
Submitted:
31 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
The European corn borer (ECB) and in a lesser extent the western corn rootworm (Diabrotica virgifera virgifera LeConte) are a threat to maize in the Po Valley (Northern Italy), and its control can require insecticide applications. The results of a study to assess the effects of insecticide sprays on the beneficial insect Trichogramma brassicae (Hymenoptera Trichogrammatidae) and aphid predators are reported. A three-year research was carried out in two study areas, in Lombardy. In area 1, crop rotation was a common practice, while in area 2, maize monoculture was practiced. The ordinary trend of ECB egg masses attacked by T. brassicae was affected and parasitism rates were reduced as a result of exposure to insecticides (chlorpyriphos methyl, cypermethrin, alphacypermethrine, teflubenzuron). Repeated maize crop and insecticides spraying increased the aphid population abundance and negatively affected the aphid predator community, which mainly included ladybirds, hoverflies, bugs and lacewings. In sprayed fields, managed according to repeated crop protocols, the predator community was dominated by hoverflies, whereas ladybirds and Orius spp. prevailed in maize fields managed according to crop rotation protocols. Crop rotation protocols help to prevent ECB outbreaks; when threshold limit can be at risk, inundative release of T. brassicae or microbial control (i.e. use of preparation of Bacillus thuringiensis) can integrate natural biocontrol, a valuable alternative to chemical insecticides.
Keywords:
Trichogramma brassicae
; aphids
; predators
; parasitoids
; insecticide side effects
; Northern Italy
1. Introduction
Maize (Zea mays L.) is a widespread cereal crop throughout the World, including Italy. The majority of Italian maize is cultivated in the Po floodplain (Northern Italy) [1]. The European corn borer (ECB - Ostrinia nubilalis Hübner, Lepidoptera: Crambidae) is historically the main entomological threat to maize in this area [2,3,4]. More recently the Po Valley, such as many other European agricultural districts, was invaded by the new maize pest: the Western Corn Rootworm (WCR - Diabrotica virgifera virgifera LeConte, Coleoptera: Chrysomelidae) [5].
Yield losses caused by WCR are mainly due to larvae feeding on roots. With the exception of spiders, which can prey on WCR adults, the effectiveness of natural enemies is poor [5]. Damage caused by this pest can be conveniently prevented by planning appropriate crop rotation protocols: a precaution that helps to keep the pest below the threshold limit [6].
Crop rotation and post-harvest stubble shredding help limit ECB populations; however, the damage caused by ECB larvae can be significant, especially in those agricultural districts where maize is the dominant cereal and area-wide-crop and repeated maize crop are common. In the Po Valley ECB usually develops two or three generations: adults coming from the overwintering larvae start flying in May [7]. First generation larvae pupate in July, generating moths which, in turn, will produce larvae responsible for more severe damage [1]. Probably due to the climate change, a third adult flight in mid-September and third generation larvae, which normally do not reach the maturity, were observed [4].
In the Po Valley, maize farms rely mainly on crop rotation and wild natural enemies to reduce ECB. Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) and the larval parasitoid Lydella thompsoni Herting (Diptera: Tachinidae) are useful to support such a strategy. However, where the threshold limit is particularly low (i.e. sweet maize and maize for seed) ECB is limited by insecticide sprays or inundative T. brassicae releases [2].
Natural control by Trichogramma egg parasitoids is active against ECB throughout its life cycle. [3,8,9,10].
Aphids (Homoptera: Aphidinae), in the Po Valley, are considered as maize secondary pests. Their natural enemies are mainly ladybirds (Coleoptera: Coccinellidae), hoverflies (Diptera: Syrphidae) predatory bugs (Hemiptera: Anthocoridae) and lacewings (Neuroptera: Chrysopidae).
Here we report the results of comparative observations carried out in maize fields that were located in farms which adopted chemical control protocols against WCR and ECB compared to untreated fields.
Side effects of insecticide on non-target species were assessed. T. brassicae and aphid predators were chosen as indicators of acute toxicity induced by chemical treatments.
2. Materials and Methods
2.1. Study areas
Two areas were studied. In both agricultural districts the maize is traditionally sown in March: maize hybrids usually sown belong to FAO classes 400, 500 and 600.
The first study area (study area 1) was located at 45°05’23’’N 9°04’58’’E in the municipality of Bastida Pancarana (Pavia Province, central western Po flood plain). The landscape is dominated by arable land (cereals) interspersed with legumes and alfalfa. Maize is grown for the production of feed grain or sweet corn. Maize is usually grown without irrigation and the majority of farms are managed according to crop rotation protocols. Insecticide use in maize is not the rule. Both sprayed and unsprayed fields were studied in the 2008 and 2009 seasons to assess the impact of insecticides on aphid predators and T. brassicae. In addition, aphid predators were surveyed in an unsprayed maize field in 2010. In this study area water pan traps (N =16), baited with ECB sex pheromone (bait = 3:97 Z:E-14 tda rubber stopper 0.1 mg renewed every 14 days) and phenylacetaldehyde (370 mg on 25 × 25 mm filter paper 2.7 mm thick) were installed and regularly checked (seasons 2008-2010) in order to monitor the diel flight rhythms of both ECB adult males and females [7].
The second study area (study area 2) is located in the central Po floodplain (Santo Stefano Ticino 45°29’02’’N 8°53’57’’E - Robecco sul Naviglio - 45°26’50’’N 8°52’42’’E). The degree of urbanization is significantly higher than in study area 1. Another difference concerns the final destination of the crop; maize biomass is used entirely for silage. The maize field used for the trials belonged to a farm that did not applied seasonal crop rotation. A network of canals provided plenty of water for crop irrigation. It is important to note that the maize in Robecco sul Naviglio area was not subjected to chemical spraying throughout the research period (2008-2010) and was therefore considered as a control sample in 2008 and 2009. On the other hand, Santo Stefano Ticino was sprayed in 2008 and 2009, while no insecticide was applied in 2010.
2.2. Fieldwork methods
2.2.1. Aphids
Aphids and their predators were studied in Bastida Pancarana (study area 1) and in Santo Stefano Ticino and Robecco sul Naviglio (study area 2). The first research season (2008) aimed to analyze aphid community composition, phenology and distribution on maize plant organs. Italian countryside is usually made of small sized fields. The extent of edges can affect pests and natural enemies distribution, and maize cultivations are no exception [11]. For this reason, samples included groups of maize plants located both in the inner sector of fields and along edges. Rectangular plots were used as study sites and this choice made it easier to place samples. Edge and inner samples were made visible by fixing a beam to a maize plant on top of which a red tape was tied. The edge samples were located in the middle of the minor sides, at a distance of 8 meters from the field edge. In 2008 presence/absence of aphids on stalk, ears and on ear axil leaves were assessed.
In study area 1 and 2, a sample of 160 maize plant was examined; such a sample comprised 4 subsamples of 40 plants; two subsamples were placed at opposite edges of the field. Other two subsamples were placed across the inner section of the field. Subsamples included 4 groups of subsequent 10 plants placed on a row.
In 2009 and 2010 a more informative survey was carried out: aphid abundance on each maize plant was measured according to 4 abundance classes: no presence, 1-10 specimens, 11-100 specimens, >100 specimens. Predators found on aphid-infested plants were also surveyed and identified. A sample of hoverfly pupae was collected and transferred to the lab for rearing.
Aphids and predator abundance was surveyed basically 4 weeks after insecticide application in both sprayed and unsprayed fields.
In 2008 a chlorpyriphos methyl - cypermethrin mixture (“Daskor”, 2kg/ha) – was sprayed in area 1, while in study area 2 the insecticides “Contest” (0.4 kg/ha - active ingredient: alpha-Cypermethrin) and “Cresit” (0.55 kg/ha - Teflubenzuron) were applied on 21th July. In 2009 “Contest” (0.5 kg/ha - active ingredient: alpha-Cypermethrin) was applied on 11th July in study area 2.
2.2.2. ECB egg masses parasitism by T. brassicae
ECB egg masses sampled for analysis of parasitism by T. brassicae consisted of two subsamples of at least 10 egg masses each. One subsample was collected from the edge of the maize field and the other from the center of the field. Egg masses were detected by careful examination of ears and leaves. Egg masses were sampled in both sprayed and untreated fields in study area 1.
ECB egg masses found in maize fields were transferred to a plastic Petri dish (35 mm diameter) and then reared under laboratory conditions (16:8 L:D - temperature 24-25 °C - RH 60-70%). Fresh eggs are whitish; this color does not change until hatching, except for the appearance of a central black spot (black head stage) in the absence of parasitism. If the eggs are parasitized, their color changes from whitish to blackish; if only part of the egg mass is parasitized, both colors can be observed.
Egg masses kept under laboratory conditions were checked daily using a stereomicroscope to detect any color changes. Eggs were counted when embryo development was clearly defined. Finally, the ratio of parasitized/non-parasitized eggs was calculated and expressed as a percentage.
In 2008 T. brassicae was exposed to the insecticide "Daskor" (2kg/ha) sprayed on 29 July. In 2009, insecticides were sprayed twice: a first application of "Daskor" (2 kg/ha - 29.07.2009) followed by a spray of alpha-cypermethrin on 11.08.2009 ("Contest" - 0.5 kg/ha).
2.3. Statistical analysis
The data were statistically analyzed using Biostat software. Data sets were first analyzed to assess their normal distribution using five specific tests: Kolmogorov-Smirnov/Lilliefor, Shapiro-Wilibuitk, D'Agostino skewness, D'Agostino kurtosis, D'Agostino omnibus.
ANOVA was used to compare the difference between means from different groups of data.
Contingency tables were used to analyse the relationship between two or more categorical variables.
3. Results
3.1. Impact of insecticides on aphids and their predators
3.1.1. Aphid community composition
Maize fields were colonised by aphids in June-July; at this stage of plant development they were mainly distributed at the top of the plant, next to the male inflorescence. In August aphid colonies were mainly concentrated on the ears.
Table 1 summarises the data on the frequency distribution of aphid species. The contingency table shows no significant difference between the study areas (p>0.05). The dominant role of Rhopalosiphum padi (L.) is confirmed by data obtained as a result of a survey (2008-2009) in other maize fields located around study area 1 (R. padi = 64.8% - aphid colonies N=55).
The number of maize plants hosting aphids in 2008 in the inner samples does not significantly exceed that observed in the edge samples. On the contrary, the data recorded in 2009 (Table 2) show a higher abundance of aphids in the edge samples compared to those in the inner part of the maize fields (contingency table - P<0.01). No difference was observed in 2010 (contingency table - P>0.05).
3.1.2. Aphids on maize and their predators (years 2008-2010)
Figure 1 shows data on the percentage of maize plants hosting aphid colonies (year 2008). The acronyms Spr1 and Spr2 denote samples exposed to insecticide sprays in study area 1 and study area 2, respectively.
The comparison between sprayed (Spr) and unsprayed (Uspr) fields in both study areas showed a highly significant difference (Contingency tables - P<0.001).
The same result (P<0.01) was obtained when Spr and Uspr data from the two study areas were compared (P<0.01).
In summary, aphid colony density in unsprayed samples is significantly lower than in samples from sprayed fields and both study areas show such a trend; aphid density in study area 2 is significantly higher than in study area 1; this trend affects both Spr and Uspr samples.
Regarding the natural enemies of aphids (Figure 2), the proportion of maize plants infested by aphids hosting predators in the Uspr samples is significantly higher than that recorded in the Spr fields (contingency table), both in study area 1 (P<0.01) and in area 2 (P<0.05).
It should be noted that the natural enemy samples from Uspr1, Uspr2 and Spr1 were dominated by ladybirds (>50%), whereas the natural enemy community in Spr2 was dominated by hoverflies (97%).
In 2009, insecticide applications in study area 2 induced a significantly higher abundance of aphid colonies on sprayed maize (Figure 3) compared to the control sample (contingency table - P<0.001). On the other hand, the frequency of predators on plants hosting aphid colonies was not significantly different in the two study areas (Figure 4 - contingency table - P>0.05).
In 2010, no insecticide was applied in either study area. As a result, there was a sharp decrease in aphid abundance in study area 2 compared to 2008 and 2009 (contingency table - P<0.001), although the level of infestation remained higher than that recorded in the study area 1 sample (contingency table - P<0.001).

Table 3.
Percentage of infested plants and aphid-hosting plants with predators (year 2010). Each sample included N=160 maize plants (S.E. shown).
Table 3.
Percentage of infested plants and aphid-hosting plants with predators (year 2010). Each sample included N=160 maize plants (S.E. shown).
| Sample | Plants with aphids (%) | Plants with predators (%) |
|---|---|---|
| Area 1 | 48.8±6 | 97.1±1.9 |
| Area 2 (Robecco S.N.) | 15.6±1.2 | 61.7± 7.8 |
| Area 2 (S. Stefano T.) | 57.5±2.5 | 44± 4.9 |
3.2.2. Predator community: composition and response to aphid density
The predator community included ladybirds, hoverflies, lacewings (Chrysoperla sp.), bugs (Orius sp., Nabis sp.), earwigs (Forficula sp.), beetles (Staphylinidae).
The proportion of maize plants colonized by predators tended to increase with increasing aphid infestation according to a density-dependent pattern (contingency table - P<0.01) (Figure 5).
Data on the composition of the predator community in the study areas are summarized in Table 4. Ladybirds and bugs (Orius sp.) were the most abundant predators in study area 1 from year to year. This was also the case in study area 2 in 2010, when no insecticide was applied. A significant dominance of hoverflies (97% and 56.9% in 2008 and 2009, respectively) was recorded in (Spr2), where maize monoculture was practiced and insecticides were sprayed.
Adult ladybird populations (N=191) included Hippodamia variegata Goeze (51.8%), Harmonia axyridis (Pallas) (16.8%), Propylaea quatuordecimpunctata (L.) (16.2%), Scymnus sp. (8.4%), Coccinella septempunctata L. (6.8%).
The sample of reared hoverfly pupae (N= 35) consisted of Episyrphus balteatus (De Geer) (80%) and Sphaerophoria sp. (20%). It should be noted that 80% of the hoverfly pupae were parasitised by Pachyneuron sp. (Hymenoptera: Pteromalidae) and Diplazon laetatorius (Fabricius) (Hymenoptera: Ichneumonidae).
3.2. Impact of insectides on Trichogramma brassicae
The typical development of second generation ECB egg parasitism by T. brassicae is shown in Figure 6 (unsprayed sample): the parasitism rate was moderate at the beginning of oviposition by second flight ECB females, but it tends to increase exponentially in August and usually peaks between the end of August and the first ten days of September, when parasitism rates can exceed 90% [12].
Figure 6 includes data from both unsprayed and sprayed samples recorded in 2008 and shows the parasitism evolution from the beginning of August after insecticide spraying. Parasitism in the control sample is significantly higher in the first two weeks of August (contingency table - P<0.05).
The effect of chemical spraying on sweet maize (2009 season) can also be deduced from Figure 7.
4. Discussion
This case study investigated the impact of insecticide side effects on some natural enemies of maize pests. These effects depend mainly on the chemical nature of the active ingredients and the dose applied, and can cause lethal and/or sublethal effects [13,14,15,16,17].
The aphid community is dominated by R. padi. This species has a cosmopolitan distribution and is a common pest of various cereal crops [18]. The survey of aphid predators provided further evidence of the effects of insecticide application. The abundance of aphid populations in maize was significantly lower in study area 1 than in study area 2, and such a discrepancy can reasonably be related to a different cropping pattern. Unlike study area 1, where maize was part of a rotation protocol, study area 2 was managed according to repeated maize crops. The use of insecticides in combination with repeated maize crop may have the side effect of increasing aphids, which are usually secondary maize pests in chemical-free maize crops [19,20].
The aphid predator community in chemically sprayed samples in study area 2 in 2008 and 2009 was dominated by hoverflies, while ladybirds, Orius sp. and other natural enemies were rare or absent. Such a compositional spectrum differed significantly from that recorded in unsprayed fields. In addition, the aphid predator community (Table 2) changed profoundly and rapidly when the maize field in study area 2 was sprayed: ladybirds and Orius sp. became dominant over other predators. Hoverflies are strong flyers: such an ability may suggest that they can colonize maize fields more quickly after insecticide spraying. Orius sp. is recognized as an ECB common natural enemy in maize grown in the Po Valley [21].
With regard to T. brassicae, there is an extensive bibliography on the effects of insecticide application, given the importance of this egg parasitoid as a biocontrol agent in many countries [22]. Sub-lethal effects have been demonstrated in several cases [13,14,15,16,17,23,24,25,26,27,28,29,30,31,32,33,34,35].
Our research has shown an effect of insecticide application on T. brassicae, resulting in a change in the typical parasitism trend. In 2008, a negative effect (Figure 6) was observed in the first fortnight of August, while no significant difference in parasitism rates was observed later in the season. The effect of insecticide spraying was much more pronounced in 2009 due to the double chemical application (Figure 7). Contrary to the trend of natural wild parasitism, which implies a progressive increase (Figure 6, Uspr sample), the percentage of ECB egg mortality due to T. brassicae drops sharply after the first spray and the parasitism levels recorded in the first ten days of July are not recovered by the second insecticide application (Figure 7).
The sensitivity of T. brassicae to sub-lethal doses of chlorpyrifos has been documented [13,17]: both sex pheromone communication and sex ratio were negatively affected. Parsaeyan et al. [30] demonstrated the effects of cypermethrine on T. brassicae: recommended field concentrations of this pesticide resulted in 80.7% mortality of immatures under laboratory conditions. In addition, life history parameters of the parasitoid were negatively affected compared to controls.
5. Conclusions
Selectivity is one of the desired properties of an insecticide, but often the target pest is not the only victim of spraying, and beneficial insects are usually affected to some extent [36,37].
Even if natural enemies can survive to insecticide application, sub-lethal doses can cause chronic toxicity, affecting the behavior and fitness of organisms in subsequent generations [38]. For these reasons, the conservation strategy for beneficial insects in crops where insecticide use is part of the pest management protocol is a difficult one.
Farm management is crucial in promoting or preventing pest outbreaks: for example, as this case study shows, maize monoculture tends to encourage pest density, which in turn may require insecticide treatments. Another consequence of chemical applications can be the increase of a pest other than the one targeted. Indeed, this case study reports an increase in aphid populations as a result of insecticide use and a reduction in egg parasitism by T. brassicae.
Although monoculture has been discouraged for decades, it is still a fairly common practice that tends to act as a predisposing factor for pest outbreaks.
Biological control and IPM can be a viable alternative to synthetic chemical sprays. Recent advances in research on sustainable management of ECB demonstrate that a biological control strategy based on the inundative release of T. brassicae can be a viable approach to limit the second generation of ECB larvae without increasing mycotoxin levels in grain and yield losses [1]. Such an approach is in line with the aim of reducing pesticide use, which is a core strategy of public policy aimed at restoring the environmental quality of agricultural land [39].
Author Contributions
Conceptualization, G.C., S.M. and L.L.; methodology, G.C., S.M. and L.L.; software, G.C., S.M. and L.L.; validation, G.C., S.M. and L.L.;.; formal analysis, , G.C., S.M. and L.L.; resources, , G.C., S.M. and L.L.; data curation, , G.C., S.M. and L.L.; writing—original draft preparation, , G.C., S.M. and L.L.; writing—review and editing, , G.C., S.M. and L.L.; visualization, , G.C., S.M. and L.L.; supervision, , G.C., S.M. and L.L.; project administration, L.L.; funding acquisition, L.L. All authors have read and agreed to the published version of the manuscript
Funding
This research received no external funding.
Acknowledgments
Hoverflies and their parasitoids were identified by Daniele Sommaggio. Costanza Jucker kindly supplied Diabrotica virgifera virgifera photo.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Magagnoli, S.; Lanzoni, A.; Masetti, A.; Depalo, L.; et al. Sustainability of strategies for Ostrinia nubilalis management in Northern Italy: Potential impact on beneficial arthropods and aflatoxin contamination in years with different meteorological conditions. Crop Prot. 2021, 142, 105529. [Google Scholar] [CrossRef]
- Burgio, G.; Maini, S. Control of European corn borer in sweet corn by Trichogramma brassicae Bezd. (Hym., Trichogrammatidae). J. Appl. Ent. 1995, 119, 83–87. [Google Scholar] [CrossRef]
- Camerini, G.; Groppali, R.; Tschorsnig, H.P.; Maini, S. Influence of Ostrinia nubilalis larval density and location in the maize plant on the tachinid fly Lydella thompsoni. Bull. Insectology, 2016, 69 (2), 301-306.
- Camerini, G.; Masanta, M.; Maini, S. Ostrinia nubilalis in northern Italy: egg parasitoids and control trials on three sweet pepper landraces in open field cultivations. Bull. Insectology, 2023, 76(2): 305-313.
- Camerini, G.; Franzini, L.; Groppali, R. Response of spiders (Araneae) to the introduction of an exotic pest (Diabrotica virgifera LeConte, 1868- Coleoptera, Chrysomelidae) in corn cultivations in Northern Italy. Maydica 2021, 66, 9. [Google Scholar]
- Toepfer, S.; Kiss, J.; Turoczi, G.; Komaromi, J.; Kuhlmann, U. Options for developing integrated control measures against the maize pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) in Europe. IOBC/ WPRS Bulletin 2005, 28: 1-7.
- Camerini, G.; Groppali, R.; Rama, F.; Maini, S. Semiochemicals of Ostrinia nubilalis: diel response to sex pheromone and phenylacetaldehyde in open field. Bull. Insectology, 2015, 68(1), 45-50.
- Smith, S.M. Biological control with Trichogramma: advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41(1), 375–406. [Google Scholar] [CrossRef] [PubMed]
- Pintureau, B. Les espèces europèennes de Trichogrammes. 2008. ILV Edition, 1-95.
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status, and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Caballero-López, B.; Bommarco, R.; Blanco-Moreno, J.M.; Sans, F.X.; Pujade-Villar, J.; Rundlöf, M.; Smith, H.G. Aphids and their natural enemies are differently affected by habitat features at local and landscape scales. Biol. Control, 2012, 63, 222-229. 63. [CrossRef]
- Camerini, G.; Maini, S.; Riedel, M. Ostrinia nubilalis parasitoids in Northern Italy: Past and present. Biol. Control, 2018, 12, 76. [CrossRef]
- Delpuech, J.M.; Froment, B.; Fouillet, P.; Pompanon, F.; Janillon, S.; Boulétreau, M. Inhibition of sex pheromone communications of Trichogramma brassicae (Hymenoptera) by the insecticide chlorpyrifos. Environ. Toxicol. Chem. 1998, 17(6), 1107. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Gareau, E.; Terrier, O.; Fouillet, P. Sublethal effects of the insecticide chlorpyrifos on the sex pheromonal communication of Trichogramma brassicae. Chemosphere. 1998, 36(8), 1775–1785. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Legallet, B.; Terrier, O.; Fouillet, P. Modifications of the sex pheromonal communication of Trichogramma brassicae by a sublethal dose of deltamethrin. Chemosphere. 1999, 38(4), 729–739. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Legallet, B.; Fouillet, P. Partial compensation of the sublethal effect of deltamethrin on the sex pheromonal communication of Trichogramma brassicae. Chemosphere 2001, 42, 985–991. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Meyet, J. Reduction in the sex ratio of the progeny of a parasitoid wasp (Trichogramma brassicae) surviving the insecticide chlorpyrifos. Arch. Environ. Contam. Toxicol. 2003, 45, 203–208. [Google Scholar] [CrossRef]
- Toft, S. Value of the aphid Rhopalosiphum padi as food for cereal spiders. J. Appl. Ecol. 1995, 552–560. [Google Scholar] [CrossRef]
- Dedryver, C.A. Les pucerons dans le rotations cerealieres. Perspectives Agricoles, 1980, V (37): 54-64.
- Mestres, R.; Cabanettes, J.P. Effets intentionnels ou non des traitements chimiques dans la lutte contre la pyrale. Phytoma, 1985, 369, 19-21.
- Vasileiadis, V.P.; Veres, A.; Loddo, D.; Masin, R.; Sattin, M.; Furlan, L. Careful choice of insecticides in integrated pest management strategies against Ostrinia nubilalis (Hübner) in maize conserves Orius spp. in the field. Crop prot. 2017, 97, 45-51. Crop prot. [CrossRef]
- Ko, K.; Liu, Y., Hou, M.; Babendreier, D.; Zhang, F.; Song, K. Toxicity of insecticides targeting rice planthoppers to adult and immature stages of Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Econ. Entomol., 2015, 108(1), 69-76. [CrossRef]
- Afshari, A.; Hamzepour Chenari, E.; Iraji, A.; Asghari Larimi, M. Lethal and sublethal effects of thiodicarb and hexaflumuron on egg parasitoid, Trichogramma brassicae Bezdenko under laboratory conditions. Plant Prot., 2018, 41(1), 75-89. [CrossRef]
- Delpuech, J.M.; Delahaye, M. The sublethal effects of deltamethrin on Trichogramma behaviors during the exploitation of host patches. Sci. Total Environ. 2013, 447, 274–27. [Google Scholar] [CrossRef]
- Hewa-Kapuge, S.; McDougall, S.; Hoffmann, A.A. Effects of methoxyfenozide, indoxacarb, and other insecticides on the beneficial egg parasitoid Trichogramma nr. brassicae (Hymenoptera: Trichogrammatidae) under laboratory and field conditions. J. Econ. Entomol. 2003, 96(4), 1083-1090. [CrossRef]
- Jalal, S.; Arghavan, B.; Mohammadreza, R. Side Effects of Five Pesticides on Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae). J. Agric. For. 2018, 66(1), 57–68. [Google Scholar]
- Jin, T.; Lin, Y.; Ma, G.; Liu, J.; Hao, Z.; Han, S.; Peng, Z. Biocontrol potential of Trichogramma species against Spodoptera frugiperda and their field efficacy in maize. Crop Prot. 2021, 150, 105790. [Google Scholar] [CrossRef]
- Liu, T.X.; Zhang, Y. Side effects of two reduced-risk insecticides, indoxacarb and spinosad, on two species of Trichogramma (Hymenoptera: Trichogrammatidae) on cabbage. Ecotoxicology 2012, 21, 2254–2263. [Google Scholar] [CrossRef]
- Nozad-Bonab, Z.; Hejazi, M.J.; Iranipour, S.; Arzanlou, M.; Biondi, A. Lethal and sublethal effects of synthetic and bio-insecticides on Trichogramma brassicae parasitizing Tuta absoluta. Plos one 2021, 6, e0243334. [Google Scholar] [CrossRef] [PubMed]
- Parsaeyan, E.; Safavi, S.A.; Saber, M.; Poorjavad, N. Effects of emamectin benzoate and cypermethrin on the demography of Trichogramma brassicae Bezdenko. Crop Prot. 2018, 110, 269–274. [Google Scholar] [CrossRef]
- Parsaeyan, E.; Saber, M.; Safavi, S.A.; Poorjavad, N.; Biondi, A. Side effects of chlorantraniliprole, phosalone and spinosad on the egg parasitoid, Trichogramma brassicae. Ecotoxicology. 2020, 29(7), 1052–1061. [Google Scholar] [CrossRef]
- Saber, M. Sublethal effects of methoxyfenozide and emamectin benzoate on Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae). Agric. Pest Manag., 2015, 2(1), 12-20.
- Saber, M.; Ghorbani, M.; Vaez, N.; Armak, A. Effects of diazinon and fipronil on functional response of Trichogramma brassicae Bezdenko (Hym.; Trichogrammatidae) in the laboratory conditions. J. Crop prot., 2020, 9 (2), 275-283.
- Thubru, D.P.; Firake, D.M.; Behere, G.T. Assessing risks of pesticides targeting lepidopteran pests in cruciferous ecosystems to eggs parasitoid, Trichogramma brassicae (Bezdenko). Saudi J. Biol. Sci. 2018, 25(4), 680–688. [Google Scholar] [CrossRef]
- Vaez, N.; Iranipour, S.; Hejazi, M.J. Effect of treating eggs of cotton bollworm with Bacillus thuringiensis Berliner on functional response of Trichogramma brassicae Bezdenko. Arch. Phytopathol. Plant Prot. 2013, 46(20), 2501–2511. [Google Scholar] [CrossRef]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? Proc. R. Soc. B, 2020, 287(1935), 20201265. [CrossRef]
- Helps, J.C.; Paveley, N.D.; Van Den Bosch, F. Identifying circumstances under which high insecticide dose increases or decreases resistance selection. J. Theor. Biol. 2017, 428, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Farm to Fork Strategy, 2020. 1-20.
Figure 1.
Aphid infested maize plants in sprayed (Spr) and control (Uspr) fields in 2008. Study area is indicated by 1 or 2 (S.E. is shown). Number of inspected plants/sample = 160.
Figure 1.
Aphid infested maize plants in sprayed (Spr) and control (Uspr) fields in 2008. Study area is indicated by 1 or 2 (S.E. is shown). Number of inspected plants/sample = 160.
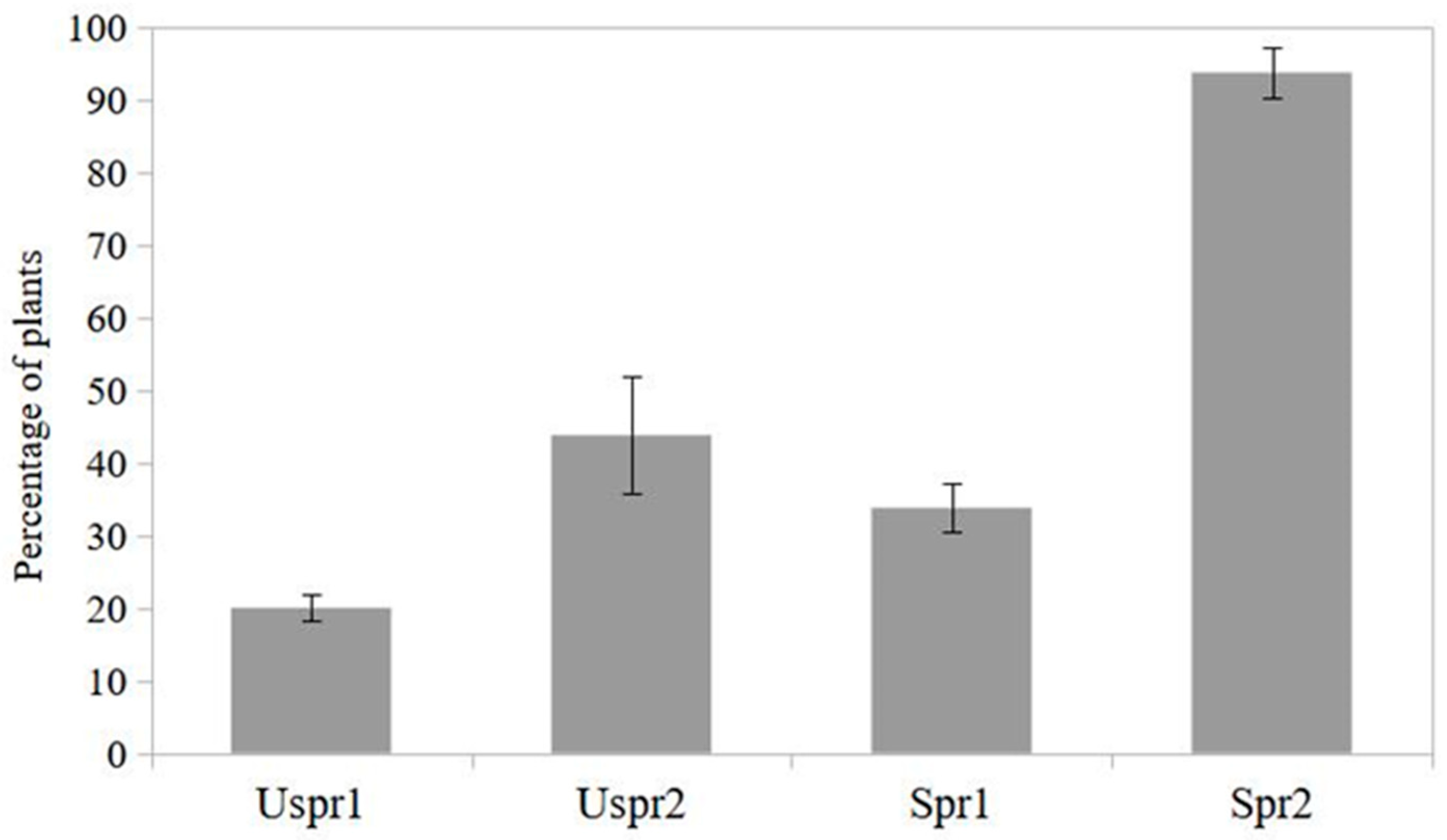
Figure 2.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
Figure 2.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
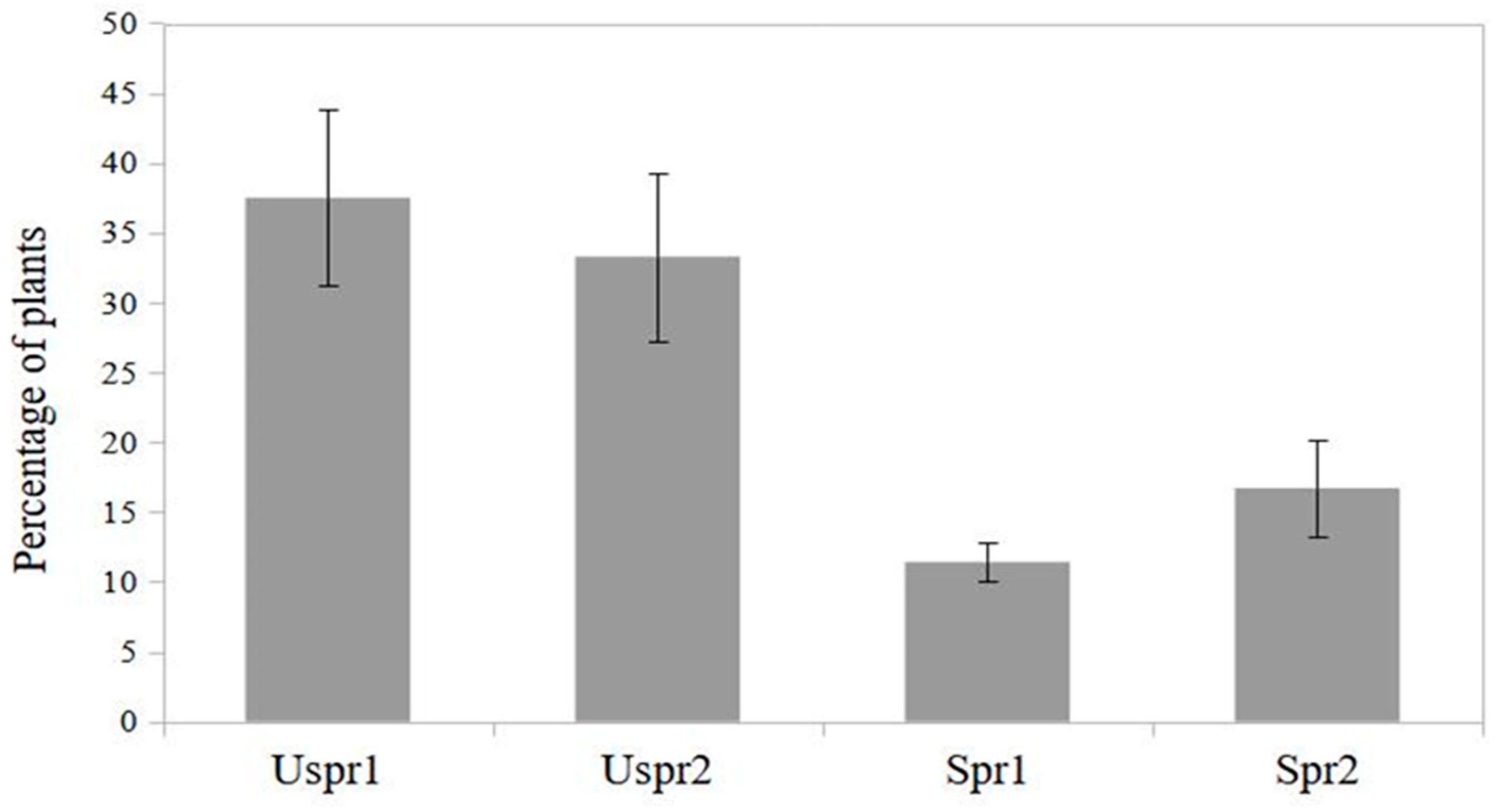
Figure 3.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
Figure 3.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
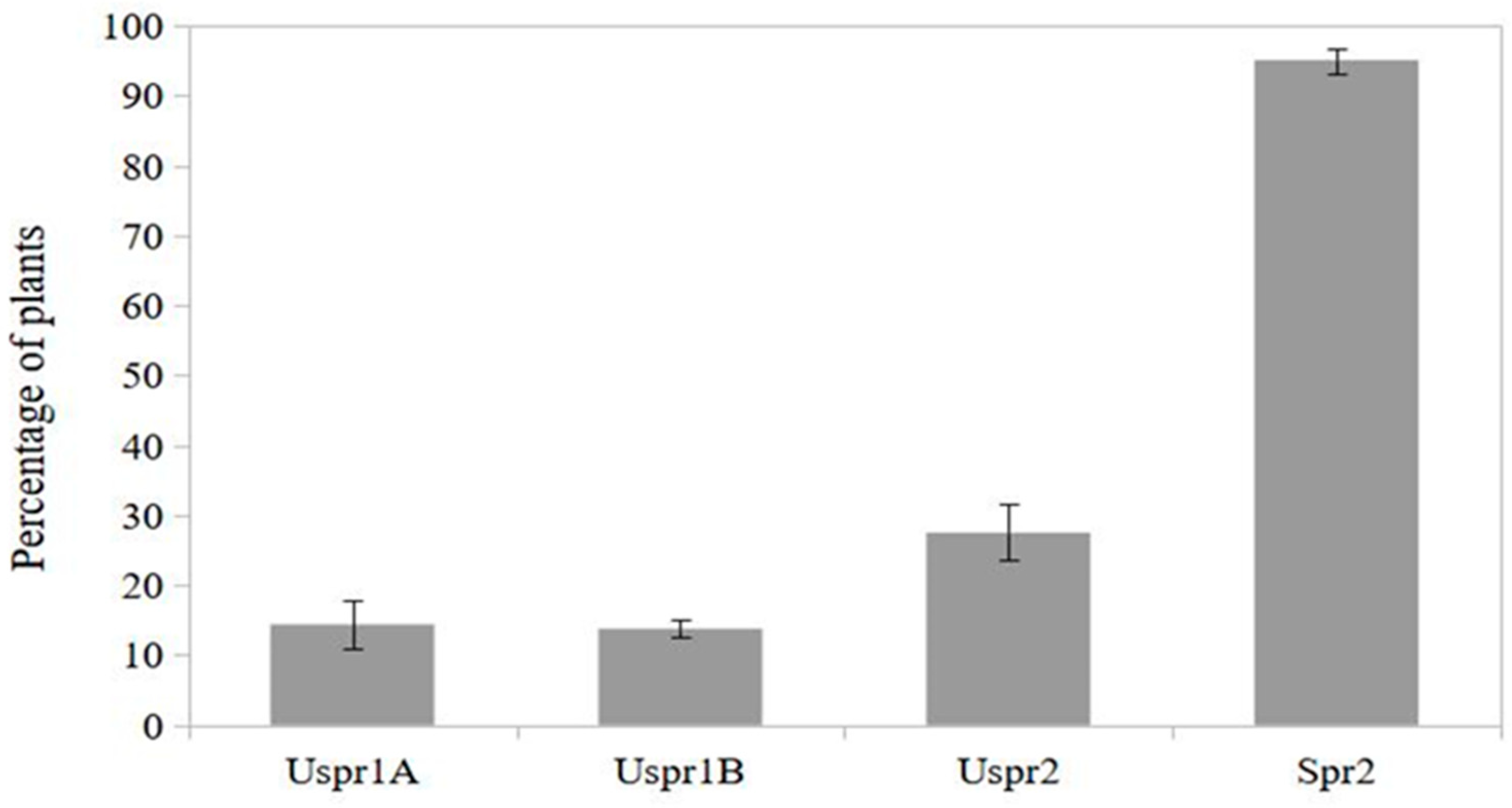
Figure 4.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
Figure 4.
Maize plants infested by aphids hosting natural enemies (%) in sprayed (Spr) and control (Uspr) samples in 2008. Study area is indicated by 1 or 2 (S.E. is shown).
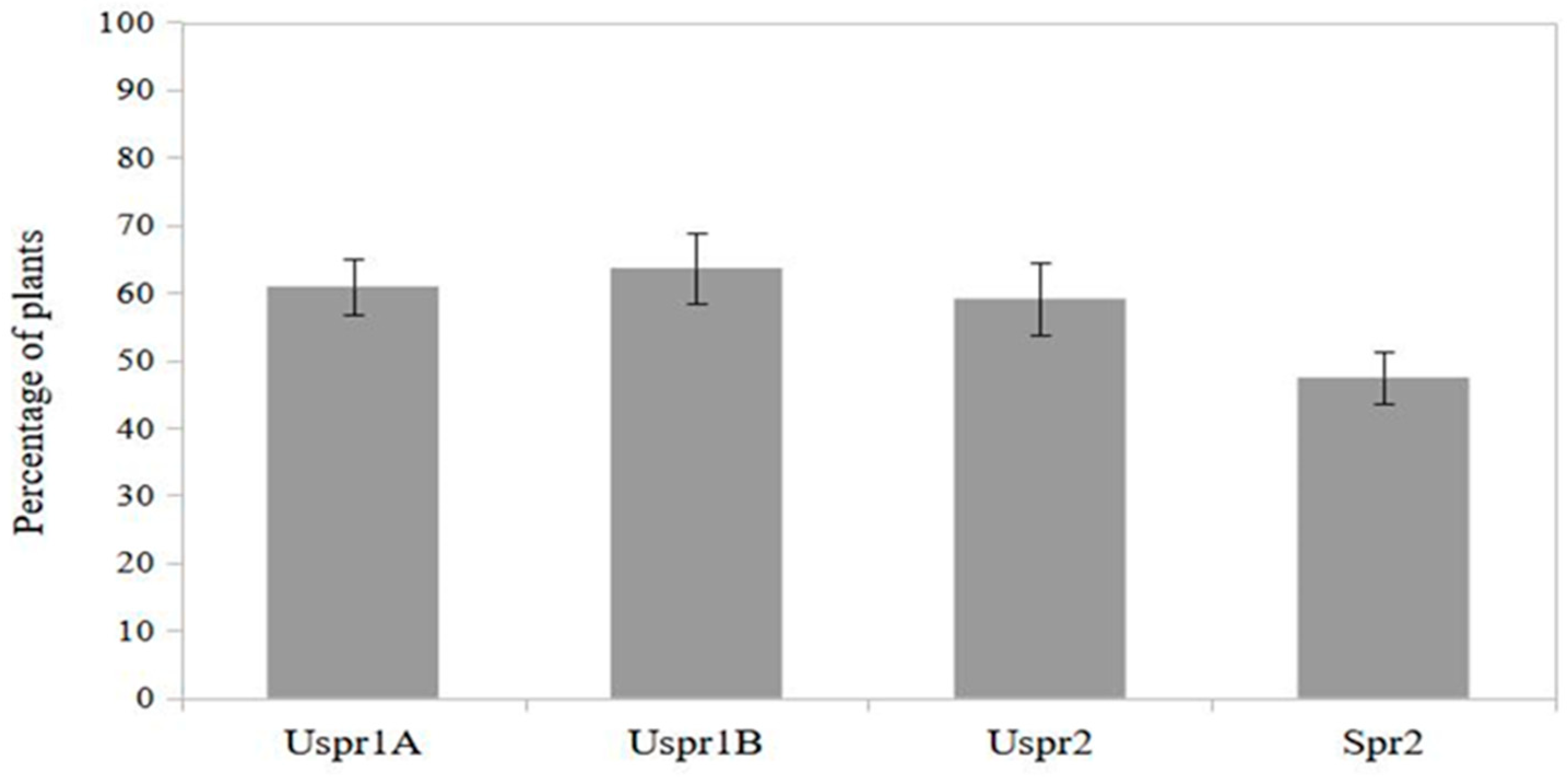
Figure 5.
Frequency distribution of maize plants colonized by predators in relation to aphid abundance; data from 2009 (N=241) and 2010 (N =195) (S.E. is shown).
Figure 5.
Frequency distribution of maize plants colonized by predators in relation to aphid abundance; data from 2009 (N=241) and 2010 (N =195) (S.E. is shown).
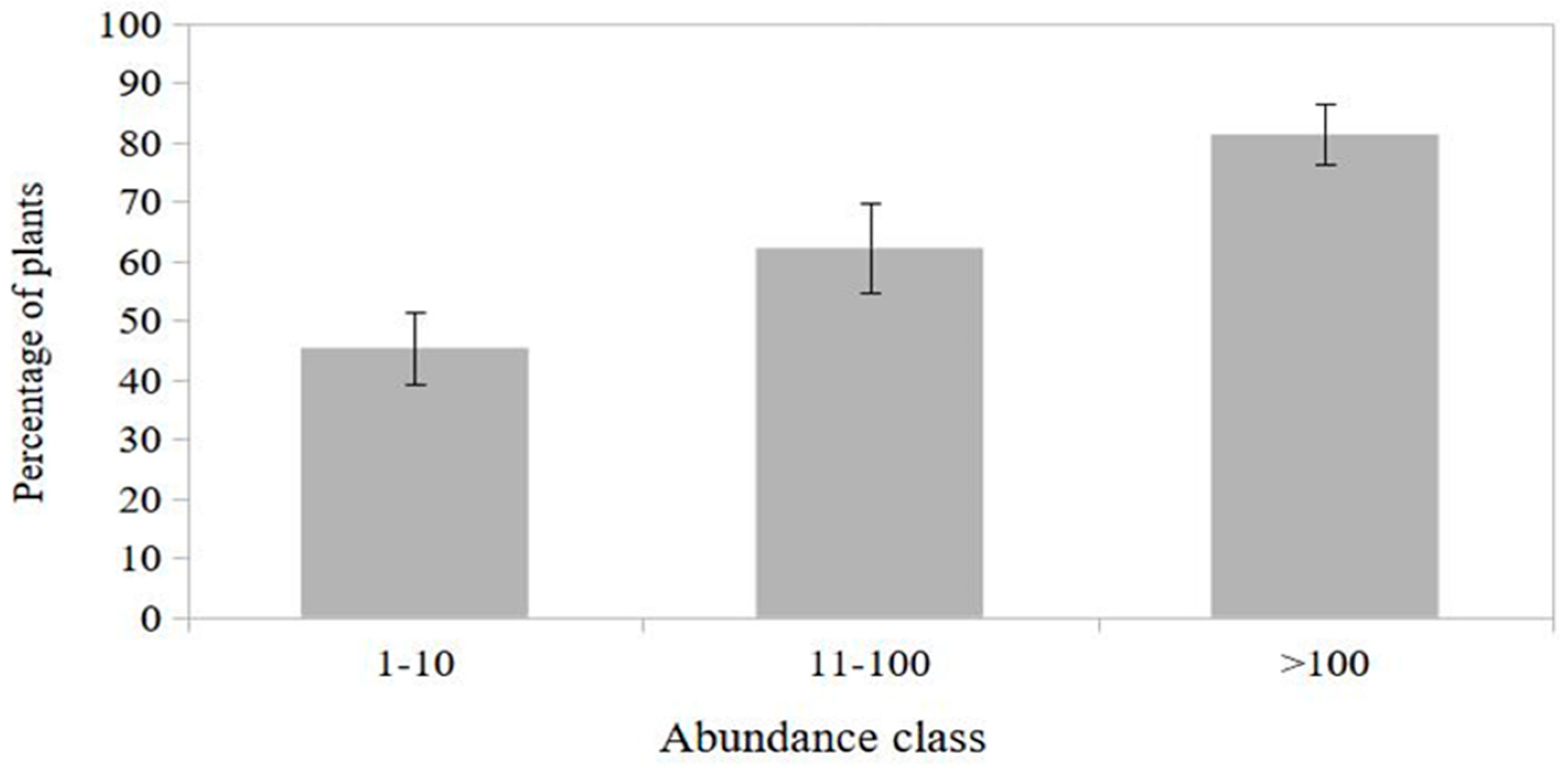
Figure 6.
Parasitism (%) of ECB eggs recorded in 2008 in unsprayed and sprayed maize fields (year 2008). The arrow points to the day (29 July) of insecticide application (Uspr: egg masses N= 88, eggs/egg mass 33.8±0.98 (S.E.); Spr: egg masses N= 91, eggs/egg mass 34±0.68 (S.E.)).
Figure 6.
Parasitism (%) of ECB eggs recorded in 2008 in unsprayed and sprayed maize fields (year 2008). The arrow points to the day (29 July) of insecticide application (Uspr: egg masses N= 88, eggs/egg mass 33.8±0.98 (S.E.); Spr: egg masses N= 91, eggs/egg mass 34±0.68 (S.E.)).

Figure 7.
Parasitism rates (%) recorded in sweet maize (year 2009). The arrows indicate the days (29 July - 11 August) when the insecticide was applied. (Uspr: egg masses N= 48, eggs/egg mass 33.83 ±8.7 S.D.); Spr: egg masses N= 41, eggs/egg mass 32.56 ±9.1 (S.D.).
Figure 7.
Parasitism rates (%) recorded in sweet maize (year 2009). The arrows indicate the days (29 July - 11 August) when the insecticide was applied. (Uspr: egg masses N= 48, eggs/egg mass 33.83 ±8.7 S.D.); Spr: egg masses N= 41, eggs/egg mass 32.56 ±9.1 (S.D.).
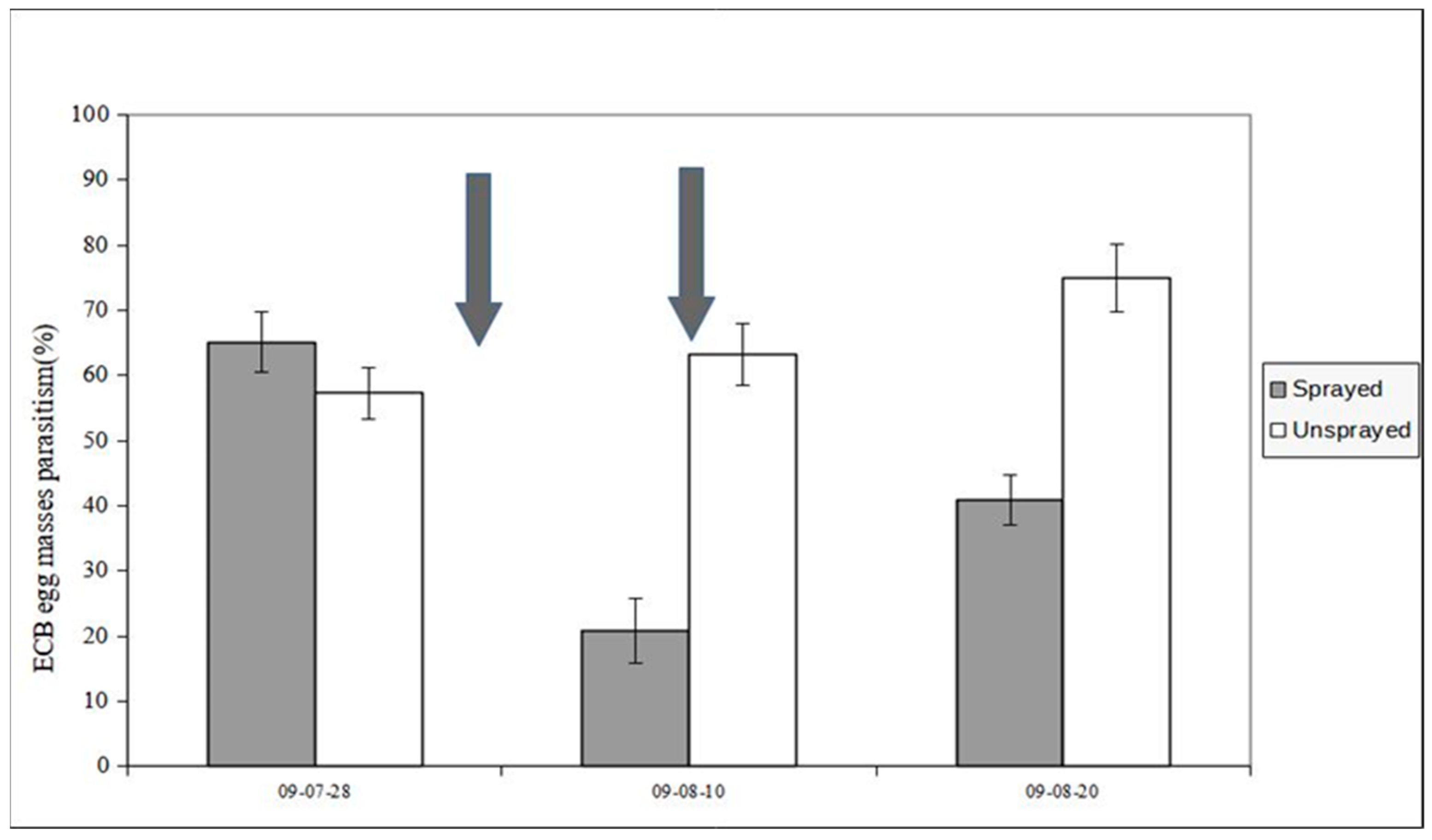
Table 1.
Frequency distribution of aphid species from samples collected in study areas 1 (N=46) and 2 (N=35) - (years 2008-2010; weighted average; samples from both sprayed and unsprayed fields).
Table 1.
Frequency distribution of aphid species from samples collected in study areas 1 (N=46) and 2 (N=35) - (years 2008-2010; weighted average; samples from both sprayed and unsprayed fields).
| Species | Area 1 | Area 2 |
|---|---|---|
| Rhopalosiphum padi (L.) | 63.8 | 59.9 |
| Rhopalosiphum maidis (Fitch) | 20.8 | 25.6 |
| Sipha maidis Passerini | 10.4 | 11.3 |
| Sitobion avenae (F.) | 4 | 3.2 |
| Aphis sp. | 0 | 0 |
Table 2.
Frequency distribution of aphid abundance in the edge and inner samples in 2009.
| Sample | Year | 1-10 | 11-100 | >100 | N |
|---|---|---|---|---|---|
| Edge | 2009 | 30.1 | 24.8 | 45.1 | 133 |
| Inner | 2009 | 27.8 | 42.6 | 29.6 | 108 |
| Edge | 2010 | 64.6 | 30.2 | 5.2 | 96 |
| Inner | 2010 | 71 | 20.4 | 8.6 | 89 |
Table 4.
Predator frequency (%) in study areas (year 2008 - N=96; year 2009 - N= 200; year 2010 - N= 185). Data from 2008 and 2009 include sprayed and unsprayed samples. No insecticide was sprayed in 2010 in either study area.
Table 4.
Predator frequency (%) in study areas (year 2008 - N=96; year 2009 - N= 200; year 2010 - N= 185). Data from 2008 and 2009 include sprayed and unsprayed samples. No insecticide was sprayed in 2010 in either study area.
| Predators | Year 2008 | Year 2009 | Year 2010 | |||
|---|---|---|---|---|---|---|
| Area 1 | Area 2 | Area 1 | Area 2 | Area 1 | Area 2 | |
| Ladybirds | 84.3 | 26.4 | 38.4 | 26.4 | 78.3 | 26.7 |
| Orius | 9.1 | 18.5 | 37.8 | 19.2 | 13 | 63.9 |
| Hoverflies | 6.2 | 49.9 | 9.7 | 33.4 | 1.7 | 7.5 |
| Lacewings | 0.3 | 0.2 | 9.4 | 17.7 | 4.4 | 1.9 |
| Others | 0.1 | 5 | 4.7 | 3.3 | 2.6 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



