
Submitted:
31 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Intratumoral immune cytolytic activity (CYT), measured as the geometric mean of the expression of Granzyme-A (GZMA) and Perforin-1 (PRF1), has emerged as a critical factor in cancer immunotherapy, with significant implications for patient prognosis and treatment outcomes. Here, we review how different immune checkpoint pathways, the composition of Tumor Microenvironment (TME), antigen presentation and metabolic pathways, all regulate CYT. We also discuss the various methods through which CYT can be assessed. The detection and analysis of tumor-infiltrating lymphocytes using flow cytometry or immunohistochemistry provide important information on the immune cell populations within the TME. Gene expression profiling and spatial analysis techniques, such as multiplex immunofluorescence and imaging mass cytometry, allow researchers to study CYT in the context of the TME. We further report the significant clinical implications that CYT has, as its increased levels are associated with positive clinical outcomes and a favorable prognosis. Moreover, CYT can be used as a prognostic biomarker and aid in patient stratification. Measuring CYT through different methods offers promising paths for improving treatment responses. Overall, we highlight that the understanding and modulation of CYT levels is critical for improving cancer immunotherapy. Research into CYT and the factors that influence it has the potential to transform cancer treatment and improve patient outcomes.
Keywords:
Cytolytic Activity (CYT)
; Tumor-Infiltrating Lymphocytes (TILs)
; Gene Expression Profiling
; Spatial Analysis Techniques
; Immune Checkpoint Pathways
; Tumor Microenvironment (TME)
; Antigen Presentation
; Metabolic Pathways
1. Introduction
1.1. Background on Cancer and the Immune System
Cancer is the second leading cause of death, worldwide and both the incidence of cancer and cancer-related mortality rates are constantly increasing. It is estimated that until 2060, cancer will be the main cause of death [1].
The immune system is a defence mechanism against infected antigens and self-antigens via a suitable balance between inhibition and activation of immune responses [2]. It also actively participates in cancer prevention, progression and therapy [3]. Cytotoxic T-cells (CTLs) via their T-cell Receptors (TCRs) recognize antigens derived from cancer cells and bound to Major Histocompatibility Complex (MHC) molecules on the surface of Antigen Presenting Cells (APCs) [4]. Moreover, the immune system creates immunological memory through the adaptive immune responses for a future more rapidly and effective response against the same antigens [5,6].
Tumors can evade the immune response, using various mechanisms, including the inhibition of the immune system, inducing T-cells exhaustion and restricting antigen recognition [7]. Evasion of cancer cells from the immune system can be defined as one of the cancer hallmarks [8]. Furthermore, cancer cells proliferate and grow faster than the immune system can manage, resulting in the escape of an attack from immune cells. To achieve this, cancer cells create in their surrounding the TME which affects the efficacy of immune cell activation [9].
The TME plays a crucial role in cancer progression and cell migration [10]. It is composed of cancer cells, blood vessels, Extracellular Matrix (ECM), stromal cells and immune cells, including Natural Killer (NK) cells, B-cells, T-cells, Neutrophils, Myeloid-Derived Suppressor Cells (MDMCs) and Tumor-Associated Macrophages (TAMs) [11]. The interplay between the cellular and extracellular components of TME contributes to early diagnosis of cancer [12].
Cancer immunotherapy has enabled never-before-seen success rates in durable tumor control and enhanced survival benefit in patients with advanced cancers. There are two different forms of immunotherapy, the Immune Checkpoint Inhibitors (ICIs) and Adoptive Cellular Therapy (ACT) [13]. ICIs using specific monoclonal antibodies (mabs), including anti-programmed death-1 (PD-1)/programmed death-ligand 1 (PD-L1) and/or anti-cytotoxic T lymphocyte antigen-4 (CTLA-4) to treat several cancers. Specifically, anti-PD-1 and/or anti-CTLA-4 have increased the overall survival of patients with melanoma [14].
There are currently eight (8) approved ICIs to treat 18 different types of cancers, comprising tumors with deficient mismatch repair system/microsatellite instability—high (dMMR/MSI-H), lung cancer, renal cell carcinoma (RCC), ovarian cancer, gastric cancer (GC) and Hodgkin’s lymphoma [15,16,17]. Nevertheless, many cancers create resistance to ICI therapies and do not respond to them.
1.2. The Role and Significance of Immune Cytolytic Activity in Tumor Control
CTLs and NK cells can target and kill target cells, using various mechanisms. One of the major mechanisms that CTLs kill cancer cells, is the granzyme/perforin pathway [18]. More specifically, upon recognition of the targeted cells, CTLs and NK cells release granzymes and perforins to enter and lead them to apoptosis [19].
Perforins are responsible to open pores in the membrane of the targeted cancer cells, facilitating the entry of the granzymes into them, which eventually eliminate cancer cells in an apoptotic manner (Figure 1) [20,21]. CYT is a new index of immune activation within a tumor and it is calculated by the expression levels of Granzyme-A (GZMA) and Perforin-1 (PRF1) mRNA expression levels [20,22].
In 2015, CYT levels were quantified and presented by Rooney et al. as a new index of local immune infiltrate , across 18 tumor types [23]. CYT index varies across different cancer types. Roufas et al. quantified the transcription levels of GZMA and PRF1 and acute myeloid leukemia, pleural mesothelioma, sarcoma, and stomach cancer are associated with high affinity in CYT. On the other hand, ovarian, liver, thyroid, esophageal, and prostate cancers, glioblastoma, glioma and adrenocortical carcinoma and uveal melanoma, shown the lowest CYT levels. Finally, head cancer, melanoma and neck cancer had considerably higher CYT levels, compared to the equivalent normal tissues [22].
In CRC, CYT is associated with different mutational events and high CYT tumors were related with increased Tumor Mutational Burden (TMB) [24,25,26] and upregulated the expression levels of immune checkpoint inhibitors, including PD-1, PD-L1/2, CTLA-4, Lymphocyte Activation Gene 3 (LAG-3) and Indoleamine 2, 3-Dioxygenase 1 (IDO-1) [22,24].
Moreover, high CYT levels were associated with high MSI status and better survival rates of patients with CRC, hepatocellular carcinoma (HCC) and triple-negative breast cancer (TNBC) [24,27,28,29].
CYT has a significant role in cancer control and progression and can be also used as a prognostic biomarker for reflecting immune status in hepatocellular carcinoma (HCC) [30], immunity and clinical outcome [25]. In addition, CYT-high skin cutaneous melanoma (SKCM) patients have better prognosis [31].
Although available therapies improved the prognosis for many patients with cancer, most patients still have a poor prognosis, or they develop resistance to them. Therefore, the mechanisms being responsible for the aberrations taking place within the TME need to be investigated further. Herein, we investigated the factors influencing CYT in cancer in order to develop better therapeutic approaches, identify new therapeutic targets or potential biomarkers and eventually help cancer patients manage better their disease, enhance their life span and increase their life span.
2. Results
2.1. Methods for Assesing CYT
2.1.1. Tumor-Infiltrating Lymphocytes
- Isolation and Characterization of TILs
TILs have the ability to recognize and kill autologous cancer cells, both in vitro and in vivo [32] and their isolation, especially in early stages of cancer is very important [33].
Isolation and characterizations of TILs have been evolved as TILs can be used for prognsis and therapeutic effects in tumors. Phenotypic analysis of TILs isolated from renal cell carcinoma (RCC), showed that mostly of TILs were composed of CD4+, CD8+ and CD56+ cells in different numbers [34]. As it is already mentioned, CYT is strictly related with the population and activity of CTLs. Therefore, CYT can be related also with the populations of TILs.
Moreover, isolation of TILs from tumor and non-tumor liver tissues of resected HCC patients was used to analyse the immune gene expression profiles and the results shown that the activation and proliferation of TILs at inflammatory immune microenvironment enhance the survival of HCC patients and the tumor progression [35].
Currently, immunotherapies using TILs have earned attention due to their efficacy and the failure of conventional immunotherapies [36]. Spatial characterization of TILs have been demonstrated as a key prognostic biomarker to predict treatment response in breast cancer [37].
- Analysis of TILs by flow cytometry or immunohistochemistry
TILs were expanded and analysed using different in vitro methodologies, including flow cytometry analysis and immunohistochemistry. In a study in 2017 TILs were surgically expanded from 19 patients with pancreatic adenocarcinoma in the presence of IL-2 and were characterized phenotypically by flow cytometry analysis. The results showed that the majority of TILs were composed of CD4+ T cells and CD8+ T cells [38].
In 1994, expanded TILs derived from NSCLC have been used in adoptive immunotherapy following surgery with promising results [32]. Flow cytometric immunophenotypic analysis of TILs immunophenotypes can predict both clinical effectiveness of immunotherapy [39] and early relapse in localized clear cell RCC [40].
2.1.2. Gene Expression Profiling
- Identification of Cytolytic Markers
Upon activation, CTLs and NK cells mediate cytolysis of targeted cells using various mechanisms. One of this, is the granzyme-perforin pathway which is strictly associated with the CYT. Specifically, CTLs and NK cells upon their expose to infected/cancer cells, release cytolytic markers, such as granzymes and PRF1 to open pores, enter to the targeted cells and lead them to apoptosis [19,41].
GZMA and PRF1 are two key cytolytic effectors whose transcription levels are used for the quantification measurement of immune CYT. The expression of both GZMA and PRF1 is significantly affected upon CD8+ T cell activation [42].
- Measurement of CYT through RNA expression
2.1.3. Spatial Analysis Techniques
- Multiplex Immunofluorescence (mIF) and Imaging Mass Cytometry (IMC)
mIF and IMC are tissue imaging technologies used to detect multiple markers on a single tissue slide and their combination has been recently demonstrated as an accurate imaging method for single-cell segmentation [44].
mIF technique allows the simultaneous accurate detection of a variety of immune markers in different tissues types [45]. It has the unique ability to study the spatial interaction between immune cells and tumor cells without affecting the architectural features of the tumor [46]
IMC is based on the principles of flow cytometry analysis and Mass Spectrometry (MS) and it uses metal isotope-labelled antibodies that enables highly multiplexed imaging analysis of proteins, isotopes and markers within tumor tissues [47,48]. In 2021, it was used to characterize the TME of metastatic melanoma patients who received immunotherapy in order to find indicative biomarkers of treatment response with very promising results [49].
Among available spatial analysis techniques, both mIF and IMC are very significant in the field of immuno-oncology and they are used to study and understand better the TME in order to create more effective therapies and identify predictive biomarkers of response to immunotherapy [50].
- Assessment of CYT within the TME
Immune CYT and immune cells within the TME are strictly related. CTLs and NK cells secret granzymes and perforins such as GZMA and PRF1 to kill cancer cells. The population of the above-mentioned immune cell types within the TME is strictly associated with high levels of immune CYT and better survival [23,30,51]. On top of CD8+ T cells and NK cells, in GC the CYT score was associated with high infiltration of macrophages and low infiltration of Tregs [25].
Furthermore, CYT-high primary and metastatic skin melanoma tumors were enriched in CD8+ T cells, B cells and M1 macrophages, while CYT-low tumors were enriched in CD4+ T cells, monocytes and NK cells [52]. In glioblastoma, high CYT levels were related with higher infiltration of immunosuppressive cells including neutrophils and M2 macrophages and worst survival [43].
As immune CYT and immune cells are closely associated within the TME, it is considered that immune CYT can be assessment within the TME.
2.2. Factor Influencing CYT
2.2.1. Tumor Intrinsic Factors
- Tumor Mutational Burden
The TMB is defined as the total number of genetic mutations in the DNA of cancer cells [53]. It is very significant index for doctors in the selection of the most effective treatment for each cancer patient.
Calculation of TMB is done by using different methods. The TMB was first measured from Whole-Exome Sequencing (WES), included only non-somatic mutations by the Next-Generation Sequencing (NGS) technology [54]. Both WES and NGS are used to measure the TMB [55].
Accumulating evidence suggests that TMB is a key predictive biomarker in cancer therapy. From the majority of studies it is clear that a high-TMB is related with clinical efficacy of ICI therapy in multiple cancer types [53,56]. It is worth mentioning that, between of all available immunotherapies and 27 different tumor types or subtypes, high-TMB is associated with better response to anti-PD-1 [57].
In 2015, a high TMB was strongly associated with better response of NSCLC patients to anti-PD-1 immunotherapy and longer Progression-Free Survival (PFS) [58]. Moreover, NSCLC patients with high TMB score, who received nivolumab (anti-PD-L1) had longer PFS and higher Objective Response Rate (ORR), than the patients who received chemotherapy [59]. On top of anti-PD-L1 alone, combination immunotherapy of nivolumab and ipilimumab in high-TMB patients with advanced NSCLC, showed a longer PFS than chemotherapy treatment [60].
The TMB is also linked with other emerging biomarkers. Specifically, MSI-H/dMMR tumors have been identified to exhibit high TMB and they also associated with better response to ICI therapy [61]. Besides of MSI-H, high TMB correlated with increased CYT and downregulated of various of immune checkpoint inhibitors in colon cancer [24].
More and more studies support that the TMB is a promising predictive biomarker, and its evaluation plays a key role in immuno-oncology. In addition, TMB could be very valuable in treatment selection for ICI therapy [55].
- Antigen Presentation and MHC Expression
APCs constitute an heterogenous group of immune cells that are responsible to process and present antigens from recognition to T cells through MHC [62]. APCs can be divided into professional, with haematopoietic origin and include B lymphocytes, macrophages and DCs and nonprofessional APCs that are not bone marrow derived and include hepatocytes and fibroblasts [63].
MHC proteins are key components of adaptive immunity and categorized in two different classes [64]. Both classes of proteins present antigens on the surface of APCs immune system and specifically to T cells [65].
MHC class I present antigens from virally infected cells to CD8+ T lymphocytes to kill them [66], however many human viruses develop proteins that interfere in the presenting procedure and enhance virally infected cells to escape the detection and destruction from immune system [67].
As opposed to MHC class I, MHC class II molecules are responsible to present antigens to CD4(+) T lymphocytes [68]. Moreover, the ability of MHC class I to present to APCs derived-peptides from cells, allow to CD8+ T cells to recognize and kill cells, e.g. cancer cells that synthesized abnormal proteins. It has been reported that loss of MHC class I antigen presentation leads to evasion of cancer cells from immune system [69]. It has been observed that some human cancers, including gastric cancer and melanoma are due to loss of MHC I expression [70,71].
Moreover, CD4+ T lymphocytes have a pivotal role in preventing tumor growth and targeting of the MHC class II antigen presentation pathway can be used to develop efficient vaccines to activate the immune system and enhance the immune responses against cancer [72]. There are also some types of tumors, such as solid tumors do not express MHC class II molecules, and the participation of CD4+ T lymphocytes depends exclusively on infiltrating APCs [73].
Both MHC I and II pathways are taking into consideration in order to improve immunotherapy.
Figure 2.
Factors Influencing CYT.

2.2.2. Tumor Microenvironment Factors
- Immunosuppressive cells
The TME has a pivotal role both in cancer progression [74] and affect the response of cancer patients to therapies [75]. It is composed by both cellular and extracellular components, including the category of immunosuppressive cells [11].
Regulatory T cells (Tregs) and Myeloid-Derived Suppressor Cells (MDSCs) are critical cells related with immune resistance [76], and they enhance the escape of cancer cells from the immune system [77]. Immunosuppressive cells have significant clinical value and targeting of them can enhance the effectiveness of immunotherapies and converse the immune resistance [76].
MDSCs are immature progenitor cells that suppress the immune responses and categorized into 2 subcategories depending on the mechanism of their action: the Monocytic (M-) MDSCs and the Granulocytic (G-) MDSCs [78]. Moreover, a subpopulation of T cells, the Tregs promote tumor progression [79] by suppressing both the proliferation of T-cells and the production of cytokines, to maintain immune homeostasis and self-tolerance [80].
More studies suggest that the elimination of immunosuppressive cells within the TME can inhibit the tumor progression. Specifically, inactivation or decrease in the number of MDSCs enhance the anti-tumor immunity and reverse the status of TME from immunosuppressive to immune-activate [81,82]. In addition, ICI therapies using anti-PD-1/anti-PD-L1 and or anti-CTLA-4 mabs decrease the number of Tregs by increasing to the number of CD8+ T cells [83]. Chemotherapy can eliminate the number of Tregs and MDCSs in the TME [84].
Immune evasion constitutes one of the hallmarks of cancer progression [81]. Immunosuppressive cells inhibit antitumor immune responses and enhance tumor immune escape, resulting to tumor cell extravasation and metastasis [77]. Additionally, in order to avoid immune cells, cancer cells enlist Tregs to upregulate tumor antigen expression, active inhibitory immune checkpoint molecules and create an immunosuppressive TME [85,86]. Immunosuppressive cells are critical in cancer progression and metastasis.
- Immune checkpoints
Immune Checkpoint Proteins (ICPs) are expressed on the surface of cancer cells to inhibit T cell-mediated immune responses and they are using to treat many different types of cancer [87].
ICI immunotherapy is used to bind to ICP, active T cells and allow them to kill cancer cells. It is currently one of the most novel and promising cancer therapies [88].
There are different immune checkpoint inhibitors that used to treat a range of cancer types. The most well-studied ICIs consists of anti-CTLA-4, anti-PD-1 and anti-PD-L1 [88,89].
In 2011, Ipilimumab, an anti-CTLA-4 ICI, was successfully approved and introduced a promising new form for cancer therapy [90]. Anti-PD-1 (nivolumab, pembrolizumab and cemiplimab) and anti-PD-L1 (atezolizumab avekumab and durvalumab) were then received the Food and Drugs Administration’s (FDA) approval and they are using to treat about 15 different types of tumor [91] (Figure 3).
Moreover, other studies have been focused to study for other antibodies against other immune checkpoint proteins, including V-domain immunoglobulin-containing suppressor of T-cell activation (VISTA), lymphocyte activation gene-3 (LAG-3), B7-homolog 3 (B7-H3) and T cell immunoglobulin, and mucin domain 3 (TIM-3), [89].
ICI therapy is a vital and very promising form of therapy to treat cancer [92], however only a small number of cancer patients respond to this form of immunotherapy. Most of the ICPs are expressed on different type of immune cells and their expression varies between a cell type [88]. In addition, it was acknowledged that the TMB, MSI-H, dMMR, neoantigen expression and certain gene mutations are predictive biomarkers for the efficacy of ICI therapies to cancer patients [56,93]
ICPs influence CYT and high CYT increases the expression of immune checkpoint inhibitors and for this, CYT-high tumors are better candidates to respond to ICI therapies [26,27,94].
ICI therapies targeting CTLA-4 and/or PD-1/PD-L1 improved outcomes and survival for many patients with different types of cancer. Despite the challenges, additional inhibitory pathways and immune checkpoint proteins must be explored in order to apply the ICI therapies to more cancer patients and improve the responses of cancer patients to it.
- Stromal Cells and Extracellular Matrix Components
Stromal cells have fundamental roles in health and disease and they are responsible for the building and infrastructure of organs [95]. Both the presence of stromal cells in TME and their interplay with cancer cells are crucial for the cancer initiation. More studies increasingly show that stromal cells and their products, actively participate and promote tumorigenesis [96].
In pancreatic cancer (PC), neoplastic and cancer cells create a specific environment which enhance the malignant properties of cancer cells [97]. Multipotent Stromal Cells (MSCs) a subpopulation of stromal cells is present in multiple tissues and have the ability to differentiate in different lineages, encompassing osteoblasts, chondrocytes and adipocytes. In cancer, MSCs can migrate from primary tumors to metastatic organs, contributing to the progression of carcinogenesis. Moreover, cancer cells enhance the migration of MSCs and their crosstalk is crucial in tumor development and can be targeted for therapeutic approaches [98].
Another form of stromal cells that are present in TME are the cancer associated fibroblasts (CAFs). They promote tumor growth, invasion, angiogenesis and metastasis [99]. Moreover, CAFs can be activated by TAMs and enhance the development of malignant tumors [100]. In breast cancer, CAFs were associated with therapeutic resistance [101].
The crosstalk of immune cells and stromal cells is crucial in both cancer progression and anticancer activity. In addition, immune cells and stromal cells conduce to immune suppression within the TME and induce ani-tumor immunity [102].
Extracellular Matrix (ECM) is a three-dimensional highly dynamic structural network that has a pivotal role in normal development as well as in disease development and progression [103]. It is composed of different macromolecules, including collagens, glycoproteins, elastin, proteoglycans, fibronectin, etc., which join each other as well as cell-surface receptors, creating a scaffold for cells and present in all tissues and organs [104]. The interaction between ECM components is crucial in different cellular processes and functions, including proliferation, differentiation, migration and survival [105].
2.2.3. Host Factors
- Genetic Variations and Polymorphisms
Genetic sequences called polymorphisms and/or variants are different DNA sequences among individuals compared to the reference genome with different functional significance. DNA polymorphisms occur in ≥ 1% of the population and the majority of them do not have impact on protein or gene function.
However, some polymorphisms affect the function of proteins and/or genes, called mutations with significant value on cancer progression. There are four types of DNA polymorphisms: The Single Nucleotide Polymorphism (SNP), the Restriction Fragment Length Polymorphism (RFLP), the Simple Sequence Repeat (SSR) and the Variable Number or Tandem Repeats (VNTR) [106].
The SNPs are the most common polymorphism and it is estimated that they occur every 1000 base pairs usually found in protein-coding regions [107]. SNPs are also used as a tool to identify determinant of different diseases [108]. In addition, SSRs are the most frequently used in mapping studies due to they are highly polymorphic, very informatic and consistent [109].
The role of polymorphisms in cancer has been evaluated and it is found that gene polymorphisms are implicated in different stages of tumor development and cancer progression. Specifically, polymorphisms affect tumor growth, invasion, metastasis and respond to cancer therapy [110].
- Immune System Composition and Function
The immune system is very important in cancer management. Its failure to recognize and kill cancer cells contributes to cell migration and cancer progression [10,92].
It is composed of different types of cells, cytokines, proteins and soluble bioactive molecules. Each of them has different role to recognize and defend against ‘’foreign’’ antigens or proteins [5]. NK cells and phagocytes, including macrophages, neutrophils and monocytes are the key players in cell-mediated innate immune responses. NK cells act using the MHC I complex proteins and phagocytes facilitate immune protection either by swallowing cells that express non-self-antigens or by using lysosomal enzymes [111]. Innate immune responses also include other cells such as basophils and eosinophils which using different inflammatory mediators to attract more immune cells to the inflammation/injured site [112].
The adaptive immunity is antigen-dependent and antigen-specific [113] and it is comprised of lymphocytes and APCs. Lymphocytes are categorized into two major types: the B cells that mature in the bone marrow and the T cells that mature in the thymus and their function is to recognize specific antigen that present on the surface of APCs [112].
Other immune cells that participate to the cellular innate immunity are the granulocytes, consists of neutrophils, eosinophils and basophils and they are responsible to release inflammatory mediators and attract more immune cells at inflammation and/or infection sites [5].
CYT index is strictly associated with the composition of immune system. Specifically, CD8+ T cells release perforin and granzymes to kill targeted cells [114]. Therefore, since CYT is based on the expression of granzymes and perforin, the composition of immune system and the percentage CD8+ T cells can affect the index of CYT.
The immune system is a defence mechanism against infected and/or self-antigens [2] and its role is very important both for prognosis and treatment of cancer. Unfortunately, cancer cells evolve different mechanisms to escape effective immunosurveillance. They produce immunosuppressive cytokines and prostaglandins to inhibit the division of NK cells as well as the proliferation and function of T helper and CTLs. Moreover, antigen-processing of mutant proteins through MHC complexes by immune-resistant cancer cells, reducing antigenicity and reinforce the destruction of malignant cells from immune system [115].
The interplay between the immune system and the cancer pathogenesis enhances understanding of immune activation and response against cancer. Immune-oncology is a fiend that is rapidly constantly evolved and knowledge of immune composition especially in the TME will significantly contribute to create more effective therapies [5].
2.3. Clinical Implications and Prognostic Significance
2.3.1. Association between CYT and Patient Outcomes
CYT is closely related with patient prognosis and outcomes and the expression of both GZMA and PRF1 genes synergistically, or alone can be affect the overall survival of cancer patients. In Adrenocortical Carcinoma (ACC), skin melanoma and bladder cancer, high CYT levels associated with better patient outcome and survival. Roufas et. al. reported that high CYT skin melanomas, activate immune-related genes and increase the levels of CD8+ T cells, NK cells, B cells, M1 macrophages and DCs [22].
Increase of T-cells and M1 macrophages within CRC and HCC patients in combination with high CYT, correlating with better survival, longer OS and Disease-Specific Survival (DSS) [27,29].
In endometrial carcinoma (EC), and high-grade serous carcinoma (HGSC), high expression levels of both GZMA and PRF1 or alone, associated with higher overall survival (OS) rates. Conversely, low CYT levels related to worst clinical outcome [26,116]. In addition, high CYT lung adenocarcinoma had significantly longer PFS and OS [117]. Unlike ACC, skin melanoma, bladder cancer, EC, HGSC and lung adenocarcinoma, in glioblastoma (GBM) high CYT associated with poor prognosis and worse survival [43,118].
In a study of 7533 breast cancer patients, TNBC group had the highest CYT levels and better survival, compared to the estrogen receptor (ER)-positive and Human Epidermal Growth Factor Receptor 2 (HER2)-negative subgroups [28]. On the other hand, breast tumors with high mutant allele tumor heterogeneity (MATH) score are related with low CYT levels, less immune responses and worse survival [119].
Recently, in Breast Cancer (BC), was reported that overexpression of GZMA alone, increased the infiltration of DCs and CD8 + T cells and related with better OS, DSS and Progress Free Interval (PFI) [120], while in 2019, high expression levels of GZMA, were significantly correlated with better survival in HCC [121]. On top of that, high CYT levels within TNBC were significantly associated with high inflammatory scores, better DSS and OS [122]. On the contrary, late recurrence BC patients have lower CYT index compared with survivors and low CYT levels in late recurrence patients were associated with worse Disease free survival (DFS) [123].
CYT was also evaluated between tumor and tumor-free tongue patients with Squamous Cell Carcinoma of the Oral Tongue (SCCOT) and it is remarkable that, tumor-free tongue patients had higher CYT levels with better survival, contralateral to the tumor patients that CYT is not predictive for their survival [51].
2.3.2. Predictive Value of CYT for Immunotherapy Response
In recent years, cancer immunotherapy has enabled never-before seen success rates in durable tumor control and enhanced survival benefits in patients with advanced cancers. Importantly, ICI therapies using monoclonal antibodies (mabs), including anti-PD-1 to treat cancer patient has spawned overwhelming enthusiasm for immunotherapy in the disease [124,125].
CYT is identified as a new immunotherapy biomarker that reflects the antitumor immune responses and activity of CD8+ cytotoxic T lymphocytes and macrophages [23] and it can be used to predict the respond of the patient to ICI therapies. It is indicative that the expression levels of both GZMA and PRF1 can predict favourable survival of cancer patients following ICI immunotherapy [126].
High CYT levels within CRC, EC and Non-small cell lung cancer (NSCLC) were associated with high expression of immune checkpoint proteins, including PD-L1 and CTLA-4 and may be respond better to ICI therapies [26,27,94]. It is also reported that, CYT-high GC tumors respond better respond to anti-PD-1 therapy [25] and CRC patients with high CYT levels showed a more sensitivity to ICI therapies, compared to CYT-low CRC patients [24]
CYT-high skin melanomas patients who receive anti-CTLA-4 and/or anti-PD-1 therapy, have better clinical results due to higher immunophenoscore, compare to the patients with low CYT levels [52]. Metastatic melanoma patients with high GZMA expression levels, exhibited significantly higher expression levels of CTLA-4, PD-L1 and PD-L2 and have better respond to anti PD-L1 (nivolumab) treatment, resulting to better clinical benefit and long-term survival [127,128].
In addition of nivolumab, in melanoma patients that treated with anti-CTLA4 antibody (ipilimumab), intratumoral immune CYT increases the infiltration of CD8+ T cells and the expression of MHC class I [129].
In cutaneous melanoma, high GZMA expression levels lead to immune activation and infiltration of CD8+ T cells, making the patients more sensitive to anti-PD-L1 immunotherapy [130]. Furthermore, CYT-high Prostate Cancer (PCa) patients are more sensitive to ICIs, due to the fact that, high GZMA and PRF1 levels, increase the number of CD8+ T cells and the expression of immune checkpoints inhibitors, including PD-L1 compared with the low CYT patients [131].
Although, the different forms of immunotherapy have achieved success in treating a lot of cancer types, only a small number of patients respond to these therapies and some patients create resistance to them. Therefore, the evaluation of CYT as a prognostic factor in treatment selection and immunotherapy response will help cancer patients to receive the most appropriate therapy for their cancer, with the best results.
2.3.3. Integration of CYT Assessment into Clinical Practice
- Potential Biomarkers for CYT Evaluation
CYT is a biomarker of antitumor immunity, and it is calculated based on the expression levels of GZMA and PRF1, released from CTLs through granzyme/perforin pathway [23]. CYT is also significantly related with the expression of other different biomarkers.
The TME plays a crucial role in cancer progression and its composition may predict patient’s prognosis. CYT depends on the proportion of immune cells within the TME which can be used as a biomarker for CYT evaluation. Notably, it has been recorded that the presence of T cells within the TME relates with favourable prognosis [132], and CYT is positively correlated with the proportion of tumor-infiltrating CD8+ T cells and M1 macrophages.
Additionally, CYT-high levels are strongly related with high infiltration of the above mentioned immune cells [22,29,129,130,131,133]. Further of GZMA and PRF1, GZMB has been also to be present in high amounts of CD8+ T cells [134,135].
CYT is positively associated with the status of MSI, the mutational burden and the rate of tumor-mutated peptides, called neoepitopes. Rooney et al, shown that neoepitopes can handle CYT in a number of tumors [23]. CYT is also correlated with increased mutational burden and neoepitope load in GC and colon tumors [24,25].
Moreover, MSI-high colon and colorectal tumors are related with increased levels of both CYT and mutations [24] and dMMR/MSI-high patients with colorectal adenocarcinoma respond better to anti-PD-1 immunotherapy [136]. In contrast, in Pancreatic ductal adenocarcinoma (PDA) increase of CYT levels does not correlated with increase in the mutational burden or neoepitope load [20].
Checkpoint inhibitors, including CTLA-4 and PD-L1 are important indicators for both CYT-low and CYT-high cancer patients. Most studies supports that high levels of PD-L1 and CTLA-4 are exhibited in CYT-high cancer patients [26,27,94] and these patients are more sensitive to ICI mab therapies [24].
- Role of CYT in Patient Stratification and Treatment Selection
In past decades, standard treatment option for cancer therapy includes immunotherapy, chemotherapy, radiotherapy, surgical removal, and/or targeted therapy, depending on the tumor’s stage [137]. CYT plays a pivotal role in patient stratification and its levels has significant value to choose the suitable treatment for each patient with different types of advanced cancer.
As it is already mentioned, CYT-high and CYT-low patients have different respond to therapies and high CYT levels increase the activation and infiltration of CD8+ T cells [22,129,130,131,133].
CYT is significantly related with the amount of active CTLs that can be used in the treatment selection. In vitro studies of Paclitaxel (TAX), Doxorubicin (DOX) and Cisplatin (CIS) showed that all of the above mentioned drugs enhance the cell-mediated killing by CTLs [138,139,140]. It has been also reported that, low non-cytotoxic doses of 5-Fluorouracil (5-FU), Taxotere, Cisplatin and Irinotecan (CPT-11), and CIS induce antitumor responses of CD8+ T cells which they are connected with high CYT levels in colon cancer [141]. On the contrary, CYT-low Androgen Receptor (AR) and ER-positive BC were associated with low infiltration and CD8+ T cells, less overall anticancer immunity and survival [142].
In many patients with different types of cancer, conventional therapies do not provide satisfactory clinical results. Therefore, in recent years, the combinational therapy of cancer immunotherapy and chemotherapy is gaining credibility due to its high efficacy, and it is used to treat patients with advanced types of cancer. CYT can be used to predict the response of cancer patients to ICI therapies. Specifically, CYT-high cancer patients were exhibited significantly higher levels of immune checkpoint inhibitors, including PD-L1 and CTLA-4 and respond better to ICI therapies with mabs [25,26,27,52,94,127,128,130,131].
Furthermore, anti-CTLA-4 mab therapy alone and/or with anti-PD-1 therapy has been offered important therapeutic benefits to cancer patients [143], and it enhances the immune anti-tumor activity due to increase the number of tumor-infiltrating CD8+ and CD4+ T cells. Moreover, anti-CTLA-4 therapy combined with CIS chemotherapy was significantly more effective, increases the expression of perforins and Granzyme B and improved the survival of patients, compared to the anti-CTLA-4 and CIS alone therapies in murine mesothelioma [144].
Even though, there is an improvement in the prognosis for many patients depending on the stage of cancer, most cancer patients still have a poor prognosis, or they develop resistance to immunotherapy. CYT as a relatively new index, and its index can be used as a key tool in cancer -patient treatment plan.
2.4. Modulation of CYT
2.4.1. Current and Emerging Immunotherapy Strategies
- Immune Checkpoint Inhibitors
ICIs have made tremendous progress and they approved and widely used in cancer immunotherapy [145]. In addition, ICIs targeting CTLA-4 and/or PD-1/PD-L1 have shown unparalleled clinical efficacy in multiple types of cancer [146].
Ipilimumab (anti-CTLA-4 antibody) constitutes the first checkpoint inhibitor that approved for cancer treatment and it showed improvements in the OS of metastatic melanoma patients [147,148]. Moreover, the use of pidilizumab (anti-PD-1 antibody) in metastatic melanoma patients that 51% of them, they previously treated with ipilimumab showed 1-year better OS [149].
- Adoptive Cell Therapies
ACTs refer to a type of personalized cancer immunotherapy in which lymphocytes are expanded and grown ex-vivo and re-infused into the cancer patients [150,151] and they give a possibly dynamic general treatment for cancer [152].
ACTs with TILs or gene-modified T cells expressing TCRs or chimeric antigen receptors (CARs) are alternative type of therapies that trigger the immune system in order to recognize and kill cancer cells and they have shown promising results in many cancer types [153].
- Vaccines and Oncolytic Viruses
OVs are created to target and kill cancer cells directly, without harming the normal cell [154] and constitute a promising category of cancer therapy. They enhance anticancer immune responses either by the production of tumor associated antigens or inflammatory signals [155]. OVs also modulate the TME by increase the maturation of tumor-specific T cells within it dye to the high number innate immune cells including of DCs [156].
OVs can be used as immunotherapeutic anti-cancer vaccines to enhance tumor-specific T -responses and mediate the killing of unharmed cancer cells [157]. Moreover, it has been shown through both in-vitro, in-vivo and pre-clinical studies that the combination of OVs with anti-CTLA-4 and/or anti-PD-1 ICIs has high oncolytic efficacy [154,158].
2.4.2. Combination Approaches to Enhance CYT
- Dual Immune Checkpoint Blockade
Despite that ICB therapies have received heightened interest in strategies, the efficacy of them alone is restricted by low response. Dual ICB constitutes one of the most promising form of immunotherapy for cancer treatment that enhance the antitumor activity of Cytotoxic T cells and it is already used to treat solid tumors, including melanoma and RCC as well as advanced HCC with great efficacy and promising results [159].
Different combinatorial therapies with mabs are used to treat cancer both in preclinical models and clinical trials. Combination of anti-CTLA-4 and anti-PD-1 mabs enhance antitumor activity by increased the infiltration of CD8+ T cells and decreased the number of Tregs. As a results of this, the TME converts from immunosuppressive to inflammatory with better clinical results [83,160]. Moreover, in NSCLC patients, immunotherapy using nivolumab (anti-PD-1) and ipilimumab (anti-CTLA-4) mabs showed survival benefit [161].
On the contrary, in head and neck squamous cell carcinoma, combinatorial immunotherapy of PD-1/CTLA4 mabs causes decrease in the activation of CD8+ T cells and increase the activation of Tregs [162]. In addition, phase II clinical trials of concurrent treatment with the above-mentioned mabs showed higher response rate and PFS, compared to the monotherapies in melanoma patients [163,164].
Dual ICB is a powerful therapy, and it can be considered almost certain that in the near future the use of this type of immunotherapy will increase with greater success rates and in more types of cancer.
- Modulation of the TME
TME involves in different biological processes that implicated with the initiation and treatment and it constitutes an important tool for both prevention and treatment of cancer [165,166]. Modulation of the TME is crucial in different stages of cancer including of primary tumor growth, development of metastasis and immune evasion and it plays a leading role in strategies for cancer treatment [74].
The TME is modulated from cancer cells by the production due to their high consumption of glucose. This metabolic modulation of the TME increases to ICPs and infiltration of TILs [167]. Moreover, the treatment of cancer through immune modulation of the TME makes headway and studies focuses to understand more about the TME components and their interactions between themselves in order to use it as a prognostic tool. TME is already a prognostic tool for enhancing cancer immunotherapy and it used to predict its efficacy [168].
2.4.3. Challenges and Future Directions in CYT Modulation
As previously mentioned, CYT is a measure of immune activation based on the expression of GZMA and PRF1 and it varies across different cancer types [23]. Moreover, the expression of GZMA was either lowly expressed or absent, while PRF1 was not detected from 20 different tumor types [22]. CYT is also associated with the TMB events as well as with different mutational signatures and it deregulates the expression of immune checkpoint inhibitors [24].
It is remarkable that, CYT-high skin melanoma tumors express immune-related genes and vice versa, and it is suggesting that CYT-high skin melanoma patients are more sensitive to ICI therapies due to their higher immunophenoscore [52].
CYT constitutes a significant prognostic biomarker for cancer treatment, and it can be used to predict the respond of cancer patients to ICI immunotherapies, as well as measure the immune activation within the tumor. Therefore, its use is expected to solve the problem that occurs with many cancer patients who are not sensitive, or they show resistance to this type of immunotherapy.
As CYT described first from Rooney et al., in 2015 and it is relatively new index, more research is needed in future to investigate how this important prognostic/predictive tool can be used in cancer strategies to treat different tumor types.
3. Conclusions
3.1. Recapitulation of Key Findings
In this review, we summarize the role of intratumoral immune CYT in cancer progression and its implication for cancer patient prognosis and treatment outcomes. CYT, as measured by the geometric mean of GZMA and PRF1, reflects a tumor’s immune activation and plays a significant role in its control. Along with GZMA and PRF1, GZMB can also be used to calculate CYT levels. Different methods are used to evaluate CYT, including of isolation, characterization and analysis of TILs by flow cytometry, due to their ability to recognize and kill autologous cancer cells, and gene expression profiling. In addition, immune CYT can be calculated within the TME by mIF and IMC and the immune cells which composed the TME, constitute an indicator to assess the CYT.
CYT is influenced by different factors, such as the TMB, where cancer patients with high TMB (especially MSI-H/dMMR ones) show better response to ICI immunotherapies. Finally, CYT is strictly related by the antigen presentation and MHC I and II complexes, as well as by TME factors, including immunosuppressive cells, immune checkpoints, stomal cells and ECM components.
CYT is importantly related with the composition of immune system, mainly within the TME. It is important to refer that the interplay between the immune cells and cancer cells is crucial for primary growth of tumor and treatment. Most studies observed that an increase in the CYT index enhances the antitumor immune response by increasing the infiltration of CD8+ T cells and decreasing the number of immunosuppressive cells (Tregs and MDSCs). On the contrary, CYT-low tumors are characterised by higher number of immunosuppressive cells and inhibition of antitumor immune responses and immune checkpoint molecules, resulting in cancer progression, metastasis and immune evasion.
3.2. Potential Clinical Implications and Future Directions
Although, in recent years, a tremendous progress has been recorded in the field of cancer therapy, there is a big number of patients that still have poor prognosis and they develop resistance to therapies. Therefore, it is considered imperative to find out accurate prognostic and predictive biomarkers in order to create the most effective therapy for each cancer patient. As inferred from this review, CYT can constitute one of these biomarkers.
CYT is expected to take a central role in cancer treatment plans in the near future due to its high clinical value. Currently, it is an important prognostic biomarker for patient response in cancer therapies. In ACC, skin melanoma, bladder cancer, BC, HCC, EC, CRC, HGSC, TNBC, lung adenocarcinoma and SCCOT, high levels of CYT showed longer OS, PFS, PFI and DSS [15,22,26,27,28,29,51,116,117,122]. On the contrary, only CYT-high GBM patients were related with poor prognosis and worse survival [43,118].
Moreover, CYT is an accurate immunotherapy biomarker that it is strictly related with the expression of immune checkpoint inhibitors. Importantly, CYT-high tumors exhibit significantly higher levels of immune checkpoint inhibitors, too, whereas CYT-high patients are more sensitive to ICI immunotherapies, using mabs with higher respond rates. It is indicated that high expression levels of GZMA and PRF1 synergistically or alone in different types of cancer, including of CRC, EC, NSCLC, GC and cutaneous melanoma, increased the levels of CTLA-4 and/or PD-1/PD-L1, resulting to better clinical outcome and survival for CYT-high patients who received anti-CTLA-4 and/or anti-PD-1/PD-L1 immunotherapies (Figure 4).
Most studies showed that CYT-high patients are related with better clinical outcome and long-term survival in different types of cancer. In the near future, more research into CYT is needed, that will focus to recapitulate its importance in cancer immunotherapy, either with dual ICB or by targeting their metabolic pathways to promote an immune-stimulatory microenvironment.
3.3. Overall Significance of Understanding CYT in Cancer
Considering all of the above parts, current review highlights the significance of understanding the implication of intratumoral immune CYT in cancer, from primary growth to distant metastasis in order to use it for therapeutic purposes.
Intratumoral immune CYT arises as an indicator of immune activation based on the expression of granzymes and perforins and through research, it becomes a critical prognostic and predictive factor in cancer immunotherapy. Importantly, increased levels of CYT are related with high expression levels of immune checkpoint molecules like CTLA-4, PD-1 and PD-L1 and blocking of them by using mabs, enhances antitumor immunity. Finally, CYT-high patients are related with positive clinical outcomes and a favourable prognosis.
On top of that, TME composition directly affects the intratumoral immune CYT and the full known and understanding of immune cells and cancer cells within the TME can modulate in in order to improve CYT, create an immune-stimulatory microenvironment and enhance antitumor immune responses against cancer cells.
In summary, understanding and modulating CYT is crucial for improving cancer immunotherapy. Research into CYT and the factors that influence it eventually transform cancer treatment, improve patient outcomes, and increase their life span.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mattiuzzi C, Lippi G. Current Cancer Epidemiology. J Epidemiol Glob Health 2019;9:217–22. [CrossRef]
- Naidoo J, Page DB, Wolchok JD. Immune checkpoint blockade. Hematol Oncol Clin North Am 2014;28:585–600. [CrossRef]
- M. Candeias S, S. Gaipl U. The Immune System in Cancer Prevention, Development and Therapy. Anticancer Agents Med Chem 2016;16:101–7. [CrossRef]
- Houghton AN, Guevara-Patiño JA. Immune recognition of self in immunity against cancer. J Clin Invest 2004;114:468. [CrossRef]
- Pandya PH, Murray ME, Pollok KE, Renbarger JL. The Immune System in Cancer Pathogenesis: Potential Therapeutic Approaches. J Immunol Res 2016;2016. [CrossRef]
- Charles A Janeway J, Travers P, Walport M, Shlomchik MJ. Immunological memory 2001.
- Kim SK, Cho SW. The Evasion Mechanisms of Cancer Immunity and Drug Intervention in the Tumor Microenvironment. Front Pharmacol 2022;13. [CrossRef]
- Cavallo F, De Giovanni C, Nanni P, Forni G, Lollini PL. 2011: the immune hallmarks of cancer. Cancer Immunol Immunother 2011;60:319. [CrossRef]
- Labani-Motlagh A, Ashja-Mahdavi M, Loskog A. The Tumor Microenvironment: A Milieu Hindering and Obstructing Antitumor Immune Responses. Front Immunol 2020;11. [CrossRef]
- Salvatore V, Teti G, Focaroli S, Mazzotti MC, Mazzotti A, Falconi M. The tumor microenvironment promotes cancer progression and cell migration. Oncotarget 2017;8:9608. [CrossRef]
- Anderson NM, Simon MC. Tumor Microenvironment. Curr Biol 2020;30:R921. [CrossRef]
- Wei R, Liu S, Zhang S, Min L, Zhu S. Cellular and Extracellular Components in Tumor Microenvironment and Their Application in Early Diagnosis of Cancers. Anal Cell Pathol (Amst) 2020;2020. [CrossRef]
- O’Donnell JS, Teng MWL, Smyth MJ. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat Rev Clin Oncol 2019;16:151–67. [CrossRef]
- Schadendorf D, Hodi FS, Robert C, Weber JS, Margolin K, Hamid O, et al. Pooled Analysis of Long-Term Survival Data From Phase II and Phase III Trials of Ipilimumab in Unresectable or Metastatic Melanoma. J Clin Oncol 2015;33:1889–94. [CrossRef]
- Lee JB, Kim HR, Ha SJ. Immune Checkpoint Inhibitors in 10 Years: Contribution of Basic Research and Clinical Application in Cancer Immunotherapy. Immune Netw 2022;22. [CrossRef]
- Franzin R, Netti GS, Spadaccino F, Porta C, Gesualdo L, Stallone G, et al. The Use of Immune Checkpoint Inhibitors in Oncology and the Occurrence of AKI: Where Do We Stand? Front Immunol 2020;11:574271. [CrossRef]
- Shiravand Y, Khodadadi F, Kashani SMA, Hosseini-Fard SR, Hosseini S, Sadeghirad H, et al. Immune Checkpoint Inhibitors in Cancer Therapy. Curr Oncol 2022;29:3044. [CrossRef]
- Grossman WJ, Verbsky JW, Barchet W, Colonna M, Atkinson JP, Ley TJ. Human T regulatory cells can use the perforin pathway to cause autologous target cell death. Immunity 2004;21:589–601. [CrossRef]
- Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol 2002 210 2002;2:735–47. [CrossRef]
- Balli D, Rech AJ, Stanger BZ, Vonderheide RH. Immune Cytolytic Activity Stratifies Molecular Subsets of Human Pancreatic Cancer. Clin Cancer Res 2017;23:3129–38. [CrossRef]
- Woodsworth DJ, Dreolini L, Abraham L, Holt RA. Targeted Cell-to-Cell Delivery of Protein Payloads via the Granzyme-Perforin Pathway. Mol Ther Methods Clin Dev 2017;7:132. [CrossRef]
- Roufas C, Chasiotis D, Makris A, Efstathiades C, Dimopoulos C, Zaravinos A. The expression and prognostic impact of immune cytolytic activity-related markers in human malignancies: A comprehensive meta-analysis. Front Oncol 2018;8:332192. [CrossRef]
- Rooney MS, Shukla SA, Wu CJ, Getz G, Hacohen N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell 2015;160:48–61. [CrossRef]
- Zaravinos A, Roufas C, Nagara M, De Lucas Moreno B, Oblovatskaya M, Efstathiades C, et al. Cytolytic activity correlates with the mutational burden and deregulated expression of immune checkpoints in colorectal cancer. J Exp Clin Cancer Res 2019;38. [CrossRef]
- Hu Q, Nonaka K, Wakiyama H, Miyashita Y, Fujimoto Y, Jogo T, et al. Cytolytic activity score as a biomarker for antitumor immunity and clinical outcome in patients with gastric cancer. Cancer Med 2021;10:3129. [CrossRef]
- Chen Q, Wang C, Lei X, Huang T, Zhou R, Lu Y. Immune Cytolytic Activity for Comprehensive Insights of the Immune Landscape in Endometrial Carcinoma. J Oncol 2022;2022. [CrossRef]
- Narayanan S, Kawaguchi T, Yan L, Peng X, Qi Q, Takabe K. Cytolytic Activity Score to Assess Anticancer Immunity in Colorectal Cancer. Ann Surg Oncol 2018;25:2323–31. [CrossRef]
- Oshi M, Kawaguchi T, Yan L, Peng X, Qi Q, Tian W, et al. Immune cytolytic activity is associated with reduced intra-tumoral genetic heterogeneity and with better clinical outcomes in triple negative breast cancer. Am J Cancer Res 2021;11:3628.
- Takahashi H, Kawaguchi T, Yan L, Peng X, Qi Q, Morris LGT, et al. Immune Cytolytic Activity for Comprehensive Understanding of Immune Landscape in Hepatocellular Carcinoma. Cancers 2020, Vol 12, Page 1221 2020;12:1221. [CrossRef]
- Wakiyama H, Masuda T, Motomura Y, Qingjiang HU, Tobo T, Eguchi H, et al. Cytolytic Activity (CYT) Score Is a Prognostic Biomarker Reflecting Host Immune Status in Hepatocellular Carcinoma (HCC). Anticancer Res 2018;38:6631–8. [CrossRef]
- Zhang H, Liu Y, Hu D, Liu S. Identification of Novel Molecular Therapeutic Targets and Their Potential Prognostic Biomarkers Based on Cytolytic Activity in Skin Cutaneous Melanoma. Front Oncol 2022;12:844666. [CrossRef]
- Melioli G, Meta M, Semino C, Casartelli G, Pasquetti W, Biassoni R, et al. Isolation and In vitro expansion of lymphocytes infiltrating non-small cell lung carcinoma: Functional and molecular characterisation for their use in adoptive immunotherapy. Eur J Cancer 1994;30:97–102. [CrossRef]
- Crossey F, Marx S, Hölters S, Schmitt K, Bohle RM, Schmidt T, et al. Robust method for isolation of tumor infiltrating lymphocytes with a high vital cell yield from small samples of renal cell carcinomas by a new collagenase-free mechanical procedure. Urol Oncol Semin Orig Investig 2018;36:402.e1-402.e10. [CrossRef]
- Finke JH, Rayman P, Alexander J, Edinger M, Tubbs RR, Connelly R, et al. Characterization of the Cytolytic Activity of CD4+ and CD8+ Tumor-infiltrating Lymphocytes in Human Renal Cell Carcinoma1. CANCER Res 1990;50:2363–70.
- Chew V, Tow C, Teo M, Wong HL, Chan J, Gehring A, et al. Inflammatory tumour microenvironment is associated with superior survival in hepatocellular carcinoma patients. J Hepatol 2010;52:370–9. [CrossRef]
- Kazemi MH, Sadri M, Najafi A, Rahimi A, Baghernejadan Z, Khorramdelazad H, et al. Tumor-infiltrating lymphocytes for treatment of solid tumors: It takes two to tango? Front Immunol 2022;13:1018962. [CrossRef]
- Fassler DJ, Torre-Healy LA, Gupta R, Hamilton AM, Kobayashi S, Van Alsten SC, et al. Spatial Characterization of Tumor-Infiltrating Lymphocytes and Breast Cancer Progression. Cancers (Basel) 2022;14. [CrossRef]
- Hall ML, Liu H, Malafa M, Centeno B, Hodul PJ, Pimiento J, et al. Expansion of tumor-infiltrating lymphocytes (TIL) from human pancreatic tumors. J Immunother Cancer 2016;4:1–12. [CrossRef]
- Kobayashi T, Kumagai S, Doi R, Afonina E, Koyama S, Nishikawa H. Isolation of tumor-infiltrating lymphocytes from preserved human tumor tissue specimens for downstream characterization. STAR Protoc 2022;3:101557. [CrossRef]
- Giraldo NA, Becht E, Vano Y, Petitprez F, Lacroix L, Validire P, et al. Tumor-infiltrating and peripheral blood T-cell immunophenotypes predict early relapse in localized clear cell renal cell carcinoma. Clin Cancer Res 2017;23:4416–28. [CrossRef]
- Peters PJ, Borst J, Oorschot V, Fukuda M, Krahenbuhl O, Tschopp J, et al. Cytotoxic T lymphocyte granules are secretory lysosomes, containing both perforin and granzymes. J Exp Med 1991;173:1099–109. [CrossRef]
- Johnson BJ, Costelloe EO, Fitzpatrick DR, Haanen JBAG, Schumacher TNM, Brown LE, et al. Single-cell perforin and granzyme expression reveals the anatomical localization of effector CD8+ T cells in influenza virus-infected mice. Proc Natl Acad Sci U S A 2003;100:2657–62. [CrossRef]
- Haddad AF, Chen JS, Oh T, Pereira MP, Joshi RS, Aghi MK. Higher cytolytic score correlates with an immunosuppressive tumor microenvironment and reduced survival in glioblastoma. Sci Reports 2020 101 2020;10:1–9. [CrossRef]
- Kim EN, Chen PZ, Bressan D, Tripathi M, Miremadi A, Pietro M di, et al. Dual-modality imaging of immunofluorescence and imaging mass cytometry for whole slide imaging with accurate single-cell segmentation. BioRxiv 2023. [CrossRef]
- Viratham Pulsawatdi A, Craig SG, Bingham V, McCombe K, Humphries MP, Senevirathne S, et al. A robust multiplex immunofluorescence and digital pathology workflow for the characterisation of the tumour immune microenvironment. Mol Oncol 2020;14:2384–402. [CrossRef]
- Lee CW, Ren YJ, Marella M, Wang M, Hartke J, Couto SS. Multiplex immunofluorescence staining and image analysis assay for diffuse large B cell lymphoma. J Immunol Methods 2020;478:112714. [CrossRef]
- Giesen C, Wang HAO, Schapiro D, Zivanovic N, Jacobs A, Hattendorf B, et al. Highly multiplexed imaging of tumor tissues with subcellular resolution by mass cytometry. Nat Methods 2014 114 2014;11:417–22. [CrossRef]
- Schlecht A, Boneva S, Salie H, Killmer S, Wolf J, Hajdu RI, et al. Imaging mass cytometry for high-dimensional tissue profiling in the eye. BMC Ophthalmol 2021;21:1–9. [CrossRef]
- Martinez-Morilla S, Villarroel-Espindola F, Wong PF, Toki MI, Aung TN, Pelekanou V, et al. Biomarker discovery in patients with immunotherapy-treated melanoma with imaging mass cytometry. Clin Cancer Res 2021;27:1987–96. [CrossRef]
- Mori H, Bolen J, Schuetter L, Massion P, Hoyt CC, VandenBerg S, et al. Characterizing the Tumor Immune Microenvironment with Tyramide-Based Multiplex Immunofluorescence. J Mammary Gland Biol Neoplasia 2020;25:417–32. [CrossRef]
- Gu X, Boldrup L, Coates PJ, Fahraeus R, Wang L, Wilms T, et al. High immune cytolytic activity in tumor-free tongue tissue confers better prognosis in patients with squamous cell carcinoma of the oral tongue. J Pathol Clin Res 2019;5:240–7. [CrossRef]
- Roufas C, Georgakopoulos-Soares I, Zaravinos A. Distinct genomic features across cytolytic subgroups in skin melanoma. Cancer Immunol Immunother 2021;70:3137. [CrossRef]
- Lawlor RT, Mattiolo P, Mafficini A, Hong SM, Piredda ML, Taormina S V., et al. Tumor mutational burden as a potential biomarker for immunotherapy in pancreatic cancer: Systematic review and still-open questions. Cancers (Basel) 2021;13:3119. [CrossRef]
- Meléndez B, Van Campenhout C, Rorive S, Remmelink M, Salmon I, D’Haene N. Methods of measurement for tumor mutational burden in tumor tissue. Transl Lung Cancer Res 2018;7:661. [CrossRef]
- Steuer CE, Ramalingam SS. Tumor Mutation Burden: Leading Immunotherapy to the Era of Precision Medicine? J Clin Oncol 2018;36:631–2. [CrossRef]
- Sha D, Jin Z, Budczies J, Kluck K, Stenzinger A, Sinicrope FA. Tumor Mutational Burden (TMB) as a Predictive Biomarker in Solid Tumors. Cancer Discov 2020;10:1808. [CrossRef]
- Yarchoan M, Hopkins A, Jaffee EM. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. N Engl J Med 2017;377:2500. [CrossRef]
- Rizvi NA, Hellmann MD, Snyder A, Kvistborg P, Makarov V, Havel JJ, et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 2015;348:124–8. [CrossRef]
- Carbone DP, Reck M, Paz-Ares L, Creelan B, Horn L, Steins M, et al. First-Line Nivolumab in Stage IV or Recurrent Non-Small-Cell Lung Cancer. N Engl J Med 2017;376:2415–26. [CrossRef]
- Hellmann MD, Ciuleanu T-E, Pluzanski A, Lee JS, Otterson GA, Audigier-Valette C, et al. Nivolumab plus Ipilimumab in Lung Cancer with a High Tumor Mutational Burden. N Engl J Med 2018;378:2093–104. [CrossRef]
- Schrock AB, Ouyang C, Sandhu J, Sokol E, Jin D, Ross JS, et al. Tumor mutational burden is predictive of response to immune checkpoint inhibitors in MSI-high metastatic colorectal cancer. Ann Oncol Off J Eur Soc Med Oncol 2019;30:1096–103. [CrossRef]
- McWilliam HEG, Villadangos JA. MR1 antigen presentation to MAIT cells and other MR1-restricted T cells. Nat Rev Immunol 2023:1–15. [CrossRef]
- Bruce Sundstrom J, A. Ansari A. Comparative study of the role of professional versus semiprofessional or nonprofessional antigen presenting cells in the rejection of vascularized organ allografts. Transpl Immunol 1995;3:273–89. [CrossRef]
- Wieczorek M, Abualrous ET, Sticht J, Álvaro-Benito M, Stolzenberg S, Noé F, et al. Major histocompatibility complex (MHC) class I and MHC class II proteins: Conformational plasticity in antigen presentation. Front Immunol 2017;8:248429. [CrossRef]
- Falk K, Rötzschke O, Stevanovié S, Jung G, Rammensee HG. Allele-specific motifs revealed by sequencing of self-peptides eluted from MHC molecules. Nature 1991;351:290–6. [CrossRef]
- Hewitt EW. The MHC class I antigen presentation pathway: strategies for viral immune evasion. Immunology 2003;110:163. [CrossRef]
- Tortorella D, Gewurz BE, Furman MH, Schust DJ, Ploegh HL. Viral subversion of the immune system. Annu Rev Immunol 2000;18:861–926. [CrossRef]
- Holling TM, Schooten E, Van Den Elsen PJ. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum Immunol 2004;65:282–90. [CrossRef]
- Dhatchinamoorthy K, Colbert JD, Rock KL. Cancer Immune Evasion Through Loss of MHC Class I Antigen Presentation. Front Immunol 2021;12:636568. [CrossRef]
- Schrörs B, Lübcke S, Lennerz V, Fatho M, Bicker A, Wölfel C, et al. HLA class I loss in metachronous metastases prevents continuous T cell recognition of mutated neoantigens in a human melanoma model. Oncotarget 2017;8:28312–27. [CrossRef]
- Erdogdu IH. MHC Class 1 and PDL-1 Status of Primary Tumor and Lymph Node Metastatic Tumor Tissue in Gastric Cancers. Gastroenterol Res Pract 2019;2019. [CrossRef]
- Thibodeau J, Bourgeois-Daigneault MC, Lapointe R. Targeting the MHC Class II antigen presentation pathway in cancer immunotherapy. Oncoimmunology 2012;1:908. [CrossRef]
- Schietinger A, Philip M, Liu RB, Schreiber K, Schreiber H. Bystander killing of cancer requires the cooperation of CD4+ and CD8+ T cells during the effector phase. J Exp Med 2010;207:2469. [CrossRef]
- Hinshaw DC, Shevde LA. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res 2019;79:4557. [CrossRef]
- Wang Q, Shao X, Zhang Y, Zhu M, Wang FXC, Mu J, et al. Role of tumor microenvironment in cancer progression and therapeutic strategy. Cancer Med 2023;12:11149–65. [CrossRef]
- Tie Y, Tang F, Wei Y quan, Wei X wei. Immunosuppressive cells in cancer: mechanisms and potential therapeutic targets. J Hematol Oncol 2022;15:61. [CrossRef]
- Liu Y, Cao X. Immunosuppressive cells in tumor immune escape and metastasis. J Mol Med (Berl) 2016;94:509–22. [CrossRef]
- Sceneay J, Griessinger CM, Hoffmann SHL, Wen SW, Wong CSF, Krumeich S, et al. Tracking the fate of adoptively transferred myeloid-derived suppressor cells in the primary breast tumor microenvironment. PLoS One 2018;13. [CrossRef]
- Sasidharan Nair V, Elkord E. Immune checkpoint inhibitors in cancer therapy: a focus on T-regulatory cells. Immunol Cell Biol 2018;96:21–33. [CrossRef]
- Kondělková K, Vokurková D, Krejsek J, Borská L, Fiala Z, Ctirad A. Regulatory T cells (TREG) and their roles in immune system with respect to immunopathological disorders. Acta Medica (Hradec Kral 2010;53:73–7. [CrossRef]
- Qin H, Lerman B, Sakamaki I, Wei G, Cha SC, Rao SS, et al. Generation of a new therapeutic peptide that depletes myeloid-derived suppressor cells in tumor-bearing mice. Nat Med 2014;20:676–81. [CrossRef]
- Iclozan C, Antonia S, Chiappori A, Chen DT, Gabrilovich D. Therapeutic regulation of myeloid-derived suppressor cells and immune response to cancer vaccine in patients with extensive stage small cell lung cancer. Cancer Immunol Immunother 2013;62:909–18. [CrossRef]
- Curran MA, Montalvo W, Yagita H, Allison JP. PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc Natl Acad Sci U S A 2010;107:4275–80. [CrossRef]
- Herber DL, Nagaraj S, Djeu JY, Gabrilovich DI. Mechanism and therapeutic reversal of immune suppression in cancer. Cancer Res 2007;67:5067–9. [CrossRef]
- Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E, et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol 2015;35 Suppl:S185–98. [CrossRef]
- Whiteside TL. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008;27:5904–12. [CrossRef]
- Sadeghi Rad H, Monkman J, Warkiani ME, Ladwa R, O’Byrne K, Rezaei N, et al. Understanding the tumor microenvironment for effective immunotherapy. Med Res Rev 2021;41:1474–98. [CrossRef]
- Gaikwad S, Agrawal MY, Kaushik I, Ramachandran S, Srivastava SK. Immune checkpoint proteins: Signaling mechanisms and molecular interactions in cancer immunotherapy. Semin Cancer Biol 2022;86:137–50. [CrossRef]
- Mehdizadeh S, Bayatipoor H, Pashangzadeh S, Jafarpour R, Shojaei Z, Motallebnezhad M. Immune checkpoints and cancer development: Therapeutic implications and future directions. Pathol Res Pract 2021;223. [CrossRef]
- Ledford H. Melanoma drug wins US approval. Nature 2011;471:561. [CrossRef]
- Wang Y, Tong Z, Zhang W, Zhang W, Buzdin A, Mu X, et al. FDA-Approved and Emerging Next Generation Predictive Biomarkers for Immune Checkpoint Inhibitors in Cancer Patients. Front Oncol 2021;11. [CrossRef]
- Marin-Acevedo JA, Kimbrough EMO, Lou Y. Next generation of immune checkpoint inhibitors and beyond. J Hematol Oncol 2021 141 2021;14:1–29. [CrossRef]
- Otoshi T, Nagano T, Tachihara M, Nishimura Y. Possible Biomarkers for Cancer Immunotherapy. Cancers (Basel) 2019;11. [CrossRef]
- Yu X, Wang X. Tumor immunity landscape in non-small cell lung cancer. PeerJ 2018;2018:e4546. [CrossRef]
- Manetti M. Molecular Morphology and Function of Stromal Cells. Int J Mol Sci 2021;22:13422. [CrossRef]
- Tlsty TD. Stromal cells can contribute oncogenic signals. Semin Cancer Biol 2001;11:97–104. [CrossRef]
- Zhan H xiang, Zhou B, Cheng Y gang, Xu J wei, Wang L, Zhang G yong, et al. Crosstalk between stromal cells and cancer cells in pancreatic cancer: New insights into stromal biology. Cancer Lett 2017;392:83–93. [CrossRef]
- Lazennec G, Jorgensen C. Concise Review: Adult Multipotent Stromal Cells and Cancer: Risk or Benefit? Stem Cells 2008;26:1387–94. [CrossRef]
- Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer 2006;6:392–401. [CrossRef]
- Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms to therapy. Immunity 2014;41:49–61. [CrossRef]
- Mao Y, Keller ET, Garfield DH, Shen K, Wang J. Stromal cells in tumor microenvironment and breast cancer. Cancer Metastasis Rev 2012 321 2012;32:303–15. [CrossRef]
- Mun JY, Leem SH, Lee JH, Kim HS. Dual Relationship Between Stromal Cells and Immune Cells in the Tumor Microenvironment. Front Immunol 2022;13:864739. [CrossRef]
- Bosman FT, Stamenkovic I. Functional structure and composition of the extracellular matrix. J Pathol 2003;200:423–8. [CrossRef]
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK. Extracellular matrix structure. Adv Drug Deliv Rev 2016;97:4–27. [CrossRef]
- Karamanos NK, Theocharis AD, Piperigkou Z, Manou D, Passi A, Skandalis SS, et al. A guide to the composition and functions of the extracellular matrix. FEBS J 2021;288:6850–912. [CrossRef]
- Trent RJ. DNA Genetic Testing. Mol Med 2012:81–115. [CrossRef]
- Aerts J, Wetzels Y, Cohen N, Aerssens J. Data mining of public SNP databases for the selection of intragenic SNPs. Hum Mutat 2002;20:162–73. [CrossRef]
- Chanock S. Candidate genes and single nucleotide polymorphisms (SNPs) in the study of human disease. Dis Markers 2001;17:89–98. [CrossRef]
- Broman KW, Murray JC, Sheffield VC, White RL, Weber JL. Comprehensive human genetic maps: individual and sex-specific variation in recombination. Am J Hum Genet 1998;63:861–9. [CrossRef]
- Loktionov A. Common gene polymorphisms, cancer progression and prognosis. Cancer Lett 2004;208:1–33. [CrossRef]
- Sun JC, Lanier LL. Natural killer cells remember: an evolutionary bridge between innate and adaptive immunity? Eur J Immunol 2009;39:2059–64. [CrossRef]
- Charles A Janeway J, Travers P, Walport M, Shlomchik MJ. The components of the immune system 2001.
- Warrington R, Watson W, Kim HL, Antonetti FR. An introduction to immunology and immunopathology. Allergy Asthma Clin Immunol 2011;7 Suppl 1. [CrossRef]
- Koretzky GA. Multiple roles of CD4 and CD8 in T cell activation. J Immunol 2010;185:2643–4. [CrossRef]
- Loose D, Van De Wiele C. The Immune System and Cancer. Https://HomeLiebertpubCom/Cbr 2009;24:369–76. [CrossRef]
- Yang L, Wang S, Zhang Q, Pan Y, Lv Y, Chen X, et al. Clinical significance of the immune microenvironment in ovarian cancer patients. Mol Omi 2018;14:341–51. [CrossRef]
- Park C, Na KJ, Choi H, Ock CY, Ha S, Kim M, et al. Tumor immune profiles noninvasively estimated by FDG PET with deep learning correlate with immunotherapy response in lung adenocarcinoma. Theranostics 2020;10:10838. [CrossRef]
- Diaz MJ, Fadil A, Kleinberg G, Root KT, Ladehoff L, Batchu S, et al. Omics analysis of uveal melanoma: Leukocyte gene signatures reveal novel survival distinctions and indicate a prognostic role for cytolytic activity scoring. Neurosci Chronicles 2022;3:6.
- McDonald K-A, Kawaguchi T, Qi Q, Peng X, Asaoka M, Young J, et al. Tumor Heterogeneity Correlates with Less Immune Response and Worse Survival in Breast Cancer Patients. Ann Surg Oncol 2019 267 2019;26:2191–9. [CrossRef]
- Huo Q, Ning L, Xie N. Identification of GZMA as a Potential Therapeutic Target Involved in Immune Infiltration in Breast Cancer by Integrated Bioinformatical Analysis. Breast Cancer (London) 2023;15:213. [CrossRef]
- Mauriello A, Zeuli R, Cavalluzzo B, Petrizzo A, Tornesello ML, Buonaguro FM, et al. High Somatic Mutation and Neoantigen Burden Do Not Correlate with Decreased Progression-Free Survival in HCC Patients not Undergoing Immunotherapy. Cancers 2019, Vol 11, Page 1824 2019;11:1824. [CrossRef]
- Oshi M, Newman S, Tokumaru Y, Yan L, Matsuyama R, Endo I, et al. Inflammation Is Associated with Worse Outcome in the Whole Cohort but with Better Outcome in Triple-Negative Subtype of Breast Cancer Patients. J Immunol Res 2020;2020. [CrossRef]
- Takeshita T, Yan L, Asaoka M, Rashid O, Takabe K. Late recurrence of breast cancer is associated with pro-cancerous immune microenvironment in the primary tumor. Sci Reports 2019 91 2019;9:1–15. [CrossRef]
- Brahmer JR, Tykodi SS, Chow LQM, Hwu W-J, Topalian SL, Hwu P, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med 2012;366:2455–65. [CrossRef]
- Turnis ME, Andrews LP, Vignali DAA. Inhibitory receptors as targets for cancer immunotherapy. Eur J Immunol 2015;45:1892–905. [CrossRef]
- Vanmeerbeek I, Borras DM, Sprooten J, Bechter O, Tejpar S, Garg AD. Early memory differentiation and cell death resistance in T cells predicts melanoma response to sequential anti-CTLA4 and anti-PD1 immunotherapy. Genes Immun 2021;22:108–19. [CrossRef]
- Van Allen EM, Miao D, Schilling B, Shukla SA, Blank C, Zimmer L, et al. Genomic correlates of response to CTLA-4 blockade in metastatic melanoma. Science (80- ) 2015;350:207–11. [CrossRef]
- Inoue H, Park JH, Kiyotani K, Zewde M, Miyashita A, Jinnin M, et al. Intratumoral expression levels of PD-L1, GZMA, and HLA-A along with oligoclonal T cell expansion associate with response to nivolumab in metastatic melanoma. Oncoimmunology 2016;5. [CrossRef]
- Patel SJ, Sanjana NE, Kishton RJ, Eidizadeh A, Vodnala SK, Cam M, et al. Identification of essential genes for cancer immunotherapy. Nature 2017;548:537–42. [CrossRef]
- Wu X, Wang X, Zhao Y, Li K, Yu B, Zhang J. Granzyme family acts as a predict biomarker in cutaneous melanoma and indicates more benefit from anti-PD-1 immunotherapy. Int J Med Sci 2021;18:1657. [CrossRef]
- Gao Z, Tao Y, Lai Y, Wang Q, Li Z, Peng S, et al. Immune Cytolytic Activity as an Indicator of Immune Checkpoint Inhibitors Treatment for Prostate Cancer. Front Bioeng Biotechnol 2020;8:552034. [CrossRef]
- Jiang Y, Li Y, Zhu B. T-cell exhaustion in the tumor microenvironment. Cell Death Dis 2015 66 2015;6:e1792–e1792. [CrossRef]
- Takeshita T, Asaoka M, Katsuta E, Photiadis SJ, Narayanan S, Yan L, et al. High expression of polo-like kinase 1 is associated with TP53 inactivation, DNA repair deficiency, and worse prognosis in ER positive Her2 negative breast cancer. Am J Transl Res 2019;11:6507.
- Mellor-Heineke S, Villanueva J, Jordan MB, Marsh R, Zhang K, Bleesing JJ, et al. Elevated Granzyme B in Cytotoxic Lymphocytes is a Signature of Immune Activation in Hemophagocytic Lymphohistiocytosis. Front Immunol 2013;4. [CrossRef]
- Verschoor CP, Picard E, Andrew MK, Haynes L, Loeb M, Pawelec G, et al. NK- and T-cell granzyme B and K expression correlates with age, CMV infection and influenza vaccine-induced antibody titres in older adults. Front Aging 2022;3:1098200. [CrossRef]
- Wang QX, Qu CH, Gao YH, Ding PR, Yun JP, Xie D, et al. The degree of microsatellite instability predicts response to PD-1 blockade immunotherapy in mismatch repair-deficient/microsatellite instability-high colorectal cancers. Exp Hematol Oncol 2021;10:1–4. [CrossRef]
- Saini A, Kumar M, Bhatt S, Saini V, Malik A. CANCER CAUSES AND TREATMENTS. Int J Pharm Sci Res 2020;11:3121. [CrossRef]
- Rao S, Horwitz SB, Ringel I. Direct photoaffinity labeling of tubulin with taxol. J Natl Cancer Inst 1992;84:785–8. [CrossRef]
- Sartiano G, Darrell Bullington W, Lynch W. Mechanism of action of the anthracycline anti-tumor antibiotics, doxorubicin, daunomycin and rubidazone: preferential inhibition of DNA polymerase alpha. J Antibiot (Tokyo) 1979;32:1038–45. [CrossRef]
- Reedijk J, Lohman PHM. Cisplatin: synthesis, antitumour activity and mechanism of action. Pharm Weekbl Sci 1985;7:173–80. [CrossRef]
- Bergmann-Leitner ES, Abrams SI. Treatment of human colon carcinoma cell lines with anti-neoplastic agents enhances their lytic sensitivity to antigen-specific CD8+ cytotoxic T lymphocytes. Cancer Immunol Immunother 2001;50:445–55. [CrossRef]
- Okano M, Oshi M, Butash AL, Asaoka M, Katsuta E, Peng X, et al. Estrogen Receptor Positive Breast Cancer with High Expression of Androgen Receptor has Less Cytolytic Activity and Worse Response to Neoadjuvant Chemotherapy but Better Survival. Int J Mol Sci 2019, Vol 20, Page 2655 2019;20:2655. [CrossRef]
- Seidel JA, Otsuka A, Kabashima K. Anti-PD-1 and anti-CTLA-4 therapies in cancer: Mechanisms of action, efficacy, and limitations. Front Oncol 2018;8:330851. [CrossRef]
- Wu L, Yun Z, Tagawa T, Rey-McIntyre K, De Perrot M. CTLA-4 blockade expands infiltrating T cells and inhibits cancer cell repopulation during the intervals of chemotherapy in murine mesothelioma. Mol Cancer Ther 2012;11:1809–19. [CrossRef]
- Li B, Chan HL, Chen P. Immune Checkpoint Inhibitors: Basics and Challenges. Curr Med Chem 2017;26:3009–25. [CrossRef]
- Jenkins RW, Barbie DA, Flaherty KT. Mechanisms of resistance to immune checkpoint inhibitors. Br J Cancer 2018 1181 2018;118:9–16. [CrossRef]
- Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med 2010;363:711–23. [CrossRef]
- Wolchok JD, Hodi FS, Weber JS, Allison JP, Urba WJ, Robert C, et al. Development of ipilimumab: a novel immunotherapeutic approach for the treatment of advanced melanoma. Ann N Y Acad Sci 2013;1291:1–13. [CrossRef]
- Atkins MB, Kudchadkar RR, Sznol M, McDermott DF, Lotem M, Schachter J, et al. Phase 2, multicenter, safety and efficacy study of pidilizumab in patients with metastatic melanoma. Https://DoiOrg/101200/Jco20143215_suppl9001 2014;32:9001–9001. [CrossRef]
- Jiang X, Xu J, Liu M, Xing H, Wang Z, Huang L, et al. Adoptive CD8+ T cell therapy against cancer:Challenges and opportunities. Cancer Lett 2019;462:23–32. [CrossRef]
- Cohen JE, Merims S, Frank S, Engelstein R, Peretz T, Lotem M. Adoptive cell therapy: past, present and future. Http://DxDoiOrg/102217/Imt-2016-0112 2017;9:183–96. [CrossRef]
- Hawkins RE, Gilham DE, Debets R, Eshhar Z, Taylor N, Abken H, et al. Development of Adoptive Cell Therapy for Cancer: A Clinical Perspective. Https://HomeLiebertpubCom/Hum 2010;21:665–72. [CrossRef]
- Rohaan MW, Wilgenhof S, Haanen JBAG. Adoptive cellular therapies: the current landscape. Virchows Arch 2019;474:449–61. [CrossRef]
- Ylösmäki E, Cerullo V. Design and application of oncolytic viruses for cancer immunotherapy. Curr Opin Biotechnol 2020;65:25–36. [CrossRef]
- de Graaf JF, de Vor L, Fouchier RAM, van den Hoogen BG. Armed oncolytic viruses: A kick-start for anti-tumor immunity. Cytokine Growth Factor Rev 2018;41:28–39. [CrossRef]
- Bartlett DL, Liu Z, Sathaiah M, Ravindranathan R, Guo Z, He Y, et al. Oncolytic viruses as therapeutic cancer vaccines. Mol Cancer 2013 121 2013;12:1–16. [CrossRef]
- Elsedawy NB, Russell SJ. Oncolytic vaccines. Expert Rev Vaccines 2013;12:1155–72. [CrossRef]
- Engeland CE, Grossardt C, Veinalde R, Bossow S, Lutz D, Kaufmann JK, et al. CTLA-4 and PD-L1 checkpoint blockade enhances oncolytic measles virus therapy. Mol Ther 2014;22:1949–59. [CrossRef]
- Carloni R, Sabbioni S, Rizzo A, Ricci AD, Palloni A, Petrarota C, et al. Immune-Based Combination Therapies for Advanced Hepatocellular Carcinoma. J Hepatocell Carcinoma 2023;10:1445. [CrossRef]
- Selby M, Engelhardt J, Lu L-S, Quigley M, Wang C, Chen B, et al. Antitumor activity of concurrent blockade of immune checkpoint molecules CTLA-4 and PD-1 in preclinical models. Https://DoiOrg/101200/Jco20133115_suppl3061 2013;31:3061–3061. [CrossRef]
- Mariniello A, Novello S, Scagliotti G V., Ramalingam SS. Double immune checkpoint blockade in advanced NSCLC. Crit Rev Oncol Hematol 2020;152:102980. [CrossRef]
- van der Leun AM, Traets JJH, Vos JL, Elbers JBW, Patiwael S, Qiao X, et al. Dual Immune Checkpoint Blockade Induces Analogous Alterations in the Dysfunctional CD8+ T-cell and Activated Treg Compartment. Cancer Discov 2023;13:2212–27. [CrossRef]
- Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD, et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N Engl J Med 2015;373:23–34. [CrossRef]
- Postow MA, Chesney J, Pavlick AC, Robert C, Grossmann K, McDermott D, et al. Nivolumab and Ipilimumab versus Ipilimumab in Untreated Melanoma. N Engl J Med 2015;372:2006–17. [CrossRef]
- Casey SC, Amedei A, Aquilano K, Azmi AS, Benencia F, Bhakta D, et al. Cancer prevention and therapy through the modulation of the tumor microenvironment. Semin Cancer Biol 2015;35:S199–223. [CrossRef]
- Chen L xun, Zeng S jie, Liu X dong, Tang H bin, Wang J wu, Jiang Q. Cell-cell communications shape tumor microenvironment and predict clinical outcomes in clear cell renal carcinoma. J Transl Med 2023;21. [CrossRef]
- Kolb D, Kolishetti N, Surnar B, Sarkar S, Guin S, Shah AS, et al. Metabolic Modulation of the Tumor Microenvironment Leads to Multiple Checkpoint Inhibition and Immune Cell Infiltration. ACS Nano 2020;14:11055–66. [CrossRef]
- Devaud C, John LB, Westwood JA, Darcy PK, Kershaw MH. Immune modulation of the tumor microenvironment for enhancing cancer immunotherapy. Oncoimmunology 2013;2. [CrossRef]
Figure 1.
CTLs release GZMA and PRF1, leading cancer cells to apoptosis.
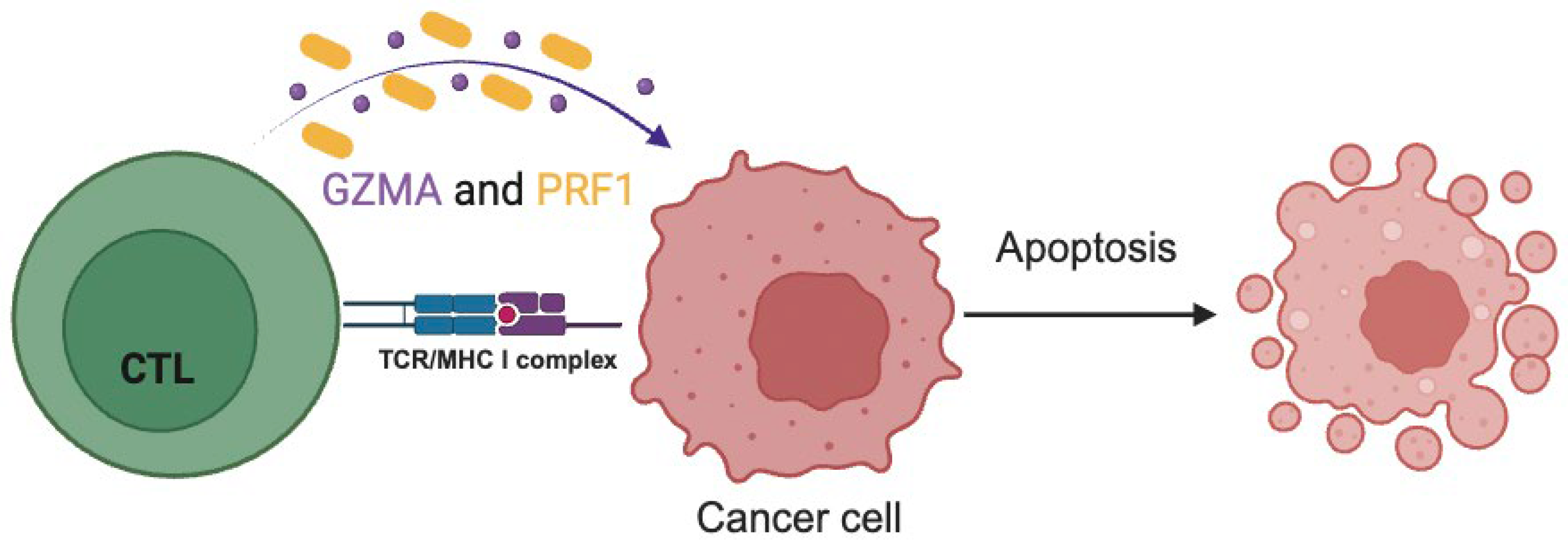
Figure 3.
Immune checkpoint Inhibitors.
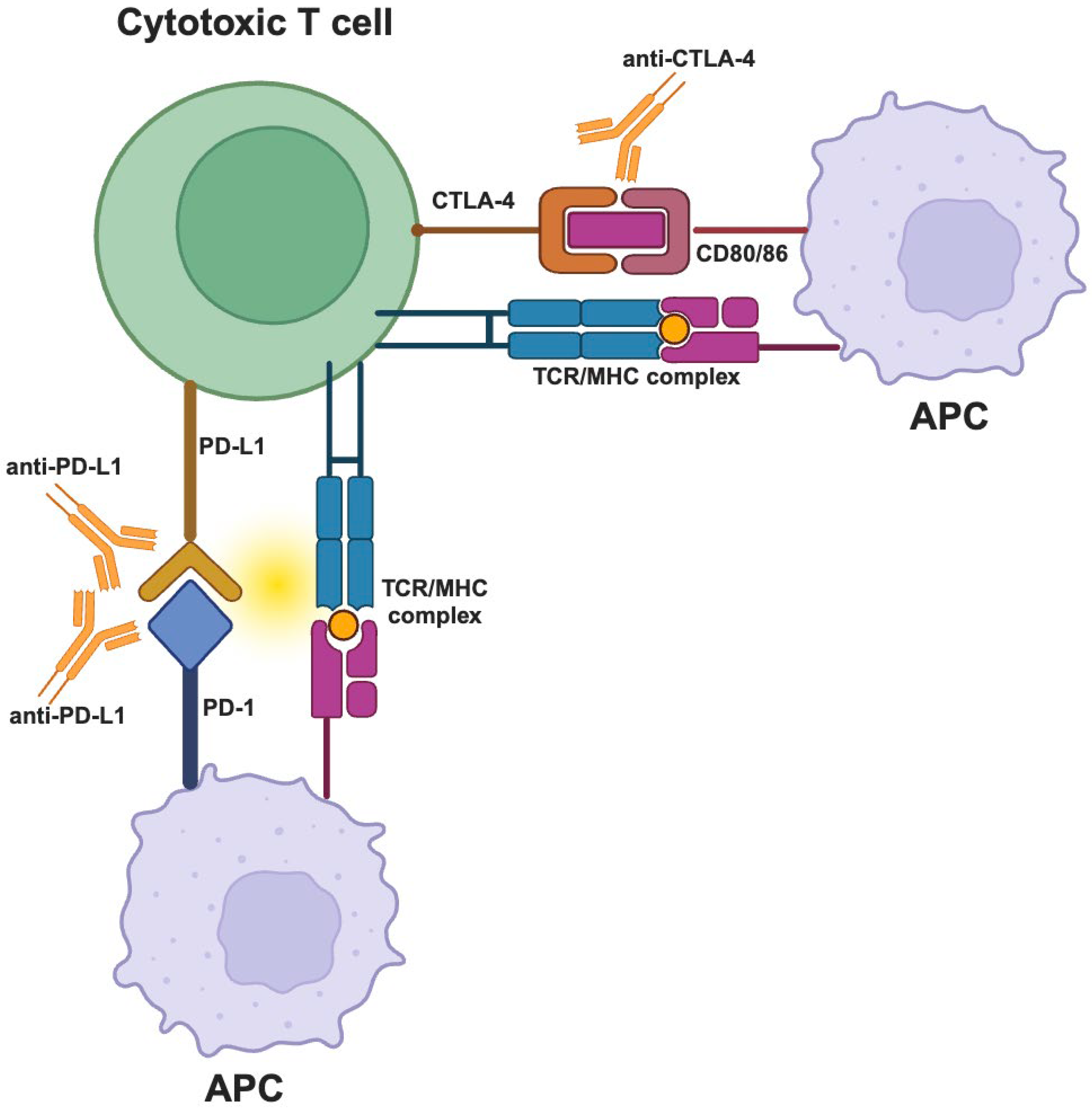
Figure 4.
CYT-high and CYT-low levels affect infiltration of immune cells, patient’s prognosis and survival.
Figure 4.
CYT-high and CYT-low levels affect infiltration of immune cells, patient’s prognosis and survival.
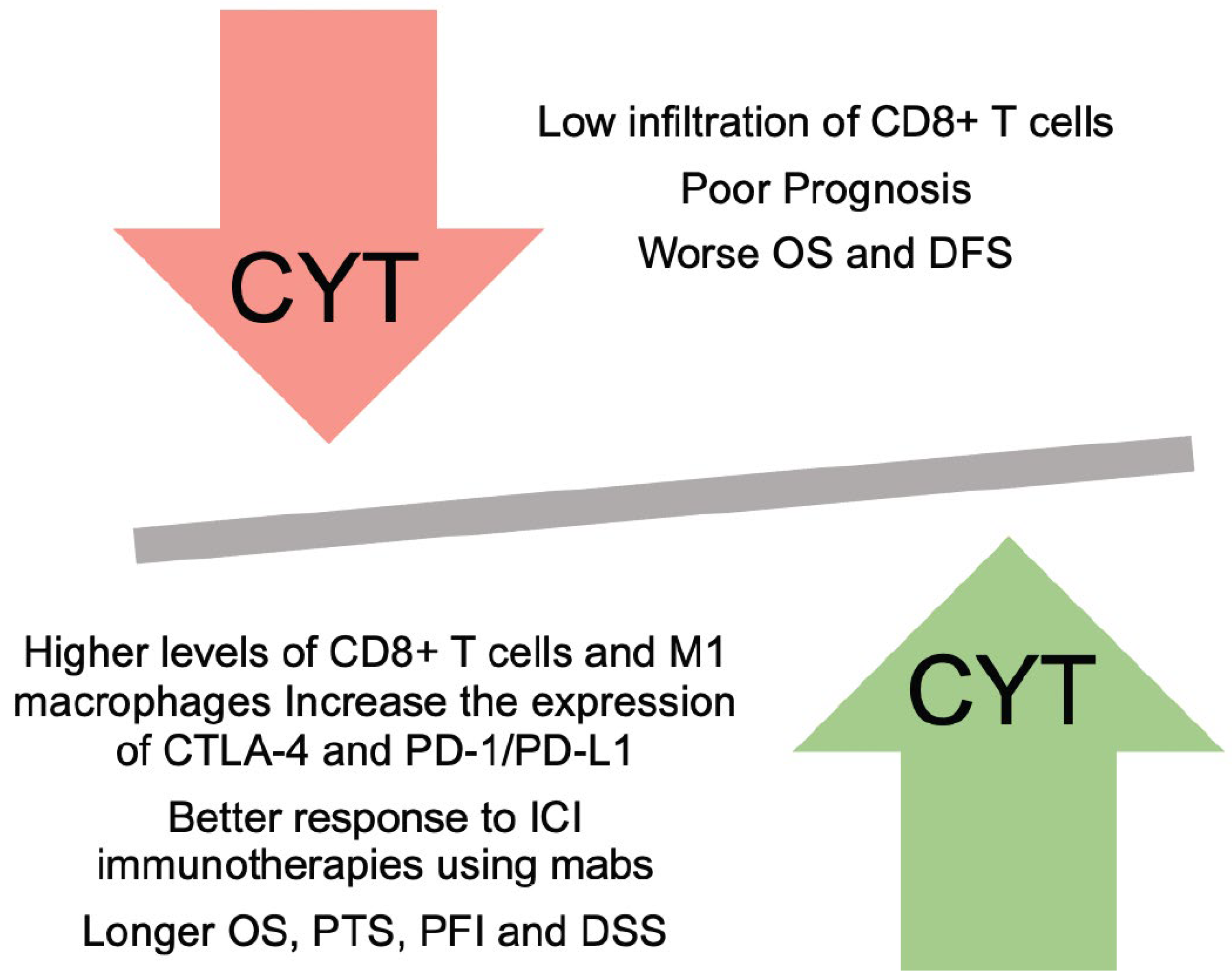
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



