
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Abstract: The worldwide spread of SARS-CoV-2 has led to a significant economic and social burden on a global scale. Even though the pandemic has concluded, there remains apprehension regarding the emergence of highly transmissible variants capable of evading immunity induced by either vaccination or prior infection. The success of viral penetration is the specific amino acid residues of the Receptor Binding Motif (RBM) involved in viral attachment. This region interacts with the cellular receptor ACE2, triggering a neutralizing antibody (nAb) response. In this study, we have evaluated the immunogenicity of serum from individuals who received either a single dose or a combination of different vaccines against the original SARS-CoV-2 strain and mutated linear RBM. Despite a modest antibody response to the wild-type SARS-CoV-2 RBM, the Omicron variants exhibit four mutations in the RBM (S477N, T478K, E484A, F486V) that result in even lower antibody titers. The primary immune responses observed were directed towards IgA and IgG. While nAbs typically target the RBD, our investigation has unveiled a reduced seroreactivity within the RBD's crucial subregion, the RBM. This deficiency may have implications for the generation of protective nAbs. Evaluation of S1WT and S2WT RBM peptides binding to nAbs using microscale thermophoresis revealed a higher affinity (35 nM) for the S2WT sequence (GSTPCNGVEGFNCYF), which includes the FNCY patch. Our findings suggest that the linear RBM of SARS-CoV-2 is not an immunodominant region in vaccinated individuals. Comprehending the intricate dynamics of the humoral response, its interplay with viral evolution, and host genetics is crucial for formulating effective vaccination strategies, not only targeting SARS-CoV-2 but also anticipating potential future coronaviruses.
Keywords:
SARS-CoV-2
; variants
; spike glycoprotein
; receptor binding motif
; receptor binding domain
; neutralizing antibodies
; Ig subclasses
; IgG
; IgA
1. Introduction
COVID-19 is attributed to the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which initially emerged in Wuhan, China, and quickly disseminated globally, leading to substantial economic and societal challenges. [1]. Despite the recent announcement by the World Health Organization declaring the end of the COVID-19 pandemic [2], the virus continues to evolve through mutations and recombination. The emergence of new SARS-CoV-2 variants raises concerns regarding their potential to evade host immunity. For instance, the Omicron variants have been reported to evade neutralizing antibodies (nAbs) induced by vaccination or prior infection [3]. Mutations, especially in the Spike (S) protein, threaten the efficacy of current vaccines due to their capacity to evade antibody recognition, a critical component of the immune response [4]. The study of the humoral response to SARS-CoV-2 is crucial for understanding population-wide and individual responses to viral infection and vaccination. The variation in the antibody response to linear B-cell epitopes among individuals is particularly important for serodiagnosis and vaccine development.
Neutralizing antibodies can target different regions of the Spike protein, with the majority binding to the N-terminal domain (NTD) and the receptor-binding domain (RBD) [5]. The binding modes of Receptor Binding Domain (RBD)-specific nAbs can be classified into four primary categories, depending on their target epitopes. Neutralization can take place through various mechanisms, including competition with the ACE-2 receptor, direct binding to the RBD, and the induction of steric hindrance, thereby restraining conformational changes in the Spike protein neutralization can occur through competition with the ACE-2 receptor, direct binding to RBD, and inducing steric hindrance and restraining conformational changes in the S protein [6].
The RBD, situated in the S1 domain of the S protein, has undergone extensive examination owing to its marked variability and pivotal function in interacting with the virus entry receptor in mammalian cells, namely the angiotensin-converting enzyme 2 (ACE2). Previous research has shown that the S protein is highly immunogenic, and protective nAb responses are predominantly directed against the RBD domain [7,8,9].
It is estimated that approximately 90% of the neutralizing activity in convalescent sera is attributed to nAbs binding to the RBD. Within the RBD, a receptor-binding motif (RBM) plays a crucial role in interacting with the ACE2 receptor. The RBM can be divided into three flexible coil regions: knob (aa 444-449 and 496-505), base (aa 490-494 and 450-456), and tip (aa 473-489) [8].
Neutralization is primarily mediated by IgG, IgA, and IgM, with IgG being the most abundant neutralizing antibody [9]. IgM and IgG are produced simultaneously or sequentially in response to SARS-CoV-2 infection, reaching peak concentrations within the first two weeks and remaining in the bloodstream for at least six months [10]. IgG can be subdivided into four subclasses (IgG1, IgG2, IgG3, and IgG4) with diverse effector functions based on the constant region [11]. Viral proteins predominantly elicit IgG1 and IgG3 responses, while IgG2 and IgG4 have been associated with polysaccharide antibacterial responses [11,12]. Regarding binding capacity, convalescent sera IgG3 has been shown to have the most pronounced ability to bind to the SARS-CoV-2 RBD [13]. High levels of anti-RBD IgG4 subclass have been correlated with increased mortality and severe disease [14].
While some SARS-CoV-2 mutations may be neutral or harmful, a subset can enhance viral fitness and enable immune evasion [4]. The S protein and ACE2 interaction is predominantly facilitated by amino acid residues within the RBM, which are the focal points of nAbs [15,16,17]. Contact points of potent nAbs like B38 and CC12.1 overlap with the Spike-ACE2 interaction residues [15]. Mutations in the RBM, including E484K and N501Y, whether alone or in combination, have been associated with decreased binding of nAbs [8,17]. Significantly, the N501Y mutation compromises the affinity of neutralizing antibodies while concurrently enhancing the binding affinity to ACE2.[18]. The most nAb appear to bind to conformational epitopes within the RBD, and antibody responses depend on structurally folded S protein and RBD [19]. Protein conformation plays a role in immunization, folded but not with unfolded RBD, induced antibodies against conformational epitopes with high virus-neutralizing activity [19].
Though numerous research groups have identified nAbs targeting the RBM, more understanding of their impact on the antibody pool in the serum of individuals infected with or vaccinated against SARS-CoV-2 should be understood. However, evidence suggests they are produced in lower frequencies [15,20]. Given the critical role of the RBM in viral entry and ACE2 interaction, it is reasonable to assume that it is highly immunogenic and can provide a structure-based framework for rational vaccine design and the selection of immunotherapeutic agents. Consequently, in this work the RBM sequences were selected for synthesizing and evaluating the humoral response in a panel of vaccinated individuals. To understand the impact of SARS-CoV-2 mutations on host antibody responses, sequences from Omicron BA.4 and BA.5 were also analyzed.
Additionally, IgG class and subclasses specific to the RBM region were characterized in vaccinated individuals, and the interaction of antibodies with the RBM region was assessed using Microscale thermophoresis (MST). The comprehensive linear peptide analysis of the RBM tip revealed that this region is not immunodominant but is still present in a subset of immunized individuals.
2. Materials and Methods
2.1. Serum Samples
A panel of 31 serum samples was obtained from individuals who received a single dose of the Oxford/AstraZeneca vaccine, and 15 serum samples were collected from individuals who received four doses. Serum from healthy donors, collected before the pandemic, was provided by HEMORIO, a centralized network of blood donor facilities in Rio de Janeiro, Brazil. To ensure patient privacy, the sera samples were supplied without identifying information.
2.2. Peptides Synthesis
SARS-CoV-2 RBM 15-mer peptides [(S1WT) FERDISTEIYQAGST); S2WT (GSTPCN GVEGFNCYF), and S3WT (YFPLQSYGFQPTNGV)] were chosen for synthesis using the F-moc strategy in a synthesizer machine (MultiPep-1 CEM Corp, Charlotte, NC, USA) [21]. Following sequence assembly, F-moc groups were removed, and the peptide-resin (Wang resin) was cleaved and fully deprotected with TFA/H2O/EDT/ TIS (94/2.5/2.5/1.0 v/v, 90 min). The peptide was precipitated with chilled diethyl ether, centrifuged, and the pellet was reconstituted in aqueous AcOH (10% v/v), dried, and stored as a lyophilized powder. Peptides, when needed, were dissolved in water and centrifuged, and the supernatant was filtered using a centric-10 filter. Single peptides were used without prior purification, with their identity confirmed by MS (MALDI-TOF or electrospray).
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
In-house ELISA was conducted using Immunolon 4HB plates (Immunochemistry Technologies, Bloomington, MN, USA). The coating process involved overnight incubation at 4 °C, wherein each well received 500 ng of peptides in a coating buffer (50 mM carbonate-bicarbonate buffer, pH 9.6). After washing with PBS-T (phosphate-buffered saline plus 0.05% Tween® 20), plates were incubated for 1 h at 37 °C with 1% BSA (200 µl) in PBS-T to block free binding sites. Subsequently, patient sera were diluted (1:25) in coating buffer, and 100 µl were applied onto immunosorbent plates, incubating for 1 h at 37 °C. After several washes with PBS-T, plates were incubated with 100 µl goat anti-human IgG-HRP (1:10.000, Sigma-Aldrich, St Louis, MO, USA) for 1 h at 37 °C. Finally, 3,3′,5,5′-tetramethylbenzidine (1-Step™ Ultra TMB-ELISA, Science Biotech Ltda, Lages, SC, Brazil) was added for 15 min, and the reaction was stopped with 0.5 M sulfuric acid. The plate was read within 2 h of adding the Stop Solution. Values of blank wells containing only peptides were subtracted from the sample's optic density.
2.4. Purification of RBM Antibodies
Antibodies anti-RBM were purified from a pool (n=10) of serum from individuals who received a single dose of the vaccine through an affinity column (3 cm × 1 cm i.d.) prepared using an RBD SARS-CoV2 recombinant protein [22] coupled on Sepharose-4B beads, following previously described procedures [23]. Anti-RBM antibodies were eluted with glycine buffer pH 2.8 and collected in Eppendorf tubes containing Tris-HCl pH 10 buffer, subsequently concentrated on centricon-P30 filters (Sigma Chemical Co, Saint Louis, MO, U.S.A.) (Figure S1). Protein concentration was determined by measuring absorbance at 280 nm and an extinction coefficient of 13.4. In-house ELISA analyzed the activity of purified antibodies. Immunolon 4HB plates were coated overnight at 4 °C with 200 ng of multiepitope (Dx-SARS-RBD) recombinant protein containing RBM peptides or non-SARS-CoV-2 sequences (Control). In-house ELISA followed the above-mentioned methods, but incubation was performed using purified antibodies (100 µg/mL) diluted at 1:1.000.
2.5. Microscale Thermophoresis (MST)
Purified nAb to the RBM regions (Figure S1) were labeled with the fluorescent dye NT-647 using Monolith NT™ Protein Labeling Kits and mixed at a 3:1 molar ratio with unlabeled protein. For the interaction experiments, the fluorescent anti-RBM antibodies were kept at a constant concentration (12.5 µg/mL), while the concentration of the unlabeled S1WT and S2WT peptides varied from 0.5 µg/mL–0.12 ng/mL. The assay was performed in PBS containing 0.05% Tween 20, and after a short incubation, the samples were loaded into standard MST NT.115 glass capillaries. Kd data were calculated using the NanoTemper software package [24].
2.6. Protein Thermal Shift Assay
Protein thermal shift assays were conducted using the QuantStudio 3 PCR System (Applied Biosystems, Foster City, California, USA). The measurements were performed in a MicroAmp® optical 8 strip (0.2 mL) from Applied Biosystems. All steps were performed on ice. The samples were assayed as duplicates or quadruplicates in each measurement. All measurements contained a no protein negative control, positive control with Lysozyme (10 µg), and peptide S8WT (5µg). A carbonate-bicarbonate buffer (50 mM, pH 9.6) and a protein thermal shift dye (Applied Biosystems) were used in a 4X dye concentration. After pipetting all components into the wells, the plate was sealed with MicroAmp™ Optical Adhesive Film (Applied Biosystems, Foster City, California, USA) and incubated on ice for 15 min. Applied Biosystems QuantStudio™ 3 Real-Time PCR System using melt curve filter setting at x2-m2, continuous data collection, and a ramp rate of 0.05°C/sec from 25°C through 99°C. Melting curves were analyzed using the TSAR application in R software.
2.7. Statistical Analysis
Data were analyzed using Prism software (GraphPad version 6, San Diego, CA, USA). The Kruskal–Wallis test was applied to identify statistical differences, followed by Dunn's multiple comparisons tests. A p-value < 0.05 was considered a significant difference. The reactivity index (RI) reflects absorbance divided by the cut-off determined by the receiver operation curve (ROC) analysis of each protein. Results >1.1 were considered positive, and <0.90 were deemed negative. Samples with a relative index (RI) value of 1.0 ±10% were defined as being in a gray zone and deemed inconclusive [22].
3. Results
The Spike sequences from Variants of Concern (VoCs) were aligned and subjected to phylogenetic analysis. The Omicron variants exhibited twelve mutations in the RBM, including mutations previously reported in other VoCs such as L452R, T478K, E484, and N501Y. Additionally, Omicron presented new mutations, namely N440K, G446S, L452Q/R, S477N, T478K, E484A, F486V, Q493R, G496S, and Y505H (Figure 1a). Most of these mutations are known to impact viral fitness and contribute to immune evasion.
The interaction of various human neutralizing antibodies with the binding residues on RBM was assessed (Figure 1b). Most of the binding occurred on the RBM tip (aa 473-489), where six amino acid residues (37.5%) were mutated in Omicron variants (S477N, T478K, E484A, F486V, Q493R, G496S). Interactions of Spike with ACE-2 were highlighted, encompassing residues between 438–506 of the RBM, which make maximal contact with the N-terminal PD domain of ACE2. The evaluated human nAbs displayed contact residues in the RBM that overlapped with amino acids interacting with ACE2.
Given the strength of most Omicron mutations in the RBM tip, we synthesized peptides with varying lengths to analyze antibody production and binding affinity. Initially, the RBM, including the tip region, was fragmented into three 15-mer peptides: S1WT (464FERDISTEIYQAGST478), S2WT (476GSTPCNGVEGFNCYF490), and S3WT (489YF PLQSYGFQPTNGV503) (Figure 1c).
To analyze the folding state of the synthesized peptide S8WT, a Thermal Shift Assay (TSA) was performed. Thermal stability was evaluated in carbonate-bicarbonate buffer at pH 9.6. Although peptides can assume many flexible conformations, no transition state was observed (Figure S1), indicating a linear conformation of the peptide S8WT. In contrast, Lysozyme used as control showed a melting curve with a Tm of 72.7 °C (Figure S1)
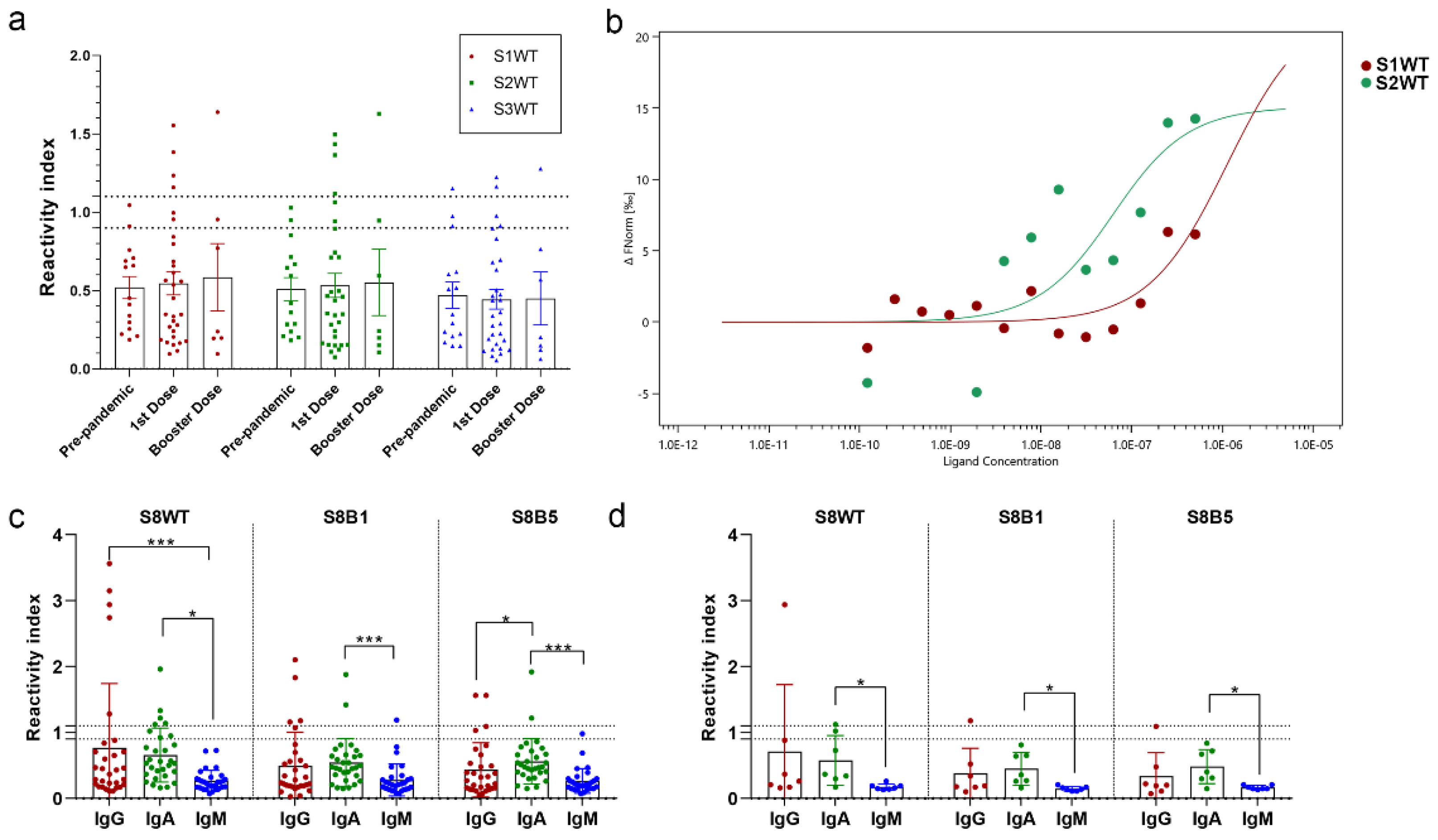
Seroreactivity of these three regions was tested by peptide ELISA, using a serum panel of vaccinated individuals with one dose of Oxford/AstraZeneca (ChAdOx1-S) (Table S1) and four doses of heterologous boosting (Oxford/AstraZeneca - ChAdOx1-S; Pfizer-BioNTech - BNT162b2 or Janssen - Ad26.COV2.S) (Figure 2 and Table S2). A panel of pre-pandemic sera was used as a control for the cut-off calculation. Few individuals responded for IgG to the RBM tip peptides, with reactivity index >1.1; seroreactivity for peptides S1WT and S2WT were similar for prime immunization 4/30 (13.3%) and for booster heterologous doses 1/7 (14.3%). Peptide S3WT had a lower performance, with only two positive samples for first-dose vaccination and one for booster heterologous doses (Figure 2a). Although low antibody concentration, one open question is the affinity of produced Spike nAbs to these regions. Microscale thermophoresis was used to measure the peptide-antibody interactions' Kd. Specific purified antibodies to the RBM were separated using a multiepitope recombinant protein affinity column (Figure S1), and then antibodies were tagged with a fluorescent probe; interactions of antibodies with different concentrations of peptides (S1WT and S2WT) were measured in glass capillaries and MST traces recorded (Figure S2). The dissociation constant calculated from the dose-response curve was strikingly different from peptides S1WT (Kd= 640,73 nM) and S2WT (Kd=35 nM) (Figure 2b). Results demonstrated that the binding affinity of RBM antibodies to peptide S2WT was stronger. Aiming to increase the sensitivity of the analysis, a 37-mer peptide was synthesized (S8WT, 452LFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFP488). Also two other peptides from Omicron BA.1 (S8BA1, 452LFRKSNLKPFERDISTEIYQAGNKPC NGVAGFNCYFP488) and Omicron BA.5 (S8BA5, 452LFRKSNLKPFERDISTEIYQAGNKP CNGVAGVNCYFP488) included these VoCs mutations (Figures 1c). There was a slight increase in seroreactivity for IgG of the first dose vaccination 6/30 (20%) for peptide S8WT in comparison with fragmented residues (13-14%) (Figures 2a and c). Mutation at the RBM region of Omicron BA.1 (S8BA1) and BA.5 (S8BA5) reduced the reactivity index mean and the number of positive samples 4/30 and 2/30, respectively. IgM levels were low and significantly different from IgG and IgA in one or heterologous doses of vaccinated individuals (Figures 2c and d). Additionally, some individuals with one dose of vaccine produced IgA response to the RBM tip of S wild type (5/30) and reduced response to Omicron BA.1 (2/30) and BA.5 (2/30) (Figure 2c). Analyzing the group of heterologous vaccination booster doses, only one sample had seroreactivity for IgG and IgA against wild-type sequence S8WT (Figure 2d). Mutations in Omicron variants reduced the reactivity index of this sample almost three times. Subclasses of IgG-positive samples were also analyzed for the group with the first dose vaccination; a predominance of IgG1 was found (5/12), but reactivity significantly decreased for Omicron variants (Figure S3).
4. Discussion
The humoral response against the S protein can be elicited through natural infection or immunization, and immune protection against symptomatic SARS-CoV-2 hinges on the levels of neutralizing antibodies [25]. It is suggested that nAbs against the S protein may be both low in quantity and short duration due to the structural properties of coronaviruses [26]. Notably, the N-terminal and the receptor-binding domain are considered immunodominant and are the primary targets of nAbs. However, nAbs can also target regions such as the S2 stem helix (SH) and the S2 fusion peptide (FP) [17,27]. Antibodies binding to the RBD constitute over 90% of the neutralizing activity in convalescent sera [7]. nAbs exhibit four main classes based on the location of their epitopes within the Spike protein [3]. Classes 1 and 2 nAbs primarily target the RBD, including the receptor-binding motif, and can compete with ACE2 binding. Class 3 RBD nAbs bind to regions flanking the ACE2-binding region, while class 4 nAbs are highly conserved in the RBD but do not directly block ACE2-RBD binding [3]. Various monoclonal antibodies have been identified to bind to the RBM region [15]. However, there remains limited knowledge regarding the diversity of antibodies produced. The RBM region is pivotal for ACE2 receptor binding and comprises the knob (aa 444-449 and aa 496-505), base (aa 490-494 and aa 450-456), and tip (aa 473-489) [8]. Many nAbs against SARS-CoV-2 are found to bind to the tip of the RBM, particularly in or around the FNCY patch [15]. The MST analysis conducted in this study corroborates the significance of these residues in antibody binding affinity. A recent study highlighted the importance of RBD folding to antibody binding and immunization. Most IgG responses were produced for folded RBD; conformational epitopes play major roles in this region [19] and correlate with high virus neutralization. In the same study, authors reported that out of the 253 COVID-19 patients, 20% lacked RBD-specific IgG antibodies, possibly due to genetic factors such as human leukocyte antigen (HLA) restriction and insufficient T/B-cell responses [19]. Despite the dominance of conformational epitopes in the RBD, our data showed that immunization also produced in some individuals (~20%) the development of antibodies to linear RBM epitopes. Linear epitopes to RBD showed a low neutralization profile [19] but have diagnostic significance since many proteins are not designed to be correctly folded in diagnostic tests. Immune evasion by variants of concern is primarily driven by mutations in the S protein that compromise both natural and vaccine-induced immunity. The Omicron variant accumulates mutations in the RBM, including ten mutations in this region. Some of these mutations directly interfere with ACE2 binding, affecting important residues in the RBM, such as those at positions Q493, Q498, N501, and Y505 [28,29]. A comparison of the immunogenicity of RBM, RBD, and the entire S protein reveals that the RBM region displays reduced seroreactivity [15,20]. However, mouse immunization with RBD or RBM vaccines has induced nAbs [30,31]. According to our data, adenovirus vaccines containing Se mRNA, such as Astra-Zeneca-Oxford or combined heterologous mRNA vaccines, seem to induce low titers of anti-RBM antibodies. The diminished sera reactivity observed in our study may be attributed to Major Histocompatibility Complex class II (MHC-II) restriction [32]. A study examining common MHC-II alleles predicted poor binding for the RBM, suggesting a lack of MHC-II support in T-B cooperation, which impacts the production of nAbs in the general population [31]. Effective antibody responses necessitate cooperation between a B cell and a CD4 T cell (helper cell) activated by an epitope on the same antigen recognized by the B cell (T-B cooperation). Nonetheless, T-B cooperation is limited by MHC-II molecules [33]. Thus, studying B and T cell epitopes and the HLA polymorphism in different populations is relevant for vaccine design [34]. A recent study demonstrated that the HLA profile influences the variability of both humoral and cellular responses to the mRNA vaccine. An association was found between HLA haplotype and high antibody concentration and or low humoral responses to Spike antigens [35]. Another study revealed that variation in humoral responses against SARS-CoV-2 spike and the RBD at 28 days after the first vaccination (ChAdOx1-S) shows genome-wide significant association with major MHC-II alleles [36].
5. Conclusions
The humoral response, elicited by both SARS-CoV-2 infection and vaccination, primarily targets the Spike protein's RBD. Despite the production of neutralizing antibodies directed to Spike, the immunodominance of the RBD region does not guarantee long-term protective immunity. The emergence of SARS-CoV-2 variants, such as the Omicron variant, with mutations in the RBD, presents a challenge to the efficacy of current vaccines. Our findings suggest that the RBM region, crucial for viral entry and ACE2 interaction, displays reduced seroreactivity, potentially impacting the production of neutralizing antibodies in vaccinated individuals. The interplay between B and T cell epitopes, MHC-II restriction, and HLA polymorphism underscores the need for comprehensive population-based studies for effective vaccine design. The intricate interplay among B and T cell epitopes, MHC-II restriction, and HLA polymorphism highlights the need for thorough population-based studies to design vaccines effectively. Additionally, our results confirm the importance of RBD conformation to antibody recognition and highlight the significance of linear epitopes in a small subset of individuals. Comprehending the intricacies of the humoral response and recognizing the impact of genetic variability on vaccine efficacy is essential for developing resilient and inclusive vaccination strategies and diagnostic tests for SARS-CoV-2 and other emerging coronaviruses
6. Patents
The protein receptacle, method for receptacle production, and antigenic peptide sequences described in this study are protected under Brazilian (BR10.2019.017792.6), USA (PCT/BR2020/ 050341), Europe. (PCT: 26/06/2023), India (PCT: 26/06/2023) and China. (PCT: 26/03/2023) Provisional patents, respectively, were filed by FIOCRUZ. They may serve as a future source of funding.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Thermal stability of S8WT peptide. Lysozyme (10 ug) was used as positive control, and peptide (5 ug) was mixed with the Protein Thermal Shift dye. Duplicate and quadruplicate reactions were run on an Applied Biosystems QuantStudio™ 3 Real-Time PCR System using melt curve filter setting at x2-m2, continuous data collection, and a ramp rate of 0.05°C/sec from 25°C through 99°C. Data were analyzed using the TSAR application in R software. Data showed Fluorescence vs. temperature (°C). FigureS2: Purification of human antibodies against the RBM region of the SARS-CoV-2 spike protein using a recombinant RBM Sepharose 4B affinity column (3 x 1 cm, inner diameter) (a) Evaluation of absorbance at 280 nm of the eluate different fractions, highlighted in green the eluate fractions used to concentrate polyclonal antibodies (b) In-house ELISA showing the specificity of purified antibodies to the multiepitope Dx-SARS-RBD containing RBM fragments. Figure S3: Microscale thermophoresis (MST) traces of anti-RBM antibodies binding to different concentrations of S1WT (red) and S2WT (green) peptides by MST. Relative Fluorescence (RF) between the bound and unbound states was determined over 21 seconds with 20 seconds of MST-on time for evaluation. The blue bar indicates the ΔRF before the temperature gradient was applied, whereas the red bar shows the ΔRF during the thermophoresis. The amount of NT.647-labeled antibodies was kept constant for interaction experiments, while the concentration of unlabeled peptides varied from 0.5 µg/mL to 0.12 ng/mL. The assay was performed in PBS containing 0.05% Tween- 20, and after a short incubation period, the samples were analyzed in standard glass MST NT.115 capillaries. Table S1: One-dose regime AstraZeneca-Oxford vaccinated serum information. Table S2: Heterologous booster dose vaccinated serum information. Figure S4: IgG subclass neutralizing response against RBM peptides in vaccinated sera. Subclass Immunoglobulin subclasses reactivity of RBM peptides S1WT, S2WT, S8WT, S8BA1, and S8BA5 using a cohort of vaccinated individuals positives for IgG and with the first dose of Oxford-AstraZeneca (n=12) and booster doses. For analysis purposes, multiple comparisons were made using Tukey's multiple comparisons test, where a p < 0.05 was considered a significant difference (* = p < 0.05).
Author Contributions
Conceptualization, S.G.D-S.; G.C.L.; C.M.M.; methodology, M.E.S.M.; L.R.G.; J.P.R. S.C.; P.N-P. software, M.E.S.M.; G.C.L.; D.W.P.; formal analysis and data curation, G.C.L.; writing—original draft preparation, M.E.S.M; G.C.L.; writing—review and editing, G.C.L.; D.W.P. and S.G.D-S.; project administration, S.G.D.-S; C.M.M. funding acquisition, C.M.M; S..G.D-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Carlos Chagas Filho Foundation for Research Support of the State of Rio de Janeiro/FAPERJ (#110.198-13; #SEI21078002021; #SEI26000306611722021), Brazilian Council for Scientific Research (CNPq, #467.488/2014-2 and #301744/2019-0) and FIOCRUZ (INOVA-FIOCRUZ #VPPIS-005-FIO-20-2-51). Funding was also provided by FAPERJ (#210.003/2018) through the National Institutes of Science and Technology Program (INCT-IDPN) to C.M.M.
Institutional Review Board Statement
The study was approved by the Human Research Ethical Committee of the Oswaldo Cruz Institute/FIOCRUZ (CAAE:49971421.8.0000.5248), the University of Estacio de Sá (CAAE: 3309 0820.8. 0000.5284) and UNIGRANRIO (CAAE: 21362220.1.0000.5283) study center ethics committee and conducted under good clinical practice and applicable regulatory requirements, including the Declaration of Helsinki.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
FIOCRUZ/Biomanguinhos (Rio de Janeiro, Brazil) for the purification of the peptides. P.N-P. and G.C.L. are post doc fellows from the INOVA-FIOCRUZ Program and L.R.G. from FAPERJ. M.E.M. and J.P.R.S.C. are CAPES/DSc fellows from the Post-Graduation Program on Parasitic Biology from FIOCRUZ and Science and Biotechnology, Federal Fluminense University, respectively.
Conflicts of Interest
The authors declare no dispute involving the research reported. The funding agencies had no role in the study design, data collection, data analysis, publication decision, or manuscript preparation.
References
- Pal: M.; Berhanu, G.; Desalegn, C.; Kandi, V. Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2): An Update. Cureus 2020, e7423. [CrossRef]
- Lenharo, M. WHO declares end to COVID-19’s emergency phase. Nature 2023, d41586-023-01559-z. [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat Rev Microbiol 2021, 19, 409–424. [CrossRef]
- Liu, L.; Iketani, S.; Guo, Y.; Chan, J.F.-W.; Wang, M.; Liu, L.; Luo, Y.; Chu, H.; Huang, Y.; Nair, M.S.; et al. Striking antibody evasion manifested by the omicron variant of SARS-CoV-2. Nature 2022, 602, 676–681. [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A Neutralizing human antibody binds to the N-terminal domain of the spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [CrossRef]
- Huang, Q.; Han, X.; Yan, J. Structure-based neutralizing mechanisms for SARS-CoV-2 antibodies. Emerg Microbes Infect 2022, 11, 2412–2422. [CrossRef]
- Piccoli, L.; Park, Y.-J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping neutralizing and immunodominant sites on the SARS-CoV-2 Spike receptor-binding domain by structure-guided high-resolution serology. Cell 2020, 183, 1024-1042.e21. [CrossRef]
- Deshpande, A.; Harris, B.D.; Martinez-Sobrido, L.; Kobie, J.J.; Walter, M.R. Epitope classification and RBD binding properties of neutralizing antibodies against SARS-CoV-2 variants of concern. Front Immuno 2021, 12, 691715. [CrossRef]
- Gasser, R.; Cloutier, M.; Prévost, J.; Fink, C.; Ducas, É.; Ding, S.; Dussault, N.; Landry, P.; Tremblay, T.; Laforce-Lavoie, A.; et al. Major role of IgM in the neutralizing activity of convalescent plasma against SARS-CoV-2. Cell Rep 2021, 34, 108790. [CrossRef]
- Long, Q.-X.; Liu, B.-Z.; Deng, H.-J.; Wu, G.-C.; Deng, K.; Chen, Y.-K.; Liao, P.; Qiu, J.-F.; Lin, Y.; Cai, X.-F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat Med 2020, 26, 845–848. [CrossRef]
- Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J Allergy Clin Immunol 2010, 125, S41–S52. [CrossRef]
- Ferrante, A.; Beard, L.J.; Feldman, R.G. IgG subclass distribution of antibodies to bacterial and viral antigens. Ped Infect Dis J 1990, 9, 516.
- Korobova, Z.R.; Zueva, E.V.; Arsentieva, N.A.; Batsunov, O.K.; Liubimova, N.E.; Khamitova, I.V.; Kuznetsova, R.N.; Rubinstein, A.A.; Savin, T.V.; Stanevich, O.V.; et al. Changes in anti-SARS-CoV-2 IgG subclasses over time and in association with disease severity. Viruses 2022, 14, 941. [CrossRef]
- Rubio-Casillas, A.; Redwan, E.M.; Uversky, V.N. Does SARS-CoV-2 induce IgG4 synthesis to evade the immune system? Biomolecules 2023, 13, 1338. [CrossRef]
- Almanza, G.; Clark, A.E.; Kouznetsova, V.; Olmedillas, E.; Castro, A.; Tsigelny, I.F.; Wu, Y.; Gao, G.F.; Leibel, S.L.; Bray, W.; et al. Structure-selected RBM immunogens prime polyclonal memory responses that neutralize SARS-CoV-2 variants of concern. PLoS Pathog 2022, 18, e1010686. [CrossRef]
- Pratesi, F.; Errante, F.; Pacini, L.; Peña-Moreno, I.C.; Quiceno, S.; Carotenuto, A.; Balam, S.; Konaté, D.; Diakité, M.M.; Aréva-lo-Herrera, M.; et al. A SARS–CoV-2 spike receptor binding motif peptide induces anti-spike antibodies in mice and is recognized by COVID-19 patients. Front. Immunol 2022, 13, 879946. [CrossRef]
- Chen, Y.; Zhao, X.; Zhou, H.; Zhu, H.; Jiang, S.; Wang, P. Broadly neutralizing antibodies to SARS-CoV-2 and other human coronaviruses. Nat Rev Immunol 2023, 23, 189–199. [CrossRef]
- Liu, Y.; Liu, J.; Plante, K.S.; Plante, J.A.; Xie, X.; Zhang, X.; Ku, Z.; An, Z.; Scharton, D.; Schindewolf, C.; et al. The N501Y spike substitution enhances SARS-CoV-2 transmission. Nature 2022, 602, 294-299. [CrossRef]
- Gattinger, P.; Niespodziana, K.; Stiasny, K.; Sahanic, S.; Tulaeva, I.; Borochova, K.; Dorofeeva, Y.; Schlederer, T., Sonnweber, T.; Hofer, G.; et al. Neutralization of SARS-CoV-2 requires antibodies against conformational receptor-binding domain epitopes. Allergy 2022, 77, 230–242. [CrossRef]
- Bachmann, M.F.; Mohsen, M.O.; Zha, L.; Vogel, M.; Speiser, D.E. SARS-CoV-2 Structural features may explain limited neutralizing-antibody responses. npj Vaccines (Basel) 2021, 6, 1–5. [CrossRef]
- De-Simone, S.G.; Gomes, L.R.; Napoleão-Pêgo, P.; Lechuga, G.C.; de Pina, J.S.; da Silva, F.R. Epitope mapping of the diphtheria toxin and development of an ELISA-specific diagnostic assay. Vaccines (Basel) 2021, 9, 313. [CrossRef]
- Gomes, L.R.; Durans, A.M.; Napoleão-Pêgo, P.; Waterman, J.A.; Freitas, M.S.; De Sá, N.B.R.; Pereira, L.V.; Furtado, J.S.; Aquino, R.G.; Machado, M.C.R.; et al. Multiepitope proteins for the differential detection of IgG antibodies against RBD of the spike protein and non-RBD regions of SARS-CoV-2. Vaccines (Basel) 2021, 9, 986. [CrossRef]
- Lechuga, G.C.; Napoleão-Pêgo, P.; Bottino, C.C.G.; Pinho, R.T.; Provance-Jr, D.W.; De-Simone, S.G. Trypanosoma cruzi presenilin-like transmembrane aspartyl protease: characterization and cellular localization. Biomolecules 2020, 10, 1564. [CrossRef]
- De-Simone, S.G.; Nascimento, H.J.; Prado, I.C.; Aguiar, A.S.; Melgarejo, A.R.; Pina, J.L.S.; Ferreira, P.F.; Provance, D.W. Puri-fication of equine IgG3 by lectin affinity and an interaction analysis via microscale thermophoresis. Anal Biochem 2018, 561–562, 27–31. [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat Med 2021, 27, 1205–1211. [CrossRef]
- Bachmann, M.F.; Mohsen, M.O.; Zha, L.; Vogel, M.; Speiser, D.E. SARS-CoV-2 Structural features may explain limited neutralizing-antibody responses. npj Vaccines (Basel) 2021, 6, 1–5. [CrossRef]
- Li, Y.; Lai, D.; Zhang, H.; Jiang, H.; Tian, X.; Ma, M.; Qi, H.; Meng, Q.; Guo, S.; Wu, Y.; et al. Linear epitopes of SARS-CoV-2 spike protein elicit neutralizing antibodies in COVID-19 patients. Cell Mol Immunol 2020, 17, 1095–1097. [CrossRef]
- Sun, C.; Xie, C.; Bu, G.-L.; Zhong, L.-Y.; Zeng, M.-S. Molecular characteristics, immune evasion, and impact of SARS-CoV-2 variants. Sig Transduct Target Ther 2022, 7, 202. [CrossRef]
- Lupala, C.S.; Ye, Y.; Chen, H.; Su, X.-D.; Liu, H. Mutations on RBD of SARS-CoV-2 Omicron variant result in stronger binding to human ACE2 receptor. Biochem Biophys Res Communi 2022, 590, 34–41. [CrossRef]
- Chen, W.-H.; Hotez, P.J.; Bottazzi, M.E. Potential for developing a SARS-CoV receptor-binding domain (RBD) recombinant protein as a heterologous human vaccine against coronavirus infectious disease (COVID)-19. Hum Vaccine Immunother 2020, 16, 1239–1242. [CrossRef]
- Liu, X.; Chang, X.; Rothen, D.; Derveni, M.; Krenger, P.; Roongta, S.; Wright, E.; Vogel, M.; Tars, K.; Mohsen, M.O.; et al. AP205 VLPs based on dimerized capsid proteins accommodate RBM domain of SARS-CoV-2 and serve as an attractive vaccine candidate. Vaccines (Basel) 2021, 9, 403. [CrossRef]
- Castro, A.; Ozturk, K.; Zanetti, M.; Carter, H. In silico analysis suggests less effective MHC-II Presentation of SARS-CoV-2 RBM Peptides: Implication for neutralizing antibody responses. PLoS ONE 2021, 16, e0246731. [CrossRef]
- Vainio, O.; Toivanen, P.; Toivanen, A. Major histocompatibility complex and cell cooperation. Poul Sci 1987, 66, 795–801. [CrossRef]
- Lani, R.; Senin, N.A.; AbuBakar, S.; Hassandarvish, P. Knowledge of SARS-CoV-2 epitopes and population HLA types is important in the design of COVID-19 vaccines. Vaccines (Basel) 2022, 10, 1606. [CrossRef]
- Bertinetto, F. E.; Magistroni, P.; Mazzola; G. A., Costa, C.; Elena, G.; Alizzi, S.; Scozzari, G.; Migliore, E.; Galassi, C.; Ciccone, G.; et al. The humoral and cellular response to mRNA SARS-CoV-2 vaccine is influenced by HLA polymorphisms. HLA 2023, 102, 301–315. [CrossRef]
- Mentzer, A. J.; Connor, D.; Bibi, S.; Chelysheva, I.; Clutterbuck, E.A.; Demissie, T.; Dinesh, T.; Edwards, N. J.; Felle, S.; Feng, S.; et al. Human leukocyte antigen alleles associate with COVID-19 vaccine immunogenicity and risk of breakthrough infection. Nat Med 2023, 29, 147–157. [CrossRef]
Figure 1.
Structural organization of Spike protein and receptor binding motif (RBM), phylogenetic analysis of VoCs, and interactions of nAbs to residues in RBM. (a) Organization of Spike protein domains and phylogenetic analysis of VoCs, highlighting mutations (yellow) in RBM. (b) Interactions of RBM residues (437-503) with ACE-2 and human nAbs. (c) The tridimensional model of the S protein trimer shows RBD (green), RBM (yellow), and the synthetic peptide S8WT (magenta) comprising the tip portion of RBM.
Figure 1.
Structural organization of Spike protein and receptor binding motif (RBM), phylogenetic analysis of VoCs, and interactions of nAbs to residues in RBM. (a) Organization of Spike protein domains and phylogenetic analysis of VoCs, highlighting mutations (yellow) in RBM. (b) Interactions of RBM residues (437-503) with ACE-2 and human nAbs. (c) The tridimensional model of the S protein trimer shows RBD (green), RBM (yellow), and the synthetic peptide S8WT (magenta) comprising the tip portion of RBM.

Figure 2.
RBM tip humoral response and affinity in vaccinated individuals. (a) In-house peptide-ELISAS performed with peptides S1WT, S2WT, and S3WT and a court of pre-pandemic sera (n=15), individuals vaccinated with the first dose of Oxford-AstraZeneca after 15 days (n=30) and booster heterologous doses (n=7). (b) Representative dos dose–responsive showing the binding of antibodies from vaccinated serum with peptides S1WT and S2WT, the Kd of 640.7 nM and 35 nM, respectively. A serial dilution of the peptide from 0.5 µg/mL–0.12 ng/mL was used. (c) Immunoglobulin classes reactivity of RBM peptides S8WT, S8B1, and S8B5 using the court of vaccinated individuals with the first dose of Oxford-AstraZeneca (n=30) and (d) booster doses (n=7). A Kruskal–Walli’s test was applied to identify statistical differences, followed by Dunn's multiple comparisons tests. A p < 0.05 was considered a significant difference. *p<0.05; **p<0.01 and ***p<0.001.
Figure 2.
RBM tip humoral response and affinity in vaccinated individuals. (a) In-house peptide-ELISAS performed with peptides S1WT, S2WT, and S3WT and a court of pre-pandemic sera (n=15), individuals vaccinated with the first dose of Oxford-AstraZeneca after 15 days (n=30) and booster heterologous doses (n=7). (b) Representative dos dose–responsive showing the binding of antibodies from vaccinated serum with peptides S1WT and S2WT, the Kd of 640.7 nM and 35 nM, respectively. A serial dilution of the peptide from 0.5 µg/mL–0.12 ng/mL was used. (c) Immunoglobulin classes reactivity of RBM peptides S8WT, S8B1, and S8B5 using the court of vaccinated individuals with the first dose of Oxford-AstraZeneca (n=30) and (d) booster doses (n=7). A Kruskal–Walli’s test was applied to identify statistical differences, followed by Dunn's multiple comparisons tests. A p < 0.05 was considered a significant difference. *p<0.05; **p<0.01 and ***p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



