbes Infect., 2006, 8, 810–822. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Sacks, D. L.; Brown, D. R.; Reiner, S. L.; Charest, H.; Glaichenhaus, N.; Seder, R. A. Vaccination with DNA Encoding the Immunodominant LACK Parasite Antigen Confers Protective Immunity to Mice Infected with Leishmania Major. J. Exp. Med., 1997, 186, 1137–1147. [Google Scholar] [CrossRef]
- Maasho, K.; Satti, I.; Nylén, S.; Guzman, G.; Koning, F.; Akuffo, H. A Leishmania Homologue of Receptors for Activated C-Kinase (LACK) Induces Both Interferon-Gamma and Interleukin-10 in Natural Killer Cells of Healthy Blood Donors. J. Infect. Dis., 2000, 182, 570–578. [Google Scholar] [CrossRef]
- Bourreau, E.; Pascalis, H.; Prévot, G.; Kariminia, A.; Jolly, N.; Milon, G.; Buffet, P.; Michel, R.; Meynard, J.-B.; Boutin, J.-P.; et al. Increased Production of Interferon-Gamma by Leishmania Homologue of the Mammalian Receptor for Activated C Kinase-Reactive CD4+ T Cells among Human Blood Mononuclear Cells: An Early Marker of Exposure to Leishmania? Scand. J. Immunol., 2003, 58, 201–210. [Google Scholar] [CrossRef]
- Carvalho, L. P.; Passos, S.; Dutra, W. O.; Soto, M.; Alonso, C.; Gollob, K. J.; Carvalho, E. M.; Ribeiro de Jesus, A. Effect of LACK and KMP11 on IFN-Gamma Production by Peripheral Blood Mononuclear Cells from Cutaneous and Mucosal Leishmaniasis Patients. Scand. J. Immunol., 2005, 61, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Azeredo-Coutinho, R. B. G.; Matos, D. C. S.; Armôa, G. G. R.; Maia, R. M.; Schubach, A.; Mayrink, W.; Mendonça, S. C. F. Contrasting Human Cytokine Responses to Promastigote Whole-Cell Extract and the Leishmania Analogue Receptor for Activated C Kinase Antigen of L. Amazonensis in Natural Infection versus Immunization. Clin. Exp. Immunol., 2008, 153, 369–375. [Google Scholar] [CrossRef]
- Stober, C. B.; Jeronimo, S. M. B.; Pontes, N. N.; Miller, E. N.; Blackwell, J. M. Cytokine Responses to Novel Antigens in a Peri-Urban Population in Brazil Exposed to Leishmania Infantum Chagasi. Am. J. Trop. Med. Hyg., 2012, 87, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Julia, V.; Glaichenhaus, N. CD4(+) T Cells Which React to the Leishmania Major LACK Antigen Rapidly Secrete Interleukin-4 and Are Detrimental to the Host in Resistant B10.D2 Mice. Infect. Immun., 1999, 67, 3641–3644. [Google Scholar] [CrossRef]
- Afonso, L. C. C.; Scharton, T. M.; Vieira, L. Q.; Wysocka, M.; Trinchieri, G.; Scott, P. The Adjuvant Effect of Interleukin-12 in a Vaccine Against Leishmania Major. Science, 1994, 263, 235–237. [Google Scholar] [CrossRef]
- Mougneau, E.; Altare, F.; Wakil, A. E.; Zheng, S.; Coppola, T.; Wang, Z. E.; Waldmann, R.; Locksley, R. M.; Glaichenhaus, N. Expression Cloning of a Protective Leishmania Antigen. Science, 1995, 268, 563–566. [Google Scholar] [CrossRef]
- Melby, P. C.; Yang, J.; Zhao, W.; Perez, L. E.; Cheng, J. Leishmania Donovani P36(LACK) DNA Vaccine Is Highly Immunogenic but Not Protective against Experimental Visceral Leishmaniasis. Infect. Immun., 2001, 69, 4719–4725. [Google Scholar] [CrossRef] [PubMed]
- Marques-da-Silva, E. A.; Coelho, E. A. F.; Gomes, D. C. O.; Vilela, M. C.; Masioli, C. Z.; Tavares, C. A. P.; Fernandes, A. P.; Afonso, L. C. C.; Rezende, S. A. Intramuscular Immunization with P36(LACK) DNA Vaccine Induces IFN-γ Production but Does Not Protect BALB/c Mice against Leishmania Chagasi Intravenous Challenge. Parasitol. Res., 2005, 98, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Fernández Cotrina, J.; Iniesta, V.; Monroy, I.; Baz, V.; Hugnet, C.; Marañon, F.; Fabra, M.; Gómez-Nieto, L. C.; Alonso, C. A Large-Scale Field Randomized Trial Demonstrates Safety and Efficacy of the Vaccine LetiFend® against Canine Leishmaniosis. Vaccine, 2018, 36, 1972–1982. [Google Scholar] [CrossRef] [PubMed]
- Sjölander, A.; Baldwin, T. M.; Curtis, J. M.; Bengtsson, K. L.; Handman, E. Vaccination with Recombinant Parasite Surface Antigen 2 from Leishmania Major Induces a Th1 Type of Immune Response but Does Not Protect against Infection. Vaccine, 1998, 16, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Sparwasser, T.; Lipford, G. B. Consequences of Bacterial CpG DNA-Driven Activation of Antigen-Presenting Cells. Curr. Top. Microbiol. Immunol., 2000, 247, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Iborra, S.; Carrión, J.; Anderson, C.; Alonso, C.; Sacks, D.; Soto, M. Vaccination with the Leishmania Infantum Acidic Ribosomal P0 Protein plus CpG Oligodeoxynucleotides Induces Protection against Cutaneous Leishmaniasis in C57BL/6 Mice but Does Not Prevent Progressive Disease in BALB/c Mice. Infect. Immun., 2005, 73, 5842–5852. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.; Alonso, C.; Requena, J. M. The Leishmania Infantum Acidic Ribosomal Protein LiP2a Induces a Prominent Humoral Response in Vivo and Stimulates Cell Proliferation in Vitro and Interferon-Gamma (IFN-Gamma) Production by Murine Splenocytes. Clin. Exp. Immunol., 2000, 122, 212–218. [Google Scholar] [CrossRef]
- Fernandes, A. P.; Coelho, E. A. F.; Machado-Coelho, G. L. L.; Grimaldi, G.; Gazzinelli, R. T. Making an Anti-Amastigote Vaccine for Visceral Leishmaniasis: Rational, Update and Perspectives. Curr. Opin. Microbiol., 2012, 15, 476–485. [Google Scholar] [CrossRef]
- Lage, D. P.; Ludolf, F.; Silveira, P. C.; Machado, A. S.; Ramos, F. F.; Dias, D. S.; Ribeiro, P. A. F.; Costa, L. E.; Vale, D. L.; Tavares, G. S. V.; et al. Screening Diagnostic Candidates from Leishmania Infantum Proteins for Human Visceral Leishmaniasis Using an Immunoproteomics Approach. Parasitology, 2019, 146, 1467–1476. [Google Scholar] [CrossRef]
- Ribeiro, P. A. F.; Dias, D. S.; Lage, D. P.; Martins, V. T.; Costa, L. E.; Santos, T. T. O.; Ramos, F. F.; Tavares, G. S. V.; Mendonça, D. V. C.; Ludolf, F.; et al. Immunogenicity and Protective Efficacy of a New Leishmania Hypothetical Protein Applied as a DNA Vaccine or in a Recombinant Form against Leishmania Infantum Infection. Mol. Immunol., 2019, 106, 108–118. [Google Scholar] [CrossRef]
- Oliveira-da-Silva, J. A.; Machado, A. S.; Tavares, G. S. V.; Ramos, F. F.; Lage, D. P.; Ludolf, F.; Steiner, B. T.; Reis, T. A. R.; Santos, T. T. O.; Costa, L. E.; et al. Biotechnological Applications from a Leishmania Amastigote-Specific Hypothetical Protein in the Canine and Human Visceral Leishmaniasis. Microb. Pathog., 2020, 147, 104283. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C. B. Nucleoside Hydrolase NH 36: A Vital Enzyme for the Leishmania Genus in the Development of T-Cell Epitope Cross-Protective Vaccines. Front. Immunol., 2019, 10, 813. [Google Scholar] [CrossRef]
- Alves-Silva, M. V.; Nico, D.; Morrot, A.; Palatnik, M.; Palatnik-de-Sousa, C. B. A Chimera Containing CD4+ and CD8+ T-Cell Epitopes of the Leishmania Donovani Nucleoside Hydrolase (NH36) Optimizes Cross-Protection against Leishmania Amazonesis Infection. Front. Immunol., 2017, 8, 100. [Google Scholar] [CrossRef]
- Iborra, S.; Soto, M.; Carrión, J.; Nieto, A.; Fernández, E.; Alonso, C.; Requena, J. M. The Leishmania Infantum Acidic Ribosomal Protein P0 Administered as a DNA Vaccine Confers Protective Immunity to Leishmania Major Infection in BALB/c Mice. Infect. Immun., 2003, 71, 6562–6572. [Google Scholar] [CrossRef]
- Chenik, M.; Louzir, H.; Ksontini, H.; Dilou, A.; Abdmouleh, I.; Dellagi, K. Vaccination with the Divergent Portion of the Protein Histone H2B of Leishmania Protects Susceptible BALB/c Mice against a Virulent Challenge with Leishmania Major. Vaccine, 2006, 24, 2521–2529. [Google Scholar] [CrossRef]
- Carrión, J. Mechanisms of Immunity to Leishmania Major Infection in Mice: The Contribution of DNA Vaccines Coding for Two Novel Sets of Histones (H2A–H2B or H3–H4). Comp. Immunol. Microbiol. Infect. Dis., 2011, 34, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Baharia, R. K.; Tandon, R.; Sahasrabuddhe, A. A.; Sundar, S.; Dube, A. Nucleosomal Histone Proteins of L. Donovani: A Combination of Recombinant H2A, H2B, H3 and H4 Proteins Were Highly Immunogenic and Offered Optimum Prophylactic Efficacy against Leishmania Challenge in Hamsters. PLOS ONE, 2014, 9, e97911. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Ghoshal, A.; Mandal, C.; Shaha, C. Leishmania Cell Surface Prohibitin: Role in Host-Parasite Interaction. Cell. Microbiol., 2010, 12, 432–452. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.; Ko, D.; Ordonez-Ercan, D.; Chellappan, S. P. A Putative Coiled-Coil Domain of Prohibitin Is Sufficient to Repress E2F1-Mediated Transcription and Induce Apoptosis. Biochem. Biophys. Res. Commun., 2003, 312, 459–466. [Google Scholar] [CrossRef]
- Lage, D. P.; Ribeiro, P. A. F.; Dias, D. S.; Mendonça, D. V. C.; Ramos, F. F.; Carvalho, L. M.; de Oliveira, D.; Steiner, B. T.; Martins, V. T.; Perin, L.; et al. A Candidate Vaccine for Human Visceral Leishmaniasis Based on a Specific T Cell Epitope-Containing Chimeric Protein Protects Mice against Leishmania Infantum Infection. NPJ Vaccines, 2020, 5, 75. [Google Scholar] [CrossRef]
- Dias, D. S.; Ribeiro, P. A. F.; Martins, V. T.; Lage, D. P.; Ramos, F. F.; Dias, A. L. T.; Rodrigues, M. R.; Portela, Á. S. B.; Costa, L. E.; Caligiorne, R. B.; et al. Recombinant Prohibitin Protein of Leishmania Infantum Acts as a Vaccine Candidate and Diagnostic Marker against Visceral Leishmaniasis. Cell. Immunol., 2018, 323, 59–69. [Google Scholar] [CrossRef]
- Kedzierski, L.; Montgomery, J.; Bullen, D.; Curtis, J.; Gardiner, E.; Jimenez-Ruiz, A.; Handman, E. A Leucine-Rich Repeat Motif of Leishmania Parasite Surface Antigen 2 Binds to Macrophages through the Complement Receptor 3. J. Immunol. Baltim. Md 1950, 2004, 172, 4902–4906. [Google Scholar] [CrossRef]
- Lincoln, L. M.; Ozaki, M.; Donelson, J. E.; Beetham, J. K. Genetic Complementation of Leishmania Deficient in PSA (GP46) Restores Their Resistance to Lysis by Complement. Mol. Biochem. Parasitol., 2004, 137, 185–189. [Google Scholar] [CrossRef]
- Chamakh-Ayari, R.; Bras-Gonçalves, R.; Bahi-Jaber, N.; Petitdidier, E.; Markikou-Ouni, W.; Aoun, K.; Moreno, J.; Carrillo, E.; Salotra, P.; Kaushal, H.; et al. In Vitro Evaluation of a Soluble Leishmania Promastigote Surface Antigen as a Potential Vaccine Candidate against Human Leishmaniasis. PLoS ONE, 2014, 9, e92708. [Google Scholar] [CrossRef] [PubMed]
- Handman, E.; Osborn, A. H.; Symons, F.; van Driel, R.; Cappai, R. The Leishmania Promastigote Surface Antigen 2 Complex Is Differentially Expressed during the Parasite Life Cycle. Mol. Biochem. Parasitol., 1995, 74, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Kemp, M.; Handman, E.; Kemp, K.; Ismail, A.; Mustafa, M. D.; Kordofani, A. Y.; Bendtzen, K.; Kharazmi, A.; Theander, T. G. The Leishmania Promastigote Surface Antigen-2 (PSA-2) Is Specifically Recognised by Th1 Cells in Humans with Naturally Acquired Immunity to L. Major. FEMS Immunol. Med. Microbiol., 1998, 20, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Petitdidier, E.; Pagniez, J.; Papierok, G.; Vincendeau, P.; Lemesre, J.-L.; Bras-Gonçalves, R. Recombinant Forms of Leishmania Amazonensis Excreted/Secreted Promastigote Surface Antigen (PSA) Induce Protective Immune Responses in Dogs. PLoS Negl. Trop. Dis., 2016, 10, e0004614. [Google Scholar] [CrossRef] [PubMed]
- Ommen, G.; Chrobak, M.; Clos, J. The Co-Chaperone SGT of Leishmania Donovani Is Essential for the Parasite’s Viability. Cell Stress Chaperones, 2010, 15, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Dias, D. S.; Ribeiro, P. A. F.; Martins, V. T.; Lage, D. P.; Portela, Á. S. B.; Costa, L. E.; Salles, B. C. S.; Lima, M. P.; Ramos, F. F.; Santos, T. T. O.; et al. Recombinant Small Glutamine-Rich Tetratricopeptide Repeat-Containing Protein of Leishmania Infantum: Potential Vaccine and Diagnostic Application against Visceral Leishmaniasis. Mol. Immunol., 2017, 91, 272–281. [Google Scholar] [CrossRef]
- Zhang, W.-W.; Matlashewski, G. Loss of Virulence in Leishmania Donovani Deficient in an Amastigote-Specific Protein, A2. Proc. Natl. Acad. Sci. U. S. A., 1997, 94, 8807–8811. [Google Scholar] [CrossRef]
- Almeida, A. P. M. M.; Machado, L. F. M.; Doro, D.; Nascimento, F. C.; Damasceno, L.; Gazzinelli, R. T.; Fernandes, A. P.; Junqueira, C. New Vaccine Formulations Containing a Modified Version of the Amastigote 2 Antigen and the Non-Virulent Trypanosoma Cruzi CL-14 Strain Are Highly Antigenic and Protective against Leishmania Infantum Challenge. Front. Immunol., 2018, 9, 465. [Google Scholar] [CrossRef]
- Toepp, A.; Larson, M.; Wilson, G.; Grinnage-Pulley, T.; Bennett, C.; Leal-Lima, A.; Anderson, B.; Parrish, M.; Anderson, M.; Fowler, H.; et al. Randomized, Controlled, Double-Blinded Field Trial to Assess Leishmania Vaccine Effectiveness as Immunotherapy for Canine Leishmaniosis. Vaccine, 2018, 36, 6433–6441. [Google Scholar] [CrossRef]
- Azam, S. S.; Abro, A.; Raza, S.; Saroosh, A. Structure and Dynamics Studies of Sterol 24-C-Methyltransferase with Mechanism Based Inactivators for the Disruption of Ergosterol Biosynthesis. Mol. Biol. Rep., 2014, 41, 4279–4293. [Google Scholar] [CrossRef] [PubMed]
- Palatnik-de-Sousa, C. B.; Nico, D. The Delay in the Licensing of Protozoal Vaccines: A Comparative History. Front. Immunol., 2020, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Brito, R. C. F.; Guimarães, F. G.; Velloso, J. P. L.; Corrêa-Oliveira, R.; Ruiz, J. C.; Reis, A. B.; Resende, D. M. Immunoinformatics Features Linked to Leishmania Vaccine Development: Data Integration of Experimental and In Silico Studies. Int. J. Mol. Sci., 2017, 18, 371. [Google Scholar] [CrossRef] [PubMed]
- Khan, M. A. A.; Ami, J. Q.; Faisal, K.; Chowdhury, R.; Ghosh, P.; Hossain, F.; Abd El Wahed, A.; Mondal, D. An Immunoinformatic Approach Driven by Experimental Proteomics: In Silico Design of a Subunit Candidate Vaccine Targeting Secretory Proteins of Leishmania Donovani Amastigotes. Parasit. Vectors, 2020, 13, 196. [Google Scholar] [CrossRef]
- Brasil. Manual de vigilância e controle da leishmaniose visceral - 1a Edição.
Figure 1.
Schematic picture of promastigote and amastigote forms of Leishmania spp. and location of proteins described in patents related to vaccines composed of chimeric proteins against visceral leishmaniasis.
Figure 1.
Schematic picture of promastigote and amastigote forms of Leishmania spp. and location of proteins described in patents related to vaccines composed of chimeric proteins against visceral leishmaniasis.
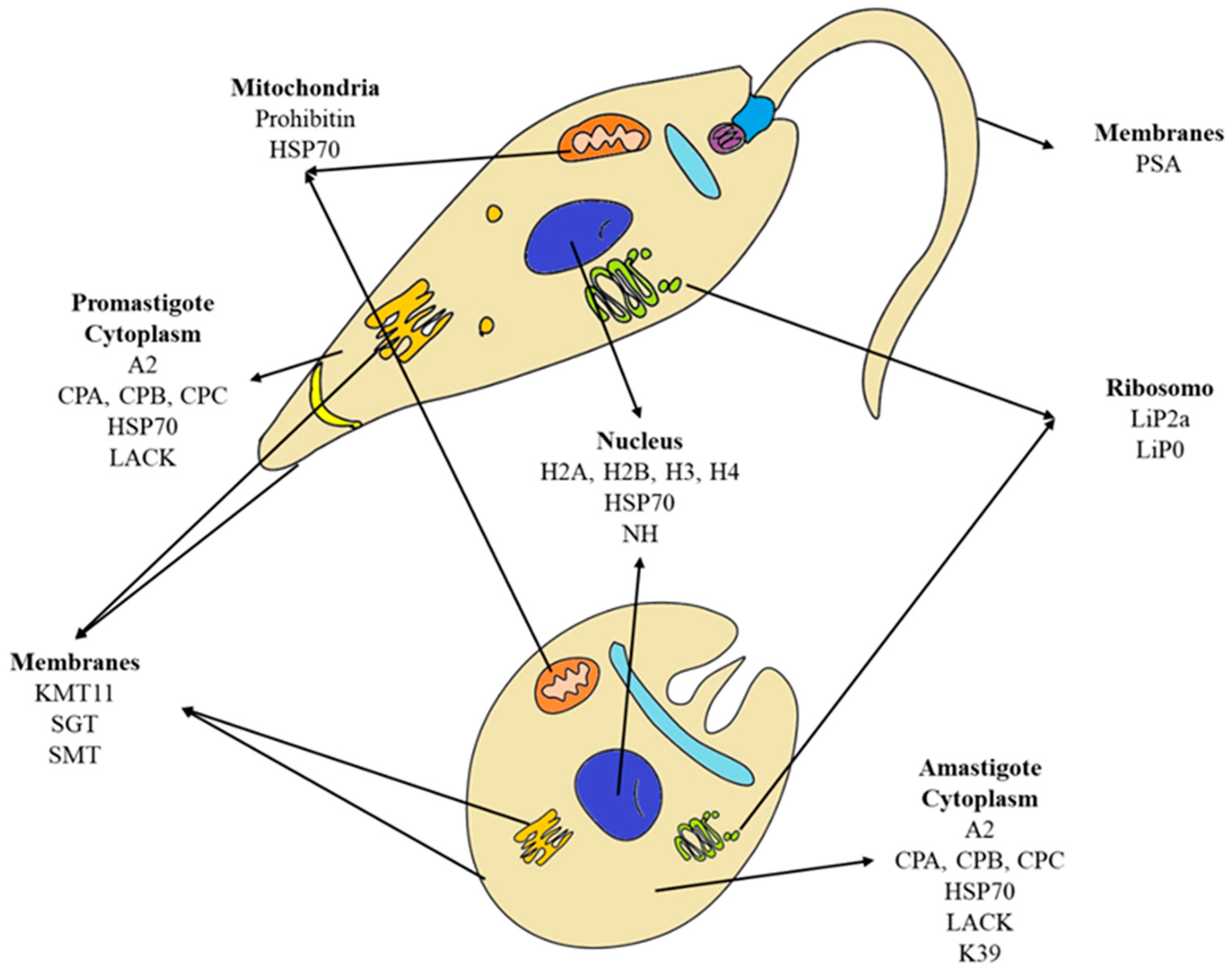

Table 1.
Identification of patents related to vaccines composed of chimeric proteins to VL.
| Original patent title | Publication number | Priority Country | Deposit year | Chimera composition | In silico analysis | In vitro analysis | In vivo analysis |
|---|---|---|---|---|---|---|---|
| “Proteína quimérica, kit, método para diagnóstico de leishmaniose, uso de proteína quimérica, composição vacinal contra leishmaniose visceral, e, uso de uma composição vacinal” | BR 10 2021 00079 4 | Brazil | 2021 | A2 and K39 | - | ↑IFN-γ in spleen cell culture | ↑IgG e IgG2, in BALB/c mice. ↓spleen and liver, parasite load by limiting dilution, in BALB/c mice |
| “Quimera sintética multiepitópica como vacuna y tratamiento frente a leishmaniosis en mamíferos” | ES2795149 | Spain | 2020 | H2A, H2B, H3, and H4 | Human and mice MHC* class I and II alleles prediction | ↑IFN-γ and IL-12 after culture stimulation. ↑leishmanicidal effect in infected BMDC** |
↓spleen and liver, parasite load by limiting dilution, in BALB/c mice |
| “Vacinas compostas de proteínas quiméricas poliepítopos contra a leishmaniose visceral humana e/ou canina” | BR 10 2018 008197 7 | Brazil | 2018 | VAC-1: H2A, LACK LiP2a, LiP0, and CPC | MHC class I and II alleles prediction | ↑IFN-γ, TNF-α, CD4+ T cells, and CD8+ T cells, after culture stimulation ↑CD4+ T lymphocytes with central memory phenotype in VAC-1 and VAC-2 ↑CD8+ T lymphocytes with central memory phenotype in VAC-1 and VAC-2. Effector memory phenotype in VAC-1 |
↓spleen parasite load by Real Time PCR (qPCR), in BALB/c mice |
| VAC-2: CPA, CPB, PSA-50S, and A2 | |||||||
| “Proteína quimérica recombinante, vacina contra leishmanioses e uso” | BR 10 2017 025621 9 | Brazil | 2017 | Prohibitin; SGT; LiHyp5 | Human MHC class I and II alleles prediction | ↑ IFN-γ, IL-12, and GM-CSF in spleen cell culture ↑ PBMCs proliferation in human and dog cells |
↓spleen, liver, draining lymph nodes, and bone marrow parasite load by limiting dilution, in BALB/c mice |
| “Proteína quimérica, composição vacinal contra leishmanioses e usos” | BR 10 2016 006121 0 | Brazil | 2016 | LiHyp1, LiHyp6, LiHyV, and HRF | Human and mice MHC class I and II alleles prediction | ↑ IFN-γ, IL-12, and GM-CSF in spleen cell culture | ↓spleen, liver, draining lymph nodes, and bone marrow parasite load by limiting dilution, in BALB/c mice |
| Recombinant polyprotein vaccines for the treatment and diagnosis of leishmaniasis | US20130177584 | United States | 2013 | KSA (KM11, SMT e A2) | - | - | ↓liver by limiting dilution, in C57BL/6 (L. infantum challenge), and BALB/c mice (L. donovani challenge) |
| Vaccines comprising leishmania polypeptides for the treatment and diagnosis of leishmaniasis | WO 2014/160987 | United States | 2013 | NS and NSC | - | ↑IFN-γ in spleen cell culture | ↓liver by qPCR, in BALB/c mice |
| “Quimera multicomponente para su uso como vacuna frente a la infección por Leishmania spp. En mamíferos” | WO 2013/110824 | Spain | 2011 | HISA70 (H2A, H2B, H3, H4, A2, HSP70) | - | - | ↓spleen and liver by limiting dilution, in BALB/c mice |
| Vaccines comprising non-specific nucleoside hydrolase and sterol 24-c-methyltransferase (SMT) polypeptides for the treatment and diagnosis of Leishmaniasis | WO 2012064659 | United States | 2010 | NS | - | ↑IFN-γ by spleen cell culture, in BALB/c mice ↑IFN-γ, and ↑IgG in non-human primates |
↓liver by limiting dilution, in BALB/c mice ↑IgG1 and IgG2 titration |
The arrows (↓ and ↑) indicate the decrease and increase in biomarker levels and/or parasite load. * Major Histocompatibility complex (MHC). ** Bone marrow-derived dendritic cells (BMDC). A2: specific amastigote protein; CPA, CPB, and CPC: cysteine peptidase proteins; HRF: IgE-dependent histamine-releasing factor; HSP70: heat shock protein; H2A, H2B, H3, H4: nucleosomal histones; K39: kinesin-related protein; KSA and KSAC: kinetoplastid membrane protein (KM11), sterol 24-c-methyltransferase (SMT), specific amastigote protein A2 and cysteine peptidase proteins B; LACK: activated kinase C receptor homologous Leishmania protein; LiHyp 1, LiHyp5, LiHyp6, and LiHyV: hypothetical protein; LiP2a and LiP0: acidic ribosomal protein; NS: nonspecific nucleoside hydrolase (NH) and sterol 24-c-methyltransferase (SMT); NSC: nonspecific nucleoside hydrolase (NH), sterol 24-c-methyltransferase (SMT) and cysteine polypeptidase B (CPB); Prohibitin: surface protein; PSA-50S: promastigote surface antigen; SGT: Small glutamine-rich TPR proteins.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



