Submitted:
31 January 2024
Posted:
01 February 2024
You are already at the latest version
Abstract
The NF-κB (nuclear factor K-light-chain-enhancer of activated B cells) transcription factor family is critical for modulating the immune proinflammatory response throughout the body. During the resting state, inactive NF-κB is sequestered by IκB in the cytoplasm. Proteasomal degradation of IκB activates NF-κB, mediating its translocation into the nucleus to act as a nuclear transcription factor in the upregulation of proinflammatory genes. Stimuli that initiate NF-κB activation are diverse, but are canonically attributed to proinflammatory cytokines and chemokines. Downstream effects of NF-κB are cell type specific and, in the majority of cases, result in activation of pro-inflammatory cascades. Acting as the primary immune responders of the central nervous system, microglia exhibit upregulation of NF-κB upon activation in response to pathological conditions. Under such circumstances, microglial crosstalk with other cell types in the central nervous system can induce cell death, further exacerbating the disease pathology. In this review, we will emphasize the role of NF-κB in triggering neuroinflammation mediated by microglia.
Keywords:
NF-κB
; inflammation
; microglia
1. NF-κB Signaling Pathway
The nuclear factor kappa B (NF-κB) family are the principal transcription factors involved in the regulation of immune-mediated inflammatory responses. Several hundred NF-κB target genes have been identified, with great diversity in function including cytokine/chemokine modulation, apoptosis, and cell proliferation [1,2]. The NF-κB family comprises 15 identified homo or heterodimers originating from five protein subunits: p65 (RelA), c-Rel, RelB, p50/p105 (NF-κB1), and p52/p100 (NF-κB2) [1,3]. All NF-κB subunits are structurally similar and possess a Rel Homology Domain (RHD) [4,5]. The highly conserved RHD is composed of a DNA binding domain at the N-terminal and a site for interaction with inhibitors at the C-terminal [5]. The NF-κB subunits can be broadly categorized into Class I precursors (p105, p100) and Class II Rel proteins (RelA, RelB, c-Rel) [1]. Class I precursors undergo C-terminal cleavage during processing before maturation into active p50 (NF-κB1) and p52 (NF-κB2) [6] (Figure 1).
Under homeostatic conditions, NF-κB is inactivated and sequestered to the cytoplasm by natural inhibitors of nuclear factor kappa B (IkB) proteins (primarily IκBα, IκBβ, and IκBε) [7]. Upon NF-κB activation, two discrete signaling pathways including canonical (classical) and non-canonical (alternative) pathways can be stimulated in many cell populations (Figure 2). In canonical signaling, RelA-p50 heterodimers are translocated to the nucleus to regulate transcription of target genes, whereas RelB-p52 heterodimers are the counterpart in non-canonical signaling [7,8]. The canonical NF-κB signaling pathway is regarded as the central regulator of the inflammatory response in normal physiology and it has been extensively studied in the context of human disorders [7]. In contrast, the non-canonical signaling pathway is characteristic of immune disorders, namely of autoimmune or inflammatory nature [8]. While the pathways differ in stimuli and mechanism of action, both canonical and non-canonical NF-κB signaling are represented in inflammation.
Within immune cells, the canonical NF-κB pathway diverges depending on innate versus adaptive responses. Innate immune cells express pattern recognition receptors (PRRs), which can recognize pathogen-associated molecular patterns (PAMPS) or damage-associated molecular patterns (DAMPS) released by damaged or dying cells [9,10]. PAMPs can be subcategorized depending on their bacterial, viral, or eukaryotic origin and can range from lipopolysaccharide (found in bacterial cell walls) to viral nucleic acids and proteins [11]. DAMP agents are diverse and can include proinflammatory cytokines (primarily TNF-ɑ, IL-1, and IL-6), chemokines, and other signals related to physical or chemical stress [12].
In the canonical NF-κB signaling pathway, diverse stimuli converge at the stimulation of transforming growth factor-beta-activated kinase 1 (TAK1) [13]. TAK1 subsequently activates the IkB kinase (IKK) complex, prevalently considered the master regulator of the NF-κB pathway. The IKK complex mediates the phosphorylation of IkBɑ at a serine residue for ubiquitination. This leads to the degradation of IkBɑ, thus triggering NF-κB nuclear translocation and binding to its specific DNA consensus site, typically located at gene promoters or enhancers [13,14]. Once in the nucleus, NF-κB can bind to the κB site of various genes participating in inflammation or mediating the immune response. The gene products of NF-κB activation in one cell type can initiate an adaptive immune response, triggering another NF-κB signaling pathway in another cell type [15]. Beyond immune response, NF-κB1 is implicated in many processes depending on cell type, including differentiation, regulation of apoptosis, and even metabolic responses [16].
Noncanonical NF-κB signaling is typically slower to activate and sustained in its response [17]. The noncanonical pathway is triggered by more specific stimuli, namely the tumor necrosis factor receptor (TNFR) family (which also induces canonical NF-κB signaling) [18]. The mechanism of action for noncanonical NF-κB signaling is also unique. Distinct stimuli converge with activation of NF-κB-inducing kinase (NIK), which subsequently activates IKKɑ [8,19]. The precursor molecule for NF-κB2 is inactivated by the p100 subunit [8]. Much like in canonical signaling, IKKɑ mediates p100 phosphorylation and eventual ubiquitination for proteasomal degradation [8]. At this point, the active NF-κB2 molecule (comprising p52 and RelB subunits) is translocated into the nucleus for differential regulation of genes involved in inflammation, cell survival, and development [18].
2. NF-κB in the Central Nervous System
NF-κB plays a multifaceted role within the central nervous system, with diverse roles in each cell type that vary depending on physiological conditions [16]. Within neurons, NF-κB activation can produce both neuroprotective and neurodegenerative effects [20,21,22,23]. NF-κB activation in oligodendrocytes also has potential to promote cell survival [24,25,26]. However, NF-κB activation in astrocytes and microglia typically leads to a more detrimental effect [27]. An overview of the functions of NF-κB signaling are outlined in Table 1.
2.1. Neurons
Broadly, NF-κB is intricately involved in several disparate aspects of neuron cell function, comprising both neural behavior under normal physiological conditions and neuron survival under disease conditions. NF-κB involvement has been documented in synaptic plasticity, growth factor signaling, and even higher order cognition (including learning and memory) [28]. The function of NF-κB in regulating the function of neurons under physiologic conditions is outside the scope of this review; however several recent reviews [16,27,28] offer a more detailed exploration of this topic. NF-κB is a key modulator of neuron survival, with a dual capacity to be both neuroprotective and neurodegenerative [20,21,22,23].
Typically, neuronal NF-κB signaling promotes cell survival. NF-κB selectively induces anti-apoptotic genes such as TNF receptor associated factors (namely TRAF-1 and TRAF-2), caspase inhibitors, apoptosis regulators in the Bcl-2 family, and superoxide dismutase (SOD) [20,29,30,31,32]. The neuroprotective effect of NF-κB is more explicitly represented in neurons derived from rodent models and subjected to apoptosis-promoting conditions. Within cultured embryonic rat hippocampal neurons, NF-κB activation by TNF-α and ceramide increased neuron survival under oxidative stress (FeSO4 and amyloid beta peptide treatment) [20]. These results were further confirmed when the neuroprotective effects were lost after introduction of decoy DNA to deactivate NF-κB [20]. The anti-apoptotic role of NF-κB is further corroborated in a transgenic mouse model with forebrain neuron specific expression of a dominant negative mutant of IκB resulting in inhibition of NF-κB. As examined in organotypic hippocampal slices, there was a subsequent increase in neuron cell death when exposed to neurotoxic insults (namely FeSO4 or kainate) [21].
Moreover, constitutive NF-κB activation has been shown to be necessary for neuron survival in select cases. Within primary cultured cortical neurons, an adenovirus encoding an IκB super-repressor was introduced for inhibition of neuronal NF-κB; under these circumstances, there was a significant reduction in cell survival. Adenovirus-induced overexpression of p50 (Rel A) in the same model resulted in increased cell survival due to accumulation of protein protectors against neuronal apoptosis due to etoposide and camptothecin [33]. A similar effect was observed in primary cultured embryonic mouse motor neurons. When interference RNA was used to selectively downregulate IKKα, IKKβ, or RelA proteins necessary for NF-κB activation, motor neuron apoptosis was induced [34]. NF-κB activation is also critical to the survival of developing sensory neurons. Embryos lacking p65 yield significantly fewer sensory neurons in culture [35]. Therefore, NF-κB activity may be intrinsic to the survival of particular neuron populations.
Conversely, some studies describe a neurodegenerative role of NF-κB signaling. A potential mechanism for neuronal cell death via endogenous NF-κB can be attributed to the induction of p53 via TNF-α activated NF-κB [36,37]. Tumor suppressor p53 has been associated with neuronal cell death: p53 knockout mice displayed decreased neuronal apoptosis in response to kainic acid excitation [22]. Furthermore, the TNF-α/NF-κB/p53 axis was identified by transcriptomic analysis as a contributor to cell death in human pluripotent stem cell (hPSC) derived dopaminergic neurons engrafted into the striatum of mice. When TNF-α/NF-κB signaling was chemically inhibited, there was enhanced survival of engrafted neurons, experimentally validating the pathway as a mediator of neuronal cell death [23].
The negative effect of NF-κB within neurons has been primarily observed in rodent models of ischemia. Nuclear NF-κB p50 and p65 localization was heightened in hippocampal neurons at the time of cell death in a rat model of global ischemia [38]. These results were further corroborated in human stroke patients, wherein activated NF-κB was detected in neurons at penumbral sites of the sampled brain sections [39]. However, there is conflicting evidence in studies evaluating p50 knockout mice for neuronal degeneration, wherein NF-κB was found to have a negative effect in a model of ischemia and a negative effect in a model of stroke [40,41]. These opposing results could be attributed to potential differences in the subunit composition of expressed neuronal NF-κB. Within cerebellar granule cells, the p65 subunit was implicated in neuronal cell death, whereas c-Rel was found to be essential for cell survival in the case of glutamate-induced excitotoxicity [42]. Therefore, the causal relationship between NF-κB and neuronal cell death needs further investigation.
2.2. Glia
NF-κB signaling in glia is primarily associated with inflammation secondary to a pathologic condition, whether as disease or trauma. Within the body, inflammation is the prototypic immune response to any fluctuation from homeostasis in efforts to eradicate abnormal stimuli and promote recovery. The CNS follows the same parameters, wherein microglia, astrocytes, and infiltrating leukocytes (in response to inflammatory conditions) primarily induce inflammation via NF-κB.
2.2.1. Oligodendrocytes
Present research remains divided regarding the effect of NF-κB activation in both healthy and diseased oligodendrocytes. NF-κB demonstrates a protective role in some studies, while other studies suggest a more dispensable role for NF-κB. Activation of NF-κB in oligodendrocyte precursor cells was found to decrease cell apoptosis and promote cell maturation, thereby indirectly contributing to myelination in the CNS [43]. In contrast, in a study involving CNS-wide deletion of RelA, histological and electron microscopic analysis of the optic nerve showed unimpaired oligodendrocyte densities and normal myelin sheath formation, suggesting that NF-κB is expendable in oligodendrocyte function [44]. Moreover, chronic NF-κB activation in mature oligodendrocytes is found to promote inflammatory conditions in the CNS similar to those in an aging brain [45]. In transgenic mice with constitutively active IKK2, RNA-Seq analysis revealed that the primary oligodendrocytes had gene expression signatures associated with increased post mitotic cellular senescence. These mice had increased white matter degeneration and myelination deficits characteristic of an aging brain [45]. Overall, existing studies demonstrate conflicting effects for NF-κB in healthy oligodendrocytes; additional studies must be conducted to establish a causal relationship of NF-κB in oligodendrocyte survival.
The NF-κB pathway has been more robustly explored in the context of multiple sclerosis (MS), particularly with respect to protection against inflammation, remyelination, and oligodendrocyte survival. In vitro studies in oligodendroglial cell lines revealed that plasmid based activation of NF-κB decreased apoptosis, whereas inhibition of NF-κB increased cytotoxicity under inflammatory conditions caused by TNF-α, IFN- γ, or reactive chemical species [24,25]. These results are corroborated in an experimental autoimmune encephalomyelitis (EAE) mouse model of MS characterized by ectopic expression of IFN-γ in the CNS and an oligodendrocyte specific expression of a dominant negative mutant of IκB for NF-κB super-repression [26]. Inactivation of NF-κB exacerbated oligodendrocyte death and induced hypomyelination in developing mice or remyelination failure in adult mice [26]. However, one study evaluating oligodendrocyte specific deletion of IKK2 for NF-κB inhibition in an EAE mouse model shows comparable numbers of oligodendrocyte progenitor cells and mature oligodendrocytes to the wildtype control, suggesting a more expendable role for NF-κB [46]. A potential cause for the conflicting evidence is that NF-κB activation in oligodendrocytes has also been traced to pancreatic endoplasmic reticulum kinase (PERK) signaling, an IKK independent pathway in models of EAE and MS [24,47,48]. Therefore, in models of NF-κB inhibition via deletion of upstream proteins in the canonical pathway, there is still potential for residual NF-κB activation via the PERK pathway. Taken together, the data suggest a protective role for NF-κB activity in oligodendrocytes under inflammatory conditions.
2.2.2. Astrocytes
Mirroring the multifaceted functions of astrocytes in the central nervous system, the activation of NF-κB in astrocytes exhibits a similar diversity. NF-κB has been implicated in astrocyte dependent clearing of synaptic glutamate, metabolic control, and modulation of astrocyte structural plasticity, as further described in two recent reviews [16,49]. In disease conditions, astrocytic NF-κB signaling is predominantly associated with increased inflammation [50,51,52]. In models of brain and spinal cord injury, NF-κB activity is upregulated. Immunohistochemical staining for p50 and p65 subunits in astrocytes after focal brain injury revealed increased NF-κB expression in the early days following the injury [50]. Often, this increase in NF-κB expression is associated with adverse outcomes, particularly in the case of chronic activation within astrocytes. Many studies have investigated the effect of inhibiting NF-κB activation in astrocytes. Within a mouse model of contusive spinal cord injury, selective inactivation of astrocytic NF-κB via transgenic IKKBα inhibition resulted in reduced lesion volume, white matter preservation, and expression of proinflammatory cytokines [51]. In a similar study of spinal cord injury, decreased NF-κB activity promoted sparing of spinal circuits involved in locomotion with improved functional outcomes [52]. Generally, astrocytic NF-κB activation is associated with increased inflammation within the brain, often worsening pathologic outcomes.
2.2.3. Microglia
As the resident macrophages of the CNS, microglia are typically regulated by NF-κB in the context of inflammation. While microglia can adopt both pro- and anti-inflammatory phenotypes, NF-κB activation is primarily associated with a pro-inflammatory state. During the next section of this review, we will provide an in-depth exploration of microglial NF-κB activity and subsequent crosstalk with other cell types in several pathologies common to the CNS.
As recent research expounds on microglia from a nonimmune standpoint in the healthy brain, the NF-κB pathway has been highlighted in microglial differentiation and homeostasis. In zebrafish, programmed cell death protein 11 (PDCD11) knockout yielded decreased macrophage differentiation into microglia that was reversable by co-over-expression of c-Rel and p105 [32]. Another study revealed the critical role of NF-κB in establishing homeostatic density of microglia: conditional knockout of IKKβ (NF-κB inhibition) in a microglia depleted mouse model showed impaired microglial repopulation and inability to regain homeostatic density [53]. At present, microglial NF-κB activation in brain development and microglial differentiation represents an under-researched niche, with potential applications in regulating microglial number within the CNS to remediate pathological conditions.
3. Role of Microglial NF-κB in CNS Disease
3.1. Alzheimer’s Disease
Alzheimer's disease (AD) is primarily defined by gradual loss of memory, leading to a spectrum of cognitive and behavioral deficits. Primary contributors to AD pathogenesis are amyloid beta aggregates and tau tangles, neurofibrillary tangles composed of insoluble tau fibrils [51]. AD is associated with widespread neuroinflammation, largely proposed to be mediated by microglia [52]. Pathway analysis has identified NF-κB signaling as one of the most perturbed pathways in late onset AD [54]. NF-κB and beta amyloid plaques have been proposed to have a positive feedback relationship, wherein presence of plaques activates NF-κB in neurons leading to further production of plaques [55,56]. Emerging research has implicated NF-κB activation in microglia as a key driver of tau tangles [57]. Within a comprehensive study of primary microglia derived from a mouse model of tauopathy, NF-κB was among the top affected pathways, associated with upregulation in proinflammatory cytokine profile (IL-1B, TNF, IL-12B) [57]. Single cell RNA sequencing studies revealed that in disease-associated microglia (DAM), NF-κB target genes were differentially expressed with IKB kinase identified as the top upstream regulator [57]. NF-κB activation in microglia is associated with improved tau fibril clearance. Within microglia derived from NF-κB inactivated mice (due to IKKB inhibition), internalized tau fibril clearance was significantly slower than in NF-κB activated conditions [57]. Fundamentally, microglial NF-κB activation is associated with worsened AD outcomes, representing a potential avenue for neurotherapeutic development.
3.2. Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease marked by progressive motor neuron death. Clinical presentation of the disease includes gradual impairments in voluntary muscle movements, muscular atrophy, and eventual death due to dysphagia and dyspnea [58]. ALS has been associated with mutations in several genes: superoxide dismutase 1 (SOD1), TAR DNA-binding protein 43 (TARDBP), fused in sarcoma (FUS), TANK-binding kinase 1 (TBK1), and chromosome 9 open reading frame 72 (C9orf72) [58]. While motor neurons are the ultimate effectors of ALS, glial cells have been explored as potential mediators for neurodegeneration, with a particular focus on microglial activation in recent years [59,60].
Microglial NF-κB interactions have been documented in several models of ALS. Spinal cord extracts from sporadic ALS human patients were used to identify TARDBP colocalization with the p65 NF-κB subunit in microglial cells, suggesting that the canonical NF-κB pathway may contribute to TARDBP deregulation characteristic of ALS [61]. Furthermore, within a mouse model of ALS characterized by mutant SOD1, inhibition of microglial NF-κB rescued motor neurons in a subsequent ALS co-culture model in vitro and significantly increased mouse survival rates in vivo [62]. Selective NF-κB inhibition in this model decreased pro-inflammatory microglia markers (CD68, CD86, and iNOS), suggesting a direct role of NF-κB in ALS pathogenesis [62]. Remarkably, within the SOD1 mouse model, astrocytic NF-κB inhibition via both transgenic and viral IκB suppression was not sufficient to improve motor neuron survival or ALS progression, highlighting role of microglial NF-κB in inducing ALS presentation [62,63]. Overall, NF-κB remains a promising target for attenuation of microglia-mediated inflammation, which may improve outcomes for ALS patients.
3.3. Hypoxic Ischemic Encephalopathy
Perinatal hypoxic-ischemic encephalopathy (HIE) is a major cause of neonatal death and long-term disability in term infants due to resultant cerebral palsy, epilepsy, learning delays, and visual impairments [64]. In this condition, the brain is injured as a result of lack of oxygen to the CNS due to complications during birth (such as cord prolapse, uterine rupture, breech presentation, chorioamnionitis) [64]. However, inflammation is a major driver of the secondary energy failure that occurs about 6-120 hours after the initial insult and is primarily driven by microglia and macrophages [65,66]. Interestingly, the NF-κB pathway becomes activated in hypoxic conditions [67]. Primary cultured microglia derived from a neonatal hypoxia-exposed rat model revealed upregulation in Toll-Like Receptor 4 (TLR4, a significant PAMP) and phosphorylated NF-κB p65 proteins [68]. Within in vivo models of HIE, specifically, NF-κB is a major mediator of microglia-induced inflammation. Selective inhibition of NF-κB in microglia in a rodent model of neonatal hypoxic ischemic injury attenuated brain injury as evidenced through decreased ventriculomegaly and proinflammatory cytokine presentation in combination with improved locomotor outcomes [69]. This suggests that NF-κB mediated signaling could be harnessed to reduce injury to the CNS in hypoxic ischemic encephalopathy.
4. Conclusion
The intricate relationship between the NF-κB pathway and microglial activation is gradually receiving more attention within the scope of research into neuroinflammation. The response of microglia to NF-κB induced activation is particularly emphasized, especially in the context of cell and animal models representative of various neurodegenerative conditions. Microglia exhibit proinflammatory functional changes upon NF-κB activation, ultimately leading to the release of proinflammatory cytokines and chemokines which act on other cells in the central nervous system. Microglia specific NF-κB inhibition is typically associated with positive downstream results, but more extensive research must be conducted into the intercellular and molecular interactions underlying this beneficial effect. Overall, the diverse effect of NF-κB on cell types within the central nervous system reviewed here supports the need for continued investigation into manipulating this molecular pathway for future neuroprotective therapies.
Author Contributions
Conceptualization, S.A. and E.W-J.; writing—original draft preparation, S.A.; writing—review and editing, E.W-J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol 2009, 1, a000034. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [PubMed]
- Christian, F.; Smith, E.L.; Carmody, R.J. The Regulation of NF-κB Subunits by Phosphorylation. Cells 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Ghosh, S. Rel/NF-kappa B and I kappa B proteins: an overview. Semin Cancer Biol 1997, 8, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Molecular Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Yin, Q.; Wu, H. Structural studies of NF-κB signaling. Cell Res 2011, 21, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduction and Targeted Therapy 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C. Non-canonical NF-κB signaling pathway. Cell Res 2011, 21, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Ernst, O.; Vayttaden, S.J.; Fraser, I.D.C. Measurement of NF-κB Activation in TLR-Activated Macrophages. Methods Mol Biol 2018, 1714, 67–78. [Google Scholar]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol Rev 2012, 249, 158–175. [Google Scholar] [CrossRef]
- Patel, S. Danger-Associated Molecular Patterns (DAMPs): the Derivatives and Triggers of Inflammation. Curr Allergy Asthma Rep 2018, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: mechanism and clinical study. Signal Transduction and Targeted Therapy 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Mussbacher, M.; Derler, M.; Basílio, J.; Schmid, J.A. NF-κB in monocytes and macrophages - an inflammatory master regulator in multitalented immune cells. Front Immunol 2023, 14, 1134661. [Google Scholar] [CrossRef] [PubMed]
- Mussbacher, M.; Salzmann, M.; Brostjan, C.; Hoesel, B.; Schoergenhofer, C.; Datler, H.; Hohensinner, P.; Basílio, J.; Petzelbauer, P.; Assinger, A.; et al. Cell Type-Specific Roles of NF-κB Linking Inflammation and Thrombosis. Front Immunol 2019, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, E.C.; Meffert, M.K. Cellular Specificity of NF-κB Function in the Nervous System. Front Immunol 2019, 10, 1043. [Google Scholar] [CrossRef]
- Sun, S.-C. Non-canonical NF-κB signaling pathway. Cell Research 2011, 21, 71–85. [Google Scholar] [CrossRef]
- Cildir, G.; Low, K.C.; Tergaonkar, V. Noncanonical NF-κB Signaling in Health and Disease. Trends Mol Med 2016, 22, 414–429. [Google Scholar] [CrossRef]
- Sun, S.-C. The non-canonical NF-κB pathway in immunity and inflammation. Nature Reviews Immunology 2017, 17, 545–558. [Google Scholar] [CrossRef]
- Mattson, M.P.; Goodman, Y.; Luo, H.; Fu, W.; Furukawa, K. Activation of NF-kappaB protects hippocampal neurons against oxidative stress-induced apoptosis: evidence for induction of manganese superoxide dismutase and suppression of peroxynitrite production and protein tyrosine nitration. Journal of neuroscience research 1997, 49, 681–697. [Google Scholar] [CrossRef]
- Fridmacher, V.; Kaltschmidt, B.; Goudeau, B.; Ndiaye, D.; Rossi, F.M.; Pfeiffer, J.; Kaltschmidt, C.; Israël, A.; Mémet, S. Forebrain-specific neuronal inhibition of nuclear factor-kappaB activity leads to loss of neuroprotection. J Neurosci 2003, 23, 9403–9408. [Google Scholar] [CrossRef]
- Morrison, R.S.; Wenzel, H.J.; Kinoshita, Y.; Robbins, C.A.; Donehower, L.A.; Schwartzkroin, P.A. Loss of the p53 tumor suppressor gene protects neurons from kainate-induced cell death. J Neurosci 1996, 16, 1337–1345. [Google Scholar] [CrossRef]
- Kim, T.W.; Koo, S.Y.; Riessland, M.; Cho, H.; Chaudhry, F.; Kolisnyk, B.; Russo, M.V.; Saurat, N.; Mehta, S.; Garippa, R.; et al. TNF-NFkB-p53 axis restricts in vivo survival of hPSC-derived dopamine neuron. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lin, Y.; Jamison, S.; Lin, W. Interferon-γ activates nuclear factor-κ B in oligodendrocytes through a process mediated by the unfolded protein response. PLoS ONE 2012, 7, e36408. [Google Scholar] [CrossRef]
- Hamanoue, M.; Yoshioka, A.; Ohashi, T.; Eto, Y.; Takamatsu, K. NF-kappaB prevents TNF-alpha-induced apoptosis in an oligodendrocyte cell line. Neurochem Res 2004, 29, 1571–1576. [Google Scholar] [CrossRef]
- Stone, S.; Jamison, S.; Yue, Y.; Durose, W.; Schmidt-Ullrich, R.; Lin, W. NF-κB Activation Protects Oligodendrocytes against Inflammation. J Neurosci 2017, 37, 9332–9344. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Kaltschmidt, C. NF-kappaB in the nervous system. Cold Spring Harb Perspect Biol 2009, 1, a001271. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Meffert, M.K. Roles for NF-κB in nerve cell survival, plasticity, and disease. Cell Death & Differentiation 2006, 13, 852–860. [Google Scholar]
- Marchetti, L.; Klein, M.; Schlett, K.; Pfizenmaier, K.; Eisel, U.L. Tumor necrosis factor (TNF)-mediated neuroprotection against glutamate-induced excitotoxicity is enhanced by N-methyl-D-aspartate receptor activation. Essential role of a TNF receptor 2-mediated phosphatidylinositol 3-kinase-dependent NF-kappa B pathway. J Biol Chem 2004, 279, 32869–32881. [Google Scholar] [CrossRef]
- Tamatani, M.; Che, Y.H.; Matsuzaki, H.; Ogawa, S.; Okado, H.; Miyake, S.; Mizuno, T.; Tohyama, M. Tumor necrosis factor induces Bcl-2 and Bcl-x expression through NFkappaB activation in primary hippocampal neurons. J Biol Chem 1999, 274, 8531–8538. [Google Scholar] [CrossRef]
- Wang, C.Y.; Mayo, M.W.; Korneluk, R.G.; Goeddel, D.V.; Baldwin, A.S., Jr. NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science 1998, 281, 1680–1683. [Google Scholar] [CrossRef]
- Yang, R.; Zhan, M.; Guo, M.; Yuan, H.; Wang, Y.; Zhang, Y.; Zhang, W.; Chen, S.; de The, H.; Chen, Z.; et al. Yolk sac-derived Pdcd11-positive cells modulate zebrafish microglia differentiation through the NF-κB-Tgfβ1 pathway. Cell Death Differ 2021, 28, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Bhakar, A.L.; Tannis, L.L.; Zeindler, C.; Russo, M.P.; Jobin, C.; Park, D.S.; MacPherson, S.; Barker, P.A. Constitutive nuclear factor-kappa B activity is required for central neuron survival. J Neurosci 2002, 22, 8466–8475. [Google Scholar] [CrossRef] [PubMed]
- Mincheva, S.; Garcera, A.; Gou-Fabregas, M.; Encinas, M.; Dolcet, X.; Soler, R.M. The canonical nuclear factor-κB pathway regulates cell survival in a developmental model of spinal cord motoneurons. J Neurosci 2011, 31, 6493–6503. [Google Scholar] [CrossRef]
- Middleton, G.; Hamanoue, M.; Enokido, Y.; Wyatt, S.; Pennica, D.; Jaffray, E.; Hay, R.T.; Davies, A.M. Cytokine-induced nuclear factor kappa B activation promotes the survival of developing neurons. J Cell Biol 2000, 148, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Grilli, M.; Memo, M. Possible role of NF-kappaB and p53 in the glutamate-induced pro-apoptotic neuronal pathway. Cell Death Differ 1999, 6, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lozano, G. NF-kappa B activation of p53. A potential mechanism for suppressing cell growth in response to stress. J Biol Chem 1994, 269, 20067–20074. [Google Scholar] [CrossRef] [PubMed]
- Clemens, J.A.; Stephenson, D.T.; Smalstig, E.B.; Dixon, E.P.; Little, S.P. Global ischemia activates nuclear factor-kappa B in forebrain neurons of rats. Stroke 1997, 28, 1073–1080. [Google Scholar] [CrossRef]
- Nurmi, A.; Lindsberg, P.J.; Koistinaho, M.; Zhang, W.; Juettler, E.; Karjalainen-Lindsberg, M.L.; Weih, F.; Frank, N.; Schwaninger, M.; Koistinaho, J. Nuclear factor-kappaB contributes to infarction after permanent focal ischemia. Stroke 2004, 35, 987–991. [Google Scholar] [CrossRef]
- Duckworth, E.A.; Butler, T.; Collier, L.; Collier, S.; Pennypacker, K.R. NF-kappaB protects neurons from ischemic injury after middle cerebral artery occlusion in mice. Brain Res 2006, 1088, 167–175. [Google Scholar] [CrossRef]
- Schneider, A.; Martin-Villalba, A.; Weih, F.; Vogel, J.; Wirth, T.; Schwaninger, M. NF-κB is activated and promotes cell death in focal cerebral ischemia. Nature Medicine 1999, 5, 554–559. [Google Scholar] [CrossRef]
- Pizzi, M.; Goffi, F.; Boroni, F.; Benarese, M.; Perkins, S.E.; Liou, H.C.; Spano, P. Opposing roles for NF-kappa B/Rel factors p65 and c-Rel in the modulation of neuron survival elicited by glutamate and interleukin-1beta. J Biol Chem 2002, 277, 20717–20723. [Google Scholar] [CrossRef]
- Nicholas, R.S.; Wing, M.G.; Compston, A. Nonactivated microglia promote oligodendrocyte precursor survival and maturation through the transcription factor NF-kappa B. Eur J Neurosci 2001, 13, 959–967. [Google Scholar] [CrossRef]
- Kretz, A.; Herrmann, K.H.; Fischer, S.; Engelmann, C.; Witte, O.W.; Reichenbach, J.R.; Weih, F.; Haenold, R. Dysfunctional NF-κB and brain myelin formation. Eur J Hum Genet 2014, 22, 724–725. [Google Scholar] [CrossRef]
- Schlett, J.S.; Mettang, M.; Skaf, A.; Schweizer, P.; Errerd, A.; Mulugeta, E.A.; Hein, T.M.; Tsesmelis, K.; Tsesmelis, M.; Büttner, U.F.G.; et al. NF-κB is a critical mediator of post-mitotic senescence in oligodendrocytes and subsequent white matter loss. Molecular Neurodegeneration 2023, 18, 24. [Google Scholar] [CrossRef]
- Raasch, J.; Zeller, N.; van Loo, G.; Merkler, D.; Mildner, A.; Erny, D.; Knobeloch, K.P.; Bethea, J.R.; Waisman, A.; Knust, M.; et al. IkappaB kinase 2 determines oligodendrocyte loss by non-cell-autonomous activation of NF-kappaB in the central nervous system. Brain 2011, 134, 1184–1198. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Yue, Y.; Stone, S.; Wu, S.; Lin, W. NF-κB Activation Accounts for the Cytoprotective Effects of PERK Activation on Oligodendrocytes during EAE. J Neurosci 2020, 40, 6444–6456. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, G.; Jamison, S.; Li, J.; Harding, H.P.; Ron, D.; Lin, W. PERK activation preserves the viability and function of remyelinating oligodendrocytes in immune-mediated demyelinating diseases. Am J Pathol 2014, 184, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Yates, D. Factoring in astrocytes. Nature Reviews Neuroscience 2015, 16, 67–67. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, Y.; Yamamoto, M.; Fukushima, T.; Kimura, H.; Ohshima, K.; Tomonaga, M. Expression of Nuclear Factor κB and Tumor Necrosis Factor α in the Mouse Brain after Experimental Thermal Ablation Injury. Neurosurgery 2001, 48, 158–166. [Google Scholar]
- Brambilla, R.; Bracchi-Ricard, V.; Hu, W.H.; Frydel, B.; Bramwell, A.; Karmally, S.; Green, E.J.; Bethea, J.R. Inhibition of astroglial nuclear factor kappaB reduces inflammation and improves functional recovery after spinal cord injury. J Exp Med 2005, 202, 145–156. [Google Scholar] [CrossRef]
- Brambilla, R.; Hurtado, A.; Persaud, T.; Esham, K.; Pearse, D.D.; Oudega, M.; Bethea, J.R. Transgenic inhibition of astroglial NF-κB leads to increased axonal sparing and sprouting following spinal cord injury. Journal of Neurochemistry 2009, 110, 765–778. [Google Scholar] [CrossRef]
- Zhan, L.; Krabbe, G.; Du, F.; Jones, I.; Reichert, M.C.; Telpoukhovskaia, M.; Kodama, L.; Wang, C.; Cho, S.H.; Sayed, F.; et al. Proximal recolonization by self-renewing microglia re-establishes microglial homeostasis in the adult mouse brain. PLoS Biol 2019, 17, e3000134. [Google Scholar] [CrossRef]
- Li, X.; Long, J.; He, T.; Belshaw, R.; Scott, J. Integrated genomic approaches identify major pathways and upstream regulators in late onset Alzheimer's disease. Scientific reports 2015, 5, 12393. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Uherek, M.; Volk, B.; Baeuerle, P.A.; Kaltschmidt, C. Transcription factor NF-kappaB is activated in primary neurons by amyloid beta peptides and in neurons surrounding early plaques from patients with Alzheimer disease. Proceedings of the National Academy of Sciences of the United States of America 1997, 94, 2642–2647. [Google Scholar] [CrossRef]
- Chen, C.H.; Zhou, W.; Liu, S.; Deng, Y.; Cai, F.; Tone, M.; Tone, Y.; Tong, Y.; Song, W. Increased NF-κB signalling up-regulates BACE1 expression and its therapeutic potential in Alzheimer's disease. The international journal of neuropsychopharmacology 2012, 15, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fan, L.; Khawaja, R.R.; Liu, B.; Zhan, L.; Kodama, L.; Chin, M.; Li, Y.; Le, D.; Zhou, Y.; et al. Microglial NF-κB drives tau spreading and toxicity in a mouse model of tauopathy. Nature Communications 2022, 13, 1969. [Google Scholar] [CrossRef]
- Masrori, P.; Van Damme, P. Amyotrophic lateral sclerosis: a clinical review. Eur J Neurol 2020, 27, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Boillée, S.; Vande Velde, C.; Cleveland, D.W. ALS: a disease of motor neurons and their nonneuronal neighbors. Neuron 2006, 52, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Clement, A.M.; Nguyen, M.D.; Roberts, E.A.; Garcia, M.L.; Boillée, S.; Rule, M.; McMahon, A.P.; Doucette, W.; Siwek, D.; Ferrante, R.J.; et al. Wild-type nonneuronal cells extend survival of SOD1 mutant motor neurons in ALS mice. Science 2003, 302, 113–117. [Google Scholar] [CrossRef]
- Swarup, V.; Phaneuf, D.; Dupré, N.; Petri, S.; Strong, M.; Kriz, J.; Julien, J.P. Deregulation of TDP-43 in amyotrophic lateral sclerosis triggers nuclear factor κB-mediated pathogenic pathways. J Exp Med 2011, 208, 2429–2447. [Google Scholar] [CrossRef]
- Frakes, A.E.; Ferraiuolo, L.; Haidet-Phillips, A.M.; Schmelzer, L.; Braun, L.; Miranda, C.J.; Ladner, K.J.; Bevan, A.K.; Foust, K.D.; Godbout, J.P.; et al. Microglia induce motor neuron death via the classical NF-κB pathway in amyotrophic lateral sclerosis. Neuron 2014, 81, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Crosio, C.; Valle, C.; Casciati, A.; Iaccarino, C.; Carrì, M.T. Astroglial inhibition of NF-κB does not ameliorate disease onset and progression in a mouse model for amyotrophic lateral sclerosis (ALS). PLoS One 2011, 6, e17187. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Yang, S.N. Perinatal hypoxic-ischemic encephalopathy. J Biomed Biotechnol 2011, 2011, 609813. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; McCullough, L.D. Inflammatory responses in hypoxic ischemic encephalopathy. Acta Pharmacol Sin 2013, 34, 1121–1130. [Google Scholar] [CrossRef]
- Fumagalli, S.; Perego, C.; Pischiutta, F.; Zanier, E.R.; De Simoni, M.G. The ischemic environment drives microglia and macrophage function. Front Neurol 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- D'Ignazio, L.; Rocha, S. Hypoxia Induced NF-κB. Cells 2016, 5. [Google Scholar] [CrossRef]
- Yao, L.; Kan, E.M.; Lu, J.; Hao, A.; Dheen, S.T.; Kaur, C.; Ling, E.-A. Toll-like receptor 4 mediates microglial activation and production of inflammatory mediators in neonatal rat brain following hypoxia: role of TLR4 in hypoxic microglia. Journal of Neuroinflammation 2013, 10, 785. [Google Scholar] [CrossRef]
- Zaghloul, N.; Kurepa, D.; Bader, M.Y.; Nagy, N.; Ahmed, M.N. Prophylactic inhibition of NF-κB expression in microglia leads to attenuation of hypoxic ischemic injury of the immature brain. Journal of Neuroinflammation 2020, 17, 365. [Google Scholar] [CrossRef]
Figure 1.
Structure of NF-κB dimers. All proteins in the NF-κB family are comprised of 2 subunits. Class I precursor proteins are proteolyzed into active NF-κB1 (p50) and NF-κB2 (p52) subunits. Upon proteolysis, these active subunits can bind to 3 known Class II proteins in the Rel family (p65/RelA, c-Rel, and p52/RelB).
Figure 1.
Structure of NF-κB dimers. All proteins in the NF-κB family are comprised of 2 subunits. Class I precursor proteins are proteolyzed into active NF-κB1 (p50) and NF-κB2 (p52) subunits. Upon proteolysis, these active subunits can bind to 3 known Class II proteins in the Rel family (p65/RelA, c-Rel, and p52/RelB).
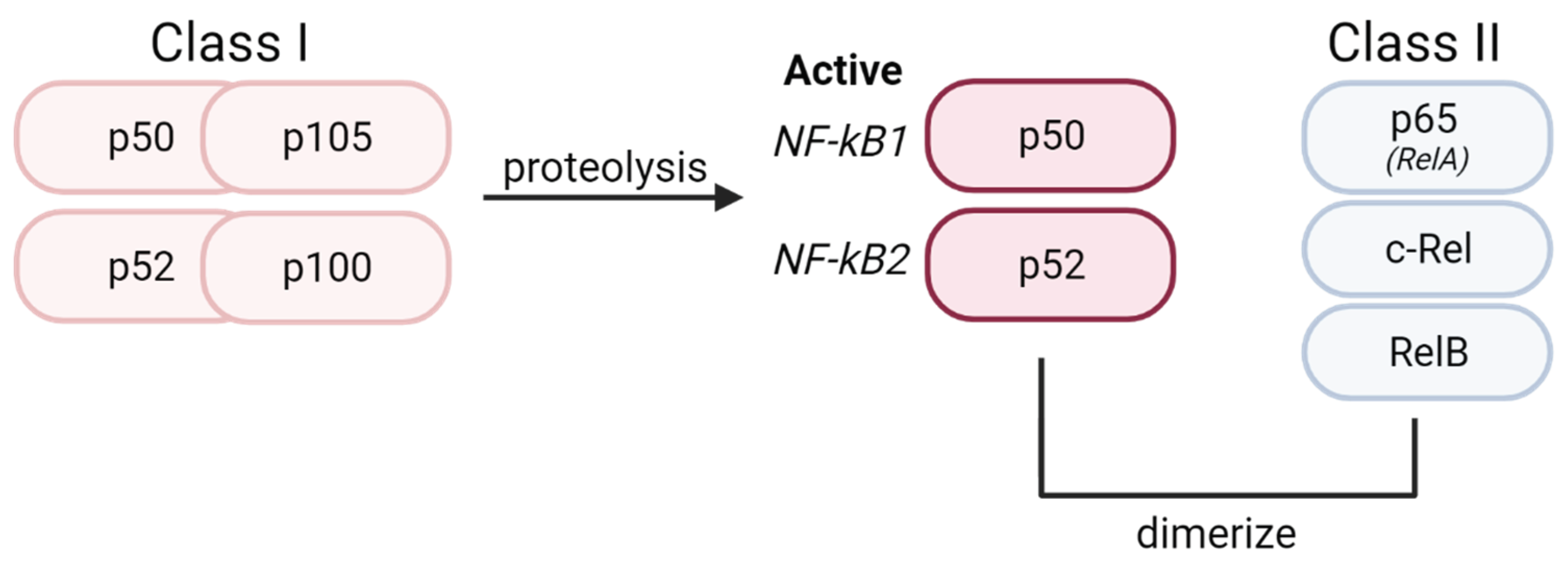
Figure 2.
Canonical and noncanonical NF-κB signaling. Under homeostatic conditions, NF-κB exists in an inactivated state and is sequestered in the cytoplasm by IκB proteins. Upon immune activation, pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs) stimulate canonical NF-kB signaling. PAMP and DAMP agents are diverse; including lipopolysaccharides (LPS), proinflammatory cytokines, and cellular stress molecules. These stimuli converge in TAK1 stimulation, which facilitates activation of the IKK complex, a tetramer composed of IKKα, IKKβ, and the regulatory IKKγ subunit. The activated IKK complex mediates phosphorylation of the IκB protein, enabling its eventual ubiquitination to allow the release of active NF-κB. Stimuli inducing noncanonical signaling include proteins within the TNFR and RANKL families. In noncanonical signaling, the first major activated protein is NIK, which stimulates IKKα to phosphorylate the p100 subunit, leading to the eventual ubiquitination of the p100 subunit. This allows activated NF-kB transcription factor to translocate into the nucleus to influence gene expression.
Figure 2.
Canonical and noncanonical NF-κB signaling. Under homeostatic conditions, NF-κB exists in an inactivated state and is sequestered in the cytoplasm by IκB proteins. Upon immune activation, pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs) stimulate canonical NF-kB signaling. PAMP and DAMP agents are diverse; including lipopolysaccharides (LPS), proinflammatory cytokines, and cellular stress molecules. These stimuli converge in TAK1 stimulation, which facilitates activation of the IKK complex, a tetramer composed of IKKα, IKKβ, and the regulatory IKKγ subunit. The activated IKK complex mediates phosphorylation of the IκB protein, enabling its eventual ubiquitination to allow the release of active NF-κB. Stimuli inducing noncanonical signaling include proteins within the TNFR and RANKL families. In noncanonical signaling, the first major activated protein is NIK, which stimulates IKKα to phosphorylate the p100 subunit, leading to the eventual ubiquitination of the p100 subunit. This allows activated NF-kB transcription factor to translocate into the nucleus to influence gene expression.
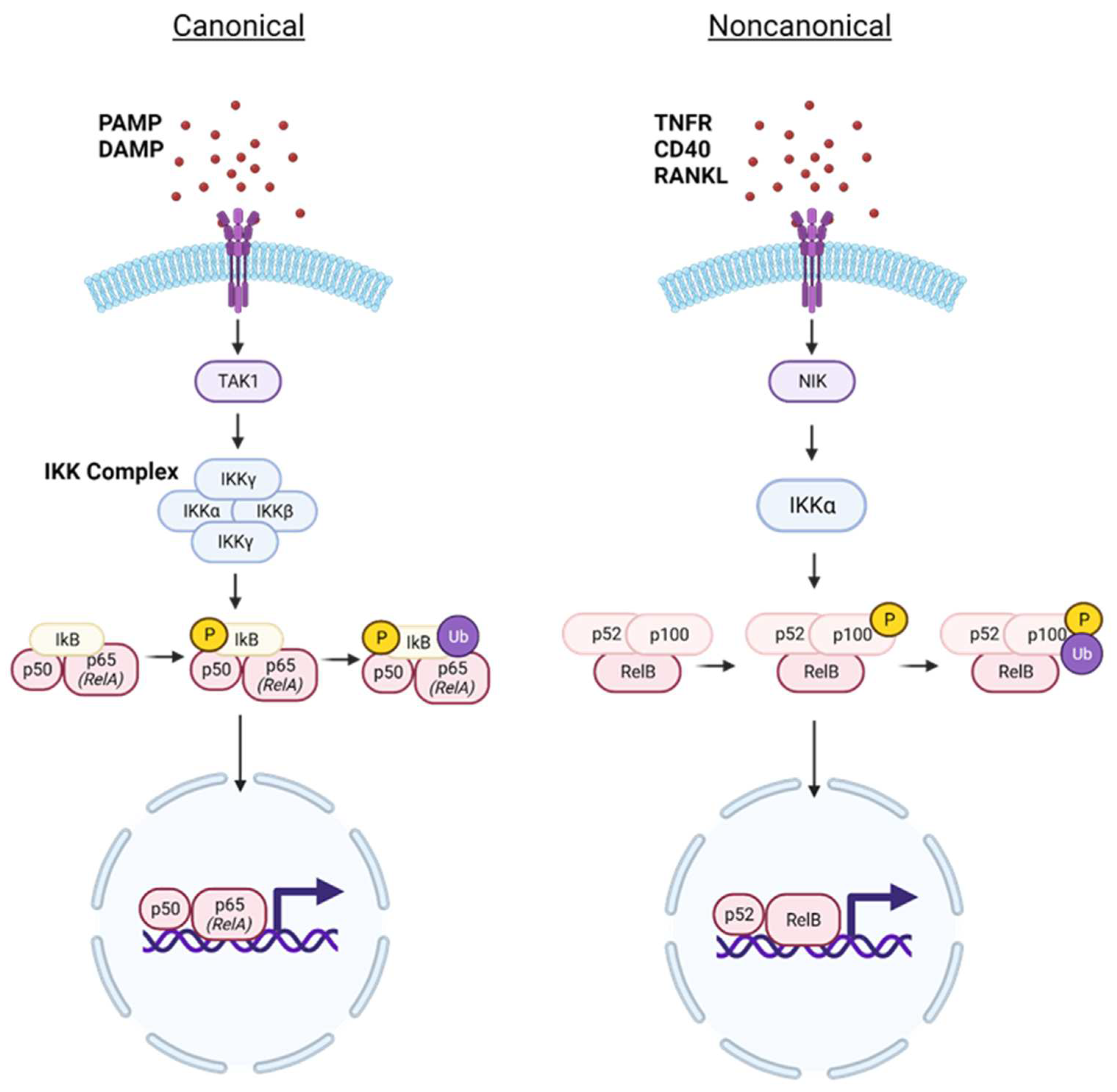

Table 1.
Role of NF-κB activation in cell types found in the central nervous system.
| Cell Type | NF-κB | ||
| Protective (Against) | Detrimental (In) |
Constitutively Active |
|
| Neuron | Oxidative Stress | Ischemia | In survival |
| In development | |||
| Oligodendrocyte | Inflammatory Cytokines | In survival (possible) | |
| Multiple Sclerosis (EAE) | |||
| Astrocyte | Spinal Cord Injury | ||
| Microglia | Alzheimer’s Disease | ||
| Amyotrophic Lateral Sclerosis (SOD1 mutant) Hypoxic Ischemic Encephalopathy |
|||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



