
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
We investigated the interaction between the insect-specific virus, Piura virus (PIUV) and the arbovirus Zika virus (ZIKV) in Aedes albopictus cells. We performed coinfection experiments in C6/36 cells. Piura virus (Cor 33 strain, Colombia) and ZIKV (PRVABC58 strain, Puerto Rico) were co-inoculated into C6/36 cells using two MOI combinations: 0.1 for both viruses and 1.0 for ZIKV, 0.1 for PIUV. Wells were infected in triplicate with either PIUV and ZIKV coinfection, ZIKV-only, or PIUV-only. Mock infected cells served as control wells. The cell suspension was collected daily through 7 days post-infection. Zika virus load was titrated by TCID50 on Vero 76 cells. The ZIKV-only infection and PIUV and ZIKV coinfection experiments were also quantified by RT-qPCR. We also investigated whether ZIKV interfered in the PIUV replication. PIUV suppressed the replication of ZIKV resulting in a 10,000-fold reduction in ZIKV titers within 3 days post-infection. PIUV viral loads were not reduced in the presence of ZIKV. We conclude that, when concurrently infected, PIUV suppresses ZIKV in C6/36 cells while ZIKV does not interfere in PIUV replication.
Keywords:
Zika virus
; Piura virus
; coinfection
1. Introduction
Arboviruses are viruses transmitted to vertebrates that may cause disease in humans and animals leading to outbreaks and epidemics. Dengue (DENV), Zika (ZIKV), and Chikungunya (CHIKV) viruses are primarily adapted to humans and these three viruses have extended their geographic range due to the dissemination of Ae. aegypti and adaptation to the ubiquitous Ae. albopictus [1,2]. ZIKV, of the family Flaviviridae, genus Orthoflavivirus, caused several outbreaks in the Pacific Islands between 2007 and 2014 and, starting in 2015, causing an epidemic of febrile illness associated with microcephaly in infants and Guillain-Barré Syndrome in many countries. With transmission reported in 86 countries, ZIKV poses an ongoing threat to the developing fetus by causing neurological impairment with congenital infection (congenital ZIKV syndrome) [1,2,3,4]. Although the cases of ZIKV congenital syndrome are rare at this time, the virus is still circulating and there is no approved therapeutics or vaccine to treat or prevent the infection. Given the everexpanding range of Ae. aegypti and Ae. albopictus and increased frequency of outbreaks of anthrophilic arboviruses, it is important to maintain active surveillance and develop interventional strategies for control of outbreaks [2].
Insect-specific viruses (ISVs) are a diverse group of viruses that naturally infect hematophagous insects such as mosquitoes, ticks, phlebotomine sandflies, and others. Unlike arboviruses, ISVs do not appear to replicate in vertebrates and their cells but instead infect only invertebrates [5,6]. ISVs are mainly transmitted vertically and may share the same mosquito vector with arboviruses. Some ISVs, mainly those belonging to the viral families Togaviridae and Flaviviridae, are phylogenetically related to certain arboviruses, indicating that ISVs may be the ancestors of the arboviruses gaining the capacity to infect vertebrates or vice versa [5,6].
While some ISVs are more phylogenetically related to arboviruses, others are closer to plant viruses, such as those belonging to the taxon Negevirus and the family Tymoviridae [7,8]. The taxon Negeviruses comprises several viruses which are classified into two genera: Nelorpivirus and Sandewavirus [9]. Negeviruses replicate well in mosquito and sandfly cell lines, reaching high titers, demonstrating that these viruses are well adapted to these insect cells [10]. Phylogenetically, negeviruses are more related to plant-infecting viruses of the genera Cilevirus, Higrevirus, and Blunervirus. In addition to these ISVs demonstrating phylogenetic similarities to plant viruses, viruses have been recently isolated from plants that are more similar to Negeviruses or “nege-like”; future work with these and yet to be discover negeviruses will contribute to our understanding of the evolution of these viruses [10,11,12,13,14]. Furthermore, metagenomics conducted on the mosquito virome will likely identify new ISVs as well as add to our knowledge of the worldwide distribution of these viruses [10,15].
One of the viruses belonging to the genus Nelorpivirus is Piura virus (PIUV), which was first isolated from Culex sp. mosquitoes in the city of Piura in Peru in 1996. In the subsequent years, PIUV was detected in Mexico, Colombia, and the United States. The virus has been isolated from mosquitoes of different genera such as Anopheles, Psorophora, Aedes, Uranotaeneia, Lutzomyia, and Culicidae. PIUV has three proteins encoded in three open reading frames (ORF) called ORF 1, ORF 2, and ORF 3. Phylogenetically, PIUV is more related to Brejeira virus, isolated in Brazil [10].
Recent studies have investigated how ISVs and arboviruses interact within insects. This previous work demonstrated that certain ISVs, mainly orthoflaviviruses and alphaviruses, inhibit arbovirus replication in cells and reduce vector competence [16,17,18,19]. Bolling and colleagues (2012) conducted experiments using a laboratory colony of Culex pipiens mosquitoes naturally infected with Culex flavivirus (CxFV). The mosquitoes were infected with West Nile virus (WNV) and, then the authors evaluated the mosquitoes’ vector competence indicating that the suppression of WNV replication was caused by the persistent previous infection with CxFV.
This observation, that ISVs in modulate the replication of arboviruses highlights their potential as a future tool in the biological control of these viruses. ISVs have been used as the backbone for the development of vaccines against arboviruses. In particular, a candidate Chikungunya vaccine using the Eilat virus as a backbone was 100% protective in two different mouse models and blocked viremia in infected nonhuman primates [20]. Other chimera’s developed with this backbone include the inclusion of structural genes from Eastern Equine Encephalitis virus (EEEV) and Venezuelan equine encephalitis virus (VEEV) [17]. These ISV chimera’s also have potential for use as diagnostic reagents allowing work on highly pathogenic reagents at biosafety level-1 [21,22].
Despite these studies showing the potential biotechnological use of orthoflaviviruses and alphaviruses ISVs, little information is known about negeviruses and arboviruses interaction. One recently published study showed that the negeviruses Negev (NEGV) and PIUV inhibited VEEV virus and CHIKV replication in mosquito cells [23]. To further understand the interaction of PIUV with arboviruses, we investigated the interaction between the PIUV, and Zika virus (ZIKV) in Ae. albopictus cells. We hypothesized, that like the interaction observed between ISV and alphaviruses, PIUV would inhibit the replication of vertebrate viruses. To this end, we investigated the interactions between Piura and Zika viruses in vitro.
2. Materials and Methods
2.1. Cells and viruses
African green monkey kidney cells (Vero 76) (ATCC, Manassas, VA) and Ae. albopictus cells (clone C6/36) (ATCC, Manassas, VA) were used for virus infections. Vero 76 cells were cultured in media consisting of advanced DMEM (Gibco, Waltham, Massachusetts, United States) supplemented with 5% Fetal Bovine Serum (FBS), 1% Hepes, 1% Penicillin/Streptomycin, 0.5% amphotericin and maintained at 37°C in 5% CO2. The C6/36 cells were cultured in DMEM (1x) (Gibco, Waltham, Massachusetts, United States) supplemented with 10% FBS, 1% Glutamine, 1% Sodium Pyruvate, 1% Tryptose Phosphate Broth, 1% MEM Non-Essential amino acids, 1% Penicillin/Streptomycin, 0.5% amphotericin. C6/36 cells were maintained at 28°C in 5% CO2.
ZIKV (strain PRVABC59, accession number KU501215.1) and PIUV (strain Cor 33, accession number KX518787.1) (both at passage 3) were grown in Vero 76 and C6/36, respectively. The cell suspension was harvested when approximately 80% cytopathic effect (CPE) was observed. The cell suspension deriving from ZIKV and PIUV stock was concentrated using a Centricon Plus-70 Centrifugal Filter (Millipore, Burlington, MA). Virus infection was titrated by TCID50 for ZIKV and PIUV on Vero 76 cells and C6/36 cells, and ZIKV and PIUV titers were 107 TCID50/mL and 1010 TCID50/mL, respectively [24].
2.2. Coinfection of ZIKV and PIUV
Confluent C6/36 cells seeded in 24-well plates were inoculated with PIUV and ZIKV, PIUV only, or ZIKV only. We used two MOI combinations consisting of 0.1 for both viruses and then a combination of 1.0 for ZIKV and 0.1 for PIUV. Wells were inoculated in triplicate in the C6/36 cells. Negative controls consisted of mock-challenged cells. The cell suspension was collected daily through 7 days post-infection and frozen at −80°C. ZIKV viral load was determined for each well using a TCID50 performed in Vero 76 cells to determine if PIUV interfered in ZIKV replication.
2.3. TCID50 (Tissue Culture Infectious Dose 50%)
Vero 76 cells were seeded in 48-well plates. When the cells were confluent, the media was removed and 100 uL of 10x serial dilutions was inoculated into four wells for each dilution. The plates were incubated at 37°C for one hour; plates were rocked every 15 minutes to distribute virus across the cell monolayer. After incubation, 0.5 mL of Advanced DMEM maintenance media with 2% FBS was added to each well. The plates were incubated at 37°C and monitored for CPE for seven days.
On day 7 post-inoculation, the plates were fixed using cold 25% methanol for 10 minutes. The methanol was removed, and the cells were stained with 0.5% crystal violet and incubated at room temperature for 30 minutes. The plates were rinsed in water and then allowed to dry. The CPE was graded in each well and the TCID50 calculated using the Reed-Muench method (1938) [24].
2.4. Intracellular replication
To assess the effect of PIUV on ZIKV intracellular replication, cells were co-infected with PIUV and ZIKV, both at a MOI of 0.1, in confluent C6/36 cells. These viruses were inoculated in triplicate; mock-challenged wells and positive controls were also included. Cells were collected every 24 hours post-inoculation for 72 hours. At each time point, the supernatant was removed, the cells were washed with PBS and then guanidinium-isothiocyanate-chloroform (TRIzol Reagent, Invitrogen, Carlsbad, CA, USA) was added directly to the cells. The cells were scraped and RNA was extracted as previously described [25]. The RNA was quantified in each sample by spectrophotometry (Nanodrop, Thermo, MA, United States). Real-time RT-qPCR was used to compare viral loads.
Indirect immunofluorescence was used to detect viruses in the cells. For this, we seeded C6/36 cells into culture slides (Millicell EZ Slide, Millipore, Burlington, MA, USA) and inoculated the cells with the viruses as previously described. Cells were fixed every 24 hours post-inoculation for 72 hours. After fixation, anti-orthoflavivirus antibody (4G2) was applied to each well and incubated for 30 minutes at 28°C. After washing, goat anti-mouse IgG Fc-FITC (Southern Biotech, Birmingham, AL, USA) was added to each well. Finally, the cells were stained with DAPI (BD Pharmingen, San Diego, CA, USA) and read with fluorescence microscope (all-in-one, BZ-X810) (Keyence, Osaka, Japan).
2.5. ZIKV and PIUV RNA Quantification
ZIKV RNA from the ZIKV-only and PIUV/ZIKV coinfection wells were tested by RT-qPCR one-step RT-qPCR protocol for ZIKV using the forward primer (5’ CCGCTGCCCAACACAAG 3’), reverse primer (5’ CCACTAACGTTCTTTTGCAGACAT 3’) and probe (5’ FAM- AGCCTACCTTGACAAGCAGTCAGACACTCAA – BHQ-1 3’) previously described [26].
To investigate whether ZIKV interfered with PIUV replication, we developed an RT-qPCR for PIUV. First, we sequenced the PIUV full-length genome (Cor 33_TVP 20159 strain/Genbank accession number: KX518787) and used Primer3 software to design several PIUV primers and probes. Oligonucleotides were synthesized by LGC Biosearch Technologies (Petaluma, CA, USA). Primers were first optimized using SYBR green chemistry (Fast SYBR Green Master Mix; Applied Biosystems, Foster City, CA). Once primer specificity and concentrations of each were optimized against the PIUV Cor 33 strain, a probe-based system was developed (TaqMan, Applied Biosystems, Foster City, CA).
Based on this initial optimization, the following primers and probes were utilized and included the forward primer CO_Cor33-F1, ATCGGTGCGAATACCATAGC and reverse primer CO_Cor33-R1, GTTTACTCCTGACCCCGTGA. The probe sequence CO_Cor33_Probe was FAM-ATCTTTCGTGGATACCGTGC-BHQ-1. The product length of the RT-PCR is 178 bp and the target is the ORF 1 (hypothetical protein 1). The total reaction volume was 20 uL consisting of 1.6 uL forward primer (400 nM), 1.6 uL reverse primer (400 nM), 5 uL TaqMan Fast Virus 1-Step Master Mix, 1 uL probe (250 nM), 8.8 uL nuclease-free water and 2 uL RNA template.
We developed a one-step RT-PCR protocol using the following cycling conditions: 5 minutes at 50 °C (reverse transcription) and 20 seconds at 95 °C (RT inactivation/initial denaturation) followed by 40 cycles of denaturation/ anneal/extension (3 seconds at 95 °C and 30 seconds at 60 °C). To determine the reaction efficiency and linearity, a standard curve was prepared using 10-fold dilution (10-1 to 10-5) of the PIUV RNA (1010 TCID50/mL, 1.17x1015 copies of RNA) in triplicate. Our newly developed PIUV RT-qPCR assay showed efficiency of 96 %, a correlation coefficient (R2) of 0.996, and a slope value of -3.408.
2.6. Statistics
The TCID50 and RT-qPCR results were analyzed using a two-way ANOVA using posteriori Bonferroni correction for multiple comparisons using commercially available statistics software (GraphPad Prism 9.3.1).
3. Results
3.1. PIUV causes intense CPE in C6/36 cells
In our initial experiments, we performed a pilot study with PIUV at different MOI (0.1, 1, 3, 5) in C6/36 cells and we observed that PIUV caused approximately 90% of CPE by day 1 post-infection at all MOI, thus a co-infection experimental design was pursued. Suprainfection was not possible due to the highly cytopathic nature of PIUV in C6/36 cells.
3.2. PIUV inhibits ZIKV replication in C6/36, but ZIKV does not interfere with PIUV replication
To determine if PIUV interferes with ZIKV replication and vice versa, we performed coinfection experiments in C6/36 cells using MOI 0.1 for both viruses. Our results showed that when PIUV is present, ZIKV replication is suppressed up to a 10,000-fold reduction in titers by 3 days post-infection (p < 0.0001) (Figure 1A). The RT-qPCR also showed a reduction in ZIKV viral load in the coinfection samples (p<0.0001) (Figure 1B). PIUV viral loads were not reduced in the co-infection wells compared to the PIUV-only wells (Figure 1C).
3.3. PIUV inhibition of ZIKV replication is dose-dependent
To determine if ZIKV cultivated at a greater MOI is inhibited by PIUV, we performed co-infection experiments with ZIKV at a MOI of 1 and PIUV at a MOI of 0.1. Our results showed that PIUV inhibition of ZIKV was sustained and resulted in a 1,000-fold reduction in ZIKV titers by 3 days post-infection (p < 0.0001) (Figure 2A). While the inhibition was less at a 10x higher dose, the difference was still significant. Viral transcripts of ZIKV were also significantly reduced in the coinfection samples (p < 0.0001) (Figure 2B). On the other hand, PIUV viral loads, as detected by RT-qPCR were not reduced in the co-infection wells compared to the PIUV-only wells (most of the time points were non-significant) (Figure 2C). These findings indicate that the interference of PIUV in ZIKV replication is dose-dependent.
3.4. PIUV CPE was predominant in PIUV/ZIKV coinfection
Both viruses caused CPE in C6/36 cells. ZIKV caused the formation of clumps of cells with consecutive cell destruction. PIUV induced rapid cell death characterized by a decrease in cell size and destruction of the cell monolayer. The predominant CPE visualized in co-infected C6/36 cells was that of PIUV, which consisted of rapid cell shrinkage and cellular death without clumping by the first day of infection (Figure 3).
3.5. PIUV does not inhibit ZIKV cellular entry and likely inhibits intracellular replication in C6/36 cells
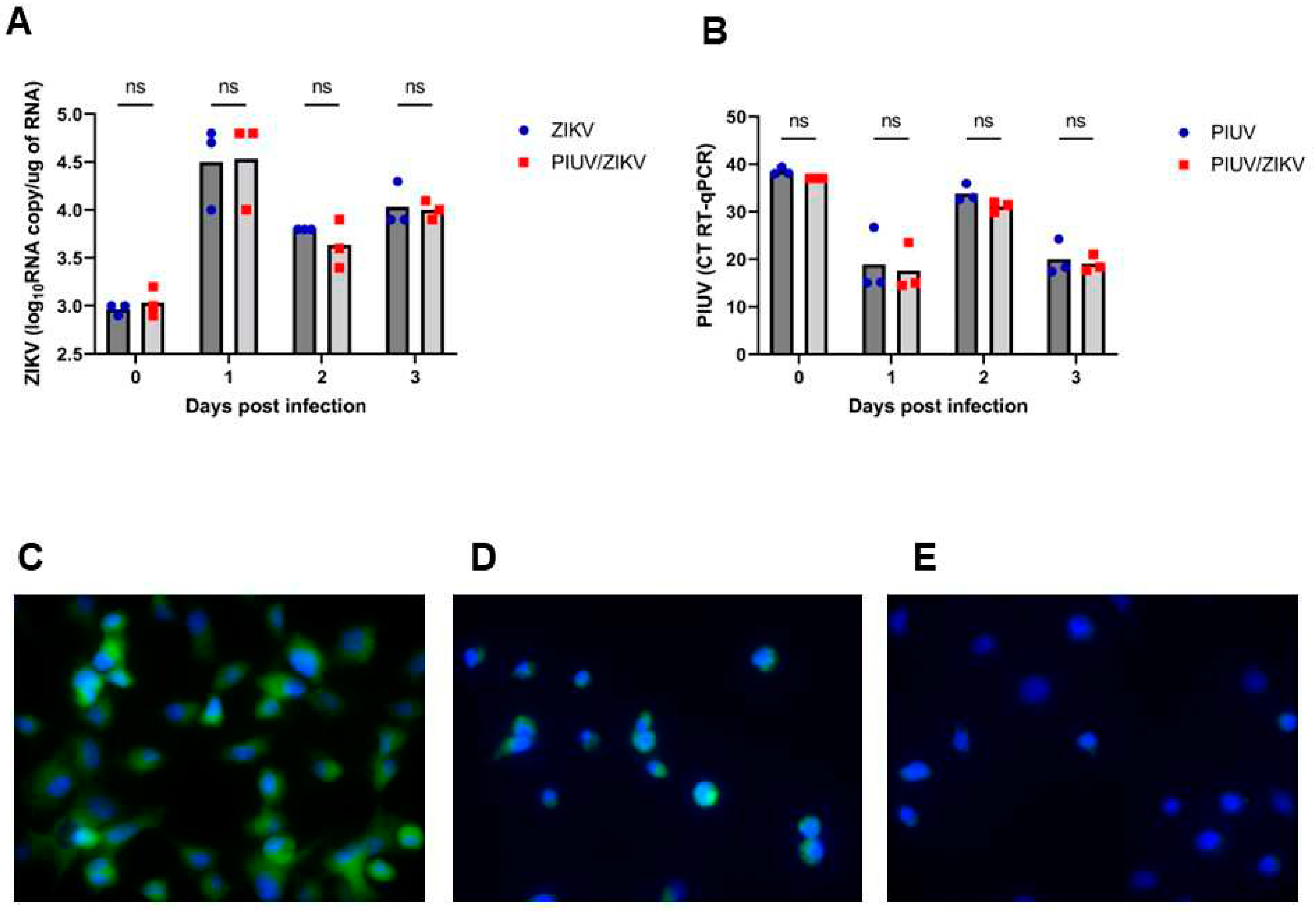
We performed experiments to determine the stage at which PIUV interfered with ZIKV infection. To investigate whether PIUV interference occurred at the level of the intracellular replication of ZIKV, we examined cellular viral load every 24 hours over 3 days by removing the supernatant followed by rinsing the cells and adding TriZol directly on the cells (see section 2.4). There was no difference in the ZIKV load between cells infected only with ZIKV and those coinfected with ZIKV/PIUV and in the PIUV load between infected with “PIUV-only” in comparison with ZIKV/PIUV coinfection (Figure 4A, 4B). The results of immunofluorescence using the anti-orthoflavivirus antibody (4G2) showed the presence of ZIKV antigen in the cytoplasm of both cells infected only with ZIKV and ZIKV/PIUV co-infected cells, however, there was greater fluorescence intensity in the cells infected only by ZIKV (Figure 4C, 4D, 4E).
4. Discussion
Our study showed that PIUV inhibits ZIKV replication when both viruses are simultaneously inoculated in C6/36 cells. These findings are consistent with the evolutive hypothesis that ISVs originated before arboviruses, since ISVs cannot replicate in vertebrates. ISVs likely have a long-term coexistence with mosquitoes and they are well adapted to them, which may explain their replicative advantage in comparison with many arboviruses. These results are promising since this provides initial data demonstrating that PIUV has potential as a biological control agent against the ZIKV.
Our study is the first or one of the first to report the interference of a negevirus in the replication cycle of an arbovirus belonging to the family Flaviviridae. In our experiments, PIUV caused intense CPE in C6/36 cells 24 hours post-infection, which precluded the ability to perform superinfection experiments. This intense pathogenicity has been shown by others wherein Negeviruses reached very high titers in mosquito cells such as C6/36 cells with rapid cell death and loss of the monolayer [10,12]. In previous work, other ISVs belonging mainly to the Flaviviridae and Togaviridae families, inhibited the replication of arboviruses from the same viral families. Limited information exists regarding the interaction between ISVs of the Negevirus genus and those from other arboviral families. A recent study showed for the first time that negeviruses such as Negev virus (NEGV) and PIUV strain EVG 7-47 (PIUV-Culex) (isolated from a pool of Culex nigripalpus mosquitoes from Florida, USA in 2013) induced superinfection exclusion of the arboviruses VEEV and CHIKV (Togaviridae, Alphavirus) in mosquito cells [23].
We performed coinfection experiments (concomitant infection) with PIUV and ZIKV in Ae. albopictus cells each at an MOI of 0.1. PIUV suppressed ZIKV replication in these mosquito cells, as ZIKV titers decreased 10,000-fold when it was in the presence of PIUV. Interestingly, we did not observe interference in the PIUV replication by ZIKV. Bolling and colleagues (2012), investigating the relationship of the CxFV and WNV, also observed that ISVs decreased WNV titers, while the WNV did not interfere with CxFV titers [16].
We observed that the inhibition in the ZIKV replication is dose-dependent even with a 10-fold higher MOI as there was a significant reduction in the ZIKV titers, although less inhibition was present at the lower MOI of 0.1 for both viruses. The dominance was also evident phenotypically since the primary CPE was most similar to PIUV CPE in the co-infection experiments. The study performed by Patterson and cols. [23] demonstrated that PIUV reduced CHIKV titers between 2.4 log10 and 5.3 log10 PFU/mL, also leading to significant reductions in the VEEV titers. Our results and those reported by Patterson and cols. [23], show the heterologous interference of negeviruses, especially PIUV in the replication cycle of arboviruses from the families Flaviviridae and Togaviridae. In other studies of homologous interference, Nhumirim virus (NHUV) of the Flaviviridae family inhibited DENV serotype 2 (DENV-2), ZIKV, and WNV by 4, 5, and 6 logs, respectively. No interference of NHUV replication occurred with co-infection [18,27,28]. Another study demonstrated that the ISV Palm Creek (Orthoflavivirus) interfered in WNV and Murray Valley encephalitis virus in vitro [29] and, in WNV transmission in Culex annulirostris mosquitoes [30].
Once it was verified that PIUV inhibits cellular replication of ZIKV, we investigated if this interference occurred at the level of the entry or beyond in the replication cycle. Our results showed that ZIKV still enters mosquito cells, suggesting that the interference is an intracellular event. These results point to possible interference of the PIUV at some stage of replication other than the synthesis of the genome since intracellularly, the virus load was similar between the ZIKV-only and the co-infection cells. Since the viral load measured in the supernatant combined with cells was significantly less with co-infection, inhibition likely occurs during downstream events such as ZIKV assembly, release, protein synthesis, or other stages.
Future studies are important to clarify which mechanisms are involved in the interference process and confirm these finding in vivo. In particular, given the defective RNAi pathway in C6/36 cells, it is necessary to perform experimental infections in mosquitoes in vivo to analyze differences in the vector competence, even though many studies have shown concordance of results between in vitro and in vivo experiments.
Further, although most of the studies show that several insect-specific viruses inhibit the replication of arboviruses in mosquitoes and their cells, each case must be evaluated carefully, considering the different species of mosquitoes, insects from different locations, and the virus strain, which may lead to different, and sometimes contradictory results [30,31,32,33]. A study carried out by Kent and colleagues (2010) [30] demonstrated that ISVs can be suppressed by co-infection with other viruses. The researchers observed that the ISV CxFV Izabal did not suppress WNV replication in C6/36 cells and Culex quinquefasciatus mosquitoes. Most importantly, WNV transmission was actually enhanced in CxFV infected Culex quinquefasciatus mosquitoes from Honduras [30].
The studies of the interaction between the mosquito microbiota and arboviruses provide essential information to better understand their relationship and how we can use this in favor of public health. More recently, ISVs have gained attention as potential as biotechnological tools for the biological control of arboviruses, as platforms for vaccines, and the development of safe laboratory diagnostics. Additionally, understanding the virome of nonpathogenic viruses in mosquitoes can be leveraged to investigation mosquito ecology [34].
Our study demonstrates that ZIKV, an important virus in terms of public health, is inhibited by the ISV of the taxon Negevirus, PIUV, in mosquito cells in vitro. These findings may provide opportunities for development of new tools to block or decrease virus transmission. Although the number of cases of disease caused by ZIKV has decreased in recent years, the virus is still circulating at low levels, causing disease and it is also spreading to new areas around the world (WHO, 2022). In a recent study of cumulative global ZIKV burden since 2015, ZIKV infections demonstrated an increasing trend from 2011 to 2015 followed by a decreasing trend [35]. Importantly, when events occur, women between the ages of 15 to 49 are most affected in this age group. This indicates that the potential for ZIKV congenital transmission still exists. Nonetheless, these years of low incidence provides an opportune moment to look for new tools to prevent, limit, or combat new outbreaks, and the emergence of the virus in new areas with vulnerable populations.
Author Contributions
Conceptualization, V.L.C and M.T.L.; Methodology: D.P, E.R.S, T.D.L, B.T.D.N, S.E.B; Validation: V.L.C, D.P., B.T.D.N; Formal Analysis: V.L.C, B.T.D.N; Investigation: V.L.C, MT.L., Resources: M.T.L; Writing—original draft preparation: V.L.C.; Writing—review and editing: M.T.L; Visualization, V.L.C and M.T.L.; Supervision: M.T.L.; Project Administration: V.L.C and M.T.L.; Funding acquisition, M.T.L.
Funding
This review is supported in part by the scholarship granted by the Brazilian Federal Foundation for Support and Evaluation of Graduate Education (Capes), in the scope of Procad-Amazonia-DRI, process number: 88881.369357/2019-01 as well as by the Fern-Audette Endowment and University of Florida.
Acknowledgments
We would like to thank the Instituto Evandro Chagas, especially the Program of Post-graduation in Virology, and the University of Florida for allowing us the opportunity to develop and collaborate research with arboviruses and ISVs. We also would like to thank Mario Basara for the English review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vasconcelos, P.F.C.; Calisher, C.H. Emergence of Human Arboviral Diseases in the Americas, 2000-2016. Vector-Borne Zoonotic Dis. 2016, 16, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Soni S, Gill VJS, Anusheel, Singh J, Chhabra J, Gill GJS, Bakshi R. Dengue, Chikungunya, and Zika: The Causes and Threats of Emerging and Re-emerging Arboviral Diseases. Cureus. 2023 Jul 11;15(7):e41717. [CrossRef]
- Masmejan, S.; Musso, D.; Vouga, M.; Pomar, L.; Dashraath, P.; Stojanov, M.; Panchaud, A.; Baud, D. Zika Virus. Pathogens 2020, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- CDC. Centers for Disease Control and Prevention. 2022. Available at: https://www.cdc.gov/campylobacter/guillain-barre.html.
- Bolling, B.G.; Weaver, S.C.; Tesh, R.B.; Vasilakis, N. Insect-specific virus discovery: Significance for the arbovirus community. Viruses 2015, 7, 4911–4928. [Google Scholar] [CrossRef] [PubMed]
- Gómez M, Martinez D, Muñoz M, Ramírez JD. Aedes aegypti and Ae. albopictus microbiome/virome: new strategies for controlling arboviral transmission? Parasit Vectors. 2022 Aug 9;15(1):287. [CrossRef]
- Carvalho, V.L.; Long, M.T. Insect-Specific Viruses: An overview and their relationship to arboviruses of concern to humans and animals. Virology 2021, 557, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.; Tangudu, C.S.; Hurt, S.L.; Tumescheit, C.; Firth, A.E.; Garcia-Rejon, J.E.; Machain-Williams, C.; Blitvich, B.J. Discovery of a novel Tymoviridae-like virus in mosquitoes from Mexico. Arch. Virol. 2019, 164, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Kallies, R.; Kopp, A.; Zirkel, F.; Estrada, A.; Gillespie, T.R.; Drosten, C.; Junglen, S. Genetic characterization of goutanap virus, a novel virus related to negeviruses, cileviruses and higreviruses. Viruses 2014, 6, 4346–4357. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.R.T.; Contreras-Gutierrezb, M.A.; Guzman, H.; Martins, L.C.; Barbiratoh, M.F.; Saviti, C.; Baltaj, V.; Uribec, S.; Viverob, R.; Suaza, J.D.; et al. Genetic characterization, molecular epidemiology, and phylogenetic relationships of insect-specific viruses in the taxon Negevirus. Virology 2017, 504, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Ye, Z.-X.; He, Y.-J.; Zhang, Y.; Wang, X.; Huang, H.-J.; Zhuo, J.-C.; Sun, Z.-T.; Yan, F.; Chen, J.-P.; et al. Discovery of Two Novel Negeviruses in a Dungfly Collected from the Arctic. Viruses 2020, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis N, Forrester NL, Palacios G, Nasar F, Savji N, Rossi SL, Guzman H, Wood TG, Popov V, Gorchakov R, González AV, Haddow AD, Watts DM, da Rosa AP, Weaver SC, Lipkin WI, Tesh RB. Negevirus: a proposed new taxon of insect-specific viruses with wide geographic distribution. J Virol. 2013 Mar;87(5):2475-88. [CrossRef]
- Ramos-Gonzalez, P.L.; Dos Santos, G.F.; Chabi-Jesus, C.; Harakava, R.; Kitajima, E.W.; Freitas-Astua, J. Passion Fruit Green Spot Virus Genome Harbors a New Orphan ORF and Highlights the Flexibility of the 5′-End of the RNA2 Segment Across Cileviruses. Front. Microbiol. 2020, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Lenz, O.; Pribylova, J.; Franova, J.; Koloniuk, I. Fragaria vesca-associated virus 1, a new virus related to negeviruses. Arch. Virol. 2020, 165, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Potter-Birriel JM, Pollio AR, Knott BD, Chunashvili T, Fung CK, Conte MA, Reinbold-Wasson DD, Hang J. Metagenomics analysis reveals presence of the Merida-like virus in Gorgia. Front Microbiol. 2023 Oct 12;14:1258810. [CrossRef]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 5, 427(2):90-7. [CrossRef]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; Melville, L.; Hunt, N.; Webb, R.I.; Blitvich, B.J.; Whelan, P.; Hall, R.A. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLoS One 2013, (2): e56534. [CrossRef]
- Romo, H.; Kenney, J.L.; Blitvich, B.J.; Brault, A.C. Restriction of Zika virus infection and transmission in Aedes aegypti mediated by an insect-specific flavivirus. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nasar, F.; Erasmus, J.H.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus induces both homologous and heterologous interference. Virology 2015, 484:51-58. [CrossRef]
- Erasmus, J.H.; Auguste, A.J.; Kaelber, J.T.; Luo, H.; Rossi, S.L.; Fenton, K.; Leal, G.; Kim, D.Y.; Chiu, W.; Wang, T.; Frolov, I.; Nasar, F.; Weaver, S.C. A chikungunya fever vaccine utilizing an insect-specific virus platform. Nat Med 2017, 23(2):192-199. [CrossRef]
- Erasmus JH, Needham J, Raychaudhuri S, Diamond MS, Beasley DW, Morkowski S, Salje H, Fernandez Salas I, Kim DY, Frolov I, Nasar F, Weaver SC. Utilization of an Eilat Virus-Based Chimera for Serological Detection of Chikungunya Infection. PLoS Negl Trop Dis. 2015 Oct 22;9(10):e0004119. [CrossRef]
- Erasmus JH, Weaver SC. Biotechnological Applications of an Insect-Specific Alphavirus. DNA Cell Biol. 2017 Dec;36(12):1045-1049. [CrossRef]
- Patterson, E.I.; Kautz, T.F.; Contreras-Gutierrez, M.A.; Guzman, H.; Tesh, R.B.; Hughes, G.L.; Forrester, N.L. Negeviruses Reduce Replication of Alphaviruses during Coinfection. J. Virol. 2021, 95, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5439. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Corrêa A, Solberg O, Couto-Lima D, Kenney J, Serra-Freire N, Brault A, Nogueira R, Langevin S, Komar N. Nhumirim virus, a novel flavivirus isolated from mosquitoes from the Pantanal, Brazil. Arch Virol. 2015 Jan;160(1):21-7. [CrossRef]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; van den Hurk, A.F. The insect-specific Palm Creek virus modulates West Nile virus infection in and transmission by Australian mosquitoes. Parasit Vectors 2016, 25;9(1):414. [CrossRef]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile virus by Culex quinquefasciatus say infected with Culex Flavivirus Izabal. PLoS Negl Trop Dis 2010, 4;4(5): e671. [CrossRef]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex flavivirus and West Nile virus mosquito coinfection and positive ecological association in Chicago, United States. Vector Borne Zoonotic Dis 2011, 11(8):1099-105. [CrossRef]
- Newman, C.M.; Krebs, B.L.; Anderson, T.K.; Hamer, G.L.; Ruiz, M.O.; Brawn, J.D.; Brown, W.M; Kitron, U.D.; Goldberg, T.L. Culex Flavivirus During West Nile Virus Epidemic and Interepidemic Years in Chicago, United States. Vector Borne Zoonotic Dis 2017, 17(8):567-575. [CrossRef]
- Crockett, R.K.; Burkhalter, K.; Mead, D.; Kelly, R.; Brown, J.; Varnado, W.; Roy, A.; Horiuchi, K.; Biggerstaff, B.J.; Miller, B.; Nasci, R. Culex flavivirus and West Nile virus in Culex quinquefasciatus populations in the southeastern United States. J Med Entomol 2012, 49(1):165-74. [CrossRef]
- Hollingsworth BD, Grubaugh ND, Lazzaro BP, Murdock CC. Leveraging insect-specific viruses to elucidate mosquito population structure and dynamics. PLoS Pathog. 2023 Aug 31;19(8):e1011588. [CrossRef]
- Guo Z, Jing W, Liu J, Liu M. The global trends and regional differences in incidence of Zika virus infection and implications for Zika virus infection prevention. PLoS Negl Trop Dis. 2022 Oct 21;16(10):e0010812. [CrossRef]
Figure 1.
PIUV reduces ZIKV titer in C6/36 cells (ZIKV and PIUV both at MOI 0.1). A. ZIKV TCID50 titers in the “ZIKV-only” and “ZIKV and PIUV coinfection” wells. B. ZIKV RNA copy number “ZIKV-only” and “ZIKV and PIUV coinfection” wells. C. Threshold cycle (CT) of PIUV (RT-qPCR), comparing “PIUV-only” and “ZIKV and PIUV coinfection” wells. ns: not-significant (p≥ 0.05); *: significant (p 0.01 to 0.05), **: very significant (p 0.001 to 0.01), ***: extremely significant (p 0.0001 to 0.001).
Figure 1.
PIUV reduces ZIKV titer in C6/36 cells (ZIKV and PIUV both at MOI 0.1). A. ZIKV TCID50 titers in the “ZIKV-only” and “ZIKV and PIUV coinfection” wells. B. ZIKV RNA copy number “ZIKV-only” and “ZIKV and PIUV coinfection” wells. C. Threshold cycle (CT) of PIUV (RT-qPCR), comparing “PIUV-only” and “ZIKV and PIUV coinfection” wells. ns: not-significant (p≥ 0.05); *: significant (p 0.01 to 0.05), **: very significant (p 0.001 to 0.01), ***: extremely significant (p 0.0001 to 0.001).
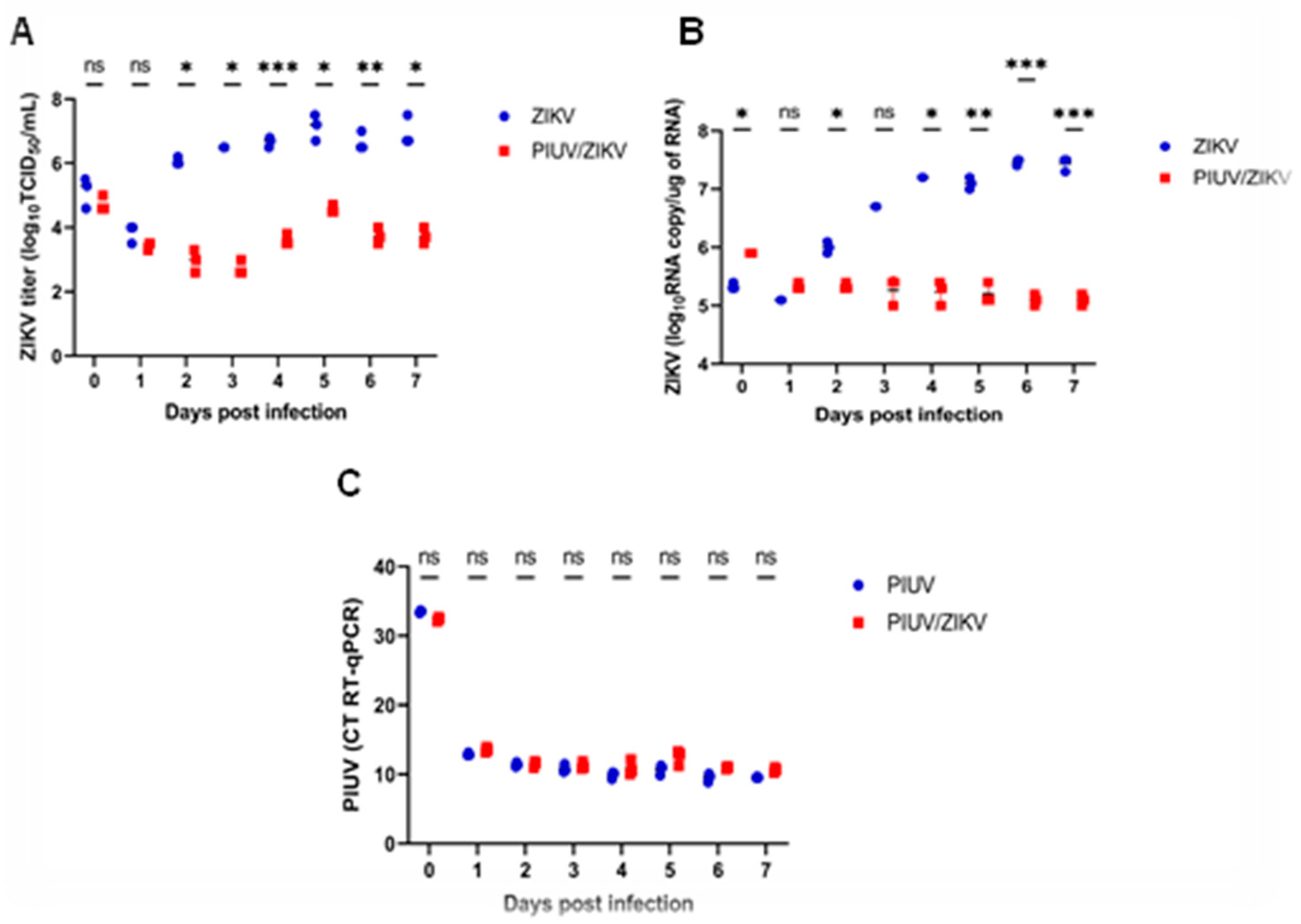
Figure 2.
PIUV reduces ZIKV titer in C6/36 cells (ZIKV at MOI 1 and PIUV at MOI 0.1). A. ZIKV TCID50 titers in “ZIKV-only” and “ZIKV and PIUV coinfection” wells. B. ZIKV RNA copy number in “ZIKV-only” and “ZIKV and PIUV coinfection” wells. C. Threshold cycle (CT) of PIUV (RT-qPCR), comparing “PIUV-only” and “ZIKV and PIUV coinfection” wells. ns: not-significant (p≥ 0.05); *: significant (p 0.01 to 0.05), **: very significant (p 0.001 to 0.01), ***: extremely significant (p 0.0001 to 0.001), ****: extremely significant (p < 0.0001).
Figure 2.
PIUV reduces ZIKV titer in C6/36 cells (ZIKV at MOI 1 and PIUV at MOI 0.1). A. ZIKV TCID50 titers in “ZIKV-only” and “ZIKV and PIUV coinfection” wells. B. ZIKV RNA copy number in “ZIKV-only” and “ZIKV and PIUV coinfection” wells. C. Threshold cycle (CT) of PIUV (RT-qPCR), comparing “PIUV-only” and “ZIKV and PIUV coinfection” wells. ns: not-significant (p≥ 0.05); *: significant (p 0.01 to 0.05), **: very significant (p 0.001 to 0.01), ***: extremely significant (p 0.0001 to 0.001), ****: extremely significant (p < 0.0001).
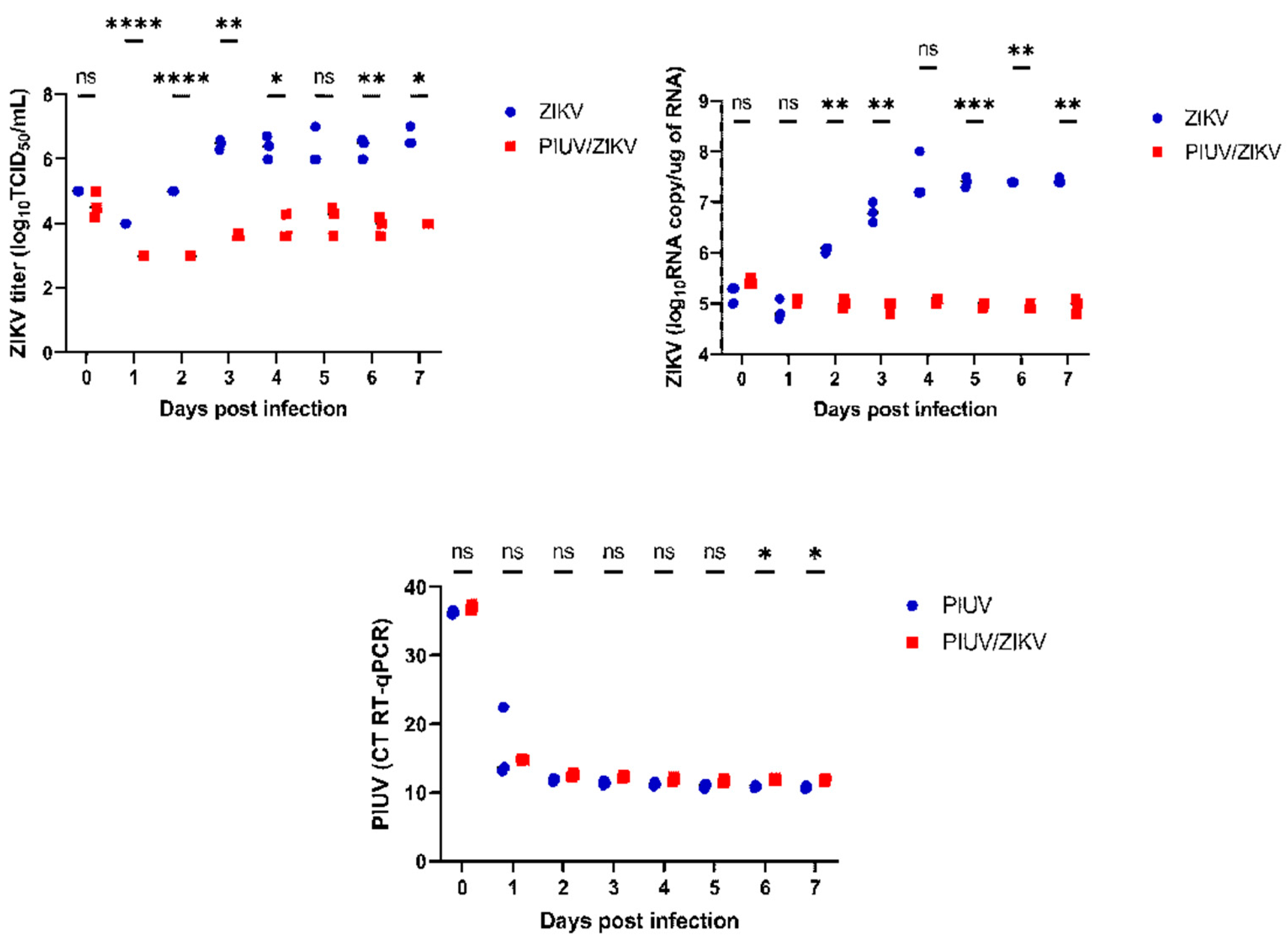
Figure 3.
Infected C6/36 cells showing Cytopathic Effect (CPE) (black arrows) and mock-challenged cells. A-H: ZIKV and PIUV MOI 0.1. A. Cells infected with ZIKV (3º dpi.) showing clumps of cells and cell destruction (100x). B. Cells infected with PIUV (3º dpi.) showing dead cells and spaces between cells (100x). C. Cells coinfected with ZIKV and PIUV (3º dpi.) presenting dead cells and spaces between cells (100x). D. Mock-challenged cells (100x). E. Cells infected with ZIKV (7º dpi.) presenting more prominent clumps of cells and cell destruction (100x). F. Cells infected with PIUV (7º dpi.) showing considerable dead cells and spaces between cells (100x). G. Cells coinfected with ZIKV and PIUV (7º dpi.) displaying the evolution of the cell death and destruction of the monolayer (100x). H. Mock-challenged cells (100x). I-P: ZIKV MOI 1 and PIUV MOI 0.1. I. Cells infected with ZIKV (3º dpi.) showing initial discrete CPE (100x). J. Cells infected with PIUV (3º dpi.) displaying dead cells and destruction of the monolayer (100x). K. Cells coinfected with ZIKV and PIUV (3º dpi.) presenting cell death and destruction of the monolayer (100x). L. Mock-challenged cells (100x). M. Cells infected with ZIKV (7º dpi.) (showing the evolution of the CPE) (200x). N. Cells infected with PIUV (7º dpi.) (showing the evolution of the CPE) (200x). O. Cells coinfected with ZIKV and PIUV (7º dpi.) (showing the evolution of the CPE) (200x). P. Mock-challenged cells (200x).
Figure 3.
Infected C6/36 cells showing Cytopathic Effect (CPE) (black arrows) and mock-challenged cells. A-H: ZIKV and PIUV MOI 0.1. A. Cells infected with ZIKV (3º dpi.) showing clumps of cells and cell destruction (100x). B. Cells infected with PIUV (3º dpi.) showing dead cells and spaces between cells (100x). C. Cells coinfected with ZIKV and PIUV (3º dpi.) presenting dead cells and spaces between cells (100x). D. Mock-challenged cells (100x). E. Cells infected with ZIKV (7º dpi.) presenting more prominent clumps of cells and cell destruction (100x). F. Cells infected with PIUV (7º dpi.) showing considerable dead cells and spaces between cells (100x). G. Cells coinfected with ZIKV and PIUV (7º dpi.) displaying the evolution of the cell death and destruction of the monolayer (100x). H. Mock-challenged cells (100x). I-P: ZIKV MOI 1 and PIUV MOI 0.1. I. Cells infected with ZIKV (3º dpi.) showing initial discrete CPE (100x). J. Cells infected with PIUV (3º dpi.) displaying dead cells and destruction of the monolayer (100x). K. Cells coinfected with ZIKV and PIUV (3º dpi.) presenting cell death and destruction of the monolayer (100x). L. Mock-challenged cells (100x). M. Cells infected with ZIKV (7º dpi.) (showing the evolution of the CPE) (200x). N. Cells infected with PIUV (7º dpi.) (showing the evolution of the CPE) (200x). O. Cells coinfected with ZIKV and PIUV (7º dpi.) (showing the evolution of the CPE) (200x). P. Mock-challenged cells (200x).
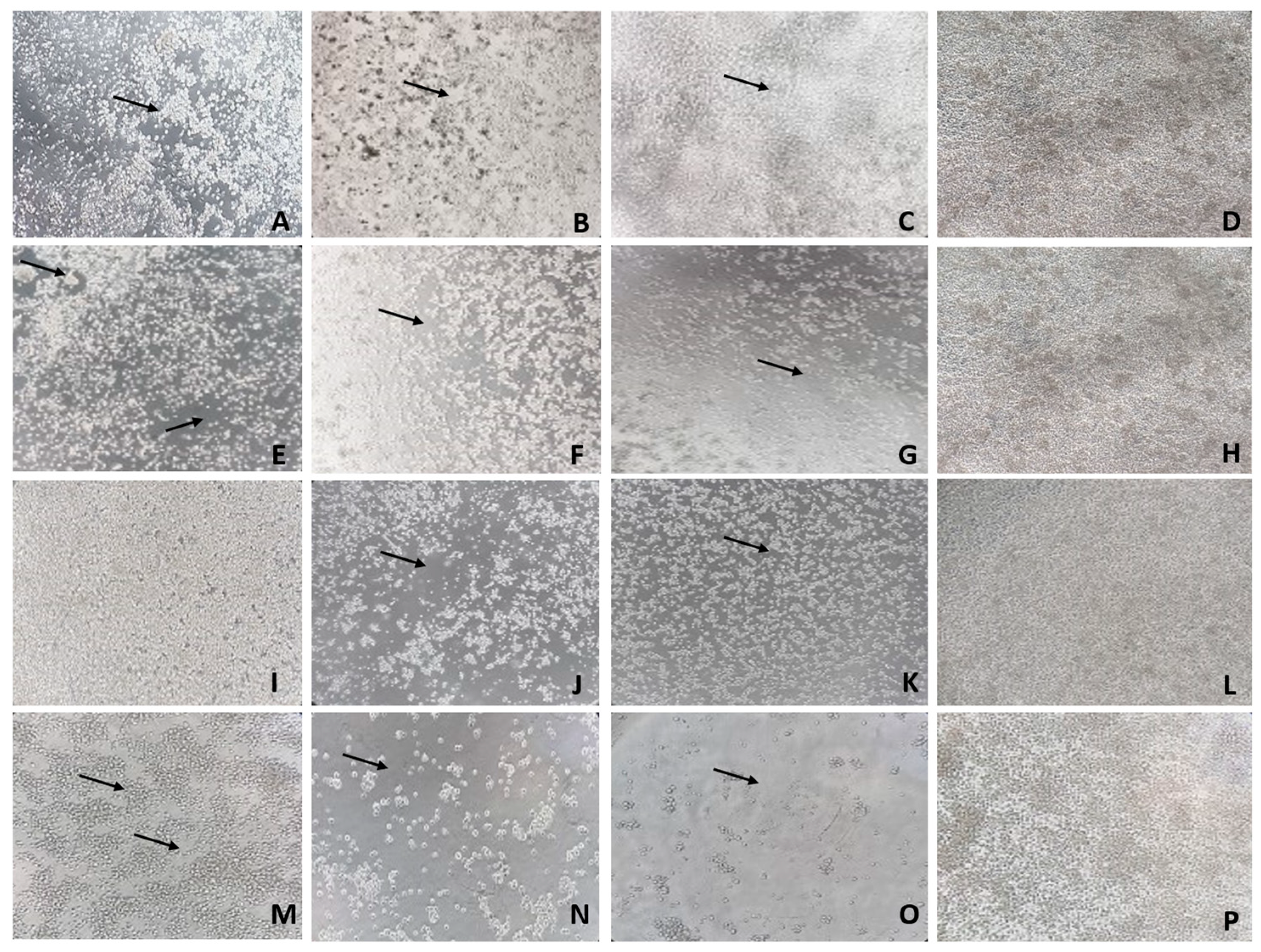
Figure 4.
PIUV does not inhibit ZIKV cellular entry but inhibits intracellular replication in C6/36 cells (ZIKV and PIUV at MOI 0.1). Investigation of the intracellular replication. A. ZIKV RNA copy number in the condition “ZIKV-only” and “ZIKV and PIUV coinfection”. B. Threshold cycle (CT) of PIUV (RT-qPCR), comparing the condition “PIUV-only” and “ZIKV and PIUV coinfection”. C. Positive Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: ZIKV (100x) (3º dpi.). D. Positive Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: ZIKV and PIUV coinfection (100x) (3º dpi.) (less fluorescence intensity in comparison with C). E. Negative Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: mock-challenge (100x). ns: not-significant.
Figure 4.
PIUV does not inhibit ZIKV cellular entry but inhibits intracellular replication in C6/36 cells (ZIKV and PIUV at MOI 0.1). Investigation of the intracellular replication. A. ZIKV RNA copy number in the condition “ZIKV-only” and “ZIKV and PIUV coinfection”. B. Threshold cycle (CT) of PIUV (RT-qPCR), comparing the condition “PIUV-only” and “ZIKV and PIUV coinfection”. C. Positive Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: ZIKV (100x) (3º dpi.). D. Positive Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: ZIKV and PIUV coinfection (100x) (3º dpi.) (less fluorescence intensity in comparison with C). E. Negative Indirect Immunofluorescence using 4G2 anti-orthoflavivirus antibodies: mock-challenge (100x). ns: not-significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



