
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Avian Influenza viruses (AIVs) has posed a significant pandemic threat since their discovery. This review mainly focuses on the epidemiology, virology, and pathogenesis, and treatments of avian influenza viruses. We delve into the global spread, past pandemics, clinical symptoms, severity, and immune response related to AIVs. The review also discusses various control measures, including antiviral drugs, vaccines, and potential future directions in influenza treatment and prevention. Lastly, by summarizing the insights from the previous pandemic control, this review aims to direct effective strategies for managing future influenza pandemics.
Keywords:
Avian Influenza
; Pandemic Management
; Epidemiology
; Virology
; Pathogenesis
; Antiviral Drugs
; Influenza Vaccines
; Immune Response
; Antigenic Drift
; Antigenic Shift
1. Introduction
Influenza is a common illness that affects a great number of people around the globe. History accounts of influenza pandemics date back to ancient times, with Hippocrates and Livy mentioning them as early as 412 B.C. [1]. Starting from 17th century, as the disease gained widespread recognition and investigation, influenza received significant attention from both scientific and lay publications [2]. During the Russian influenza pandemic of 1889, Haemophilus influenzae was mistakenly identified as the cause of influenza, but this paved the way for modern medicine to be used to study influenza [3]. Following that, a British physiologist named Walter Morley Fletcher isolated the influenza virus as the causative agent of flu in 1933 [3]. Until 21st century, seasonal influenza still affects about 10% of the global population and nearly half a million of people still die from the disease each year globally [4].
The causative agent of influenza is the influenza virus. This family contains four genera: Influenza A Virus, Influenza B Virus, Influenza C Virus, and Influenza D Virus [5]. Both influenza A and B viruses have a lipid envelope containing two surface proteins: hemagglutinin (HA) and neuraminidase (NA) [6]. While influenza C and D viruses have hemagglutinin-esterase-fusion (HEF) glycoproteins on their surfaces [7]. In addition, both the influenza A and B viruses genomes contain eight negative-sense, single-stranded RNA segments, while the influenza C and D virus genome contains seven segments instead [7].
Among four genera influenza viruses, influenza A virus is regarded as the most important type of influenza virus to be studied due to its potential to cause severe pandemics with high morbidity and mortality rates [8]. Humans have experienced several severe pandemics caused by the influenza A virus, including the Spanish Flu in 1918, the Asian flu in 1957, and the American swine flu in 1976 [3,8]. Influenza A viruses also have high mutation rates, resulting in their unpredictability and the ability to cause new epidemics or pandemics [8]. So, this review will mainly focus on the characteristics of the influenza A virus and its subtypes.
The RNA genome segments in the influenza A virus encode eight different viral proteins. These include three membrane-bound proteins responsible for antigen-antibody interaction: hemagglutinin, neuraminidase, and matrix protein 2 (M2) [9]. Glycoprotein HA facilitates viral entry into host cells by binding to sialic acid receptors on the surface of target cells, whereas NA is essential for viral particle release from infected cells [8]. Ion channel M2 acidifies the interior of the viral particle, causing the viral ribonucleoprotein complex to dissociate from the matrix protein and allowing the viral genome to enter the host cell [8]. In the ribonucleoprotein core, polymerase basic protein 1 (PB1), polymerase basic protein 2 (PB2), polymerase acidic protein (PA), and NP form polymerase complexes responsible for viral replication [9]. Additionally, there are nuclear export protein/non-structural protein 2 (NEP/NS2) and matrix protein 1 (M1) exist in the virus, while non-structural protein 1 (NS1) is not packaged in virions [9]. The function of NEP/NS2 and NS1 is to promote viral RNA replication, whereas M1 has multiple functions such as providing structural support and stability to the virus particle, facilitating viral assembly, participating in viral RNA synthesis regulation, etc [8].
The influenza A virus subtypes can be differentiated by the H and N antigens on the virus surface [10]. Each influenza virus expresses one H antigen and one N antigen, which can appear in any combination [10]. Currently, 16 different H subtypes (H1-H16) and 9 N subtypes (N1-N9) have been identified [10]. The influenza B viruses are classified into two lineages, Victoria and Yamagata, based on the genetic and antigenic differences in H and N [11]. Influenza C virus does not have HA or NA, but rather a single surface glycoprotein HEF, so it is not classified into subtypes or lineages [11]. Only some of the Influenza A Virus genus is known to infect birds, which are called avian influenza [10]. The first report of avian influenza infection was in northern Italy, in 1878 [12]. Researchers discovered that the pathogen of avian influenza was an influenza A virus subtype by identifying the presence of type A influenza virus type-specific ribonucleoprotein, and it was officially named Avian Influenza Virus in 1955 [12]. Different strains of avian influenza have varying levels of lethality and infectivity for humans and birds. Most of the avian influenza viruses belong to Low Pathogenic Avian Influenza (LPAI) [10]. Only specific subtypes carrying H5 or H7 belong to Highly Pathogenic Avian Influenza (HPAI), which is highly dangerous to susceptible species, including birds and humans [10]. The first reported HPAI was in Scotland in 1959 [12]. Over the years, HPAI outbreaks have occurred in various countries and regions including North America, Europe, the Middle East, East and Southeast Asia (Figure 1). One of the prevalent avian influenza strains in Asia is human avian influenza H7N9. Human infections of H7N9 were first reported in China in 2013 and have continued to occur almost annually, with a total of 1568 reported infections and a high mortality rate (Figure 1). From 15th May 2023, there have been a recent confirmed outbreaks of highly pathogenic avian influenza (HPAI) in the state of Paraná, which is Brazil's primary region for poultry production (Figure 1) [13]. Until 3rd July 2023, there have been 48 cases of HPAI infection, all involving wild birds [14]. Notably, there have been no reports of HPAI cases in commercial or subsistence poultry farms [13]. The Brazilian government has also implemented effective measures to prevent the disease spreading from wild birds to commercial herds [13].
2. Virus Dissemination
Because it involves the transmission of potentially zoonotic and pandemic viruses between different hosts, such as wild birds, poultry, and humans, the spread of avian influenza viruses (AIVs) is a major topic in public health and animal health. Avian species, particularly waterfowl and shorebirds, serve as natural reservoirs for AIVs to grow and mutate [25].
2.1. Global Spread of AIVs
The dissemination of AIVs within and across regions is largely influenced by the migratory movements of wild birds, which can carry and exchange different virus strains along their flyways [26]. Flyways are large corridors that connect migratory birds' breeding and wintering grounds across continents [26]. Studies have shown, for example, that the Pacific Flyway, a migratory path connecting western North America to eastern Asia via breeding grounds in Beringia, has the potential to introduce or exchange avian influenza viruses between North America and Asia [26].
2.2. Regional & Local Spread of AIVs
AIVs can spread and exchange between host species and populations through various pathways, including direct contact with infected live birds or corpses, inhalation of virus-containing aerosols, and consumption of contaminated meat [14,27].
The live poultry market plays a crucial role in the spread of AIVs, as it is a place where various bird species and populations come into contact. These markets could be a source of virus transmission between domestic and wild bird populations, as well as between human and birds [14]. Apart from live poultry market, commercial poultry farms, family backyard farms, and zoos are all places for AIVs to reproduce, mutate, and spread among birds, mammals and humans [28].
2.3. Antigenic Drift
Antigenic drift is the main cause of AIV mutation [25]. Antigenic drift is a gradual evolutionary process driven by the selective pressure to evade host immunity that occurs as a result of frequent mutations within the antibody-binding sites of the influenza virus's HA and NA proteins [29]. The mutations reduce host antibody binding, allowing the virus to spread more efficiently and widely among the population [29]. For example, the H3N2 subtype of influenza A virus has exhibited antigenic drift since it was introduced into humans in 1968 [30]. These mutations can lead to altered antigenic properties of hemagglutinin and neuraminidase, causing the virus to be recognized differently by the immune system, thus escaping neutralization by pre-existing antibodies [30]. Some antigenic variants resulting from drift may possess increased fitness or enhanced pathogenicity, leading to more severe illness or higher rates of transmission [30]. This necessitates the regular update of influenza vaccines to match the predominant circulating strains and maintain their effectiveness [30].
2.4. Antigenic Shift
Antigenic shift is another cause of mutation that happens less frequently than antigenic drift [31]. It happens when several different influenza A viruses, each containing a different combination of gene segments, infect the same host cell [31]. The gene segments of the different viruses can mix and match during replication, producing new viral particles with novel combinations of genes, including changes in their HA and NA proteins [31,32]. The new virus after antigenic shift may have the ability to infect new host, such as human [31]. Since human immune system does not recognize the novel virus, allowing it to spread rapidly through the population, leading to the risk of pandemic [31]. Several pandemics in the history were proved to be caused by antigenic shift, such as the Spanish flu in 1918, and H5N1 outbreak in Hong Kong, 1997 [31,32]. The H1N1 influenza A virus responsible for the Spanish flu had a novel combination of HA and NA genes, which likely emerged through antigenic shift, involving reassortment between swine, human, and avian influenza viruses over several years [33].
2.5. The Mechanism of Human Infections by Influenza Viruses
Once inside the body, influenza viruses often attack the respiratory system, specifically the upper respiratory tract, including the nose, throat, and bronchi [34]. Influenza virus enters the host cell by first recognizing a terminal α-sialic acid that is linked to saccharides anchored on the host cell surface, which serves as a specific receptor molecule, allowing it to gain access to the host cell where it can replicate [15]. The virus particle then induces endocytosis, which creates an endosome that encapsulates the virus [34]. The endosome is transported near the nucleus, and the virus membrane fuses with the host membrane, releasing the eight segments of the RNA genome into the nucleus [15]. This initiates virus transcription and replication, which allow the virus to propagate within the host cell [15]. After binding to and entering the targeted epithelial cells, the virus can cause damage to them, resulting in cell death and viral particle shedding into the airway, resulting in fever, cough, and sore throat symptoms [34]. The infection mechanism will trigger different levels of pathogenic effect in human bodies, which will be discussed in the next section.
3. Clinical Features and Severity of Disease
AIVs primarily affects birds, while a small number of strains of currently discovered over 100 subtypes can infect humans and cause respiratory illness with varying clinical features and severity [35]. However, since 2013, a newly emerged strain of AIV, known as H7N9, has caused over 1500 human infections and 600 deaths in China, while another strain, H5N1, has resulted in over 800 cases and 400 deaths worldwide since its initial emergence in Hong Kong, China [19,35].
The severity of symptoms can vary depending on the individual's immune response and other factors. Human infection of AIVs ranges from mild to severe, in which mild cases of influenza-like illness is more common [36]. Signs and symptoms of mild AIV infection include nasal obstruction, cough, sore throat, fever, shortness of breath, etc. [36] In these mild cases, individuals often experience self-limiting illness and recover in a week [36].
Severe cases, usually caused by HPAI strains like H7N9 or H5N1, have symptoms including acute respiratory distress syndrome (ARDS), pneumonia, multi-organ failure, etc. [37,38] The outbreaks of H7N9 and H5N1 in China have especially high case-fatality rate [37]. The data shows that the rate is 18.7% for H7N9, and 59% for H5N1 [37]. In addition, the severity of AIV infection can be different for some other factors, including age, immune status, and viral load [39]. Patients with pre-existing health conditions such as diabetes, cardiovascular disease, or respiratory diseases are at higher risk of developing severe AIV infection [39]. H5N1 infection may also result in viremia at high levels and for prolonged periods [40]. This allows the virus to spread to extra-respiratory tissues, such as the brain, intestine, liver, lymph nodes, spleen, bone marrow, placenta, fetus, and kidneys [40]. These tissues often exhibit associated lesions, including edema, demyelination, necrosis, accumulation of reactive histiocytes, hepatic necrosis, lipidosis, cholestasis, and hemophagocytic activity in lymph nodes [40].
Individuals infected with avian influenza may exhibit rare or atypical symptoms in addition to the typical respiratory signs, including gastrointestinal upset (diarrhea and vomiting), and neurological symptoms (headache, confusion, seizures, and altered mental status) [38,40,41]. Cardiovascular manifestations, such as myocarditis, have been observed in some severe cases. Additionally, coagulation abnormalities and disseminated intravascular coagulation (DIC) have been reported, which can result in abnormal bleeding and organ damage [41,42].
4. Characteristics of Influenza Virus
Avian influenza viruses, belonging to the influenza A virus family, are characterized by several unique features in their genetic makeup and protein composition, which play crucial roles in their replication and interaction with host cells [9]. Avian influenza is composed of eight segments of single-stranded negative-sense RNA, encoding for different viral proteins [9].
Among the viral proteins, hemagglutinin (HA) is significant in the life cycle of avian influenza viruses among the viral proteins (Figure 2). HA is responsible for mediating the entry of the virus into host cells and determining the host range and tissue tropism of the virus [43]. The HA protein binds to specific receptors on the surface of host cells, primarily sialic acid receptors, allowing the virus to enter the correct cell types [43]. Avian influenza viruses typically show a preference for α(2,3)-linked sialic acid receptors, which are abundant in bird intestine [43]. For example, the HA protein tends to interact with N-acetylneuraminic acid (NeuAc) linked to galactose through an α(2,3) linkage (NeuAcα2-3Gal), a linkage primarily found on the surface of duck intestinal epithelial cells [44]. This receptor specificity restricts the natural transmission of avian influenza viruses to certain species. However, the genetic changes in the HA protein can enable the virus to switch its receptor binding preference to α(2,6)-linked sialic acid receptors, which are predominantly found in human lungs [43]. Similarly, mutated HA protein exhibits a preference for NeuAcα2-6Gal, which is the main type of sialic acid linkage found on the surface of human tracheal epithelial cells [44]. Such receptor binding adaptations are critical for avian influenza viruses to cross the species barrier and potentially infect humans and other mammals.
In addition to HA, another important protein in the life cycle of avian influenza viruses is neuraminidase (NA) (Figure 2). NA acts as an enzyme that cleaves sialic acid residues, which are present on the surface of host cells and are also used by HA to attach to [45]. During the later stages of viral replication, newly formed influenza viruses bud from the host cell membrane. However, the viral particles remain attached to the infected cell surface through their binding to sialic acid receptors [45]. This attachment can hinder the release of viral particles and limit the spread of the virus to neighboring cells and tissues [45]. NA can facilitate the release of newly formed viral particles from infected cells and prevent viral aggregation by cleaving sialic acid residues from glycoproteins on the host cell membrane, thus helping viruses to infect more healthy cells [45].
5. Role of Immune System in the Pathology of Influenza Virus
5.1. Innate Immunity
The innate immune response is the first line of defense against influenza virus infection. It involves epithelial cells in the respiratory tract that detect the virus and initiate an immune response [46].
Three main categories of pattern recognition receptors (PRRs) are activated when detect the pathogen-associated molecular patterns (PAMPs) in innate immune response involved in this process: Toll-like receptors (TLRs), retinoic acid-inducible gene-I (RIG-I) receptors, and nucleotide oligomerization domain-like receptor family pyrin domain containing 3 (NLRP3) [46]. Alveolar macrophages become activated during infection, enhancing their inflammatory response and limiting viral spread through phagocytosis, while potentially contributing to severe pathology through the production of nitric oxide synthase 2 (NOS2) and TNF-α [46]. Natural Killer (NK) cells can bind to HA surface proteins on influenza viruses using sialylated NKp44 and NKp46 receptors, leading to the lysis of infected cells [46]. Dendritic cells (DCs), as antigen-presenting cells, bridge the innate and adaptive immune systems by presenting viral antigens to T and B lymphocytes, surveying the lungs for the presence of pathogens, and acquiring antigens through direct infection or phagocytosis [46]. Each of these cell types plays a critical role in the immune response to influenza, contributing to both inhibiting the virus infection and initiating the adaptive immune response.
5.2. Adaptive Immunity
Adaptive immunity develops late and is specific to the influenza virus strain. The adaptive immune response is composed of two main branches: humoral immunity, which involves virus-specific antibodies, and cellular immunity, which includes virus-specific CD4+ and CD8+ T cells [46].
5.2.1. Humoral Immunity
There are two kinds of humoral immune response, primary and secondary B cell response. The primary response involves both innate-like and conventional B-1 cells, producing IgM antibodies [47]. During the infection's progression, B-2 cells produce IgG, IgA, and IgE antibodies, tailored to specific influenza antigens and have higher affinity compared to IgM antibodies [47]. During secondary B cell activation, which occurs upon subsequent exposure to the same or similar influenza strains, memory B cells are rapidly activated to produce high-affinity antibodies more quickly and in greater quantities [48].
The initial infection with the influenza virus triggers the production of IgM, IgA, and IgG antibodies [46]. While secondary exposures predominantly involve IgA and IgG responses, with IgM typically absent [46]. A key aspect of antibody response, especially in mucosal areas like the respiratory tract, is the production of secretory IgA (sIgA) antibodies. sIgA is produced by plasma cells in mucosal tissues and secreted into the lumen [49]. It provides frontline defence by neutralizing pathogens at mucosal surfaces, preventing their entry into the body [49]. It effectively clears viruses from infected epithelial cells and can redirect antigens from the lamina propria to the lumen without triggering inflammatory responses [49].
The humoral immune response produces antigens that target specific epitopes, including HA, NA, M2, and NP viral proteins [46]. It is important to find out the conserved epitopes of influenza virus. By focusing on conserved epitopes, such as those found in the hemagglutinin stalk and the M2 protein, vaccines can potentially provide protection against a wide range of influenza strains, reducing the need for frequent vaccine updates and offering better defence against pandemic and seasonal influenza outbreaks [50].
5.2.2. Cell-mediated Immunity
T cells, including CD8+ cytotoxic T cells and CD4+ helper T cells, are critical for controlling and clearing AIV infections [46,48]. CD4+ T cells become activated when they identify viral epitopes presented by MHC II and co-stimulatory molecules on antigen-presenting cells (APCs) [46]. When naive CD4+ T cells are activated, they can differentiate into either Th1 or Th2 cells. Th1 cells generate IFN-γ and IL-2, which can enhance responses of cytotoxic T lymphocytes (CTLs) and are vital in developing memory CD8+ T cells [46]. While Th2 cells secrete IL-4, IL-5, and IL-13, which are crucial in stimulating B cells to differentiate and produce antibodies [46]. In addition, memory CD4+ T cells, especially those residing in the lungs, are key in managing reinfections by the influenza A virus and preventing the body against secondary infections [46].
Naive CD8+ T cells can differentiate to CTLs and destroy infected cell, providing immunity against different strains of influenza, as shown in historical patterns of infection and immunity [46]. For example, people who experienced symptomatic H1N1 infection before the 1957 pandemic showed some resistance to the H2N2 strain [46]. The reason for this cross-reactivity against various strains is because of the targets of CTLs, which are highly conserved M1, NP, PA, and PB2 viral proteins [46].
Apart from CD4+ and CD8+ T cells, regulatory T cells (Tregs) and T helper 17 cells (Th17) also plays an important role in cell-mediated immunity against influenza viruses [46]. Tregs can regulate immune responses to prevent excessive tissue damage, while Th17 are essential in countering secondary bacterial infections, such as S. aureus pneumonia [46].
6. Control Measures
Management and control of the influenza virus potentially involve several measures, from early detection of cases and case alerts, and quarantine measurement, to antiviral drugs and vaccines development.
6.1. Antiviral Drugs
6.1.1. Neuraminidase Inhibitors
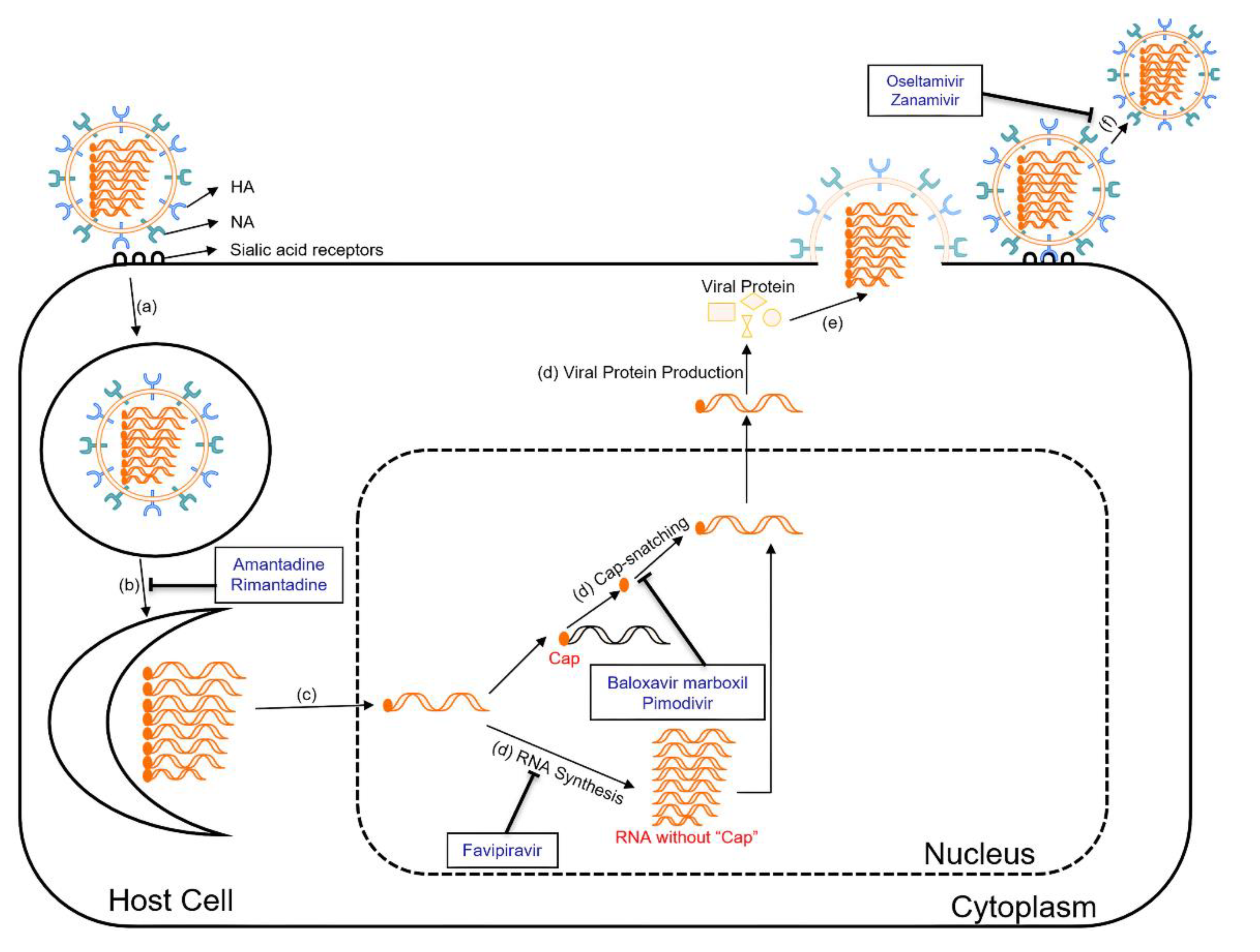
Neuraminidase inhibitors, such as oseltamivir (Tamiflu), zanamivir (Relenza), are commonly used antiviral drugs for the treatment of influenza [45]. They work by inhibiting the NA on the surface of the influenza virus (Figure 2) [45]. By preventing the cleavage of sialic acid receptors, these drugs can cause the accumulation of viral particles on the infected cell surface [45]. As a result, the spread of the virus is impeded, reducing the severity and duration of influenza symptoms.
6.1.2. M2 Ion Channel Inhibitors
The M2 protein forms an ion channel in the viral envelope, facilitating the viral genome to enter the host cell [8]. M2 ion channel inhibitors, including amantadine and rimantadine, target the M2 protein of influenza A viruses (Figure 2) [51]. They block the function of the M2 protein by binding to the pore of the ion channel, which prevents the influx of protons into the viral particle, disrupting the uncoating process and subsequent viral replication [51]. This inhibition of the M2 ion channel effectively reduces the production of infectious viral particles. However, the widespread emergence of resistant strains has significantly limited the usefulness of M2 ion channel inhibitors in recent years [52].
6.1.3. Polymerase Inhibitors
Polymerase inhibitors are a newer class of antiviral drugs that target the viral polymerase complex involved in viral replication. The main polymerase inhibitors currently approved for influenza A virus treatment are baloxavir marboxil, pimodivir, and favipiravir, while favipiravir and baloxavir marboxil are also effective on influenza B viruses [53]. Baloxavir marboxil targets the cap-dependent endonuclease activity of the polymerase acidic (PA) protein of influenza A and B viruses (Figure 2) [53]. The PA protein plays a crucial role in viral replication by cleaving host mRNA molecules in order to "cap-snatch" the capped fragments and produce primers for viral mRNA synthesis [54]. Once inside the host cell, baloxavir marboxil is converted to its active form, which is baloxavir acid [54]. Baloxavir acid binds to the conserved active site of the PA protein and inhibits its endonuclease activity [54]. By blocking the endonuclease activity, baloxavir prevents the cleavage of host mRNA, disrupting the cap-snatching process and inhibiting viral mRNA synthesis [54]. Pimodivir is an inhibitor of the polymerase basic protein 2 (PB2) of influenza A viruses (Figure 2) [55]. It works by preventing the binding of PB2 to the 7-methyl GTP cap structures of host mRNA, thereby inhibiting the early stages of viral transcription [55]. Favipiravir targets at viral RNA-dependent RNA polymerase (RdRp) and introduces errors in the genetic code of the virus (Figure 2) [56]. These mutations can lead to the generation of non-functional or less viable viral particles, thus reducing viral load in the body [56]. Favipiravir also has broad-spectrum antiviral activity against RNA viruses like influenza, Ebola, Rabies, etc. [56] Its precise antiviral mechanism and effects are still being investigated [56].
Apart from the antiviral drugs mentioned above, there are some new potential development directions for influenza antiviral therapy. First, utilizing anti-influenza virus antibodies as a therapeutic approach shows promise in preventing the binding of virions to target cells [57]. Convalescent blood products, immunoglobulins, and monoclonal antibodies have demonstrated efficacy in protecting animals from lethal infections and improving the condition of severely ill patients [57]. Second, sequence-based therapies involving antisense DNA oligomers and short interfering RNA (siRNA) molecules targeting viral mRNA hold the potential to block viral replication and protein synthesis [57]. What is more, combining drugs with different mechanisms of action could enhance antiviral effects and reduce the risk of drug resistance [57]. Combinations of adamantanes and NA inhibitors, like rimantadine plus zanamivir, have shown synergistic effects in animal models [57]. Therefore, exploring these alternative approaches in influenza antiviral therapy holds great promise in the future.
6.2. Vaccines
There are mainly two ways of vaccine production, which are egg-based and recombinant [58]. Egg-based influenza vaccines are commonly produced using fertilized chicken eggs as a substrate for growing the influenza virus. These vaccines can be either inactivated (containing dead viruses post chemical treatment like formaldehyde or beta-propiolactone) or attenuated (containing weakened live viruses) [58,59]. Egg-based vaccines have a potential risk for individuals with severe egg allergies. However, advances in vaccine manufacturing have significantly reduced the amount of egg protein present in the final vaccine product, making it generally safe for most people with egg allergies [58]. While recombinant influenza vaccines (RIVs) are produced using recombinant DNA technology, where specific genes encoding viral proteins are inserted into host cells, such as insect cells or mammalian cells [58,60]. The host cells then produce large quantities of the desired viral proteins to make the vaccines [58]. Although recombinant protein vaccine is safer, faster, and cheaper than egg-based vaccine, it has some challenges like requirement of a high-yielding re-assorted viruses, and potential genetic changes in the virus during production, which may lead to less effectiveness of the vaccine [60,61].
6.2.1. Live Attenuated Influenza Vaccines
Live attenuated influenza vaccines (LAIV), such as FluMist, are quadrivalent vaccines that are administered through nasal spray [58]. LAIV is derived from live influenza viruses that have been modified to be less virulent [58]. After administration via nasal spray, the attenuated viruses replicate in the nasal passages, sinuses, and upper respiratory tract, which can induce mucosal immunity [62]. The immune response induced by LAIV leads to the production of antibodies that can recognize and neutralize the actual influenza viruses [62].
6.2.2. Inactivated Influenza Vaccines
6.2.3. Subunit Influenza Vaccines
Subunit Influenza Vaccines contain specific components of the influenza virus. These vaccines offer a high level of safety as they do not contain infectious viruses [62]. Promising targets for subunit vaccines against influenza viruses include viral structural proteins like M2e, HA, and NP [64]. In addition, proteins like M1 and NA hold promise for future development as subunit vaccines against influenza [64]. Flublok, a kind of recombinant protein subunit vaccine, has been approved by FDA and used for many years [58]. Flublok is an influenza vaccine that contains full-length HA protein antigens derived from three selected influenza virus strains [60]. These antigens are produced in a “non-transformed, non-tumorigenic continuous cell line from Sf9 cells of the fall armyworm”, and they are “full length proteins containing the transmembrane domain and the HA1 and HA2 regions” [60]. The vaccine aims to induce the production of HA inhibition (HAI) antibodies and prevent influenza infection [60].
6.2.4. Epitope-Based Influenza Vaccines
Epitope-based influenza vaccines target specific antigenic sites on the influenza virus that are conserved and immunogenic regions, reducing the impact of antigenic variation and potentially providing broader and long-lasting protection against diverse influenza strains [65]. Epitope-based influenza vaccines can stimulate the immune system to produce antibodies against critical epitopes on the viral proteins [66]. Potential targets include conserved regions of the HA, NP, M1 and M2 proteins of influenza viruses [65,66]. Several epitope-based influenza vaccine candidates are in various stages of development, such as “M001 (BiondVax Pharmaceuticals Ltd., Jerusalem, Israel), Flu-V (hVivo, London, UK) ,FP-01.1 (Immune Targeting Systems Ltd., London, UK), rMVA-k1-k2 (Federal Medical-Biological Agency, Moscow, Russia)” [66].
6.2.5. mRNA Influenza Vaccines
The mRNA influenza vaccine is a new approach to influenza immunization, leveraging the same technology as the highly successful COVID-19 mRNA vaccines [67]. It work by introducing a small piece of synthetic messenger RNA (mRNA) into the body, encoding specific virus proteins, such as HA protein of influenza viruses [67,68]. Once inside cells, the mRNA instructs them to produce harmless fragments of the target protein, inducing immune response and antibody production [68]. mRNA technology, proven successful in COVID-19 vaccines, offers hope for more effective influenza vaccines [67]. It eliminates egg-based production, potentially improving strain matching and manufacturing [67]. Clinical trials have shown promise, although robust seroconversion across strains remains a challenge. mRNA allows multiple antigen combinations, potentially broadening protection [67]. Recent research expanded this to a quadrivalent influenza A group 2 mRNA formulation, offering protection for multiple influenza A viruses, which demonstrates broad protection of mRNA vaccines [67]. Currently, mRNA influenza vaccines from Moderna and Pfizer have begun phase 3 clinical trials, and vaccines from Sanofi/Translate Bio and GlaxoSmithKline (GSK)/CureVac are under phase 1 [67].
7. Conclusions and Future Directions
This review summarizes the multifaceted nature of avian influenza, covering its epidemiology, virology, hosts, vectors, symptoms, immune responses, and treatments of AIVs. The ability of the virus to mutate, including antigenic drift and shift, enhances its ability to evade host immunity and adapt to new hosts. Vaccines and antiviral drugs are the two main ways to combat AIV infection. However, the influenza virus can co-evolve with preventive and treatment measures, making the control of the disease difficult.
Looking back on the COVID-19 pandemic, we have learned a lot in controlling this airborne and zoonotic virus that can be applied to future pandemics. Future approaches emphasize developing new models for public sector funding and research, increasing testing capacity with new technologies, and building a connected diagnostic system for surveillance [69]. Addressing social inequalities is highlighted as a critical intervention. Control measures like social distancing can have a negative impact on the most vulnerable groups in society, widening socio-economic gaps between the richest and poorest. Future policies should consider the interests of vulnerable groups to increase the overall effectiveness of policies and minimize their negative impacts [70]. Additionally, understanding the current challenges and potential limitations of COVID-19 vaccination in this population is critical, not only to inform decisions about additional vaccine doses but also to advise on non-pharmacologic prevention strategies and prophylactic approaches. Because of the potential for COVID-19 to transition from a pandemic to an endemic, establishing effective healthcare interventions to protect immunocompromised individuals from the ongoing threat of infection remains a public health priority [71]. Adapting and applying these approaches will enable us to better prepare for and respond to the risk of future outbreaks, including potential influenza pandemics.
Here, we propose some possible AIV control measures in two aspects: biology and public policies. Firstly, developing avian-specific vaccines and antiviral drugs is crucial to prevent the spread of influenza among birds. Concurrently, advancing human vaccines, particularly leveraging rapid advancements in mRNA technology, is essential for more effective and timely protection. Secondly, it's important to establish and enhance surveillance systems for wildlife and livestock diseases, providing early warning for potential influenza risks. Implementing stringent laws to limit wild bird hunting and live bird market activities can reduce human-bird interaction, thereby controlling infection sources. Finally, intensifying public awareness and education, particularly about the risks of consuming poultry during high-risk influenza periods, is vital for preventing infections.
In conclusion, comprehensive strategies are needed to prevent and respond to future pandemics caused by AIVs. The joint efforts of governments across the world, and of various health organizations, will enable the effective implementation of these strategies and achieve even better results.
Author Contributions
“Conceptualization, Y.-W.K.; data curation, J.H.; writing—original draft preparation, J.H.; writing—review and editing, J.H. and Y.-W.K.; visualization, J.H.; supervision, Y.-W.K.; project administration, Y.-W.K.; funding acquisition, Y.-W.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the Startup Fund from the Division of Natural and Applied Sciences, Duke Kunshan University (00AKUG0130). Y.-W.K. acknowledges funding from Duke Kunshan University. This research is also supported in part by the Summer Research Scholar (SRS) program, Signature Work Research Grant (SWRG), and Signature Fund (SF) from Duke Kunshan University. J. H. acknowledges funding from Duke Kunshan University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created in this review article.
Acknowledgments
This research was supported in part by the Startup Fund from the Division of Natural and Applied Sciences, Duke Kunshan University (00AKUG0130). Y.-W.K. acknowledges funding from Duke Kunshan University. This research is also supported in part by the Summer Research Scholar (SRS) program, Signature Work Research Grant (SWRG), and Signature Fund (SF) from Duke Kunshan University. J. H. acknowledges funding from Duke Kunshan University. We thank Heng Yu (Hazel) for critically reading and editing the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kuszewski K, Brydak L. The epidemiology and history of influenza. Biomedicine & pharmacotherapy 2000, 54, 188–95. [Google Scholar]
- Potter, CW. A history of influenza. Journal of applied microbiology 2001, 91, 572–9. [Google Scholar] [CrossRef]
- Barnett, R. Influenza. Lancet 2019, 393, 396. [Google Scholar] [CrossRef]
- Layne SP, Monto AS, Taubenberger JK. Pandemic influenza: an inconvenient mutation. Science 2009, 323, 1560–1. [Google Scholar] [CrossRef] [PubMed]
- Asha K, Kumar B. Emerging influenza D virus threat: what we know so far! Journal of Clinical Medicine 2019, 8, 192. [Google Scholar] [CrossRef]
- Palese P, Young JF. Variation of influenza A, B, and C viruses. Science 1982, 215, 1468–74. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu S, Murakami S, Shindo K, et al. Influenza C and D viruses package eight organized ribonucleoprotein complexes. Journal of virology 2018, 92, 10–1128. [Google Scholar] [CrossRef]
- Cheung TK, Poon LL. Biology of influenza a virus. Annals of the New York Academy of Sciences 2007, 1102, 1–25. [Google Scholar] [CrossRef]
- Suarez DL, Schultz-Cherry S. Immunology of avian influenza virus: a review. Developmental & Comparative Immunology 2000, 24, 269–83. [Google Scholar]
- Alexander, DJ. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–44. [Google Scholar] [CrossRef]
- Types of Influenza Viruses. March 30 2023. https://www.cdc.gov/flu/about/viruses/types.htm (accessed April 3, 2023).
- Lupiani B, Reddy SM. The history of avian influenza. Comparative Immunology, Microbiology and Infectious Diseases 2009, 32, 311–23. [Google Scholar] [CrossRef]
- Azevedo, D. Brazil confirms bird flu in main poultry-producing state. July 3 2023. https://www.poultryworld.net/health-nutrition/health/brazil-confirms-avian-influenza-in-main-poultry-producing-state/ (accessed July 17, 2023).
- Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiological reviews 1992, 56, 152–79. [Google Scholar] [CrossRef]
- Luo, M. Influenza virus entry. Viral Molecular Machines 2012, 201–21. [Google Scholar]
- Skowronski D, Tweed S, Petric M, Booth T, Li Y, Tam T. Human illness from avian influenza H7N3. British Columbia Emerg Infect Dis 2004, 10, 219. [Google Scholar]
- Barrera-Badillo G, Ramirez-Gonzalez E, Aparicio-Antonio R, Nuñez-Garcia T, Arellano-Suarez D. Highly Pathogenic Avian Influenza A (H7N3) Virus Infection in Two Poultry Workers—Jalisco, Mexico, July 2012. MMWR 2012, 61, 36. [Google Scholar]
- Stegeman A, Bouma A, Elbers AR, et al. Avian influenza A virus (H7N7) epidemic in The Netherlands in 2003: course of the epidemic and effectiveness of control measures. The Journal of Infectious Diseases 2004, 190, 2088–95. [Google Scholar] [CrossRef] [PubMed]
- Technical Report: Highly Pathogenic Avian Influenza A(H5N1) Viruses. February 24 2023. https://www.cdc.gov/flu/avianflu/spotlights/2022-2023/h5n1-technical-report.htm (accessed April 12, 2023).
- Reported Human Infections with Avian Influenza A Viruses. March 17 2023. https://www.cdc.gov/flu/avianflu/reported-human-infections.htm (accessed April 3, 2023).
- Kullman G, Delaney LJ, Decker J, MacMahon K. Protecting poultry workers from avian influenza (bird Flu). 2008.
- Belser JA, Bridges CB, Katz JM, Tumpey TM. Past, present, and possible future human infection with influenza virus A subtype H7. Emerging infectious diseases 2009, 15, 859. [Google Scholar] [CrossRef] [PubMed]
- El-Shesheny R, Franks J, Marathe BM, et al. Genetic characterization and pathogenic potential of H10 avian influenza viruses isolated from live poultry markets in Bangladesh. Scientific Reports 2018, 8, 10693. [Google Scholar] [CrossRef] [PubMed]
- Bird flu case confirmed at farm. BBC NEWS. 2007 24 May.
- Blagodatski A, Trutneva K, Glazova O, et al. Avian influenza in wild birds and poultry: dissemination pathways, monitoring methods, and virus ecology. Pathogens 2021, 10, 630. [Google Scholar] [CrossRef] [PubMed]
- Prosser DJ, Chen J, Ahlstrom CA, et al. Maintenance and dissemination of avian-origin influenza A virus within the northern Atlantic Flyway of North America. PLoS Pathogens 2022, 18, e1010605. [Google Scholar]
- Kilpatrick AM, Chmura AA, Gibbons DW, Fleischer RC, Marra PP, Daszak P. Predicting the global spread of H5N1 avian influenza. Proceedings of the National Academy of Sciences 2006, 103, 19368–73. [Google Scholar] [CrossRef]
- Abdelwhab E, Hafez HM. An overview of the epidemic of highly pathogenic H5N1 avian influenza virus in Egypt: epidemiology and control challenges. Epidemiology & Infection 2011, 139, 647–57. [Google Scholar]
- Carrat F, Flahault A. Influenza vaccine: the challenge of antigenic drift. Vaccine 2007, 25, 6852–62. [Google Scholar] [CrossRef]
- Smith DJ, Lapedes AS, De Jong JC, et al. Mapping the antigenic and genetic evolution of influenza virus. science 2004, 305, 371–6. [Google Scholar] [CrossRef] [PubMed]
- Zambon, MC. Epidemiology and pathogenesis of influenza. Journal of Antimicrobial Chemotherapy 1999, 44 (suppl_2), 3–9. [Google Scholar]
- Bouvier NM, Palese P. The biology of influenza viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef] [PubMed]
- Smith GJ, Bahl J, Vijaykrishna D, et al. Dating the emergence of pandemic influenza viruses. Proceedings of the National Academy of Sciences 2009, 106, 11709–12. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger JK, Morens DM. The 1918 influenza pandemic and its legacy. Cold Spring Harbor perspectives in medicine 2019, a038695.
- Philippon DA, Wu P, Cowling BJ, Lau EH. Avian influenza human infections at the human-animal interface. The Journal of Infectious Diseases 2020, 222, 528–37. [Google Scholar] [CrossRef] [PubMed]
- Avian influenza A viruses: Clinical features. May 4 2022. https://www.cdc.gov/flu/avianflu/avian-inhumans.htm (accessed June 17, 2023).
- Mei Z, Lu S, Wu X, et al. Avian influenza A (H7N9) virus infections, Shanghai, China. Emerging infectious diseases 2013, 19, 1179. [Google Scholar]
- Yuen K-Y, Chan P, Peiris M, et al. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. The Lancet 1998, 351, 467–71. [Google Scholar] [CrossRef]
- Wang Z, Loh L, Kedzierski L, Kedzierska K. Avian influenza viruses, inflammation, and CD8+ T cell immunity. Frontiers in immunology 2016, 7, 60. [Google Scholar]
- Kuiken T, Taubenberger JK. Pathology of human influenza revisited. Vaccine 2008, 26, D59–D66. [Google Scholar] [CrossRef]
- Davis LE, Koster F, Cawthon A. Neurologic aspects of influenza viruses. Handbook of clinical neurology: Elsevier, 2014, 619-45.
- Chen E, Wang F, Lv H, et al. The first avian influenza A (H7N9) viral infection in humans in Zhejiang Province, China: a death report. Frontiers of medicine 2013, 7, 333–44. [Google Scholar] [CrossRef] [PubMed]
- Skehel JJ, Wiley DC. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annual review of biochemistry 2000, 69, 531–69. [Google Scholar] [CrossRef]
- Kobasa D, Kodihalli S, Luo M, et al. Amino acid residues contributing to the substrate specificity of the influenza A virus neuraminidase. Journal of virology 1999, 73, 6743–51. [Google Scholar] [CrossRef] [PubMed]
- Gubareva LV, Kaiser L, Hayden FG. Influenza virus neuraminidase inhibitors. The Lancet 2000, 355, 827–35. [Google Scholar] [CrossRef] [PubMed]
- Van de Sandt CE, Kreijtz JH, Rimmelzwaan GF. Evasion of influenza A viruses from innate and adaptive immune responses. Viruses 2012, 4, 1438–76. [Google Scholar] [CrossRef] [PubMed]
- Lam JH, Baumgarth N. The multifaceted B cell response to influenza virus. The journal of immunology 2019, 202, 351–9. [Google Scholar] [CrossRef]
- Murphy K, Weaver C. Janeway's immunobiology: Garland science; 2016.
- van Riet E, Ainai A, Suzuki T, Hasegawa H. Mucosal IgA responses in influenza virus infections; thoughts for vaccine design. Vaccine 2012, 30, 5893–900. [Google Scholar] [CrossRef] [PubMed]
- Staneková Z, Varečková E. Conserved epitopes of influenza A virus inducing protective immunity and their prospects for universal vaccine development. Virology journal 2010, 7, 1–13. [Google Scholar]
- Moorthy N, Poongavanam V, Pratheepa V. Viral M2 ion channel protein: a promising target for anti-influenza drug discovery. Mini reviews in medicinal chemistry 2014, 14, 819–30. [Google Scholar]
- Deyde VM, Xu X, Bright RA, et al. Surveillance of resistance to adamantanes among influenza A (H3N2) and A (H1N1) viruses isolated worldwide. The Journal of infectious diseases 2007, 196, 249–57. [Google Scholar] [CrossRef]
- Hayden FG, Shindo N. Influenza virus polymerase inhibitors in clinical development. Current opinion in infectious diseases 2019, 32, 176. [Google Scholar] [CrossRef]
- Noshi T, Kitano M, Taniguchi K, et al. In vitro characterization of baloxavir acid, a first-in-class cap-dependent endonuclease inhibitor of the influenza virus polymerase PA subunit. Antiviral research 2018, 160, 109–17. [Google Scholar] [CrossRef]
- Patel MC, Chesnokov A, Jones J, et al. Susceptibility of widely diverse influenza a viruses to PB2 polymerase inhibitor pimodivir. Antiviral research 2021, 188, 105035. [Google Scholar] [CrossRef] [PubMed]
- Shiraki K, Daikoku T. Favipiravir, an anti-influenza drug against life-threatening RNA virus infections. Pharmacology & therapeutics 2020, 209, 107512. [Google Scholar]
- Beigel J, Bray M. Current and future antiviral therapy of severe seasonal and avian influenza. Antiviral research 2008, 78, 91–102. [Google Scholar] [CrossRef]
- Nypaver C, Dehlinger C, Carter C. Influenza and influenza vaccine: a review. Journal of midwifery & women's health 2021, 66, 45–53. [Google Scholar]
- Bonnafous P, Nicolaï M-C, Taveau J-C, et al. Treatment of influenza virus with beta-propiolactone alters viral membrane fusion. Biochimica et Biophysica Acta (BBA)-Biomembranes 2014, 1838, 355–63. [Google Scholar] [CrossRef] [PubMed]
- Cox MM, Patriarca PA, Treanor J. FluBlok, a recombinant hemagglutinin influenza vaccine. Influenza and other respiratory viruses 2008, 2, 211–9. [Google Scholar] [CrossRef]
- Wang K, Holtz KM, Anderson K, Chubet R, Mahmoud W, Cox MM. Expression and purification of an influenza hemagglutinin—one step closer to a recombinant protein-based influenza vaccine. Vaccine 2006, 24, 2176–85. [Google Scholar] [CrossRef]
- Gasparini R, Amicizia D, Lai P, Panatto D. Live attenuated influenza vaccine–a review. J Prev Med Hyg 2011, 52, 95–101. [Google Scholar]
- Sridhar S, Brokstad KA, Cox RJ. Influenza vaccination strategies: comparing inactivated and live attenuated influenza vaccines. Vaccines 2015, 3, 373–89. [Google Scholar] [CrossRef]
- Goncalves P, Young J. Influenza Vaccine. Journal of Asthma & Allergy Educators 2011, 2, 44–6. [Google Scholar]
- Gottlieb T, Ben-Yedidia T. Epitope-based approaches to a universal influenza vaccine. Journal of autoimmunity 2014, 54, 15–20. [Google Scholar] [CrossRef]
- Mintaev RR, Glazkova DV, Orlova OV, Bogoslovskaya EV, Shipulin GA. Development of a Universal Epitope-Based Influenza Vaccine and Evaluation of Its Effectiveness in Mice. Vaccines 2022, 10, 534. [Google Scholar] [CrossRef]
- Pecetta S, Rappuoli R. mRNA, the beginning of a new influenza vaccine game. Proceedings of the National Academy of Sciences 2022, 119, e2217533119. [Google Scholar] [CrossRef]
- Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines—a new era in vaccinology. Nature reviews Drug discovery 2018, 17, 261–79. [Google Scholar] [CrossRef]
- Peeling RW, Sia SK. Lessons from COVID-19 for improving diagnostic access in future pandemics. Lab on a Chip 2023, 23, 1376–88. [Google Scholar] [CrossRef]
- Li L, Taeihagh A, Tan SY. A scoping review of the impacts of COVID-19 physical distancing measures on vulnerable population groups. Nature communications 2023, 14, 599. [Google Scholar] [CrossRef] [PubMed]
- Antinori A, Bausch-Jurken M. The burden of COVID-19 in the immunocompromised patient: Implications for vaccination and needs for the future. The Journal of Infectious Diseases 2023, 228 (Supplement_1), S4–S12.
Figure 1.
Global Distribution of AIV Infections among Humans from 1997 to 2023 [14,15,16,17,18,19,20,21,22,23,24]. (A) The world map illustrates the geographical distribution of human cases of various subtypes of avian influenza viruses reported from 1997 to 2021. Each marker on the map corresponds to a country with the number of cases and the year of the first reported case beside it. (B) A summary table categorizes the avian influenza virus subtypes based on reported human infections, the pathogenicity of the virus (classified as either HPAI or LPAI), and the affected areas. The table provides a quick reference to the spread and severity of each subtype, showing the total number of human cases reported since the year the first case was detected for each subtype, as well as the geographical regions where these cases were reported.
Figure 1.
Global Distribution of AIV Infections among Humans from 1997 to 2023 [14,15,16,17,18,19,20,21,22,23,24]. (A) The world map illustrates the geographical distribution of human cases of various subtypes of avian influenza viruses reported from 1997 to 2021. Each marker on the map corresponds to a country with the number of cases and the year of the first reported case beside it. (B) A summary table categorizes the avian influenza virus subtypes based on reported human infections, the pathogenicity of the virus (classified as either HPAI or LPAI), and the affected areas. The table provides a quick reference to the spread and severity of each subtype, showing the total number of human cases reported since the year the first case was detected for each subtype, as well as the geographical regions where these cases were reported.
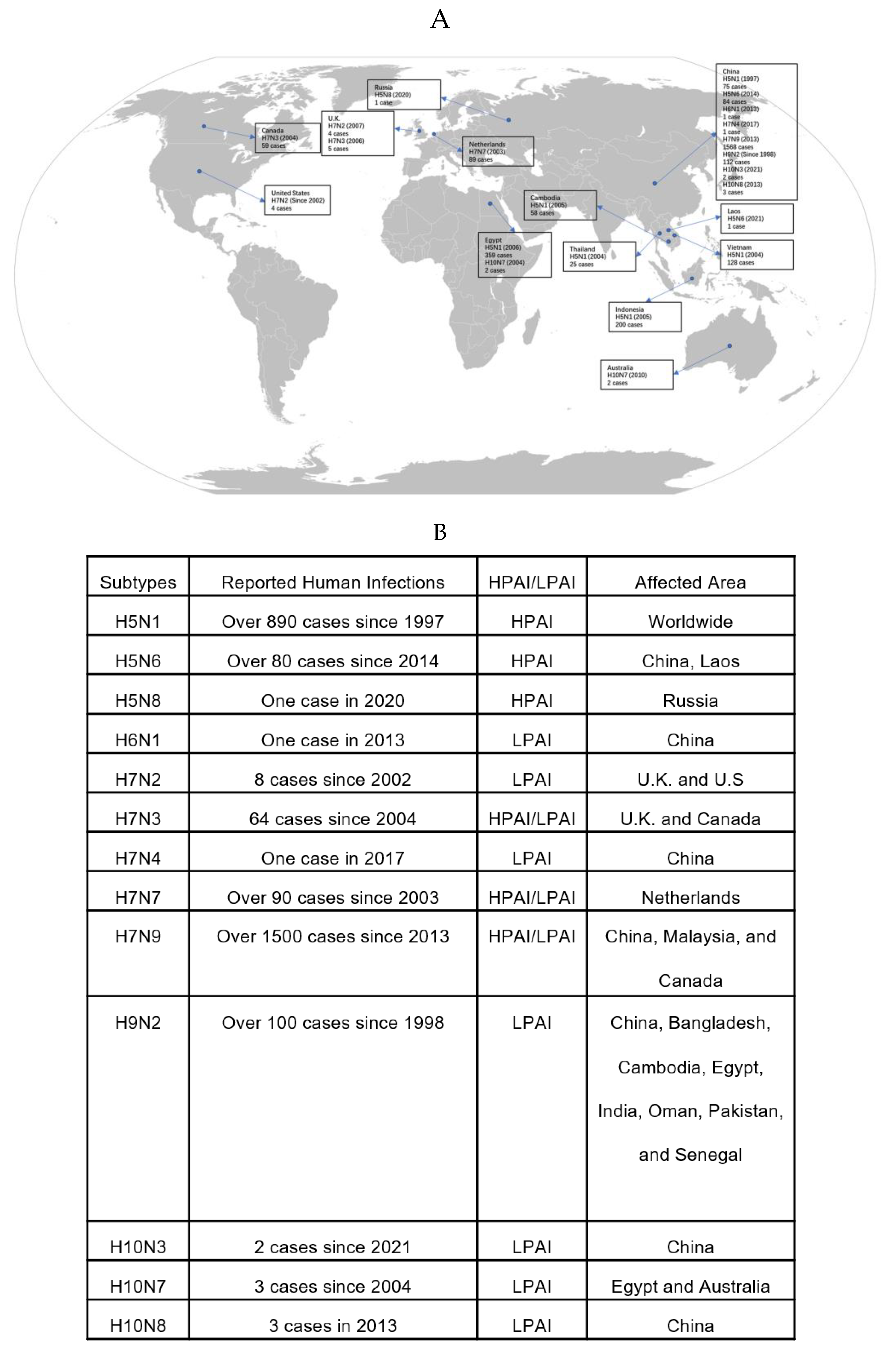
Figure 2.
Life cycle of influenza virus and sites of action of antiviral drugs. The influenza virus life cycle involves a series of steps, including (a) HA binding to sialic acid on the cell surface, leading to endocytosis; (b) M2 protein facilitating membrane fusion using M2 ion channels; (c) The viral RNA and RdRp complex releasing into the cytoplasm through fusion, followed by transportation to the nucleus for replication; (d) Viral mRNA synthesis and Cap-snatching, forming viral mRNA, leading to viral protein production; (e) Viral proteins and genomic RNA move to the cell surface assemble into new virion, bud from the membrane, and form new viral particles; (f) Neuraminidase (NA) cleaves the sialic acid-HA bond, releasing virus particles from infected cells for the next round of infection.
Figure 2.
Life cycle of influenza virus and sites of action of antiviral drugs. The influenza virus life cycle involves a series of steps, including (a) HA binding to sialic acid on the cell surface, leading to endocytosis; (b) M2 protein facilitating membrane fusion using M2 ion channels; (c) The viral RNA and RdRp complex releasing into the cytoplasm through fusion, followed by transportation to the nucleus for replication; (d) Viral mRNA synthesis and Cap-snatching, forming viral mRNA, leading to viral protein production; (e) Viral proteins and genomic RNA move to the cell surface assemble into new virion, bud from the membrane, and form new viral particles; (f) Neuraminidase (NA) cleaves the sialic acid-HA bond, releasing virus particles from infected cells for the next round of infection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



