
Submitted:
31 January 2024
Posted:
01 February 2024
You are already at the latest version
Abstract
Kodamaea ohmeri is an environmental yeast considered a rare emerging pathogen. In clinical settings, the correct identification of this yeast is relevant since some isolates are associated with resistance to antifungals. There is a lack of available data regarding the geographical distribution, virulence, and drug resistance profile of K. ohmeri. To contribute to the knowledge of this yeast, this study aimed to describe in depth three isolates of K. ohmeri associated with fungemia in Honduras. The identification of the isolates was carried out by sequencing the ribosomal ITS region. In addition, the sensitivity profile to antifungals was determined, and some properties associated with virulence were evaluated (exoenzyme production, biofilm formation, cell adhesion, and invasion). The isolates showed strong protease, phospholipase, and hemolysin activity, in addition to being biofilm producers. Adherence and invasion capacity were evident in the HeLa and Raw 264.7 cell lines respectively. This study expands the understanding of the underlying biological traits associated with virulence in K. ohmeri and is the first report of the detection and identification of K. ohmeri in Honduras as a cause of human infection.
Keywords:
Kodamaea ohmeri
; Honduras
; virulence
; diagnosis
; adhesion
; macrophages
1. Introduction
Fungi represent one of the most varied kingdoms of eukaryotes and it is believed that they constitute between 1.5 and 5 million fungal species [1], with nearly 300 species of fungi associated with infections in humans [2,3]. The selection, adaptation, and dissemination of novel fungal infections have been influenced by multiple factors in recent years, including climate change, the widespread use of antifungals in agriculture, and large-scale migrations [4,5,6,7]. These novel organisms have been successfully identified due to advancements in methods utilizing molecular biology and phylogenetic analysis.
Kodamaea ohmeri (homotypic synonym: Yamadazyma ohmeri/Pichia ohmeri) is a ubiquitous yeast strain frequently employed in the food industry to facilitate the fermentation process of different vegetables and fruits [8]. Presently, this fungus is regarded as a rare emerging pathogen. It was originally documented as a cause of fungemia in 1994 [9] and has subsequently been increasingly reported as the cause of human infections [3,10,11].
This yeast has caused concerns regarding its potential impact on public health and its significance in epidemiology worldwide, primarily due to its high mortality rate (40% to 50%) [10,12,13], its propensity to cause outbreaks within healthcare facilities, causing infections of various anatomical sites [12,14,15], and its resistance to certain antifungal medications, such as echinocandins and fluconazole, as evidenced by high Minimum Inhibitory Concentrations (MICs) [10].
The accurate geographic distribution of this microorganism as a causative agent of human infections remains uncertain. This yeast can be mistakenly identified phenotypically as Candida tropicalis, C. albicans, and C. glabrata using conventional laboratory tests, which implies that the number of cases may have been underestimated [12,13]. Moreover, there is a substantial lack of data regarding the virulence and drug resistance of this emerging pathogen, requiring the gathering of additional information to enhance our comprehension. To improve our understanding of the epidemiology and diagnosis of this emerging pathogen, this study aimed to provide a comprehensive description of three K. ohmeri isolates responsible for fungemia cases in Honduras as part of an ongoing epidemiological surveillance program. As far as we know, this is the first report of K. ohmeri causing fungemia in Honduras.
2. Materials and Methods
Collection of isolates
As a component of a surveillance program in Honduras, tertiary care laboratories under the National Health System were instructed to send all Candida isolates from sterile anatomical sites to the National Surveillance Laboratory (NSL) of the Ministry of Health. The purpose was to verify the identification of the Candida species and assess their sensitivity to antifungal drugs.
Identification of isolates by PCR-RFLP of the ribosomal ITS region
The yeasts were cultured individually in 4 mL of YPD broth at 30°C for 24 h with constant shaking at 200 rpm. The DNA of all isolates was extracted according to a previously published protocol [16]. The amplifications were performed under the following conditions in a volume of 50 μL: 25 of PCR Master Mix (Promega Corp. Madison, WI, USA), 1 μL of each primer, ITS1 and ITS4: 5′-TCC GTA GGT GAA CCT GCG G-3 / 5′-TCC TCC GCT TAT TGA TAT GC- 3′ and 1 μL of DNA (40 ng/μL). Reactions were carried out with an initial denaturation step at 95°C for 5 min, 37 cycles of 95°C for 30 s, 56°C for 30 s, and 72°C for 30 s, with a final extension at 72°C for 5 min. Amplicons were separated by 1.5% agarose gel electrophoresis with ethidium bromide. Four microliters of each PCR product were digested for 2 h at 37 °C with 2 μL buffer, 0.2 μL of acetylated BSA (10 μg/μL), and 0.5 μL of the restriction enzyme MspI (10 U/μL) (Promega Corp. Madison, WI, USA). The restriction fragments were analyzed on a 2% agarose gel.
Identification of Candida auris
Isolates that produced bands of around 400 base pairs using primers ITS1 and ITS4, which is in line with the predicted pattern for the C. haemulonii complex and its related species C. auris [17], were chosen for a targeted PCR assay based on the GPI gene for the diagnosis of C. auris [18]. A mixture was prepared with 12.5 µL of 2× Master Mix (Promega Corp. Madison, WI, USA), 9 µL of nuclease-free water, 1.5 µL of DNA (40 ng/µL) and 1 µL of each 10 µM primer: 03410_F (5′-GCC GCT AGA TTG ATC ACC GT-3′ and 03410_R:5′-TAG GTG TGG GTA CCC TTG GT-3′). The PCR program consisted of one cycle at 94 °C for 3 min, 35 cycles at 94 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s, and one cycle at 72 °C for 3 min. A band size of 137 bp indicated C. auris and no amplification product was expected for the rest of the Candida species. Positive and negative controls were included in each PCR reaction.
Sequence analysis and construction of cladograms
Isolates that presented PCR products of around 400 bp for the ITS marker, but failed to amplify the GPI gene, which is critical to confirming the presence of C. auris, were selected for sequencing. The PCR products were purified and sequenced in both directions according to the protocols of the Psomagen company (https://lims.psomagen.com/). The quality of the sequences was analyzed with Geneious prime® 2023.1.2 software and international databases contained in NCBI were queried to confirm the identity of the sequences using the BLAST tool. The sequences were compared to sequences found in GenBank, and the result with the highest percentage of similarity was recorded as the most likely identification for each isolate. The sequences were submitted to GenBank, and accession numbers were assigned to each strain.
More than 110 homologous sequences from the ITS region of K. ohmeri isolated from 17 countries in 4 continents were downloaded and aligned with the sequences obtained in this study. The ClustalW tool of the Geneious software was used to align sequences with the same length. The Tamura–Nei genetic distance model and the Neighbor-Joining method, with a bootstrap of 1,000 replicates, were used to construct a cladogram. Homologous sequences from Candida albicans and Schizosaccharomyces japonicus were also downloaded and included in the alignment as an outgroup.
Phenotypic identification
Microbiological identification and chromogenic culture media
The isolates were cultured on Sabouraud dextrose agar (SDA, Becton Dickinson, NJ, USA), at 30 °C for 48 h, and subsequently cultured in three chromogenic media for the identification of yeasts: CHROMagar (CHROMagar CandidaTM, France), CHROMagarTM (Becton, Dickinson and Company, NJ, USA), ChromaticTM (Candida, Liofilchem®, Teramo, Italy). These media were incubated at 37 °C for 48 h and evaluated based on the color of the colonies according to the manufacturers’ instructions. Furthermore, all isolates were identified by the systems BD Phoenix BD PhoenixTM (Becton, Dickinson and Company, NJ, USA), and VITEK® 2 (bioMerieux, Craponne, Francia), following the manufacturers’ instructions.
Also, morphological characteristics such as chlamydospore formation were evaluated in corn flour agar and germ tube production or early filamentation following the protocols proposed by Giusiano et al., 2016 [19]. Both tests serve for the presumptive identification of Candida albicans complex. Candida albicans ATCC 10231 and C. parapsilosis ATCC 22019 were used as positive and negative controls, respectively.
Antifungal sensitivity testing
The antifungal sensitivity profile of the isolates was determined using the SensititreTM Yeast-OneTM system (Thermo Fisher Scientific, Waltham, MA, USA). SensititreTM Yeast-OneTM is a commercial microdilution method used to determine the Minimum Inhibitory Concentrations (MICs) of yeasts and some filamentous fungi to the following antifungals: anidulafungin (0.015 – 8 mg/L), micafungin (0.008 – 8 mg/L), caspofungin (0.008 – 8 mg/L), fluconazole (0.12 – 256 mg/L), posaconazole (0.008 – 8 mg/L), voriconazole (0.008 – 8 mg/L), itraconazole (0.015 – 16 mg/L), amphotericin B (0.12 – 8 mg/L) and 5-fluorocytosine (0.06 – 64 mg/L). Antifungal susceptibility testing was performed following the manufacturer’s recommendations. Briefly, all isolates were cultured in SDA at 37°C for 24 h. A suspension was prepared by mixing isolated colonies with sterile distilled water until it reached a density equivalent to the McFarland Nº 0.5 standard. The accuracy of the density was confirmed using a nephelometer. C. parapsilosis ATCC 22019 was used as a reference strain for quality control. The MICs were evaluated after 24 h of incubation at 37 °C. The yeast’s growth was demonstrated by a transition in color from blue (negative, indicating absence of growth) to red (positive, indicating growth).
Assays for hydrolytic enzyme activity of Kodamaea ohmeri
The phospholipase and hemolytic activities of the K. ohmeri isolates were assessed using the established techniques described in earlier publications [20]. Proteinase activity was assessed using the determination of caseinase and gelatinase enzymes, as well as the hydrolysis of bovine serum albumin (BSA) [20]. The enzymatic activity was evaluated by the enzymatic activity coefficient (Pz). Pz was calculated by dividing the diameter of the colony (A) by the sum of the colony diameter and the hydrolysis/precipitation zone (B), [Pz = A/B] [21]. The isolates were classified according to the Pz value into four categories: Pz = 1.0 no enzymatic activity, Pz = 0.99 to 0.90 weak enzymatic activity, Pz = 0.89 to 0.70 moderate activity, and Pz ≤ to 0.69 strong activity. The Pz values were calculated by taking the average of two independent tests, with each experiment being conducted twice.
Assessment of Kodamaea ohmeri’s ability to produce biofilms
Biofilm formation was determined according to the protocol previously published by Saiprom et al., 2023 [22] with modifications. Briefly, the isolates were cultured in a YPD medium overnight at 37 °C. Afterwards, a yeast suspension was prepared in YPD medium with a density corresponding to the McFarland Nº 0.5 standard. A volume of 100 μL of the suspension was distributed into a 96-well flat-bottom plate and underwent incubation at 30 °C for 48 hours. The wells were rinsed thrice with 1× PBS to eliminate cells that were not adhered. The empty wells were allowed to dry for 45 min, and 200 μL of 0.1% crystal violet was added to each well and incubated for 45 min at room temperature. After that, the crystal violet was eliminated, and the plate was left to dry for 10 minutes at room temperature. Subsequently, the wells were gently rinsed twice with 200 μL of sterile distilled water. A volume of 300 μL of 100% ethanol was added to remove color from the biofilm, and the plate was kept at room temperature for 45 minutes. A 150 μL volume of eluted crystal violet was transferred to a new 96-well plate and the optical density (OD) was measured at 590 nm in a spectrophotometer (Thermo ScientificTM Genesys 20). Sterile YPD was used as a negative control. Two independent experiments were performed for each isolate and each experiment was repeated three times. The biomass of each isolate was calculated as the average OD value based on the two independent experiments. The production capacity of biofilm was calculated by averaging the OD of each sample (ODs). The OD of the negative control (ODnc) was also determined. The strains were classified using the following criteria: Non-producer (ODs ≤ ODnc); weak producer (ODnc < ODs ≤ 2× ODnc); moderate producer (2× ODnc < ODs ≤ 4× ODnc), and strong producer (ODs > 4× ODnc) [23].
Invasion and adherence assays
The invasion assays were carried out according to the methodology proposed by Faria-Gonçalves et al., 2022 and Saiprom et al., 2023 [22,24] with modifications. Briefly, RAW 264.7 macrophages were reactivated in culture dishes with Dulbecco’s modified Eagle’s medium (DMEM) (Merck, Darmstadt, Germany) supplemented with 5% fetal bovine serum (FBS) (GIBCOTM, Fisher Scientific Inc., Madrid, Spain). The cultured cells were placed in an incubator at 37 °C and 5% CO2 until they reached 80% confluency. Subsequently, in six-well polystyrene plates with coverslips, a cell suspension equal to 50,000 cells/mL was prepared in 2 mL of DMEM supplemented with 5% FBS and antibiotics (Penicillin-Streptomycin). Plates were incubated overnight at 37°C in 5% CO2. RAW 264.7 cell monolayers were washed with sterile 1× PBS, and after washing, 2 mL of fresh DMEM supplemented with 5% FBS and antibiotics was added to each well. From a 48-hour culture of the three isolates under study, a suspension was made in DMEM containing 5% FBS with a density equivalent to the McFarland Nº 0.5 standard (approximately 1 × 106 CFU/mL). The RAW 264.7 cells were infected with each K. ohmeri isolates at a multiplicity of infection (MOI) of 5 and incubated for 4 h at 37°C in 5% CO2. Uninfected cells were used as a negative control. After 4 h, the cultures were washed three times with 1× PBS to remove planktonic yeasts. The cells were fixed with methanol for 15 minutes, stained with Giemsa for 15 minutes and subsequently washed with sterile water. The coverslips were removed from each well and fixed on a slide for visualization by bright-field microscopy at 100× objectives to observe their morphology and detect the invasive yeasts. The cells were counted in 10 microscopic fields (approximately five cells per field), the number of macrophages invaded with yeast was determined and the percentage of invasion was obtained. Each strain was evaluated in triplicate.
Adherence tests were performed following previously published protocols with modifications [22,24]. For the adhesion assays, the HeLa cell line was used, which was cultured on plates with DMEM medium supplemented with 5% FBS, which was incubated at 37 °C in 5% CO2 until 80% confluency was observed. Subsequently, in six-well polystyrene plates with coverslips, a cell suspension of 50,000 cells/mL was prepared in 2 mL of DMEM supplemented with 5% FBS and antibiotic (Penicillin-Streptomycin). Plates were incubated for 24 hours at 37°C in 5% CO2. HeLa cell monolayers were washed with sterile 1× PBS, and after washing, 2 mL of fresh DMEM was added to each well. From a 48-hour culture of the yeast under study, and in DMEM culture medium, a cell suspension equivalent to the McFarland standard Nº 0.5 (approximately 1×106 CFU/mL) was made.
HeLa cells were infected with each K. ohmeri isolate at a multiplicity of infection (MOI) of 10 and incubated for 2 h at 37°C in 5% CO2. Uninfected cells were implemented as a negative control. After 4 h, cultures were washed three times with 1× PBS to remove non-adherent yeast, and adhered cells were fixed with methanol for 15 min. To visualize the adhered cells, Giemsa staining was performed, followed by three washes with 1× PBS and finally, they were observed under the microscope using the 40× and 100× objectives. The HeLa cells present per field were counted in 10 fields (approximately 200 HeLa per field), the yeasts adhered to the HeLa cells and the number of cells with adherence was quantified. The results were expressed as the percentage of cells with adhered yeasts and the number of adhered yeasts per HeLa cell. Each isolate was analyzed in independent experiments in triplicate.
For invasion and adherence assays standard deviation (SD) was calculated from counting a total of 10 fields per slide. A statistical analysis was performed using one-tailed ANOVA and Tukey’s multiple comparisons test.
3. Results
A total of seventy-four yeast isolates, which were phenotypically identified as Candida spp and obtained from blood cultures, were submitted to the Laboratory of National Health Surveillance (LNS) of the Ministry of Health for molecular identification and assessment of their sensitivity to antifungal drugs. Three isolates, identified as C-29, C-35, and C-38, were initially believed to be Candida auris. These isolates were obtained from two public hospitals in San Pedro Sula (n=2) and Tegucigalpa (n=1).
The molecular identification of the three yeast isolates was conducted using the PCR-RFLP technique targeting the ribosomal ITS region and the MspI enzyme. This approach is commonly employed for the identification of Candida species and complexes [25,26,27,28]. The PCR products from the three isolates had a length of around 400 base pairs, and no cleavage occurred when the products underwent digestion with the MspI enzyme (Supplementary material). The amplified size obtained using the ITS1 and ITS4 primers matched the predicted size for the species grouping in the C. haemulonii complex (C. haemulonii s.s., C. haemulonii var. vulnera, C. duobushaemulonii, C. pseudohaemulonii) and its related species C. auris [17]. Consequently, a targeted PCR was conducted to detect C. auris using the GPI gene, but no amplification result was detected in the three isolates (Supplementary material). Subsequently, the ITS region was amplified and sequenced [17], and the resulting sequences of the three isolates were edited and analyzed using the NCBI BLAST tool, and all isolates were identified as Kodamaea ohmeri. The sequences obtained were deposited in GenBank under accession numbers OR791936-OR791938. The nucleotide sequencing of the three isolates showed no discrepancies. A phylogenetic tree was created using the ITS sequences of this study and comparing them with sequences of K. ohmeri isolated from different geographical locations. The three sequences obtained in this study showed a 100% similarity with no single nucleotide polymorphisms (SNPs) (Figure 1). A phylogenetic tree was constructed by aligning the ITS sequences obtained from this study with more than 110 sequences of K. ohmeri isolated from various geographical areas. The K. ohmeri isolates from Honduras exhibited a high degree of similarity, forming a cohesive cluster with most sequences. No clear phylogenetic clustering was seen based on the geographic origin of the isolates (Figure 1).
In this work, we assessed three isolates of K. ohmeri using phenotypic identification methods. This was done since there is insufficient information available on clinical isolates of K. ohmeri and their identification profile in routine laboratory conditions. The three isolates cultivated in CHROMagarTM Candida, CHROMagarTM, ChromaticTM Candida, produced colonies with an iridescent-pink appearance within the initial 18 to 24 hours, which then became green after 48 hours of incubation in both media (Figure 2).
Similarly, the BD PhoenixTM and VITEK® 2 automated identification systems were used to analyze these isolates. The VITEK® 2 system successfully identified the isolates as K. ohmeri, with identity percentages of 96% or higher. However, the BD PhoenixTM system failed to identify two isolates and one of them was mistakenly identified as C. albicans with an identity percentage of 95%.
Phenotypic tests were conducted to examine the morphology of the isolates. Specifically, the creation of chlamydospores in corn flour agar and the development of germ tubes were assessed. However, no similarity to C. albicans was seen in either of these tests (i.e., no chlamydospores or germ tubes were produced). The MICs of the three isolates for nine antifungals are shown in Table 1. The MICs for voriconazole, posaconazole, and itraconazole varied between 0.03 to 0.12 mg/L. However, all isolates showed a MIC of 4 mg/L for fluconazole. The MIC range for the echinocandin family ranged from 0.12 to 2 mg/L, with the highest value recorded in an isolate for caspofungin. The MIC range for amphotericin B was between 0.25 and 0.5 mg/L, whereas all isolates showed a MIC of ≤ 0.06 mg/L for 5-fluorocytosine.
The three isolates exhibited biofilm generation and extracellular enzyme synthesis after 48 hours of incubation at 37 °C. The absorbance values ranged from 0.727 to 1.245. Based on the measured absorbances, all isolates demonstrated a high level of biofilm production. In addition, all isolates of K. ohmeri had a Pz value ≤ 0.69 for the synthesis of extracellular enzymes, indicating significant hemolytic, phospholipase, and protease (gelatin, casein, BSA) activity. The absorbances and Pz values are shown in Table 2.
In addition, the capacity of isolates to adhere to and invade cells was assessed using the HeLa and RAW 264.7 cell monolayer infection models. All isolates exhibited adhesion to HeLa cells, whereas C-29 and C-38 demonstrated the highest level of adherence. There was a statistically significant difference in the number of HeLa cells with yeast in strain C-29 compared to the other two (p < 0.05). Each isolate had a minimum of two pseudohyphae per HeLa cell. Similarly, the rate of invasion in RAW 264.7 cells ranged from 30% to 35%. Figure 3 displays the outcomes of adhesion in HeLa cells and invasion in RAW 264.7. All isolates of K. ohmeri exhibited a significant ability to form filaments in both experiments.
4. Discussion
Fungal infections pose a significant public health concern. Approximately 1.5 million individuals perish each year due to fungal infections, while a much larger number have serious fungal diseases [29]. The prospect of fungal infections appears unfavorable in the foreseeable future due to several factors. These include the rising number of individuals vulnerable to fungal infections, the growing prevalence of resistance to various antifungal medications, limited accessibility to treatment options, and the restricted availability of methods for detecting and identifying fungi. Additionally, there is a concerning emergence of new human fungal pathogens that possess inherent resistance to currently available antifungal drugs [4,5,7,30,31].
The dominant theory regarding the emergence of new fungal infections attributes it to the influence of climate change on microorganisms in the environment [7,32,33,34]. For instance, Candida auris has been proposed as the first human pathogenic fungus that emerged as a result of climate change [33]. Historically, only a limited number of fungal species were capable of prospering at temperatures over 37 °C, hence impeding their capacity to infect and establish within mammals [4]. However, during the last century, global warming has been promoting the emergence of new fungal lineages that can tolerate high temperatures. This enables them to surpass the temperature limits of mammals, known as the "thermal restriction zone" [7]. Likewise, permafrost thawing could release uncharacterized microbes with harmful potential [34]. Similarly, the destruction of high biodiversity areas, such as tropical forests and untouched jungles, increases the likelihood of contacting novel microbes that could eventually develop into harmful illnesses for both humans and animals [32].
As a result, environmental and human-induced factors are greatly impacting the evolution, selection, and adaptation of previously harmless fungal species, which are now posing a growing threat to the health of humans and animals [35]. This is the case of Candida viswanathii, C. auris, C. blankii, C. palmioleophila, C. vulturna, C. massiliensis, Trichophyton indotineae, Pseudogymnoascus destructans, Batrachochytrium dendrobatidis and Kodamaea ohmeri, all of them environmental fungi that over time have been successfully adapted to animals or humans [12,18,31,36,37,38,39].
The genus Kodamaea consists of six species: K. anthophila, K. kakaduensis, K. laetipori, K. nitidulidarum, K. samutsakhonensis, and K. ohmeri. However, only K. ohmeri has been proven to have clinical significance [8]. K. ohmeri is commonly associated with insects and flowers [8,40], however, it is now considered a rare emerging pathogen. In recent years, its significance has increased due to the decreased effectiveness of certain antifungal medications like echinocandins and azoles. Additionally, it is associated with a mortality rate of approximately 50% [12,13].
Since its initial report of causing fungemia in a 64-year-old male patient in 1994, cases of human infections attributed to this yeast have been on the rise in multiple nations across Asia, America, Europe, and Africa [10,12,41]. While China and India, report the largest number of cases, K. ohmeri has also been documented in the Americas: USA (n=7), Brazil (n=3), Colombia (n=1), and Mexico (n=1) [12]. A national surveillance program in Honduras has detected three isolates of K. ohmeri in the cities of San Pedro Sula and Tegucigalpa in response to recent reports of emergent yeasts worldwide. Initially, the three isolates were misidentified as Candida albicans due to their morphological traits. This finding is highly significant in the local context as it emphasizes the importance of implementing an ongoing surveillance program that utilizes molecular tests and/or mass spectrometry to accurately identify filamentous fungi and yeasts.
K. ohmeri isolates were cultivated on three commercially available chromogenic agars specifically formulated for yeast species identification based on colony color. After 48 hours, the three isolates exhibited green colonies, which might have led to a misidentification as C. albicans. Agrawal et al., 2014 described different morphotypes of K. ohmeri in HiCromeTM Candida, where the colors of the colonies showed pink during the first 24 h, and then turned green or blue at 48 h until evolving to metallic blue at 72 hours [42]. In their 2007 study, Lee et al, reported the growth of pink and green colonies in CHROMagar CandidaTM, which transitioned into blue colonies after 72 hours [43]. Similar results were reported by Biswa et al., 2015, Yu et al., 2019 and Mtibaa et al., 2019 in this medium [41,44,45]. In this sense, it seems that chromogenic agars play an important role in the identification of the most important species of the genus Candida (C. albicans, C. tropicalis, C. parapsilosis, and C. krusei), but do not represent a viable approach for the identification of emerging yeasts.
K. ohmeri is frequently misdiagnosed as C. tropicalis, C. glabrata, or C. albicans during routine testing [13,43,46], which would have an impact on under-reporting of clinical cases due to this species. The study also utilized traditional phenotypic assays for assessing K. ohmeri isolates. None of the three strains exhibited the ability to generate either a germ tube or chlamydoconidia on corn flour agar. Consequently, these techniques can differentiate between species within the C. albicans complex, but they lack sufficiency in identifying additional Candida species or those belonging to other genera of yeast. Regarding automated identification methods, VITEK® 2 demonstrated successful identification of the three isolates. This is not surprising, as this system can identify up to 50 different yeasts, including K. ohmeri. However, it has been observed that in this identification system, certain isolates of Candida palmioleophila can be mistakenly identified as K. ohmeri [36]. The BD Phoenix BD PhoenixTM yeast identification system, on the other hand, could not correctly identify any isolate. This is in line with what the manufacturer states, since this equipment can identify up to 64 species of yeasts, but not the Kodamaea species.
Sequencing of ITS regions has been recognized as a reference method for the characterization of yeasts, including K. ohmeri [10,12]. After comparing the sequences obtained in this study with over 110 ITS sequences of clinical and environmental isolates of K. ohmeri from different regions worldwide, it became clear that they formed a monophyletic group, without any cluster formation. Our findings, along with the existing genomic data, suggest that the genetic diversity of this emerging species is restricted. In contrast, one study used the MLST approach with six K. ohmeri isolates causing infections in Bangladesh, and the authors reported five different allelic profiles [47]. The authors present a dendrogram displaying eight distinct clusters. This dendrogram was created using ITS sequences from their six isolates, as well as homologous sequences from various geographical locations. However, it is important to note that the sequences labeled under accession numbers OQ606981-OQ606985 do not exhibit any variations among themselves. Consequently, these findings contradict our results.
Concerning the susceptibility profiles of the antifungal drugs, the isolates exhibited low MICs for amphotericin B, triazoles, 5-Flucytosine, anidulafungin, and micafungin. However, the MICs for caspofungin were found to be high. These results are consistent with the evidence collected in two recent reviews [10,12]. While there are no accepted epidemiological cutoff values or clinical breakpoints for K. ohmeri according to EUCAST [48] or CLSI [49], it is crucial to determine the sensitivity profile to antifungals in emerging yeasts. This information assists the clinician in determining the most suitable course of therapy. Furthermore, the calculation of MICs has previously contributed to the identification of novel or emerging pathogens [50,51,52]. It is imperative to allocate resources toward incorporating antifungal susceptibility testing into routine laboratories, especially in low- and middle-income countries where access to these tests is limited [12]. We hope that our data advances the epidemiological understanding of the susceptibility profile of K. ohmeri.
Yeast pathogenesis seems to be influenced significantly by the production of biofilms and the presence of extracellular hydrolytic enzymes [53,54]. The isolates we tested exhibited strong biofilm production, which aligns with the findings of Giacabino et al (2015). In their study, they investigated yeasts linked with fungal peritonitis and identified one of the isolates as K. ohmeri. This strain exhibited a high biofilm production capability, ranking as the second most proficient isolate in biofilm production [55]. Similarly, our findings align with the data presented by Maciel et al., 2019, who examined potentially harmful yeasts obtained from beaches in Brazil. These authors assessed five strains of K. ohmeri, which exhibited the maximum biofilm production among all the strains examined [56]. Several fungi of the phylum Ascomycota have been shown to form biofilms. Among yeasts, Candida species have been extensively investigated as one of the main models [57,58]. Biofilms affect the long-term survival of microbes on non-living surfaces and tissues. They provide protection against host immune cells and are strongly linked to resistance to antifungal treatments [59]. Further research is required to establish the correlation between the development of K. ohmeri biofilm and its resistance to antifungal agents, as well as its ability to persist on non-living surfaces.
The isolates had a high potential for producing extracellular enzymes, specifically proteases, phospholipase, and hemolysin. Hydrolytic enzymes have an important role in adhering to and infiltrating host cells [20,22,53]. Proteases facilitate the entry into cells by breaking down proteins like albumin, collagen, and mucin. They also contribute to the breakdown of antibodies, complement factors, and cytokines [20,54]. Phospholipases alter the cell membrane, whereas hemolysins assist in the destruction of cells and the uptake of iron [20,53,54].
This study also aimed to assess the adhesive potential of three K. ohmeri isolates to the HeLa cell line. This methodology has previously been employed to demonstrate the capacity of several Candida species to adhere [24,60,61,62]. For yeasts to successfully colonize and infiltrate tissues, they need to adhere themselves to the epithelium. This attachment is facilitated by adhesins, integrins, and cadherins, which are located on the surface of the fungal cell wall [60,63]. After adhering, yeasts can penetrate host cells either through induced endocytosis or active penetration [60,64]. Our results demonstrate a significant index of K. ohmeri’s adherence to this specific cell line. Furthermore, all three isolates exhibited notable morphological transformations from yeast to pseudohyphae, indicating that these modifications may facilitate host invasion through active penetration. Further research is required to investigate the adhesion processes of K. ohmeri in more depth.
In addition, we demonstrated the effectiveness of the RAW 264.7 macrophage line in effectively engulfing all three strains of K. ohmeri. Macrophages are cells specialized in the detection, phagocytosis, and destruction of fungal propagules [65,66], that are recognized through receptors that interact with molecular patterns associated with pathogens, which are generally arranged along the cell surface of fungal cells [65]. Mannans, galactomannans, glucosylceramides, mannoproteins, chitin, melanin, and β-glucans are key molecules that interact with and activate the immune system [65,66]. Our results highlight the need to elucidate the molecular mechanisms by which immune cells interact with K. ohmeri. However, this study solely assessed the extent of cellular internalization without examining the lethal activity of macrophages. Therefore, further investigations are required to ascertain and comprehend the survival rate and the processes via which yeasts interact with macrophage phagosomes. This information would enhance our comprehension of potential strategies for escape and immune recognition, while also aiding in the identification of novel treatment targets for this emerging pathogen.
5. Conclusions
To our knowledge, this is the first report of K. ohmeri in Honduras causing human infection. Our finding highlights the importance of strengthening the capacities of mycology reference laboratories and epidemiological surveillance in the country. On the other hand, our results demonstrate that the identification of emerging pathogens such as K. ohmeri is a challenge for clinical laboratories and that the implementation of tools based on molecular biology and/or mass spectrometry is urgently needed in the main hospital centers of the main cities of the country since some pathogens could be being misdiagnosed. The implementation of these tools would contribute to decision-making for the containment of outbreaks and the spread of emerging fungal pathogens. Furthermore, here we present important information about some characteristics that would be mediating the virulence of K. ohmeri.
6. Limitations
The patients’ medical records, with key data on risk factors, underlying diseases, and treatment procedures, were inaccessible. SensititreTM Yeast-OneTM system is a strongly recommended approach for assessing the susceptibility of yeasts, especially Candida species, to antifungal drugs [67,68,69], however, Zhou et al. (2019) found that for uncommon yeasts like K. ohmeri, the MIC values may vary due to the absence of standardized tests. Therefore, it is advisable to employ the broth microdilution method, which offers greater reliability and precision [12]. Due to the unavailability of broth microdilution in Honduras, it was not feasible to estimate the minimum inhibitory concentrations (MICs) using this method.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
B.O, M.B.M and P.M.F conceptualized the study; R.L, C.M, I.L.A performed the mycological diagnosis; B.O, F.P, F.C, C.G, L.R, K.A, M.B.M, P.M.F and D.V performed the experiments; B.O., K.A., M.B.M, and G.F. organized and cured the data; writing and original draft preparation, B.O, K.A, and G.F.; All the authors contributed with writing, review, and editing the manuscript; supervision, project administration, and funding acquisition, G.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. The experiments were conducted with the resources provided by the Genetic Research Center of the National University of Honduras and Laboratorio de Biología Celular y Molecular of Universidad de Sonora, Campus Caborca.
Institutional Review Board Statement
Regarding ethical considerations, the study was conducted on anonymous biological samples. Furthermore, this research did not involve any personal data that could directly or indirectly identify a specific individual. Therefore, a statement of consent from the ethics committee of the hospital or university was not necessary. Clinical samples were collected according to ethical standards and due to the retrospective nature of the study, the consent form was not applicable to the study.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Choi, J.; Kim, S.-H. A genome Tree of Life for the Fungi kingdom. Proceedings of the National Academy of Sciences 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [PubMed]
- Wickes, B.L.; Wiederhold, N.P. Molecular diagnostics in medical mycology. Nature communications 2018, 9, 5135. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P. Emerging fungal infections: New species, new names, and antifungal resistance. Clinical Chemistry 2022, 68, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Nnadi, N.E.; Carter, D.A. Climate change and the emergence of fungal pathogens. PLoS Pathogens 2021, 17, e1009503. [Google Scholar] [CrossRef] [PubMed]
- van Rhijn, N.; Bromley, M. The consequences of our changing environment on life threatening and debilitating fungal diseases in humans. Journal of Fungi 2021, 7, 367. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.W.; Rossato, L.; Goldman, G.H.; Santos, D.A. Fungicide effects on human fungal pathogens: Cross-resistance to medical drugs and beyond. PLoS Pathogens 2021, 17, e1010073. [Google Scholar] [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the emergence of Candida auris: climate change, azoles, swamps, and birds. MBio 2019, 10. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The yeasts: a taxonomic study; Elsevier: 2011.
- Jin, S.; Jin, Z. A case of septicemia caused by Kodamaea ohmeri. Chin J Clin Lab Sci 1994, 3, 167. [Google Scholar]
- Ioannou, P.; Papakitsou, I. Kodamaea ohmeri infections in humans: A systematic review. Mycoses 2020, 63, 636–643. [Google Scholar] [CrossRef]
- Bergman, M.; Gagnon, D.; Doern, G.V. Pichia ohmeri fungemia. Diagnostic microbiology and infectious disease 1998, 30, 229–231. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.; Kudinha, T.; Xu, Y.; Liu, Z. Kodamaea ohmeri as an emerging human pathogen: a review and update. Frontiers in Microbiology 2021, 12, 736582. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Rudramurthy, S.; Kale, P.; Hariprasath, P.; Dhaliwal, M.; Singhi, S.; Rao, K. Epidemiological study of a large cluster of fungaemia cases due to Kodamaea ohmeri in an Indian tertiary care centre. Clinical Microbiology and Infection 2014, 20, O83–O89. [Google Scholar] [CrossRef] [PubMed]
- Otag, F.; Kuyucu, N.; Erturan, Z.; Sen, S.; Emekdas, G.; Sugita, T. An outbreak of Pichia ohmeri infection in the paediatric intensive care unit: case reports and review of the literature. Mycoses 2005, 48, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, J.; Dong, L.; Mai, J.; Zhang, L.; Zhu, J. Clinical features and homological analysis of Pichia ohmeri-caused hospital-acquired fungemia in premature infants. Zhonghua yi xue za zhi 2013, 93, 285–288. [Google Scholar] [PubMed]
- Ortiz, B.; Aguilar, K.; Galindo, C.; Molina, L.; Fontecha, G. Candida species isolated from clinical samples in a tertiary hospital in Honduras: Where is Candida auris? Current Medical Mycology 2022, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Fontecha, G.; Montes, K.; Ortiz, B.; Galindo, C.; Braham, S. Identification of cryptic species of four Candida complexes in a culture collection. Journal of Fungi 2019, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, M.; Bartolomé Álvarez, J.; Lockhart, S.R.; Valentín, E.; Ruiz-Gaitán, A.C.; Eraso, E.; de Groot, P.W.J. Identification of Candida auris and related species by multiplex PCR based on unique GPI protein-encoding genes. Mycoses 2021, 64, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Giusiano, G.; Piontelli, E. Hongos oportunistas levaduriformes y filamentosos comunes en clínica. 2016.
- Neji, S.; Hadrich, I.; Trabelsi, H.; Abbes, S.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Virulence factors, antifungal susceptibility and molecular mechanisms of azole resistance among Candida parapsilosis complex isolates recovered from clinical specimens. Journal of biomedical science 2017, 24, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Price, M.F.; Wilkinson, I.D.; Gentry, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia: Journal of Medical and Veterinary Mycology 1982, 20, 7–14. [Google Scholar] [CrossRef]
- Saiprom, N.; Wongsuk, T.; Oonanant, W.; Sukphopetch, P.; Chantratita, N.; Boonsilp, S. Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand. Journal of Fungi 2023, 9, 353. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Faria-Gonçalves, P.; Oliveira, A.S.; Gaspar, C.; Rodrigues, L.; Palmeira-de-Oliveira, R.; Martinez-de-Oliveira, J.; Gonçalves, T.; Palmeira-de-Oliveira, A.; Rolo, J. Vulvovaginal Candida albicans Clinical Isolates’ Resistance to Phagocytosis In-Vitro. Life 2022, 12, 838. [Google Scholar] [CrossRef]
- Farooq, H.; Monowar, T.; Chinni, S.V.; Latt, S.S.; Zainol, N.H.; Sabesan, G.S. Epidemiology and molecular identification of mixed yeast isolates in Malaysia: A way forward. Current medical mycology 2022, 8, 35. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mirhendi, H.; Rezaei-Matehkolaei, A.; Ghahri, M.; Shidfar, M.R.; Jalalizand, N.; Makimura, K. Molecular identification and distribution profile of Candida species isolated from Iranian patients. Medical Mycology 2013, 51, 657–663. [Google Scholar] [CrossRef]
- Kord, M.; Elmimoghaddam, A.; Hashemi, S.J.; Reziae, S.; Daie Ghazvini, R.; Salehi, M.; Abdollahi, A.; Ahmadi, A.; Getso, M.I.; Boekhout, T.; et al. Comparison of PCR-RFLP with 21-plex PCR and rDNA Sequencing for Identification of Clinical Yeast Isolates. Mycopathologia 2021, 186, 213–220. [Google Scholar] [CrossRef]
- Montes, K.; Ortiz, B.; Galindo, C.; Figueroa, I.; Braham, S.; Fontecha, G. Identification of Candida species from clinical samples in a Honduran tertiary hospital. Pathogens 2019, 8, 237. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Frontiers in microbiology 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A. Fungal diseases in the 21st century: the near and far horizons. Pathogens & immunity 2018, 3, 183. [Google Scholar]
- Johnson, E.M. Rare and emerging Candida species. Current Fungal Infection Reports 2009, 3, 152–159. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Chies, J.A.B. Candida auris emergence as a consequence of climate change: Impacts on Americas and the need to contain greenhouse gas emissions. The Lancet Regional Health–Americas 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Akinbobola, A.B.; Kean, R.; Hanifi, S.M.A.; Quilliam, R.S. Environmental reservoirs of the drug-resistant pathogenic yeast Candida auris. PLoS pathogens 2023, 19, e1011268. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Trubl, G.; Taş, N.; Jansson, J.K. Permafrost as a potential pathogen reservoir. One Earth 2022, 5, 351–360. [Google Scholar] [CrossRef]
- Morales-López, S.E.; Garcia-Effron, G. Infections due to rare Cryptococcus species. A literature review. Journal of fungi 2021, 7, 279. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.L.d.; Negri, M.; Miranda, R.P.R.d.; Corrêa-Moreira, D.; Pinto, T.C.A.; Ramos, L.d.S.; Ferreira, D.G.; Salomão, B.; Fumian, T.M.; Mannarino, C.F. Candida palmioleophila: A New Emerging Threat in Brazil? Journal of Fungi 2023, 9, 770. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Bing, J.; Xu, X.; Zheng, Q.; Hu, T.; Hao, Y.; Li, S.; Nobile, C.J.; Zhan, P.; Huang, G. Candida vulturna outbreak caused by cluster of multidrug-resistant strains, China. Emerging Infectious Diseases 2023, 29, 1425. [Google Scholar] [CrossRef] [PubMed]
- Kabtani, J.; Boulanouar, F.; Militello, M.; Cassagne, C.; Ranque, S. Candida massiliensis sp. nov. Isolated from a Clinical Sample. Mycopathologia 2023, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kano, R.; Kimura, U.; Kakurai, M.; Hiruma, J.; Kamata, H.; Suga, Y.; Harada, K. Trichophyton indotineae sp. nov.: a new highly terbinafine-resistant anthropophilic dermatophyte species. Mycopathologia 2020, 185, 947–958. [Google Scholar] [CrossRef]
- Rosa, C.A.; Lachance, M.-A.; Starmer, W.T.; Barker, J.S.F.; Bowles, J.M.; Schlag-Edler, B. Kodamaea nitidulidarum, Candida restingae and Kodamaea anthophila, three new related yeast species from ephemeral flowers. International Journal of Systematic and Evolutionary Microbiology 1999, 49, 309–318. [Google Scholar] [CrossRef]
- Mtibaa, L.; Souid, H.; Jemli, B.; Hajjej, Z.; Halweni, C. Kodamaea ohmeri. An Emerging Yeast in Tunisia: First Identification in Three Case Reports and Literature Review. J Med Microb Diagn 2019, 8, 1000299. [Google Scholar]
- Agrawal, V.; Bhagwat, A.; Vishalakshi, V.; Gode, V.; Sawant, C. Exploring the potential of chromogenic medium for the identification of medically important yeast species other than Candida. Int J Pharm Pharm Sci 2014, 6, 291–294. [Google Scholar]
- Lee, J.S.; Shin, J.H.; Kim, M.-N.; Jung, S.-I.; Park, K.H.; Cho, D.; Kee, S.J.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Kodamaea ohmeri isolates from patients in a university hospital: identification, antifungal susceptibility, and pulsed-field gel electrophoresis analysis. Journal of clinical microbiology 2007, 45, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Yan, J.; Gao, Z.; Yang, H.; Tang, Y.; Yang, L. Subcutaneous granuloma caused by Kodamaea ohmeri in an immunocompromised patient in China. Australasian Journal of Dermatology 2020, 61, e213–e216. [Google Scholar] [CrossRef] [PubMed]
- Biswal, D.; Sahu, M.; Mahajan, A.; Advani, S.H.; Shah, S. Kodameae ohmeri–An emerging yeast: two cases and literature review. Journal of Clinical and Diagnostic Research: JCDR 2015, 9, DD01. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yu, S.; Kudinha, T.; Xiao, M.; Wang, H.; Xu, Y.; Zhao, H. Identification and antifungal susceptibility profiles of Kodamaea ohmeri based on a seven-year multicenter surveillance study. Infection and drug resistance 2019, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Sathi, F.A.; Aung, M.S.; Paul, S.K.; Nasreen, S.A.; Haque, N.; Roy, S.; Ahmed, S.; Alam, M.M.; Khan, S.; Rabbany, M.A. Clonal Diversity of Candida auris, Candida blankii, and Kodamaea ohmeri Isolated from Septicemia and Otomycosis in Bangladesh as Determined by Multilocus Sequence Typing. Journal of Fungi 2023, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Overview of antifungal ECOFFs and clinical breakpoints for yeasts, m.a.d.u.t.E.E.D., E.Def 9.4 and E.Def 11.0 procedures. Version 4.0, 2023. http://www.eucast.org.
- Clinical and Laboratory Standards Institute. Reference method for broth dilution antifungal susceptibility testing of yeasts 4th ed CLSI standard M27. Wayne, P. 2017.
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiology and immunology 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Cendejas-Bueno, E.; Gomez-Lopez, A.; Mellado, E.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Identification of pathogenic rare yeast species in clinical samples: comparison between phenotypical and molecular methods. Journal of clinical microbiology 2010, 48, 1895–1899. [Google Scholar] [CrossRef]
- Sipiczki, M.; Tap, R.M. Candida vulturna pro tempore sp. nov., a dimorphic yeast species related to the Candida haemulonis species complex isolated from flowers and clinical sample. International Journal of Systematic and Evolutionary Microbiology 2016, 66, 4009–4015. [Google Scholar] [CrossRef]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Brunke, S.; Mogavero, S.; Kasper, L.; Hube, B. Virulence factors in fungal pathogens of man. Current opinion in microbiology 2016, 32, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Giacobino, J.; Montelli, A.C.; Barretti, P.; Bruder-Nascimento, A.; Caramori, J.T.; Barbosa, L.; Bagagli, E. Fungal peritonitis in patients undergoing peritoneal dialysis (PD) in Brazil: molecular identification, biofilm production and antifungal susceptibility of the agents. Medical Mycology 2016, 54, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Maciel, N.O.; Johann, S.; Brandão, L.R.; Kucharíková, S.; Morais, C.G.; Oliveira, A.P.; Freitas, G.J.; Borelli, B.M.; Pellizzari, F.M.; Santos, D.A. Occurrence, antifungal susceptibility, and virulence factors of opportunistic yeasts isolated from Brazilian beaches. Memórias do Instituto Oswaldo Cruz 2019, 114. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Johnson, A.D. Candida albicans biofilms and human disease. Annual review of microbiology 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Malinovská, Z.; Čonková, E.; Váczi, P. Biofilm Formation in medically important Candida species. Journal of Fungi 2023, 9, 955. [Google Scholar] [CrossRef]
- Ramage, G.; Borghi, E.; Rodrigues, C.F.; Kean, R.; Williams, C.; Lopez-Ribot, J. Our current clinical understanding of Candida biofilms: where are we two decades on? APMIS 2023. [Google Scholar] [CrossRef] [PubMed]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PloS one 2012, 7, e36952. [Google Scholar] [CrossRef]
- de Souza, C.; Perini, H.; Caloni, C.; Furlaneto-Maia, L.; Furlaneto, M. Adhesion of Candida tropicalis to polystyrene and epithelial cell lines: Insights of correlation of the extent of adherent yeast cells among distinct surfaces. Journal de Mycologie Médicale 2020, 30, 101043. [Google Scholar] [CrossRef]
- de Souza, C.M.; Perini, H.F.; Verri, W.A.; Zaninelli, T.H.; Furlaneto-Maia, L.; Furlaneto, M.C. Changes in adhesion of Candida tropicalis clinical isolates exhibiting switch phenotypes to polystyrene and HeLa cells. Mycopathologia 2021, 186, 81–91. [Google Scholar] [CrossRef]
- Pote, S.T.; Sonawane, M.S.; Rahi, P.; Shah, S.R.; Shouche, Y.S.; Patole, M.S.; Thakar, M.R.; Sharma, R. Distribution of Pathogenic Yeasts in Different Clinical Samples: Their Identification, Antifungal Susceptibility Pattern, and Cell Invasion Assays. Infection and Drug Resistance 2020, 13, 1133–1145. [Google Scholar] [CrossRef]
- Dalle, F.; Wächtler, B.; L’Ollivier, C.; Holland, G.; Bannert, N.; Wilson, D.; Labruère, C.; Bonnin, A.; Hube, B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cellular microbiology 2010, 12, 248–271. [Google Scholar] [CrossRef]
- Erwig, L.P.; Gow, N.A.R. Interactions of fungal pathogens with phagocytes. Nature Reviews Microbiology 2016, 14, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Immunity to fungal infections. Nature Reviews Immunology 2011, 11, 275–288. [Google Scholar] [CrossRef]
- Trovato, L.; Calvo, M.; Scalia, G.; Oliveri, S. A Comparative Prospective Study in Evaluating Candida spp. In Vitro Susceptibility through Micronaut-AM and Sensititre Yeast-One. Microbiology Research 2023, 14, 1077–1088. [Google Scholar] [CrossRef]
- Altinbaş, R.; BARIŞ, A.; Şen, S.; Öztürk, R.; Kiraz, N. Comparison of the Sensititre YeastOne antifungal method with the CLSI M27-A3 reference method to determine the activity of antifungal agents against clinical isolatesof Candida spp. Turkish Journal of Medical Sciences 2020, 50, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Turnidge, J.; Alastruey-Izquierdo, A.; Botterel, F.; Canton, E.; Castro, C.; Chen, Y.-C.; Chen, Y.; Chryssanthou, E.; Dannaoui, E. Method-dependent epidemiological cutoff values for detection of triazole resistance in Candida and Aspergillus species for the Sensititre YeastOne colorimetric broth and Etest agar diffusion methods. Antimicrobial agents and chemotherapy 2019, 63. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic analysis of K. ohmeri based on the internal transcribed spacer (ITS) regions of the 18S rDNA gene of isolates from different geographical regions. The Tamura– Nei genetic distance model and the Neighbor-Joining method, with a bootstrap of 1,000 replicates, were used to construct a cladogram. The sequences obtained in this study are shown inside the light blue box.
Figure 1.
Phylogenetic analysis of K. ohmeri based on the internal transcribed spacer (ITS) regions of the 18S rDNA gene of isolates from different geographical regions. The Tamura– Nei genetic distance model and the Neighbor-Joining method, with a bootstrap of 1,000 replicates, were used to construct a cladogram. The sequences obtained in this study are shown inside the light blue box.
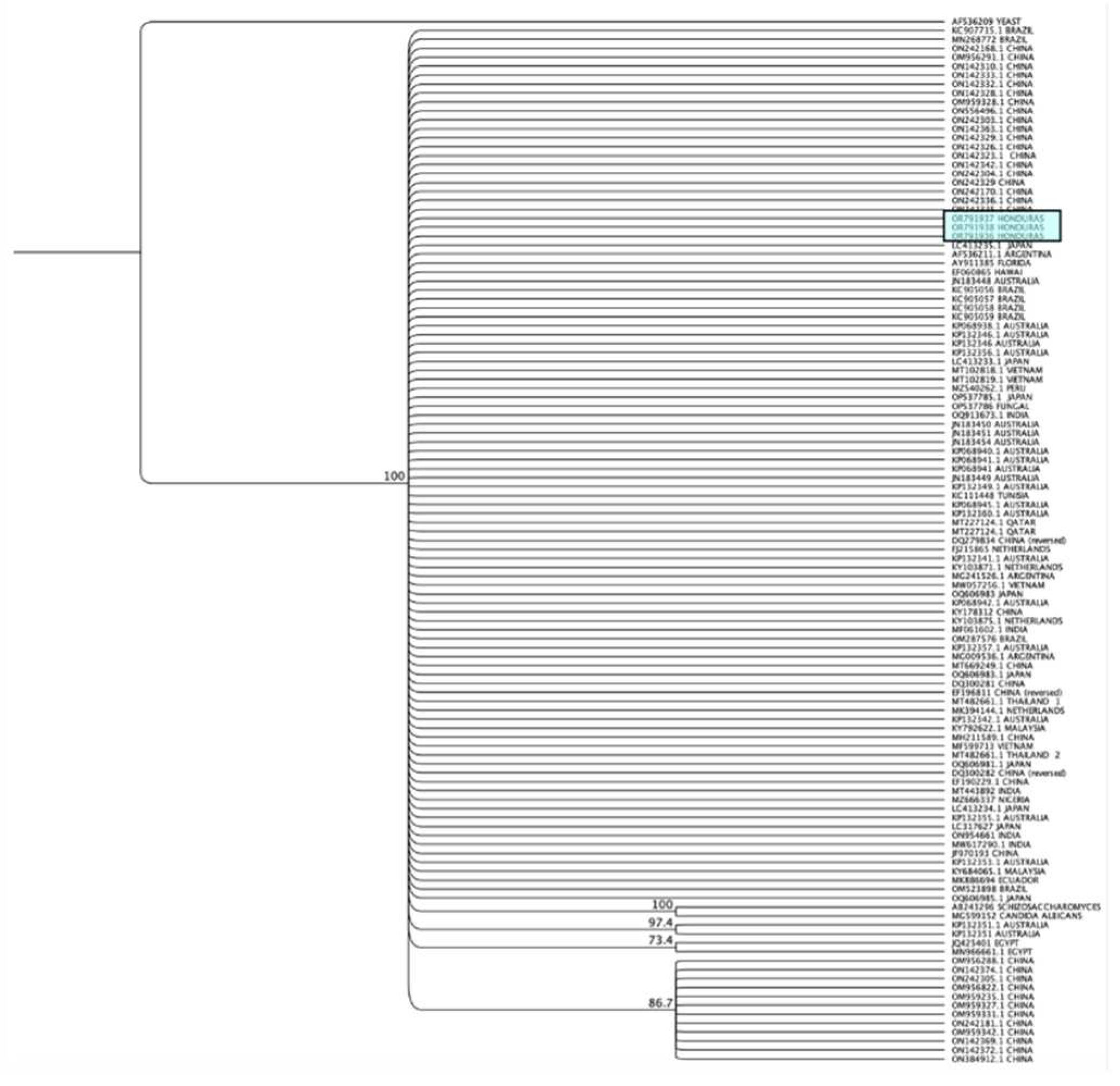
Figure 2.
Growth of Kodamaea ohmeri in (A) CHROMagar medium (CHROMagar CandidaTM, France) (B) CHROMagarTM (Becton, Dickinson and Company, NJ, USA), (C) ChromaticTM Candida, Liofilchem®. (D1) Germ tube production of the positive control (C. albicans ATCC 10231). (D2) Germ tube test in K. ohmeri (non-producer), (E) Photomicrograph with 40× objective of fungal structures obtained from culture on corn flour agar at 30 °C, for 48 h of (E1) the positive control (C. albicans ATCC 10231), and (E2) K. ohmeri showing pseudohyphae with ovoid and elongated blastoconidia on the sides.
Figure 2.
Growth of Kodamaea ohmeri in (A) CHROMagar medium (CHROMagar CandidaTM, France) (B) CHROMagarTM (Becton, Dickinson and Company, NJ, USA), (C) ChromaticTM Candida, Liofilchem®. (D1) Germ tube production of the positive control (C. albicans ATCC 10231). (D2) Germ tube test in K. ohmeri (non-producer), (E) Photomicrograph with 40× objective of fungal structures obtained from culture on corn flour agar at 30 °C, for 48 h of (E1) the positive control (C. albicans ATCC 10231), and (E2) K. ohmeri showing pseudohyphae with ovoid and elongated blastoconidia on the sides.
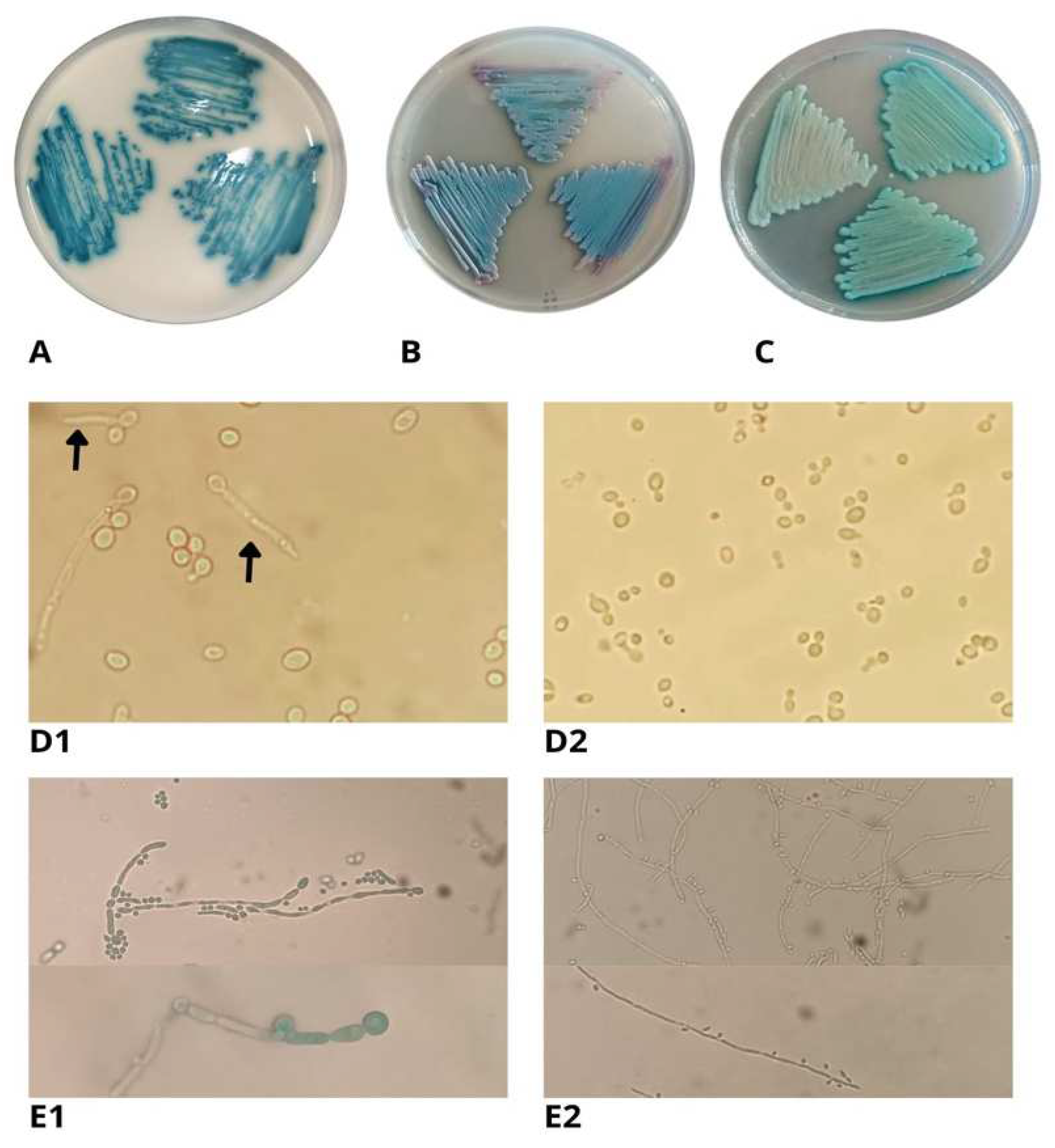
Figure 3.
Adhesion and invasion assays in HeLa cells. (A) Percentage of HeLa cells that exhibited adherent yeasts; (A1) Micrograph obtained by brightfield microscopy with 40× objective. Giemsa stain showing yeasts and pseudohyphae of strain C-29 adhered to HeLa cells; (B) Number of pseudohyphae per HeLa cell; (B1) Micrograph obtained by brightfield microscopy at 100× objective. Giemsa stain. Pseudohyphae adhered to HeLa cells; (C) Percentage of macrophages per field invaded by strain (C1) Micrograph obtained by brightfield microscopy at 100× objective. Giemsa stain. Macrophage invaded by yeasts of strain C-29.
Figure 3.
Adhesion and invasion assays in HeLa cells. (A) Percentage of HeLa cells that exhibited adherent yeasts; (A1) Micrograph obtained by brightfield microscopy with 40× objective. Giemsa stain showing yeasts and pseudohyphae of strain C-29 adhered to HeLa cells; (B) Number of pseudohyphae per HeLa cell; (B1) Micrograph obtained by brightfield microscopy at 100× objective. Giemsa stain. Pseudohyphae adhered to HeLa cells; (C) Percentage of macrophages per field invaded by strain (C1) Micrograph obtained by brightfield microscopy at 100× objective. Giemsa stain. Macrophage invaded by yeasts of strain C-29.
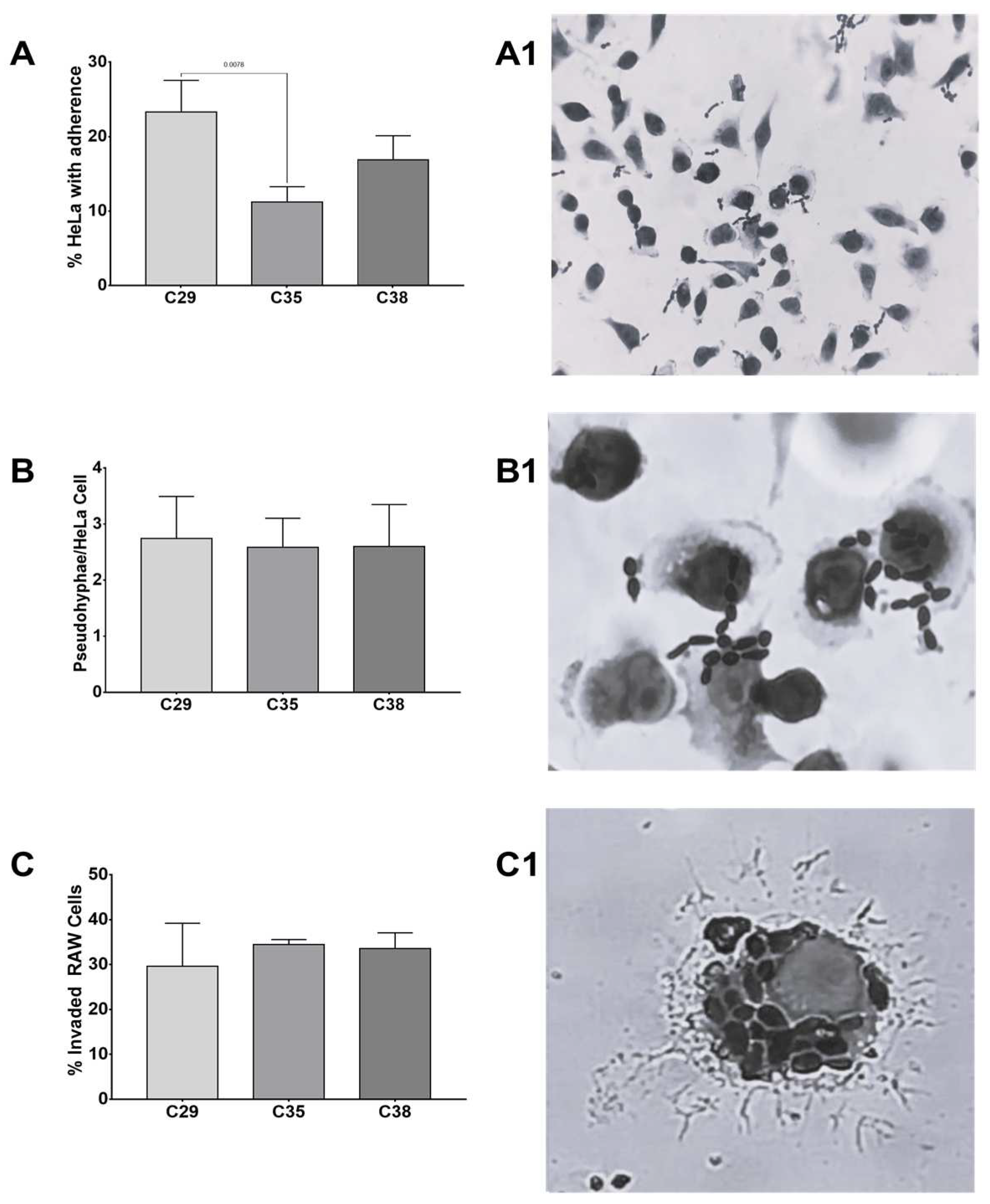

Table 1.
Evaluation of the Minimum Inhibitory Concentration (MIC) of nine antifungal drugs on three isolates of Kodamaea ohmeri using the SensititreTM Yeast-OneTM system.
Table 1.
Evaluation of the Minimum Inhibitory Concentration (MIC) of nine antifungal drugs on three isolates of Kodamaea ohmeri using the SensititreTM Yeast-OneTM system.
| Antifungal drug | Isolate (MIC) | ||
|---|---|---|---|
| C-29 (mg/L) | C-35 (mg/L) | C-38 (mg/L) | |
| Amphotericin B | 0.5 | 0.25 | 0.5 |
| Anidulafungin | 0.25 | 0.12 | 0.12 |
| Caspofungin | 2.0 | 1.0 | 0.25 |
| Micafungin | 0.12 | 0.25 | 0.12 |
| Fluconazole | 4.0 | 4.0 | 4.0 |
| Itraconazole | 0.12 | 0.12 | 0.12 |
| Posaconazole | 0.06 | 0.12 | 0.06 |
| Voriconazole | 0.03 | 0.03 | 0.03 |
| 5-Flucytosine | ≤ 0.06 | ≤ 0.06 | ≤ 0.06 |
Table 2.
Analysis of biofilm production and coefficient of extracellular enzymatic activity (Pz).
| (Pz)** | ||||||
|---|---|---|---|---|---|---|
| Isolate | Biofilm production* | Gelatin | BSA | Casein | Phospholipase | Hemolisin |
| C-29 | 0.831 ± 0.12 Strong producer |
0.57 ± 0.04 | 0.48 ± 0.02 | 0.83 ± 0.06 | 0.67 ± 0.03 | 0.30 ± 0.02 |
| C-35 | 1.245 ± 0.16 Strong producer |
0.51 ± 0.05 | 0.38 ± 0.05 | 0.89 ± 0.01 | 0.62 ± 0.03 | 0.30 ± 0.04 |
| C-38 | 0.727 ± 0.15 Strong producer |
0.56 ± 0.02 | 0.46 ± 0.04 | 0.89 ± 0.02 | 0.67 ± 0.05 | 0.30 ± 0.02 |
* Non-producer (ODs ≤ ODnc); weak producer (ODnc < ODs ≤ 2× ODnc); moderate producer (2× ODnc < ODs ≤ 4× ODnc), and strong producer (ODs > 4× ODnc). ** Pz = 1.0 no enzymatic activity; Pz [0.99 — 0.90] = weak; Pz [0.89 — 0.70] moderate; Pz ≤ to 0.69 = strong.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



